Изобретение относится к лечению желудочной инфекции у млекопитающих, включая людей. Более конкретно, оно относится к методу лечения геликобактерной инфекции, к составам вакцин и к антителам пригодным для использования при таком лечении.
Инфекция геликобактерных пироли (Г.пилори) желудочного эпителия человека имеет существенное значение при развитии гастрита и язвы желудка и может стать фактором риска развития рака желудка. Это небольшой, имеющий форму буквы S, грамм-отрицательный микроорганизм обычно получается из желудочной ткани взрослых и детей с гистологическими признаками гастрита или пептического изъявления. Свидетельство причинной связи между Г.пилори и гастродуоденальной болезнью получены из изучения людей-добровольцев, гнотобиотических свинок и свободных от бактерий грызунов, при этом были выполнены постулаты Коха путем создания гистологически подтвержденного гастрита после поглощения жизнеспособных микроорганизмов. Хотя эту болезнь трудно лечить, после искоренения лежащей в основе болезни гастрит исчезает и у пациентов с заболеванием дуоденальной язвы резко снижается повторяемость язвенных заболеваний. Peterson W. L Helicobacter pylori and peptic ulcor disease., N.Enge.J.Med. 1991, 324, 1034-1048.
Несмотря на подверженность многим антимикробным факторам in vitro, трудно достичь долгосрочность искоренения установившихся инфекций Г.пилори с антимикробными факторами in vivo Микроорганизм обнаруживается внутри слизистого покрытия, лежащего над эпителием желудка. Это то размещение, которое, казалось бы, не позволяет достичь адекватных антимикробных уровней при оральном лечении. В настоящее время большая часть специалистов рекомендует "тройную терапию", а именно соль висмута в сочетании с тетрациклином и метронидазолом в течение 2-4 недель. Однако эффективность этих или других химиотерапевтических режимов остается субоптимальной.
В настоящее время мало известно о роли мукозальной (слизистой) иммунной системы желудка. Распределение Ig - производящих клеток в нормальной желудочной полости указывает на то, что клетки IgA плазмы достигают до 80% общей популяции клеток плазмы. Кроме того, число клеток IgA плазмы, присутствующих в полости, сравнимо с другими слизистыми мембранами. Хотя ряд исследований был посвящен уровням иммуноглобулина в различных эндокринных жидкостях, нет данных относительно концентрации иммуноглобулинов в желудочных секрециях. Более того, имеется только ограниченное количество данных, говорящих о том, что пациенты, инфецированные Г.пилори, вырабатывают специфические Ig G или/и Ig A антитела в аспиаратах желудка. Таким образом, если инфекция установилась, в ее искоренении не эффективны на антитела, ни антибиотики.
Зинн и др. показали, что повторная оральная иммунизация с помощью антигенов Г.пилори и холерного токсина приводит к стимулированию резкого желудочного-кишечного Ig A анти- Г.пилори реагирования у мышей и хорьков (Infect. Immun., 1991, 59, 2359-2363). Однако, поскольку линии и корьки стойки по отношению к инфекции Г.пилори и поскольку в то время не имелось небольшой животной модели для оценки степени защиты, было неизвестно, насколько могут предохранить сформированные таким образом антитела.
Ли и др. сообщили о возможности инфицировать свободных от бактерий грызунов посредством Г.фелис и воспроизвели документально гистологический гастрит. Однако не было сообщено об оценке степени защиты. (Gastroenterology., 1990, 99, 1315-1323; Infect. Immun., 1991, 59, 785-791.).
Поэтому имеется необходимость эффективного лечения желудочной инфекции Г.пилори, особенно у людей.
Задачей настоящего изобретения является создание способа вызывания иммунной реакции у млекопитающих, обеспечивающей защитные функции от геликобактерной инфекции, а также создание вакцины для выполнения этого способа. Задачей изобретения является также создание способа наделения млекопитающего пассивной защитой от геликобактерной инфекции.
Изобретатели обнаружили, что, как это не удивительно, оральная иммунизация "хозяина" (паразитирующих организмов) антигеном геликобактера приводит к образованию антител, которые защищают от острой инфекции микроорганизма геликобактера. Образование таких защитных антител нельзя было предсказать на основе предыдущих исследований, поскольку до настоящего изобретения не существовало подходящей модели для оценки уровня защиты.
Согласно одному аспекту настоящего изобретения, имеется способ вызывания в "хозяине"-млекопитающем защитного иммунного реагирования на инфекцию геликобактером, состоящий в оральном введении в "хозяина" иммуногенного эффективного количества антигена геликобактера для вызывания желательной защитной иммунной реакции.
Согласно другому аспекту настоящего изобретения, создана вакцина, содержащая некоторое количество антигена геликобактера, эффективное для вызывания защитной реакции человека в сочетании с фармацевтически приемлемым разбавителем.
