Настоящее изобретение относится к методу вакцинации домашних птиц против кокцидиоза. В частности, изобретение относится к введению in ovo живых спорозоитов или мерозоитов видов Eimeria или их смесей в развивающиеся яйца домашней птицы с целью иммунизации вылупившихся птенцов против кокцидиоза.
Кокцидиоз является кишечным заболеванием домашних птиц, вызванным заражением внутриклеточными простейшими паразитами рода Eimeria. Кокцидиоз является паразитарным заболеванием домашних птиц, которое наносит наиболее ощутимый экономический ущерб. Установлено, что лечение кокцидиоза и потери из-за него обходятся птицеводству в сотни миллионов долларов ежегодно.
Известно, что уже с начала 1950-х годов предпринимались различные попытки вакцинировать домашних птиц против кокцидиоза. Применяющиеся в настоящее время методы вакцинации включают введение живых ооцист Eimeria птицам с кормом или водой. Однако эти методы неудобны и неэффективны, поскольку не все птицы получают предполагаемую дозу ооцист и многие либо остаются защищены вакциной, либо получают патогенное заражение.
В J. M. Sharma и B.R. Burmester, в Avian Dis., 26:134-149, 1981, авторы констатируют, что куры, вакцинированные in ovo вирусом герпеса индейки, выработали иммунитет против последующего контрольного заражения вирусом болезни Марека. В европейской патентной публикации N 291173 описывается процесс иммунизации, в котором нереплицирующий иммуноген вводят in ovo. Иммуногенами, указанными конкретно в европейском патенте, являются полученный методом генной инженерии антиген Eimeria и экстракт ооцисты Eimeria. В Европейском патенте подчеркивается, что живые стадии паразита, такие как те, которые используются в методе вакцинации, заявленном здесь, исключаются.
Предлагаемый метод вакцинации заключается в введении in ovo живых спорозоитов или мерозоитов Eimeria или их смеси в развивающиеся яйца домашних птиц. В известной литературе по данному вопросу содержится указание, что такой метод вакцинации будет неэффективным in ovo и его следует применять после вылупления птенцов. В Т. К. Jeffers и G.E. Wagenbach в J. Parasit. 56/4/: 656-662, 1970, сообщается, что инъекция in ovo спорозоитов Е. tenella на 10 день инкубации не дала какой-либо значительной иммунологической защиты против последующего контрольного заражения ооцистами Е. tenella. Фактически они констатировали, что птенцы, не прошедшие такое лечение, имели лучшие показатели выживания против последующего контрольного заражения ооцистами Е. tenella, чем птенцы, которым спорозоиты вводились in ovo. B K.L. Watkins и др., Proc. VI, International Coccidiosis Conf., Abstract Е1-2, Ontario, Canada, 1993, авторы описали инокуляцию in ovo живыми спороцистами и спорулированными ооцистами Е. maxima и пришли к выводу, что их исследование не дало доказательств того, что введение in ovo обеспечивает защиту против последующего кокцидиального контрольного заражения ооцистами Е. maxima через 10 дней после вылупления. Watkins и др. сделали вывод, что существенная иммунологическая защита обеспечивается, когда инокуляция делается вскоре после вылупления птенцов, а не in ovo. В противоположность этому утверждению метод вакцинации in ovo по настоящему изобретению неожиданно обеспечивает иммунитет, который защищает вылупившихся птиц против последующего контрольного заражения кокцидиозом.
Настоящее изобретение, именуемое здесь как "данный метод вакцинации", относится к способу вакцинирования домашней птицы против кокцидиоза, включающему введение in ovo во время последней четверти периода инкубации эффективной иммунизирующей дозы живых спорозоитов или мерозоитов Eimeria или их смеси.
Термин "домашняя птица /ы/" здесь, если не указано иначе, включает кур, индеек, уток, охотничье-промысловую птицу /включая, (но не ограничиваясь) перепелов, фазанов и гусей/ и бескилевых /включая (но не ограничиваясь ими) страусов/;
термин "in ovo", здесь, если не указано иначе, означает "в яйцо домашней птицы", содержащее живой развивающийся эмбрион.
