Область техники, к которой относится изобретение
Изобретение относится к микроорганизмам как носителям гетерогенных нуклеотидных последовательностей, кодирующих антигены и белковые токсины, способу их получения, а также соответствующим плазмидам или векторам экспрессии. Данные микроорганизмы можно использовать как лекарственные средства, в частности, как противоопухолевые вакцины для лечения различных опухолей.
Уровень техники
Иммунотерапия рака дает многообещающую возможность лечения опухолей. Множество клинических исследований с использованием различных подходов сосредоточивается на ее эффективности у пациентов. В принципе показано различие между пассивной и активной иммунотерапией.
Активная иммунотерапия направлена на индукцию связанного с вакциной опухолеспецифического иммунного ответа. Последний в настоящее время исследуют в клинических условиях с использованием ряда различных подходов. Например, имеются так называемые цельноклеточные вакцины, исходным материалом для которых являются опухолевые клетки, которые либо получают непосредственно от пациента (аутологичные) или выделяют из соответствующих клеточных линий (гетерологичные). Затем данные клетки, как правило, инактивируют, дифференцированно проводят манипуляции и вводят (обратно вводят) пациенту.
Напротив, антигенспецифические вакцины включают один или более опухолеспецифических антигенов, частей антигенов или ДНК, кодирующую специфический антиген, а также так называемые антиидиотипические вакцины. Обычно данные вакцины не являются выделенными, но их инъецируют в комбинации с подходящим носителем. Следовательно, с одной стороны, используют различные классические адъюванты, но также используют комбинации с биологическими иммуностимуляторами, такими как цитокины.
С целью иммуностимуляции используют подходы, которые включают антиген-ассоциированные иммуностимуляторы, такие как столбнячный токсин. Более того, имеются попытки использования антигенов в комбинации с дендритными клетками. И, наконец, имеется ряд попыток использования рекомбинантных живых вакцин с вирусными или бактериальными носителями.
Слитые белки бактериальных токсинов, такие как столбнячный токсин, шига-токсин, летальный токсин или холерный токсин, в качестве адъювантов с антигеном используют как вакцины, особенно противоинфекционные, уже давно (см. статью Freytag и Clements, 1999). Кроме того, нативные токсины, часто слитые со специфической в отношении клетки молекулой-мишенью, такой как молекула клеточной поверхности опухолевых клеток, также используют с целью разрушения клеток-мишеней.
В таком случае, слитые белки с нативным токсином, который, как правило, включает ферментную группу и белок-связывающий домен, проявляют свой оптимальный эффект при использовании в качестве вспомогательных лекарственных веществ (см. статью Freytag и Clements, 1999). С помощью данных вакцин получают удовлетворительный иммунный ответ даже после иммунизации через слизистые и особенно после пероральной иммунизации. Сложности с данными слитыми белками заключаются в том, что нативные токсины являются высокотоксичными и, вследствие этого, не могут быть использованы для человека (см. статью Holmgren et al., 2005).
Целое направление исследований, таким образом, занимается детоксификацией токсинов, которые в то же время сохраняют эффект в качестве вспомогательных лекарственных веществ. Однако, поскольку в большинстве случаев данный эффект совпадает с ферментной активностью, которая ответственна за токсический эффект (см. статью Lycke et al., 1992), детоксификация не может быть проведена прямым путем, даже если это представляется возможным для некоторых токсинов, которые не теряют своей ферментной адъювантной активности (см. статьи Hormozi et al., 1999; Lycke et al., 1992).
В случае холерного токсина (СТ) предпринят ряд попыток детоксификации (см. статьи Agren et al., 1999; Byun et al., 2001; Eriksson et al., 2004; Kweon et al., 2002; Sanchez et al., 2002), в которых, однако, преобладает использование вспомогательного лекарственного вещества для слизистых (см. работу Freytag и Clements). Вследствие этого главным образом эффективная индукция ответа антител (в основном IgA слизистых), которые получают усиленную связанную с токсином поддержку Т-клеток с рестрикцией МНС (главного комплекса гистосовместимости) класса II, является основным предварительным условием для вспомогательного лекарственного средства для слизистых для вакцины, которая состоит из белка-антигена и слитого или совместно используемого токсина (см. статью Freytag и Clements).
Что касается холерного токсина, особенно его В-субъединицы (CtxB), то его тестируют как адъювант, поскольку он является ответственным за связывание с рецептором GM-1 и не проявляет токсических эффектов, будучи выделенным (см. статью Holmgren et al.). Слияния белка с CtxB характеризуются в основном индукцией так называемых Th2 иммунных ответов. Они представляют собой Т-клеточные ответы, которые главным образом характеризуются цитокинами, такими как ИЛ-4 или ИЛ-6, и которые в основном вызывают индукцию антител, но которые совсем не инициируют или наиболее ограниченно инициируют клеточный иммунный ответ, в частности, цитотоксических Т-клеток (CTL) (см. статью Holmgren et al.).
Кроме того, CtxB в качестве адъюванта для слизистых индуцирует системную толерантность к белку-антигену. Системная толерантность описывает истощение или инактивацию антиген-специфических лимфоцитов, в частности, Т-клеток или В-клеток. Подход данного типа, вследствие этого, неприменим для индукции системного иммунного ответа (см. статью Holmgren et al.).
В противоположность применению на слизистых внутрибрюшинное или подкожное применение слитого белка токсин-антиген способно индуцировать системный, а также низкий цитотоксический ответы. Это в действительности используют для противоопухолевой вакцинации в модельных системах (см., например, работу (Becerra et al., 2003)). Однако данный ответ также получают с самим очищенным антигеном и главным образом в зависимости от используемого адъюванта. За исключением того факта, что измеренные ответы CTL на модели являются очень низкими, отсутствуют доказательства того, что защита в той же мере зависит от данных эффектов. Более того, антиген не применяют перорально, но только путем прямой инъекции (s.c. (подкожной), i.d. (внутрикожно), i.m. (внутримышечно), i.p. (внутрибрюшинно)) антигена.
Антигенные белки, слитые с детоксифицированным токсином, как правило, неэффективны при применении в качестве противоопухолевой вакцины. Основными причинами являются, если они вообще имеются, лишь низкая индукция системного иммунного ответа или даже, в случае CtxB, индукция системной толерантности, а также индукция ограниченных слизистой антител и иммунных ответов типа Th2.
В статье McSorley et al., например, показано, что назальная иммунизация слитым белком CtxB-антиген (который для противоопухолевой вакцины представляет предпочтительный путь индукции системного ответа) предпочтительно переносится и, вследствие этого, инактивирует Th1-клетки, тогда как Th2-клетки не подвергаются воздействию (см. статью McSorley et al., 1998). Ответы Th2 характеризуются Т-хелперными клетками, которые преимущественно продуцируют ИЛ-4 или ИЛ-6. Данные цитокины особенно ответственны за инициацию продукции антител В-клетками, которые обеспечивают защиту в случае самых распространенных вакцин. Напротив, Т-клетки Th1 в основном секретируют ИЛ-2 и ИФН-γ соответственно цитокины, которые играют роль в клеточном иммунном ответе. В зависимости от цели стратегии иммунизации имеет большое значение, инициируется ли вызываемый антителами иммунный ответ Th2 (так называемый сдвиг Th2) или вызываемый клетками ответ Th1 (так называемый сдвиг Th1).
Напротив, системная индукция Th1-доминирующего клеточного иммунного ответа с помощью ИФН-γ, дающая Т-клетки хелперы, и индукция цитотоксических Т-клеток (CTL) является необходимой для иммунотерапии опухолей.
Предшествующий уровень техники показывает, что токсины могут действовать как адъюванты. В частности, холерный токсин (СТ) показывает сильный адъювантный эффект. Однако данный эффект существенно ослабляется, как только токсин детоксифицируют. Что касается его субъединицы CtxB, пероральное применение даже индуцирует системную толерантность. Данной проблемы можно частично избежать путем назального применения. Однако если используют назальное введение, возникают другие проблемы, в частности, такие как ассоциированные с субъединицей CtxB, затрагивающие экспрессию рецептора GM-1 в головном мозге (см. статью van Ginkel et al., 2000), которые выражаются в повышенном риске развития определенных тяжелых побочных эффектов (см. статью Mutsch et al., 2004).
Для преодоления данных проблем возможно генерировать рекомбинантные живые вакцины, которые соэкспрессируют токсины и (гетерологичные) антигены. Данная возможность уже рассматривалась в связи с противоинфекционными вакцинами, т.е. вакцинами, которые направлены на специфический патоген, например на возбудитель туберкулеза, и предназначены для индукции иммунитета против данного патогена.
В процессе разработки данных противоинфекционных вакцин уже получены рекомбинантные токсины, слитые с соответствующими (гетерологичными) антигенами, с использованием ряда рекомбинантных бактериальных штаммов, которые вводят перорально как в случае живой, так и инактивированной вакцины. В большинстве случаев генерированные штаммы экспрессируют только рекомбинантный токсин. Вследствие этого данные подходы в основном направлены на иммунизацию исключительно против самого токсина (индукцию ответа антител) и, следовательно, штамма, экспрессирующего токсина (патогена). Данные типы вакцин непригодны для применения в качестве потенциальных противоопухолевых вакцин при лечении опухолей и неудивительно, что в соответствующих испытаниях не упоминают о какой-либо возможности данного применения (см. статью Reveneau et al., 2002), (см. статью Vertiev et al., 2001), (см. статьи Freytag и Clements, 1999; Jackson et al., 1996).
Что касается данных противоинфекционных вакцин, в процессе индукции ответа антител токсин не работает как адъювант, в данном случае преобладает индукция иммунного ответа антител слизистых. Данные испытания противоинфекционных вакцин не включают или включают только очень слабый системный иммунный ответ. В противоположность противоопухолевым вакцинам данные вакцины не осуществляют индукцию клеточного иммунного ответа, особенно цитотоксических Т-клеток. Бактериальные патогены, экспрессирующие токсин, обычно относятся к внеклеточному живому типу и, вследствие этого, не участвуют в активации CTL, т.е. защита, представляемая данными противоинфекционными вакцинами, является CTL-независимой.
В упомянутых испытаниях вакцин использованы следующие бактериальные штаммы: рекомбинантные Lactobacilli (см. статью Reveneau et al., 2002), Listeria (см. статью Vertiev et al., 2001), Bacillus anthracis (см. статьи Mendelson et al., 2005; Mesnage et al., 1999), Shigellae (см. статьи Andersen et al., 2000; Tzschaschel et al., 1996b), E. coli (см. статью Walker et al., 1992), Vibrio (см. статьи Butterton et al., 1995; Chen et al., 1998; Thungapathra et al., 1999) и Salmonella (см. статью Jackson et al., 1996).
Кроме того, пытались усилить иммунные ответы слизистых путем поверхностной презентации (см. статью Konieczny et al., 2000) или секреции токсина в качестве гетерологичного антигена (Salmonella, Shigellae (см. статьи Garmory et al., 2003; Orr et al., 1999; Su et al., 1992; Tzschaschel et al., 1996a; Tzschaschel et al., 1996b), Yersinia (см. статью Sory и Cornelis, 1990), Vibrio (cm. статью Ryan et al., 1997a), E. coli (см. статью Zhu et al., 2006)). Однако в данных дополнительных случаях сами токсины представляют антигены, в отношении которых направлен иммунный ответ.
Таким образом, следует отметить, что токсины не действуют как адъюванты и в описанных исследованиях не планируют экспрессировать токсины в виде слитых белков с дополнительным отдельным (гетерологичным) антигеном. Более того, слитые белки с СТ или CtxB не были генерированы с данными системами, и это не предполагалось.
Следовательно, слитые белки, упомянутые в вышеприведенных документах, представляют собой слитые белки пептидного сигнала секреции, такого как HlyA, и белка токсина, а не слитые белки, состоящие из токсина и (гетерологичного) антигена.
Главной целью данных исследований является просто получить оптимальный иммунный ответ слизистых (см. статью Tzschaschel et al., 1996a). Индукцией системного иммунного ответа не занимаются. До настоящего времени, если сравнивать в целом, анализы системных иммунных ответов были ограничены только антителами (в частности, системными IgA), поскольку защита в моделях данного типа главным образом опосредуемая антителами (ср., например, [31]). Индукция системного клеточного иммунного ответа, в частности ответа цитолитических Т-клеток, не описана. Слияние с сигналом секреции в основном использовали для повышения растворимости токсинов и повышения их стабильности соответственно, поскольку сильная абсолютная цитоплазматическая экспрессия часто приводит в результате к продукции нерастворимых агрегатов (см. статью Gentschev et al., 2002a).
В данном аспекте ряд авторов обращает внимание на то, что белки с сигналом секреции вызывают быстрое цитоплазматическое разложение (см. статью Tzschaschel et al., 1996a), тогда как другие наблюдают стабилизацию (см. статью Orr et al., 1999). Предшествующий уровень техники, следовательно, в данном случае имеет противоречия. До настоящего времени предшествующие эксперименты с секретируемыми токсинами (токсин + сигнал секреции), прежде всего, направлены на повышение стабильности, которая очевидным образом не достигается во всех случаях.
Одна причина безусловно обнаружена в изменяющейся интенсивности экспрессии и стабильности плазмид. В статье Tzschaschel et al. описано, что используемая плазмидная система высоко нестабильна и, кроме того, без селекции просто найдена в нескольких бактериях. В качестве возможного решения авторы используют хромосомную интеграцию посредством минитранспозона (см. статьи Tzschaschel et al., 1996a; Tzschaschel et al., 1996b).
Однако данная система имеет ряд недостатков. С одной стороны, точка интеграции не определена, что может привести к нежелательному фенотипическому изменению штамма-хозяина (например, повышенной/пониженной экспрессии фланкирующих генов). С другой стороны, ожидаемый уровень экспрессии при использовании одной геномной копии только низкий, что оказывает отрицательное воздействие на иммуногенность. Кроме прочего, хромосомная интеграция транспозона относительно нестабильна, поскольку она очень часто приводит к спонтанным эксцизиям по сторонам повторяющихся элементов.
Как отмечено выше, предшествующий уровень техники является противоречивым в отношении стабильности секретируемого гетерологичного токсина.
В статье Garmory et al. даже предполагают, что секреция гетерологичного антигена не обладает каким-либо особенным преимуществом в отношении иммуногенности (см. статьи Garmory et al., 2002; Roland et al., 2005). В других случаях повышенный системный ответ антител на секретируемый токсин после внутривенного введения в действительности наблюдают, но не после перорального введения. Напротив, пероральное введение, пусть даже косвенно, является частью проблемы (см. статью Roland et al., 2005).
В конечном счете исследования с грамположительным штаммом (Lactobacillus plantarum), который продуцирует столбнячный токсин, не показывают существенных различий в индукции системного ответа антител по сравнению со штаммами, которые секретируют токсин, присутствует ли он в связанном с их мембраной виде, или они содержат токсин в цитоплазме (см. статью Reveneau et al., 2002).
Системные клеточные иммунные ответы, в частности ответы цитотоксических Т-клеток, и полученная в результате защита не исследованы и не описаны.
Таким образом, на основании вышеприведенных данных по предшествующему уровню техники, касающихся противоинфекционных вакцин на основе токсинов, нельзя утверждать, представляет ли секреция токсина, используемого в качестве адъюванта, преимущество в плане индукции системного (клеточного) иммунного ответа. Напротив, вышеприведенные исследования скорее указывают на проблему стабильности и отмечают потерю положительного эффекта секретируемого гетерологичного токсина. Слитые белки, состоящие из сигнала, токсина и гетерологичного антигена, совсем не описаны или не предполагаются.
В вышеприведенных абзацах представлен предшествующий уровень техники, который описывает бактериальные носители, которые гетерологично экспрессируют токсины и могут быть использованы в качестве противоинфекционных вакцин. В приведенных примерах осуществлены в основном модификации, касающиеся экспрессии или стабильности токсинов и их растворимости, например инсерция сильного экспрессирующего промотора или слияния токсина с сигналом секреции.
Другие авторы получают также исследованные генетические слияния токсинов с гетерологичными антигенами в живых вакцинах. В данных случаях токсин в основном используют в качестве адъюванта. В ряде случаев (например, см. статью Brassier et al., 2000) гетерологичный антиген действует как адъювант, а токсин как подходящий антиген.
Однако важно отметить, что в упомянутых случаях экспрессия конструкции слитых генов токсина-антигена происходит исключительно цитоплазматически или периплазматически. Конструкция токсин-антиген не слита с дополнительным сигналом секреции (который может привести к его полной секреции) и не секретируется непосредственно.
В процессе получения данных конструкций слитых генов токсина-антигена используют рекомбинантную Е. coli (см. статью Clemens et al., 2004), Bacillus anthracis (см. статью Brassier et al., 2000), Shigella (см. статьи Koprowski et al., 2000; Ranallo et al., 2005; Zheng et al., 2005) и штаммы Vibrio (см. статью Silva et al., 2003). Для Salmonella (см. обзор в статье (Garmory et al., 2002)) описаны также слияния вариантов СТ с антигенами (см. статьи Hajishengallis et al., 1996; Huang et al., 2000) или других токсинов с антигенами (см. статьи Barry et al., 1996; Cardenas и Clements, 1993; Chabalgoity et al., 1997; Chabalgoity et al., 1996; Chabalgoity et al., 2000; Chabalgoity et al., 1995; Chacon et al., 1996; Jagusztyn-Krynicka et al., 1993; Khan et al., 1994a; Khan et al., 1994b; Lee et al., 2000; Pogonka et al., 2003; Schodel et al., 1990; Smerdou et al., 1996; Ward et al., 1999; Wu et al., 2000).
В большинстве данных случаев основное внимание сосредоточено на индукции иммунного ответа слизистых (антител), и для индукции системного иммунного ответа выбирают только подкожное, но не пероральное применение [36].
Несколько работ с Salmonella в качестве поддерживающего штамма ограничены простой характеризацией штамма (см. статьи Gomez-Duarte et al., 1995; Jagusztyn-Krynicka et al., 1993), в других только анализируют ответ антител слизистых и/или системный ответ антител и/или защиту (см. статьи Barry et al., 1996; Cardenas и Clements, 1993; Dunstan et al., 2003; Hajishengallis et al., 1996; Harokopakis et al., 1997; Khan et al., 1994a; Khan et al., 1994b; Pogonka et al., 2003; Smerdou et al., 1996; Somner et al., 1999). Во всех данных случаях, где попутно используют слияние антигена и столбнячного токсина и которые используют исключительно в качестве противоинфекционных вакцин, системные клеточные иммунные ответы, особенно ответы цитотоксических Т-клеток, не изучали.
Вследствие этого, с иммунологической точки зрения из данных исследований нельзя сделать никаких заключений относительно потенциального применения в качестве противоопухолевой вакцины, поскольку они сосредоточены на опосредованных антителами эффектах в качестве противоинфекционных вакцин.
Исследования, которые включают анализ изотипа исследуемого иммунного ответа, по-видимому, являются более подходящими. В действительности в данных случаях непосредственно измеряют клеточные иммунные ответы, но изотипический профиль ответа антител позволяет сделать заключение относительно сдвига Th1/Th2 иммунного ответа. Изотипы антител, подобных lgGI, связаны с ответами Th2, и изотипы, подобные lgG2a, ассоциированы с ответами TM. Как уже отмечали, ответы ТМ представляют собой преобладающие клеточные иммунные ответы, тогда как ответы Th2 в основном представляют гуморальные ответы, направляемые антителами. Кроме того, в данных исследованиях не описывают противоопухолевые вакцины, и в них не предполагают какого-либо применения противоопухолевого агента.
Одно исследование противоинфекционной вакцины на основе Salmonella в качестве носителя, которая экспрессирует слитый белок столбнячного токсина и антигена, осуществлено на собаках. Низкие уровни ответов антител, которые индуцированы у собак, показывают сдвиг ТМ, касающийся профиля антител, следовательно, ответа, который сильно коррелирует с иммунным ответом клеточного типа (см. статью Chabalgoity et al., 2000). Надо отметить, что иммунология собак недостаточно изучена и, вследствие этого, неясно, в какой степени профиль антител собаки может дать информацию относительно сдвига Th1.
Исследования на мышах, проведенные той же группой с использованием сравнимых конструкций, показали тем не менее профиль антител с одинаковым уровнем для lgG1, что для lgG2a, что будет указывать на смешанный ответ Th1/Th2.
Интересно, что имеющийся иммунитет к столбнячному токсину, который обнаружен у большинства людей вследствие предшествующих иммунизации, вызывает иную относительно сильную индукцию lgG1, тогда как lgG2a почти не индуцируется. Это ясно указывает на другое соотношение Th2 (см. статью Chabalgoity et al., 1995). По данной причине живая вакцина на основе столбнячного токсина в качестве противоопухолевой вакцины является очень вредной для использования на человеке. Вследствие этого можно ожидать, что сильный вызываемый антителами ответ Th2 индуцируется у большинства данных пациентов, которые демонстрируют специфический ответ на столбнячный токсин.
Только в некоторых исследованиях также анализируют клеточный иммунный ответ и сравнивают генетические конструкции со слитым токсином и без него. В одном случае, например, слияние антигена со столбнячным токсином и без него сравнивают на Salmonella (см. статью Lee et al.). В данном случае слитая конструкция столбнячного токсина и антигена главным образом повышает общий уровень антител, тогда как профиль Th1/Th2 мало изменяется. Даже антиген-специфическая секреция Т-клетками CD4+ типичных цитокинов Th1, таких как ИФН-γ и ИЛ-2 соответственно, показывает только слабое отклонение. В более раннем исследовании той же группы измеряют также уровни ИФН-γ. Однако не представлено никакого сравнения с конструкциями без столбнячного токсина (см. статью Chabalgoity et al.). В других исследованиях с различными грамположительными бактериальными носителями, подобными Shigella (см. статьи Koprowski et al., Ranallo et al., Zheng et al.) или Vibrio (см. статьи Campos et al., Ryan et al.), не анализируют ни изотипы, ни клеточные иммунные ответы. Подводя итоги, можно утверждать, что упомянутые исследования ясно сосредоточены на индукции направляемого антителами гуморального иммунного ответа. В действительности используют генетические конструкции токсина и антигена, но они не имеют сигнала секреции и они непосредственно не секретируются. Однако ни при каких условиях не анализировали системные клеточные иммунные ответы, в частности ответы цитотоксических Т-клеток. К тому же данные клеточные ответы цитотоксических Т-клеток не могут быть выведены из гуморального ответа антител и не могут быть определены, если ответ антител включает Th1/Th2.
Однако, это именно те клеточные иммунные ответы цитотоксических Т-клеток, которые важны для использования в лечении путем противоопухолевой вакцинации.
Тем не менее, в плане слияний токсин-антиген, которые ограничены только противоинфекционными вакцинами для слизистых, состояние уровня техники не допускает никакого утверждения относительно возможного использования каких-либо данных конструкций в качестве противоопухолевых вакцин.
Как уже отмечено, экспрессию генетических слитых конструкций осуществляют без помощи системы секреции. Токсины и конструкции токсин-антиген, соответственно, как правило, расположены цитоплазматически, а также периплазматически, т.е. между двумя мембранами. Для индукции эффективного клеточного иммунного ответа токсин должен быть свободно доступен для антиген-презентирующей клетки (АРС). Как правило, нативный токсин продуцируется в периплазме грамотрицательных бактерий. Это достаточно для полных иммунных ответов слизистых, поскольку периплазматические токсины могут выйти из периплазмы в толстой кишке и, следовательно, также являются доступными (см. статью Hunt и Hardy, 1991). Однако это не принимают во внимание, если носитель направлен на антигенпрезентирующие клетки вне толстой кишки, такие как, например, пейеровы бляшки или лимфатические органы, такие как лимфатические узлы или селезенка.
В принципе, два фактора являются важными в плане эффективности противоопухолевой вакцины: индукция клеточного иммунного ответа Th1-типа и участие компонентов врожденной иммунной системы, таких как NK-клетки, NKT-клетки и γ-δ Т-клетки, которые играют важную роль в плане эффективности противоопухолевой терапии (см. статью Dunn et al., 2004).
Важность данных компонентов врожденных иммунных систем лежит на множестве уровней. Должным образом активированные NK- и γ-δ Т-клетки способны локально продуцировать большие количества ИФН-γ. Данный интерферон, который также продуцируется специфическими Th1-поляризованными Т-клетками, имеет множество функций, относящихся к терапии опухолей. Одной из его главных функций является ингибирование ангиогенеза, которое перекрывает снабжение опухоли кислородом и питательными веществами и приводит фактически к голоданию опухоли. Кроме того, NK-клетки имеют рецепторы, которые распознают молекулы МНС класса I. Если данные молекулы присутствуют на клетке, NK-клетки ингибируются.
Что касается вакцины, которая индуцирует специфические цитотоксические Т-клетки, то опухолевые клетки могут быть уничтожены данными CTL. Если опухолевая клетка теряет свою способность экспрессировать молекулы МНС класса I, что очень часто имеет место в опухолях, специфические цитотоксические Т-клетки неэффективны. Вследствие этого в данном случае ингибирование NK-клеток прекращается, и они способны непосредственно элиминировать опухолевые клетки.
Следовательно было бы идеальным, если бы противоопухолевая вакцина эффективно индуцировала оба компонента. Имеются противоречивые данные относительно сдвига Th1-Th2 слитого токсина-адъюванта. Как обсуждалось ранее, слитые конструкции токсин-антиген, используемые в выделенном виде, очевидным образом индуцируют сильный Th2-поляризованный иммунный ответ. Ряд авторов, кроме того, описывает низкий уровень сдвига для живых носителей; другие авторы отмечают небольшой Th1-сдвиг.
Однако данные результаты снова основаны исключительно на несекретируемых конструкциях. Индукцию врожденного иммунитета с помощью данных типов противоинфекционных вакцин никогда не сравнивали и не предусматривали.
Как уже отмечено, главная причина состоит в том, что имеющиеся вакцины представляют собой противоинфекционные вакцины для слизистых, а не противоопухолевые вакцины. Вследствие этого индукция Th1-иммунных ответов, иммунных ответов CTL и ответов врожденной иммунной системы не были в центре внимания. Напротив, что касается противоопухолевых вакцин, индукция данных иммунных ответов обязательна.
Интересно, что в биологии клетки нативные токсины обычно используют как ингибиторы путей передачи сигнала. Так, среди прочего показано, что нативный токсин коклюша, но не холерный токсин, способен ингибировать определенный тип апоптоза NK-клеток (см. статью Ramirez et al., 1994). Другое научное исследование способно показать, что холерный токсин, но не его В-субъединица, блокирует специфические функции NK-клеток (см. статью Poggi et al., 1996). Нативный токсин коклюша используют для ингибирования хемотаксиса лимфоцитов (см. статью Spangrude et al., 1985). Даже если данные исследования не относятся к противоопухолевой вакцинации, компетентный специалист в области техники мог бы сделать заключение о том, что использование токсинов в качестве противоопухолевых вакцин было бы вредным, поскольку ответ врожденной иммунной системы, важный для терапии опухолей, более ингибируется, чем индуцируется.
Другие научные исследования способны показать, что токсины, подобные нативному токсину коклюша (не инактивированному токсину коклюша), эффективно индуцируют компоненты врожденной иммунной системы. Касательно иммунотерапии опухолей это будет означать, что если их вообще будут использовать, то следует использовать нативные токсины. Однако по причинам токсичности данный тип введения недопустим. Кроме того, данная индукция неизбежно приводила бы к Тh2-направляемому вторичному иммунному ответу, который в свою очередь будет вредным для лечения опухолей (см. статью Boyd et al., 2005). Следовательно, касательно индукции врожденного иммунного ответа, в предшествующем уровне техники не описана и не предусмотрена противоопухолевая вакцинация. Напротив, критический анализ литературы даже мешает использованию токсинов при лечении опухолей.
Интересным, тем не менее, является анализ синергического эффекта токсинов или их субъединиц с другими стимуляторами, такими как иммуностимулирующие олигонуклеотиды ДНК с гипометилированными мотивами CpG (CpG ODN) (см. статью Holmgren et al., 2005) или липосахариды (LPS). В случае LPS в основном индукция моноцитов, по-видимому, повышается главным образом посредством В-субъединицы токсинов, тогда как она ингибируется токсином в целом (голотоксином) (см. статью Hajishengallis et al., 2004). Однако данные исследования основаны исключительно на использовании очищенных слитых конструкций токсин-антиген, к которым добавляют субстанции, подобные LPS или CpG в качестве адъювантов. Кроме того, анализ в данном случае проводят только на макрофагах, которые индуцируют адаптивный иммунный ответ, но непосредственно не действуют на опухоли. Следовательно, данные исследования не имеют существенного значения касательно индукции компонентов врожденной иммунной системы, в частности, NK-клеток, которые могут непосредственно действовать на опухоли.
Другие исследования, однако, показывают, что NK-клетки могут быть активированы и хемотаксически привлечены токсинами, подобными экзотоксину A Pseudomonas aeruginosa (см. статью Muhlen et al., 2004). В зависимости от экспериментальной системы можно также индуцировать Th1-ответы, хотя вследствие этого главным образом происходит супрессия NK-клеток и Th1-ответов (см. статью Michalkiewicz et al., 1999). Тем не менее, данные исследования в основном направлены на анализ гепатотоксичности энтеротоксина А и не относятся к противоопухолевой вакцинации. Интересно, что эффекты являются в высокой степени дозозависимыми, и только незначительное изменение дозы может инвертировать эффекты. Хотя авторы не могли показать, какой ответ эффективно имеет место in vivo. Вследствие этого данные не дают предсказания, какие эффекты могут иметь место, если токсин или даже детоксифицированный токсин используют в качестве адъюванта.
В общем и целом иммунный ответ в сильной степени зависит от используемой конкретной системы. В большинстве случаев противоинфекционая вакцина для слизистых направлена на локальную манипуляцию с иммунной системой в слизистой с целью индукции эффективного иммунного ответа в слизистых (см. статью Lycke, 2005). Однако данные исследования не включают разработку противоопухолевой вакцины. Кроме того, в данных исследованиях отсутствует информация об индукции системных клеточных иммунных ответов, в частности ответов цитотоксических Т-клеток, которые являются основными для противоопухолевой вакцинации.