Согласно еще одному аспекту настоящего изобретения, создан способ наделения "хозяина"-млекопитающего пассивной защиты против инфекции геликобактера, состоящий в оральном введении в хозяина иммунологически эффективного количества специфического IgA антитела геликобактера для создания желательного пассивной защиты.
Согласно еще одному аспекту этого изобретения, создана клеточная цепочка #71-G5-A8.
Изобретение будет далее описано со ссылками на сопровождающие чертежи, в которых:
фиг. 1 - гистограмма титров антитела в различных погонышах каролинских и секреции свободных от бактерий мышей после оральной иммунизации лизатом Г. фелис в ассоциации с холерным токсином;
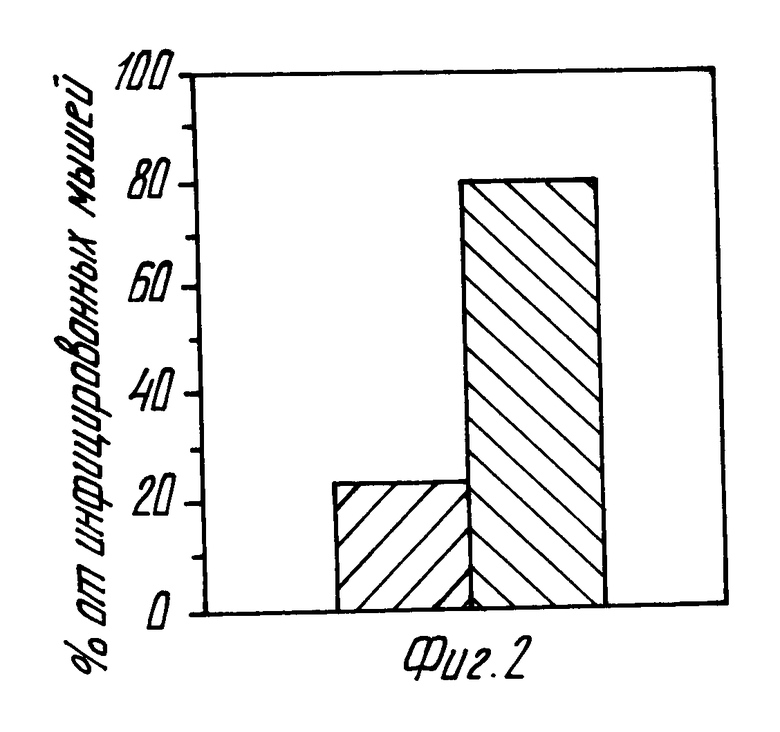
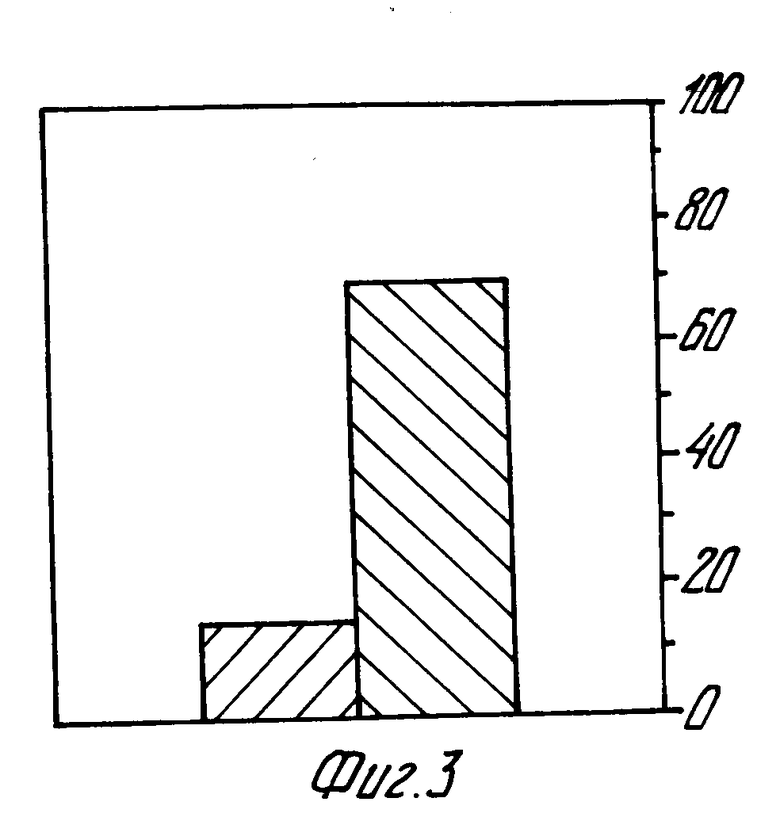
фиг. 2 и 3 - это гистограммы процентного отношения мышей, инфецированных Г. фелис после активной иммунизации (фиг. 2) и пассивной иммунизации в сравнении с контрольными данными.
