Настоящее изобретение относится к области генной инженерии и биотехнологии, в частности к производству вакцин против вируса гепатита B.
Сущность изобретения.
Объектом изобретения является способ получения поверхностного антигена вируса гепатита В (HBsAg). Способ заключается в культивировании штамма дрожжей Pichia pastoris C 226, трансформированного рекомбинантной плазмидой TAО 906, содержащей ген HBsAg, лизисе клеток, осаждении контактирующих примесей при pH 2,5 - 3,0, отделении недостаточного слоя, содержащего антигенную фракцию, смешивании его с адсорбентом на основе диатомовой земли, из которой элюируют контаминирующие примеси при значении pH 3-4, отделении антигенной фракции от адсорбента при pH 9,0 - 9,5, очистке данной фракции с помощью иммуноаффинной хроматографии при использовании моноклональных антител CB-HEP I и элюции буфером, содержащим хаотропный агент, гельфильтрации с последующей анионообменной хроматографией в присутствии детергента, элюции антигенной фракции, фильтровании через мембрану с размером пор 0,2 мкм и сборе целевого продукта после хроматографии высокого разрешения в виде иммуногенных частиц. Полученный указанным способом HBsAg обладает следующими признаками: имеет мол.массу 24000 дальтон при определении методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия, агрегирован в частицы размером 22 нм, обладает гомогенностью более 3700 мкг специфического белка по отношению к 1 мг общего белка, примеси нуклеиновых кислот составляют менее 3 пг на исследуемую дозу антигена и контаминация белками дрожжей составляет менее 3%. Используемые для иммуноаффинной очистки HBsAg моноклональные антитела CB-HEP I продуцируются гибридным клоном 48/1/574, относятся к изотопу IgG2b или IgM, специфически взаимодействуют с полученным, как указано выше, и природным HBsAg на поверхности твердой фазы и в растворе и распознают непрерывный эпитоп на общей антигенной детерминанте "а" поверхностного антигена всех подвидов вируса гепатита B. Гибридный клон 48/1/574, продуцирующий моноклональные антитела CB-HEP, получают традиционным способом, который заключается в иммунизации мышей линии BaPb/c HBsAg, слиянии клеток селезенки иммунизированных животных с клетками миеломы SP2/0/Ag14, культивировании гибридных клонов на селективной питательной среде и отбором клона 48/1/574, который продуцирует нужные антитела CB-HEP I.
Уровень техники.
Существуют два основных направления производства вакцины против вируса гепатипа B: на основе плазмы крови и с помощью методов генной инженерии.
Получение вакцины из плазмы основывается на выделении в чистом виде поверхностного антигена вируса гепатита B (HBsAg) из плазмы хронических носителей вируса с помощью различных физических методов и последующей его инактивации (Zuckerman A.J. et al. British Medical Journal 1985, 290:492). Эффективность и надежность этих вакцин прошла проверку временем (Prince A.M. et al. Annual Clinical Research 1982, 14:225), при этом не было отмечено сопутствующего риска передачи вируса иммунодефицита человека или других возбудителей инфекции (Deinhardt F. et al. Journal of Medical Virology 1985, 17: 209).
Тем не менее широкое применение этой вакцины затрудняется ограниченными запасами сыворотки носителей вируса для получения HBsAg, используемого для производства вакцины, необходимостью строгого соблюдения серьезных требований, предъявляемых к очистке и инактивации вируса гепатита B, и отделения других возбудителей инфекции, присутствующих в плазме, а также продолжительными испытаниями на безвредность, необходимыми для допуска партий этой продукции к применению. Кроме того, существует особая озабоченность тем, что возбудители инфекции, которые передаются через кровь, могут "избежать", процесса инактивации в ходе выработки вакцины. Поэтому наблюдается общая тенденция к более широкому применению методов генной инженерии для получения вакцин против гепатита B.
Ген поверхностного вируса гепатита B клонируется и экспрессируется в прокариотических и эукариотических клетках.
Уровни экспрессии, получаемые при использовании бактерий, бывают низкими, и не происходит эффективной сборки антигена в иммуногенные частицы размером 22 нм, в силу чего данная система обычно не используется в качестве источника антигена для производства вакцин (Valensuela P. et al., Nature 1980, 280:815; Burrell C.J. et al., Nature 1979, 279:43).
Эукариотические клетки также используются для получения HBsAg (Michel M. L. et al., Biotechnology 1985, 3:561; Macnab G.M. et al., British Journal of Cancer 1976, 36:509), тем не менее производство вакцин на этой основе требует применения дорогостоящих и сложных методов, оборудования и питательных сред, что затрудняет организацию крупномасштабного производства. Кроме того, существуют опасения относительно надежности вакцин, полученных на основе клеточных линий млекопитающих, из-за возможного наличия ретровирусов. Поэтому данный подход также не всегда используется для широкого производства гепатитной вакцины.
Существующие коммерческие рекомбинантные вакцины против гепатита B основаны главным образом на HBsAg, полученном в генетически трансформированных дрожжах.
Антиген производится внутриклеточно, извлекается с помощью различных способов лизиса клеток и очищается при использовании разнообразных физико-химических методов, позволяющих добиться степени чистоты более 97% и получить продукт, антигенные свойства которого, как это было подтверждено на животных и человеке, соответствуют антигену из плазмы крови (Hauser P. et al. Posgraduate Medical Journal 1987, 63 suppl 2:83).
Недавно была описана эффективная система экспрессии, где в качестве клеток-хозяев используются клетки метилотрофных дрожжей Pichia pastoris. Система основана на применении промотора гена фермента алькогольоксидазы I (AOX1). Указанный фермент катализирует первую реакцию утилизации метанола дрожжами Pichia pastoris. Промотор AOX1 жестко регулируется и обеспечивает высокие уровни экспрессии фермента (до 30%), когда клетки выращивают в присутствии метанола, но не в присутствии глюкозы (Ellis S.B. et al. Molecular and Cellular Biology, 1985, 5:1111; Couderc R.et al. Agric Biol Chem. 1980, 44:2259).
Дрожжи P. pastoris используются для экспрессии различных чужеродных белков, в том числе HBsAg (Cregg J.M. et al. Biotechnology 1987, 5: 479). При этом ген, кодирующий HBsAg клонируется под контролем промотора алкогольоксидазы I (AOX1) в кассете экспрессии, которая интегрируется в хромосому мутантного штамма указанных дрожжей. Это обуславливает тот факт, что при активации системы в присутствии метанола от 2 до 3% полученных в растворенном виде белков приходится на HBsAg.
