РОДСТВЕННАЯ ЗАЯВКА
По настоящей заявке испрашивается приоритет временной заявки на патент США № 62/266199, поданной 11 декабря 2015 года, которая в полном объеме включена в качестве ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Объект изобретения, описываемый в настоящем описании, относится к респираторно-синцитиальному вирусу (РСВ) и его аттенуированным мутантным штаммам, подходящим для применения в качестве вакцин.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Респираторно-синцитиальный вирус (РСВ) человека заражает почти каждого в мире на раннем этапе жизни и несет ответственность за значительную смертность и заболеваемость. Только в Соединенных Штатах Америки РСВ несет ответственность за 75000-125000 госпитализаций каждый год, и консервативные оценки указывают, что РСВ несет ответственность во всем мире за 64 миллиона педиатрических инфекций и 160000 или более педиатрических смертей каждый год. Еще одна заметная особенность РСВ заключается в том, что тяжелая инфекция в младенчестве часто сопровождается затяжной дисфункцией дыхательных путей, в том числе предрасположенностью к реактивности дыхательных путей, что у некоторых индивидуумов длится годами и может продолжаться в подростковом возрасте и за его пределами. Инфекция РСВ усугубляет астму и может участвовать в инициации астмы.
РСВ является членом семейства Paramyxoviridae, и как таковой, является оболочечным вирусом, который реплицируется в цитоплазме и созревает, отпочковываясь от плазматической мембраны клетки-хозяина. Геном РСВ представляет собой единую минус-цепь РНК размером 15,2 килобазы, которая транскрибируется вирусной полимеразой в 10 мРНК с помощью последовательного механизма стоп-старта, который инициируется у одного вирусного промотора на 3'-конце гена. Каждая мРНК кодирует один основной белок, за исключением мРНК M2, которая имеет две перекрывающихся открытых рамки считывания (ORF), кодирующих два отдельных белка M2-1 и M2-2. У РСВ 11 белков: РНК-связывающий нуклеопротеин (N), фосфопротеин (P), большой белок полимеразы (L), гликопротеин присоединения (G), белок слияния (F), малый гидрофобный (SH) поверхностный гликопротеин, белок внутреннего матрикса (M), два неструктурных белка NS1 и NS2, и белки M2-1 и M2-2. Порядок генов РСВ следующий: 3'-NS1-NS2-N-P-M-SH-G-F-M2-L. Каждый ген фланкирован короткими консервативными транскрипционными сигналами, называемыми сигналом «старт гена» (GS), которые присутствуют на конце против хода транскрипции каждого гена и участвуют в инициировании транскрипции соответствующего гена и сигнала «конца гена» (GE), который присутствует на конце по ходу транскрипции каждого гена и участвует в управлении синтезом полиА-хвоста с последующим высвобождением мРНК.
Разработка вакцин на основе РСВ продолжается с 1960-х годов, но осложняется рядом факторов. Например, было показано, что иммунизация младенцев, интактных по РСВ, инактивированным РСВ вызывает усиленное заболевание после последующего естественного инфицирования РСВ, а исследования на экспериментальных животных свидетельствуют о том, что усиление заболевания также связано с вакцинами с очищенными субъединицами РСВ.
Еще одним препятствием для иммунной защиты является то, что РСВ реплицируется и вызывает заболевание в поверхностных клетках просвета дыхательных путей, где иммунная защита имеет сниженную эффективность. Таким образом, иммунный контроль инфекции РСВ является неэффективным и часто неполным, и важно, чтобы вакцина РСВ была максимально иммуногенной. Еще одним препятствием для вакцин РСВ является то, что величина защитного иммунного ответа примерно пропорциональна степени вирусной репликации (и продукции антигенов). Таким образом, ослабление РСВ, необходимое для создания живой вакцины, как правило, сопровождается уменьшением репликации и синтеза антигенов, и сопутствующим снижением иммуногенности, и, таким образом, полезно идентифицировать уровень репликации, который хорошо переносится, но остается достаточно иммуногенным.
Еще одним препятствием является то, что РСВ растет только до умеренных титров в клеточной культуре и часто присутствует в длинных нитях, которые трудно очистить. РСВ может легко потерять инфекционность во время обработки. Еще одним препятствием является трудность в выявлении и разработке аттенуирующих мутаций. Соответствующие мутации должны ослаблять in vivo, но должны минимально ограничивать репликацию in vitro, так как это предпочтительно для эффективного производства вакцин. Еще одним препятствием является генетическая нестабильность, характерная для РНК-вирусов, при которой аттенуирующие мутации могут вернуться к состоянию дикого типа (wt) или к альтернативному состоянию, которое придает деаттенуирующий фенотип. Неустойчивость и деаттенуация особенно проблематичны для точечных мутаций.
В целом, существует необходимость в живых аттенуированных штаммах РСВ, которые эффективно реплицируются in vitro, максимально иммуногенны, удовлетворительно аттенуированы, и устойчивы к деаттенуации.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к мутациям, отдельно или в комбинациях, которые могут включать другие известные мутации, которые подходят для получения рекомбинантных штаммов РСВ человека, проявляющих ряд аттенуирующих фенотипов. Мутации мешают экспрессии открытой рамки считывания (ORF), кодирующей вирусный белок M2-2. Также изобретение относится к рекомбинантным штаммам РСВ, которые содержат такие мутации, либо отдельно, либо в комбинации с одной или несколькими дополнительными мутациями в других локусах, которые могут уменьшать или, альтернативно, повышать величину фенотипа аттенуации. Таким образом, настоящее изобретение относится к новым живым аттенуированным штаммам РСВ с рядом фенотипов аттенуации, пригодным для использования в качестве вакцин РСВ. В некоторых примерах показаны раскрытые варианты осуществления рекомбинантного РСВ, которые являются инфекционными, атенуированными и самореплицирующимися, и вызывают удивительно высокий титр нейтрализующих антител у людей.
В некоторых вариантах осуществления изобретение относится к рекомбинантному РСВ, который ослаблен посредством одной или нескольких модификаций в геноме вируса, таких как модификация, которая препятствует экспрессии открытой рамки считывания (ORF), кодирующей вирусный белок M2-2. В некоторых вариантах осуществления геном рекомбинантного РСВ содержит одну или более модификаций, включающих делецию в M2-2 ORF, соответствующую делеции 241 нуклеотида в положениях 8189-8429 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями, такими как замены T на C, в положениях, соответствующих T8161, T8167 и T8179 SEQ ID NO: 1 («ΔM2-2»). Нуклеотидные мутации разрушают кодоны начала трансляции в этих положениях. В некоторых вариантах осуществления геном рекомбинантного РСВ содержит одну или более модификаций, включающих делецию в ORF M2-2, соответствующую делеции 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями T8198A и C8200G в положениях, соответствующих T8198 и C8200 SEQ ID NO: 1 («ΔM2-2-AclI»). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит одну или более модификаций, включающих делецию в ORF M2-2, соответствующую делеции 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями T8198A и C8199G в положениях, соответствующих T8198 и C8199 SEQ ID NO: 1 («ΔM2-2-HindIII»), где рекомбинантный геном РСВ представляет собой геном D46, аттенуированный посредством одной или нескольких модификаций.
В дополнение к модификации, которая препятствует экспрессии открытой рамки считывания (ORF), кодирующей вирусный белок M2-2, геном рекомбинантного РСВ может содержать дополнительные модификации для увеличения или уменьшения аттенуации вируса или других свойств рекомбинантного вируса. В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ могут дополнительно содержать делецию 112 нуклеотидов, соответствующую положениям 4499-4610 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями C4489T, C4492T, A4495T, A4497G, и G4498A в положениях, соответствующих C4489, C4492, A4495, A4497, и G4498 SEQ ID NO: 1 («6120»). Например, в некоторых вариантах осуществления рекомбинантный РСВ аттенуирован посредством одной или нескольких геномных модификаций, включающих делецию в ORF M2-2, соответствующую делеции 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями T8198A и C8199G в положениях, соответствующих T8198 и C8199 SEQ ID NO: 1 («ΔM2-2-HindIII»), и делецию 112 нуклеотидов соответствующую положениям 4499-4610 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих C4489T, C4492T, A4495T, A4497G, и G4498A SEQ ID NO: 1 («6120»).
В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат нуклеотидные мутации, кодирующие замены аминокислот V267I в белке N, E218A и T523I в белке F, и C319Y и H1690Y в белке L РСВ («cp»). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат нуклеотидные мутации для введения замены Y1321K в белок L РСВ, и где белок L содержит остаток S1313, где кодоны, кодирующие замену Y1321K и остаток S1313, представляют собой кодоны AAA и TCA, соответственно («1030s»). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат нуклеотидные мутации, кодирующие замену аминокислоты K51R в белке NS2 РСВ («NS2»). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат нуклеотидные мутации, кодирующие замену аминокислоты T24A в белке N РСВ («N»). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат нуклеотидные мутации, кодирующие замены аминокислот K51R в белке NS2 и T24A в белке N РСВ («NS2/N»). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат делецию в SH ORF, соответствующую делеции 419 нуклеотидов в положениях 4198-4616 SEQ ID NO: 1 («ΔSH»). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат замену нуклеотидной последовательности, кодирующей белок G РСВ, соответствующей кодон-оптимизированной нуклеотидной последовательностью, кодирующей белок G из клинического изолята A/Maryland/001/11 (такого как SEQ ID NO: 8, G001BB). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат замену нуклеотидной последовательности, кодирующей белок F РСВ, соответствующей кодон-оптимизированной нуклеотидной последовательностью, изложенной в виде SEQ ID NO: 9 (FBB). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат замену нуклеотидной последовательности, кодирующей белок F РСВ, соответствующей нуклеотидной последовательностью, изложенной в виде SEQ ID NO: 10 (F001), которая кодирует белок F из клинического изолята A/Maryland/001/11. В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат замену нуклеотидной последовательности, кодирующей белок F РСВ, соответствующей кодон-оптимизированной нуклеотидной последовательностью, которая кодирует белок F из клинического изолята A/Maryland/001/11 (такой как SEQ ID NO: 11, F001BB). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат нуклеотидные мутации, кодирующие замены аминокислот K66E и Q101P в белке F РСВ («HEK»). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат нуклеотидные мутации, кодирующие замены аминокислот E218A и T523I в белке F РСВ (замены cp в F). В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат обратный порядок генов, кодирующих белки G и F в геноме РСВ.
В некоторых вариантах осуществления одна или более модификаций в геноме рекомбинантного РСВ дополнительно содержат комбинацию или состоят из комбинации мутаций, выбранных из любой из: ΔM2-2, cp/ΔM2-2, cp/ΔM2-2/HEK, ΔM2-2/1030s, NS2/N/ΔM2-2, NS2/ΔM2-2, N/ΔM2-2, ΔSH/ΔM2-2, cp/ΔSH/ΔM2-2, 6120/ΔM2-2, 6120/cp/ΔM2-2, 6120/ΔM2-2/1030s, 6120/NS2/N/ΔM2-2, 6120/G001BB/FBB/ΔM2-2, 6120/FBB/G001BB/ΔM2-2, 6120/G001BB/F/ΔM2-2, 6120/G/FBB/ΔM2-2, 6120/G/FBBHEK/ΔM2-2, 6120/G/FBBcpHEK/ΔM2-2, 6120/FBB/G/ΔM2-2, 6120/G001BB/F001BB/ΔM2-2, 6120/NS2/ΔM2-2, или 6120/N/ΔM2-2; или ΔM2-2-AclI, cp/ΔM2-2-AclI, cp/ΔM2-2-AclI/HEK, ΔM2-2-AclI/1030s, NS2/N/ΔM2-2-AclI, NS2/ΔM2-2-AclI, N/ΔM2-2-AclI, ΔSH/ΔM2-2-AclI, cp/ΔSH/ΔM2-2-AclI, 6120/ΔM2-2-AclI, 6120/cp/ΔM2-2-AclI, 6120/ΔM2-2-AclI/1030s, 6120/NS2/N/ΔM2-2-AclI, 6120/G001BB/FBB/ΔM2-2-AclI, 6120/FBB/G001BB/ΔM2-2-AclI, 6120/G001BB/F/ΔM2-2-AclI, 6120/G/FBB/ΔM2-2-AclI, 6120/G/FBBHEK/ΔM2-2-AclI, 6120/G/FBBcpHEK/ΔM2-2-AclI, 6120/FBB/G/ΔM2-2-AclI, 6120/G001BB/F001BB/ΔM2-2-AclI, 6120/NS2/ΔM2-2-AclI, или 6120/N/ΔM2-2-AclI; или ΔM2-2-HindIII; cp/ΔM2-2-HindIII, cp/ΔM2-2-HindIII/HEK, ΔM2-2-HindIII/1030s, NS2/N/ΔM2-2-HindIII, NS2/ΔM2-2-HindIII, N/ΔM2-2-HindIII, ΔSH/ΔM2-2-HindIII, cp/ΔSH/ΔM2-2-HindIII, 6120/ΔM2-2-HindIII, 6120/cp/ΔM2-2-HindIII, 6120/ΔM2-2-HindIII/1030s, 6120/NS2/N/ΔM2-2-HindIII, 6120/G001BB/FBB/ΔM2-2-HindIII, 6120/FBB/G001BB/ΔM2-2-HindIII, 6120/G001BB/F/ΔM2-2-HindIII, 6120/G/FBB/ΔM2-2-HindIII, 6120/G/FBBHEK/ΔM2-2-HindIII, 6120/G/FBBcpHEK/ΔM2-2-HindIII, 6120/FBB/G/ΔM2-2-HindIII, 6120/G001BB/F001BB/ΔM2-2-HindIII, 6120/NS2/ΔM2-2-HindIII, или 6120/N/ΔM2-2-HindIII.
В некоторых вариантах осуществления геном рекомбинантного РСВ содержит одну или более мутаций, как указано выше, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 1 (последовательность D46). В некоторых вариантах осуществления геном рекомбинантного РСВ представляет собой геном D46, модифицированный одной или несколькими мутациями, как указано выше. В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации 6120 и ΔM2-2, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 5 (последовательность LID/ΔM2-2). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации cp и ΔM2-2, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 1 (последовательность D46). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации cp и ΔM2-2, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 15 (последовательность D46/cp/ΔM2-2). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации 6120, ΔM2-2, и 1030s, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 16 (последовательность LID/ΔM2-2/1030s). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации 6120, cp и ΔM2-2, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 17 (последовательность LID/cp/ΔM2-2). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации NS2, N, ΔM2-2-HindIII, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 4 (последовательность D46/ΔM2-2-HindIII). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации NS2, N, ΔM2-2-HindIII, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 18 (последовательность D46/NS2/N/ΔM2-2-HindIII). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации NS2, N, ΔM2-2-AclI, и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 3 (последовательность D46/ΔM2-2-AclII). В некоторых вариантах осуществления геном рекомбинантного РСВ содержит мутации NS2, N, и ΔM2-2-AclI, следующие нуклеотидные мутации в положениях относительно SEQ ID NO: 1: 404C, 779G, делеция C1099, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T и 13634A; и нуклеотидную последовательность, соответствующую последовательности плюс-цепи по меньшей мере на 90% (такую как по меньшей мере на 95% или по меньшей мере 99%) идентичную SEQ ID NO: 19 (последовательность 276).
В некоторых вариантах осуществления геном рекомбинантного РСВ представляет собой геном D46/cp/ΔM2-2, геном LID/ΔM2-2/1030s, геном LID/cp/ΔM2-2, геном D46/NS2/N/ΔM2-2-HindIII, геном LID/ΔM2-2 или геном 276. В некоторых вариантах осуществления геном рекомбинантного РСВ содержит или состоит из нуклеотидной последовательности, соответствующей последовательности плюс-цепи, изложенной в виде любой из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18 или SEQ ID NO: 19.
В некоторых вариантах осуществления геном рекомбинантного РСВ содержит нуклеотидную последовательность, соответствующую последовательности плюс-цепи, изложенной в виде SEQ ID NO: 3, дополнительно модифицированную путем введения следующих нуклеотидных мутаций относительно SEQ ID NO: 1: 404C, 779G, 1099T, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T, и 13634A. геном рекомбинантного РСВ содержит нуклеотидную последовательность, соответствующую последовательности плюс-цепи, изложенной в виде SEQ ID NO: 3, дополнительно модифицированную путем введения следующих нуклеотидных мутаций относительно SEQ ID NO: 1: 404C, 779G, делеция C1099, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T, и 13634A. В некоторых вариантах осуществления геном рекомбинантного РСВ содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3 и SEQ ID NO: 4, дополнительно модифицированную путем введения одной или нескольких из следующих нуклеотидных замен в положениях относительно SEQ ID NO: 1: 404C, 779G, 1099T, 1139A, 1140G, 1182G, 1210G, 1938A, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T, 13634A, 13901T.
Варианты осуществления рекомбинантного РСВ, описываемого в настоящем описании, могут быть подтипом A РСВ или подтипом B РСВ. Варианты осуществления рекомбинантного РСВ, описываемого в настоящем описании, являются инфекционными, аттенуированными и самореплицирующимися.
Также настоящее изобретение относится к способам и композициям, связанным с экспрессией описанных вирусов. Например, изобретение относится к выделенным полинуклеотидным молекулам, которые включают последовательность нуклеиновой кислоты, кодирующей геном или антигеном описанных вирусов.
Изобретение также относится к фармацевтическим композициям, включающим рекомбинантный РСВ. Композиции могут дополнительно включать адъювант. Изобретение также относится к способам индукции иммунного ответа у индивидуума путем введения иммуногенно эффективного количества описанного рекомбинантного РСВ индивидууму. В некоторых вариантах осуществления индивидуум является человеком, например, человеком в возрасте между 1 и 6 месяцами, или между 1 и 12 месяцами, или между 1 и 18 месяцами, или старше.
Вышеуказанные и другие признаки и преимущества этого изобретения станут более очевидными из следующего подробного описания нескольких вариантов осуществления, которые приводятся со ссылкой на прилагаемые чертежи.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
ФИГ. 1A и 1B. Мутация «ΔM2-2» в геноме РСВ подавляет экспрессию M2-2 ORF в остове РСВ путем введения делеции в 241 нуклеотид и удаления трех потенциальных стартовых кодонов трансляции для белка M2-2. Нумерация последовательности приведена в соответствии с полной последовательностью штамма A2 дикого типа РСВ человека, который представлен посредством номера доступа GeneBank M74568, включенного в настоящий документ в качестве ссылки. (фиг. 1A) Организация генома РСВ и перекрывание открытых рамок считывания M2-1 и M2-2. Показаны три потенциальных стартовых кодона трансляции ATG M2-2 ORF. (фиг. 1B) Подробности мутации ΔM2-2. Верхняя нуклеотидная последовательность представляет собой биологический дикий тип (wt) РСВ (M74568) и показывает нуклеотиды, соответствующие 8150-8204 и 8247-8440 SEQ ID NO: 1. Нумерация нуклеотидов между M74568 и SEQ ID NO: 1 прерывается посредством одного нуклеотида, из-за вставки одного нуклеотида в положении 1099 SEQ ID NO: 1 по сравнению с M74568. Аминокислотная последовательность сразу под ней представляет собой C-конец белка M2-1 (SEQ ID NO: 12). Следующая аминокислотная последовательность представляет собой N-концевые остатки белка M2-2 (SEQ ID NO: 13). Три потенциальных инициирующих кодона ATG для M2-2 ORF заключены в рамку и выделены жирным шрифтом. Вторая нуклеотидная последовательность (для мутации ΔM2-2) показывает мутации, которые подавляют экспрессию M2-2 ORF (показаны нуклеотиды 8150-8188 и 8430-8440 SEQ ID NO: 1, с аминокислотной последовательностью SEQ ID NO: 13, показанной непосредственно снизу). Для мутации ΔM2-2 каждый из трех потенциальных стартовых сайтов трансляции ATG был заменен на ACG, а для M74568 нуклеотиды от 8188 до 8428 были делетированы, с удалением 241 нуклеотидов. TER, кодон терминации трансляции.
ФИГ. 2. Изображение генетических карт двух примеров аттенуированных рекомбинантных РСВ с использованием остова D46 и включением мутации ΔM2-2. Эти карты представляют собой вирусы D46/ΔM2-2 (содержащий мутацию ΔM2-2) и LID/ΔM2-2 (содержащий мутацию ΔM2-2 и мутацию «6120», см. ФИГ. 3). Антигеномные последовательности кДНК этих двух конструкций указаны посредством SEQ ID NO: 2 и SEQ ID NO: 5, соответственно.
ФИГ. 3A и 3B. Мутация «6120» в геноме РСВ содержит делецию в 112 нуклеотидов в 3'-нетранслируемой области гена SH. Нумерация последовательности приведена в соответствии с полной последовательностью штамма A2 дикого типа РСВ человека, который представлен посредством номера доступа GeneBank M74568. Карта генома РСВ (фиг. 3A) и 3'-конец гена SH (фиг. 3B), содержащий мутацию «6120», которая присутствует в LID/ΔM2-2. В этом изобретении, использование «6120» в названии вируса указывает на присутствие мутации 6120. (фиг. 3A) Карта генома РСВ. Стрелка указывает на расположение мутации 6120 в 3'-нетранслируемой области гена SH. (фиг. 3B) Подробности мутации 6120. Верхняя нуклеотидная последовательность представляет собой биологический дикий тип (wt) РСВ (M74568) и показывает нуклеотиды 4481-4498 и 4611-4632 SEQ ID NO: 1, которые соответствуют 3'-концу гена SH. Показаны последние пять кодонов SH ORF с соответствующей представленной аминокислотной последовательностью (ARVNT, SEQ ID NO: 14), с последующим кодоном терминации трансляции (TER). После чего справа следует 3'-нетранслируемая область гена SH (нуклеотиды 4611-4632 SEQ ID NO: 1), с подчеркнутым сигналом окончания гена SH. Три точки представляют 112 нуклеотидов 3'-нетранслируемой области (нуклеотиды 4498-4609 в M74568), которые удалены в мутации 6120. Нижняя нуклеотидная последовательность (показывающая нуклеотиды 4481-4520 SEQ ID NO: 5, с соответствующей представленной аминокислотной последовательностью (ARVNT, SEQ ID NO: 14)) показывает мутацию 6120, которая включает делецию в 112 нуклеотидов, а также пять молчащих точечных мутаций (выделены жирным) в нижележащих трех кодонах и терминирующем кодоне SH ORF. Природные участки рестрикции XhoI и PacI показаны курсивом.
ФИГ. 4. Схематические диаграммы геномов трех примеров производных D46/ΔM2-2 РСВ, каждый из которых содержит одну или более дополнительных аттенуирующих мутаций. Показана мутация ΔM2-2. Другие аттенуирующие мутации включают набор мутаций «cp» (пять замен аминокислот в белках N, F, и L: N (V267I), F (E218A и T523I), и L (C319Y и H1690Y)), и стабилизированную мутацию 1030 («1030s») в белке L, которая включает замену Y1321K, созданную путем введения кодона AAA для аминокислоты 1321L, и стабилизирующую версию серинового кодона в положении 1313L (nt 12435-12437 SEQ ID NO:1; AGC заменен на TCA) (1321K(AAA)+S1313(TCA); Luongo, et al. 2012. J Virol 86:10792-10804). Дополнительные мутации представляют собой изменения HEK в белке F (K66E и Q101P)(Connors, et al. 1995. Virology 208:478-484; Whitehead, et al. 1998. J Virol 72:4467-4471).
ФИГ. 5. Схематические диаграммы геномов четырех примеров производных LID/ΔM2-2 РСВ, каждый из которых содержит одну или более дополнительных аттенуирующих мутаций. Остов «LID» представляет собой геном на основе D46, содержащий мутацию «6120». Указаны мутация ΔM2-2 и мутация 6120. Другие аттенуирующие мутации включают набор мутаций «cp» (пять замен аминокислот в белках N, F, и L: N (V267I), F (E218A и T523I), и L (C319Y и H1690Y)), делецию гена SH (см. ФИГ. 6), и мутацию «1030s» в белке L. Следует отметить, что вирусы, из которых полностью был удален ген SH (РСВ ΔSH/ΔM2-2 и РСВ cp/ΔSH/ΔM2-2) не обозначаются как «LID», поскольку делеция SH удаляет мутацию 6120.
ФИГ. 6. Подробности мутации «ΔSH», которая представляет собой делецию в 419 нуклеотидов, подавляющую экспрессию гена SH. Нумерация последовательности приведена в соответствии с полной последовательностью штамма A2 дикого типа РСВ человека, который представлен посредством номера доступа GeneBank M74568. В верхней последовательности (показывающей нуклеотиды 4191-4229 и 4617-4637 SEQ ID NO: 1), элементы слева направо включают: сигнал конца гена M (подчеркнут), межгенную последовательность (IGS) M/SH, сигнал начала гена SH (подчеркнут), три точки, которые указывают на основную часть гена SH (нуклеотиды 4229-4615 в M74568, которые не показаны), сигнал конца гена SH (подчеркнут), и часть IGS SH/G. Нижняя последовательность (нуклеотиды 4191-4197 и 4617-4637 SEQ ID NO: 1) показывает делецию в 419 нуклеотидов, которая создает мутацию ΔSH. Следует отметить, что, хотя в рабочем порядке она называется делецией гена SH, делеция фактически покрывает последовательность непосредственно перед 5'-сигналом конца гена M и непосредственно до 3'-сигнала конца гена SH.
ФИГ. 7. Репликация примера рекомбинантного РСВ у мышей BALB/c. Показаны результаты репликации РСВ D46 (рекомбинант родительского wt), РСВ LID/ΔM2-2, РСВ ΔSH/ΔM2-2, РСВ LID/ΔM2-2/1030s, и РСВ cp/ΔSH/ΔM2-2 в дыхательных путях мышей BALB/c, с репликацией в носовых ходах и легких по оценке RT-qPCR. Мыши в группах по 10 особей (или 12 для РСВ D46) были заражены интраназальным путем (IN) 5,8 log10 БОЕ указанного вируса в 0,1 мл. Пять мышей на вирус (или шесть для РСВ D46) умерщвляли на сутки 4 и 5 (D4, D5), и удаляли носовые ходы и легкие, гомогенизировали и оценивали путем RT-qPCR, специфичной для гена M РСВ, число копий определяли относительно клонированной кДНК M, которую оценивали параллельно. LoD: предел детекции.
ФИГ. 8A и 8B. Репликация примера рекомбинантных вирусов РСВ в дыхательных путях африканских зеленых мартышек (АЗМ). Показаны данные репликации РСВ LID/ΔM2-2, РСВ ΔSH/ΔM2-2, РСВ LID/ΔM2-2/1030s, и РСВ cp/ΔSH/ΔM2-2 в дыхательных путях АЗМ с выделением вируса в среду по оценке RT-qPCR. АЗМ в группах из четырех особей 4 заражали, сочетая интраназальный и интратрахеальный (IT) пути при помощи (на участок) 6,0 log10 БОЕ указанного вируса в 0,1 мл. Мазки из носоглотки (NP) и смывы с трахеи на указанные сутки оценивали посредством RT-qPCR, специфичной для гена M РСВ, число копий определяли относительно клонированной кДНК M, которую оценивали параллельно. Результаты в виде графиков получены из анализов, описанных в примере 2, таблицах 1 и 2.
ФИГ. 9A и 9B. Пиковые титры иллюстративных рекомбинантных РСВ у серонегативных младенцев и детей. Показаны пиковые титры РСВ MEDI/ΔM2-2 и РСВ rA2cp248/404/1030ΔSH (фиг. 9A) или РСВ LID/ΔM2-2 (фиг. 9B) в смывах из носа у серонегативных младенцев и детей (возрастом 6-24 месяцев) после однократного интраназального посева. Результаты для РСВ MEDI/ΔM2-2 и rA2cp248/404/1030ΔSH получены из Karron, et al. 2015. Science Transl Med 2015 7(312):312ra175. Вирусные титры определяли путем титрования бляшек в образцах назальных смывов: образцы из исследований РСВ MEDI/ΔM2-2 и РСВ rA2cp248/404/1030ΔSH оценивали параллельно, в то время как образцы из исследования РСВ LID/ΔM2-2 оценивали раздельно. Символы указывают пиковые величины для отдельных индивидуумов. Показано среднее пиковых титров. Вакцинированные пациенты получали каждый одну интраназальную дозу 5,0 log10 БОЕ (РСВ MEDI/ΔM2-2, РСВ LID/ΔM2-2) или 5,3 log10 PFU (РСВ rA2cp248/404/1030ΔSH) вируса в вакцине. Исходное описание кандидатной вакцины rA2cp248/404/1030ΔSH было у Karron, et al. 2005. J Infect Dis 191:1093-1104.
ФИГ. 10A и 10B. Мутации «ΔM2-2-AclI» и «ΔM2-2-HindIII» в геноме РСВ. Мутация ΔM2-2-AclI подавляет экспрессию M2-2 ORF путем сайт-направленного мутагенеза для удаления 234 нуклеотидов и введения двух точечных мутаций, создающих участок рестрикции AclI и сайт терминации трансляции (TER). Мутация ΔM2-2-HindIII подавляет экспрессию M2-2 ORF путем сайт-направленного мутагенеза для удаления 234 нуклеотидов и введения двух точечных мутаций, создающих участок рестрикции HindIII и сайт терминации трансляции (TER). Нумерация последовательности приведена в соответствии с полной последовательностью штамма A2 дикого типа РСВ человека, который представлен посредством номера доступа GeneBank M74568. ФИГ. 10A показывает организацию генома РСВ и перекрывание M2-1 и M2-2 ORF. Показаны три потенциальных старт-кодона трансляции ATG M2-2 ORF, но не модифицированные мутациями ΔM2-2-AclI и ΔM2-2-HindIII. ФИГ. 10B показывает подробности мутаций ΔM2-2-AclI и ΔM2-2-HindIII. Верхняя нуклеотидная последовательность (нуклеотиды 8150-8204 и 8427-8440 SEQ ID NO: 1) представляет собой биологический wt РСВ (M74568). Аминокислотные последовательности непосредственно снизу представляют собой C-конец белка M2-1 (SEQ ID NO: 12). Следующая аминокислотная последовательность представляет собой N-концевую часть белка M2-2 (SEQ ID NO: 13). Три потенциальных инициирующих кодона ATG для M2-2 ORF заключены в рамку и выделены жирным шрифтом. Вторая нуклеотидная последовательность (нуклеотиды 8150-8202 SEQ ID NO: 3) показывает последовательность мутации ΔM2-2-AclI, которая является результатом делеции нуклеотидов 8202-8435 и введения точечных мутаций T8197A и C8199G для создания сайта для AclI, а также кодона терминации TAA в кодоне 13 в M2-2 ORF. Аминокислотные последовательности 12 и 13 показаны под последовательностью ΔM2-2-AclI. Третья нуклеотидная последовательность (нуклеотиды 8150-8202 SEQ ID NO: 4) показывает последовательность мутации ΔM2-2-HindIII, которая является результатом делеции нуклеотидов 8202-8435 и введения точечных мутаций T8197A и C8198G для создания сайта для HindIII, а также кодона терминации TAA в кодоне 13 в M2-2 ORF. Как показано, мутированная M2-2 ORF и при ΔM2-2-AclI, и при ΔM2-2-HindIII имеет потенциал к кодированию пептида из 12 аминокислот, представляющего N-конец белка M2-2. Аминокислотные последовательности 12 и 13 показаны под последовательностью ΔM2-2-HindIII.
ФИГ. 11. Схематические диаграммы геномов D46 и LID РСВ, несущих мутацию ΔM2-2-AclI, обозначаемые D46/ΔM2-2-AclI и LID/ΔM2-2-AclI. Атигеномные последовательности кДНК этих двух конструкций обозначены посредством SEQ ID NO: 3 и SEQ ID NO: 6, соответственно.
ФИГ. 12A-12C. Схематические диаграммы геномов примеров вирусов ΔM2-2, в которые были введены мутации K51R (ген NS2) и T24A (ген N). В названиях вирусов, K51R и T24A сокращены как «NS2» и «N», соответственно. ФИГ. 12A показывает совместное введение мутаций K51R и T24A в остов D46/ΔM2-2-AclI и остов LID/ΔM2-2-AclI. ФИГ. 12B и 12C показывают введение мутаций K51R и T24A в остов D46/ΔM2-2 (фиг. 12B) по отдельности или вместе, или в остов LID/ΔM2-2 (фиг. 12C) по отдельности или вместе.
ФИГ. 13. Схематические диаграммы геномов примеров производных от РСВ D46 и LID, несущих мутацию «ΔM2-2-HindIII», а также производных, несущих геном ΔM2-2-HindIII в комбинации с мутациями K51R и T24A. Введение мутации ΔM2-2-HindIII по отдельности в остовы D46 или LID приводит к геномам D46/ΔM2-2-HindIII и LID/ΔM2-2-HindIII (первый и третий геномы сверху). Введение мутации ΔM2-2-HindIII в остовы D46 или LID в комбинации с мутациями K51R и T24A, приводит к геномам D46/NS2/N/ΔM2-2-HindIII и LID/NS2/N/ΔM2-2-HindIII (второй и четвертый геномы сверху). Последовательности конструкций D46/ΔM2-2-HindIII и РСВ LID/ΔM2-2-HindIII обозначены посредством SEQ ID NO: 4 и SEQ ID NO: 7, соответственно.
ФИГ. 14A и 14B. Схематические диаграммы геномов примеров производных от РСВ LID/ΔM2-2 (т.е., содержащих мутацию ΔM2-2, показанную на ФИГ. 1 и мутацию 6120, показанную на ФИГ. 3), содержащих дополнительные модификации генов F и G. Для этих конструкций, идентификатор «6120» применяют вместо «LID» по отношению к мутации 6120. G001 и F001 относятся к генам G и F, соответственно, из клинического изолята подгруппы A штамма (не A2), называемого РСВ A/Maryland/001/11. Все остальные гены относятся к штамму A2. «BB» относится к кодон-оптимизированной последовательности. HEK относится к двум заменам аминокислот в белке F, K66E и Q101P. В этом случае, «FBBcpHEK» относится к кодон-оптимизированной последовательности F, дополнительно включающей замены HEK, и замены «cp», которые находятся внутри белка F, а именно E218A и T523I.
ФИГ. 15. Урожай вирусов в клетках Vero для конструкций, показанных на ФИГ. 14A и 14B. Титр P1 представляет собой урожай первого пассажа (сделан вслепую, т.е., без количественной оценки первичной множественности заражения, MOI) после трансфекции. Титр P2 представляет собой урожай второго пассажа, сделанный с первичной MOI 0,01 БОЕ/клетку; следует заметить, что один вирус представлен посредством P3, урожая третьего пассажа при MOI 0,001. Wt LID представляет собой D46, содержащий мутацию 6120.
ФИГ. 16. Схематическая диаграмма, иллюстрирующая геном РСВ D46/276/ΔM2-2-AclI.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Нуклеотидные и аминокислотные последовательности, перечисленные в прилагаемом списке последовательностей, показаны с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот, как определено в 37 C.F.R. 1.822. Список последовательностей подан в виде текстового файла ASCII в форме файла с названием «Sequence.txt» (~240 т.п.н.), который был создан 8 декабря 2016 года и включен в настоящий документ в качестве ссылки. В прилагаемом списке последовательностей:
SEQ ID NO: 1 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма D46 РСВ.
