Область техники, к которой относится изобретение
Настоящее изобретение относится к области иммунологии и, в частности, к созданию вакцин для предупреждения или лечения синдрома приобретенного иммунодефицита (СПИД). Раскрыты химерные гены и экспрессирующий их вирус оспы кур, пригодные для лечения и предупреждения СПИДа.
Предшествующий уровень техники
ВИЧ является этиологическим агентом СПИДа (Popovic M., Sarngadharan M, Read G. and Gallo RC. Science 1984, 224:497-500). Данный вирус инфицирует не только CD4+-Т-клетки (Klatzman D., Barre Sinoussi F., Nugeyre M.T., Dauguet C., Vilmer E., Griscelli C., Brun-Vezinet F., Rouzioux C., Gluckman J.D., Chermann J.C. and Montagnier L. Science 1984, 225:59-63), но также и другие типы клеток, такие как макрофаги, дендритные клетки, клетки микроглии и эпителиальные клетки.
ВИЧ может уклониться от иммунного ответа хозяина, несмотря на высокий уровень антител, который сохраняется на протяжении всего периода заражения. Впоследствии ВИЧ вызывает у данного хозяина сильный иммунодефицит, который определяет высокую чувствительность к условно-патогенной инфекции.
Свыше 36 миллионов человек живут с ВИЧ/СПИДом и 94% из 16000 ежедневно инфицированных находятся в развивающихся странах (UNAIDS. Report on the global HIV/AIDS epidemic (Сообщение о всемирном эпидемическом ВИЧ/СПИДе), июнь 2000 г.). В связи с этими тревожными цифрами и в отсутствие эффективного и доступного по средствам лечения данного заболевания существует безотлагательная необходимость создания ВИЧ-вакцины.
Из нескольких показателей ВИЧ, которые затрудняют данную задачу, по-видимому, наиболее важным является высокий уровень генетической вариабельности его антигенов, особенно оболочечных гликопротеинов (gp160), в которых располагаются основные домены, участвующие в инфекционном процессе и являющиеся сайтами для нейтрализующих антител.
Вакцины-кандидаты на основе нейтрализующих антител оказались способны защитить шимпанзе от ВИЧ (Berman P.W., Gregory T.J., Lavon R., Nakamura G.R., Champe M.A., Porter J.P., Wurm F.M., Hershberg R.D., Cobb G.K. and Eichberg J.W. Nature 1990, 345:622-625; Girard M., Kieny M.P., Pinter A; Barre-Sinoussi F., Nara P., Kolbe H., Kusumi K., Chaput A., Rainhart T., Muchmore E., Ronco J., Kaczorek M., Gomard E., Gluckman J.C. and Flutz P.N. PNAS 1991, 88:542-546). Однако эти эксперименты были выполнены в почти идеальных условиях, где дозировка, путь введения и продолжительность вирусного заражения сильно отличались от естественного инфицирования. Более того, эти иммуногены не могут защитить от дивергентных изолятов ВИЧ, а полученные антитела оказываются неспособными нейтрализовать первичные изоляты ВИЧ.
Различные вакцины-кандидаты были оценены в клинических опытах Фазы I и II (Johnston M.I. AIDS vaccine development: status and future directions (Создание СПИД-вакцин: состояние и будущие направления). 1999. XII Colloque des Cent Gardes. Ed. Girard M. and Dodet B. 161-163). Большинство из них базируется на оболочечных белках: gp160 и gp120. Лишь одна вакцина, созданная на основе рекомбинантного gp120, проходит в настоящее время испытание на эффективность в опытах Фазы III в Таиланде и США. Результаты предварительных опытов дают основание полагать, что от данной вакцины можно ожидать лишь весьма ограниченную защиту, если таковая вообще имеется.
В связи с этими существенными ограничениями в получении гуморального ответа, способного обеспечить защиту от разных изолятов и субтипов ВИЧ, усилия исследователей в последние годы переключились, в основном, на создание вакцин-кандидатов, способных стимулировать главным образом клеточную часть иммунной системы и особенно цитотоксические Т-клетки, направленные против антигенов ВИЧ.
Экспериментальные данные, которые в значительной мере подтверждают клиническую значимость CTl против ВИЧ, представляют собой следующее: введение моноклональных антител против CD8 макакам, предварительно инокулированным вирусом иммунодефицита обезьяна-человек (SHIV), заметно повышает содержание данного вируса в крови (Matano T., Shibata R., Simeon C., Connors M., Lane C., Martin M. Administration of the Anti-CD8 monoclonal antibody interferes with the clearance of chimeric Simian/Human Immunodeficiency virus during primary infections of rhesus macaques (Введение моноклональных антител к CD8 препятствует выведению химерного вируса иммунодефицита обезьяна/человек при первичном инфицировании макак-резус), J. Virol., 1998, 72, 1:164-169); варианты вируса, способные избегать распознавания CD8+-T-клетками, отбираются у ВИЧ-инфицированных индивидов (Borrow P., H. Lewicki, X. Wei, M.S. Horwitz, N. Peffer, H. Meyers, J.A. Nelson, J.E. Gairin, B.H. Hahn, M.B.A. Oldstone, and G.M. Shaw. 1997. Antiviral pressure exerted by HIV-1-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of CTL escape virus (Противовирусное давление, производимое специфичными для ВИЧ-1 цитотоксическими Т-лимфоцитами (CTL) в течение первичного инфицирования, проявляется в быстром отборе вируса, избегающего CTL). Nature Med. 3:205-211) и у макак, инфицированных SIV (Allen T.M., O.C.DH, P. Jing, J.L. Dzuris, B.R. Mothe, T.U. Vogel, E. Dunphy, M.E. Liebl, C. Emerson, N. Wilson, K.J. Kunstman, X. Wang, D.B. Allison, A.L. Hughes, R.C. Desrosiers, J.D. Altman, S.M. Wolinsky, A. Sette, and D.I. Watkins. 2000. Tat-specific cytotoxic T lymphocytes select for SIV escape variants during resolution of primary viraemia (Tat-специфичные цитотоксические Т-лимфоциты отбирают избегающие SIV-варианты в процессе разрешения первичной виремии). Nature, 407:386-90); совокупность SCID-мышей, содержащих PBMC от добровольцев, инъецированных рекомбинированным вирусом коровьей оспы (VV), кодирующий ВИЧ-1, были защищены от заражения в отсутствие нейтрализующих антител (Van Kuyk R., Torbett B., Gulizia R. et al., Human CTL specific for the nef protein of HIV protect hu-PBL-SCID mice from HIV infection (Человеческие CTL, специфичные для nef-белка ВИЧ, защищают hu-PBL-SCID-мышей от ВИЧ-инфекции). AIDS Res. Hum. Retroviruses, 1993; 9 (suppl. 1:S77); существенная часть подверженных воздействию, но неинфицированных индивидов, проявляет клеточный иммунный ответ, специфичный для ВИЧ-белков, это установлено для шести проституток из Африки (Rowland-Jones S.L., J. Sotton, K. Ariyoshi, T. Dong, F. Gotch, S. McAdams, D. Whitby, S. Sabally, A. Gallimore, T. Corrah, M. Takigushi, T. Schltz, A. McMichael, H. Whittle. 1995. HIV-specific cytotoxic T cells in HIV-exposed but uninfected Gambian woman (ВИЧ-специфичные цитотоксические Т-клетки у подверженных воздействию ВИЧ, но неинфицированных женщин из Гамбии) Nature Medicine, 1:59-64) и для детей, рожденных от сероположительных матерей (Rowland-Jones S.L., D.F. Nixon, M.C. Aldhous, F. Gotch, K. Aroyoshi, N. Hallam, J.S. Kroll, K. Froebel, A. McMichael. HIV specific cytotoxic T-cell activity in an HIV-exposed but uninfected infant (ВИЧ-специфичная цитотоксическая Т-клеточная активность у подвергнутых воздействию ВИЧ, но не инфицированных новорожденных). Lancet, 1993, 341:860-861). Кроме того, длительно непрогрессирующие [индивиды] обнаруживают сильный CTL-ответ (Cao Y., Qin L., Zhang I., Safrit J. and Ho D.D., New Engl. J. Med., 1995, 332:201-208; Riviere Y., McChesney M.B., Porrot E., et al., AIDS Res. Hum. Retroviruses, 11:903-990); а тип HLA класса I ассоциируется с прогрессирующим темпом заболевания у инфицированных ВИЧ-1 индивидов (Carrington M., G.W. Nelson, M.P. Martin, T. Kissner, D. Vlahov, J.J. Goedert, R. Kaslow, S. Buchbinder, K. Hoots, and O.B. SJ. 1999. HLA and HIV-1: heterozygote advantage and B*35-Cw*04 disadvantage (HLA и ВИЧ-1: гетерозиготное преимущество и отсутствие преимущества у B*35-Cw*04). Science, 283:1748-52). CTL-ответ предшествует появлению нейтрализующих антител при естественной инфекции и ассоциируется с регуляцией виремии при острой инфекции (Koup R.A., Safrit J.T., Cao Y., et al., Temporal association of cellular immune responses with the initial control of viremia in primary human immunodeficiency syndrome (Временная ассоциация клеточных иммунных ответов с начальной регуляцией виремии при первичном синдроме иммунодефицита человека), J. Virol., 1994; 68:4650-4655), а развитие СПИДа существенно коррелирует с ослаблением CTL-активности (Harrer T., Harrer E., Kalams S., Elbeik T., Staprans S., Feinberg M.B., Cao Y., Ho D.D., Yilma T., Caliendo A., Jonson R.P., Buchbinder S., and Walker B. HIV-specific CTL-response in healthy long-term asymptomatic HIV infection (ВИЧ-специфичный CTL-ответ у здоровых длительно асимптоматичных по ВИЧ-инфекции). AIDS Res. Hum. Retroviruses, 1996, 12, 7:585-592). Наконец, вакцины, которые индуцируют вирус-специфичный CD8+-Т-клеточный ответ, могут благоприятно повлиять на исход инфекции в SIV-моделях ВИЧ-инфекции (Barouch D.H., S. Santa, J.E.Schmitz, M.J. Kuroda, T.M. Fu, W. Wagner, M. Bilska, A. Craiu, X.X. Zheng, G.R. Krivulka, K. Beaudry, M.A. Lifton, C.E. Nickerson, W.L. Trigona, K. Punt, D.C. Freed, L. Guan, S. Dubey, D. Casimiro, A. Simon, M.E. Davies, M. Chastain, T.B. Strom, R.S. Gelman, D.C. Montefiori, M.G. Lewis, E.A. Emini, J.W. Shiver, and N.L. Letvin. 2000. Control viremia and prevention of clinical AIDS in rhesus monkeys by cytokine-augmented DNA vaccination (Контроль за виремией и предотвращение клинического проявления СПИДа у макак-резус с помощью усиленной цитокином ДНК-вакцинации). Science. 290:486-92; Gallimore A., M. Cranage, N. Cook, N. Almond, J. Bootman, E. Rud, P. Silvera, M. Dennis, T. Corcoran, J. Stott, A. McMichael, and F. Gotch. 1995. Early suppression of SIV replication by CD8+ nef-specific cytotoxic T cells in vaccinated macaques (Ранняя супрессия репликации SIV с помощью nef-специфичных цитотоксичных CD8+-Т-клеток). Nature Med. 1:1167-1173).