Другим достоинством данной системы по сравнению с S. cerevisiae является эффективная сборка антигена в частицы размером 22 нм, и в результате этого практически весь полученный антиген оказывается агрегированным в антигенные частицы в отличие от того, что происходит в системе S. serevisiae, где только незначительная часть мономеров размером 24 kd собирается в антигенные частицы (Valenzuela P. et al. Nature 1982, 298:397; Hitzeman R.A. et al. Nucleic Acids Research 1983, 11: 2745; Miyanohara A. et al. Proceedings of the National Academy of Sciences USA 1983, 80:1). Тем не менее, для вакцинных препаратов, основанных на рекомбинантном HBsAg, которые в настоящее время имеются на рынке, в качестве источника антигена по-прежнему используют дрожжи S. cerevisiae. Известен способ получения HBsAg при культивировании дрожжей P. pastoris с последующим выделением HBsAg из культуральной жидкости (заявка на Европейский патент 0337492, C 12 P 21/00, 1989).
С помощью заявленного способа, включающего соблюдение специфических условий ферментации и очистки, рекомбинантный антиген получают при культивировании дрожжей Pichia pastoris; при этом он отличается высокой степенью агрегирования в частицы и большой однородностью, что придает ему более высокие иммуногенные свойства.
Особенности процесса ферментации и состава примененной питательной среды позволяют эффективно создавать оптимальные физиологические условия, необходимые для роста микроорганизмов и для предпочтительной экспрессии клонируемого гена, а также добиваться высокой степени агрегирования получаемого продукта в частицы.
Для получения агрегированного в частицы антигена высокой степени чистоты авторами изобретения разработана специальная процедура очистки, при которой получают антигенный препарат, контролируемые характеристики которого отвечают установленным требованиям.
Сведения, подтверждающие возможность осуществления изобретения
Пример 1.
Для получения гена поверхностного антигена вируса гепатита B проводят клонирование вирусной ДНК по сайту рестриктазы EcoRI в векторе pBR 322. Вирусный геном изолируют из частиц Дейна, выделенных из сыворотки бессимптомного носителя вируса и очищенных при помощи процедуры, сходной с описанной в литературе (Valenzuela P. et al. Nature 1979, 280: 815). Полученную плазмиду, обозначенную pHBS1, расщепляют при помощи рестрикционных эндонуклеаз EcoRI и HpaI, в результате чего получают фрагмент, который в свою очередь расщепляют эндонуклеазой TagI. Затем по данному сайту присоединяют линкер EcoRI и осуществляют субклонирование по сайту рестрикции EcoRI в pBP 322. Полученную плазмиду обозначают pHBS2, из нее путем использования рестриктазы EcoRI и обработки S1-нуклеазой получают фрагмент, содержащий ген HBsAg.
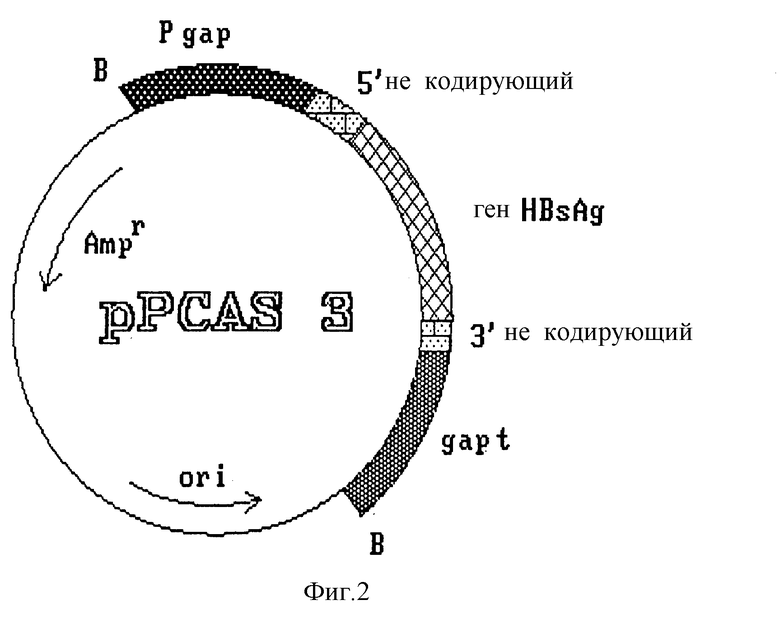
Этот фрагмент субклонируют в вектор pBS 5 (фиг. 1), предварительно линеаризованный с помощью NcoI и обработанный S1-нуклеазой и щелочной фосфатазой. Полученную плазмиду обозначают pRCAS 3 (фиг. 2). Из нее удаляют некодирующие вирусные участки на концах 5' (25 pb) и 3' (125 pb) по обе стороны от гена HBsAg с помощью полимеразной цепной реакции (PCR), которую осуществляют по стандартной методике (Saiki R.K. et al. Science 1985, 230:1350).
