Инфекция человека респираторно-синцитиальным вирусом (RSV) варьирует от бессимптомного до тяжелого заболевания дыхательных путей. У младенцев и детей респираторно-синцитиальный вирус (RSV) рассматривается как одна из наиболее важных причин заболевания нижних дыхательных путей во всех географических зонах мира. RS вирус превосходит все другие микробные патогены в качестве причины пневмонии и бронхиолита у младенцев до 1 года и является главной причиной летального заболевания дыхательных путей у таких младенцев. Фактически все дети инфицируются в возрасте двух лет. Реинфекция имеет место с заметной частотой у детей старшего возраста и у молодых взрослых людей (Chanock et al. , in Viral Infection of Humans, 3d ed., A.S. Evans. ed., Plenum Press, N.Y. (1989)). Хотя большая часть взрослых людей не имеет серьезных заболеваний, вызываемых инфекцией RS вирусом, пожилые больные и индивидуумы с нарушенной иммунной системой могут иметь тяжелые и, возможно, угрожающие жизни инфекции.
Лечение RSV инфекции было проблематичным. Маленькие дети имеют ответные реакции в виде образования сывороточных и секреторных антител на антигены RSV и поэтому страдают от более тяжелых инфекций, тогда как кумулятивный иммунитет, по-видимому, защищает более старших детей и взрослых от серьезных форм этой инфекции. Было показано, что одно антивирусное соединение, рибавирин, может быть перспективным в лечении инфицированных младенцев в тяжелом состоянии, хотя не получены доказательства того, что оно сокращает продолжительность или уменьшает необходимость поддерживающей терапии.
Механизмы иммунитета к инфекции RSV недавно стали центром внимания исследователей. По-видимому, секреторные антитела являются наиболее важными в защите верхних дыхательных путей, тогда как высокие уровни сывороточных антител, возможно, играют главную роль в устойчивости к инфекции RSV в нижних дыхательных путях. Очищенный человеческий иммуноглобулин, содержащий высокий титр нейтрализующих антител к RSV, может оказаться применимым в иммунотерапевтических подходах к лечению тяжелого заболевания нижних дыхательных путей у младенцев и детей младшего возраста. Однако препараты иммуноглобулина имеют серьезные недостатки, такие как возможность передачи находящихся в крови вирусов и трудности и большие расходы при их приготовлении и хранении.
Несмотря на настоятельную потребность в эффективной вакцине против RS вируса, в частности, для младенцев и детей младшего возраста, прежние попытки получения надежной и эффективной вакцины были неудачными. Вакцина с инактивированным формалином вирусом, исследованная в середине 1960-х годов, не давала защиты против инфекции RS вирусом или заболеванием. Вместо этого заболевание обострялось при последующей инфекции RS вирусом. Kim et al., Am. J. Epidemiol 89:422-434; Chim et al., Am.J. Epidemiol. 89: 449-463 (1969); Kapikian et al., Am. J. Epidemiol. 89:405-421 (1969).
Для того чтобы обойти проблемы, связанные с инактивированными вакцинами и возможным изменением вируснейтрализующей антигенной детерминанты, усилия были направлены на получение аттенуированных RS мутантов. Friedewald et al., J. Amer. Med. Assoc. 204:690-694 (1968) сообщили о получении низкотемпературного пассированного мутанта RS вируса, который, по-видимому, обладал достаточной аттенуацией для того, чтобы быть кандидатом на вакцину. Этот мутант проявлял слегка увеличенную эффективность роста при 26oC по сравнению с родительским вирусом дикого типа, но его репликация была нечувствительной к температуре и не была адаптирована к холоду. Однако пассированный на холоду мутант был аттенуированным (ослабленным) для взрослых. Хотя этот мутант был достаточно аттенуированным и иммуногенным для младенцев и детей, ранее инфицированных RSV (например, серопозитивных индивидуумов), он сохранял низкий уровень вирулентности для верхних дыхательных путей серонегативных младенцев. Этот мутант был пассирован в культуре клеток почки быка при низкой температуре (26oC) и в результате приобрел аттенуирующие мутации круга хозяев. Приобретение этих мутаций позволяло мутанту эффективно реплицироваться в бычьих тканях, тогда как те же самые мутации ограничивали рост мутанта в дыхательных путях человека по сравнению с родительским штаммом A2 RVS.
Подобно этому Garpure et al. , J. Virol. 3:414-421 (1969) сообщили о выделении чувствительных к температуре (ts) мутантов, которые также были перспективными кандидатами на вакцины. Один мутант, ts-1, был подвергнут детальному исследованию в лаборатории и на добровольцах. Мутант вызвал бессимтомную инфекцию у взрослых добровольцев и устойчивость к введению вируса дикого типа через 45 дней после иммунизации. Опять-таки, в то время как серопозитивные младенцы и дети подвергались бессимптомной инфекции, серонегативные младенцы обнаруживали признаки ринита и другие легкие симптомы. Кроме того, была обнаружена нестабильность ts фенотипа, хотя вирус, проявляющий частичную или полную потерю чувствительности к температуре, представлял небольшую часть извлекаемого из вакцин вируса и не был ассоциирован с иными признаками заболевания, чем легкий ринит.
Таким образом, эти исследования выявили, что пассированные на холоду и чувствительные к температуре штаммы были недостаточно аттенуированы и вызывали легкие симптомы заболевания у некоторых вакцинированных индивидуумов, в частности у серонегативных младенцев, тогда как другие штаммы избыточно аттенуированы и не реплицируются в достаточном количестве для индуцирования защитных иммунных ответных реакций. (Wright et al., Infect. Immun. 37:397-400 (1982)). Генетическая нестабильность, позволяющая мутантам-кандидатам на вакцины терять их устойчивый к температуре фенотип, также была нарушающим планы открытием. См. Hodes et al., Proc. Soc. Exp. Biol. Med. 145:1159-1164 (1974). Mc Intosh et al., Pediatr. Res. 8: 689-696 (1974) и Belshe et al., J. Med. Virol. 3:101-110 (1978).
Оставив подход, предусматривающий применение вакцин с аттенуированным RS вирусом, исследователи испытали потенциальные субъединичные вакцины, основанные на гликопротеинах оболочки RS вируса, очищенных из лизатов инфицированных клеток. Эти гликопротеины индуцировали устойчивость к инфекции RS вирусом в легких хлопковых (cotton) крыс, Walsh et al., J. Infect. Dis. 155: 1198-1204 (1987), но индуцированные антитела имели очень слабую вируснейтрализующую активность и иммунизация грызунов очищенной субъединичной вакциной приводила к усилению заболевания (Murphy et al., Vaccine 8: 497-502 (1990)).
Исследовались такие вакцины, основанные на рекомбинантном вирусе осповакцины, который экспрессирует гликопротеин оболочки F или G. Эти рекомбинанты экспрессируют гликопротеины RVS, которые неотличимы от аутентичной вирусной копии, и мелкие грызуны, инфицированные внутрикожно рекомбинантными вирусами осповакцины, экспрессирующими F- и G-RSV, обнаружили высокие уровни специфических антител, которые нейтрализовали инфекционность вируса. Действительно, инфицирование хлопковых крыс рекомбинантами вируса осповакцины-F стимулировала почти полную устойчивость к репликации RSV в нижних дыхательных путях и значительную устойчивость в верхних дыхательных путях. Olmsted et al. , Proc. Natl. Asad. Sci. USA. 83:7462-7466 (1986). Однако иммунизация шимпанзе рекомбинантами вирус осповакцины -F и вирус осповакцины -G почти не давала защиты против введения RSV/Collins et al., Vaccine 8: 164-168 (1990)). Это привело к заключению, что этот подход, по-видимому, не сможет обеспечить удачной вакцины.
В то время как исследователи испытывали несколько различных подходов для получения эффективной и надежной RS вакцины в течение ряда лет. RS вирус оставался наиболее обычной причиной тяжелого вирусного заболевания нижних дыхательных путей у младенцев и детей. В результате остается настоятельная потребность в надежной вакцине, которая способна предотвращать серьезное заболевание в этой популяции, требующее часто госпитализации, и предотвращать заболевание у других индивидуумов. Совершенно удивительно, данное изобретение решает эти и близкие к ним проблемы.
Данное изобретение обеспечивает вакцинные композиции аттенуированного респираторно-синцитиального вируса. Аттенуированный вирус обеспечен в количестве, достаточном для индуцирования иммунного ответа в хозяине-человеке, в соединении с физиологически приемлемым носителем и может иногда содержать адъювант для усиления иммунного ответа хозяина. Изобретение рассматривает несколько различных антигенных подгрупп аттенуированного RS вируса, которые произведены из неполностью ослабленного RS вируса и обладают свойствами, до сих пор не обнаруживаемыми аттенуированными RS вирусами, описанными ранее в литературе. В одном из вариантов изобретения аттенуированный вирус содержит RS вирус с ограниченным кругом хозяев, неполностью ослабленный пассированием на холоду (cp RSV), в который введены по меньшей мере одна или несколько дополнительных мутаций для получения вируса и его потомства, имеющих чувствительный к температуре фенотип (ts), обозначаемый далее cpts RSV. В другом варианте RS вирус с ограниченным кругом хозяев, неполностью ослабленный пассированием на холоду (cp RSV), адаптируют к холоду (ca) пассированием при более пониженных температурах для введения дополнительных ограничивающих рост мутаций. Еще в одном варианте неполностью ослабленные ts мутанты RSV, такие как ts-4 и ts-1 NGI RSV, ослаблены далее введением дополнительных мутаций. Аттенуированные производные ts или cp штаммов получают несколькими путями, но предпочтительно введением дополнительных чувствительных к температуре мутаций при помощи химического мутагенеза, дальнейшим пассированием в культуре при ослабляющих температурах 20-24oC или введением мутаций малых бляшек (Small plaque) (cp) и отбором производных, которые более ограничены в репликации, чем неполностью аттенуированный родительский мутантный штамм. Этот аттенуированный вирус изобретения принадлежит к антигенной подгруппе либо A, либо B и вирус из обеих подгрупп может быть объединен в вакцинных препаратах для большего охвата преобладающих RSV инфекций. Вакцину обычно готовят в дозе 103 - 106 бляшкообразующих единиц (PFU) или в более высокой дозе для максимальной эффективности.