Уже показано, что секреция гетерологичного антигена дает преимущества системному иммунному ответу (см. статью Hess et al., 1996). Однако описанные секретируемые антигены не являются секретируемыми конструкциями токсин-антиген; токсины или их субъединицы не используют. Достигаемые таким образом иммунные ответы в модели трансгенной опухоли также в высокой степени ограничены (см. статью Gentschev et al., 2005). Действительно, в данном случае могут быть индуцированы слабые ответы антител и цитотоксических Т-клеток, которые частично защищают от развития опухоли. Однако, не только сами иммунные ответы, но и сама защита ограничена. Аналогично в данных исследованиях противоопухолевой вакцинации отсутствует сравнение с несекретируемыми конструкциями. Более того, сравнительные исследования не проводили в контексте противоопухолевой вакцинации (см. статью Hess et al., 1996) и, они противоречат дальнейшим исследованиям, в которых не наблюдают какого-либо преимущества в отношении секреции (см. статьи Garmory et al., 2002; Roland et al., 2005). В итоге и как отмечено ранее, предшествующий уровень техники очень противоречив в плане секреции и в общем и целом не дает никакого намека касательно потенциальных преимуществ секретируемых конструкций токсин-антиген при лечении опухолей. Напротив, критический анализ имеющейся литературы в значительной мере противоречит данному типу применения.
Бактериальные токсины (см. статью Todar, 2002): на химическом уровне существует два типа бактериальных токсинов, липополисахариды, которые связаны с клеточными стенками грамотрицательных бактерий, и белки, которые высвобождаются из бактериальных стенок и могут действовать на тканевые участки, удаленные от области бактериального роста. Связанные с клеткой липополисахаридные (LPS) токсины называют эндотоксинами, и внеклеточные способные к диффузии токсины называют экзотоксинами.
Экзотоксины, как правило, представляют собой растворимые белки, секретируемые живыми бактериями в период экспоненциального роста, но в ряде случаев они высвобождаются путем лизиса бактериальной клетки. Продукция токсина, как правило, специфична в отношении определенных видов бактерий, которые вызывают заболевание, ассоциированное с токсином (например, только Clostridium tetani продуцирует столбнячный токсин; только Corynebacterium diphtheriae продуцирует дифтерийный токсин). Как грамположительные, так и грамотрицательные бактерии продуцируют растворимые белковые токсины.
В общем имеется три класса белковых (экзо)токсинов: (i) токсины типа I (суперантигены), которые связываются с поверхностью клетки-хозяина и модулируют иммунный ответ, но не транслоцируются в клетку, (ii) токсины типа II (порообразующие токсины), которые действуют на мембрану клетки-хозяина и вызывают вытекание из клетки-хозяина и ее гибель, и (iii) токсины типа III (A-B токсины), которые связываются с клеткой-хозяином посредством специфического рецептора и транслоцируются в клетку, становятся в ней активными и модифицируют белки или другие компоненты клетки-хозяина.
Как показано выше, токсины типа III, действующие внутриклеточно в отношении клеток-хозяев, состоят из двух компонентов: один компонент (субъединица А) отвечает за ферментную активность токсина, другой компонент (субъединица В) участвует в связывании со специфическим рецептором на мембране клетки-хозяина и переносе фермента через мембрану. Ферментный компонент неактивен до тех пор, пока не высвободится из нативного (А+В) токсина. Выделенные А-субъединицы ферментативно активны, но лишены способности связывания и входа в клетку. Выделенные В-субъединицы могут связываться с клетками-мишенями (и даже блокировать связывание нативного токсина), но они нетоксичны.
Существует ряд путей, которыми можно синтезировать и собрать субъединицы токсина: А+В показывает, что токсин синтезирован и секретируется в виде двух отдельных белковых субъединиц, которые взаимодействуют на поверхности клетки-мишени; А-В или А-5В или АВ5 показывает, что А- и В-субъединицы синтезируются раздельно, но связаны нековалентными связями во время секреции и связываются со своей мишенью; 5В ил В5 показывает, что домен связывания белка состоит из 5 идентичных субъединиц. АВ или А/В показывает токсин, синтезированный в виде одного полипептида, разделенного на домены А и В, которые могут быть разделены путем протеолитического расщепления. Примерами токсинов АВ или А/В являются дифтерийный токсин, экзотоксин А, ботулинический токсин и столбнячный токсин. Примерами токсинов А-5В или АВ5 являются холерный токсин и шига-токсин, тогда как токсин сибирской язвы LF и токсин сибирской язвы EF являются примерами токсинов А-В.
Дальнейшие имеющие отношение к делу документы предшествующего уровня техники включают следующие:
Michl et al. описывают использование бактерий и бактериальных токсинов в качестве лечебных факторов для солидных опухолей. Раскрывают слитые конструкции токсин-антиген, а также бактериальную направленность данной конструкции. Исследуют использование дифтерийного токсина (DT), экзотоксина A Pseudomonas (РЕ) и энтеротоксина Clostridium perfringens (СРЕ). Однако авторы не упоминают использование холерного токсина и не показывают или не представляют заметную секрецию слитых конструкций токсин-антиген, выделенных из бактерий (см. статью Michl и Gress, 2004).
Lahiri представляет обзор по различным бактериальным токсинам и обсуждает их разнообразное применение. Хотя автор упоминает слитые белки токсин-антиген, он не говорит о холерном токсине и бактериальной направленности секретируемых слитых конструкций токсин-антиген (см. статью Lahiri, 2000).
Lavelle et al. раскрывает молекулы инфекционных агентов в качестве иммуномодулирующих лекарственных препаратов. Авторы также отмечают слитые белки холерного токсин-антигена, поданные конструкции используют только непосредственно как белки и не в качестве генетически модифицированных живых вакцин (см. статью Lavelle et al., 2004).
Заявка WO 01/74383 направлена на химерные иммуногены для слизистых типа антиген-энтеротоксин и, кроме того, в ней упоминают использование субъединиц холерного токсина А2 и В. Однако, данные химерные иммуногены всегда включают субъединицы А2 и В одновременно и предназначены для использования в иммунизации через слизистые, но не в лечении опухолей.
Заявка WO 02/077249 описывает авирулентные мутантные штаммы Yersinia enterocolitica, предназначенные для доставки гетерологичных белков к специфическим мутантным клеткам-мишеням. Кроме того, упоминают использование субъединицы А1 холерного токсина, но патентный документ не говорит о секреции и относится только к лечению инфекций и инфекционных состояний.
Заявка WO 2004/018630 раскрывает рекомбинантные двухцепочечные PHK-фаги, кодирующие двухцепочечную эукариотическую экспрессирующую кассету. Хотя упоминают субъединицу А холерного токсина, документ не имеет дальнейшей значимости. Holmgren et al. дают краткий обзор, касающийся области иммунизации слизистых и адъювантов. Авторы обсуждают в числе прочего эффекты холерного токсина как адъювантов для слизистых, но не раскрывают информацию о генетических экспрессирующих системах или живых вакцинах и, кроме того, не говорят о противоопухолевой терапии (см. статью Holmgren et al., 2003).
Holmgren и Czerkinsky приводят также обзор по иммунитету слизистых и вакцинам. Однако данная статья ограничена только противоинфекционными факторами и в ней не обсуждают или не представляют очевидное возможное применение в области противоопухлевой терапии (см. статью Holmgren и Czerkinsky, 2005).
В другом обзоре, написанном Freytag и Clements, обсуждают адъюванты для слизистых в плане применения в противоинфекционной иммунотерапии. Хотя холерный токсин отмечают в качестве адъювантов для слизистых, авторы не говорят о секретируемых конструкциях токсин-антиген и противоопухолевой терапии как возможной области применения (см. статью Freytag и Clements, 2005).
Shaw и Starnbach описывают использование модифицированных бактериальных токсинов для доставки вакцинных антигенов. Однако в статье не упоминают холерный токсин и, кроме того, ограничиваются прямым применением слитых белков токсин-антиген для целей вакцинации (см. статью Shaw и Starnbach, 2003).
Заявка WO 03/072789 направлена на микроорганизмы как носители нуклеотидных последовательностей, кодирующих клеточные антигены, используемые для лечения опухолей. Хотя патентный документ упоминает секрецию и использование в области противоопухолевой терапии, он совсем не говорит о бактериальных токсинах и слитых белках. В работах Gentschev, Dietrich и Goebel, а также Gentschev et al. описывают бактериальную направленность и ее использование в разработке противоопухолевой вакцины. Однако в двух данных документах не упоминают о применении бактериальных токсинов и слитых белков в противоопухолевой терапии (см. статьи Gentschev et al., 2002a; Gentschev et al., 2002b).
Заявка WO 98/23763 раскрывает клетки Vibrio cholerae, экспрессирующие субъединицы В и D гемолизина E. coli наряду со слитым полипептидом, который включает гетерологичный антиген, слитый с hylA. Далее описан вакцинный штамм Vibrio cholerae, который экспрессирует субъединицу В холерного токсина и слитый полипептид секреторной сигнальной последовательности, гетерологичного антигена и субъединице А2 холерного токсина. Наконец, описан слитый полипептид, который включает субъединицу В холерного токсина, слитую с антигенной частью субъединицы А или В токсина С.difficile. Однако в патентной заявке не упоминают использование слитого белка белкового токсина с антигеном гетерологичного небелкового токсина в противоопухолевой терапии.
Dietrich с соавт. обсуждают два инструмента для доставки вакцины - гемолизин А и Ns-териолизин, которые можно использовать для получения опосредованного клетками иммунитета. Однако не упоминают никаких слитых белков типа белковый токсин - гетерологичный антиген или совместной экспрессии (см. статью Dietrich et al., 2003).
Gentschev et al. описывают использование системы секреции α-гемолизина Escherichia coli для доставки антигена в вакцинном штамме Salmonella typhi Ty21. Однако авторы не упоминают об использовании бактериальных токсинов и слитых белков в противоопухолевой терапии (см. статью Gentschev et al., 2004).
Заявка WO 02/47727 направлена на лечебные факторы, включающие В-субъединицу белкового токсина. Документ раскрывает только слитые белки CtxB и EtxB с вирусным антигеном. Не упоминают никакую бактериальную вакцину или доставку бактериальной вакцины.
Cheng-hua S и соавт. описывают слияние генов В-субъединицы холерного токсина и эпитопа HBV PreS2 и антигенность слитого белка в исследованиях прямой иммунизации. Однако не упоминают никакую бактериальную вакцину или доставку бактериальной вакцины (см. статью Cheng-hua et al., 1995).
Sanchez et al. раскрывают, что ген В-субъединицы холерного токсина повышает цитотоксические ответы иммуноглобулина А, Th1-типа и CD8+ при внутрикожном совместном введении с ДНК-вакциной (см. статью Sanchez et al., 2004). Однако в данном подходе авторы используют ДНК в качестве носителя, который действует как стимулирующий Th1 адъювант для самой себя. Кроме того, белки продуцируются клетками-хозяевами и не доставляются непосредственно или с помощью бактериального носителя и, вследствие этого, непосредственно доступны для эукариотических клеток.
Заявка WO 01/29233 направлена на химерные иммуногенные композиции и кодирующие их нуклеиновые кислоты. Однако не упоминают никакую бактериальную вакцину или доставку бактериальной вакцины.
Заявка WO 2007/044406 относится к способам стимуляции иммунного ответа с использованием системы доставки бактериального антигена, которая основана на SopE, несущем сигнал секреции типа III. Однако в патентной заявке не упоминают использование бактериальных токсинов и слитых белков в противоопухолевой терапии.
В итоге, из предшествующего уровня техники можно заключить, что невозможно создать нативные токсины для использования у человека вследствие их сильной токсичности. Кроме того, их использование в противоопухолевой терапии было бы вредным, поскольку подавлялся бы ответ врожденной иммунной системы, в частности NK-клеток. Однако, это тот иммунный ответ, который является важным компонентом успешной противоопухолевой терапии, поскольку опухолевые клетки очень часто утрачивают свою способность экспрессировать молекулы МНС класса I и вследствие этого устойчивы к распознаванию и атаке CTL.
Использование субъединиц детоксифицированного токсина, с другой стороны, отдельно или слитых с (гетерологичными) белками антигенов приводит в результате к сильно аттенюированному адъювантному эффекту и/или даже индуцированной системной толерантности иммунной системы, а также ограниченным слизистыми иммунным ответам антител и Th2-типа.
Кроме того, секреция слитых белков (гетерологичный) антиген-токсин, которая описана только в разделе противоинфекционных вакцин (т.е. затрагивает антиген или даже сам токсин), как считают, не только не проявляет преимуществ относительно цитоплазматической экспрессии, но, в общем, скорее является неприемлемой. В общем и в целом не представлен ни подход противоопухолевой терапии, ни системная индукция Th1-преобладающего клеточного иммунного ответа с использованием Т-клеток-хелперов, продуцирующих ИФН-γ, ни индукция CTL, и не описана или не достигнута активация врожденной иммунной системы, все аспекты, которые необходимы для противоопухолевой терапии.
Раскрытие изобретения
Целью настоящего изобретения является получение новых противоопухолевых вакцин, посредством которых индуцируют сильный системный клеточный ответ иммунной системы и может быть достигнута эффективная противоопухолевая терапия.
Цель настоящего изобретения неожиданно осуществлена в одном аспекте путем получения микроорганизма в качестве носителя нуклеотидных последовательностей, кодирующих антигены и белковые токсины, включающего следующие компоненты:
первый компонент (I) в виде по меньшей мере одной нуклеотидной последовательности, кодирующей по меньшей мере один полный или неполный антиген из по меньшей мере одного белка дикого типа или мутантного белка,
второй компонент (II) в виде по меньшей мере одной нуклеотидной последовательности, кодирующей по меньшей мере один белковый токсин и/или по меньшей мере одну субъединицу белкового токсина,
третий компонент (III), состоящий по меньшей мере из одного первого подкомпонента (III-а) в виде по меньшей мере одной нуклеотидной последовательности, кодирующей по меньшей мере одну систему транспорта, которая обеспечивает возможность экспрессии продуктов экспрессии компонента (I) и компонента (II) на наружной поверхности микроорганизма и/или обеспечивает возможность секреции продуктов экспрессии компонента (I) и компонента (II) и/или кодирующую по меньшей мере одну сигнальную последовательность, которая обеспечивает возможность секреции продуктов экспрессии компонента (I) и компонента (II) и/или, необязательно,
второго подкомпонента (III-б) в виде по меньшей мере одной нуклеотидной последовательности, кодирующей по меньшей мере один белок для лизиса микроорганизма в цитозоле клеток млекопитающих и для внутриклеточного высвобождения плазмид или векторов экспрессии, которые содержатся в лизированном микроорганизме, и
и четвертый компонент (IV) в виде по меньшей мере одной нуклеотидной последовательности для по меньшей мере одной последовательности активации для экспрессии по меньшей мере одного компонента (I)-(III), причем указанная последовательность активации может быть активирована в микроорганизме и/или является специфической для клетки ткани, специфической для опухолевой клетки, макрофаг-специфической, дендрит-специфической, лимфоцит-специфической, функция-специфической или неспецифической в отношении клетки.
Вышеперечисленные любые из компонентов (I)-(IV) могут присутствовать либо один, либо несколько раз и, если какой-либо из компонентов (I)-(IV) присутствует несколько раз, он может независимо друг от друга быть либо идентичным, либо отличающимся.
В предпочтительном варианте осуществления предусматривают микроорганизм, включающий вышеописанные компоненты (I)-(IV), причем компонент (I) и компонент (II) неидентичны, т.е. компонент (I) не кодирует по меньшей мере одну нуклеотидную последовательность, кодирующую по меньшей мере один белковый токсин и/или по меньшей мере одну субъединицу белкового токсина.
Термин "специфический в отношении клетки ткани" в сочетании с компонентом (IV) на протяжении настоящего изобретения относится к последовательности(ям) активации, которая специфически активирована в клетках ткани-мишени, таких как, например, гормон-зависимые промоторы в тканях простаты.
Термин "специфический в отношении опухолевых клеток" в сочетании с компонентом (IV) на протяжении настоящего изобретения относится к последовательности(ям) активации, которая специфически активирована в опухолевых клетках, таких как промоторные элементы, активированные действием опухолеспецифических онкогенов.
Термин "макрофаг-специфический" в сочетании с компонентом (IV) на протяжении настоящего изобретения относится к последовательности(ям) активации, которая специфически активирована в макрофагах, таких как промоторные элементы, которые кодируются макрофаг-специфическими генами, например, геном, кодирующим F4/80.
Термин "дендрит-специфический" в сочетании с компонентом (IV) на протяжении настоящего изобретения относится к последовательности(ям) активации, которая специфически активирована в дендритных клетках, таких как промоторные элементы, контролирующие экспрессию В7.1. Термины "дендрит-специфический" и "специфический в отношении дендритных клеток" эквиваленты, т.е. они имеют одинаковое значение и оба относятся к дендритным клеткам.
Термин "лимфоцит-специфический" в сочетании с компонентом (IV) на протяжении настоящего изобретения относится к элементам, которые специфически активированы в клетках лимфоцитарной линии, таким как промоторные элементы, регулирующие экспрессию молекул CD3 в Т-клетках, или промоторные элементы, регулирующие экспрессию CD20 в зрелых В-клетках.
Термин "функция-специфический" в сочетании с компонентом (IV) на протяжении настоящего изобретения относится к последовательности(ям) активации, которая специфически активирована в клеточном контексте, например, в опухолевых клетках, которые утратили экспрессию р53, или последовательности(ям) активации, которая активирована в бактериях в зависимости от контекста, например клеточной локализации или давления кислорода.
Термин "неспецифический в отношении клетки" в сочетании с компонентом (IV) на протяжении настоящего изобретения относится к последовательности(ям) активации, которая повсеместно активна, таким как конститутивно активные бактериальные промоторы.
Термин "нуклеотидная последовательность" на протяжении настоящего изобретения относится к dsDNA, ssDNA, dsRNA, ssRNA или гибридам dsDNA/RNA. Предпочтительной является dsDNA.
Термин "антиген" на протяжении настоящего изобретения относится к молекулам, которые взаимодействуют с антителами, т.е. которые способны генерировать антитела. Некоторые антигены сами по себе не вызывают продукцию антител; только те, которые могут индуцировать продукцию антител, называют иммуногенами. Для цели настоящего изобретения предусматривают включение всех известных антигенов. В уровень знаний компетентного специалиста в области техники входит найти необходимую информацию относительно потенциальных антигенов с помощью баз данных и/или экспериментального скрининга без излишних усилий. Примерами антигенов являются в числе прочих клеточных антигенов специфические антигены клеток ткани (например, клеток ткани, из которых происходит опухоль), клеточные белковые антигены, вирусные антигены, вирусные белковые антигены и т.п. Предпочтительными являются белковые антигены. Кроме того, предпочтительны гетерологичные антигены или чужеродные антигены, т.е. антигены, которые не являются эндогенными для соответствующего микроорганизма, представленного в изобретении, или антигены, которые не экспрессируются соответствующим микроорганизмом, представленным в изобретении, в естественных условиях, но интродуцированы с помощью стандартных молекулярных биотехнологических способов.
Термин "полный антиген" на протяжении настоящего изобретения относится к полным молекулам, которые реагируют с антителами согласно вышеприведенному определению. Примерами полных антигенов являются, например, белки полной длины (непроцессированные), которые также предпочтительны.
Термин "неполный антиген" на протяжении настоящего изобретения относится к специфическим частям молекул, которые реагируют с антителами согласно вышеприведенному определению. Частичные антигены могут представлять собой, например, белковые мотивы, такие как аминокислотные петли в белках, домены протеинкиназы, эпитопы и т.п. Предпочтительными являются домены протеинкиназы и эпитопы, последние из которых представляют собой специфические центры антигена, распознаваемого антителом (называемые также антигенными детерминантами).
Термины "дикий тип" и "мутантный" в сочетании с термином "белок" на протяжении настоящего изобретения относятся к белкам, состоящим из их любой "природной" преобладающей последовательности аминокислот (кодируемой соответствующей нуклеотидной последовательностью), и белкам, которые включают одну или более мутаций в их последовательностях аминокислот (кодируемых соответствующей нуклеотидной последовательностью) по сравнению с последовательностью дикого типа соответственно. Предпочтительно, когда белки дикого типа и/или мутантные белки выделены из опухолевых клеток. Что касается неполных антигенов, предпочтительно, кроме того, чтобы последовательность охватывала мутации, т.е. выбирают эпитоп, который предпочтительно включает одну или более мутаций, например, эпитоп B-Raf V600E.
Микроорганизмы в понимании изобретения представляют собой бактерии, грамположительные бактерии, грамотрицательные бактерии и эукариотические клетки, из которых последние включают одноклеточные паразиты, дрожжи, опухолевые клетки и клетки клеточных линий, такие как Sacharomyces cerevisiae, Leishmania spp., аутологичные выделенные у пациента опухолевые клетки и опухолевые клеточные линии. Данные микроорганизмы, как правило, используют в качестве носителей для переноса нуклеотидных последовательностей, являющихся чужеродными (гетерологичными или гетерогенными) для микроорганизма. Предпочтительно, когда используют бактерии, которые аттенюированы по вирулентности, например бактерии, которые несут делецию или инактивированный ген aroA, aro, asd, gal, pur, cya, crp, phoP/Q, omp или представляют собой температурочувствительные мутанты (см. статью Cardenas и Clements, 1992). Кроме того, предпочтительной в качестве микроорганизма, включающего вышеуказанные компоненты (I)-(IV), является грамотрицательная, аттенюированная, факультативно внутриклеточная бактерия в качестве носителя, который способен преодолевать слизистую оболочку кишки (например, Salmonella spp. или Shigella spp.).
В предпочтительном варианте осуществления предусматривают микроорганизм, включающий вышеуказанные компоненты (I)-(IV), причем микроорганизм выбран из группы, состоящей из "бактерии, грамположительной бактерии, грамотрицательной бактерии, эукариотической клетки" и предпочтительно выбран из группы, состоящей из "Escherichia spp., Escherichia coli, Salmonella spp., Salmonella typhi, Salmonella typhimurium, Yersinia spp., Yersinia enterocolitica, Vibrio spp., Vibrio cholerae, Listeria spp., Listeria monocytogenes, Shigella spp., Shigella flexneri", причем предпочтительно, когда вирулентность микроорганизма аттенюирована. Кроме того, предпочтительный Vibrio cholerae исключен из определенных выше микроорганизмов.
Термин "spp." в связи с любым микроорганизмом предусматривает включение для цели настоящего изобретения всех членов данного рода, включая виды, подвиды и др. Термин "Salmonella spp.", например, предусматривает включение всех членов рода Salmonella, таких как Salmonella typhi и Salmonella typhimurium.
В другом предпочтительном варианте осуществления предусматривают микроорганизм, соответствующий вышеприведенным определениям, в котором по меньшей мере один полный или неполный антиген по меньшей мере одного белка дикого типа или мутантного белка, соответствующий компоненту (I), выбран из группы, состоящей из следующих белков дикого типа и их известных мутантов: "рецептор; внеклеточная, трансмембранная или внутриклеточная часть рецептора; молекула адгезии; внеклеточная, трансмембранная или внутриклеточная часть молекулы адгезии; белок сигнальной трансдукции; белок клеточного цикла; фактор транскрипции; белок дифференцировки; эмбриональный белок; вирусный белок; аллерген; белок микробного патогена; белок эукариотического патогена; белок антигена рака яичка; белок опухолевого антигена и/или специфический белок клетки ткани", причем клетка ткани выбрана из группы, состоящей из "щитовидной железы, молочной железы, слюнной железы, лимфатического узла, слизистой оболочки желудка, почки, яичника, простаты, шейки матки, серозной оболочки мочевого пузыря и невуса".
Что касается мутантного белка, то мутация может быть онкогенной и может быть вызывать утрату или расширение его исходных клеточных функций.
Данные антигены осуществляют в клетке контроль клеточного роста и деления клетки и представлены на клеточной мембране нормальных клеток, например, посредством молекулы МНС класса I. В опухолевых клетках данные гены часто сверхэкспрессируются или специфически мутируют. Данные мутации могут иметь ограничения функций супрессоров онкогенов или, как следствие, активации протоонкогенов в онкогены и могут быть включены сами по себе или, обыкновенно, со сверхэкспрессией в рост опухоли. Данные клеточные антигены представлены на мембране опухолевых клеток и, таким образом, представляют антигены на опухолевых клетках, однако, не вызывая иммунную реакцию, воздействующую на опухолевое заболевание пациента. Rapp (US 5156841) уже описал использование онкопротеинов, т.е. продуктов экспрессии онкогенов в качестве иммуногена для противоопухолевых вакцин.
Примерами антигенов и их (онкогенных) мутаций, соответствующих изобретению, являются i) рецепторы, такие как Her-2/neu, андрогенный рецептор, эстрогенный рецептор, лактоферриновый рецептор, midkine-рецептор, рецептор EGF, рецептор ERBB2, ERBB4, рецептор TRAIL, FAS, рецептор TNFα, рецептор TGF-β; ii) белки сигнальной трансдукции, такие как c-Raf (Raf-1), A-Raf, B-Raf, B-Raf V599E, B-Raf V600E, B-Raf KD, домен киназы B-Raf V600E, домен киназы KD B-Raf V600E, B-Raf V600E, домен киназы B-Raf, домен киназы KD B-Raf, Ras, Bcl-2, Bcl-X, Bcl-W, Bfl-1, Brag-1, Mcl-1, A1, Bax, BAD, Bak, Bcl-Xs, Bid, Bik, Hrk, Bcr/abl, Myb, C-Met, IAP1, IAO2, XIAP, ML-IAP LIVIN, сурвивин, APAF-1; iii) белки контроля клеточного цикла, такие как циклин D(1-3), циклин Е, циклин А, циклин В, циклин Н, Cdk-1, Cdk-2, Cdk-4, Cdk-6, Cdk-7, Cdc25C, p16, p15, p21, p27, p18, pRb, p107, p130, E2F(1-5), GAAD45, MDM2, PCNA, ARF, PTEN, APC, BRCA, p53 и гомологи; iv) факторы транскрипции, такие как С-Мус, NFκB, c-Jun, ATF-2, Sp1; v) эмбриональные белки, такие как карциноэмбриональный антиген, α-фетопротеин, MAGE, MAGE-1, MAGE-3, NY-ESO-1, PSCA; vi) антигены дифференцировки, такие как MART, Gp100, тирозиназа, GRP, TCF-4, основной миелин, α-лактальбумин, GFAP, простатаспецифический антиген (PSA), фибриллярный кислый белок, тирозиназа, EGR-1, MUC1; vii) вирусные антигены, такие как следующие вирусы: ВИЧ, HPV, HCV, HPV, EBV, CMV, HSV, вируса гриппа, вируса гриппа типа А, вируса гриппа типа A (H5N1) и (H3N2), вируса гриппа типа В, вируса гриппа типа С; гемагглютининов, гемагглютинина Н1, гемагглютинина Н5, гемагглютинина Н7, гемагглютинина НА1 (предпочтительно из вируса гриппа A (A/Thailand/1(KAN-1)2004(H5N1), гемагглютинина НА12 (предпочтительно из вируса гриппа A (A/Thailand/1(KAN-1)2004(H5N1), гемагглютинина НА12С (предпочтительно из вируса гриппа А (A/Thailand/1(KAN-1)2004(H5N1), нейрамидаза, микробные антигены: р60, LLO, уреаза и т.п., антигены эукариотических патогенов: CSP (малярия), калфлагин (трипаносома), СРВ (Leishmania major) и т.п.
В еще одном предпочтительном варианте осуществления предусматривают микроорганизм, соответствующий вышеприведенным определениям, причем по меньшей мере один полный или неполный антиген по меньшей мере одного белка дикого типа или мутантного белка, соответствующий компоненту (I), выбран из группы, состоящей из следующих белков дикого типа и их известных мутантов: Her-2/neu, андрогенный рецептор, эстрогенный рецептор, midkine-рецептор, рецептор EGF, рецептор ERBB2, ERBB4, рецептор TRAIL, FAS, рецептор TNFα, рецептор TGF-β, лактоферриновый рецептор, основной миелин, α-лактальбумин, GFAP, фибриллярный кислый белок, тирозиназа, EGR-1, MUC1, c-Raf (Raf-1), A-Raf, B-Raf, B-RafV599E, B-RafV600E, B-Raf KD, домен киназы B-RafV600E, B-Raf V600E KD, домен киназы KD B-RafV600E, домен киназы В-Raf, домен киназы KD B-Raf, N-Ras, K-Ras, H-Ras, Bcl-2, Bcl-X, Bcl-W, Bfl-1, Brag-1, Mcl-1, A1, Bax, BAD, Bak, Bcl-Xs, Bid, Bik, Hrk, Bcr/abl, Myb, C-Met, IAP1, IA02, XIAP, ML-IAP LIVIN, сурвивин, APAF-1, циклин D(1-3), циклин Е, циклинА, циклин В, циклин Н, Cdk-1, Cdk-2, Cdk-4, Cdk-6, Cdk-7, Cdc25C, p16, p15, p21, p27, p18, pRb, p107, p130, E2F(1-5), GAAD45, MDM2, PCNA, ARF, PTEN, APC, BRCA, Akt, PI3K, mTOR, p53 и гомологи, C-Myc, NFκB, c-Jun, ATF-2, Sp1, простатаспецифический антиген (PSA), карциноэмбриональный антиген, α-фетопротеин, PAP; PSMA; STEAP; MAGE, MAGE-1, MAGE-3, NY-ESO-1, PSCA, MART, Gp100, тирозиназа, GRP, TCF-4, вирусные антигены вирусов ВИЧ, HPV, HCV, HPV, EBV, CMV, HSV, вируса гриппа, вируса гриппа типа А, вируса гриппа типа A (H5N1) и (H3N2), вируса гриппа типа В, вируса гриппа типа С; гемагглютининов, гемагглютинина Н1, гемагглютинина Н5, гемагглютинина Н7, гемагглютинина НА1 (предпочтительно из вируса гриппа A (A/Thailand/1 (KAN-1)2004(H5N1), гемагглютинина НА12 (предпочтительно из вируса гриппа A (A/Thailand/1(KAN-1)2004(H5N1), гемагглютинина НА12С (предпочтительно из вируса гриппа А (A/Thailand/1(KAN-1)2004(H5N1), нейрамидазы, р60, LLO, уреазы, CSP, калфлагина и/или СРВ".
