Область техники, к которой относится изобретение
Настоящее изобретение относится к области иммунологии, в частности к вакцинной антигенной композиции, способной вызывать мощный и смешанный иммунный ответ против вируса гепатита С (HCV), и к комбинированным вакцинам против патогенных организмов, включающим эту вакцинную композицию.
Предпосылки к созданию изобретения
Созданию эффективной вакцины против HCV препятствовали различные проблемы, поскольку, являясь РНК-содержащим вирусом, этот вирус может быстро мутировать при адаптации к окружающей среде. Это вносит свой вклад в высокое разнообразие последовательностей множества вирусных изолятов, идентифицированных по всему миру. Значительная гетерогенность сосредоточена в гипервариабельном участке, заключенном в белке Е2 HCV, определенно включающем в себя нейтрализующий эпитоп (Bukh et al. US 6,110,465). Несмотря на наличие активной иммунной реакции, HCV вызывает персистентные инфекции у иммунокомпетентных индивидуумов. (Lechmann et al. (2000) Vaccine development for hepatitis C. Semin Liver Dis. 20:211-226). Не существует эффективной модели на животных или клеточной культуральной системы in vitro для поддержания репликации HCV и определения наличия нейтрализующих антител. Иммунологические модели, имеющие отношение к развитию заболевания или защитной реакции, не были определены полностью. Вероятно для избавления от HCV инфекции необходимы сильные, мультиспецифичные, долговременные как гуморальные, так и клеточные иммунные реакции (Lechmann et al. (2000) Vaccine development for hepatitis C. Semin Liver Dis. 20:211-226).
Для разработки вакцины против HCV было использовано несколько подходов, среди которых были всесторонне апробированы рекомбинантные белки, синтетические пептиды, вирусоподобные частицы, депротеинизированная ДНК и рекомбинантный вирус (Depla et al. US 6635257; Liang et al. US 6387662; Pachuk et al. US 6235888; Berzofsky et al. US 6685944).
Одной из первых стратегий, апробированных для HCV, была разработка вакцины на основе белковых субъединиц (Min et al. US 5985609), поскольку антитела к белкам оболочки могут обеспечивать защиту от некоторых флавивирусов. Некоторые стратегии, основанные на структурных антигенах HCV, вызывали ограниченную защиту против вируса в моделях на животных. Например, в случае с шимпанзе, иммунизированными олигомером E1 и E2, были вакцинированы семь шимпанзе, у 5 из 7 была защитная реакция, а 2 заболели, но выздоровление наступило до достижения хронической фазы (Choo et al. (1994) Vaccination of chimpanzees against infection by hepatitis C virus. PNAS USA, 91:1294-98). Такая защитная реакция коррелировала с наличием антител (Ат), способных ингибировать связывание E2 с клетками человека (Rosa et al. (1996) A quantitative test to estimate neutralizing antibodies to the hepatitis C virus: Cytofluorimetric assessment of envelope glycoprotein 2 binding to target cells. PNAS USA, 93:1759-63).
Рекомбинантный белок E1 из генотипа изолята 1b был очищен в виде гомодимеров, объединенных в частицы размером приблизительно 9 нм (Maertens et al. (2000) Improvement of chronic active hepatitis C in chronically infected chimpanzees after therapeutic vaccination with the HCV E1 protein. Acta Gastroenterol Belg. 63:203). Два шимпанзе, хронически инфицированных HCV, получали 9 доз по 50 мкг рекомбинантного белка E1. Вакцинация улучшала гистологическую картину печени, было установлено исчезновение вирусных антигенов в печени и снижение уровней аланинаминотрансферазы (ALT). Хотя уровни ARN в сыворотке не изменялись в течение лечения, после завершения лечения вновь появилось воспаление ткани печени и вирусные антигены. Наблюдалась корреляция между высоким уровнем антител к E1 и улучшением течения заболевания (Maertens et al. (2000) Improvement of chronic active hepatitis C in chronically infected chimpanzees after therapeutic vaccination with the HCV E1 protein. Acta Gastroenterol. Belg. 63:203).
Очень эффективным является получение вирусоподобных частиц из рекомбинантных белков и их использование в качестве вакцин, поскольку эти структуры зачастую имитируют свойства вируса (Liang et al. US 6,387,662). Частицы такой природы, полученные из клеток насекомых, инфицированных рекомбинантным бакуловирусом, несущим последовательность структурных антигенов HCV, обладали способностью вызывать гуморальный и клеточный иммунный ответ на такие антигены (Baumert et al. (1999) Hepatitis C virus-like particles synthesized in insect cells as a potential vaccine candidate. Gastroenterology 117:1397-407; Lechmann et al. (1999) Induction of humoral and cellular immune responses in mice by immunization with insect-cell derived HCV-like particles. Hepatology 30:423-29).
С другой стороны, для разработки рекомбинантной вакцины против HCV было апробировано несколько рекомбинантных вирусных векторов. В частности, интересными кандидатами являются рекомбинантные дефектные аденовирусы, благодаря их гепатотропизму, потенциальной возможности индуцировать гуморальный и клеточный иммунитет, а также возможности введения парентеральным и пероральным путем. Рекомбинантный аденовирус, содержащий гены, кодирующие структурные белки HCV, индуцирует антительный ответ на эти белки (Makimura et al. (1996). Induction of antibodies against structural proteins of hepatitis C virus in mice using recombinant adenovirus. Vaccine 14:28-36). Кроме того, после иммунизации мышей рекомбинантным аденовирусом, содержащим ядерный и E1 антиген, на эти антигены определялся T-цитотоксический специфический ответ (Bruna-Romero et al. (1997) Induction of cytotoxic T-cell response against hepatitis C virus structural antigens using a defective recombinant adenovirus. Hepatology 25:470-77). Хотя эти результаты были обнадеживающими, недавние проблемы, касающиеся применения рекомбинантных аденовирусов в генной терапии, подняли несколько вопросов в отношении их применения у людей. При использовании других рекомбинантных вирусов, подобных вирусу осповакцины, несущих различные гены HCV, у мышей индуцировался сильный Т-цитотоксический и хелперный ответ (Shirai et al. (1994) An epitope in hepatitis C virus core region recognized by cytotoxic T cells in mice and humans. J. Virol. 68:3334-42; Large et al. (1999) Suppression of host immune response by the core protein of hepatitis C virus: possible implications for hepatitis C virus persistence. J. Immunol. 162:931-38). Тем не менее, на применение этих рекомбинантных вирусов, а также других вариантов альфа вирусов, подобных вирусу леса Семлики, влияют нормативно-правовые вопросы и вопросы безопасности, связанные с их применением (Vidalin et al. (2000) Use of conventional or replicating nucleic acid-based vaccines and recombinant Semliki forest virus-derived particles for the induction of immune responses against hepatitis C virus core and E2 antigens. Vaccine 276:259-270).
Идентификация некоторых эпитопов для T CD4+ и CD8+ в вирусном полипротеине HCV, которые могут быть важны для распознавания вируса, поддерживает стратегию использования синтетических пептидов в качестве вакцин-кандидатов против этого патогена (Berzofsky et al. US 5980899). Различные пептиды в чистом виде или липидированные, содержащие эпитопы ядерных, NS4 и NS5 белков HCV, индуцировали мощный Т цитотоксический ответ у мышей (Shirai et al. (1996) Use of intrinsic and extrinsic helper epitopes for in vivo induction of anti-hepatitis C virus cytotoxic T lymphocytes (CTL) with CTL epitope peptide vaccines. J. Infect. Dis. 173:24-31; Hiranuma et al. (1999) Helper T cell determinant peptide contributes to induction of cellular immune responses by peptide vaccines against hepatitis C virus. J. Gen. Virol. 80:187-193; Oseroff et al. (1998) Pools of lipidated HTL-CTL constructs prime for multiple HBV and HCV CTL epitope responses. Vaccine, 16:823-833).
Другая стратегия, используемая для разработки вакцины против HCV, основана на возможности образования антител к линейным эпитопам. Эта альтернатива была опробована главным образом в отношении образования антител к гипервариабельному участку 1 HCV (HVR-1) у кроликов и шимпанзе с некоторыми обнадеживающими результатами (Esumi et al. (1999) Experimental vaccine activities of recombinant E1 and E2 glycoproteins and hypervariable region 1 peptides of hepatitis C virus in chimpanzees. Arch Virol. 144:973-980; Shang et al. (1999) Broadly cross-reactive, high-affinity antibody to hypervariable region 1 of the hepatitis C virus in rabbits. Virology 258:396-405). Основной проблемой выбора HVR в качестве мишени для HCV вакцины является существование псевдовидов в этом участке генома.