В других вариантах изобретение обеспечивает способы стимулирования иммунной системы индивидуума для индуцирования защиты против респираторно-синцитиального вируса. Способы предусматривают введение индивидууму иммунологически достаточного количества RSV, аттенуированного введением мутаций, которые придают характер ts, ca и (или) sp фенотипа RSV, исходно неполностью аттенуированному ts мутацией (мутациями) или пассированием при низкой температуре, например, при 26oC. Ввиду потенциальных тяжелых последствий инфекции RSV у новорожденных, серонегативных и серопозитивных младенцев и детей младшего возраста, а также у пожилых людей, иммунизация в соответствии с данными способами обычно наиболее полезна для этих индивидуумов. В большинстве случаев аттенуированный вирус вводят в дыхательные пути индивидуума, предпочтительно интраназально в виде аэрозоля или капель.
В дальнейших вариантах изобретение обеспечивает чистые культуры аттенуированного RS вируса, в которых вирус был более полно аттенуирован дальнейшей дериватизацией идентифицированных ранее ts или cp мутантов. Этот аттенуированный вирус способен вызывать защитный иммунный ответ в инфицированном хозяине-человеке, но в то же время достаточно ослаблен, так что не вызывает нежелательных симптомов тяжелого респираторного заболевания в иммунизированном хозяине. Аттенуированный вирус может находиться в супернатанте клеточной культуры, может быть выделен из культуры или частично, либо полностью очищен. Вирус может быть лиофилизирован и может быть соединен со множеством других компонентов для хранения или доставки хозяину, по желанию.
Описание характерных вариантов
Данное изобретение обеспечивает RS вирус, пригодный для применения в качестве вакцины для человека. RS вирус, описанный здесь, получают введением дополнительных мутаций в штаммы во время роста вируса в клеточных культурах, к которым добавляют химический мутаген, отбором вируса, подвергнутого пассированию при субоптимальной температуре для введения ограничивающих рост мутаций, или отбором подвергнутого мутагенезу вируса, который продуцирует малые бляшки в клеточной культуре.
Таким образом, вакцина данного изобретения содержит аттенуированный RS вирус и физиологически приемлемый носитель. Вакцину вводят в иммуногенно достаточном количестве индивидууму, нуждающемуся в иммунологической защите против RS вируса, например, младенцу, ребенку, пожилым людям или взрослым для иммуносупрессивной терапии. Вакцина вызывает иммунный ответ, защищающий против серьезного заболевания нижних дыхательных путей, например, пневмонии и бронхиолита, при последующем инфицировании индивидуума RS вирусом дикого типа. Хотя циркулирующий в природе вирус еще способен вызывать инфекцию, в частности, в верхних дыхательных путях, в результате вакцинации очень сильно снижается возможность ринита и возможно повышение устойчивости при последующей инфекции вирусом дикого типа. После вакцинации образуются детектируемые уровни сывороточных и секреторных антител у хозяина, которые способны нейтрализовать гомологичный (той же самой подгруппы) вирус дикого типа in vitro и in vivo. Во многих случаях антитела хозяина будут также нейтрализовать вирус дикого типа другой, невакционной подгруппы. Для достижения более высоких уровней перекрестной защиты, т.е. защиты против гетерологичных штаммов другой подгруппы, предпочтительно вакцинировать индивидуумов аттенуированным вирусом по меньшей мере из одного предпочтительного штамма как подгруппы A, так и B.
Аттенуированный вирус, являющийся компонентом вакцины, находится в ней в выделенном и обычно очищенном виде. Под словом "выделенный" имеют ввиду, что аттенуированный модифицированный RS вирус находится в другой среде по сравнению с обычной природной средой обитания вируса дикого типа, такой как носоглотка инфицированного индивидуума. Более конкретно, "выделенный" обозначает, что аттенуированный вирус находится в виде гетерологичного компонента в клеточной культуре или иной системе. Например, аттенуированный RS вирус данного изобретения может быть продуцирован инфицированной клеточной культурой, отделен от нее и добавлен к стабилизатору, содержащему другие, не встречающиеся в природе RS вирусы, например, вирусы, которые выбраны как аттенуированные по устойчивости к нейтрализующим моноклональным антителам к F-белку, как описано в одновременно представленном U.S. patent application attorney docket 15280-11-2, даваемом здесь в виде ссылки.
Аттенуированный RS вирус данного изобретения обнаруживает очень заметное уменьшение вирулентности при сравнении с вирусом дикого типа, циркулирующего природно в людях. Аттенуированный вирус достаточно ослаблен, так что симптомы инфекции не наблюдаются в большинстве иммунизированных индивидуумов. В некоторых случаях аттенуированный вирус еще способен диссеминировать к невакционированным индивидуумам. Однако его вирулентность достаточно подавлена, так что в вакцинированном или случайном хозяине не бывает серьезных инфекций нижних дыхательных путей.
Уровень аттенуации можно определить, например, определением количества вируса, присутствующего в дыхательных путях иммунизованного хозяина и сравнением этого количества с количеством, продуцируемым RS вирусом дикого типа или другими аттенуированными RS вирусами, которые оценивались как кандидаты вакцинных штаммов. Например, аттенуированный вирус данного изобретения имеет более высокую степень ограничения репликации в верхних дыхательных путях высокочувствительного хозяина, такого как шимпанзе, по сравнению с уровнями репликации вируса дикого типа, например, в 10-1000 раз меньше. Также уровень репликации аттенуированного вакцинного штамма RSV в верхних дыхательных путях шимпанзе был ниже, чем уровень репликации неполностью аттенуированного мутанта A2ts-1RSV. Для дальнейшего снижения развития ринореи, связанной с репликацией вируса в верхних дыхательных путях, идеальный вакцинный вирус-кандидат должен проявлять пониженный уровень репликации как в верхних, так и в нижних дыхательных путях. Однако аттенуированные вирусы данного изобретения должны быть достаточно инфекционными и иммуногенными в человеке для выработки защиты в вакцинированных индивидуумах. Способы определения уровней RS вируса в носоглотке инфицированного хозяина хорошо известны в литературе. Пробы получают аспирацией или вымыванием носоглоточных секреций (выделений) и количество вируса определяют при помощи лабораторного способа. См., например, Belshe et al., J. Med. Virology 1:157-162 (1977). Friedewald et al., J. Amer Med. Assoc. 204: 690-694 (1968); Gharpure et al., J. Virol. 3: 414-421 (1969) и Wright et al., Arch. Ges Virusforsch. 41: 238-247 (1973). Вирус может быть измерен пригодным для этого способом в носоглотке хозяев-животных, таких как шимпанзе.
Для получения удовлетворительно аттенуированного производного вируса данного изобретения мутации вводят в родительский вирусный штамм, который неполностью или частично аттенуирован, такой как ts-1 или ts-4 мутант или cp RSV. Для вируса подгруппы A предпочтительным неполностью аттенуированным родительским вирусом является ts-1 или ts-1 NG-1 или cp RSV, которые представляют собой мутанты штамма A2 подгруппы A или их производные или субклоны.
Частично ослабленные мутанты вируса подгруппы B моно получить биологическим клонированием вируса подгруппы B дикого типа в приемлемом клеточном субстрате и получением из него мутантов пассированием на холоду, а также при помощи химического мутагенеза с образованием ts мутантов или отбором мутантов, образующих малые бляшки. Различные способы отбора можно также комбинировать для получения частично аттенуированных мутантов подгрупп A или B, пригодных для дальнейшей описанной здесь дериватизации.
Как только отобран (отобраны) желаемый частично аттенуированный родительский штамм (штаммы), дальнейшее аттенуирование, достаточное для получение вакцины, приемлемой для применения в человеке, согласно данному изобретению выполняется несколькими путями, как описано здесь.
Согласно данному изобретению, cp мутант может быть подвергнут дальнейшему мутагенезу различными путями. В одном из вариантов способ предусматривает пассирование частично ослабленного вируса в клеточной культуре при прогрессивно более низких, ослабляющих температурах. Например, в то время как вирус дикого типа в типичном случае культивируют приблизительно при 34-35oC, частично аттенуированные мутанты получают пассированием в клеточных культурах (например, в первичных клетках почек быков) при субоптимальных температурах, например, при 26oC. Эти мутанты имеют слабое, но четко выраженное доказательство адаптации к холоду (ca), т.е. повышенную эффективность роста при 26oC по сравнению с родительским вирусом дикого типа, но обычно не ts. Так, в одном способе данного изобретения cp мутант или другой частично ослабленный штамм, например, ts-1 или sp, адаптируют для эффективного роста при пониженной температуре пассированием в клетках MRС-5 или Vero до температуры приблизительно 20-24oC, предпочтительно 20-22oC. Этот отбор мутантного RS вируса во время холодного пассирования в основном исключает какую-либо остаточную вирулентность в производных штаммах по сравнению с частично аттенуированным родителем.
В другом варианте изобретения неполностью аттенуированные штаммы подвергают химическому мутагенезу для введения ts мутаций или, в случае вирусов, которые уже ts (чувствительны к температуре), дополнительные ts мутации достаточны для увеличения стабильности ts фенотипа аттенуированного производного. Способы для введения ts мутаций в RS вирус предусматривают репликацию вируса в присутствии мутагена, такого как 5-фторуридин или 5-фторурацил в концентрации приблизительно 10-3 - 10-5 М, предпочтительно приблизительно 10-4 М, или экспонирование вируса с нитрозогуанидином при концентрации приблизительно 100 мкг/мл в соответствии с общим способом, описанным, например, в Gharpure et al., J. Virol. 3: 414-421 (1969) и Richardson et al. , J. Med. Virol. 3: 91-100 (1978). Можно использовать и другие химические мутагены. Аттенуирование может быть результатом ts мутации почти в любом гене RS вируса. Уровень температурной чувствительности репликации аттенуированного RS вируса изобретения определяют сравнением его репликации при разрешающей температуре с репликацией при нескольких ограничивающих температурах. Самая низкая температура, при которой репликация вируса снижается в 100 раз или более по сравнению с его репликацией при пермиссивной температуре, названа температурой выключения (shutoff). В экспериментальных животных и людях как репликация, так и вирулентность RS вируса коррелируют с температурой выключения мутанта. Репликация мутантов с температурой выключения 39oC умеренно ограничена, тогда как мутанты с температурой выключения 38oC размножаются менее хорошо и симптомы болезни в основном ограничиваются верхними дыхательными путями. Вирус с температурой выключения 35-37oC должен быть полностью ослаблен в человеке. Так, аттенуированный RS вирус изобретения, который чувствителен к температуре, имеет температуру выключения в диапазоне приблизительно 35-39oC, предпочтительно 35-38oC. Добавление свойства чувствительности к температуре частично аттенуированному штамму создает полностью аттенуированный вирус, применимый в вакцинных композициях данного изобретения.