При использовании двух искусственных олигонуклеотидов со следующими последовательностями:
Область 5': 5′GCCATGGAGAACATCACAT3′
Область 3': 3′GACCCATATGTAAATTTGCAGCTGG5′
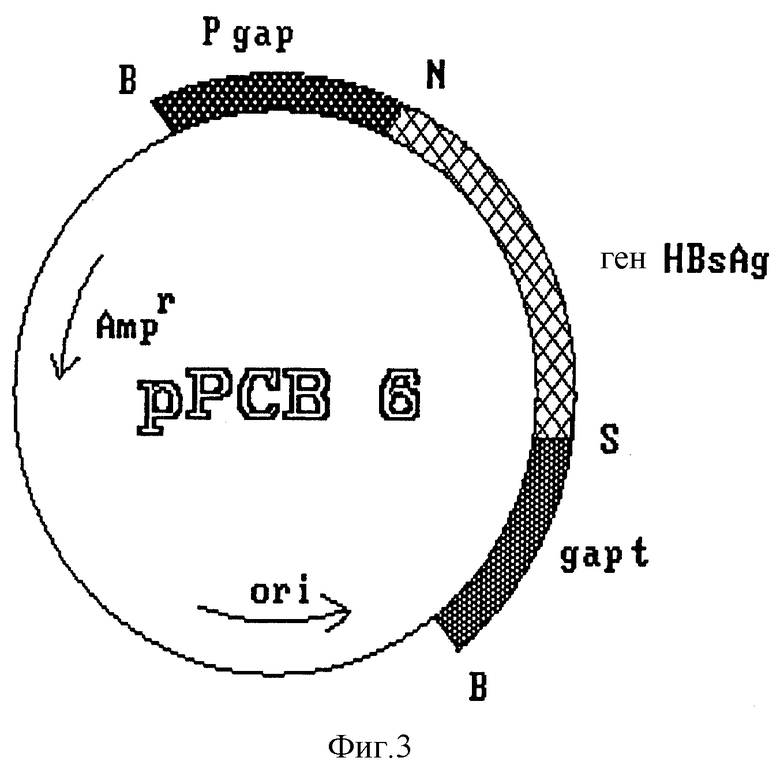
создают сайты рестрикции Ncol и Sall на 5'- и 3'-концах гена соответственно, что обеспечивает удобство дальнейшего манипулирования и удаление некодирующих участков после разрезания амплифицированного фрагмента упомянутыми ферментами. Полученный фрагмент размером 678 pb, содержащий полный ген HBsAg, субклонируют в плазмиде pBS 5. Полученная плазмида, обозначенная pPCB6 (фиг. 3), содержит упомянутый ген, контролируемый сигналами регуляции транскрипции гена фермента глицеральдегид-3-фосфат-дегидрогеназы (GAP) дрожжей S. cerevisiae.
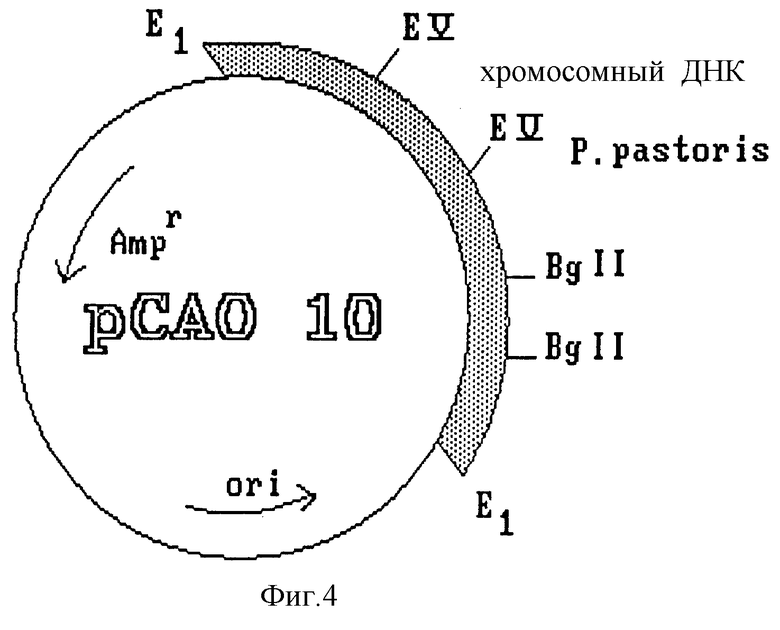
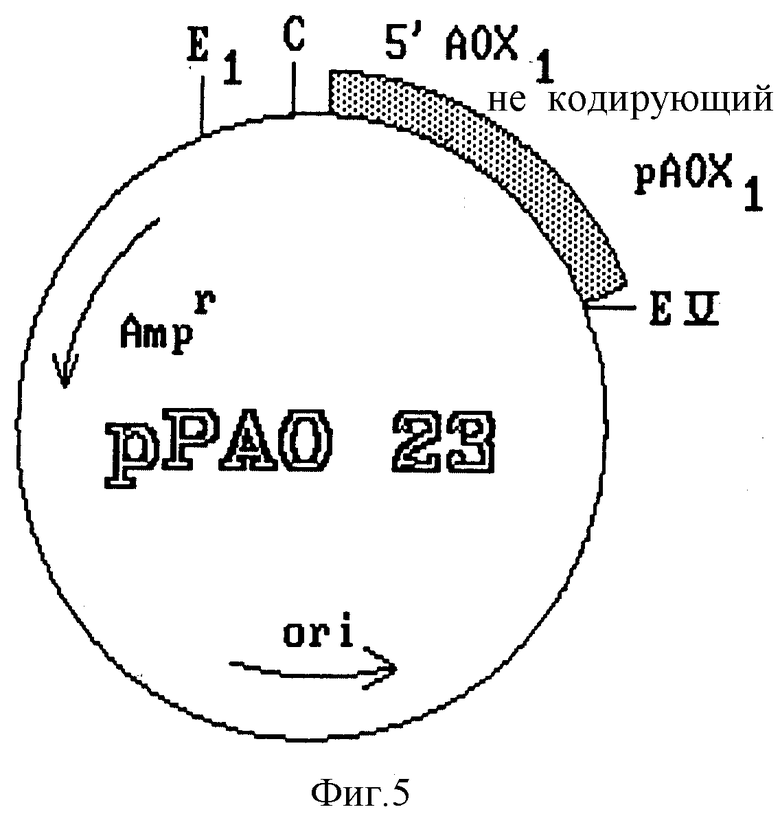
Из плазмиды pCAO 10, полученной из библиотеки ДНК Pichia pastoris (фиг. 4), изолируют фрагмент длиной 1,1 kb, содержащий промотор гена фермента AOX1, а также некодирующую область 5'. Этот фрагмент субклонируют в плазмиде PBR 322, и в результате получают плазмиду pPAO 23 (фиг. 5). К последней применяют полимеразную цепную реакцию (PCR) в целях элиминации 18 pb структурного гена фермента AOX1, которые соответствуют нуклеотидам, кодирующим первые шесть аминокислот этого белка, присутствующие в данной конструкции.
Используют следующие олигонуклеотиды:
Область 5': 5′GTATCACGAGGCCCT3′
Область 3': 3′TTGATTAATAAGGTTTGGTACGGC5′
Олигонуклеотид области 3' позволяет генерировать сайт рестрикции NcoI на 3' - конце промотора гена фермента AOX1 для элиминации последовательности длиной 18 pb из структурного гена указанного фермента и достижения точного соединения с геном HBsAg. Олигонуклеотид области 5' обеспечивает сохранение сайта рестрикции в PBR 322.