Основным препятствием для концепции пептидных вакцин является то, что эти пептиды без хелперной функции могут быть низкоиммуногенными, и очень часто эффективность этой вакцины основана на индукции мультивалентного ответа с очень широким спектром в отношении различных антигенов. Эти ограничения являются недостатками этой стратегии.
Для разработки вакцины против HCV широко изучалась ДНК-иммунизация (Donnelly et al. US 6653125). Белок ядра включали в несколько векторов экспрессии, используя полную длину или усеченные варианты этого белка (Lagging et al. (1995) Immune responses to plasmid DNA encoding the hepatitis C virus core protein. J. Virol. 69:5859-5863; Chen et al. (1995) Genetic immunization of mice with plasmid containing hepatitis C virus core protein-encoding DNA. Vaccine Res. 4:135-144).
Другие генетические конструкции включали также 5ґ нетранслируемую область (Tokushige et al. (1996) Expression and immune response to hepatitis C virus core DNA-based vaccine constructs. Hepatology 24:14-20). Также были изучены слитые варианты в отношении поверхностного антигена HB и других вирусов (Major et al. (1995) DNA-based immunization with chimeric vectors for the induction of the immune responses against the hepatitis C virus nucleocapsid. J. Virol. 69:5798-805). Иммунизация этими векторами главным образом индуцировала заметные лимфопролиферативные и Т-клеточные цитотоксические реакции.
Для этого типа технологии белки оболочки HCV также представляли интересный объект исследования. В случае E2, гуморальный иммунный ответ, по-видимому, направлен на HVR-1 (Lee et al. (1998) Hepatitis C virus envelope DNA-based immunization elicits humoral and cellular immune responses. Mol. Cells 8:444-451). При проведении иммунизации векторами для секретируемых вариантов белков E1 и E2 никаких отличий в иммунном ответе по сравнению с несекретируемыми вариантами обнаружено не было (Lee et al. (1998) Optimal induction of hepatitis C virus envelope-specific immunity by bicictronic plasmid DNA inoculation with the granulocyte-macrophage colony-stimulating factor gene. J. Virol. 72:8430-8436). Заражение бицистронными плазмидами, экспрессирующими независимо гены, кодирующие GM-CSF, белки E1 и E2, вызывает и наращивает гуморальный и клеточный иммунный ответ. В последнее время иммуногенное действие этих белков было изучено при включении обоих в бицистронные векторы, где возможность образования гетеродимера in vivo была активизирована или исключена. Утверждалось, что при образовании таких комплексов антительный ответ не наблюдался. В полной противоположности с этим высокие титры антител к конформационным и линейным детерминантам вырабатывались при иммунизации животных плазмидами, экспрессирующими усеченные формы этих двух структурных белков. Следовательно, по-видимому, для получения хорошего антительного ответа при проведении иммунизации и тем, и другим антигенами необходимо избегать образования гетеродимеров (Fournillier et al. (1999) Expression of noncovalent hepatitis C virus envelope E1-E2 complexes is not required for the induction of antibodies with neutralizing properties following DNA immunization. J. Virol. 73:497-504).
По этой методике также были апробированы неструктурные белки. Кодирующую область C-концевого домена белка NS3 включали в вектор, обеспечивающий либо одновременную, либо независимую экспрессию этого белка и IL-2, с получением хороших результатов (Papa et al. (1998) Development of multigenetic plasmid vector for HCV DNA immunization. Res. Virol. 149:315-319). Белки NS4 и NS5 вызывали Т-цитотоксический и антительный ответ аналогичным путем (Encke et al. (1998) Genetic immunization generates cellular and humoral immune responses against the nonstructural proteins of the hepatitis C virus in murine model. J. Immunol. 161:4917-4923).
В последнее время подтвердилось, что использование генной конструкции, кодирующей вирусные неструктурные белки (NS3, NS4 и NS5, включая ген, кодирующий GM-CSF) вызывало мощный антительный ответ, а также усиление Т-клеточного пролифиративного ответа в отношении каждого из неструктурных белков (Cho et al. (1999) Enhanced cellular immunity to hepatitis C virus nonstructural proteins by codelivery of granulocyte macrophage-colony stimulating factor gene in intramuscular DNA immunization. Vaccine 17:1136-1144).
В целом сообщалось об эффективной экспрессии, а также выработке антител к различным антигенам HCV после ДНК иммунизации. Уровни располагались в интервале от 1:100 до 1:100000 в соответствии с исследуемой комбинацией (Inchauspe et al. (1997) DNA vaccination for the induction of immune responses against hepatitis C virus proteins. Vaccine 15:853-856). Также подтвердилось усиление специфической цитотоксичности и лимфопролиферации (Inchauspe et al. (1997) Plasmid DNA expressing a secreted or a nonsecreted form of hepatitis comparative studies of antibody and T-helper responses following genetic immunization. DNA and Cell Biology 16:185-195). Однако необходимо усовершенствование этой методики для достижения достаточно мощного гуморального и клеточного иммунного ответа на различные белки HCV. В этом смысле были опробованы некоторые варианты для улучшения индуцированного иммунного ответа после ДНК вакцинации, поскольку в них используют липосомы в качестве адъювантов (Gramzinski et al. (1998) Immune response to a hepatitis B DNA vaccine in Aotus Monkey: A comparison of vaccine formulation, route and method of administration. Mol. Medicine 4:109-118), монофосфорил липид А и сапонин QS-21 (Sasaki et al. (1998) Induction of systemic and mucosal immune responses to human immunodeficieny virus type 1 by a DNA vaccine formulated with QS-21 saponin adjuvant via intramuscular and intranasal routes. J. Virol. 72:4931-4939). С другой стороны, в качестве биологического адъюванта при ДНК иммунизации были изучены дендритные клетки. Были использованы вакцины, которые составляют дендритные клетки, полученные из костного мозга мышей, генетически модифицированные ex vivo для экспрессии опухолевых антигенов с использованием вирусных векторов, и была доказана их способность стимулировать специфический Т-клеточный ответ и опосредованный клетками профилактический иммунитет против опухолей у мышей (Specht et al. (1997) Dendritic cells retrovirally transduced with a model antigen gene are therapeutically effective against established pulmonary metastases. J. Exp. Med. 186:1213-1221; Brossart et al. (1997) Virus-mediated delivery of antigenic epitopes into dendritic cells as a means to induce CTL. J. Immunol. 158:3270-3276; Song et al. (1997) Dendritic cells genetically modified with an adenovirus vector encoding the cDNA for a model antigen induce protective and therapeutic antitumor immunity. J. Exp. Med. 186:1247-1256), o ARN (Boczkowski et al. (1996) Dendritic cells pulsed with RNA are potent antigen-presenting cells in vitro and in vivo. J. Exp. Med. 184:465-472).
В настоящее время усовершенствование векторов, в том числе вставка CpG мотивов для усиления иммунного ответа в отношении экспрессируемых антигенов и систем для высвобождения из плазмиды являются ключевыми целями изучения путей преодоления ограничений этой технологии (Hasan et al. (1999) Nucleic acid inmunization: concepts and techniques associated with third generation vaccines. J. Immunol. Meth. 229:1-22).
В результате трудностей, создаваемых HCV, и отсутствия четких определений иммунологических параметров, которые коррелируют с защитным действием против этого патогена, возможно, что эффективная вакцина против HCV требовала многоаспектного подхода, который стимулирует различные проявления иммунной реакции. Возможно решение этой проблемы лежит в объединении нескольких подходов к созданию вакцин, изученных к настоящему времени. В этом смысле были опробованы схемы иммунизации, которые объединяют начальные дозы ДНК вакцины и ревакцинирующие дозы либо с белками, либо с рекомбинантными вирусными векторами, даже при положительных результатах которых требуются дополнительные исследования для демонстрации того, что объединение этих подходов могло вызывать защитный иммунитет против HCV (Hu et al. (1999) Characterization of the humoral and cellular immune responses against hepatitis C virus core induced by DNA-based immunization. Vaccine 17:3160-3170; Pancholi et al. (2000) DNA prime-canarypox boost with polycistronic hepatitis C virus (HCV) genes generates potent immune responses to HCV structural and nonstructural proteins. J. Infect. Dis. 182:18-27).