SEQ ID NO: 2 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма D46/ΔM2-2 РСВ.
SEQ ID NO: 3 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма D46/ΔM2-2-AclI РСВ.
SEQ ID NO: 4 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма D46/ΔM2-2-HindIII РСВ.
SEQ ID NO: 5 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма LID/ΔM2-2 РСВ.
SEQ ID NO: 6 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма LID/ΔM2-2-AclI РСВ.
SEQ ID NO: 7 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма LID/ΔM2-2-HindIII РСВ.
SEQ ID NO: 8 представляет собой пример полинуклеотидной последовательности, кодирующей G001BB.
SEQ ID NO: 9 представляет собой пример полинуклеотидной последовательности, кодирующей FBB.
SEQ ID NO: 10 представляет собой пример полинуклеотидной последовательности, кодирующей F001.
SEQ ID NO: 11 представляет собой пример полинуклеотидной последовательности, кодирующей F001BB.
SEQ ID NO: 12 представляет собой C-концевую аминокислотную последовательность белка M2-1 (SDTNDHAKNNDTT).
SEQ ID NO: 13 представляет собой N-концевую аминокислотную последовательность белка M2-2 (MTMPKIMILPDKYPC).
SEQ ID NO: 14 представляет собой C-концевую аминокислотную последовательность белка SH (ARVNT).
SEQ ID NO: 15 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма D46/cp/ΔM2-2 РСВ.
SEQ ID NO: 16 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма LID/ΔM2-2/1030s РСВ.
SEQ ID NO: 17 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма LID/cp/ΔM2-2 РСВ.
SEQ ID NO: 18 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма D46/NS2/N/ΔM2-2-HindIII РСВ.
SEQ ID NO: 19 представляет собой антигеномную последовательность кДНК для рекомбинантного штамма «276» РСВ.
SEQ ID NO: 20 и 21 представляют собой нуклеотидные последовательности транскрипционных сигналов начала гена (GGGGCAAATA и GGGGCAAACA, соответственно).
ПОДРОБНОЕ ОПИСАНИЕ
Изобретение относится к рекомбинантным штаммам РСВ, подходящим для применения в качестве аттенуированных живых вакцин у людей. Штаммы РСВ получают путем введения мутаций, которые блокируют экспрессию белка M2-2 и наделяют аттенуацией. Дополнительно изобретение относится к рекомбинантным штаммам РСВ, в которых мутации, блокирующие экспрессию белка M2-2, присутствуют в комбинации с одной или несколькими дополнительно сконструированными мутациями в одном или нескольких других локусах, которые увеличивают или уменьшают величину аттенуации, обеспечивая кандидатные вакцины степенями дифференцировки по аттенуации.
Мутации, которые снижают или удаляют экспрессию M2-2 ORF, приводят к повышенной экспрессии вирусных генов, включающих гены, кодирующие защитные антигены (Bermingham и Collins. 1999. Proc Natl Acad Sci USA 96:11259-11264), и имеют потенциал наделять повышенной иммуногенностью. Однако повышенная иммуногенность не была продемонстрирована ранее и не выявлялась у экспрериментальных животных, включая шимпанзе (Teng, et al. 2000. J Virol 74:9317-9321). Клиническая оценка описанных на сегодняшний день штаммов, которые содержат мутации M2-2, описываемые в настоящем описании, демонстрировала, что эти штаммы проявляют повышенную иммуногенность. Исследования у серонегативных младенцев и детей, которые представляют первичную целевую популяцию для вакцины, показали, что штаммы, описываемые в настоящем описании, были аттенуированными и, в основном, хорошо переносились, и вызывали существенный титр РСВ-нейтрализующих сывороточных антител, который был значимо выше, чем титр для предыдущей кандидатной вакцины на основе РСВ (rA2cp248/404/1030ΔSH), которая имела другую основу аттенуации (т.е., серии точечных мутаций в нескольких вирусных генах, включая полимеразу, и представляла более распространенный тип кандидатной живой вакцины) и которую оценивали в сходной популяции индивидуумов (Karron, et al. 2005. J Infect Dis 191:1093-1104).
Таким образом, это изобретение относится к новым улучшенным аттенуированным штаммам РСВ, несущим новые делеционные мутации M2-2, которые обладают повышенной иммуногенностью у людей. Использование большой делеции, которая удаляет большинство вирусных ORF, обеспечивает генетическую стабильность. Дополнительно, это изобретение относится к партиям новых аттенуированных штаммов РСВ, несущих делеционные мутации M2-2 в комбинации с дополнительными мутациями, которые модифицируют фенотип. Это обеспечивает вирусные штаммы с фенотипами с дифференцированной аттенуацией.
Рекомбинантные штаммы РСВ по настоящему изобретению содержат геном или антигеном РСВ дикого типа, содержащий дополнительные модификации или мутации, как подробно описано далее. Вирусный геном или антигеном РСВ дикого типа кодирует следующие 11 белков: РНК-связывающий нуклеопротеин (N), фосфопротеин (P), большой белок полимеразы (L), гликопротеин прикрепления (G), белок слияния (F), малый гидрофобный (SH) поверхностный гликопротеин, белок внутреннего матрикса (M), два не структурных белка NS1 и NS2, и белки M2-1 и M2-2. Порядок генов РСВ является следующим: 3'-NS1-NS2-N-P-M-SH-G-F-M2-L. Полные аминокислотные последовательности эти белков известны в данной области.
Учитывая, что существует ряд штаммов РСВ (например, РСВ A2, РСВ B1, РСВ Long), специалистам в данной области будет ясно, что определенные штаммы РСВ могут иметь вставки или делеции нуклеотидов или аминокислот, которые меняют положение указанного остатка. Например, если белок другого штамма РСВ имел, в сравнении со штаммом A2, две дополнительных аминокислоты на 5'-конце белка, это может вызвать увеличение нумерации аминокислотных остатков, расположенных далее, на +2 относительно штамма A2. Однако, поскольку эти штаммы обладают высокой степенью идентичности последовательностей, специалисты в данной области будут способны определить расположение соответствующих последовательностей путем простого выравнивания нуклеотидной или аминокислотной последовательности референсного штамма A2 с интересующим штаммом. Таким образом, следует понимать, что аминокислотные и нуклеотидные положения, описываемые в настоящем описании, хотя конкретно пронумерованы в отношении этого описания, могут соответствовать другим положениям, когда произошел сдвиг последовательности или из-за вариабельности последовательности между штаммами. При сравнении белка, или сегмента белка, или гена, или генома, или сегмента генома между двумя или более родственными вирусами, «соответствующий» аминокислотный или нуклеотидный остаток представляет собой тот остаток, который считается точно или приблизительно эквивалентным по функции у раличных видов.
Если контекст не указывает на иное, нумерация, использованная в настоящем изобретении основана на последовательности штамма A2 дикого типа РСВ (номер доступа GeneBank M74568) и описанные геномные последовательности вирусов представлены в виде «плюс»-цепи.
В некоторых вариантах осуществления настоящего изобретения, рекомбинантные штаммы РСВ были получены из рекомбинантной версии штамма A2, которая называется D46. Полная последовательность D46 показана в патенте США 6790449 и приведена в настоящем описании в виде SEQ ID NO: 1. (В некоторых случаях и публикациях, родительские вирус и последовательность называются D53, а не D46, различие, сохранившееся в книгах, которое относится к штамму бактерий, используемому для размножения антигеномной кДНК и не имеет другого известного значения или эффекта. Для целей настоящего изобретения, D46 и D53 являются взаимозаменяемыми). SEQ ID NO: 1 (нуклеотидная последовательность D46) отличается от последовательности штамма А2 РСВ M74568 по двадцати пяти нуклеотидным положениям, которые включают однонуклеотидную вставку в положении 1099. Таким образом, нумерация последовательностей относительно SEQ ID NO: 1 отличается от нумерации относительно M74568 на +1 нуклеотид, который расположен в положении после нуклеотида 1099.
В некоторых вариантах осуществления геном или антигеном РСВ модифицирован делецией в M2-2 ORF. Белок M2-2 РСВ кодируется второй, дистальной ORF в мРНК M2, которая слегка перекрывается 5'-проксимальной, M2-1 ORF (фиг. 1A). Существует три потенциальных стартовых кодона трансляции, которые приводят к появлению продуктов в 90, 88, и 84 аминокислоты в длину для штамма A2 (в рамке на ФИГ. 1B). Мутации М2-2, описываемые в настоящем описании, включают делецию большого числа нуклеотидов (как правило, более чем 200 нуклеотидов каждая), вовлекающую большую часть M2-2 ORF, и, таким образом, значительно снижают или полностью прекращают экспрессию белка M2-2. Мутации М2-2, таким образом, устойчивы к компенсации или реверсии, которые могли бы привести к потере аттенуации. Эту генетическую стабильность подтверждали в клиническом исследовании. Большинство предыдущих кандидатных вакцин на основе РСВ содержали аттенуирующие точечные мутации, которые подвержены реверсии или компенсации, приводящим к деаттенуации (например, Karron, et al. 2005. J Infect Dis 191:1093-1104; Malkin, et al. 2013. PLoS One 8:e77104; Karron, Buchholz, Collins. 2013. Curr Top Microbiol Immunol 372:259-284). Де-аттенуация обладает потенциалом для повышенной репликации вируса в вакцине, что может привести к реактогенности, и также потенциалом для распространения производных со сниженной аттенуацией у предрасположенных индивидуумов. Таким образом, мутации М2-2, описываемые в настоящем описании, устраняют серьезную проблему разработки вакцин на основе РСВ.
В некоторых вариантах осуществления мутация М2-2 содержит делецию из 241 нуклеотида, расположенную в положениях 8188-8428 (8189-8429 SEQ ID NO: 1), и мутации T8160C, T8166C и T8178C (T8161C, T8167C и T8179C SEQ ID NO: 1), которые удаляют три потенциальных стартовых кодона. Эта мутация объяснена на ФИГ. 1B и называется в настоящем описании как мутация «ΔM2-2».
В некоторых вариантах осуществления мутация М2-2 содержит делецию из 234 нуклеотидов, расположенную в положениях 8202-8435 (8203-8436 SEQ ID NO: 1), в сочетании с наличием 8197A и 8199G (8198A и 8200G SEQ ID NO: 1), соответствующим присутствию участка распознавания фермента рестрикции AclI. Эта мутация объяснена на ФИГ. 10 и называется в настоящем описании как мутация «ΔM2-2-AclI».
В некоторых вариантах осуществления мутация М2-2 содержит делецию из 234 нуклеотидов, расположенную в положениях 8202-8435 (8203-8436 SEQ ID NO: 1), в сочетании с наличием 8197A и 8198G (8198A и 8199G SEQ ID NO: 1), соответствующим присутствию участка распознавания фермента рестрикции HindIII. Эта мутация объяснена на ФИГ. 10 и называется в настоящем описании как мутация «ΔM2-2-HindIII». В некоторых вариантах осуществления штамм РСВ, используемый для конструирования штамма, может быть D46 (SEQ ID NO: 1). В этом случае, полученный рекомбинантный штамм называется D46/ΔM2-2-HindIII.
Присутствие термина «ΔM2-2» в названии вируса в этом описании указывает на присутствие мутации ΔМ2-2, показанной на ФИГ. 1, в этом вирусе, за исключением случая «MEDI/ΔM2-2», который обозначает другую мутацию, которая описана в примерах далее. Остальные мутации определяются терминами ΔM2-2-AclI или ΔM2-2-HindIII.
Можно вводить дополнительные мутации в комбинации с одной из мутаций М2-2, определенных выше, для конструирования дополнительных вирусных штаммов с желаемыми характеристиками. Например, добавленные мутации могут определять различные величины аттенуации, и, таким образом, давать постепенное увеличение аттенуации. Таким образом, штаммы для кандидатных вакцин могут быть дополнительно аттенуированы путем введения по меньшей мере одной, и предпочтительно двух или более различных аттенуирующих мутаций, например, мутаций, выявленных из панели известных, полученных биологическим путем штаммов РСВ. Ряд таких мутаций обсуждается в настоящем описании в виде примеров. Из этой иллюстративной панели можно создать большое «меню» аттенуирующих мутаций, в котором каждую мутацию можно комбинировать с любой другой мутацией/мутациями в пределах панели для калибровки уровня аттенуации и других желаемых фенотипов. Дополнительные аттенуирующие мутации можно идентифицировать у РНК-вирусов (не РСВ) с «минус»-цепью и вводить в мутантов РСВ по изобретению путем картирования мутации в соответствующий гомологичный сайт в реципиентный геном или антигеном РСВ и мутирования существующей последовательности реципиента в мутантный генотип (путем или идентичной или консервативной мутации). Дополнительные полезные мутации можно определять эмпирическим путем посредством мутационного анализа с использованием систем рекомбинантных минигеномов и инфекционного вируса, как описано в ссылках, включенных в настоящий документ.
В некоторых вариантах осуществления описанные рекомбинантные штаммы РСВ для вакцин можно получать с использованием способов на основе рекомбинантной ДНК, называемых «обратной генетикой» (Collins, et al. 1995. Proc Natl Acad Sci USA 92:11563-11567). Эта система позволяет полностью восстанавливать инфекционный вирус de novo из кДНК на субстрате компетентных клеток при определенных условиях. Обратная генетика обеспечивает способы для введения предопределенных мутаций в геном РСВ через промежуточную кДНК. Конкретные аттенуирующие мутации были охарактеризованы в доклинических исследованиях и скомбинированы для получения желаемых уровней аттенуации. Получение вирусов для вакцины из кДНК минимизирует риск загрязнения случайными агентами и помогает сохранить историю пассажей краткой и хорошо документированной. После восстановления сконструированные вирусные штаммы размножаются также как биологически полученный вирус. В результате пассажа и амплификации, вирусы для вакцины не содержат рекомбинантной ДНК из исходного восстановления.
Рекомбинантные вирусные штаммы, которые содержат различные комбинации мутаций, обсуждаемых в настоящем описании, приведены только с иллюстративными целями и не ограничивают объем настоящего изобретения. Другие аттенуирующие мутации, не описанные в настоящем описании, также можно использовать в комбинации с описанной мутацией М2-2 (такой как ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII мутация).
Например, в некоторых вариантах осуществления рекомбинантные штаммы РСВ по настоящему изобретению дополнительно содержат делецию нетранслируемых последовательностей. В одном из вариантов осуществления такая делеция происходит на 3'-конце гена SH, приводя к мутации, которая в настоящем описании называется «6120». Мутация «6120» показана на ФИГ. 3. Она включает делецию из 112 нуклеотидов 3'-нетранслируемой области гена SH и введение пяти трансляционно-молчащих точечных мутаций в последних трех кодонах и кодоне терминации гена SH (Bukreyev, et al. 2001. J Virol 75:12128-12140). Наличие термина «LID» или «6120» в названии рекомбинантного вируса указывает на то, что рекомбинантный вирус содержит мутацию 6120.
Мутация 6120 стабилизирует антигеномную кДНК в бактериях, и, таким образом, ее можно более легко обрабатывать и готовить. У wt РСВ, ранее было обнаружено, что эта мутация вызывает 5-кратное повышение эффективности репликации in vitro (Bukreyev, et al. 2001. J Virol 75:12128-12140), в то время как считают, что она не повышает эффективность репликации in vivo. Когда РСВ LID/ΔM2-2 оценивали на возможность повышения репликации, ассоциированной с мутацией 6120, наблюдали умеренное, но непостоянное повышение роста эффективности.
Мутация 6120 была ассоциирована с повышенной репликацией у серонегативных младенцев и детей. Таким образом, мутация 6120 обеспечивает другие способы для сдвига уровня аттенуации. Хотя использование этой стратегии демонстрируется в настоящем описании в сочетании с мутацией ΔМ2-2, ее можно применять на других аттенуированных штаммах с этой же целью. Также, делеция последовательности проиллюстрирована мутацией 6120 в 3'-нетранслируемой области гена SH, но в принципе могла бы включать любую сравнимую геномную последовательность, которая не содержит важного цис-действующего сигнала (Collins и Karron. 2013. Fields Virology 6th Edition, pp 1086-1123). Геномные области, которые являются кандидатами для делеции, в качестве неограничивающих примеров включают, нетранслируемые области в других генах, в межгенных областях, и в трейлерной области.
В некоторых вариантах осуществления рекомбинантные штаммы РСВ могут содержать мутацию «cp». Эта мутация относится к набору из пяти замен аминокислот в трех белках (N (V267I), F (E218A и T523I), и L (C319Y и H1690Y)), которые вместе (самостоятельно) обеспечивают 10-кратное снижение репликации у серонегативных шипманзе, и уменьшение заболевания (Whitehead, et al. 1998. J Virol 72:4467-4471). Ранее было показано, что мутация cp ассоциирована с умеренно аттенуированным фенотипом (Whitehead, et al. 1999. J Virol 72:4467-4471).
Кроме того, предшествующий анализ шести биологических вирусов, которые были получены путем химического мутагенеза cpРСВ и выбраны по фенотипу, чувствительному к температуре (ts), выявил в общем 6 независимых мутаций, каждая из которых обеспечивала аттенуирующий ts-фенотип и могла бы быть использована в различных комбинациях. Пять из них были заменами аминокислот в белке L, которые были названы на основе номера вируса, а не положения в последовательности: "955" (N43I), «530» (F521L), «248» (Q831L), «1009» (M1169V), и «1030» (Y1321N) (Juhasz, et al. 1999. Vaccine 17:1416-1424; Collins, et al. 1999. Adv Virus Res 54:423-451; Firestone, et al. 1996. Virology 225:419-422; Whitehead, et al. 1999. J Virol 73:871-877). Шестая мутация (под названием "404") представляла собой однонуклеотидную замену в транскрипционном сигнале начала гена в гене M2 (GGGGCAAATA (SEQ ID NO: 20) на GGGGCAAACA (SEQ ID NO: 21), мРНК-чувствительная) (Whitehead, et al. 1998. Virology 247:232-239). Для повышения генетической стабильности мутаций 248 и 1030 была использована обратная генетика (Luongo, et al. 2009. Vaccine 27:5667-5676; Luongo, et al. 2012. J Virol 86:10792-10804). Кроме того, была получена новая аттенуирующая мутация путем делеции кодона 1313 в белке L, и в сочетании с заменой I1314L обеспечивала повышенную генетическую стабильность (Luongo, et al. 2013. J Virol 87:1985-1996).
В некоторых вариантах осуществления рекомбинантные штаммы могут содержать одну или более замен в белке F, например, мутацию «HEK», которая содержит две замены аминокислот в белке F, а именно K66E и Q101P (описаны у Connors, et al. 1995. Virology 208:478-484; Whitehead, et al. 1998. J Virol 72:4467-4471). Введение аминокислотных замен HEK в последовательность F штамма A2 настоящего изобретения приводит к аминокислотной последовательности белка F, которая идентична последовательности раннего пассажа (пассаж 7 клеток эмбриональной почки человека, HEK-7) исходного клинического изолята штамма A2 (Connors, et al. 1995. Virology 208:478-484; Whitehead, et al. 1998. J Virol 72:4467-4471). Это приводит к белку F, который гораздо менее способен к слиянию, и считают, что он представляет фенотип исходного клинического изолята штамма A2 (Liang et al. J Virol 2015 89:9499-9510). Белок F с HEK также формирует более стабильный тример (Liang et al. J Virol 2015 89:9499-9510). Это может обеспечивать более аутентичную и имууногенную форму белка F РСВ, возможно обогащенную по высокоиммуногенной конформации до слияния (McLellan et al. Science 2013 340(6136):1113-7; Science 2013 342(6158):592-8). Таким образом, можно вводить мутации с эффектами, дополнительными к воздействиям на величину репликации вируса.
В некоторых вариантах осуществления рекомбинантные штаммы могут содержать одну или более замен в белке L, например, стабилизированную мутацию 1030 или «1030s», которая содержит 1321K(AAA)/1313S(TCA) (Luongo, et al. 2012. J Virol 86:10792-10804).
В некоторых вариантах осуществления рекомбинантные штаммы могут содержать делеции одного или нескольких генов РСВ. Было показано, что делеция генов SH, NS1 и NS2 по отдельности и в сочетании дает вирусы, которые сохраняют способность реплицироваться в клеточной культуре, аттенуированы in vivo в следующем порядке повышения величины: SH<NS2<NS1 (Bukreyev, et al. 1997. J Virol 71:8973-8982; Whitehead, et al. 1999. J Virol 73:3438-3442; Teng, et al. 2000. J Virol 74:9317-9321). Таким образом, делецию или другие мутации генов SH, NS2, или NS1, или частей их ORF, можно комбинировать с описанной мутацией М2-2 (такой как мутация ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII). Например, в некоторых вариантах осуществления рекомбинантные штаммы могут содержать одно или несколько изменений в белке SH, включая разрушение или удаление белка SH. В некоторых вариантах осуществления вирусные штаммы содержат делецию в гене SH. Например, в некоторых вариантах осуществления вирусные штаммы содержат делецию из 419 нуклеотидов в положениях 4197-4615 (4198-4616 SEQ ID NO: 1), обозначенную в настоящем описании как мутация «ΔSH». Эта делеция приводит к делеции конца гена М, межгенной области M/SH и делеции SH ORF, как показано на ФИГ. 6. В некоторых вариантах осуществления рекомбинантные штаммы могут содержать одно или несколько изменений в белке NS1 или белке NS2, которые могут включать разрушение или удаление белка. В некоторых вариантах осуществления мутация может быть заменой аминокислоты, такой как K51R в белке NS2. В некоторых вариантах осуществления рекомбинантные штаммы могут содержать одно или несколько изменений в белке N, например, замену аминокислоты, такую как T24A.
Можно вводить различные признаки в штаммы РСВ, несущие описанную мутацию М2-2 (такую как мутация ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII), которые изменяют характеристики вируса иными способами, чем аттенуация. Например, можно проводить оптимизацию кодонов ORF, кодирующих белки. Основные протективные антигены F и G могут приводить к повышенному синтезу антигенов. Ген белка F и/или G можно сдвинуть в 5'-направлении (ближе к промотору) для повышения экспрессии. Однако настоящее изобретение также описывает неожиданные ограничения этой стратегии в случае рекомбинантных вирусных штаммов ΔM2-2. Аминокислотные последовательности белка F и/или G можно модифицировать для представления штаммов, циркулирующих в настоящее время, что может быть применимо в случае различающегося белка G, или для представления клинических изолятов из ранних пассажей. Делеции или замены можно вводить в белок G для получения улучшенной иммуногенности или других желаемых свойств. Например, фракталкиновый мотив CX3C в белке G можно удалять для улучшения иммуногенности (Chirkova et al. J Virol 2013 87:13466-13479).
Например, в некоторых вариантах осуществления нуклеотидная последовательность, кодирующая белок G РСВ, может быть замещена соответствующей нуклеотидной последовательностью из клинического изолята A/Maryland/001/11. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая белок F РСВ, может быть замещена соответствующей нуклеотидной последовательностью из клинического изолята A/Maryland/001/11, например, F001 (SEQ ID NO: 10).
В некоторых вариантах осуществления нативная или природная нуклеотидная последовательность, кодирующая белок РСВ, может быть замещена кодон-оптимизированной последовательностью, разработанной для повышения экспрессии в выбранном хозяине, в частности, в человеке. Например, в некоторых вариантах осуществления нуклеотидная последовательность, кодирующая белок F РСВ, может быть замещена кодон-оптимизированной последовательностью FBB («FBB») (SEQ ID NO: 9). В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая белок F РСВ, может быть замещена кодон-оптимизированной последовательностью из клинического изолята A/Maryland/001/11 («F001BB») (SEQ ID NO: 11). В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая белок G РСВ, может быть замещена кодон-оптимизированной последовательностью G001BB (SEQ ID NO: 8) из клинического изолята A/Maryland/001/11 («G001BB»).
Еще одни дополнительные аспекты изобретения включают изменение положения гена или изменения порядка генов для создания или модификации мутантного РСВ с делецией M2-2. Например, гены NS1, NS2, SH и G можно удалять по отдельности, или гены NS1 и NS2 можно удалять вместе, таким образом, смещая положение каждого гена с 3'-конца относительно вирусного промотора. Например, если NS1 и NS2 удалены вместе, N смещается с положения гена 3 до положения гена 1, P с положения гена 4 до положения гена 2, и так далее. Альтернативно, делеция любого другого гена в пределах генетического порядка будет изменять положение (относительно промотора) только тех генов, которые расположены далее в направлении 3'. Например, SH занимает положение 6 в вирусе дикого типа, и его делеция не сдвинет M с положения 5 (или любой другой ранее стоящий ген), но сдвинет G с положения 7 до 6 относительно промотора. Следует отметить, что делеция гена также происходит (редко) в биологических мутантных вирусах. Например, у подгруппы B РСВ, которая интенсивно размножается в клеточной культуре, спонтанно делетируются гены SH и G (Karron et al. Proc. Natl. Acad. Sci. USA 94:13961 13966, 1997; включен в настоящий документ в качестве ссылки).
Модификации, связанные со смещением порядка генов (т.е., позиционные модификации, смещающие один или несколько генов в более проксимальное или более дистальное по отношению к промотору положение в рекомбинантном вирусном геноме) приводит к вирусам с измененными биологическими свойствами. Например, было показано, что РСВ с отсутствием NS1, NS2, SH, G, NS1 и NS2 вместе, или SH и G вместе, аттенуирован in vitro, in vivo, или и там, и там. В частности, гены G и F могут быть сдвинуты по отдельности или вместе, в более проксимальное по отношению к промотору положение по сравнению с порядком генов в вирусе дикого типа. Эти два белка обычно занимают положения 7 (G) и 8 (F) в генетическом порядке РСВ (NS1-NS2-N-P-M-SH-G-FM2-L). В некоторых вариантах осуществления порядок нуклеотидных последовательностей, кодирующих белки G и F белки может менять направление по отношению к природному порядку.
Известно, что белки F и G РСВ индуцируют РСВ-нейтрализующие антитела, и являются основными протективными антигенами. Считают, что белок F, как правило, является более эффективным нейтрализационным и протективным антигеном, чем белок G. F также является относительно консервативным в штаммах РСВ, в то время как белок G может быть по существу дивергентным. Дивергенция в G является основным фактором распределения штаммов РСВ по двум антигенным подгруппам, A и B (~53% и ~90% идентичности аминокислотных последовательностей между двумя подгруппами для G и F, соответственно). Инструменты и способы настоящего изобретения сфокусированы на штамме A2 подгруппы A РСВ, но могут быть легко применимы к любым штаммам другой подгруппы.
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит рекомбинантный геном РСВ, содержащий мутацию ΔM2-2, ΔM2-2-AclI или ΔM2-2-HindIII в комбинации с одной или несколькими из вышеописанных мутаций. В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит рекомбинантный геном РСВ, содержащий D46 (SEQ ID NO: 1), геном которого был модифицирован мутацией ΔM2-2, ΔM2-2-AclI или ΔM2-2-HindIII, а также одной или несколькими из вышеописанных мутаций.
В некоторых вариантах осуществления рекомбинантный штамм может быть штаммом РСВ на основе D46, включающим мутации «276», и дополнительно включающим одну из описанных мутаций М2-2, такую как мутация ΔM2-2, ΔM2-2-AclI или ΔM2-2-HindIII. Как обсуждается в примере 8, мутации «276» включают следующие нуклеотидные мутации: 404C, 779G, делеция C1099, 1138A, 1139G, 1181G, 1209G, 5611A, 5615A, 5639G, 6215C, 6221C, 6386T, 7214C, 7481T, 7559A, 7701G, 10514T, и 13633A (относительно SEQ ID NO: 1, эти мутации являются следующими: 404C, 779G, делеция C1099, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T и 13634A). Пример антигеномной последовательности кДНК для штамма РСВ на основе ΔM2-2-HindIII, включающего мутации «276» предоставлен в виде SEQ ID NO: 19.
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной (такую как по меньшей мере на 95% идентичной или по меньшей мере на 99% идентичной) антигеномной последовательности кДНК, изложенной в виде SEQ ID NO: 1, которая была модифицирована, чтобы содержать мутации ΔM2-2, ΔM2-2-AclI или ΔM2-2-HindIII, как описано выше, по отдельности или в комбинации с одной или несколькими аттенуирующими мутациями, предлагаемыми в настоящем описании.
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации cp и ΔМ2-2, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 1 (последовательность D46).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутацию ΔМ2-2, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 2 (последовательность D46/ΔM2-2).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутацию ΔM2-2-AclI, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 3 (последовательность D46/ΔM2-2-AclI).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутацию ΔM2-2-HindIII, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 4 (последовательность D46/ΔM2-2-HindIII).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации LID и ΔМ2-2, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 5 (последовательность LID/ΔM2-2).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации LID и ΔM2-2-AclI, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 6 (последовательность LID/ΔM2-2-AclI).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации LID и ΔM2-2-HindIII, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 7 (последовательность LID/ΔM2-2-HindIII).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации cp и ΔМ2-2, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 15 (последовательность D46/cp/ΔM2-2).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации 6120, ΔM2-2, и 1030s, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 16 (последовательность LID/ΔM2-2/1030s).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации 6120, cp, и ΔМ2-2, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 17 (последовательность LID/cp/ΔM2-2).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации NS2, N, ΔM2-2-HindIII, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 4 (последовательность D46/ΔM2-2-HindIII).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации NS2, N, ΔM2-2-HindIII, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 18 (последовательность D46/NS2/N/ΔM2-2-HindIII).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации NS2, N, ΔM2-2-AclI, как описано в настоящем описании, и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной, и/или по меньшей мере на 99% идентичной SEQ ID NO: 3 (последовательность D46/ΔM2-2-AclII).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий мутации NS2, N и ΔM2-2-AclI, как описано в настоящем описании, следующие нуклеотидные мутации в положениях относительно SEQ ID NO: 1: 404C, 779G, делеция C1099, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T, и 13634A; и нуклеотидную последовательность, соответствующую антигеномной последовательности кДНК, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 19 (последовательность 276).
В некоторых вариантах осуществления рекомбинантный штамм РСВ содержит геном, содержащий нуклеотидную последовательность, соответствующую последовательность «плюс»-цепи, изложенную в виде любой из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18 или SEQ ID NO: 19.
В некоторых вариантах осуществления, геном рекомбинантного РСВ содержит одну или более мутаций, описанных в настоящем описании, и любое отличие оставшейся последовательности генома рекомбинантного РСВ по сравнению с геномной последовательностью D46 РСВ (SEQ ID NO: 1) является биологически незначимым (например, отличия остальной последовательности не включают изменений относительно геномной последовательности дикого типа, которые модифицируют известный сигнал, действующий в цис-положении, или изменение кодирования аминокислоты, или измеряемым образом воздействуют на репликацию in vitro или размер бляшек вируса).
В дополнение к вышеописанным мутациям, инфекционные мутанты с делецией M2-2 (такие как мутант с ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII) могут включать гетерологичные, кодирующие или некодирующие нуклеотидные последовательности из любого РСВ или РСВ-подобного вируса, например, человеческого, бычьего, овечьего, мышиного (вирус пневмонии мышей), или птичьего (вирус ринотрахеита индюков) пневмовируса, или из другого оболочечного вируса, например, вируса парагриппа (PIV). Примеры гетерологических последовательностей включают последовательности РСВ из одного штамма РСВ человека в сочетании с последовательностями из другого штамма РСВ человека. Альтернативно, мутанты с делецией M2-2 (такие как мутант с ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII) могут включать последовательности от двух или более, дикого типа или мутантных подгрупп РСВ человека, например, сочетание последовательностей подгруппы A и подгруппы B РСВ человека. Еще в одних дополнительных аспектах, один или несколько кодирующих или некодирующих полинуклеотидов РСВ человека замещены аналогичной последовательностью из гетерологичного РСВ или не-РСВ вируса для получения новых аттенуированных штаммов для вакцин.
В дополнение к рекомбинантным РСВ с конкретными мутациями и комбинациями таких мутаций, описываемых в настоящем описании, описанные вирусы можно модифицировать дополнительно, как это было бы понятно специалистам в данной области. Например, у рекомбинантных РСВ можно удалять или иным образом мутировать один или несколько из его белков или можно добавлять к геному или антигеному гетерологичный ген из другого организма, таким образом, что рекомбинантный РСВ экспрессирует или включает этот белок после инфицирования клетки и репликации. Кроме того, специалистам в данной области будет понятно, что другие ранее определенные мутации, о которых известно, что они имеют влияние на РСВ, можно комбинировать с одной или несколькими из любых мутаций, описываемых в настоящем описании, для получения рекомбинантного РСВ с желаемыми характеристиками аттенуации или стабильности.
В некоторых вариантах осуществления мутации, описываемые в настоящем описании, при использовании по отдельности или в комбинации с другой мутацией, могут обеспечивать различные уровни аттенуации вируса, обеспечивая способность корректировать баланс между аттенуацией и иммуногенностью, и обеспечивая более стабильный генотип, чем у родительского вируса.
Что касается последовательности нумерации положений в нуклеотидной и аминокислотной последовательности для описанных вирусов, применяли конвенцию, согласно которой каждый нуклеотид или аминокислотный остаток в данной вирусной последовательности сохранял номер положения в последовательности, которое он имеет в исходном 15222-нуклеотидном биологическом штамме A2 вируса дикого типа (номер доступа GeneBank M74568), независимо от каких-либо модификаций. Таким образом, хотя ряд геномов содержат делеции и/или вставки, которые вызывают изменения в нуклеотидной длине, и в некоторых случаях аминокислотной длине, нумерация всех остальных остатков (нуклеотид или аминокислота) в геноме и кодируемых белков остается неизменной. Также признано, что даже без целесообразности этой конвенции специалист в данной области может легко идентифицировать соответствующие положения в последовательности между вирусными геномами или белками, которые могут различаться по длине, руководствуясь выравниванием последовательностей, а также положениями открытых рамок считывания, хорошо известных особенностей РНК, таких как сигналы начала и конца гена, и особенностями аминокислотных последовательностей.
Дополнительные типичные вирусы из тех, что описаны в настоящем изобретении, можно оценивать в клеточной культуре на инфекционность, кинетику репликации, урожайность, эффективность экспрессии белка и генетическую стабильность при помощи способов, описываемых в настоящем описании и проиллюстрированных в примерах с использованием иллюстративных рекомбинантных штаммов. Дополнительные типичные вирусы можно оценивать у грызунов и не являющихся человеком приматов на инфекционность, кинетику репликации, урожайность, эффективность экспрессии белка и генетическую стабильность. Хотя эти полупермиссивные системы не могут надежно выявлять каждое отличие в репликации, существенные различия, в частности, можно выявлять (например, между РСВ D46/ΔM2-2 и LID/ΔM2-2, таблицы 4 и 5). Также рекомбинантные штаммы можно оценивать напрямую у серонегативных детей без предварительных шагов оценки у взрослых и сероположительных детей. Это можно производить, например, в группах из десяти реципиентов вакцины и пяти реципиентов плацебо, что является небольшим числом, которое позволяет одновременно оценивать несколько кандидатов. Кандидатов можно оценивать в период сразу после иммунизации на инфекционность, кинетику репликации, выделение вируса, переносимость, иммуногенность и генетическую стабильность вируса в вакцине, а вакцины можно подвергать надзору во время следующего сезона РСВ на безопасность, заболеваемость РСВ и изменения РСВ-специфичных сывороточных антител, как описано в Karron, et al. 2015, Science Transl Med 2015 7(312):312ra175, который в полном объеме включен в настоящий документ. Таким образом, анализ выбранных типичных вирусов может обеспечить относительно быструю сортировку, чтобы сузить число кандидатов для определения наиболее оптимальных.