Термин "введение in ovo" или "in ovo введение", здесь, если не указано иначе, означает введение описанной здесь вакцины в яйцо домашней птицы, содержащее живой развивающийся эмбрион, любым путем проникновения через скорлупу яйца и введения вакцины. Такие способы введения включают, но не ограничены до инъекции вакцины.
Термин "последняя четверть срока инкубации" здесь, если не указано иначе, означает последнюю четверть инкубации развивающегося яйца домашней птицы.
Термин "Eimeria" здесь, если не указано иначе, означает один или несколько видов рода Eimeria, которые заражают домашних птиц. Такие виды Eimeria включают те, которые выявлены у кур, это E. tenella, E. acervulina, E. maxima, E. necatrix, E. mitis, E. praecox, и E. brunetti, те, которые выявлены у индеек, такие как E. meleagrimitis, E. adenoeides, E. gallopavonis, E. dispersa, E. meleagridis, E. innocua и E. subrotunda, а также виды Eimeria, которые заражают других домашних птиц, упомянутых выше. Термин "Eimeria" включает также все штаммы указанных выше видов Eimeria, включающие, но не ограниченные до преждевременно развившихся и ослабленных штаммов, которые включают штаммы, подвергшиеся облучению или какой-либо другой обработке, в результате чего они не проходят стадии полного развития. Термин Eimeria включает также любые вновь открытые штаммы или виды Eimeria, которые заражают домашних птиц, как указывалось выше.
Термины "спорозоиты", "спороцисты", "ооцисты" и "мерозоиты" здесь, если не указано иначе, означают живых спорозоитов, спороцист, ооцист и мерозоитов Eimeria.
Термин "эффективная иммунизирующая доза" здесь, если не указано иначе, означает количество спорозоитов, или мерозоитов, или в случае смешивания количество спорозоитов и мерозоитов, достаточное, чтобы обеспечить иммунологическую защиту у вылупившихся птиц, которая сильнее врожденного иммунитета неиммунизированных птиц. Т.о. здесь термины "иммунизировать" и "вакцинировать" являются синонимами и заменяют друг друга.
Предпочтительная доза, которая вводится в соответствии с методом изобретения, содержит 10 - 106 спорозоитов, или мерозоитов, или их смесь, в которой общее количество указанных спорозоитов и мерозоитов составляет от 10 до 106.
Наиболее предпочтительная доза содержит от 103 до 106 спорозоитов, или мерозоитов, или их смесь, в которой общее количество указанных спорозоитов и мерозоитов колеблется от 103 до 106.
Другая предпочтительная доза включает 102 - 105 спорозоитов, или мерозоитов, или их смесь, в которой их общее количество составляет от 102 до 105.
Метод по изобретению наиболее предпочтителен для вакцинации кур.
Лучше, если доза для введения in ovo в яйца кур будет включать спорозоиты, или мерозоиты, либо их смесь, двух или нескольких видов Eimeria, выбранных из группы, состоящей из E. tenella, E. acervulina, E. maxima, E. necatrix, E. mitis, E. praecox, E. brunetti. Предпочтительно доза включает от 10 до 106 спорозоитов или мерозоитов или их смесь для каждого вида, который входит в дозу.
Данный метод изобретения также хорош для вакцинации индеек.
Предпочтительная доза для введения in ovo в яйца индеек включает спорозоиты, или мерозоиты, или их смесь, двух или нескольких видов Eimeria, выбранных из группы, состоящей из E. meleagrimitis, E. adenoeides, E. gallopavonis, E. dispersa, E. meleagridis, E. innocua и E. subrotunda. Доза предпочтительно включает от 10 до 106 спорозоитов, или мерозоитов, или их смесь для каждого вида, который входит в дозу.
Другими предпочтительными одомашненными птицами для вакцинации в соответствии с методом изобретения являются охотничье-промысловые птицы, утки и бескилевые.
Метод изобретения включает далее введение in ovo иммунного стимулятора в комбинации с методом вакцинации в любое время в течение периода инкубации.
Иммунный стимулятор лучше вводить одновременно с введением in ovo дозы спорозоитов, или мерозоитов, либо их смеси в последней четверти инкубационного периода.
