Изобретение относится к профилактике и лечению желудочной инфекции у млекопитающих, в том числе у человека. Более конкретно данное изобретение касается вакцины, пригодной для применения для профилактики и лечения инфекции Helicobacter у млекопитающих, в том числе у человека, и способа лечения больных, страдающих от желудочной инфекции, ее последствий, таких, как хронический гастрит или пептическая язва, и профилактики рака желудка.
Инфекция микроорганизмов Helicobacter эпителия желудка человека, вызывающая гастрит, является основным фактором в развитии пептических язв и лимфомы желудка и может быть фактором риска для развития рака желудка /1 - 3/. Наиболее частным инфекционным агентом является Helicobater pylori, за ним следует с гораздо меньшей частотой Helicobater heilmanii. H.pylori представляет собой тонкий S-образный грамотрицательный микроорганизм, который рутинно получают из желудочных биопсий взрослых и детей с гистологическим подтверждением гастрита или пептической язвы. Доказательство причинной связи между H. pylori и гастродуоденальным заболеванием получено в исследованиях, проведенных на добровольцах, больных с язвами и раком желудка, гнотобиотических свиньях и не содержащих возбудителей инфекций грызунах. Что касается этиологии, постулаты Коха были удовлетворены развитием гистологически подтвержденного гастрита в ранее не инфицированных индивидуумах после поглощения жизнеспособных микроорганизмов /4 - 11/ и лечением для ликвидации H. pylori с исчезновением гастрита, у больных с пептической язвой, снижение скорости рецидивов /12/.
Вопреки чувствительности in vitro ко многим антимикробным агентам очень часто трудно добиться ликвидации установленных инфекций H. pylori при помощи антимикробных агентов in vitro /13/. Микроорганизм обнаружен внутри слизистой оболочки, покрывающей эпителей желудка, и в желудочных ямках. Эти местоположения, по-видимому, не позволяют достичь адекватных антимикробных уровней, даже при пероральном введении антибиотиков в высокой дозе. В настоящее время большая часть специалистов рекомендует "тройную терапию", а именно соль висмута в сочетании с лекарственными средствами, такими, как тетрациклин и метронидазол, в течение 2 - 4 недель. Однако эффективность этого или других химиотерапевтических режимов остается субоптимальной. Кроме того, такое лечение может вызывать серьезные побочные реакции.
В настоящее время мало известно относительно роли иммунной системы слизистой оболочки в желудке. Распределение продуцирующих иммуноглобулин /Ig/ клеток в нормальном желудочном пространстве указывает, что продуцирующие IgA клетки плазмы составляют 80% всей популяции клеток плазмы. Кроме того, число продуцирующих IgA клеток плазмы, присутствующих в желудочном пространстве, сравнимо с другими покрытыми слизью мембранами /14,15/. Ряд исследований на человеке /16/ и животных моделях /8, 10/ продемонстрировали специфические IgG и IgA ответные реакции в сыворотке и в желудочных секрециях в ответ на инфекцию Helicobacter. Однако наблюдение, что инфекция H. pylori сохраняется в виде хронической инфекции годами вопреки индукции местного и общего иммунного ответа, не вдохновляет на развитие стратегий иммунизации.
Lee et al. сообщили о способности инфицировать не содержащих возбудителей заболеваний грызунов бактерии Helicobacter felis, близко родственной H. pylori, и воспроизводимо документировали гистологический гастрит /9, 10/. С тех пор эта пара бактерия-хозяин была принята в качестве хорошей модели для исследования вызываемого Helicobacter гастрита и инициирующих его факторов /17/. Czinn et al. показали, что повторяемая пероральная иммунизация неочищенным лизатом H. pylori с холерным токсином в качестве адьюванта индуцирует сильный желудочно-кишечный у мышей и хорьков в виде образования антител против H. pylori /13/. Кроме того, Chen et al и Czinn et al недавно сообщил, что пероральная иммунизация неочищенным лизатом H. felis индуцировала защиту против инфекции H. felis у мышей /21, 22/. Однако точная природа антигена /антигенов/, ответственных за индикацию этой защиты, не была определена, и ни в одной из информаций нет предположений о том, что защитный антиген /антигены/ H. felis, которые индуцируют защиту против этого патогена, будут индуцировать перекрестно-реактивную защиту против другого вида Helicobacter.
Мы впервые показали, что H. pylori и H. felis, разрушение ультразвуком, вызывают образование антител и некоторые из антител против H. pylori могут давать перекрестную реакцию с H. felis и наоборот /24, 25/. Основа этой перекрестной реактивности неизвестна.
На основе гемологии, существующей между различными аминокислотными последовательностями различных известных уреаз, было сделано предположение, что уреаза могла бы применяться в качестве вакцины против H. pylori /26/. Тем не менее кросс-реактивность не является правилом. Guo и Liu показали несколько лет назад, что уреазы Proteus vulgaris, Proteus mirabilis и Providencia rettgeri обнаруживают кросс-реактивность друг с другом, тогда как уреазы канавалии мечевидной и Morganella morganii иммуннологически отличаются перекрестная реактивность уреазы H. pylori с другими уреазами других Helicobacter была разумным требованием, не было данных, демонстрирующих, что она имеет место, до тех пор, пока мы не показали, что часть моноклональных антител против H. felis прекрасно реагировали с уреазой H.pylori /25/. В дальнейшем J.Pappo показал, что мыши, инфицированные H.felies, продуцируют антитела, которые прекрасно реагируют с уреазой H.pylori, но не с уреазой канавалии мечевидной (J.Pappo, неопубликованные данные, 1993).
Применение уреазы H. pylori или родственных уреаз в качестве вакцины против инфекции H.pylori было предложено ранее A.Labigne в ЕРО 367 644 /28/. Однако эта заявка не содержит доказательства вакцинации какого-либо млекопитающего против какой-либо инфекции Helicobacter при помощи уреазы.
Более того, в то время как гомология последовательности с другими бактериальными уреазами может поддерживать применение уреазы в качестве кандидата на применение в вакцине против H.pylori, текущие знания об инфекции H.pylori у человека не поддерживают возможность применение уреазы. Во-первых, вопреки факту, что инфицированные индивидуумы часто дают сильную ответную реакцию с образованием антител на уреазу, иммунный ответ в виде антител против уреазы не приводит к удалению бактерий или контролю инфекции. Во-вторых, H.pylori способна выводить уреазу из клетки и сбрасывать ее с поверхности клетки /19, 20/. Поэтому уреаза может не представлять собой подходящую мишень для развития защитного иммунного ответа слизистой оболочки. Действительно считают, что иммунная защита слизистой оболочки опосредована в основном секреторным IgA, агглютинирующая активность которого будет ослаблена, если узнаваемый антиген может быть сброшен патогеном-мишенью и вследствие этого служить ловушкой для защитных антител. В-третьих, уреаза, по-видимому, токсична для эпителиальных клеток в культуре, и предполагают, что она принимает участие в разрушении слизистой оболочки и в образовании пептических язв in vivo. Следовательно, ее применение в качестве антигена может быть токсичным.
Несмотря на это мы решили, что этот антиген не мог бы быть потенциально эффективной вакциной, если:
- во-первых, мы вводили бы ее перорально в достаточно высокой дозе, чтобы вызвать более сильный иммунный ответ, чем природно наблюдаемый ответ,
- во-вторых, количество продуцируемых антител было бы достаточно высоким, чтобы связывать всю уреазу, сброшенную и несброшенную микроорганизмом,
- в-третьих, мы применяли бы субъединицы уреазы или вид молекул, которые не являются токсичными.
Короче говоря, остается потребность в эффективном лечении и профилактике вызываемой H.pylori желудочной инфекции у человека. Полученные недавно данные предполагают возможность получения вакцины против этого заболевания, но не дают ясной идентификации определенного антигена /антигенов/, общей для всех штаммов H. pylori, которые могли бы быть включены в надежную и эффективную вакцину.
В данном изобретении мы идентифицировали уреазный антиген H.pylori как кандидат для вакцины и показали его эффективность на модели животных. Эти результаты были неожиданными в свете естественной истории инфекций, называемых Helicobacter.
Было обнаружено, что можно индуцировать иммунитет у млекопитающих, восприимчивых к желудочно-кишечной инфекции, вызываемой Helicobacter, путем использования эпитопов уреазы, проявляющихся на или около поверхностей организмов Helicobacter, и применения их в качестве вакцины-мишени /целевой вакцины/. Иммунитет можно было индуцировать при помощи природной уреазы, но также субъединицы рекомбинатной уреазы, продуцируемой в виде ферментативно неактивной, т. е. нетоксичной формы. Изобретение обеспечивает способ индуцирования иммунитета к инфекции Helicobacter путем введения на поверхность слизистой оболочки млекопитающего полиаминокислотного препарата, т.е. смеси пептидов и/или белков, вместе с подходящим адьювантом. Этот полиаминокислотный препарат представляет множество эпитопов, характерных для уреазного фермента, эндогенного для инфицированного организма Helicobacter. Введение полиаминокислотного препарата можно проводить пероральным путем.
Активный ингредиент препарата может содержать природные или биосинтетические эпитопы и быть в различных формах. Неисчерпывающий перечень возможных препаратов включает в себя очищенные, встречающиеся в природе или рекомбинантные препараты бактериального или иного происхождения, переваренные ферментами препараты уреазы, слитые белки, содержащие эпитопы уреазы, усеченные формы уреазы или пептиды, гомологичные аминокислотной последовательности уреазы. Поскольку развитие иммунитета зависит от индукции гуморальных или/и клеточных иммунных ответных реакций, которые связывают инфицирующий организм Helicobacter, предпочтительными препаратами являются те, которые наиболее точно воспроизводят эпитопы уреазы, эндогенной для инфицирующего организма. Например, препараты, обнаруживающие эпитопы уреазы H.pylori, предпочтительны для введения больным, восприимчивым к H.pylori. Однако согласно важному аспекту данного изобретения, было обнаружено, что можно применять также и уреазу из других видов. Например, мы показали, что инфекцию H. felis у мышей можно предупредить введением уреазы из H.pylori.