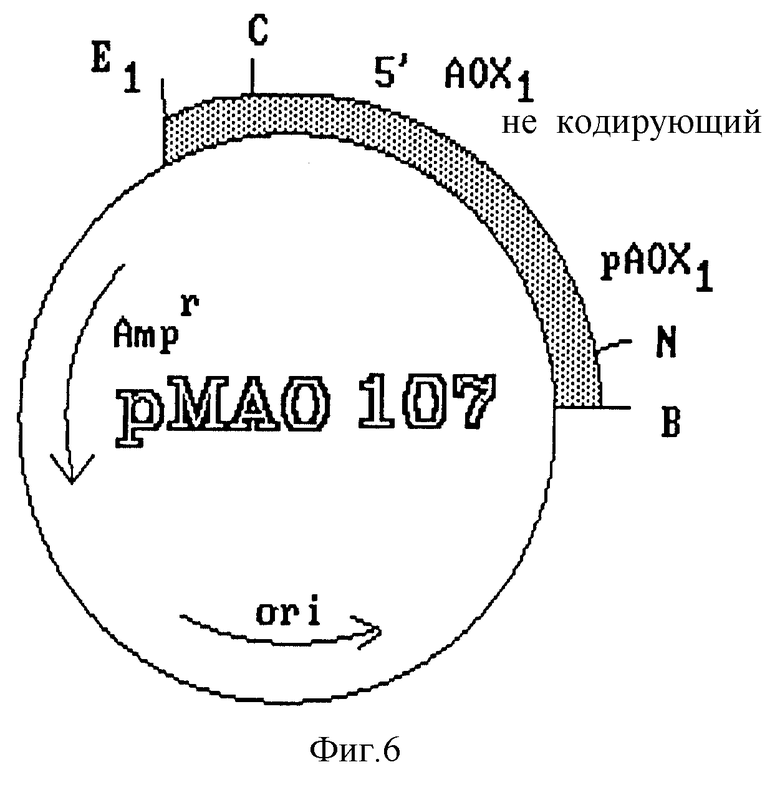
Амплифицированный фрагмент длиной 1115 pb субклонируют в плазмиде PUS 19, и в результате получают плазмиду, обозначенную pMAO 107 (фиг. 6), содержащую усовершенствованный промотор гена фермента AOXI дрожжей P. pastoris.
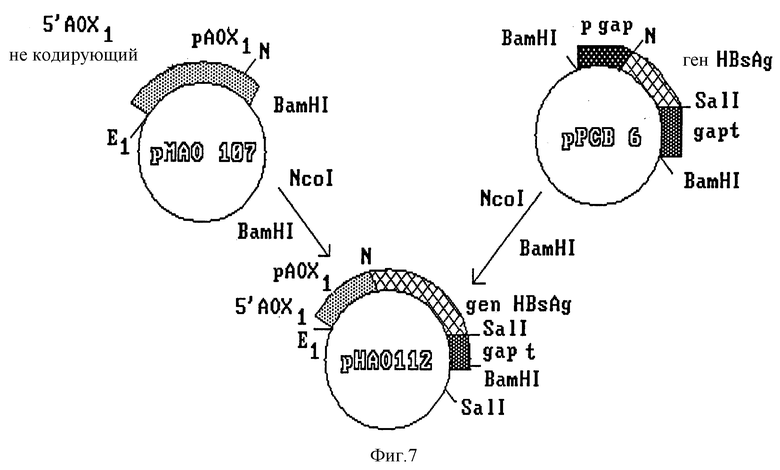
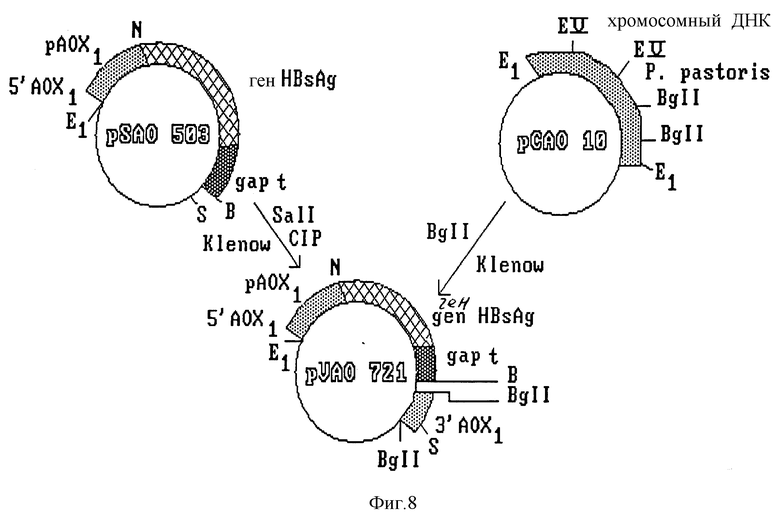
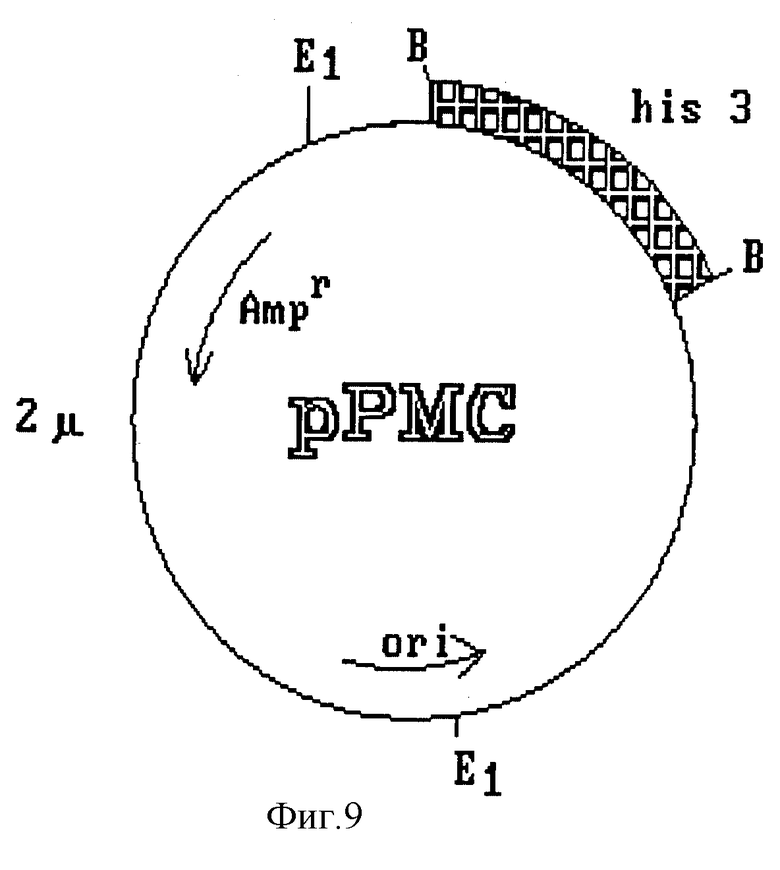
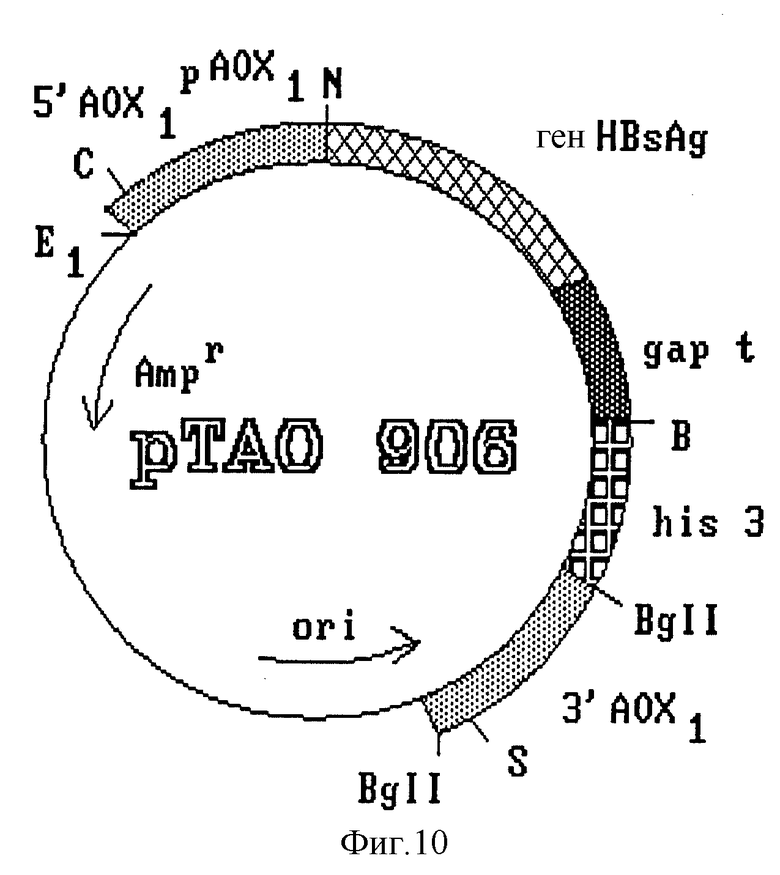
Конечный вектор интеграции получают путем субклонирования полного гена HBsAg под контролем усовершенствованного промотора фермента AOX1 (фиг. 7). Полученный клон, обозначенный pHAO 112, содержит ген HBsAg, контролируемый сигналом регуляции промотора AOX1 и сигналом терминации гена дрожжей S. cerevisiae. В нем также имеется участок хромосомной ДНК из P. pastoris, необходимый для гомологичной рекомбинации с дрожжевой ДНК. Путем частичного расщепления и обработки этой плазмиды S1-нуклеазой в ней разрушают сайт рестриктазы Sall, находящийся между 3'-концом гена HBsAg и терминатором фермента глицеральдегид-3-фосфат-дегидрогеназы. Полученный клон обозначают PSAO 503 (фиг. 8). В нем субклонируют фрагмент длиной 2,4 kb, содержащий участок структурного гена фермента AOX1, и некодирующую область 3'-конца; фрагмент изолизируют из плазмиды pCAO 10. Полученную в результате плазмиду обозначают pVAO 721 (фиг. 8), в ней клонируют выделенный из плазмиды pPMC фрагмент длиной 1,8 kb, содержащий клеточный ген his 3 дрожжей S. cerevisiae (фиг. 9).
Полученный клон pTAO 906 (фиг. 10), представляющий собой конечную интеграционную плазмиду, используют для трансформации штамма MP-36 дрожжей P. pastoris. Трансформанты с интегрированной плазмидой используются для оценки уровней экспрессии антигена. Штамм, отобранный для производства антигена, обозначают C 226.
Пример 2.
Для синтеза HBsAg трансформированный штамм C 226 предварительно культивируют в соляной среде следующего состава:
K2HPO4 - 5 г/л
MgSO4 - 4,6 г/л
NH4SO4 - 22 г/л
CaCl2 - 0,5 г/л
Глицерин - 20 мл/л
Витамины 200 x - 10 мл/л
Соли в следовых количествах 200 x - 5 мл/л
Выращивание производят при температуре 30oC и pH 4,5, причем в таких условиях аэрации и перемешивания, которые гарантировали бы парциальное давление кислорода на 30% выше давления насыщения. Культуру выращивают в течение 10-12 часов.
На этапе инокуляции производят замену соляной среды на обогащенную среду следующего состава:
Дрожжевой экстракт - 1%
Пептон - 2%
Глицерин - 2%
Соли в следовых количествах 200 x - 10 мл/л
Витамины 200 x - 5 мл/л
Культуру выдерживают 10-12 часов в этих условиях, пока не будет достигнута плотность клеток около 10-12 г/л (в пересчете на сухое вещество), а затем переносят в ферментационную среду (в ферментер), причем ферментацию осуществляют в той же обогащенной среде при тех же условиях температуры и pH. Культуру выдерживают в течение 8-10 часов до достижения плотности клеток около 10-12 г/л (в пересчете на сухое вещество).
С этого момента начинают непрерывное добавление метанола, чтобы индуцировать экспрессию антигена, которая составляет примерно 3% от всего белка через 90-100 часов ферментации. Добавление метанола время от времени увеличивают в зависимости от того, как увеличивается плотность клеток в среде. В конце периода индукции начинается прирост культуры за счет обогащенной среды следующего состава:
Экстракт дрожжевой - 3%
Пептон - 6%
Прирост сохраняется до конца ферментации, которая продолжается около 180-200 часов, при этом 5-кратно увеличивается скорость роста, повышаются уровни экспрессии. Благодаря данному методу также обеспечивается сохранение экспрессированного таким образом белка (более 80%) в растворимом виде. Это обуславливает большую эффективность на дальнейших этапах очистки, а также лучшее агрегирование антигена в частицы.