Настоящий метод вакцинации включает введение in ovo в последней четверти инкубации живых спорозоитов, или мерозоитов, либо их смеси в яйца домашних птиц. Если это куры, введение in ovo лучше делать на 15 - 20 дни инкубации, предпочтительно на 18 день. Если это индейки, введение in ovo делают на 21 - 26 дни периода инкубации.
При этом можно использовать любой приемлемый способ введения in ovo. Лучше данную вакцину вводить инъекцией. В соответствии с одним способом инъекции на большем конце яйца в яичной скорлупе делают отверстие иглой номер 18 с тем, чтобы открыть воздушную полость яйца. Иглу 22 номера длиной 1.0-1.5 дюйма (2.5-3.8 см), надетую на шприц соответствующего объема (1 - 3 мл), можно ввести через отверстие и через мембрану воздушной ячейки. Нужное количество спорозоитов, или мерозоитов, либо их соответствующее количество в смеси суспендируют в подходящем жидком носителе, например, 10 - 500 мкл физраствора с фосфатным буфером и затем вводят в яйцо. Нужный объем зависит от размера яйца, естественно, яйца страуса потребуют большего объема, чем яйца кур. Участок инъекции может быть любым участком яйца или эмбриона. Инъекцию лучше делать аксиально через центр большого конца яйца в амнион.
В настоящем методе вакцинации возможно применение автоматизированной системы инъекции яйца. Такие системы описаны в патентах США N 4681063, 4040388, 4469047 и 4593646, которые включены в качестве ссылки. Другие подходящие методы инъекций известны специалистам данной области.
Ооцисты можно получить любым из нескольких методов, известных специалистам. Такие методы включают методы, описанные J.E. Ryley и др. в Parasitology, 73:311 - 326, 1976 и P.L.Long и др., Folia veterinaria Latina, У1 # 3, 201 - 217, 1976, которые также включены для ссылки. По одному из методов, промышленных бройлерных цыплят возрастом приблизительно в 2 недели, инфицируют нужными видами Eimeria оральным введением соответствующей дозы спорулированных ооцист. Например, типичная доза для E. tenella составляет 200,000 спорулированных ооцист/птицу. Затем следуют хорошо известные процедуры сбора и очистки ооцист из инфицированных птиц. Для большинства видов Eimeria фекалии от инфицированных птиц собирают на 5-7 дни после заражения, их смешивают и фильтруют для удаления остатков органических веществ, затем подвергают центрифугированию со скоростью, достаточной, чтобы получить осадок остающегося фекального материала. Для Е. tenella используют аналогичную процедуру, за исключением того, что содержимое из слепой кишки берут на 6 день после заражения. Осадок вновь суспендируют в насыщенном солевом растворе, в котором ооцисты всплывают и большую часть примесей органических остатков можно удалить центрифугированием. Затем суспензию ооцист разбавляют до более низкой солевой концентрации. Ооцисты промывают несколько раз, чтобы удалить соль, и вновь суспендируют в растворе бихромата калия /2,5% вес/об./. Суспензию ооцист инкубируют при 29oC при встряхивании /например, 140 об/мин/ в течение приблизительно 72 часов для индуцирования споруляции ооцист. В другом варианте ооцисты можно обработать гипохлоритом натрия, а затем спорулировать. Число спорулированных ооцист на мл определяют прямым подсчетом, используя гемоцитометр, и культуру хранят до применения, желательно на холоде.
Чтобы получить спороцисты, бихромат калия удаляют из суспензии ооцист, описанной выше, повторным промыванием ооцист, которое предполагает сбор ооцист центрифугированием и повторное суспендирование в деионизированной или дистиллированной воде. Когда бихромат удален, о чем можно судить по отсутствию желтовато-оранжевой окраски, суспензию ооцист смешивают с равным объемом гипохлорита натрия /отбеливатель/ и инкубируют при комнатной температуре 15 минут. Потом отбеливатель удаляют повторными промываниями и ооцисты вновь суспендируют в физрастворе или деионизированной воде. Для высвобождения спороцист ооцисты разрушают, используя множество известных способов. Например, ооцисты можно разрушить и высвободить спороцисты, смешивая ооцисты со стеклянными шариками диаметром 1-4 мм встряхиванием рукой, вихревой мешалкой или встряхиванием инкубатора либо с применением ручного гомогенизатора. Неразрушенные ооцисты и их оболочки можно отделить от освобожденных спороцист дифференциальным центрифугированием в 50% Percoll (от Pharmacia Biotech) или 1 М сахарозе, как описано у Dulski и др., Avian Diseases, 32:235-239, 1988.