Согласно одному аспекту изобретения обеспечен способ индуцирования в млекопитающем-хозяине защитного иммунного ответа на инфекцию Helicobacter, предусматривающий введение к поверхности слизистой оболочки хозяина иммунологически эффективного количества антигена уреазы, способного индуцировать защитный иммунный ответ, предпочтительно уреазы H.pylori или B субъединицы уреазы H.pylori.
Согласно другому аспекту данного изобретения обеспечена композиция вакцины, пригодная для предупреждения инфекции Helicobacter, содержащая эффективное количество антигена, предпочтительно уреазы H.pylori или субъединицы B уреазы H.pylori, способного вызывать в хозяине защитный иммунный ответ на инфекцию Helicobacter, в соединении с фармацевтически приемлемыми носителем или разбавителем.
Согласно дальнейшему аспекту данного изобретения, обеспечен способ наделения млекопитающего-хозяина пассивной защитой против инфекции Helicobacter, предусматривающий введение к поверхности слизистой оболочки хозяина иммунологически эффективного количества антител, специфических для уреазы, продуцируемых в хозяине, иммунизированном уреазой, предпочтительно уреазой H. pylori или субъединицей B H.pylori, способными вызывать защитный иммунный ответ на инфекцию Helicobacter.
Изобретение будет описано далее со ссылкой на сопутствующий рисунки, в которых фиг. 1 - фиг. 6 являются графическими представлениями результатов, даваемых в табл. 1 - 6.
Авторы данного изобретения обнаружили, что пероральное введение полиаминокислотных препаратов, имеющих эпитопы уреазы H.pylori, дает повышение защитного иммунологического ответа против H.felis у мышей, модели животных общепринятой значимости для исследования иммунного ответа на инфекцию Helicobacter /9/. Эффект защитного иммунного ответа заключается в том, что иммунизированные животные при введении патогена обнаруживают значительно сниженную частоту инфицирования по сравнению с неиммунизированными животными. Кроме того, авторы изобретения обнаружили, что пероральная иммунизация мышей с применением субъединицы B H.pylori, полученной в виде ферментативно неактивного рекомбинантного белка, дает повышение защитного иммунологического ответа у мышей против H. felis. Эффект этого защитного иммунного ответа заключается в том, что иммунизированные животные при введении патогена обнаруживают также значительно сниженную частоту инфицирования по сравнению с неиммунизированными животными, которые действительно заражаются.
Таким образом, в первом аспекте данное изобретение обеспечивает способ индуцирования в млекопитающем-хозяине защитного иммунного ответа против инфекции Helicobacter. Способ предусматривает стадию введения к поверхности слизистой оболочки млекопитающего, в том числе человека, иммунологически эффективного количества антигена уреазы, предпочтительно уреазы H.pylori, способного индуцировать такой защитный иммунный ответ.
Во втором аспекте данное изобретение обеспечивает способ индуцирования в млекопитающем-хозяине защитного иммунного ответа против инфекции Helicobacter. Способ предусматривает стадию введения к поверхности слизистой оболочки животного, в том числе человека, иммунологически эффективного количества антигена субъединицы B рекомбинантной ферментативно неактивной уреазы, предпочтительно субъединицы B рекомбинантной уреазы H.pylori, способного индуцировать такой защитный иммунный ответ.
Изобретение предусматривает также внутри сферы его действия лечение или профилактику млекопитающих, в том числе человека, от вызываемой Helicobacter инфекции, иммунологически эффективное количество уреазы или ее единицы, способных индуцировать защитный иммунный ответ на инфекцию Helicobacter, вводят к поверхности слизистой оболочки больного. Предпочтительной уреазой является уреаза H.pylori или субъединица B H.pylori. Уреаза может вводиться отдельно или в соединении с гидроксилированным фосфатом кальция, например гидроксиапатитом, в качестве частицы-носителя. Кроме того, предпочтительно введение уреазы H.pylori в соединении с адьювантом для слизистой оболочки, субъединицей B холерного токсина, мурамилдипептидом или другими подобными адьювантами.
Не будучи связанными какой-либо теорией, авторы данного изобретения убеждены, что введение антигена уреазы или ее субъединицы B к поверхности слизистой оболочки стимулирует общую иммунную систему слизистой оболочки и, возможно, локальные сайты в слизистой оболочке желудка, в том числе появление IgA антител, специфических для H.pylori, в желудочных секрециях, которые предупреждают инфекцию Helicobacter. Поскольку проведение предклинических испытаний кандидатов-вакцин для применения в лечении человека на моделях животных является рутинным, авторы считают, что методология данного изобретения эффективна для человека, в частности, в профилактике и лечении пептических язв, гастрита, злокачественных заболеваний желудка и других состояний, возникающих как результат присутствия H.pylori и/или H.heilmanii. A-Бактериальные культуры и очистка уреазы.
Штамм H.pylori, применяемый в этом исследовании, происходил из больного с язвой двеннадцатиперстной кишки и был субкультивирован на чашках с BHI агарозой до гомогенности. H.pylori культивировали в подходящей среде, обычно в BHI /Brain-Heart Ifusion/ среде, содержащей 0,25% дрожжевого экстракта и 10% эмбриональной сыворотки теленка с добавлением 0,4% селективного комплемента Campylobacter/ конглютинин Skirrow; Oxoid 69/. Бактерии инкубировали в течение ночи при микроаэрофильных условиях при 37oC в колбах, которые закрывали герметично и встряхивали при 37oC в течение 2 - 3 дней для получения жидкой культуры. Культуру можно также приготовить на чашках с агарозой, содержащей BHI с 0,25% дрожжевого экстракта и 5% овечьей крови при микроаэрофильных условиях при 37oC в течение 3 дней. Количество бактерий определяли по оптической плотности раствора BHI при 660 нм, причем одна единица оптической плотности соответствовала 108 бактерий. Культуры на чашках с агарозой перед этим ресуспендировали в 154 мл NaCl.
Одним из предпочтительных в настоящее время источников полиаминокислоты, несущей эпитопы уреазы, является очищенная уреаза, например уреаза H.pylori, полученная по особому способу Duann et al., J.Biol. Chem. 265, 9464 - 9469, модифицированному как описано ниже. После культивирования H.pylori собирали в воду, перемешивали вращением и откручивали еще раз для получения супернатанта. Затем раствор, содержащий уреазную активность H.pylori /оцениваемую по быстрому тесту на уреазу, см. ниже/, хроматографировали на колонке CL-6B и фракции, обладающие сильной уреазной активностью, собирали и диализовали в течение ночи и вновь хроматографировали на анионо-обменном геле. Фракции элюировали в буфере с NaCl с увеличивающейся концентрацией и собранные фракции с сильной уреазной активностью подвергали раздельно гель-электрофорезу с DC-Na с последующим окрашиванием Кумасси. Две отдельные полосы с мол. массами приблизительно 63 и 29 кДа идентифицировали как уреазу. Фракции, содержащие уреазу, соединяли вместе, получая очищенную уреазу H.pylori с чистотой 95 - 99%.
B-Пероральная иммунизация уреазой, очищенной из H.pylori.
Хотя предпочтительно применять очищенную уреазу H.pylori, полученную, как описано, в качестве антигенного материала, следует понимать, что также можно использовать в качестве антигенного материала любую уреазу или ее субъединицу, либо встречающиеся в природе либо полученные техникой рекомбинантной ДНК, а также переваренный фрагмент уреазы, слитые белки, содержащие фрагменты целой уреазы, усеченные конструкции уреазы и другие пептидные или белковые препараты, проявляющие эпитопы уреазы, которые способны индуцировать защитный иммунный ответ на инфекцию Helicobacter /см. ниже/. Таким образом, можно применять уреазу, имеющую существенную гомологию относительно уреазы H.pylori и эффективную в индуцировании защитной перекрестной иммунной ответной реакции на Helicobacter. Примером такой уреазы является уреаза канавалии мечевидной, которая имеет 70% гомологии с уреазой H.pylori. Изобретение, следовательно, не лимитировано применением интактной уреазы и охватывает применение любого полиаминокислотного препарата, который имеет эпитопы уреазы и эффективен в генерировании защитного иммунологического ответа у хозяина на инфекцию Helicobacter. Обычно уреаза, имеющая гомологию 70 - 95%, например гомологию 80 - 90%, относительно уреазы H.pylori, может применяться в качестве уреазы в данном изобретении.
Неограничивающий перечень источников потенциально применимых препаратов уреазы включает в себя эндогенные уреазы разных видов Helicobacter, уреазу из других бактерий, таких, как Klebsiella Pneumoniae или Proteus mirabilis, и по аналогии любую другую уреазу при условии, что эти уреазы имеют общие перекрестно реактивные эпитопы с уреазой H.pylori. Гены уреазы всех упомянутых выше организмов являются потенциальным инструментом для экспрессии рекомбинантных уреазных продуктов в виде целого белка или в виде его части.
Неограничивающий перечень потенциально применимых препаратов уреазы включает в себя пептиды, полученные из очищенной уреазы /источники упомянутые выше) при помощи физических и/или химических способов расщепления/ с применением протеаз, например V8-протеазы, трипсина или др./, или пептиды, синтезированные химически и содержащие общие последовательные эпитопы с уреазой.
Другими потенциально применимыми эпитопами являются эпитопы, идентифицированные по их перекрестной реактивности с уреазой в результате скрининга при помощи антител против уреазы. Такими пептидами могут быть природные пептиды или полученные химическим синтезом. Кроме того, такие пептиды могут быть результатом экспрессии рекомбинантного произвольного олигонуклеотида.