Все эти экспериментальные данные убедительно свидетельствует о том, что терапевтические и профилактические подходы должны включать индукцию/сохранение/восстановление этой части иммунного ответа, по меньшей мере, в качестве одной из их целей.
Для выработки CTL у животных и человека были разработаны разные методы. До сих пор наиболее эффективными были рекомбинантные живые векторы. В данном способе используют безвредные вирусы или бактерии для переноса избранных генов из данного патогена в определенные клетки реципиента для наработки там выбранных антигенов. Данная методика доставки гена в клетки максимизирует процессинг CTL-эпитопов и их презентацию в молекулах MHC-I и последующую эффективную стимуляцию CTL-клонов в данном хозяине.
Вирусы, которые наиболее успешно использовались в качестве векторов, представляли собой поксвирусы (семейство Poxviridae). Наиболее известным членом данного семейства является вирус коровьей оспы (VV), который широко использовали на людях во время проведения операции по искоренению оспы.
Осуществлено несколько клинических исследований рекомбинированных VV, кодирующих белки ВИЧ (Corey L., McElroth, Weihold K., Matthewa T., Stablein D., Grahm B., Keefer M., Schwartz D., Gorse G. Cytotoxic T Cell and Neutralizing Antibody Responses to Human Immunodeficiency Virus Type 1 Envelope with a combination vaccine regimen (Ответы в виде Цитотоксических Т-клеток и нейтрализующих антител на оболочку вируса иммунодефицита человека типа 1 при комбинированной схеме вакцинации). J. Infectious Dis., 1998, 177:301-9; Graham B.S., Matthews T.J., Belshe R., Clements M.L., Dolin R., Wright P.F., Gorse G.L., Schwartz D.H., Keefer M.C., Bolognesi D.P., Corey L., Stablein D., Esterlitz J.R., Hu S.L., Smith G.E., Fast P., Koff W., J. Infectious Dis., 1993, 167:533-7). Однако VV обладает двумя основными ограничениями для использования на человеке: (1) небольшой процент вакцинированных лиц демонстрировал сильные неблагоприятные реакции, которые могут привести к летальному исходу в случае индивидов с нарушенным иммунитетом; (2) лица с VV-вакцинацией в прошлом слабо реагируют на гетерологичные антигены.
Для преодоления этих недостатков вместо VV использовали представителей Avipoxvirus. Они являются членами семейства поксвирусов, но их репликация ограничена клетками птиц, а в человеческих клетках их цикл репликации прерывается. Для этих целей использовали два представителя поксвирусов птиц: вирус оспы канарейки (CPV) и вирус оспы кур (FPV).
Рекомбинантные поксвирусы птиц, кодирующие разные патогены человека опухолеспецифичных антигенов, индуцируют CTL-ответ у животных (Limbach K.J. and E. Paoletti. 1996. Non-replicating expression vectors: applications in vaccines development and gene therapy (Нереплицирующиеся экспрессионные векторы: применение для создания вакцин и генной терапии). Epidemiol. Infect. 116:241-256). Использование рекомбинантного поксвируса птиц для создания вакцины запатентовано в США (Paoletti E. y cols 1992 US5174993, Paoletti E. и соавт., 1993, US5505941), и специальная патентная заявка, относящаяся к использованию рекомбинантных поксвирусов птиц, кодирующих лентивирусные антигены, были представлены в Европе (Paoletti E. и соавт., ЕР0956360).
CPV, рекомбинантный по gag, pol и env ВИЧ-1, оценивали в испытаниях Фазы I и II на здоровых добровольцах (Clements-Mann M.L., K. Weinhold, T.J. Matthews, B.S. Graham, G.L. Gorse, M.C. Keefer, M.J. McElrath, R.-H. Hsieh, J. Mestecky, S. Zolla-Pazner, J. Mascola, D. Schwartz, R. Siliciano, L. Corey, P.F. Wright, R. Belshe, R. Dolin, S. Jackson, S. Xu, P. Fast, M.C. Walker, D. Stablein, J.-L. Excler, J. Tartaglia, A.-M. Duliege, F. Sinangil, E. Paoletti. 1998. Immune responses to Human Immunodeficiency Virus (HIV) Type 1 induced by Canarypox expressing HIV-1MN gp120, HIV-1SF2 recombinant gp120, or both vaccines in seronegative adults (Иммунные ответы у серонегативных взрослых на вирус иммунодефицита человека (ВИЧ) типа 1, индуцированные поксвирусом оспы канарейки, экспрессирующим gp120 ВИЧ-1MN, рекомбинантным gp120 ВИЧ-1SF2, или обоими вакцинами). J. Infect. Dis. 177:1230-1246; Egan M.A., W.A. Pavlat, J. Tartaglia, E. Paoletti, K.J. Weinhold, M.L. Clements, R.F. Siliciano. 1955. Induction of Human Immunodeficiency Virus Type 1 (HIV-1)-specific cytolytic T lymphocyte responses in seronegative adults by a nonreplicating, host-range-restricted canarypox vector (ALVAC) carrying the HIV-1MN env gene (Индукция у серонегативных взрослых ответов в виде цитолитических Т-лимфоцитов, специфичных к вирусу иммунодефицита человека (ВИЧ) типа 1, с помощью нереплицирующегося, ограниченного рядом хозяев вектора, связанного с поксвирусом оспы канарейки (ALVAC), несущего ген env ВИЧ-1MN). J. Infect. Dis. 171:1623-1627). CTL против, по меньшей мере, одного ВИЧ-антигена были обнаружены у 50% вакцинированных в испытании Фазы I, у 30% в испытании Фазы II и менее 10% в испытании последней Фазы I в Уганде. Этот rCPV (vCP205) создали путем встраивания ВИЧ-генов в три разные несущественные области данного генома для получения CTL-ответа против более чем одной ВИЧ-мишени.