Чтобы подготовить спорозоиты или препарат, богатый спорозоитами, для применения в соответствии с изобретением, спорозоиты извлекают (эксцистируют) из препарата спороцист, описанного выше. По одной процедуре, спороцисты, полученные как описано выше, осаждают центрифугированием, повторно суспендируют в буфере для эксцистирования /0.5% тауродеоксихолевая кислота и 0.25% трипсина в физрастворе с фосфатным буфером, pH 8.0/ и инкубируют со встряхиванием в течение одного часа при 41oC. В пробе полученной суспензии подсчитывают количество спорозоитов, спорозоиты промывают один раз для удаления буфера эксцистирования и вновь суспендируют в физрастворе с фосфатным буфером с нужной концентрацией для введения in ovo. Этот препарат содержит спорозоиты, спороцисты и ооцисты и без дальнейшей очистки может применяться в настоящем методе вакцинации. Очищенные спорозоиты, отделенные от спороцист и ооцист, можно получить анионообменной хроматографией на DE-52, как описано у D.M. Schmatz и др. в J. Protozool 31:181-183, 1984, которая также включена сюда для ссылки. Доза спорозоитов для применения в данном методе вакцинации будет различна, в зависимости от вида вакцинируемых птиц и применяемых в вакцине видов Eimeria. В общем, доза может быть в пределах от 10 до 106 спорозоитов на яйцо. Предпочтительно пределы дозы составляют от 10 до 105 спорозоитов на яйцо, а еще лучше - от 102 до 105 спорозоитов на яйцо.
Мерозоиты можно получить различными методами, известными специалистами. По одному из них спорозоитами инфицируют первичные почечные клетки цыпленка /PCK/, которые выращивают в культуре в виде агрегатов клеток, с использованием модификации метода, описанного у D.J. Doran, в J. Parasit., 57:891-900, 1971, который приведен здесь для ссылки. PCK клетки выращивают при 40oС в 3% СО2 в модифицированной RK2 среде - DMEM/F12 (DMEM - модифицированная по способу Дульбекко Среда Игла) с L-глутамином и 15 мМ HEPES (N-2-гидроксиэтилпиперазин гидроксиэтилпиперазин -N'-2-этансульфоновая кислота), дополненной фетальной околоплодной коровьей сывороткой, пенициллином-стрептомицином, 15 мМ бикарбоната натрия, 10 нг/мл эпидермального фактора роста, 5 мкг/мл инсулина, 5 мкг/мл трансферрина, 5 нг/мл селенистой кислоты и 0.01 мкМ гидрокортизона в HCl, как описано у S.D. Chung и др. в J. Cell Biol., 95:118-126, 1982. Первичные почечные клетки готовят из почек 2 - 3-недельных цыплят измельчением почки и затем обработкой ткани при 37oС 0.02 мг/мл коллагеназы /от Worthington Biochemical Corp., Freehold, NJ/ в физрастворе с фосфатным буфером в несколько циклов. Клеточные агрегаты в надосадочном слое промывают, вновь суспендируют в модифицированной RK2 среде, содержащей 5% фетальную коровью сыворотку, и применяют для высевания тканевых культур с плотностью 105 агрегатов на 1 см2. Клетки PCK инкубируют 18 часов при 40oС в 3% СО2, а потом инфицируют 4 • 105 спорозоитов/см2. Инфицированные культуры выращивают в модифицированной RК2 среде, содержащей 2% фетальную коровью сыворотку. После 24 часов инкубации, времени, достаточного для инвазии, неинвазированные спорозоиты удаляют взбалтыванием колбы и удалением среды культуры. Клеточный слой один раз промывают модифицированной RК2 средой, содержащей 2% фетальную коровью сыворотку, и культуральную среду вновь удаляют. Добавляют свежую RК2 среду и культуры инкубируют еще 48 - 54 часа до тех пор, пока мерозоиты не высвободятся в культуральную среду.