Другим источником потенциально применимых эпитопов являются эпитопы, подобные уреазе и полученные как результат генерирования антиидиотипических антител в уреазе. Такие антиидиотипические антитела, генерируемые в любом иммуннокомпетентном хозяине, получают иммунизацией этого хозяина антителами противоуреазы для генерирования антител против антител против уреазы, которые сохраняют структурные гомологии с уреазой.
Обсуждение здесь фокусируется на применении уреазы, природно продуцируемой H. pylori /раздел B/. Однако должно быть понятно, что уреаза или ее субъединицы или конструкции, упомянутые выше, способные индуцировать желаемый защитный иммунный ответ, могут быть получены техникой рекомбинантной ДНК, хорошо известной исследователям. Эффективность отдельных препаратов можно определить рутинным введением с применением моделей животных, пероральными введением кандидата-вакцины и введением патогена с применением протокола, подобного описанной ниже процедуре или идентичного этой процедуре.
Табл. 1 и 2 ниже и фиг. 1 - 5 описывают результаты, полученные при пероральной иммунизации мышей очищенной уреазой H.pylori. В этом первом эксперименте введение антигена H. pylori проводили путем перорального введения мышам уреазы H. pylori, очищенной как описано в разделе A, и соединенной с кристаллами гидроксиапатита, применяемыми в качестве носителя для усиления связывания M-клеток и поглощения. Холерный токсин /Sigma/ давали в качестве адьюванта для слизистой оболочки. В этом эксперименте группы самок SPF BALB/c шестинедельных мышей перорально иммунизировали 30 мкг очищенной уреазы H. pylori, соединенной с 1 мг гидроксиапатита и 10 мкг холерного токсина в качестве адьюванта, во временных точках 0, 7, 14 и 21 (дни). Затем мышам вводили дважды 108 H.felis на 28-ой и на 30-ый день. Для сравнительныхцелей подобные самки SPF BALB/c шестинедельных мышей перорально иммунизировали полным лизатом /гомогенатом, полученным при разрушении клеток ультразвуком/ H.pylori и 10 мкг холерного токсина в те же дни 0, 7-ой, 14-ый и 21-ый. Мышам вводили на 28-ой и 30-ый день H.felis. Лизат H.pylori получали сбором клеток из клеточных культур, осаждением их центрифугированием, ресуспендированием осадка в 0,9% NaCl с последующей обработкой ультразвуком.
В качестве контроля самки SPF BALB/c шестинедельных мышей были перорально ложно иммунизированы 10 мкг холерного токсина и 1 мг гидроксиапатита в те же дни 0, 7-ой, 14-ый и 21-ый. Всех мышей содержали, иммунизировали и заражали параллельно. Всех мышей убивали на 35-ый день.
C - Пероральная иммунизация субъединицами рекомбинантной уреазы H.pylori.
Гены, колирующие структурные единицы A и B уреазы H.pylori, были получены при помощи клонирования с применением полимеразной цепной реакции /ПЦР/ в соответствии со стандартными процедурами, основывающимися на ранее опубликованных последовательностях /29/. Эти гены встраивали в вектор /названный pEV 40/, предназначенный для высокоэффективной экспрессии и легкой очистки чужеродных генов в E.coli. Вкратце чужеродный ген встраивали справа от терморепрессированного промотора и в рамке последовательности, кодирующей повтор из шести гистидинов. Для отбора трансформаторов на векторе присутствовал ampR ген. При подходящих температурных условиях полученный рекомбинантный белок имел шесть гистидинов на N-конце, что делало возможной ее одноступенчатую очистку на содержащей никель колонке. Как субъединицу A, так и субъединицу B рекомбинантной уреазы H.pylori экспрессировали раздельно в E.coli и очищали на содержащей никель колонке до чистоты 95%.
Хотя предпочтительно применение рекомбинантной уреазы H.pylori, полученной как описано выше, следует понимать, что можно также применять в качестве антигенного материала любую уреазу или субъединицу уреазы, полученные рекомбинантной техникой /например, слитый белок/, имеющие антигенные сайты уреазы, которые способны вызвать защитный иммунный ответ на инфекцию Helicobacter. Так, можно применять конструкцию гена уреазы, имеющей существенную гомологию с уреазой H.pylori и эффективный в индуцировании перекрестного защитного иммунного ответа на Helicobacter. Примером такой уреазы является уреаза канавалии мечевидной, которая имеет 70% гомологии с уреазой H.pylori, или уреазой H. felis, которая имеет 88% гомологии с уреазой H.pylori. Следовательно, изобретение не лимитировано применением генов H.pylori и их генных продуктов и включает в себя применение любой рекомбинантной уреазы или ее субъединицы, которые эффективны в генерации защитного иммунологического ответа в хозяине на инфекцию Helicobacter. Как правило, рекомбинантная уреаза, имеющая 70 - 95% гомологии, например 80 - 90% гомологии, с уреазой H. pylori может применяться в данном изобретении в качестве антигена рекомбинантной уреазы.
Обсуждение здесь фокусируется на применении субъединиц A и B рекомбинантной уреазы H.pylori, продуцируемой E.coli /раздел C/. Однако должно быть понятно, что рекомбинантная уреаза или ее субъединицы или конструкции, упомянутые выше, способные вызывать желаемый защитный иммунный ответ, могут быть получены с применением других способов рекомбинантной ДНК и других эукариотических или прокариотических экспрессирующих векторов, хорошо известных в данной области исследований.
Табл. 3, 4 и 5 ниже на фиг. 6 описывают результаты, полученные при пероральной иммунизации мышей субъединицами уреазы H.pylori, полученными E.coli. В этом эксперименте введение антигена H.pylori проводили пероральным введением мышам субъединиц A или B рекомбинантной уреазы H.pylori, полученных в E.coli и очищенных, как описано выше, и соединенных с кристаллами гидроксиапатита, применяемых в качестве носителя для усиления связывания M-клеток и поглощения. Холерный токсин /Sigma/ давали в качестве адьюванта слизистой оболочки. В этом эксперименте группы самок SPF/BALB/c шестинедельных мышей перорально иммунизировали 30 мкг субъединиц A и B рекомбинантной уреазы H.pylori, соединенных с 1 мг гидроксиапатита и 10 мкг холерного токсина в качестве адьюванта, в дни 0, 8-ой, 14-ый и 21-ый. Затем мышам трижды вводили 108 H.felis на 32-ой, 34-ый и 36-ой день. Для сравнения подобных самок SPF BALB/c шестинедельных мышей перорально иммунизировали 30 мкг субъединицы B рекомбинантной уреазы H.pylori, соединенной с гидроксиапатитом и 10 мкг холерного токсина, на 0, 8-ой, 14-ый и 21-ый день. Мышей заражали трижды на 32-ой, 34-ый и 36-ой день H.felis. В качестве контроля самок SPF BALB/c шестинедельных мышей перорально ложно иммунизировали 10 мкг холерного токсина и 1 мг гидроксиапатита на 0, 8-ой, 14-ый и 21-ый день. Затем мышам вводили H.felis на 32-ой, 34-ый и 36-ой день. Всех мышей иммунизировали и заражали в параллелях. Животных убивали на 48-ой день /через 12 дней после заражения/ или через 10 недель после заражения.
D-Анализ желудочных биопсий, крови и кишечных секреций
Биопсии брали из желудка, кровь получали из сердца. Кишечник удаляли и промывали 1 мМ ФМСФ /Boeringer/ в PBS буфере для получения кишечных секреций для твердофазного иммунноферментного анализа /ELISA/. Для оценки защиты против колонизации H.felis желудочные биопсии из каждого животного подвергали скринингу на присутствие H.felis путем быстрого определения уреазной активности по тесту Tatrox HP/Rohm Pharma/. Вкратце желудочные биопсии погружали в 0,5 мл смеси поставщика из мочевины и фенолового красного, индикатора pH. Уреазная активность продуцирует аммиак и бикарбонат из мочевины и сопровождается калориметрическим изменением раствора в сторону более высокого поглощения при 550 нм. Уреазную активность измеряют количественно спектрофотометрически.
Желудочные биопсии каждого экспериментального животного раздела B также культивировали в чашках с BHI агарозой с описанными выше добавками для обнаружения H. felis. После 3 - 10 дней инкубации в микроаэрофильных условиях присутствие H. felis подтверждали окрашиванием на грамотрицательный или грамположительные бактерии и определением уреазной активности. Поскольку наблюдали значительную корреляцию при детектировании культур H.felis в первой серии экспериментов /см. табл. 3/, то для обнаружения H.felis в эксперименте, описанном в разделе C, проводили только уреазные тесты желудочных биопсий. Обнаружение H.felis подтверждалось микроскопией двумя независимыми исследователями с применением двух разных красителей /акридинового оранжевого и крезилового фиолетового/.
Пробы крови коагулировали в течение 3 ч при RT (комнатной температуре), собирали сыворотки и замораживали их при -20oC до проведения анализа. Кишечные инфекции откручивали в течение 5 мин при 4oC для удаления остатков клеток и хранили замороженными при -20oC. Затем пробы сыворотки и кишечные пробы каждого животного анализировали при помощи ELISA для оценки активности против Helicobacter согласно стандартным процедурам. Вкратце пластинки с 96 ячейками покрывали гомогенатом, полученным после обработки клеток H.pylori ультразвуком с последующим насыщением 5%-ным обезжиренным молоком. Пробы сериально разбавляли от 1:1 до 1:1000 и инкубировали в течение ночи при 4oC на пластинках ELISA. Биотинилированные IgA против мышей /сыворотка/ и IgA, обработанные затем стрептавидином - пероксидазой хрена, были использованы для определения уровней антител.
Результаты введений H.felis после иммунизаций очищенной уреазы H.pylori представлены в табл. 1 - 3 и фиг. 1 - 4, а результаты введения H.felis после иммунизации субъединицами A и B рекомбинантной уреазы H.pylori представлены в табл. 4 - 6 и фиг. 5 и 6.