Ссылка на белок или пептид включает его природную форму, а также любой фрагмент, домен, или гомолог такого белка. В рамках изобретения, термин "гомолог" применяют для ссылки на белок или пептид, который отличается от природного белка или пептида (т.е., "прототипа" или белка "дикого типа") путем незначительных модификаций в природном белке или пептиде, но который сохраняет основную структуру белка и боковой цепи природной формы. Такие изменения в качестве неограничивающих примеров включают изменения в одной или нескольких боковых цепях аминокислот; изменения в одной или нескольких аминокислотах, включая делеции (например, укороченную версию белка или пептида) вставки и/или замены; изменения в стереохимии одного или нескольких атомов; и/или вторичные производные, включая в качестве неограничивающих примеров метилирование, гликозилирование, фосфорилирование, ацетилирование, миристоилирование, пренилирование, пальмитирование, амидирование. Гомолог может иметь повышенные, сниженные, или по существу аналогичные свойства по сравнению с природным белком или пептидом. Гомолог данного белка может содержать, состоять по существу из, или состоять из аминокислотной последовательности, которая по меньшей мере приблизительно на 50% или по меньшей мере приблизительно на 55%, или по меньшей мере приблизительно на 60%, или по меньшей мере приблизительно на 65%, или по меньшей мере приблизительно на 70%, или по меньшей мере приблизительно на 75%, или по меньшей мере приблизительно на 80%, или по меньшей мере приблизительно на 85%, или по меньшей мере приблизительно на 90%, или по меньшей мере приблизительно на 95%, или по меньшей мере приблизительно на 96%, или по меньшей мере приблизительно на 97%, или по меньшей мере приблизительно на 98%, или по меньшей мере приблизительно на 99% идентична (или любой процент идентичности между 45% и 99%, в полностью целочисленном приращении), с аминокислотной последовательностью референсного белка.
В одном из аспектов изобретения, сегмент выбранного гена, такого как ген, кодирующий выбранный белок или область белка (например, цитоплазматический хвост, трансмембранный домен или эктодомен, эпитопный участок или область, участок или область связывания, активный центр или область, содержащая активный центр, и т.д.) из одного РСВ, может быть замещен аналогичным сегментом гена из такого же или различного РСВ или другого источника, для получения новых рекомбинантов с желаемыми фенотипическими изменениями по сравнению со штаммами дикого типа или родительскими штаммами РСВ. Например, рекомбинанты этого типа могут экспрессировать химерный белок с цитоплазматическим хвостом и/или трансмембранным доменом из одного РСВ, слитый с эктодоменом другого РСВ. Другие примеры рекомбинантов этого типа экспрессируют удвоенные области белка, такие как удвоенные иммуногенные области. В рамках изобретения, «аналогичные» гены, сегменты генов, белки или области белков, как правило, происходят из гетерологичных источников (например, из других генов РСВ, или представляют тот же самый (т.е., гомологичный или аллельный) ген или генетический сегмент в других штаммах РСВ). Типичные аналоги, выбираемые в этом контексте, обладают общими структурными особенностями, например, каждый аналог может кодировать сравнимый структурный «домен», такой как цитоплазматический домен, трансмембранный домен, эктодомен, участок или область связывания, эпитопный участок или область, и т.д. Аналогичные домены и кодирующие их сегменты генов охватывают совокупность видов, имеющих диапазон размеров и вариаций аминокислотной (или нуклеотидной) последовательностей, при этом диапазон определяется общей биологической активностью между вариантами доменов и сегментами генов. Например, два выбранных белковых домена, кодируемые сегментами гена-аналога в пределах изобретения, могут иметь по существу одинаковую по качеству активность, такую как обеспечение трансмембранной функции, специфическую активность связывания, иммунологический участок распознавания, и т.д. Более типично, специфическая биологическая активность, разделяемая между аналогами, например, между выбранными сегментами белка или белками, будет по существу аналогичной в количественных терминах, т.е., они не будут варьировать по относительным количественным профилям активности более чем на 30%, предпочтительно, не более, чем на 20%, более предпочтительно, не более, чем на 5-10%.
В альтернативных аспектах изобретения, инфекционный РСВ, полученный из генома или антигенома, экспрессируемого кДНК, может быть любым из штаммов РСВ или РСВ-подобных штаммов, например, человеческим, бычьим, мышиным, и т.д., или любым пневмовирусом или метапневмовирусом, например, вирусом пневмонии мышей или птичьим метапневмовирусом. Чтобы создать защитный иммунный ответ, штамм РСВ может быть штаммом, который является эндогенным по отношению к индивидууму, которого будут иммунизировать, таким как РСВ человека, используемый для иммунизации людей. Геном или антигеном эндогенного РСВ может быть модифицирован, однако, для экспрессии генов или генетических сегментов РСВ из комбинации из различных источников, например, комбинации генов или генетических сегментов из различных видов РСВ, подгрупп, или штаммов, или из РСВ и другого респираторного патогена, такого как вирус парагриппа человека (PIV) (см., например, Hoffman et al. J. Virol. 71:4272-4277 (1997); Durbin et al. Virology 235(2):323-32 (1997); Murphy et al. патентная заявка США с серийным № 60/047575, поданная 23 мая 1997 года, и следующие плазмиды для получения инфекционных клонов PIV: p3/7(131) (ATCC 97990); p3/7(131)2G (ATCC 97889); и p218(131) (ATCC 97991); каждая была размещена на хранение 18 апреля 1997 года на условиях Будапештского договора с Американской коллекцией типовых культур (ATCC) 10801 University Blvd., Manassas, Va. 20110-2209, USA., и получила вышеуказанные номера доступа.
В определенных вариантах осуществления изобретение относится к рекомбинантным РСВ, где отдельные внутренние гены РСВ человека замещены, например, бычьим или другим аналогом РСВ, или аналогом или чужеродным геном из другого респираторного патогена, такого как PIV. Замены, делеции, и т.д. генов или генетических сегментов РСВ в этом контексте могут включать часть или все из одного или нескольких генов NS1, NS2, N, P, M, SH, и L, или открытые рамки считывания M2-1, или неиммуногенные части генов G и F. Также, последовательности РСВ человека, действующие в цис-положении, такие как промотор или сигналы транскрипции, можно замещать, например, их аналогами из бычьего РСВ. Аналогично, предлагаются способы для создания живого аттенуированного бычьего РСВ посредством вставки аттенуирующих генов человека или последовательностей, действующих в цис-положении, в основу генома или антигенома бычьего РСВ.
Таким образом, инфекционный рекомбинантный РСВ, предназначенный для введения людям, может быть РСВ человека, который был модифицирован для содержания генов, например, из бычьего РСВ или PIV, с целью аттенуации. Например, путем вставки сегмента гена или гена из PIV предлагается двухвалентная вакцина как для PIV, так и для РСВ. Альтернативно, гетерологичные виды, подгруппа или штамм РСВ, или отдельный респираторный патоген, такой как PIV, могут быть модифицированы, например, для содержания генов, которые кодируют эпитопы или белки, которые вызывают защиту от инфекции РСВ человека. Например, гены гликопротеина РСВ человека могут быть заменены генами гликопротеина быка, так что полученный бычий РСВ, который теперь несет поверхностные гликопротеины РСВ человека и сохраняет ограниченную способность реплицироваться в человеческом хозяине из-за оставшейся генетической бычьей основы, вызывает защитный иммунный ответ у людей против штаммов РСВ человека.
Способность анализировать и включать другие типы аттенуирующих мутаций в инфекционные РСВ для разработки вакцин распространяется на широкую совокупность целевых изменений в клонах РСВ. Например, любой ген РСВ, который не является существенным для роста, может быть удален или иным образом модифицирован, чтобы получить желаемые воздействия на вирулентность, патогенез, иммуногенность и другие фенотипические признаки. Кроме того, ряд других генетических изменений можно получить в рекомбинантном геноме или антигеноме РСВ для включения в инфекционный рекомбинантный РСВ, по отдельности или вместе с одной или несколькими аттенуирующими точечными мутациями, адаптированными из биологически полученных мутантов РСВ.
В рамках изобретения, «гетерологичные гены» относится к генам, взятым из разных штаммов РСВ или типов или не-РСВ источников. Эти гетерологичные гены могут быть вставлены целиком или частично, порядок генов может быть изменен, удалено перекрытие генов, промотор генома РСВ заменен на его антигеномный аналог, части генов удалены или замещены, и даже целые гены удалены. Различные или дополнительные модификации в последовательности могут быть сделаны для облегчения манипуляций, таких как вставка уникальных участков рестрикции в различные межгенные области (например, уникальный сайт StuI между генами G и F) или в другом месте. Нетранслируемые последовательности гена можно удалить, чтобы увеличить емкость для вставки чужеродных последовательностей.
Делеции, вставки, замены и другие мутации, включающие изменения целых вирусных генов или генетических сегментов в рекомбинантных РСВ по изобретению дают высокостабильных кандидатов для вакцин, которые могут быть подходящими в случае индивидуумов с иммуносупрессией. Многие из этих мутаций будут приводить к аттенуации полученных штаммов для вакцины, в то время как другие будут определять различные типы желаемых фенотипических изменений. Например, известно, что определенные вирусные гены кодируют белки, которые специфически препятствуют иммунитету хозяина (см., например, Kato et al., EMBO. J. 16:578-87 (1997). Ожидается, что удаление таких генов у вирусов для вакцины снизит вирулентность и патогенез и/или улучшит иммуногенность.
Другие мутации в РСВ по настоящему изобретению включают замену 3'-конца генома его аналогом из антигенома, что связано с изменениями в репликации и транскрипции РНК. Кроме того, межгенные области (Collins et al., Proc. Natl. Acad. Sci. USA 83:4594-4598 (1986)) можно укорачивать, удлинять или менять в содержимом последовательности, и можно удалять перекрывание природных генов или менять (Collins et al., Proc. Natl. Acad. Sci. USA 84:5134-5138 (1987)) на другую межгенную область способами, описываемыми в настоящем описании.
В другом варианте осуществления модифицируют последовательность, окружающую сайт начала трансляции (предпочтительно, включающую нуклеотид в положении -3) выбранного гена РСВ, по отдельности или в комбинации с введением 5'-стартового кодона, для модулирования экспрессии гена РСВ, устанавливая положительную или отрицательную регуляцию трансляции.
Альтернативно, или в комбинации с другими модификациями РСВ, описываемыми в настоящем описании, экспрессию гена РСВ можно модулировать, изменяя транскрипционный сигнал GS выбранного гена/генов вируса. В одном иллюстративном варианте осуществления сигнал GS NS2 модифицирован для включения определенной мутации для наложения ограничения по температурной чувствительности (ts) на вирусную репликацию.
Еще одни дополнительные клоны РСВ в пределах изобретения включают модификации транскрипционного сигнала GE. Например, предлагаются клоны РСВ, которые замещают или мутируют сигнал GE из генов NS1 и NS2 на сигнал из гена N, что приводит к сниженным уровням прочитывания мРНК и повышенной экспрессии белков нижерасположенных генов. Полученный рекомбинантный вирус демонстрирует повышенную кинетику роста и увеличенный размер бляшек, обеспечивая лишь один пример изменения ростовых свойств РСВ за счет модификации цис-действующего регуляторного элемента в геноме РСВ.
В другом аспекте экспрессия белка G может быть увеличена модификацией мРНК G. Белок G экспрессируется как в мембрано-связанной, так и в секретируемой форме, причем последняя форма экспрессируется путем инициации трансляции в точке старта внутри открытой рамки считывания трансляции гена G. Секретируемая форма может составлять до половины экспрессированного белка G. Удаление внутреннего стартового сайта (например, путем изменения последовательности, делеции и т. д.), отдельно или вместе с изменением контекста последовательности 5'-стартового сайта приводит к желаемым изменениям в экспрессии белка G. Удаление секретируемой формы белка G также улучшит качество иммунного ответа хозяина на иллюстративный рекомбинантный РСВ, поскольку считается, что растворимая форма белка G выступает в качестве «приманки» для улавливания нейтрализующих антител. Кроме того, растворимый белок G участвует в усиленной иммунопатологии из-за его предпочтительной стимуляции Th2-смещенного ответа.
В родственных аспектах уровни экспрессии гена РСВ могут быть изменены на уровне транскрипции. В одном из аспектов положение выбранного гена на генетической карте РСВ может быть изменено на положение, более проксимальное или более дистальное по отношению к промотору, при этом ген будет экспрессироваться более или менее эффективно, соответственно. В соответствии с этим аспектом может быть достигнута модуляция экспрессии для конкретных генов, приводящая к уменьшению или увеличению экспрессии гена от двух раз, более типично в четыре раза, до десяти раз или более по сравнению с уровнями дикого типа. В одном примере положение гена NS2 (второй по порядку на генетической карте РСВ) заменяют на положение для гена SH (шестой по порядку), что дает прогнозируемое снижение экспрессии NS2. Повышенная экспрессия выбранных генов РСВ из-за позиционных изменений может быть достигнута до 10-кратного, 30-кратного, 50-кратного, 100-кратного повышения или более, часто сопровождаемого соразмерным снижением уровней экспрессии для взаимно, позиционно замещенных генов.
В некоторых иллюстративных вариантах осуществления гены F и G могут быть перенесены по отдельности или вместе на участок, более проксимальный или более дистальный по отношению к промотору в пределах (рекомбинантной) генетической карты РСВ для достижения более высоких или более низких уровней экспрессии гена, соответственно. Эти и другие транспозиционные изменения приводят к появлению новых клонов РСВ, имеющих аттенуированные фенотипы, например из-за уменьшения экспрессии выбранных вирусных белков, участвующих в репликации РНК. В других вариантах осуществления РСВ, полезные в составе вакцины, могут быть соответствующим образом модифицированы для размещения антигенной изменчивости в циркулирующем вирусе. Как правило, модификация будет в белках G и/или F. Весь ген G или F, или сегменты, кодирующие его специфические иммуногенные области, включают в геном или антигеном кДНК РСВ путем замены соответствующей области в инфекционном клоне или путем добавления одной или нескольких копий гена таким образом, что представлено несколько антигенных форм.
Потомство вируса, полученное из модифицированной кДНК РСВ, затем используют в протоколах вакцинации против возникающих штаммов. Кроме того, включение гена белка G подгруппы В РСВ в качестве добавочного гена расширит ответ, чтобы охватить более широкий спектр относительно разнообразных штаммов подгрупп A и B, присутствующих в популяции людей.
Инфекционный клон РСВ по изобретению можно также сконструировать в соответствии со способами и композициями, описываемыми в настоящем описании, для повышения иммуногенности индуцирования уровня защиты, большего чем, тот, который обеспечивается при заражении РСВ дикого типа или неполностью аттенуированным родительским вирусом или клоном. Например, иммуногенный эпитоп из гетерологичного штамма или типа РСВ или из не-РСВ источника, такого как PIV, можно добавлять путем соответствующих нуклеотидных замен в полинуклеотидную последовательность, кодирующую геном или антигеном РСВ. Рекомбинантный РСВ можно также сконструировать для выявления и удаления (например, путем вставки аминокислоты, замены или делеции) эпитопов, ассоциированных с нежелательными иммунопатологическими реакциями. В других вариантах осуществления можно вставлять дополнительный ген внутрь или перед геномом или антигеномом РСВ, который находится под контролем независимого набора транскрипционных сигналов. Гены, представляющие интерес, могут в качестве неограничивающих примеров включать гены, кодирующие цитокины (например, от IL-2 до IL-15, особенно IL-2, IL-6 и IL-12, и т.д.), гамма-интерферон, и белки, богатые эпитопами для клеток Т-хелперов. Дополнительный белок можно экспрессировать или в виде отдельного белка или в качестве химеры, сконструированной из второй копии одного из белков РСВ, такого как SH. Это обеспечивает способность модифицировать и улучшать иммунный ответ против РСВ в количественном и качественном отношении.
В дополнение к вышеописанным модификациям рекомбинантных РСВ, можно производить различные или дополнительные модификации в клонах РСВ для облегчения манипуляций, такие как вставка уникальных участков рестрикции в различных межгенных областях (например, уникальный сайт StuI между генами G и F) или в другом месте. Нетранслируемые последовательности гена можно удалить, чтобы увеличить емкость для вставки чужеродных последовательностей.
Введение вышеуказанных определенных мутаций в инфекционный клон РСВ производят при помощи ряда хорошо известных способов. Под «инфекционным клоном» подразумевают кДНК или ее продукт, синтетический или иной, который можно транскрибировать в геномную или антигеномную РНК, способную производить инфекционный вирус. Термин «инфекционный» относится к вирусу или вирусной структуре, которая способна реплицироваться в культивируемой клетке или животном или человеке-хозяине для производства вирусного потомства или вирусных структур, способных к такой же активности. Таким образом, определенные мутации можно вводить общепринятыми способами (например, сайт-специфическим мутагенезом) в кДНК копию генома или антигенома. Использование субфрагментов кДНК антигенома или генома для сборки полного генома или антигенома кДНК хорошо известно специалистам в данной области и имеет то преимущество, что с каждой областью можно работать отдельно (с более маленькими кДНК легче работать, чем с более крупными), а затем легко собрать в полную кДНК. Таким образом, полную антигеномную или геномную кДНК, или любой ее фрагмент, можно использовать в качестве матрицы для сайт-специфического мутагенеза. Мутированный субфрагмент можно затем собрать в полную антигеномную или геномную кДНК. Мутации могут варьироваться от однонуклеотидных замен до замен крупных кусков кДНК, содержащих один или несколько генов или областей генов.
Рекомбинантный РСВ можно получать путем внутриклеточной коэкспрессии кДНК, которая кодирует геномную РНК РСВ, вместе с теми вирусными белками, которые необходимы для получения транскрибирующегося, реплицирующегося нуклеокапсида. Плазмиды, кодирующие другие белки РСВ, можно также включать вместе с этими незаменимыми белками. Альтернативно, РНК можно синтезировать в реакции транскрипции in vitro и трансфицировать культивируемые клетки.
Таким образом, в настоящем описании также описаны выделенные полинуклеотиды, которые кодируют описанные мутированные вирусы, составляют описанные геномы или антигеномы, экспрессируют описанные геномы или антигеномы, или кодируют различные белки, подходящие для получения рекомбинантного РСВ in vitro. Полинуклеотиды, содержащие последовательности любой из SEQ ID NO, описываемых в настоящем описании, включены в настоящее изобретение. Дополнительно включены полинуклеотиды, содержащие последовательности, которые состоят или по существу состоят из любой из вышеуказанных последовательностей, которые обладают по меньшей мере приблизительно 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100 процентами идентичности (или любым процентом идентичности между указанными) с любой из вышеуказанных SEQ ID NO, а также полинуклеотиды, которые гибридизуются с вышеуказанными молекулами или комплементарны им.
Эти полинуклеотиды могут быть включены в состав векторов или экспрессироваться векторами для того, чтобы производить рекомбинантный РСВ. Таким образом, клетки, трансфицированные выделенными полинуклеотидами или векторами, также находятся в объеме изобретения и приведены в качестве примеров в настоящем описании.
В родственных аспектах изобретение относится к композициям (например, выделенным полинуклеотидам и векторам, содержащим ДНК, кодирующую РСВ) и способам получения выделенных инфекционных рекомбинантных РСВ, несущих аттенуирующую мутацию с делецией M2-2 (таких как мутант ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII). В эти аспекты изобретения включены новые, выделенные полинуклеотидные молекулы и векторы, содержащие такие молекулы, которые включают геном или антигеном РСВ, модифицированный, как описано в настоящем описании. Также изобретение относится к такому же или другому экспрессирующему вектору, содержащему одну или более выделенных полинуклеотидных молекул, кодирующих белки РСВ. Эти белки также можно экспрессировать напрямую с геномной или антигеномной кДНК. Вектор(-ы) предпочтительно экспрессируются или коэкспрессируются в клеточном или бесклеточном лизате, таким образом, производя инфекционную частицу делеционного мутанта РСВ M2 ORF2 или нокаутного мутанта РСВ или субвирусную частицу.
В одном из аспектов изобретение относится к способу получения одного или более очищенных белков РСВ, который включает заражение клетки-хозяина, пермиссивной по отношению к инфекции РСВ, рекомбинантным штаммом РСВ в условиях, который позволяют РСВ размножаться в инфицированной клетке. После периода репликации в культуре, клетки лизируют и из них выделяют рекомбинантный РСВ. Один или несколько желаемых белков РСВ очищают после выделения вируса, получая один или несколько белков РСВ для вакцины, диагностики и других применений.
Вышеописанные способы и композиции для получения мутантов по делеции M2-2 (таких как мутант ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII) производят инфекционные вирусные или субвирусные частицы, или их производные. Инфекционный вирус сопоставим с аутентичной вирусной частицей РСВ, и сам по себе является инфекционным. Он может напрямую заражать свежие клетки. Инфекционная субвирусная частица, как правило, является субкомпонентом вирусной частицы, который может инициировать инфекцию при соответствующих условиях. Например, нуклеокапсид, содержащий геномную или антигеномную РНК и белки N, P, L и M2-1, является примером субвирусной частицы, которая может инициировать инфекцию при введении в цитоплазму клеток. Предлагаемые в изобретении субвирусные частицы включают вирусные частицы, в которых отсутствует один или несколько белков, белковый сегмент (-ы), или другой вирусный компонент(-ы), не существенные для инфекционности.
В других вариантах осуществления изобретение относится к клеточному или бесклеточному лизату, содержащему экспрессирующий вектор, который содержит выделенную полинуклеотидную молекулу, кодирующую геном или антигеном мутанта РСВ по делеции M2-2 (такого как мутант ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII), как описано выше, и экспрессирующий вектор (такой же или другой вектор), который содержит одну или более выделенных полинуклеотидных молекул, кодирующих N, P, L и белковые факторы элонгации для РНК-полимеразы РСВ. Один или несколько из этих белков также можно экспрессировать с геномной или антигеномной кДНК. После экспрессии генома или антигенома белки N, P, L, и факторов элонгации РНК-полимеразы комбинируются для получения инфекционных вирусных или субвирусных частиц РСВ.
Рекомбинантные РСВ по изобретению подходят для различных композиций для получения желаемого иммунного ответа против РСВ у хозяина, чувствительного к инфекции РСВ. Аттенуированные мутантные штаммы РСВ с делецией M2-2 (такие как мутант ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII), описываемые в настоящем описании, способны вызывать защитный иммунный ответ у инфицированного человека-хозяина, но достаточно аттенуированы, чтобы не вызывать неприемлемых симптомов тяжелого респираторного заболевания у иммунизированного хозяина. Аттенуированная вирусная или субвирусная частица могут присутствовать в супернатанте клеточной культуры, могут быть выделены из культуры и частично или полностью очищены. Вирус можно также лиофилизировать, и можно комбинировать с рядом других компонентов для хранения или доставки хозяину, если необходимо.
В другом аспекте мутанты с делецией M2-2 (такие как мутант ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII) можно использовать в качестве «векторов» для протективных антигенов других патогенов, в частности, патогенов дыхательных путей, таких как вирус парагриппа (PIV). Например, можно сконструировать рекомбинантный РСВ с делецией M2-2 (такой как мутант ΔM2-2, ΔM2-2-AclI, или ΔM2-2-HindIII), коорые включает последовательности, которые кодируют протективные антигены из PIV, для получения инфекционного аттенуированного вируса для вакцины.
В родственных аспектах, изобретение относится к способу стимуляции иммунной системы индивидуума для индукции иммунного ответа против РСВ у млекопитающего индивидуума. Способ включает введение иммуногенного состава с иммунологически достаточным количеством аттенуированного мутанта РСВ с делецией M2-2, как описано в настоящем описании, в физиологически приемлемом носителе и/или адъюванте.
Изобретение дополнительно относится к новым вакцинам, содержащим физиологически приемлемый носитель и/или адъювант и выделенную аттенуированую частицу или субвирусную частицу мутантного РСВ с делецией M2-2. В предпочтительных вариантах осуществления вакцина состоит из мутанта РСВ с делецией M2-2, имеющего, по меньшей мере одну, и предпочтительно две или более аттенуирующих мутации или другие нуклеотидные модификации, как описано выше, для достижения подходящего баланса между аттенуацией и иммуногенностью.
Для выбора вирусов-кандидатов для вакцин из хозяина рекомбинантных штаммов РСВ, предлагаемых в настоящем описании, определяют критерии жизнеспособности, эффективной репликации in vitro, аттенуации in vivo, иммуногенности и фенотипической стабильности хорошо известными способами. Вирусы, которые будут наиболее желательными для вакцин по изобретению, должны сохранять жизнеспособность, должны успешно реплицироваться in vitro в пермиссивных условиях, чтобы сделать возможным производство вакцины, должны иметь стабильный фенотип аттенуации, должны хорошо переноситься, должны демонстрировать репликацию у иммунизированного хозяина (хотя бы и на более низких уровнях), и должны эффективно вызывать выработку иммунного ответа в вакцине, достаточного для обеспечения защиты от серьезного заболевания, вызванного последующей инфекцией вируса дикого типа.
Для размножения вируса РСВ для применения в вакцине и других целей, можно использовать ряд клеточных линий, в которых может расти РСВ. РСВ растет в ряде клеток человека и животного. Предпочтительные клеточные линии для размножения аттенуированного RS вируса для использования в вакцине включают клетки DBSFRhL-2, MRC-5 и Vero. Наибольшие урожаи вируса, как правило, получают с эпителиальными клеточными линиями, такими как клетки Vero. Клетки, как правило, заражают вирусом с мультиплетностью заражения в диапазоне приблизительно от 0,001 до 1,0, или больше, и культивируют в условиях, пермиссивных для репликации вируса, например, приблизительно при 30-37°C и в течение приблизительно 3-10 суток, или при условии, что вирусу необходимо достичь адекватного титра. Температуро-чувствительные вирусы часто выращивают при 32°C в качестве «пермиссивной температуры». Вирус удаляют из клеточной культуры и отделяют от клеточных компонентов, как правило, путем хорошо известных способов очищения, например, центрифугирования, и его можно дополнительно очищать при необходимости с использованием способов, хорошо известных специалистам в данной области.
РСВ, который был аттенуированн, как описано в настоящем описании, можно исследовать на различных хорошо известных и общепринятых моделях in vitro и in vivo для подтверждения адекватной аттенуации, устойчивости к обратимости фенотипа, и иммуногенности для применения в качестве вакцины. В анализах in vitro, модифицированный вирус, который может быть многократно аттенуированным, биологически полученным или рекомбинантным РСВ, тестируют на чувствительность репликации вируса к температуре «фенотип ts» и на фенотип с малыми бляшками. Модифицированные вирусы дополнительно тестируют на моделях инфекции РСВ на животных. Ряд моделей на животных (например, мыши, хлопковый хомяк и примат) описаны и известны специалистам в данной области.
В соответствии с вышеуказанным описанием и на основании примеров, изложенных далее, изобретение также относится к композициям выделенного инфекционного РСВ для применения в качестве вакцин. Аттенуированный вирус, который является компонентом вакцины, находится в выделенной и, как правило, очищенной форме. Под выделенным подразумевается ссылка для обозначения РСВ, который находится в окружении ином, чем природное окружение вируса дикого типа, такое как носоглотка зараженного индивидуума. В более общем смысле, выделенный означает включение аттенуированного вируса в виде компонента клеточной культуры или другой искусственной среды. Например, аттенуированный РСВ по изобретению можно получать путем заражения клеточной культуры, выделения из клеточной культуры и добавления стабилизатора.
Вакцины с РСВ по изобретению содержат в качестве активного ингредиента иммуногенно эффективное количество РСВ, полученного, как описано в настоящем описании. Биологически произведенный или рекомбинантный РСВ можно использовать непосредственно в составах вакцин. Биологически произведенный или рекомбинантно модифицированный вирус можно вводить хозяину с физиологически приемлемым носителем и/или адъювантом. Подходящие носители хорошо известны в данной области, и включают, например, воду, забуференную воду, 0,4% физиологический раствор, 0,3% глицин, гиалуроновую кислоту и т.п. Полученные водные растворы можно упаковывать для применения как есть, или в замороженной форме, которую размораживают перед применением, или лиофилизировать, лиофилизированный препарат комбинируют со стерильным раствором перед введением, как указано выше. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, которые требуются для приближения к физиологическим условиям, и которые в качестве неограничивающих примеров включают средства для коррекции pH и буферные средства, средства для коррекции тоничности, увлажнители и т.п., например, ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция, сахарозу, сульфат магния, фосфатные буферы, буфер HEPES (4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота), сорбитанмонолаурат и триэтаноламин олеат. Приемлемые адъюванты включают неполный адъювант Фрейнда, алюминий фосфат, гидроксид алюминия, или квасцы, которые хорошо известны в данной области. Предпочтительные адъюванты также включают Stimulon™ QS-21 (Aquila Biopharmaceuticals, Inc., Worchester, Mass.), MPL™ (3-0-деацетилированный монофосфорил липид A; RIBI ImmunoChem Research, Inc., Hamilton, Mont.), и интерлейкин-12 (Genetics Institute, Cambridge, Mass.).
После иммунизации вакцинной композицией с РСВ, хозяин отвечает на вакцину, продуцируя антитела, специфичные для вирусных белков РСВ, например, гликопротеинов F и G. Кроме того, индуцируются врожденный иммунный ответ и клеточно-опосредованный иммунный ответ, которые могут обеспечить противовирусные эффекторы, а также регуляцию иммунного ответа. В результате вакцинации хозяин становится, по крайней мере, частично или полностью невосприимчивым к инфекции РСВ, либо устойчив к развивающимся умеренному или тяжелому заболеванию РСВ, особенно нижних дыхательных путей.
Вакцинные композиции, содержащие аттенуированный РСВ по изобретению вводят индивидууму, предрасположенному к инфекции РСВ или иным образом имеющему риск инфекции РСВ, в «иммуногенно эффективной дозе», которая является достаточной для индуцирования или усиления способностей иммунного ответа индивидуума против РСВ. Вакцинную композицию с РСВ можно вводить любым подходящим способом, включая в качестве неограничивающих примеров, инъекцию, аэрозоль, назальный спрей, капли в нос, пероральную вакцинацию, или местное применение. В случае людей, аттенуированный вирус по изобретению вводят в соответствии с хорошо разработанными протоколами вакцинирования РСВ человека (Karron et al. JID 191:1093-104, 2005). В кратком изложении, взрослые или дети получают интраназально путем капель иммуногенно эффективную дозу вакцины РСВ, как правило, в объеме 0,5 мл физиологически приемлемого разбавителя или носителя. Это имеет преимущество простоты и безопасности по сравнению с парентеральной иммунизацией не реплицирующейся вакциной. Это также обеспечивает прямую стимуляцию местного иммунитета дыхательных путей, который играет ключевую роль в устойчивости к РСВ. Кроме того, этот способ вакцинации эффективно обходит иммуносупрессивные эффекты РСВ-специфических сывороточных антител, полученных от матери, которые, как правило, встречаются у очень маленьких детей. Также, хотя парентеральное введение антигенов РСВ может иногда быть ассоциировано с иммунопатологическими осложнениями, этого не наблюдают с живым вирусом.
В некоторых вариантах осуществления вакцину можно вводить интраназально или подкожно, или внутримышечно. В некоторых вариантах осуществления ее можно вводить в верхние дыхательные пути. Введение можно проводить любым подходящим способом, включая в качестве неограничивающих примеров, спрей, капли или аэрозоль. Часто, композицию будут вводить индивидууму, серонегативному по антителам к РСВ или обладающему материнскими антителами к РСВ, полученными через плаценту
У всех индивидуумов точное количество вводимой вакцины РСВ и время, и повторность введения будут определяться различными факторами, в том числе состоянием здоровья пациента и его массой, способом введения, природой состава, и т.д. Дозировкм будут, как правило, находиться в диапазоне приблизительно от 3,0 log10 до приблизительно 6,0 log10 бляшкообразующих единиц («БОЕ») или более вируса на пациента, более часто приблизительно от 4,0 log10 до 5,0 log10 БОЕ вируса на пациента. В одном из вариантов осуществления приблизительно от 5,0 log10 до 6,0 log10 БОЕ на пациента можно вводить младенцам, таким как в возрасте от 1 до 6 месяцев, и одну или более дополнительных бустерных доз можно дать через 2-6 месяцев или значительно позже. В другом варианте осуществления младенцы могут получить дозу приблизительно от 5,0 log10 до 6,0 log10 БОЕ на пациента приблизительно в возрасте 2, 4, и 6 месяцев, что является рекомендуемым сроком введения ряда других детских вакцин. В еще одном варианте осуществления дополнительную бустерную дозу можно вводить приблизительно в возрасте 10-15 месяцев. В любом случае, составы вакцин должны обеспечивать количество аттенуированного РСВ по изобретению, достаточное для эффективного стимулирования или индуцирования иммунного ответа против РСВ («эффективное количество»).
В некоторых вариантах осуществления вакцина может содержать аттенуированный вирус с делецией M2-2, который вызывает иммунный ответ против одного штамма или антигенной подгруппы РСВ, например, A или B, или против нескольких штаммов или подгрупп РСВ. В отношении этого, мутанты РСВ с делецией M2-2 можно комбинировать в составах вакцин с другими штаммами или подгруппами РСВ для вакцин, имеющими другие иммуногенные характеристики для более эффективной защиты против одного или нескольких штаммов или подгрупп РСВ. Их можно вводить в смеси вакцин или вводить раздельно по координированному протоколу лечения для индукции более эфективной защиты против одного штамма РСВ, или против нескольких штаммов или подгрупп РСВ.
Полученный иммунный ответ можно характеризовать рядом способов. Эти способы включают забор образцов носовых смывов или сыворотки для анализа на РСВ-специфические антитела, которые можно выявлять тестами, включая в качестве неограничивающих примеров, фиксацию комплементом, подавление бляшкообразования, твердофазный иммуноферментный анализ, анализ иммунопреципитации с люциферазой, и проточную цитометрию. Кроме того, иммунные ответы можно выявлять путем анализа цитокинов в носовых смывах или сыворотке, анализа ELISPOT иммунных клеток из другого источника, количественной ОТ-ПЦР или микропанельного анализа образцов носовых смывов или сыворотки, и повторной стимуляции иммунных клеток из носовых смывов или сыворотки путем повторного воздействия вирусного антигена in vitro и анализа выработки или наличия цитокинов, поверхностных маркеров, или других иммунных коррелятов, измеренных проточной цитометрией или для цитотоксической активности против индикаторных клеток-мишеней, несущих антигены РСВ. В отношении этого, индивидуумов также проверяют на признаки и симптомы заболевания верхних дыхательных путей.