Изобретатели показали, что иммунизация мышей с использованием антигена Г. фелис вызывала защитную иммунную реакцию, когда антигенные специфические антитела присутствуют в желудочных секрециях. Эффект защитной иммунной реакции состоит в том, что иммунизированные животные при воздействии патогена не инфицируются в сравнении с неиммунизированными животными, которые реально инфицируются. Изобретатели считают, что оральная иммунизация с помощью антигена Г. фелис стимулирует общую слизистую систему и, вероятно, локальные участки в слизистой оболочке желудка, что приводит к появлению Г. фелис - специфических - IgA антител в желудочных секрециях что предотвращает инфекцию Г. фелис. Поскольку Г. фелис и Г. пилори это сходные виды из одного рода (геликобактер), можно прийти к выводу, что иммунизация, например, свободной от бактерий свинки, антигеном Г. пилори плюс слизистое вспомогательное вещество, такое, как холерный токсид, будет эффективной в предотвращении Г.пилори - инфекции желудка. Поскольку обычно практикой является проведение доклинических испытаний вакцин-кандидатов для использования человека на животных моделях, считается, что методология настоящего изобретения эффективна для людей, особенно при лечении инфекции Г. пилори у людей.
Изобретатели обнаружили, что свободная от бактерий модель мыши Г.фелис может быть использована для оценки уровней защиты антитела после иммунизации антителом Г.фелис. Фиг. 1 показывает результаты, полученные в экспериментах со свободной от бактерий моделью мыши Г.фелис. Оральная иммунизация модели бактериальными антигенами в ассоциации с холерным токсином приводила к повышенным титрам сыворотки, желудочного и кишечного антитела анти- Г.фелис и к защите от стройной инфекции желудка патогенном Г.фелис. В этих экспериментах группы свободных от бактерий мышей Суисс-Уэбстрера (таконических) были орально иммунизированы 4 или 5 раз в течение одного месяца с помощью 2 - 4 мг соницированного Г. фелис - лизата плюс 10 мгм холерного токсина. Затем в мышей орально вводилось приблизительно 10 жизнеспособных бактерий Г. фелис. Затем мыши жертвовались, и собирались их кишечные и желудочные секреции, как описано в следующих рабочих примерах. Титры анти-фелис антитела определялись методом ELISA.
Черные полосы на фиг.1 представляют средине титры (+S.D.) от иммунизированных мышей, а белые полосы представляют средние титры (+S.D.) от контрольные неиммунизированных мышей. Эти результаты, графически представлены на фиг. 1, суммарно представлены в табл.1.
Из вышеперечисленных результатов можно видеть, что значительно более высокие титры антитела наблюдаются для иммунизированных мышей, чем для контрольных животных.
На фиг. 2 и 3 приведены результаты исследований по определению защиты против инфекции Г. фелис посредством проведения экспериментов по активной и пассивной иммунизации. При активной иммунизации собирались желудочные биопсии при жертвовании от мышей, инфецированных Г. фелис в экспериментах, описанных выше в связи с фиг. 1. Биопсии подсчитывались на присутствии Г. фелис быстрым тестом на мочевину или/и положительную культуры, как описано в нижеследующих рабочих примерах.
Фиг. 2 - показывает результаты суммированных данных 3-х экспериментов (17 иммунизированных животных и 18 контрольных животных). Черные полосы на столбике слева представляют инфецированных иммунизированных мышей, а полосы (столбик справа) - контрольных неиммунизированных мышей.
Как видно, из общего числа 17 иммунизированных животных только 4 стали инфецированными по сравнению с 14 из 18 контрольных животных. Другими словами, 78% иммунизированных животных были защищены от инфекции Г. фелис по сравнению с 23 % неиммунизированных животных.
То, что защита явилась непосредственным результатом антител IgA, было определено пассивной иммунизацией свободных от бактерий мышей специфическими IgA моноклонными антителами Г. фелис и сравнением результирующей защиты с той, которая проявлялась у мышей, которым не дали антитела или дали неподходящее антитело (например, специфическое IgA моноклонное антитело вирус Сендай). Результаты представлены на фиг. 3.