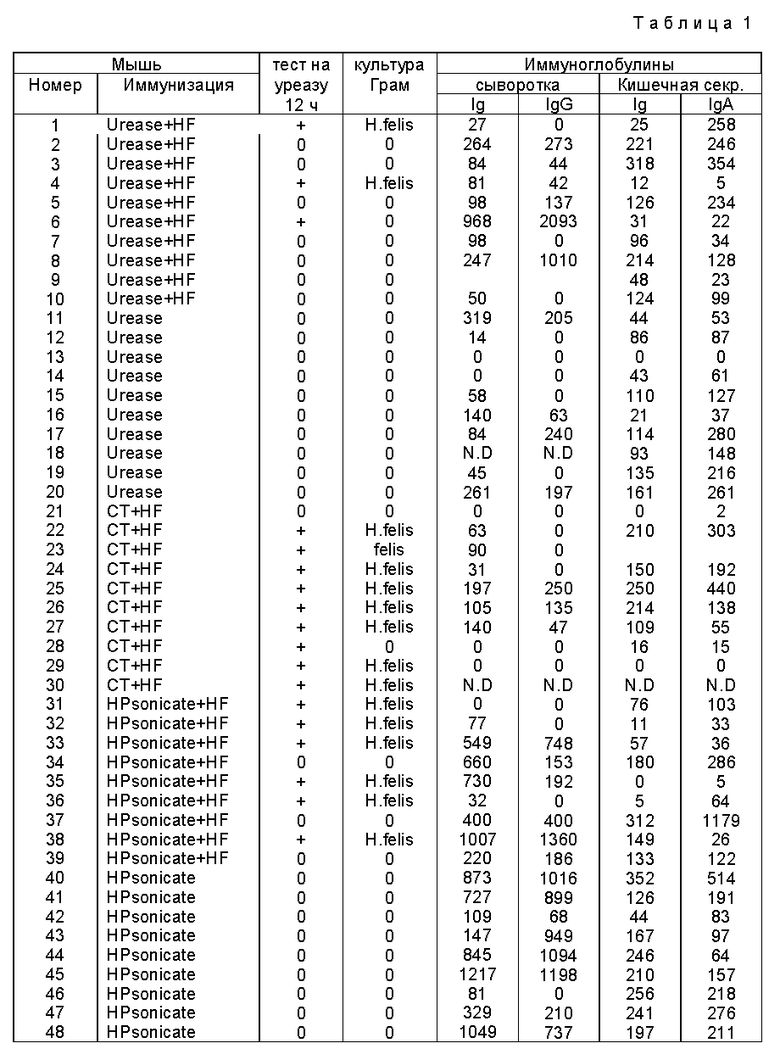
В табл. 1, относящейся к эксперименту, описанному в разделе B, "ч" обозначает часы, "Ig" обозначает иммуноглобулин, "ND" обозначает "не определяли", "уреаза + HF" обозначает, что мышей иммунизировали уреазой /соединенной с гидроксиапатитом и холерным токсином/ и затем заражали H.felis, "уреаза" обозначает, что мышей иммунизировали уреазой (соединенной с гидроксиапатитом и холерным токсином) и не заражали, "CT + HF"обозначает, что мышей ложно иммунизировали холерным токсином и заражали H.felis, "HP Sonicate+ HF" обозначает, что мышей иммунизировали гомогенатом, полученным при обработке клеток H.pylori ультразвуком, с холерным токсином и заражали H.felis, а "HF Sonicate" обозначает, что мышей иммунизировали гомогенатом, полученным обработкой клеток H. pylori ультразвуком, с холерным токсином и не заражали. В табл. 1 числа для результатов с антителами даются как измерение поглощения при 595 нм, умноженное на 1000. Измеренный фон в отсутствие антител вычитали.
Результаты эксперимента, описанного в разделе B, полученные на основе уреазы тестов желудочных биопсий и Грам-окрашивания культур H.felis, представлены в табл. 2. Инфекцию определяли с применением одного или нескольких маркеров колонизации H. felis, в том числе при помощи уреазного теста или Грам-окрашивания культур.
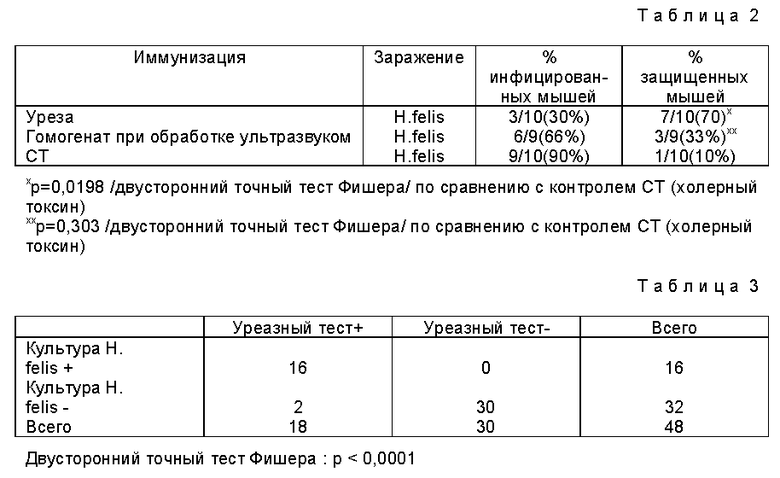
Из результатов, представленных в табл. 1 и 2, видно, что при пероральной иммунизации уреазой H. pylori по сравнению с вариантами, в которых вводили либо гомогенат, полученный обработкой клеток H.pylori ультразвуком, либо холерный токсин, получали статистически значимую защиту против заражения H.felis. Как видно из табл. 2, из 10 иммунизированных животных только 3 инфицировались, по сравнению с 6 животными, иммунизированными гомогенатом клеток H. pylori, полученными при обработки ультразвуком, и 9 животными, иммунизированными холерным токсином. Табл. 2 показывает, что 70% животных были защищены от заражения H.felis по сравнению с 33% животных, иммунизированных гомогенатом клеток при обработке ультразвуком, и 10% животных, иммунизированных холерным токсином и подвергнутых затем введению H.felis. Другими словами, 90% контрольных мышей, экспонированных с H.felis, были инфицированы патогеном, тогда как у мышей, иммунизированных уреазой H.pylori за 28 дней до экспозиции с H.felis, скорость инфицирования была только 30%. Это означает значительное снижение заражения /p = 0,0198 в точном тесте Фишера по сравнению с контрольными мышами/. При пероральной иммунизации мышей гомогенатом клеток H.pylori, полученным при обработке клеток ультразвуком, скорость инфицирования была 67% /незначительно отличалась от контроля/. Защита, полученная с применением уреазы H.pylori, была неожиданной и не могла быть предсказана на основании результатов, наблюдаемых при использовании гомогената клеток H.pylori, полученного при обработке клеток ультразвуком.
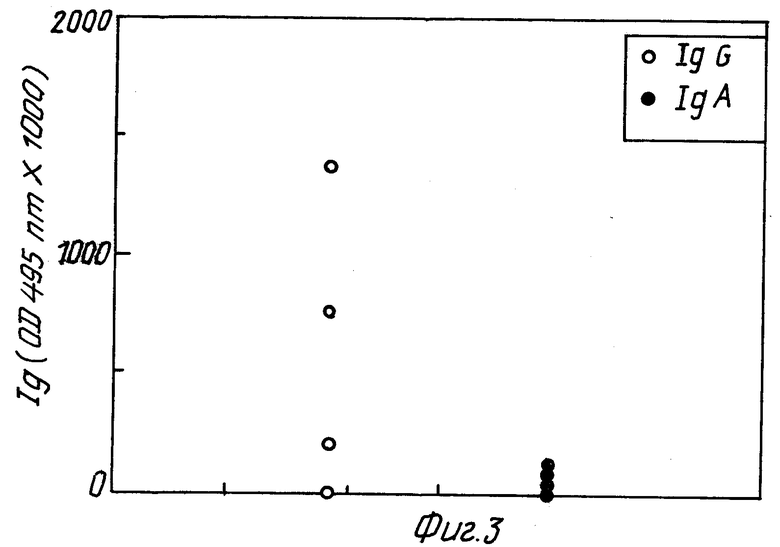
Фиг. 1 представляет графически результаты тестов на антитела в сыворотке /IgG/ и кишечной секреции /IgA/ у мышей, не защищенных после иммунизации уреазой. Эти мыши соответствуют нормам 1, 4 и 6 в табл. 1 и составляют группу A.
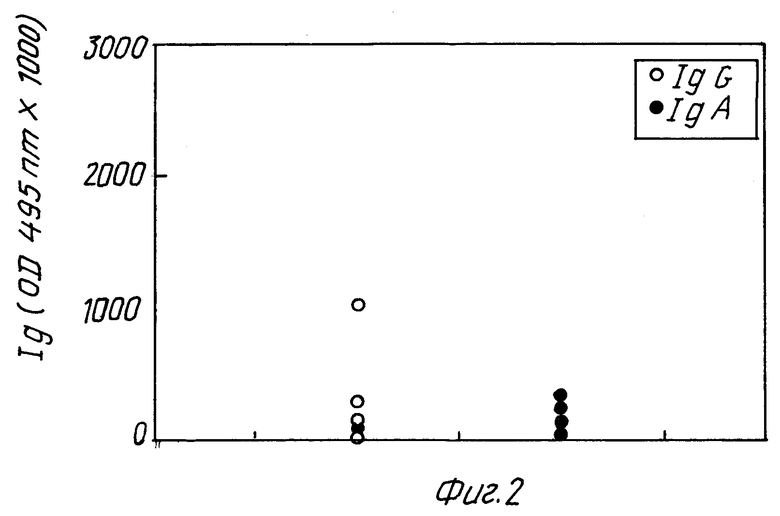
Фиг. 2 показывает ответ в виде образования антител у мышей, которые были защищены после иммунизации уреазой /группа B/, т.е. мышей 2, 3, 5 и 7 - 10.