В дополнение к критериям жизнеспособности, ослабленности и иммуногенности свойства производного, которое отбирают, должны также быть несколько возможно стабильными, так чтобы желаемые признаки сохранялись. Генетическая нестабильность ts фенотипа после репликации in vivo была правилом для ts вирусов (Murphy et al., Infect. and Immun. 37: 235-242 (1982)). Затем идеально, если вирус, применяемый в вакцинах данного изобретения, сохраняет жизнеспособность, ослабленность, способность размножаться в иммунизированном хозяине (хотя и с низкими уровнями) и способность эффективно индуцировать иммунный ответ в вакцинированных индивидуумах, достаточный для защиты против серьезного заболевания, вызываемого последующей инфекцией вирусом дикого типа. Очевидно, что известные до сих пор и описанные мутанты RS вируса не отвечают всем этим критериям. Действительно, вопреки ожиданиям, основанным на результатах, сообщенных для известных аттенуированных RS вирусов, некоторые из вирусов данного изобретения, имеющие минимально две-три разные мутации, не только жизнеспособны и более ослаблены, чем прежние мутанты, но и более стабильны генетически in vivo, чем исследованные ранее мутанты, и сохраняют способность стимулировать защитный иммунный ответ и в некоторых случаях расширять защиту, полученную в результате множественных модификаций, например, индуцировать защиту против различных вирусных штаммов или подгрупп или защиту на различной иммунологической основе, например, на основе секреторных, а не сывороточных иммуноглобулинов, на основе клеточного иммунитета и т.п.
Аттенуированный вирус данного изобретения может быть размножен в ряде клеточных линий, которые пригодны для роста RS вируса. RS вирус растет во многих человеческих и животных клетках. Предпочтительными клеточными линиями для размножения аттенуированного RS вируса для применения в вакцинах являются клетки DBS-FRhL-2, MRC-3 и Vero. Наибольшие выходы вируса обычно достигаются в гетероплоидных линиях, таких как клетки Vero. Обычно клетки инокулируют вирусом при множественности заражения в диапазоне 0,001-1,0 или более. Клетки культивируют при условиях, разрешающих репликацию вируса, например, при 30-37oC в течение 3-5 дней или так долго, как это необходимо для достижения требуемого титра. Вирус удаляют из клеточной культуры и отделяют от клеточных компонентов, обычно хорошо известными способами, например, центрифугированием, и, если нужно, очищают далее при помощи способов, известных специалистам данной области.
Вирус, аттенуированный, как описано здесь, может быть испытан в моделях in vitro и in vivo для подтверждения адекватного аттенуирования, генетической стабильности и иммуногенности для применения в вакцинах. В тестах in vitro модифицированный вирус тестируют на sp фенотип. Далее модифицированные вирусы тестируют в животных моделях RS инфекции. Описаны многие животные модели, которые суммированы в Meignier et al., eds., Animal Models of Respiratory Synсytial Virus Infection, Merieux Foundation Publication, (1991), даваемой в виде ссылки. Модель инфекции RSV хлопковых крыс описана в U.S. 4800078 и Prince et al., Virus Res. 3: 193-206 (1985), включенных в ссылки. Эта модель, как считают, предсказывает аттенуирование и эффективность в человеке. Модель RS инфекции приматов с применением шимпанзе предсказывает аттенуирование и эффективность в человеке и описана в деталях в Richardson et al. , J. Med. Virol. 3: 91-100 (1978); Wright et al., Infect. Immun. 37: 397-400 (1982); Crowe et al. , Vaccine (1993) (in press), включенных в ссылки.
Например, было показано, что терапевтическое действие RSV нейтрализующих антител в инфекционных крысах было очень близким к последующему опыту с иммунотерапией обезьян и человека, инфицированных RSV. Действительно, хлопковая (cotton) крыса, по-видимому, является надежным экспериментальным заменителем для изучения ответа инфицированных обезьян и людей на иммунотерапию RSV нейтрализующими антителами. Например, количество RSV нейтрализующих антител, дающих терапевтический эффект на крысах, измеренный по уровню таких антител в сыворотке обработанных животных (т.е. титр RSV нейтрализующей сыворотки 1: 302 - 1:518) лежит в том же самом диапазоне, который продемонстрирован для обезьян (т.е. титр 1:539) или младенцев человека или детей младшего возраста (т.е. 1:877). Терапевтическое действие в крысах проявлялось в виде 100-кратного или большего снижения титра вируса в легком (Prince et al. , J. Virol. 61: 1851-1854), тогда как в обезьянах терапевтическое действие проявлялось в виде 50-кратного снижения титра легочного вируса. (Hemming et al., J. Infect. Dis. 152: 1083-1087 (1985)). Наконец, терапевтическое действие в младенцах и детях младшего возраста, госпитализированных с тяжелым RSV бронхиолитом или пневмонией, проявлялось в виде значительного увеличения оксигенации в подвергнутой лечению группе и значительном снижении количества RSV, извлекаемого из верхних дыхательньых путей прошедших лечение больных. (Hemming et al. , Antimicrob. Agents _ Chemother. 31: 1882-1886 (1987)). Следовательно, на основе этих исследований видно, что хлопковая крыса представляет собой удобную модель для предсказания успеха RSV вакцины у младенцев и детей младшего возраста. Другие грызуны, в том числе хомяки и мыши, должны быть также применимыми, поскольку эти животные могут допускать репликацию в них RSV и имеют температуру внутри тела, подобную температуре человека (Wight et al. , J. Infect. Dis. 122: 501-512 (1970) и Anderson et al., J. Gen. Virol. 71: (1990)).
Для применения в вакцинах аттенуированный вирус данного изобретения можно использовать непосредственно в препарате вакцины или он может быть лиофилизирован, если желательно, при помощи известных протоколов лиофилизации. Лиофилизированный вирус обычно хранят приблизительно при 4oC. Перед использованием лиофизилированный вирус воссоздают в стабилизирующем растворе, например, солевом растворе или в растворе, содержащем SPG, Mg++ и HEPES, с адъювантом или без него, как описано ниже.
Таким образом, RS вирусные вакцины изобретения содержат в качестве активного ингредиента иммунногенно эффективное количество аттенуированного RS вируса, как описано здесь. Аттенуированный вирус может быть введен в хозяина, в частности человеку, с физиологически приемлемым носителем и(или) адъювантом. Применимые носители хорошо известны в этой области и представляют собой, например, воду, содержащую буфер воду, 0,4%-ный солевой раствор, 0,3%-ный глицин, гиалуроновую кислоту и т.п. Полученные водные растворы могут быть упакованы для применения или лиофилизированы, причем лиофилизированный препарат соединяют со стерильным раствором перед введением, как упомянуто выше. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, необходимые для создания близких к физиологическим условий, такие как корректоры pH и буферные средства, корректоры тоничности, смачивающие вещества и т.п., например, ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция, сорбитанмонолаурат, олеат триэтаноламина и т.п.
При инокуляции композицией аттенуированного RS вируса, как описано здесь, через аэрозоль, капли, грубый спрей, пероральным, топическим или иным путем, наиболее предпочтительным для интраназальной доставки, иммунная система хозяина отвечает на вакцину образованием антител, как секреторных, так и сывороточных, специфических для белков RS вируса. В результате вакцинации хозяин становится по меньшей мере частично или полностью иммунным к инфекции RS вирусом или устойчивым к развитию умеренной или тяжелой RS вирусной инфекции, в частности, нижних дыхательных путей.
Вакцинные композиции, содержащие аттенуированный RS вирус данного изобретения, вводят лицу, восприимчивому к инфекции RS вирусом или по иной причине подвергающемуся опасности инфекции RS вирусом, для усиления возможностей собственной иммунной системы индивидуума. Такое количество определено как "иммуногенно эффективная доза". При этом точные количества зависят от состояния здоровья больного и его веса, способа введения, природы препарата и т. д. , но находятся в диапазоне приблизительно 103 - 106 бляшкообразующих единиц (PFU) или более вируса на больного, более типично, приблизительно 104 - 105 PFU вируса на больного. В любом случае вакцинные препараты должны обеспечить количество аттенуированного RV вируса данного изобретения, достаточное для эффективной защиты больного против тяжелой или угрожающей жизни инфекции RS вирусом.
Аттенуированный RS вирус изобретения одной определенной RS подгруппы или одного штамма может быть соединен с аттенуированными вирусами другой подгруппы или других штаммов для получения защиты против множественных RS вирусов. В типичном случае различные модифицированные вирусы должны находиться в смеси и вводиться одновременно, но они могут вводиться и раздельно. Благодаря феномену перекрестной защиты среди определенных штаммов вируса иммунизация одним штаммом может защищать против нескольких различных штаммов той же самой или другой подгруппы.
В некоторых случаях может быть желательным комбинирование вакцин аттенуированного вируса данного изобретения с вакцинами, которые индуцируют защитные ответные реакции на другие агенты, в частности, на другие поражающие детей вирусы. Например, вакцину данного изобретения можно вводить одновременно (в типичном случае раздельно) или последовательно с вакциной против вируса парагриппа, как это описано в Clements et al., J. Clin. Microbiol. 29: 1175-1182 (1991).
Можно проводить одноразовые или множественные введения вакцинных композиций изобретения. Для новорожденных и младенцев до 2 лет множественное введение может быть необходимым для индуцирования достаточных уровней иммунитета. Введение следует начинать на первом месяце жизни и продолжать с интервалами, например, в 2 месяца, шесть месяцев, один год и два года, что необходимо для поддержания достаточных уровней защиты против нативной (дикого типа) RS вирусной инфекции. Подобным образом, взрослым, которые особенно восприимчивы к повторяющейся или тяжелой RS вирусной инфекции, например, работникам медико-санитарной помощи, дневной медицинской помощи, членам семей с детьми младшего возраста, пожилым людям, индивидуумам с нарушенной кардиолегочной функцией и т.д. могут быть необходимы многократные иммунизации для создания и(или) поддержания иммунных ответных реакций. Уровни индуцированного иммунитета могут прослеживаться путем измерения количеств нейтрализующих секреторных или сывороточных антител, и корректированные дозировки или вакцинации могут повторяться при необходимости для поддержания желаемых уровней защиты.