IgA моноклонное антитело, реактивное с Г.фелис, изолировалось и субклонировалось после процесса иммунизации, аналогично тому, что описан на фиг. 1, Асцит, содержащий Г. фелис специфическое IgA моноклонное антитело, полученное из клеточной цепочки #71-G-A, приготовленное как это описано в рабочих примерах, или специфическое IgA моноклонное антитело вирус Сендай или соль, были орально введены в свободных от бактерий мышей во время инфецирования Г.фелис и 4, 8 и 24 ч. спустя. Семь дней после инфецирования мыши были пожертвованы, и желудочные биопсии подсчитывались на Г.фелис (7 мышей получили Г.фелис специфическое моноклонное антитело и 13 мышей не получили антитела или получили специфическое моноклонное антитело вирус Сендай). Четыре полосы (столбик слева) представляют мышей, которые получили Г.фелис специфическое моноклонное антитело, полосы (столбик справа) представляют мышей, которые получили либо специфическое моноклонное антитело вирус Сендай, либо соль (без антитела).
Эти результаты показывают то, что только IgA защищает от инфекции Г.фелис желудочной слизистой оболочки.
Также обнаружено, что оральное введение антигена Г.фелис приводит к значительно повышенным уровням анти-Г.фелис IgA (антител), а также IgA антител. Для этого феномена имеется ряд возможных объяснений. Во-первых, обнаруживалось, что холерный токсин может в некоторых случаях повышать как антиген-специфическую IgA реакцию, так и антиген-специфическую IgG реакцию 22. Во-вторых, исследование движения клеток показали, что лимфоциты мезентерического узла IgG является компонентом слизистой иммунной системы и могут вызывать слизистые клетки плазмы IgG , которые наблюдались в слизистой оболочки желудка. В-третьих, по меньшей мере часть наблюдаемого желудочного IgG может быть результатом просачивания сывороточного антитела в полость клеток желудка вторично в слишком слабом виде для того, чтобы смягчить воспаление, наблюдаемое как у контрольных, так и иммунизированных животных.
Обсуждение, приведенное выше, было посвящено использование антигена Г. фелис при лечении инфекции Г.фелис. Однако желательно, чтобы настоящее изобретение не ограничивалось лечением инфекции Г.фелис.
Таким образом, в объем настоящего изобретения входит также лечение или профилактика млекопитающих, включая людей, в отношении инфекции Г.пилори, когда пациент орально иммунизируется посредством иммунологически эффективного количества антигена Г.пилори с тем, чтобы вызвать образование защитных антител против патогена Г.пилори. Желательно, чтобы Г.пилори вводились в ассоциации со слизистым вспомогательным веществом, например с холерным токсином.
Кроме того, в объем настоящего изобретения входит пассивная иммунизация млекопитающих, включая людей, против инфекции Г.пилори. Это достигается оральным введением эффективного количества специфического антитела Г.пилори в пациента. Желательно, чтобы орально в пациента вводилось специфическое IgA моноклонное антитело Г.пилори.
Вакцина согласно данному изобретению вводится орально в количествах, которых легко могут определить обычные специалисты-медики. Для взрослых людей подходящая дозировка будет находится в диапазоне от 10 мкг до 10 мг, например, от 5 мкг до 5 мг. Сходные дозировки пригодны и для детей.
Как отмечалось выше, подходящим вспомогательным мукозальным (слизистым) веществом является холерный токсин. Другие вещества, которые можно применять, это нетоксичные производные холерного токсина, включая его B подразделение или/и сопряженные элементы антигена плюс холерный токсин или его B подразделение, микрокапсулы, или иммунные стимулирующие комплексы (ИСКОМы) или липосомы и ослабленные живые векторы, такие, как вирусы или бактерии Салмонеллы. Количество используемого слизистого вспомогательного вещества зависит от типа используемого вещества. Например, если слизистое вспомогательное вещество - это холерный токсин, он удобно используется в количестве от 5 мкг до 50 мкг, например от 10 мкг до 35 мкг. При использовании в виде микрокапсул используемое количество будет зависеть от количества вещества в матрице микрокапсулы для достижения желаемой дозировки. Все это может решить обычный специалист-медик.
Подходящие носители и разбавители это энтерические покрытые капсулы или/и 0,2N NaHCO3 или/и соль.
Примеры.
Далее изобретение будет описано с помощью следующих неограниченных его примеров.
(а) Мыши
Мыши, используемые в экспериментах, это свободные от бактерий мыши Суисс-Уэбстера (в возрасте 8 недель), полученные из Таконика (Джермантаун, штат Нью-Йорк). Животные были помещены в микроизолированных клетках в условиях отсутствия бактерий и им был предоставлен свободный доступ к помещенной в лабораторный автоклав пище и воде. За исключением времени от время изолирующих дифтероидов, животные поддерживались в свободном от микробов состоянии на протяжении всего процесса иммунизации.
(б) Бактериальные штаммы
Бактерии, полученные из образцов желудочной биопсии кошки, были идентифицированы как Г.фелис, основываясь на морфологии. Грамм-штамме и на производстве мочевины, катализа и оксидаза 9. Организмы хранились в 50% фосфатно-буферной соли (ФБС), 25% глицерина, 25% нагретой утробной сыворотки теленка при - 70oC. Бактерии, использованные в приведенных примерах, переносились in vitro от 2-х до 3-х раз после изоляции.