Кроме того, для модели гепатита В был опробован вакцинный препарат, который составлен комплексом поверхностного антигена вируса гепатита В с моноклональным антителом против HBsAg и плазмиды, кодирующей этот антиген (Wen et al. US 6,221,664). Эта композиция позволяла осуществлять презентацию антигена различными путями и быструю индукцию иммунного ответа, более высокую, чем при использовании отдельных активных компонентов. Таким образом, разработка эффективных вакцин является крайне необходимой и помимо заметного успеха все еще остается текущей проблемой.
В настоящем изобретении описан вакцинный препарат, который составляют структурные антигены вируса гепатита С. В отличие от предыдущих вакцинных препаратов на основе белка, в которых использовались только белки Е1 и Е2 (отдельно или в комбинации), в вакцинный препарат по данному изобретению также включен ядерный антиген HCV. Универсальность этой вакцинной композиции лежит в интервалах количества антигена, используемого в этом вакцинном препарате, что дополнительно позволяет вызывать мощные иммунные реакции (гуморальные и клеточные) на различные антигены одновременно, а также защитную реакцию на заражение рекомбинантным вирусом осповакцины в моделе на мышах.
Подробное описание изобретения
По литературным данным хорошо известно, что существует крайняя необходимость в разработке надежной и эффективной терапевтической вакцины против HCV. Таким образом, настоящее изобретение направлено на получение вакцинного препарата, составленного из смеси структурных антигенов вируса гепатита С. В этом случае ядерный белок, структурный белок E1 и структурный белок E2 обладают способностью индуцировать гуморальный и клеточный специфический иммунный ответ против HCV, в частности, (но не только) у пациентов, с хронической формой HCV.
Новизна изобретения заключается в получении интервала антигенных соотношений и защитного эффекта, достигаемого после иммунизации смесями по изобретению. Изучаемые антигены являются мишенями, представляющими особый интерес, с помощью которых можно вызвать иммунный ответ на HCV.
В конкретном варианте осуществления, в вакцинной композиции используют интервал соотношений для антигенов, присутствующих в смеси, и составленный из числа (1,5-2,5):(1-2,5):(1-2) для белка ядра, Е1 и Е2 соответственно для формирования оптимального гуморального иммунного ответа. В другом варианте осуществления антигены используют в соотношениях (1,5:1,1:1) для белка ядра Е1 и Е2 соответственно.
В одном варианте осуществления изобретения в вакцинной композиции используется интервал соотношений для антигенов, присутствующих в смеси, составляющий (1-50):(120-180):(120-180) для белка ядра, Е1 и Е2 соответственно для формирования оптимального клеточного иммунного ответа. В другом варианте осуществления белковые антигены используют в соотношении (1:160:160) для белка ядра, Е1 и Е2 соответственно.
Вакцинные препараты по настоящему изобретению можно вводить в виде твердого продукта, жидкого продукта или аэрозоля любым традиционным способом введения вакцин, в том числе перорально и путем парентеральной инъекции (т.е. внутривенно, подкожно или внутримышечно), кроме того, эта композиция может быть получена либо путем смешивания белковых антигенов до их усиления или усилением антигенов отдельно и последующего их смешивания.
Антигены по изобретению могут быть получены из природного источника или технологией рекомбинантных ДНК в любой экспрессионной системе, подобной описанной ниже.
В одном варианте осуществления настоящего изобретения вакцинные препараты, включенные в это изобретение, можно вводить по различным схемам иммунизации.
Аналогичным образом иммунизация этими антигенными комбинациями, смешанными с другими полисахаридными антигенами (конъюгированными или нет) и/или вирусными антигенами и/или бактериальными антигенами, формирует суммарный иммунный ответ, следовательно, в отношении широкого спектра инфекционных организмов.
В другом варианте осуществления настоящего изобретения вакцинная композиция включает комбинации структурных антигенов HCV с плазмидами, которые кодируют антигены инфекционных организмов, против вирусных и/или бактериальных заболеваний.
Предпочтительные вакцинные композиции по изобретению представляют собой препараты, в которых другие вирусные антигены, включенные в них, представляют собой ядерные и/или поверхностные антигены вируса гепатита В, протеолипосому из N. meningitidis или очищенные белки наружной мембраны N. meningitidis в качестве бактериальных антигенов, и аналогичные полисахаридные антигены, конъюгированные или неконъюгированные с белками-носителями. Они могут быть использованы в качестве активных компонентов в композициях против вирусных и бактериальных организмов.
Описанные здесь вакцинные композиции можно вводить по схемам, объединенным с другими кандидатными вакцинами на основе ДНК вакцины; живыми рекомбинантными векторами, пептидами и белками. Их также можно использовать для индукции иммунитета против HCV у больных хроническим гепатитом С с циррозом и раком печени.
Краткое описание чертежей
Фиг.1: гуморальный иммунный ответ на структурные белки HCV по данным ELISA; (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2.
Фиг.2: лимфопролиферативный иммунный ответ на структурные белки HCV в клетках селезенки мышей, иммунизированных исследуемыми комбинациями. HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.3: Функциональный ответ изучаемых препаратов, оцениваемый после заражения рекомбинантным вирусом осповакцины.
Фиг.4: Анализ влияния компонентов, присутствующих в вакцинных препаратах против HCV, на (A) гуморальный иммунный ответ, (B) лимфопролиферативный иммунный ответ, (C) заражение рекомбинантным вирусом осповакцины.
Фиг.5: влияние на гуморальный иммунный ответ интервалов соотношений антигенов, используемых в вакцинных препаратах против HCV: (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2.
Фиг.6: влияние интервала соотношений антигенов, используемых в вакцинных препаратах против HCV на лифопролиферативный иммунный ответ на структурные белки HCV в клетках селезенки иммунизированных мышей. HCcAg ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.7: влияние на функциональный ответ, определяемый у мышей на заражение рекомбинантным вирусом осповакцины, интервала соотношений антигенов, используемых в вакцинных препаратах против HCV.
Фиг.8: гуморальный иммунный ответ на структурные белки HCV по данным ELISA после иммунизации в различные промежутки времени (различные схемы иммунизации), (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2.
Фиг.9: лимфопролиферативный иммунный ответ на структурные белки HCV в клетках селезенки мышей, иммунизированных в различные промежутки времени (различные схемы иммунизации), HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.10: функциональный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных в различные промежутки времени.
Фиг.11: гуморальный иммунный ответ на структурные белки HCV по данным ELISA, у мышей, иммунизированных вакцинными препаратами против HCV, введенными различными путями. (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2.
Фиг.12: лимфопролиферативный иммунный ответ на структурные белки HCV в клетках селезенки мышей, иммунизированных вакцинными препаратами против HCV, введенными различными путями. HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.13: функциональный иммунный ответ, определяемый на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных вакцинными препаратами против HCV, введенными различными путями.
Фиг.14: гуморальный иммунный ответ на структурные белки HCV по данным ELISA, у мышей, иммунизированных вакцинными препаратами против HCV с использованием различных адъювантов. (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2.
Фиг.15: влияние различных адъювантов на лимфопролиферативный иммунный ответ против структурных белков HCV в клетках селезенки иммунизированных мышей. HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.16: влияние различных адъювантов на функциональный иммунный ответ на заражение рекомбинантным вирусом осповакцины иммунизированных мышей.
Фиг.17: гуморальный иммунный ответ по данным ELISA после иммунизации мышей смесью вакцинного препарата против HCV и вирусных антигенов, (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2, (D) против HBsAg и (E) против HBcAg.
Фиг.18: лимфопролиферативный иммунный ответ на структурные белки HCV (ядерный, E1, E2), HBsAg и HBcAg в клетках селезенках мышей, иммунизированных комбинациями вакцинного препарата против HCV и вирусных антигенов. HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.19: функциональный иммунный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных комбинациями вакцинного препарата против HCV и вирусных антигенов.
Фиг.20: гуморальный иммунный ответ по данным ELISA после иммунизации смесью вакцинного препарата против HCV и бактериальных антигенов (белок наружной мембраны N. meningitidis), (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2 и (D) против белков наружной мембраны N. meningitidis.
Фиг.21: лимфопролиферативный иммунный ответ на структурные белки HCV (ядерный, E1, E2) и белки наружной мембраны N. meningitidis в клетках селезенки мышей, иммунизированных комбинациями вакцинного препарата против HCV и бактериальных антигенов (OMP, белки наружной мембраны N. meningitidis). HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.22: функциональный иммунный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных комбинациями вакцинного препарата против HCV и бактериальных антигенов (белки наружной мембраны N. meningitidis).
Фиг.23: гуморальный иммунный ответ по данным ELISA у мышей, иммунизированных смесью вакцинного препарата против HCV и конъюгированного полисахарида капсулы, (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2 и (D) против полисахарида капсулы N. meningitidis серогруппы С.