На конечном этапе ферментации получают биомассу концентрацией от 60 до 80 г/л (в пересчете на сухое вещество), а уровни экспрессии антигена составляют от 5 до 6% всего белка.
По завершении ферментации клетки собирают центрифугированием и вплоть до дальнейшей обработки хранят при температуре -70oC.
Пример 3.
Для получения препарата на основе HBsAg используют колоночную хроматографию по следующей схеме.
Сырой экстракт, приготовленный путем механического разрушения клеток биомассы, полученной центрифугированием культуры дрожжей, трансформированной геном HBsAg, подвергают разработанной авторами процедуре, основанной на физико-химических и биологических особенностях антиген.
С этой целью около 40-60 граммов клеточной биомассы, содержащей поверхностный антиген, полученный внутриклеточным путем при использовании рабочего штамма C 226 и имеющий форму агрегата, состоящего из частиц размером 22 нм, ресуспендируют в 1000 мл дистиллированной воды. Эту суспензию непрерывно центрифугируют со скоростью 5000-6000 g, удаляют надосадочную жидкость и получают таким образом дрожжевую массу концентрацией 120-150 г/л (в пересчете на сухое вещество).
Данную процедуру повторяют вновь для отмывания клеточной суспензии, которую затем доводят до концентрации 100 г/л при помощи дистиллированной воды, и добавляют в нее специфические вещества, образующие детергентный буферный раствор следующего состава, г/л:
Трис - 12,1
Сахароза - 100
NaCl - 17,5
EDTA - 1,86
pH доводят до 7,5-8,0 при помощи NCl 1N.
Этот раствор гомогенизируют в течение 5 минут, после чего осуществляют механическое разрушение клеток, и затем клеточный дебрис и оставшиеся нетронутыми клетки отделяют центрифугированием со скоростью 15000 g для получения сырого экстракта. Последний подвергают процессу осветления, основанному на устойчивости HBsAg в условиях экстремальных значений pH, при которых многочисленные контаминирующие белки дрожжей-хозяина выпадают в осадок, в то время как антиген остается в растворе.
Осветление экстракта осаждением контаминирующих веществ при кислом pH осуществляют по следующей схеме.
Сырой экстракт, охлажденный до температуры 0-4oC, при постоянном перемешивании и контроле pH быстро смешивают (избегая образования пены) с раствором HCl 1N при температуре 4oC и в количестве, достаточном для снижения pH до 2,5-3,0. После дополнительного перемешивания в течение 5 минут смесь помещают в центрифугу непрерывного действия, охлажденную до температуры 4oC, и она находится там в течение 45 минут. При этом сосуды, из которых извлекают и в которые собирают материал, оставляют на холоде. PH надосадочной жидкости доводят до 7,5-8,5 сразу же после центрифугирования при непрерывном перемешивании и постоянном контроле.
Осветленный надосадочный слой, полученный на предыдущем этапе, примерно в течение часа гомогенизируют с матрицей диатомовой (инфузорной) земли в приблизительном соотношении 0,15-0,35 г антигена на килограмм земли. Антиген адсорбируется матрицей, затем посредством центрифугирования, фильтрации или отстаивания отделяют неадсорбированный материал, который удаляют. Далее осуществляют две или три промывки антигена, закрепившегося на подложке, буферным раствором следующего состав, г/л:
KSCN - 291,54
Сахароза - 100
Трис - 2,42
EDTA - 1,12
pH доводят до 3,0-4,0 при помощи HCl 1N.
После последовательных промываний производят десорбцию антигена из матрицы с помощью элюирующего буферного раствора следующего состава, г/л:
Трис - 2,42
EDTA - 1,12
Сахароза - 100
pH доводят до 9,0-9,5 при помощи NaOH 1N.
Полученный элюат вводят в концентрирующий аппарат типа AMICON в патроне с порами размером 0,1 мкм для повышения концентрации его начального объема примерно в 55 раз. Весь процесс осуществляют при температуре 4oC.
С целью получения материала, пригодного для хроматографических процессов, смешивают две части вышеупомянутого концентрата с одной частью 95%-ного этанола класса A, предварительно охлажденного до температуры -70oC, при постоянном перемешивании и температуре 0-4oC. Эту смесь центрифугируют со скоростью 15000 об/мин в течение 30 мин. Собирают надосадочную жидкость и сразу же в три раза разводят ее холодным буферным раствором Трис-HCl, 30 mM, pH 8,0 + 5% сахарозы. Концентрацию этанолового экстракта повышают в 4-5 раз, как было описано на предыдущем этапе.
Пример 4.
Для полной очистки HBsAg, полученного в соответствии с процедурой, описанной в предыдущем примере, осуществляют ряд хроматографических операций согласно нижеследующему описанию.
500-1000 мл концентрата этанолового экстракта обессоливают, пропуская через набухшую гельфильтрационную колонку с Сефадексом G-25, уравновешенную 20 мМ буфером Трис-HCl, pH 8,0. Прохождение через колонку контролируют посредством измерения проводимости и коэффициента поглощения при длине волны 280 нм, причем оба пика на хроматограмме должны быть полностью разделенными.
Приготавливают иммуноаффинную матрицу, которая содержит около 100 мл Сефарозы C1-4B, активированной бромистым цианом (CNBr), и примерно 500 мг сшитого с этой смолой моноклональных антител CB-HEP I (см. пример 5), обладающих особыми характеристиками, которые обеспечивают выделение частиц антигена высокой иммуногенности. Далее иммуноаффинную колонку, заполненную вышеупомянутой матрицей, уравновешивают в 20 мМ в буфере Трис-HCl, pH 8,0, вводят в колонку один объем обессоленного этанолового экстракта, который содержит антиген в примерном соотношении 0,1 мг HBsAg на 1 мл подложки, и пропускают его через колонку со скоростью около 400 мл/час при комнатной температуре, промывают уравновешивающим буферным раствором и контролируют прохождение по коэффициенту поглощения при длине волны 280 нм с регистрацией на графике. Элюат собирают до тех пор, пока кривая на хроматограмме не достигнет базовой линии, затем пропускают 20 мМ буферного раствора Трис-HCl, pH 8,0 + 1 M NaCl в целях элиминации неспецифической адсорбции до тех пор, пока кривая на хроматограмме вновь не достигнет базовой линии.