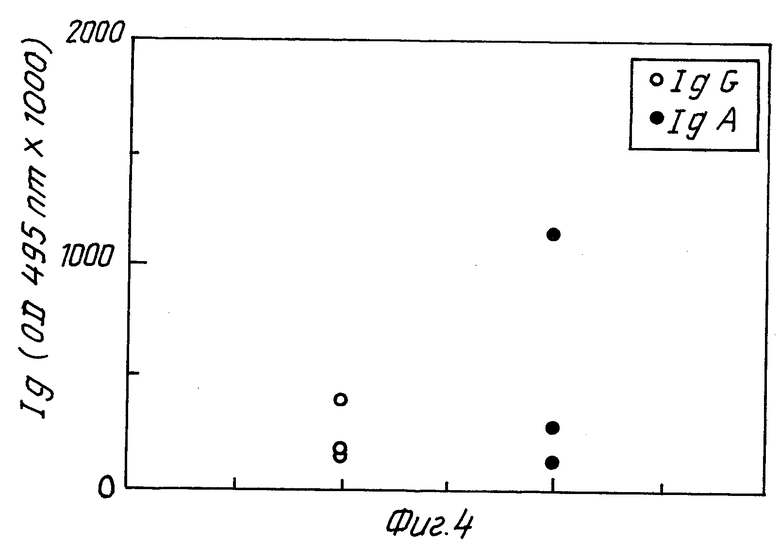
Фиг. 3 и 4 относятся к результатам, полученным с мышами 31 - 39. Фиг. 3 /группа C/ изображает образование антител у мышей, не защищенных после иммунизации гомогенатом H.pylori /номера мышей 31, 32, 33, 35, 36 и 38/. Фиг. 4 /группа D/ изображает образование антител у мышей, защищенных после иммунизации гомогенатом H.pylori /номера мышей 34, 37 и 39/. Интересно отметить, в отношении фиг. 3 и 4, что обозначение IgA антител /но не IgG/ выше у мышей защищенных, чем у незащищенных мышей, что предполагает корреляцию между защитой и образованием IgA антител. Образование IgG не обнаруживает подобной корреляции. Как известно, IgA слизистой оболочки, а не IgG сыворотки играют роль в защите против бактериальных инфекций кишечника.
Результаты корреляции между детектированием H.felis в желудочных биопсиях при помощи уреазных тестов и при помощи культур представлены в табл. 3.
Табл. 3 показывает, что существует очень значительная корреляция между результатами уреазных тестов, проведенных на желудочных биопсиях, и идентификацией H.felis в культурах. В дальнейших экспериментах предпочтение отдавали уреазным тестам для диагностики инфекции H.felis в мышах вследствие большей чувствительности этих тестов. Этот подход позволил проводить уреазные тесты с большими фрагментами желудка каждой мыши и увеличить далее чувствительность уреазного теста. Кроме того, применение способа с наивысшей чувствительностью предохраняет от завышенной оценки защиты, полученной с испытуемыми препаратами. При использовании положительной культуры в качестве стандарта для инфицированная защита, индуцированная после иммунизации уреазой во время эксперимента, представленного в разделе B, так же значительна, как при применении сочетания уреазного теста и культуры /p = 0,021 против p = 0,019/.
Результаты экспериментов, описанных в разделе C /субъединицы рекомбинантной уреазы/, полученные на основании уреазных тестов желудочных биопсий, представлены в табл. 1, 5 и 6 и изображены на фиг. 6.
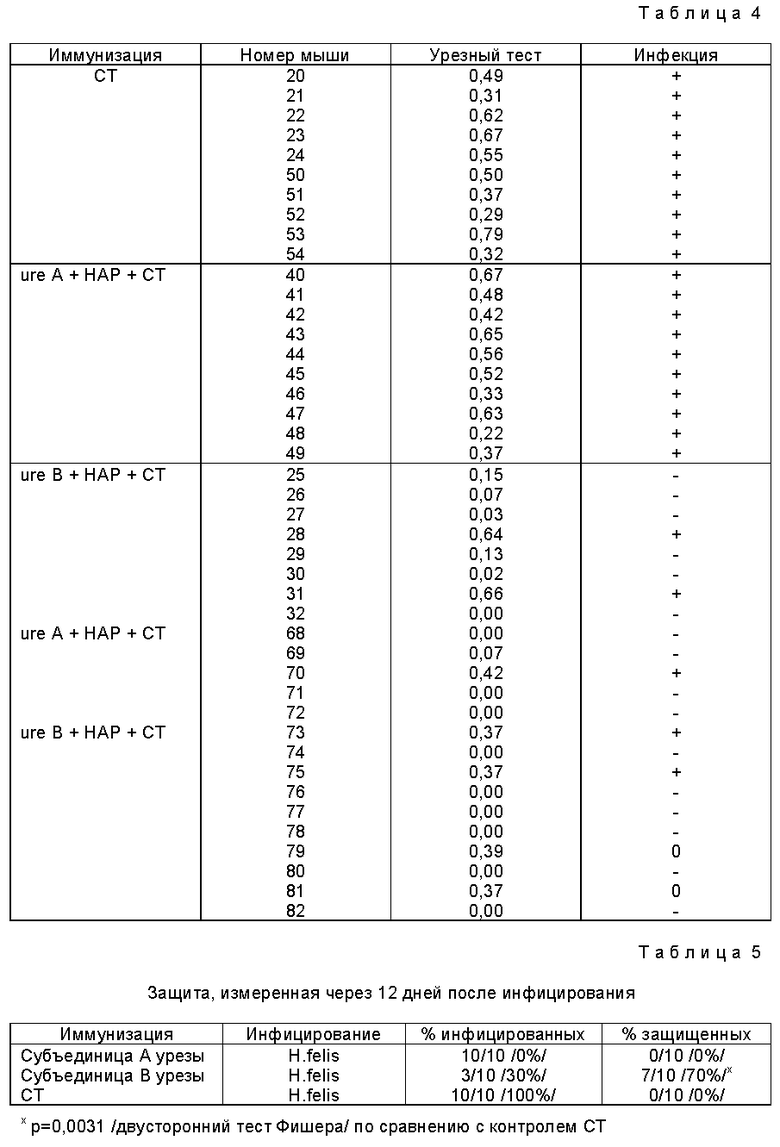
В табл. 4 "CT" обозначает холерный токсин, "ureA" обозначает субъединицу A уреазы H.pulori, "ureB" обозначает субъединицу B уреазы H.pylori и "HAP" обозначает кристаллы гидроксиапатита. Мышей 20 - 54 убивали через 12 дней после инфицирования, а мышей 68 - 82 - через 10 недель /106 дней/ после инфицирования. Результаты уреазного теста, проведенного на биопсиях желудка каждого животного, представлены в виде величин OD при 550 нм. Знаки + и - обозначают конечное состояние инфекции каждого животного в соответствии с положительным или отрицательным уреазным тестом для обнаружения H.felis. Положительная реакция соответствует OD 550 > 0,2.
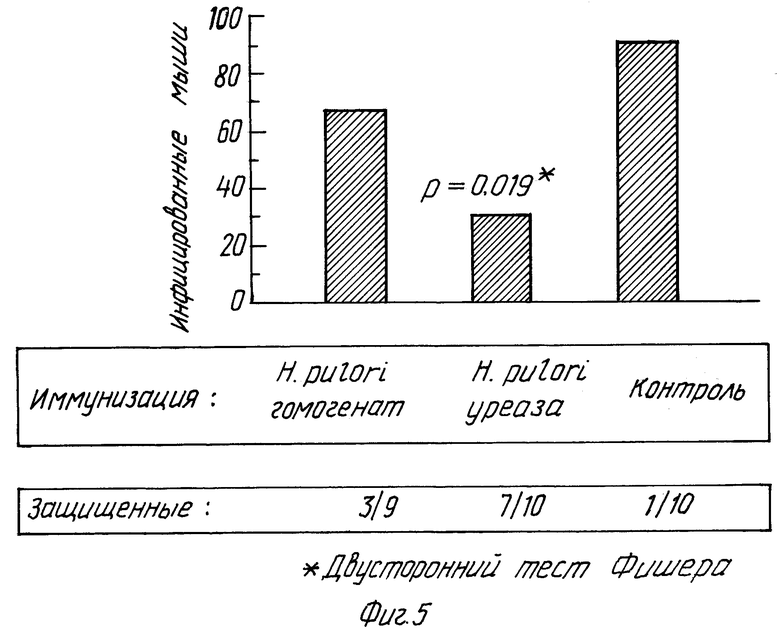
Из результатов, представленных в табл. 4, 5 и 6, видно, что статистически значимую защиту против инфицирования H.felis получали при пероральной иммунизации B субъединицей рекомбинантной уреазы H.pylori по сравнению с защитой, полученной с применением либо субъединицы A H.pylori, либо холерного токсина. Из табл. 4 видно, что через 12 дней после инфицирования из 10 иммунизированных животных только 3 были инфицированы в группе с субъединицей B уреазы по сравнению с 10 животными, иммунизированными субъединицей A уреазы H.pylori, и 10 из 10 животных, иммунизированных холерным токсином. Табл. 4 показывает, что 70% животных были защищены от инфицирования H.felis по сравнению с 0% животных, иммунизированных субъединицей A H.pylori и 0% животных, иммунизированных холерным токсином и затем подвергнутых инфицированию H.felis. Другими словами, 100% контрольных мышей, инфицированных H.felis, заражались, тогда как у мышей, иммунизированных субъединицей B рекомбинантной уреазы H.pylori, скорость (коэффициент) инфицирования была только 30%. Это свидетельствует о значительном снижении инфицирования /p = 0,0031, точный тест Фишера/, по сравнению с контрольными мышами.