Фиг.24: лимфопролиферативный иммунный ответ на структурные белки HCV (ядерный, E1, E2) в клетках селезенки мышей, иммунизированных смесью вакцинного препарата против HCV и конъюгированного полисахарида капсулы. HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.25: функциональный иммунный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных смесью вакцинного препарата против HCV и конъюгированного полисахарида капсулы.
Фиг.26: гуморальный иммунный ответ по данным ELISA после иммунизации смесью вакцинного препарата против HCV и плазмид, кодирующих HBsAg и HBcAg, (A) против ядерного антигена HCV, (B) против белка E1, (C) против белки E2, (D) против HBsAg, и (E) против HBcAg.
Фиг.27: лимфопролиферативный иммунный ответ на структурные белки HCV (ядерный, E1, E2), HBsAg и HBcAg, в клетках селезенки мышей, иммунизированных комбинациями вакцинного препарата HCV и плазмид, кодирующих HBsAg и HBcAg. HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.28: функциональный иммунный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных комбинациями вакцинного препарата против HCV и плазмид, кодирующих антигены HBsAg и HBcAg.
Фиг.29: гуморальный иммунный ответ по данным ELISA у мышей, иммунизированных комбинациями ДНК и ревакцинирующей дозы с вакцинным препаратом против HCV, (A) против ядерного антигена HCV, (B) против белка E1, (C) против белка E2.
Фиг.30: лимфопролиферативный иммунный ответ на структурные белки HCV (ядерный, E1, E2), в клетках селезенки мышей, иммунизированных комбинациями ДНК и ревакцинирующей дозы с вакцинным препаратом против HCV. HCcAg, белок ядра HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.31: функциональный иммунный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных комбинациями ДНК и ревакцинирующей дозы с вакцинным препаратом против HCV.
Фиг.32: гуморальный иммунный ответ на структурные белки HCV (ядерный, E1, E2), у мышей, иммунизированных вакцинными препаратами против HCV, полученными разными способами.
Фиг.33: лимфопролиферативный иммунный ответ на структурные белки HCV (ядерный, E1, E2) в клетках селезенки мышей, иммунизированных вакцинным препаратом против HCV, полученным различными способами. HCcAg, ядерный антиген HCV, E2-coli, E2, полученный и очищенный из Escherichia coli, E2-lev, E2, полученный и очищенный из дрожжей Pichia pastoris, E1-coli, E1, полученный и очищенный из Escherichia coli.
Фиг.34: функциональный иммунный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных вакцинным препаратом против HCV, полученным различными способами.
Подробное описание способов осуществления/Примеры
Пример 1: гуморальный, лимфопролиферативный и функциональный ответ на заражение рекомбинантным вирусом осповакцины у мышей, иммунизированных белковым вакцинным препаратом против HCV
Для демонстрации формирования специфического и функционального иммунного ответа на HCV после введения вакцинного препарата самкам мышей BALB/c (группы из 10 животных) интраперитонеально вводили ядерный антиген HCV, E1 и E2 антигены после перемешивания и усиления гидроксидом алюминия. Схема иммунизации включала в себя 3 инокуляции в 0, 7, 14 дни. Гуморальный и клеточный (лимфопролиферативный ответ) иммунные ответы, а также защитную реакцию на заражение рекомбинантным вирусом осповакцины vvRE (который включал в себя структурные белки HCV) изучали через 15 дней после последней инокуляции. Группы этого исследования приведены в Таблице 1.

Гуморальный иммунный ответ на структурные белки HCV определяли с помощью иммуноферментного анализа (ELISA). Для статистического анализа результатов использовали однофакторный дисперсионный анализ Крускал-Валлиса и метод Данна в качестве повторной проверки. Результаты с p<0,05 считались статистически значимыми. На Фиг.1 показано, что возможно получить специфический гуморальный иммунный ответ на три структурных антигена HCV при их введении в определенных соотношениях. Некоторые комбинации давали высокие титры антител против белков HCV (ядерного, E1 и E2), и они были статистически выше, чем титры, полученные в контрольных группах, в которых мышей иммунизировали отдельными белками или одним адъювантом.
Было показано характерное изменение лимфопролиферативного ответа (Фиг.2) на структурные антигены HCV (ядерный, E1 и E2) для каждой комбинации. Эти данные впоследствии использовали для расчета оптимального варианта (соотношения антигенов) для формирования наилучшего иммунного ответа. Результаты представлены в виде индекса стимуляции клеток селезенки иммунизированных животных (рассчитан путем включения 3H-тимидина). Экспериментальные группы, представленные на Фиг.2, соответствуют группам, представленным в Таблице 1, также имеется отрицательный контроль (группа 17), который представляют мыши, иммунизированные только гидроксидом алюминия.
Функциональный иммунный ответ, вызываемый исследуемыми препаратами, оценивали после заражения vvRE. Животных заражали интраперитонеальным введением 106 бляшкообразующих единиц (pfu) vvRE через 15 дней после окончания проведения иммунизации, а присутствие вируса в яичниках мышей оценивали через 5 дней после инокуляции вируса. Полученные результаты представлены на Фиг.3, на которой в группе 13 показано 2-Log снижение титра вируса по сравнению с группой отрицательного контроля. Экспериментальные группы, представленные на Фиг.3, соответствовали группам, указанным в Таблице 1.
Проводя анализ поверхности отклика, полученного согласно функциональным свойствам используемых параметров (антительный ответ, лимфопролиферативный ответ и защитная реакция на заражение вирусом осповакцины (Фиг.4)), были рассчитаны наилучшие соотношения антигенов для получения оптимального ответа для каждого параметра. Для антительного ответа оптимальное используемое соотношение антигенов соответствует (1,5:1,1:1):(ядерный:E1:E2), тогда как для получения оптимального клеточного ответа наилучшее соотношение антигенов, используемое для иммунизации, составило (1:160:160):(ядерный:E1:E2).
Пример 2: Оценка иммунного ответа у мышей BALB/c после иммунизации препаратами белков HCV с различными интервалами соотношений антигенов
Для оценки интервала антигенов, используемых в вакцинных препаратах для получения хорошего иммунного ответа, мышей BALB/c иммунизировали внутрибрюшинно по той же схеме иммунизации, которая описана в примере 1. Соотношения антигенов HCV (ядерного, E1 и E2) приведены в Таблице 2. Количества антигена, используемого в этом исследовании, представляли собой результаты анализа поверхности отклика из примера 1. Для гуморального иммунного ответа исследуемые интервалы соотношений составили: (1,5-2.5):(1-2,5):(1-2):(ядерный:E1:E2), тогда как для клеточного иммунного ответа исследуемые интервалы соотношений составили: (1-50):(120-180):(120-180): (ядерный:E1:Е2).
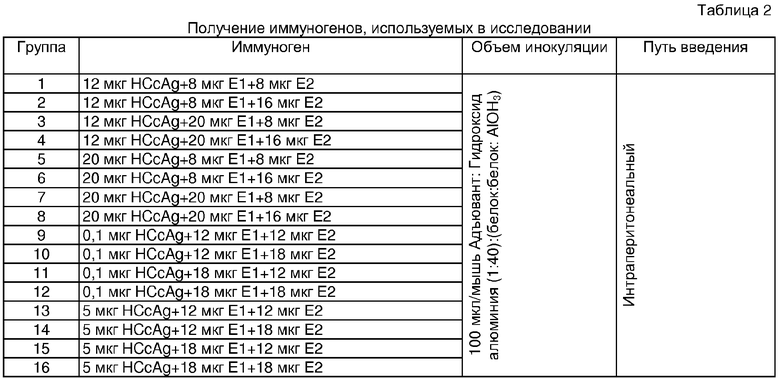
Гуморальный иммунный ответ на структурные антигены HCV представлен на Фиг.5. Изучаемые соотношения вызывали специфический антительный ответ на антигены HCV (ядерный, Е1 и Е2). Не было обнаружено статистически значимых различий в гуморальном ответе, вызываемом различными вариантами (оптимальным вариантом для клеточного ответа и оптимальным вариантом для гуморального ответа) несмотря на изменения, наблюдаемые для каждого конкретного антигена. Экспериментальные группы, отмеченные на Фиг.5, соответствовали группам, указанным в Таблице 2.