С другой стороны, для индукции у макак CTL-ответа против ВИЧ-антигенов также использовали FPV в сочетании с ДНК-иммунизацией (Robinson H.L., D.C. Montefiori, R.P. Johnson, K.H. Manson, M.L. Kalish, J.D. Lifson, T.A. Rizvi, S. Lu, S.-L. Hu, G.P. Mazzara, D.L. Panicali, J.G. Herndon, R. Glickmanm, M.A. Candido, S.L. Lydy, M.S. Wyand and H.M. McClure. 1999. Nature Medicine, 5:526-534). Данное сочетание иммуногенов создавало определенный уровень защиты для инфекционной модели ВИЧ-1/macaca nemestrina (Kent S.J., A. Zhao, S.J. Best, J.D. Chandler, D.B. Boyle, I.A. Ramshaw. Enhanced T-Cell immunogenicity and protective efficacy of a human immunodeficiency virus type 1 vaccine regime consisting of a consecutive priming with DNA and boosting with recombinant fowlpox virus (Усиление Т-клеточной иммуногенности и защитной эффективности схемы вакцинации против вируса иммунодефицита типа 1, состоящей из последовательного праймирования с помощью ДНК и повторной иммунизации с помощью рекомбинантного поксвируса оспы кур). 1998. J. Virol., 72:10180-10188). Однако данная животная модель содержит существенные ограничения, так как ВИЧ-инфицирование у M.nemestrina является неэффективным и трудным для воспроизведения.
Сообщено также о получении CTL-ответа в результате иммунизации минигенами, состоящими из ряда точных CTL-эпитопов нескольких патогенов (Whitton L., Sheng N., Oldstone M.B., and McKee T. A "string of beads" vaccine, comprising linked minigenes, confers protection from lethal-dose virus challenge (Вакцина "нитка бус", содержащая связанные минигены, обеспечивает защиту от вирусного заражения летальной дозой), J. Virol., 1993, 67, 1:348-352; A multivalent minigene vaccine, containing B-cell, cytotoxic T-lymphocyte and Th epitopes from several microbes, induces appropriate responses in vivo and confers protection against more than one pathogen (Мультивалентная минигенная вакцина, содержащая В-клетки, цитотоксические Т-лимфоциты и Th-эпитопы от нескольких микробов, индуцирует соответствующие ответы in vivo и обеспечивает защиту от более чем одного патогена). J. Virol., 71, 3:2292-2302).
Модифицированный рекомбинантный вирус коровьей оспы Анкара (MVA), кодирующий gag-производное минигена вместе с целым gag-геном, использовали для индукции CTL-ответа у мышей (Hanke T., R.V. Samuel, T.J. Blanchard, V.C. Neumann, T.M. Allen, J.E. Boyson, S.A. Sharpe, N. Cook, G.L. Smith, D.I. Watkins, M.P. Cranage, A.J. McMichael. 1999. Effective induction of simian immunodeficiency virus-specific cytotoxic T lymphocytes in macaques by using a multiepitope gene and DNA prime-Modified Vaccinia Virus Ankara boost vaccination regimen (Эффективная индукция у макак цитотоксических Т-лимфоцитов, специфичных к вирусу иммунодефицита обезьян, путем использования модифицированного с помощью мультиэпитопного гена и ДНК-затравки вируса коровьей оспы Анкара в режиме повторной вакцинации). J. Virol., 73, 9:7524-7532). Эти минигены состоят из цепочки дискретных CTL-эпитопов gag.
Основное ограничение минигенного подхода заключается в том, что сочетание индивидуальных CTL-эпитопов охватывает лишь ограниченный ряд HLA-антигенов и поэтому возбуждаемый CTL-ответ является по определению более ограниченным.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Сущность настоящего изобретения заключается в создании конструкции химерных генов, составленной из участков ВИЧ-белков, богатых CTL-эпитопами, где эти участки выбраны из внутренних консервативных белков и регуляторных белков, экспрессируемых очень рано в жизненном цикле вирусов.
Данный метод решения проблемы обладает преимуществами перед описанными ВИЧ-минигенами, так как позволяет одновременно осуществлять процессинг перекрывающихся CTL-эпитопов, представленных многими HLA-аллелями. Другое преимущество данного метода решения, в сравнении с другими рекомбинантными поксвирусами птиц, кодирующими несколько белков ВИЧ-1, заключается в том, что сосредоточение иммунологически значимых областей из нескольких белков в одном гене способствует образованию рекомбинантных вирусов и исключает необходимость в использовании нескольких систем резистентности к антибиотикам в одном и том же рекомбинантном вирусе. Кроме того, облегчается объединение эпитопов от нескольких субтипов ВИЧ в единственном рекомбинантном вирусе. Выбранные области принадлежат наиболее консервативным вирусным белкам и рано экспрессируемым регуляторным продуктам. Эти области, богатые CTL-эпитопами, объединяют с консервативными эпитопами Т-хелперных клеток, фланкированными с помощью двух лизинов, чтобы облегчить их процессинг клеточными протеазами. И в заключение добавляют В-клеточный эпитоп, распознаваемый моноклональным антителом, для облегчения детекции данного полипептида иммунохимическими методами.
Данный химерный ген собирают, соединяя вместе разные фрагменты ДНК, некоторые из них получены химическим синтезом, а другие амплифицированы с помощью полимеразной цепной реакцией (ПЦР) с использованием ВИЧ-генов в качестве матриц. Фрагменты ДНК клонируют совместно в соответствующий плазмидный вектор, секвенируют и переносят в поксвирусный рекомбинантный вектор.
Более конкретно, настоящее изобретение относится к гену cr3, который содержит Th-клеточные эпитопы белков ВИЧ-1 - gp120, gp41 и Vpr, эпитоп V3-петли gp120, распознаваемый Mab 2С4 (Duarte C.A., Pérez L., Vázquez J., Duecas M., Vilarubia O.L., Navea L., Valdés R., Reyes O., Montero M., Ayala M., and Gavilondo J. Epitope mapping, V region DNA sequence, and neutralizing Fab fragments of two monoclonal antibodies against HIV-1 V3 loop (Картирование эпитопов, ДНК-последовательность V-области и нейтрализующие Fab-фрагменты двух моноклональных антител к V3-петле ВИЧ-1). Immunotechnology 1996, 2:11-20), и богатые CTL-эпитопами области на белках RT, Gag и Nef.
Эти химерные гены встраивают в геном бактериального или вирусного живого вектора (например, поксвируса, герпесвируса, альфавируса, полиовируса, аденовируса, BCG, Salmonella), предпочтительно данный вектор представляет собой поксвирус, более точно поксвирус птиц и точнее FPV. Эти рекомбинантные живые векторы используют для индукции у животных и человека TH1-иммунного ответа и цитотоксических Т-клеток против ВИЧ.
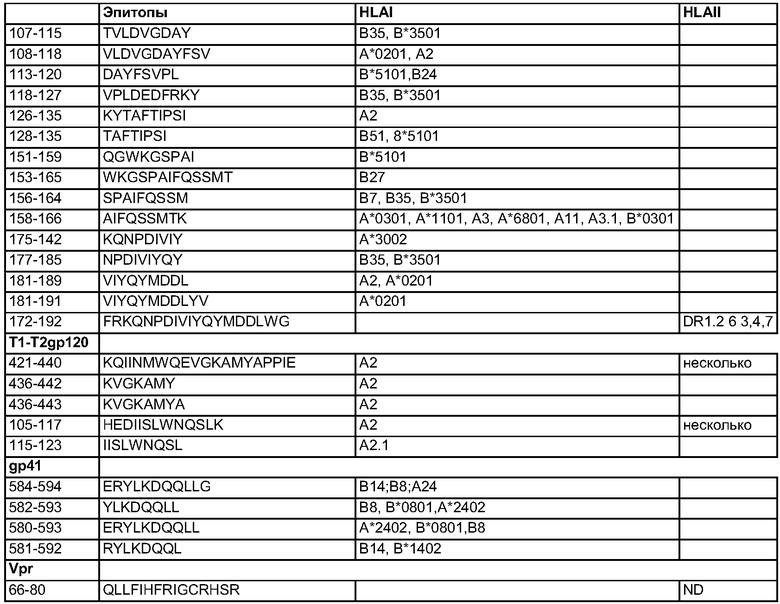
Совсем точно, настоящее изобретение относится к рекомбинантному FPV, кодирующему эти химерные белки, и, в частности, к рекомбинантным штаммам FPV, названным FPCR3 и FPSCR3gpt, которые содержат химерный ген cr3. После сборки, как указано выше, cr3 клонируют в рекомбинантный поксвирусный вектор, в частности FPV-рекомбинантный вектор. В данном конкретном случае в качестве рекомбинантных векторов использовали плазмиды pEFL29 и pFP67xgpt. pEFL29 представляет области, гомологичные терминальному BamHI фрагменту размером 6 т.п.н. из генома FPB, который фланкирует транскрипционный элемент, в который встроен гетерологичный ген под контролем 7.5K-промотора VV, и содержит также репортерный ген lacZпод контролем 4b-промотора FPV. pFP67xgpt использует открытые рамки считывания 6 и 7 из BamHI-области размером 11.2 т.п.н. в качестве сигналов гомологичной рекомбинации. Эти области фланкируют транскрипционный элемент, в котором данный гетерологичный ген помещен под синтетический поксвирусный E/L-промотор, и он также содержит gpt-ген, который придает устойчивость к микофеноловой кислоте, что позволяет селектировать рекомбинантные вирусы.