Уровень аттенуации вируса в вакцине можно определять, например, путем оценки количества вируса, присутствующего в дыхательных путях иммунизированного хозяина и сравнения количества с количеством, производимым РСВ дикого типа или другими аттенуированными РСВ, которые оценивали как кандидатные штаммы для вакцин. Например, аттенуированный вирус по изобретению будет иметь ограниченную в большей степени репликацию в верхних дыхательных путях высоковосприимчивого хозяина, такого как шимпанзе, по сравнению с уровнями репликации вируса дикого типа, например, от 10 до 1000 раз меньше. Для того чтобы дополнительно снизить развитие насморка, который связан с репликацией вируса в верхних дыхательных путях, идеальный вирус-кандидат для вакцины должен демонстрировать ограниченный уровень репликации и в верхних, и в нижних дыхательных путях. Однако, аттенуированные вирусы по изобретению должны быть достаточно инфекционными и иммуногенными у людей, чтобы обеспечить защиту у вакцинированных индивидуумов. Способы для определения уровней РСВ в носоглотке инфицированного хозяина хорошо известны в литературе. Образцы получают путем аспирации или вымывания носоглоточного секрета, и вирус оценивают в тканевой культуре или иным лабораторным способом. См., например, Belshe et al., J. Med. Virology 1:157-162 (1977), Friedewald et al., J. Amer. Med. Assoc. 204:690-694 (1968); Gharpure et al., J. Virol. 3:414-421 (1969); и Wright et al., Arch. Ges. Virusforsch. 41:238-247 (1973). Вирус можно удобно измерять в носоглотке животных-хозяев, таких как шимпанзе.
Дополнительные варианты осуществления
Пункт 1. Выделенная полинуклеотидная молекула, кодирующая рекомбинантный вариант респираторно-синцитиального вируса (РСВ) с аттенуированным фенотипом, содержащая геномную или антигеномную последовательность РСВ, где
(a) геном или антигеном РСВ модифицирован путем делеции в M2-2 ORF, соответствующей делеции, содержащей делецию из 241 нуклеотида, расположенных в положениях 8189-8429 SEQ ID NO: 1 в сочетании с мутациями в положениях T8161, T8167 и T8179 SEQ ID NO: 1; или
(b) геном или антигеном РСВ модифицирован путем делеции в M2-2 ORF, соответствующей делеции, содержащей делецию из 234 нуклеотидов, расположенных в положениях 8203-8436 SEQ ID NO: 1 в сочетании с присутствием 8198A и 8200G SEQ ID NO: 1; или
(c) геном или антигеном РСВ имеет последовательность «плюс»-цепи, обозначенную в SEQ ID NO: 1, модифицированную путем делеции в M2-2 ORF, содержащей делецию из 234 нуклеотидов, расположенных в положениях 8203-8436 SEQ ID NO: 1 в сочетании с присутствием 8198A и 8199G SEQ ID NO: 1.
Пункт 2. Выделенная полинуклеотидная молекула по п.1, где геном или антигеном РСВ, изложенный в a и b, имеет последовательность «плюс»-цепи, обозначенную последовательностью, которая по меньшей мере на 90% идентична последовательности, обозначенной SEQ ID NO: 1.
Пункт 3. Выделенная полинуклеотидная молекула по п.1, где геном или антигеном РСВ дополнительно модифицирован делецией из 112 нуклеотидов, расположенных в положениях 4499-4610 SEQ ID NO: 1 в сочетании с мутациями C4489T, C4492T, A4495T, A4497G, и G4498A SEQ ID NO: 1 («6120»).
Пункт 4. Выделенная полинуклеотидная молекула, кодирующая рекомбинантный вариант респираторно-синцитиального вируса (РСВ) с аттенуированным фенотипом, содержащая геномную или антигеномную последовательность РСВ
с последовательностью «плюс»-цепи, обозначенной последовательностью, которая по меньшей мере на 90% идентична последовательности, обозначенной SEQ ID NO: 1, где геном или антигеном РСВ модифицирован путем делеции в M2-2 ORF, соответствующей делеции, содержащей делецию из 234 нуклеотидов, расположенных в положениях 8203-8436 SEQ ID NO: 1,
где геном или антигеном РСВ дополнительно модифицирован делецией из 112 нуклеотидов, расположенных в положениях 4499-4610 SEQ ID NO: 1 в сочетании с мутациями C4489T, C4492T, A4495T, A4497G, и G4498A SEQ ID NO: 1.
Пункт 5. Выделенная полинуклеотидная молекула по пп.1, 2, 3 или 4, где геном или антигеном РСВ дополнительно модифицирован введением одной или нескольких следующих замен в SEQ ID NO: 1:
мутации, кодирующие замены аминокислот V267I в белке N, E218A и T523I в белке F, и C319Y и H1690Y в белке L РСВ («cp»);
мутации, кодирующие замены аминокислот K66E и Q101P в белке F РСВ («HEK»);
делеция из 419 нуклеотидов, расположенных в положениях 4198-4616 SEQ ID NO: 1, которая кодирует делецию белка SH РСВ (ΔSH);
мутация, кодирующая замену аминокислоты K51R в белке NS2 РСВ («NS2»);
мутация, кодирующая замеу аминокислоты T24A в белке N РСВ («N»);
нуклеотидная последовательность, кодирующая белок G РСВ замещена соответствующей кодон-оптимизированной нуклеотидной последовательностью, кодирующей белок G из клинического изолята A/Maryland/001/11;
нуклеотидная последовательность, кодирующая белок F РСВ, замещена соответствующей нуклеотидной последовательностью, кодирующей белок F из клинического изолята A/Maryland/001/11; или соответствующей кодон-оптимизированной нуклеотидной последовательностью, кодирующей белок F из клинического изолята A/Maryland/001/11; или кодон-оптимизированной последовательностью FBB («FBB»); и
порядок нуклеотидных последовательностей, кодирующих белки G и F РСВ SEQ ID NO: 1 является обратным.
Пункт 6. Выделенная полинуклеотидная молекула по пункту 1(a) или 2, содержащая нуклеотидную последовательность SEQ ID NO: 2.
Пункт 7. Выделенная полинуклеотидная молекула по пункту 1(b) или 2, содержащая нуклеотидную последовательность SEQ ID NO: 3.
Пункт 8. Выделенная полинуклеотидная молекула по пункту 1(c), содержащая нуклеотидную последовательность SEQ ID NO: 4.
Пункт 9. Выделенная полинуклеотидная молекула по пункту 3, содержащая нуклеотидную последовательность SEQ ID NO: 5.
Пункт 10. Выделенная полинуклеотидная молекула по пункту 3 или 4, содержащая нуклеотидную последовательность SEQ ID NO: 6.
Пункт 11. Выделенная полинуклеотидная молекула по пункту 3 или 4, содержащая нуклеотидную последовательность SEQ ID NO: 7.
Пункт 12. Выделенная полинуклеотидная молекула по пункту 5, где модифицированный геном или антигеном РСВ содержит комбинацию мутаций, выбранных из группы, состоящей из:
cp/ΔM2-2, cp/ΔM2-2/HEK, ΔM2-2/1030s, NS2/N/ΔM2-2, NS2/ΔM2-2, N/ΔM2-2, NS2/N/ΔM2-2-AclI, ΔSH/ΔM2-2, cp/ΔSH/ΔM2-2, 6120/cp/ΔM2-2, 6120/ΔM2-2/1030s, 6120/NS2/N/ΔM2-2, 6120/G001BB/FBB/ΔM2-2, 6120/FBB/G001BB/ΔM2-2, 6120/G001BB/F/ΔM2-2, 6120/G/FBB/ΔM2-2, 6120/G/FBB/HEK/ΔM2-2, 6120/G/FBB/cp/HEK/ΔM2-2, 6120/FBB/G/ΔM2-2, 6120/G001BB/F001BB/ΔM2-2, 6120/NS2/ΔM2-2, 6120/N/ΔM2-2, 6120/NS2/N/ΔM2-2-Acl-I, NS2/N/ΔM2-2-HindIII, и 6120/NS2/N/ΔM2-2-HindIII.
Пункт 13. Выделенная полинуклеотидная молекула по пункту 1, где геном или антигеном РСВ содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3 и SEQ ID NO: 4, дополнительно модифицированную введением одной или нескольких следующих нуклеотидных замен в SEQ ID NO: 2, SEQ ID NO: 3 и SEQ ID NO: 4: 404C, 779G, 1099T, 1139A, 1140G, 1182G, 1210G, 1939A, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T, 13634A, 13901T.
Пункт 14. Выделенная полинуклеотидная молекула по любому из пунктов 1-12, дополнительно содержащая ранее охарактеризованную мутацию или делецию РСВ.
Пункт 15. Вектор, содержащий выделенную полинуклеотидную молекулу по любому из пунктов 1-13.
Пункт 16. Клетка, содержащая выделенный полинуклеотид по любому из пунктов 1-13.
Пункт 17. Фармацевтическая композиция, содержащая иммунологически эффективное количество рекомбинантного варианта РСВ, кодируемого выделенной полинуклеотидной молекулой по любому из пунктов 1-13.
Пункт 18. Способ вакцинации индивидуума против РСВ, включающий введение фармацевтической композиции по пункту 16.
Пункт 19. Способ по пункту 17, где фармацевтическую композицию вводят интраназально.
Пункт 20. Способ по пункту 17, где респираторно-синцитиальный вирус вводят путем инъекции, аэрозоля, назального спрея или капель в нос.
Примеры последовательностей
Антигеномная последовательность кДНК D46 (SEQ ID NO: 1)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTCAACACATAGCATTCATCAATCCAACAGCCCAAAACAGTAACCTTGCATTTAAAAATGAACAACCCCTACCTCTTTACAACACCTCATTAACATCCCACCATGCAAACCACTATCCATACTATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAATGACCATGCCAAAAATAATGATACTACCTGACAAATATCCTTGTAGTATAACTTCCATACTAATAACAAGTAGATGTAGAGTTACTATGTATAATCAAAAGAACACACTATATTTCAATCAAAACAACCCAAATAACCATATGTACTCACCGAATCAAACATTCAATGAAATCCATTGGACCTCTCAAGAATTGATTGACACAATTCAAAATTTTCTACAACATCTAGGTATTATTGAGGATATATATACAATATATATATTAGTGTCATAACACTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК D46/ΔM2-2 (SEQ ID NO: 2)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTCAACACATAGCATTCATCAATCCAACAGCCCAAAACAGTAACCTTGCATTTAAAAATGAACAACCCCTACCTCTTTACAACACCTCATTAACATCCCACCATGCAAACCACTATCCATACTATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAACGACCACGCCAAAAATAACGATACTACCTAACACTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК D46/ΔM2-2-AclI (SEQ ID NO: 3)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTCAACACATAGCATTCATCAATCCAACAGCCCAAAACAGTAACCTTGCATTTAAAAATGAACAACCCCTACCTCTTTACAACACCTCATTAACATCCCACCATGCAAACCACTATCCATACTATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAATGACCATGCCAAAAATAATGATACTACCTGACAAATAACGTTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК D46/ΔM2-2-HindIII (SEQ ID NO: 4)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTCAACACATAGCATTCATCAATCCAACAGCCCAAAACAGTAACCTTGCATTTAAAAATGAACAACCCCTACCTCTTTACAACACCTCATTAACATCCCACCATGCAAACCACTATCCATACTATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAATGACCATGCCAAAAATAATGATACTACCTGACAAATAAGCTTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК LID/ΔM2-2 (SEQ ID NO: 5)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTTAATACTTGATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAACGACCACGCCAAAAATAACGATACTACCTAACACTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК LID/ΔM2-2-AclI (SEQ ID NO: 6)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTTAATACTTGATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAATGACCATGCCAAAAATAATGATACTACCTGACAAATAACGTTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК LID/ΔM2-2-HindIII (SEQ ID NO: 7)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTTAATACTTGATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAATGACCATGCCAAAAATAATGATACTACCTGACAAATAAGCTTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Полинуклеотидная последовательность, кодирующая G001BB (SEQ ID NO: 8)
ATGTCTAAGACAAAGGATCAGCGGACAGCCAAAACACTGGAACGGACATGGGATACCCTGAATCACCTCCTCTTCATCAGCAGTTGCCTGTACAAGCTCAATCTGAAGTCCATCGCCCAGATCACTCTCTCCATCCTTGCCATGATCATCTCTACAAGCCTCATCATTGCCGCAATTATCTTCATCGCCAGCGCTAACCACAAGGTCACCCTTACCACAGCCATTATTCAGGATGCCACCAACCAGATCAAGAACACAACCCCTACCTACCTGACACAGAACCCTCAGCTTGGAATTTCACTGAGCAACCTGTCCGAAACCACATCTAAACCTACAACCATCTTGGCTCTGACCACACCAAACGCCGAGTCCACCCCACAAAGTACCACAGTGAAGACCAAAAACACCACAACCACACAGATTCAGCCAAGCAAGCCTACAACTAAGCAAAGGCAGAACAAGCCACAGAACAAACCCAACAACGACTTTCACTTTGAGGTGTTCAACTTTGTGCCCTGCTCCATTTGCTCCAACAACCCTACCTGTTGGGCTATCTGCAAGAGGATCCCCAACAAGAAGCCCGGCAGGAAGACTACTACTAAGCCTACTAAACAGCCAGCCATTAAGACCACTAAGAAGGACCCAAAGCCACAGACAACCAAGCCAAAGGAGGTGCTCACTACCAAGCCCACTGAGAAGCCCACCATTAACACCACTAAAACCAACATCCGCACAACATTGCTGACATCAAACATTACAGAGAACCAGGAGCACACAAGCCAGAAGGAGACACTGCATAGCACTACATCCGAAGGCAATCCCAGCCCAAGCCAGGTCTATACTACCTCAGAGTACCTGTCCCAGAGCCTGAGCCCTAGCAACACTACTAGATGGTAG
Полинуклеотидная последовательность, кодирующая FBB (SEQ ID NO: 9)
ATGGAGCTCCTCATTCTCAAAGCCAACGCAATCACAACAATTCTGACCGCCGTCACATTCTGCTTTGCCTCCGGACAGAACATCACAGAAGAGTTTTACCAAAGTACATGCAGCGCCGTGAGCAAAGGCTACCTGTCCGCCCTGAGGACAGGGTGGTACACATCCGTGATTACCATTGAGCTGAGTAATATCAAGAAGAACAAGTGCAACGGCACTGATGCCAAAGTGAAGCTCATTAAACAGGAACTCGATAAGTACAAGAACGCCGTGACTGAGCTCCAGCTGCTGATGCAGTCAACTCAGGCTACAAACAACAGAGCCCGGAGGGAGCTGCCCAGGTTTATGAACTACACCCTGAACAACGCCAAGAAGACCAACGTGACATTGAGCAAGAAGAGGAAGCGGCGGTTCCTGGGGTTCTTGCTAGGTGTGGGCAGCGCTATTGCTTCTGGCGTCGCCGTCTCCAAGGTGCTGCACCTGGAAGGCGAAGTGAATAAGATTAAGTCCGCACTGCTTAGCACCAATAAGGCCGTCGTGAGCCTGTCTAACGGAGTGAGTGTGCTCACAAGCAAGGTCCTCGATCTCAAGAACTACATTGATAAGCAGCTCCTGCCCATCGTCAACAAGCAGTCATGCTCCATTAGTAACATCGAGACCGTGATTGAATTTCAACAGAAGAACAACCGGCTCCTGGAGATTACTAGGGAGTTCAGCGTGAACGCCGGGGTGACAACACCAGTCTCCACCTATATGCTTACCAACAGCGAGTTGCTCTCCCTGATTAACGATATGCCAATTACAAACGACCAGAAGAAGCTGATGTCAAACAACGTCCAGATTGTCCGGCAGCAGTCCTACTCAATCATGTCCATTATTAAGGAGGAGGTCCTGGCTTACGTCGTGCAGCTGCCTCTTTATGGGGTGATCGACACCCCTTGCTGGAAGCTCCATACATCCCCTCTGTGCACTACCAACACCAAGGAGGGGTCCAACATCTGCTTGACAAGAACCGATCGCGGCTGGTACTGCGATAACGCAGGCAGTGTCTCCTTCTTTCCCCAGGCCGAGACTTGTAAGGTGCAGTCTAACCGCGTCTTCTGCGACACCATGAACAGCCTGACCCTTCCCAGCGAGGTGAACCTTTGTAACGTGGACATCTTCAACCCAAAGTATGATTGTAAGATTATGACTAGCAAAACCGATGTCAGCAGCAGCGTGATCACTAGCCTGGGCGCTATCGTCAGCTGCTACGGAAAGACTAAGTGCACCGCCAGCAACAAGAACAGAGGCATCATCAAGACCTTCAGTAATGGATGTGACTACGTGTCCAACAAAGGGGTGGATACAGTGAGCGTGGGAAACACATTGTACTACGTGAACAAACAGGAGGGGAAGTCCTTGTACGTGAAGGGTGAGCCCATTATCAACTTCTACGACCCTCTCGTGTTCCCATCAGACGAGTTTGACGCCTCCATCTCCCAGGTGAACGAGAAGATCAATCAGTCACTGGCCTTTATTAGGAAATCCGACGAGCTGCTGCACAACGTCAACGCCGGAAAGTCTACCACTAACATCATGATCACCACAATCATCATTGTGATCATCGTCATCCTCCTGAGCTTGATCGCTGTCGGGTTGCTGTTGTACTGCAAGGCCCGGTCCACACCCGTGACTCTGAGCAAGGACCAGCTGTCTGGCATTAACAACATCGCCTTTAGCAACTAA
Полинуклеотидная последовательность, кодирующая F001 (SEQ ID NO: 10)
ATGGATTTGCCAATCCTCAAGACAAATGCTATTACCACAATCCTTGCTGCAGTCACACTCTGTTTCGCTTCCAGTCAAAATATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTAAGAACTGGTTGGTATACTAGTGTTATAACTATAGAATTAAGTAATATCAAAGAAAATAAGTGTAATGGAACAGACGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAACTGCAGTTGCTCATGCAAAGCACGCCAGCATCCAACAATCGAGCCAGAAGAGAACTACCAAGATTTATGAATTATACACTCAACAATACCAAAAACACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGCTTTTTGTTAGGGGTTGGATCTGCAATCGCCAGTGGCATTGCTGTATCTAAGGTCCTGCACTTAGAAGGGGAAGTGAACAAAATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCTAATGGAGTCAGTGTCTTAACCAGCAAAGTGTTAGATCTCAAAAACTATATAGATAAACAGTTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAACATTGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGTGTAACTACACCTGTAAGCACTTATATGTTAACTAATAGTGAATTATTATCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCAATAATAAAGGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTAATAGATACACCTTGTTGGAAACTGCACACATCCCCTCTATGTACAACCTACACAAAGGAAGGGTCCAACATCTGCTTAACAAGAACCGACAGGGGATGGTACTGTGACAATGCAGGATCAGTATCTTTTTTCCCACAAGCTGAAACATGTAAAGTTCAATCGAATCGGGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAGGTAAATCTCTGCAACATTGACATATTCAACCCCAAATATGATTGCAAAATTATGACTTCAAAAACAGATGTAAGTAGCTCTGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACCAAATGTACAGCATCCAATAAAAATCGTGGGATCATAAAGACATTTTCTAATGGGTGTGATTATGTATCAAATAAGGGGGTGGATACTGTGTCTGTAGGTAATACATTATATTATGTAAATAAGCAAGAAGGCAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGATCCATTAGTTTTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAATGAGAAGATTAACCAGAGTCTAGCATTTATCCGTAAATCAGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTAATTGTAGTAATATTGTTATCATTAATTGCAGTTGGACTGCTTCTATACTGCAAGGCCAGAAGCACACCAGTCACACTAAGTAAGGATCAACTGAGTGGTATAAACAATATTGCATTTAGTAGCTGA
Полинуклеотидная последовательность, кодирующая F001BB (SEQ ID NO: 11)
ATGGACCTTCCAATCCTGAAGACCAACGCTATCACCACCATCCTCGCAGCTGTGACTCTTTGTTTCGCATCCTCCCAAAACATCACCGAAGAGTTCTACCAGTCCACCTGTTCCGCAGTGTCTAAGGGATACCTTAGCGCTCTCAGAACCGGATGGTACACATCCGTGATCACTATCGAACTGAGCAACATCAAGGAGAACAAGTGCAACGGCACCGACGCTAAGGTGAAGCTCATCAAGCAGGAACTGGACAAGTACAAGAACGCCGTGACCGAACTTCAGCTCCTTATGCAGTCTACCCCAGCTTCCAACAACAGAGCCAGAAGGGAGCTCCCAAGGTTTATGAACTACACCCTCAACAACACCAAGAACACCAACGTGACCCTGTCCAAGAAGAGGAAGAGGCGGTTCCTTGGATTCCTCTTGGGAGTCGGATCTGCTATCGCCTCAGGCATTGCCGTCAGTAAAGTGTTGCATTTGGAGGGCGAGGTCAACAAAATCAAGTCCGCCTTGTTGTCCACTAACAAGGCCGTCGTGTCTTTGTCCAACGGGGTGTCTGTCTTGACAAGTAAGGTGTTGGACTTGAAGAACTACATCGACAAGCAGCTGCTGCCTATCGTCAACAAGCAGTCCTGCTCTATCAGCAACATCGAGACCGTGATCGAGTTCCAGCAGAAGAACAACCGGCTGCTGGAGATCACAAGGGAGTTCAGTGTCAACGCCGGCGTCACAACACCTGTGTCAACTTATATGCTGACAAACTCAGAGCTGCTGTCACTGATCAACGACATGCCTATCACCAACGACCAGAAGAAGCTGATGAGCAACAACGTGCAGATCGTGAGGCAGCAGTCATACAGCATCATGTCCATCATCAAGGAGGAAGTCCTGGCCTACGTGGTCCAACTGCCTCTGTACGGCGTGATTGATACTCCATGTTGGAAGCTGCACACATCACCACTGTGTACCACTTACACCAAGGAGGGGAGTAACATCTGCCTGACTCGGACAGATAGAGGGTGGTATTGCGATAATGCCGGCAGTGTCTCCTTTTTCCCCCAGGCCGAGACTTGCAAAGTCCAGAGCAATCGCGTGTTTTGCGATACAATGAATAGCCTGACACTCCCCAGCGAGGTGAATCTCTGCAATATTGATATTTTCAACCCCAAGTACGACTGCAAGATCATGACCAGCAAGACCGACGTCAGCAGCAGCGTGATTACTAGCCTCGGAGCCATTGTGAGCTGCTATGGGAAAACAAAATGCACAGCCTCCAACAAAAACAGAGGCATTATCAAGACTTTCTCCAACGGGTGCGATTACGTGTCCAACAAGGGCGTGGATACTGTGAGCGTGGGGAACACACTCTACTACGTGAACAAACAGGAGGGGAAAAGCCTGTACGTGAAAGGCGAGCCCATTATTAACTTTTACGACCCTCTGGTGTTTCCCAGCGATGAGTTTGATGCCAGCATCTCCCAGGTGAACGAGAAGATTAACCAGTCCCTCGCCTTTATTCGCAAGAGCGATGAGCTGCTGCACAACGTGAACGCCGGCAAGTCCACTACAAACATTATGATTACAACAATTATTATTGTCATTGTCGTCATTCTGCTCAGCCTGATTGCCGTCGGCCTGCTGCTCTACTGCAAGGCCAGGTCCACACCCGTGACACTCAGCAAGGATCAGCTGTCCGGCATTAACAACATTGCCTTTAGCAGCTAA
Антигеномная последовательность кДНК D46/cp/ΔM2-2 (SEQ ID NO: 15)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGATTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTaACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTCAACACATAGCATTCATCAATCCAACAGCCCAAAACAGTAACCTTGCATTTAAAAATGAACAACCCCTACCTCTTTACAACACCTCATTAACATCCCACCATGCAAACCACTATCCATACTATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGcAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCAtAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAAcGACCAcGCCAAAAATAAcGATACTACCTAACACTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTacATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAATACATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК LID/ΔM2-2/1030s (SEQ ID NO: 16)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTTAATACTTGATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAACGACCACGCCAAAAATAACGATACTACCTAACACTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCTCAATAGGAACCCTTGGGTTAACAAAAGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК LID/cp/ΔM2-2 (SEQ ID NO: 17)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAAACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACACCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGATTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTTAATACTTGATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGCAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCATAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAACGACCACGCCAAAAATAACGATACTACCTAACACTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTACATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAATACATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК D46/NS2/N/ΔM2-2-HindIII (SEQ ID NO: 18)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAATGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAGACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAACTTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAACCATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAAGATCAACTTCTGTCATCCAGCAAATACGCCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTCAACACATAGCATTCATCAATCCAACAGCCCAAAACAGTAACCTTGCATTTAAAAATGAACAACCCCTACCTCTTTACAACACCTCATTAACATCCCACCATGCAAACCACTATCCATACTATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAGGCCTTGACCAACTTAAACAGAATCAAAATAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTTAGTGTTTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGCGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCTGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATCGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGCATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCCCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAATGACCATGCCAAAAATAATGATACTACCTGACAAATAAGCTTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAACTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACCCTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Антигеномная последовательность кДНК генома РСВ 276 (SEQ ID NO: 19)
ACGGGAAAAAATGCGTACAACAAACTTGCATAAACCAAAAAAATGGGGCAAATAAGAATTTGATAAGTACCACTTAAATTTAACTCCCTTGGTTAGAGATGGGCAGCAATTCATTGAGTATGATAAAAGTTAGATTACAAAATTTGTTTGACAATGATGAAGTAGCATTGTTAAAAATAACATGCTATACTGATAAATTAATACATTTAACTAATGCTTTGGCTAAGGCAGTGATACATACAATCAAATTGAATGGCATTGTGTTTGTGCATGTTATTACAAGTAGTGATATTTGCCCTAATAATAATATTGTAGTAAAATCCAATTTCACAACAATGCCAGTACTACAAAATGGAGGTTATATATGGGAAATGATGGAATTAACACATTGCTCTCAACCTAACGGTCTACTAGATGACAATTGTGAAATTAAATTCTCCAAAAAACTAAGTGATTCAACAATGACCAATTATATGAATCAATTATCTGAATTACTTGGATTTGATCTTAATCCATAAATTATAATTAATATCAACTAGCAAATCAATGTCACTAACACCATTAGTTAATATAAAACTTAACAGAAGACAAAAATGGGGCAAATAAATCAATTCAGCCAACCCAACCATGGACACAACCCACAATGATAATACACCACAAAGACTGATGATCACAGACATGAGACCGTTGTCACTTGAGACCATAATAACATCACTAACCAGAGACATCATAACACACAAATTTATATACTTGATAAATCATGAATGCATAGTGAGAAGACTTGATGAAAGACAGGCCACATTTACATTCCTGGTCAACTATGAAATGAAACTATTACACAAAGTAGGAAGCACTAAATATAAAAAATATACTGAATACAACACAAAATATGGCACTTTCCCTATGCCAATATTCATCAATCATGATGGGTTCTTAGAATGCATTGGCATTAAGCCTACAAAGCATACTCCCATAATATACAAGTATGATCTCAATCCATAAATTTCAACACAATATTCACACAATCTAAAACAACAACTCTATGCATAACTATACTCCATAGTCCAGATGGAGCCTGAAAATTATAGTAATTTAAAATTAAGGAGAGATATAAGATAGAAGATGGGGCAAATACAAAGATGGCTCTTAGCAAAGTCAAGTTGAATGATACACTCAACAAGGATCAACTTCTGTCATCCAGCAAATACGCCATCCAACGGAGCACAGGAGATAGTATTGATACTCCTAATTATGATGTGCAGAAACACATCAATAAGTTATGTGGCATGTTATTAATCACAGAAGATGCTAATCATAAATTCACTGGGTTAATAGGTATGTTATATGCGATGTCTAGGTTAGGAAGAGAAGACACCATAAAAATACTCAGAGATGCGGGATATCATGTAAAAGCAAATGGAGTAGATGTAACAACACATCGTCAAGACATTAATGGAAAAGAAATGAAATTTGAAGTGTTAACATTGGCAAGCTTAACAACTGAAATTCAAATCAACATTGAGATAGAATCTAGAAAATCCTACAAAAAAATGCTAAAAGAAATGGGAGAGGTAGCTCCAGAATACAGGCATGACTCTCCTGATTGTGGGATGATAATATTATGTATAGCAGCATTAGTAATAACTAAATTAGCAGCAGGGGACAGATCTGGTCTTACAGCCGTGATTAGGAGAGCTAATAATGTCCTAAAAAATGAAATGAAACGTTACAAAGGCTTACTACCCAAGGACATAGCCAACAGCTTCTATGAAGTGTTTGAAAAACATCCCCACTTTATAGATGTTTTTGTTCATTTTGGTATAGCACAATCTTCTACCAGAGGTGGCAGTAGAGTTGAAGGGATTTTTGCAGGATTGTTTATGAATGCCTATGGTGCAGGGCAAGTGATGTTACGGTGGGGAGTCTTAGCAAAATCGGTTAAAAATATTATGTTAGGACATGCTAGTGTGCAAGCAGAAATGGAACAAGTTGTTGAGGTTTATGAATATGCCCAAAAATTGGGTGGTGAAGCAGGATTCTACCATATATTGAACAACCCAAAAGCATCATTATTATCTTTGACTCAATTTCCTCACTTCTCCAGTGTAGTATTAGGCAATGCTGCTGGCCTAGGCATAATGGGAGAGTACAGAGGTACACCGAGGAATCAAGATCTATATGATGCAGCAAAGGCATATGCTGAACAACTCAAAGAAAATGGTGTGATTAACTACAGTGTACTAGACTTGACAGCAGAAGAACTAGAGGCTATCAAACATCAGCTTAATCCAAAAGATAATGATGTAGAGCTTTGAGTTAATAAAAAATGGGGCAAATAAATCATCATGGAAAAGTTTGCTCCTGAATTCCATGGAGAAGATGCAAACAACAGGGCTACTAAATTCCTAGAATCAATAAAGGGCAAATTCACATCACCCAAAGATCCCAAGAAAAAAGATAGTATCATATCTGTCAACTCAATAGATATAGAAGTAACCAAAGAAAGCCCTATAACATCAAATTCAACTATTATCAACCCAACAAATGAGACAGATGATACTGCAGGGAACAAGCCCAATTATCAAAGAAAACCTCTAGTAAGTTTCAAAGAAGACCCTACACCAAGTGATAATCCCTTTTCTAAACTATACAAAGAAACCATAGAAACATTTGATAACAATGAAGAAGAATCCAGCTATTCATACGAAGAAATAAATGATCAGACAAACGATAATATAACAGCAAGATTAGATAGGATTGATGAAAAATTAAGTGAAATACTAGGAATGCTTCACACATTAGTAGTGGCAAGTGCAGGACCTACATCTGCTCGGGATGGTATAAGAGATGCCATGGTTGGTTTAAGAGAAGAAATGATAGAAAAAATCAGAACTGAAGCATTAATGACCAATGACAGATTAGAAGCTATGGCAAGACTCAGGAATGAGGAAAGTGAAAAGATGGCAAAAGACACATCAGATGAAGTGTCTCTCAATCCAACATCAGAGAAATTGAACAACCTATTGGAAGGGAATGATAGTGACAATGATCTATCACTTGAAGATTTCTGATTAGTTACCAATCTTCACATCAACACACAATACCAACAGAAGACCAACAAACTAACCAACCCAATCATCCAACCAAACATCCATCCGCCAATCAGCCAAACAGCCAACAAAACAACCAGCCAATCCAAAACTAACCACCCGGAAAAAATCTATAATATAGTTACAAAAAAAGGAAAGGGTGGGGCAAATATGGAAACATACGTGAACAAGCTTCACGAAGGCTCCACATACACAGCTGCTGTTCAATACAATGTCTTAGAAAAAGACGATGACCCTGCATCACTTACAATATGGGTGCCCATGTTCCAATCATCTATGCCAGCAGATTTACTTATAAAAGAACTAGCTAATGTCAACATACTAGTGAAACAAATATCCACACCCAAGGGACCTTCACTAAGAGTCATGATAAACTCAAGAAGTGCAGTGCTAGCACAAATGCCCAGCAAATTTACCATATGCGCTAATGTGTCCTTGGATGAAAGAAGCAAACTAGCATATGATGTAACCACACCCTGTGAAATCAAGGCATGTAGTCTAACATGCCTAAAATCAAAAAATATGTTGACTACAGTTAAAGATCTCACTATGAAGACACTCAACCCTACACATGATATTATTGCTTTATGTGAATTTGAAAACATAGTAACATCAAAAAAAGTCATAATACCAACATACCTAAGATCCATCAGTGTCAGAAATAAAGATCTGAACACACTTGAAAATATAACAACCACTGAATTCAAAAATGCTATCACAAATGCAAAAATCATCCCTTACTCAGGATTACTATTAGTCATCACAGTGACTGACAACAAAGGAGCATTCAAATACATAAAGCCACAAAGTCAATTCATAGTAGATCTTGGAGCTTACCTAGAAAAAGAAAGTATATATTATGTTACCACAAATTGGAAGCACACAGCTACACGATTTGCAATCAAACCCATGGAAGATTAACCTTTTTCCTCTACATCAGTGTGTTAATTCATACAAACTTTCTACCTACATTCTTCACTTCACCATCACAATCACAAACACTCTGTGGTTCAACCAATCAAACAAAACTTATCTGAAGTCCCAGATCATCCCAAGTCATTGTTTATCAGATCTAGTACTCAAATAAGTTAATAAAAAATATACACATGGGGCAAATAATCATTGGAGGAAATCCAACTAATCACAATATCTGTTAACATAGACAAGTCCACACACCATACAGAATCAACCAATGGAAAATACATCCATAACAATAGAATTCTCAAGCAAATTCTGGCCTTACTTTACACTAATACACATGATCACAACAATAATCTCTTTGCTAATCATAATCTCCATCATGATTGCAATACTAAACAAACTTTGTGAATATAACGTATTCCATAACAAAACCTTTGAGTTACCAAGAGCTCGAGTCAACACATAGCATTCATCAATCCAACAGCCCAAAACAGTAACCTTGCATTTAAAAATGAACAACCCCTACCTCTTTACAACACCTCATTAACATCCCACCATGCAAACCACTATCCATACTATAAAGTAGTTAATTAAAAATAGTCATAACAATGAACTAGGATATCAAGACTAACAATAACATTGGGGCAAATGCAAACATGTCCAAAAACAAGGACCAACGCACCGCTAAGACATTAGAAAGGACCTGGGACACTCTCAATCATTTATTATTCATATCATCGTGCTTATATAAGTTAAATCTTAAATCTGTAGCACAAATCACATTATCCATTCTGGCAATGATAATCTCAACTTCACTTATAATTGCAGCCATCATATTCATAGCCTCGGCAAACCACAAAGTCACACCAACAACTGCAATCATACAAGATGCAACAAGCCAGATCAAGAACACAACCCCAACATACCTCACCCAGAATCCTCAGCTTGGAATCAGTCCCTCTAATCCGTCTGAAATTACATCACAAATCACCACCATACTAGCTTCAACAACACCAGGAGTCAAGTCAACCCTGCAATCCACAACAGTCAAGACCAAAAACACAACAACAACTCAAACACAACCCAGCAAGCCCACCACAAAACAACGCCAAAACAAACCACCAAGCAAACCCAATAATGATTTTCACTTTGAAGTGTTCAACTTTGTACCCTGCAGCATATGCAGCAACAATCCAACCTGCTGGGCTATCTGCAAAAGAATACCAAACAAAAAACCAGGAAAGAAAACCACTACCAAGCCCACAAAAAAACCAACCCTCAAGACAACCAAAAAAGATCCCAAACCTCAAACCACTAAATCAAAGGAAGTACCCACCACCAAGCCCACAGAAGAGCCAACCATCAACACCACCAAAACAAACATCATAACTACACTACTCACCTCCAACACCACAGGAAATCCAGAACTCACAAGTCAAATGGAAACCTTCCACTCAACTTCCTCCGAAGGCAATCCAAGCCCTTCTCAAGTCTCTACAACATCCGAGTACCCATCACAACCTTCATCTCCACCCAACACACCACGCCAGTAGTTACTTAAAAACATATTATCACAAAAAGCCATGACCAACTTAAACAGAATCAAAGTAAACTCTGGGGCAAATAACAATGGAGTTGCTAATCCTCAAAGCAAATGCAATTACCACAATCCTCACTGCAGTCACATTTTGTTTTGCTTCTGGTCAAAACATCACTGAAGAATTTTATCAATCAACATGCAGTGCAGTTAGCAAAGGCTATCTTAGTGCTCTGAGAACTGGTTGGTATACCAGTGTTATAACTATAGAATTAAGTAATATCAAGAAAAATAAGTGTAATGGAACAGATGCTAAGGTAAAATTGATAAAACAAGAATTAGATAAATATAAAAATGCTGTAACAGAATTGCAGTTGCTCATGCAAAGCACACAAGCAACAAACAATCGAGCCAGAAGAGAACTACCAAGGTTTATGAATTATACACTCAACAATGCCAAAAAAACCAATGTAACATTAAGCAAGAAAAGGAAAAGAAGATTTCTTGGTTTTTTGTTAGGTGTTGGATCTGCAATCGCCAGTGGCGTTGCTGTATCTAAGGTCCTGCACCTAGAAGGGGAAGTGAACAAGATCAAAAGTGCTCTACTATCCACAAACAAGGCTGTAGTCAGCTTATCAAATGGAGTCAGTGTCTTAACCAGCAAAGTGTTAGACCTCAAAAACTATATAGATAAACAATTGTTACCTATTGTGAACAAGCAAAGCTGCAGCATATCAAATATAGAAACTGTGATAGAGTTCCAACAAAAGAACAACAGACTACTAGAGATTACCAGGGAATTTAGTGTTAATGCAGGTGTAACTACACCTGTAAGCACTTACATGTTAACTAATAGTGAATTATTGTCATTAATCAATGATATGCCTATAACAAATGATCAGAAAAAGTTAATGTCCAACAATGTTCAAATAGTTAGACAGCAAAGTTACTCTATCATGTCCATAATAAAAGAGGAAGTCTTAGCATATGTAGTACAATTACCACTATATGGTGTTATAGATACACCCTGTTGGAAACTACACACATCCCCTCTATGTACAACCAACACAAAAGAAGGGTCCAACATCTGTTTAACAAGAACTGACAGAGGATGGTACTGTGACAATGCAGGATCAGTATCTTTCTTCCCACAAGCTGAAACATGTAAAGTTCAATCAAATCGAGTATTTTGTGACACAATGAACAGTTTAACATTACCAAGTGAAGTAAATCTCTGCAATGTTGACATATTCAACCCCAAATATGATTGTAAAATTATGACTTCAAAAACAGATGTAAGCAGCTCCGTTATCACATCTCTAGGAGCCATTGTGTCATGCTATGGCAAAACTAAATGTACAGCATCCAATAAAAATCGTGGAATCATAAAGACATTTTCTAACGGGTGCGATTATGTATCAAATAAAGGGGTGGACACTGTGTCTGTAGGTAACACATTATATTATGTAAATAAGCAAGAAGGTAAAAGTCTCTATGTAAAAGGTGAACCAATAATAAATTTCTATGACCCATTAGTATTCCCCTCTGATGAATTTGATGCATCAATATCTCAAGTCAACGAGAAGATTAACCAGAGCCTAGCATTTATTCGTAAATCCGATGAATTATTACATAATGTAAATGCCGGTAAATCCACCACAAATATCATGATAACTACTATAATTATAGTGATTATAGTAATATTGTTATCATTAATTGCTGTTGGACTGCTCTTATACTGTAAGGCCAGAAGCACACCAGTCACACTAAGCAAAGATCAACTGAGTGGTATAAATAATATTGCATTTAGTAACTAAATAAAAATAGCACCTAATCATGTTCTTACAATGGTTTACTATCTGCTCATAGACAACCCATCTGTCATTGGATTTTCTTAAAATCTGAACTTCATTGAAACTCTCATCTATAAACCATCTCACTTACACTATTTAAGTAGATTCCTAGTTTATAGTTATATAAAACACAATTGAATGCCAGATTAACTTACCATCTGTAAAAATGAAAACTGGGGCAAATATGTCACGAAGGAATCCTTGCAAATTTGAAATTCGAGGTCATTGCTTAAATGGTAAGAGGTGTCATTTTAGTCATAATTATTTTGAATGGCCACCGCATGCACTGCTTGTAAGACAAAACTTTATGTTAAACAGAATACTTAAGTCTATGGATAAAAGTATAGATACCTTATCAGAAATAAGTGGAGCTGCAGAGTTGGACAGAACAGAAGAGTATGCTCTTGGTGTAGTTGGAGTGCTAGAGAGTTATATAGGATCAATAAACAATATAACTAAACAATCAGCATGTGTTGCCATGAGCAAACTCCTCACTGAACTCAATAGTGATGATATCAAAAAGCTGAGGGACAATGAAGAGCTAAATTCACCCAAGATAAGAGTGTACAATACTGTCATATCATATATTGAAAGCAACAGGAAAAACAATAAACAAACTATCCATCTGTTAAAAAGATTGCCAGCAGACGTATTGAAGAAAACCATCAAAAACACATTGGATATCCATAAGAGCATAACCATCAACAACCCAAAAGAATCAACTGTTAGTGATACAAATGACCATGCCAAAAATAATGATACTACCTGACAAATAACGTTCAATTCTAACACTCACCACATCGTTACATTATTAATTCAAACAATTCAAGTTGTGGGACAAAATGGATCCCATTATTAATGGAAATTCTGCTAATGTTTATCTAACCGATAGTTATTTAAAAGGTGTTATCTCTTTCTCAGAGTGTAATGCTTTAGGAAGTTACATATTCAATGGTCCTTATCTCAAAAATGATTATACCAACTTAATTAGTAGACAAAATCCATTAATAGAACACATGAATCTAAAGAAACTAAATATAACACAGTCCTTAATATCTAAGTATCATAAAGGTGAAATAAAATTAGAAGAACCTACTTATTTTCAGTCATTACTTATGACATACAAGAGTATGACCTCGTCAGAACAGATTGCTACCACTAATTTACTTAAAAAGATAATAAGAAGAGCTATAGAAATAAGTGATGTCAAAGTCTATGCTATATTGAATAAACTAGGGCTTAAAGAAAAGGACAAGATTAAATCCAACAATGGACAAGATGAAGACAACTCAGTTATTACGACCATAATCAAAGATGATATACTTTCAGCTGTTAAAGATAATCAATCTCATCTTAAAGCAGACAAAAATCACTCTACAAAACAAAAAGACACAATCAAAACAACACTCTTGAAGAAATTGATGTGTTCAATGCAACATCCTCCATCATGGTTAATACATTGGTTTAACTTATACACAAAATTAAACAACATATTAACACAGTATCGATCAAATGAGGTAAAAAACCATGGGTTTACATTGATAGATAATCAAACTCTTAGTGGATTTCAATTTATTTTGAACCAATATGGTTGTATAGTTTATCATAAGGAACTCAAAAGAATTACTGTGACAACCTATAATCAATTCTTGACATGGAAAGATATTAGCCTTAGTAGATTAAATGTTTGTTTAATTACATGGATTAGTAACTGCTTGAACACATTAAATAAAAGCTTAGGCTTAAGATGCGGATTCAATAATGTTATCTTGACACAACTATTCCTTTATGGAGATTGTATACTAAAGCTATTTCACAATGAGGGGTTCTACATAATAAAAGAGGTAGAGGGATTTATTATGTCTCTAATTTTAAATATAACAGAAGAAGATCAATTCAGAAAACGATTTTATAATAGTATGCTCAACAACATCACAGATGCTGCTAATAAAGCTCAGAAAAATCTGCTATCAAGAGTATGTCATACATTATTAGATAAGACAGTGTCCGATAATATAATAAATGGCAGATGGATAATTCTATTAAGTAAGTTCCTTAAATTAATTAAGCTTGCAGGTGACAATAACCTTAACAATCTGAGTGAACTATATTTTTTGTTCAGAATATTTGGACACCCAATGGTAGATGAAAGACAAGCCATGGATGCTGTTAAAATTAATTGCAATGAGACCAAATTTTACTTGTTAAGCAGTCTGAGTATGTTAAGAGGTGCCTTTATATATAGAATTATAAAAGGGTTTGTAAATAATTACAACAGATGGCCTACTTTAAGAAATGCTATTGTTTTACCCTTAAGATGGTTAACTTACTATAAACTAAACACTTATCCTTCTTTGTTGGAACTTACAGAAAGAGATTTGATTGTGTTATCAGGACTACGTTTCTATCGTGAGTTTCGGTTGCCTAAAAAAGTGGATCTTGAAATGATTATAAATGATAAAGCTATATCACCTCCTAAAAATTTGATATGGACTAGTTTCCCTAGAAATTACATGCCATCACACATACAAAACTATATAGAACATGAAAAATTAAAATTTTCCGAGAGTGATAAATCAAGAAGAGTATTAGAGTATTATTTAAGAGATAACAAATTCAATGAATGTGATTTATACAACTGTGTAGTTAATCAAAGTTATCTCAACAACCCTAATCATGTGGTATCATTGACAGGCAAAGAAAGAGAACTCAGTGTAGGTAGAATGTTTGCAATGCAACCGGGAATGTTCAGACAGGTTCAAATATTGGCAGAGAAAATGATAGCTGAAAACATTTTACAATTCTTTCCTGAAAGTCTTACAAGATATGGTGATCTAGAACTACAAAAAATATTAGAATTGAAAGCAGGAATAAGTAACAAATCAAATCGCTACAATGATAATTACAACAATTACATTAGTAAGTGCTCTATCATCACAGATCTCAGCAAATTCAATCAAGCATTTCGATATGAAACGTCATGTATTTGTAGTGATGTGCTGGATGAACTGCATGGTGTACAATCTCTATTTTCCTGGTTACATTTAACTATTCCTCATGTCACAATAATATGCACATATAGGCATGCACCCCCCTATATAGGAGATCATATTGTAGATCTTAACAATGTAGATGAACAAAGTGGATTATATAGATATCACATGGGTGGCATCGAAGGGTGGTGTCAAAAACTATGGACCATAGAAGCTATATCACTATTGGATCTAATATCTCTCAAAGGGAAATTCTCAATTACTGCTTTAATTAATGGTGACAATCAATCAATAGATATAAGCAAACCAATCAGACTCATGGAAGGTCAAACTCATGCTCAAGCAGATTATTTGCTAGCATTAAATAGCCTTAAATTACTGTATAAAGAGTATGCAGGCATAGGCCACAAATTAAAAGGAACTGAGACTTATATATCACGAGATATGCAATTTATGAGTAAAACAATTCAACATAACGGTGTATATTACCCAGCTAGTATAAAGAAAGTCCTAAGAGTGGGACCGTGGATAAACACTATACTTGATGATTTCAAAGTGAGTCTAGAATCTATAGGTAGTTTGACACAAGAATTAGAATATAGAGGTGAAAGTCTATTATGCAGTTTAATATTTAGAAATGTATGGTTATATAATCAGATTGCTCTACAATTAAAAAATCATGCATTATGTAACAATAAACTATATTTGGACATATTAAAGGTTCTGAAACACTTAAAAACCTTTTTTAATCTTGATAATATTGATACAGCATTAACATTGTATATGAATTTACCCATGTTATTTGGTGGTGGTGATCCCAACTTGTTATATCGAAGTTTCTATAGAAGAACTCCTGACTTCCTCACAGAGGCTATAGTTCACTCTGTGTTCATACTTAGTTATTATACAAACCATGACTTAAAAGATAAACTTCAAGATCTGTCAGATGATAGATTGAATAAGTTCTTAACATGCATAATCACGTTTGACAAAAACCCTAATGCTGAATTCGTAACATTGATGAGAGATCCTCAAGCTTTAGGGTCTGAGAGACAAGCTAAAATTACTAGCGAAATCAATAGACTGGCAGTTACAGAGGTTTTGAGTACAGCTCCAAACAAAATATTCTCCAAAAGTGCACAACATTATACTACTACAGAGATAGATCTAAATGATATTATGCAAAATATAGAACCTACATATCCTCATGGGCTAAGAGTTGTTTATGAAAGTTTACCCTTTTATAAAGCAGAGAAAATAGTAAATCTTATATCAGGTACAAAATCTATAACTAACATACTGGAAAAAACTTCTGCCATAGACTTAACAGATATTGATAGAGCCACTGAGATGATGAGGAAAAACATAACTTTGCTTATAAGGATACTTCCATTGGATTGTAACAGAGATAAAAGAGAGATATTGAGTATGGAAAACCTAAGTATTACTGAATTAAGCAAATATGTTAGGGAAAGATCTTGGTCTTTATCCAATATAGTTGGTGTTACATCACCCAGTATCATGTATACAATGGACATCAAATATACTACAAGCACTATATCTAGTGGCATAATTATAGAGAAATATAATGTTAACAGTTTAACACGTGGTGAGAGAGGACCCACTAAACCATGGGTTGGTTCATCTACACAAGAGAAAAAAACAATGCCAGTTTATAATAGACAAGTCTTAACCAAAAAACAGAGAGATCAAATAGATCTATTAGCAAAATTGGATTGGGTGTATGCATCTATAGATAACAAGGATGAATTCATGGAAGAACTCAGCATAGGAACCCTTGGGTTAACATATGAAAAGGCCAAGAAATTATTTCCACAATATTTAAGTGTCAATTATTTGCATCGCCTTACAGTCAGTAGTAGACCATGTGAATTCCCTGCATCAATACCAGCTTATAGAACAACAAATTATCACTTTGACACTAGCCCTATTAATCGCATATTAACAGAAAAGTATGGTGATGAAGATATTGACATAGTATTCCAAAACTGTATAAGCTTTGGCCTTAGTTTAATGTCAGTAGTAGAACAATTTACTAATGTATGTCCTAACAGAATTATTCTCATACCTAAGCTTAATGAGATACATTTGATGAAACCTCCCATATTCACAGGTGATGTTGATATTCACAAGTTAAAACAAGTGATACAAAAACAGCATATGTTTTTACCAGACAAAATAAGTTTGACTCAATATGTGGAATTATTCTTAAGTAATAAAACACTCAAATCTGGATCTCATGTTAATTCTAATTTAATATTGGCACATAAAATATCTGACTATTTTCATAATACTTACATTTTAAGTACTAATTTAGCTGGACATTGGATTCTGATTATACAACTTATGAAAGATTCTAAAGGTATTTTTGAAAAAGATTGGGGAGAGGGATATATAACTGATCATATGTTTATTAATTTGAAAGTTTTCTTCAATGCTTATAAGACCTATCTCTTGTGTTTTCATAAAGGTTATGGCAAAGCAAAGCTGGAGTGTGATATGAACACTTCAGATCTTCTATGTGTATTGGAATTAATAGACAGTAGTTATTGGAAGTCTATGTCTAAGGTATTTTTAGAACAAAAAGTTATCAAATACATTCTTAGCCAAGATGCAAGTTTACATAGAGTAAAAGGATGTCATAGCTTCAAATTATGGTTTCTTAAACGTCTTAATGTAGCAGAATTCACAGTTTGCCCTTGGGTTGTTAACATAGATTATCATCCAACACATATGAAAGCAATATTAACTTATATAGATCTTGTTAGAATGGGATTGATAAATATAGATAGAATACACATTAAAAATAAACACAAATTCAATGATGAATTTTATACTTCTAATCTCTTCTACATTAATTATAACTTCTCAGATAATACTCATCTATTAACTAAACATATAAGGATTGCTAATTCTGAATTAGAAAATAATTACAACAAATTATATCATCCTACACCAGAAACACTAGAGAATATACTAGCCAATCCGATTAAAAGTAATGACAAAAAGACACTGAATGACTATTGTATAGGTAAAAATGTTGACTCAATAATGTTACCATTGTTATCTAATAAGAAGCTTATTAAATCGTCTGCAATGATTAGAACCAATTACAGCAAACAAGATTTGTATAATTTATTCCCTATGGTTGTGATTGATAGAATTATAGATCATTCAGGCAATACAGCCAAATCCAACCAACTTTACACTACTACTTCCCACCAAATATCCTTAGTGCACAATAGCACATCACTTTACTGCATGCTTCCTTGGCATCATATTAATAGATTCAATTTTGTATTTAGTTCTACAGGTTGTAAAATTAGTATAGAGTATATTTTAAAAGATCTTAAAATTAAAGATCCCAATTGTATAGCATTCATAGGTGAAGGAGCAGGGAATTTATTATTGCGTACAGTAGTGGAACTTCATCCTGACATAAGATATATTTACAGAAGTCTGAAAGATTGCAATGATCATAGTTTACCTATTGAGTTTTTAAGGCTGTACAATGGACATATCAACATTGATTATGGTGAAAATTTGACCATTCCTGCTACAGATGCAACCAACAACATTCATTGGTCTTATTTACATATAAAGTTTGCTGAACCTATCAGTCTTTTTGTCTGTGATGCCGAATTGTCTGTAACAGTCAACTGGAGTAAAATTATAATAGAATGGAGCAAGCATGTAAGAAAGTGCAAGTACTGTTCCTCAGTTAATAAATGTATGTTAATAGTAAAATATCATGCTCAAGATGATATTGATTTCAAATTAGACAATATAACTATATTAAAAACTTATGTATGCTTAGGCAGTAAGTTAAAGGGATCGGAGGTTTACTTAGTCCTTACAATAGGTCCTGCGAATATATTCCCAGTATTTAATGTAGTACAAAATGCTAAATTGATACTATCAAGAACCAAAAATTTCATCATGCCTAAGAAAGCTGATAAAGAGTCTATTGATGCAAATATTAAAAGTTTGATACCCTTTCTTTGTTACCCTATAACAAAAAAAGGAATTAATACTGCATTGTCAAAACTAAAGAGTGTTGTTAGTGGAGATATACTATCATATTCTATAGCTGGACGTAATGAAGTTTTCAGCAATAAACTTATAAATCATAAGCATATGAACATCTTAAAATGGTTCAATCATGTTTTAAATTTCAGATCAACAGAACTAAACTATAACCATTTATATATGGTAGAATCTACATATCCTTACCTAAGTGAATTGTTAAACAGCTTGACAACCAATGAACTTAAAAAACTGATTAAAATCACAGGTAGTCTGTTATACAACTTTCATAATGAATAATGAATAAAGATCTTATAATAAAAATTCCCATAGCTATACACTAACACTGTATTCAATTATAGTTATTAAAAATTAAAAATCATATAATTTTTTAAATAACTTTTAGTGAACTAATCCTAAAGTTATCATTTTAATCTTGGAGGAATAAATTTAAACCCTAATCTAATTGGTTTATATGTGTATTAACTAAATTACGAGATATTAGTTTTTGACACTTTTTTTCTCGT
Таким образом, материалы, информация и способы, описанные в этом изобретении, обеспечивают множество аттенуированных штаммов с фенотипами с дифференцированной аттенуацией, и обеспечивают рекомендации по выбору подходящих штаммов-кандидатов для вакцин на основе клинических тестов. Следующие примеры предоставлены с целью иллюстрации, а не ограничения.
ПРИМЕРЫ
1.Рекомбинантные штаммы РСВ, приведенные в качестве примеров в этом изобретении, были получены из рекомбинантной версии штамма A2 дикого типа, которая называется D46 (Collins, et al. 1995. Proc Natl Acad Sci USA 92:11563-11567). Полная нуклеотидная последовательность D46 показана в виде SEQ ID NO: 1. Вирус РСВ MEDI/ΔM2-2 не является производным D46.
2. Далее в примерах, когда название вируса включает термин «LID» или «6120», это указывает на то, что его остов содержит мутацию «6120», показанную на ФИГ. 3.
3. Вирусы в настоящем описании названы путем перечисления комбинации мутаций, присутствующих в них. Использование символа «/» в названии вируса (такого как в РСВ D46/cp/ΔM2-2, который обозначает РСВ D46, содержащий мутации cp и ΔM2-2) не имеет никакого значения, кроме присутствия для упрощения чтения имени, в частности, если оно присутствует в тексте. Таким образом, РСВ D46/cp/ΔM2-2 означает то же самое, что и РСВ D46cpΔM2-2. Также, РСВ D46/cp/ΔM2-2 означает то же самое, что и РСВ D46cpΔM2-2 или РСВ D46 cpΔM2-2, и т.д. Также, название вируса, как правило, начинается с РСВ, так как в РСВ D46/cp/ΔM2-2.
4. Как указано ранее, мутация «ΔM2-2» относится к делеции в 241 нуклеотид вместе с тремя точечными мутациями, как показано на ФИГ. 1. Другие мутации, которые подавляют экспрессию M2-2 ORF, обозначены другими именами, например, ΔM2-2-AclI и ΔM2-2-HindIII, показанными на ФИГ. 10. Присутствие «ΔM2-2» в названиях вирусов в этом изобретении указывает на присутствие мутации «ΔM2-2».
5. Величину репликации вируса in vivo применяют в качестве указания на аттенуацию вируса: конкретно, сниженную репликацию in vivo применяют как индикатор повышенной аттенуации, и наоборот. Это отражает общее наблюдение о том, что повышенная репликация РСВ связана с повышенной заболеваемостью, как и для заражения wt РСВ (например, El Saleeby, et al. 2011. J Infect Dis 204:996-1002; DeVincenzo, et al. 2010. Am J Respir Crit Care Med 182:1305-1314) и для аттенуированных кандидатов РСВ в клинических исследованиях (например, Karron, et al. 1997. J Infect Dis 176:1428-1436; Karron, et al. 2005. J Infect Dis 191:1093-1104). Эти термины применяют для описательных целей, а не для ограничительных.
Пример 1
Этот пример иллюстрирует дизайн и конструкцию новых вариантов РСВ, несущих делецию в M2-2 ORF (ΔM2-2), по отдельности и в комбинации с дополнительными мутациями.
Типичные вирусы конструировали и оценивали до клинического исследования. Один типичный вирус из этой панели, и второй вирус, мутантный по M2-2 ORF, из другого источника, оценивали в фазе 1 клинического исследования у серонегатичных младенцев и маленьких детей, которые составляли первичную педатрическую целевую группу для вакцины с РСВ. Этот пример предлагает новые штаммы для вакцины вместе с клиническими критериями для репрезентативных примеров в наиболее подходящей популяции людей.
Вирус РСВ rA2-K5. Штамм РСВ под названием РСВ rA2-K5 был сконструирован ранее (из родительского wt D46 вируса, полученного из кДНК), в котором экспрессия M2-2 ORF была подавлена за счет комбинации трех типов мутаций: (i) введения сдвига рамки в середине M2-2 ORF, (ii) изменения трех потенциальных старт-кодонов трансляции ATG M2-2 ORF (см. ФИГ. 1A для диаграммы генома РСВ и перекрывания M2-1 и M2-2 ORF) на кодоны ACG, и (iii) введения стоп-кодонов во все три регистра последовательности M2-2 вскоре после конца M2-1 ORF (Bermingham и Collins. 1999. Proc Natl Acad Sci USA 96:11259-11264). Этот вирус rA2-K5 (который не показан в настоящем изобретении) оценивали по репликации в дыхательных путях серонегативных шимпанзе, и показали, что он был ограничен, по меньшей мере в 2800 раз в верхних дыхательных путях и не выявлялся в нижних дыхательных путях (представляя снижение, по меньшей мере в 55000 раз) (Teng, et al. 2000. J Virol 74:9317-9321).
Создание РСВ D46/ΔM2-2 и РСВ LID/ΔM2-2. Конструировали дополнительный рекомбинантный вирус, в котором большая часть M2-2 ORF была удалена. кДНК wt D46 была модифицирована таким образом, что каждый из трех старт-кодонов ATG для M2-2 ORF был заменен на ACG, и нуклеотиды 8188-8428 были удалены (полная делеция 241 нуклеотида), с удалением большей части M2-2 ORF (фиг. 1). Таким образом, с мутацией всех известных сайтов начала трансляции ATG и удаленной большей частью ORF, экспрессия полного белка M2-2 белок не должна быть возможной, и экспрессия любых укороченных фрагментов M2-2 должна быть незначительной или отсутствовать. Генетическая карта полученного вируса РСВ D46/ΔM2-2 вирус показана на ФИГ. 2.
Конструировали дополнительные мутанты ΔM2-2, которые могли бы иметь диапазон аттенуирующих фенотипов. Поскольку вирус-прототип РСВ rA2-K5 был высоко аттенуирован у шимпанзе, как указано выше (Teng, et al. 2000. J Virol 74:9317-9321), было вероятно, что вирус, в котором не экспрессируется M2-2, может быть сверхаттенуированным. С другой стороны, также было вероятно, что он может быть недоаттенуированным, в частности у серонегатичных младенцев и маленьких детей. Таким образом, конструировали дополнительные вирусные варианты для выявления производных с повышенной, а также сниженной репликацией.
Не существует установленного способа увеличения репликации штамма РСВ, и, в частности, мутанта ΔM2-2. Ранее сообщалось, что перемещение генов G и F с 6-го и 7-го места в генном порядке (это было сделано в вирусе, в котором SH ген был удален, и таким образом G и F были 6-м и 7-м генами, а не на своих нативных позициях 7-ой и 8-ой) до 1-го и 2-го места, соответственно, приводили к 10-кратному увеличению репликации in vitro, хотя не было статистически значимого увеличения репликации у мышей (Krempl, et al., 2002). J Virol 76: 11931-11942) или АЗМ. Одно из ограничений заключается в том, что общепринятые доклинические анализы для оценки репликации и аттенуации РСВ (например, репликация в клеточных линиях, грызунах, и не являющихся человеком приматах, кроме шимпанзе) могут быть относительно полупермиссивными и нечувствительными, что затрудняет демонстрацию статистически значимых изменений в эффективности репликации, и, таким образом, любое изменение в репликации кажется примечательным, даже если оно не обнаруживается в каждом анализе. Таким образом, было предпринята попытка модифицировать вирус ΔM2-2/ΔSH для перемещения генов G и F гены в положения, более проксимальные к промотору. (Делеция ΔSH была включена в исходное исследование в остов дикого типа [Krempl, et al., 2002. J Virol 76: 11931-11942], чтобы избежать нестабильности в этой последовательности во время амплификации плазмиды в бактериях и рассматривалась как случайная, а также делецию ΔSH применяли с остовом ΔM2-2). Оценивали несколько перестановок, таких как те, в которых гены G и F размещали как первый и второй гены, соответственно, или как второй и первый гены, соответственно. Однако эти модификации уменьшали вирусную репликацию в 100-1000 раз, указывая, что эти конкретные изменения плохо переносятся инфекционным вирусом. Может быть, увеличение экспрессии G и F, которое, как известно, связано с движением их генов в области, проксимально близкие к промотору, (Krempl, et al., 2002. J Virol 76: 11931-11942), в сочетании с увеличением экспрессии белка, связанной с мутацией ΔМ2-2, не переносится РСВ, по крайней мере, в этом остове ΔM2-2/ΔSH.
Дополнительно, известно, что изменения в длине генома могут влиять на эффективность репликации. Конкретно, показано, что увеличение длины генома парамиксовируса может снижать эффективность его репликации. Например, увеличение длины генома РСВ на 140 или 160 нуклеотидов, когда это не нарушает экспрессию гена, приводит к 5-25-кратному снижению репликации у мышей (Bukreyev, Murphy, Collins. 2000. J Virol 74:11017-11026). В другом исследовании с родственным вирусом, а именно вирусом парагриппа человека типа 3 (PIV3), увеличение длины генома или путем добавления дополнительных генов или путем увеличения длины генома за счет вставок нетранслируемых областей (которые, таким образом, не меняют число генов) сохраняет эффективную репликацию in vitro и является аттенуирующим у хомяков (Skiadopoulos, et al. 2000. Virology 272:225-234). Предполагают, что аттенуация, связанная с увеличенной длиной генома, происходит из-за повышенной репликативной нагрузки для более длинного генома. Наблюдение, что увеличение длины генома снижает эффективность репликации, приводит к обратной идее, чтоуменьшение длины генома может повысить эффективность репликации. С этой целью, вирус РСВ D46/ΔM2-2 модифицировали для внесения мутации под названием «6120», приводящей к вирусу под названием РСВ LID/ΔM2-2 (диаграмма генома показана на ФИГ. 2, диаграмма снизу). В этом документе, «LID» в названии вируса указывает на присутствие мутации 6120.
Мутация «6120» (фиг. 3) включает делецию из 112 нуклеотидов в 3'-нетранслируемой области гена SH и введение пяти трансляционно-молчащих точечных мутаций в последние три кодона и кодон терминации гена SH (Bukreyev, et al. 2001. J Virol 75:12128-12140). Основной целью изначального дизайна этой мутации была стабилизация антигеномной кДНК в бактериях, таким образом, что бы с ней можно было легко манипулировать и легко получать, что фактически имело место. Ранее было обнаружено, что в wt РСВ эта мутация приводит к 5-кратному увеличению эффективности репликации in vitro (Bukreyev, et al. 2001. J Virol 75:12128-12140), хотя, по-видимому не повышает эффективность репликации у мышей. Когда РСВ LID/ΔM2-2 оценивали на возможность повышения репликации in vitro, связанной с мутацией 6120, в некоторых экспериментах, но не во всех, наблюдали умеренное увеличение эффективности роста.
Включение дополнительных мутаций в РСВ D46/ΔM2-2 и РСВ LID/ΔM2-2. Конструировали серии дополнительных производных вирусов РСВ D46/ΔM2-2 и РСВ LID/ΔM2-2, в которых одна или более дополнительных мутаций были различным образом вставлены в один или оба вируса, с целью получения спектра дополнительно аттенуированных вирусов.
Примеры производных РСВ D46/ΔM2-2 показаны на ФИГ. 4. Например, производное вируса РСВ D46/cp/ΔM2-2 (фиг. 4, верхняя диаграмма) сочетает мутацию ΔМ2-2 (фиг. 1) с мутациями «cp», которые представляют собой серию из пяти замен аминокислот в трех белках (N (V267I), F (E218A и T523I), и L (C319Y и H1690Y)), что совместно (их силами) приводит приблизительно к 10-кратному снижению репликации у сегонегативных шимпанзе и и уменьшению заболевания (Whitehead, et al. 1998. J Virol 72:4467-4471). Наличие фенотипических данных у шимпанзе примечательно тем, что это экспериментальное животное приближается к людям по его пермиссивности по отношению к репликации РСВ и к заболеваемости РСВ. Следует отметить, что конструкция D46/cp/ΔM2-2 имела единственное случайное изменение нуклеотида в остове D46, на уровне ДНК: в частности, существовала молчащая замена нуклеотида G3878A, присутствующая в M ORF.
Другое производное, вирус РСВ D46/ΔM2-2/1030s (фиг. 4, вторая диаграмма сверху), содержало мутацию ΔМ2-2 в комбинации с генетически стабилизированной мутацией 1030 («1030s»), которая состоит из 1321K(AAA)/S1313(TCA) (Luongo, et al. 2012. J Virol 86:10792-10804). Мутация 1030s приводит в среднем к 0,6 и 1,5 log10 снижению репликации РСВ в верхних и нижних дыхательных путях, соответственно, у мышей. Ее также оценивали у серонегативных шимпанзе, но только в комбинации с рядом дополнительных аттенуирующих мутаций (Luongo, et al. 2012. J Virol 86:10792-10804).
Другое производное, вирус РСВ D46/cp/ΔM2-2/HEK (фиг. 4, диаграмма снизу), сочетает мутации cp и ΔМ2-2 с мутациями «HEK». Мутации HEK состоят из двух замен аминокислот в белке F РСВ, K66E и Q101P, которые соответствуют на аминокислотном уровне последовательности раннего пассажа того же штамма (A2) под названием HEK-7, который получен путем семикратного пересевания клинического изолята исходного штамма A2 на клетках эмбриональной почки человека (HEK) (Connors, et al. 1995. Virology 208:478-484; Whitehead, et al. 1998. J Virol 72:4467-4471), и который, как считают, наиболее близок (и вероятно будет идентичен) клиническому изоляту исходного штамма A2 (Liang, et al. 2014. J Virol 88:4237-4250; Liang, et al. 2015. J Virol 89:9499-9510). Ранее было показано, что мутации HEK стабилизируют тример белка F и обеспечивают фенотип с низким уровнем слияния, который, как полагают, близок к фенотипу исходного клинического изолята (Liang, et al. 2014. J Virol 88:4237-4250; Liang, et al. 2015. J Virol 89:9499-9510). В дополнение к вероятно встречающимся в клиническом изоляте исходного штамма А2, замены HEK найдены почти во все клинических изолятах подгруппы А РСВ, присутствующих в GenBank (Liang, et al. 2015. J Virol 89:9499-9510). Таким образом, мутации HEK могут обеспечить более аутентичную и иммуногенную форму белка F РСВ, возможно обогащенную высоко иммуногенной конформацией «до слияния» (McLellan et al., Science 2013 340(6136):1113-7; Science 2013 342(6158):592-8.). Таким образом, вместо того, чтобы непосредственно быть ассоциированными с аттенуацией, мутации HEK обеспечивают версию белка F, которая более точно отражает клинический изолят исходного штамма А2, а также клинические изоляты других штаммов РСВ.
Примеры производных РСВ LID/ΔM2-2 показаны на ФИГ. 5. Одно из производных, вирус РСВ LID/cp/ΔM2-2 (фиг. 5, диаграмма сверху), сочетает мутации ΔM2-2 и cp. Другое, вирус РСВ LID/ΔM2-2/1030s (фиг. 5, вторая диаграмма сверху), сочетает мутации ΔM2-2 и 1030s. Еще одно, вирус РСВ ΔSH/ΔM2-2 (фиг. 5, третья диаграмма сверху), сочетает мутацию ΔМ2-2 с делецией гена SH (см. ФИГ. 6 для деталей конструкции делеции SH). Ранее было показано, что делеция гена SH приводит к 40-кратному снижению репликации РСВ у серонегативных шимпанзе, и уменьшению заболевания (Whitehead, et al. 1999. J Virol 73:3438-3442). Другое производное, вирус РСВ cp/ΔSH/ΔM2-2 (фиг. 5, диаграмма снизу), сочетает мутации ΔM2-2 и cp с делецией гена SH. Следует заметить, что вирусы, в которых удален полностью ген SH, не обозначают как «LID», поскольку делеция гена SH удаляет мутацию 6120.