Лимфопролиферативный ответ оценивали путем включения 3H-тимидина в клетки селезенки иммунизированных мышей после их стимуляции. У иммунизированных мышей формировался специфический лимфопролиферативный ответ на изучаемые антигены (Фиг.6). Группы, отмеченные на Фиг.6, соответствовали группам, указанным в Таблице 2, плюс другой отрицательный контроль (группа 17), который представляют неиммунизированные мыши. Между индексами стимуляции не наблюдалось статистически достоверных различий для каждого из изученных вариантов (оптимального варианта для клеточного ответа и оптимального варианта для гуморального ответа соответственно).
Иммунизированных мышей заражали внутрибрюшинным введением 106 pfu vvRE. Титр вируса оценивали в яичниках иммунизированных мышей через 5 дней после инокуляции вируса. На Фиг.7 показаны полученные защитные уровни. Снижение вирусной нагрузки было статистически значимым у мышей, иммунизированных смесями, по сравнению с отрицательным контролем, главным образом в тех группах, в которых животных иммунизировали оптимальным вариантом для формирования клеточного иммунного ответа (приблизительно 2,5-Log снижение титра вируса). Исследуемые группы, представленные на Фиг.7, соответствовали группам, указанным в Таблице 2, плюс дополнительный отрицательный контроль (группа 17), который представлен неиммунизированными мышами.
Пример 3: Оценка иммунного ответа, вызываемого у мышей BALB/c иммунизацией композициями белков HCV после использования различных схем иммунизации
Для анализа влияния времени между иммунизациями на формирование иммунного ответа самок мышей BALB/c иммунизировали внутрибрюшинно в различные интервалы времени вариантом вакцины для формирования оптимального клеточного ответа, как указано в примере 1. Группу 1 иммунизировали на 0, 1 и 2 неделях. Группу 2 иммунизировали на 0, 2 и 4 неделях. Группу 3 иммунизировали на 0, 3 и 6 неделях, тогда как группу 4 иммунизировали на 0, 4 и 8 неделях. Титры антител против ядерного белка HCV, белков E1 и E2 оценивали с помощью ELISA. Результаты представлены на Фиг.8. При оценке через 15 дней после последней иммунизации статистически достоверных различий в титрах антител против E1 и E2 не было. Статистически достоверные различия наблюдались в титрах антител против ядерного антигена между группой 1 (схема иммунизации на 0, 1, 2 неделе) и остальными группами при их сравнении.
Лимфопролиферативный ответ иммунизированных мышей оценивали через 15 дней после последней иммунизации. Аналогичным образом различия в индексах стимуляции между каждым из исследуемых вариантов не были статистически значимыми. Полученные результаты показаны на Фиг.9, где также была включена дополнительная группа отрицательного контроля (Группа 5), которую представляли неиммунизированные мыши.
Защитную реакцию иммунизированных мышей против вирусного заражения оценивали как описано ранее в других примерах. Животных заражали через 15 дней после окончания проведения иммунизации внутрибрюшинным введением 106 pfu vvRE, и присутствие вируса в яичниках мышей оценивали через 5 дней после инокуляции вируса. Полученные результаты показаны на Фиг.10, где наблюдаемые различия титров вирусов между группами, иммунизированными по различным схемами, не были статистически значимыми. На этой фигуре также был включен другой отрицательный контроль (Группа 5), который представляли неиммунизированные мыши.
Пример 4: Оценка иммунного ответа у мышей BALB/c после иммунизации препаратами белков HCV, введенными различными путями
Также одним из изучаемых аспектов было применение вакцинного препарата посредством различных путей иммунизации. Самок мышей BALB/c иммунизировали препаратами HCV для формирования оптимального клеточного и гуморального иммунного ответа различными путями: Группа 1: оптимальный состав для гуморального ответа, введенный подкожно (п.к.), Группа 2: оптимальный состав для гуморального ответа, введенный внутрибрюшинно (и.п.), Группа 3: оптимальный состав для формирования гуморального ответа, введенный внутримышечно (в.м.), Группа 4: оптимальный состав для гуморального ответа, введенный интраназально (и.н.), Группа 5: оптимальный состав для клеточного ответа, введенный подкожно, Группа 6: оптимальный состав для клеточного ответа, введенный внутрибрюшинно, Группа 7: оптимальный состав для клеточного ответа, введенный внутримышечно, Группа 8: оптимальный состав для клеточного ответа, введенный интраназально.
Гуморальный иммунный ответ на структурные антигены HCV (ядерный, E1 и E2) изучали через 15 дней после завершения проведения иммунизации (0, 1 и 2 недели). Полученные результаты показывают, что, несмотря на статистически значимые различия между уровнями антител, на используемые антигены формировался специфический иммунный ответ. Эти различия обусловлены различными схемами иммунизации, где формирование антительного ответа является предпочтительным для некоторых вариантов вследствие кинетики иммунного ответа. Не наблюдалось статистически значимых различий при подкожном, внутрибрюшинном и внутримышечном путях введения. Однако результаты были выше при использовании внутрибрюшинного пути введения и ниже при использовании интраназального пути введения в этом случае со статистически достоверными отличиями. Результаты показаны на Фиг.11.
Через 15 дней после последней иммунизации оценивали лимфопролиферативный иммунный ответ мышей на вирусные антигены. Подобно гуморальному иммунному ответу различия индексов стимуляции между исследуемыми группами находятся в соответствии с количеством антигенов, используемых при интраназальном введении, и в целом (для остальных путей иммунизации) исходя из кинетики иммунологически сформированного иммунного ответа. Специфический лимфопролиферативный ответ формировался на каждый антиген HCV. Тем не менее, индексы стимуляции, полученные при интраназальном введении, были статистически достоверно ниже по сравнению с другими путями иммунизации. Полученные результаты показаны на Фиг.12. В это исследование также была включена другая группа отрицательного контроля (Группа 9), которую представляли неиммунизированные мыши.
Защитную реакцию иммунизированных мышей на вирусное заражение оценивали через 15 дней после окончания проведения иммунизации путем инокуляции 106 pfu vvRE. Через пять дней после заражения определяли количество вируса в яичниках иммунизированных и зараженных мышей. Полученные результаты представлены на Фиг.13, где не наблюдалось различий между различными схемами иммунизации, хотя при интраназальном введении показано 1-Log снижение титра вируса. В этот эксперимент была включена другая группа отрицательного контроля (Группа 9), которую представляли неиммунизированные мыши.
Пример 5: Оценка иммунного ответа у мышей BALB/c после иммунизации препаратами белков HCV с использованием различных адъювантов
Также было изучено применение адъювантов для усиления иммунного ответа, вызываемого вакцинным препаратом. Самок мышей BALB/c иммунизировали внутрибрюшинно либо вакцинным препаратом для формирования оптимального клеточного, либо оптимального гуморального иммунного ответа следующим образом: Группа 1: оптимальный вариант для гуморального ответа в гидроксиде алюминия (AlOH3), Группа 2: оптимальный вариант для гуморального иммунного ответа в фосфате алюминия, Группа 3: оптимальный вариант для гуморального иммунного ответа в адъюванте Фрейнда, Группа 4: оптимальный вариант для гуморального иммунного ответа в масляном адъюванте (Montanide ISA 51), Группа 5: оптимальный вариант для гуморального иммунного ответа без адъюванта, Группа 6: оптимальный вариант для клеточного иммунного ответа в гидроксиде алюминия (AlOH3), Группа 7: оптимальный вариант для клеточного иммунного ответа в фосфате алюминия, Группа 8: оптимальный вариант для клеточного иммунного ответа в адъюванте Фрейнда, Группа 9: оптимальный вариант для клеточного иммунного ответа в масляном адъюванте (Montanide ISA 51), Группа 10: оптимальный вариант для клеточного иммунного ответа без адъюванта. Гуморальный ответ на структурные антигены HCV (ядерный, Е1 и Е2) изучали через пятнадцать дней после окончания проведения иммунизации (недели 0, 1 и 2). Результаты показывают, что несмотря на различия титров антител во всех группах, против антигенов HCV формировался специфический ответ. Уровень антител был сходным в группах, иммунизированных вариантами вакцин в гидроксиде алюминия (AlOH3) и фосфате алюминия. Наивысшие титры антител достигались в группах, иммунизированных препаратом в адъюванте Фрейнда, за которыми следуют группы, иммунизированные препаратом в Montanide ISA 51. Несмотря на то, что имел место специфический антительный ответ в группах, иммунизированных препаратом без адъюванта, полученные титры антител были наименьшими. Эти результаты показаны на Фиг.14.