(в) Бактериальные антигены
Испытательный штамм прививался колумбийскому агару (Дифко, Детройт, штат Мичиган), содержащему 7% лошадиной крови, и культивировался микроаэрофилически при 37oC в течение 5-7 дней. Организмы собирались в ФСБ, и результирующие взвеси были соницированы для лизирования бактерий при 40oC; очищались от клеточных остатков низкоскоростным центрифугированием и стерильно фильтровались. Эти полноклеточные соникаты хранились как 100 мкл алигвоты при - 70oC до применения в оральной иммунизации животных.
(г) Внешние мембраны
Внешние мембраны подготавливались как описывалось. Говоря кратко, бактериальные суспензии обрабатывались 1 мг рибонуклеазы и деоксирибонуклеазы (Сигма Кемикал, Сент-Луис) в 0,5 M Tris - EDTA Буфера (pH 7,8) при 4oC непосредственно перед соникацией и низкоскоростным центрифугированием, как описано выше. Затем бактериальные обвертки отделялись от очищенного лизата ультрацентрифугированием при 1500000 хг в течение 1 ч. внешние мембраны отделялись от клеточных обверток путем дифференциальной солюбилизации в натрий п-лауройлсарконине и восстанавливались ультрацентрифугированием. Получившиеся в результате гранулы взвешивались в 0,05 M фосфатном буфере (pH 7,0), разделялись на алигваты и хранились при - 70oC. Концентрация белка определялась по методу Лоури и др. для использования в ELISA (20).
Пример 1.
Мыши слегка анестезировались i. p. инъекцией 1,0 мг кетамина до внутрижелудочной иммунизации. Затем полноклеточные соникатные препараты плюс 10 мг холерного токсина (Лист Байолоджикалс, Кэмпбелл, штат Калифорния) взвешивались в 0,2 M NaHCO3, 0,5 мл были введены в желудки мышей интубацией через полиэтиленовые трубки, присоединенные к шприцу для подкожных впрыскиваний. Эту процедуру мы называем оральной иммунизацией.
Для изучения возможности развития функциональной невосприимчивости было оценено три процесса оральной иммунизации. Процесс 1 включал в себя 4 оральные иммунизации в течение 1 месяца, включая 2 мг лизата Г.фелис плюс холерный токсин (известное слизистое вспомогательное вещество). Процесс 2 увеличивал количество Г. Феликс до 1 мг на иммунизацию плюс холерный токсин, а процесс 3 состоял из 5 оральных иммунизаций в течение 6 недель, каждая из которых включая 4 мг лизата Г.фелис плюс холерный токсин. За исключением тех случаев, когда отмечено иначе, животные инфицировались спустя 7 - 10 дней после последней иммунизации и жертвовались спустя 7 дней.
Собирались следующие жидкости тканей: сыворотка, желудочные секреции и кишечные секреции. Затем эти образцы титровались на присутствие антител анти-Г. пилори посредством энзимосвязанного иммуносорбентного анализа (ELISA). Кроме того, были получены желудочные биопсии для быстрого теста мочевины и культуры. Инфекция определялась как положительная, если либо культура, либо культура, либо быстрый тест мочевины (смотрите ниже) были положительными. Сыворотка получалась путем вскрытия хвостовой вены и свертывания крови при комнатной температуре. Желудочные и кишечные секреции мышей собрались раздельно путем модификации процедуры Элсона и др. Желудки и кишечники удалялись и инъецировались 2,0 мл. орошением на основе полиэтилен гликоля плюс анти-протеазный раствор. Желудочное орошение содержало Tris буфер для нейтрализации желудочной кислоты.
Процедура ELISA выполнялась следующим образом. Анализировались образцы мышей на антитела Г.Феликс следующим образом. 96 полистиреновых микротитрных пластинок покрывались 100 мкл/well соответствующими белками внешней мембраны (20 мкг/мл) в течение ночи (или накануне вечером) при 4oC. Неспецифические связующие участки блокировались 1% BSA в ФСБ на 90 мин при комнатной температуре и затем пластинки промывались с помощью 0,1% BSA в ФСБ. Образцы испытывались в дубликате при разбавлении в диапазоне от неразбавленности до 1: 512000 и 100 мкл для каждого разбавления/well добавлялось к пластинкам, покрытым антигеном. После культивации при комнатной температуре в течение 90 мин пластинки промывались 3 раза с 0,1% BSA в ФБС и 100 мкл 1:1000 раствора козлиного антимышиного IgA или IgG сопряженного щелочного фосфата (Займед, Сан-Франциско, штат Калифорния) добавлялось к каждому углублению (well) в течение 90 мин. После промывания пластинки проявлялись посредством 100 мкл на углубление (well) раствора 1 мг/мл р-нитрофенил фосфата в глицинном буфере (pH 9,6) в течение 1 ч. Абсорбируемость при 410 нм измерялась в каждом углублении (well) с использованием устройства для чтения пластинок с микротитрами MR 700 фирмы Дайнатек. Титр антитела определялся как обратная величина самого высокого разбавления, дающего оптическую плотность 0,05 над углублениями (well), которые содержали антиген и которые были культивированы сопряженным антителом, но без первичного образца антитела (Infeet. Immun., 1991, 59, 785-79).