В еще одном предпочтительном варианте осуществления предусматривают микроорганизм, соответствующим вышеприведенным определениям, причем по меньшей мере один полный или неполный антиген по меньшей мере одного белка дикого типа или мутантного белка, соответствующий компоненту (I), выбран из группы киназ, состоящей из следующих белков дикого типа и их известных мутантов (регистрационные номера приводят в скобках: AAK1 (NM 014911), AATK (NM 004920), ABU (NM 005157), ABL2(NM 005158), ACK1 (NM 005781), ACVR1 (NM 001105), ACVR1 В (NM 020328), ACVR2 (NM 001616), ACVR2B (NM 001106), ACVRL1 (NM 000020), ADCK1 (NM 020421), ADCK2 (NM 052853), ADCK4 (NM 024876), ADCK5 (NM 174922), ADRBK1 (NM 001619), ADRBK2 (NM 005160), AKT1 (NM 005163), AKT2 (NM 001626), AKT3 (NM 005465), ALK (NM 004304), ALK7 (NM 145259), ALS2CR2 (NM 018571), ALS2CR7 (NM 139158), AMHR2 (NM 020547), ANKK1 (NM 178510), ANKRD3 (NM 020639), APEG1 (NM 005876), ARAF (NM 001654), ARK5 (NM 014840), ATM (NM 000051), ATR (NM 001184), AURKA (NM 003600), AURKB (NM 004217), AURKC (NM 003160), AXL (NM 001699), BCKDK (NM 005881), BCR (NM 004327), BIKE(NM 017593), BLK (NM 001715), BMPR1 A(NM 004329), BMPR1 В (NM 001203), BMPR2 (NM 001204), BMX (NM 001721), BRAF (NM 004333), BRD2 (NM 005104), BRD3 (NM 007371), BRD4 (NM 014299), BRDT (NM 001726), BRSK1 (NM 032430), BRSK2 (NM 003957), ВТК (NM 000061), BUB1 (NM 004336), BUB1 В (NM 001211), CABC1 (NM 020247), CAMK1 (NM 003656), CaMK1b (NM 198452), CAMK1 D (NM 020397), CAMK1 G (NM 020439), CAMK2A (NM 015981), CAMK2B (NM 001220), CAMK2D (NM 001221), CAMK2G (NM 001222), CAMK4 (NM 001744), CAMKK1 (NM 032294), CAMKK2 (NM 006549), CASK (NM 003688), CCRK (NM 012119), CDC2 (NM 001786), CDC2L1 (NM 001787), CDC2L5 (NM 003718), CDC42BPA (NM 014826), CDC42BPB (NM 006035), CDC7L1 (NM 003503), CDK10 (NM 003674), CDK11 (NM 015076), CDK2 (NM 001798), CDK3 (NM 001258), CDK4 (NM 000075), CDK5 (NM 004935), CDK6 (NM 001259), CDK7 (NM 001799), CDK8 (NM 001260), CDK9 (NM 001261), CDKL1 (NM 004196), CDKL2 (NM 003948), CDKL3 (NM 016508), CDKL4 (NM 001009565), CDKL5 (NM 003159), CHEK1 (NM 001274), CHUK (NM 001278), CIT (NM 007174), CLK1 (NM 004071), CLK2 (NM 003993), CLK3 (NM 003992), CLK4 (NM 020666), CRK7 (NM 016507), CSF1 R (NM 00521 1), CSK (NM 004383), CSNK1 A1 (NM 001892), CSNK1 D (NM 001893), CSNK1 E (NM 001894), CSNK1 G1 (NM 022048), CSNK1 G2 (NM 001319), CSNK1 G3 (NM 004384), CSNK2A1 (NM 001895), CSNK2A2 (NM 001896), DAPK1 (NM 004938), DAPK2 (NM 014326), DAPK3 (NM 001348), DCAMKL1 (NM 004734), DCAMKL2(NM 152619), DCAMKL3 (XM 047355), DDR1 (NM 013993), DDR2 (NM 006182), DMPK (NM 004409), DMPK2 (NM 017525.1), DYRK1 A (NM 001396), DYRK1 В (NM 006484), DYRK2 (NM 006482). DYRK3 (NM 003582), DYRK4 (NM 003845), EEF2K (NM 013302), EGFR (NM 005228), EIF2AK3 (NM 004836), EIF2AK4 (NM_001013703), EPHA1 (NM 005232), EPHA10 (NM 001004338), EPHA2 (NM 004431), EPHA3 (NM 005233), EPHA4 (NM 004438), EPHA5 (NM 004439), EPHA6 (XM 114973), EPHA7 (NM 004440), EPHA8 (NM 020526), EPHB1 (NM 004441), EPHB2 (NM 017449), EPHB3 (NM 004443), EPHB4 (NM 004444), EPHB6 (NM 004445), ERBB2 (NM 004448), ERBB3 (NM 001982), ERBB4 (NM 005235), ERK8 (NM 139021), ERN1 (NM 001433), ERN2 (NM 033266), FASTK (NM 025096), FER (NM 005246), FES (NM 002005), FGFR1 (NM 000604), FGFR2 (NM 022970), FGFR3 (NM 000142), FGFR4 (NM 022963), FGR (NM 005248), FLJ23074 (NM 025052), FLJ23119 (NM 024652), FLJ23356 (NM 032237), FLT1 (NM 002019), FLT3 (NM 0041 19), FLT4 (NM 002020), FRAP1 (NM 004958), FRK (NM 002031), FYN (NM 002037), GAK (NM 005255), GPRK5 (NM 005308), GPRK6 (NM 002082), GPRK7 (NM 139209), GRK4 (NM 005307), GSG2 (NM 031965), GSK3A (NM 019884), GSK3B (NM 002093), GUCY2C (NM 004963), GUCY2D (NM 000180), GUCY2F (NM 001522), H11 (NM 014365), HAK(NM 052947), HCK (NM 0021 10), HIPK1 (NM 152696), HIPK2 (NM 022740), HIPK3 (NM 005734), HIPK4 (NM 144685), HRI (NM 014413), HUNK(NM 014586), ICK (NM 016513), 1GF1 R (NM 000875), IKBKB (NM 001556), IKBKE (NM 014002), ILK (NM 004517), INSR (NM 000208), INSRR (NM 014215), IRAKI (NM 001569), IRAK2 (NM 001570), IRAK3 (NM 007199), IRAK4 (NM 016123), ITK (NM 005546), JAK1 (NM 002227), JAK2 (NM 004972), JAK3 (NM 000215), KDR (NM 002253), KIS (NM 144624), KIT (NM 000222), KSR (XM 290793), KSR2 (NM 173598), LAK(NM 025144), LATS1 (NM 004690), LATS2 (NM 014572), LCK (NM 005356), LIMK1 (NM 016735), LIMK2 (NM 005569), LMR3 (XM 055866), LMTK2 (NM 014916), LOC149420 (NM 152835), LOC51086 (NM 015978), LRRK2 (XM 058513), LTK (NM 002344), LYN (NM 002350), MAK (NM 005906), MAP2K1 (NM 002755), MAP2K2 (NM 030662), MAP2K3 (NM 002756), MAP2K4 (NM 003010), MAP2K5 (NM 002757), MAP2K6 (NM 002758), MAP2K7 (NM 005043), MAP3K1 (XM 042066), MAP3K10 (NM 002446), MAP3K11 (NM 002419), MAP3K12 (NM 006301), MAP3K13 (NM 004721), MAP3K14 (NM 003954), MAP3K2 (NM 006609), MAP3K3 (NM 002401), MAP3K4 (NM 005922), MAP3K5 (NM 005923), MAP3K6 (NM 004672), MAP3K7 (NM 003188), MAP3K8 (NM 005204), MAP3K9 (NM 033141), MAP4K1 (NM 007181), MAP4K2 (NM 004579), MAP4K3 (NM 003618), MAP4K4 (NM 145686), MAP4K5 (NM 006575), MAPK1 (NM 002745), MAPK10 (NM 002753), MAPK11 (NM 002751), MAPK12 (NM 002969), MAPK13 (NM 002754), MAPK14 (NM 001315), MAPK3 (NM 002746), MAPK4 (NM 002747), MAPK6 (NM 002748), MAPK7 (NM 002749), MAPK8 (NM 002750), MAPK9 (NM 002752), MAPKAPK2 (NM 032960), MAPKAPK3 (NM 004635), MAPKAPK5 (NM 003668), MARK (NM 018650), MARK2 (NM 017490), MARK3 (NM 002376), MARK4 (NM 031417), MAST1 (NM 014975), MAST205 (NM 0151 12), MAST3 (XM 038150), MAST4 (XM 291 141), MASTL (NM 032844), MATK (NM 139355), MELK (NM 014791), MERTK (NM 006343), MET (NM 000245), MGC33182 (NM 145203), MGC42105 (NM 153361), MGC43306 (C9orf96), MGC8407 (NM 024046), MIDORI (NM 020778), MINK (NM 015716), MKNK1 (NM 003684), MKNK2 (NM 017572), MLCK (NM 182493), MLK4 (NM 032435), MLKL (NM 152649), MOS (NM 005372), MST1 R (NM 002447), MST4 (NM 016542), MUSK (NM 005592), MYLK (NM 053025), MYLK2 (NM 0331 18). MY03A (NM 017433), MY03B (NM 138995), NEK1 (NM 012224), NEK10 (NM 152534), NEK11 (NM 024800), NEK2 (NM 002497), NEK3 (NM 002498), NEK4 (NM 003157), NEK5 (MGC75495), NEK6 (NM 014397), NEK7 (NM 133494), NEK8 (NM 178170), NEK9 (NM 0331 16), NLK (NM 016231), NPR1 (NM 000906), NPR2 (NM 003995), NRBP (NM 013392), NRBP2 (NM 178564), NRK (NM 198465), NTRK1 (NM 002529), NTRK2 (NM 006180), NTRK3 (NM 002530), OBSCN (NM 052843), OSR1 (NM 005109), PACE-1 (NM 020423), PAK1 (NM 002576), PAK2 (NM 002577), PAK3 (NM 002578), PAK4 (NM 005884), PAK6 (NM 020168), PAK7 (NM 020341), PASK (NM 015148), PCTK1 (NM 006201), PCTK2 (NM 002595), PCTK3 (NM 212503), PDGFRA (NM 006206), PDGFRB (NM 002609), PDK1 (NM 002610), PDK2 (NM 00261 1), PDK3 (NM 005391), PDK4 (NM 002612), PDPK1 (NM 002613), PFTK1 (NM 012395), PHKG1 (NM 006213), PHKG2 (NM 000294), PIK3R4 (NM 014602), PIM1 (NM 002648), PIM2 (NM 006875), PIM3 (NM 001001852), PINK1 (NM 032409), PKE (NM 173575), PKMYT1 (NM 004203), pknβ (NM 013355), PLK (NM 005030), PLK3 (NM 004073), PRKAA1 (NM 006251), PRKAA2 (NM 006252), PRKACA (NM 002730), PRKACB (NM 002731), PRKACG (NM 002732), PRKCA (NM 002737), PRKCB1 (NM 002738), PRKCD (NM 006254), PRKCE (NM 005400), PRKCG (NM 002739), PRKCH (NM 006255), PRKCI (NM 002740), PRKCL1 (NM 002741), PRKCL2 (NM 006256), PRKCM (NM 002742), PRKCN (NM 005813), PRKCQ (NM 006257), PRKCZ (NM 002744), PRKD2 (NM 016457), PRKDC (NM 006904), PRKG1 (NM 006258), PRKG2 (NM 006259), PRKR (NM 002759), PRKWNK1 (NM 018979), PRKWNK2 (NM 006648), PRKWNK3 (NM 020922), PRKWNK4 (NM 032387), PRKX (NM 005044), PRKY (NM 002760), PRPF4B (NM 003913), PSKH1 (NM 006742), PSKH2 (NM 033126), PTK2 (NM 005607), PTK2B (NM 004103), PTK6 (NM 005975), PTK7 (NM 002821), PTK9 (NM 002822), PTK9L(NM 007284), PXK (NM 017771), QSK (NM 025164), RAD53 (NM 007194), RAF1 (NM 002880), RAGE (NM 014226), RET (NM 020975), RHOK (NM 002929), R10K1 (NM 031480), RIOK2 (NM 018343), RIPK1 (NM 003804), RIPK2 (NM 003821), RIPK3 (NM 006871), RIPK5 (NM 015375), RNASEL(NM 021133), ROCK1 (NM 005406), ROCK2 (NM 004850), ROR1 (NM 005012), ROR2 (NM 004560), ROS1 (NM 002944), RPS6KA1 (NM 002953), RPS6KA2 (NM 021135), RPS6KA3 (NM 004586), RPS6KA4 (NM 003942), RPS6KA5 (NM 004755), RPS6KA6 (NM 014496), RPS6KB1 (NM 003161), RPS6KB2 (NM 003952), RPS6KC1 (NM 012424), RPS6KL1 (NM 031464), RYK (NM 002958), SBK (XM 370948), SCYL1 (NM 020680), SCYL2 (NM 017988), SGK (NM 005627), SgK069 (SU SgK069), SgK085 (XM 373109), SgK110 (SU SgKHO), SGK2 (NM 016276), SgK223 (XM 291277), SgK269 (XM 370878), SgK424 (CGP SgK424), SgK493 (SU_SgK493), SgK494 (NM 144610), SgK495 (NM 032017), SGKL(NM 013257), SK681 (NM 001001671), SLK (NM 014720), SMG1 (NM 015092), SNARK (NM 030952), SNF1 LK (NM 173354), SNF1 LK2 (NM 015191), SNK (NM 006622), SNRK (NM 017719), SRC (NM 005417), SRMS (NM 080823), SRPK1 (NM 003137), SRPK2 (NM 003138), SSTK (NM 032037), STK10 (NM 005990), STK11 (NM 000455), STK16 (NM 003691), STK17A (NM 004760), STK17B (NM 004226), STK18 (NM 014264), STK19 (NM 032454), STK22B (NM 053006), STK22C (NM 052841), STK22D (NM 032028), STK23 (NM 014370), STK24 (NM 003576), STK25 (NM 006374), STK3 (NM 006281), STK31 (NM 031414), STK32B (NM 018401), STK33 (NM 030906), STK35 (NM 080836), STK36 (NM 015690), STK38 (NM 007271), STK38L (NM 015000), STK39 (NM 013233), STK4 (NM 006282), STLK5 (NM 001003787), STYK1 (NM 018423), SUDD (NM 003831), SYK (NM 003177), TAF1 (NM 138923), TAF1 L (NM 153809), TA01 (NM 004783), TAOK1 (NM 020791), TAOK3 (NM 016281), TBCK (NM 0331 15), TBK1 (NM 013254), TEC (NM 003215), TEK (NM 000459), TESK1 (NM 006285), TESK2 (NM 007170), ТЕХ14 (NM 031272), TGFBR1 (NM 004612), TGFBR2 (NM 003242), TIE (NM 005424), TIF1 (NM 003852), TLK1 (NM 012290), TLK2 (NM 006852), TNIK (NM 015028), TNK1 (NM 003985), TOPK (NM 018492), TP53RK (NM 033550), TRAD (NM 007064), TRIB1 (NM 025195), TRIB2 (NM 021643), TRIB3 (NM 021 158), TRIM28 (NM 005762), TRIM33 (NM 015906), TRIO (NM 0071 18), TRPM6 (NM 017662), TRPM7 (NM 017672), TRRAP (NM 003496), TSSK4 (NM 174944), TTBK1 (NM 032538), TTBK2 (NM 173500), TTK (NM 003318), TTN (NM 003319), TXK (NM 003328), TYK2 (NM 003331), TYR03 (NM 006293), ULK1 (NM 003565), ULK2 (NM 014683), ULK3 (NM 015518), ULK4 (NM 017886), VRK1 (NM 003384), VRK2 (NM 006296), VRK3 (NM 016440), WEE1 (NM 003390), Wee1 B (NM 173677), YANK1 (NM 145001), YES1(NM 005433), ZAK (NM 016653), и/или ZAP70 (NM 001079)".
Термин "аллерген" в контексте настоящего изобретения относится к полным или неполным антигенам, как определено в данном контексте, которые вызывают гиперчувствительность и/или аллергические реакции. Примерами являются Der p 5 (клещи), Bet v1 (пыльца березы), Phl p 1 (пыльца злаков), Asp f I/a (Aspergillus), PLA 2 (пчела), Hev b (латекс) (см. статью Schmid-Grendelmeier и Crameri, 2001).
Антигены микробных и эукариотических патогенов и антигены рака яичка приведены в перечне выше.
На протяжении настоящего изобретения белковые токсины и/или их субъединицы, соответствующие компоненту (II), предпочтительно представляют собой бактериальные белковые токсины, более предпочтительно - экзотоксины. Примерами бактериальных экзотоксинов являются токсины типа I (суперантигены), токсины типа II (порообразующие токсины) и (iii) токсины типа III (A-B токсины). В предпочтительном варианте осуществления предусматривают микроорганизм, соответствующий вышеприведенным описаниям, причем компонент (II) выбран из группы, состоящей из "бактериального токсина, энтеротоксина, экзотоксина, токсина типа I, токсина типа II, токсина типа III, токсина типа IV, токсина типа V, токсина RTX, токсина АВ, токсина A-B, токсина А/В, токсина А+В, токсина А-5В и/или токсина АВ5.
В еще одном предпочтительном варианте осуществления предусматривают микроорганизм, соответствующий вышеприведенным описаниям, причем компонент (II) выбран из группы, состоящей из "токсина аденилатциклазы, токсина сибирской язвы, токсина сибирской язвы (EF), токсина сибирской язвы (LF), ботулинического токсина, холерного токсина (СТ, Ctx), субъединицы холерного токсина В (СТВ, CtxB), дифтерийного токсина (DT, Dtx), токсина Е. coli LT, термолабильного энтеротоксина Е. coli (LT), субъединицы В термолабильного энтеротоксина Е. coli (LTB), токсина Е. coli ST, термостабильного энтеротоксина Е. coli (ST), эритрогенного токсина, токсина эксфолиатина, экзотоксина А, энтеротоксина Perfringens, токсина коклюша (РТ, Ptx), шига-токсина (ST, Stx), субъединицы В шига-токсина (STB, StxB), шига-подобного токсина, энтеротоксинов Staphylococcus, столбнячного токсина (ТТ), токсина синдрома токсического шока (TSST-1), веро-токсина (VT), токсина А (ТА) и токсина В (ТВ) Clostridium difficile, летального токсина (LT) и геморрагического токсина (НТ) Clostridium sordellii, α-токсина (AT) Clostridium novyi".
Однако если холерный токсин или его субъединицу CtxB используют в качестве токсинов, соответствующих компоненту (II), представленному в изобретении, предпочтительно не использовать Vibrio cholerae в качестве бактериального носителя (микроорганизма).
В предпочтительном варианте осуществления предусматривают микроорганизм, соответствующий вышеприведеным определениям, причем компонент (I) и компонент (II) связаны друг с другом, чтобы обеспечить возможность экспрессии и/или секреции слитого белка, кодируемого обоими компонентами. Более предпочтительно, когда данный слитый белок выбран из группы, состоящей из "CtxB-PSA, CtxB-B-Raf V600E KD (киназа dead), CtxB-домен киназы B-Raf V600E, CtxB-домен киназы KD B-Raf V600E (киназа dead), CtxB-B-Raf, CtxB-B-Raf KD (киназа dead), CtxB-домен киназы KD B-Raf (киназа dead), CtxB-HA1 (субъединицы 1 гемагглютинина вируса гриппа), CtxB-HA12C".
Секреция представляет собой процесс отделения, превращения и высвобождения химических агентов из клетки, либо секретируемой химической субстанции, либо количества субстанции. Секреция не уникальна только для эукариот; она присутствует также у бактерий и архей. Транспортеры типа АТФ-связывающей кассеты распространены во всех трех типах живых организмов. Система Sec также является другой консервативной системой, которая гомологична транслокону в эукариотическом эндоплазматическом ретикулюме, состоящему из комплекса транслокона Sec 61 у дрожжей и комплексу Sec Y-E-G у бактерий. Грамотрицательные бактерии имеют две мембраны, делая, таким образом, секрецию более сложной. Вследствие этого у грамотрицательных бактерий имеется по меньшей мере пять специализированных систем секреции:
(1) Система секреции типа I: Такая же, как вышеупомянутые транспортеры АТФ-связывающей кассеты.
(2) Система секреции типа II: от системы Sec зависит, перейдет ли белок внутреннюю мембрану, и от другой специальной системы, перейдет ли он наружную мембрану. Бактериальные пили используют модификации системы sec, но отличаются от системы типа I.
(3) Система секреции типа III (T3SS): она гомологична бактериальному флагеллярному базальному телу. Она похожа на молекулярный шприц, через который бактерия (например, Shigella или Yersinia) может инъецировать белки в эукариотические клетки. Низкая концентрация Са2+ в цитозоле отрывает вход, который регулирует T3SS. Система Hrp у патогенов растений инъецирует в растения шпильки через подобные механизмы.
(4) Система секреции типа IV: Она гомологична механизму конъюгации бактерий (и архейных жгутиковых). Она способна транспортировать как ДНК, так и белки. Она обнаружена у Agrobacterium tumefaciens, которая использует данную систему для интродукции Ti-плазмиды и белков хозяину, у которого развивается корончатый галл (опухоль). Helicobacter pylori использует систему секреции IV, чтобы ввести Cag А в эпителиальные клетки желудка. Bordetella pertussis, возбудитель коклюша, секретирует токсин коклюша частично посредством системы типа IV.
(5) Система секреции типа V, также называемая системой аутотранспортеров: Она использует систему sec для перехода через внутреннюю мембрану. Белки, которые используют данный путь, обладают способностью формировать β-бочку на С-конце и встраиваться в наружную мембрану для транспорта остальной части пептида из клетки. Наконец, β-бочка может отщепляться и оставаться в наружной мембране. Некоторые считают, что из данных остатков аутотранспортеров образуются порины, которые подобны β-бочкам.
Бактерии, а также митохондрии и хлоропласты также используют много других специальных транспортных систем, таких как путь двойной транслокации аргинина (Tat), который, в противоположность Sec-зависимому экспорту, транспортирует полностью уложенные белки через мембрану. Название данной системы происходит из необходимости присутствия двух следующих друг за другом аргининов в сигнальной последовательности, требующихся для направленности на данную систему. Секреция у грамотрицательных бактерий включает преодоление внутренней и наружной мембран с помощью подходящей системы секреции, подобной, например, системе секреции Hly типа I или типа III или аутоторанспортеру AIDA. У грамположительных бактерий система секреции должна преодолевать внутреннюю мембрану и клеточную стенку, что у большинства штаммов может достигаться слиянием с подходящим сигналом секреции.
Подкомпонент (III-а) представляет собой по меньшей мере одну нуклеотидную последовательность, кодирующую по меньшей мере одну транспортную систему, которая обеспечивает возможность экспрессии продуктов экспрессии компонента (I) и компонента (II) на наружной поверхности микроорганизма и/или обеспечивает возможность секреции продуктов экспрессии компонента (I) и компонента (II). Соответствующий компонент необязательно может либо секретироваться, либо экспрессироваться на мембране микроорганизма, т.е. быть связанным с мембраной. Данные транспортные системы представляют собой, например i) систему секреции типа I, систему секреции типа II, систему секреции типа III, систему секреции типа IV, систему секреции типа V, ii) гемолизиновую транпортную систему (сигнал) Е. coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем hly-специфического промотора); следует использовать следующие транспортные сигналы: для секреции - С-концевой транспортный сигнал HlyA в присутствии белков HlyB и HlyD; для связанной с мембраной экспрессии - С-концевой транспортный сигнал HlyA в присутствии белка HlyB, iii) гемолизиновую транспортную систему (сигнал) Е. coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD, под контролем не-hly-специфического бактериального промотора), iv) транспортный сигнал для белка S-слоя (RsaA) Caulobacter crescentus; следует использовать следующие транспортные сигналы: для секреции и связанной с мембраной экспрессии - С-концевой транспортный сигнал RsaA, v) транспортный сигнал для белка TolC Escherichia coli; следует использовать следующие транспортные сигналы: связанной с мембраной экспрессии - N-концевой транспортный сигнал TolC (полный мембранный белок TolC Е. coli представляет собой многофункциональный порообразующий белок наружной мембраны Е. coli, который служит в дополнение к таким функциям, как прием колицина Е1 (см. статью Morona et al., 1983) и секреция колицина V (см. статью Fath et al., 1991), так же как рецептор для фага U3 (см. статью Austin et al., 1990); данный белок обнаружен не только в Е. coli, но также во множестве грамотрицательных бактерий (см. статью Wiener, 2000); локализация в наружной мембране и широкое распространение делают TolC идеальным кандидатом для представления гетерологичных антигенов, чтобы, например, вызвать иммунную реакцию.
Грамположительные бактерии не заключены в наружную мембрану. Секреция в данных случаях происходит проще и обычно не требует специального механизма секреции для транспорта через клеточную мембрану и клеточную стенку. В случае грамположительных бактерий сигналы секреции слиты по N-концу с гетерологичным белком и, как правило, необходимы и достаточны для секреции. Белки, для которых описаны данные сигнальные последовательности, включают в основном все секретируемые бактериальные белки. Примерами для листерий являются сигналы секреции, выделенные из листериолизина, р60 или ActA. В общем аналогичный подход применяют к эукариотическим клеткам, где сигнальная последовательность (например, универсальный сигнал секреции Vtgss), направляющая белок на эндоплазматический ретикулюм в отсутствие сигнала удерживания, необходима и достаточна для секреции.
Необязательный второй подкомпонент (III-б) представляет собой по меньшей мере одну нуклеотидную последовательность, кодирующую по меньшей мере один белок для лизиса микроорганизма в цитозоле клеток млекопитающих и для внутриклеточного высвобождения плазмид или векторов экспрессии, которые содержатся в лизированном микроорганизме. Данные литические белки (эндолизины) представляют собой, например, белки лизиса, специфические для Listeria, такие как PLY551 (см. статью Loessner et al., 1995) и/или специфический для Listeria холин под контролем промотора листерий. Предпочтительный вариант осуществления данного изобретения представляет собой комбинацию различных подкомпонентов (III-b), например комбинацию белка лизиса и холина.
Оба подкомпонента (III-а) и/или (III-б) могут быть независимо друг от друга конститутивно активными.
В предпочтительном варианте осуществления предусматривают микроорганизм, соответствующий вышеприведенным определениям, причем подкомпонент (III-а) выбран из группы, состоящей из "системы секреции типа I, системы секреции типа II, системы секреции типа III, системы секреции типа IV, системы секреции типа V, гемолизиновой системы транспорта (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем hly-специфического промотора), гемолизиновой системы транспорта (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем не-hly-специфического промотора), транспортного сигнала для белка S-слоя (Rsa A) Caulobacter crescentus, транспортного сигнала для белка TolC Escherichia coli, сигнала секреции Vtgss и/или сигналов секреции, выделенных из листериолизина, р60 и/или ActA", и причем подкомпонент (III-б) выбран из группы, состоящей из "эндолизинов, литического белка грамположительных бактерий, литического белка Listeria monocytogenes, PLY551 Listeria monocytogenes и/или холина Listeria monocytogenes".
В следующем предпочтительном варианте осуществления такой же подкомпонент (III-а) обеспечивает возможность экспрессии продуктов экспрессии компонента (I) и компонента (II) на наружной поверхности микроорганизма и/или обеспечивает возможность секреции продуктов экспрессии компонента (I) и компонента (II). Именно в данном предпочтительном варианте осуществления подкомпонент (III-а) представляет собой по меньшей мере одну нуклеотидную последовательность, кодирующую только одну транспортную систему, которая обеспечивает возможность сопутствующей экспрессии продуктов экспрессии компонента (I) и компонента (II) на наружной поверхности микроорганизма и/или обеспечивает возможность сопутствующей секреции продуктов экспрессии компонента (I) и компонента (II), причем данный предпочтительный подкомпонент (III-а) представляет собой по меньшей мере одну нуклеотидную последовательность, кодирующую гемолизиновую транспортную систему (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем hly-специфического промотора) или гемолизиновую транспортную систему (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем не-hly-специфического бактериального промотора).
В еще одном предпочтительном варианте осуществления предусматривают микроорганизм, соответствующий вышеприведенным определениям, причем в соответствии с подкомпонентом (III-а) продукты экспрессии компонентов (I) и компонента (II) секретируются. Более предпочтительно, когда компонент (I) и компонент (II) связаны друг с другом, экспрессируются вместе в виде слитого белка, кодируемого обоими компонентами. Наиболее предпочтительно, когда данный слитый белок выбран из группы, состоящей из "CtxB-PSA, CtxB-B-Raf V600E KD (киназа dead), CtxB-домен киназы B-Raf V600E, CtxB-B-домен киназы KD Raf V600E (киназа dead), CtxB-B-Raf, CtxB-B-Raf KD (киназа dead), CtxB-B-домен киназы KD Raf (киназа dead), CtxB-HA1 (субъединица 1 гемагглютинина вируса гриппа), CtxB-HA12C".
На протяжении настоящего изобретения, термин "секреция" относится к секреции белкового антигена, белкового токсина и/или слитого белка токсин-антиген через обе мембраны грамотрицательной бактерии или через внутреннюю мембрану и клеточную стенку грамположительной бактерии в прилегающую среду посредством надлежащей системы секреции, такой как проиллюстрированные выше.
Поскольку известно, что при использовании системы секреции, такой как гемолизиновая система секреции типа I из Е. coli, продукт секреции, как правило, обнаруживают во всех клеточных фракциях: цитоплазматически, ассоциированным с мембраной и локализованным в аутомембранных пузырьках и полностью секретируемым в прилегающую среду (см. статью Balsalobre et al., 2006), на протяжении настоящего изобретения не требуется, чтобы секреция была полной. Однако желательна и предпочтительна полная или почти полная секреция, т.е. большая часть полностью секретируемого продукта секреции.
Компонент (IV) представляет собой по меньшей мере одну нуклеотидную последовательность для по меньшей мере одной последовательности активации для экспрессии одного или более компонентов (I) - (III), причем указанная последовательность активации может быть активирована в микроорганизме и/или является специфической для клетки ткани, специфической для опухолевой клетки, макрофаг-специфической, дендрит-специфической, лимфоцит-специфической, функция-специфической или неспецифической в отношении клетки.