Очистку мерозоитов с целью удаления остатков клеток хозяина можно провести любыми известными специалистам методами. В соответствии с одним из них, как описано у J.A. Olson, Antimicrob. Agents Chemother., 34:1435-1439, 1990, культуральную среду, содержащую высвобожденные мерозоиты, собирают и вращают при 450 оборотах в минуту 10 минут для концентрирования мерозоитов. Пластинки осадка, содержащего мерозоиты и органические остатки клеток хозяина, суспендируют в 0.1 М NaСl-0.05 М КСl-20% альбумине коровьей сыворотки и подают на анионообменную колонну DE-52, уравновешенную 75 мМ Трис-40 мМ NaH2PO4-86 мМ NаСl-100 мМ глюкозы при рН 8.2. Мерозоиты протекают через колонну. Собранные из колонны мерозоиты можно проверить на чистоту электронной микроскопией, как описано у A. Kilejian, в J. Biol. Chem., 249:4650-4655, 1974. В общем, доза мерозоитов может составлять от 10 до 106 мерозоитов на яйцо. Предпочтительно она колеблется от 10 до 105 мерозоитов на яйцо, а лучше, если она составляет 102 - 105 мерозоитов на яйцо. Если спорозоиты и мерозоиты смешивают, в общем, доза, включающая общее количество мерозоитов и спорозоитов, может составлять от 10 до 106 на яйцо. Лучше, если доза включает от 10 до 105 мерозоитов и спорозоитов на яйцо, а еще лучше - от 102 до 105 мерозоитов и спорозоитов на яйцо.
Спорозоиты, или мерозоиты, либо их смесь можно ввести in ovo в любой физиологически приемлемой среде. Их желательно суспендировать в физиологически сбалансированном физрастворе, таком как физраствор с фосфатным буфером. Выбранная среда может включать один или несколько суспендирующих агентов из ряда физиологически приемлемых гелей, желатинов, гидрозолей, целлюлоз или полисахаридных смол.
В настоящем методе вакцинации спорозоиты, или мерозоиты, либо их смесь двух или нескольких видов Eimeria лучше вводить in ovo одновременно. При этом методе вакцинации спорозоиты, или мерозоиты, либо их смесь, всех идентифицированных видов Eimeria, которые заражают конкретную домашнюю птицу, такую как курица, можно ввести in ovo одновременно или серийно, с тем чтобы обеспечить иммунологическую защиту против всех видов.
В методе вакцинации настоящего изобретения можно использовать иммунные стимуляторы. Подходящие иммунные стимуляторы включают, но не ограничиваются следующими: цитокины факторы роста, химокины, надосадочные слои клеточных культур лимфоцитов, моноцитов или клеток от лимфоидных органов, клеточных препаратов или клеточных экстрактов /Staphylococcus aureus или препаратов липополисахарида/, митогены или адъюванты, включающие низкомолекулярные фармацевтические средства. Иммунный стимулятор можно вводить in ovo в любое время в период инкубации. Иммунный стимулятор желательно вводить in ovo в среде, содержащей дозу спорозоитов или мерозоитов Eimeria или их смесь.
Эффективность настоящего изобретения в вакцинации против кокцидиоза иллюстрируется в следующих примерах. Каждую дозу вводили инъекцией in ovo в физиологически приемлемом растворе соли, как описано выше. Эффективность конкретного препарата определяли, наблюдая его действие на показателе выведения птенцов и вес вылупившихся птенцов, и после контрольного заражения - образование ооцист, привес и патогенность /показатель поражения/. Определения показателя поражения производили по протоколу, описанному у J.K. Johnson и W.M. Reid в Exp. Parasitol., 28:30-36, 1970, в соответствии с которым величина 0 соответствует отсутствию заболевания и величина 4 представляет максимальную патологию.
Пример 1.