Тот факт, что защита, наблюдаемая при применении уреазы H.pylori, полностью зависела от иммунизации субъединицей B уреазы и субъединица A не давала такого эффекта, не был предсказуем на основании этого эксперимента с очищенной уреазой. Следовательно, это определение роли 2 структурных субъединиц уреазы в развитии защитного иммунного ответа является новым. Защита, полученная с применением субъединицы B рекомбинантной уреазы, которая ферментативно неактивна, свидетельствует также о том, что нетоксичные формы уреазы могут быть использованы в качестве пероральной вакцины против инфекции Helicobacter. Кроме того, эти результаты являются сильным свидетельством в пользу того, что для защиты не требуется узнавание активного сайта, поскольку едва ли можно думать, что неактивная субъединица B уреазы способна индуцировать антитела,которые будут узнавать и ингибировать каталитический сайт нативной уреазы.
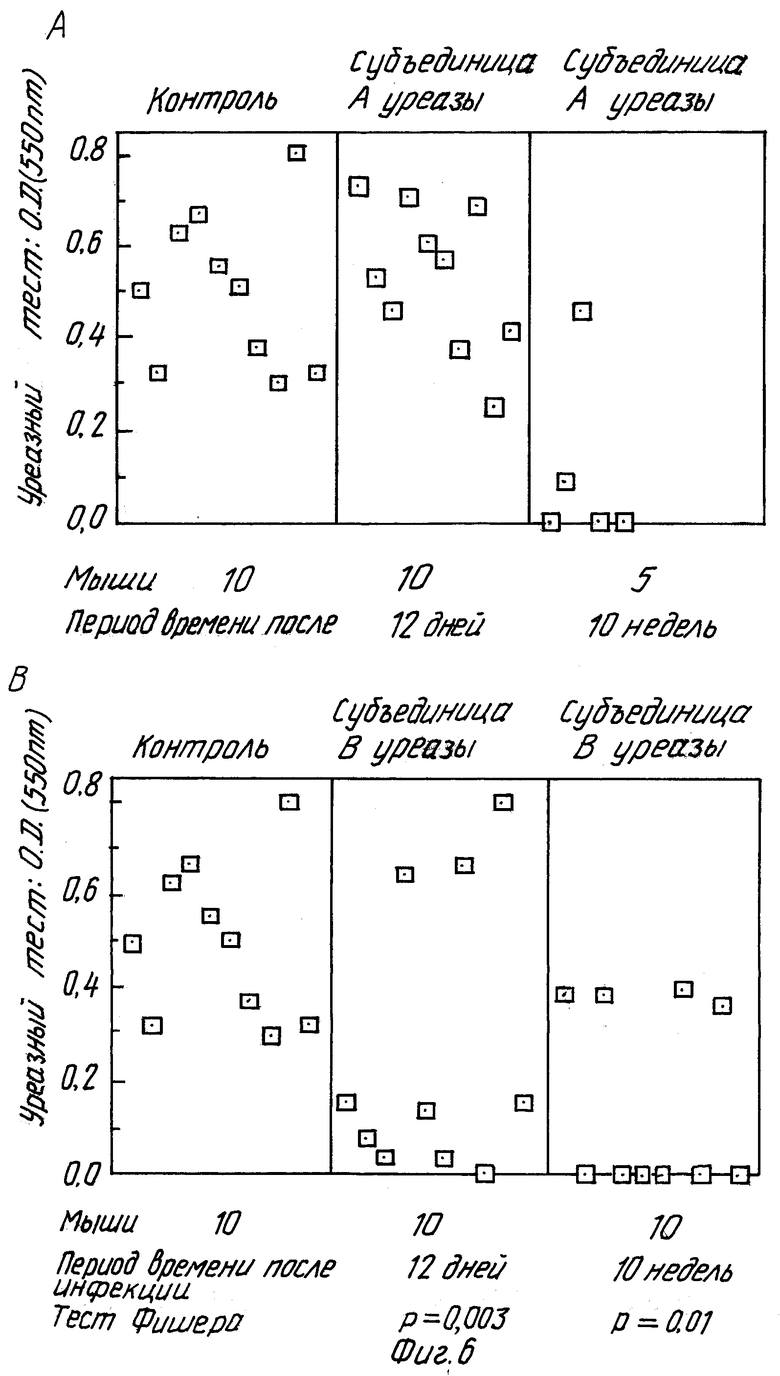
Из табл. 6 видно, что в случае, когда мышей убивали через 10 недель после инфицирования, 60% /6 мышей из 10/ животных, иммунизированных субъединицей B уреазы, и 80% /4 мыши из 5/ животных, иммунизированных субъединицей B H. pylori, были защищены против инфекции H.felis. Тот факт, что защита, полученная иммунизацией субъединицей B уреазы, сохраняется с течением времени и что иммунизация субъединицей A уреазы вызывает защиту, которая исчезает со временем по сравнению с защитой, вызываемой субъединицей B уреазы, был неожиданным после нашего эксперимента с очищенной уреазой или других проведенных ранее экспериментов. Факт, что иммунизация субъединицей B уреазы вызывает защиту, определенно доказывает, что для защиты не требуется узнавания активного сайта. Фиг. 6 суммирует результаты, полученные после пероральной иммунизации субъединицами A и B рекомбинантной уреазы /описанные в табл. 5 и 6/.
Данное изобретение также обеспечивает композиции вакцин, пригодные для профилактики инфекции Helicobacter. Эти композиции содержат эффективное количество антигена уреазы, предпочтительно субъединиц уреазы H.pylori или рекомбинантной уреазы H.pylori, способного индуцировать в хозяине защитный иммунный ответ на инфекцию Helicobacter, в соединении с фармацевтически приемлемым носителем или разбавителем.
Вакцины вводят в количествах, легко определяемых лицами с обычной квалификацией в этой области. Так, для взрослых подходящая доза лежит в диапазоне 10 мкг - 100 мг, например 50 мкг - 50 мг. Близкие диапазоны доз применимы для детей. Системы носителя могут представлять собой капсулы с высвобождением активного компонента в желудке, защищающие антиген от кислой среды в желудке и содержащие антиген уреазы в нерастворимой форме в виде слитых белков. Вакцина может вводиться в качестве первичного профилактического средства для взрослых и детей, в качестве вторичного профилактического средства, после успешной ликвидации H.pylori в инфицированном хозяине или в качестве терапевтического средства с целью индуцирования иммунного ответа в хозяине для уничтожения H.pylori.
Как отмечалось выше, подходящим адьювантом для слизистой оболочки является холерный токсин. Можно использовать также мурамилдипептид или его производные, нетоксичные производные холерного токсина, в том числе его B-субъединицу, и/или конъюгаты или генетически сконструированные слияния (слитые белки) антигена уреазы с холерным токсином или его B-субъединицей. Другими подходящими средствами доставки являются биодеградируемые микрокапсулы или иммуностимулирующие комплексы /IS COM'S/ или липосомы, генетически сконструированные ослабленные живые векторы, такие, как вирусы или бактерии, и рекомбинантные /химерные/ подобные вирусу частицы, например вирусу блутанга /"синего языка"/. Количество адьюванта слизистой оболочки зависит от типа этого адьюванта. Например, в случае холерного токсина его применяют в количестве 5 мкг - 50 мкг, в частности 10 мкг - 35 мкг. В случае микрокапсул применяемое количество будет зависеть от количества, применяемого в матриксе микрокапсул для достижения желаемой дозы. Определение этого количества вполне доступно лицам с обычной квалификацией в этой области. Пригодными носителями для вакцин данного изобретения являются желудочные капсулы с покрытием и поликислотные-гликолидные микросферы. В качестве разбавителей пригодны 0,2N NaHCO3 и/или солевой /физиологический/ раствор.
Гидроксилированный фосфат кальция /HCP/ в виде частиц особенно применим в качестве носителя для уреазы H. pylori для нанесения на поверхность слизистой оболочки. Считается, что конъюгат уреазы H. pylori с гидроксилированным фосфатом кальция транспортируется через эпителий, где он вызывает иммунный ответ в виде образования поли - Ig. Предпочтительно применение гидроксилированного фосфата кальция в форме микрочастиц, пригодных для транспорта через эпителий, в частности клетками, специализированными для этой цели /М - клетками/. Предпочтительной формой гидроксилированного фосфата кальция является гидроксиапатит, коммерчески доступный кристаллический фосфат кальция Ca10(PO4)6(OH)2.
Коммерческий гидроксиапатит обычно состоит из стержнеподобных кристаллов, которые химически и физически аналогичны неорганическому гидроксиапатиту в нормальной костной ткани. Поэтому прием внутрь гидроксиапатита является безопасным, что подтверждается существованием пищевых кальций-фосфорных добавок, получаемых из размолотых костей и предназначенных для глотания. Коммерческий очищенный гидроксиапатит /из Cal Biochem/ состоит из кристаллов, широко варьирующих по размеру. Кристаллы длиной более 1 мкм, вероятно, не поглощаются М - клетками. Поэтому для применения в данном изобретении кристаллы коммерческого гидроксиапатита разбивают на небольшие, относительно однородные кристаллические фрагменты, например, разрушением ультразвуком. Предпочтительно основная часть гидроксиапатита присутствует в виде фрагментов приблизительно 0,01 - 0,0 мкм. Фрагментация может быть изменена либо при помощи электронной микроскопии, либо по рассеиванию света обычными способами.
Предпочтительными способами введения антигена H.pylori является пероральный, назальный, ректальный и глазной способы. Пероральный способ введения может обеспечить доставку к слизистой оболочки не только желудка, но и кишечника.
Вакцины данного изобретения можно вводить к поверхности слизистой оболочки в форме аэрозоля, суспензии, капсулы и/или суппозитория. Способ введения легко определяют лица с обычной квалификацией в этой области.
Данное изобретение предусматривает также пассивную иммунизацию млекопитающих, в том числе человека, против инфекции Helicobacter. Это достигается введением к поверхности слизистой оболочки больного эффективного количества специфических для уреазы антител, предпочтительно IgA моноклональных антител, специфических для H.pylori.