Если экспрессия является мембрана-связанной на наружной поверхности микроорганизма, то предпочтительно, когда последовательность активации выбирают так, чтобы она была способна активироваться в микроорганизме. Данные последовательности активации представляют собой, например: i) конститутивно активные промоторные участки, такие как промоторный участок с "сайтом связывания рибосом" (RBS) гена β-лактамазы Е. coli или гена tetA (см. статью Busby и Ebright, 1994), эндогенный промотор локуса hly Е. coli; ii) промоторы, которые способны индуцироваться, предпочтительно промоторы, которые становятся активными после входа в клетку. К ним принадлежит промотор actA L monocytogenes (см. статью Dietrich et al., 1998) или промотор pagC S. typhimurium (см. статью Bumann, 2001).
Если плазмиды высвобождаются из микроорганизма после его лизиса в цитозоле клетки млекопитающего, последовательность активации не является специфической в отношении клетки, но специфической в отношении клетки ткани, специфической в отношении клеточного цикла или функция-специфической. Предпочтительно, когда выбирают такие последовательности активации, которые особенно активируются в макрофагах, дендритных клетки и лимфоцитах.
В следующем предпочтительном вариант осуществления предусматривают микроорганизм, соответствующий вышеприведенным определениям, в котором компонент (I) выбран из группы, состоящей из B-Raf V600E, домена киназы B-Raf V600E, B-Raf V600E KD (киназа dead), домена киназы KD B-Raf V600E (киназа dead), B-Raf KD (киназа dead), домена киназы B-Raf, домена киназы KD B-Raf (киназа dead), простата-специфического антигена (PSA), гемагглютинина НА1 (предпочтительно из вируса гриппа А (A/Thailand/1(KAN-1)2004(H5N1), гемагглютинина НА12 (предпочтительно из вируса гриппа А (A/Thailand/1(KAN-1)2004(Н5М1), гемагглютинина НА12С (предпочтительно из вируса гриппа A (A/Thailand/1 (KAN-1)2004(H5N1)"; компонент (II) выбран из группы, состоящей из "субъединицы В холерного токсина (СТВ, CtxB), субъединицы В термолабильного энтеротоксина Е. coli (LTB), столбнячного токсина (ТТ)"; подкомпонент (III-а) выбран из группы, состоящей из "сигнала гемолизинового транспорта HlyA Escherichia coli вместе с компонентами системы секреции Hly (нуклеотидными последовательностями, включающими HlyA, HlyB и HlyD под контролем hly-специфического промотора)"; компонент (IV) выбран из группы, состоящей из "эндогенного промотора локуса hly Е. coli", причем компонент (I) и компонент (II) связаны друг с другом, чтобы обеспечить возможность экспрессии слитого белка, кодируемого обоими компонентами, и причем слитый белок секретируется.
Проиллюстрированные выше микроорганизмы, соответствующие компонентам (I)-(IV), а также предпочтительные варианты осуществления далее называют микроорганизмами, соответствующими изобретению.
Микроорганизмы, соответствующие изобретению, преимущественно подходят для использования в противоопухолевой терапии в качестве живых вакцин в ходе получения направленности на опухоль. Именно посредством микроорганизмов, соответствующих изобретению, которые действуют как носители генетической информации, гетерологичные антигены совместно с белковыми токсинами транспортируются в область опухоли, экспрессируются как слитые белки и секретируются in situ.
Микроорганизмы, соответствующие изобретению, неожиданно и преимущественно характеризуются посредством эффективной и повышенной экспрессии и секреции транспортируемых, кодируемых и экспрессируемых слитых белков токсин-антиген, т.е. никакие цитоплазматические и/или периплазматические агрегаты не присутствуют.
Более того, путем введения микроорганизмов, соответствующих изобретению, неожиданно вызывают сильный системный клеточный ответ иммунной системы по сравнению с перорально вводимыми белковыми вакцинами, которые, напротив, индуцируют системную толерантность иммунной системы.
Наиболее заметно, что, кроме индукции системного клеточного ответа иммунной системы Th1-типа, микроорганизмы, соответствующие изобретению, вызывают активацию цитотоксических Т-клеток (CTL) CD8+ с рестрикцией МНС класса I и сильно повышают данный иммунный ответ CTL.
Кроме того, в дополнение к индукции и/или повышению уровня сильных клеточных системных Th1- и CTL-ответов иммунной системы врожденная иммунная система, например NK-клетки, NKT-клетки и/или γ-δ T-клетки, также неожиданно активируются микроорганизмами, соответствующими изобретению, синергическим образом.
При использовании холерного токсина в качестве компонента токсина, микроорганизмы, соответствующие изобретению, обладают преимуществом в том, что у человека отсутствует обычно существующий конституциональный иммунитет против токсина (в отличие от столбнячного токсина, например, вследствие противостолбнячной вакцинации в детстве). Вследствие этого предпочтительно использование холерного токсина и/или его субъединиц, в частности, CtxB.
Микроорганизмы, соответствующие изобретению, особенно подходят для перорального применения в направленной противоопухолевой (иммуно)терапии на основе живой вакцины. Вследствие этого можно улучшить соблюдение пациентами режима и схемы лечения.
Однако микроорганизмы, соответствующие изобретению, не ограничены применением только в противоопухолевой терапии. В принципе микроорганизмы, соответствующие изобретению, подходят также для лечения и/или профилактики всех данных заболеваний, для которых требуется терапия, в которой обязательна индукция системного клеточного Th1-ответа иммунной системы. Примеры данных инфекционных заболеваний включают, но без ограничения перечисленным, ВИЧ, грипп, HCV и другие вирусные болезни, Mycobacterium tuberculosis, Listeria monocytogenes и другие бактериальные заболевания.
Микроорганизмы, соответствующие изобретению, в полной мере подходят для лечения заболеваний, подобных гриппу, при которых необходима оптимальная защита посредством комбинации ответа иммунной системы слизистых и системного клеточного иммунного ответа. Кроме того, их можно было бы использовать для специфической индукции Th1-подобного иммунитета при аллергических заболеваниях, например аллергическом рините. В данном случае антиген представлял бы собой аллерген, который сливают с компонентом белкового токсина, и принцип действия соответствующей вакцины состоял бы в сдвиге иммунного ответа с Th2-преобладающего иммунного ответа в отношении аллергена при аллергических реакциях (заболеваниях) к Th1-иммунному ответу.
Предпочтительным способом введения является пероральное введение. В предпочтительном варианте осуществления штамм Salmonella, соответствующий данному изобретению, ферментируют в подходящей среде, собирают и промывают центрифугированием и затем получают и стабилизируют с использованием соответствующих субстанций и лиофилизируют. Лиофилизированной субстанцией наполняют устойчивые в желудке капсулы, содержащие количества живых клеток предпочтительно от 109 до 1010 бактерий. Капсулы перорально принимают с жидкостью.
Альтернативно лиофилизированные бактерии, как описано выше, распространяют вместе с пакетиками, содержащими буфер, который может нейтрализовать желудочную кислоту (фармацевтический набор). В предпочтительном варианте осуществления данный буфер представляет собой карбонатный буфер. Непосредственно перед применением буфер готовят с водой и принимают, сразу после этого принимают лиофилизированные бактерии, смешанные с водой.
Еще одной альтернативой является использование замороженных бактерий. В данном случае после промывания бактерии стабилизируют с помощью стабилизатора, предпочтительно сахарозы или глицерина, и затем замораживают и хранят при -80°C, предпочтительно в концентрациях от 109 до 1010 бактерий/дозу. Предпочтительно, когда данный препарат используют в фармацевтическом наборе, как описано выше, в сочетании с карбонатным буфером.
В предпочтительном варианте осуществления предусматривают фармацевтическую композицию, включающую по меньшей мере один микроорганизм, соответствующий изобретению, предпочтительно по меньшей мере один лиофилизированный микроорганизм, соответствующий изобретению, и фармацевтически приемлемый носитель, предпочтительно капсулы.
Компоненты (I)-(IV), соответствующие настоящему изобретению, вводят в микроорганизмы, соответствующие изобретению, способами, хорошо известными специалисту в области техники. Если микроорганизмы представлены бактериями, компоненты вводят в плазмиды или векторы экспрессии, и плазмиды или векторы экспрессии переносят в бактерии. Способы молекулярно-биологического клонирования и трансформации, пригодные для получения плазмид, векторов экспрессии и микроорганизмов, соответствующих изобретению, хорошо известны специалисту в области техники и представляют собой рутинную экспериментальную работу.
Другим объектом изобретения является введение лекарственного препарата, включающего микроорганизмы, соответствующие изобретению. Введение проводят местно или системно, например орально, перорально, ректально, в эпидермис, подкожно, внутримышечно, в полость тела, в орган, в опухоль или в кровоток.
Данные лекарственные препараты представляют собой, например, суспензии микроорганизмов, соответствующих изобретению, в растворах, известных фармацевтам, подходящих для инъекций.
В частном варианте осуществления, предметом настоящего изобретения является пероральное или ректальное введение лекарственного препарата, соответствующего изобретению, для лечения и/или профилактики заболеваний. Введение проводят однократно или несколько раз. При каждом введении вводят приблизительно 10-1011 микроорганизмов, соответствующих изобретению. Если введение данного количества микроорганизмов, соответствующих изобретению, не вызывает достаточной иммунной реакции, количество, предназначенное для инъекции, увеличивают.
После введения микроорганизмов, соответствующих изобретению, толерантность в отношении клетки, презентирующей компонент (I), например, опухолевой клетке или клетке ткани, из которой происходит опухоль, нарушается и запускается сильный системный иммунный ответ, направленный на опухоль и/или клетки ее ткани. В зависимости от выбора компонента (I) данная клеточная иммунная реакция направлена либо исключительно в отношении опухоли, либо, кроме того, в отношении опухолевых клеток, включающих клетки ткани, из которой происходят опухолевые клетки.
В другом аспекте задачей настоящего изобретения является получение лекарственного препарата, включающего по меньшей мере один микроорганизм, соответствующий изобретению, включающий проиллюстрированные выше генетические компоненты (I) - (IV) или по меньшей мере одну фармацевтическую композицию, как определено в данном контексте.
В предпочтительном варианте осуществления микроорганизмы, соответствующие изобретению, могут быть использованы для получения лекарственного препарата для лечения и/или профилактики физиологических и/или патофизиологических состояний, выбранных из группы, состоящей из "неконтролируемого деления клеток, злокачественных опухолей, доброкачественных опухолей, солидных опухолей, сарком, карцином, гиперпролиферативных нарушений, карциноидов, сарком Эвинга, сарком Капоши, опухолей головного мозга, опухолей, происходящих из головного мозга, и/или нервной системы, и/или мозговых оболочек, глиом, нейробластом, рака желудка, рака почки, карциномы почечных клеток, рака простаты, карцином простаты, опухолей соединительных тканей, сарком мягких тканей, опухолей поджелудочной железы, опухолей печени, опухолей головы, опухолей шеи, рака пищевода, рака щитовидной железы, остеосарком, ретинобластом, тимомы, рака яичка, рака легкого, бронхиальных карцином, рака молочной железы, карцином молочной железы, рака кишки, колоректальных опухолей, карцином толстой кишки, карцином прямой кишки, гинекологических опухолей, опухолей яичника/овариальных опухолей, рака матки, рака шейки матки, карцином шейки матки, рака тела матки, карцином тела, эндометриальных карцином, рака мочевого пузыря, рака полости, рака кожи, базалиом, спиналиом, меланом, внутриглазных меланом, лейкоза, хронического лейкоза, острого лейкоза, лимфом, инфекции, вирусной или бактериальной инфекции, гриппа, хронического воспаления, отторжения органа и/или аутоиммунных заболеваний".
Бактериальные инфекции включают, но без ограничения перечисленным, сибирскую язву, бактериальный менингит, ботулизм, бруцеллез, камплобактериоз, болезнь кошачьих царапин, холеру, дифтерию, эпидемический тиф, импетиго, легиогеллез, лепру (болезнь Гансена), лептоспироз, листериоз, лаймскую болезнь, мелиоидоз, инфекцию MRSA (метициллин-резистентных стафилококков), нокардиоз, коклюш (судорожный кашель), чуму, пневмококковую пневмонию, пситтакоз, Ку-лихорадку, пятнистую лихорадку Скалистых гор (RMSF), сальмонеллез, скарлатину, шигеллез, сифилис, столбняк, трахому, туберкулез, туляремию, брюшной тиф, тиф, инфекции мочевых путей, вызываемые бактериями сердечные заболевания.
Вирусные инфекции включают, но без ограничения перечисленным, СПИД, связанный со СПИДом комплекс (ARC), ветрянку (ветряную оспу), простуду, цитомегаловирусную инфекцию, колорадскую клещевую лихорадку, лихорадку Денге, геморрагическую лихорадку Эбола, ящур, гепатит, простой герпес, опоясывающий лишай, HPV, инфлюэнцу (грипп), лихорадку Ласса, корь, марбургскую геморрагическую лихорадку, инфекционный мононуклеоз, паротит, полиомиелит, прогрессирующую многоочаговую лейкоэнцефалопатию, бешенство, краснуху, САРС (атипичную пневмонию), оспу (натуральную оспу), вирусный энцефалит, вирусный гастроэнтерит, вирусный менингит, вирусную пневмонию, лихорадку Западного Нила, желтую лихорадку.
Хронические воспаления или хронические воспалительные заболевания включают, но без ограничения перечисленным, хронический холецистит, бронхектаз, ревматоидный артрит, тиреоидит Хашимото, воспалительную болезнь кишки (язвенный колит и болезнь Крона), силикоз и другой пневмокониоз.
Аутоиммунные заболевания включают, но без ограничения перечисленным, системные синдромы, такие как SLE, синдром Шегрена, склеродерму, ревматоидный артрит и полимиозит, а также местные синдромы, такие как IDDM (инсулинозависимый сахарный диабет), тиреоидит Хашимото, болезнь Аддисона, обыкновенная пузырчатка, псориаз, атопический дерматит, атопический синдром, астма, аутоиммунная гемолитическая анемия, рассеянный склероз.
Соответствующие лекарственные препараты, включающие по меньшей мере один микроорганизм, как описано в данном контексте, или по меньшей мере одну фармацевтическую композицию, как определено в данном контексте согласно всем описанным в данном контексте вариантам осуществления, предназначенным для применения для лечения и/или профилактики физиологических и/или патофизиологических состояний, как описано и определено в данном контексте, также включены в настоящее изобретение.
В другом аспекте задачей настоящего изобретения является получение плазмиды или вектора экспрессии, включающих компоненты (I)-(IV), как проиллюстрировано в данном контексте. Предпочтительным является микроорганизм, соответствующий изобретению, который несет по меньшей мере одну плазмиду или вектор экспрессии, включающие компоненты (I)-(IV), как проиллюстрировано в данном контексте.
В другом аспекте задачей настоящего изобретения является осуществление способа получения микроорганизма, соответствующего изобретению, причем получают плазмиду или вектор экспрессии, как проиллюстрировано в данном контексте, и трансформируют микроорганизм данной плазмидой или вектором экспрессии.
В другом аспекте задачей настоящего изобретения является способ получения фармацевтического набора, включающего по меньшей мере один микроорганизм, соответствующий изобретению, или фармацевтическую композицию, как описано выше, либо лекарственный препарат, как описано выше, и фармакологически приемлемый буфер, предпочтительно карбонатный буфер.
Краткое описание чертежей
На фигуре 1 представлена нуклеотидная последовательность (5'>3') сигнала секреции гемолизина A (hlyA) E. coli. Сайт Nsil подчеркнут.
На фигуре 2 представлена нуклеотидная последовательность (5'>3') гемолизина В (NyB) E. coli транспортной системы типа 1 гемолизина E. coli.
На фигуре 3 представлена нуклеотидная последовательность (5'>3') гемолизина D (MyD) E. coli транспортной системы типа 1 гемолизина E. coli.
На фигуре 4 представлена нуклеотидная последовательность (5'-3') человеческого простата-специфического антигена (PSA) без сигнального пептида, регистрационный номер М26663 (мРНК простата-специфического антигена Homo, полные cds), участок 100-807 (соответствующий пептид: АА 26-261).
На фигуре 5 представлена нуклеотидная последовательность (5'>3') субъединицы В холерного токсина (CtxB) без сигнального пептида, охватывающий участок 204-494, регистрационный номер K01170 (гены toxA и toxB Vibrio cholerae для субъединиц А2 холерного энтеротоксина А2 (γ).
На фигуре 6 представлена нуклеотидная последовательность (5'-3') домена киназы B-Raf (B-Raf KD) человеческого белка B-raf (BRAF), регистрационный номер М95712 (мРНК белка B-raf Homo sapiens (BRAF), полные cds), участок 1403-2359 без участка 1448-1477 (соответствующий пептид: АА 448-766 без АА 463-472), включающий мутацию V600E.
На фигуре 7 представлена нуклеотидная последовательность (5'>3') эпитопа человеческого B-Raf V600E с рестрикцией HLA B27.
На фигуре 8 представлена нуклеотидная последовательность (5'>3') генетически слитой конструкции CtxB - PSA - HlyA.
На фигуре 9 представлена нуклеотидная последовательность (5'>3') генетически слитой конструкции Ctx - B-Raf V600E - HlyA.
На фигуре 10 представлена нуклеотидная последовательность (5'>3') генетически слитой конструкции CtxB - домен киназы KD B-Raf V600E - HlyA.
На фигуре 11 представлена функциональная экспрессия и секреция слитых белков PSA-CtxB в различных видах бактерий, рекомбинантных по pMKhly-PSA-CtxB.
На фигуре 12 представлены спленоциты, секретирующие ИФН-γ, в качестве меры ответов Т-клеток CD8+ и врожденной иммунной системы (NK-клеток) у мышей, иммунизированных различными живыми вакцинными носителями.
На фигуре 13 представлено уменьшение объема опухоли в ответ на иммунизацию различными живыми вакцинными носителями.
На фигуре 14 представлены спленоциты, секретирующие ИФН-γ, в качестве меры ответов Т-клеток CD8+ и врожденной иммунной системы (NK-клеток) у мышей, иммунизированных различными живыми вакцинами, несущими различные иммунологические адъюванты со слитыми конструкциями CtxB-OVA.
На фигуре 15 представлена эффективная экспрессия и функциональная секреция слитых белков гемагглютинин Н1 куриного гриппа - CtxB в ветеринарном вакцинном штамме, включающем полную последовательность гемагглютинина H5N1 (НА1+НА2, представлено, НА12) без трансмембранного участка (НА12с). Живая аттенюированная Salmonella enterica, серовар Ty-phimurium vacT (gyrA D87G) и живая аттенюированная Salmonella enterica серовар Enteritidis vacE (штамм Sm24/Rif12/Ssq), обе фирмы Lohmann Animal Health GMBH & CO KG.
На фигуре 16 представлена нуклеотидная последовательность, кодирующая слитый белок CtxB-HA12c-HlyA.
На фигуре 17 представлена экспрессия и секреция слитого белка с использованием вектора pMBKDC, электропорированного в штамм Salmonella typhi Ty21a и другие бактериальные штаммы:
A) ряды А1-А3: Анализ супернатанта согласно ТР StMoBKDC с использованием антитела к B-Raf для детекции. Ряд А1: S. typhi Ty21a pMoBKDC, А2: S. typhi Ty21a, A3: S. typhimurium aroA. Слитый белок BRaf-V600E KD-CtxB отмечен вверху стрелкой и имеет предполагаемый размер приблизительно 48 кД. Белковый маркер: предварительно окрашенная белковая лестница Bench-Mark, No 10748-010.
B) анализ, аналогичный А с использованием недавно полученного антитела к HlyA в качестве антитела для детекции. Ряды 1 и 2: конструкции StMoPC, ряд 3: S. typhi Ty21a MoBKDC; ряд 4: Е. coli MoBKDC, ряд 5: S. typhi Ty21a, ряд 6: S. typhimurium aroA. Белковый маркер: предварительно окрашенная белковая лестница Bench-Mark, No 10748-010 Invitrogen BenchMark, партия 1315592.
На фигуре 18 представлена полная нуклеотидная последовательность (5'>3') ненагруженного вектора pMKhlyi.
Содержание всех приведенных ссылок и патентов таким образом включено в данное описание в виде ссылки во всей их полноте. Изобретение более подробно раскрывается с помощью следующих примеров, однако без ограничения ими.
Осуществление изобретения
Примеры
Пример 1: Конструирование, экспрессия и секреция слитых белков PSA-CtxB в различных штаммах бактериальных носителей
Для доказательства осуществимости и демонстрации эффективности системы секреции гемолизина типа I E. coli секретировать слитые белки опухолевых антигенов, в данном случае простата-специфического антигена (PSA), и компонентов белковых токсинов, в данном случае CtxB, в качестве адъюванта конструируют слитый белок PSA-CtxB, как описано в данном контексте. Экспрессию и секрецию исследуют на различных грамотрицательных бактериальных штаммах, которые потенциально эффективны в качестве живых вакцинных штаммов в противоопухолевой терапии. Молекулярно-биологическое клонирование основано на плазмиде/векторе экспрессии pMOhlyl, которые описаны ранее (см. статьи Gentschev et al., 2005; Gentschev et al., 1996, WO 03/072789).
1A Конструирование устойчивого к к анамицину производного вектора экспрессии pMOhlyl
Замену кассеты устойчивости кампициллину pMOhlyl осуществляют, как описано в статье Datsenko и Wanner, 2000.
Вкратце, смысловой праймер Р1 (5'-GAGTATTCAACATTTCCGTGTCGC CCTTATTCCCTTTTTTGGTGTAGGCTGGAGCTGCTTC-S') и антисмысловой праймер Р2 (5'-GCGATCTGTCTATTTCGTTCATCCATAGTTGCCTGACTCCCCATATGAATATCCTCCTTA-S') и плазмиду pKD4 в качестве матрицы используют для ПЦР с целью получения фрагмента, несущего ген устойчивости к канацицину (KanR), фланкированный участками, гомологичными гену устойчивости к ампициллину (неподчеркнутые). Штамм Е. coli BW251 14, несущий плазмиду pKD46 и направленную плазмиду pMOhlyl, выращивают при 37°C с среде LB (Difco) с добавлением 0,2% L-(+)-арабинозы в течение 3-4 часов перед трансформацией ПЦР-фрагментом.
После трансформации бактериальные клетки распределяют на пластинках агара LB, содержащего 25 мкг/мл канамицина, и инкубируют при 37°C в течение ночи. На следующий день клоны KanR снимают и инкубируют в течение дополнительных 48 ч в среде LB, содержащей 50 мкг/мл канамицина, чтобы избавиться от всех плазмид, обусловливающих устойчивость к ампициллину.
Наконец, селектируют клоны с KanR и чувствительным к ампициллину фенотипом. Замену гена ApR кассетой KanR подтверждают с помощью ПЦР и секвенирования. Полученную в результате плазмиду называют pMKhlyl.
1В Клонирование плазмиды pMKhly-PSA
Смысловой праймер PSA-Nsi1 (5'-GATTGGTGATGCATCCCTCAT-S'; рестрикционные сайты Nsil подчеркнуты) и антисмысловой праймер PSA-Nsi2 (5'-GGTGCTCATGCATTGGCCACG-3') используют для амплификации фрагмента ДНК, кодирующуго PSA, с помощью ПЦР. ПЦР проводят в циклере Thermal Cycler 60 (Biometra, Göttingen, Germany) в течение 30 циклов при 94°C в течение 1 мин, 54°C в течение 1 мин и 72°C в течение 2 мин.
После разрезания рестрикционным ферментом Nsil фрагмент ДНК, несущий ген psa вводят в единственный сайт Nsil экспортирующего вектора pMKhly1. Полученную в результате плазмиду pMKhly-PSA выделяют из Е. coli DH5α (Invitrogene, Germany), анализируют с использованием рестрикционного анализа и секвенируют.
1C Клонирование плазмиды pMKhly-PSA/CtxB
Смысловой праймер Ptac-Sall (5'-AAAAAAGTCGACGGCTGTGCAGGTCGTAAATCACTGC-3') и антисмысловой праймер Ptac-NotI (5'-AAAAAAGCGGCCGCGAAATTGTTATCCGCTCACAATTCC-3') используют для амплификации с помощью ПЦР фрагмента ДНК из 201 пары оснований, кодирующего промотор Ptac из плазмиды pGEX-6p-1 (Amersham Bioscience, Germany). ПЦР проводят в термоциклере T3 (Biometra, Germany): в течение 30 циклов при 95°C в течение 30 сек, 55°C в течение 30 сек и 72°C в течение 90 сек. Смысловой праймер Rbs-Notl-прямой (5'-AAAAAAGCGGCCGCTAAGGATGAATTATGATTAAATTAAAATTTGG-S') и антисмысловой праймер ctb-Sall обратный (5'-TTTATAGTCGACTTAATTTGCCATACTAATTGCGGCAATCGC-S') используют для амплификации с помощью ПЦР фрагмента ДНК из 413 пар оснований, кодирующего сайт связывания рибосом, и полной кодирующей CtxB из V. cholerae El tor. ПЦР проводят в термоциклере ТЗ (Biometra, Germany): стадия 1-30 циклов при 95°C в течение 30 сек, 50°C в течение 30 сек и 72°C в течение 2 мин. После очистки с помощью набора Qiaquick PCR Purification Kit (Qiagen, Germany) и разрезания обоих фрагментов рестрикционным ферментом NotI два фрагмента лигируют, получая в результате фрагмент из 594 пар оснований Ptac-cfxB. Полученный в результате фрагмент также очищают и после разрезания рестрикционным ферментом Sail фрагмент Ptac-cfxB вводят в единственный сайт Sail экспортирующего вектора pMKhly-PSA. Полученную в результате плазмиду pMKhly-PSA/CtxB выделяют из Е. coli DH5α (Invitrogene, Germany), анализируют и секвенируют.
1D Клонирование слитой конструкции PSA-CtxB-HlyAs
Смысловой праймер 5'ctxB Nsil (5'-GCATATGCACATGCATCACCTCAAAATATTACTGAT-3') и антисмысловой праймер 3' ctxB Srfl Nsil (5'-GGCTTTTTTATATCTTATGCATGCCCGGGCATTGCGGCAATCGC-3') (сайт Srfl выделен жирным шрифтом) используют для амплификации посредством ПЦР фрагмента ДНК из ~300 пар оснований, представляющего ген ctxB из V. cholerae El tor. После очистки с помощью набора Qiaquick PCR Purification Kit (Qiagen, Germany) и разрезания рестрикционным ферментом Nsil фрагмент ДНК, несущий целый ген ctxB без N-концевой сигнальной последовательности вводят в единственный сайт Nsil экспортирующего вектора pMKhly1. Полученную в результате плазмиду pMKhly-CtxB выделяют из Е. coli DH5α (Life Technologies), анализируют и секвенируют. Человеческий ген psa амплифицируют из плазмиды pCDNA3PSA с помощью ПЦР с использованием праймеров 5-PSA-Blunt (5'-GTGGGAGGCTGGGAGTGC-3') и 3-PSA-Blunt (5'-GGGGTTGGCCACGATGGT-S'). ПЦР проводят в циклере Thermal Cycler 60 (Biometra, Göttingen, Germany) в течение 30 циклов при 94°C в течение 1 мин, 56°C в течение 1 мин и 72°C в течение 2 мин. Затем продукт ДНК 0,7 т.п.н. клонируют в сайт Srfl pMKhly-CtxB. Полученную в результате плазмиду pMKhly-CtxB-PSA проверяют с помощью рестрикционного анализа и секвенирования.
1Е Экспрессия и секреция слитых белков в различных бактериальных штаммах
Используя стандартные способы электропорации плазмидой pMKhly-CtxB-PSA трансформируют электрокомпетентные бактериальные штаммы. Устойчивые к канамицину отдельные колонии снимают и выращивают в среде BHI (Beckton Dickinson, USA: телячий головной мозг, экстракт из 200 г, 6,0 г/л; бычье сердце, экстракт из 250 г, 9,8 г/л; протеозопептон 10,0 г/л; хлорид натрия 5,0 г/л; декстроза 2,0 г/л; динатрийфосфат 2,5 г/л) до густоты 1×109 клеток/мл. После выращивания 20 мл культуры центрифугируют в течение 30 мин при 4000 об/мин (3000 g) в центрифуге Heraeus при 4°C. 18 мл супернатанта переносят в свежую пробирку. Затем добавляют 1,8 мл ТСА (трихлоруксусной кислоты, Applichem, Germany), жидкость перемешивают и инкубируют в течение по меньшей мере 1 часа на льду. После инкубирования суспензию центрифугируют в течение 30 мин при 4000 об/мин (3000 g) в центрифуге Heraeus при 4°C. Супернатант сливают с осадка и осадок промывают 1 мл ацетона р.а. (чистого для анализа) (Applichem, Germany). Осадок центрифугируют в течение 10 минут при 4000 об/мин (3000 g) в центрифуге Heraeus при 4°C. Осадок сушат на воздухе и помещают в 150 мкл 5х буфера Леммли (70 мМ Трис-HCl, рН 6,8, 40% (об./об.) глицерина, 3% (об./об.) додецил сульфата натрия, 5% (об./об.) 2-меркаптоэтанола и 0,05% (масс./об.) бромфенолового синего) с добавлением или без добавления β-меркаптоэтанола (см. статью Laemmli, 1970). Для каждой дорожки при SDS PAGE (электрофорез в полиакриламидном геле с использованием додецилсульфата натрия) используют 20 мкл раствора. Разделенные белки электрофоретически переносят (вестерн-блот) на нитроцеллюлозную мембрану Hybond ECL (Amersham-Pharmacia, Little Chalfont, UK) и блокируют в течение ночи PBS (хлорид калия 0,20 г/л, дигидрофосфат калия 0,20 г/л, хлорид натрия 8,00 г/л, дигидрофосфат динатрий безводный 1,15 г/л), содержащим 1% BSA (бычий сывороточный альбумин). Мембрану промывают в PBS-Твин 0,05%, инкубируют с поликлональным кроличьим антителом к PSA (1:750, DAKO, Denmark), антителом k CtxB (1:1000, Zytomed, Berlin, Germany) или антителом к HlyAs (см. статью Gentschev et al., 1996)) и затем инкубируют с HRP (пероксидза хрена)-связанным IgG к кролику (1/2000; Dianova, Hamburg, Germany) в течение 1 часа. Вестерн-блот проявляют с помощью набора с усиленной флуоресценцией (GE Healthcare Life Science, Germany).