Все мутанты на фиг. 2, 4, и 5 легко восстанавливаются путем обратной генетики с использованием стандартных способов. Однако, в отличие от ожиданий, основанных на предыдущей работе (например, Bukreyev et al., J Virol 1997 71:8973-8982; Whitehead et al. J Virol 73:3438-3442 1999), вирусы, которые содержали мутацию ΔSH в контексте с мутацией ΔМ2-2, реплицировались приблизительно в 10 раз менее эффективно, чем другие мутанты ΔM2-2. Это открытие иллюстрирует, как могут возникнуть непредвиденные, но важные эффекты, когда вирусы уже фактически сделаны и оценены.
Что касается аттенуирующих мутаций, указанных на фиг. 4 и 5, предшествующие исследования мутаций cp, ΔSH, 1030 (родительская для 1030s), и ΔМ2-2 на шимпанзе указывают, что их порядок повышения аттенуации следующий: cp≈ΔSH<1030s<ΔM2-2 (Whitehead, et al. 1999. J Virol 73:3438-3442; Whitehead, et al. 1999. J Virol 73:871-877; Teng, et al. 2000. J Virol 74:9317-9321). Таким образом, мутации cp, ΔSH, и 1030s, при объединении по отдельности или в комбинации с мутацией ΔМ2-2, обеспечивают диапазон повышающихся уровней добавленной аттенуации. Их можно оценивать в клиническом исследовании в подходящей целевой популяции для вакцины (серонегативные младенцы и маленькие дети).
Пример 2
Этот пример описывает доклиническую оценку вирусов LID и D46 ΔM2-2.
Типичные вирусы по изобретению оценивали по репликации в дыхательных путях мышей BALB/c, а именно: РСВ D46 wt, РСВ LID/ΔM2-2, РСВ ΔSH/ΔM2-2, РСВ LID/ΔM2-2/1030s, и РСВ cp/ΔSH/ΔM2-2 (фиг. 7). Животных заражали интраназально 5,8 log10 указанного вируса и умерщвляли на 4-5 сутки после заражения, собирали носовые ходы и легкие, гомогенизировали, и оценивали путем ОТ-кПЦР, которая обеспечивает более чувствительную детекцию, чем оценка инфекционных частиц, и, таким образом, подходит для полупермиссивных экспериментальных животных. Оценка показала, что все вирусы, содержащие аттенуирующие мутации, были более ограничены, чем вирус wt D46. Таким образом, дополнительное добавление других аттенуирующих мутаций будет дополнительно аттенуирующим, хотя уровень репликации у этой полупермиссивной модели грызуна был так ограничен, что подробные сравнения не были возможны (фиг. 7).
Те же самые четыре ΔM2-2-содержащих вируса были исследованы по репликации в дыхательных путях АЗМ, а именно: РСВ LID/ΔM2-2, РСВ ΔSH/ΔM2-2, РСВ LID/ΔM2-2/1030s, и РСВ cp/ΔSH/ΔM2-2 (фиг. 8A и 8B, таблицы 1-3). АЗМ представляет собой более аутентичную модель для репликации РСВ, чем грызуны, из-за своей филогенетической близости и анатомического сходства с природным человеческим хозяином для РСВ. Кроме того, АЗМ поддерживает несколько более высокие уровни репликации РСВ по сравнению с макаками резус и яванскими макаками, и таким образом кажется наиболее подходящей доступной обезьяньей моделью. Однако, АЗМ, тем не менее, являются только полупермиссивными для репликации РСВ, а уровень репликации РСВ у АЗМ по существу меньше, чем у шимпанзе или людей. АЗМ в группах по четыре прививали комбинированными путями (интраназально и интратрахеально) 6 log10 БОЕ на мл на каждый из двух участков (IN и IT). Носоглоточные мазки брали ежедневно на сутки 1-10 и 12, а трахеальные смывы брали на сутки 2, 4, 6, 8, 10, и 12 (Таблицы 1 и 2). Было показано, что все три вируса, которые содержали одну или более дополнительных аттенуирующих мутаций, были более аттенуированными, чем РСВ LID/ΔM2-2. В частности, вирус РСВ ΔM2-2/1030s оказался самым аттенуированным. Все три из этих вирусов индуцировали титры РСВ-нейтрализующих сывороточных антител, которые были приблизительно в 2 раза (РСВ ΔSH/ΔM2-2 и РСВ cpΔSH/ΔM2-2) или в 4 раза (РСВ LID/ΔM2-2/Δ1030s) меньше чем для РСВ LID/ΔM2-2 (Таблица 3), что согласуется с общим ожиданием того, что снижение уровня репликации и полученное снижение антигенной нагрузки могут быть связаны со сниженной иммуногенностью, и, таким образом, следует позаботиться об идентификации кандидата для вакцины РСВ, который бы хорошо переносился, но был достаточно иммуногенным.
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 6 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное).
bКомбинированные мазки с носоглотки помещали в 2 мл среды L-15 с сахарозно-фосфатным буфером в качестве стабилизатора. Титрование вируса проводили на клетках Vero при 37°C. Нижний предел детекции составил 0,7 log10 БОЕ/мл. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,35 применяли для образцов без детектируемого вируса.
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 6 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное). Исследование на АЗМ было одобрено Комитетом по уходу и использованию животных NIAID, NIH.
bНа сутки 2, 4, 6, 8, 10, и 12, проводили трахеальные смывы в 3 мл PBS. Титрование вируса проводили на клетках Vero при 37°C. Нижний предел детекции составил 1,0 log10 БОЕ/мл раствора смыва. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,7 применяли для образцов без детектируемого вируса.
aАЗМ были привиты i.n. и i.t. 6,0 log10 указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное).
bПолучали сыворотку на сутки 0, 21, и 28 после прививки. Титры нейтрализующих антител определяли в анализе нейтрализации с 60% уменьшением бляшек. Нижний предел детекции составил 3,3 (1:10).
Еще один эксперимент проводили на АЗМ для сравнения РСВ LID/ΔM2-2 с тремя следующими вирусами: D46/ΔM2-2, РСВ D46/cp/ΔM2-2, D46/cp/ΔM2-2/HEK (таблицы 4-6). Было показано, что вирус РСВ LID/ΔM2-2 реплицировался по существу более эффективно в верхних (Таблица 4) и нижних (Таблица 5) дыхательных путях, чем РСВ D46/ΔM2-2. Важно, что поскольку единственным различием, между этими вирусами была мутация 6120, присутствующая в РСВ LID/ΔM2-2, это показывает, что мутация 6120 приводит к повышению репликации у примата-хозяина. Она, таким образом, обеспечивает способы для постепенного снижения ограничения вируса и аттенуации. Таким образом, остовы LID и D46 обеспечивают существенное различие в эффективности репликации, таким образом, что включение дополнительных мутаций в любой из остовов может обеспечить диапазон фенотипов аттенуации. Вирусы РСВ D46 с дополнительными мутациями, а именно РСВ D46/cp/ΔM2-2/HEK и РСВ D46/cp/ΔM2-2, имеют по существу сниженную репликацию, что свидетельствует об увеличении аттенуации. Все вирусы вызывали существенные титры РСВ-нейтрализующих сывороточных антител (табл. 6). РСВ LID/ΔM2-2 вызывал самые высокие титры; титры, индуцированные РСВ D46/ΔM2-2 и РСВ D46/cp/ΔM2-2, были ниже менее чем в два раза, и титр, вызванный РСВ D46/cp/ΔM2-2/HEK, был почти в 6 раз ниже. Это указывает на то, что включение мутаций, определяющих различные степени аттенуации, дает диапазон фенотипов аттенуации. Это также дает дополнительное указание на то, что сниженная репликация может приводить к снижению иммуногенности.
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 6,7 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 7,0 log10 БОЕ на животное). Исследование на АЗМ было одобрено Комитетом по уходу и использованию животных NIAID, NIH.
bКомбинированные мазки с носоглотки помещали в 2 мл среды L-15 сахарозно-фосфатным буфером в качестве стабилизатора. Титрование вируса проводили на клетках Vero при 32°C. Нижний предел детекции составил 0,7 log10 БОЕ/мл. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,35 применяли для образцов без детектируемого вируса.
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 6,7 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 7,0 log10 БОЕ на животное).
bНа сутки 2, 4, 6, 8, 10, и 12, проводили трахеальные смывы в 3 мл PBS. Титрование вируса проводили на клетках Vero при 32°C. Нижний предел детекции составил 1,0 log10 БОЕ/мл раствора смыва. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,7 применяли для образцов без детектируемого вируса.
aАЗМ были привиты i.n. и i.t. 6,7 log10 указанного вируса в 1 мл прививочного материала на участок (общая доза: 7,0 log10 БОЕ на животное).
bПолучали сыворотку на сутки 0, 21, и 28 после прививки. Титры нейтрализующих антител определяли в анализе нейтрализации с 60% уменьшением бляшек. Нижний предел детекции составил 3,3 (1:10).
Пример 3
Этот пример описывает клиническую оценку вируса РСВ MEDI/ΔM2-2.
Ранее был описан другой мутант РСВ с модифицированной M2-2, обозначенный как РСВ MEDI/ΔM2-2 (Jin, et al. 2000. J Virol 74:74-82). Вирус РСВ MEDI/ΔM2-2 был получен путем введения сайтов для HindIII в нуклеотидных положениях 8197-8201 и 8431-8436 в антигеномную кДНК, с последующим рестрикционным расщеплением HindIII и лигированием для делеции находящихся между сайтами 234 нуклеотидов из M2-2 ORF (Jin, et al. 2000. J Virol 74:74-82). Таким образом, вирус РСВ MEDI/ΔM2-2 не содержит мутацию «ΔM2-2», как описано в настоящем описании (см., например, ФИГ. 1). Это вирус был получен не из D46 (который был разработан из препаратов штаммов A2 РСВ Коллинзом с коллегами (Collins et al. Proc Natl Acad Sci USA 1995 92:11563-11567); вместо этого, он был получен из другого штамма A2 РСВ (Пример 5 далее описывает различие между этими остовами). Был изготовлен материал для клинического испытания (CTM) из РСВ MEDI/ΔM2-2, подходящий для оценки у человека в виде живой аттенуированной интраназальной вакцины. Определяли нуклеотидную последовательность CTM из РСВ MEDI/ΔM2-2 и была обнаружено, что она идентична последовательности исходного клона кДНК, за исключением диморфизмов (смеси двух различных нуклеотидов) в трех положениях: (i) нуклеотид 285 в гене NS1 представляет собой смесь A/G в CTM по сравнению с A в кДНК, что приводит к смеси аминокислот S/G в CTM по сравнению с S в кДНК; (ii) нуклеотид 900 в гене NS2 представляет собой смесь C/T, по сравнению с C в кДНК, не влияя на аминокислотный код; и (iii) нуклеотид 4311 в гене SH представляет собой смесь T/G в CTM по сравнению с T в кДНК, что приводит к смеси аминокислот N/K в CTM, по сравнению с N в кДНК. Обнаружение полиморфизмов в РНК-вирусе является распространенным, и поскольку CTM имел высокий уровень инфекционности (определяемый анализом бляшек), не проявлял диморфизма в фенотипе бляшек и эффективно реплицировался in vitro, эти диморфизмы считались скорее несущественными. Можно проводить оценку последовательности вируса, выделяемого от экспериментальных животных и клинических субъектов, чтобы определить, было ли какое-либо из этих различий в последовательности предпочтительным in vivo, что будет указывать, является ли какое-либо из них значительным. Получали последовательность от одного клинического изолята вакцины и следы диморфизмов все еще присутствовали во всех указанных сайтах, показывая, что ни одно из этих изменений не было значительным.
РСВ MEDI/ΔM2-2 оценивали в виде интраназальной кандидатной вакцины в фазе 1 клинических исследований последовательно у взрослых, детей, сероположительных по РСВ, и младенцев и детей в возрасте 6-24 месяцев, серонегативных по РСВ (ClinicalTrials.gov NCT01459198; Karron, et al. 2015. Science Transl Med 2015 7(312):312ra175). Исследование у взрослых было открытым, а исследования у сероположительных и серотрицательных младенцев и детей были двойными слепыми, рандомизируемыми и плацебо-контролируемыми. Исследование проводили в Центре по исследованию иммунизации (CIR) в университете Джона Хопкинса в школе здравоохранения Блумберга (JHU).
При оценке у взрослых и детей, сероположительных по РСВ, эта экспериментальная вакцина была очень слабо инфекционной, очень слабо иммуногенной, и хорошо переносилась, как и ожидалось для аттенуированного штамма РСВ. У младенцев и маленьких детей, серонегативных по РСВ, 20 человек получили однократную дозу 5,0 log10 БОЕ вакцины РСВ MEDI/ΔM2-2 и 10 человек получили плацебо. Уровни лихорадки и кашля были сходными у вакцинированных и получивших плацебо, в то время как заболевание верхних респираторных путей произошло в два раза чаще у вакцинированных, чем с плацебо (85% по сравнению с 44%), хотя это не было статистически достоверным. Часто выделялись различные случайные респираторные вирусы из обеих групп, которые, по сей видимости, вызывали большую часть заболеваний и которые затрудняли определение переносимости вакцины в этом конкретном исследовании. Частота заражения и заболевания случайными вирусами может непредсказумо варьировать между различными исследованиями, и в этом случае частота была необычно высокой и требует необходимости дальнейших исследований для оценки переносимости РСВ MEDI/ΔM2-2. Выделение вакцинного вируса в назальных мазках было выявлено при помощи анализа бляшкообразования у 12/20 реципиентов и путем ОТ-кПЦР у 17/20 реципиентов. Средний титр выделенного вируса у тех детей, которые выделяли инфекционный вирус, составил 1,5 log10 БОЕ/мл (фиг. 9, левая верхняя сторона). Эти данные повысили вероятность того, что вакцина могла быть слишком ограничена по репликации, поскольку исследования, в основном, были нацелены на >90% выделение вируса на основании анализа бляшкообразования, и средний титр инфекционного выделяемого вируса приблизительно 2,5 log10 БОЕ/мл. Что касается антительного ответа, 19/20 серонегативных детей имели увеличение титров сывороточных антител, нейтрализующих РСВ, ≥4 раза со средним титром 6,6 log2 (1:97). Это дает основания предполагать, что вирус РСВ MEDI/ΔM2-2 был по существу иммуногенным. Однако, наблюдение, что только 12/20 индивидуумов выделили инфекционный вирус, в сочетании с низкими титрами выделенного вакцинного вируса, повышает вероятность того, что вирус РСВ MEDI/ΔM2-2 имел субоптимальную репликацию, и что вирус, мутантный по M2-2, который более эффективно реплицируется, может быть более эффективным. Это актуальная проблема, поскольку иммунная защита поверхностного эпителия дыхательных путей, где РСВ реплицируется и вызывает заболевание, является недостаточной, и, таким образом, желательно, чтобы вакцина РСВ была настолько иммуногенна, насколько это практически возможно.
Пример 4
Этот пример иллюстрирует клиническую оценку РСВ LID/ΔM2-2.
Как указано выше, сравнение РСВ D46/ΔM2-2 и РСВ LID/ΔM2-2 у АЗМ указывает, что наличие мутации «6120» в РСВ LID/ΔM2-2 было ассоциировано с повышенной репликацией (таблицы 4 и 5). Было сделано дополнительное сравнение репликации у АЗМ для РСВ LID/ΔM2-2 по сравнению с CTM из РСВ MEDI/ΔM2-2, параллельно с wt РСВ (таблицы 7-9). Анализ выделения инфекционного вируса в носоглоточных мазках (Таблица 7) или образцах трахеальных смывов (Таблица 8) показал, что оба вируса были более аттенуированными, чем wt РСВ, который оценивали параллельно. Однако не было заметной разницы в выделении вируса, и, таким образом, в репликации, между РСВ LID/ΔM2-2 и РСВ MEDI/ΔM2-2. Все три вируса (РСВ LID/ΔM2-2, РСВ MEDI/ΔM2-2, и wt РСВ) индуцировали сходные титры РСВ-нейтрализующих сывороточных антител (Таблица 9).
Вирус РСВ LID/ΔM2-2 оценивали в клиническом исследовании для того чтобы определить, может ли он реплицироваться более эффективно у людей, чем РСВ MEDI/ΔM2-2 и может ли он быть более иммуногенным. Была произведена партия CTM для РСВ LID/ΔM2-2, подходящая для введения человеку в виде экспериментальной интраназальной вакцины РСВ. Анализ нуклеотидной последовательности показал, что CTM имел такую же последовательность, что и его клон кДНК, что указывало на отсутствие выявляемых посторонних мутаций во время производства. Его эффективность репликации в клетках Vero (которые применяют для производства вакцин) была по существу такой же, что и у РСВ MEDI/ΔM2-2. CTM РСВ LID/ΔM2-2 оценивали параллельно с wt РСВ на репликацию и иммуногенность у АЗМ (таблицы 10-12). Титрование инфекционного вируса с мазком из носоглотки (Таблица 10) и образцов трахеальных смывов (Таблица 11) подтвердило аттенуированный фенотип CTM РСВ LID/ΔM2-2. Тем не менее, титр РСВ-нейтрализующих сывороточных антител, индуцируемый CTM был практически таким же, что и титр, индуцируемый wt РСВ (Таблица 12), указывая на то, что эта экспериментальная вакцина сохранила большую часть иммуногенности своего родителя дикого типа.
CTM РСВ LID/ΔM2-2 оценивали у РСВ-серонегативных младенцев и детей в возрасте 6-24 месяцев в двойном слепом плацебо-контролируемом исследовании, которое проводили в CIR/JHU (ClinicalTrials.gov NCT02040831) и в семи клинических участках Международной группы клинических испытаний по СПИДу у матерей, детей и подростков (IMPAACT, ClinicalTrials.gov NCT02237209). В целом, 20 индивидуумов получили однократную дозу 5,0 log10 БОЕ вакцины, и девять - получили плацебо. Что касается респираторных заболеваний после вакцинации, респираторные заболевания встречались часто и у вакцинированных, и у реципиентов плацебо. Уровни лихорадки, воспаление среднего уха, заболевание верхних дыхательных путей, заболевание нижних дыхательных путей, кашель, или любое респираторное или лихорадочное заболевание были по существу одинаковыми между двумя группами. Дополнительные вирусы, в том числе риновирус, аденовирус, вирус парагриппа, и коронавирус, выявлялись часто и у вакцинированных, и у реципиентов плацебо. У одного вакцинированного развился короткий эпизод мягкого заболевания нижних дыхательных путей (хрипы) на сутки 9, который разрешился на сутки 11 и совпал с выделением вакцинного вируса, детекцией риновируса и энтеровируса в качестве дополнительных агентов. Таким образом, причина этого клинического заболевания остается неясной. Выделение инфекционного вакцинного вируса было получено у 19/20 вакцинированных, со средним пиковым титром 3,4 log10 БОЕ/мл (фиг. 9B).
Таким образом, вирус РСВ LID/ΔM2-2 был более инфекционным, чем РСВ MEDI/ΔM2-2 у человека-хозяина, на основании числа индивидуумов, выделявших инфекционный вирус (19/20 по сравнению с 12/20) и на основании среднего пикового титра (3,4 log10 БОЕ/мл по сравнению с 1,5 log10 БОЕ/мл, что было статистически значимым). Вирус РСВ LID/ΔM2-2 также реплицировался более эффективно, чем предыдущий ведущий кандидат под названием rA2cp248/404/1030ΔSH, который оценивали в предыдущем клиническом исследовании (Karron, et al. 2005. J Infect Dis 191:1093-1104): ряд образцов из этого предыдущего исследования анализировали параллельно с образцами из исследования MEDI/ΔM2-2, и было показано, что rA2cp248/404/1030ΔSH имеет средний пиковый титр 2,5 log10 (фиг. 9A, правая панель). Вирус РСВ LID/ΔM2-2 также индуцировал более высокий титр РСВ-нейтрализующих сывороточных антител (1:137) по сравнению с РСВ MEDI/ΔM2-2 (1:97) и rA2cp248/404/1030ΔSH (1:34), которые анализировали параллельно (Karron, et al. 2015. Science Transl Med 2015 7(312):312ra175).
Вышеописанное клиническое исследование показало, что вирус РСВ LID/ΔM2-2 был более инфекционным и реплицировался более эффективно, чем РСВ MEDI/ΔM2-2 у человека-хозяина. Он также был более иммуногенным. Как отмечалось, различие в эффективности вирусной репликации между этими двумя вирусами не было воспроизводимо показано на клеточных линиях или у АЗМ, и также не была продемонстрирована более высокая иммуногенность для LID/ΔM2-2 по сравнению с MEDI/ΔM2-2 у АЗМ. Таким образом, в отличие от доклинических исследований, РСВ LID/ΔM2-2 обеспечивает более уверенную репликацию, и большую иммуногенность по сравнению с РСВ MEDI/ΔM2-2.
Были разработаны и сконструированы дополнительные производные РСВ LID/ΔM2-2, которые обладают одной или несколькими дополнительными аттенуирующими мутациями. Примеры этих штаммов включают: РСВ LID/cp/ΔM2-2 (см. Пример 8), РСВ LID/ΔSH/ΔM2-2, РСВ LID/cp/ΔSH/ΔM2-2, и РСВ LID/ΔM2-2/1030s. На основании предшествующей оценки мутаций cp, ΔSH, 1030s, и ΔМ2-2 у серонегативных шимпанзе, ожидается, что порядок увеличения аттенуации этих штаммов будет: РСВ LID/cp/ΔM2-2≈РСВ LID/ΔSH/ΔM2-2<РСВ LID/cp/ΔSH/ΔM2-2<РСВ LID/ΔM2-2/1030s. Также предлагаются дополнительные штаммы, такие как штаммы, которые включают мутации, описанные в Примерах 5-8.
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 6,0 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное). Исследование на АЗМ было одобрено Комитетом по уходу и использованию животных NIAID, NIH.
bКомбинированные мазки с носоглотки помещали в 2 мл среды L-15 сахарозно-фосфатным буфером в качестве стабилизатора. Титрование вируса проводили на клетках Vero при 37°C. Нижний предел детекции составил 0,7 log10 БОЕ/мл. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,35 применяли для образцов без детектируемого вируса.
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 6,0 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное).
bНа сутки 2, 4, 6, 8, 10, и 12, проводили трахеальные смывы в 3 мл PBS. Титрование вируса проводили на клетках Vero при 37°C. Нижний предел детекции составил 1,0 log10 БОЕ/мл раствора смыва. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,7 применяли для образцов без детектируемого вируса.
aАЗМ были привиты i.n. и i.t. 6,0 log10 указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное).
bПолучали сыворотку на сутки 0, 21, и 28 после прививки. Титры нейтрализующих антител определяли в анализе нейтрализации с 60% уменьшением бляшек. Нижний предел детекции составил 3,3 (1:10).
wt РСВ
aМартышки были привиты i.n. и i.t. 5,9 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,2 log10 БОЕ/АЗМ).
bТитрование вируса проводили на клетках Vero при 37°C. Нижний предел детекции составил 0,7 log10 БОЕ/мл. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты. Результаты показывают, что РСВ LID ΔM2-2 значительно ограничен в верхних дыхательных путях АЗМ по сравнению с РСВ rA2.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,35 применяли для образцов без детектируемого вируса.
wt РСВ
aМартышки были привиты i.n. и i.t. 5,9 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,2 log10 БОЕ/АЗМ).
bТитрование вируса проводили на клетках Vero при 32°C. Нижний предел детекции составил 1,0 log10 БОЕ/мл раствора смыва. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты. Как и ожидалось, высокочувствительный к температуре вирус РСВ LID ΔM2-2 не реплицировался в нижних дыхательных путях АЗМ (температура тела: 39°C). TL, трахеальный смыв.
cСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,7 применяли для образцов без детектируемого вируса.
wt РСВ
aМартышки были привиты i.n. и i.t. 5,9 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,2 log10 БОЕ/АЗМ).
bНижний предел детекции в анализе 60% уменьшения бляшкообразования составил 3,3 (Log2 от обратного разведения). Образцы ниже предела детекции обозначены как «-».
Пример 5
Этот пример иллюстрирует различия между РСВ LID/ΔM2-2, РСВ D46/ΔM2-2, и РСВ MEDI/ΔM2-2, которые могут влиять на эффективность репликации in vivo.
Как указано выше, вирусы РСВ D46/ΔM2-2 и РСВ LID/ΔM2-2 идентичны по последовательности, за исключением мутации 6120 в гене SH РСВ LID/ΔM2-2, которая удаляет 112 нуклеотидов из 3'-нетранслируемой области и производит молчащие нуклеотидные замены в последних трех кодонах и стоп-кодоне SH ORF (фиг. 3). РСВ LID/ΔM2-2 реплицировался значительно более эффективно, чем РСВ D46/ΔM2-2 в верхних (Таблица 4) и нижних (Таблица 5) дыхательных путях АЗМ. Поскольку эти вирусы в остальном идентичны, это показывает, что мутация 6120 связана с повышенной репликацией у приматов, и эффект был достаточно велик, чтобы однозначно обнаружить его у АЗМ.
Как указано выше, вирусы РСВ LID/ΔM2-2 и РСВ MEDI/ΔM2-2 не различаются в отношении эффективности репликации в клеточной культуре и у АЗМ, хотя последний был значительно более эффективен в репликации у серонегативных младенцев и детей, и был более иммуногенным. Таким образом, эти вирусы также отличаются по эффективности репликации, но это был единственный случай у полностью пермиссивного человека-хозяина. РСВ LID/ΔM2-2 имеет мутацию 6210, а РСВ MEDI/ΔM2-2 - нет. Хотя, в этом случае мутация 6120 не была ассоциирована с повышенной репликацией у полу-пермиссивных АЗМ, она была ассоциирована с повышенной репликацией у серонегативных младенцев и детей, пермиссивного природного хозяина. Эти наблюдения позволяют предположить, что, хотя РСВ LID/ΔM2-2 явно имеет более высокую эффективность репликации, чем РСВ MEDI/ΔM2-2, это может быть несколько снижено некоторым другим различием между вирусами LID и MEDI. Существует два таких различия дополнительно к мутации 6120.
Одно из различий в том, что детали мутаций, которые выключают экспрессию M2-2 ORF, различны между РСВ LID/ΔM2-2 и РСВ MEDI/ΔM2-2. В РСВ LID/ΔM2-2 (и РСВ D46/ΔM2-2), делеция представляет собой 241 нуклеотид в длину и начинается после нуклеотида 8187 и, кроме того, все три старт-кодона трансляции ATG изменены на ACG, так что должен быть очень низкий уровень или отсутствие трансляции любого из пептидов M2-2 (фиг. 1). Напротив, в РСВ MEDI/ΔM2-2, делеционная мутация включает вставку чужеродного участка рестрикции для HindIII, начинающегося в нуклеотиде 8196, включает делецию из 234 нуклеотидов, и будет кодировать пептид из 12 аминокислот, представляющий N-конец наиболее длинной версии белка M2-2 (Jin, et al. 2000. J Virol 74:74-82).
Второе из различий в том, что кДНК РСВ MEDI/ΔM2-2 и РСВ LID/ΔM2-2 различаются по 21 дополнительному положению в нуклеотидной последовательности, рассеянному по двум остовам (Таблица 13). Из них, 6 нуклеотидных различий (включая однонуклеотидную вставку в положении 1099 РСВ LID/ΔM2-2) связаны с участками рестрикции, которые добавляли в процессе конструирования клона кДНК D46 (Collins, et al. 1995. Proc Natl Acad Sci USA 92:11563-11567). Считают, что эти шесть изменений являются фенотипически молчащими, поскольку биологический РСВ дикого типа и рекомбинантный wt D46 РСВ реплицируются со сходной эффективностью у шимпанзе и вызывают аналогичный уровень заболевания (например, Whitehead, et al. 1998. J Virol 72:4467-4471). Эти изменения также присутствуют в ряде кандидатных вакцин, которые в настоящее время оценивают на людях. Таким образом, кажется невероятным, чтобы эти 6 нуклеотидов и связанные с ними участки рестрикции влияли на репликацию, хотя нельзя утверждать это однозначно. Остальные 15 нуклеотидных различий между РСВ MEDIΔM2-2 и РСВ LID/ΔM2-2 представляют собой точечные мутации, которые, как полагают, отражают дополнительные различия, присутствующие в двух различных родительских биологических источниках штамма A2, из которых были получены две независимых системы с обратной генетикой. Нередко можно найти множество нуклеотидных различий между двумя препаратами одного штамма РСВ, которые имели различные истории пересевов. Два из этих пятнадцати нуклеотидных различий приводят к различиям по аминокислотам, одному в белке NS2 (K51R), и одному в белке N белок (T24A) (аминокислотные обозначения даны сначала для LID, а потом для MEDI). Недавнее исследование указывает, что ни одно из этих двух аминокислотных различий не влияет на эффективность репликации in vitro (Lawlor, Schickli, и Tang. 2013. J Gen Virol 94:2627-2635).
V
1Геномное положение пронумеровано относительно WT штамма A2 РСВ (номер доступа GeneBank M74568). Все последовательности являются плюс-цепями.
2ncr, не-кодирующая область
4ig, межгенная область
*Изменения, сконструированные в качестве маркеров в исходном LID в антигеномном клоне кДНК (Collins et al. PNAS 92:11563-7 1995 PMID 8524804). Эти изменения присутствуют в большинстве рекомбинантных кандидатных вакцин РСВ, которые оценивали на людях.
Таким образом, предлагаются новые реагенты и информация, которые указывают, что:
1. РСВ LID/ΔM2-2 реплицируется по существу более эффективно у АЗМ, чем РСВ D46/ΔM2-2. Поскольку эти вирусы отличаются только наличием мутации 6120 в РСВ LID/ΔM2-2, был сделан вывод, что эта мутация приводит к фенотипу с повышенной репликацией у приматов, отличие, которое выявляется даже на полупермиссивной модели АЗМ. Это существенное отличие в эффективности репликации между РСВ LID/ΔM2-2 и РСВ D46/ΔM2-2 обеспечивает два остова, которые значительно отличаются по аттенуации. Таким образом, можно вводить общий набор аттенуирующих мутаций (например, cp, ΔSH, 1030s) в каждый остов и получать широкий спектр аттенуирующих фенотипов, которые напрямую связаны с контрольными данными у людей (т.е., клиническое исследование РСВ LID/ΔM2-2).
2. РСВ LID/ΔM2-2 не реплицировался более эффективно, чем РСВ MEDI/ΔM2-2 у АЗМ, но делал это у более пермиссивного натурального хозяина, а именно, у серонегативных младенцев и детей. Это позволяет предположить, что РСВ LID/ΔM2-2 имеет преимущество в репликации по сравнению с РСВ MEDI/ΔM2-2, но меньшее, чем по сравнению с РСВ D46/ΔM2-2. Таким образом, порядок эффективности репликации у людей для РСВ LID/ΔM2-2 и РСВ MEDI/ΔM2-2 представляет собой РСВ LID/ΔM2-2>РСВ MEDI/ΔM2-2, и данные от АЗМ поддерживают дополнительный вывод, что РСВ D46/ΔM2-2 даже более аттенуирован, давая порядок аттенуации: РСВ LID/ΔM2-2>РСВ MEDI/ΔM2-2>D46/ΔM2-2. Это позволяет предположить, что одно или несколько отличий в остовах РСВ MEDI/ΔM2-2 по сравнению с D46/LID несет ответственность за промежуточный фенотип. Наиболее вероятными кандидатами являются мутации K51R и T24A в NS2 и N, соответственно, несмотря на то, что опубликованные данные, отмеченные сразу же выше, полагают, что они являются фенотипически молчащими (Lawlor, Schickli, и Tang. 2013. J Gen Virol 94:2627-2635). Эти две замены аминокислот считаются наиболее вероятными кандидатами, поскольку аминокислотные замены наиболее вероятно воздействуют на фенотип - через влияние на структуру и функцию белка - чем молчащие нуклеотидные сигналы, не содержащиеся в цис-действующем сигнале. Альтернативной или дополнительной возможностью является то, что играет роль отличие в деталях конструкции мутаций ΔМ2-2. Дополнительной возможностью является то, что играют роль одно или несколько из остальных 19 транскрипционно молчащих нуклеотидных отличий. Эти возможности можно различить с использованием штаммов, выбранных из панели, описанной ниже.
Пример 6
Этот пример иллюстрирует дополнительные конструкции ΔM2-2 в сочетании с признаками из РСВ LID/ΔM2-2, РСВ D46/ΔM2-2 и РСВ MEDI/ΔM2-2.
Дополнительные вирусы на основе ΔM2-2 конструировали с использованием вышеизложенных результатов в качестве руководства для получения различных комбинаций, содержащих одну из нескольких мутаций ΔМ2-2, мутацию 6120, мутации K51R/T24A, и другие случайные различия между остовами MEDI и D46/LID.
Была создана дополнительная делеция M2-на основе сайт-специфического мутагенеза, которая удаляла 234 нуклеотида в M2-2 ORF (нуклеотиды 8202-8435), вводила точечные мутации T8197A и C8199G, которые создавали сайт для AclI, и вводила стоп-кодон в кодон 13 M2-2 ORF (фиг. 10). Эта модификация была сделана для обоих остовов РСВ D46 и РСВ LID и приводила к получению РСВ D46/ΔM2-2-AclI и РСВ LID/ΔM2-2-AclI (фиг. 11). Эти вирусы сочетали остов D46 или LID (т.е., без мутации и с мутацией 6120, соответственно, но в остальном идентичные) с мутацией ΔМ2-2 (ΔM2-2-AclI), что напоминает РСВ MEDI/ΔM2-2, в том числе потенциал для экспрессии пептида, представляющего 12 N-концевых аминокислот M2-2 (фиг. 11).
кДНК РСВ D46/ΔM2-2-AclI и РСВ LID/ΔM2-2-AclI были дополнительно модифицированы включением мутации K51R в белке NS2 и мутации T24A в белке N (фиг. 12A). Таким образом, вводили в остовы D46/ΔM2-2 и LID/ΔM2-2 два самых значительных отличия по сравнению с остовом MEDI.
В качестве дополнительных примеров, мутацию K51R в белке NS2 и мутацию T24A в белке N также вводили в остов D46/ΔM2-2 по отдельности (фиг. 12B, первая и вторая конструкция сверху) и вместе (третья конструкция). Эти мутации также вводили в остов LID/ΔM2-2 (фиг. 12C) по отдельности (четвертая и пятая конструкции) и вместе (нижняя конструкция).
Кроме того, мутацию ΔM2-2-HindIII (описана на ФИГ. 10) вводили в D46 для получения остовов D46/ΔM2-2-HindIII и LID/ΔM2-2-HindIII (фиг. 13, первая и третья конструкции сверху). Дополнительные производные включали дополнительные вставки замен аминокислот K51R и T24A в белках NS2 и N (фиг. 13, вторая и четвертая конструкции сверху).
Пример 7
Этот пример иллюстрирует дополнительные конструкции ΔM2-2 с дополнительными модификациями в генах F и/или G.