Быстрый тест на мочевину выполняется следующим образом. 2 образца желудочной биопсии по 10 мг веса во влажном состоянии от каждой мыши были немедленно помещены в 0,2 мл испытательно питательной среды мочевины Стьюарта 28 и культивировались при комнатной температуре. Присутствие мочевины определялось по изменению цвета от желтого до розового в испытательной среде после 4-х часов (J. Pediatr., 1987, 11-569-570).
Культуры получались следующим образом. Биопсии желудочной полости были гомогенизированы и помещены на колумбийского агара, содержащего 5% овечьей крови, и культивировались при 37oC в микроаэрофильных условиях (набор для выработки газа; Оксолд Лдт, Лондон, Англия). Положительная культура определялась визуально как рост спустя 5 дней. Все изоляты идентифицировались как Г.фелис, основываясь на морфологии, грамм-штамме и выработке мочевины, катализа и оксидазы.
Несмотря на небольшие изменения в устройстве эксперимента в трех группах, не было отмечено значительного различия в иммунной реакции. Данные были собраны таким образом, и геометрические средства желудочного орошения, кишечного орошения и титры антител сыворотки от исследованных 13 контрольных и 12 иммунизированных животных представлены в табл. 1 и фиг. 1.
Хотя эти животные были иммунизированы и инфицированы, титры антитела значительно не отличались от мышей, которые были иммунизированы и не инфицированы. В этих экспериментах титры IgA и IgG антитела желудка, кишечника и сыворотки были значительно выше, чем те, которые наблюдались у неиммунизированных контрольных животных. Конкретно, имелось 4-кратное увеличение в желудочном IgA (p-0,001), 8-кратное увеличение в кишечном IgA (p=0,0038) и 350-кратное увеличение в сыворотке IgA (p=0,0001) по сравнению с неиммунизированными контрольными животными. Аналогичным образом, наблюдалось значительное повышение желудочного IgG (p=0,0009), кишечного IgG (p=0,0001) и IgG сыворотки (p=0,0001).
Для того чтобы оценить защиту от инфекции Г.фелис, были взяты биопсии желудка при жертвовании всех животных и оценены быстрым тестом на мочевину и на культуру, как описано выше. Кроме того, для определения того, проявляют ли контрольные животные хроническую инфекцию, и являются ли иммунизированные животные определенно Г.фелис отрицательными, дополнительные иммунизированные и контрольные животные были инфицированы, как описано выше но не были убиты до 4-х недель после инфецирования. Степень защиты среди всех иммунизированных групп животных не была заметно разной.
Для того, чтобы не исключить возможность нижнего уровня, не производился подсчет образцов желудочной биопсии как положительной или отрицательной для роста Г. фелис до конца 5-ти после размещения. После размещения последовательных растворов известных количеств (по гемацитомерной размерности) выращенной культуры Г.фелис было обнаружено, что чувствительность этого конечного пункта это приблизительно 10 организмов. В последующих экспериментах пластинки с культурой биопсии иногда сохранялись даже дольше, чем 5 дней, и когда пластинки, остающиеся отрицательными в произрастании при визуальном осмотре, были очищены и исследованы на влажном заключение среза, можно было видеть изолированный организм в форме спирали. Нельзя было подтвердить идентичность этих изолированных организмов и нельзя было определить их жизнеспособность. Во всяком случае, основываясь на результатах культуры для последовательно разбавленных Г. фелис, считается, что образцы биопсии, оставшиеся отрицательными к росту при 5-ти днях, содержали 10 или менее бактерий.
Пример 2.