Полученные плазмиды были названы, соответственно, pFPCR3 и pFPSCR3gpt. Этими плазмидами трансфицируют первичную культуру эмбриональных фибробластов цыпленка (CEF) с использованием одной из нескольких методик трансфекции, доступных в данной области техники. В данном конкретном случае трансфекцию осуществляют с использованием липофектина (Sigma, США) в CEF, предварительно инфицированных штаммом FP29 FPV, но могут использоваться и другие способы, такие как электропорация и DEAE-декстран. В результате гомологичной рекомбинации между плазмидой и соответствующими несущественными областями FPV-генома, в случае с pFPCR3 можно выделить рекомбинантные вирусы, которые экспрессируют β-галактозидазу, или в случае pFPSCR3gpt - рекомбинантные вирусы, устойчивые к микофеноловой кислоте. Присутствие селективного маркера позволяет идентифицировать рекомбинантные вирусные бляшки и очистить их с помощью нескольких пассажей в CEF. Наличие данного гетерологичного гена в отселектированных вирусах можно проверить с помощью ПЦР, а экспрессию данного белка можно проверить с помощью Вестерн-блота.
Настоящее изобретение относится также к использованию рекомбинантного FPV, полученного как указано, чтобы индуцировать TH1-иммунный ответ вместе с CTL-активностью у Balb/c-мышей, либо отдельно, либо в сочетании с фармацевтически приемлемой композицией, выбранной из известных в данной области техники композиций.
Настоящее изобретение относится также к терапевтическому или профилактическому сочетанию рекомбинантного по описанным химерным генам FPV, в частности FPCR3 и FPSCR3gpt, и иммуномодуляторов или адъювантов, в частности цитокинов, таких как IL2, IL12, IFNγ, GMSCG, GSCF, которые предпочтительно стимулируют TH1-иммунный ответ.
В особенности, настоящее изобретение относится к сочетанию вирусов FPCR3 или FPSCR3gpt с ежедневными дозами IL2 в диапазоне между 102 и 107 МЕ для животных и человека. Ежедневное введение IL2 Balb/c-мышам начинают со дня введения FPV или после стимулирования клеточного иммунного ответа против CR3.
Хотя настоящее изобретение относится, в частности, к CR3, сущность настоящего изобретения состоит в том, что можно также использовать CTL-богатые фрагменты, отличные от CTL-богатых фрагментов CR3, или фрагменты, эквивалентные CTL-богатым фрагментам CR3, но из других изолятов ВИЧ-1.
Точно также, хотя настоящее изобретение относится, в частности, к FP9-штамму FPV, сущность данного изобретения состоит в том, что можно использовать и другие родительские штаммы FPV для создания рекомбинантных вирусов, а также другой поксвирус птиц, такой как CPV, другой поксвирус, такой как VV или MVA, либо совсем другие вирусы, например, такие как герпесвирус, альфавирус, аденовирус, полиовирус, либо даже бактерии, такие как BCG или Salmonella.
В другом варианте осуществления настоящего изобретения данный ген можно клонировать в подходящий плазмидный вектор для экспрессии в клетках млекопитающих и инъецировать млекопитающему для индукции TH1-иммунного ответа и CTL-активности в сочетании с фармацевтически приемлемым носителем.
Еще в одном варианте осуществления настоящего изобретения оно также включает терапевтическую или профилактическую комбинацию вышеописанных рекомбинантных плазмид с иммуномодуляторами или адъювантами, которые описаны, или с иными, такими как липосомы, полисахариды, липопептиды, липиды, протеолипосомы или с их сочетаниями.
Еще в одном варианте осуществления настоящего изобретения указанные гены можно клонировать в другие плазмиды для экспрессии рекомбинантных белков в клетках бактерий, дрожжей, грибов, насекомых или млекопитающих, растений или в молоке трансгенных животных. Белки, выделенные из этих систем, могли бы также использоваться для индукции TH1-иммунного ответа и CTL-активности у животных или человека, при введении в фармацевтически приемлемом носителе.
Еще в одном варианте осуществления настоящего изобретения терапевтические или профилактические сочетания CR3-белка с иммуномодуляторами или адъювантами, такими как описанные выше, или и вовсе с другими, такими как липосомы, полисахариды, липиды, протеолипосомы или иными адъювантами, имеющимися в данной области техники, способны усилить у животных или человека иммунный ответ типа TH1 и CTL-активность.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1. Плазмида pEFL-cr3 для гомологичной рекомбинации в вирусе оспы кур с использованием ORF-1 из терминальной BamH1-области размером 6 т.п.н. в качестве вставочного сайта. Ген cr3 находится под контролем p7.5K-промотора VV, а репортерный ген LacZ - под 4b-промотором FPV.
Фигура 2. Плазмида pFP67xgpt для гомологичной рекомбинации в FPV с использованием участка ДНК между ORF-6 и ORF-7 из BamHI-фрагмента размером 11.2 т.п.н. в качестве вставочного сайта. Ген cr3 помещен под контроль синтетического промотора E/L, а ген Ecogpt - под контроль 7.5K-промотора VV.
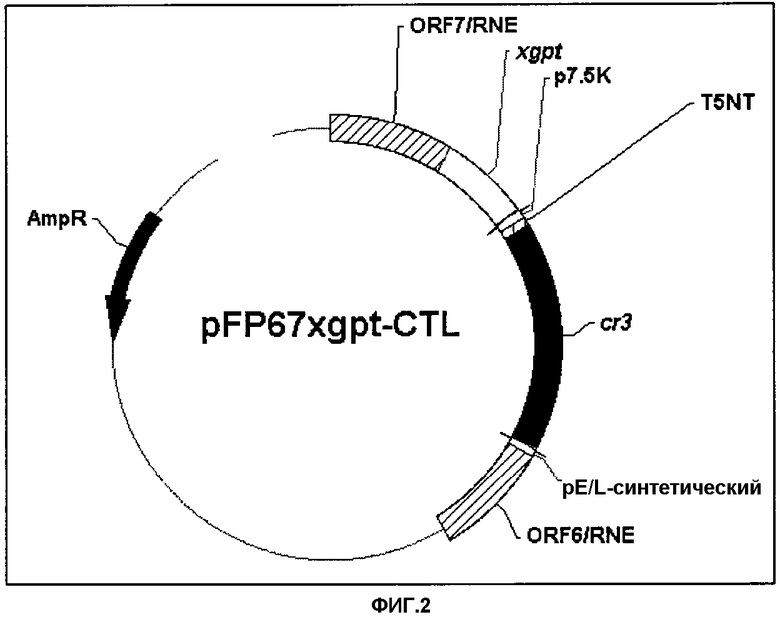
Фигура 3. (А) ПЦР и (В) Вестерн-блот трех независимых cr3-рекомбинантных FPV: (1) FPCR3.1; (2) FPCR3.2; (3) FPCR3.3; (4) FPL29; (5) маркер молекулярной массы ДНК.
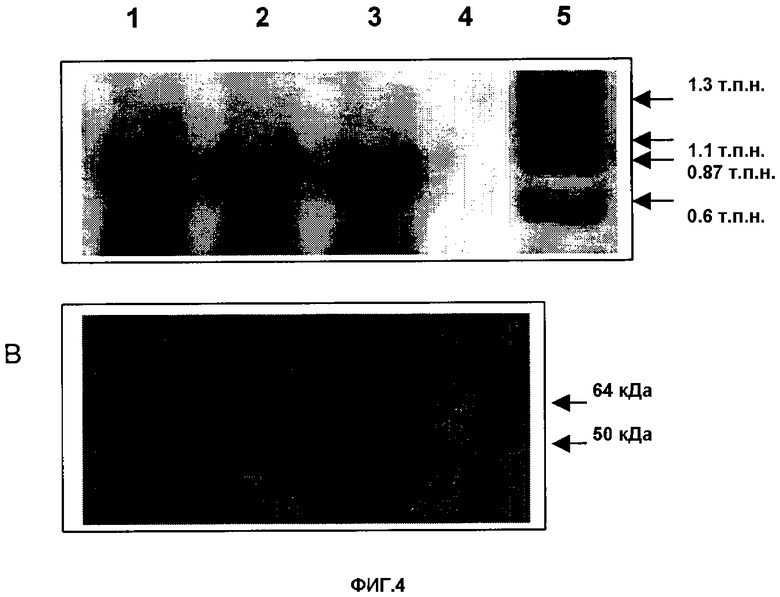
Фигура 4. (А) ПЦР с cr3-внутренними олигонуклеотидами (В) Вестерн-блот трех независимых cr3-рекомбинантов FPV (1) FPSCR3GPT.1; (2) FPSCR3GPT.2; (3) FPSCR3GPT.3; (4) родительский вирус; (5) маркер молекулярной массы ДНК.