Для иллюстрации (но не для ограничения) приведены следующие примеры.
Пример I
Выделение и характеристика полученных в результате мутагенеза производных пассированного на холоду RSV.
Этот пример описывает химический мутагенез неполностью аттенуированого cp RSV с ограниченным кругом хозяев для получения производных ts и cp штаммов, которые более сильно ослаблены и, следовательно, предпочтительны для применения в вакцинных препаратах RSV.
Был приготовлен родительский исходный запас пассированного на холоду RSV (cp RSV). Вирус Flow Laboratories Lot 3131, родительский cp RSV, который неполностью аттенуировали в человеке, пассировали дважды в клетках MRC-5 при 25oC, в конце разбавляли в два раза в клетках MRC-5 при 25oC, затем пассировали три раза в клетках MRC-5 для получения суспензии cp RSV для мутагенеза.
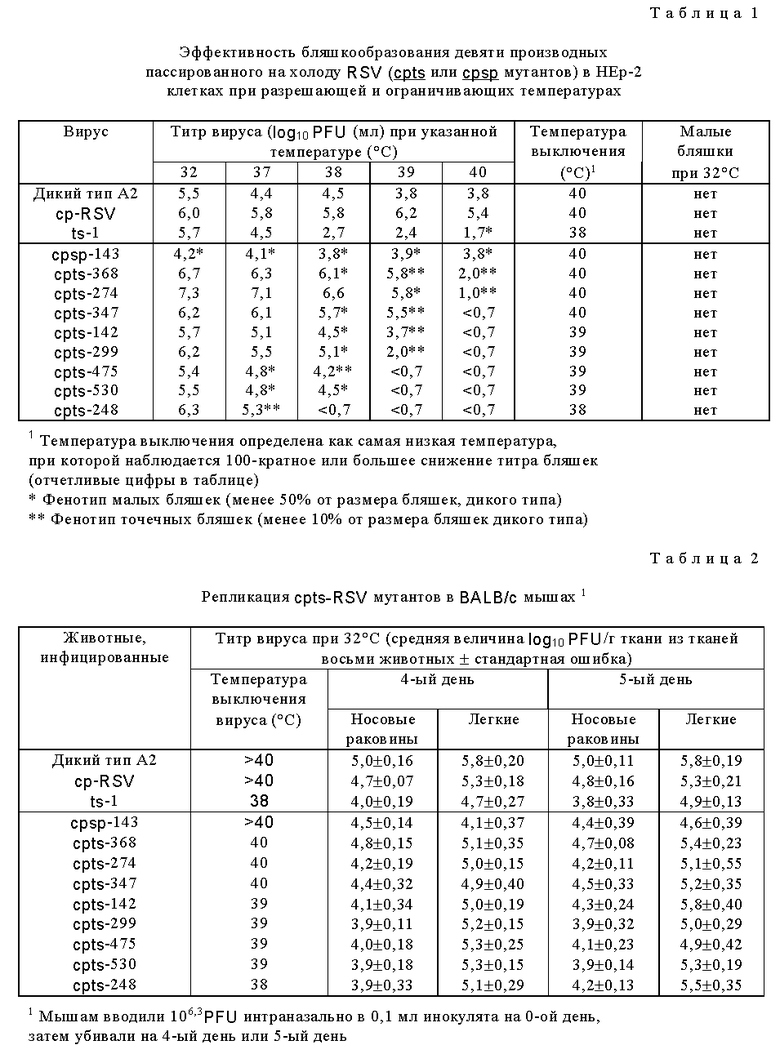
cp RSV подвергали мутагенезу выращиванием родительской исходной популяции в MRC-5 клетках при 32oC в присутствии 5-фторурацила в среде при концентрации 4•10-4 М. В предварительных исследованиях было показано, что эта концентрация является оптимальной, поскольку она вызывала 100-кратное снижение титра вируса на 5-й день роста в культуре клеток по сравнению со средой без 5-фторурацила. Затем подвергнутую мутагенезу исходную популяцию анализировали методом бляшек (пятен) на клетках Vero, которые поддерживались под верхним слоем агара, и после определенного интервала инкубирования бляшки окрашивали красителем нейтральным красным. 854 бляшки извлекали и потомство каждой бляшки отдельно размножали путем выращивания на свежих монослоях клеток Vero. Содержимое каждой из тканевых культур инокулировали потомством одной бляшки подвергнутого мутагенезу cp RSV, собирали отдельно, когда эффекты подтверждения на клетках Vero казались максимальными. Потомство вируса, обнаруживающее чувствительный к температуре (ts) или содержащий малые бляшки (sp) фенотип, находили титрованием пулов бляшек на HEp-2 клетках при 32oC и 38oC. Каждый вирус, обнаруживающий sp фенотип (размер бляшек был уменьшен на 50% или более по сравнению с родительским вирусом при 32oC) или ts фенотип (100-кратное снижение титра при ограничивающей температуре 37o-40oC по сравнению с 32oC) оценивали далее. Эти штаммы биологически клонировали серийной очисткой бляшек на клетках Vero три раза и затем размножали на клетках Vero. Клонированные штаммы титровали при 32o, 37o, 38o, 39o и 40o (в тесте эффективности бляшкообразования (ECP)) для подтверждения их sp и ts фенотипов. Поскольку титры некоторых клонированных штаммов были относительно низкими даже при пермиссивной температуре (32oC), эти вирусы пассировали один раз в HEp-2 клетках, получая вирусные суспензии для анализа in vitro. Фенотипы потомства подвергнутого мутагенезу cp RSV представлены в табл. 1 (табл. 1-17 см. в конце описания).
Одно из мутантных потомств имело фенотип малых бляшек, RSV cpsp-143 (sp обозначает фенотип малых бляшек (sp), остальные мутантные потомства имели ts фенотип. RSV cpts мутанты обнаруживают вариации в способности продуцировать бляшки в монослойных культурах in vitro в диапазоне температур 37oC - 40oC. Так, cpts 368 сохраняет способность продуцировать бляшки при 40oC, тогда как наиболее чувствительный к температуре (ts) вирус, cpts 248, не мог продуцировать бляшки при 38oC. Таким образом, некоторые из подвергнутых мутагенезу cp RSV потомств обнаруживают заметное отличие от родительского cp RS вируса в отношении чувствительности к температуре бляшкообразования.
Исследование репликации и генетической стабильности в мышах
Уровень репликации cp RSV потомства вируса в верхних и нижних дыхательных путях BALB/c мышей изучали на следующем этапе исследований (табл. 2). Было обнаружено, что cpts 530 и cpts 248, два из наиболее чувствительных к температуре ts вирусов (см. табл. 1), имели в 7-12 раз сниженную репликацию в носовых раковинах мышей (табл. 2). Однако ни один из этих вирусов не был ограничен в репликации в легких по сравнению с cp RSV родительским вирусом. Большее ограничение размножения в носовых раковинах чем в легких, не является характерным для ts мутантов, которые обычно имеют более ограниченное размножение в более теплых нижних дыхательных путях (Richman and Murphy, Rev. Jnfect. Dis. 1: 413-433 (1979). Вирус, образовавшийся в легких и носовых раковинах, сохранял ts характер вошедшего вируса (данные не представлены). Эти открытия предполагают, что комбинация ts мутаций на фоне мутаций родительского cp вируса привела к ts потомству cp RSV с более высоким уровнем стабильности ts фенотипа после репликации in vivo, чем это было показано для изучаемых ранее ts мутантов.
Для дальнейшего изучения уровня генетической стабильности ts фенотипа потомств cp RSV эффективность бляшкообразования вируса, присутствующего в легких, и в носовых раковинах мышей исследовали для двух потомств подвергнутого мутагенезу cp RSV, которые были среди наиболее чувствительных к температуре (ts) потомств, а именно ts 248 и ts 530. Голые мыши были выбраны, поскольку их иммунная система нарушена вследствие врожденного отсутствия функциональных T-клеток, и вирус может реплицироваться в этом хозяине в течение более длительного периода времени. Более длительный период репликации благоприятствует появлению вирусных мутантов с измененным фенотипом. Вирус, присутствующий на 12-й день (примечание: в нормальных мышах в это время уже нельзя детектировать вирус), был охарактеризован и обнаружили, что он сохраняет неизмененный ts фенотип (табл. 3). Как и ожидали, ts-1 мутант, включенный в тест в качестве положительного контроля, обнаруживал нестабильный ts фенотип in vivo. Таким образом, в противоположность прежней оценке ts мутантных вирусов в грызунах, эти результаты показывают, что после пролонгированной репликации в грызунах был достигнут высокий уровень стабильности ts фенотипа, что является важным и до сих пор не достигаемым очень желательным свойством вирусов данного изобретения.
В шимпанзе
Далее уровень аттенуирования ts потомства cp RSV оценивали в серонегативных шимпанзе, хозяине, наиболее близком к человеку. Опыты на шимпанзе или совинолицых мартышках проводили согласно общему протоколу Richardson et al., J. Med. Virol. 3: 91/100 (1979); Crowe et al., Vaccine (1993) (in press). Один мл суспензии, содержащей приблизительно 104 бляшкообразующих единиц/PFU/мутантного аттенуированиого вируса, вводили интраназально каждому животному. Альтернативным способом является инокуляция RSV как в верхние, так и в нижние дыхательные пути в дозе 104 PFU к каждому сайту. Шимпанзе брали для взятия проб ежедневно в течение 10 дней, затем каждые 3-4 дня до 20-го дня. Пробы из нижних дыхательных путей шимпанзе брали при помощи трахеального лаважа согласно протоколу Shyder et al., J. Infect. Dis. 154: 370-371 (1986) и Crowe et al., Vaccine (1993) (in press). Некоторых животных заражали спустя 4-6 недель вирусом дикого типа. Животных оценивали на признаки респираторного заболевания каждый день при взятии проб из носоглотки. Ринорея, оцениваемая от 0 до 4+, считалась сильным заболеванием верхних дыхательных путей при оценке + 2+ или выше.
Вирус выделяли из проб-мазков из носа и горла и жидкостей трахеального лаважа путем инокуляции в чувствительные к RSV HEp-2 клетки, как описано выше. Количество вируса можно также определять непосредственно по методу бляшек с применением HEp-2 клеток, как описано Schnitzer et al., J. Virol. 17: 431-433 (1976). Пробы крови брали перед введением вируса и через 3-4 недели после инокуляции для определения RSV нейтрализующих антител, как описано в Mills et al., J. Immunol. 107: 123-130 (1970).