Поскольку уреаза H. pylori представляет антиген, участвующий в индуцировании защитного иммунитета, дальнейшим аспектом данного изобретения является применение уреазы H.pylori в качестве диагностического реагента для измерения иммунного ответа лиц, получивших вакцину на основе уреазы, или для определения иммунности или восприимчивости /т.е. необходимости вакцинации/ индивидуума. Данное изобретение также предусматривает применение уреазы или специфических для уреазы антител для составления реакционных смесей и наборов /китов/ для диагностики иммунитета к инфекции Helicobacter, оценки восприимчивости к инфекции Helicobacter и определения иммунного ответа на вакцину.
Примеры. Далее изобретение будет описано со ссылкой на следующие неограничивающие изобретение примеры.
a) Бактериальные штаммы
H. felis был получен у J Fo /division of Cooperative Medicine, Mass. Institute of Technology, Boston. USA/. H.pylori был выделен из больных с язвенными заболеваниями /CHUV, Lausanne, Switzer Land/.
b) Бактериальные культуры
Жидкая культура - бактерии культивировали на BHI /Brian Heart Infusion, BioMerieux/ жидкой среде, содержащей 0,25% дрожжевого экстракта /Difco/ и 10% эмбриональной сыворотки теленка Inotech/ с добавлением 0,4% выбранного комплемента Campylobacter /Oxoid/. Бактерии инкубировали в течение ночи при микроаэрофильных условиях при 37oC и затем качали при 37oC в течение 2 - 3 дней.
Культура на агарозных чашках - бактерии культивировали на агарозных чашках, содержащих BHI, 0,25% дрожжевого экстракта и 50 овечьей крови, при микроаэрофильных условиях в течение 3 дней при 37oC.
Определение количества бактерий - количество бактерий определяли по оптической плотности раствора BHI при 660 нм /1 единица оптической плотности соответствует 108 бактерий/.
c) Приготовление гомогентов разрушенных ультразвуком клеток
H.pylori собирали из 31 чашки, содержащих агарозу с кровью в 0,15 М NaCl и откручивали 5 мин при 1400 g при 4oC. Остаток ресуспендировали в 3 мл NaCl и обрабатывали звуком /"озвучивали в течение 4 мин. Количество белка определяли по Бредфорду /BioRadKit в соответствии с поставщиком/.
d) Соединение иммуногена с гидроксиапатитом
Иммуноген /уреазу или ее субъединицу/ инкубировали в течение часа при 4oC с гидроксиапатитом. 1,0 г гидроксиапатита применяли для 30 мкг иммуногена на мышь. В конце инкубирования добавляли 10 мкг холерного токсина в конечном объеме 200 мкл PBS.
Пример 1.
a) Экстракция
H. pylori из 30 чашек с кровью и агаром собирали в 0,15 М NaCl на льду. Раствор откручивали 5 мин при 1400 g при 4oC. Осадок ресуспендировали в 20 мл H2O и откручивали 45 с при максимальной скорости. Затем экстракт откручивали 20 мин при 6700 g при 4oC. Супернатант извлекали и оценивали количества белка /см. выше/, после чего осаждали 70%-ным сульфатом аммония.
b) Очистка уреазы
Раствор хроматографировали на колонке Sepharose CL-6B /Pharmacia/ с PBS /физиологический раствор с фосфатным буфером/ в качестве подвижной фазы. 22 собранные фракции, которые обнаружили активную уреазную активность, соединяли вместе и диализировали в течение ночи при 4oC против 3 л PEB (20 нМ фосфатный буфер, pH 7) и затем хроматографировали на Q Sepharose fast flow /Pharmacia/ с PEB в качестве подвижной фазы. Фракции элюировали при помощи градиента 0 - 500 нМ NaCl. 10 собранных фракций с сильной уреазной активностью отдельно наносили на гель с DC-Na с последующим окрашиванием Кумасси. 6 фракций обнаружили 2 отдельные полосы, соответствующие мол.массам 63 и 28 КДа. Эти фракции соединяли и рассматривали как очищенную уреазу.
Пример 2 /см. также раздел B/.
Мышей, применяемых в исследовании иммунизации, подвергали легкой анестезии эфиром перед внутрижелудочной иммунизацией. Затем озвученный препарат или очищенную уреазу, гидроксиапатит и холерный токсин суспендировали в PBS и 200 мкл доставляли в желудок соответствующих мышей при помощи полиэтиленовой трубки, соединенной со шприцем для подкожных инъекций. Этот способ назван пероральной иммунизацией.
Оценивались протоколы трех пероральных иммунизаций. Они описаны ниже.
Протокол B1 - Вакцинация очищенной уреазой
Самок BALB/c шестинедельных мышей /20/ перорально иммунизировали 30 мкг очищенной уреазы H.pylori и 1 мг гидроксиапатита и 10 мкг холерного токсина на 0, 7-ой, 14-ый и 21-ый дни. 10 мышей инфицировали на 28-ой и 30-ый дни 5•107 и 108 H.felis из жидкой культуры.
Протокол B2 - Вакцинация озвученными гомогенатами He
Самок BALB/c шестинедельных мышей /20/ перорально иммунизировали 2 мг раствора после обработки ультразвуком клеток H.pylori на 0, 7-ой, 14-ый и 21-ый дни. 10 мышей инфицировали на 28-ой и 30-ый дни 5•107 и 108 H.felis.
Протокол B3 - Контроль
Самок BALB/c шестинедельных мышей /20/ перорально иммунизировали 1 мг гидроксиапатита и 10 мкг холерного токсина на 0, 7-ой, 14-ый и 21-ый дни. Мышей инфицировали на 28-ой и 30-ый дни 5•107 и 108 H.felis.
На 35-ый день мышей убивали и были взяты биопсии из желудка, а также кишечные секреции и кровь.
Защита и оценка
Для оценки защиты биопсии скринировали на уреазную активность при помощи теста Tetrox HP /Rohm Pharma/ в соответствии с инструкциями поставщика. Уреазу определяли количественно путем спектрофотометрического измерения при 550 нм. Затем биопсии культивировали в присутствии H.felis и оценивали по Грам-окрашиванию. Биопсии полости желудка гомогенизировали и разбавляли /1: 100 и 1:1000/ в 0,15 М NaCl и высевали в чашки с кровью и агаром и инкубировали при микроаэрофильных условиях при 37oC в течение 4 - 10 дней.
ELISA /твердофазный иммуноферментный анализ/
Кишечные секреции и кровь анализировали при помощи ELISA для оценки титра антител. Анализ проводили следующим образом. Планшеты из полистирола /96 ячеек/ покрывали 1 мкг/ячейку очищенной уреазой при 37oC в течение 2 ч. Сайты неспецифического связывания блокировали 5%-ным порошкообразным молоком в PBS - 0,1% Tween при 37oC в течение 30 мин. Планшеты промывали один раз PBS - 0,1% Tween. Пробы крови тестировали при разведении 1:100, а кишечные секреции при разведении 1:1. 100 мкл каждой пробы добавляли к покрытым антигеном планшетам. После 2 ч инкубации планшеты промывали 3 раза PBS - 0,1% Tween. Антимышиные биотинилированные целые антитела из козы и антимышиные биотинилированные IgA, IgG и IgM /Amersham/ добавляли /100 мкл/ при разбавлении 1: 500 за исключением IgA /1:250/ и инкубировали при 37oC в течение 1 ч. Планшеты промывали 3 раза PBS - 0,1% Tween, добавляли 100 мкл разведения 1:1000 стрептавидин-пероксидазы хрена в PBS - 0,1% Tween и инкубировали при 37oC в течение 30 мин. Планшеты промывали 3 раза, добавляли 50 мкл разведения 1:50 о-фенилдиамина в цитратном буфере pH 5,0 с 1 мкл/мл 30%-ной H2O2 и инкубировали при комнатной температуре в течение 20 мин. В каждой ячейке определяли поглощение при 495 нм.
Пример 3 /см. также раздел C/.
Мышей, применяемых в исследованиях по иммунизации, подвергали легкой анестезии эфиром перед внутрижелудочной иммунизацией. Затем 30 мкг субъединиц A и B рекомбинантной уреазы H.pylori, продуцировали в E.coli, связанной с гидроксиапатитом и дополненной холерным токсином, суспендировали в PBS и 200 мкл доставляли в желудок соответствующих мышей при помощи полиэтиленовой трубки, соединенной со шприцем для подкожных инъекций. Эту процедуру называют пероральной иммунизацией.
Оценивали протоколы трех пероральных иммунизаций. Протоколы описаны ниже.
Протокол C1 - Вакцинация субъединицей A рекомбинантной уреазы
Самок BALB/c шестинедельных мышей /10/ перорально иммунизировали 30 мкг очищенной субъединицы A рекомбинантной уреазы H.pylori и 1 мг гидроксиапатита и 10 мкг холерного токсина на 0, 8-ой, 14-ый и 21-ый дни. 10 мышей инфицировали на 32-ой, 34-ый и 36-ой дни 108 H.felis из жидкой культуры.
Протокол C2 - Вакцинация субъединицей B рекомбинантной уреазы
Самок BALB/c шестинедельных мышей /10/ перорально иммунизировали 30 мкг очищенной субъединицы B рекомбинантной уреазы H.pylori и 1 мг гидроксиапатита и 10 мкг холерного токсина на 0, 8-ой, 14-ый и 21-ый дни. Мышей инфицировали на 32-ой, 34-ый и 36-ой дни 108 H.felis из жидкой культуры.
Протокол C3 - Контроль
Самок BALB/c шестинедельных мышей /10/ перорально иммунизировали 1 мг гидроксиапатита и 10 мкг холерного токсина на 0, 8-ой, 14-ый и 21-ый дни. Мышей инфицировали на 32-ой, 34-ый и 36-ой дни 108 H.felis.
На 42-ой 106-ой день мышей убивали и брали множество биопсий их желудка.