РСВ LID/ΔM2-2 (фиг. 2) был модифицирован дополнительными изменениями в генах F и/или G. В основном, эти модификации разрабатывались изначально не для влияния на аттенуацию, а для влияния на другие параметры, такие как эффективность экспрессии антигенов или включение генов из другого штамма. Следует отметить, что эти штаммы используют терминологию «6120», а не «LID» для указания на наличие мутации 6120. Эти конструкции являются следующими:
РСВ 6120/G001BB/FBB/ΔM2-2 (фиг. 14A): содержит кодон-оптимизированный ген G (G001BB) из недавнего (2011 год), низко пассированного клинического изолята A/Maryland/001/11. Эта конструкция также содержит кодон-оптимизированный ген F штамма A2 (FBB). Следует отметить, что доказано, что нативная последовательность G0001 является нестабильной во время клонирования в бактериях. Оптимизация кодонов, приводящая к G001BB, оказала воздействие на придание стабильности.
РСВ 6120/FBB/G001BB/ΔM2-2 (фиг. 14A): содержит кодон-оптимизированный ген F A2 (FBB) и кодон-оптимизированный ген G из недавнего клинического изолята (G001BB), но их порядок в генетической карте был изменен на обратный, с G-F на F-G, для получения повышенной экспрессии белка F, основного нейтрализующего и протективного антигена РСВ.
РСВ 6120/G001BB/F/ΔM2-2 (фиг. 14A): содержит G001BB и нативный ген F A2.
РСВ 6120/G/FBB/ΔM2-2 (фиг. 14A, четвертая конструкция сверху): содержит нативный ген G A2 и кодон-оптимизированный ген F A2 (FBB).
РСВ 6120/G/FBBHEK/ΔM2-2 (фиг. 14B): содержит нативный ген G A2 и кодон-оптимизированный ген F A2 (FBB), который также имеет две мутации HEK, K66E и Q101P.
РСВ 6120/G/FBBcpHEK/ΔM2-2 (фиг. 14C): содержит нативный ген G A2 и кодон-оптимизированный ген F A2 (FBB), который также имеет две мутации HEK, K66E и Q101P, и две мутации cp, содержащиеся в гене F, а именно, E218A и T523I.
РСВ 6120/FBB/G/ΔM2-2 (фиг. 14C): содержит кодон-оптимизированный F A2 (FBB) и нативный ген G A2, но их порядок в генетической карте изменен на обратный, от G-F до F-G.
РСВ 6120/G001BB/F001BB/ΔM2-2 (фиг. 14C): содержит ген G001BB ген и ген F001, которые были кодон-оптимизированы (G001BB, F001BB).
Кратко, гены G и/или F из недавнего клинического изолята под названием A/Maryland/001/11 (который выделяли в 2011 у медицинского работника с существенным респираторным заболеванием) были использованы для того чтобы исследовать, могут ли эти комбинации создать повышенную репликацию и/или иммуногенность. Это также показало бы, что живую вакцину РСВ можно было бы быстро обновлять для содержания поверхностных белков от более поздних штаммов. Использование оптимизации кодонов (BB) производили для повышения экспрессии одного или обоих главных протективных антигенов. Изменения в порядке генов G и F от G-F до F-G было разработано для повышения экспрессии антигенов, и было сделано со знанием того, что перемещение F и G в положения, более проксимальные к промотору на генетической карте, в отношении мутации ΔМ2-2, приведет к вирусам, которые неожиданно покажут сниженный уровень репликации in vitro, как описано в примере 1. Использование мутаций HEK, с наличием или отсутствием двух мутаций cp в F, было сделано для получения более стабильного белка F, который мог бы иметь улучшенную иммуногенность из-за сохранения эпитопов нейтрализации. Это было основано на идее, что метастабильная природа белка F РСВ может вносить вклад в уклонение от иммунной системы путем презентирования денатурированного антигена denatured антиген (Sakurai, et al. 1999. J Virol 73:2956-2962; Collins и Graham, 2008. J Virol 82:2040-2055), и, таким образом, обеспечение более стабильной формы могло бы индуцировать качественно превосходящий иммунный ответ.
Каждый из вирусов, показанный на Фиг. 14A и 14B был легко восстановлен из кДНК. Пассаж P1 представлял собой первый пассаж после трансфекции, и его проводили без титрования посевной среды (слепой пассаж). Полученные урожаи P1 конструкций на Фиг. 14A и 14B сравнивали с wt РСВ, содержащим мутацию 6120(wt LID), и с РСВ LID/ΔM2-2, который являлся родительским штаммом для конструкций (фиг. 15). Это показывает, что титры P1 всех вирусов были сопоставимы благоприятно с двумя контролями, с единственным исключением, что титр P1 РСВ 6120/G001BB/F001BB/ΔM2-2 был снижен приблизительно до 3,0 log10. Однако, этот титр во время пассажа P2 вернулся к уровням, соспоставимым с другими конструкциями (фиг. 15). В основном, это показало, что все модификации хорошо переносились, в том числе новые гены, изменения порядка генов, оптимизация кодонов, и введение мутаций HEK и/или cp.
Пример 8
Этот пример иллюстрирует оценку дополнительных конструкций РСВ ΔM2-2.
Как описано выше, было обнаружено, что РСВ D46/cp/ΔM2-2 (фиг. 4, верхний геном), реплицируется эффективно в клетках Vero, необходимых для производства вакцин, и является высоко аттенуированным и все еще высоко иммуногенным у АЗМ (таблицы 4-6). Таким образом, получали посевной вирус для вакцины и использовали для производства материала для клинических испытаний (CTM) из D46/cp/ΔM2-2. Как уже отмечалось, эта конструкция имела единичную случайную нуклеотидную замену в остове D46, на уровне ДНК: конкретно, G3878A, которая присутствовала в M ORF и была молчащей на аминокислотном уровне. Автоматический анализ последовательности показал, что последовательность CTM была идентична последовательности кДНК. Анализ репликации и иммуногенности этого CTM у АЗМ подтвердил, что он высоко аттенуирован (таблицы 14 и 15) все еще высоко иммуногенный (Таблица 16). Эту кандидатную вакцину оценивали в двойном слепом плацебо-контролируемом исследовании у 15 детей, сероположительных по РСВ, в возрасте от 12 до 59 месяцев (10 реципиентов вакцины, 5 реципиентов плацебо), проводимом у CIR/JHU (ClinicalTrials.gov identifier NCT02601612). После интраназального введения в дозе 106 БОЕ, выделение вакцины не детектировалось, и D46/cp/ΔM2-2 был слабо иммуногенным у сероположительных детей. Это указывает на то, что вакцина сильно рестриктирована и аттенуирована, предсказывая, что она будет безопасной и подходит для оценки у серонегативных младенцев и детей. Оценка у РСВ-серонегативных младенцев и детей в возрасте 6-24 месяцев продолжается в настоящее время. Она обеспечит информацию о перспективной кандидатной вакцине, указав, подходит ли она для расширенных исследований. Эта информация также послужит дополнительным контролем, связывающим доклинические и клинические исследования.
Кроме того, конструировали аналог этого вируса LID, LID/cp/ΔM2-2 (фиг. 5, геном сверху). Было обнаружено, что он реплицируется эффективно в клетках Vero, и анализ репликации и иммуногенности этого вируса у АЗМ показал, что он высоко аттенуирован (таблицы 17 и 18) и все еще высоко иммуногенный (Таблица 19). Это показывает, что добавление мутаций cp к LID/ΔM2-2, который был неполностью аттенуирован у серонегативных младенцев и детей (фиг. 9B), для получения LID/cp/ΔM2-2, привело к повышенной аттенуации у АЗМ (т.е., сравнивая данные для LID/ΔM2-2, таблицы 1 и 2, 4 и 5, 7 и 8, и 10 и 11, с данными для LID/cp/ΔM2-2, таблицы 17 и 18). Это позволяет предположить, что LID/cp/ΔM2-2 должен давать повышенную аттенуацию у серонегативных людей, хотя для подтверждения этого необходимы клинические испытания, чтобы определить величину повышения аттенуации и подтвердить безопасность.
Затем был произведен материал для клинического испытания для LID/cp/ΔM2-2 с использованием антигеномной кДНК, чья последовательность показана на SEQ ID NO: 17. Последовательность материала для клинического испытания (LIDcpΔM2-2, Lot РСВ#009B) подтверждали анализом консенсусной последовательности, что она соответствует последовательности кДНК, из которой был получен рекомбинантный вирус за исключением точечной мутации C/T в нуклеотидном положении 9972 (следует отметить, что все последовательности описаны в виде плюс-цепи или антигеномной цепи). Эта мутация была молчащей на аминокислотном уровне и также присутствовала в посевном вирусе, который использовали для получения LIDcpΔM2-2, Lot РСВ#009B. Случайные мутации могут возникать во время пассажа РСВ, как и для большинства РНК-вирусов, из-за высокого уровня ошибок РНК-зависимой РНК-полимеразы. Когда такие замены не влияют на известный цис-действующий сигнал или не меняют кодирование аминокислоты, и если они не имеют измеримого влияния на репликацию in vitro и размер бляшек вируса, они считаются биологически несущественными. Молчащая точечная мутация C9972T в материале для клинического испытания LIDcpΔM2-2, вероятно, несущественна, но будет контролироваться.
Вакцину с вирусом LID/cp/ΔM2-2 оценивают у серонегативных детей в возрасте 6-24 месяцев в двойном слепом плацебо-контролируемом исследовании. По меньшей мере 5 индивидуумов получили вакцину, без признаков реактогенности во время периода, когда ожидается выделение вируса, позволяя предположить, что эта вакцина хорошо переносится.
Как уже отмечалось, получали партию вируса LID/ΔM2-2/1030s не для клинических экспериментов (фиг. 5, второй геном сверху), и было обнаружено, что он реплицируется эффективно в клетках Vero. Его оценивали у АЗМ и было показано, что он высоко аттенуированый (таблицы 1 и 2) и высоко иммуногенный (Таблица 3).
Получали препарат CTM для LID/ΔM2-2/1030s с использованием антигеномной кДНК, показанной в SEQ ID NO: 16, и автоматический анализ последовательности показал, что последовательность материала для клинического испытания(CTM) была идентична последовательности кДНК. Эту вакцину оценивали у серонегативных детей в возрасте 6-24 месяцев в двойном слепом плацебо-контролируемом исследовании. Всего участвовало 33 индивидуума, с предполагаемым соотношением вакцина:плацебо 2:1. Назальные смывы от 30 участников оценивали по анализу бляшкообразования (культивирование вируса), а также путем количественной ОТ-ПЦР (кПЦР) для выделения вакцинного вируса LID/ΔM2-2/1030s, как измерение аттенуации. Было показано, что 17 индивидуумов имели явное выделение вакцинного вируса (предполагается, что всего 20 индивидуумов получили вакцину). 15/30 индивидуумов в исследовании LID/ΔM2-2/1030s были положительными в анализе бляшкообразования, и эти 15 плюс два дополнительных индивидуума были положительными по кПЦР, которая представляет собой более чувствительный анализ. Предполагается, что индивидуумы, которые выделяют вирус в течение 14-18 суток после введения вакцины, будут обнаружены как реципиенты вакцины, и таким образом эти данные можно использовать в качестве предполагаемой оценки выделения вакцинного вируса и, таким образом, аттенуации. Для сравнения, аналогичный анализ выделения вируса РСВ LID/ΔM2-2 в сопоставимой когорте серонегативных детей в возрасте 6-24 месяцев, ClinicalTrials.gov NCT02040831, обнаружил выделение вируса у 19 из 20 реципиентов вакцины посредством анализа бляшкообразования и кОТ-ПЦР. В исследовании LID/ΔM2-2/1030s, предполагаемые средние пиковые титры были: 5,1 log10 копий/мл путем ПЦР, и 2,9 log10 БОЕ/мл путем культивирования, в то время как для исследования LID/ΔM2-2, средние пиковые титры были 5,9 log10 копий/мл путем ПЦР, и 3,4 log10 БОЕ/мл путем культивирования. Таким образом, вирус LID/ΔM2-2/1030s, по-видимому, более аттенуирован, чем вирус LID/ΔM2-2 на основании уровня инфекции и титров выделяемого вируса. С вирусом LID/ΔM2-2/1030s, три самых высоких индивидуальных пиковых титра были 4,7, 4,5, и 4,1 log10 БОЕ/мл, по сравнению с 5,4, 5,3, и 5,1 log10 БОЕ/мл для LID/ΔM2-2. Кроме того, для вируса LID/ΔM2-2/1030s, шесть индивидуумов выделили инфекционный вирус только в одни сутки, по сравнению с двумя сутками для вируса LID/ΔM2-2. Таким образом, по каждому из этих измерений, вставка мутации 1030s в LID/ΔM2-2 обеспечила измеряемое, постоянное снижение выделения вируса у серонегативных детей, которые являются мишенью вакцины.
Вирус РСВ LID/ΔM2-2/1030s оценивали на температурно-чувствительный фенотип, поскольку мутация 1030s представляет собой мутацию температурной чувствительности (Таблица 20). Этот анализ показал, что РСВ LID/ΔM2-2/1030s имеет выключающую температуру (TSH) 40°C и температуру малых бляшек (TSP) 38°C, в то время как РСВ дикого типа, LID/ΔM2-2, MEDI/ΔM2-2, LID/cp/ΔM2-2, и D46/cp/ΔM2-2 имеют TSH и TSP >40°C. Таким образом, РСВ LID/ΔM2-2/1030s, но не другие вирусы дикого типа и вирусы на основе ΔM2-2, имеют температурно-чувствительный фенотип. Два остальных известных температурно-чувствительных вируса, РСВ ΔNS2/Δ1313/I1314L и РСВ cps2, были включены в качестве положительных контролей, и подтвердили, что анализ был точным. Таким образом, введение мутации 1030s в РСВ LID/ΔM2-2 создает температурно-чувствительный фенотип. Это важно, поскольку, полагают, что температурно-чувствительный фенотип предпочтительно ограничивает репликацию в более теплых нижних дыхательных путях по сравнению с более прохладными верхними дыхательными путями, и, таким образом, придает дополнительную безопасность против реактогенности. Полагают, что это так, даже если TSH и/или TSP выше, чем физиологическая температура.
Таким образом, это обеспечивает спектр аттенуированных вирусов со все более снижающейся репликацией: LID/ΔM2-2>LID/cp/ΔM2-2>LID/ΔM2-2/1030s, имеющих разные балансы аттенуации и иммуногенности.
aМартышки были привиты i.n. и i.t. 6,0 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ/АЗМ).
bКомбинированные мазки с носоглотки помещали в 2 мл среды L-15 с сахарозно-фосфатным буфером в качестве стабилизатора. Титрование вируса проводили на клетках Vero при 32°C. Нижний предел детекции составил 0,7 log10 БОЕ/мл. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты. Значение 0,35 применяли для образцов с недетектируемым вирусом. Результаты показывают, что D46/c/pΔM2-2 значительно ограничен в верхних дыхательных путях (ВДП) АЗМ.
aМартышки были привиты i.n. и i.t. 6,0 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ/АЗМ).
bНа сутки 2, 4, 6, 8, 10, и 12, проводили трахеальные смывы (TL) в 3 мл PBS. Титрование вируса проводили на клетках Vero при 32°C. Нижний предел детекции составил 1,0 log10 БОЕ/мл раствора смыва. Образцы с недетектируемым вирусом представлены как «-». Величину 0,7 применяли для образцов без детектируемого вируса. D46/cp/ΔM2-2 значительно ограничен в нижних дыхательных путях (НДП) АЗМ.
aАЗМ были привиты i.n. и i.t. 6,0 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное).
bПолучали сыворотку на сутки 0, 21, и 29 после прививки. Титры нейтрализующих антител определяли в анализе нейтрализации с 60% уменьшением бляшек. Нижний предел детекции составил 3,3 (1:10).
aМартышки были привиты i.n. и i.t. 6,0 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ/АЗМ). Значение 0,35 применяли для образцов с недетектируемым вирусом.
bКомбинированные мазки с носоглотки помещали в 2 мл среды L-15 с сахарозно-фосфатным буфером в качестве стабилизатора. Титрование вируса проводили на клетках Vero при 32°C. Нижний предел детекции составил 0,7 log10 БОЕ/мл. Образцы с недетектируемым вирусом представлены как «-». Значение 0,35 применяли для образцов с недетектируемым вирусом. Результаты показывают, что LID/cp/ΔM2-2 значительно ограничен в ВДП АЗМ.
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 6,7 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 7,0 log10 БОЕ на животное).
bНа сутки 2, 4, 6, 8, 10, и 12, проводили трахеальные смывы в 3 мл PBS. Титрование вируса проводили на клетках Vero при 32°C. Нижний предел детекции составил 1,0 log10 БОЕ/мл раствора смыва. Образцы с недетектируемым вирусом представлены как «-».Величину 0,7 применяли для образцов без детектируемого вируса. LID/cp/ΔM2-2 значительно ограничен в НДП АЗМ.
aАЗМ были привиты i.n. и i.t. 6,0 log10 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 6,3 log10 БОЕ на животное).
bПолучали сыворотку на сутки 0, 21, и 29 после прививки. Титры нейтрализующих антител определяли в анализе нейтрализации с 60% уменьшением бляшек. Нижний предел детекции составил 3,3 (1:10).
aФенотип ts для каждого вируса оценивали по анализу бляшкообразования на клетках Vero при указанных температурах. Для вирусов с фенотипом ts, титры при выключающих температурах (TSH) помечены (жирный шрифт, подчеркивание). См. примечание b для определения TSH.
bTSH (жирный шрифт, подчеркивание) определяют как наименьшую ограничивающую температуру, при которой уменьшение по сравнению с 32°C составляет 100раз или больше, чем то, что наблюдают для wt РСВ при двух температурах. Фенотип ts определяют как фенотип, имеющий TSH 40°C или меньше.
cTSP, Температуру малых бляшек определяют как наименьшую ограничивающую температуру, при которой наблюдают фенотип малых бляшек. Титры при наименьшей ограничивающей температуре помечены звездочкой.
dКурсив: Температуру микробляшек определяют как наименьшую ограничивающую температуру, при которой наблюдают фенотип микробляшек. Титры при наименьшей ограничивающей температуре для фенотипа микробляшек помечены звездочкой.
XXконтрольные вирусы ts
Пример 9
Этот пример описывает конструирование РСВ D46/276/ΔM2-2-AclI, и его сравнение у африканских зеленых обезьян с РСВ D46/NS2/N/ΔM2-2-HindIII и выбранными контрольными вирусами.
Конструировали дополнительный мутантный вирус M2-2 для представления дополнительной комбинации признаков из остовов D46/ΔM2-2 и MEDI/ΔM2-2, получая вирус под названием РСВ D46/276/ΔM2-2-AclI, который также называется в настоящем описании «РСВ 276» или «276».
Структура РСВ 276 обобщена на ФИГ. 16 и в таблице 21 и представлена в SEQ ID NO: 19. По сравнению с D46 (т.е., полной антигеномной кДНК дикого типа, SEQ ID NO: 1), РСВ 276 отличается в целом на 21 нуклеотид (включая делецию одного нуклеотида в положении 1099) плюс делеция нуклеотидов 8202-8435 включительно, дающая делецию из 234 нуклеотидов). Нуклеотидные замены удаляют четыре не-природных участка рестрикции (AflII, NcoI, StuI, и SphI), которые были намеренно введены в D46 во время его исходного конструирования (Collins, et al. 1995 Proc Natl Acad Sci USA 92:11563-11567), и, кроме того, вводят сайт AclI покрывающий делецию M2-2 (таким образом, делеция ΔM2-2-AclI такая же, как описано в ФИГ. 10). Эти замены в участках рестрикции включают восемь нуклеотидов. Большинство из остальных 13 замен вводят выбранные замены из РСВ MEDI/ΔM2-2 в остов нового вируса РСВ 276 на основе D46.
(234 nt)
1В таблице 21, нумерация нуклеотидных и аминокислотных последовательностей дана относительно биологического штама дикого типа РСВ A2 (номер доступа GeneBank M74568), который был первой полной последовательностью штамма РСВ A2. Этот геном имеет 15222 нуклеотида в длину. Таким образом, делеции или вставки в вирусах не изменяют нумерацию последовательности оставшихся нуклеотидов (или аминокислот). Названия в нуклеотидной и аминокислотной последовательности даны относительно РСВ D46 WT (SEQ ID NO:1), если не указано иначе. D46 представляет собой вторую, рекомбинантно полученную версию штамма A2, которая отличается нуклеотидной длиной из-за однонуклеотидной вставки в положении 1099 (как указано), приводящей к длине генома в 15223 нуклеотида. Эту вставку удаляли в РСВ 276, и название в этом положении стало T.
2ncr, некодирующая область.
3Аминокислоты в РСВ 276, которые отличаются от РСВ D46 затенены серым.
4ig, межгенная область
*Изменения, сконструированные в D46 для создания четырех маркерных участков рестрикции (Collins et al PNAS 92:11563-7 1995 PMID 8524804). Их удаляли в РСВ 276.
**Нуклеотидные замены, которые создают сайт AclI в РСВ 276.
Вирус 276 конструировали с использованием антигеномной кДНК D46 (SEQ ID NO: 1) в комбинации с синтетическими фрагментами ДНК. Конкретно, синтезировали кДНК, которая простиралась от уникального сайта NotI в плазмидном векторе перед лидерной областью до уникального сайта AvrII в положениях 2129-2134 в гене N в D46. Синтезировали вторую кДНК, которая простиралась от уникального сайта XhoI в D46 (положения 4481-4486) до уникального сайта BamHI (положения 8499-8505). Этот последний кусок также содержал мутацию ΔM2-2-AclI, за исключением того, что на месте желаемого сайта AclI был HindIII (участок рестрикции, который отличался инверсией порядка двух нуклеотидов, не показан). Эти два куска вводили в D46 путем способов общепринятого молекулярного клонирования, таким образом, достигая большинства желаемых нуклеотидных замен, показанных наФИГ. 16 и в таблице 21. Затем, проводили три этапа сайт-специфического мутагенеза: сайт HindIII был заменен желаемым сайтом AclI (включая замену двух соседних нуклеотидов), и были сделаны замены C10514T и C13633A в L. Это привело к антигеномной кДНК для РСВ 276 (SEQ ID NO: 19).
Вирус РСВ 276 был легко восстановлен в виде экспериментальных партий, и была подтверждена эффективность его репликации на клетках Vero. Кроме того, партия материала для клинического испытания из вируса РСВ 276 была восстановлена и произведена в условиях, подходящих для применения у человека, в препарате для клинического испытания. Было подтверждено, что его последовательность свободна от случайных мутаций.
Кроме того, партия материала для клинического испытания была произведена для вируса D46/NS2/N/ΔM2-2-HindIII (см. ФИГ. 13, второй вирус сверху). Последовательность антигеномной кДНК D46/NS2/N/ΔM2-2-HindIII показана в SEQ ID NO: 18. Последовательность материала для клинического испытания D46/NS2/N/ΔM2-2-HindIII (Lot РСВ#011B) совпадала с последовательностью плазмидной кДНК, из которой был получен рекомбинантный вирус, за исключением трех полиморфизмов: (1) G2485A (~20-40% A); кодон: GAT на AAT; аминокислота: D47N в P ORF; (2) однонуклеотидная вставка тимидина в политимидиновом участке (нуклеотиды 4537-39) в 3'-некодирующей области гена SH ген (+ 1 нуклеотид; присутствует в субпопуляции примерно 30%); (3) двухнуклеотидная аденинозиновая вставка в полиаденозиновом участке (нуклеотиды 14830-35) в сигнале окончания гена L (+1A приблизительно у 30% популяции,+2A приблизительно у 70% популяции). Эти полиморфизмы считаются биологически несущественными.
В серии исследований, экспериментальную партию РСВ 276 и три различных партии РСВ D46/NS2/N/ΔM2-2-HindIII оценивали на репликацию и иммуногенность у африканских зеленых обезьян. Каждый препарат оценивали раздельно из-за ограничений по времени и доступности животных. Результаты сравнивали совместно в таблицах 22, 23, и 24 параллельно с данными два препаратов для сравнения (LID/ΔM2-2 и LID/ΔM2-2/1030s), взятых из таблицы 1. Репликацию вируса оценивали путем количественной оценки выделения вируса из образцов мазков носоглотки (Таблица 22) и трахеальных смывов (Таблица 23), которые оценивали в анализе бляшкообразования. Было показано, что, в верхних дыхательных путях (по мазкам из носоглотки) две из трех партий D46/NS2/N/ΔM2-2-HindIII (исследования 2 и 3) выделялись заметно (средние пиковые титры 1,1-1,6 log10 БОЕ/мл) в течение ериода 5,5-7,3 суток, в то время как выделение для третьей партии (исследование 1) было минимальным. В нижних дыхательных путях (трахеальный смыв), результаты для трех партий были очень похожи, с умеренными уровнями выделения (2,2-2,6 log10 БОЕ/мл) в течение 7,8-9,2 суток. Для сравнения, выделение для РСВ 276 было очень схоже с выделением для D46/NS2/N/ΔM2-2-HindIII в исследованиях 2 и 3 в верхних дыхательных путях, и было очень схоже со всеми тремя исследованиями по выделению вируса в нижних дыхательных путях. Для сравнения, выделение LID/ΔM2-2 в обоих анатомических отделах было по существу больше, хотя выделение LID/ΔM2-2/1030s было по существу меньше. Титр РНБ60 для этих вирусов на сутки 28 составил 8,3 и 8,5 обратного log2 для двух из партий D46/NS2/N/ΔM2-2-HindIII, и 6,3 обратного log2 для третьей партии, и 8,5 обратного log2 для РСВ 276. Эти титры, в основном, равны или превышают титры, показанные для LID/ΔM2-2 и LID/ΔM2-2/1030s (Таблица 24). Таким образом, эти вирусы обеспечивают дополнительный спектр аттенуирующих фенотипов на основе остовов ΔM2-2.
(исследование 1)
(исследование 2)
(исследование 3)
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 106 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 2×106 БОЕ на животное). Исследование на АЗМ было одобрено Комитетом по уходу и использованию животных NIAID, NIH. Результаты из предыдущего исследования LID ΔM2-2 и LID ΔM2-2 1030s показаны для сравнения.
bКомбинированные мазки с носоглотки помещали в 2 мл среды L-15 с сахарозно-фосфатным буфером в качестве стабилизатора. Титрование вируса проводили на клетках Vero при 37°C. Нижний предел детекции составил 0,7 log10 БОЕ/мл. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,35 применяли для образцов без детектируемого вируса.
(исследование 1)
(исследование 2)
(исследование 3)
aАЗМ были привиты комбинированными путями (интраназально и интратрахеально) 106 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 2×106 БОЕ на животное). Результаты из предыдущего исследования LID ΔM2-2 и LID ΔM2-2 1030s показаны для сравнения.
bНа сутки 2, 4, 6, 8, 10, и 12, проводили трахеальные смывы в 3 мл PBS. Титрование вируса проводили на клетках Vero при 37°C. Нижний предел детекции составил 1,0 log10 БОЕ/мл раствора смыва. Образцы с недетектируемым вирусом представлены как «-». Пиковые титры для каждого животного подчеркнуты.
cПериод в сутках с первых до последних суток, на которые выявлялся вирус, включая негативные сутки (если были) в промежутке.
dСумму дневных титров применяют для оценки величины выделения вируса (площадь под кривой). Величину 0,7 применяли для образцов без детектируемого вируса.
(исследование 1)
(исследование 2)
aАЗМ были привиты i.n. и i.t. 106 БОЕ указанного вируса в 1 мл прививочного материала на участок (общая доза: 106,3 БОЕ на животное). Результаты из предыдущего исследования LID ΔM2-2 и LID ΔM2-2 1030s показаны для сравнения.
bПолучали сыворотку на сутки 0, 21, и 28 после прививки. Титры нейтрализующих антител определяли в анализе нейтрализации с 60% уменьшением бляшек. Нижний предел детекции составил 3,3 (1:10).
Очевидно, что точные подробности описанных способов или композиций можно варьировать или модифицировать в пределах сущности описанных вариантов осуществления. Мы претендуем на все такие модификации и вариации, которые находятся в пределах объема и сущности прилагаемой формулы изобретения.