На фигуре 11 представлена функциональная экспрессия и секреция слитых белков PSA-CtxB в ряде бактериальных штаммов, включая Escherichia coli spp., Salmonella dublin, Citrobacter spp, Salmonella typhi Ty21a, Salmonella typhimurium spp, Erwinia spp и Shigella flexneri spp.
Штамм Salmonella typhi Ty21a, экспрессирующий и секретирующий слитый белок PSA-CtxB депонирован в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ) под номером DSM 19244.
Пример 2: Иммунный ответ и защита от прививания опухолевых клеток у мышей, иммунизированных штаммами Salmonella, секретирующими слитые белки опухолевых антигенов и CtxB или CtxB в виде монокомпонента
Посредством следующих экспериментов повышенная защитная эффективность секретируемых слитых белков опухолевых антигенов и белковых токсинов продемонстрирована на модели опухолей у животных. Для данной модели штамм Salmonella typhimurium aroA (SL7207) pMKhly-CtxB-PSA, экспрессирующий и секретирующий слитый белок CtxB-PSA сравнивают с другими контрольными штаммами.
2А Способы иммунизации
Мышей DBA/2 иммунизируют три раза с интервалом 3 недели. Для иммунизации бактериями животных предварительно лечат введением 50 мкл 7% NaHCO3 внутрижелудочно с целью повышения внутрижелудочного рН. Через 5-10 минут после предварительного лечение вводят 5×108 живых нечувствительных к канамицину бактерий в объеме 100 мкл PBS внутрижелудочно. В качестве контроля мышей иммунизируют внутримышечно голой (депротеинизированной) плазмидной ДНК, кодирующей PSA (рсДНК-PSA), как описано в статье Fensterle et al., 2005.
2В Т-клеточные ответы
Через семь дней после последней иммунизации спленоциты иммунизированных мышей получают, как опубликовано ранее (см. статью Fensterle et al., 1999), пропусканием селезенки через сетку с последующим лизисом эритроцитов. Анализ ELISPOT для детекции PSA-специфических Т-клеток CD8+ проводят согласно способу, опубликованному в статье Fensterle et al., 1999. Вкратце, для анализа ex vivo PSA-специфических Т-клеток CD8+ 96-луночные нитроцеллюлозные планшеты (Millititer HA; Millipore, Bedford, MA) покрывают 5 мкг/мл моноклонального антитела (mAb) к мышиному ИФН-γ R4 (PharMingen) в 100 мкл карбонатного буфера (см. статью Fensterle et al., 1999), рН 9,6. После инкубирования в течение ночи при 4°C и блокирования 1% BSA в PBS добавляют 1×105 спленоцитов, полученных от вакцинированных мышей в 100 мкл RP10 (см. статью Fensterle et al., 1999)/лунку.
Для анализа ответа Т-клеток CD8+ используют экспрессирующий PSA клон клеток Р815 PPSA 24; вкратце, данный клон экспрессирует PSA полной длины посредством промотора CMV, кодируемый плазмидой pCDNA3 (см. статью Fensterle et al., 2005). После инкубирования в течение 20-22 ч при 37°C, 5% CO2 в присутствии 30 ед./мл ИЛ-2 планшеты промывают и инкубируют в течение дополнительных 2 ч со 100 мкл биотинилированного mAb XMB1.2 к мышиному ИФН-γ (0,25 мкг/мл, PharMingen). Планшеты промывают и инкубируют в течение 1 ч при 37°C в присутствии 100 мкл разведения 1/20000 связанного с щелочной фосфатазой стрептавидина (PharMingen). Пятна визуализируют добавлением 50 мкл готового к применению субстрата BCIP/NBT (Sigma, St. Louis, МО), растворенного в воде. Пятна подсчитывают под препаровальным микроскопом при 3-кратном увеличении.
Частоту пептид-специфических Т-клеток (CTL) выражают как число ИФН-γ-секретирующих клеток/105 спленоцитов. Для анализа ответов Т-клеток после повторной стимуляции 2,5×107 спленоцитов повторно стимулируют 2×106 облученных экспрессирующих PSA клеток Р815 (см. статью Fensterle et al., 2005) в среде RP10 (см. статью Fensterle et al., 1999) в присутствии 60 ед./мл рекомбинантного ИЛ-2 в течение 5 дней. Анализ ELISPOT проводят, как описано выше, используя различные количества повторно стимулированных клеток (105, 3×104, 104 или 3×103/лунку), смешанных с 4×105 фидерных клеток (= свежеполученных спленоцитов от непримированных (наивных) мышей DBA/2) и 105 клеток PPSA24.
Индукция клеточного иммунного ответа, особенно цитотоксических Т-клеток CD8+, играет важную роль в плане эффективности противоопухолевой терапии (см. статьи Boon et al., 2006; Rosenberg, 2001). Вследствие этого сначала тестируют эффективность рекомбинантного бактериального штамма Salmonella typhimurium (SL7207), несущего вектор экспрессии pMKhly-CtxB-PSA, в индукции PSA-специфического иммунного ответа Т-клеток CD8+.
Для этой цели 64 самок мышей DBA-2 в возрасте 10-14 недель иммунизируют перорально рекомбинантным SL7207/pMKhly-CtxB (n=10), SL7207/pMKhly-CtxB-PSA (n=10), SL7207/pMKhly-PSA (n=10), SL7207/pMKhly-PSA/CxtB (n=10), SL7207/pMKhly1 (n=7) и голой рсДНК3-PSA (n=10) в качестве положительного контроля, как описано в данном контексте.
На фигуре 12 представлены данные ELISPOT, показывающие, что иммунизированные ДНК мыши демонстрируют выраженные ответы Т-клеток CD8+. После повторной стимуляции явные иммунные ответы определяют у животных, вакцинированных Salmonella, секретирующей белок PSA, слитый с CtxB, но не штаммами, секретирующими PSA в виде монокомпонента или раздельно PSA и CtxB. Интересно, что животные, вакцинированные Salmonella, секретирующей токсин CtxB в виде монокомпонента, также проявляют существенные иммунные ответы после повторной стимуляции. Данные ответы наиболее вероятно обусловлены NK-клетками или другими клетками врожденной иммунной системы, которая неспецифически распознает клеточную линию-мишень. Следовательно, на основании данных по секретируемому CtxB в виде монокомпонента можно заключить, что иммунный ответ, наблюдаемый для секретируемого слитого белка CtxB-PSA (секретирующие ИФН-γ спленоциты) состоит из ответа Т-клеток CD8+, а также выраженного ответа врожденной иммунной системы (наиболее вероятно, NK-клеток).
2С Защита от прививания опухолевых клеток
Для анализа защитной способности иммунизации 6-7 мышей/группу иммунизируют согласно вышеописанной схеме. Через три недели после третьей иммунизации мышам прививают PPSA 24 (см. выше) путем подкожной инъекции 1×106 клеток с каждого бока выбритой кожи живота. Мышей мониторируют в течение 14 дней на предмет появления опухолей и оценивают объем опухолей путем измерения самого большого (а) и самого маленького (b) диаметра опухоли. Объем опухоли рассчитывают как тело, образуемое вращением эллипса, используя следующую формулу:
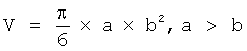
Анализируют значимость результатов посредством одностороннего ANOVA и повторного теста множественного сравнения Даннетта с использованием пакета программ Graph Pad Prism. Повторный тест проводят только в случае, когда ANOVA показывает значимость результатов.
На фигуре 13 представляют результаты как среднее ±SD (стандартная ошибка). В день 6, 9, 12 и 14 наблюдают существенные защитные эффекты. Как ожидают, вакцинация голой ДНК включает в качестве контроля мышей, полностью защищенных от роста опухолей. Однако вакцинация голой ДНК показывает самое большее среднюю эффективность у человека. Что касается бактериальных конструкций, то вакцинный штамм SL7207/pMKhly-CtxB-PSA (экспрессирующий и секретирующий слитый белок CtxB-PSA) оказывается наиболее эффективным. Он существенно уменьшает объем опухоли в 9, 12 и 14 после прививания опухоли. Известно также, что штамм SL7207/pMKhly-CtxB уменьшает рост опухоли на величины, сравнимые с SL7207/pMKhly-CtxB-PSA, в день 14. Хотя недостоверно, данный задерживающий эффект хорошо согласуется с клеточным ответом, который измеряют с помощью анализа ELISPOT. Более того, штамм SL7207/pMKhly-PSA также дает значительную защиту в день 14, что показывает, что данный штамм также индуцирует Т-клеточный ответ, который ниже порога определения. Напротив, штамм SL7207/pMKhly-PSA/CxtB не индуцирует соответствующий эффект и остается в том же диапазоне, что SL7207/pMKhly1 как монокомпонент.
Пример 3: Экспрессия и секреция слитого белка онкоген-токсин в S. typhi Ty21a
Аналогично примеру 1 используют другой опухолевый антиген, домен киназы онкогена B-Raf V600E с делецией 10 аа (аминокислот) в домене киназы (домен киназы BRaf V600E киназа dead (KD) или, сокращенно, BRaf* KD, более коротко BKD). Слитый белок данного онкогена и компонентов белкового токсина, в данном случае CtxB, в качестве адъюванта, конструируют, как описано в данном контексте. Экспрессию и секрецию демонстрируют на человеческом вакцинном штамме S. typhi Ty21 а. Молекулярное клонирование основано на использовании плазмиды/вектора экспрессии pMOhlyl, который описан ранее (см. статьи Gentschevet al., 2005; Gentschev et al., 1996), и способ клонирования аналогичен описанному в примере 1 данной заявки. Полученный в результате вектор называют pMBKDC. Последовательность слитого белка приведена на фигуре 10.
На фигуре 17 показана экспрессия и секреция слитого белка с использованием вектора pMBKDC, электропорированного в штамм Salmonella typhi Ty21a и другие бактериальные штаммы.
Штамм Salmonella typhi Ty21a, экспрессирующий и секретирующий слитый белок BKD-CtxB, депонирован в Немецкой коллекции Микроорганизмов и клеточных культур (DSMZ) под номером DSM 19245.
Пример 4: Сравнение слитой конструкции OVA-CtxB с другими генетически кодируемыми иммунологическими адъювантами
Для сравнения иммунологической эффективности слитых белков токсин-антиген, секретируемых живыми вакцинами, конструируют слитый белок, включающий широко используемый для иммунологических исследований антиген, куриный овальбумин (OVA), и CtxB.
Смысловой праймер Nsil-OVA-прямой 5' - CAT GTA TGC ATT AGC CAT GGT ATA CCT GG-3' и антисмысловой праймер Nsil-OVA-обратный 5'-TTT TTT ATG CAT AAG GG AAA CAC CAC ATC TGC C-3' используют для амплификации посредством ПЦР фрагмента ДНК из 1033 пар оснований, представляющий собой ген ova (NM205152). После очистки с помощью набора QIAquick PCR Purification Kit (Qiagen, Germany) и разрезания рестрикционным ферментом Nsil фрагмент ДНК, несущий целый ген ova без N-концевой сигнальной последовательности, вводят в единственный сайт Nsil экспортирующего вектора pMKhlyl. Полученную в результате плазмиду pMKhly-Ova выделяют из Е. coli DH5α (Life Technologies), анализируют и секвенируют.
Смысловой праймер 5'cfxB Nsil (5'-GCATATGCAATGCATCACCTCAAAATATTACTGAT-3') и антисмысловой праймер 3' ctxB Srfl Nsil (5'-GGCTTTTTTATATCTTATGCATGCCC GGGCATTGCGGCAATCGC-3') (сайт Srfl выделен жирным шрифтом) используют для амплификации с помощью ПЦР фрагмента ДНК из ~300 пар оснований, представляющий собой ген ctxB из V. cholerae El tor. После очистки с помощью набора QIAquick PCR Purification Kit (Qiagen, Germany) и разрезания рестрикционным ферментом Nsil фрагмент ДНК, несущий целый ген ctxB без N-концевой сигнальной последовательности, вводят в единственный сайт Nsil экспортирующего вектора pMKhly1. Полученную в результате плазмиду pMKhly-CtxB выделяют из Е. coli DH5α (Life Technologies), анализируют и секвенируют. Человеческий ген ova амплифицируют из плазмиды pCl- OVA с помощью ПЦР с использованием праймеров 5-OVA-Sfrl GCC АТС ATG ТСА GCT СТА и 3-OVA-Sfrl AGG GGA ААС АСА ТСТ GCC. ПЦР проводят в циклере Thermal Cycler 60 (Biometra, Göttingen, Germany) в течение 30 циклов 1 мин при 94°C, 1 мин при 49°C и 2 мин 30 сек при 72°C. Затем продукт ДНК 1,1 т.п.н. клонируют в сайт Srfl pMKhly-CtxB. Полученную в результате плазмиду pMKhly-CtxB-OVA проверяют с помощью рестрикционного анализа и секвенирования.
Способность данного слитого белка вызывать иммунные ответы Th1 сравнивают со способностью кодируемого вектором pMKhly OVA в виде монокомпонента в комбинации с плазмидами, доставляющими ДНК (кодирующими белки под контролем эукариотического промотора), кодирующими интерфероны (ИФН-γ), интерлейкины (ИЛ-12) и хемокины (IP-10).
Способы иммунизации и анализ ELISPOT осуществляют, как описано выше. Вкратце, мышей C57BL/6 иммунизируют перорально три раза различными штаммами. Через 7 дней после конечной иммунизации спленоциты удаляют, повторно стимулируют в течение 5 дней и исследуют с помощью анализа ELISPOT. Для повторной стимуляции спленоцитов используют клетки, обработанные в импульсном режиме пептидом SIINFEKL (буквы означают аминокислоты), эпитопом овальбумина Н-2b с рестрикцией МНС-1.
На фигуре 14 показана повышенная эффективность слитого белка, включающего куриный овальбумин (OVA) и CtxB, в индукции OVA-специфических ответов Т-клеток CD8+ и потенциальных ответов врожденной иммунной системы по сравнению с конструкциями, совместно доставляющими ИФН-γ или IP-10. Он проявляет эффективность, близкую к эффективности штамма с конструкцией совместной доставки ИЛ-12.
Пример 5: Экспрессия и секреция вирусных антигенов
Пригодность микроорганизмов, соответствующих изобретению, для экспрессии и секреции какого-либо (неопухолевого) антигена демонстрируют посредством экспрессии белка гемагглютинина H1 вируса куриного гриппа H5N1, слитого с CtxB в ветеринарном вакцинном штамме Salmonella typhimurium VacT.
Смысловой праймер 5'-HA-G: 5'-ATC TGT CAA ATG GAG AAA-3' и антисмысловой праймер 3-НА12С: 5'-TAC TCC ACT TAT TTC CTC TCT-3' используют в ПЦР, амплифицируя фрагмент ДНК, кодирующий Н5 без С-концевого мембранного домена (Н12С). Затем продукт ПЦР клонируют в сайт Srfl pMKhly-CtxB. Полученную в результате плазмиду pMKhly-CtxB-H12C исследуют посредством рестрикционного анализа и секвенирования. Штамм S. typhimurium VacT/pMKhly-CtxB-H12C эффективно экспрессирует и секретирует гибридный белок Н5, как показано с помощью иммуноблоттинга с поликлональными антителами, образующимися к CtxB и HlyA (см. фиг.2). Количество секретируемого Н5 составляет 2-3 мкг белка/мл супернатанта в условиях эксперимента.
Бактерии выращивают в среде ВНI до густоты 1×109 клеток/мл. 20 мл культуры центрифугируют в течение 30 мин при 4000 об/мин (3000 g) и 4°C в центрифуге Hereaus. 18 мл супернатанта переносят в свежую пробирку. Затем добавляют 1,8 мл ТСА (трихлоруксусной кислоты, Applichem, Germany), жидкость перемешивают и инкубируют на льду в течение по меньшей мере 1 часа. После инкубирования суспензию центрифугируют в течение 30 мин при 4000 об/мин (3000 g) в центрифуге Heraeus при 4°C. Супернатант сливают с осадка и осадок промывают 1 мл ацетона р.а. (чистого для анализа) (Applichem, Germany); осадок центрифугируют в течение 10 минут при 4000 об/мин (3000 g) при 4°C в центрифуге Heraeus. Осадок сушат на воздухе и помещают в 150 мкл 5х буфера Леммли. Для каждой дорожки при SDS PAGE используют 20 мкл раствора. Разделенные белки электрофоретически переносят (вестерн-блот) на нитроцеллюлозную мембрану Hybond ECL (Amersham-Pharmacia, Little Chalfont, UK) и блокируют в течение ночи PBS, содержащим 1% BSA. Мембрану промывают в РВS-Твин 0,05%, инкубируют с поликлональным кроличьим антителом к CtxB (1:1000, Zytomed, Berlin, Germany) или антителом к HlyAs (см. статью Gentschev et al., 1996)) и затем инкубируют с HRP-связанным IgG к кролику (1/2000; Dianova, Hamburg, Germany) в течение 1 часа. Вестерн-блот проводят с использованием набора с усиленной флуоресценцией (GE Healthcare Life Science, Germany).
На фигуре 15 представлена эффективная экспрессия и функциональная секреция слитого белка HA12C-CtxB в ветеринарном вакцинном штамме Salmonella typhimurium VacT.
Пример 6: Экспрессия и секреция слитых белков токсинных адъювантов и опухолевых антигенов посредством систем секреции типа III
Некоторые грамотрицательные бактерии несут системы секреции типа III, которые можно использовать для секреции антигенов. Как правило, данные системы можно использовать для непосредственной инъекции гетерологичных антигенов в клетки. В данном примере токсинный компонент Yersinia, который служит в качестве сигнала секреции (YopE), генетически слит с фрагментом термолабильной субъединицы энтеротоксина В (LT-B), служащим в качестве адъюванта, и опухолевым антигеном PSA. Данная конструкция должна экспрессироваться в подходящих, предпочтительно аттенюированных штаммах Yersinia, которые приводят к секреции слитого белка путем эндогенного механизма секреции типа III с помощью сигнальной последовательности YopE.
В данном примере генетическое слияние YopE18-LT-B-PSA клонируют в сайт EcoRI плазмиды paCYC1 84 для использования конструкции с такой же ориентацией, в которой используют ген, с получением продукта, экспрессируемого плазмидой, кодирующей промотор cat (http://www.fermentas.com/techinfo/nucleicacids/mappacyc184.htm). Для повышенного уровня экспрессии можно использовать любой подходящий промотор.
В дальнейшем иллюстрируют способ клонирования. Хромосомную ДНК из Е. coli, включающую субъединицу В термолабильного энтеротоксина, или вектор, включающий данную последовательность (GenBank регистрационный номер AF242418), амплифицируют, используя следующие праймеры в реакции ПЦР:
Eco-yope18-f (включающий сайт EcoRI site + сигнальную последовательность YopE18):
5'-CTGAATTCATGAAAATATCATCATTTATTTCTACATCACTGCCCCTGCCGGCATCAGTGTCAATAAAGTAAAATGTTATGTTTTAT3' (5'-участок показан черным: сайт EcoRI+3 основания 5', показан красным: последовательность, кодирующая сигнал для секреции типа III белка YopE (aa 1-18), GenBank регистрационный No M92066; участок, выделенный курсивом: участок гомологии гена LT-B (GB регистрационный No AF242418), основания 4-28).
Фосфорилированный обратный праймер LT-B-r: 5'GTTTTCCATACTGATTGCCGCAATTGAATTGG-3' (обратная цепь 372-341 GB AF242418).
Параллельно последовательность PSA, как описано в данной заявке, из вектора pMKCtxB-PSA амплифицируют, используя следующие праймеры: фосфорилированный прямой праймер psa-f: 5'-GTGGGAGGCTGGGAGTGCGAG-3', обратный праймер psa-eco-r: 5'-CCTGAATTCTTAGACGTGATACCTTGAAGCAC (5', показано черным: сайт EcoRI+3 основания 5', показаны синим: стоп-кодон, показан черным: 3'-участок последовательности PSA, данная заявка).
Затем два фрагмента лигируют с помощью ДНК-лигазы в подходящих условиях. После лигирования новый фрагмент амплифицируют во второй реакции ПЦР, используя праймеры Eco-yope18-f и PSA-Eco-R, описанные выше.
Полученный в результате фрагмент очищают, используя подходящий набор очистки для ПЦР, с целью удаления буфера и олигонуклеотидов и затем разрезают ферментом EcoRI в подходящих условиях реакции. Полученный в результате фрагмент из 1040 пар оснований очищают, используя электрофорез в агарозном геле, и лигируют в разрезанный EcoRI вектор paCYC184. После секвенирования выбирают устойчивые к тетрациклину, чувствительные к cm (канамицину) клоны, которые имеют правильную последовательность, интегрированную в той ориентации, что ген cm, и трансформируют подходящий штамм Yersinia, используя электропорацию. Данный вакцинный штамм будет секретировать слияние LT-B-PSA посредством эндогенной системы секреции типа III.
Пример 7: Экспрессия и секреция слияний токсин-антиген в грамположительных бактериях
Грамположительные и отрицательные бактерии имеют различные предварительные условия для секреции. Грамотрицательные бактерии имеют две мембраны и, вследствие этого, механизм секреции в дополнение к сигналам секреции является необходимым для полной секреции. В случае грамположительных бактерий достаточно сигнальной последовательности секреции.
В данном примере описана система для секреции слияния столбнячный токсоид-PSA в Listeria с использованием сигнала секреции р60. На первой стадии вектор pUC18(PS)actAOVATinlA (см. статью Loeffler et al., 2006) разрезают, используя Nsil, и затем обрабатывают, 3'-5' экзонуклеазой в подходящих условиях. Наконец, фрагмент очищают электрофорезом на агарозном геле.
Параллельно фрагмент С столбнячного токсоида (GenBank регистрационный номер No. M 12739) амплифицируют из подходящего источника, например, геномной ДНК из штамма Clostridium tetanii, с использованием следующих фосфорилированных праймеров:
tetc прямой: 5'-TAAAAATCTGGATTGTTGGGTTG-3', где подчеркнутое дополнительное основание введено, чтобы обеспечить правильную рамку слияния.
tetc обратный: 5'-ATCATTTGTCCATCCTTCATCTG-3'.
Фрагмент BRAF KD амплифицируют из плазмиды рМО BKDC, описанной в данной заявке, с использованием фосфорилированных праймеров:
br прямой: 5'-GATTGGGAGATTCCTGATG-3'
br обратный: 5'-CCCGTGG ACAGG AAACGCACC-3'.
Оба фрагмента очищают с использованием подходящего набора для ПЦР-очистки и затем лигируют с помощью ДНК-лигазы в подходящих условиях. После лигирования фрагмент очищают снова, используя подходящий набор для ПЦР-очистки, и амплифицируют, используя tetc прямой и br обратный праймер. После амплификации полученный в результате фрагмент из 2280 пар оснований очищают электрофорезом на агарозном геле и лигируют в очищенный фрагмент 3'-5' экзонуклеазы Nsil pUC18(PS)actAOVATinlA. После лигирования и трансформации E. coli отбирают клон, несущий фрагмент в рамке считывания. Затем плазмиду разрезают, используя PstI и SacI, и фрагмент из 2837 пар оснований очищают, используя гель-электрофорез, и лигируют в соответствующим образом разрезанный PstI и SacI и очищают вектор рSР118 (см. статью Loeffler et al., 2006). Полученным в результате вектором pSP118-act-TetC-BRaf*KD трансформируют штамм Listeria, несущий делецию trpS, например Listeria monocytogenes у trps у aroA/В (см. статью Loeffler et al., 2006). В данном исследовании плазмиду стабилизируют с помощью trpS на основе плазмиды ("Сбалансированная летальная система"), и плазмида кодирует фаговый лизин, что приводит к внутриклеточному лизису штамма. Кассету act-TetC-BRaf*KD экспрессируют главным образом в эукариотических клетках-хозяевах под контролем промотора actA, в которых происходит некоторое внеклеточное вытекание (Loeffler et al., 2006). Сигнальная последовательность кассеты actA приводит к секреции слитого белка TetC-BRaf*KD.
Пример 8: Эукариотические клетки, секретирующие слияния токсин-антиген
В данном примере конструируют эукариотическую клеточную линию, например, опухолевую клеточную линию, которая секретирует слитый белок CtxB-PSA. Для этой цели используют универсальный сигнал секреции (US 6733997), который в принципе обеспечивает секрецию соответствующей экспрессирующей кассеты в клетках различной природы, включая клетки млекопитающих, дрожжи и прокариотические клетки.
На первом этапе слияние CtxB-BRaf* KD амплифицируют из плазмиды рМО BKDC (см. данную заявку), используя следующие праймеры:
СВ-прямой: 5'-atcGGATCCTCAAAATATTACTGATTTGTGTGC-3' (строчные буквы: спейсер подчеркнут: сайт BamHI, прописные буквы: CtxB 5')
СВ-обратный: 5'-tagGGATCCTTAGTGGACAGGAAACGCACCATATCC-3' (строчные буквы: спейсер подчеркнут: сайт BamHI, курсив: стоп-кодон, прописные буквы: BRaf* KD 3'). Затем продукт ПЦР очищают, используя подходящий набор для ПЦР-очистки, и потом частично разрезают (фрагмент включает внутренний сайт BamHI) с помощью BamHI. Фрагмент частичного разрезания из 1231 пары оснований выделяют с помощью электрофореза на агарозном геле и затем лигируют в разрезанную BamHI и гель-очищенную плазмиду pVtgEGFP (патентная заявка США 6733997). Потом отбирают клон, несущий в ориентации рамки считывания с Vtgss, и называют pVtgCtxBRAf. Данной плазмидой можно трансформировать посредством электропорации эукариотическую клеточную линию, которую можно выбрать с помощью селекционной кассеты kan/neo. Клеточные линии могут представлять собой стабилизированные клеточные линии, подобные раковым клеточным линиям, предназначенным для использования в качестве гетерологичной противораковой вакцины, или опухолевым клеткам, выделенным у пациента, предназначенным для использования в качестве аутологичных противораковых вакцин. В любом случае сигнал секреции Vtgss, кодируемый вектором pVtgEGFP, генетически слитый с CtxB-BRaf*KD, будет приводить в результате к секреции слитого белка.
Используя аналогичный подход, слитые белки можно также экспрессировать в дрожжах. В качестве примера демонстрируют модифицированную стратегию клонирования, как показано на фиг.11 заявки US 6733997. На первом этапе кассету, включающую слияние Vtgss-CtxB-BRaf*KD из вышеописанной плазмиды pVtgCtxBRAf, вырезают с помощью Eagl и Eco47III и вводят в вектор pBSPGK (см. US 6733997, фиг.11). Затем полученный в результате вектор разрезают Sacl и HindIII и фрагмент, включающий элемент PGK и слияние Vtgss-CtxB-BRaf*KD интегрируют в соответствующий участок вектора pYEX-S1 аналогично описанию заявки US 6733997 (см. фиг.11). Полученной в результате плазмидой можно трансформировать путем электропорации штаммы дрожжей, такие как Saccharomyces cerevisiae. Дрожжи будут экспрессировать и секретировать слитый белок и могут быть использованы для целей вакцинации.
Сокращения
Библиография
Agren, L С, Ekman, L, Lowenadler, В., Nedrud, J. G., and Lycke, N. Y. (1999). Adjuvanticity of the cholera toxin A1-based gene fusion protein, CTA1-DD, is critically dependent on the ADP-ribosyltransferase and Ig-binding activity (Адъювантность слитого белка на основе гена холерного токсина A1, СТА1-DD, в существенной степени зависит от активности АДФ-рибозилтрансферазы и lg-связывающей активности. J Immunol 162, 2432-2440.
Anderson, R.J., Pasetti, M.F., Sztein, M.В., Levine, M.M., and Noriega, F.R. (2000). Delta guaBA attenuated Shigella flexneri 2a strain CVD 1204 as a Shigella vaccine and as a live mucosal delivery system for fragment С of tetanus toxin. (Дельта guaBA аттенюированный штамм Shigella flexneri 2a CVD 1204 как вакцина против Shigella и как живая система доставки через слизистые для фрагмента С столбнячного токсина). Vaccine 18, 2193-2202.
Austin, E.A., Graves, J.F., Hite, L.A., Parker, С.Т., and Schnaitman, C.A. (1990). Genetic analysis of lipopolysaccharide core biosynthesis by Escherichia coli K-12: insertion mutagenesis of the rfa locus. (Генетический анализ биосинтеза липополисахаридного ядра Escherichia coli K-12: инсерционный мутагенез локуса rfa). J Bacteriol 172, 5312-5325.
Balsalobre, С, Silvan, J.M., Berglund, S., Mizunoe, Y., UhNn, B.E., and Wai, S.N. (2006). Release of the type I secreted alpha-haemolysin via outer membrane vesicles from Escherichia coll. (Высвобождение секретируемого α-гемолизина типа I через пузырьки наружной мембраны из Escherichia coli). Mol Microbiol 59, 99-112.
Barry, E. M., Gomez-Duarte, O., Chatfield, S., Rappuoli, R., Pizza, M., Losonsky, G., Galen, J., and Levine, M.M. (1996). Expression and immunogenicity of pertussis toxin 81 subunit-tetanus toxin fragment С fusions in Salmonella typhi vaccine strain CVD 908. (Экспрессия и иммуногенность слияний субъединица S1 токсина коклюша-фрагмента С столбнячного токсина). Infect Immun 64, 4172-4181.
Becerra, J.С, Arthur, J.F., Landucci, G.R., Forthal, D.N. and Theuer, C.P. (2003). CD8+ T-cell mediated tumor protection by Pseudomonas exotoxin fused to ovalbumin in C57BL/6mice. (Опосредованная Т-клетками CD8+ противоопухолевая защита посредством экзотоксина Pseudomonas, слитого с овальбумином, у мышей C57BL/6). Surgery 133, 404-410.
Boon, Т., Coulie, P. G., Van den Eynde, B. J., and van der Bruggen, P. (2006). Human Т cell responses against melanoma. (Ответы человеческих Т-клеток в отношении меланомы). Annu Rev Immunol 24, 175-208.
Boyd, A.P., Ross, P.J., Conroy, H., Mahon, N., Lavelle, E.C, and Mills, K.H. (2005). Bordetella pertussis adenylate cyclase toxin modulates innate and adaptive immune responses: distinct roles for acylation and enzymatic activity in immunomodulation and cell death, (токсин аденилатциклаза Bordetella pertussis модулирует врожденный и адаптивный иммунный ответы: особая роль ацилирования и ферментной активности в иммуномодуляции и гибели клетки). J Immunol 175, 730-738.