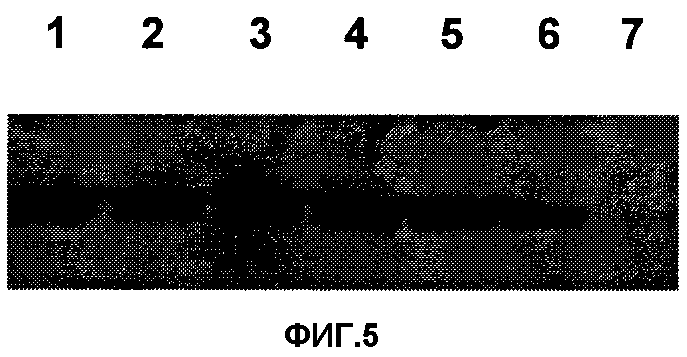
Фигура 5. Оценка стабильности экспрессии CR3 с помощью Вестерн-блота. Дорожки соответствуют трем независимым образцам CEF, инфицированным FPSCR3GPT из вирусного запаса (1, 2, 3) или выделенного очисткой через сахарозную подушку (4, 5, 6). Дорожка 7 представляет CEF, инфицированные родительским вирусом.
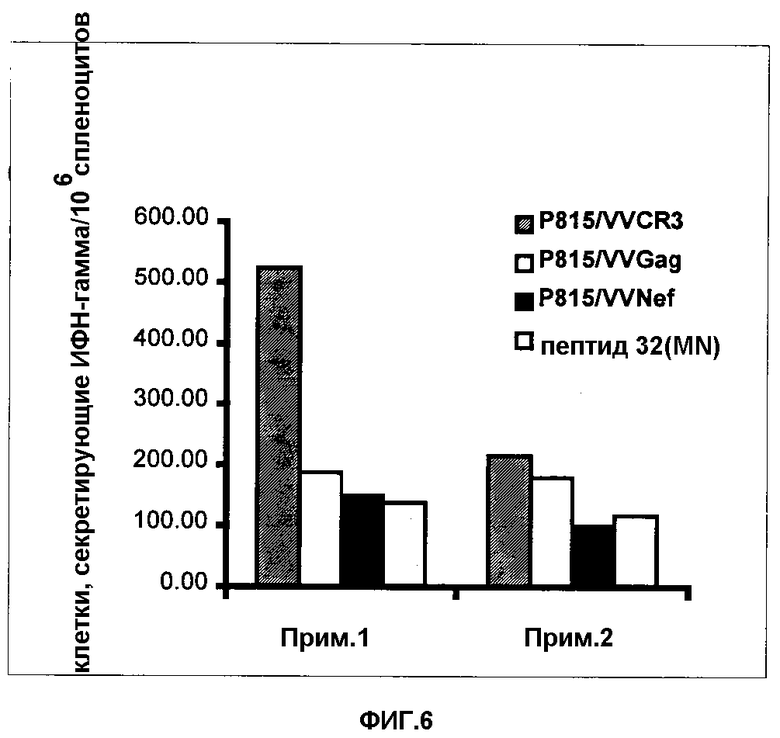
Фигура 6. Результаты двух независимых ELISPOT-экспериментов с использованием спленоцитов мышей, иммунизированных FPSCR3gpt, и клеток P815, нагруженных пептидом 32 или инфицированных VV, рекомбинантным по CR3, Gag или Nef. Данные результаты представлены в виде числа клеток, секретирующих гамма-ИФН, на 106 спленоцитов. Значения соответствующих негативных контролей (VV инфицированный WR или только P815) вычитались.
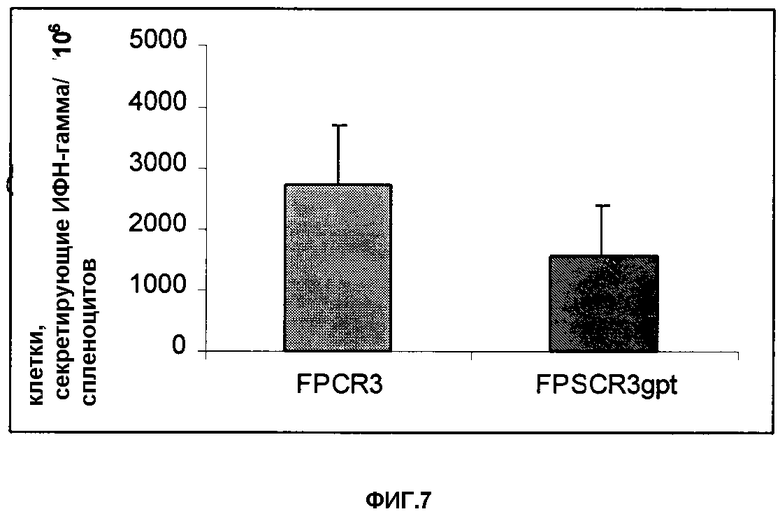
Фигура 7. Гамма-ИФН-ELISPOT-эксперименты с использованием спленоцитов мышей, иммунизированных FPCR3 или FPSCR3gpt, и Р815, стабильно трансфицированные геном cr3. Полученные результаты представлены в виде числа клеток, секретирующих гамма-ИФН на 106 спленоцитов. Значения негативных контролей (родительские Р815) вычитались.
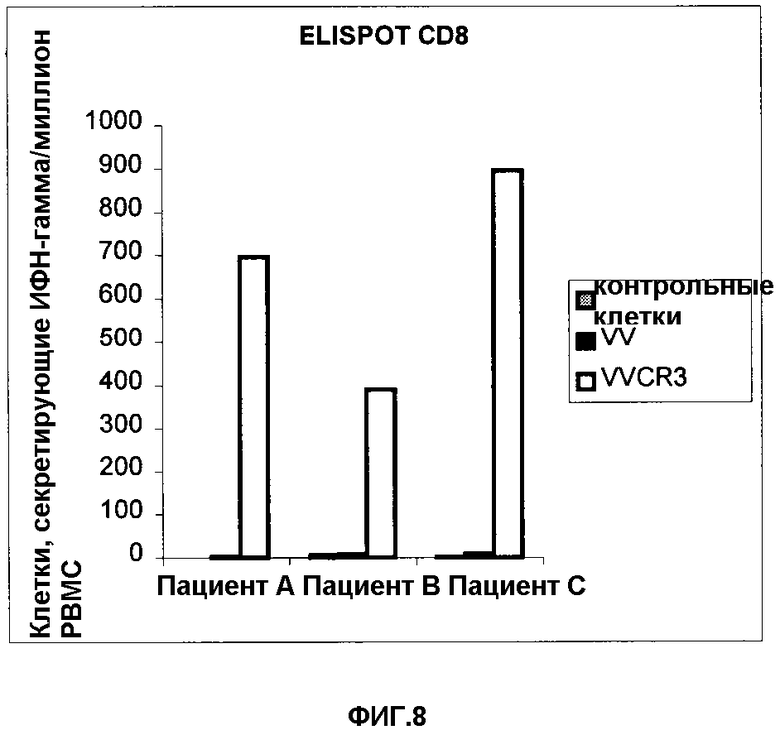
Фигура 8. Распознавание VVCR3-инфицированных аутологичных В-клеток Т-лифоцитами больных СПИДом. Результаты IFNγ-ELISPOT представлены в виде числа клеток, секретирующих гамма-ИФН на 106 мононуклеарных клеток периферической крови.