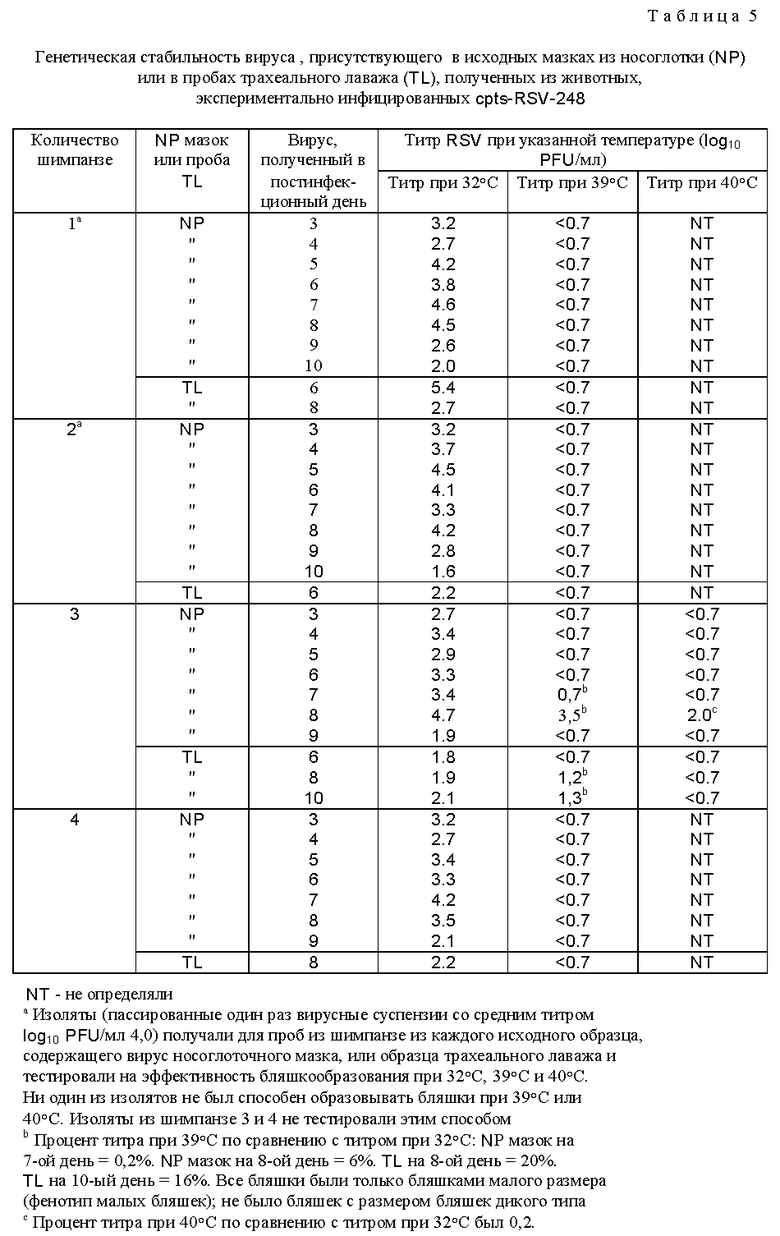
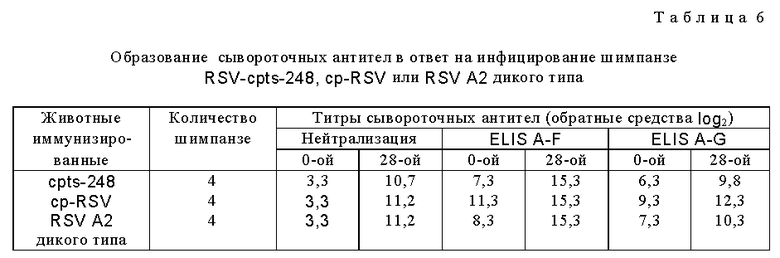
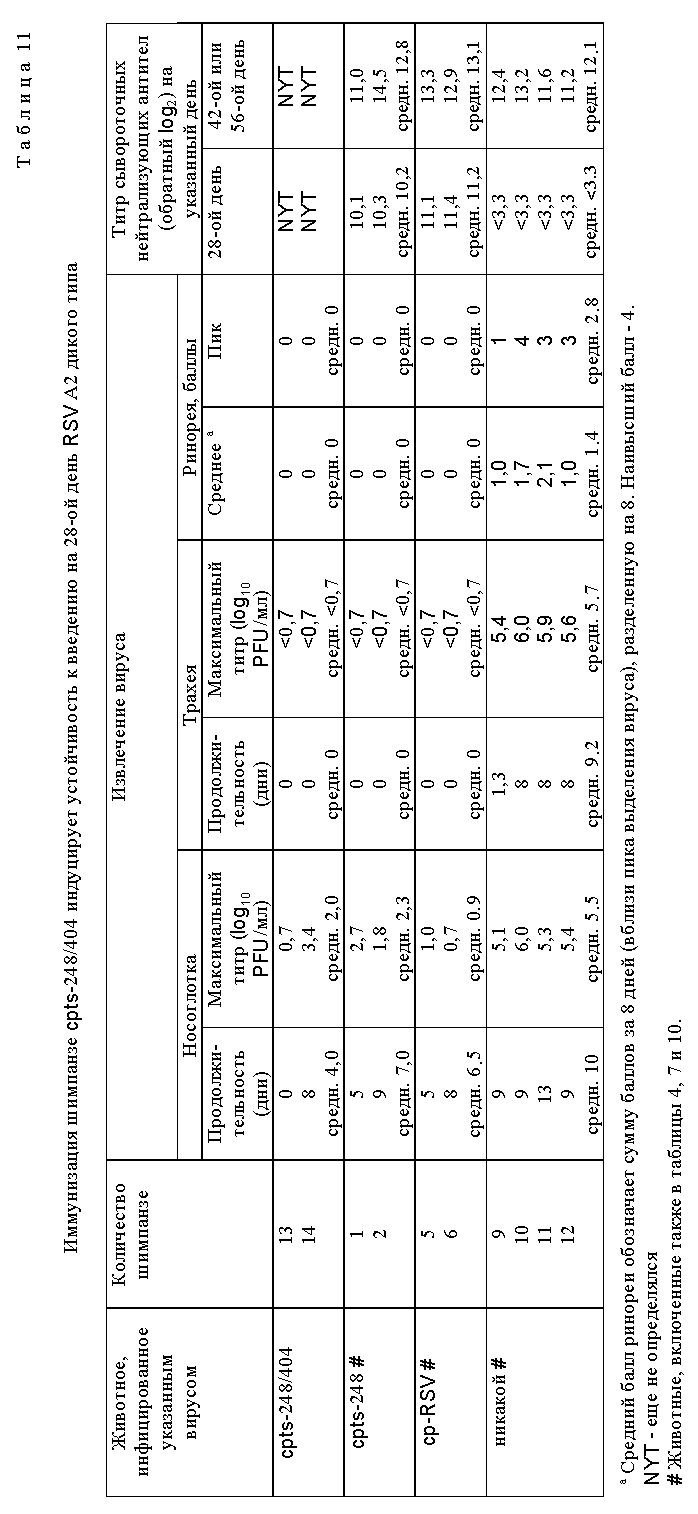
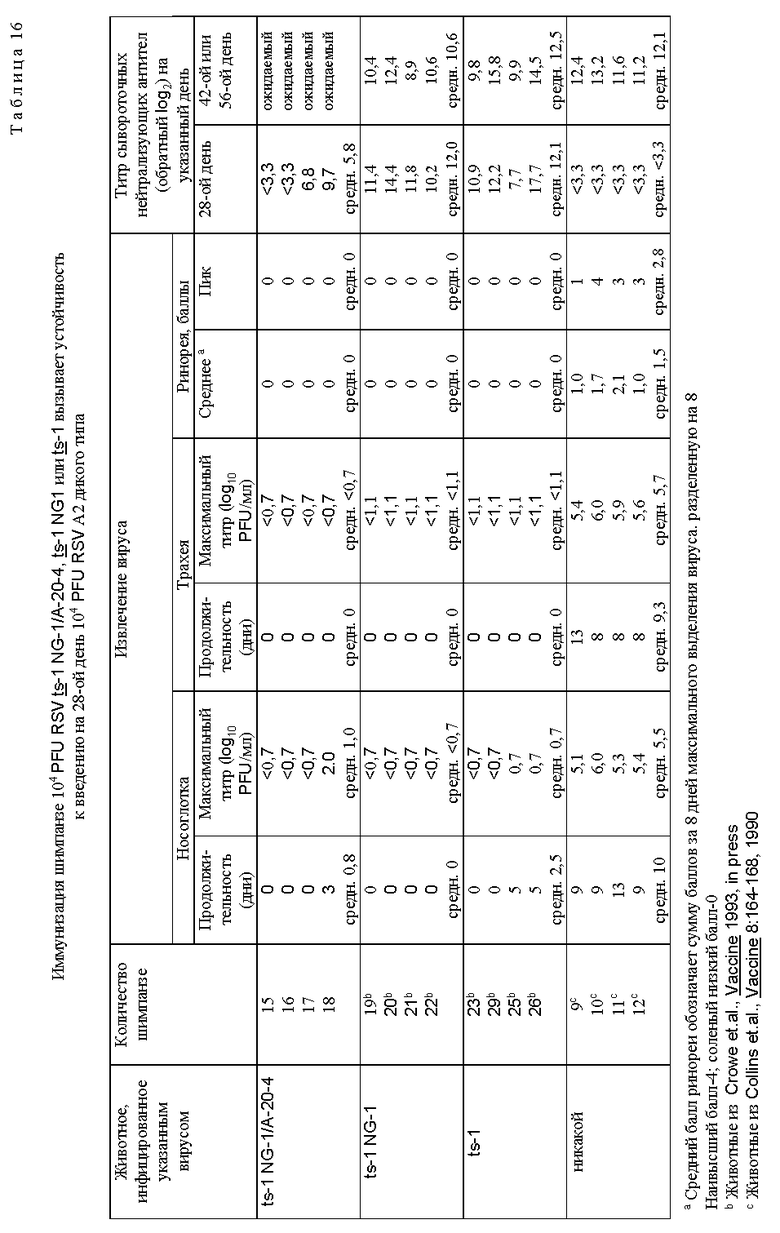
Наиболее ts и аттенуированное потомство cp RSV (cp RSV 248) исследовали и сравнивали с RSV дикого типа и родительским cp RSV (табл. 4). Репликация родительского cp RSV была слегка понижена в носоглотке по сравнению с диким типом, наблюдали снижение ринореи по сравнению с вирусом дикого типа, а также приблизительно 600-кратное уменьшение репликации вируса в нижних дыхательных путях по сравнению с вирусом дикого типа. Ясно, что cp вирус имел значительное ограничение репликации в нижних дыхательных путях шимпанзе, что является очень желательным признаком, не обнаруживаемым ранее при прежних оценках cp RSV в животных и человеке. Более важно то, что cp RSV 248 вирус имел 10-кратное ограничение размножения в носоглотке по сравнению с вирусом дикого типа и это ограничение было связано с заметным уменьшением ринореи. Эти результаты показали, что это производное cp RSV обладает двумя крайне желательными свойствами для живой RSV вакцины, а именно аттенуацией как в верхних, так и в нижних дыхательных путях высокочувствительных серонегативных шимпанзе. Далее оценивали уровень генетической стабильности вируса, присутствующего в дыхательных путях шимпанзе (табл. 5). Вирус, присутствующий в выделениях дыхательных путей, сохранял ts фенотип, и это было видно даже для вируса из шимпанзе N 3 на 8-й день, имеющего 100-кратное снижение титра при 40oC и обнаруживающего фенотип малых бляшек при 40oC, что свидетельствует о том, что его размножение все еще было чувствительным к температуре. Это наиболее генетически стабильный ts мутант, идентифицированный до настоящего времени. Увеличенная стабильность ts фенотипа вирусов cp RSV 248 и cp RSV 530 отражает влияние cp мутаций на генетическую стабильность тех мутаций, которые важны для ts фенотипа in vivo. Таким образом, ts мутации в сочетании с мутациями, присутствующими в родительском вирусе cp 3131, по-видимому, являются более стабильными, чем можно было бы ожидать в их отсутствие. Это важное свойство не было обнаружено и сообщено ранее. Инфицирование шимпанзе вирусом cpts 248 индуцировало высокий титр нейтрализующих антител, а также антител к F и G гликопротеинам (табл. 6). Существенно, что иммунизация при помощи cpts 248 защищала животных от заражения RSV (табл. 7), что указывает на то, что этот мутант действует как эффективный вакцинный вирус в хозяине, близкородственном человеку.