Brassier, F., Weber-Levy, M., Mock, M., and Sirard, J.C. (2000). Protective antigen-mediated antibody response against a heterologous protein produced in vivo by Bacillus anthracis. (Защитный опосредованный антигеном ответ антител в отношении гетерологичного белка, продуцируемого in vivo Bacillus anthracis). Infect Immun 68, 5731-5734.
Bumann, D. (2001). Regulated antigen expression in live recombinant Salmonella enterica serovarTyphimurium strongly affects colonization capabilities and specific CD4(+)-T-cell responses. (Регулируемая экспрессия антигена в живой рекомбинантной Salmonella enterica серовар Typhimurium сильно влияет на способности колонизации и специфические ответы Т-клеток CD4+). Infect Immun 69, 7493-7500.
Busby, S., and Ebright, R. H. (1994). Promoter structure, promoter recognition, and transcription activation in prokaryotes. (Структура промотора, распознавание промоторов и активация транскрипции у прокариот). Cell 79, 743-746.
Butterton, J.R., Beattie, D.Т., Gardel, C. L, Carroll, P.A., Hyman, Т., Killeen, K.P., Mekalanos, J.J., and Calderwood, S.B. (1995). Heterologous antigen expression in Vibrio cholerae vector strains. (Экспрессия гетерологичного антигена в векторных штаммах Vibrio cholerae) Infect Immun 63, 2689-2696.
Byun, Y., Ohmura, M., Fujihashi, K., Yamamoto, S., McGhee, J. R., Udaka, S., Kiyono, H., Takeda, Y., Kohsaka, Т., and Yuki, Y. (2001). Nasal immunization with E. coli vero-toxin 1 (VT1)-B subunit and a nontoxic mutant of cholera toxin elicits serum neutralizing antibodies. (Назальная иммунизация В-субъединицей веро-токсина 1 (VT1) E. coli и нетоксичный мутантхолерного токсина вызывает образование сывороточных нейтрализующих антител). Vaccine 19, 2061-2070.
Campos, J., Martinez, E., Marrero, K., Silva, Y., Rodriguez, B. L, Suzarte, E., Ledon, Т., and Fando, R. (2003). Novel type of specialized transduction for CTX phi or its satellite phage RS1 mediated by filamentous phage VGJ phi in Vibrio cholerae. (Новый тип специализированной трансдукции для CTX phi или его сателлитного фага RS1, опосредованной филаментозным фагом VGJ phi в Vibrio cholerae) J Bacteriol 185, 7231-7240.
Cardenas, L, and Clements, J.D. (1992). Oral immunization using live attenuated Salmonella spp. as carriers of foreign antigens. (Пероральная иммунизация с использованием живой аттенюированной Salmonella spp. как носителя чужеродных антигенов). Clin Microbiol Rev 5, 328-342.
Cardenas, L, and Clements, J.D. (1993). Development of mucosal protection against the heat-stable enterotoxin (ST) of Escherichia coli by oral immunization with a genetic fusion delivered by a bacterial vector. (Развитие защиты слизистой от термостабильного энтеротоксина (ST) Escherichia coli путем пероральной иммунизации генетическим слиянием, доставляемым бактериальным вектором). Infect Immun 61, 4629-4636.
Chabalgoity, J.A., Harrison, J.A., Esteves, A., Demarco de Hormaeche, R., Ehrlich, R., Khan, С.М., and Hormaeche, С.Е. (1997). Expression and immunogenicity of an Echinococcus granulosus fatty acid-binding protein in live attenuated Salmonella vaccine strains. (Экспрессия и иммуногенность связывающего жирные кислоты белка Echinococcus granulosus в живых аттенюированных вакцинных штаммах Salmonella). Infect Immun 65, 2402-2412.
Chabalgoity, J.A., Khan, С.М., Nash, A.A., and Hormaeche, С.Е. (1996). A Salmonella typhimurium htrA live vaccine expressing multiple copies of a peptide comprising amino acids 8-23 of herpes simplex virus glycoprotein D as a genetic fusion to tetanus toxin fragment С protects mice from herpes simplex virus infection. (Живая вакцина на основе Salmonella typhimurium htrA, экспрессирующая множество копий пептида, содержащего аминокислоты 8-23 гликопротеина D вируса простого герпеса в виде генетического слияния с фрагментом С столбнячного токсина, защищает мышей от инфекции вируса простого герпеса). Mol Microbiol 19, 791-801.
Chabalgoity, J. A., Moreno, M., Carol, H., Dougan, G., and Hormaeche, C. E. (2000). Salmonella typhimurium as a basis for a live oral Echinococcus granulosus vaccine. (Salmonella typhimurium как основа для живой пероральной вакцины Echinococcus granulosus). Vaccine 19, 460-469.
Chabalgoity, J.A., Villareal-Ramos, В., Khan, С.M., Chatfield, S.N., de Hormaeche, R.D., and Hormaeche, C.E. (1995). Influence of preimmunization with tetanus toxoid on immune responses to tetanus toxin fragment C-guest antigen fusions in a Salmonella vaccine carrier. (Влияние предварительной иммунизации столбнячным токсоидом на иммунные ответы в отношении слияний фрагмент С столбнячного токсина - чужеродный антиген и вакцинном носителе Salmonella). Infect Immun 63, 2564-2569.
Chacon, M.R., Londono, P., Dougan, G., and Selkirk, M.E. (1996). Heterologous expression of the cuticular-glutathione peroxidase of lymphatic filariae in an attenuated vaccine strain of Salmonella typhimurium abrogates H-2 restriction of specific antibody responses. (Гетерологичная экспрессия кутикулярной глутатион-пероксидазы лимфатических фибрилл в аттенюированном вакцинном штамме Salmonella typhimurium отменяет Н-2-рестрикцию ответов специфических антител). Parasite Immunol 18, 307-316.
Chen, I., Finn, Т. M., Yanqing, L, Guoming, Q., Rappuoli, R., and Pizza, M. (1998). A recombinant live attenuated strain of Vibrio cholerae induces immunity against tetanus toxin and Bordetella pertussis tracheal colonization factor. (Рекомбинантный живой аттенюированный штамм Vibrio cholerae индуцирует иммунитет в отношении столбнячного токсина и трахеального фактора колонизации Bordetella pertussis). Infect Immun 66, 1648-1653.
Cheng-hua, S., Cheng, C, Jing-sheng, Z., Jiezhi., L, and Qing-jun, M. (1995). Gene fusion of cholera toxin В subunit and HBV PreS2 epitope and the antigenicity of fusion protein. (Слияние генов В-субъединицы холерного токсина и эпитопа HBV PreS2 и антигенность слитого белка). Vaccine 13, 933-937.
Clemens, J., Savarino, S., Abu-Elyazeed, R., Safwat, M., Rao, M., Wierzba, Т., Svennerholm, A.M., Holmgren, J., Frenck, R., Park, E., and Naficy, A. (2004). Development of pathogenicity-driven definitions of outcomes for a field trial of a killed oral vaccine against enterotoxigenic Escherichia coli in Egypt: application of an evidence-based method. (Разработка обусловливаемых патогенностью определений конечных итогов для полевых испытаний убитой пероральной вакцины против энтеротоксигенной Escherichia coli в Египте, применение метода на основе доказательств). J Infect Dis 189, 2299-2307.
Datsenko, К.A., and Wanner, В.L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. (Одностадийная инактивация хромосомных генов в Escherichia coli K-12 с использованием продуктов ПЦР). Proc Natl Acad Sci USA 97, 6640-6645.
Dietrich, G., Bubert, A., Gentschev, I., Sokolovic, Z., Simm, A., Catic, A., Kaufmann, S.H., Hess, J., Szalay, A.A., and Goebel, W. (1998). Delivery of antigen-encoding plasmid DNA into the cytosol of macrophages by attenuated suicide Usteria monocytogenes. (Доставка кодирующей антиген плазмидной ДНКв цитозоль макрофагов посредством аттенюированной суицидной Usteria monocytogenes). Nat Biotechnol 16, 181-185.
Dietrich, G., Viret, JF, and Gentschev, I. (2003). Haemolysin A and listeriolysin - two vaccine delivery tools for the induction of cell-mediated immunity. (Гемолизин А и листериолизин - два инструмента доставки вакцины для индукции опосредованного клетками иммунитета). Int. J. Parasitol 33, 495-505.
Dunn, G.P., Old, L.J., and Schreiber, R.D. (2004). The immunobiology of cancer immunosurveillance and immunoediting. (Иммунобиология иммуновыживаемости и иммунокоррекции рака) Immunity 21, 137-148.
Dunstan, S.J., Simmons, C.P., and Strugnell, R.A. (2003). In vitro and in vivo stability of recombinant plasmids in a vaccine strain of Salmonella enterica var. typhimurium. (Стабильность in vitro и in vivo рекомбинантных плазмид в вакцинном штамме Salmonella enterica var. typhimurium). FEMS Immunol Med Microbiol37, 111-119.
Eriksson, A.M., Schon, K.M., and Lycke, N.Y. (2004). The cholera toxin-derived CTA1-DD vaccine adjuvant administered intranasallydoes not cause inflammation or accumulate in the nervous tissues. (Вакцинный адъювант CTA1-DD, полученный из холерного токсина, вводимый интраназально, не вызывает воспаления или накопления в нервных тканях). J Immunol 173, 3310-3319.
Fath, M.J., Skvirsky, R.C, and Kolter, R. (1991). Functional complementation between bacterial MDR-like export systems: colicin V, alpha-hemolysin, and Erwinia protease. (Функциональная комплементация между бактериальными MDR-подобными системами экспорта: колицином V, α-гемолизином и протеазой Erwinia). J Bacteriol 173, 7549-7556.
Fensterle, J., Grode, L, Hess, J., and Kaufmann, S. H. E. (1999). Effective DNA vaccination against listeriosis by prime/boost inoculation with the gene gun. (Эффективная ДНК-вакцинация против листериоза путем первичной/бустерной инокуляции гена gun). J Immunol 163, 4510-4518.
Fensterle, J., Schwartz, V., Riedmiller, H., and Rapp, U. R. (2005). Animal models for DNA vaccination against prostate cancer using PSA encoding plasmids. (Модели на животных ДНК-вакцинации против рака простаты с использованием PSA-кодирующих плазмид). Onkologie 28 (доп. 2), 52.
Freytag, L.С, and Clements, J.D. (1999). Bacterial toxins as mucosal adjuvants. (Бактериальные токсины как адъюванты для слизистых). Curr Top Microbiol Immunol 236, 215-236.
Freytag, L.С., and Clements, J.D. (2005). Mucosal adjuvants (Адъюванты для слизистых). Vaccine 23, 1804-1813.
Garmory, H.S., Brown, K.A., and Titball, R.W. (2002). Salmonella vaccines for use in humans: present and future perspectives. (Вакцины на основе Salmonella, предназначенные для применения у человека, настоящее и будущие перспективы). FEMS Microbiol Rev 26, 339-353.
Garmory, H.S., Titball, R.W., Griffin, K.F., Hahn, U., Bohm, R., and Beyer, W. (2003). Salmonella enterica serovar typhimurium expressing a chromosomally integrated copy of the Bacillus anthracis protective antigen gene protects mice against an anthrax spore challenge. (Salmonella enterica серовар typhimurium, экспрессирующая хромосомно интегрированную копию гена защитного антигена Bacillus anthracis, защищает мышей от заражения спорами сибирской язвы). Infect Immun 71, 3831-3836.
Gentschev, I., Dietrich, G., and Goebel, W. (2002a). The E. coli alpha-hemolysin secretion system and its use in vaccine development. (Система секреции α-гемолизина E. coli и ее использование в разработке вакцин) Trends Microbiol 10, 39-45.
Gentschev, I., Dietrich, G., Spreng, S., Pilgrim, S., Stritzker, J., Kolb-Maurer, A., and Goebel, W. (2002b). Delivery of protein antigens and DNA by attenuated intracellular bacteria. (Доставка белковых антигенов и ДНК посредством аттенюированных внутриклеточных бактерий). Int J Med Microbiol 291, 577-582.
Gentschev, I., Fensterle, J., Schmidt, A., Potapenko, Т., Troppmair, J., Goebel, W., and Rapp, U.R. (2005). Use of a recombinant Salmonella enterica serovar typhimurium strain expressing C-Raf for protection against C-Raf induced lung adenoma in mice. (Использование рекомбинантного штамма Salmonella enterica serovar typhimurium, экспрессирующего C-Raf для защиты от индуцированной С-raf аденомы легкого у мышей). ВМС Cancer 5, 15.
Gentschev, I., Mollenkopf, H., Sokolovic, Z., Hess, J., Kaufmann, S.H., and Goebel, W. (1996). Development of antigen-delivery systems, based on the Escherichia coli hemolysin secretion pathway. (Разработка систем доставки антигена на основе пути секреции гемолизина Escherichia coli) Gene 179, 133-140.
Gentschev, I., Dietrich, G., Spreng, S., Neuhaus, В., Maier, E., Benz, R., Goebel, W., Fensterle, J., and Rapp, UR. (2004). Use of the alpha-hemolysin secretion system of Escherichia coli for antigen delivery in the Salmonella typhi Ty21a vaccine strain. (Использование системы секреции α-гемолизина Escherichia coli для доставки антигена в вакцинном штамме Salmonella typhi Ty21a). Int J Med Microbiol 294, 363-371.
Gomez-Duarte, О.G., Galen, J., Chatfield, S.N., Rappuoli, R., Eidels, L, and Levine, M.M. (1995). Expression of fragment С of tetanus toxin fused to a carboxyl-terminal fragment of diphtheria toxin in Salmonella typhi CVD 908 vaccine strain. (Экспрессия фрагмента С столбнячного токсина, слитого в карбоксил-концевым фрагментом дифтерийного токсина в вакцинном штамме Salmonella typhi CVD 908). Vaccine 13, 1596-1602.
Hajishengallis, G., Harokopakis, E., Hollingshead, S.K., Russell, M.W., and Michaiek, S.M. (1996). Construction and oral immunogenicity of a Salmonella typhimurium strain expressing a streptococcal adhesin linked to the A2/B subunits of cholera toxin. (Конструирование и пероральная иммуногенность штамма Salmonella typhimurium, экспрессирующего стрептококковый адгезин, связанный с субъединицами A2/B холерного токсина). Vaccine 14, 1545-1548.
Hajishengallis, G., Nawar, H., Tapping, R.I., Russell, M.W., and Connell, T.D. (2004). The Type II heat-labile enterotoxins LT-IIa and LT-IIb and their respective В pentamers differentially induce and regulate cytokine production in human monocytic cells. (Термолабильные энтеротоксины типа II LT-IIa и их соответствующие В-пентамеры дифференцирование индуцируют и регулируют продукцию человеческих моноцитарных клеток). Infect Immun 72, 6351-6358.
Harokopakis, E., Hajishengallis, G., Greenway, T.E., Russell, M.W., and Michaiek, S.M. (1997). Mucosal immunogenicity of a recombinant Salmonella typhimurium-cloned heterologous antigen in the absence or presence of coexpressed cholera toxin A2 and В subunits. (Иммуногенность для слизистых клонированного в рекомбинантную Salmonella typhimurium гетерологичного антигена в отсутствие или в присутствии соэкспрессированных A2- и В-субъединиц холерного токсина) Infect Immun 65, 1445-1454.
Hess, J., Gentschev, I., Miko, D., Welzel, M., Ladel, C, Goebel, W., and Kaufmann, S.H. (1996). Superior efficacy of secreted over somatic antigen display in recombinant Salmonella vaccine induced protection against listenosis. (Повышенная эффективность представления секретированного относительно соматического антигена в вакцине на основе рекомбинантной Salmonella, индуцирующей защиту от листериоза). Proc Natl Acad Sci USA 93, 1458-1463.
Holmgren, J., Adamsson, J., Anjuere, F., Clemens, J., Czerkinsky, C, Eriksson, K., Flach, C.F., George-Chandy, A., Harandi, A.M., Lebens, M., et al. (2005). Mucosal adjuvants and anti-infection and anti-immunopathology vaccines based on cholera toxin, cholera toxin В subunit and CpG DNA. (Адъюванты для слизистых и вакцины против инфекций и иммунопатологий на основе субъединицы В холерного токсина и ДНК CpG). Immunol Lett 97, 181-188.
Holmgren, J., and Czerkinsky, C. (2005). Mucosal immunity and vaccines. (Иммунитет слизистых и вакцины). Nat Med 11, S45-53.
Holmgren, J., Czerkinsky, C, Eriksson, K., and Mharandi, A. (2003). Mucosal immunisation and adjuvants: a brief overview of recent advances and challenges. (Иммунизация и адъюванты для слизистых - краткий обзор современных достижений и проблем). Vaccine 21 доп. 2, S89-95.
Hormozi, K., Parton, R., and Coote, J. (1999). Adjuvant and protective properties of native and recombinant Bordetella pertussis adenylate cyclase toxin preparations in mice. (Адъювантные и защитные свойства препаратов токсина аденилатциклазы нативной и рекомбинантной Bordetella pertussis у мышей). FEMS Immunol Med Microbiol 23, 273-282.
Huang, Y., Hajishengallis, G., and Michaiek, S.M. (2000). Construction and characterization of a Salmonella enterica serovar typhimurium clone expressing a salivary adhesin of Streptococcus mutans under control of the anaerobically inducible nirB promoter. (Конструирование и характеризация клона Salmonella enterica серовар typhimurium, экспрессирующего слюнный адгезин Streptococcus mutans под контролем энаэробного индуцируемого промотора nirB). Infect Immun 68, 1549-1556.
Hunt, P.D., and Hardy, S.J. (1991). Heat-labile enterotoxin can be released from Escherichia coli cells by host intestinal factors. (Термолабильный энтеротоксин может быть высвобожден из клеток Escherichia coli посредством кишечных факторов хозяина). Infect Immun 59, 168-171.
Jackson, R.J., Marinaro, M., VanCott, J.L, Yamamoto, M., Okahashi, N., Fujihashi, K., Kiyono, H., Chatfield, S.N., and McGhee, J.R. (1996). Mucosal immunity: regulation by helper Т cells and a novel method for detection. (Иммунитет слизистых - регуляция Т-клетками-хелперами и новый метод детекции). J Biotechnol 44, 209-216.
Jagusztyn-Krynicka, E.K., Clark-Curtiss, J.E., and Curtiss, R., 3rd (1993). Escherichia coli heat-labile toxin subunit В fusions with Streptococcus sobrinus antigens expressed by Salmonella typhimurium oral vaccine strains: importance of the linker for antigenicity and biological activities of the hybrid proteins. (Слияния В-субъединицы термолабильного токсина Escherichia coli с антигенами Streptococcus sobrinus, экспрессируемыми пероральными вакцинными штаммами Salmonella typhimurium - важность линкера для антигенности и биологической активности гибридных белков). Infect Immun 61, 1004-1015.
Khan, С.M., Villarreal-Ramos, В., Pierce, R.J., Demarco de Hormaeche, R., McNeNI, H., Ali, Т., Chatfield, S., Capron, A., Dougan, G., and Hormaeche, C. E. (1994a). Construction, expression, and immunogenicity of multiple tandem copies of the Schistosoma mansoni peptide 115-131 of the P28 glutathione S-transferase expressed as C-terminal fusions to tetanus toxin fragment С in a live aro-attenuated vaccine strain of Salmonella. (Конструирование, экспрессия и иммуногенность множества тандемных копий пептида 115-131 глутатион-трансферазы S Schistosoma mansoni P28, экспрессируемой в виде С-концевых слияний с фрагментом С столбнячного токсина в живом aro-аттенюированном вакцинном штамме Salmonella). J Immunol 153, 5634-5642.
Khan, С.М., Villarreal-Ramos, В., Pierce, R.J., Riveau, G., Demarco de Hormaeche, R., McNeill, H., Ali, Т., Fairweather, N., Chatfield, S., Capron, A., and et al. (1994b). Construction, expression, and immunogenicity of the Schistosoma mansoni P28 glu-tathione S-transferase as a genetic fusion to tetanus toxin fragment С in a live Aro attenuated vaccine strain of Salmonella. (Конструирование, экспрессия и иммуногенность глутатион-трансферазы S Schistosoma mansoni P28 как генетического слияния с фрагментом С столбнячного токсина в живом Aro-аттенюированном вакцинном штамме Salmonella). Proc Natl Acad Sci USA 91, 11261-11265.
Konieczny, М.P., Suhr, М., Noll, A., Autenrieth, I.B., and Alexander Schmidt, М. (2000). Cell surface presentation of recombinant (poly-) peptides including functional T- cell epitopes by the AIDA autotransporter system. (Презентация на клеточной поверхности рекомбинантных полипептидов, включающих функциональные Т-клеточные эпитопы, посредством системы аутотранспортеров AIDA). FEMS Immunol Med Microbiol 27, 321-332.
Koprowski, H., 2nd, Levine, М.М., Anderson, R.J., Losonsky, G., Pizza, М., and Barry, E.М. (2000). Attenuated Shigella flexneri 2a vaccine strain CVD 1204 expressing colonization factor antigen I and mutant heat-labile enterotoxin of enterotoxigenic Escherichia coli. (Аттенюированный вакцинный штамм Shigella flexneri 2a CVD 1204, экспрессирующий антиген фактора колонизации и мутантный термолабильный энтеротоксин энтеротоксигенной Escherichia coli). Infect Immun 68, 4884-4892.
Kweon, M.N., Yamamoto, M., Watanabe, F., Tamura, S., Van Ginkel, F.W., Miyauchi, A., Takagi, H., Takeda, Y., Hamabata, Т., Fujihashi, K., et al. (2002). A nontoxic chimeric enterotoxin adjuvant induces protective immunity in both mucosal and systemic compartments with reduced IgE antibodies. (Адъювант на основе нетоксичного химерного энтеротоксина индуцирует защитный иммунитет в обоих, слизистом и системном отделах с пониженным уровнем антител IgE). J Infect Dis 186, 1261-1269.
Laemmli, U.К. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. (Расщепление структурных белков во время сборки бактериофага T4). Nature 227, 680-685.
Lahiri, S.S. (2000). Bacterial toxins - an overview. (Бактериальные токсины - обзор) J Nat Toxins 9, 381-408.
Lavelle, E.C, McGuirk, P., and Mills, K.H. (2004). Molecules of infectious agents as immunomodulatory drugs. (Молекулы инфекционных агентов как иммуномодулирующие лекарственные препараты). Curr Top Med Chem 4, 499-508.
Lee, J.J., Sinha, K.A., Harrison, J.A., de Hormaeche, R.D., Riveau, G., Pierce, R.J., Capron, A., Wilson, R.A., and Khan, C.M. (2000). Tetanus toxin fragment C expressed in live Salmonella vaccines enhances antibody responses to its fusion partner Schistosoma haematobium glutathione S-transferase. (Фрагмент С столбнячного токсина, экспрессируемый в живых вакцинах Salmonella, усиливает ответы антител на его партнера слияния глутатион S-трансферазу Schistosoma haematobium). Infect Immun 68, 2503-2512.
Loeffler, D.I., Schoen, C.U., Goebel, W., and Pilgrim, S. (2006). Comparison of different live vaccine strategies in vivo for delivery of protein antigen or antigen-encoding DNA and mRNA by virulence-attenuated Listeria monocytogenes. (Сравнение различных стратегий живых вакцин in vivo для доставки белкового антигена или кодирующей антиген ДНК и мРНК с использованием Listeria monocytogenes с аттенюированной вирулентностью). Infect Immun 74, 3946-3957.
Loessner, M.J., Wendlinger, G., and Scherer, S. (1995). Heterogeneous endolysins in Listeria monocytogenes bacteriophages: a new class of enzymes and evidence for conserved holin genes within the siphoviral lysis cassettes. (Гетерогенные эндолизины бактерифагов Listeria monocytogenes - новый класс ферментов и подтверждение присутствия консервативных генов холина в сифовирусных лизирующих кассетах). Mol Microbiol 16, 1231-1241.
Lycke, N. (2005). From toxin to adjuvant: basic mechanisms for the control of mucosal IgA immunity and tolerance. (От токсина к адъюванту - основные механизмы контроля иммунитета IgA слизистых и толерантности). Immunol Lett 97, 193-198.
Lycke, N., Tsuji, Т., and Holmgren, J. (1992). The adjuvant effect of Vibrio cholerae and Escherichia coli heat-labile enterotoxins is linked to their ADP-ribosyltransferase activity. (Адъювантный эффект термолабильных энтеротоксинов Vibrio cholerae и Escherichia coli связан с активностью АДФ-рибозилтрансферазы). Eur J Immunol 22, 2277-2281.
McSorley, S.J., Rask, C, Pichot, R., Julia, V., Czerkinsky, C, and Glaichenhaus, N. (1998). Selective tolerization of Th1-like cells after nasal administration of a cholera toxoid-LACK conjugate. (Избирательная толеризация Th1-подобных клеток после назального введения конъюгата холерный токсоид-LACK). Eur J Immunol 28, 424-432.
Mendelson, 1., Gat, O., Aloni-Grinstein, R., Altboum, Z., Inbar, I., Kronman, C, Bar-Haim, E., Cohen, S., Velan, В., and Shafferman, A. (2005). Efficacious, nontoxigenic Bacillus anthracis spore vaccines based on strains expressing mutant variants of lethal toxin components. (Эффективные нетоксигенные споровые вакцины Bacillus anthracis на основе штаммов, экспрессирующих мутантные варианты компонентов летального токсина). Vaccine, 23, 5688-5697.
Mesnage, S., Weber-Levy, M., Haustant, M., Mock, M., and Fouet, A. (1999). Cell surface-exposed tetanus toxin fragment С produced by recombinant Bacillus anthracis protects against tetanus toxin. (Фрагмент С столбнячного токсина, представляемого на клеточной поверхности, продуцируемый рекомбинантным Bacillus anthracis, защищает от столбнячного токсина). Infect Immun 67, 4847-4850.
Michalkiewicz, J., Stachowski, J., Barth, C, Patzer, J., Dzierzanowska, D., and Madalinski, K. (1999). Effect of Pseudomonas aeruginosa exotoxin A on IFN-gamma synthesis: expression of costimulatory molecules on monocytes and activity of NK cells. (Эффект эндотоксина A Pseudomonas aeruginosa на синтез ИФН-γ: экспрессия костимулирующих молекул на моноциты и активность NK-клеток). Immunol Lett 69, 359-366.
Michl, P., and Gress, T.M. (2004). Bacteria and bacterial toxins as therapeutic agents for solid tumors. (Бактерии и бактериальные токсины в качестве лечебных факторов для солидных опухолей). Curr Cancer Drug Targets 4, 689-702.
Morona, R., Manning, P.A., and Reeves, P. (1983). Identification and characterization of the TolC protein, an outer membrane protein from Escherichia coli. (Идентификация и характеризация белка TolC, белка наружной мембраны из Escherichia coli). J Bacteriol 153, 693-699.
Muhlen, K.A., Schumann, J., Wittke, F., Stenger, S., Van Rooijen, N., Van Kaer, L, and Tiegs, G. (2004). NK cells, but not NKT cells, are involved in Pseudomonas aeruginosa exotoxin A-induced hepatotoxicity in mice. (NK-клетки, но не NKT-клетки участвуют в гепатотоксичности, индуцируемой экзотоксином A Pseudomonas aeruginosa, у мышей). J Immunol 172, 3034-3041.
Mutsch, M., Zhou, W., Rhodes, P., Bopp, M., Chen, R.Т., Under, Т., Spyr, C, and Steffen, R. (2004). Use of the inactivated intranasal influenza vaccine and the risk of Bell's palsy in Switzerland. (Использование инактивированной интраназальной вакцины против гриппа и риск развития паралича Белла в Швейцарии). N Engl J Med 350, 896-903.
Orr, N., Galen, J.E., and Levine, M.M. (1999). Expression and immunogenicity of a mutant diphtheria toxin molecule, CRM(197), and its fragments in Salmonella typhi vaccine strain CVD 908-htrA. (Экспрессия и иммуногенность мутантной молекулы дифтерийного токсина CRM(197) и ее фрагментов в вакцинном штамме Salmonella typhi CVD 908-htrA). Infect Immun 67, 4290-4294.
Poggi, A., Spada, F., Costa, P., Tomasello, E., Revello, V., Pella, N., Zocchi, M.R., and Moretta, L. (1996). Dissection of lymphocyte function-associated antigen 1-dependent adhesion and signal transduction in human natural killer cells shown by the use of cholera or pertussis toxin. (Анализ ассоциированной с функцией лимфоцитов антиген 1-зависимой адгезии и сигнальной трансдукции в человеческих натуральных клетках-киллерах, показанной с использованием холерного токсина или токсина коклюша) Eur J Immunol 26, 967-975.
Pogonka, Т., Klotz, С, Kovacs, F., and Lucius, R. (2003). A single dose of recombinant Salmonella typhimurium induces specific humoral immune responses against heterologous Eimeria tenella antigens in chicken. (Однократная доза рекомбинантной Salmonella typhimurium индуцирует специфические гуморальные иммунные ответы в отношении гетерологичных антигенов Eimeria tenella у кур). Int J Parasitol 33, 81-88.
Ramirez, R., Carracedo, J., Zamzami, N., Castedo, M., and Kroemer, G. (1994). Pertussis toxin inhibits activation-induced cell death of human thymocytes, pre-B leukemia cells and monocytes. (Токсин коклюша ингибирует индуцированную активацией гибель человеческих тимоцитов, предлейкозных В-клеток и моноцитов). J Exp Med 180, 1147-1152.
Ranallo, R.Т., Fonseka, С.P., Cassels, F., Srinivasan, J., and Venkatesan, M.M. (2005). Construction and characterization of bivalent Shigella flexneri 2a vaccine strains SC608 (pCFAI) and SC608(pCFAI/LTB) that express antigens from enterotoxigenic Escherichia coli. (Конструирование и характеризация бивалентных вакцинных штаммов Shigella flexneri 2a SC608(pCFAI) и SC608(pCFAI/LTB), которые экспрессируют антигены из энтеротоксигенной Escherichia coli). Infect Immun 73, 258-267.