Моноклонные антитела IgA и IgG, специфические для Г.фелис, были получены посредством модификации процедуры Мазанека и др. (15). Мыши BALB/c, полученные из лаборатории Джексона (Бар Харбор, штат Мейн) были иммунизированы внутрижелудочно 4 раза на протяжении периода в 6 недель, первые 3 раза с 2 мг соницированного Г.фелис плюс 10 мкг холерного токсина (Сигма Кемикал Компани, Сент-Луис). Для последней иммунизации холерный токсин не использовался, и мыши также получили внутривенное форсирование с 2 мг белка Г.фелис. Три дня спустя мыши были убиты, и их клетки селезенки были гибридизированы до SP2/0 клеток миеломы. Клоны, полученные ограничивающим разведением, были проведены на секрецию антитела анти-Г.фелис IgA методом ELISA. Обнаруживалось, что получившаяся в результате клеточная цепочка, идентифицированная как # 71-G5. A8, является стабильной выделяющей IgA гибридомой. После ряда субклонирований стабильные секреторы IgA и IgG инъецировались внутрибрюшинно в лучших мышей BALB/c, собиралась и просветлялась асцитическая жидкость.
Клеточная цепочка #71-G5-A8 по состоянию на 13 апреля 1992 г. сдавалась на хранение и поддерживалась в жизнеспособном состоянии в лаборатории Стивена Дж. Зинна, доктора медицины, детская больница Рейнбору, комната 465, университет Кейс-Вестерн, 2074 Эбингтон Роуд, Кливленд, штат Огайо, США 44106. Доступ к депозиту будет предоставлен лицу, назначенному главным уполномоченным по патентам и товарным знакам США и имеющему соответствующие полномочия во время рассмотрения настоящей заявки, и все ограничения на предоставление вниманию публики будут окончательно сняты после выдачи патента по данной заявке.
Клеточная цепочка #71-G5-A8 сдается на хранение в Американском собрании типов культуры, размещенном в 12301 Парклон Драйв, Роквил, штат Мериленд 20852, США, под идентификационным номером #71-G5-A8. Номер доступа в ACTK и время сдачи на хранение HB11514 и 23.12.93 соответственно.
Пример 3.
Исследования по пассивной иммунизации проводились следующим образом. Асцит, содержащий IgA моноклональное антитело, полученное из #71-G5-A8 (200 мкл), вводился внутрижелудочно одновременно с 10 жизнеспособными организмами. Предварительные исследования указывали на то, что желудочные IgA тиры животных, которые приняли одну дозу 200 мкл моноклонального IgA антитела, снизились до уровней ниже тех, которые наблюдаются у активно иммунизированных животных через 8 ч. Поэтому 3 дополнительные дозы MAb давались через следующие 24 ч. Контрольные животные подвергались воздействию идентичным образом, но принимали либо соль, либо специфическое IgA моноклонального антитело вирус Сендай (несоответствующее IgA моноклональное антитело). Спустя неделю мыши были пожертвованы. Желудочная ткань прививалась на пластинки с кровью колумбийского агара и культивировалась в течение 5 дней при 37oC. Инфекция определялась как положительная культура или положительный быстрый тест Стьюарта на мочевину в питательной среде.
Для того, чтобы исследовать, могут ли IgA антитела, отличительный признак слизистой иммунной системы, сами защищать от инфекции Г.фелис слизистой оболочки желудка, как описано выше, генерировались Г.фелис IgA моноклональные антитела. Одно из этих антител (#71-G5-A8) затем пассивно орально вводилось свободным от бактерий мышам при и после воздействия посредством Г. фелис. Контрольные животные получали либо соль, либо специфическое IgA моноклональное антитело вирус Сендай, специфическое для хемагглутининнейраминидазы гликопротеина вирус Сендай.
Результаты представлены в табл. 2.
Г. фелис или специфическое IgA моноклональное антитело вирус Сендай давались внутрижелудочно 4 раза в течение 24 ч. совместно с воздействием 10 жизнеспособных Г.фелис. Желудочные биопсии были получены спустя 1 неделю после воздействия, и инфекция определялась по культуре или/и быстрому тесту на мочевину.
Из 13 контрольных животных, которые принимали антитела или антитела вируса Сендай, 70% были инфицированы (фиг. 3). Из 7-ми экспериментальных животных 6 были защищены и только одно (14%) было инфицировано. По анализу "хи квадратов" разница была значительной (p = 0,19).
Сравнивание титров антитела среди экспериментальных групп оценивалось путем анализа методом дисперсии и путем защищенного Т-теста Фишера. Для защиты отсутствие или присутствие экспериментальной инфекции среди групп определялось анализом "хи квадратов".