Защита и оценка
Для оценки защиты биопсии корпуса и полости желудка скринировали на уреазную активность при помощи теста Tatrox HB /Rohm Pharma/ в соответствии с инструкциями поставщика. Уреазу определяли количественно путем спектрофотометрического измерения при 55% нм. Тотальные величины OD корпуса и полости желудка складывали для получения конечной величины OD для каждой мыши.
Литература
1. Blaser, M.J. "Gastric Campylobacter-like organisms, gastritis and peptic ulcer disease" Gastroenterology. 1987, 93, 371-383.
2. Graham, D.Y. "Campylobacter pylori and peptic ulcer disease" Gastroenterology. 1989, 196, 615-625.
3. Parsonnet, J. et al "Helicobacter pylori infection in intestinal and diffuse-type gastric adenocarcinomas" J.Natl. Cancer Inst. 1991, 93, 640-643.
4. Marshall, B.J. et al "Attempt to fulfill Koch's postulate for pyloric Campylobacter" Med. J. Aust. 1985, 142, 436-439.
5. Morris, A. et al "Ingestion of Campylobacter pyloridis causes gastritis and fasting gastric pH" Am. J. Gastroenterology, 1987, 82, 192-199.
6. Engstrand, L. et al "Inoculation of barrier-born pigs with Helicobacter pylori: a useful animal model for gastritis type B" Infect. Immun. 1990, 53, 1763-1768.
7. Fox, J.G. et al "Gastric colonization by campylobacter pylori subsp. mustelae in ferrets" Infect. Immun. 1988, 56, 2994-2996.
8. Fox, J.G. et al "Helicobacter mustelae-associated gastritis in ferrets: an animal model of Helicobacter pylori gastritis in humans" Gastroenterology 1990, 99, 352-361.
9. Lee, A, et al "A small animal model of human Helicobacter pylori active chronic gastritis" Gastroeneteroloy 1990, 99, 1315-1323.
10. Fox, J.G. et al "Helicobacter Felis gastritis in gnotobiotic rats: an animal model of Helicobacter pylori gastritis" Infect. Immun 1991, 59, 785-791.
11. Eaton, K.A. et al "Campylobacter pylori virulence factors in gnotobiotic piglets" Infect. Immun. 1989, 57, 1119-1125.
12. Peterson, W. L. "Helicobacter pylori and peptic ulcer disease" N. Engl. J. Med. 1991, 324, 1043-1048.
13. Czinn, S.J. and Nedrud, J.G. "Oral Immunization against Helicobacter pylori" Infect. Immun. 1991, 2359-2363.
14. Brandtzaeg, P. "Role of H chain and secretory component in receptor-mediated glandular and hepatic transport of immunoglobulins in man" Scand. J. Immunol. 1985, 22, 111-146.
15. Brandtzaeg. P. "Production and secreion of immunoglobulins in the gastrointestinal tract" Ann. Allergy 1987, 59, 21-39.
16. Wyatt, J. I. "Local immune response to gastritic campylobacter in non-ulcer dyspepsin" J. Clin. Path. 1986, 39, 863-870.
17. Lee, A. et al "Pathogenicity of Helicobacter pylori: A perspective" Infect. Immun. 1993, 61, 1601-1610.
18. Pallen, M. J. and Clayton, C.L. "Vaccination against Helicobacter pylori urease, letter" Lancet, 1990, 336, 186.
19. Evans, D.J. et al "Urease-associated heat shock protein of Helicobacter pylori Infect. Immun. 1992, 60, 2125-2127.
20. Ferrero, R.L. and Lee, A "The importance of urease in acid protection for the gastric-colonizing bacteria Helicobacter pylori and Helicobacter felis sp. nov." Microb. Ecol. Health Dis. 1991, 4, 121-134.
21. Chem, et al "Immunization against gastric Helicobacter infection in a mouse/Helicobacter felis model, letter" Lancet, 1992, 339, 1120-1121.
22. Czinn, S. et al "Oral immunization protects germ-free mice against infection from Helicobacterl felis". Proceedings of the DDW, American Gastroenterological Assocoation. May 10-13, 1992, 1321, A-331.
23. Guo, M. and Liu, P.V. "Serological specificities of ureases of Proteus species" J. Gen. Microbiol. 1965, 136, 1995-2000.
24. Michetti, P. et al "Specificity of mucosal IgAa response in Balb/c mice following H. felis or H. pylori challenges" Proceedings of the DDW, American Gastroenterology Association. May 10-13, 1992, 1001, A-251.
25. Davin, C. et al "H. pylori urease elicits protection against H. felis infection in mice" Proceedings of the DDW, American Gastroenterology Association, May 16-19, 1993, 1213, A-304.
26. Pallen, M. J. and Clayton, C.L. "Vaccination against Helicobacter pylori urease". Lancet 1990, 336, 186-7.
27. Pimentel, J. L. and Cook, M.E. "Improved growth in the progeny of hens immunized with jackbean urease" Poultry Sci. 1988, 64, 434-439.
28. Labigne, A. "Sequences of nucleotides coding for a protein having an urease activity". EPO patent application #EPO 0 367 644 A1, 1989 Intl publication # WO90/04030, 1990.
29. Clayton, C.L. et al. "Nucleotide sequence of two genes from Helicobacter pylori encoding for urease subunits". Nucleic Acid Res. 1990, 18, 362.
30. McGhee, J.R. and Kyono, H. "New perspectives in vaccine development: mucosal immunity to infections". Infect Agents Dis. 1993, 2, 55-73.9
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕН HELICOBACTER PYLORI И ВАКЦИННАЯ КОМПОЗИЦИЯ | 1996 |
|
RU2195463C2 |
| ЛАКТОСЫВОРОТКА ИММУННАЯ СПЕЦИФИЧЕСКАЯ ПРОТИВ HELICOBACTER PYLORI И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2000 |
|
RU2201256C2 |
| ОРАЛЬНОЕ ЛЕЧЕНИЕ ГЕЛИКОБАКТЕРНОЙ ИНФЕКЦИИ | 1993 |
|
RU2107513C1 |
| КОМПОЗИЦИЯ ДЛЯ ИНДУКЦИИ ИММУННОЙ РЕАКЦИИ У МЛЕКОПИТАЮЩЕГО И СПОСОБ ВВЕДЕНИЯ МЛЕКОПИТАЮЩЕМУ АКТИВНОГО ИНГРЕДИЕНТА | 1991 |
|
RU2126270C1 |
| СПОСОБ ИММУНОДИАГНОСТИКИ ЗАБОЛЕВАНИЙ, ВЫЗВАННЫХ HELICOBACTER PYLORI-ИНФЕКЦИЕЙ | 2014 |
|
RU2572717C1 |
| МИКРООРГАНИЗМ-НОСИТЕЛЬ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, КОДИРУЮЩИХ АНТИГЕНЫ И БЕЛКОВЫЕ ТОКСИНЫ | 2007 |
|
RU2447145C2 |
| Рекомбинантный белок для иммунизации против холеры | 2019 |
|
RU2723705C1 |
| ВНЕШНИЙ КОРОВОЙ ЭПИТОП ЛИПОПОЛИСАХАРИДА Н. pylori | 2010 |
|
RU2558257C2 |
| СПОСОБ ДОСТАВКИ БИОАКТИВНОГО АГЕНТА ЖИВОТНОМУ ДЛЯ ИНИЦИАЦИИ ИММУННОГО ОТВЕТА (ВАРИАНТЫ) | 1989 |
|
RU2127118C1 |
| СПОСОБ УСИЛЕНИЯ ИММУННОГО ОТВЕТА | 2010 |
|
RU2442604C1 |

Изобретение относится к профилактике и лечению желудочной инфекции у млекопитающих, в том числе у человека. В млекопитающем - хозяине индуцируют защитный иммунный ответ против инфекции Helicobacter путем введения иммуногенно эффективного количества уреазы или субъединиц уреазы Нelicobacter в качестве антигена. Для индуцирования защитного иммунного ответа на инфекцию Helicobacter используют вакцину, содержащую в качестве антигена полиаминокислотный препарат, представляющий эпитопы, проявляемые уреазой, эндогенной для организма. Оценку иммунного ответа у млекопитающих, инфицированных Helicobacter, осуществляют путем определения в пробе, отобранной из желудочно-кишечного тракта, присутствия антител, реагирующих с эпитопами, проявляемыми уреазой, эндогенной для Helicobacter. 4 с. и 23 з.п.ф-лы, 6 ил., 6 табл.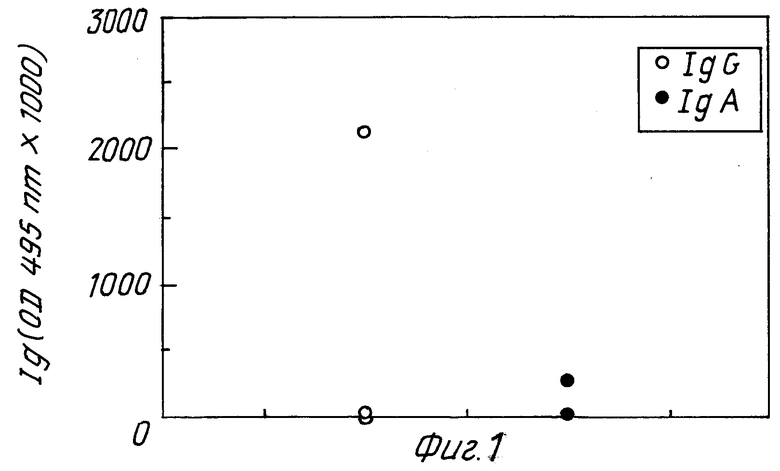
Приоритет по пунктам:
03.11.92 по пп.1 - 3, 5, 6, 8, 10 - 27;
06.07.93 по пп.4, 7, 9.
| Jnfection Jmmunidation, 1991, 2359-2363 | |||
| Dastroenterology, 1990, 99, 1315 - 1323 | |||
| EP 0367644 А1, 09.05.90. |