Представленные здесь результаты свидетельствуют о том, что cpts 248 вирус обладает многими свойствами, желательными для живой RSV вакцины, в том числе: 1) аттенуацией для верхних и нижних дыхательных путей; 2) повышенной генетической стабильностью после репликации in vivo даже после пролонгированной репликации в животных с супрессией иммунного ответа; 3) удовлетворительной иммуногенностью; 4) значительной защитной эффективностью против заражения RSV дикого типа. Вирус cpts 530 имеет подобную чувствительность бляшкообразования к температуре по сравнению с cpts 248, подобную степень ограничения размножения в носовых раковинах мышей и высокий уровень генетической стабильности в иммунонедостаточной голой мыши. Поэтому он также представляет собой вакцинный штамм RSV.
Дальнейшее аттенуирование
Поскольку RS вирус вызывает больше симптомов заболевания нижних дыхательных путей у человека, чем у шимпанзе, и поскольку мутанты, которые удовлетворительно ослаблены для шимпанзе, могут быть недостаточно ослабленными для серонегативных младенцев и детей, производные cpts 248 и 530, обладающие нехарактерными свойствами ts мутантов, а именно, ограниченным размножением и аттенуированием в верхних дыхательных путях и высоким уровнем генетической стабильности, были подвергнуты дальнейшему мутагенезу.
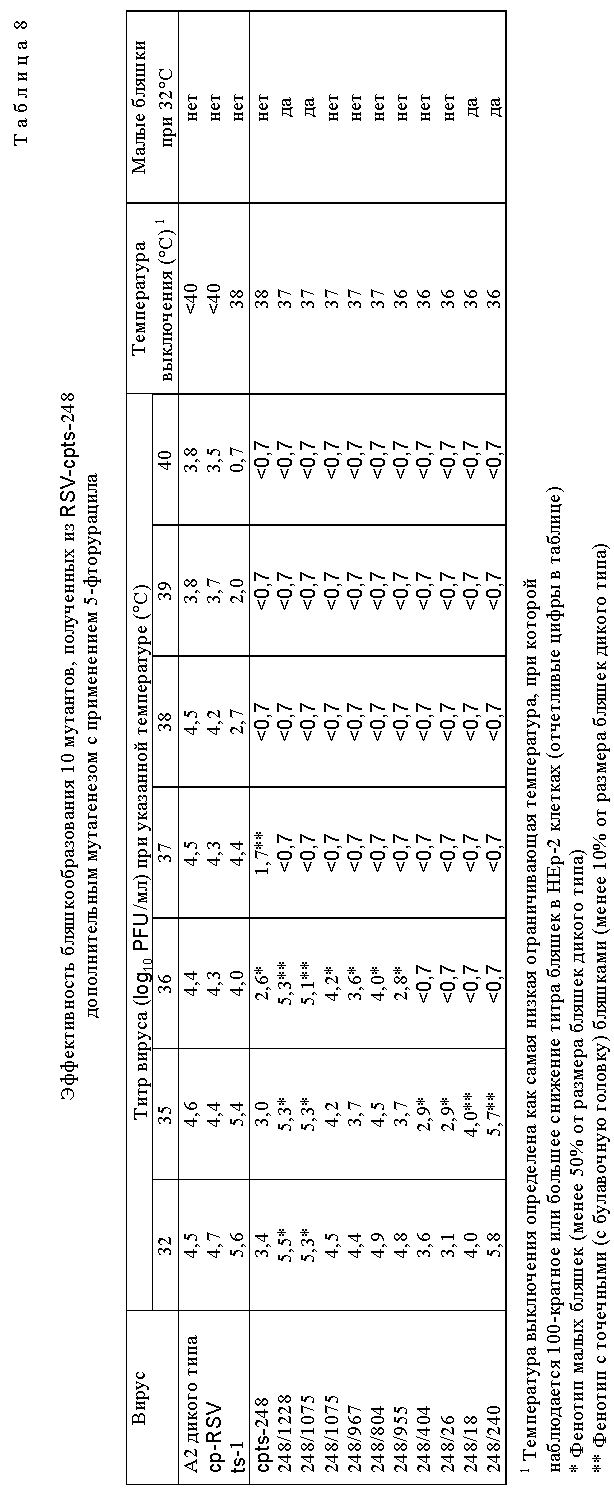
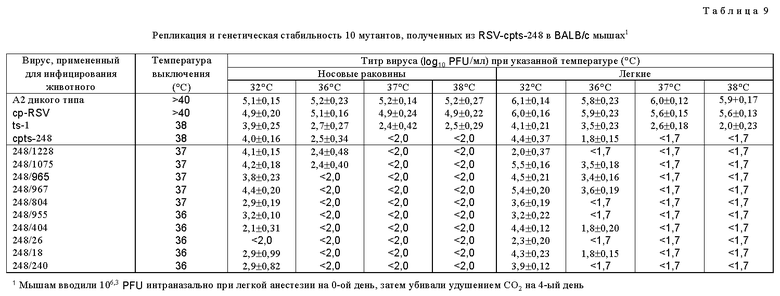
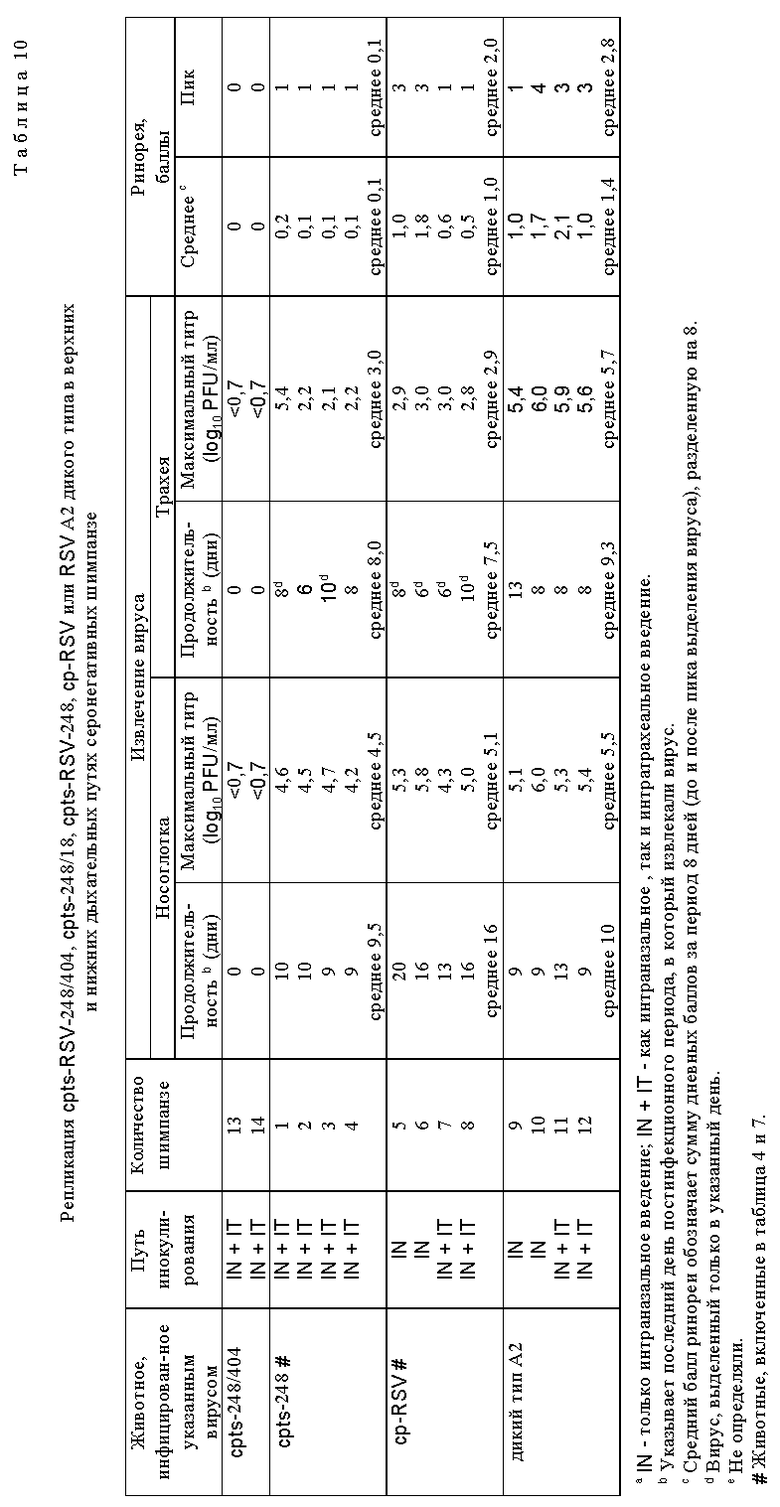
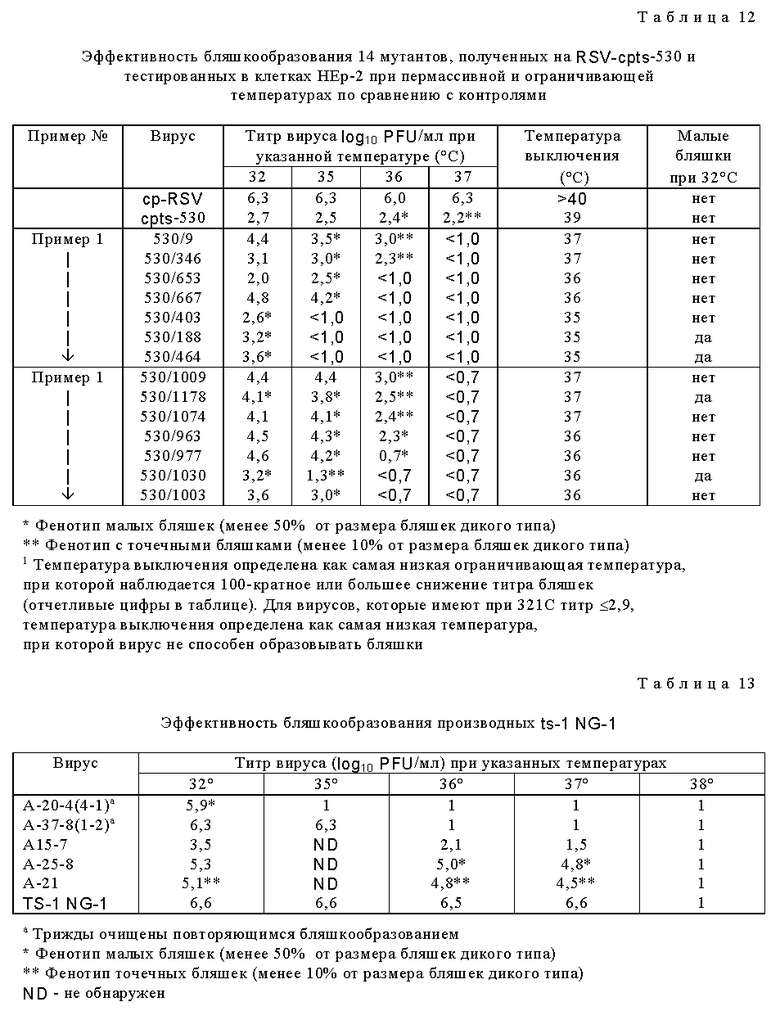
Для дальнейшего исследования были выбраны потомства вирусов, которые обнаружили более высокую степень чувствительности к температуре in vitro, чем cpts 248, или которые имели фенотип малых бляшек. Мутантные производные cpts 248, обладающие одной или несколькими дополнительными ts мутациями, получали при помощи мутагенеза в присутствии 5-фторурацила (табл. 8). Ts мутанты, более чувствительные к температуре (ts), чем cpts 248, были идентифицированы и некоторые из них имели фенотип малых бляшек (sp). Эти производные cpts 248 вводили мышам. Мутанты cpts 248/804, 248/955, 248/404, 248/26, 248/18 и 248/240 были более ограничены в размножении в верхних и нижних дыхательных путях мышей, чем их родительский вирус cpts 248 (табл. 9). Таким образом, были идентифицированы жизнеспособные мутанты cpts 248, которые были более аттенуированы, чем cpts 248, и эти производимые cpts 248 обнаружили широкий диапазон действия на размножение в мышах, причем ts 248/26 имел наибольшее ограничение размножения. Ts фенотип вируса, присутствующий в носовых раковинах и легких мышей, был почти идентичен ts фенотипу введенного вируса, что свидетельствует о генетической стабильности. Высокоаттенуированное производное cpts 248, вирус cpts 248/404, был в 100 раз более ограничен в размножении в носоглотке по сравнению с вирусом дикого типа. Мутант cpts 248/404, обладающий по меньшей мере тремя аттенуирующими мутациями, был также значительно ограничен в размножении в верхних и нижних дыхательных путях двух серонегативных шимпанзе и инфекция не вызывала ринореи (табл. 10). Этот вирус также обнаруживал высокую степень ограничения размножения по сравнению с диким типом, имея сниженную в 60000 раз репликацию в носоглотке и в 100000 раз - в легких. Тем не менее эти два шимпанзе были высокоустойчивы к последующему заражению RS вирусом дикого типа (табл. 11). Кроме того, были получены ts производные вируса cpts 530 (табл. 12). Эти результаты представляют собой дальнейшее усовершенствование в свойствах RS вирусов данного изобретения и также являются очень важным и значительным успехом в разработке вакцинных штаммов RS вируса.
Эти результаты были совершенно неожиданными, если основываться на опыте, приобретенном во время прежних исследований. Например, результаты более раннего исследования указывали на то, что in vivo свойства ts мутантов RSV, полученных в одном цикле мутагенеза с применением 5-фторурацила, не могут быть предсказаны a priori. Более того, хотя один из первых четырех ts мутантов, полученных этим способом, обнаруживал ту же самую температуру выключения бляшкообразования, что и другие мутанты, он был чрезмерно ослаблен при тестировании в восприимчивых шимпанзе и младенцах и деталях младшего возраста (Wright et al. , Infect. Immun. 