Лимфопролиферативный иммунный ответ на вирусные структурные антигены HCV оценивали в клетках селезенки иммунизированных мышей через 15 дней после окончания проведения иммунизации. Самые высокие индексы пролиферации наблюдались в группах, иммунизированных оптимальным вариантом для клеточного ответа в адъюванте Фрейнда и Montanide ISA 51, соответственно. Лимфопролиферативные индексы в группах, иммунизированных препаратами в гидроксиде алюминия и фосфате алюминия, были сходными. Самые низкие уровни клеточного иммунного ответа наблюдались при введении препаратов без адъюванта. Эти результаты показаны на Фиг.15, где был включен отрицательный контроль (Группа 11), который представляли неиммунизированные мыши.
Защитную реакцию против вирусного заражения оценивали путем инокуляции 106 pfu vvRE через 15 дней после окончания проведения иммунизации. Через пять дней после заражения определяли титр вируса в яичниках мышей. Результаты показаны на Фиг.16, в том числе дополнительный отрицательный контроль (Группа 11), который представляли неиммунизированные мыши. У мышей, иммунизированных оптимальным вариантом для гуморального иммунного ответа и оптимальным вариантом для клеточного иммунного ответа, и тот, и другой в адъюванте Фрейнда, наблюдалось соответственно 1,7 и 2,4-Log снижение титра вируса. У мышей, иммунизированных оптимальным вариантом для гуморального иммунного ответа и оптимальным вариантом для клеточного иммунного ответа, усиленного в Montanide ISA 51, наблюдалось соответственно 1,9 и 2,5-Log снижение титра вируса. У мышей, иммунизированных препаратами в солях алюминия, снижение титров вируса в яичниках мышей составило 1,4 Log и 2,1 Log в случае варианта вакцины для индукции оптимального гуморального иммунного ответа и оптимального варианта вакцины для клеточного иммунного ответа соответственно. У животных, иммунизированных без адъювантов, наблюдалось только небольшое снижение титра вируса.
Пример 6: Оценка иммунного ответа у мышей BALB/c после иммунизации препаратами белков HCV, смешанными с другими вирусными антигенами
У мышей оценивали формирование или усиление иммунного ответа другими вирусными антигенами (поверхностным антигеном вируса гепатита B - HBsAg, ядерным антигеном вируса гепатита B - HBcAg), смешанными с препаратами HCV (препарат HVC для формирования оптимального клеточного иммунного ответа). Мышей BALB/c иммунизировали внутрибрюшинно: Группа 1 вакцинный препарат HBsAg-HCV, группа 2 - вакцинный препарат HBcAg-HCV, группа 3 -вакцинный препарат HCV, группа 4 - HBsAg и группа 5 - HBcAg. В используемой схеме иммунизации инокуляцию проводили на 0, 2 и 4 неделе, а иммунный ответ исследовали через 15 дней после последней иммунизации. Оценивали антительный ответ на структурные антигены HCV (ядерный, E1 и E2), а также на HBcAg и HBsAg. Наблюдали формирование антиген-специфического иммунного ответа. Образовались специфические антитела против HBcAg и HBsAg. Хотя титры антител против HBcAg по группе 2 (вакцинный препарат HBcAg-HCV) были несколько ниже по сравнению с титрами антител, полученными в контрольной группе (группа 5 - только HBcAg), эти различия не были статистически значимыми. Титры против HBsAg были выше в группе 1 (вакцинный препарат HBsAg-HCV) по сравнению с контрольной группой 4 (только HBsAg), указывая на тенденцию усиления иммунного ответа на этот антиген, хотя наблюдаемые различия не были статистически значимыми. Эти результаты показаны на Фиг.17.
Оценивали лимфопролиферативную иммунную реакцию на структурные вирусные антигены HCV в клетках селезенки иммунизированных мышей, а также на HBcAg и HBsAg через 15 дней после окончания проведения иммунизации. Как показано на Фиг.18, не наблюдалось различий в индексах стимуляции, при сравнении смеси вакцинного препарата HCV и вирусных антигенов HBcAg или HBsAg с соответствующими контролями. В каждом случае формировался антиген-специфичный иммунный ответ. При сравнении индексов стимуляции исследуемых групп статистически достоверных различий не наблюдалось. В этот эксперимент была включена дополнительная группа отрицательного контроля (Группа 6), которую представляли неиммунизированные мыши.
Защитную реакцию на вирусное заражение иммунизированных мышей оценивали путем инокуляции 106 pfu vvRE через 15 дней после окончания проведения иммунизации. Через пять дней после заражения в яичниках мышей определяли титр вируса. На Фиг.19 показаны полученные результаты, причем наблюдалось, что только в группах мышей, иммунизированных вакцинным препаратом HCV, смешанным или не смешанным с вирусными антигенами HBV (HBsAg или HBcAg), определялось приблизительно 2,5-Log снижение титра вируса. При сравнении титров вирусов статистически значимых различий не наблюдалось. В группах отрицательного контроля для HCV снижения вирусной нагрузки в яичниках зараженных мышей не определялось. В этот эксперимент был включен дополнительный отрицательный контроль (Группа 6), который представляли неиммунизированные мыши.
Пример 7: Оценка иммунного ответа у мышей BALB/c после иммунизации препаратами белков HCV, смешанными с бактериальными антигенами
Формирование или усиление иммунного ответа комбинацией вакцинного препарата против HCV (препарата для формирования оптимального клеточного иммунного ответа на HCV) с бактериальными антигенами (белками наружной мембраны - OMP - Neisseria meningitidis) оценивали после иммунизации мышей. Мышей BALB/c иммунизировали внутрибрюшинно: Группу 1 вакцинным препаратом OMP-HCV, группу 2 - вакцинным препаратом HCV и группу 3 - OMP, действие которого усилено гидроксидом алюминия. Мышей иммунизировали на 0, 2 и 4 неделях, а иммунный ответ изучали через 15 дней после завершения иммунизации. Оценивали гуморальный иммунный ответ на структурные антигены HCV (ядерный, E1 и E2), а также на препарат OMP Neisseria meningitidis, кубинского штамма CU 385/83. На Фиг.20 показано, что уровни антител, образовавшихся под действием комбинации вакцинного препарата HCV с белками наружной мембраны, были выше уровней антител, образовавшихся под действием одного OMP и вакцинного препарата HCV; определяя ответ на HCV независимые антигены и OMP. Эти различия не были статистически значимыми.
Оценивали лимфопролиферативную реакцию в клетках селезенки иммунизированных мышей на структурные антигены HCV, а также на OMP Neisseria meningitides CU 385/83 Cuban isolate, через 15 дней после окончания проведения иммунизации. Как показано на Фиг.21, существует повышение индексов стимуляции в клетках селезенки мышей, иммунизированных вакцинным препаратом OMP-HCV, при их стимуляции структурными антигенами HCV и OMP, по сравнению с наблюдаемыми в контрольных группах. Эта стимуляция была антигенспецифичной. В этот эксперимент была включена дополнительная группа отрицательного контроля (Группа 4), которую представляли неиммунизированные мыши.
Защитную реакцию иммунизированных мышей на вирусное заражение оценивали путем инокуляции 106 pfu vvRE, через 15 дней после окончания проведения иммунизации. Через пять дней после заражения определяли титр вируса в яичниках зараженных мышей. Полученные результаты показаны на Фиг.22. В обеих группах было 2-Log снижение титра вируса (вакцинный препарат HCV и вакцинный препарат OMP-HCV). В контрольной группе (мыши, иммунизированные OMP) снижения титра вируса не наблюдалось. То же происходило в другой группе отрицательного контроля (Группа 4), включенной в этот эксперимент, в которой мыши были неиммунизированы.
Функциональную активность антител, образовавшихся против менингококков, оценивали с помощью анализа бактерицидной активности сыворотки. В этом анализе определяют способность антител уничтожать бактерии in vitro, в присутствии экзогенного источника комплемента. Результаты, приведенные в Таблице 3, указывают на то, что смесь вакцинного препарата HCV с OMP ни ингибировала, ни усиливала образование функциональных антител, специфичных к Neisseria meningitidis, поскольку бактерицидные титры сыворотки были сходными в группах 1 и 3.
Бактерицидная активность сыворотки в отношении штамма CU 385/83 Neisseria menigitidis
Пример 8: Оценка иммунного ответа мышей BALB/c после иммунизации препаратом белков HCV, смешанным с полисахаридами
Влияние на иммунный ответ, который вырабатывается под действием полисахаридов капсулы (полисахарид капсулы Neisseria meningitidis серогруппы C - конъюгированный с белком-носителем P64k), в смеси с вакцинным препаратом (вакцинный препарат, который индуцирует оптимальный клеточный иммунный ответ) оценивали после внутрибрюшинной иммунизации мышей BALB/c: группу 1: конъюгатом (MenC-P64k)-вакцинный препарат против HCV, группу 2: вакцинным препаратом против HCV, группу 3: конъюгатом (MenC-P64k). Схема иммунизации включала в себя инокуляции на 0, 2, 4 неделях, а иммунный ответ изучали через 15 дней после последней иммунизации.