Reveneau, N., Geoffroy, M. С, Locht, С, Chagnaud, P., and Mercenier, A. (2002). Comparison of the immune responses induced by local immunizations with recombinant Lactobacillus plantarum producing tetanus toxin fragment С in different cellular locations. (Сравнение иммунных ответов, индуцированных местными иммунизациями рекомбинантной Lactobacillus plantarum, продуцирующей фрагмент С столбнячного токсина в различных местоположениях клетки). Vaccine 20, 1769-1777.
Roland, K. L, Tinge, S. A., Killeen, K. P., and Kochi, S. K. (2005). Recent advances in the development of live, attenuated bacterial vectors. (Современные достижения в создании живых аттенюированных бактериальных векторов). Curr Opin Mol Ther 7, 62-72.
Rosenberg, S. A. (2001). Progress in human tumour immunology and immunotherapy. (Развитие иммунологии и иммунотерапии опухолей человека). Nature 411, 380-384.
Ryan, E.Т., Butterton, J.R., Smith, R.N., Carroll, P.A., Crean, T.I., and Calderwood, S.B. (1997a). Protective immunity against Clostridium difficile toxin A induced by oral immunization with a live, attenuated Vibrio cholerae vector strain. (Защитный иммунитет против токсина A Clostridium difficile, индуцируемый пероральной иммунизацией живым аттенюированным векторным штаммом Vibrio cholerae). Infect Immun 65, 2941-2949.
Ryan, E.Т., Butterton, J.R., Zhang, Т., Baker, M.A., Stanley, S.L., Jr., and Calderwood, S.B. (1997b). Oral immunization with attenuated vaccine strains of Vibrio cholerae expressing a dodecapeptide repeat of the serine-rich Entamoeba histolytica protein fused to the cholera toxin B subunit induces systemic and mucosal antiamebic and anti-V. cholerae antibody responses in mice. (Пероральная иммунизация аттенюированными вакцинными штаммами Vibrio cholerae, экспрессирующими додекапептидный повтор богатого серином белка Entamoeba histolytica, слитый с В-субъединице и холерного токсина, индуцирует системный и слизистый ответы антиамебных антител и антител κ V. cholerae у мышей). Infect Immun 65, 3118-3125.
Sanchez, J., Wallerstrom, G., Frednksson, M., Angstrom, J., and Holmgren, J. (2002). Detoxification of cholera toxin without removal of its immunoadjuvanticity by the addition of (STa-reIated) peptides to the catalytic subunit. A potential new strategy to generate immunostimulants for vaccination. (Детоксификация холерного токсина без удаления его иммуноадъювантности путем добавления (STa-родственных) пептидов к каталитической субъединице. Потенциальная новая стратегия генерации иммуностимуляторов для вакцинации). J Biol Chem 277, 33369-33377.
Sanchez, AE., Aquino, G., Ostoa-Saloma, P., Laclette, JP., and Rocha-Zavaleta, L (2004). Cholera toxin B-subunit gene enhances mucosal Immunoglobulin A, Th1-type and CD8+ cytotoxic responses when co-administered intradermally with a DNA vaccine. (Ген В-субъединицы холерного токсина усиливает ответы иммуноглобулина А слизистых, Th1-типа и цитотоксических ответов CD8+ при совместном введении внутрикожно с ДНК-вакциной). Clinic Diagnost Laborat Immun 11, 711-719.
Schmid-Grendelmeier, P., and Crameri, R. (2001). Recombinant allergens for skin testing. (Рекомбинантные аллергены для кожного тестирования). Int Arch Allergy Immunol 125, 96-111.
Schodel, F., Enders, G., Jung, M.C, and Will, H. (1990). Recognition of a hepatitis В virus nucleocapsid T-cell epitope expressed as a fusion protein with the subunit В of Escherichia coli heat labile enterotoxin in attenuated salmonellae. (Распознавание Т-клеточного эпитопа нуклеокапсида вируса гепатита С, экспрессируемого в виде слитого белка с субъединицей В термолабильного энтеротоксина Escherichia coli в аттенюированных сальмонеллах). Vaccine 8, 569-572.
Shaw, C.A., and Starnbach, M.N. (2003). Using Modified Bacterial Toxins to Deliver Vaccine Antigens. ASM News 69, 384-389. Silva, A.J., Mohan, A., and Benitez, J.A. (2003). Cholera vaccine candidate 638: intranasal immunogenicity and expression of a foreign antigen from the pulmonary pathogen Coccidioides immitis. (Кандидат холерной вакцины 638 - интраназальная иммуногенность и экспрессия чужеродного антигена, выделенного из легочного патогена Coccidioides immitis). Vaccine 21, 4715-4721.
Smerdou, С, Anton, I.M., Plana, J., Curtiss, R., 3rd, and Enjuanes, L (1996). A continuous epitope from transmissible gastroenteritis virus S protein fused to E. coli heat-labile toxin В subunit expressed by attenuated Salmonella induces serum and secretory immunity. (Постоянный эпитоп из S-белка трансмиссивного вируса гастроэнтерита, слитый с субъединицей В термолабильного токсина E. coli, экспрессируемый аттенюированной Salmonella, индуцирует сывороточный и секреторный иммунитет). Virus Res 41, 1-9.
Somner, E.A., Ogun, S.A., Sinha, K.A., Spencer Valero, L.M., Lee, J.J., Harrison, J.A., Holder, A.A., Hormaeche, C.E., and Khan, C.M. (1999). Expression of disulphide - bridge-dependent conformational epitopes and immunogenicity of the carboxy-terminal 19 kDa domain of Plasmodium yoelii merozoite surface protein-1 in live attenuated Salmonella vaccine strains. (Экспрессия зависимых от дисульфидных мостиков конформационных эпитопов и иммуногенность карбокси-концевого домена 19 кД поверхностного белка-1 мерозитов Plasmodium yoelii в живых аттенюированных вакцинных штаммах Salmonella. Microbiology 145 (часть 1), 221-229.
Sory, M.P., and Cornelis, G.R. (1990). Delivery of the cholera toxin В subunit by using a recombinant Yersinia enterocolitica strain as a live oral carrier. (Доставка субъединицы В холерного токсина с использованием рекомбинантного штамма Yersinia enterocolitica в качестве живого перорального носителя). Res Microbiol 141, 921-929.
Spangrude, G.J., Sacchi, F., Hill, H.R., Van Epps, D.E., and Daynes, R.A. (1985). Inhibition of lymphocyte and neutrophil chemotaxis by pertussis toxin. (Ингибирование хемотаксиса лимфоцитов и нейтрофилов посредством токсина коклюша). J Immunol 135, 4135-4143.
Su, G.F., Brahmbhatt, H.N., de Lorenzo, V., Wehland, J., and Timmis, K.N. (1992). Extracellular export of Shiga toxin B-subunit/haemolysin A (C-terminus) fusion protein expressed in Salmonella typhimurium aroA-mutant and stimulation of B-subunit specific antibody responses in mice. (Внеклеточный экспорт слитого белка В-субъединицы шига-токсина/гемолизина А (С-конца), экспрессируемого в мутанте Salmonella typhimurium aroA, и стимуляция ответов В-субъединица-специфических антител у мышей). Microb Pathog 13, 465-476.
Thungapathra, M., Sharma, С, Gupta, N., Ghosh, R.K., Mukhopadhyay, A., Koley, H., Nair, G.В., and Ghosh, A. (1999). Construction of a recombinant live oral vaccine from a non-toxigenic strain of Vibrio cholerae 01 serotype inaba biotype E1 Tor and assessment of its reactogenicity and immunogenicity in the rabbit model. (Конструирование рекомбинантной живой пероральной вакцины из нетоксигенного штамма Vibrio cholerae 01 серотип inaba биотип E1 Tor и оценка его реактогенности и иммуногенности на модели на кроликах). Immunol Lett 68, 219-227.
Todar, K. (2002). Mechanism of bacterial pathogenicity: protein toxins. Todar's Online Textbook of Bacteriology. (Механизм бактериальной патогенности: белковые токсины. Основное руководство по бактериологии Тодара): http://textbookofbacteriology.net/proteintoxins.html.
Tzschaschel, В.D., Guzman, С.A., Timmis, K.N., and de Lorenzo, V. (1996а). An Escherichia coli hemolysin transport system-based vector for the export of polypeptides: export of Shiga-like toxin NeB subunit by Salmonella typhimurium aroA. (Система транспорта гемолизина Escherichia coli на основе вектора для экспорта полипептидов: экспорт субъединицы NeB шига-подобного токсина посредством Salmonella typhimurium aroA). Nat Biotechnol 14, 765-769.
Tzschaschel, В.D., Klee, S.R., de Lorenzo, V., Timmis, K.N., and Guzman, C.A. (1996b). Towards a vaccine candidate against Shigella dysenteriae 1: expression of the Shiga toxin B-subunit in an attenuated Shigella flexneri aroD carrier strain. (К созданию потенциальной вакцины против Shigella dysenteriae 1: экспрессия В-субъединицы шига-токсина в аттенюированном штамме-носителе Shigella flexneri aroD). Microb Pathog 21, 277-288.
van Ginkel, F.W., Jackson, R.J., Yuki, Y., and McGhee, J.R. (2000). Cutting edge: the mucosal adjuvant cholera toxin redirects vaccine proteins into olfactory tissues. (Дающее преимущество свойство - адъювантный холерный токсин для слизистых перенаправляет вакцинные белки в обонятельные ткани). J Immunol 165, 4778-4782.
Vertiev, Y.V., Zdanovsky, A.G., Shevelev, А.В., Bonnskaya, S. A., Gening, E.L., Martin, Т., Ivanov, P.A., and Yankovsky, N.K. (2001). Recombinant Listeria strains producing the nontoxic L-chain of botulinum neurotoxin A in a soluble form. (Рекомбинантные штаммы Listeria, продуцирующие нетоксичную L-цепь ботулинического нейротоксина А в растворимой форме). Res Microbiol 152, 563-567.
Walker, M.J., Rohde, M., Timmis, K.N., and Guzman, C.A. (1992). Specific lung mucosal and systemic immune responses after oral immunization of mice with Salmonella typhimurium aroA, Salmonella typhi Ty21a, and invasive Escherichia coli expressing recombinant pertussis toxin S1 subunit. (Специфические легочные ответы слизистых и системные иммунные ответы после пероральной иммунизации мышей Salmonella typhimurium aroA, Salmonella typhi Ty21 а и инвазионной Escherichia coli, экспрессирующей рекомбинантную S1-субъединицу токсина коклюша). Infect Immun 60, 4260-4268.
Ward, S.J., Douce, G., Figueiredo, D., Dougan, G., and Wren, B.W. (1999). Immunogenicity of a Salmonella typhimurium aroA aroD vaccine expressing a nontoxic domain of Clostridium difficile toxin А. (Иммуногенность вакцины Salmonella typhimurium aroA aroD, экспрессирующей нетоксичный домен токсина A Clostridium difficile). Infect Immun 67, 2145-2152.
Wiener, M.С. (2000). Bacterial export takes its Tol. (Бактериальный экспорт использует свой Tol). Structure 8, FM 71-175.
Wu, S., Beier, M., Sztein, M.В., Galen, J., Pickett, Т., Holder, A.A., Gomez-Duarte, O.G., and Levine, M.M. (2000). Construction and immunogenicity in mice of attenuated Salmonella typhi expressing Plasmodium falciparum merozoite surface protein 1 (MSP-1) fused to tetanus toxin fragment (Конструирование и иммуногенность у мышей аттенюированной Salmonella typhi, экспрессирующей поверхностный белок 1 мерозоитов Plasmodium falciparum (MSP-1), слитый с фрагментом С столбнячного токсина). J Biotechnol 83, 125-135.
Zheng, J.P., Zhang, Z.S., Li, S.Q., Liu, X.X., Yuan, S. L1 Wang, P., Zhan, D.W., Wang, L C, and Huang, C.F. (2005). Construction of a novel Shigella live-vector strain co-expressing CS3 and LTB/STm of enterotoxigenic E. coli. (Конструирование нового живого векторного штамма Shigella, соэкспрессирующего CS3 и LTB/STm энтеротоксигенной E. coli). World J Gastroenterol 11, 3411-3418.
Zhu, C, Ruiz-Perez, F., Yang, Z., Мао, Y., Hackethal, V.L, Greco, K.М., Choy, W., Davis, K., Butter-ton, J.R., and Boedeker, E.C. (2006). Delivery of heterologous protein antigens via hemolysin or autotransporter systems by an attenuated ler mutant of rabbit enteropathogenic Escherichia coli. (Доставка гетерологичных белковых антигенов посредством систем гемолизина или аутотранспортеров с помощью ler-мутанта или кроличьей энтеропатогенной Escherichia coli). Vaccine 24, 3821-3831.
| название | год | авторы | номер документа |
|---|---|---|---|
| МИКРООРГАНИЗМЫ В КАЧЕСТВЕ НОСИТЕЛЕЙ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, КОДИРУЮЩИХ КЛЕТОЧНЫЕ АНТИГЕНЫ, ДЛЯ ЛЕЧЕНИЯ ОПУХОЛЕЙ | 2003 |
|
RU2319741C2 |
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |
| Рекомбинантный белок для иммунизации против холеры | 2019 |
|
RU2723705C1 |
| Рекомбинантный штамм Mycobacterium bovis rBCG-1 на основе вакцинного штамма Mycobacterium bovis BCG strain Russia 368 | 2022 |
|
RU2807732C1 |
| СЛИТЫЕ БЕЛКИ КАРЦИНОЭМБРИОНАЛЬНОГО АНТИГЕНА | 2005 |
|
RU2380375C2 |
| СПОСОБ ИНДУЦИРОВАНИЯ У МЛЕКОПИТАЮЩИХ ЗАЩИТНОГО ИММУННОГО ОТВЕТА НА ИНФЕКЦИЮ HELICOBACTER, ВАКЦИНА ДЛЯ ИНДУЦИРОВАНИЯ ЗАЩИТНОГО ИММУННОГО ОТВЕТА НА ИНФЕКЦИЮ HELICOBACTER У МЛЕКОПИТАЮЩИХ, СПОСОБ ИНДУЦИРОВАНИЯ ПАССИВНОЙ ЗАЩИТЫ У МЛЕКОПИТАЮЩИХ ПРОТИВ ИНФЕКЦИИ HELICOBACTER И СПОСОБ ОЦЕНКИ ЭНДОГЕННОГО ИММУННОГО ОТВЕТА У МЛЕКОПИТАЮЩИХ, ИНФИЦИРОВАННЫХ HELICOBACTER | 1993 |
|
RU2125891C1 |
| ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА С УЛУЧШЕННОЙ ЭФФЕКТИВНОСТЬЮ | 2004 |
|
RU2342400C2 |
| МУТАНТ ЭНТЕРОТОКСИНА, ЭФФЕКТИВНЫЙ В КАЧЕСТВЕ НЕТОКСИЧНОГО ОРАЛЬНОГО СТИМУЛЯТОРА | 1995 |
|
RU2160606C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ И СПОСОБ РАЗРАБОТКИ ВАКЦИНЫ, ОСНОВАННОЙ НА СЛИТОМ БЕЛКЕ | 2004 |
|
RU2407749C2 |
| ПРОТИВОЧУМНАЯ ВАКЦИНА | 1996 |
|
RU2197988C2 |


























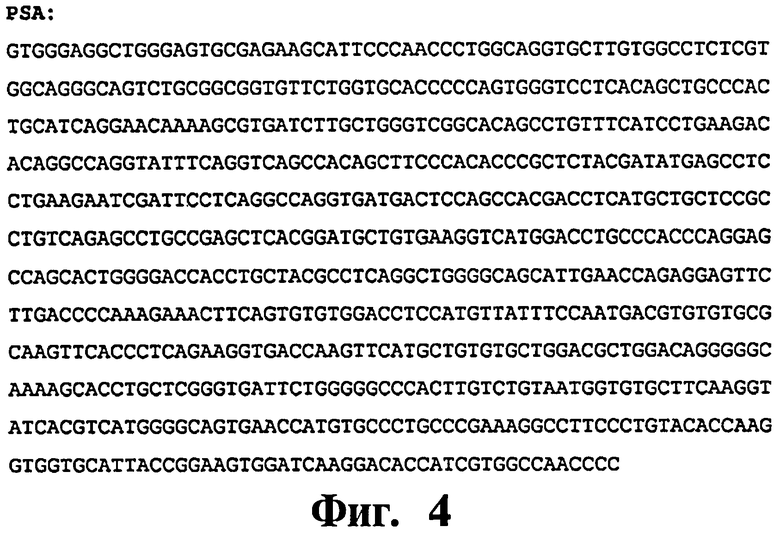






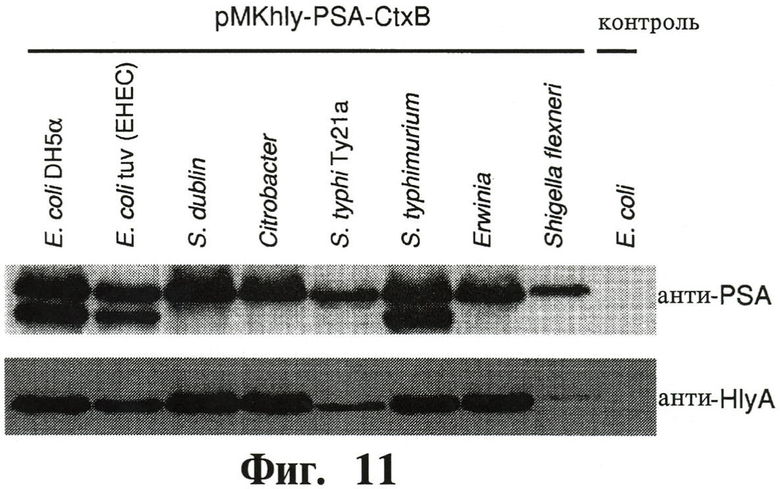
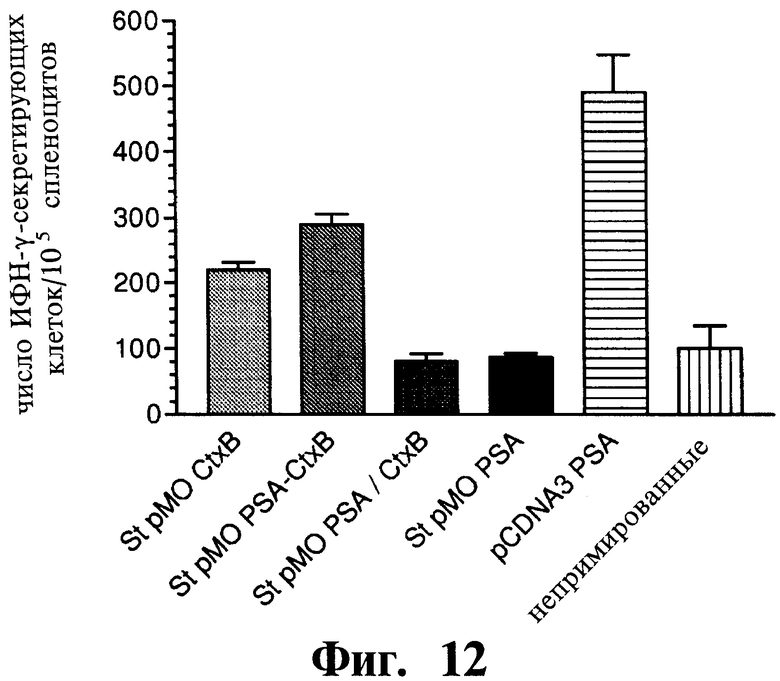
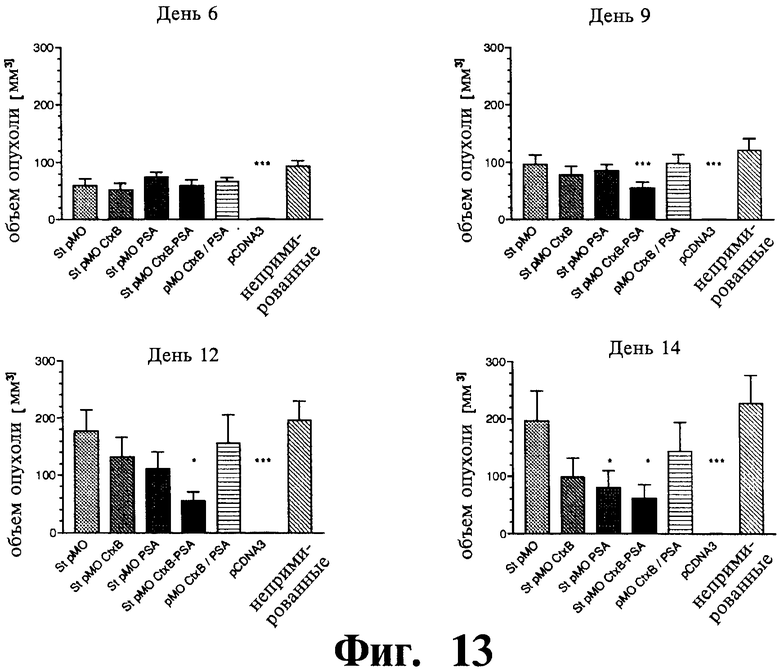
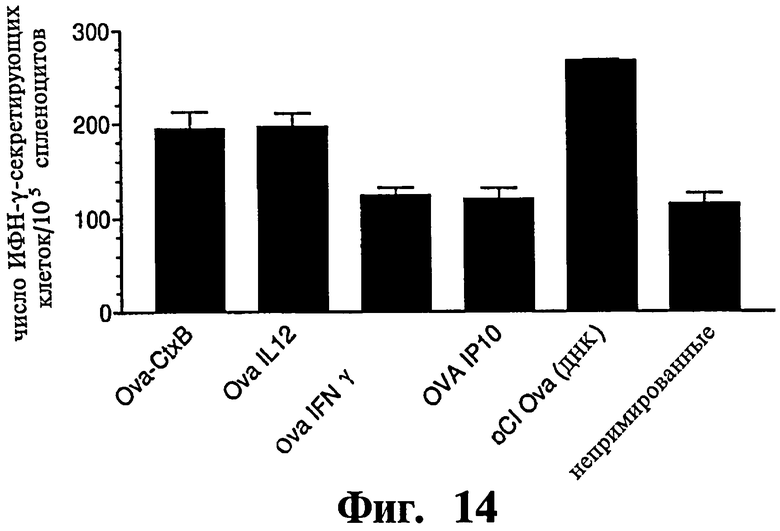
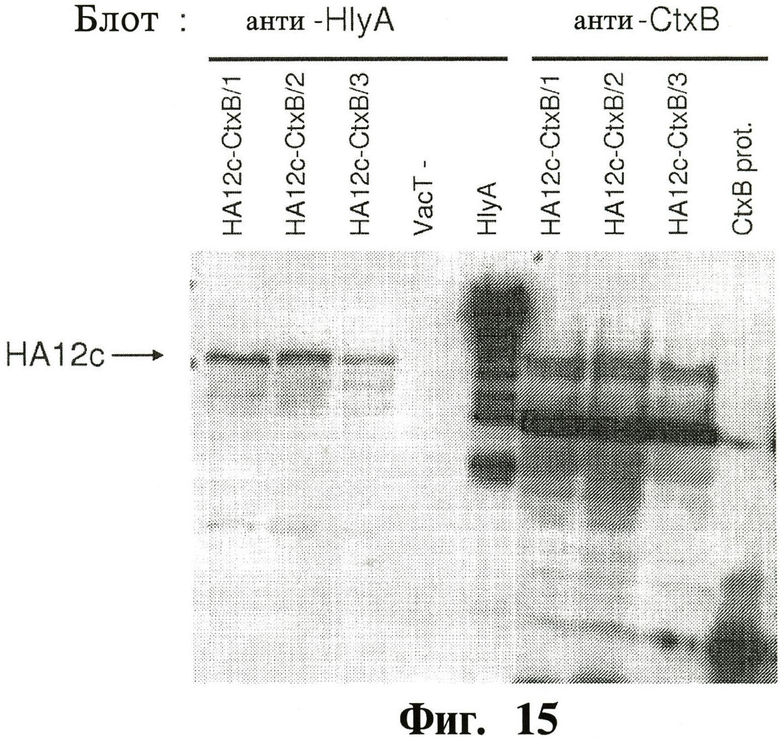

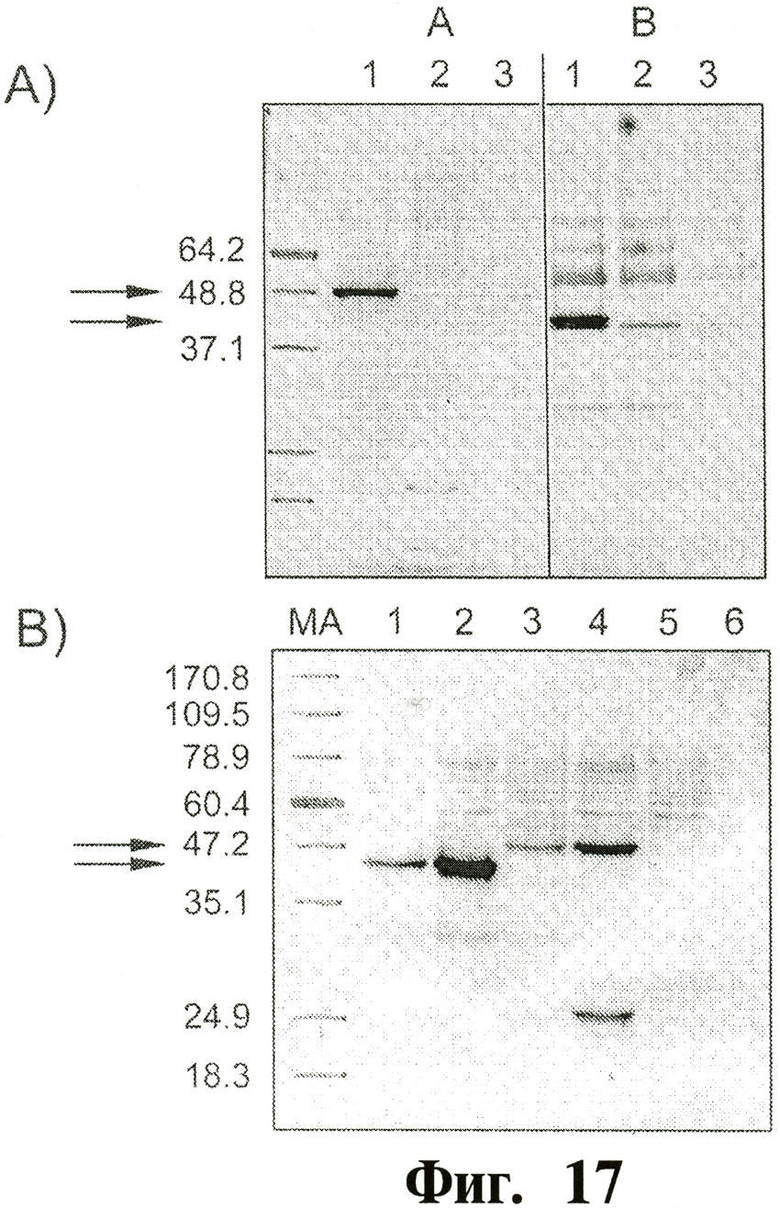





Изобретение относится к микроорганизму-носителю нуклеотидных последовательностей, кодирующих антигены и белковые токсины, включающему первый компонент, представленный по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере один полный или неполный антиген по меньшей мере одного белка дикого типа или мутантного белка, второй компонент, представленный по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере один белковый токсин и/или по меньшей мере одну субъединицу белкового токсина, третий компонент, состоящий по меньшей мере из одного первого подкомпонента, представленного по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере одну систему транспорта, которая облегчает экспрессию продуктов экспрессии первого и второго компонентов на наружной поверхности микроорганизма и/или облегчает секрецию продуктов экспрессии первого и второго компонентов, и/или кодирующей по меньшей мере одну сигнальную последовательность, которая облегчает секрецию продуктов экспрессии первого и второго компонентов, и/или, необязательно, из второго подкомпонента, представленного по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере один белок для лизиса микроорганизма в цитозоле клеток млекопитающих и для внутриклеточного высвобождения плазмид или векторов экспрессии, которые содержатся в лизированном микроорганизме, и четвертый компонент, представленный по меньшей мере одной нуклеотидной последовательностью для по меньшей мере одной последовательности активации для экспрессии по меньшей мере одного из первого, второго и третьего компонентов, где указанная последовательность активации может быть активирована в микроорганизме и/или является специфической для клетки ткани, специфической для опухолевой клетки, макрофаг-специфической, дендрит-специфической, лимфоцит-специфической, функция-специфической или неспецифической в отношении клетки, причем любой первый, второй, третий или четвертый компоненты, представленные в микрорганизме более одного раза, являются независимо друг от друга идентичными или отличающимися, при этом первый и второй компонент отличны друг от друга. Кроме того, описаны лекарственное средство для стимулирования иммунного ответа и фармацевтическая композиция на основе указанного микроорганизма, способы получения указанного микроорганизма, соответствующие экспрессионная плазмида и вектор экспрессии для получения указанного микроорганизма. Изобретение позволяет получать новые противоопухолевые вакцины, которые индуцируют сильный системный клеточный ответ иммунной системы, что повышает эффективность противоопухолевой терапии. 9 н. и 12 з.п. ф-лы, 18 ил., 8 пр.
1. Микроорганизм-носитель нуклеотидных последовательностей, кодирующих антигены и белковые токсины, включающий первый компонент, представленный по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере один полный или неполный антиген из по меньшей мере одного белка дикого типа или мутантного белка, второй компонент, представленный по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере один белковый токсин и/или по меньшей мере одну субъединицу белкового токсина, третий компонент, состоящий по меньшей мере из одного первого подкомпонента, представленного по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере одну систему транспорта, облегчающую экспрессию продуктов экспрессии первого и второго вышеупомянутых компонентов на наружной поверхности микроорганизма и/или облегчающую секрецию продуктов экспрессии первого и второго компонентов, и/или кодирующей по меньшей мере одну сигнальную последовательность, которая облегчает секрецию продуктов экспрессии первого и второго компонентов, и/или, необязательно, из второго подкомпонента, представленного по меньшей мере одной нуклеотидной последовательностью, кодирующей по меньшей мере один белок для лизиса микроорганизма в цитозоле клеток млекопитающих и для внутриклеточного высвобождения плазмид или векторов экспрессии, которые содержатся в лизированном микроорганизме, и четвертый компонент, представленный по меньшей мере одной нуклеотидной последовательностью для по меньшей мере одной последовательности активации для экспрессии по меньшей мере одного первого, второго или третьего компонентов, при этом указанная последовательность активации активируется в микроорганизме и/или является специфической для клетки ткани, специфической для опухолевой клетки, макрофаг-специфической, дендрит-специфической, лимфоцит-специфической, функция-специфической или неспецифической в отношении клетки, причем любой первый, второй, третий или четвертый компоненты, представленные в указанном микроорганизме более одного раза, являются независимо друг от друга идентичными или отличающимися, при этом первый и второй компоненты отличны друг от друга.