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИНДУЦИРОВАНИЯ У МЛЕКОПИТАЮЩИХ ЗАЩИТНОГО ИММУННОГО ОТВЕТА НА ИНФЕКЦИЮ HELICOBACTER, ВАКЦИНА ДЛЯ ИНДУЦИРОВАНИЯ ЗАЩИТНОГО ИММУННОГО ОТВЕТА НА ИНФЕКЦИЮ HELICOBACTER У МЛЕКОПИТАЮЩИХ, СПОСОБ ИНДУЦИРОВАНИЯ ПАССИВНОЙ ЗАЩИТЫ У МЛЕКОПИТАЮЩИХ ПРОТИВ ИНФЕКЦИИ HELICOBACTER И СПОСОБ ОЦЕНКИ ЭНДОГЕННОГО ИММУННОГО ОТВЕТА У МЛЕКОПИТАЮЩИХ, ИНФИЦИРОВАННЫХ HELICOBACTER | 1993 |
|
RU2125891C1 |
| МУТАНТ ЭНТЕРОТОКСИНА, ЭФФЕКТИВНЫЙ В КАЧЕСТВЕ НЕТОКСИЧНОГО ОРАЛЬНОГО СТИМУЛЯТОРА | 1995 |
|
RU2160606C2 |
| ТЕРАПЕВТИЧЕСКИЕ АГЕНТЫ И АУТОИММУННЫЕ ЗАБОЛЕВАНИЯ | 1996 |
|
RU2203088C2 |
| АНТИГЕН HELICOBACTER PYLORI И ВАКЦИННАЯ КОМПОЗИЦИЯ | 1996 |
|
RU2195463C2 |
| ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ДЛЯ ВВЕДЕНИЯ ЧЕРЕЗ СЛИЗИСТУЮ И СПОСОБ ПРЕДОТВРАЩЕНИЯ ГРИППА | 2004 |
|
RU2390351C2 |
| ИНДУКТОР СЕКРЕТОРНЫХ АНТИТЕЛ IgA И IgG | 2006 |
|
RU2440138C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВАКЦИННОЙ КОМПОЗИЦИИ ПРОТИВ КИШЕЧНОЙ ИНФЕКЦИИ | 1992 |
|
RU2127121C1 |
| СИНТЕТИЧЕСКИЙ INVAPLEX | 2007 |
|
RU2440136C2 |
| Фармацевтическая композиция на основе PLGA для индукции эффективного мукозального иммунного ответа | 2018 |
|
RU2742580C2 |
| СПОСОБ ДОСТАВКИ БИОАКТИВНОГО АГЕНТА ЖИВОТНОМУ ДЛЯ ИНИЦИАЦИИ ИММУННОГО ОТВЕТА (ВАРИАНТЫ) | 1989 |
|
RU2127118C1 |



Способ может быть использован в области медицины, в частности в иммунологии и гастроэнтерологии для лечения желудочной инфекции у млекопитающих, в частности геликобактерной инфекции, составами вакцин и антител. Способ позволяет создать вакцины, содержащие некоторое количество антигена геликобактера, эффективное для вызывания защитной иммунной реакции. Способ вызывания у "хозяина" защитной иммунной реакции на инфекцию Геликобактер, заключается в введении в "хозяина" иммуногенически эффективного количества антигена Геликобактер для использования при получении указанной защитной реакции у человека. Указанный антиген Геликобактер представляет собой антиген Геликобактер пилори и фелис. Вакцина для лечения инфекции Геликобактер включает в себя иммуногенически эффективное количество антигена Геликобактер в ассоциации с фармацевтически приемлемым носителем или разбавителем, а способ наделения "хозяина" - млекопитающего пассивной защитной от инфекции Геликобактер также заключается в введении в "хозяина" иммунологически эффективного количества Геликобактер - специфического антитела, представляющего собой JgA антитело. 3 с и 13 з.п.ф-лы, 3 ил., 2 табл.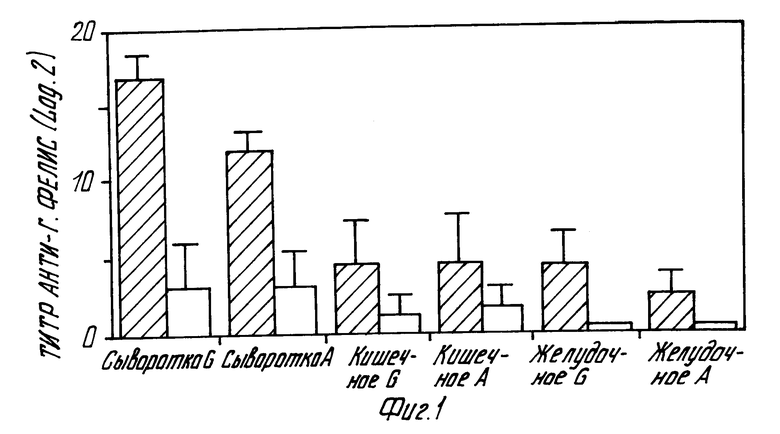
| Czinn S.J | |||
| and Nedrud J.G | |||
| Oral Immunization against Helicobacter pylori., Infect | |||
| Immun., 1991, 59, p | |||
| Приспособление для обточки карандашей | 1925 |
|
SU2359A1 |