Оценивали гуморальный иммунный ответ на структурные антигены HCV (ядерный, белки E1 и E2), а также на полисахарид капсулы Neisseria meningitidis серогруппы C.
На Фиг.23 показано, что уровни антител, вырабатываемых под действием комбинации вакцинного препарата против HCV, смешанного с конъюгатом, были сходны с уровнями, полученными под действием обоих компонентов независимо друг от друга, при оценке ответов на структурные белки HCV, а также на полисахарид капсулы Neisseria meningitidis серогруппы C.
Оценивали лимфопролиферативную реакцию в клетках селезенки иммунизированных мышей на структурные белки HCV. Как показано на Фиг.24, клетки селезенки мышей, иммунизированных вакцинной комбинацией MenC-P64k-вакцинный препарат против HCV и вакцинным препаратом против HCV, стимулировали антигенами структурной области HCV в отличие от контрольных групп, иммунизированных только конъюгатом. В этот эксперимент была включена другая группа отрицательного контроля, группа 4, включающая в себя неиммунизированных мышей.
Защитную реакцию иммунизированных мышей после вирусного заражения оценивали путем инокуляции 106 pfu vvRE, через 15 дней после окончания проведения иммунизации. Через пять дней после заражения определяли титрование вирусной нагрузки в яичниках иммунизированных и зараженных мышей. На Фиг.25 показаны результаты, в которых нет различий в вирусной нагрузке, определенной в яичниках мышей, иммунизированных вакцинным препаратом HCV с конъюгатом (MenC-P64k) или без него. В обеих группах наблюдалось 2,1-Log снижение титра вируса по сравнению с контрольной группой. Различия были статистически значимыми, когда титры вируса в этих группах сравнивали с отрицательным контролем (мыши, иммунизированные только конъюгатом MenC-P64k). В этот эксперимент был включен дополнительный отрицательный контроль (Группа 4), который представляли неиммунизированные мыши.
Функциональную активность антител, образовавшихся против Neisseria meningitidis серогруппы C, оценивали с помощью бактерицидного анализа, измеряя in vitro способность антител к уничтожению бактерий, в присутствии источника экзогенного комплемента. В этом отношении результаты, показанные в Таблице 4, указывают на то, что смесь вакцинного препарата HCV-MenC-P64K конъюгата не влияла на выработку специфичных и функциональных антител против Neisseria meningitidis, продуцируемых под действием конъюгата MenC-P64K.
Бактерицидные титры сыворотки против Neisseria meningitidis серогруппы C
Пример 9: Оценка иммунного ответа мышей BALB/c после иммунизации препаратами белков HCV, смешанными с плазмидами, кодирующими вирусные антигены
Влияние плазмид для ДНК иммунизации, кодирующих поверхностный антиген (pAEC-M7-HBsAg) и ядерный антиген (pAEC-M7-HBcAg) вируса гепатита B, на формирование иммунного ответа при смешивании с вакцинным препаратом против HCV (оптимальным вариантом для клеточного ответа) изучали после внутримышечной иммунизации мышей BALB/c: Группу 1 - pAEC-M7, группу 2 - pAEC-M7-HCV вакцинным препаратом, группу 3 - pAEC-M7-HBsAg, группу 4 - pAEC-M7-HBsAg-HCV вакцинным препаратом, группу 5 - pAEC-M7-HBcAg, группу 6 - pAEC-M7-HBcAg-HCV вакцинным препаратом, группу 7 - вакцинным препаратом против HCV. Животных иммунизировали на 0, 2, 4 и 6 неделях. Оценивали гуморальный иммунный ответ на структурные антигены HCV (ядерный, E1 и E2), а также на поверхностный (HBsAg) и ядерный антигены (HBcAg) вируса гепатита B. Полученные результаты представлены на Фиг.26, на которой видно формирование специфического ответа на каждый из изучаемых антигенов. В основном, при смешивании плазмид с вакцинным препаратом против HCV, титры антител к структурным антигенам HCV были выше титров, полученных под действием только вакцинного препарата против HCV. Несмотря на эту тенденцию, наблюдаемые различия не были статистически значимыми. При иммунизации животных плазмидами, кодирующими HBsAg и HBcAg, либо в отдельности, либо в смеси с вакцинным препаратом против HCV, титры антител к антигенам были схожими.
Оценивали лимфопролиферативную реакцию в клетках селезенки иммунизированных мышей на структурные антигены HCV, а также на HBsAg и HBcAg, через 15 дней после окончания проведения иммунизации. Как показано на Фиг.27, при сравнении вакцинного препарата против HCV, смешанного с плазмидами, кодирующими антигены вируса гепатита B (HBsAg или HBcAg), с соответствующими контролями статистически значимого различия индексов стимуляции, не наблюдалось. В этот эксперимент была включена дополнительная группа отрицательного контроля (Группа 8), включающая в себя неиммунизированных мышей. В каждом случае формировался антиген-специфичный ответ. При сравнении исследуемых групп выявленные различия индексов стимуляции не были статистически значимыми. В то же время, при стимуляции клеток селезенки структурными антигенами HCV, не наблюдалось статистически значимых различий между группами мышей, иммунизированных вакцинным препаратом против HCV, в смеси с плазмидами или без них.
Защитную реакцию после вирусного заражения иммунизированных мышей оценивали путем инокуляции 106 pfu vvRE, через 15 дней после окончания проведения иммунизации. Вирусную нагрузку в яичниках зараженных мышей оценивали через пять дней после заражения. Полученные результаты показаны на Фиг.28, где только в группах, иммунизированных вакцинным препаратом против HCV, либо в смеси с плазмидами, либо без них, наблюдалось приблизительно 2-Log снижение титра вируса. Не было статистически значимых различий между этими группами при сравнении титров вируса. В группах отрицательного контроля HCV снижения вирусной нагрузки в яичниках иммунизированных мышей не было. В этот эксперимент была включена дополнительная группа отрицательного контроля (Группа 8), которую представляли неиммунизированные мыши.
Пример 10: Оценка иммунного ответа у мышей BALB/c после первичной иммунизации ДНК композициями, кодирующими структурную область HCV, и повторной иммунизации изучаемыми препаратами белков HCV
Комбинацию первичной иммунизации ДНК и реиммунизации с вакцинным препаратом против HCV (для оптимального клеточного иммунного ответа) изучали у мышей BALB/c. Схема иммунизации приведена в Таблице 5.


Мышей BALB/c иммунизировали композицией плазмидной ДНК (Duecas-Carrera, et al. (2002). Enhancement of the immune response generated against the hepatitis C virus envelope proteins after DNA vaccination with polyprotein-encoding plasmids. Biotechnol Appl Biochem, 35, 205-212) внутримышечно, и реиммунизирующие дозы вводили внутримышечно.
Оценивали гуморальный иммунный ответ на структурные антигены HCV (ядерный, E1 и E2). Полученные результаты показаны на Фиг.29. Никаких подтверждений усиления антительного ответа на ядерный антиген HCV обнаружено не было. Уровни антител, определенные в группах, иммунизированных белковыми вариантами, были выше уровней антител в группах, иммунизированных ДНК, хотя эти различия не были статистически значимыми.
Гуморальный иммунный ответ на белок Е1 усиливался при введении вторичной инъекции антигена с вакцинным препаратом против HCV. В этом случае более высокие уровни антител определяли в группах мышей, иммунизированных белковыми вариантами, по сравнению с группами, иммунизированными ДНК. Различия, наблюдаемые в уровнях антител против Е1 до и после реиммунизирующей дозы, не были статистически значимыми.
Уровни антител, определяемые к белку Е2 с использованием этой схемы иммунизации, были выше у животных, иммунизированных белковыми составами, по сравнению с антительным ответом, полученным у мышей, иммунизированных ДНК. После реиммунизирующей дозы уровни антител, определяемые против белка Е2, не изменились.
Оценивали лимфопролиферативную реакцию в клетках селезенки иммунизированных мышей на структурные антигены HCV через 15 дней после последней иммунизации. Как показано на Фиг.30, несмотря на повышение лимфопролиферативного индекса в клетках селезенки мышей, ревакцинированных белковыми составами, различия не были статистически значимыми. Подобным образом лимфопролиферативная реакция на структурные антигены HCV у мышей, иммунизированных ДНК и реиммунизированных белками, заметно усиливалась на белок Е1. Различия между группами не были статистически значимыми.