ПРИМЕРЫ
Пример 1. Получение cr3.
cr3 представляет собой химерный ген, собранный из фрагментов разных ВИЧ-генов. Его собрали в плазмиде ТАВ11, которая фактически идентична рТАВ9 (Gymez C.E., Navea L., Lobaina L., Dubed M., Expysito N., Soto A. and Duarte C.A. The V3 loop based Multi-Epitope Polypeptide TAB9 Adjuvated with Montanide ISA720 is Highly Immunogenic in Nonhuman Primates and Induces Neutralizing Antibodies Against Five HIV-1 isolates (V3-петля мультиэпитопного полипептида ТАВ9, дополненная Montanide ISA720, у приматов, не принадлежащих человеческому роду, является весьма иммуногенной и индуцирует нейтрализующие антитела против пяти ВИЧ-изолятов). Vaccine 17:2311-2319, 1999), но обладает эпитопами Т-хелперных клеток Т1 и Т2 из gp120 в самом 5'-конце данного гена вместо фрагмента, кодирующего N-терминальную часть белка Р64K. Затупленный по концам синтетический BamHI-фрагмент ДНК размером 186 п.н., кодирующий Т2-эпитоп из gp120, V3-эпитоп MN-штамма, и эпитопы Т-хелперных клеток из gp41 и vpr, клонировали в ТАВ11, предварительно расщепленную EcoRV-BamHI. ДНК-последовательности, кодирующие два последовательных лизина, встраивали между индивидуальными эпитопами, чтобы способствовать внутриклеточному процессингу. Полученную плазмиду назвали pCR1. (603 п.н.)-фрагмент, кодирующий белок (RT) р66/р51 (поз. 2663-3109 из SF2-провируса ВИЧ-1), амплифицировали с помощью ПЦР с использованием праймеров 0,2660 и 0,2661 (таблица 1). Полученный ПЦР-фрагмент, экстрагированный из легкоплавкой агарозы, расщепляли с помощью BglII-EcoRI и субклонировали в резанный BglII-EcoRI pCR1-вектор для получения pCR2-плазмиды, кодирующей CR2-белок. Затем, (324 п.н.)-фрагмент, включающий последовательность nef-гена (поз. 8516-8818 из LAI-изолята ВИЧ), подвергали ПЦР-амплификации с праймерами 0,2662 и 0,2663. Наконец, другой (267 п.н.)-сегмент в gag-гене (поз. 1451-1696 из SF2 ВИЧ) амплифицировали с использованием праймеров 0,2664 и 0,2665 (таблица 1). Затем осуществляли перекрывающуюся ПЦР с использованием 20 пмоль праймеров 0,2662 и 0,2666 (таблица 1). Равные количества каждой полосы (0,47 пмоль) смешивали в ПЦР-буфере [50 мМ KCl; 10 мМ Трис-HCl, (рН 8,3), при 25°С; 0,001 % желатин], 2,5 мМ MgCl2, 0,2 мМ dNTP (каждого) и 4 Ед. Taq-полимеразы в объеме 50 мкл. Для ускорения отжига полос по комплементарным (9 п.н.)-концам олигонуклеотидов 0,2663 и 0,2664 смесь вначале нагревали при 92°С в течение 2 мин, а затем охлаждали при 50°С. В заключение температуру повышали до 72°С в течение 5 мин для удлинения отожженных сегментов. Позже, 10 мкл вышеуказанной реакционной смеси вносили в ПЦР-буферную смесь, содержащую 2,5 мМ MgCl2, 0,2 мМ dNTPs, 20 пмоль О,2662 и 20 пмоль 0,2666 и 4 Ед. Vent-пол. в общем объеме 50 мкл. Стандартными условиями амплификации являются 92°С в течение 2 мин, с последующими 30 циклами при 92°С в течение 40 с, 50°С в течение 1 мин, и 72°С в течение 1 мин и окончательное удлинение при 72°С в течение 5 мин. Позже перекрывающуюся амплифицированную полосу nef-p24 выделяли очисткой из подвергнутой электрофорезу легкоплавкой агарозы и обрабатывали с помощью Xbal. В конечном итоге, первую затупленную Xbal-полосу клонировали в pCR2 вектор, предварительно резанный Nrul-Xbal, для получения pCR3-плазмиды. Таким образом, cr3 кодирует химерные белки, которые включают эпитопы Т-хелперных клеток и CTL-эпитопы из gp120, gp41, vpr, RT, nef и gag, представленные широким рядом HLA-антигенов (таблица 2).
ДНК-ПОСЛЕДОВАТЕЛЬНОСТИ ОЛИГОНУКЛЕОТИДОВ, ИСПОЛЬЗУЕМЫЕ В ПЦР-РЕАКЦИЯХ
Числа соответствуют позициям, связанным с HXB2-аминокислотной последовательностью каждого вирусного белка, вирусный изолят указан в скобках; ND не определен.
Пример 2. Клонирование cr3 в pFPL29.
В pCR3 ген cr3 был клонирован под контролем pTryp, с ClaI-сайтом на 5'-конце, терминатором гена 32 фага Т4 и HindIII-сайтом - на 3'-конце. Данную плазмиду расщепляли Clal и HindIII и обрабатывали фрагментом Кленова I с получением cr3-гена с ATG на 5'-конце и стоп-кодонами трансляции на 3'-конце. Данный фрагмент ДНК клонировали в поксвирусный рекомбинантный вектор pEFL29.
pEFL29 обладает терминальным BamHI-фрагментом размером 6 т.п.н. FPV в качестве несущественных областей для гомологичной рекомбинации в геноме FPV. Этот фрагмент содержит три ORF, и ORF1 разорвана. Фланкированными этими гомологичными областями являются p7.5K VV, промотор, за которым следует Smal-сайт, и репортерный ген lacZ под контролем позднего 4b-промотора из FPV. Данная плазмида включает также ген устойчивости к канамицину и бактериальный ориджин репликации.
PEFL29 расщепляли с помощью Smal, обрабатывали щелочной фосфатазой и лигировали с полосой, расщепленной Clal/HindIII, содержащей ген cr3. Отбирали несколько клонов с cr3 в правильной ориентации относительно 7.5K. Штамм DH5α E. coli (φ80dlacZΔM15, recA1, endA1, gyrA96, thi-1, hsdR17 (rK- mK+), supE44, relA1, deoR, Δ(lacZYA-argF)U169) использовали для размножения и селекции рекомбинантных плазмид в LB-среде, содержащей канамицин (25 мкг/мл). Все генетические операции осуществляли в соответствии с Sambrook и соавт. (Sambrook J., Fritsch E.F., Maniatis T. 1989. Molecular Cloning. A Laboratory Manual (Молекулярное клонирование. Лабораторное руководство). Sec. Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.).
Секвенированную ДНК клона pEFL-cr3 (Фигура 1) проверяли с использованием автоматического сиквенатора (Pharmacia). Данный клон выделяли очисткой с использованием градиента CsCl и использовали для трансфицирования фибробластов эмбриона цыпленка (CTF).
Пример 3. Клонирование cr3 в pFP67xgpt.
Ген cr3 амплифицировали с помощью ПЦР и клонировали в pMosblue-вектор (Amersham, Великобритания). Полученную плазмиду назвали pTCR3. Плазмиду pTCR3 расщепляли с помощью Hpal/BamHI и нижнюю полосу, содержащую cr3, клонировали в поксвирусный вектор pFP67xgpt.
Вектор pFP67xgpt обладает BamH1-фрагментом размером 11,2 т.п.н. из генома FPV в качестве несущественной области для гомологичной рекомбинации в геноме FPV (Tomley F., Binns M., Campwell J., Boursnell M. Sequence Analysis of an 11.2 Kilobase, near-terminal, Bam HI fragment of fowlpox virus (Анализ последовательности около концевого BamHI-фрагмента размером 11,2 т.п.н. поксвируса оспы кур), J. Gen. Virol., 1988, 69, 1025-1040). Данный фрагмент содержит открытую рамку считывания 6 и 7 из этой области, а встраивание происходит в межгенную область. Фланкированными этими гомологичными областями являются синтетический E/L-промотор (Carrol M.W., Moss B.E. E.coli β-glucoronidase (GUS) as a marker for recombinant vaccinia viruses (β-глюкоронидаза E.coli (GUS) в качестве маркера рекомбинантных вирусов коровьей оспы), Biotechniques, 1995, 19, 3:352-354), и репортерный ген Ecogpt под контролем 7.5K-промотора VV. Данная плазмида включает также ген устойчивости к канамицину и бактериальный ориджин репликации.
Плазмиду pFP67xgp расщепляли с помощью Stul/BamHI и лигировали с cr3, содержащим ДНК-фрагмент, полученный при Hpal/BamHI-расщеплении pTCR3. Отбирали несколько клонов с cr3 в надлежащей ориентации относительно синтетического E/L-промотора (Фигура 1). Штамм DH5α E. coli (φ80dlacZΔM15, recA1, endA1, gyrA96, thi-1, hsdR17 (rK- mK+), supE44, relA1, deoR, Δ(lacZYA-argF)U169) использовали для размножения и селекции рекомбинантных плазмид в LB-среде, содержащей ампициллин (50 мкг/мл). Все генетические операции осуществляли в соответствии с Sambrook и соавт. ДНК-последовательность клона рFP67xgptct подтверждали с использованием автоматического секвенатора (Pharmacia). Данный клон выделяли очисткой с использованием градиента CsCl и использовали для трансфекции CEF.
Пример 4. Создание рекомбинантных FPV
Родительский FPV, используемый для создания рекомбинантов, представлял собой аттенуированный штамм HP-438, который получали из патогенного штамма НР-1 с помощью шести последовательных пассажей в CEF, двух дополнительных пассажей в хориоаллонтойных мембран, и, в конце концов, 438 пассажей через CEF (Mayr A. and K. Malicki. 1966. Attenuierung von virulentem Huhnerpockenvirus in Zellkulturen und Eigenschaften des attenuierten Virus (Аттенуирование вирулентного куриного поксивируса в культуре и свойства аттенуированного вируса). Zentralbl. Veterinaermed, Reihe B 13:1-13). Дважды выделенный в виде бляшек изолят HP438 (FP9) пассировали затем шесть раз для создания штамма. FPV-штаммы выращивали в CEF на среде 199, содержащей 2% сыворотки новорожденного теленка (NBCS).