2. Микроорганизм по п.1, который выбран из группы, включающей бактерии, грамположительной бактерии, грамотрицательной бактерии и эукариотической клетки, а именно, Escherichia spp., Escherichia coli, Salmonella spp., Salmonella typhi, Salmonella typhimurium, Yersinia spp., Yersinia enterocolitica, Vibrio spp., Vibrio cholerae, Listeria spp., Listeria monocytogenes, Shigella spp., Shigella flexneri, Yersinia spp.H Pseudomonas spp., при этом вирулентность микроорганизма может быть аттенюирована.
3. Микроорганизм по п.1, который выбран из группы, включающей бактерии, грамположительной бактерии, грамотрицательной бактерии и эукариотической клетки, а именно, Escherichia spp., Escherichia coli, Salmonella spp., Salmonella typhi, Salmonella typhimurium, Yersinia spp., Yersinia enterocolitica, Vibrio spp., Listeria spp., Listeria monocytogenes, Shigella spp., Shigella flexneri, Yersinia spp.H Pseudomonas spp., при этом вирулентность микроорганизма может быть аттенюирована.
4. Микроорганизм по п.1, в котором по меньшей мере один полный или неполный антиген по меньшей мере одного белка дикого типа или мутантного белка, соответствующий первому компоненту, выбран из группы, включающей следующие белки дикого типа и их известные мутантные формы: рецептор, внеклеточная, трансмембранная или внутриклеточная часть рецептора, молекула адгезии, внеклеточная, трансмембранная или внутриклеточная часть молекулы адгезии, белок сигнальной трансдукции, белок клеточного цикла, фактор транскрипции, белок дифференцировки, эмбриональный белок, вирусный белок, аллерген, белок микробного патогена, белок эукариотического патогена, белок антигена рака яичка, белок опухолевого антигена и/или специфический белок клетки ткани, где клетка ткани выбрана из группы, включающей клетку щитовидной железы, молочной железы, слюнной железы, лимфатического узла, слизистой оболочки желудка, почки, яичника, простаты, шейки матки, серозной оболочки мочевого пузыря и невуса.
5. Микроорганизм по п.4, в котором по меньшей мере один полный или неполный антиген по меньшей мере одного белка дикого типа или мутантного белка, соответствующий первому компоненту, выбран из группы, включающей следующие белки дикого типа и их известные мутантные формы: Her-2/neu, андрогенный рецептор, эстрогенный рецептор, midkine-рецептор, рецептор EGF, рецептор ERBB2, ERBB4, рецептор TRAIL, FAS, рецептор TNFα, рецептор TGF-β, лактоферриновый рецептор, основной миелин, α-лактальбумин, GFAP, фибриллярный кислый белок, тирозиназа, EGR-1, MUC1, c-Raf (Raf-1), A-Raf, B-Raf, B-RafV599E, B-RafV600E, B-Raf KD, домен киназы B-RafV600E, B-Raf V600E KD, домен киназы KD B-RafV600E, домен киназы B-Raf, домен киназы KD B-Raf, N-Ras, K-Ras, H-Ras, Bcl-2, Bcl-X, Bcl-W, Bfl-1, Brag-1, Mcl-1, Al, Bax, BAD, Bak, Bcl-Xs, Bid, Bik, Hrk, Bcr/abl, Myb, C-Met, IAP1, IAO2, XIAP, ML-IAP LIVIN, сурвивин, APAF-1, циклин D(1-3), циклин E, циклин А, циклин В, циклин H, Cdk-1, Cdk-2, Cdk-4, Cdk-6, Cdk-7, Cdc25C, p16, p15, p21, p27, p18, pRb, p107, p130, E2F(1-5), GAAD45, MDM2, PCNA, ARF, PTEN, APC, BRCA, Akt, PI3K, mTOR, p53 и гомологи, C-Myc, NFkB, c-Jun, ATF-2, Sp1, простата-специфический антиген (PSA), карциноэмбриональный антиген, α-фетопротеин, PAP; PSMA; STEAP; MAGE, MAGE-1, MAGE-3, NY-ESO-1, PSCA, MART, GplOO, тирозиназа, GRP, TCF-4, вирусные антигены вирусов ВИЧ, HPV, HCV, HPV, EBV, CMV, HSV, вируса гриппа, вируса гриппа типа А, вируса гриппа типа А (H5N1) и (H3N2), вируса гриппа типа В, вируса гриппа типа С, гемагглютинины, гемагглютинин H1, гемагглютинин Н5, гемагглютинин Н7, гемагглютинин НА1 (предпочтительно из вируса гриппа А (A/Thailand/1(KAN-1)2004(H5N1)),гемагглютинин НА 12 (предпочтительно из вируса гриппа A (A/Thailand/1(KAN-1)2004(H5N1)), гемагглютинин НА12С (предпочтительно из вируса гриппа A (A/Thailand/1 (KAN-1)2004(H5N-1)), нейрамидазу, р60, LLO, уреазу, CSP, калфлагин и/или СРВ, или в котором по меньшей мере один полный или неполный антиген по меньшей мере одного белка дикого типа или мутантного белка, соответствующий первому компоненту, выбран из группы киназ, включающей следующие белки дикого типа и их известные мутантные формы: (регистрационные номера приведены в скобках): AAK1 (NM 014911), AATK (NM 004920), ABL1 (NM 005157), ABL2 (NM 005158), ACK1 (NM 005781), ACVR1 (NM 001105), ACVR1B (NM 020328), ACVR2 (NM 001616), ACVR2B (NM 001106), ACVRL1 (NM 000020), ADCK1 (NM 020421), ADCK2 (NM 052853), ADCK4 (NM 024876), ADCK5 (NM 174922), ADRBK1 (NM 001619), ADRBK2 (NM 005160), AKT1 (NM 005163), AKT2 (NM 001626), AKT3 (NM 005465), ALK (NM 004304), ALK7 (NM 145259), ALS2CR2 (NM 018571), ALS2CR7 (NM 139158), AMHR2 (NM 020547), ANKK1 (NM 178510), ANKRD3 (NM 020639), APEG1 (NM 005876), ARAF (NM 001654), ARK5 (NM 014840), ATM (NM 000051), ATR (NM 001184), AURKA (NM 003600), AURKB (NM 004217), AURKC (NM 003160), AXL (NM 001699), BCKDK (NM 005881), BCR (NM 004327), BIKE (NM 017593), BLK (NM 001715), BMPR1 A (NM 004329), BMPR1 В (NM 001203), BMPR2 (NM 001204), BMX (NM 001721), BRAF (NM 004333), BRD2 (NM 005104), BRD3 (NM 007371), BRD4 (NM 014299), BRDT (NM 001726), BRSK1 (NM 032430), BRSK2 (NM 003957), BTK (NM 000061), BUB1 (NM 004336), BUB1 В (NM 001211), CABC1 (NM 020247), CAMK1 (NM 003656), CaMK1b (NM 198452), CAMK1D (NM 020397), CAMK1G (NM 020439), CAMK2A (NM 015981), CAMK2B (NM 001220), CAMK2D (NM 001221), CAMK2G (NM 001222), CAMK4 (NM 001744), CAMKK1 (NM 032294), CAMKK2 (NM 006549), CASK (NM 003688), CCRK (NM 012119), CDC2 (NM 001786), CDC2L1 (NM 001787), CDC2L5 (NM 003718), CDC42BPA (NM 014826), CDC42BPB (NM 006035), CDC7L1 (NM 003503), CDK10 (NM 003674), CDK11 (NM 015076), CDK2 (NM 001798), CDK3 (NM 001258), CDK4 (NM 000075), CDK5 (NM 004935), CDK6 (NM 001259), CDK7 (NM 001799), CDK8 (NM 001260), CDK9 (NM 001261), CDKL1 (NM 004196), CDKL2 (NM 003948), CDKL3 (NM 016508), CDKL4 (NM 001009565), CDKL5 (NM 003159), CHEK1 (NM 001274), CHUK (NM 001278), CIT (NM 007174), CLK1 (NM 004071), CLK2 (NM 003993), CLK3 (NM 003992), CLK4 (NM 020666), CRK7 (NM 016507), CSF1R (NM 005211), CSK (NM 004383), CSNK1 A1 (NM 001892), CSNK1 D(NM 001893), CSNK1E (NM 001894), CSNK1 G1 (NM 022048), CSNK1 G2 (NM 001319), CSNK1 G3 (NM 004384), CSNK2A1 (NM 001895), CSNK2A2 (NM 001896), DAPK1 (NM 004938), DAPK2 (NM 014326), DAPK3 (NM 001348), DCAMKL1 (NM 004734), DCAMKL2 (NM 152619), DCAMKL3 (XM 047355), DDR1 (NM 013993), DDR2(NM 006182), DMPK (NM 004409), DMPK2 (NM 017525.1), DYRK1 A (NM 001396), DYRK1 В (NM 006484), DYRK2 (NM 006482), DYRK3 (NM 003582), DYRK4 (NM 003845), EEF2K (NM 013302), EGFR (NM 005228), EIF2AK3 (NM 004836), EIF2AK4 (NM_001013703), EPHA1 (NM 005232), EPHA10 (NM 001004338), EPHA2 (NM 004431), EPHA3 (NM 005233), EPHA4 (NM 004438), EPHA5 (NM 004439), EPHA6 (XM 114973), EPHA7 (NM 004440), EPHA8 (NM 020526), EPHB1 (NM 004441), EPHB2 (NM 017449), EPHB3 (NM 004443), EPHB4 (NM 004444), EPHB6 (NM 004445), ERBB2 (NM 004448), ERBB3 (NM 001982), ERBB4 (NM 005235), ERK8 (NM 139021), ERN1 (NM 001433), ERN2 (NM 033266), FASTK (NM 025096), FER (NM 005246), FES (NM 002005), FGFR1 (NM 000604), FGFR2 (NM 022970), FGFR3 (NM 000142), FGFR4 (NM 022963), FGR (NM 005248), FLJ23074 (NM 025052), FLJ23119 (NM 024652), FLJ23356 (NM 032237), FLT1 (NM 002019), FLT3 (NM 0041 19), FLT4 (NM 002020), FRAP1 (NM 004958), FRK (NM 002031), FYN (NM 002037), GAK (NM 005255), GPRK5 (NM 005308), GPRK6 (NM 002082), GPRK7 (NM 139209), GRK4 (NM 005307), GSG2 (NM 031965), GSK3A (NM 019884), GSK3B (NM 002093), GUCY2C (NM 004963), GUCY2D (NM 000180), GUCY2F (NM 001522), H11 (NM 014365), HAK (NM 052947), HCK (NM 0021 10), HIPK1 (NM 152696), HIPK2 (NM 022740), HIPK3 (NM 005734), HIPK4 (NM 144685), HRI (NM 014413), HUNK (NM 014586), ICK (NM 016513), IGF1 R (NM 000875), IKBKB (NM 001556), IKBKE (NM 014002), ILK (NM 004517), INSR (NM 000208), INSRR (NM 014215), IRAKI (NM 001569), IRAK2 (NM 001570), IRAK3 (NM 007199), IRAK4 (NM 016123), ITK (NM 005546), JAK1 (NM 002227), JAK2 (NM 004972), JAK3 (NM 000215), KDR (NM 002253), KIS (NM 144624), KIT (NM 000222), KSR (XM 290793), KSR2 (NM 173598), LAK (NM 025144), LATS1 (NM 004690), LATS2 (NM 014572), LCK (NM 005356), LIMK1 (NM 016735), LIMK2 (NM 005569), LMR3 (XM 055866), LMTK2 (NM 014916), LOC149420 (NM 152835), LOC51086 (NM 015978), LRRK2 (XM 058513), LTK (NM 002344), LYN (NM 002350), MAK (NM 005906), MAP2K1 (NM 002755), MAP2K2 (NM 030662), MAP2K3 (NM 002756), MAP2K4 (NM 003010), MAP2K5 (NM 002757), MAP2K6 (NM 002758), MAP2K7 (NM 005043), MAP3K1 (XM 042066), MAP3K10 (NM 002446), MAP3K11 (NM 002419), MAP3K12 (NM 006301), MAP3K13 (NM 004721), MAP3K14 (NM 003954), MAP3K2 (NM 006609), MAP3K3 (NM 002401), MAP3K4 (NM 005922), MAP3K5 (NM 005923), MAP3K6 (NM 004672), MAP3K7 (NM 003188), MAP3K8 (NM 005204), MAP3K9 (NM 033141), MAP4K1 (NM 007181), MAP4K2 (NM 004579), MAP4K3 (NM 003618), МАР4К4 (NM 145686), MAP4K5 (NM 006575), MAPK1 (NM 002745), MAPK10 (NM 002753), MAPK11 (NM 002751), MAPK12 (NM 002969), МАРК13 (NM 002754), MAPK14 (NM 001315), MAPK3 (NM 002746), MAPK4 (NM 002747), MAPK6 (NM 002748), MAPK7 (NM 002749), MAPK8 (NM 002750), MAPK9 (NM 002752), MAPKAPK2 (NM 032960), MAPKAPK3 (NM 004635), MAPKAPK5 (NM 003668), MARK (NM 018650), MARK2 (NM 017490), MARK3 (NM 002376), MARK4 (NM 031417), MAST1 (NM 014975), MAST205 (NM 015112), MAST3 (XM 038150), MAST4 (XM 291141), MASTL (NM 032844), MATK (NM 139355), MELK (NM 014791), MERTK (NM 006343), MET (NM 000245), MGC33182 (NM 145203), MGC42105 (NM 153361), MGC43306 (C9orf96), MGC8407 (NM 024046), MIDORI (NM 020778), MINK (NM 015716), MKNK1 (NM 003684), MKNK2 (NM 017572), MLCK (NM 182493), MLK4 (NM 032435), MLKL (NM 152649), MOS (NM 005372), MST1 R (NM 002447), MST4 (NM 016542), MUSK (NM 005592), MYLK (NM 053025), MYLK2 (NM 0331 18), MYO3A (NM 017433), MYO3B (NM 138995), NEK1 (NM 012224), NEK10 (NM 152534), NEK11 (NM 024800), NEK2 (NM 002497), NEK3 (NM 002498), NEK4 (NM 003157), NEK5 (MGC75495), NEK6 (NM 014397), NEK7 (NM 133494), NEK8 (NM 178170), NEK9 (NM 0331 16), NLK (NM 016231), NPR1 (NM 000906), NPR2 (NM 003995), NRBP (NM 013392), NRBP2 (NM 178564), NRK (NM 198465), NTRK1 (NM 002529), NTRK2 (NM 006180), NTRK3 (NM 002530), OBSCN (NM 052843), OSR1 (NM 005109), PACE-1 (NM 020423), PAK1 (NM 002576), PAK2 (NM 002577), PAK3 (NM 002578), PAK4 (NM 005884), PAK6 (NM 020168), PAK7 (NM 020341), PASK (NM 015148), PCTK1 (NM 006201), PCTK2 (NM 002595), PCTK3 (NM 212503), PDGFRA (NM 006206), PDGFRB (NM 002609), PDK1 (NM 002610), PDK2 (NM 00261 1), PDK3 (NM 005391), PDK4 (NM 002612), PDPK1 (NM 002613), PFTK1 (NM 012395), PHKG1 (NM 006213), PHKG2 (NM 000294), PIK3R4 (NM 014602), PIM1 (NM 002648), PIM2 (NM 006875), PIM3 (NM 001001852), PINK1 (NM 032409), PKE (NM 173575), PKMYT1 (NM 004203), pknβ (NM 013355), PLK (NM 005030), PLK3 (NM 004073), PRKAA1 (NM 006251), PRKAA2 (NM 006252), PRKACA (NM 002730), PRKACB (NM 002731), PRKACG (NM 002732), PRKCA (NM 002737), PRKCB1 (NM 002738), PRKCD (NM 006254), PRKCE (NM 005400), PRKCG (NM 002739), PRKCH (NM 006255), PRKCI (NM 002740), PRKCL1 (NM 002741), PRKCL2 (NM 006256), PRKCM (NM 002742), PRKCN (NM 005813), PRKCQ (NM 006257), PRKCZ (NM 002744), PRKD2 (NM 016457), PRKDC (NM 006904), PRKG1 (NM 006258), PRKG2 (NM 006259), PRKR (NM 002759), PRKWNK1 (NM 018979), PRKWNK2 (NM 006648), PRKWNK3 (NM 020922), PRKWNK4 (NM 032387), PRKX (NM 005044), PRKY (NM 002760), PRPF4B (NM 003913), PSKH1 (NM 006742), PSKH2 (NM 033126), PTK2 (NM 005607), PTK2B (NM 004103), PTK6 (NM 005975), PTK7 (NM 002821), PTK9 (NM 002822), PTK9L (NM 007284), PXK (NM 017771), QSK (NM 025164), RAD53 (NM 007194), RAF1 (NM 002880), RAGE (NM 014226), RET (NM 020975), RHOK (NM 002929), RIOK1 (NM 031480), RIOK2 (NM 018343), RIPK1 (NM 003804), RIPK2 (NM 003821), RIPK3 (NM 006871), RIPK5 (NM 015375), RNASEL (NM 021133), ROCK1 (NM 005406), ROCK2 (NM 004850), ROR1 (NM 005012), ROR2 (NM 004560), ROS1 (NM 002944), RPS6KA1 (NM 002953), RPS6KA2 (NM 021135), RPS6KA3 (NM 004586), RPS6KA4 (NM 003942), RPS6KA5 (NM 004755), RPS6KA6 (NM 014496), RPS6KB1 (NM 003161), RPS6KB2 (NM 003952), RPS6KC1 (NM 012424), RPS6KL1 (NM 031464), RYK (NM 002958), SBK (XM 370948), SCYL1 (NM 020680), SCYL2 (NM 017988), SGK (NM 005627), SgK069 (SU SgK069), SgK085 (XM 373109), SgK110 (SU_SgKHO), SGK2 (NM 016276), SgK223 (XM 291277), SgK269 (XM 370878), SgK424 (CGP SgK424), SgK493 (SU_SgK493), SgK494 (NM 144610), SgK495 (NM 032017), SGKL (NM 013257), SK681 (NM 001001671), SLK (NM 014720), SMG1 (NM 015092), SNARK (NM 030952), SNF1 LK (NM 173354), SNF1 LK2 (NM 015191), SNK (NM 006622), SNRK (NM 017719), SRC (NM 005417), SRMS (NM 080823), SRPK1 (NM 003137), SRPK2 (NM 003138), SSTK (NM 032037), STK10 (NM 005990), STK11 (NM 000455), STK16 (NM 003691), STK17A (NM 004760), STK17B (NM 004226), STK18 (NM 014264), STK19 (NM 032454), STK22B (NM 053006), STK22C (NM 052841), STK22D (NM 032028), STK23 (NM 014370), STK24 (NM 003576), STK25 (NM 006374), STK3 (NM 006281), STK31 (NM 031414), STK32B (NM 018401), STK33 (NM 030906), STK35 (NM 080836), STK36 (NM 015690), STK38 (NM 007271), STK38L (NM 015000), STK39 (NM 013233), STK4 (NM 006282), STLK5 (NM 001003787), STYK1 (NM 018423), SUDD (NM 003831), SYK (NM 003177), TAF1 (NM 138923), TAF1 L (NM 153809), TAO1 (NM 004783), TAOK1 (NM 020791), TAOK3 (NM 016281), TBCK (NM 0331 15), TBK1 (NM 013254), TEC (NM 003215), ТЕК (NM 000459), TESK1 (NM 006285), TESK2 (NM 007170), TEX 14 (NM 031272), TGFBR1 (NM 004612), TGFBR2 (NM 003242), TIE (NM 005424), TIF1 (NM 003852), TLK1 (NM 012290), TLK2 (NM 006852), TNIK (NM 015028), TNK1 (NM 003985), TOPK (NM 018492), TP53RK (NM 033550), TRAD (NM 007064), TRIB1 (NM 025195), TRIB2 (NM 021643), TRIB3 (NM 021 158), TRIM28 (NM 005762), TRIM33 (NM 015906), TRIO (NM 0071 18), TRPM6 (NM 017662), TRPM7 (NM 017672), TRRAP (NM 003496), TSSK4 (NM 174944), TTBK1 (NM 032538), TTBK2 (NM 173500), TTK (NM 003318), TTN (NM 003319), TXK (NM 003328), TYK2 (NM 003331), TYR03 (NM 006293), ULK1 (NM 003565), ULK2 (NM 014683), ULK3 (NM 015518), ULK4 (NM 017886), VRK1 (NM 003384), VRK2 (NM 006296), VRK3 (NM 016440), WEE1 (NM 003390), Wee1 В (NM 173677), YANK1 (NM 145001), YES1 (NM 005433), ZAK (NM 016653) и/или ZAP70 (NM 001079).
6. Микроорганизм по п.1, в котором второй компонент выбран из группы, включающей бактериальный токсин, энтеротоксин, экзотоксин, токсин типа I, токсин типа II, токсин типа III, токсин типа IV, токсин типа V, токсин RTX, токсин АВ, токсин А-В, токсин А/В, токсин А+В, токсин А-5 В и токсин АВ5.
7. Микроорганизм по п.6, в котором второй компонент выбран из группы, включающей токсин аденилатциклазы, токсин сибирской язвы, токсин сибирской язвы (EF), токсин сибирской язвы (LF), ботулинический токсин, холерный токсин (СТ, Ctx), субъединицу холерного токсина В (СТВ, CtxB), дифтерийный токсин (DT, Dtx), токсин Е. coli LT, термолабильный энтеротоксин Е. coli (LT), субъединицу В термолабильного энтеротоксина Е. coli (LTB), токсин Е. coli ST, термостабильный энтеротоксин Е. coli (ST), эритрогенный токсин, токсин эксфолиатина, экзотоксин А, энтеротоксин Perfringens, токсин коклюша (РТ, Ptx), шига-токсин (ST, Stx), субъединицу В шига-токсина (STB, StxB), шига-подобный токсин, энтеротоксины Staphylococcus, столбнячный токсин (ТТ), токсин синдрома токсического шока (TSST-1), веро-токсин (VT), токсин А (ТА) и токсин В (ТВ) Clostridium difficile, летальный токсин (LT) и геморрагический токсин (НТ) из Clostridium sordellii и α-токсин (AT) из Clostridium novyi.
8. Микроорганизм по п.1, в котором первый и второй компоненты связаны друг с другом с облегчением экспрессии и/или секреции слитого белка, кодируемого обоими вышеуказанными компонентами.
9. Микроорганизм по п.8, в котором слитый белок выбран из группы, включающей CtxB-PSA, CtxB-B-RafV600E KD, CtxB-домен киназы В-RafV600E, CtxB-домен киназы KD B-Raf V600E, CtxB-B-Raf, CtxB-B-Raf KD, CtxB домен киназы KD B-Raf, CtxB-HAl и CtxB-HA12C.
10. Микроорганизм по п.1, в котором первый подкомпонент выбран из группы, включающей систему секреции типа I, систему секреции типа II, систему секреции типа III, систему секреции типа IV, систему секреции типа V, гемолизиновую систему транспорта (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем hly-специфического промотора), гемолизиновую систему транспорта (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем не-hly-специфического бактериального промотора), транспортный сигнал для белка S-слоя (Rsa A) Caulobacter crescentus, транспортный сигнал для белка TolC Escherichia coli, сигнал секреции Vtgss и сигналы секреции, выделенные из листериолизина, р60 и/или ActA, а второй подкомпонент выбран из группы, включающей эндолизины, литический белок грамположительных бактерий, литический белок Listeria monocytogenes, PLY551 Listeria monocytogenes и холин Listeria monocytogenes.
11. Микроорганизм по п.10, в котором первый подкомпонент представляет собой по меньшей мере одну нуклеотидную последовательность, кодирующую только одну транспортную систему, которая облегчает сопутствующую экспрессию продуктов экспрессии первого и второго компонентов на наружной поверхности микроорганизма и/или облегчает сопутствующую секрецию продуктов экспрессии первого и второго компонентов, и по меньшей мере одну нуклеотидную последовательность, кодирующую гемолизиновую транспортную систему (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем hly-специфического промотора) или гемолизиновую транспортную систему (сигнал) Escherichia coli (нуклеотидные последовательности, включающие HlyA, HlyB и HlyD под контролем не-hly-специфического бактериального промотора).
12. Микроорганизм по п.1, в котором в присутствии первого подкомпонента секретируются продукты экспрессии первого и второго компонентов.
13. Микроорганизм по п.1, в котором первый компонент выбран из группы, включающей B-RafV600E, домен киназы B-RafV600E, В-RafV600E KD, домен киназы KD B-RafV600E, B-Raf KD, домен киназы В-Raf, домен киназы KD B-Raf, простата-специфический антиген (PSA), гемагглютинин НА1 (предпочтительно из вируса гриппа A (A/Thailand/1(KAN-1)2004(H5N1)), гемагглютинин НА 12 (предпочтительно из вируса гриппа A (A/Thailand/1(KAN-1)2004(H5N1)), гемагглютинин НА12С (предпочтительно из вируса гриппа A (A/Thailand/1(KAN-1)2004(H5N1)), второй компонент выбран из группы, включающей субъединицу В холерного токсина (СТВ, CtxB), субъединицу В термолабильного энтеротоксина Е. coli (LTB), столбнячный токсин (ТТ), первый подкомпонент выбран из группы, включающей сигнал гемолизинового транспорта HlyA Escherichia coli вместе с компонентами системы секреции Hly (нуклеотидными последовательностями, включающими HlyA, HlyB и HlyD под контролем hly-специфического промотора), четвертый компонент выбран из группы, включающей эндогенный промотор локуса hly Е. coli, при этом первый и второй компоненты связаны друг с другом с облегчением экспрессии секретируемого слитого белка, кодируемого обоими вышеуказанными компонентами.
14. Фармацевтическая композиция для стимулирования иммунного ответа, включающая по меньшей мере один микроорганизм, предпочтительно в лиофилизированной форме, по любому из пп.1-13 и фармацевтически приемлемый носитель предпочтительно в виде капсулы.
15. Лекарственное средство для стимулирования иммунного ответа, включающее по меньшей мере один микроорганизм по любому из пп.1-13 или по меньшей мере одну фармацевтическую композицию по п.14.
16. Применение микроорганизма по любому из пп.1-13 для приготовления лекарственного средства для лечения и/или профилактики физиологических и/или патофизиологических состояний, выбранных из группы, включающей неконтролируемое деление клеток, злокачественные опухоли, доброкачественные опухоли, солидные опухоли, саркому, карциному, гиперпролиферативные нарушения, карциноиды, саркому Эвинга, саркому Капоши, опухоли головного мозга, опухоли, происходящие из головного мозга, и/или нервной системы, и/или мозговых оболочек, глиом, нейробластомы, рак желудка, рак почки, карциному почечных клеток, рак простаты, карциному простаты, опухоли соединительных тканей, саркому мягких тканей, опухоли поджелудочной железы, опухоли печени, опухоли головы, опухоли шеи, рак пищевода, рак щитовидной железы, остеосаркому, ретинобластому, тимому, рак яичка, рак легкого, бронхиальные карциномы, рак молочной железы, карциному молочной железы, рак кишки, колоректальные опухоли, карциному толстой кишки, карциному прямой кишки, гинекологические опухоли, опухоли яичника/овариальные опухоли, рак матки, рак шейки матки, карциному шейки матки, рак тела матки, карциному тела, эндометриальные карциномы, рак мочевого пузыря, рак полости, рак кожи, базалиому, спиналиому, меланому, внутриглазные меланомы, лейкоз, хронический лейкоз, острый лейкоз, лимфому, инфекцию, вирусную или бактериальную инфекцию, грипп, хроническое воспаление, отторжение органа и аутоиммунные заболевания.
17. Экспрессионная плазмида, включающая первый, второй третий и четвертый компоненты микроорганизма по любому из пп.1-13.
18. Способ получения микроорганизма по любому из пп.1-13, в котором получают экспрессионную плазмиду по п.17, а затем трансформируют микроорганизм данной плазмидой.
19. Вектор экспрессии, включающий первый, второй, третий и четвертый компоненты микроорганизма по любому из пп.1-13.
20. Способ получения микроорганизма по любому из пп.1-13, в котором получают вектор экспрессии по п.19, а затем трансформируют микроорганизм данным вектором экспрессии.
21. Фармацевтический набор для стимулирования иммунного ответа, включающий по меньшей мере один микроорганизм по любому из пп.1-13 или фармацевтическую композицию по п.14, или лекарственное средство по п.15 и фармакологически приемлемый буфер, предпочтительно карбонатный буфер.
| WO 9823763 А1, 04.06.1998 | |||
| СПОСОБ ВВЕДЕНИЯ ЧУЖЕРОДНЫХ ГЕНОВ В ГЕНОМЫ ГРАМОТРИЦАТЕЛЬНЫХ МИКРООРГАНИЗМОВ, ПЛАЗМИДНЫЙ ВЕКТОР РКС47М ДЛЯ ВВЕДЕНИЯ ЧУЖЕРОДНЫХ ГЕНОВ В ГЕНОМЫ ГРАМОТРИЦАТЕЛЬНЫХ МИКРООРГАНИЗМОВ, СПОСОБ КОНСТРУИРОВАНИЯ ПЛАЗМИДНОГО ВЕКТОРА РКС47М | 1995 |
|
RU2092556C1 |
| РЕКОМБИНАНТНЫЙ АТТЕНУИРОВАННЫЙ ШТАММ БАКТЕРИЙ SALMONELLA TYPHIMURIUM T10/PKHBC - ПРОДУЦЕНТ КОРОВОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В | 2002 |
|
RU2216590C1 |