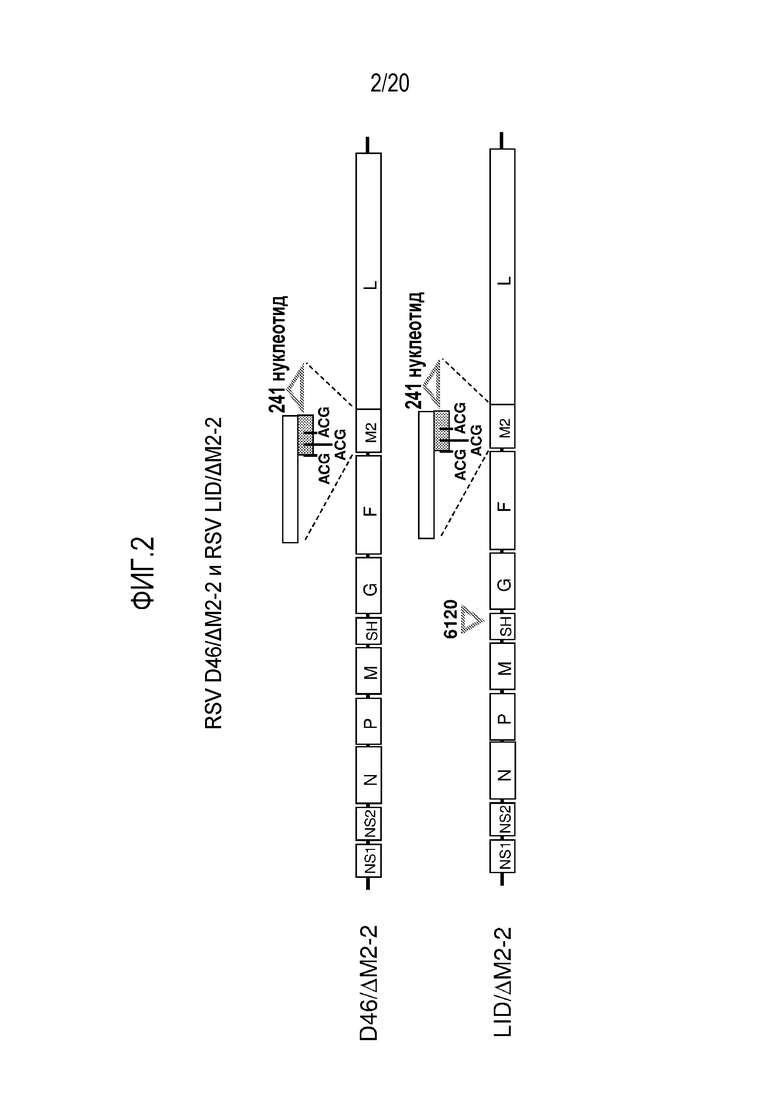

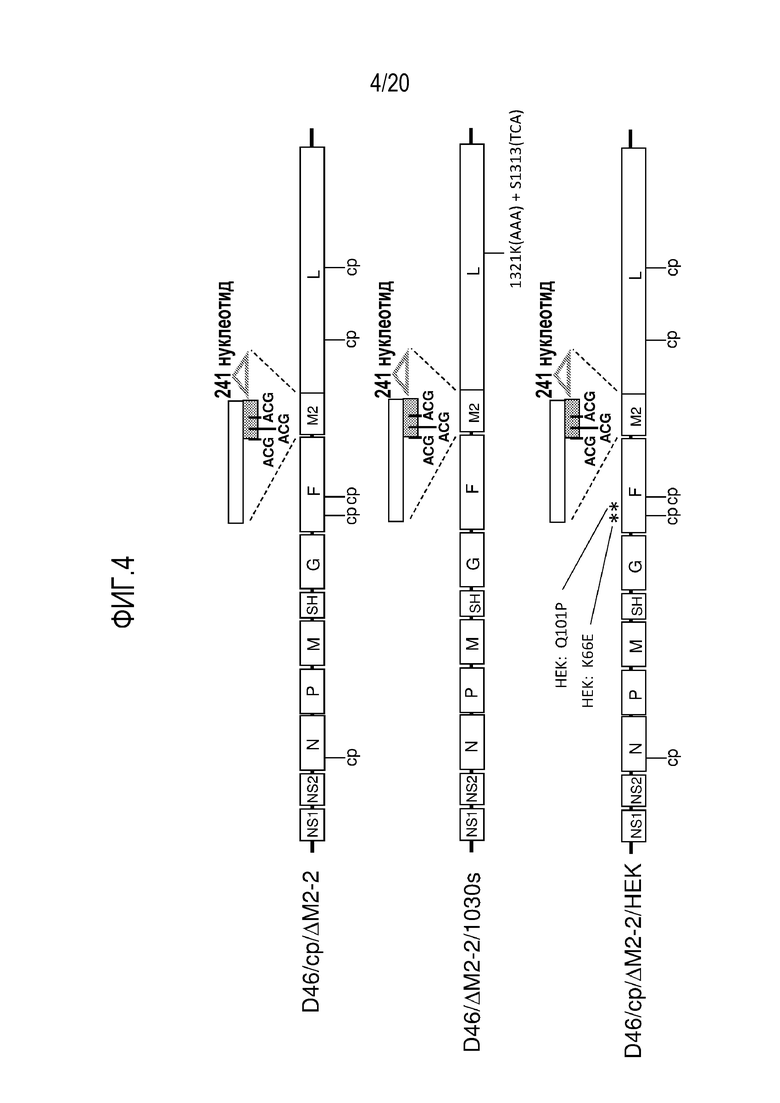


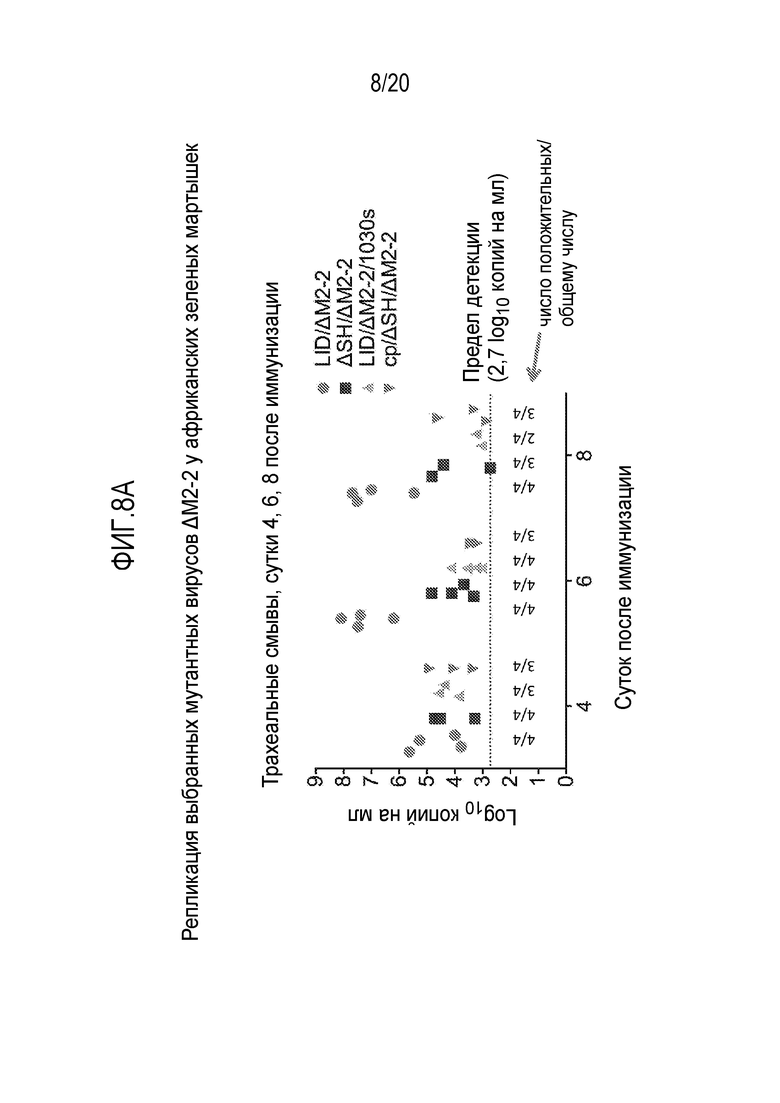
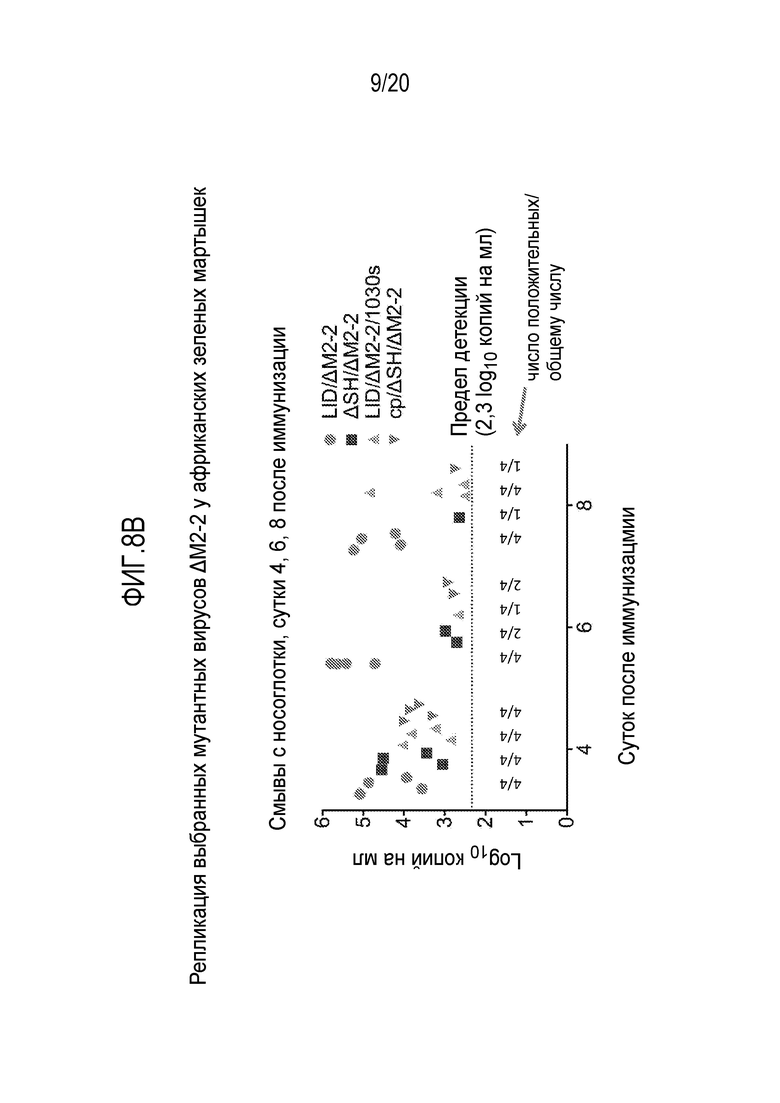
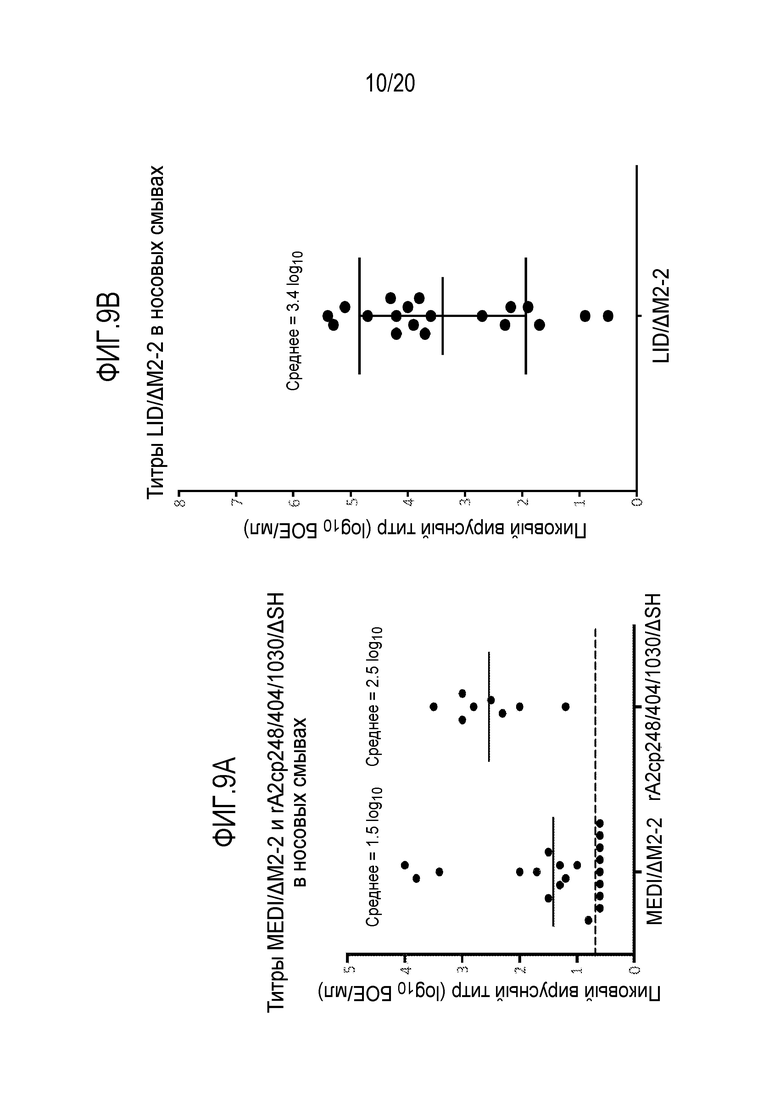





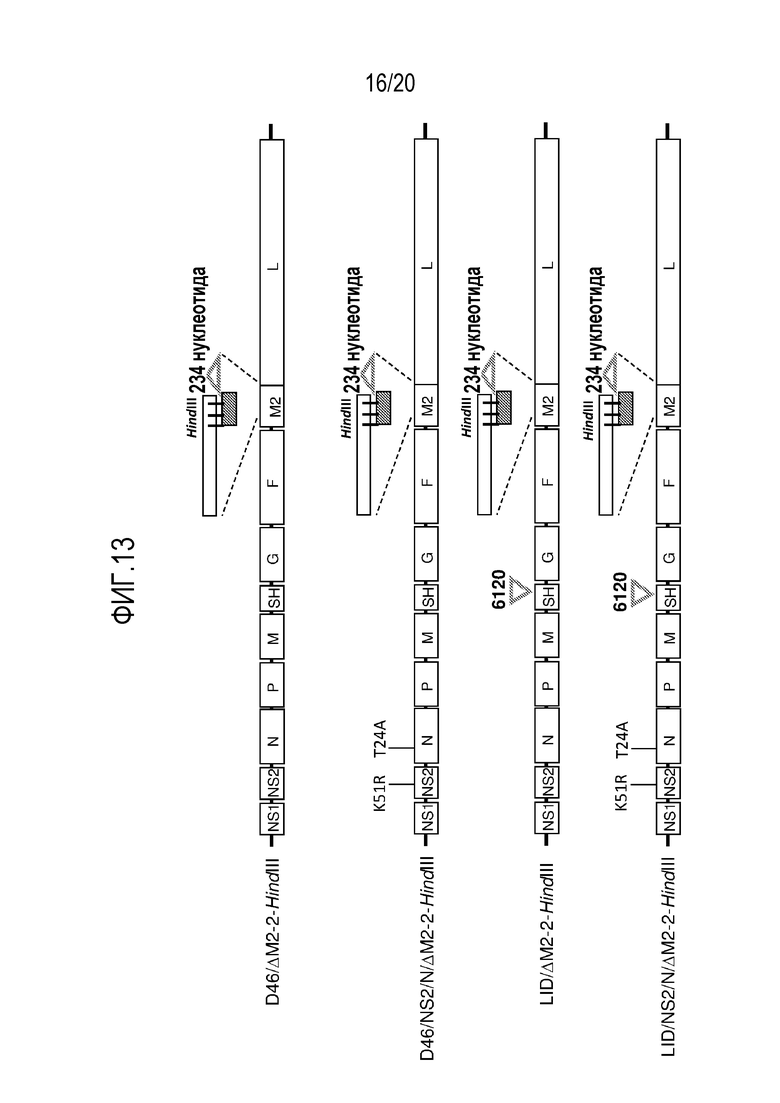
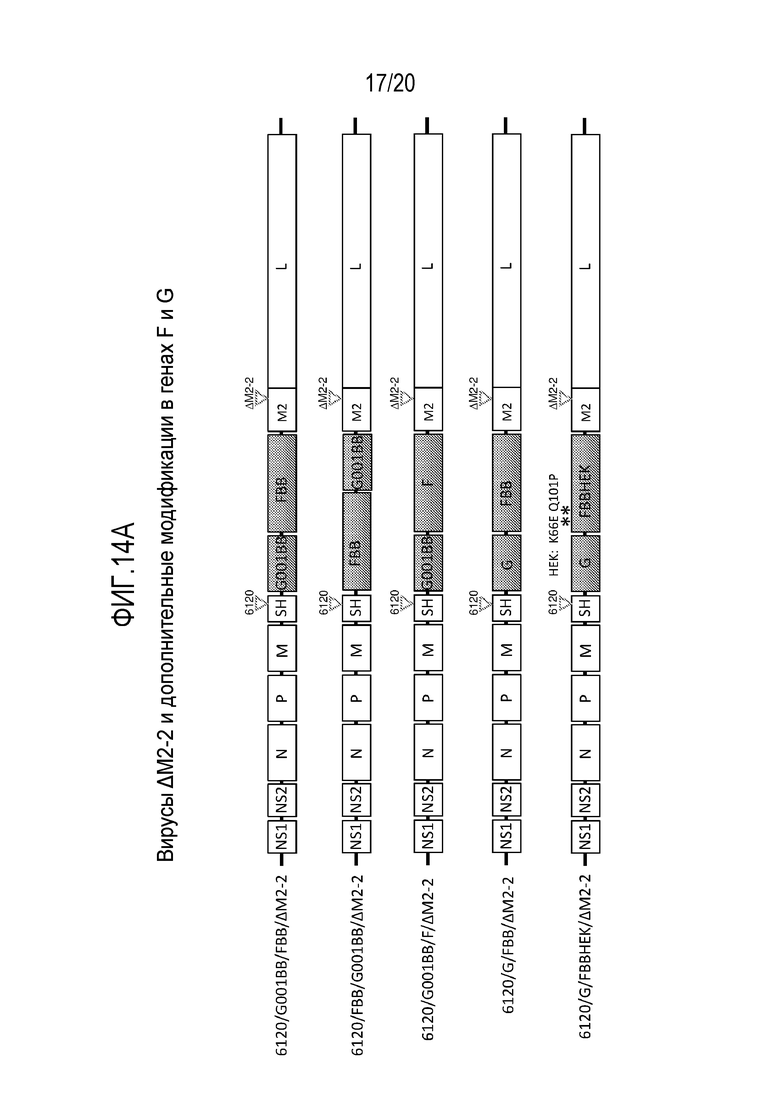

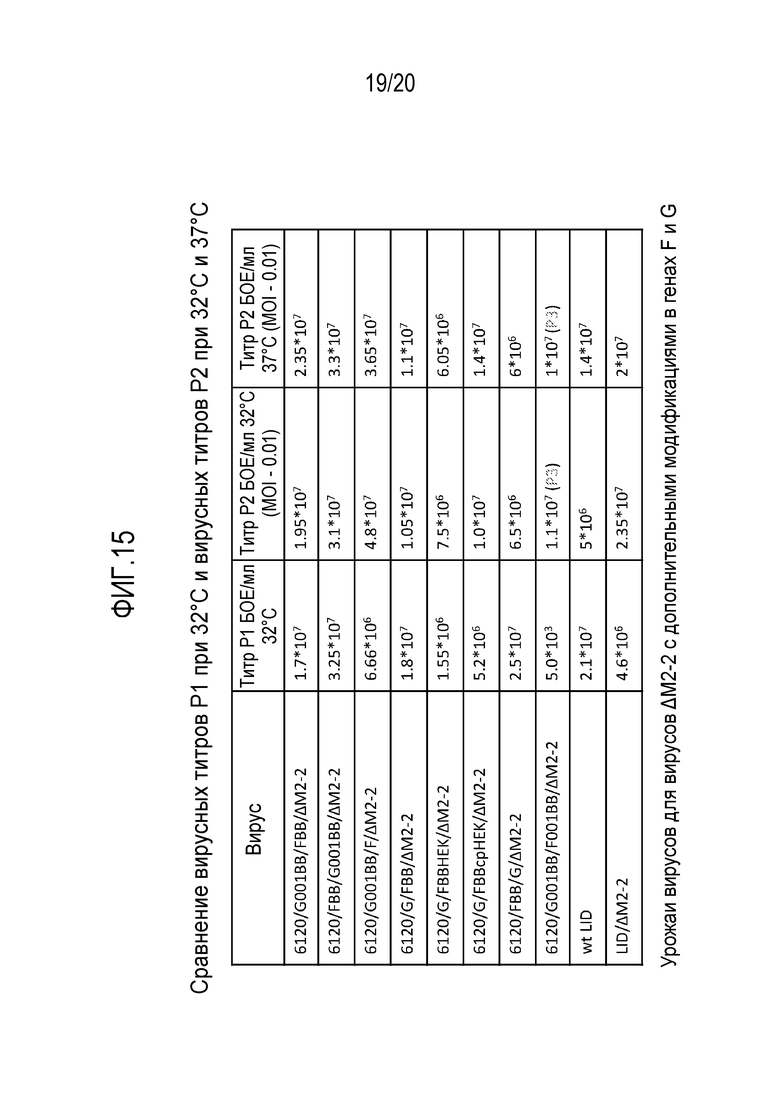
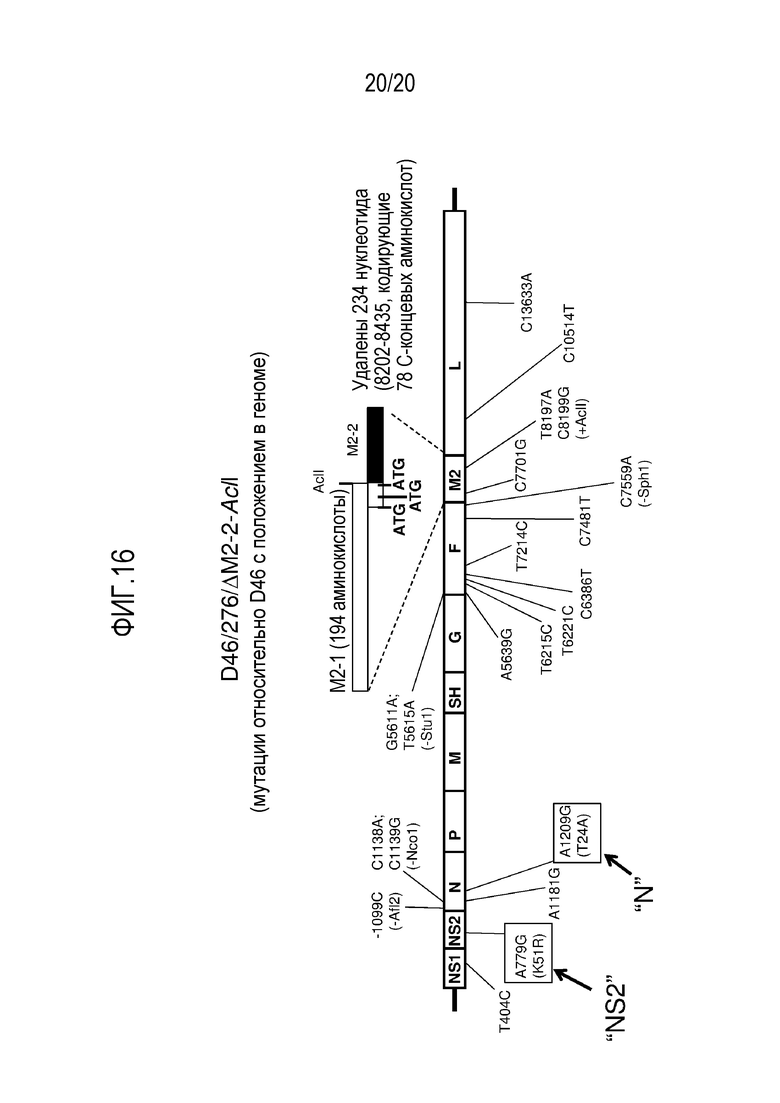
Изобретение относится к биотехнологии. Описан рекомбинантный респираторно-синцитиальный вирус (РСВ) для индукции иммунного ответа на РСВ, аттенуированный одной или несколькими модификациями до рекомбинантного генома РСВ, где одна или несколько модификаций включают: (a) делецию в открытой рамке считывания (ORF) M2-2, соответствующую делеции из 241 нуклеотида в положениях 8189-8429 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих T8161, T8167 и T8179 SEQ ID NO: 1 («ΔM2-2»); (b) делецию в M2-2 ORF, соответствующую делеции из 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих 8198A и 8200G SEQ ID NO: 1 («ΔM2-2-AclI»); или (c) делецию в M2-2 ORF, соответствующую делеции из 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих 8198A и 8199G SEQ ID NO: 1 («ΔM2-2-HindIII»), где рекомбинантный геном РСВ представляет собой геном D46, аттенуированный одной или несколькими модификациями. Также описан рекомбинантный респираторно-синцитиальный вирус (РСВ) для индукции иммунного ответа на РСВ, аттенуированный одной или несколькими модификациями до рекомбинантного генома РСВ, где одна или несколько модификаций включают: делецию в M2-2 ORF, соответствующую делеции из 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих 8198A и 8199G SEQ ID NO: 1 («ΔM2-2-HindIII»), и делецию из 112 нуклеотидов, соответствующую положениям 4499-4610 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих C4489T, C4492T, A4495T, A4497G и G4498A SEQ ID NO: 1 («6120»). Представлены соответствующие полинуклеотиды, векторы и клетки. Кроме того, представлена вакцина, содержащая указанный вирус. Описан способ получения указанного вируса. Изобретение относится к комбинациям мутаций с использованием способов, описанных в настоящем изобретении, для достижения желаемых уровней аттенуации. Рекомбинантные штаммы РСВ, описанные в настоящем изобретении, подходят для применения в качестве живых аттенуированных вакцин РСВ. 10 н. и 36 з.п. ф-лы, 16 ил., 24 табл., 9 пр.
1. Рекомбинантный респираторно-синцитиальный вирус (РСВ) для индукции иммунного ответа на РСВ, аттенуированный одной или более модификациями до рекомбинантного генома РСВ, где одна или более модификаций включают:
(a) делецию в открытой рамке считывания (ORF) M2-2, соответствующую делеции из 241 нуклеотида в положениях 8189-8429 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих T8161, T8167 и T8179 SEQ ID NO: 1 («ΔM2-2»);
(b) делецию в M2-2 ORF, соответствующую делеции из 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих 8198A и 8200G SEQ ID NO: 1 («ΔM2-2-AclI»); или
(c) делецию в M2-2 ORF, соответствующую делеции из 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих 8198A и 8199G SEQ ID NO: 1 («ΔM2-2-HindIII»), где рекомбинантный геном РСВ представляет собой геном D46, аттенуированный одной или несколькими модификациями.
2. Рекомбинантный РСВ по п. 1, где одна или более модификаций дополнительно содержат делецию из 112 нуклеотидов, соответствующую положениям 4499-4610 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих C4489T, C4492T, A4495T, A4497G, и G4498A SEQ ID NO: 1 («6120»).
3. Рекомбинантный респираторно-синцитиальный вирус (РСВ) для индукции иммунного ответа на РСВ, аттенуированный одной или более модификациями до рекомбинантного генома РСВ, где одна или более модификаций включают:
делецию в M2-2 ORF, соответствующую делеции из 234 нуклеотидов в положениях 8203-8436 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих 8198A и 8199G SEQ ID NO: 1 («ΔM2-2-HindIII»), и
делецию из 112 нуклеотидов, соответствующую положениям 4499-4610 SEQ ID NO: 1, в сочетании с нуклеотидными мутациями в положениях, соответствующих C4489T, C4492T, A4495T, A4497G, и G4498A SEQ ID NO: 1 («6120»).
4. Рекомбинантный РСВ по любому из пп. 1-3, где одна или более модификаций дополнительно содержат нуклеотидные мутации, кодирующие замены аминокислот V267I в белке N, E218A и T523I в белке F и C319Y и H1690Y в белке L РСВ («cp»).
5. Рекомбинантный РСВ по любому из пп. 1-4, где одна или более модификаций дополнительно содержат нуклеотидные мутации для введения замены Y1321K в белок L РСВ и где белок L содержит остаток S1313, где кодоны, кодирующие замену Y1321K и остаток S1313, представляют собой кодоны AAA и TCA, соответственно («1030s»).
6. Рекомбинантный РСВ по любому из пп. 1-5, где одна или более модификаций дополнительно содержат нуклеотидные мутации, кодирующие замену аминокислоты K51R в белке NS2 РСВ («NS2»).
7. Рекомбинантный РСВ по любому из пп. 1-6, где одна или более модификаций дополнительно содержат нуклеотидные мутации, кодирующие замену аминокислоты T24A в белке N РСВ («N»).
8. Рекомбинантный РСВ по любому из пп. 1-7, где одна или более модификаций дополнительно содержат нуклеотидные мутации, кодирующие замену аминокислоты K51R в белке NS2 и T24A в белке N РСВ («NS2/N»).
9. Рекомбинантный РСВ по любому из пп. 1-8, где одна или более модификаций дополнительно содержат делецию в SH ORF, соответствующую делеции из 419 нуклеотидов в положениях 4198-4616 of SEQ ID NO: 1 («ΔSH»).
10. Рекомбинантный РСВ по любому из пп. 1-9, где одна или более модификаций дополнительно содержат замену нуклеотидной последовательности, кодирующей белок G РСВ, соответствующей кодон-оптимизированной нуклеотидной последовательности, кодирующей белок G из клинического изолята A/Maryland/001/11.
11. Рекомбинантный РСВ по п. 10, где кодон-оптимизированная нуклеотидная последовательность, кодирующая белок G из клинического изолята A/Maryland/001/11, содержит нуклеотидную последовательность, соответствующую SEQ ID NO: 8 (G001BB).
12. Рекомбинантный РСВ по любому из пп. 1-11, где одна или более модификаций дополнительно содержат замену нуклеотидной последовательности, кодирующей белок F РСВ, соответствующей кодон-оптимизированной нуклеотидной последовательности, изложенной как SEQ ID NO: 9 (FBB).
13. Рекомбинантный РСВ по любому из пп. 1-11, где одна или более модификаций дополнительно содержат замену нуклеотидной последовательности, кодирующей белок F РСВ, соответствующей нуклеотидной последовательности, изложенной как SEQ ID NO: 10 (F001), которая кодирует белок F из клинического изолята A/Maryland/001/11.
14. Рекомбинантный РСВ по любому из пп. 1-11, где одна или более модификаций дополнительно содержат замену нуклеотидной последовательности, кодирующей белок F РСВ, соответствующей кодон-оптимизированной нуклеотидной последовательности, кодирующей белок F из клинического изолята A/Maryland/001/11.
15. Рекомбинантный РСВ по п. 14, где кодон-оптимизированная нуклеотидная последовательность, кодирующая белок F из клинического изолята A/Maryland/001/11, содержит нуклеотидную последовательность, соответствующую SEQ ID NO: 11 (F001BB).
16. Рекомбинантный РСВ по любому из пп. 1-15, где одна или более модификаций дополнительно содержат нуклеотидные мутации, кодирующие замены аминокислот K66E и Q101P в белке F РСВ («HEK»).
17. Рекомбинантный РСВ по любому из пп. 1-16, где одна или более модификаций дополнительно содержат нуклеотидные мутации, кодирующие замены аминокислот E218A и T523I в белке F РСВ (замены cp в F).
18. Рекомбинантный РСВ по любому из пп. 1-17, где одна или более модификаций дополнительно содержат обратный порядок генов, кодирующих белки G и F в геноме РСВ.
19. Рекомбинантный РСВ по п. 1, где одна или более модификаций включают или состоят из комбинации мутаций, выбранных из любой из:
ΔM2-2, cp/ΔM2-2, cp/ΔM2-2/HEK, ΔM2-2/1030s, NS2/N/ΔM2-2, NS2/ΔM2-2, N/ΔM2-2, ΔSH/ΔM2-2, cp/ΔSH/ΔM2-2, 6120/ΔM2-2, 6120/cp/ΔM2-2, 6120/ΔM2-2/1030s, 6120/NS2/N/ΔM2-2, 6120/G001BB/FBB/ΔM2-2, 6120/FBB/G001BB/ΔM2-2, 6120/G001BB/F/ΔM2-2, 6120/G/FBB/ΔM2-2, 6120/G/FBBHEK/ΔM2-2, 6120/G/FBBcpHEK/ΔM2-2, 6120/FBB/G/ΔM2-2, 6120/G001BB/F001BB/ΔM2-2, 6120/NS2/ΔM2-2 или 6120/N/ΔM2-2; или ΔM2-2-AclI, cp/ΔM2-2-AclI, cp/ΔM2-2-AclI/HEK, ΔM2-2-AclI/1030s, NS2/N/ΔM2-2-AclI, NS2/ΔM2-2-AclI, N/ΔM2-2-AclI, ΔSH/ΔM2-2-AclI, cp/ΔSH/ΔM2-2-AclI, 6120/ΔM2-2-AclI, 6120/cp/ΔM2-2-AclI, 6120/ΔM2-2-AclI/1030s, 6120/NS2/N/ΔM2-2-AclI, 6120/G001BB/FBB/ΔM2-2-AclI, 6120/FBB/G001BB/ΔM2-2-AclI, 6120/G001BB/F/ΔM2-2-AclI, 6120/G/FBB/ΔM2-2-AclI, 6120/G/FBBHEK/ΔM2-2-AclI, 6120/G/FBBcpHEK/ΔM2-2-AclI, 6120/FBB/G/ΔM2-2-AclI, 6120/G001BB/F001BB/ΔM2-2-AclI, 6120/NS2/ΔM2-2-AclI; или 6120/N/ΔM2-2-AclI; или ΔM2-2-HindIII; cp/ΔM2-2-HindIII, cp/ΔM2-2-HindIII/HEK, ΔM2-2-HindIII/1030s, NS2/N/ΔM2-2-HindIII, NS2/ΔM2-2-HindIII, N/ΔM2-2-HindIII, ΔSH/ΔM2-2-HindIII, cp/ΔSH/ΔM2-2-HindIII, 6120/ΔM2-2-HindIII, 6120/cp/ΔM2-2-HindIII, 6120/ΔM2-2-HindIII/1030s, 6120/NS2/N/ΔM2-2-HindIII, 6120/G001BB/FBB/ΔM2-2-HindIII, 6120/FBB/G001BB/ΔM2-2-HindIII, 6120/G001BB/F/ΔM2-2-HindIII, 6120/G/FBB/ΔM2-2-HindIII, 6120/G/FBBHEK/ΔM2-2-HindIII, 6120/G/FBBcpHEK/ΔM2-2-HindIII, 6120/FBB/G/ΔM2-2-HindIII, 6120/G001BB/F001BB/ΔM2-2-HindIII, 6120/NS2/ΔM2-2-HindIII или 6120/N/ΔM2-2-HindIII.
20. Рекомбинантный РСВ по любому из пп. 1-19, где рекомбинантный геном РСВ содержит одну или более модификаций и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичную SEQ ID NO: 1.
21. Рекомбинантный РСВ по любому из пп. 1-20, где рекомбинантный геном РСВ содержит одну или более модификаций и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 95% идентичную SEQ ID NO: 1.
22. Рекомбинантный РСВ по любому из пп. 1-21, где рекомбинантный геном РСВ содержит одну или более модификаций и остальные нуклеотиды генома РСВ содержат нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 99% идентичную SEQ ID NO: 1.
23. Рекомбинантный РСВ по любому из пп. 1-22, где рекомбинантный геном РСВ представляет собой геном D46, содержащий одну или более модификаций.
24. Рекомбинантный РСВ по п.1, где рекомбинантный геном РСВ содержит одно из:
(a) мутации 6120 и ΔМ2-2 и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 5 (последовательность LID/ΔM2-2);
(b) мутации cp и ΔМ2-2 и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 1 (последовательность D46);
(c) мутации cp и ΔМ2-2 и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 15 (последовательность D46/cp/ΔM2-2);
(d) мутации 6120, ΔM2-2 и 1030s и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 16 (последовательность LID/ΔM2-2/1030s);
(e) мутации 6120, ΔM2-2 и 1030s и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 17 (последовательность LID/cp/ΔM2-2);
(f) мутации NS2, N, ΔM2-2-HindIII и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 4 (последовательность D46/ΔM2-2-HindIII);
(g) мутации NS2, N, ΔM2-2-HindIII и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 18 (последовательность D46/NS2/N/ΔM2-2-HindIII);
(h) мутации NS2, N, ΔM2-2-AclI и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 3 (последовательность D46/ΔM2-2-AclII); или
(i) мутации NS2, N, ΔM2-2-AclI, следующие нуклеотидные мутации с положениями относительно SEQ ID NO: 1: 404C, 779G, делеция C1099, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T и 13634A; и нуклеотидную последовательность, соответствующую последовательности плюс-цепи, по меньшей мере на 90% идентичной, по меньшей мере на 95% идентичной и/или по меньшей мере на 99% идентичной SEQ ID NO: 19 (последовательность 276).
25. Рекомбинантный РСВ по п. 1, где рекомбинантный геном РСВ представляет собой один из:
генома D46/cp/ΔM2-2;
генома LID/ΔM2-2/1030s;
генома LID/cp/ΔM2-2;
генома D46/NS2/N/ΔM2-2-HindIII;
генома LID/ΔM2-2; или
генома 276.
26. Рекомбинантный РСВ по п. 1, где нуклеотидная последовательность рекомбинантного генома РСВ содержит или включает нуклеотидную последовательность, соответствующую последовательности плюс-цепи, изложенной в виде любой из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18 или SEQ ID NO: 19.
27. Рекомбинантный РСВ по п. 1, где рекомбинантный геном РСВ содержит нуклеотидную последовательность, соответствующую последовательности плюс-цепи, изложенной в виде SEQ ID NO: 3, дополнительно модифицированную введением следующих нуклеотидных мутаций относительно SEQ ID NO: 1: 404C, 779G, 1099T, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T и 13634A.
28. Рекомбинантный РСВ по п. 1, где рекомбинантный геном РСВ содержит нуклеотидную последовательность, соответствующую последовательности плюс-цепи, изложенной в виде SEQ ID NO: 3, дополнительно модифицированную введением следующих нуклеотидных мутаций относительно SEQ ID NO: 1: 404C, 779G, делеция C1099, 1139A, 1140G, 1182G, 1210G, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T и 13634A.
29. Рекомбинантный РСВ по п. 1, где рекомбинантный геном или антигеном РСВ содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3 и SEQ ID NO: 4, дополнительно модифицированную введением одного или нескольких следующих нуклеотидных замен в положениях относительно SEQ ID NO: 1: 404C, 779G, 1099T, 1139A, 1140G, 1182G, 1210G, 1938A, 5612A, 5616A, 5640G, 6216C, 6222C, 6387T, 7215C, 7482T, 7560A, 7702G, 10515T, 13634A, 13901T.
30.Рекомбинантный РСВ по любому из пп. 1-29, где РСВ представляет собой подтип A РСВ.
31. Рекомбинантный РСВ по любому из по пп. 1-19, где РСВ представляет собой подтип B РСВ.
32. Рекомбинантный РСВ по любому из пп. 1-31, где РСВ является инфекционным, аттенуированным и самореплицирующимся.
33. Выделенная полинуклеотидная молекула для получения рекомбинантного РСВ по п. 1, содержащая антигеномную последовательность кДНК генома рекомбинантного РСВ по любому из пп. 1-32.
34. Выделенная полинуклеотидная молекула для получения рекомбинантного РСВ по п. 1, содержащая антигеномную последовательность РНК генома рекомбинантного РСВ по любому из пп. 1-32.
35. Вектор для получения рекомбинантного РСВ по п.1, содержащий полинуклеотидную молекулу по п. 33 или 34.
36. Клетка для получения рекомбинантного РСВ по п.1, содержащая полинуклеотидную молекулу или вектор по любому из пп. 33-35.
37. Способ получения рекомбинантного РСВ, включающий:
трансфекцию пермиссивной клеточной культуры вектором по п. 34;
инкубирование клеточной культуры в течение достаточного периода времени для репликации вируса; и
очистку реплицированного рекомбинантного РСВ.
38. Вакцинная композиция РСВ для индукции иммунного ответа на РСВ, содержащая рекомбинантный РСВ по любому из пп. 1-32.
39. Способ индукции иммунного ответа на РСВ у индивидуума, включающий введение иммуногенно эффективного количества фармацевтической композиции по любому из пп. 1-32 индивидууму.
40. Способ по п. 39, где иммунный ответ представляет собой защитный иммунный ответ.
41. Способ по п. 39 или 40, где фармацевтическую композицию вводят интраназально.
42. Способ по п. 39 или 40, где РСВ вводят путем инъекции, аэрозоля, назального спрея или капель в нос.
43. Способ по любому из пп. 39-42, где индивидуум является человеком.
44. Способ по любому из пп. 39-43, где индивидуум имеет возраст от 1 до 6 месяцев.
45. Способ по любому из пп. 39-44, где индивидуум является серонегативным по РСВ.
46. Применение рекомбинантного РСВ по любому из пп. 1-32 для индукции иммунного ответа на РСВ у индивидуума.
| WO 2002044334 A2, 06.06.2002 | |||
| JIN H ET AL, "Respiratory Syncytial Virus That Lacks Open Reading Frame 2 of the M2 Gene (M2-2) Has Altered Growth Characteristics and Is Attenuated in Rodents", JOURNAL OF VIROLOGY, THE AMERICAN SOCIETY FOR MICROBIOLOGY, US,Vol | |||
| Приспособление в центрифугах для регулирования количества жидкости или газа, оставляемых в обрабатываемом в формах материале, в особенности при пробеливании рафинада | 0 |
|
SU74A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| US 20040005542 A1, | |||