Рекомбинантный FPV получали путем гомологичной рекомбинации между FP9 и плазмидой pEFL29 или ее производными, как описано раньше. CEF, выращенные в 25 см2 матрасах, заражали FP9 с множественностью заражения (множественности заражения) 2 бляшкообразующие единицы (pfu)/клетку, затем через 2 часа эти клетки трансфицировали 10 мкг выделенной очисткой в градиенте CsCl плазмидной ДНК (pEFL-cr3 или pFP67xgptctl) с использованием 20 мкг липофектина (Gibco BRL, США). Приливали свежую среду (3 мл среды 199, содержащей 10% триптозофосфатной питательной среды и 2% NBCS) и клетки инкубировали при 37°С в СО2-инкубаторе. Через 24 ч вновь приливали свежую среду и клетки инкубировали в течение последующих 3-4-х дней. По истечении этого времени проинкубированные клетки замораживали-оттаивали три раза. Затем полученный клеточный лизат титровали в CEF для отбора рекомбинантных вирусов. После 2-х часов адсорбции удаляли вирусный инокулюм и добавляли слой агарозы, содержащей EMEN. Данный слой готовили путем смешивания одинаковых объемов 2% легкоплавкой агарозы и EMEN 2X (Gibco, Grand Island, NY) с 4% фетальной сыворотки теленка (Gibco, Grand Island, NY). На четвертый день были видны вирусные бляшки. CEF, трансфицированные pEFL-cr3, окрашивали путем добавления к данным культурам другого слоя агарозы с 0,33% Xgal (Melford Laboratories, Великобритания). Голубые бляшки отбирали и трижды выделяли очисткой до получения 100% вирусных бляшек, позитивных по экспрессии β-галактозидазы. Затем вирусные штаммы lacZ+ амплифицировали в CEF, выращенных в 25 см2 матрасах в среде 199, содержащей 10% триптозофосфатную питательную среду и 2% NBCS. Отобранный рекомбинантный FPV назвали FPCR3.
Селективная среда для трансфекции плазмидой pFP67xgptctl содержала микофеноловую кислоту (25 мкг/мл), ксантин (250 мкг/мл) и гипоксантин (1 мкг/мл). На четвертый день были видны вирусные бляшки. Так как гены gpt и cr3 фланкированы одними и теми же гомологичными областями, выделение вирусных бляшек в селективной среде свидетельствует о том, что произошла рекомбинация и оба гена встраивались в геном FPV. Бляшки выделяли очисткой трижды подряд в CEF. Отобранный рекомбинантный вирус назвали FPSCR3GPT.
Пример 5. ПЦР-анализ FPCR3.
Чтобы удостовериться, что рекомбинанты FPCR3 содержат cr3-ген, использовали ПЦР-анализ. Рекомбинантный FPV размножали в CEF в течение 6 дней, после чего клетки собирали и осаждали. Полученный осадок клеток суспендировали и инкубировали в течение 2 ч при 55°С в 200 мкл экстрагирующего буфера (10 мМ Трис-HCl, 100 мМ NaCl, 10 мМ ЭДТА, 0,5% SDS, 2% β-меркаптоэтанола), содержащего 1,25 мг/мл протеиназы К. Затем ДНК подвергали фенольно-хлороформному экстрагированию и осаждали этанолом. ДНК каждого вируса тестировали в ПЦР с помощью описанных ниже праймеров, комплементарных, соответственно, 5'- и 3'-концам cr3-гена. Условиями используемой ПЦР были 5 мин при 94°С, с последующими 25 циклами по 1 мин при 94°С, 1 мин 30 с при 45°С и 1 мин 30 с при 72°С и заключительным удлинением при 72°С в течение 10 мин.
Праймерные последовательности были следующими:
праймер 775, 5'-TATTAACATTGCCTAGTAG-3'
праймер 776, 5'-GAAGTAGAATCATAAAGAAC-3'.
После ПЦР-реакции для трех независимых CR3-рекомбинантных вирусов (FPCR3.1; FPCR3.2; FPCR3.3) обнаружили ожидаемую (1.3 т.п.н.)-полосу. Эта полоса отсутствовала у родительского вируса FPL29 (Фигура 3А).
Пример 6. ПЦР-анализ FPSCR3GPT.
Чтобы удостовериться, что FPSCR3GPT-рекомбинанты содержат cr3-ген, использовали ПЦР-анализ. Рекомбинантный FPV размножали в CEF в течение 6 дней, а затем полученные клетки собирали и осаждали. Полученный осадок клеток суспендировали и инкубировали в течение 2 ч при 55°С в 200 мкл экстрагирующего буфера (10 мМ Трис-HCl, 100 мМ NaCl, 10 мМ ЭДТА, 0,5% SDS, 2% β-меркаптоэтанола), содержащего 1.25 мг/мл протеиназы К. Затем ДНК подвергали фенольно-хлороформному экстрагированию и осаждали этанолом. ДНК каждого вируса тестировали в ПЦР с помощью праймеров, описанных ниже, комплементарных, соответственно, 5'- и 3'-концам cr3-гена. Условиями используемой ПЦР были 5 мин при 94°С, с последующими 25 циклами по 1 мин при 94°С, 1 мин 30 с при 45°С и 1 мин 30 с при 72°С, и заключительным удлинением при 72°С в течение 10 мин.
Праймерные последовательности были следующими:
праймер 2660, (257-279) 5'-GAAGATCTGTACAGAAATGGAAAAG-3'
праймер 2663, (1029-1059)
5'-CCCTGCATGTGGCTCAACTGGTACTAGCTTG-3'.
После ПЦР-реакции для трех независимых CR3-рекомбинантных вирусов (FPSCR3gpt.1; FPSCR3gpt.2; FPSCR3gpt.3) обнаружили ожидаемую (800 п.н.)-полосу. Эта полоса отсутствовала у родительского вируса FPL29 (Фигура 4А).
Пример 7. Оценка экспрессии CR3 в CEF, инфицированных FPCR3.
Экспрессию CR3 в FPCR3 подтверждали с помощью Вестерн-блоттинга. Конфлюирующие CEF, на 60 мм чашках Петри, инфицировали рекомбинантным FPV при множественности заражения 0,5 pfu/клетку. Через 24 часа эти клетки собирали, осаждали и суспендировали в 1Х буфере с SDS для внесения образцов (50 мМ Трис-HCl рН 6,8, 100 мМ DTT, 2% SDS, 0,1% бромфенолового синего, 10% глицерина). Белки фракционировали с помощью электрофореза в ДСН-ПААГ (в 15%-м геле). Затем электропереносом их помещали на нитроцеллюлозную мембрану (Hybond-C, Amersham, Великобритания) в соответствии со стандартным протоколом. После переноса эту мембрану блокировали в течение ночи в 5% обезжиренном сухом молоке в фосфатно-солевом буфере (PBS: 2,68 мМ KCl, 1,47 мМ KH2PO4, 0,137 M NaCl, 8,06 M Na2HPO4). Затем ее инкубировали в течение 2 ч при комнатной температуре с 10 мкг/мл моноклональных антител 6,2, разбавленных в PBS, содержащем 1% сухого молока. Эти моноклональные антитела получали у мышей, иммунизированных CR3 (Iglesias E., Ruiz M., Carrazana Y., Cruz L.J., Aguilar A., Jiménez V., Carpio E., Martinez M., Pérez M., Martinez C., Cruz O., Martin A., Duarte C. Chimeric proteins containing HIV-1 epitopes (Химерные белки, содержащие эпитопы ВИЧ-1). Journal Biochemistry, Molecular Biology and Biophysics, 2001, 5:109-20). Затем данную мембрану отмывали и инкубировали с овечьими антителами, конъюгированными с пероксидазой хрена, против мышиных антител (1:2000) (HRPO) (Amersham, Великобритания). После нескольких отмывок полученные иммуноблоты проявляли с использованием системы детекции ECL Вестерн-блот (Amersham, Великобритания) в соответствии с рекомендациями производителя. Для культур, инфицированных FPCR3, детектировали специфичную полосу с молекулярной массой между 50 и 60 кДа. В CEF, инфицированных родительским вирусом FP9, белок не детектировался (Фигура 3В).
Пример 8. Оценка экспрессии CR3 в CEF, инфицированных FPSCR3gpt.
Экспрессию CR3 в FPSCR3gpt подтверждали с помощью Вестерн-блоттинга по методу, аналогичному вышеописанному в предыдущем примере. Специфичная полоса с молекулярной массой между 50 и 64 кДа также детектировалась в культурах, инфицированных FPSCR3gpt, тогда как в CEF, инфицированных родительским FP9-вирусом, белок не детектировался (Фигура 4В).