37(1): 397-400 (1982). Это свидетельствует о том, что получение ts фенотипа, приводящего к температуре выключения бляшкообразования 37o-38oC, не давало надежно мутанта с желаемым уровнем аттенуирования для восприимчивых шимпанзе, младенцев и детей. Действительно, эти результаты исследований с известными до сих пор мутантами были полностью не способны обеспечить какую-либо основу для вывода о том, что введение трех независимых мутаций (или множества мутаций) в RSV путем пассирования на холоду с последующими двумя последовательными циклами химического мутагенеза могут дать жизнеспособные мутанты, которые сохранят инфекционность для шимпанзе (и, путем экстраполяции, для младенцев) и обнаружат желаемый уровень аттенуирования, иммуногенности и защитную эффективность, требуемые для живой вирусной вакцины, которая могла бы применяться для предотвращения RSV заболевания.
Представленные выше вирусы ясно демонстрируют, что определенные ts производные cp RSV данного изобретения являются инфекционными и обнаруживают значительную степень аттенуирования для мышей и шимпанзе. Эти производные ts мутанта аттенуированы и обнаруживают высокую генетическую стабильность после репликации in vivo. Эти мутанты также индуцируют значительную устойчивость к инфекции RSV в шимпанзе. Таким образом эти производные cp RSV представляют собой вирусные штаммы, пригодные для применения в живых вакцинах, предназначенных для предотвращения тяжелого заболевания, вызываемого респираторно-синцитиальным вирусом у человека.
Пример II
Применение адаптации к холоду для аттенуирования cp RSV мутантов
Этот пример описывает введение ограничивающих рост мутаций в неполностью аттенуированные cp RSV штаммы с ограниченным кругом хозяев путем дальнейшего пассирования этих штаммов при все более сниженных температурах для получения производных штаммов, более удовлетворительно аттенуированных для использования в вакцинах для человека.
Эти подходы адаптации к холоду (ca) были применены для введения дальнейшего аттенуирования в cp RSV 3131 вируса, который неполностью ослаблен в серонегативных детях.
При первой стратегии родительский исходный запас популяции пассированного на холоду RSV (cp RSV 3131), полученного из Flow Laboratories, был приготовлен пассированием в MRС-5 клетках при 25oC, как описано в примере 1. Вкратце, пассированный на холоду вирус инокулировали в монослойную культуру MRC-5 или Vero клеток при множественности заражения ≤ 0,01 и инфицированные клетки инкубировали в течение 3-14 дней перед последующим пассированием. Вирус пассировали свыше 20 раз при 20-22oC для получения более ослабленного вируса. Способ быстрого пассирования, как только становилось очевидным первое указание на репликацию вируса (т.е. 3-5 дней), был предпочтительным для отбора мутантов, способных эффективно реплицироваться при низких температурах. Дополнительно штамм RSV подгруппы B, St. Louis (14617) 85 клон IAI, был выделен из первичных клеток почек африканской зеленой мартышки, а также пассирован и клонирован в MRC клетках (IAI-M C14) и пассирован на холоду 51 раз в этих клетках при 3-22oC. Вторая стратегия предусматривала применение биологически клонированного производного неклонированного родительского вируса cp RSV 3131. Вирус биологически клонировали в клетках почки эмбриона быка (BEK) (ткань, применяемая для исходного получения вируса cp RSV 3131, - см. Friedewold et al., J. Amer. Med. Assoc. 204: 690-694 (1968). Клонированный вирус затем пассировали с 10-дневными интервалами в Vero клетках при низкой температуре. Альтернативно, вирус cp RSV 3131 клонировали конечным разведением (TD2P4) в MRC-5 клетках и пассировали с 10-дневными интервалами в Vero клетках.
Третья стратегия предусматривала отбор мутантов, продуцирующих бляшки при низкой температуре. Было идентифицировано производное вируса RSV cp 3131, названное plaque D1, продуцирующее большие бляшки при 25oC. Этот вирус получали из уровня третьего пассирования (P3) cp 3131-1 (BEK) cp. 3131-17 (BEK) линии. Самую большую бляшку, продуцируемую P3 вирусом, размножали при 32oC, затем получали бляшки при 25oC. Опять выбирали самую большую бляшку, размножали и вновь получали бляшки. После пяти таких циклов был получен мутантный вирус D1 с большими бляшками. D1 клонировали биологически при помощи двух дополнительных циклов очистки при помощи повторения бляшкообразования.