Антиген элюируют 20 мМ буферным раствором Трис-HCl, pH 8,0 + 3 M KSCH + 1 M NaCl, и колонку сразу же промывают излишками уравновешивающего буферного раствора в присутствии 0,01% мертиолата при температуре 4oC, если она не будет использоваться в течение длительного времени. Конечный элюат, содержащий антиген, обессоливают на гельфильтрационный колонке с Сефадексом G-25, как было описано выше.
В набухшую колонку, содержащую DEAE целлюлозу (Whatman DE-52), уравновешенную 20 мМ буферным раствором Трис-NCl, pH 8,0 + 5% сахароза, вводят около 2000 мл обессоленного иммуноаффинной очисткой материала и пропускают его через колонку при температуре 4oC. Затем промывают колонку уравновешивающим буфером и контролируют процесс элюции по коэффициенту поглощения при длине волны 280 нм с графической регистрацией. Элюат собирают, пока кривая на хроматограмме не достигнет базовой линии, и затем пропускают 20 мМ буферный раствор Трис-NCl, pH 8,0 + 5% сахарозы + 150 mM NaCl; элюат же собирают до тех пор, пока кривая вновь не достигнет базовой линии на хроматограмме.
Фракцию, полученную при использовании ионообменной колонки в соответствии с описанным выше, подвергают диализу в 100 объемах стерильного PBS и фильтруют через стерилизующую мембрану с размером пор 0,2 мкм.
Для получения препарата, содержащего только антиген в виде частиц, осуществляют хроматографию с молекулярной фильтрацией высокого разрешения и определяют фракцию с высоким молекулярным весом, которая, как известно, обладает наибольшей иммуногенностью.
Поскольку очищенный антиген, как указывалось выше, необходимо соединить с гидроксидом алюминия и (адъювант) подготовить как препарат для инъекций, соблюдают самые строгие меры по выполнению требований, предъявляемых к препаратам парентерального введения, и проводят соответствующий контроль качества (Reguirements for Hepatitis B vaccines made by recombinant DNA technigues, Reguirimenst for Biological Substances N 45, World Health Ordanization, Technical Report Series, N 786, 1989).
Пример 5.
Для получения моноклонального антитела (МКА), специфичного к HBsAg, иммунизируют самцов мышей Balb/c в возрасте 8 недель поверхностным антигеном вируса B, полученным из человеческой плазмы (Krugman S. et al. 1971, J. Am. Med. Assoc. 217: 41). Слияние клеток, выращивание и контроль за гибридными клетками осуществлялись на основе базовых принципов, описанных Келлером (Kholer The Technic of Hybridoma Production, Immunologycal Methods, 1981, Vol 1, Academic Press, 285-298). Клетки селезенки гибридизуют с клетками миеломы SP2/0/Ag14 в соотношении 10:1 с использованием 50%-ного полиэтиленгликоля 1500 (BDH) и вносят в планшеты с 96 ячейками в концентрации 100000 клеток на 1 мл среды и 100 мкл на ячейку. В качестве селективной питательной среды используют RPMI 1640 с добавлением 1 г/л NaHCO3, 3 мкг/л HEPEC, 2 mM L-глутамина, 1 mM пирувата натрия, 0,05 mM 2-меркаптоэтанола, 10% бычьей сыворотки от новорожденных телят, 40 мкг/мл гентамицина, 3% надосадочной жидкости культуры эндотелиальных клеток человека (HECS), (Asta di et al. Methods of Enzymology, 1983, Vol XCII, ed J. Langone, Academic Press, 39-46), гипоксантина до 0,03 mM, тимидина до 3 мкМ и аминоптерина до 0,4 мкМ.
Начиная с третьей недели выращивания гибридных клонов надосадочную жидкость исследуют на специфические антитела против HBsAg с помощью непрямого иммуноферментного метода ELISA. Из положительных ячеек выбирают ячейку N 48 ввиду высоких значений, полученных при повторных испытаниях, и проводят многократное клонирование и вторичное клонирование с применением метода предельных разведений. В качестве целевого отбирают клон 48/1/574, продуцирующий моноклональные антитела со следующими свойствами.
Полученные антитела специфически узнают как природный поверхностный антиген вируса гепатита B (HBsAg), так и рекомбинатный (HBsAgr), полученный в дрожжах. Это подтверждается методом ELISA при использовании обоих антигенов. Антитела узнают HBsAgr как на поверхности твердой фазы, так и в растворе. Ингибирование связывания моноклонального антитела с антигеном на твердой фазе обеспечивается при его предварительной инкубации с антигеном в растворе.
Не наблюдаются перекрестные реакции при использовании ELISA в случае тестирования нескольких молекул, адсорбированных в ячейках планшет. Также регистрируется специфическое узнавание HBsAg, полученного при использовании дрожжей, в Вестерн-блоттинге. Это позволяет утверждать, что полученные моноклональные антитела обладают высокой специфичностью к HBsAg, а узнаваемый эпитоп носит непрерывный характер. Полученные моноклональные антитела узнают общую для всех подвидов вируса гепатита B антигенную детерминанту d.
Своеобразной особенностью клона 48/1/574 является то, что он синтезирует два типа тяжелых цепей: одну подкласса гамма 2b и другую класса M. Клетки секретируют как антитела класса IgG2b, так и пятимерные молекулы IgM, которые могут быть выделены с помощью гельфильтрационной хроматографии на Сефакриле S-300. Указанный факт подтверждается двойной радиальной иммунодиффузией (Ouchterlony O. et al. 1973. Handbook of Experimental Immunology. Vol 1: 19.1) и электрофорезом в полиакриламидном геле (Laemmli N.K. 1970, Nature 227: 680). Оба антитела специфичны к HBsAg, что подтверждается методом ELISA с конъюгатами, специфичными к IgG и IgM. Опыты на пластинах из поливинилхлорида с адсорбированными в ячейках антителами показывают, что моноклональные антитела способны к эффективному захвату HBsAg. Значительная часть последнего (достаточно высокий процент) может быть затем элюирована с помощью высокомолярных растворов тиоцианата калия.
Для производства моноклональных антител, обозначенных CB-HEP I, сначала получают их чистый препарат из асцитной жидкости, извлекаемой после инокуляции 3 миллионов гибридных клеток в перитонеальную полость мышей Balb/c, которым за 10 дней до этого ввели минеральное масло тем же путем. Асцит, собранный в результате повторных пункций, центрифугируют при 500 g, удаляют липиды хлороформом в соотношении 1:1 и диализуют в фосфатном буферном растворе (PBS).