Пример 9. Выделение очисткой FPCR3 и FPSCR3gpt и иммунизация мышей.
Большинство штаммов рекомбинантного FPV выращивали в CEF, полученных из флоккулированного осадка яйцеклеток, не содержащего специфичного патогена. FPV выделяли центрифугированием цитоплазматических экстрактов через 25% (w/v) сахарозную подушку на Beckman-центрифуге в роторе SW28 в течение 2-х часов при 29000 об/мин. Затем с помощью анализа бляшкообразования в монослое CEF определяли титр вируса. На Фигуре 5 показано, что после масштабирования данной культуры СR3-экспрессия не изменялась.
Молодых взрослых (в возрасте пяти-восьми недель) самок мышей Balb/c (полученных из SPF-колонии питомника в Institute for Animal Health, Compton, Великобритания или в Centro Nacional de Produccion de Animales de Laboratorio (CENPALAB), Куба) подвергали воздействию путем внутривенного (i.v.), внутрибрюшинного (i.p.) или подкожного (s.c.) введения 2.5-5х107 pfu FPCR3, FPSCR3gpt или в негативном вирусном контроле в 200 мкл стерильного PBS. Спустя две-четыре недели мышей тем же путем иммунизировали второй дозой 2,5-9х107 pfu тех же вирусов в 200 мкл стерильного PBS.
Пример 10. Детекция CTL-ответа у Balb/c-мышей на CR3.
Пятнышковый иммуноферментный анализ (ELISPOT) для выявления антигенспецифичных клеток, высвобождающих IFN-γ, осуществляли с использованием способа, базирующегося на раннее описанном (Tanguay S. And J.J. Killion. Direct comparison of ELISPOT and ELISA-based assays for detection of individual cytokine-secreting cells (Прямое сравнение методов ELISPOT и базирующегося на ELISA для выявления индивидуальных клеток, секретирующих цитокин). 1994. Lymphokine Cytokine Res., 13:259-263). Вкратце, 96-луночный планшет с мембраной из immobilon-P (Millipore, Molsheim, Франция) в течение ночи при 4°С покрывали 100 мкл/лунку мышиными 5 мкг/мл IFN-γ-специфичными моноклональными антителами R4 (Pharmingen, San Diego, California), трижды отмывали с помощью PBS и блокировали с использованием среды RPMI 1640 с добавлением 10% FBS при 37°С в течение 1 ч. Затем добавляли тестируемые клетки: которые представляли собой любую ex vivo суспензию спленоцитов (полученную как указано выше) мышей, подвергнутых первичному воздействию и повторно иммунизированных FPCR3 или FPSCR3gpt. Добавляли разные количества тестируемых клеток: 106, 2х105 и 4х104 в расчете на лунку. Клетки стимулировали добавлением клеток Р815, инкубированных с синтетическими пептидами при 1 мкМ или инфицированных рекомбинантным по CR3, Gag, или Nef VV при множественности заражения 5pfu/клетку. Для обнаружения фонового количества клеток, продуцирующих IFN-γ, включали клетки Р815 без пептида или клетки Р815, инфицированные контрольными вирусами коровьей оспы (vSC8 или штаммом коровьей оспы дикого типа WR). Каждая лунка имела конечный объем 200 мкл среды R10 с добавлением hIL-2. Все отличающиеся анализы тестировали в повторе. После инкубации в течение ночи (по меньшей мере 17 часов) данные планшеты отмывали 3Х с помощью PBS и 5Х с помощью PBS плюс 0,05% Твин 20, а затем добавляли конъюгированные с биотином вторичные антитела XMG1.2 (Pharmingen, San Diego, California) в количестве 0,5 мкг/мл и оставляли для взаимодействия при комнатной температуре на 2 ч. Далее лунки отмывали 5Х с помощью PBS плюс 0,05% Твин 20, добавляли стрептавидин, меченный щелочной фосфатазой (АР) (Vector Labs, CA, США), в разведении 1/1000 в PBS с добавлением 0,05% Твин 20 и оставляли при комнатной температуре на 1 ч. Далее лунки вновь отмывали 3Х с помощью PBS плюс 0.05% Твин 20 и 3Х с помощью PBS, а пятна проявляли с использованием набора для выявления АР-активности (Biorad, CA, США). Через 15 мин эти лунки отмывали водопроводной водой, сушили и пятна подсчитывали под стереоскопическим микроскопом (Leica Microscopy System, Heerbrugg, Швейцария). В качестве альтернативы, в некоторых анализах авторы настоящего изобретения использовали HRPO-меченный стрептавидин (Amersham, Великобритания), разбавленный 1/800; затем пятна проявляли с помощью 0,1% 3,3'-диаминобензидина (Sigma, Saint Louis, США) в 50 мМ Трис-HCl и 0,1% перекиси водорода. Полученные результаты выражали в виде числа клеток, образующих пятна (SFC) из расчета на 106 спленоцитов или фракционированных клеток. Значения, вдвое превышающие негативный контроль по каждой группе (Р815 без пептида или инфицированные контрольным VV), считали позитивными.
Результаты двух независимых ELISPOT-анализов представлены на Фигуре 6. Существенная часть спленоцитов Balb/c-мышей, иммунизированных FPCR3, но не негативным вирусом, оказывается позитивной в IFN-gamma-ELISPOT в противоположность Р815, инфицированных VVCR3 или VVgag и VVnef или подвергнутых воздействию V3 MN-пептидами (LKKKRIHIGPGRAFYERY).
В другом эксперименте Balb/c-мышей иммунизировали, как описано, FPCR3 или FPSCR3gpt, а индукцию CTL измеряли с использованием Р815, стабильно трансфицированных с помощью cr3 (P815cr3). Результаты этого эксперимента представлены на Фигуре 7. Оба рекомбинантных FPV индуцировали значительную часть специфичных для CR3 клеток, секретирующих гамма-IFB.
Пример 11. Процессинг и распознавание CR3-эпитопов лимфоцитами больных СПИДом.
Аутологичные В-клетки ВИЧ-инфицированных пациентов EBV-трансформировали и инфицировали рекомбинантным по CR3 VV (VVCR3). Эти клетки-мишени инкубировали с лимфоцитами периферической крови ВИЧ-пациентов, а количество спленоцитов, секретирующих IFNγ, подсчитывали с помощью ELISPOT. Данный эксперимент свидетельствует, что экспрессируемый поксвирусом cr3-ген способен эффективно представлять свои эпитопы CTL-лимфоцита ВИЧ-инфицированных пациентов.

Изобретение относится к области биотехнологии, иммунологии и вирусологии. Химерный ген CR3, имеющий последовательность SEQ ID NO:1, кодирует химерный полипротеин. Ген содержит фрагменты, кодирующие фрагменты, богатые эпитопами цитотоксических Т-клеток (CTL). Фрагменты выбраны из внутренних консервативных и регуляторных белков, эпитопов Т-хелперных клеток (Th) ВИЧ и В-клеточного эпитопа. Химерный полипротеин содержит фрагменты различных белков ВИЧ, индуцирующих иммунный ответ против ВИЧ-1. Изобретение может быть использовано в медицине для лечения и предупреждения ВИЧ. 4 н. и 3 з.п ф-лы, 8 ил., 2 табл.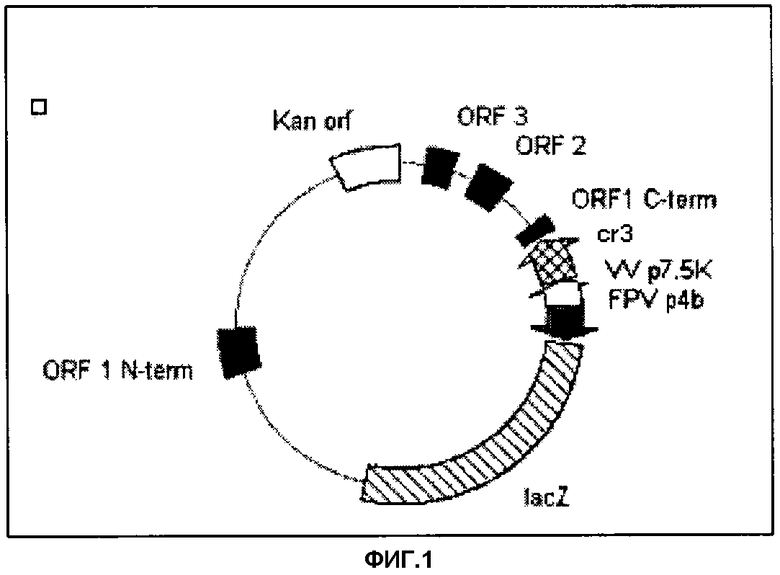
| WO 9821354, 22.05.1998 | |||
| EP 0412766, 13.02.1991. |