Биологически клонированный вирус D1 образует заметно и равномерно большие бляшки при 25oC, чем cp 3131 или вирус A2 дикого типа. Таким образом, D1 адаптирован к холоду согласно критерию большого размера бляшек при 25oC. Предварительные исследования позволили предположить, что D1 не является чувствительным к температуре. При 37oC бляшки D1 неотличимы от бляшек RSV дикого типа или cp 3131, что позволяет предположить, что D1 не ограничен в росте при этой температуре. В соответствии с этим D1 производит сильное цитопатогенное действие в монослоях Vero клеток при 37oC и 40oC (т.е. при самых высоких испытуемых температурах).
Пример III
Введение дальнейших аттенуирующих мутацией в ts-RSV.
Этот пример описывает применение ts мутантов в качестве родительских вирусов для получения более полно аттенуированных штаммов. Для этого способа выбирали два ts мутанта RSV A2, а именно ts-4 и ts-1 NG 1. Для введения дополнительных мутаций в ts мутантный RSV были выбраны два разных способа. Во-первых, неполностью аттенуированный ts мутант RSV подвергали химическому мутагенезу и мутантные потомства, которые были более чувствительны к температуре относительно бляшкообразования, отбирали для дальнейшего анализа. Во-вторых, ts мутанты RSV пассировали при низкой температуре для отбора ts мутантов RSV с ca фенотипом, т.е. с увеличенной способностью размножаться при субоптимальной температуре по сравнению с родительским вирусом дикого типа.
Родительскую исходную популяцию ts-1 NG-1 вируса готовили из ts-1 NG-1 мутанта живого RSV (A-2) из Flow Laboratories Lot M2, росшего в MRC-5 клетках. Этот мутант, полученный из ts-1 мутанта во втором цикле мутагенеза в присутствии 5-фторурацила, имеет две или более независимых ts мутаций, но все еще вызывает ринорею в восприимчивых к нему шимпанзе. Вирус пассировали дважды в Vero клетках при 32oC для получения ts - 1NG-1 суспензии для мутагенеза. Затем вирус выращивали в присутствии 4•10-4 М 5-фторурацила для индуцирования мутаций во время репликации или экспонировали с 5-азацитидином при 36oC после обработки 5-фторурацилом. Затем мутантную популяцию анализировали при помощи теста бляшкообразования на Vero клетках, которые выдерживали под верхним слоем агара и после определенного периода времени инкубации бляшки идентифицировали микроскопически. Затем извлекали 596 бляшек и потомство каждой бляшки отдельно размножали при помощи роста на свежих монослоях Vero клеток. Содержимое каждой из культур ткани, инокулированных потомством отдельной бляшки мутагенизированного вируса ts-1 NG-1, собирали отдельно при достижении максимальных цитопатогенных эффектов на Vero клетках. Потомство вируса, более чувствительное к температуре, чем ts-1 NG-1, находили титрованием пулов бляшек на HEp-2 клетках при 32oC и 36oC. Каждый вирус, обнаруживающий большую чувствительность к температуре, чем ts-1 NG-1 (т.е. 100-кратное снижение титра при ограничивающей температуре (36oC) по сравнению с 32oC), оценивали далее. Были идентифицированы шесть потомств бляшек, более ts, чем ts-1 NG-1 RSV. Эти штаммы клонировали биологически серийной очисткой с применением бляшкообразования на Vero клетках (3 раза) и затем размножали на Vero клетках. Клонированные штаммы титровали при 32oC, 35oC, 36oC, 37oC и 38oC (тестом эффективности бляшкообразования) для подтверждения их ts фенотипов. Данные эффективности бляшкообразования, полученные на HEp-2 клетках, дали дальнейшее подтверждение фенотипов шести мутантов (табл. 13).
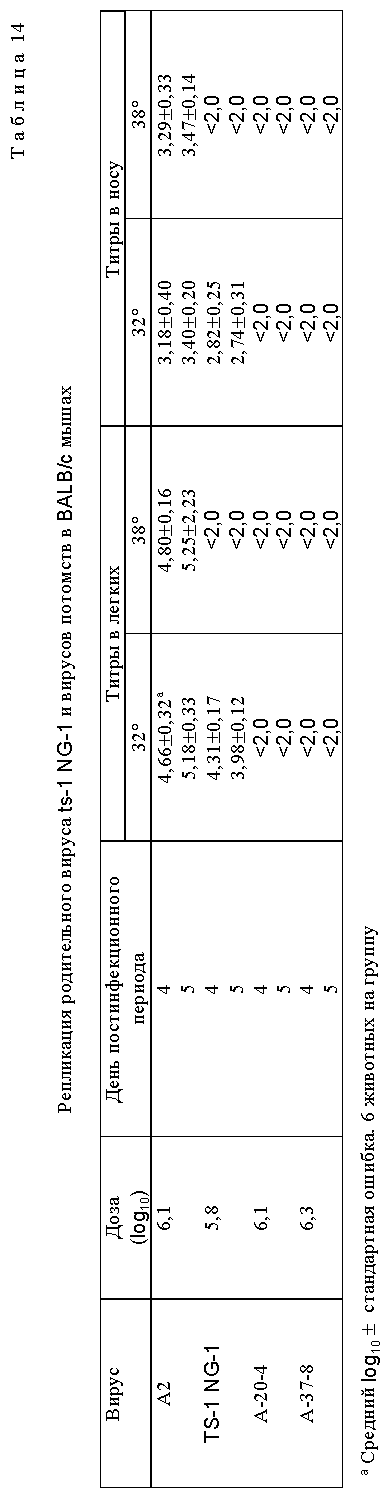
Два наиболее ts (чувствительных к температуре) вируса, A-20-4 и A-37-8, были сильно аттенуированы в мышах по сравнению с их родительским вирусом ts-1 NG-1, что свидетельствует о том, что приобретение увеличенного уровня чувствительности к температуре сопровождается увеличенной аттенуацией (табл. 14). Эти вирусы были инфекционными для мышей, так как они индуцировали образование антител. Вирус ts-1 NG 1/A-20-4 аттенуирован для шимпанзе (табл. 15) и инфицирование шимпанзе ts-1 NG 1/A-20-4 вызывало устойчивость к заражению вирусом дикого типа (табл. 16). Важным является то, что ринорея не наблюдалась.
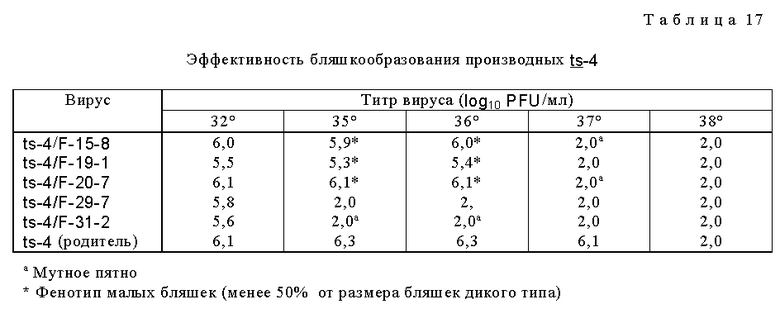
Проводили также мутагенез вируса ts-4 при помощи того же способа, который применяли для мутагенеза вируса ts-1 NG-1. Пять потомств бляшек, более чувствительных к температуре, чем родительский вирус ts-4 RSV, были идентифицированы (табл. 17).
Мутации были введены также в ts-1 NG-1 и ts-4 вирусы пассированием на холоду. Вирус ts-4 размножается до высокого титра при 22oC после 38 пассирований на холоду.
Исследования на человеке
Аттенуированный вирус изобретения вводили человеку в соответствии с хорошо упрочившимися протоколами для RS вакцин, предназначенных для человека, описанных, например, в Wright et al., Infect. Immun. 37: 397-400 (1982), Kim et al., Pediatrics 52: 56-63 (1973) и Wright et. al., J. Pediatr. 88: 931-936 (1976), которые включены в качестве ссылки. Вкратце, взрослых или детей инокулировали интраназально в виде капель с 103 - 105 PFU аттенуированного вируса на мл в объеме 0,5 мл. Образование антител оценивали фиксацией комплемента, нейтрализацией бляшек и(или) твердофазным иммуноферментным анализом. Наблюдали появление признаков и симптомов заболевания верхних дыхательных путей у индивидуумов. Как и при введении шимпанзе, аттенуированный вирус вакцины растет в носоглотке вакцинированных индивидуумов при уровнях, приблизительно более чем в 10 раз более низких по сравнению с уровнями вируса дикого типа и приблизительно в 10 раз или менее при сравнении с уровнями cp RSV, или другого неполностью аттенуированного родительского штамма. Последующие иммунизации проводили периодически по необходимости для поддержания достаточных уровней защитного иммунитета.
Из вышесказанного должно быть понятно, что хотя здесь описаны специфические варианты изобретения для иллюстрации и понимания, могут быть введены различные модификации без отхода от духа и сферы действия данного изобретения. Соответственно этому, изобретение не ограничивается за исключением ограничений, указанных в формуле изобретения.


Изобретение относится к медицине, оно обеспечивает вакцинные композиции аттенуированного респираторно-синцитиального вируса (RSV). Более конкретно, аттенуированный вирус может быть производным RSV, который был неполностью аттенуирован пассированием на холоду или введением мутаций, которые создают вирус, имеющий чувствительный к температуре (ts) или адаптированный к холоду (са) фенотип. Изобретение относится также к способу стимулирования иммунной системы индивидуума для индуцирования защиты против респираторно-синцитиального вируса путем введения аттенуированного RSV. Изобретение обеспечивает также чистые культуры аттенуированного RS вируса, в которых этот вирус более полно аттенуирован путем дальнейшей дериватизации (дальнейшего получения производных) идентифицированных ранее неполностью аттенуированных ts и ср мутантов. Изобретения обеспечивают высокие уровни защитного иммунитета. 2 с. и 27 з.п. ф-лы, 7 табл.
Приоритет по пунктам:
21.04.92 - по пп. 1-11, 13-19 и 24-29;
09.04.93 - по пп. 1-29.
| US 4122167 A, 24.10.78 | |||
| Friedewald et al, J | |||
| Amer | |||
| Med | |||
| Assoe., 1968, 204, p.690-694. |