Защитную реакцию на вирусное заражение иммунизированных мышей оценивали путем инокуляции 106 pfu vvRE через 15 дней после последней иммунизации. Через пять дней после заражения определяли титр вируса в яичниках иммунизированных и зараженных мышей. На Фиг.31 показаны полученные результаты, где установлено приблизительно 2-Log снижение вирусной нагрузки у животных, иммунизированных вакцинным препаратом для получения оптимального клеточного ответа. У мышей, иммунизированных ДНК, было показано 1,2-Log снижение титра вируса. После реиммунизирующей дозы с вакцинным составом для оптимального клеточного иммунного ответа наблюдалось 6,6-Log снижение титра вируса у животных, иммунизированных сначала составом для оптимального клеточного ответа. 1,8-Log снижение титра вируса наблюдалось у мышей, примированных ДНК и реиммунизированных вакцинным препаратом HCV. В других группах снижения вирусной нагрузки в яичниках мышей не наблюдалось.
Пример 11: Оценка иммунного ответа у мышей BALB/c после иммунизации препаратами белков HCV, с учетом различных способов получения
Индукцию иммунного ответа на рекомбинантные структурные белки HCV изучали после получения иммуногена различными способами. Были изучены следующие варианты: (1) Перемешивали три антигена и затем усиливали гидроксидом алюминия, (2) каждый антиген сначала отдельно усиливали гелем гидроксида алюминия, а затем перемешивали для получения конечной композиции. Мышей BALB/c иммунизировали обеими композициями. Использовалась вакцинная композиция для индукции оптимального клеточного ответа у мышей BALB/c.
Установленная схема иммунизации была 0, 2 и 4 недели. Иммунный ответ изучали через 15 дней после окончания проведения иммунизации.
Оценивали антительный ответ на антигены, присутствующие в вакцинном препарате против HCV (ядерный, E1 и E2). На Фиг.32 показано, что уровни антител, образовавшихся против каждого антигена, были сходными; различия между изучаемыми группами не были статистически значимыми.
Оценивали лимфопролиферативную реакцию в клетках селезенки иммунизированных мышей на структурные антигены HCV. Как показано на Фиг.33, способ получения иммуногенов HCV не влиял на антиген-специфическую лимфопролиферативную реакцию. Различия между изучаемыми группами не были статистически значимыми. В этот эксперимент была включена другая контрольная группа (группа 3), которую представляли неиммунизированные мыши.
Защитную реакцию иммунизированных мышей на вирусное заражение оценивали путем инокуляции 106 pfu vvRE, через 15 дней после последней иммунизации. Через пять дней после заражения определяли титр вируса в яичниках иммунизированных и зараженных мышей. Количество вируса, определенное в яичниках иммунизированных мышей, выраженное как Log титра вируса, указывало на то, что среди изучаемых групп различия не были статистически значимыми. Эти результаты показаны на Фиг.34. В этот эксперимент была включена другая контрольная группа (группа 3), которую представляли иммунизированные мыши.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИННЫЙ СОСТАВ, ПОТЕНЦИРОВАННЫЙ КОМБИНАЦИЕЙ ДНК И АНТИГЕНА | 2002 |
|
RU2294212C2 |
| ХИМЕРНЫЕ АНТИГЕНЫ ДЛЯ ВАКЦИНЫ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2013 |
|
RU2639504C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ТЕРАПЕВТИЧЕСКОГО ПРИМЕНЕНИЯ | 2004 |
|
RU2362586C2 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕННЫХ АГРЕГАТОВ И ИХ ПРИМЕНЕНИЕ В ПРЕПАРАТАХ | 2001 |
|
RU2266754C2 |
| ОЛИГОМЕРНАЯ ЧАСТИЦА, ИНДУЦИРУЮЩАЯ ИММУНИТЕТ ПРОТИВ ВИРУСА ГЕПАТИТА С, СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРНОЙ ЧАСТИЦЫ, КОМПОЗИЦИЯ, СПЕЦИФИЧЕСКОЕ АНТИТЕЛО, НАБОР (ВАРИАНТЫ), ИММУНОЛОГИЧЕСКИЙ АНАЛИЗ И ВАКЦИНА ПРОТИВ ВИРУСА ГЕПАТИТА С | 1999 |
|
RU2247729C2 |
| РЕКОМБИНАНТНЫЙ МОДИФИЦИРОВАННЫЙ ВИРУС ВАКЦИНЫ АНКАРА, СПОСОБНЫЙ ЭКСПРЕССИРОВАТЬ СТРУКТУРНЫЕ АНТИГЕНЫ ВИРУСА ГЕПАТИТА С | 2002 |
|
RU2270860C2 |
| ПРИМЕНЕНИЕ РАСАР В КАЧЕСТВЕ МОЛЕКУЛЯРНОГО АДЪЮВАНТА ДЛЯ ВАКЦИН | 2012 |
|
RU2580294C2 |
| СУППОЗИТОРИИ ДЛЯ ИММУНОПРОФИЛАКТИКИ ВИРУСНЫХ ИНФЕКЦИЙ | 2004 |
|
RU2296560C2 |
| ХИМЕРНЫЕ ВАКЦИННЫЕ АНТИГЕНЫ ПРОТИВ ВИРУСА КЛАССИЧЕСКОЙ ЧУМЫ СВИНЕЙ | 2007 |
|
RU2406534C2 |
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |
Изобретение относится к области вирусологии. Вакцинная композиция для лечения и профилактики гепатита С содержит в качестве компонентов структурные белки вируса гепатита С в определенных соотношениях. Композиция обладает усиливающим действием в развитии иммунного ответа против вируса гепатита С. Также описаны комбинированные вакцины против патогенных организмов, включающие эту вакцинную композицию. Изобретение может быть использовано в медицине. 3 н. и 7 з.п. ф-лы, 36 ил., 5 табл.
1. Вакцинная композиция для создания защитного клеточного иммунного ответа против инфекции вируса гепатита С, характеризующаяся оптимальной комбинацией структурных антигенов вируса гепатита С (ядерный, Е1 и Е2), где количество используемых антигенов находится в интервале соотношений [(1-50):(120-180):(120-180)]:[ядерный:Е1:Е2], и вводимая с использованием адъюванта любого типа.
2. Вакцинная композиция по п.1, где количество используемых антигенов представляет собой соотношение (1:160:160):(ядерный:Е1:Е2).
3. Вакцинная композиция по п.1 или 2, которая вводится согласно различным схемам иммунизации.
4. Комбинированные вакцинные композиции против вирусных и/или бактериальных заболеваний, где смесь структурных антигенов вируса гепатита С в качестве активных компонентов объединена, в соответствии с п.1 или 2, с полисахаридами (конъюгированными или неконъюгированными) и/или вирусными, и/или бактериальными антигенами.
5. Комбинированная вакцинная композиция против вирусных и/или бактериальных заболеваний, где смесь структурных антигенов вируса гепатита С в качестве активных компонентов объединена, в соответствии с п.1 или 2, с плазмидами, кодирующими антигены инфекционных агентов.
6. Комбинированная вакцинная композиция по п.4, где используемыми вирусными антигенами являются ядерный и/или поверхностный антиген вируса гепатита В.
7. Комбинированная вакцинная композиция по п.4, где используемыми бактериальными антигенами являются белки наружной мембраны N. meningitidis, отдельно или в составе вакцинной композиции.
8. Комбинированная вакцинная композиция по п.4, где полисахаридные антигены, конъюгированные или неконъюгированные с белками-носителями, используются в качестве активных компонентов в составах против бактериальных агентов.
9. Вакцинная композиция по п.1 или 2, которая вводится согласно комбинированным схемам с другими кандидатными вакцинами на основе ДНК конструкций, живых вирусных векторов, пептидов и белков.
10. Вакцинная композиция по п.1 или 2 для индукции иммунного ответа против вируса гепатита С у пациентов, страдающих хроническим гепатитом С, циррозом и раком печени.
| WO 9821338, 22.05.1998 | |||
| Распылитель жидкости | 1960 |
|
SU137869A1 |
| СПОСОБ РАЗДЕЛЕНИЯ ПОБОЧНЫХ ПРОДУКТОВ И ОТХОДОВ МЕТАЛЛУРГИЧЕСКОЙ ПРОМЫШЛЕННОСТИ И УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1995 |
|
RU2097091C1 |