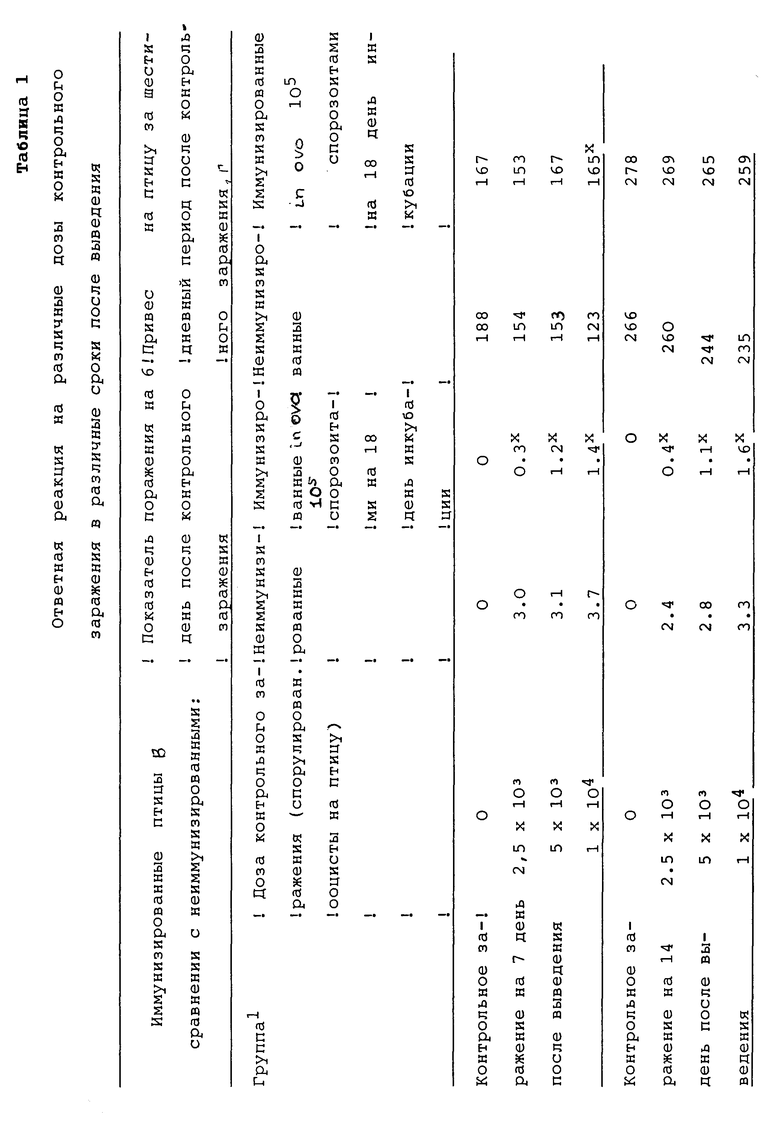
На 18-й день инкубации в яйца кур делали инъекцию препарата, содержащего 105 спорозоитов E. tenella на яйцо. Препарат не был очищен от спороцист и ооцист. В каждой дозе содержалось приблизительно 104 E. tenella спороцист и около 104 ооцист E. tenella. В качестве контроля служили яйца, в которые делали инъекции только физраствора с фосфатным буфером. В популяции птиц, обработанных спорозоитами, средний выход ооцист на 7 день после выведения цыплят составил 1,1 • 106 ооцист/птицу. Неиммунизированных птиц и птиц, обработанных спорозоитами, подвергали контрольному заражению различными дозами спорулированных ооцист E. tenella, введенных орально, на 7, 14 или 21 дни после выведения. Данные представлены в Таблице 1.
Данные в Таблице 1 ясно показывают, что иммунизированные птицы были менее подвержены заражению, чем их неиммунизированные выведенные сородичи, как указывают сниженные показатели поражений и улучшенные привесы у иммунизированных птиц. Данные также демонстрируют, что метод изобретения обеспечивает иммунитет цыплятам в относительно раннем возрасте /до семи дней после выведения/. Кроме того, данные показывают, что иммунитет сохраняется с ростом и развитием цыплят. Получение иммунитета цыплятами в раннем возрасте обеспечивает существенное преимущество для промышленности бройлерных цыплят, поскольку бройлеры, как правило, поступают в продажу по достижении 6 недель.
Пример 2.
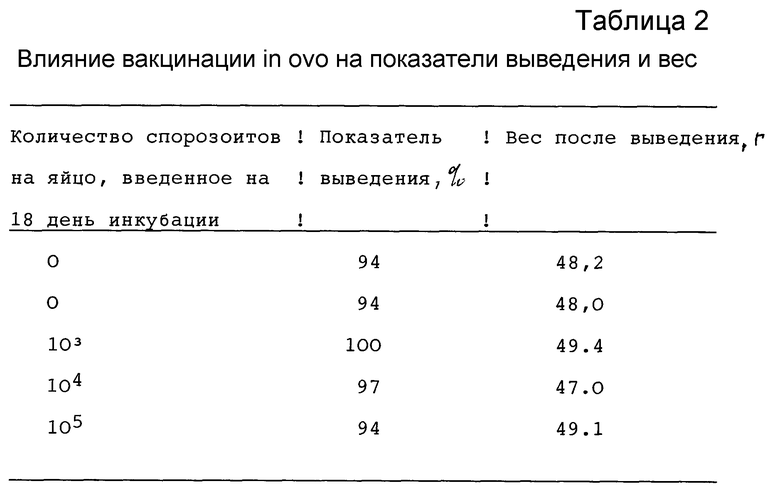
На 18 день инкубации в яйца кур делали инъекцию физраствора /контроль/ или препаратов, содержащих различные дозы спорозоитов Е. tenella как указано в Таблице 2, в которой представлены результаты, предваряющие контрольное заражение. Примененный препарат спорозоитов содержал на каждую дозу 62% спорозоитов, 9% спороцист и 29% ооцист. Каждая доза, содержащая 105 спорозоитов, включала в целом 1.6 • 105 различных стадий паразита /спорозоитов, спороцист и ооцист/.
Данные в Таблице 2 показывают, что цыплята, выведенные из яиц, в которые вводили живые спорозоиты, по своим показателям выведения и веса были фактически идентичны своим неиммунизированным сородичам. На 14 день после выведения цыплят подвергали контрольному заражению 1.25 • 104 спорулированных ооцист E. tenella на птицу оральным введением. Результаты ответной реакции на контрольное заражение представлены в Таблице 3.
Данные в Таблице 3 показывают, что по каждому параметру /привес, показатель поражения, выход ооцист/ цыплята, выведенные из яиц, в которые вводили различные дозы препарата спорозоитов, показали наличие иммунитета. В сравнении с контрольными птицами, которые обрабатывались только физраствором и которых подвергали контрольному заражению, птицы, иммунизированные препаратом спорозоитов, показали большой привес и сниженный показатель поражения. Кроме того, птицы, иммунизированные in ovo препаратом спорозоитов, имели меньший выход ооцист, чем контрольные птицы после контрольного заражения, указывая на то, что у иммунизированных птиц заражение было менее значительным.


Изобретение относится к способу вакцинирования птицы против кокцидиоза, включающему введение in ovo эффективной иммунизирующей дозы живых спорозоитов, или мерозоитов, или их смеси. В предпочтительном варианте птицей, которую подвергают вакцинации, является курица или индейка. Способ обеспечивает получение иммунитета против кокцидиоза у вылупляющихся цыплят и повышение их привесов. 9 з.п. ф-лы, 3 табл.
| СПОСОБ ИЗМЕРЕНИЯ НАПРЯЖЕННОС МАГНИТНОГО ПОЛЯ | 0 |
|
SU291173A1 |
| Watkins K.L | |||
| et al | |||
| Proc | |||
| VI, International Coccidiosis Conf., Abstract E1-2, Ontario, Canada, 1993 | |||
| US 4639372 А, 27.01.87 | |||
| US 5055292 А, 08.10.91 | |||
| Шевцов А.А | |||
| и др | |||
| Паразитология | |||
| - М.: Агропромиздат, 1985, с | |||
| Клапан | 1919 |
|
SU357A1 |