Для очистки используют осаждение 50%-ным NH2SO4 в течение 2 часов при температуре 4oC и центрифугирование со скоростью 4000 об/мин в течение 30 мин. Затем осадок дважды отмывают раствором NH2SO4 одной и той же концентрации, обессоливают на колонке с Сефадексом G-25. Обессоленную фракцию подвергают аффинной хроматографии на протеин A Сефарозе CL48 в соответствии с процедурой, предусмотренной изготовителем.
Очищенные антитела сшивают с Сефарозой CL48, активированной бромистым цианом согласно методу, предложенному изготовителем. Используют 6 мг моноклональных антител на каждый мл геля.
Пример 6.
Для получения вакцинного препарата на основе очищенного рекомбинантного антигена высокой гомогенности и высокой степени агрегирования в частицы его смешивают со стерильным раствором гидроксида алюминия.
В соответствии с указанным готовят несколько партий антигена и исследуют его свойства. В таблице приводятся результаты, полученные при оценке трех партий препарата.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ТЕРАПЕВТИЧЕСКОГО ПРИМЕНЕНИЯ | 2004 |
|
RU2362586C2 |
| ФРАГМЕНТЫ АНТИТЕЛА, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ЧЕЛОВЕЧЕСКОГО РАКОВО-ЭМБРИОНАЛЬНОГО АНТИГЕНА (CEA) | 2003 |
|
RU2294939C2 |
| ХИМЕРНЫЕ ВАКЦИННЫЕ АНТИГЕНЫ ПРОТИВ ВИРУСА КЛАССИЧЕСКОЙ ЧУМЫ СВИНЕЙ | 2007 |
|
RU2406534C2 |
| ПОЛИЭПИТОПНАЯ ВАКЦИНА 4-ГО ПОКОЛЕНИЯ ПРОТИВ ГЕПАТИТА В И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2011 |
|
RU2469741C1 |
| ХИМЕРНЫЙ ГЕН CR3 И КОДИРУЕМЫЙ ИМ ХИМЕРНЫЙ БЕЛОК CR3 (ВАРИАНТЫ), ИНДУЦИРУЮЩИЙ ИММУННЫЙ ОТВЕТ ПРОТИВ ВИЧ-1 | 2002 |
|
RU2302461C2 |
| ВАКЦИННЫЙ СОСТАВ, ПОТЕНЦИРОВАННЫЙ КОМБИНАЦИЕЙ ДНК И АНТИГЕНА | 2002 |
|
RU2294212C2 |
| КОМПОЗИЦИЯ ДЛЯ УХОДА ЗА КОЖЕЙ | 1994 |
|
RU2112503C1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕННЫХ АГРЕГАТОВ И ИХ ПРИМЕНЕНИЕ В ПРЕПАРАТАХ | 2001 |
|
RU2266754C2 |
| МЕТОД ОБНАРУЖЕНИЯ АНТИТЕЛ К ТРАНСГЛУТАМИНАЗЕ ДЛЯ ПРИМЕНЕНИЯ В ДИАГНОСТИКЕ ЦЕЛИАКИИ | 2001 |
|
RU2214835C2 |


Изобретение относится к генной инженерии, в частности к производству вакцин против гепатита В. Для осуществления способа получения поверхностного антигена вируса гепатита В (HBsAg) культивируют штамм дрожжей Pichia pastoris С 226, трансформированный рекомбинантной плазмидой ТАО 906, лизируют клетки и последовательно очищают антигенную фракцию. При очистке используют моноклональные антитела (МКА) СВ-НЕР I. Антиген получают в виде иммуногенных частиц размером 22 нм. Он имеет мол. м. 24000 Да и обладает повышенной гомогенностью. Используемые для иммуноаффинной очистки антигена МКА CB-HEP I продуцируются гибридным клоном 48/1/574, относятся к изотипу IgG2b или IgM, распознают непрерывный эпитоп на общей антигенной детерминанте "а" поверхностного антигена всех подвидов вируса гепатита В. Для получения гибридного клона мышей линии Balb/c иммунизируют HBsAg, далее проводят слияние клеток селезенки с миеломой SP2/0/Ag14, культивируют гибридные клоны в питательной среде и отбирают нужный клон, продуцирующий МКА СВ-НЕРI. 4 с.п.ф-лы, 10 ил., 1 табл.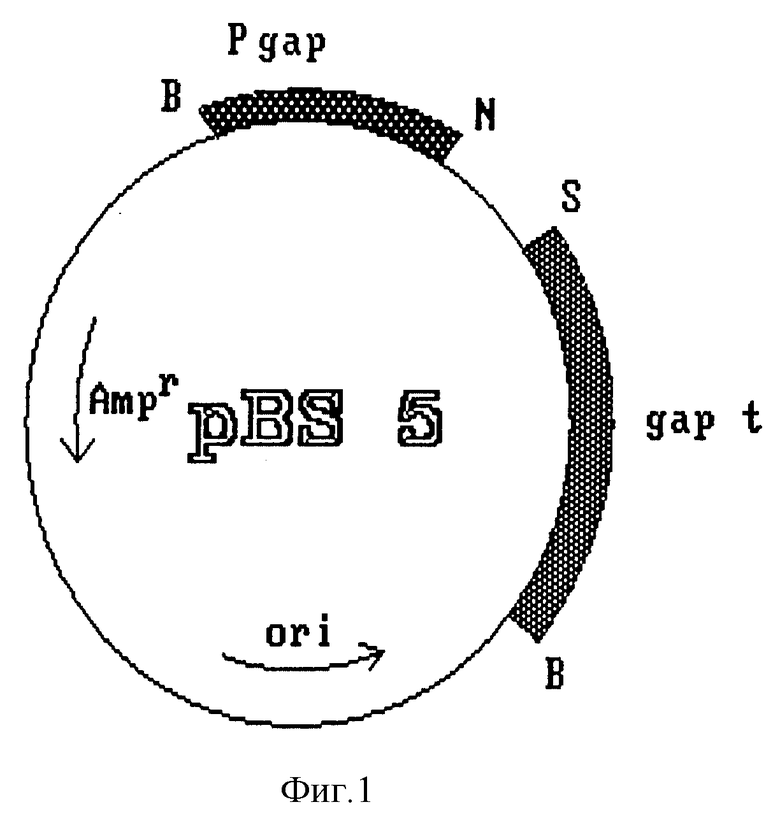
| EP, заявка, 0337492, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |

