A. Микобактерии и вызываемые ими заболевания
Mycobacterium tuberculosis (МВТ) является возбудителем туберкулеза (ТБ) - наиболее распространенного инфекционного заболевания в мире на сегодняшний день. По данным Всемирной организации здравоохранения (ВОЗ) 1,7 миллиарда людей инфицированы или были инфицированы возбудителем туберкулеза (Kochi,A. Tubercle 72: 1-6 (1991)). В 1991 году число инфицированных людей в мире возросло на 8 миллионов человек (Sudre, P. et al., Bull. W.H.O. 70:149-159 (1992)) и по прогнозу ВОЗ в девяностых годах заболеет еще 88,2 миллиона человек, а примерно 3 миллиона людей умрет только за этот период (Morbidity and Mortality Weekly Report 42 (N. 49):961- 964 (1993)). Центры по предупреждению и контролю заболеваний США в 1992 году зафиксировали 26673 случая (Morbidity and Mortality Weekly Report 42:696- 703 (1993)) и по их оценкам от 10 до 15 миллионов человек в США имеют туберкулез в латентной форме (Morbidility and Mortality Weekly Report 39(RR-8):9-12 (1990)).
Важное значение микобактерий, относящихся к комплексу MAC (преимущественно виды M.avium и M.intracellulare), как патогенов человека, было отмечено в обзоре Inderlied, C.B. et al., Clin. Microbiol. Rev. 6: 266-310 (1993). Инфекционные заболевания, вызываемые микроорганизмами комплекса MAC, получили распространение в связи с тем, что их возбудители являются оппортунистическими патогенами у больных СПИДом. Примерно у 43% больных СПИДом на выраженных стадиях заболевания присутствуют диссеминированные инфекции MAC (Nightingale et al., Jour. Infect. Dis. 165:1082-1085 (1992)). По оценкам ВОЗ сегодня примерно 3 миллиона людей больны СПИДом и примерно 15 миллионов инфицированы вирусом иммунодефицита человека (ВИЧ), а к 2000 году число инфицированных людей может подняться примерно до 40 миллионов (Всемирная организация здравоохранения (документ WHO/GPA/CNP/EVA/93. 1) Global Programme on AIDS (Всемирная программа по СПИДу) (1993)). Предполагается, что помимо инфекционных заболеваний, связанных со СПИДом, M. paratuberculosis, подвид M. avium (Thorel, M.F. et al., Int. J. Syst. Bacteriol. 40: 254-260 (1990)) ассоциирован с болезнью Крона, воспалением кишечника (Chiodini, R.J. Clin. Micro. Rev. 2:90-117 (1989)).
Микобактерии также вызывают заболевания животных. М.paratuberculosis также вызывает воспаление кишечника у жвачных животных (Thoen, C.O. et al., Rev. Infect. Dis. 3:960-972 (1981)). Это заболевание чаще известно под названием болезнь Джона. Крупный рогатый скот с положительной реакцией на болезнь Джона отбраковывается и уничтожается. В штате Висконсин, где примерно одна треть поголовья скота инфицирована (Collins, M.T., Hoard's Dairyman Feb 10:113 (1991)), экономический ущерб в 1993 году составил 52 миллиона долларов (Arnoldi, J.M. et al., Proceedings, 3rd Int. Symp. World Assoc.Vet. Lab. Diag. 2:493- 494(1983)). В целом по стране поголовье крупного рогатого скота заражено примерно от 3 до 18% (Merkal, R.S. et al., J. Am. Vet. Med. Assoc. 190: 676-680 (1987)). Экономический ущерб только от данного заболевания по молочной отрасли достигает 1,5 миллиарда долларов ежегодно (Whitlock,R. Proceedings of the Third International Colloquium on Paratuberculosis, pp. 514-522 (1991): Whitlock, R. et al., Proceedings of the 89th Annual Meeting of the United States Animal Health Association, pp. 484-490 (1985)).
Помимо упомянутых выше микроорганизмов, известен широкий круг микобактерий, также являющихся патогенными для человека, в том числе Mycobacterium leprae, Mycobacterium Kansasii, Mycobacterium marinum, Mycobacterium fortuitum комплекс и многие другие. В обзоре Wayne. L.G. et al., Clin. Micro. Rev. 5:1-25 (1992) рассматриваются различные возбудители, относящиеся к данному роду. Однако, по распространению и вреду, наносимому этими микроорганизмами, они несопоставимы с МТВ комплексом и MAC патогенами. Например, хотя предполагается, что возбудитель лепры наиболее распространен среди этой категории микроорганизмов, во всем мире насчитывается 5,5 миллионов случаев заболевания проказой, вызываемых Mycobacterium leprae (Norden, S.K. et al., Int. J. Lepr. 63:282-287 (1993)). Таким образом, в целом эта группа микроорганизмов имеет огромное социальное значение.
Данная заявка основывается на четырех первых заявках, на основании которых испрашивается конвенционный приоритет данной заявки. В большей степени она основывается на заявке США N 08/322864 от 11 октября 1994 года, частично продолжающей заявку США N 08/224592 от 7 апреля 1994 года, которая в свою очередь является продолжением заявки США N 08/222731 от 5 апреля 1994 года.
B. Культивирование и определение микобактерий
Современные методы лабораторной диагностики микобактериозов являются довольно длительными. Поскольку этим бактериям присущ медленный рост, их инкубирование является длительным процессом. Исходя из этого, требуется длительный период времени для постановки диагноза и лица, у которых предполагается наличие инфекции, подвергаются карантину, поскольку в противном случае имеется существенный риск для общества в целом.
Кроме того, лабораторное подтверждение микобактериозов требует выделения нескольких культур от пробы пациента. Каждый образец должен инкубироваться до восьми недель (в случае М. paratuberculosis - до шестнадцати недель) прежде, чем он будет признан отрицательным. Необходимость выделять много культур от каждого образца, в котором предполагается наличие возбудителя, приводит, в частности, к периодической потере определяемых штаммов, а потеря возбудителей обусловлена жесткой химической деконтаминацией, инактивирующей сапрофитные микроорганизмы. Такие действия неэффективны и зачастую приводят к гибели микобактерий, которые пытаются выделить. Например, известно, что действия с использованием N-ацетил-L-цистеин-NaOH (NALC/NaOH) (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 31-46) убивает 28-33% существующих микобактерий (Krasnow, I. et al. , Am. J. Clin. Path. 45:352-355 (1966); Kubica, G.P.W. et al., Am. Rev. Resir. Dis. 87: 775-779 (1963)). Новые современные методы исследования (Gonzales, R. et al., Diag. Microbiol. Infect. Dis. 8:69-78 (1987)) по дополнительным способам культивирования ускорили время диагностики: однако, все еще остается поле для деятельности по существенному улучшению метода.
Комбинация социальной значимости и надежности методов культивирования показывает крайнюю необходимость разработки способов определения микобактерий, которые сокращают время анализа и повышают чувствительность. Изотермальная схема, поставленная на коммерческую основу корпорацией Gen-Probe (San Diego, CA: Jonas. V. et al., J. Clin. Micro. 31:2410-2416 (1993)), и полимеразная цепная реакция (ПЦР) обе способны обнаруживать единичные молекулы (Higuchi et al., Nature (London) 332:543-546 (1988)). Более того, амплификация и обнаружение могут быть выполнены примерно за 8 часов, причем реагенты существенно не увеличивают цену анализа. Если есть возможность применения, амплификация может существенно повысить скорость и чувствительность анализа, а также снизить цену диагностики микобактерий (De Cresce, R.P. et al., Med. Laboratory Obs. 25:28-33 (1993)). Быстрота, с которой данные технологии могут способствовать диагностике микобактериозов, может иметь огромный финансовый эффект для общества.
Однако, как описывается в данной работе, исследователи сталкиваются с множеством проблем при попытке внедрения таких технологий, как ПЦР амплификация, при выявлении микобактерий. Особенно было сложно создать способ по подготовке образца для анализа таким образом, чтобы (а) гарантировать определение методом амплификации истинного положительного результата, а также (б) не дать ложноотрицательных результатов. Вариабельность результатов, с которой неожиданно сталкиваются исследователи, приводится в работе Noordhoek, G. T. et al. , J. Clin. Micro. 32:277-284 (1994). Данные авторы рассматривают независимые исследования, выполненные в семи лабораториях. Во всех лабораториях использовалась одна и та же система амплификации, но разные методы обработки и определения. Исходные краткие изложения данных результатов (Noordhoek, G.T. et al., N. Eng. J. Med. 329:2036 (1993) показывают, что при небольшом числе копий (1000 копий) корреляция варьирует от 2 до 90%, среднее значение 54%. В итоге из-за существующих проблем до сих пор не получено разрешение FDA (Food and drug administration - Департамент по контролю за качеством пищевых продуктов, медикаментов и косметических средств) на применение наборов для ТБ-амплификации.
C. Способы обработки образцов, содержащих микобактерии
Обзор научной литературы по амплификации нуклеиновых кислот микобактерий показывает в соответствии с общей идеей, методами работы с образцом и клиническими исследованиями, что существует высокая вариабельность результатов, что послужило основанием для FDA не одобрить применение наборов для ТБ-амплификации.
Использовалось две разные схемы амплификации; существует множество ПЦР методов и много клинических исследований, направленных на изучение микобактериозов, в подавляющем числе случаев на изучение МТБ. Как правило, сначала из образцов делают высев и затем материал используют для амплификации. Таким образом, работа с образцом по амплификации может быть рассмотрена, в большинстве случаев, как дополнение к методу культивирования.
Существует ряд причин, почему развитие исследований шло в этом направлении. Во-первых, получение клинических образцов является сложным. Лица, которых диагностируют на туберкулез, в ходе диагноза обязательно подвергаются медикаментозному лечению. Во-вторых, работа с МТВ образцами требует специальных мер безопасности и соответственно разработанного технического обеспечения. В-третьих, самый легкий путь - это получить результаты по "корреляции с результатами высева", т.е. корреляции положительного или отрицательного результата амплификации и одновременно положительного или отрицательного результата того же образца по высеву. Поэтому обычно исследователи сначала работают с образцом, чтобы высеять возбудитель, и затем используют методики по амплификации для "дальнейшей" работы с оставшимся материалом. При такой последовательности в работе могут использоваться подлинные клинические образцы, безопасность не нарушается, работа движется непрерывно, сохраняется забота о пациенте и корреляция с современными методиками является реальностью. Эта упомянутая выше "дальнейшая" работа включала широкий диапазон методов обработки проб и лизиса клеток.
В табл. 1 суммировано 35 публикаций, в которых образцы для работы были получены в 17 странах, и оцениваются существующие в клинических лабораториях методы по амплификации. В табл. 1 работы представлены в хронологическом порядке. Каждая ссылка представлена таким образом, чтобы точно представить суть исходной публикации. Однако, в отдельных статьях можно заметить двусмысленность в интерпретации некоторых позиций и отличительных особенностей. Далее в примечании приводятся разъяснения.
За основу при сравнительном анализе данных, представленных в табл. 1, была выбрана корреляция результатов амплификации нуклеиновых кислот и результатов культивирования. При таком подходе исследование выявило загадочное явление: согласно методам, представленным в табл. 1, образец, подвергшийся амплификации, в подавляющем большинстве случаев происходит из "пуговицы" (осадка после центрифугирования), откуда производился высев культуры. Приведенная чувствительность амплификации по отношению к культивированию, положительный результат по культивированию/отрицательный результат по амплификации (т. е. ложная отрицательная амплификация) не имеет смысла. Некоторые авторы дают "поправочные корреляции" (См. примечание C в табл. 1). Например, если были получены ложные отрицательные результаты, и результат может быть повторно определен путем дальнейшей очистки ДНК-мишени, разбавлением ингибиторов, множественной амплификацией этого же образца, множественной амплификацией разных образцов, полученных от этого же пациента, или повторной амплификацией амплифицированного образца; откорректированные результаты были представлены.
Возникает интересная дилемма. Jackson, J.B. et al., J. Clin. Micro. 31: 3123-3128 (1993)) успешно снабдили надежно защищенной ВИЧ-ПЦР панелью 11 лабораторий. Опубликованная чувствительность показывает, что все лаборатории обладают обычной возможностью идентифицировать 2 копии генома ВИЧ на фоне 106 клеток человека. Данные, представленные в табл. 1, четко показывают, что чувствительность методов ТБ-амплификации является схожей. Из анализа данных табл. 1 следует, что хотя чувствительность данных методов амплификации по выявлению микобактерий, как и ожидалось, на много порядков выше, чем чувствительность высевов или мазков для микроскопирования, однако против ожидания существуют постоянные аберрации метода. Хотя некоторые аномалии происходят из-за ингибирования и поэтому легко объяснимы, многие нарушения являются "необъяснимыми". Становится ясным, что даже если используется внутренний контроль для выявления ингибиторов, эти необъяснимые аберрации довольно обычны и являются существенным препятствием для внедрения методов амплификации в практику клинических лабораторий для выявления микобактерий. Например, если при положительном результате или предположительно положительном можно провести повторную проверку, то при отрицательном, как правило, нет. Поскольку ложноотрицательные результаты будут постоянно, сотрудникам лаборатории будет неясно, который из образцов дал действительно ложноотрицательный результат и нуждается в "перепроверке". Следовательно, если диагноз будет неправильно признан отрицательным, не будет обеспечено должной заботы о пациенте. Поэтому для обсуждения приводимых публикаций были привлечены также данные, которые не могут быть признаны корректными. Эти аберрации - ложноотрицательные результаты - находятся в центре продолжающегося обсуждения.
(A) Во всех исследованиях за исключением Hermans, P.W.M. et al., J. Clin. Micro. 28: 1204-1213 (1990), deWit, D.et al., J. Clin. Micro. 28: 2437-2441 (1990), Del Portillo, P. et al. , Clin. Micro. 29: 2163-2168 (1991), Irula, J.V. et al., J. Clin. Micro. 31:1811-1814 (1993), и, возможно, Thierry, D. et al., J. Clin. Micro. 28:2668- 2673 (1990), при работе с образцами в первую очередь производится высев. Практически все способы работы с образцами суммированы в работе Kent, P.T. et al., "Public Health Mycobacteriology" в A Guide for the Level III Laboratory, U.S. Department of Health and Human Services. Centers for Disease Control, 1985, pp. 31-46. Для описания обработки клинических образцов используются следующие сокращения: "NALC" - сокращенное название способа с использованием N-ацетил-L-цистеина/гидроксида натрия для разжижения и деконтаминации, предложенного Kubica, G. P. W. et al., Am. Rev. Resir. Dis. 87:775-779 (1963). "DTT" - сокращенное название способа, в котором используется дитиотриэтол (известный также как SputolysinTM) для работы с микобактериями по способу Hirsch, S.R. et al., J. Lab. Clin. Med. 74:346-353 (1969). "SDS" (натрий додецилсульфат) имеет отношение к способу Engbaek, H. C. et al., Scand. J. Respir. Dis. 48:268-284(1967). "Fic/Hyp" имеет отношение к использованию градиента Ficoll/Hypaque для очистки мононуклеарных клеток периферической крови (PBMC - periferal blood mononuclear cells) согласно методу Boyum (Boyum, A.J. Clin. Lab. Invest. 21 (Suppl. 97):77-109 (1968)). "NaOH" относится к способам деконтаминации с использованием гидроксида натрия, специфических для авторов, или четко указанного автором, когда детали способа деконтаминации не оговорены. "OxAc" относится к применению оксалиновой кислоты (Corper, H.J. et al. , J. Lab. Clin. Med. 15:348-369 (1930). "PEG" относится к применению преципитации полиэтиленгликолем по усмотрению авторов. "TriPO4" относится к обработке трифосфатом Collins et al., Organization and Practice in Tuberculosis Bacteriology, Buttersworth, London, 1985. Конкретно, образец центрифугируется для получения "пуговицы" (осадка). Осадок ресуспендируется в воде или солевом фосфатном буфере (PBS - phosphate buffered saline), после чего берутся аликвоты для высева и для мазка. Затем оставшийся осадок используется для амплификации. Сокращение "P" применяется для обозначения способа, исходящего из того факта, что ресуспендированный осадок центрифугировался повторно до того, как приступить к дальнейшей амплификации, и затем в работе использовался осадок в виде пилюли. Сокращение "S" применяется для обозначения способа, исходящего из того факта, что супернатант осветлялся центрифугированием и в таком виде использовался для исследования. Учитывая последующий высев, некоторые авторы сравнивают различные способы приготовления образца для амплификации: здесь учтены только те способы работы с образцами, которые представляют интерес для данного исследования. Такие исследования по сравнению способов обработки проб раскрываются в тексте, там же приводится обсуждение результатов. Приводятся следующие аббревиатуры названий буферов: "TE" = Tris/EDTA, "TX" = Tris/Triton-X100, "TEX" =Tris/EDTA/Triton-X100 и "Nonl" = Tris/Tween20 & NP-40. Сокращение "W" применяется для обозначения стадии с использованием промывочных буферов. "GenBuf", "IDEXXBuf" и "PCRBuf" применяется для обозначения суспендирования/ресуспендирования образца в Gen-Probe буфере или в IDEXX буфере (составы которых неизвестны) или в ПЦР буфере (20 mM Tris-HCl pH 8,3, 50 mM KCl, 2,5 mM MgCl2, 0,45% Tween20, 0,45% NP-40), соответственно. "CHCl3", "HCl" и "perCl" связаны с использованием хлороформа, соляной кислоты и хлорной кислоты, соответственно, по ходу работы. "PrK" и "Lz" относятся к энзиматическому расщеплению образца протеиназой K и/или лизоцимом. Любой этап, включающий кипячение или обработку при экстремальной температуре, упрощенно обозначается как 95o на протяжении указываемого отрезка времени. "Org/ppt" относится к любым способам экстракции с использованием органических растворителей или их смеси, таких как фенол, хлороформ и/или изоамиловый спирт, с последующей преципитацией нуклеиновой кислоты этанолом или изопропанолом. Существует множество вариаций на данную тему, но все методики схожи с методом Maniatis, et al., "Molecular Cloning A Laboratory Manual", Cold Spring Harbor Laboratory, New York, 1982, pp. 458-463). "TMA" относится к утилизации четвертичных аминов для преципитации ДНК или полисахаридов, которая схожа с методом Baess, I. Acta Path. Microbiol. Scand. Sect. B, 82:780-784 (1974). "GuSCN/Si" относится к очистке ДНК из образца с использованием гуанидинизотиоцианата в присутствии кремния, как описано, например, Boom, R. et al., J. Clin. Microbiol. 28:495-503 (1990). "Sonic" относится к способу высвобождения ДНК путем сонации образца (с/без использования стеклянных бус). Kubica, G.P.W. et al., Am. Rev. Resir. Dis. 87:775-779 (1963).
(B) Общее количество клинических образцов, использовавшихся в данном исследовании, первоначально сопровождается числом положительных высевов и затем числом положительных результатов амплификаций. Дополнительные ссылки в данной колонке описывают соответствующие характеристики по данному исследованию.
(C) Представленное является несоответствием корреляции между результатами амплификации и результатами высевов (например, число образцов с положительной амплификацией, которые также дали положительные результаты высевов, отделенные общим количеством положительных высевов). Некоторые авторы продолжили исследование противоречивых результатов (например, "ложноотрицательных" или "ложноположительных") или включили информацию о пациентах в обобщение и опубликовали скорректированные результаты. Корреляция, отмеченная в этой колонке, относится к неоткорректированным результатам. Если существует расхождение между опубликованными результатами и результатами, приведенными в этой колонке, число заключено в круглые скобки и снабжено соответствующей ссылкой, объясняющей различие.
(D) Shankar, P. et al., Lancet 335:423 (1990) неясно сообщает метод, использовавшийся для работы с образцами как по высевам, так и по ПЦР. Однако, Manjunath, N.et al., Tubercle 72:21-27 (1991) утверждают, что их публикация является итоговой версией предыдущей работы, представленной в исследовании Shankar, P. et al., Lancet 335:423 (1990). Следовательно, в целях дискуссии допускается, что методические условия идентичны таковым в работе Manjunath, N.et al., Tubercle 72:21-27 (1991).
(E) В исследовании Hermans, P.W.M. et al., J. Clin. Micro. 28:1204-1213 (1990) работа с образцами производится сразу, без предварительной обработки по методу амплификации (т.е. из образца высев не производился и осадок не разделялся на высев и на ПЦР).
(F) В работе Pao, C.C. et al., J. Clin. Micro. 28:1877-1880 (1990) нет неоткорректированных результатов по ПЦР. Однако, они утверждают, что: "Если первоначальный результат был отрицательный, часть (обычно одна пятая) первично амплифицированной реакционной смеси амплифицировалась еще дополнительно 32 цикла...". Следовательно, они могли быть первоначально ложноотрицательными.
(G) Культура в пробирке со скошенным агаром, соответствующая такому ложноотрицательному результату ПЦР в исследовании Sjobring et al., J. Clin. Micro. 28: 2200-2204 (1990) дала единичную колонию. Оставшиеся три образца, давшие положительный результат ПЦР, были непосредственно подвергнуты амплификации.
(H) В исследовании deWit, et al., Tubercle and Lung Dis. 73:262-267 используется только плевральная респираторная жидкость. Образцы без предварительной обработки были непосредственно осаждены PEG и подвергнуты амплифицированию.
(I) Thierry, D. et al., J. Clin. Micro. 28:2668-2673 (1990) утверждают: "ДНК экстрагировалась из 0,2-1 мл клинического образца, не прошедшего предварительной обработки, или из образца, деконтаминированного гидроксидом натрия...". Следовательно, в целях обсуждения, следует заключить, что образец мог использоваться непосредственно для амплификации.
(J) Pierre, C. et al., J. Clin. Micro. 29:712-717 (1991) рассматривают два метода исследования клинических образцов. Авторы однозначно не сообщают, какой именно использовался для этой работы. Однако, они говорят о том, что: "В некоторых случаях часть образца..." исследовалась с помощью второго метода неионного лизиса. Также они утверждают, что в этих образцах было обнаружено больше ингибиторов. Следовательно, для дискуссии допускается, что первым методом в данной работе была экстракция органическими растворителями.
(K) Pierre, C. et al., J. Clin. Micro. 29:712-717 (1991) используют метод "повторной амплификации" с открытыми пробирками, часть образца помещается в свежую ПЦР смесь и повторно амплифицируется. По их данным было 79,2% по первичной амплификации и 100% после повторной.
(L) По данным Del Portillo, P. et al., J. Clin. Micro. 29:2163-2168 (1991) непосредственно для амплификации использовалась слюна.
(M) Brisson-Noel, A. et al., Lancet 338:364-366 (1991) анализировали 514 образцов, но только 446 из них подвергались как ПЦР, так и высеву.
(N) Система, использовавшаяся Brisson-Noel, A. et al., Lancet 338:364-366 (1991), создана в отношении белка, имеющего мол.вес 65Kd, и амплифицирует большинство микобактерий. Из 141 образца, давшего положительный ответ по результату культивирования, 130 содержали МВТ и 11 - МОТТ (микобактерии другие, чем туберкулезные). При этом 126 из них дали положительную ПЦР, 6 - ложноотрицательную и 9 содержали ингибиторы. Таким образом, корреляция с результатами высевов составила 89,4% (126 : 141 = 0,894). Сообщается, что корреляция составила 97,4%, при этом были учтены как элиминация образцов ингибиторами, так и анализ состояния пациентов (например, веские клинические основания считать, что образцы содержат микобактерии, и, хотя они были ПЦР положительные, но дали отрицательный результат по высеву).
(O) Sritharan, V. et al., Mol. Cell. Probes 5:385-395 (1991) сравнивает 8 различных способов выделения ДНК для ПЦР. Для исследования был выбран метод с кипячением.
(P) Корреляция, установленная Sritharan, V. et al., Mol. Cell. Probes 5: 385-395 (1991), составляет 100% и представляет повторную амплификацию трех образцов, которые дали ложноотрицательный результат при первой ПЦР. Некорректировавшаяся корреляция составляет 96,0%.
(Q) van der Giessen, J.W.B. et al., J. Clin. Microbiol. 30:1216-1219 (1992) сравнивали три системы на основе ПЦР (McFadden, J.J. et al., Mol. Microbiol. 1:283-291 (1987); van der Giessen, J.W.B. etal., J. Med. Microbiol. 36: 255-263 (1992); Vary, P. H. et al. , J. Clin. Microbiol. 28:933-937 (1990)), разработанных для выявления М. paratuberculosis в фекалиях крупного рогатого скота. Одним из них является коммерчески доступный набор от IDEXX (Vary, P. H. et al., J. Clin. Microbiol. 28:933-937 (1990)). В этой работе они исследовали 87 образцов каждым из трех методов в двух независимых повторах. Высевы производились однократно (из них положительных было 30), в то время как каждая ПЦР проводилась дважды, поэтому представлено два числа.
(R) Buck, G.E. et al., J. Clin. Micro. 30:1331-1334 (1992) сравнивает 4 различных способа выделения ДНК для ПЦР. Для работы был выбран метод сонации. Кроме того, было известно, что все образцы в данном исследовании ТБ-положительные по результатам высевов и по микроскопированию мазков.
(S) Victor et al. , J. Clin. Micro. 30:1514-1517 (1992) разделяли все осадки и как непосредственно амплифицировали, так и очищали путем фракционирования в градиенте сахарозы. Результаты обоих методов были доступны для обсуждения.
(T) Fauville-Dufaux et al., Eur. J. Micro. Inf. Dis. 11:797-803 (1992) утверждают, что образцы обрабатывались как SDS, так и трифосфатом. В дальнейшем они не разделяют, какой образец был исследован каким методом.
(U) Праймер, использовавшийся Fauville-Dufaux et al., Eur. J. Micro. Inf. Dis. 11:797-803 (1992), был сконструирован против антигенной 85-членной последовательности. Соответственно, с помощью их системы можно амплифицировать большинство видов микобактерий. Из 206 исследовавшихся образцов 92 дали положительные результаты высева. Из этих 92 образцов о 84 изначально было известно, что они дали положительный ответ при анализе на ПЦР (91,3%). Из 92 было определено, что в 82 образцах содержится МВТ. Из этих 82 только 74 были исходно положительны по ПЦР (90,2%). Хотя, как было показано, все 8 с ложным результатом содержат ингибиторы, 3 образца можно было развести до такой концентрации, когда фиксировался положительный сигнал. Таким образом, в результате дополнительного разведения авторы утверждают, что 77 из 82 (93,9%) достоверно идентифицировались с помощью ПЦР. Авторы оценивают величину корреляции по своим данным как 93,9%.
(V) Wilson, S. M. et al., J. Clin. Micro. 31:776-782 (1993) сравнивали два разных метода для выделения ДНК для ПЦР. Оба метода использовались для всех образцов в данной работе, поэтому получено две корреляции. Кроме того, Wilson, S.M. et al., J. Clin. Micro. 31:776-782 (1993) применяют способ "из одной пробирки".
(W) Wilson, S. M. et al., J. Clin. Micro. 31:776-782 (1993) определяют степень корреляции как 75% для GuSCN/Si метода и как 92% для выделения с применением хлороформа. Представлены "результаты пациента". Полученные результаты по корреляции в данной колонке представляют опубликованные результаты корреляции "индивидуального образца". Кроме того, все образцы в данной работе были исследованы с повтором и расхождения результатов были разрешены.
(X) Folgueira, L. et al., J. Clin. Micro. 31:1019-1021 (1993) сравнивали два разных метода для выделения ДНК для ПЦР. Продемонстрированный метод с кипячением был выбран для исследования. Было известно, что все образцы в данной работе ТБ-положительные.
(Y) Folgueira, L. et al., J. Clin. Micro. 31:1019-1021 (1993) утверждают, что корреляция результатов по высеву и ПЦР составляет 100% и в том числе представляют данные по семи образцам, нуждавшимся в разбавлении ингибиторов. Неоткорректированная корреляция составляет 90,7%.
(Z) Kocagoz, Т. et al., J. Clin. Micro. 31:1435- 1438 (1993) сравнивают два различных метода для получения осадка для ПЦР. Продемонстрированный метод с кипячением был выбран для исследования.
(α) "Золотой стандарт", использовавшийся Kocagoz, Т. et al., J. Clin. Micro. 31:1435-1438 (1993), представляет собой "высокую вероятность на основе клинических признаков". Хотя по этому критерию дана корреляция 87%, величина корреляции по высевам составляет 100%.
(β) Forbes, B.A. et al., J. Clin. Micro. 32:1688-1694 (1993) исходно анализировали 173 образца для оптимизации ПЦР. Затем они проанализировали 727 образцов. Информация обоих исследований пригодна для обсуждения.
(γ) Forbes, B.A. et al., J. Clin. Micro. 32:1688-1694 (1993) установили величину корреляции 87,2% для 727 изученных образцов. Эта чувствительность предполагает включение данных о пациенте при анализе образцов ПЦК положительных/по высевам отрицательных. В этой работе в табл. 2 показано, что по результатам высевов из 80 MTB-положительных культур 67 были определены методом ПЦР. Следовательно, неоткорректированная чувствительность могла быть 83,8%.
(δ) Irula, J.V. et al., J. Clin. Micro. 31:1811-1814 (1993) в работе использовали только образцы крови. Кроме того, ценность данной работы состоит в том, что проведена идентификация M.avium и подтверждено отличие от MTB.
 Clarridge, J.E. et al., J. Clin. Micro. 31:2049-2056 (1993) установили несколько случаев по корреляции высевов. Реальное число образцов с MTB-положительным результатом высева составило 218, а число образцов с положительной ПЦР среди этой группы составило 181 (см. табл. 2 в этой работе). Следовательно, некорректная корреляция, отмеченная для обсуждения, составляет 83,0%.
Clarridge, J.E. et al., J. Clin. Micro. 31:2049-2056 (1993) установили несколько случаев по корреляции высевов. Реальное число образцов с MTB-положительным результатом высева составило 218, а число образцов с положительной ПЦР среди этой группы составило 181 (см. табл. 2 в этой работе). Следовательно, некорректная корреляция, отмеченная для обсуждения, составляет 83,0%.
(ζ) Miyazaki, Y. et al., J. Clin. Micro. 31:2228-2232 (1993) используют систему ПЦР (т.е. образец из первой реакции переносился в свежую смесь для ПЦР и повторно амплифицировался) и идентифицировали 54 из 56 образцов, которые дали положительный результат высева (корреляция 96,4%). Однако, также было 10 образцов, которые были отрицательны по высевам/положительны по микроскопированию мазков/положительны по ПЦР. Авторы включили эти 10 образцов и определили коэффициент корреляции 97,0% (64 : 66 = 0,970).
(η) В работе Jonas, V. et al., J. Clin. Micro. 31:2410-2416 (1993) исключительно использовалась запатентованная коммерческая схема Gen-Probe.
(θ) B работе Jonas, V. et al., J. Clin. Micro. 31:2410- 2416 (1993) из 119 образцов с положительным результатом высева только 95 было подтверждено путем амплификации (79,8%). 21 образец был отрицательным по высевам/положительным по амплификации. Авторы считают, что только 17 из них были истинно положительными. Включение этих 17 говорит о том, что путем амплификации идентифицировано 112 из 136 истинно положительных проб, что повышает чувствительность по данной работе до 82,4%.
(ι) В работе Abe, C. et al., J. Clin. Micro. 31:3270-3274 (1993) все образцы использовались таким образом, что результаты, полученные Gen-Probe методом (Gen), можно было сопоставить с результатами, полученными с помощью системы ПЦР, описанной Kolk, A.H.J. et al., J. Clin. Micro. 30:2567-2575 (1992).
(κ) Abe, C. et al., J. Clin. Micro. 31:3270-3274 (1993) разрешили несколько противоречивых результатов и установили, что корреляция метода Gen-Probe составляет 91,9% и 84,2% по ПЦР. Неожиданное разрешение противоречивых данных дало 90,6 и 81,3%, соответственно. Чувствительность метода Gen-Probe была выше, чем ПЦР.
(λ) Принципиальной схемой амплификации, использовавшейся Miller et al., J. Clin. Micro. 32:393-397 (1994) была коммерческая схема Gen-Probe. Однако, они сравнили эти результаты с результатами, полученными при использовании системы ПЦР Eisenach et al. , J. Clin. Micro. 26:2240-2245 (1988). После того, как все 750 образцов были амплифицированы методом Gen-Probe, 156 из них, давших положительный результат, были также проамплифицированы с помощью ПЦР.
(μ) 156 образцов в работе Miller et al., J. Clin. Micro. 32:393-397 (1994) были "...с положительным результатом высева и/или диагностированные как туберкулезные по клиническим признакам." Из этих 156 образцов только 142 были действительно положительные по результатам высевов. В последующем "повторный анализ образцов с другим объемом аликвоты из тех же самых осадков повысил число выявленных культур из 156 до 142 (91,0%)." Реальные данные представлены таким образом, что включают все образцы, но невозможно определить, представляет ли это число корреляцию с результатами высевов. Амплификация методом ПЦР 156 образцов с положительным результатом высева первоначально идентифицировала 122 (78,2%) образца. 143 (92,3%) было идентифицировано при повторном тестировании ПЦР (чувствительность ПЦР лишь незначительно превышает чувствительность метода Gen-Probe). Ряд образцов, идентифицированных как содержащие микобактерии на основании микроскопирования мазков, были затем признаны как ложноположительные на основании первичного анализа обоими методами по амплификации; все они были идентифицированы положительно при повторном тестировании.
(ν) Pfyffer et al., J. Clin. Micro. 32:918-923 (1994) проводили осаждение двумя разными способами - NALC и SDS, а амплификацию всех осадков с помощью Gen-Probe метода. Каждый образец анализировался только одним из двух методов.
(ξ) В работе Pfyffer et al., J. Clin. Micro. 32:918-923 (1994) 42 образца по методу NALC дали положительные результаты высевов и 39 образцов (92,9%) были идентифицированы путем амплификации. 36 образцов по методу SDS дали положительные результаты высевов и 35 образцов (97,2%) были идентифицированы путем амплификации. Однако, авторы учитывают положительные по амплификации/отрицательные по высеву результаты и приходят к выводу, что амплификация NALC осадков четко выявила 46 из 49 "истинно положительных" (93,9%), а амплификация SDS осадков четко выявила 38 из 39 "истинно положительных" (97,4%).
(π) Bodmer et al., J. Clin. Micro. 32:1483-1487 (1994) использовали схему амплификации Gen-Probe.
(†) Отмечены статьи, в которых авторы обсуждают ингибирование или ингибиторы как причину ложноотрицательных результатов.
 Отмечены статьи, в которых авторы обсуждают низкое число копий как причину ложноотрицательных результатов.
Отмечены статьи, в которых авторы обсуждают низкое число копий как причину ложноотрицательных результатов.
(§) Отмечены статьи, в которых авторы обсуждают явление статистических исключений или феномен, согласующийся со статистическими исключениями (что могло бы исключить "необъяснимые" результаты) или обсуждают возможность того, что появление ложных результатов усиливалось за счет того, что бактерии при росте образуют скопления групп клеток.
(¶) Отмечены статьи, в которых рассмотрены примеры образцов положительных по высеву/положительных по результату микроскопирования мазка/отрицательных по результату проведенной амплификации.
D. Исследования, представленные в таблице 1
Анализ опубликованных данных, представленных в табл. 1, говорит о том, что несмотря на общую идею или опробованное техническое воплощение в результатах наблюдается широкая вариабельность. В этих работах были использованы образцы, полученные абсолютно от всех возможных источников. Сюда входили мокрота, бронхиальные промывные воды, плевральный выпот, материалы аспирации желудка, спинномозговая жидкость (СМЖ), моча, образцы для тканевой биопсии, костный мозг, нарывы и эксудаты, кровь, сыворотка, брюшная жидкость и фекалии. Применялось две разные схемы амплификации: в 30 исследованиях использовалась исключительно ПЦР, в трех использовалась изотермальная, ретровирусного типа схема амплификации, запатентованная и коммерциализованная Gen-Probe (Jonas, V. et al., J. Clin. Micro. 31:2410-2416 (1993); Pfyffer et al., J. Clin. Micro. 32:918-923 (1994); Bodmer et al., J. Clin. Micro. 32:1483-1487 (1994)) и в двух исследованиях проводилось сравнение двух способов амплификации (Abe, C. et at., J. Clin. Micro. 31:3270-3274 (1993); Miller et al. , J. Clin. Micro. 32: 393-397 (1994)). В тридцати трех исследованиях внимание концентрируется на выявлении MTB, в одном - на выявлении M.avium (Irula, J.V. et al., J. Clin. Micro. 31:1811- 1814 (1993) и еще в одном - на выявлении M. paratuberculosis (van der Giessen, J.W.B. et al., J. Clin. Microbiol. 30: 1216-1219 (1992)). Ложноотрицательные образцы могли быть обнаружены среди большинства типов, независимо от задачи, способа обработки и метода амплификации.
Работы, упоминающиеся в табл. 1, колеблются по среднему количеству образцов от 7 до 1166. Корреляция с результатами высева была в диапазоне от 3 до 100%. В девяти из 35 публикаций (26%) сообщается о корреляции, равной 100%. Однако в большинстве работ, 7 из 9 (78%), число образцов меньше 100 (n < 100). Только в двух работах (Manjunath, N. et al., Tubercle 72:21-27 (1991): Pao, C.C. et al, J. Clin. Micro. 28:1877-1880 (1990)) использовалось более 100 образцов (следует отметить, что Pao, C.C. et al, J. Clin. Micro. 28: 1877-1880 (1990) могли повторно амплифицировать для того, чтобы перепроверить образцы, давшие отрицательный ответ: см. сноску F в табл. 1). С другой стороны, в 26 из 35 работ (74%) величина корреляции была меньше 100%. В этой группе в 20 из 26 (77%) исследований анализировалось более 100 образцов (n>100). Создается впечатление, что существует обратная зависимость между корреляцией результатов по амплификации-высеву и числом бывших в работе образцов: чем больше образцов использовалось в работе, тем ниже был коэффициент корреляции.
Показано, что в 32 из 35 исследований (91%), представленных в табл. 1, с помощью амплификации удавалось обнаружить ДНК микобактерий в образцах, давших отрицательный результат высевов (в двух из трех остальных исследований использовались только такие образцы, о которых заранее было известно, что они содержат микобактерии). В двух работах из 32 число образцов с положительными результатами высевов более чем в два раза превышало число образцов с отрицательными результатами высевов (Irula, J.V. et al., J. Clin. Micro. 31: 1811-1814 (1993): Pao, C.C. et al, J. Clin. Micro. 28:1877-1880 (1990)), и в трех работах - почти в два раза (Kolk, A.H. J. et al., J. Clin. Micro. 30: 2567-2575 (1992); Manjunath, N. et al. Tubercle 72:21-27 (1991); Miyazaki, Y. et al., J. Clin. Micro. 31:2228-2232 (1993)). В 31 работе авторы прямо утверждают или ссылаются на то, что при оптимальных условиях in vitro их системы позволяют уловить присутствие 10 копий или меньше. Чувствительность в остальных 4 работах колебалась от 15 до 40 копий (Altamirano, M. et al., J. Clin. Micro.30:2173-2176 (1992): Hermans. P.W.M. et al, J. Clin. Micro. 28: 1204-1213 (1990): Pao, C.C. et al., J. Clin. Micro. 28:1877-1880 (1990); Soini, H. et al., J. Clin. Micro. 30:2025-2028 (1992)). Образцы, показавшие отрицательные результаты высевов, но положительные по амплификации, легче поддаются объяснению, чем образцы, положительные по результатам высевов и отрицательные по данным амплификации: в отличие от высевов, для ПЦР не требуются живые микроорганизмы. Способы обработки, для которых известно, что они убивают бактерии, например химиотерапия, также могут дать результат, приведший к потере жизнеспособных бактерий, или низкое число копий в сочетании с пониженным числом жизнеспособных бактерий, все это вместе может повлиять на характер образцов. Независимо от системных параметров амплификация может иметь более высокую чувствительность, чем высевы или микроскопирование мазка.
Среди 35 исследований существует 8 разных методов, применяемых для изучения образцов, не прошедших предварительной обработки. В 17 исследованиях образцы подвергались воздействию N-ацетил-L- цистеина/NaOH (NALC), после чего использовались как для высевов, так и для амплификации, в 6 исследованиях использовался натрий додецилсульфат (SDS), в 3 исследованиях использовали гидроксид натрия (NaOH), в двух исследованиях - дитиотриэтол (DTT); оксалиновая кислота (OxAC), полиэтиленгликоль (ПЭГ) и Ficoll- hypaque (Fic/Hyp) градиентный метод - каждый из них использовался в одной из работ. Конкретно сравнивались методики с применением NALC и SDS (Pfyffer et al., J. Clin. Micro. 32:918-923 (1994). В трех публикациях избегали стадии осаждения и проводили амплификацию непосредственно в образце. Из 9 исследований, где было получено 100% соответствие, в трех использовали NALC, в трех проводили амплификацию непосредственно на образцах, в одной работе использовали SDS, еще в одной - OxAC, и еще в одной - ПЭГ. Следует отметить, что в 7 из этих работ анализировали не более 100 образцов (n<100), никаких выводов не было сделано по поводу того, как поддаются анализу образцы, не прошедшие предварительной обработки, нет четкой корреляции. В работе Pfyffer et al., J. Clin. Micro. 32: 918-923 (1994) сравнивали обработку NALC и SDS и приходят к выводу, что нельзя отдать предпочтение какому-то из методов.
Подготовка полученного осадка (или нативных образцов) для амплификации распадается на 7 основных категорий: (i) в 16 случаях используются различные способы экстракции органическими растворителями и осаждение спиртом (org/ppt), как описано Maniatis, Т. et al. ("Molecular Cloning A Laboratori Manual, " Cold Spring Harbor Laboratory, New York (1982), pp. 458-463); (ii) в двух случаях используют ферментный лизис и способ кипячения (Lz/Prk= >95o/15'), как описано у Higuchi, R. Amplifications 2:1-3 (1989), и в шести случаях просто прогревали образец; (iv) в трех случаях использовали хелатирующие вещества и стеклянные бусы (GuSCN/Si), как описано у Boom, R. et al. , J. Clin. Microbiol. 28:495-503 (1990), и в двух случаях использовали близкий способ осаждения ДНК на кремние; (v) в одном случае использовали фракционирование в градиенте сахарозы (Victor, T. et al., J. Clin. Micro. 30:1514-1517 (1992)); (vi) в одном случае использовали Chelex-100, как описано у Lamballerie, X. et al., Res. Microbiol. 143:758-790 (1992); и (vii) в одном случае (van der Giessen, J.W.B. et al., J. Clin. Microbiol. 30:1216-1219 (1992)) использовали колоночную хроматографию с учетом рекомендаций производителя (Vary, P. H. et al., J. Clin. Microbiol. 28:933-937 (1990)). Сумма превышает 35, потому что в исследованиях van der Giessen, J.W.B. et al., J. Clin. Microbiol. 30:1216-1219 (1992), Victor et al., J. Clin. Micro. 30: 1514-1517 (1992), Wilson, S.M. et al., J. Clin. Micro. 31:776-782 (1993) и Abe, C. et al., J. Clin. Micro. 31:3270-3274 (1993), Miller et al., J. Clin. Micro. 32:393-397 (1994) и Pfyffer et al., J. Clin. Micro. 32:918-923 (1994) работали с образцами более чем одним методом и дали заключение по каждому методу. В 8 исследованиях из 9, в которых авторы отмечают 100% корреляцию, ДНК выделяли методами с использованием органических растворителей и осаждали спиртом. Однако, в 8 работах, где также использовали методы экстракции органическими растворителями, величина корреляции колебалась от 55,9% до 98,4%. В одной из работ, где заявлено о 100% корреляции, применяли кипячение (Kocagoz, T. et al., J. Clin. Micro. 31:1435-1438 (1993)). В противоположность этим результатам, в 6 исследованиях, где использовали тот же метод, заявлено о корреляции от 78,9 до 95,9%. В 7 исследованиях фактически сравниваются методы приготовления осадка после высева для амплификации. Их выводы отличаются: Pierre, C. et al., J. Clin. Micro. 29:712-717 (1991) подбирал условия экстракции органическими растворителями/осаждения спиртом; Wilson, S. M. et al., J. Clin. Micro. 31:776-782 (1993) просто обрабатывали хлороформом; Folgueira, L. et al., J. Clin. Micro. 31:1019-1021 (1993) предпочитали ферментный лизис/кипячение; Kocagoz, T. et al., J. Clin. Micro. 31: 1435-1438 (1993) и Sritharan, V. et al. Mol. Cell. Probes 5:385-395 (1991) использовали простой метод кипячения; Forbes, B.A. et al., J. Clin. Micro. 32:1688-1694 (1993) и Buck, G.E. et al., J. Clin. Micro. 30:1331-1334 (1992) определили сонацию как оптимальный метод (только в работе Kocagoz, T. et al. , J. Clin. Micro. 31:1435-1438 (1993) достигнуто 100% корреляции). Могло бы показаться, что присутствие ложноотрицательных результатов не только не зависит от использовавшегося метода подготовки образца для амплификации, но спорно также все по поводу этих результатов.
Два исследования фактически сравнивают методы амплификации: Abe, C. et al. , J. Clin. Micro. 31:3270-3274 (1993) показали, что Gen-Probe метод был незначительно лучше метода ПЦР (см. сноску k в табл. 1), в то время как Miller et al., J. Clin. Micro. 32:393-397 (1994) пришли к противоположному выводу (см. сноску (μ) в табл. 1). Очевидно, что не существует способа амплификации, который мог бы дать существенный прорыв в клинической диагностике ТБ инфекций.
Е. Ингибиторы ПЦР
Среди публикаций, в которых сообщается, что корреляция составляет меньше 100%, в 17 работах ссылаются на "ингибиторы" амплификации как фактор, способствующий получению ложноотрицательных результатов (см. в табл. 1 ссылки на этих авторов, помеченные значком (†)). Кровь (Panaccio, М. et al., Nucl. Fcid Res. 19: 1151 (1991), фекалии (Allard, F. et al., J. Clin. Microbiol. 28: 2659-2667 (1990), мокрота (Hermans, P.W.M. et al., J. Clin. Micro. 28: 1204-1213 (1990): Shawar, R.M. et al., J. Clin. Micro. 31:61-65 (1993)) и моча (Khan, G. et al., J. Clin. Pathol. 44:360-365 (1991) содержат ингибиторы ПЦР. Кроме того, что касается мокроты, бронхиальных промывных вод и трахеальных выделений, наблюдается прямая корреляция между вязкостью образца (слизистая консистенция) и степенью заболевания: у пациентов с прогрессирующими стадиями туберкулеза наиболее вязкая мокрота и взятые пробы имеют наибольшую вероятность того, что в них сохранились ингибиторы амплификации. Hermans, P. W.M. et al., J. Clin. Micro. 28:1204-1213 (1990) и Shawar, R.M. et al., J. Clin. Micro. 31:61-65(1993) показали понижение чувствительности в 5 - 20 раз и, соответственно, только в 5 раз в присутствии мокроты.
Из 42 способов, представленных в табл. 1, только в двенадцати не предусмотрена замена буфера. Например, экстракция органическими растворителями/осаждение, промывка осадка, или способы, использующие хелатирующие вещества (GuSCN/Si), все нуждаются в замене буфера на некоторой стадии. Сонация осадка, однако, не требует замены буфера. Ни в одном из двенадцати методов не достигалось 100% корреляции и в девяти случаях из них ссылаются на ингибиторы как фактор, влияющий на получение ложноотрицательных результатов. Ингибиторы поступают как от образца, так и от растворов, применяемых при обработке, и оба источника являются существенным препятствием для применения этих способов идентификации в клиниках.
F. Низкое число копий, статистические выпадения и "необъяснимые" результаты
Статистические выпадения, также называемые "статистическая погрешность", обусловлены низким числом копий; для образца с очень низким числом копий, из которого должна отбираться аликвота, существует риск, что некоторые аликвоты не будут содержать копий. Например, если в затравке содержится 10 копий на миллилитр и взяты аликвоты объемом по 100 мкл, то в некоторых из них материал может отсутствовать. Такие аликвоты, которые будут интерпретироваться как ложноотрицательные, на самом деле будут "отрицательные по результату амплификации". В восьми исследованиях, представленных в табл. 1, описаны случаи, которые могли бы быть объяснены таким типом погрешности (см. ссылки на авторов, помеченные значком (§) в табл. 1). Как дискуссируется далее, это явление в значительной степени осложняется за счет агрегирования бактерий.
Среди публикаций, в которых указаны величины корреляции менее 100%, в 15 ссылаются на "низкое число копий" как на непосредственную причину ложноотрицательных результатов (см. ссылки на авторов, помеченные значком  в табл. 1). Однако, в 6 из 15, а также в 3 других, образцы были отрицательны по результату амплификации и в то же время положительны по
в табл. 1). Однако, в 6 из 15, а также в 3 других, образцы были отрицательны по результату амплификации и в то же время положительны по
результату высева и микроскопированию мазков (см. ссылки на авторов, помеченные значком  в табл. 1). Сообщалось, что предел определения методом микроскопирования при окраске мазка на кислотоустойчивость составляет от 7800 до 9500 бактерий на миллилитр мокроты (Hobby, G.L et al., Antimicrob. Ag. Chemother. 4:94-104 (1973); Yeager, H. et al., Amer. Rev. Resp. Dis. 95: 998- 1004 (1967)). Clarridge, J. E. et al., J. Clin. Micro. 31:2049-2056 (1993) представил обширный анализ ложноотрицательных результатов (см. табл. 7 по этой публикации). Из 37 детально проанализированных ложноотрицательных проб в 26 были выпадения результатов, а 11 были "истинно" ПЦР отрицательными. Девять из этих 37 дали положительный результат по микроскопированию мазков: 4 из этих 9 содержали ингибиторы, 3 не анализировались на наличие ингибиторов, 2, как было показано, не содержали ингибиторов. Из этих двух последних один был действительно ПЦР отрицательным. Shawar, R.M. et al., J. Clin. Micro. 31:61-65 (1993) также описал образцы, положительные по результату высева/положительные по микроскопированию мазков/отрицательные по ПЦР, которые не содержали ингибиторов. Если образец не содержит ингибиторов и является положительным по результатам высевов и по микроскопированию мазков, то полученный результат действительно может быть объяснен "низким числом копий". Shawar, R. M. et al. , J. Clin. Micro. 31:61-65 (1993) относит эту группу ложноотрицательных образцов к "необъяснимым".
в табл. 1). Сообщалось, что предел определения методом микроскопирования при окраске мазка на кислотоустойчивость составляет от 7800 до 9500 бактерий на миллилитр мокроты (Hobby, G.L et al., Antimicrob. Ag. Chemother. 4:94-104 (1973); Yeager, H. et al., Amer. Rev. Resp. Dis. 95: 998- 1004 (1967)). Clarridge, J. E. et al., J. Clin. Micro. 31:2049-2056 (1993) представил обширный анализ ложноотрицательных результатов (см. табл. 7 по этой публикации). Из 37 детально проанализированных ложноотрицательных проб в 26 были выпадения результатов, а 11 были "истинно" ПЦР отрицательными. Девять из этих 37 дали положительный результат по микроскопированию мазков: 4 из этих 9 содержали ингибиторы, 3 не анализировались на наличие ингибиторов, 2, как было показано, не содержали ингибиторов. Из этих двух последних один был действительно ПЦР отрицательным. Shawar, R.M. et al., J. Clin. Micro. 31:61-65 (1993) также описал образцы, положительные по результату высева/положительные по микроскопированию мазков/отрицательные по ПЦР, которые не содержали ингибиторов. Если образец не содержит ингибиторов и является положительным по результатам высевов и по микроскопированию мазков, то полученный результат действительно может быть объяснен "низким числом копий". Shawar, R. M. et al. , J. Clin. Micro. 31:61-65 (1993) относит эту группу ложноотрицательных образцов к "необъяснимым".
G. Разделение микобактерий при центрифугировании
Способность микобактерий хорошо держаться на поверхности жидкости была известна с 1924 года (Andrus, P.M. et al., Am. Rev. Tuberc. 9:99 (1924)). Затем в нескольких исследованиях была объяснена сложность осаждения микобактерий (Hanks, J.H. et al., J. Lab. Clin. Med. 23:736-746 (1938); Hata, Jr., D. et al. , Dis. Chest 18:352-362 (1950); Klein, G.C. et al., Am. J. Clin. Pathol. 22:581-585 (1952): Ratman, S. et al., J. Clin. Microbiol. 23:582-585 (1986); Rickman, T.W. et al., J. Clin. Microbiol. 11:618-620 (1980); Robinson, L et al. , J. Lab. Clin. Med. 27:84-91 (1941)) a, в ряде примеров, культивирование супернатанта было принято как стандартная практика.
Хотя в нескольких исследованиях сообщается, что фракция супернатанта содержит материал, положительно фиксируемый при микроскопировании (Hanks, J. H. et al., J. Lab. Clin. Med. 23:736- 746 (1938); Rickman, T.W. et al., J. Clin. Microbiol. 1 1:618-620 (1980)), другое исследование показывает, что у 88,8 и 82,4% от всех центрифугировавшихся образцов при скорости 2000 и 3000 об/мин, соответственно, высевы из супернатантов дали положительный ответ (Klein, G.C. et al., Am. J. Clin. Pathol. 22:581-585 (1952)). Кроме того, в этой же работе показано, что у 2,2% и 2,7% от числа образцов, центрифугировавшихся при 2000 и 3000 об/мин, соответственно, высевы из осадков давали отрицательный результат, а высевы из супернатантов - положительный. Анализирование фракций супернатанта также обсуждается в современных лабораторных руководствах (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S. Department of Health and Human Service, Center for Disease Control, (1985) pp. 31-46); Sommers, H.M. et al., "Mycobacterium", in: Manual of Clinical Microbiology, E.H. Lennette et al., eds., 4th ed., Am. Soc. Microbiol., Washington, D.C. (1985), pp.216-248).
Можно предполагать, что обратная зависимость между размером пробы и корреляцией объясняется эффектом флотации. Более крупный образец нуждается в обработке промывкой. Во время промывки возрастает временной интервал между обработкой первого и последнего образца. Если время увеличивается, на флотацию приходится больше времени, чтобы повлиять на результат. Высказано предположение, что причиной флотации является высокое содержание липидов у этих бактерий (Silverstolpe, L. Nord. Med. 40.48:2220- 2222 (1948)).
H. Влияние клеточной стенки и поверхностного натяжения на выделение микобактерий
Природа клеточной стенки микобактерий является причиной их высокой жизнестойкости. Микрофотографии показывают очень сложное строение стенки, толщина которой 30-40 нм (Rastogi, N. et al., Antimicrob. Agents Chemother. 20: 666-677 (1981)). Липиды составляют 60% сухого веса клеточной стенки (Joklik, W. K. et al., Zinsser Microbiology 20th edition, Appleton & Lange, Norwalk, CT (1992), pp.499).
Клеточная стенка микобактерий состоит из трех различных слоев: (i) пептидогликан, (i) арабиногалактан и (iii) гликолипиды (для получения полной информации по строению клеточной стенки см. McNiel, M.R. et al., Res. Microbiol. 142:451-463 (1991)). Миколовые кислоты, являющиеся чрезвычайно гидрофобными и составляющие основу углеводородных цепей (Σ = C76-C80), составляют значительную часть арабиногалактанового и гликолипидного слоев. Структура и видовое значение миколовых кислот рассматриваются в работе Takayama, K. et al. , "Structure and Synthesis of Lipids", в: The Mycobacteria: a Source Book, Part A, G. P. Kubica et al., eds., Marcel Dekker, Inc., New York, NY (1984), pp. 315-344. Клетки микобактерий в значительной степени инкапсулированы воском.
М. tuberculosis в процессе роста формирует "тяжи" (тяжи в процессе роста группируют на себе или агрегируют большое число бактерий), и существует прямая зависимость между вирулентностью MTB и способностью образовывать тяжи (Joklik, W. K. et al., Zinsser Microbiology 20th edition, Appleton & Lange, Norwalk, CT (1992), pp. 503). Бактерии, относящиеся к MAC комплексу, в составе клеточной стенки имеют дополнительные гликолипидные (C-микозидные) компоненты (различия серологических типов рассматриваются в работе Brennan, P. J. Rev. Infect. Dis. 11 (Supp. 2): s420-s430 (1989)). При культивировании в отличие от бактерий MTB комплекса, которые формируют плотные образования, M. avium растет более диффузно, в виде отдельных клеток (Dubos, R.J. et al., J. Exp. Med. 83:409-423 (1946)).
Dubos, R. J. Proc. Soc. Exp. Biol. Med. 58:361-362 (1945) наблюдал, что характерный пленчатый рост MTB может быть изменен при внесении в среду полиоксиалкильного производного моностеарата сорбитана (Tween 60: CASRN. 9005-67-8). Это наблюдение было позднее продолжено с целью показать, что другие близкие производные могут вызвать у MTB "быстрый", "диффузный" и "погруженный" рост (Dubos, R. J. et al., J. Exp. Med. 83:409-423 (1946)). Было обнаружено, что Tween 80 (CAS® N. 9005-65-6) имеет наибольшую активность. Эти авторы приходят к выводу, что погруженный рост происходил благодаря "увлажнению" поверхности клеток. Понятие "увлажнение" применяется исключительно в контексте поверхностного натяжения. Смысл данной работы состоял в том, что пленчатый рост, образующийся в результате поверхностного натяжения между восковым чехлом бактерий и водной средой, нарушался при добавлении Tween 80 за счет прерывания физического взаимодействия.
Если поверхностное натяжение удерживало бактерии на поверхности, это, в сочетании с образованием тяжей, могло объяснить ошибочные результаты: тяжело больные пациенты заражены бактериями, имеющими тенденцию образовывать тяжи. Кроме того, крупные тяжи дают положительный результат при микроскопировании мазков и при высевах, что быстро позволяет дать положительное заключение. Крупные тяжи могли бы также усилить статистическую погрешность, поскольку крупный тяж может быть воспринят как одна копия, но потенциально содержит тысячи копий. Следовательно, как считает Klein, G.C. et al., Am. J. Clin. Pathol. 22:581-585 (1952), микроорганизмы могли бы быть легко выброшены вместе с супернатантом, способствуя таким образом явлению статистической погрешности. Кроме того, образование тяжей могло бы вызвать такое разделение, при котором бы наблюдались положительные результаты микроскопирования мазков и высевов и отрицательные результаты амплификации.
Если причиной ложных результатов были поверхностное натяжение и агрегация клеток, то их можно было бы преодолеть добавлением неионных детергентов, и представляется логичным, что эти реагенты могли бы улучшить корреляцию с результатами высевов. В пяти исследованиях, представленных в табл. 1, используются неионные детергенты для того, чтобы промыть осадок до проведения амплификации (Clarndge, J. E. et al., J. Clin. Micro. 31:2049-2056 (1993); Irula, J.V. et al., J. Clin. Micro. 31:1811-1814 (1993): Kolk, A.H. J. et al. , J. Clin. Micro. 30:2567-2575 (1992): Shawar, R.M. et al., J. Clin. Micro. 31: 61-65 (1993) и Sritharan, V. et al., Mol. Cell. Probes 5: 385-395 (1991)). В этих работах корреляция с результатами высевов совпадает в диапазоне от 78,9 до 95,5%. Уже в 1941 году было обнаружено, что вещества, понижающие поверхностное натяжение, не способствовали повышению выделяемости при центрифугировании (Robinson, L. et al., J. Lab. Clin. Med. 27:84-91 (1941)). Следовательно, ясно, что изложенные здесь методические подходы не дают дополнительного преимущества, чтобы включить данные детергенты в буферы для промывки.
I. Проблемы, связанные с бактериями MAC комплекса
Из 35 исследований, представленных в табл. 1, в 27 используют MTB-специфические последовательности для амплификации и/или определения. В 6 используются ген-специфичные праймеры, но существующие разработки предпочтительно направлены на амплификацию микроорганизмов ТВ комплекса. Только в работе Irula, J.V. et al., J. Clin. Micro. 31:1811-1814 (1993) обращается специальное внимание на M. avium, и только в работе van der Giessen, J.W.B. et al., J. Clin. Microbiol. 30:1216-1219 (1992) - на M. paratuberculosis, Irula использовал специальную методику выделения, затрудняющую сравнение. Однако, когда PBMC были выделены, они были подвергнуты стадии промывки в Tris/EDTA/Triton X-100: если PBMC лизировались в ходе промывки, бактерии могли быть выброшены вместе с супернатантом. Несмотря на это, можно было ожидать, что способы выделения PBMC чрезвычайно эффективны. В работе van der Giessen сравнивались три способа на основе ПЦР (McFadden, J.J. et al., Mol. Microbiol. 1:283-291 (1987); van der Giessen, J. W. B. et al., J. Med. Microbiol. 36:255-263 (1992); Vary, P.H. et al., J. Clin. Microbiol. 28:933-937 (1990)), созданных для того, чтобы определять М. paratuberculosis в фекалиях крупного рогатого скота (один из этих способов является доступным коммерческим набором от IDEXX (Vary, P. H. et al., J. Clin. Microbiol. 28:933-937 (1990)). Эти результаты были намного хуже, чем что бы то ни было еще из представленного в табл. 1, и искусственно занижены. Эти результаты показывают, что микроорганизмы MAC комплекса с их дополнительными липофильными компонентами представляют дальнейшие неясные проблемы для проведения амплификации.
J. Естественные свойства микобактерий, мешающие внедрению способов амплификации
Очевидно, что существует две основные причины ложноотрицательных результатов. Во-первых, ингибиторы в больших количествах содержатся в образцах самых разных типов и используемые в работе растворы также играют роль в изменении эффективности реакции амплификации. Вторая категория имеет отношение к естественному свойству микобактерий. Поскольку число таких свойств огромно, их влияние на чувствительность так сильно превалирует, что Noordhoek, G. T. et al., J. Clin. Micro. 32:277-284 (1994) заключают: "...мы не будем спекулировать по поводу возможных факторов, которые могли бы объяснить крайнее расхождение данных по чувствительности ПЦР среди семи лабораторий... ".
Образование тяжей, флотация данных микроорганизмов не позволяют разделять их эффективным способом и поэтому бактерии выбрасываются вместе с супернатантом. Крайнее проявление данной ситуации вызывает "необъяснимые" результаты: образец, давший положительный ответ по результатам высева и по микроскопированию мазка, может оказаться действительно отрицательным по данным множественных амплификаций и при этом не содержать ингибиторов. Очевидно, что источник и природа таких свойств требует полного разъяснения. Однако, это явления такого порядка, причина которых кроется в сущности самих микроорганизмов, и приводимые в данной работе методы разрешают существующие проблемы.
Сущность изобретения
Очерчивание круга проблем, входящих в данные методы приготовления микобактерий для секвинирования и высевания и осознание потребности в быстром, недорогом но корректном методе для исследования биологических и неорганических образцов с целью определения микобактерий, автора привело к созданию метода экстракции образцов для выделения микобактерий.
Эти исследования нашли свое завершение в методах обработки микобактерий с целью их определения, включая определение путем высевов, и особенно определение с помощью методов, таких как амплификация, и особенно амплификация нуклеиновых кислот, которые в первую очередь эффективно преодолевают существующие в этой области проблемы, такие как агрегация и флотация, которые ответственны за ложноотрицательные результаты и статистические выпадения. Изобретатель обнаружил, что некоторые цвиттерионные детергенты, именуемые здесь "SB-18-подобные детергенты", неожиданно и необъяснимо диспергируют микобактерии и очевидно снимают несмачиваемость поверхности этих бактерий. Для микобактерий MTB комплекса, у которых агрегирование является весьма серьезной проблемой, возможность диспергировать представляет первостепенную движущую силу на пути улучшения эффективности выделения бактерий. Для других микобактерий, которые растут в виде одиночных клеток, например, для бактерий MAC комплекса, принципиальным моментом для улучшения способов выделения является противодействие флотации путем аккумулирования детергента. Для таких микобактерии, как MTB, так и MAC комплекса, включение таких детергентов в методику работы с образцом помогает избежать схожих проблем в данной области, которые приводили к ложноотрицательным результатам амплификаций.
Изобретатель также обнаружил, что когда микобактерии дегазируются, другие детергенты, которые раньше не давали положительного эффекта в снятии эффекта флотации, стали пригодными для улучшения выделения. По-видимому, дегазирование уничтожает флотацию до того состояния, когда поверхностное натяжение остается единственным фактором, связывающим микроорганизмы с поверхностью среды. При создании подходящих условий огромное большинство детергентов способно преодолеть поверхностное натяжение. Эти детергенты не снимают агрегирования бактерий MTB комплекса и, следовательно, не снимают феномена погрешности образца, но они, тем не менее, повышают эффективность осаждения центрифугированием. Изобретатель показал, что для бактерий MAC комплекса и для других бактерий, растущих преимущественно в виде отдельных клеток, есть класс детергентов, которые при комбинировании с частичным дегазированием улучшают выделение микроорганизмов с возросшей результативностью.
Способы по данному изобретению применимы в целом для любых микроорганизмов, особенно для тех, которые содержат миколовую кислоту или липиды на основе близких к ней кислот в составе клеточной стенки, например, коринобактерии, у которых липиды содержат кориномиколовую кислоту, и нокардии, у которых липиды содержат нокардиомиколовую кислоту. Способы в рамках данного изобретения также в целом применимы для работы с биологическими образцами по выявлению любых микроорганизмов.
Перечень фигур чертежей
Фиг. 1 представляет дот-блоттинг, на котором показано ингибирование ПЦР при экстракции образца NALC/NaOH раствором.
Фиг. 2 показывает схему постановки опыта: способ in vitro, описывающий кратко последовательность действий по определению микобактерий в клинических образцах.
Фиг. 3 показывает эффект от добавления к раствору для экстракции 0,1% Тритона Х-100.
Фиг.4 представляет схему исследования по агрегированию: методика разработана, чтобы оценить способность детергентов диспергировать Mycobacterium tuberculosis. Данные по трем представленным условиям показаны: вода, 0,1% Твин 80 и 2mM SB-18.
Фиг. 4A показывает результаты дот-блот гибридизации проб с целью диспергировать микобактерии. Показаны данные, полученные в трех представленных условиях опыта: вода, 0,1% Tween 80 и 2mM SB-18.
Фиг. 5 показывает результат обработки in vitro Mycobacterium avium и Mycobacterium tuberculosis, при условии включения в раствор для экстракции SB-18.
Фиг.6 показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены ионные гомологи SB-18.
Фиг.7 показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены различные детегренты SB-серии.
Фиг. 7A показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены различные детегренты SB-серии.
Фиг.8 показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены кококарбоксибетаины (т.е. SB-подобные детергенты).
Фиг.9 показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены бетаины, имеющие различия в комбинациях зарядов и структурных связях.
Фиг. 9A показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены бетаины, имеющие различия в структурах "мостиков".
Фиг. 9B показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены бетаины, имеющие различия в алкилах и структурах алкильных "связей".
Фиг. 9C показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены бетаины, имеющие гидрофобные домены, происходящие исключительно из природных масел.
Фиг. 9D показывает результаты дот-блот гибридизации in vitro при обработке Mycobacterium tuberculosis, когда в раствор для экстракции добавлены бетаины, имеющие гидрофобные домены, происходящие исключительно из природных масел.
Фиг. 10 показывает, что предсказуемые модификации в условиях анализа позволяют бетаинам, которые были или скомпрометированы, или нефункциональны, функционировать in vitro при обработке Mycobacterium tuberculosis.
Фиг. 11 представляет схему последовательных стадий эксперимента по изучению воздействия вакуума на обработку in vitro.
Фиг. 11A показывает эффект воздействия вакуума на обработку in vitro как Mycobacterium tuberculosis, так и Mycobacterium avium.
Фиг. 12 показывает, что длительное дегазирование Mycobacterium tuberculosis требуется для того, чтобы позволить Triton X-100 повысить эффективность выделения при обработке in vitro.
Фиг. 13 показывает, что Brij 96, приблизительно - октадециловый неионный структурный гомолог Tween 80, и SB-18, проявляет SB-18 подобную активности.
Фиг. 14 показывает, что ограниченное вакуумное дегазирование Mycobacterium avium раскрывает класс октадецилоподобных детергентов, которые имеют некоторую степень эффективности в улучшении сбора бактерий путем центрифугирования.
Фиг. 15 описывает устройство аппарата по дегазированию в вакууме, используемого в данных экспериментах.
Фиг. 16 представляет схему стадий экспериментального метода, созданного для сравнения способа обработки с применением SB-18, очерченного на фиг. 11, со способом обработки с использованием NALC/NaOH.
Фиг.16A представляет кривые роста Mycobacterium tiberculosis при проведении метода, представленного на фиг. 16.
Фиг. 16B представляет результаты амплификации, когда образцы, показанные на фиг. 16, обрабатываются по ПЦР.
Сведения, подтверждающие возможность осуществления изобретения
Данное изобретение представляет способ обработки образцов для определения присутствия в них или для высева из них микроорганизмов, которые характеризуются способностью к флотации (плавучесть) в жидкой среде, и/или образованием тяжей или агрегированием в процессе роста.
Под термином "образец" подразумеваются любые материалы, которые будут исследоваться или высеваться с целью определения в них микобактерий, особенно из рода Mycobacterium, включая, но не ограничиваясь этим, биологические пробы и пробы, полученные не от живых объектов.
Под термином "биологическая проба" подразумевается образец, взятый от животного (в том числе от человека) или от растения. Биологические пробы от животных, представляющие особый интерес, включают пробы, взятые у жвачных животных (таких, как коровы или овцы), рыб и птиц. Под этим термином также подразумеваются образцы пищевых продуктов, прошедшие или не прошедшие обработку (включая, например, яйца, тушки кур, молоко и другие молочные продукты), растения и культуры клеток (таких как культуры клеток моноцитов или фибробластов).
Под термином "небиологическая проба" подразумевается проба, полученная из неживого источника, например проба из окружающей среды, такая, как проба почвы, воды, опилки или проба воздуха.
Под термином "осадок" подразумевается образец, который обрабатывался или очищался таким способом, при котором происходит концентрация микобактерий, позволяющая таким образом использовать образец для последующего процесса определения.
"Промывка" означает, что анализируемый образец помещается в раствор, содержащий, как правило, октадецил-подобный детергент или SB-18-подобный детергент.
Под понятием "структуры миколовых кислот" подразумеваются β-оксикислоты, замещенные в α-положении алифатической цепью среднего размера, как описывается в статье (Goren, М.В. Bact. Rev. 36:33-64 (1966), которая включена в данную работу в виде ссылки).
Под понятием "SB-подобный детергент" подразумевается бетаин, обладающий способностью физически способствовать требуемому качественному сбору бактерий, содержащих структуры миколовых кислот, для последующего определения. Термин "SB-подобный детергент" является синонимом термину "бетаиноподобный". SB- подобные детергенты по данному изобретению обладают способностью диспергировать тяжи (и группы скопления) микобактерий и/или компенсировать плавучесть микобактерий. Диспергирование способствует определению за счет увеличения вероятности того, что аликвоты, взятые для определения, будут содержать микроорганизмы. SB-18-подобные детергенты, способные диспергировать тяжи, имеют алкильную цепочку длиной более чем 16 атомов углерода, и алкильная цепочка с 18-20 атомами углерода наиболее предпочтительная. SB-18-подобные детергенты по данному изобретению также обладают способностью облегчать сбор клеток микобактерий, таких, например, как представители MAC комплекса, которые не растут в виде пленки, за счет снижения до некоторой степени естественной плавучести, преимущественно путем облегчения попадания детергента в бактериальные клетки. SB-18-подобные детергенты, способные снизить плавучесть, предпочтительно имеют алкильную цепь, содержащую более 12 атомов углерода, и алкильная цепочка с 16-20 атомами углерода наиболее предпочтительна.
Под термином "SB-18-подобная активность" подразумевается способность как способствовать разрушению тяжей (и тем самым переходу клеток бактерий в раствор с образованием суспензии), так и способность снижать плавучесть и тем самым существенно облегчать сбор таких микроорганизмов путем центрифугирования, или и то, и другое. Например, для тех микроорганизмов, которые формируют скопления в процессе роста, как например MTB, обработка SB-подобным детергентом облегчает определение как за счет снижения плавучести, так и за счет диспергирования микроорганизмов равномерно на протяжении процесса выделения. Эта функциональная двойственность и является "SB-18-подобной активностью" детергента. Примерами SB-18- подобных детергентов с такими свойствами являются любые бетаины, которые обладают перечисленными свойствами, и описанные здесь либо в качестве примера, либо по аналогии со структурами приведенных в примерах бетаинов, включая CB-подобные, SB-подобные, HSB-подобные, PB-подобные, StB-подобные, PhB-подобные, So-подобные, Rev-В- подобные, AO-подобные, cAB-подобные и ImB-подобные детергенты, имеющие структуры, указанные в табл. 2 и 3 и/или описанные в этой работе.
Под понятием "дегазирование" подразумевается помещение образца или осадка под вакуум на такой срок и при такой температуре, чтобы созданные условия были достаточны для разрушения естественной плавучести микроорганизмов, содержащих структуры миколовых кислот, такие, например, как миколовые кислоты, нокардиомиколовые кислоты или кориномиколовые кислоты. Не предвосхищая последующие объяснения, допускается, что дегазирование удаляет заключенный в клеточных стенках CO2 и тем самым частично снижает естественную плавучесть этих организмов. Предпочтительным является помещение образца в условия: давление 600 мм рт. ст., температура 40-42oC, длительность обработки 60 мин, которые облегчают сбор микроорганизмов, если используются SB-18-подобные детергенты; однако, дегазирование в течение длительного периода при такой же величине разрежения воздуха и при такой же температуре позволит облегчить определение любым детергентам.
Таким образом, согласно способам по данному изобретению плавучесть может быть нарушена тремя различными путями: накапливанием детергента внутри клетки, дегазированием или и тем и другим одновременно. Существуют определенные детергенты, не относящиеся к бетаинам, которые облегчали определение в присутствии или в отсутствие дегазирования. Например, неионные линейные детергенты, такие как Brij 96 (полиоксиэтилен 10 олеиловый эфир (C18:1E10) ( CAS® N. 9004-98-2)) имеет диаметр головки головной группы, близкий к аналогичной величине у бетаина. Эти "палочковидные" детергенты не так пространственно защищены от проникновения в клетки (по сравнению с Твином 80), так как могут быть быстро изолированы внутри клетки, компенсируя таким образом плавучесть. Таким образом понятие "палочковидные", в данной работе, относится к неионным детергентам, которые проявляют SB-18- подобную активность без проведения дегазирования способами по данному изобретению. Можно было бы ожидать, что другие неионные палочкообразные детергенты компенсируют плавучесть таким же образом. Было отмечено, что другие детергенты, такие как октадециловые катионные детергенты бромистый октадецилтриметиламмоний (TMA-18: CAS® N. 1120-02-1) или хлористый бензилдиметилоктадециламмоний (BenzDMA-18), облегчают поиск в большей степени, чем их гомологи с более короткой цепью, но только на фоне дегазирования. Не предвосхищая последующие объяснения, допускается, что определенные детергенты, исходя из их "октадецилоподобной" структуры, более легко накапливаются в клетке.
Термин "палочкообразный", в этой работе, относится к молекуле детергента, близкого по "осевой пропорции" к бетаину. Осевая пропорция определяется как: "Соотношение большей оси к меньшей." (McGraw-Hill Dictionary of Scientific and Technical Terms, 5th ed., Parker, S.P., ed. Mcgraw-Hill, Washington, D.C. (1994), p. 168). За большую ось принимается протяженная алкильная цепь (например, гидрофобный домен: R1 как обозначено в табл. 2), а за меньшую - диаметр головной группы. Большая и меньшая оси могут, по определению, быть взаимоперпендикулярными.
Под термином "октадецилоподобные" подразумевается детергент, молекулы которого обладают октадецилоподобной частью, желательно состоящей из 12-20 атомов углерода и в наилучшем варианте - 18-20 атомов углерода, похожей на SB-18-подобную октадециловую часть, что может быть использовано в способах по данному изобретению таким образом, что проявляется активность как у SB-18, но не требуется присутствие для проявления эффективности цвиттерионной функции. Можно было бы включить, но не ограничиться этим, палочкообразные детергенты, которые не требуют дегазирования, или катионные детергенты, которые также не требуют дегазирования, для проявления SB-18-подобной активности. Октадецилоподобные детергенты, используемые в способах по данному изобретению, полезны при работе с бактериями MAC комплекса; октадецилоподобные детергенты включают SB-18-подобные детергенты. Не все октадецилоподобные детергенты являются SB-18- подобными детергентами, но все SB-18-подобные детергенты являются октадецилоподобными детергентами.
Под обозначением "CAS® Number" или "CAS® N." имеется в виду регистрационный номер Службы химических сокращений (Chemical Abstracts Service Registry Number, 2540 Olentangy River Road, PO Box 3012, Columbus, Ohio). Список всех CAS® N. и соответствующие им структуры даны в приложении в табл. 13.
Обработка образцов для определения
Как приведено здесь, так и в большинстве предпочтительных воплощений, микроорганизмами являются микобактерии. Как отражено в этой работе далее, способы по данному изобретению для обработки образца для последующего определения группирующихся между собой клеток микроорганизма, содержащего структуры миколовых кислот на наружной поверхности клеточной стенки, проиллюстрированы обработкой и определением микобактерий MTB комплекса. Далее на примере определения микобактерий MAC комплекса приводятся способы по данному изобретению для обработки образца для последующего определения группы микроорганизмов, содержащих структуры миколовых кислот на наружной поверхности клеточной стенки и способных к флотации. Хотя представители микобактерий специфически представлены здесь, следует понять, что излагаемые примеры и способы по данному изобретению применимы для работы с образцами, содержащими любой микроорганизм, имеющий сходные характеристики, а именно способность к флотации и/или образованию групп во время роста и/или содержащий структуры миколовой кислоты в составе клеточной стенки.
Согласно первому воплощению данного изобретения, образец (или, если обрабатывался, осажденный образец) обрабатывался путем помещения образца или осадка в среду, содержащую забуференный SB-18-подобный детергент или октадецилоподобный детергент, который обладает активностью SB-18, но в отсутствие дегазирования, палочкообразный детергент, например. В том случае, если микроорганизмы не растут группами, например, как микроорганизмы комплекса MAC, эта стадия облегчает сбор клеток бактерий за счет некоторой компенсации такого свойственного им признака как плавучесть. В том случае, если микроорганизмы растут группами, например, как микроорганизмы комплекса MTB, эта стадия облегчает сбор клеток бактерий как за счет некоторой компенсации плавучести, так и за счет диспергирования клеток, более равномерного на протяжении работы с раствором. В первом воплощении способов изобретения отсутствует стадия дегазирования и SB-18-подобный детергент используется для дезагрегирования скоплений микроорганизмов, особенно микобактерий, в частности относящихся к MTB комплексу.
По поводу второго воплощения изобретения следует отметить, что проба (или, если была проведена обработка, осажденный образец) обрабатывается на стадии дегазирования, например в вакууме. Во втором воплощении изобретения микобактерии, например относящиеся к MAC комплексу, просто дегазируются и затем анализируются. Состав промывного раствора во втором воплощении может быть, но не ограничивается этим, представлен водой или водным буферным раствором, как раскрывается далее.
Согласно второму воплощению изобретения, проба (или, если была проведена обработка, осажденный образец) обрабатывается как на стадии дегазирования, так и подвергается промывке с использованием детергента. В третьем воплощении в сочетании со стадией применения вакуума может быть использован любой детергент, но особенно желательно использовать октадецилоподобные детергенты, из которых лучший SB-18-подобный детергент. Кроме того, любой детергент, но особенно детергенты, относящиеся к октадецилоподобным детергентам, из которых лучший SB-18-подобный детергент, может быть использован на стадии промывки, которая может предшествовать, следовать за или проводиться одновременно со стадией дегазирования в зависимости от преследуемой цели. Третье воплощение особенно полезно для образцов, о которых неизвестно, содержатся ли в них микобактерии, или предполагается, что содержится по меньшей мере один вид, относящийся к MTB или MAC комплексу.
Жидкая среда, которая вносится к образцу или осадку, например такая, как промывочный буфер, дополнительно в случае необходимости, одновременно с желаемым детергентом, может содержать компоненты, такие как дитиотриэтол (ДТТ) и ферменты, такие, как гликозидазы и DNase, с целью облегчить растворение определенного биологического материала, такого как мокрота, и, с другой стороны, принять участие в устранении нежелательных свойств образца. ДТТ и ферменты, такие как гликозидазы и DNase, являются подходящими веществами для снижения вязкости биологических жидкостей.
Может быть выполнено более одной промывки, если в этом есть потребность, и такое многократное промывание может быть выполнено тандемно (одно за одним последовательно) или может перемежаться другими методическими операциями. На таких стадиях многократной промывки могут быть задействованы дополнительные реагенты. По мере того, как дополнительные методические стадии требуют включения дополнительных реагентов, таких, например, как реагенты для дальнейшей деконтаминации образца, подобные операции могут быть использованы. Такие многократные промывки могут использовать разные детергенты или каждый раз один и тот же. Сочетание разных детергентов, не относящихся к SB-18-подобным детергентам, на одной стадии промывки как правило приводило к снижению индивидуальной эффективности каждого из них. Однако, в случае, когда конкретная комбинация этих детергентов, содержащая SB-18-подобные или октадецилоподобные детергенты, сохраняет эффективность способов по данному изобретению, такая комбинация может быть использована.
В любом воплощении данного изобретения образец, предназначенный для анализа на присутствие микобактерий, может быть первоначально проэкстрагирован согласно любому стандартному биологическому методу, требуемому на первой стадии, например по одному из методов, рекомендованных Kent (Kent, P.T. et al. , "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 31-46), и особенно по методу выделения NALC/NaOH, как показано в примерах 2, 4 и 6. В первую очередь берется часть осадка для высева и приготовления мазка. Оставшийся осадок затем подвергается дальнейшей обработке с помощью любого из воплощений изобретения для дальнейшего определения с помощью амплификации (нуклеиновой кислоты или сигнала), или иммунодетекции или любым другим методом, способным распознать присутствие микобактерий в образце. Однако, как дискутируется далее и как изложено в примерах, такие манипуляции стандартными методами, используемыми в современной практике, весьма вероятно не способствуют должному сбору и/или определению микобактерий, находящихся в образце, особенно если микобактерии содержатся в небольшом количестве. Таким образом, хотя сами методы по данному изобретению, когда они применяются сами по себе, являются по существу количественными даже для небольшого числа микобактерий, находящихся в образце, при их комбинировании с применяемыми современными методиками, в которых не используется детергент или не производится предлагаемое в данном изобретении дегазирование, выделение микобактерий может быть неудачным. Можно модифицировать современные способы Kent, P.T. et al. [ссылка приведена выше], чтобы избежать их неэффективности при выделении микобактерий таким образом, например, как нейтрализация гидроксидом натрия или оксалиновой кислоты, или любым из способов деконтаминации, описанных Kent, P.T. et al. [ссылка приведена выше], с помощью подходящего реагента и затем добавить SB-18-подобный или октадецилоподобный детергент на стадии, предшествующей центрифугированию; или путем добавления соответствующим образом забуференного SB-18-подобного или октадецилоподобного детергента, таким образом, что он воздействует как нейтрализующий буфер на последующих стадиях согласно способам по данному изобретению до стадии центрифугирования; или путем простого дегазирования нейтрализованного образца до стадии центрифугирования. Подобные модифицированные способы могли бы за счет SB-18-подобной активности и/или за счет дегазирования улучшить эффективность выделения на стадии центрифугирования, если по-прежнему используются существующие сейчас методы.
Таким образом, в предпочтительном воплощении, как оно отражено в примере 13, для образца, анализируемого на наличие микобактерий, используется способ по данному изобретению прямого выделения микобактерий для определения путем амплификации непосредственно и/или для культивирования, без предварительной обработки какой- либо из стандартных предварительных методик. Далее, образец используется непосредственно, или просветляется, или освобождается от твердых частиц и/или ингибиторов, если это необходимо. При этом используется обычная методика разделения, известная на практике, такая как центрифугирование, фильтрация, гель-хроматография или ионнообменная хроматография. Образец непосредственно подвергается либо промывке с SB-18-подобным детергентом, вакуумной обработке для устранения плавучести, или им обоим, как описано в данной работе. Обработанные клетки эффективно собираются и осадок используется для определения присутствия искомого микроорганизма.
В первом воплощении, описанном выше, требуемое количество осадка образца помещается в подходящего размера емкость с промывочным буфером, который содержит SB-18-подобный детергент, на определенное время, при температуре и уровне перемешивания, достаточном для однородного дезагрегирования микобактерий во всем объеме раствора и для компенсирования плавучести. Такое перемешивание может, например, быть достигнуто за счет встряхивания или непосредственного помещения емкости, содержащей образец, на встряхиватель с целью ресуспендировать осадок (или суспендировать образец) в сильной степени, чтобы позволить детергенту эффективно разбить скопления микроорганизмов и добиться их диспергирования в жидкости до перехода к следующей стадии. Легкодостижимые температуры, например 37oC, способствуют дезагрегированию без ухудшения последующих методов определения и могут быть необходимы для предохранения SB-18-подобного детергента от преципитации. После получения однородной суспензии, особенно микобактерий, они могут быть собраны с использованием известных способов, таких как центрифугирование, и определены или исследованы и проанализированы, как описано далее.
Во втором воплощении образец или оставшийся осадок непосредственно ресуспендируется в воде или другой подходящей среде, не обязательно содержащей детергент, и дегазируется при нужных давлении и температуре в течение времени, достаточного для того, чтобы избежать естественной тенденции к плавучести микобактерий, особенно относящейся к комплексу MAC, в растворе. Затем микроорганизмы могут быть собраны и определены, как описывается далее.
В третьем воплощении, описанном выше, в значительной степени объединены первые два, за исключением того, что предшествует, происходит одновременно или продолжает дегазирование практически любым детергентом, но особенно такими, которые относятся к октадецилоподобным, и среди них особенное положение занимают SB- 18-подобные детергенты, которые могут применяться в составе среды для достижения диспергированности, а также для компенсации плавучести. Стадия дегазирования может предшествовать, следовать за промывкой с применением детергента, или стадия с применением детергента и дегазирование могут происходить одновременно. Применяющий данную технологию должен представлять, что для современных практикуемых технологий одновременная промывка и дегазирование образцов могут представить существенные риск с точки зрения безопасности.
Третье воплощение изобретения рассматривается далее в примере 10. Как показано в примере 10, если вакуумная обработка используется вместе с детергентной промывкой, тогда могут использоваться октадецилоподобные детергенты, предпочтительно SB-18-подобные детергенты, и самый предпочтительный - SB-18. Как показано далее в примере 10, при условиях, когда образец интенсивно дегазируется с целью в значительной степени ликвидировать естественную плавучесть, может применяться любой детергент. В данном воплощении образцы могут нуждаться (или нет) в просветлении или очистке: эти стадии могут быть как до, так и между или после желаемых стадий по вакуум-детергентной обработке. Как описано выше, микроорганизмы затем могут быть собраны и определены.
Таким образом, для выделения существенного количества микобактерий стандартные методы обработки образцов, такие, как например N-ацетил-L-цистеин/NaOH (NALC) метод, описанный в публикации Kent, P.T. et al., "Public Health Mycobacteriology, " в A Guide for the Level III Laboratory, U.S. Department of Health and Human Service. Center for Disease Control, (1985) pp. 31-46, практически не используется, или он должен быть модифицирован как описано выше, и образец или декантеминированный образец, непосредственно (i) подвергается промывке с использованием SB-18-подобного детергента, желательно SB-18, (ii) дегазируется или (iii) возможно комбинирование как промывки с использованием детергента, так и вакуумной обработки, в зависимости оттого, какого типа микобактерии предполагается исследовать.
Однако, по желанию, среди реализаций метода по данному изобретению образец, полученный от биологических жидкостей, включая, например, мокроту, цереброспинальную жидкость и мочу, на первой стадии следует центрифугировать, чтобы получить осадок. Например, образец может быть первоначально обработан любым из способов, описанных в публикации Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U. S. Department of Health and Human Service, Center for Disease Control, (1985) pp. 31-46, включенной в данную работу в виде ссылки, включая способ с использованием N-ацетил-L-цистеин/NaOH (NALC), как раскрывается в примере 12 далее и как описывалось выше.
Осадок может быть собран после второй промывки с использованием детергента путем взбалтывания в жидкости, например в воде или в буфере, содержащем желательный компонент SB-18-подобный детергент, в подходящей по размеру емкости, способной выдержать интенсивное перемешивание, например такое, которое дает встряхиватель, и в последующем при необходимости проводить очистку, используя методы, известные в данной области, например такие, как гель- хроматография, ионнообменная хроматография или способы фильтрации, проводящиеся до, в течение или после второй промывки. Бактериальный осадок, полученный в результате такой процедуры, может использоваться как источник микобактерий для определения, включая определение путем культивирования.
В предпочтительном воплощении изобретения образец непосредственно обрабатывается раствором, содержащим желаемый детергент, такой как SB-18, что отражено в примере 13 далее, помещается в подходящего размера емкость, способную выдержать интенсивное перемешивание, такое, как на встряхивателе, и последующую, в случае необходимости, обработки, используя известные в этой области методы, например, такие как гель-хроматография, ионнообменная хроматография или фильтрация, до, в процессе или после непосредственной обработки. Полученный после такой начальной процедуры бактериальный осадок может быть использован как источник микобактерий для определения, включая определение путем культивирования.
Дополнительные первичные или вторичные этапы просветления или очистки, известные в данной области, если необходимо, могут быть включены на соответствующих стадиях в процесс, если в состав среды внесен подходящий детергент, такой как SB-18-подобный или октадецилоподобный детергент. Как показано выше, когда как дегазирование, так и обработка детергентом применены, то если обработка подходящим детергентом, как описано в данной работе, предшествует, совпадает или следует за дегазированием, то не является необходимым использовать исключительно SB-18-подобные детергенты и могут быть использованы любые детергенты, особенно такие, как октадецилоподобные детергенты, как показано в этой работе и проиллюстрировано.
Конечно, если образец изначально обработан стандартным методом (таким как NALC/NaOH метод), как показано в примере 12, и затем либо (i) промыт с SB-18-подобным детергентом, предпочтительно SB-18, (ii) просто дегазируется или (iii) сочеталась промывка с использованием детергента, и вакуумная обработка для того, чтобы избавиться от плавучести, выделение остающихся микроорганизмов может быть повышено, но не обязательно в количественном отношении больше, чем потери, имевшие место в результате обработки любым из стандартных методов, которые уже подвергали риску образец.
В любой из описанных в данной работе комбинаций, включая сочетание методов по данному изобретению со стандартной NALC/NaOH методикой, обработанные клетки более эффективно собираются в осадок, что может теперь использоваться для определения исследуемых микроорганизмов в данной работе.
Применение изобретения приводит к количественной экстракции микобактерий из образца способом, который фактически уничтожает ложноотрицательные результаты, получающиеся из-за группирования бактерий при росте или из-за плавучести. Способы по данному изобретению, в первую очередь, обеспечивают уверенность в результатах определения, например по методу амплификации нуклеиновых кислот, и обеспечивают такое состояние микобактерий, которое позволяет их высевать, а также лизис таких микобактерий другими методами, например кипячением.
Способы по данному изобретению применяются для определения микобактерий в образцах, взятых от любого объекта. Помимо человека известно, что микобактериозы бывают у следующих животных: крупный рогатый скот, свиньи, домашняя птица, овцы, козы, олени, обезьяны, слоны, лошади, собаки, кошки, норки и различные другие наземные и водные животные. Образцы ткани, пригодные для использования по способу данного изобретения, обычно берутся из грануломатозных поражений, обычно бугристых и имеющих кальцинированный центр, заключенный в фиброзную капсулу. Если крупные очаги поражения отсутствуют, предпочтительнее анализировать лимфатические узлы. Если поражен пищеварительный тракт, обычно анализируется толстый кишечник.
Образцы могут быть в виде биологических жидкостей, например, мокрота, спинномозговая жидкость, моча, бронхиальные промывные воды, плевральная жидкость, материал, полученный путем аспирации желудка, кровь, сыворотка, перитонеальная жидкость, содержимое абсцессов и эксудаты или другие биологические жидкости, а также штаммы могут быть получены из государственных или частных коллекций культур микроорганизмов и из других источников, или специально из медицинских клиник или от ветеринарных служб. Жидкие культуры, замороженные
суспензии или колонии, выросшие на твердой среде, могут использоваться. Некоторые образцы могут быть полутвердые и/или нуждаться в осветлении, например такие как фекалии и стабилизированная антикоагулянтом кровь или гемокультура. Способы по данному изобретению также полезны для образцов, полученных от таких необычных объектов, как, например, рыбы или рептилии и от образцов ткани.
Нет необходимости брать образец для культивирования. Действительно, вкладом данного изобретения является то, что прямое определение искомых микроорганизмов путем амплификации нуклеиновых кислот или сигнальной амплификации может быть выполнено непосредственно от образца, взятого прямо от осадка. В данной работе показано, что микобактерии остаются живыми (например, не лизируются) после использования метода по данному изобретению, и, следовательно, высев может быть выполнен по желанию. Например, все образцы сначала могут быть обработаны способами по данному изобретению с целью обеспечить максимальную эффективность выделения микроорганизмов для определения методами амплификации, и ответ, касающийся постановки диагноза, поступает в кратчайшие сроки. Образцы с положительным ответом в дальнейшем могут быть высеяны. Таким образом, данная технология резко снижает время, требуемое для диагностики, и переводит микроорганизмы, особенно микобактерии, находящиеся в образце, в форму, пригодную для культивирования. Возрастающее число устойчивых к лекарственным препаратам штаммов микобактерий создает потребность в определении чувствительности к лекарственным веществам у всех культур, содержащихся в образцах, давших положительный ответ. По данному изобретению эффективность выделения микобактерий в жизнеспособной форме является оптимальной, предлагаемые методы облегчают постановку диагноза и определение чувствительности.
В первую очередь способы по данному изобретению существенно снижают число ложноотрицательных результатов, вызывавшихся способностью таких микроорганизмов, как микобактерий MTB комплекса, группироваться. Группирование клеток бактерий существенно усиливает неэффективность разделения таких микроорганизмов при обработке пробы. Известно, что способы по данному изобретению облегчают гомогенное поступление группирующихся микроорганизмов в требуемый раствор и таким образом повышают возможность их определения, используя методы определения, известные в данной области, например, культивирование и амплификацию нуклеиновых кислот (ДНК или РНК), за счет повышения вероятности, что все аликвоты взятого образца будут содержать микроорганизмы.
Кроме того, способы по данному изобретению преодолевают естественную плавучесть данных микроорганизмов в водных средах, облегчая тем самым их сбор с помощью известных методов, например, центрифугирования. Природная плавучесть существенно снижает способность к эффективному сбору этих микроорганизмов с помощью центрифугирования.
Представленные методы по данному изобретению существенно улучшают выделение этих микроорганизмов из образцов, потребность в длительном культивировании для получения результатов исчезает, поскольку в диагностике могут в качестве замены использоваться более технологичные современные методы; технология амплификации, основанная на использовании нуклеиновых кислот, один из примеров более усовершенствованного метода. Способы по данному изобретению, однако, не лизируют микроорганизмы, а дают жизнеспособные клетки, так что способы культивирования не исключаются как способы определения.
Способы по данному изобретению направлены на то, чтобы образец подготавливался для тестирования на наличие микроорганизма, такого как микобактерия. Следовательно, какие бактерии, в итоге определяются, зависит от процедуры определения, которая используется в сочетании с методами по данному изобретению. Такая процедура определения может быть создана для того, чтобы определить любые искомые организмы, и особенно, любые искомые группы или комплексы видов микобактерий, наиболее предпочтительными являются такие комплексы микобактерий, как М. tuberculosis (MTB) комплекс, M.avium (MAC) комплекс, MAIS комплекс и M.fortuitum комплекс, а также быстрорастущие и медленно растущие микобактерии, включая специфические и неспецифические фотохромогены, нефотохромогены, скотохромогены и, особенно, М. africanum, М. asiaticum, М. avium, М. bovis (BCG), М. butyricum, М. chelonae, М. duvalii, М. flavescens, М. fortuitum, М. gastri, M. gorgonae, М. haemophilum, М. intracellularae, М. kansasii, М. leprae, М. lepraemurium, М. linda, М. lufu, М. marinum, М. malmoense, М. microti, М. mucoscum, М. nonchromogenicum, М. paratuberculosis, М. peregrinum, М. phlei, М. rhodochrous. М. scrofulaceum, М. shimoidei, М. simiae, М. smegmatis, M.szulgai, М. terrae, М. thermoresistable, М. triviale, М. tuberculosis, М. ulcerans, М. vaccae, М. xenopi и их серологические типы.
M. kansasii, М. marinum, М. simiae и М. asiaticum являются примерами фотохромогенов. М. scrofulaceum, М. szulgai, М. xenopi, М. gordonae и М. flavescens являются примерами скотохромогенов. М. avium, М. intracellulare, М. gastri, М. malmoense, М. terrae и М. triviale все являются примерами нефотохромогенов.
М. africanum, М. avium, М. bovis, М. haemophilum, М. intracellularae, М. kansasii, М. malmoense, М. marinum, М. microti, М. paratuberculosis, М. scrofulaceum, М. simiae, М. szulgai, М. tuberculosis и М. xenopi все являются примерами медленнорастущих видов микобактерий (более семи суток). М. chelonae, М. flavescens, М. fortuitum, М. gordonae, М. leprae, М. phlei, М. smegmatis, М. terrae, М. ulcerans все являются примерами быстрорастущих видов (менее семи суток) микобактерий.
М. tuberculosis, М. africanum, М. bovis, М. bovis (BCG) и М. microti все относятся к MTB комплексу. М. avium и М. intracellularae все относятся к MAC комплексу; существует по меньшей мере три разные серологические группы М. avium и более 25 серологических вариантов М. intracellularae.
Данное изобретение особенно полезно для работы с липофильными микроорганизмами или микроорганизмами, покрытыми воскоподобной капсулой, в составе которой молекулы типа миколовой кислоты на наружной поверхности клеточной стенки (такие, например, как кориномиколовая кислота, нокардиомиколовая кислота и миколовая кислота наряду с другими). Все они характеризуются как "структуры миколовой кислоты", которая является β- окси кислотой, замещенной в α- положении алифатической цепью средней длины, как было установлено (Goren, М. В. Bact. Rev. 36:33-64 (1966), работа упоминается здесь в виде ссылки). Примером бактерии, содержащей кориномиколовую кислоту, является Corynebacterium diphteria, примером микроорганизма, содержащего нокардиомиколовую кислоту, является Nocardia asteroides; и примером микроорганизма, имеющего миколовую кислоту, является Mycobacterium tuberculosis. Такие вещества, близкие к миколовой кислоте, в данной работе все вместе называются "миколовыми компонентами". Дополнительные примеры, представляющие миколовые структуры, включая ненасыщенные, циклопропаноидные, метаксилированные и кетоновые кислоты, можно найти, например, в работах Lederer, E.Chem. Phys. Lipids 1: 294-315 (1967); Lederer, Е. Pure Appl. Chem. 25:135-165 (1971), включенных в данную работу в качестве ссылок. "Структуры миколовой кислоты" являются кислотоустойчивыми.
Примерами заболеваний и условий, при которых различные виды микобактерий очень важно определить, являются особенно возбудители туберкулеза (М. tuberculosis комплекс) и лепры (M.leprae (проказа) и M.lepraemurium (изъязвленная проказа)). Комплекс Mycobacterium avium вызывает серьезные заболевания птиц. М. avium также была выделена у пациентов, больных СПИДом, отягощенных микобактериальной суперинфекцией. М. bovis имеет значение в ветеринарной практике. М. fortuitum является почвенной бактерией, которая была выделена из язв человека и животных. М. intracellulare является оппортунистической инфекцией и чаще встречается у носителей ВИЧ. М. paratuberculosis представляет интерес при диагностике болезни Крона (регионарный илеит) у людей. Mycobacterium kansasii является редким, но разрушительно действующим возбудителем, обычно ассоциированным с легочными заболеваниями. Mycobacterium marinum инфицирует не теплокровных животных, в частности рыб; также этот возбудитель был выделен из поверхностных гранулем на конечностях у людей. Mycobacterium paratuberculosis является возбудителем болезни Джона у крупного рогатого скота; возбудитель характеризуется очень медленным ростом и культивирование должно продолжаться не менее 16 недель, после чего можно делать вывод об отрицательном результате. М. ulcerans также представляет интерес для медицины. Многие, перечисленные выше, и другие возбудители обсуждались в работе Wayne, L. G. et al., Clin. Micro. Rev. 5:1-25 (1992), которая включена в данный текст в виде ссылок.
Стадия определения может использовать любой метод, известный для определения искомого микроорганизма, особенно искомых микобактерий, включая, но не ограничиваясь, методом амплификации нуклеиновых кислот, сигнальной амплификацией, гибридизацией, культивированием и иммуноанализом. В предпочтительном воплощении изобретения, методом определения является амплификация нуклеиновых кислот и/или высевы. В наиболее предпочтительном воплощении изобретения используется амплификация, а именно амплификация полимеразной цепной реакции. В самом деле, ценностью метода по данному изобретению является то, что SB-18-подобные детергенты не ингибируют ПЦР и что бактерии сохраняют жизнеспособность и их можно культивировать. В отличие от традиционных методов в этой области, где применение NALC/NaOH раствора очень сильно ингибирует ПЦР. Кроме того, четко установлено, что если микроорганизмы сохранили жизнеспособность, то при последующей обработке NALC/NaOH 28-33% микобактерий погибало (Krasnow, I. et al., Am. J. Clin. Path. 45:352-355 (1966); Kubica, G.P.W. et al. , Am. Rev. Resir. Dis. 87:775-779 (1963)). В противоположность этому, хотя SB-18-подобные детергенты могут проявлять бактерицидную активность, в методах по данному изобретению они применяются в концентрациях, при которых микобактерии не лизируются и сохраняют жизнеспособность.
В методах по поиску микобактерий могут применяться метки, которые обычно используются при определении нуклеиновых кислот и/или при иммуноанализе антигенов микобактерий, включая, но не ограничиваясь ими, радиоактивные маркеры (например, 32P, 33P, 35S, 14C и 3H), флуоресцентные красители (например, флюоресцеин, аурамин, родамин, Texas Red и т.д.), хемилюминесцентные красители (например, зонды, меченные акридиниевым эфиром, LUMI-PHOS 530TM, Schaap реагент (4-метокси-4-(3-фосфатофенил)-спирол-[1,2-диоксетан-3,2'-адамантан), CSPD© (патент США 5112960) и т.д.), колориметрическое определение (например, 3,3',5,5'-тетраметилбензидин (ТМБ) и т.д.), электрохемилюминесцентные (например, хелат трис(2,2'- бипиридин)рутения (II) (ТБР)) и белковые или ферментные маркеры (например, антитела, щелочная фосфатаза, пероксидаза хрена и т.д.); все эти реагенты могут использоваться совместно с любой категорией веществ, упоминавшейся выше.
Способы по данному изобретению наиболее удобно использовать, получая реактивы, используемые в каждом из методов, в виде наборов. Такой набор предпочтительно содержит требуемые буферные растворы, соли, SB-18-подобный детергент (или их эквиваленты, так как детергент применяется в сочетании с дегазированием) и, если необходимо, воду требуемой чистоты. По меньшей мере один тип идентифицируемого агента также может быть включен в зависимости от типа анализа, используемого при дальнейшем определении (например, амплификации, иммуноопределения и т.д.); и "положительный" стандарт, предоставляющий либо нуклеиновую кислоту (ДНК или РНК), полезную при определении конкретного микроорганизма, или, желательно, специфический антиген (белок или другое вещество), или другое специфическое вещество, как компонент для идентификации липидного состава микроорганизмов, особенно для методов газожидкостной хроматографии. Специфические наборы могут содержать, inter alia, конкретно средства для идентификации микобактерий, такие как конкретные зонды нуклеиновых кислот, антитела или стандартные препараты-мазки для микроскопии, или материалы для высевов. Средства, с помощью которых идентифицируемый агент обнаруживается, могут быть специфичными для набора, так что набор обеспечивает, например, ферментное, флюоресцентное, радиоактивное или хемилюминесцентное определение, или любые другие соответствующие определения, известные в этой области. В таком наборе, такие средства определения как правило находятся в тесной связи с детергентом (детергентами), даже если содержатся в отдельных емкостях или пакетах.
Если в качестве метода определения используется амплификация нуклеиновых кислот, следует отметить, что известно много типов такой амплификации и все могут использоваться по способам данного изобретения. Например, типы систем амплификации нуклеиновых кислот включают полимеразную цепную реакцию (ПЦР), лигазную цепную реакцию (ligase chain reaction - LCR), Qβ репликазная амплификация, амплификация по типу замещения цепи (strand displacement amplification - SDA) или одиночная праймер амплификация (singl primer amplification - SPA), или системы амплификации на базе транскрипции, такая как NASBA (nucleic acid sequence based amplification - амплификация на базе нуклеиновой последовательности), SSSR (self sustained sequence replication - репликация с самозамещающейся последовательностью), или LAT (ligation activated transcription - транскрипция, активируемая лигированием) амплификация. Системы сигнальной амплификации, как например система разветвленной ДНК (bDNA), также используются.
Создание микобактериальных зондов ген-специфических и последовательность-специфических было описано (Fries, J.W.U. et al., Molec. Cell. Probes 4: 87-105 (1990). Определенные генные последовательности существуют не у одного вида микобактерий и могут эффективно использоваться для определения присутствия видов семейства микобактерий. Например, Crawford et al., патент США 5183737, описывают повторяющуюся последовательность ДНК, специфическую для представителей М. tuberculosis (MTB) комплекса. Определение может быть произведено прямым анализом гибридизации (например, методом Саузерна) или с использованием фрагмента консервативной последовательности в качестве праймера при проведении амплификации. В описываемом в данной работе новом методе амплификации MTB комплекс, Mycobacterium avium комплексе, Mycobacterium intracellulare, Mycobacterium paratuberculosis, Mycobacterium kansasii, Mycobacterium marinum, Mycobacterium marinum, Mycobacterium szulgai и Mycobacterium gastri также могут быть амплифицированы с использованием одного набора зондов, как описано в данной работе, используя последовательности на основе генной последовательности 16S rРНК, как опубликовано Rogall, T. et al. , Int. J. Sys. Bacteriol. 40:323-330 (1990). Праймеры были сконструированы таким образом, что они могут обеспечить оптимальную амплификацию следующих групп/видов микобактерий: М. tuberculosis (ТВ комплекс: [MTB]), М. avium - М. intracellulare и М. paratuberculosis (MAC комплекс), М. kansasii и М. marinum. Прямой праймер был сконструирован в отношении нуклеотидов 119-144 (по номенклатуре Rogall, T. et al., Int. J. Sys. Bacteriol. 40: 323-330 (1990)) второй вариабельной области (V2) генной последовательности 16S rРНК этих микобактерий. Последовательность прямого праймера (TBv2-119): 5'-AAA CTG GGT CTA ATA CCG GAT AGG A-3' [SEQ ID N.: 1:]. Обратный праймер был сконструирован в отношении нуклеотидов 431-453 третьей вариабельной области. Последовательность обратного праймера (TBv3-453): 5'-CCA CCT ACC GTC AAT CCG AGA-3' [SEQ ID N.:2:]. Продукт амплификации содержит примерно 335 пар оснований (в зависимости от амплифицируемого вида). Геноспецифичный зонд был разработан в отношении центрального участка амплифицируемого продукта и является общим для всех микобактерий. Его последовательность: 5'-GCC GGC iCA TCC CAC ACC GC-3' [SEQ ID N.:3:]. Зонд, специфичный для MTB-видов, был сконструирован в отношении другого участка амплифицируемого продукта и является специфичным для бактерий, входящих в ТВ комплекс. Его последовательность: 5'-GAC CAC GGG ATG CAT GTC TTG TG-3' [SEQ ID N.:4:].
С другой стороны, известны видоспецифичные зонды. Например, McFadden et al., патент США 5225324, описывают группу ДНК инсертных последовательностей, которые можно использовать в качестве зондов для идентификации микобактерий и для дифференциации близкородственных видов. В патенте США 5216143 описываются зонды, специфичные для M.gordonae. Зонды для М. paratuberculosis также известны: инсертная последовательность IS900 специфична для этого вида (McFadden, J.J. et al., Mol. Microbiol. 1:283-291 (1987)).
Действие SB-18-подобных детергентов
В наиболее предпочтительном воплощении изобретения предпочитаемым для среды детергентом и, особенно на стадии второй промывки, является SB-18-подобный детергент, способный разрушить тяжи микобактерий, и особенно детергент SB-18 (CAS® N. 13177-41-8), в количестве, достаточном для разрушения тяжей и для того, чтобы равномерно диспергировать микобактерии. SB-18 является предпочтительным ввиду его экономической доступности в чистой форме. C18-карбоксипропилбетаин( CAS® N. 78195-27-4) высокопредпочтителен, поскольку идеально сочетает растворимость, длину цепи и структуру мостика связи для использования в методах по данному изобретению.
SB-18 (CAS® N. 13177-41-8) имеет химический состав C23H49NO3S и химическую формулу CH3(CH2)17N(CH3)2 (CH2)3SO3. Химические названия SB-18 следующие: диметилоктадецил(3-сульфопропил)аммоний гидроксид, внутренняя соль (Aldrich N. 36712-5); N-октадецил-N,N-диметил-3-аммонио-1-пропан сульфонат (Sigma N. 08004); 3-(N, N-диметилстеариламмонио)пропаносульфонат и 3-(стеарил-диметиламмонио)пропаносульфонат (Fluka N. 41570)). Наиболее распространенное название SB-18 фирмы Sigma используется в методах по данному изобретению.
Только еще один детергент из серии SB был пригоден для разрушения тяжей: SB-16 (CAS® N. 2281-11-0; N-гексадецил-N,N-диметил-3-аммонио-1- пропан-сульфонат; пальмитил сульфобетаин; Sigma H 6883), однако, он был не так эффективен при равных концентрациях. Представленные в данной работе данные показывают, что большинство октадецилбетаинов, у которых R1, как раскрывается далее, содержит более 16 атомов углерода, обладают способностью разрушать скопления клеток М. tuberculosis.
Определенные виды детергентов не проявляют способности разрушать бактериальные тяжи и доводить до удовлетворительной степени раздробленность микроорганизмов MTB комплекса. Было обнаружено, что детергенты и другие проверявшиеся вещества не могут дезагрегировать MTB комплекс; среди этих веществ SB-10 (CAS® N. 155163-36-7), SB-12 (CAS® N. 14933-08-5), SB-14 (CAS® N. 14933-09-6), декановая кислота (CAS® N. 334-48-5), додекановая кислота (CAS® N. 143-07-7), натрий додецил сульфат (SDS: CAS® N. 151-21-3), бензалкониум хлорид (BenzAlk: CAS® N. 8001-54-5), смешанный алкилтриметил аммоний бромид (mTMA), додецилтриметиламмоний бромид (TMA-12: CAS® N. 1119-94-4), миристилтриметиламмоний бромид (TMA-14: CAS® N. 1119-97-7), октадецилтриметиламмоний бромид (TMA-18: CAS® N. 1120-02-1), дезоксихоликовая кислота (CAS® N. 302-95-4), бензилдиметилдодециламмоний бромид (BenzDMA-12: CAS® N. 7281-04-1), бензилдиметилтетрадециламмоний хлорид (BenzDMA-14: CAS® N. 139-08-2), бенздиметилоктадециламмоний хлорид (BenzDMA-18), Tween 20 (CAS® N. 9005-64-5), Tween 60 (CAS® N. 9005-67-8), Tween 80 (CAS® N.9005-65-6), TritonX-100 (CAS® N. 9002-93-1), NP-40 (CAS® N. 127087-87-0), Brij 35 (CAS® N. 9002-92-0), Brij 99 (CAS® N. 9004-98-2), Span 20 (CAS® N. 1338-39-2), Span 60 (CAS® N. 1338-41-6), Span 80 (CAS® N. 1338-43-8), SynperonicF/68, полиэтиленгликоль 1450 (CAS® N. 25322-68-3), Ficoll 400000 (CAS® N. 26873-85-8), поливинилпироллидон 360000 (CAS® N. 9003- 39-8), формамид (CAS® N. 75-12-7) и диметилформамид (CAS® N. 68-12-2).
Некоторые детергенты, несмотря на то, что они не могут разрушать бактериальные тяжи, способствуют сбору клеток. Например, в данной работе было показано, что все детергенты серии SB, особенно SB-12, SB-14, SB-16 и SB-18, предпочтительно SB-18, могут довольно эффективно использоваться для микроорганизмов, не образующих скопления групп клеток, например М. avium. Следовательно, помимо SB-18 ( CAS® N. 13177-41-8) другие детергенты серии SB, способные снимать плавучесть, могут улучшить сбор бактериальных клеток методами с использованием гравитационной силы; к ним относятся: SB-16 ( N. 2281-11-0: N-гексадецил- N, N-диметил-3-аммонио-1-пропансульфонат; пальмитилсульфабетаин; Sigma H 6883); SB-14 (CAS® N. 14933-09-6: N-тетрадецил-N, N-диметил-3-аммонио- 1-пропансульфонат; миристил сульфобетаин; Sigma T 7763); и SB-12 (CAS® N. 14933-08-5; N-додецил-N,N-диметил-3- аммонио-1-пропансульфонат; лаурилсульфобетаин; Sigma D 4516).
N. 2281-11-0: N-гексадецил- N, N-диметил-3-аммонио-1-пропансульфонат; пальмитилсульфабетаин; Sigma H 6883); SB-14 (CAS® N. 14933-09-6: N-тетрадецил-N, N-диметил-3-аммонио- 1-пропансульфонат; миристил сульфобетаин; Sigma T 7763); и SB-12 (CAS® N. 14933-08-5; N-додецил-N,N-диметил-3- аммонио-1-пропансульфонат; лаурилсульфобетаин; Sigma D 4516).
Цвиттерионные детергенты серии SB содержат как четвертичный азот, так и сульфонатную группу, разделенные пропильной группой, и каждый обладает длинной алкильной цепью; додецил, тетрадецил, гексадецил или октадецил, соответственно, у SB-12, SB-14, SB-16 и SB-18 связаны с четвертичным азотом. Как было показано в данной работе, в то время, как SB-18 проявил большую эффективность в разрушении бактериальных тяжей, серия SB-детергентов в целом была в некоторой степени эффективна при сборе клеток М. tuberculosis при центрифугировании. Кроме того, детергенты SB-серии также были эффективны при концентрировании таких микроорганизмов, которые не образуют тяжей, например М. avium. Как описано далее, без намерения вдаваться в объяснение, эти цвиттерионные детергенты, как можно предположить, имеют характеристики, позволяющие облегчить попадание детергентов в бактериальные клетки. Положительный эффект от такой аккумуляции заключается в компенсировании природной плавучести этих микроорганизмов, что позволяет существенно улучшить их концентрирование путем центрифугации.
Согласно данному изобретению, в его воплощениях, где используются детергенты, получается равный конечный результат, независимо от того, используется SB-18-подобный детергент для облегчения диспергирования тяжей, или он используются для облегчения концентрирования бактерий центрифугированием: согласно данному изобретению микобактерии помещаются в среду, которая облегчает их последующее определение, как за счет равномерного диспергирования из образца, и/или за счет улучшения их концентрирования при центрифугировании.
В данной работе такая двойственность функции называется "SB-18-подобная активность". Следовательно, SB-18-подобная активность, как здесь используется это понятие, будет подразумевать как способность SB-18-подобного детергента улучшать разрушение тяжей в ходе обработки микроорганизма, образующего скопления клеток при росте (и, таким образом, равномерно высвобождать бактерии в раствор), так и способность SB-18-подобного детергента или октадецилоподобного детергента способствовать существенному количественному сбору таких микроорганизмов при центрифугировании в связи со снижением плавучести, или способность SB-18-подобного детергента делать и то, и другое.
Бетаиноподобные детергенты
В данной работе показано, что вещества, близкие к SB-18 обладают, (а) только четвертичным амином (октадецил, катионные детергенты триметилоктадециламмоний бромид TMA-18: CAS® N. 1120-02-1) и бензилдиметилоктадециламмоний хлорид (BenzDMA-18), например; (b) только сульфатной частью (октадецил, анионный детергент натрий октадецилсульфат (SOS): CAS® N. 1120-04-3); и (c) только цвиттерионными свойствами (3-[(3-холамидопропил)-диметиламмонио] - 111-пропан-сульфонат (CHAPS): CAS® N. 75621-03-3), все они не могли повысить выделение без проведения дегазирования. Это означает, что связь между алкильной частью молекулы и цвиттерионной функциональностью способствует SB-18-подобной активности.
Детергенты SB-серии, также известные как сульфобетаины, являются частью широко представленного класса цвиттерионных детергентов, известных как бетаины. Бетаины - это цвиттерионные молекулы, содержащие, inter alia, центр положительного заряда отдельно от центра отрицательного заряда. В табл. 2 даны родовые структуры наиболее обычного класса бетаинов (n-алкилбетаинов), а в табл. 3 описываются некоторые общие их структурные варианты. Не вдаваясь подробно в какую либо теорию, предполагается, что цвиттерионная функциональность образует дипольный момент, что очевидно создает SB-18-подобную активность для целого класса веществ. Невзирая на это, в данной работе показано, что вариации в структуре бетаина создают предсказуемые результаты в плане использования бетаинов в способах по данному изобретению.
В табл. 2 приведено общее описание n-алкилбетаинов. В этом описании "R1" представляет гидрофобную часть детергента. Обычно это алкильная часть молекулы и может быть представлена всего 8 атомами углерода, а может быть длина цепи до 22 атомов углерода. Желательно, чтобы алкильная часть содержала от 12 до 20 атомов углерода, а более предпочтительно - от 16 до 18 атомов углерода. Алкильная цепь может быть ненасыщенной (алкиленовой), разветвленной и/или быть замещенной, например, гидроксилом, простым эфиром, сложным эфиром, карбонильными группами, а также другими группами.
"α" может быть представлена, или может отсутствовать, в зависимости от того, равно "n" 0 или 1. Если n = 0, α отсутствует; если n = 1, α присутствует. Там, где α присутствует, она связывает R1 с центром положительного заряда. Такой связью может быть метиленовая группа. Одной из наиболее распространенных подобных связующих групп является амидопропильная группа (-(CO)-NH-CH2CH2CH2-). Также известны другие связующие группы: эфирная (-O-), карбонильная (-CO-) или гидроксиметильная (-CH(OH)-). Весьма вероятны другие группы, например амины.
"R2" и "R3" независимо выбраны из группы, состоящей из водорода и алкила, предпочтительно низший алкил из 1-4 атомов углерода, например, метил, этил, пропил, бутил или их изомеры, и наиболее предпочтительным является метил; увеличение объема таких алкильных групп, как за счет удлинения, так и за счет разветвления, ухудшает их действие, что обсуждается в примерах 9 и 10.
"β" представляет центр положительного заряда. В подавляющем большинстве случаев β представлена четвертичным азотом (-N⊕-), однако, возможны фосфоний (-P⊕-), сульфоний (-S⊕-) и другие фрагменты молекулы. Следует отметить, что в большинстве случаев β представлена четвертичным азотом, могут быть нужны заместители "R2" и "R3".
"R4" является мостиком, разделяющим энергетические типы. Такой мостик может быть алкиленовой группой (например, m≥1), например, метилен, этилен, пропилен, бутилен, пентилен и гексилен. Состав и структура R4 не ограничивается только алкиленовой группой: также распространены примеры, когда включены гидроксильные группы, или когда R4 имеет разветвленную структуру или представлен бензильной группой. Также возможны другие модификации R4, включающие амины, например, и можно предполагать их функциональную схожесть.
Центр отрицательного заряда, "γ", образуется от большого круга групп, включающего сульфонат (-SO
Таблица 2: Структура N-алкилбетаинов
Общая структура N-алкилбетаинов представлена ниже. R1 - гидрофобная алкильная цепь, α- связь, связывающая R1 с катионом β. R2 и R3 - заместители, модифицирующие катион. R4 - мостик, связывающий катион с анионом γ.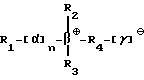
Сульфопропилбетаины были взяты из различных источников (т.е. CalBiochem, La Jolla, Ca; Sigma, St.Louis, MO; Fluka, Ronkonkoma, NY; Aldrich, Milwaukee, WI). Некоторые из них были успешно протестированы по методам изобретения и проявили SB-18-подобную активность. Такими соединениями были следующие: C12-сульфопропилбетаин (SB-12: CAS® N. 14933-08-5), C14-сульфопропилбетаин (SB-14: CAS® N. 149933-09-6), C16- сульфопропилбетаин (SB-16: CAS® N. 2281-11-0), C18-сульфопропилбетаин (SB-18: CAS® N.13171-41-8). Некоторые другие сульфобетаины были либо синтезированы, либо получены в качестве образцов, как описано ниже. Такими соединениями были следующие: C18-сульфобутилбетаин (CAS® N. 22313-73-1), C16-гидроксипропилсульфобетаин (CAS® N.7425-12-9), 3-бутокси-2-гидрокси
гидроксипропилсульфобетаин (CAS® N. 108797-84-8). Также были протестированы некоторые сульфабетаины, алкильные цепи которых получены из природных масел. Это были кокоамидопропилгидроксипропилсульфобетаин (CAS® N.68139-30-0) и эрукоамидопропил гидроксипропилсульфобетаин. Таллоуамидопропилгидроксипропилсульфобетаин.
Благодаря хорошей растворимости наиболее широко были изучены карбоксибетаины (γ-COO⊖). Некоторые из них были протестированы методами изобретения и проявили SB-18-подобную активность. Такими соединениями были следующие: C16-карбоксиметилбетаин (CAS® N. 693-33-4), C18-карбоксиэтилбетаин (CAS® N. 30612-73-8), C18:1-карбоксиметилбетаин (CAS® N. 871-37-4), C18-амидопропилкарбоксиметилбетаин (CAS® N.6179-44-8). Также были протестированы некоторые карбоксибетаины, алкильные цепи которых получены из природных масел. Такими соединениями были следующие: кокоамидопропилкарбоксиметилбетаин (CAS® N.61789-37-9 и CAS® N.61789-40-0), коко-карбоксиметилбетаин (CAS® N. 68424-94-2), рициноамидопропилкарбоксиметилбетаин (CAS® N.71850-81-2) и таллоубисгидроксиэтилглицинат (CAS® N.70750-46-8). Также были протестированы некоторые карбоксибетаины, для которых не даны CAS®-номера. Такими соединениями были следующие: бегенил-карбоксиметилбетаин, содержащий C22-цепь из масло-амидопропилкарбоксиметилбетаина (Schercjtaine WOAB: Scher Chemicals, Inc., Clifton, NJ), вавассуамидопропилкарбоксиметилбетаин (Croda, Inc. Parsippany, NJ), сояамидопропилкарбоксиметилбетаин (Chembetaine S: Chemron Corp. , Paso Robles, CA), каноламидопропилбетаин (Hetaine CLA: Heterene, Inc., Patterson, NJ).
Из-за различной природы бетаинов некоторые из них могут действовать при специальных условиях: бегенил-карбоксиметилбетаин, содержащий C22-цепь, может действовать только при минимальных концентрациях соли, a C16-гидроксипропилсульфобетаин требует присутствия иодида калия.
Два дополнительных бетаина были синтезированы, чтобы представить два других класса бетаинов. Они также были успешно применены и такими соединениями были C16-амидопропилсульфатобетаин (CAS® N. 58930-11-3) и C18-фосфоэтилбетаин (CAS® N. 126712-89-8), представляющие сульфатные и фосфатные анионы, соответственно.
Таким образом, 25 бетаинов, представляющих широкий спектр структурных различий в их молекулах, были протестированы на эффективность для увеличения сбора микобактерий, например, центрифугированием. Оказалось, что бетаины с длиной цепи менее 16 атомов углерода главным образом действовали непосредственно путем снижения плавучести без нарушения агрегации клеток, в то время как бетаины с длиной цепи 18 атомов углерода или больше действовали, по большей части разрушая тяжи, а также уменьшая плавучесть. Оказалось, что все они применимы в методах изобретения. Комбинация в молекуле бетаинов цвиттериона и гидрофобной алкильной цепи придают последним уникальные свойства. Поэтому можно ожидать, что молекулы с "бетаиноподобной структурой" проявляют SB-18-подобную активность.
Известны способы синтеза для многих структурных комбинаций, перечисленных выше. Число SB-подобные детергентов, эффективные в методах по данному изобретению, не ограничено примерами, поскольку поведение веществ этого класса в данных методах предсказуемо.
Дополнительные указания приведены в списке внизу, в частности примеры бетаинов, являющихся коммерчески доступными, или о которых известно, как их можно синтезировать, и которые могут быть полезны в способах по данному изобретению.
Большинство бетаинов имеют в качестве катиона четвертичный азот (β). Представители этого большого семейства детергентов наиболее легко перечислить, принимая в качестве катиона четвертичный N,N-диметиламмоний и затем группируя заместители, в первую очередь основываясь на анионе (γ). Каждый заместитель может быть затем поделен сначала на мостиковую структуру (R4), затем на связь (α) и затем на алкильную цепь (R1). Другие структурные сочетания, которые не подпадают под эти дискретные категории, затем легко перечисляются. Снова все могут быть обоснованно рассмотрены с точки зрения их действия по методам данного изобретения.
Примеры карбоксибетаинов, имеющих метиленовый мостик ("карбоксиметилбетаины": R4= -CH2-), метиленовую связь (α =-CH2-), и 15 варьирующихся исключительно по длине алкильной цепи: C10 (CAS® N. 2644-45-3), C11 (CAS® N. 2956-38-9), C12 (CAS® N. 683-10-3), C13 (CAS® N. 23609-76-9), C14 (CAS® N. 2601-33- 4), C15 (CAS® N. 23609-77-0), C16 (CAS® N. 693-33-4) и C18 (CAS® N. 820-66-6). C12карбоксиметилбетаин (CAS® N.6232-16-2) является примером вещества, у которого N,N,диэтил (R3=R4=-CH2CH3); пример, когда алкил имеет двойную связь: C18:1 (CAS® N. 871-37-4). В этой подгруппе находится несколько карбоксиметилбетаинов, у которых α представлена амидопропильной группой. К ним относятся: C12-амидо (CAS® N. 4292-10-8), C14-амидо (CAS® N. 59272-84-3), C16-амидо (CAS® N. 32954-43-1) и C18-амидо (CAS® N. 6179-44-8). C18-амидо (CAS® N. 6179-44-8) имеет неопределенную структуру, поскольку алкил представлен "изо" формой, которая как предполагается, что разветвляется некоторым неопределенным образом. Также там содержится несколько амидопропилкарбоксиметилбетаинов, у которых алкильная цепь происходит от масла кокосового ореха и зависит от способа приготовления. Два примера в этой категории: CAS® N. 61789-39-7 и 61789-40-0. Примером кококарбоксиметилбетаина может быть CAS® N. 68424-94-2. Производные других природных масел следующие: рицин-амидопропилкарбоксиметилбетаин ( N. 71850-81-2) и сталлоубисгидрокси бишидрохуэтилглицинат (CAS® N. 70750- 46-8). Также исследовалось еще несколько карбоксиметилбетаинов, для которых не определены CAS® номера. К ним относятся масло проростков пшеницы - амидопропил карбоксиметилбетаин (Schercotaine WOAB: Scher Chemicals, inc., Clifton, NJ), бабассу-амидопропилкарбоксиметилбетаин (Croda, Inc., Parsippany, NJ), соя-амидопропилкарбоксиметилбетаин (Chembetaine S: Chemron Corp., Paso Robles, CA) и канло-амидопропилбетаин (Hetaine CLA: Heterene, Inc., Patterson, NJ, NJ). Существует несколько примеров, в которых азот амидной связи является четвертичным азотом (например, связью (α) служит карбонил). К ним относятся: C11 (CAS® N. 66451-67-0), C15 (CAS® N. 66516-99-2) и C17 (CAS® N. 66451-68-1). Примерами карбоксибетаинов, которые содержат этилбромид ("карбоксиэтилбетаины": R4= -CH2CH2-), метиленовую связь ( α =-CH2-) и варьируются исключительно на основе длины алкильной цепи, являются C12 (CAS® N. 16527-85-8), C13 (CAS® N. 132621-79-5), C14 (CAS® N. 69725-38-3), C16 (CAS® N. 42416-43-3) и C18 (CAS® N. 30612-73-8). Примером карбоксиэтилбетаина, у которого R2 и R3 представлены атомами азота, при подходящих условиях, может быть C18 (CAS® N. 1462-54-0 (C12-β аланин). Примерами карбоксибетаинов, имеющих пропиловый мостик ("карбоксипропилбетаины": R4=-CH2CH2CH2-), метиленовую связь ( α =-CH2-) и отличающихся исключительно по длине алкильной цепи, являются: C11 (CAS® N. 150147-53-8), C12 (CAS® N.15163-30-1), C14 (CAS® N. 146959-90-2), C15 (CAS® N. 146959-91-3), C16 (CAS® N. 71695-32-4) и C18 (CAS® N. 78195-27-4). Примером карбоксибетаина, имеющего бутиловый мостик ("карбоксибутилбетаин": R4=-CH2CH2CH2CH2-) и метиленовую связь (α =-CH2-), может быть: C12 (CAS® N. 120139-51-7). Двумя примерами карбоксибетаинов, имеющих пентиловый мостик ("карбоксипентилбетаин": R4=-CH2CH2CH2CH2CH2-) и метиленовую связь (α =-CH2-), может быть: C12 (CAS® N. 76392-97-7) и C16 (CAS® N. 73565-98-7). Примером карбоксибетаина, имеющего гексиловый мостик ("карбоксигексилбетаин": R4= -CH2CH2CH2CH2CH2CH2-) и метиленовую связь (α = -CH2-), могут быть: C12 (CAS® N. 132621-80-8). Существует несколько примеров карбоксибетаинов, у которых функцию мостика выполняет бензильная группа (R4=-CH2-C6H4-). Есть два примера C12, один из которых имеет карбокси
N. 71850-81-2) и сталлоубисгидрокси бишидрохуэтилглицинат (CAS® N. 70750- 46-8). Также исследовалось еще несколько карбоксиметилбетаинов, для которых не определены CAS® номера. К ним относятся масло проростков пшеницы - амидопропил карбоксиметилбетаин (Schercotaine WOAB: Scher Chemicals, inc., Clifton, NJ), бабассу-амидопропилкарбоксиметилбетаин (Croda, Inc., Parsippany, NJ), соя-амидопропилкарбоксиметилбетаин (Chembetaine S: Chemron Corp., Paso Robles, CA) и канло-амидопропилбетаин (Hetaine CLA: Heterene, Inc., Patterson, NJ, NJ). Существует несколько примеров, в которых азот амидной связи является четвертичным азотом (например, связью (α) служит карбонил). К ним относятся: C11 (CAS® N. 66451-67-0), C15 (CAS® N. 66516-99-2) и C17 (CAS® N. 66451-68-1). Примерами карбоксибетаинов, которые содержат этилбромид ("карбоксиэтилбетаины": R4= -CH2CH2-), метиленовую связь ( α =-CH2-) и варьируются исключительно на основе длины алкильной цепи, являются C12 (CAS® N. 16527-85-8), C13 (CAS® N. 132621-79-5), C14 (CAS® N. 69725-38-3), C16 (CAS® N. 42416-43-3) и C18 (CAS® N. 30612-73-8). Примером карбоксиэтилбетаина, у которого R2 и R3 представлены атомами азота, при подходящих условиях, может быть C18 (CAS® N. 1462-54-0 (C12-β аланин). Примерами карбоксибетаинов, имеющих пропиловый мостик ("карбоксипропилбетаины": R4=-CH2CH2CH2-), метиленовую связь ( α =-CH2-) и отличающихся исключительно по длине алкильной цепи, являются: C11 (CAS® N. 150147-53-8), C12 (CAS® N.15163-30-1), C14 (CAS® N. 146959-90-2), C15 (CAS® N. 146959-91-3), C16 (CAS® N. 71695-32-4) и C18 (CAS® N. 78195-27-4). Примером карбоксибетаина, имеющего бутиловый мостик ("карбоксибутилбетаин": R4=-CH2CH2CH2CH2-) и метиленовую связь (α =-CH2-), может быть: C12 (CAS® N. 120139-51-7). Двумя примерами карбоксибетаинов, имеющих пентиловый мостик ("карбоксипентилбетаин": R4=-CH2CH2CH2CH2CH2-) и метиленовую связь (α =-CH2-), может быть: C12 (CAS® N. 76392-97-7) и C16 (CAS® N. 73565-98-7). Примером карбоксибетаина, имеющего гексиловый мостик ("карбоксигексилбетаин": R4= -CH2CH2CH2CH2CH2CH2-) и метиленовую связь (α = -CH2-), могут быть: C12 (CAS® N. 132621-80-8). Существует несколько примеров карбоксибетаинов, у которых функцию мостика выполняет бензильная группа (R4=-CH2-C6H4-). Есть два примера C12, один из которых имеет карбокси
группу в положении 2 или орто (CAS® N. 71695-34-6). Существует два C16 примера, у одного из которых карбокси группа находится в положении 4, или пара (CAS® N. 71695-33-5), а у другого карбокси группа в положении 2, или opто (CAS® N. 71695-35-7). Следовательно, "карбоксибетаин-подобные" ("CB-подобные") включают ту самую SB-18-подобную бетаиновую структуру, которая имеет карбоксильную группу в качестве аниона (γ-COO⊖), как показано в табл. 2, и включают все возможные комбинации R1, α, R2, R3, β и R4, как было показано в данной работе выше.
Второй большой группой бетаинов являются сульфобетаины (γ=SO N. 58930-08-8). Пример "сульфогексилбетаина" (R4= -CH2CH2CH2CH2CH2CH2-): C16 (CAS® N. 13262111-81-9). Несколько примеров веществ, у которых мостиком является бензильная группа (R124=-CH2-C6H4-). К ним оносятся: C12 (CAS® N.65180-40-7), C14 (CAS® N. 65180-41-8), C16 (CAS® N. 65180-42-9) и C18 (CAS® N. 65180-43-), у которых сульфонат в позиции 4 (орто). Также несколько примеров веществ, у которых последние 4 молекулы связаны алкильной цепью с четвертичным азотом амидопропильной группой: C12 (CAS® N. 65180-44-1), C14 (CAS® N. 65180-45-2), C16 (CAS® N. 65180-46-3) и C18 (CAS® N. 65180-47-4). Следовательно, "сульфобетаин-подобные" (SB-подобные) соединения имеют в составе молекулы такие SB-подобные бетаиновые структуры, у которых сульфонат выполняет функцию аниона γ = -SO
N. 58930-08-8). Пример "сульфогексилбетаина" (R4= -CH2CH2CH2CH2CH2CH2-): C16 (CAS® N. 13262111-81-9). Несколько примеров веществ, у которых мостиком является бензильная группа (R124=-CH2-C6H4-). К ним оносятся: C12 (CAS® N.65180-40-7), C14 (CAS® N. 65180-41-8), C16 (CAS® N. 65180-42-9) и C18 (CAS® N. 65180-43-), у которых сульфонат в позиции 4 (орто). Также несколько примеров веществ, у которых последние 4 молекулы связаны алкильной цепью с четвертичным азотом амидопропильной группой: C12 (CAS® N. 65180-44-1), C14 (CAS® N. 65180-45-2), C16 (CAS® N. 65180-46-3) и C18 (CAS® N. 65180-47-4). Следовательно, "сульфобетаин-подобные" (SB-подобные) соединения имеют в составе молекулы такие SB-подобные бетаиновые структуры, у которых сульфонат выполняет функцию аниона γ = -SO
Большая подгруппа сульфобетаинов, которые обычно приводятся раздельно ввиду их выраженно различных характеристик и от которых от всех было бы обоснованно ожидать проявления SB-подобной активности, представлена гидроксипропилсульфобетаинами (HSB). Эти детергенты не подвергались интенсивному исследованию ввиду их низкой растворимости. Большинство этих сульфобетаинов содержит мостик в виде 2-гидроксипропила (R4=-CH2CH(OH)CH2-). Они содержат прямые алкильные цепи: C10 (CAS® N. 34135-76-7), C12 (CAS® N. 13197-76-7), C14 (CAS® N. 13177-42-9), C15 (CAS® N. 71502-45-9), C16 (CAS® N. 7425-12-9) и C18 (CAS® N. 19223-56-4); а также молекулы с амидопропильной связью: C12-амидо (CAS® N. 19223- 55-3), C14-амидо (CAS® N. 63663-10-5), C16-амидо (CAS® N. 63663-11-6) и C18-амидо (CAS® N. 63663-12-7). Также несколько примеров молекул C14, у которых алкил не является прямой цепью: (CAS® N. 56505-82-9) и (CAS® N. 71497-51-3). Следовательно, "гидроксисульфобетаин-подобные" (HSB-подобные) соединения содержат эти SB-18-подобные бетаиновые структуры, связанные гидроксипропильным мостиком (R4=-CmH2m-1(OH)-), с сульфонатом в качестве аниона (γ = -SO
Третьей группой этого семейства детергентов, также хорошо изученной, являются фосфобетаины. Представители фосфобетаинов (γ = -OPO
метиленовую связь (α =-CH2-) и отличаются исключительно длиной алкильной цепи. К ним относятся: C10 (CAS® N. 134842-83-4), C11 (CAS® N. 134842-84-5), C12 (CAS® N. 126712-86-5), C14 (CAS® N. 126712-87-6), C16 (CAS® N. 126712-88-7), C17 (CAS® N. 145578-49-0) и C18 (CAS® N. 126712-89-8). У двух фосфоэтилбетаинов алкил имеет двойную связь: C18:1 (CAS® N. 134590-60-6 и (CAS® N. 148716-30-7). Несколько примеров веществ, относящихся к C16-фосфоэтилбетаинам, где R3 и R4 фрагменты существенно варьируют. К ним относятся: N,N-диэтил (CAS® N. 126712-90-1); N,N-дипропил (CAS® N. 126712-91-2); N,N-дибутил (CAS® N. 126712-92-3); N-этил-N-пропил (CAS® N. 126712-93-4); N- метил-N-этил (CAS® N. 134842-85-6) и N-этил-N-бутил (CAS® N. 126712-94-5). Примером фосфобетаинов, которые содержат пропиловый мостик ("фосфопропилбетаины": R4=-CH2CH2CH2-), могли бы быть: C16-фосфопропилбетаин (CAS® N. 89367-17-9). Примером фосфобетаина, содержащего бутиловый мостик ("фосфобутилбетаин": R4= -CH2CH2CH2CH2-), может быть: C16фосфобутилбетаин (CAS® N. 13842-86-7), примером фосфобетаина, содержащего гексиловый мостик ("фосфогексилбетаин": R4=-CH2CH2CH2 CH2CH2CH2-), может быть: C16-фосфогексилбетаин (CAS® N. 13482-87-8). Несколько примеров n-гидроксиалкилфосфоэтилбетаинов (например, они содержат гидроксильную связь): C12 (CAS® N. 124591-53-3), C14 (CAS® N. 124591-54-4) и C16 (CAS® N. 124591-57-7). Гидроксипропилфосфобетаины содержат амидопропильную связь:
C12-амидо (CAS® N. 73602-79-6) и C18-амидо (CAS® N. 144077-12-3). Несколько примеров фосфоэтилбетаинов, содержащих гидроксипропиловую группу в сочетании с эфирной группой, как связи: C10 (CAS® N. 128506-41-2), C12 (CAS® N. 128506-42-3) и C14 (CAS® N. 128506-46-7). Gallot, В. et al., J. Colloid Interface Sci. 121:514-521 (1988) описывает серию фосфобетаинов, у которых этильная группа была введена для модификации аниона (-OPO
В четвертой группе бетаинов содержатся вещества, имеющие в качестве аниона сульфат (-OSO = -CH2-) и отличающиеся исключительно длиной алкильной цепи, к которым относятся: C10 (CAS® N. 92764-22-2), C12 (CAS® N. 15163-35-6), C14 (CAS® N. 58930-12-4) и C16 (CAS® N. 34236-95-8). Вещества с бутиловым мостиком ("сульфатобутилбетаин": R4=-CH2CH2CH2CH2-), с метиленовой связью (α =-CH2-) и отличающиеся исключительно длиной алкильной цепи: C12 (CAS® N. 58930-14-6) и C16 (CAS® N. 58930-15-7). Несколько примеров сульфатов, имеющих амидопропильную связь: C16-амидопропилсульфатоэтилбетаин (CAS® N. 58930-11-3), C16-амидопропилсульфатопропилбетаин (CAS® N. 58930-13-5), C13-амидопропилсульфатобутилбетаин (CAS® N. 144077-11-2) и C16-амидопропилсульфатобутилбетаин (CAS® N. 58930-16-8). Таким образом, "сульфатобетаин-подобные" ("StB-подобные") вещества имеют SB-18-подобные бетаиновые структуры, представленные в табл. 2 (сульфат выступает в роли аниона (γ =-OSO3SO
= -CH2-) и отличающиеся исключительно длиной алкильной цепи, к которым относятся: C10 (CAS® N. 92764-22-2), C12 (CAS® N. 15163-35-6), C14 (CAS® N. 58930-12-4) и C16 (CAS® N. 34236-95-8). Вещества с бутиловым мостиком ("сульфатобутилбетаин": R4=-CH2CH2CH2CH2-), с метиленовой связью (α =-CH2-) и отличающиеся исключительно длиной алкильной цепи: C12 (CAS® N. 58930-14-6) и C16 (CAS® N. 58930-15-7). Несколько примеров сульфатов, имеющих амидопропильную связь: C16-амидопропилсульфатоэтилбетаин (CAS® N. 58930-11-3), C16-амидопропилсульфатопропилбетаин (CAS® N. 58930-13-5), C13-амидопропилсульфатобутилбетаин (CAS® N. 144077-11-2) и C16-амидопропилсульфатобутилбетаин (CAS® N. 58930-16-8). Таким образом, "сульфатобетаин-подобные" ("StB-подобные") вещества имеют SB-18-подобные бетаиновые структуры, представленные в табл. 2 (сульфат выступает в роли аниона (γ =-OSO3SO
Фосфоний-подобные бетаины (β = -P⊕-) были описаны Gaertner, V.R. et al. (патент США 2828332). На основании данных и гипотез, выдвинутых в этой работе, "фосфонийбетаино-подобные" ("PhB-подобные") соединения содержат все возможные SB-18-подобные комбинации бетаинов, имеющие фосфоний катион, как показано в табл. 2, и все возможные комбинации R1, α, γ, R2, R3, и R4, как было показано в данной работе.
Как фосфоний бетаины, "сульфониумбетаино-подобные" вещества ("SoB-подобные") относятся к SB-18-подобным, тем, у которых β = -S⊕-, как показано в табл. 2, включая все возможные комбинации R1, α, γ, R2, R3 и R4, как было показано в данной работе.
В табл. 3 представлены некоторые необычные бетаиноподобные структуры. К ним относятся "обратные бетаины" (RevB), оксидоподобные амины (AOx) детергенты, c-алкилбетаины (cAB) и производные имидазолинийбетаина (ImB). Они легко описываются с помощью номенклатуры в табл. 2, и, возможно, наиболее удобны для того, чтобы спрогнозировать возможные варианты структур при изменении алкила (R1): алкил может быть связан как с катионом (β), так и с анионом (γ) или с мостиком (R4), все варианты являются бетаин-подобными структурами. Например, в случае n-алкил бетаинов гидрофобный домен (R1) является ковалентно связанным с катионом, в то время как в случае обратных бетаинов алкил связан с анионом (-γ⊖-), и в случае с c-алкилбетаинами алкил связан с мостиком (R4).
Обратные бетаины схожи с n-алкилбетаинами в том, что противоположный заряд (в случае катиона -[β]⊕- соприкасается с анионом через мостик (-R4-). В типичном случае катионом является (-[β]⊕-) четвертичный азот и такие группы, как R2 и R4, связаны по этой позиции.
c-алкилбетаины структурно отличны в том, что алкил ковалентно связан с мостиком (R4) как противовес и к катиону (-[β]⊕-),, и к аниону (-γ⊖-).
В случае оксидов амина n-алкилбетаиновая структура сохраняется; однако, β определяется как четвертичный азот, и γ - как оксид. Предполагается, что мостик минимизирован: (R4) и является простой ковалентной связью N-О. Существуют другие оксидоподобные структуры, схожие с оксидом амина, как например оксиды фосфония, от которых можно обоснованно ожидать требуемого эффекта в методах по данному изобретению.
Цвиттерионы типа имидазония являются детергентами, которые также продаются как амфотерные бетаины. В свете требований, центр положительного заряда не является радикалом, но нужен модифицированный имидазолин, у которого резонантная структура образует катион. Классическая мостиковая структура сохраняется и алкил может быть атакован в любой точке кольца.
Таблица 3: Дополнительные бетаиноподобные структуры
Представлены структуры, альтернативные бетаину. Сохранена номенклатура, использовавшаяся в табл. 2
Т.к. структуры в табл. 3 были изучены незначительно, они не получили так много внимания, как n-алкилбетаины (табл. 2) и, следовательно, представляют лишь малую долю структурных исследований, выполненных на бетаинах. Невзирая на это, представители нескольких описанных выше классов использовались в методах данного изобретения. Взятый в работу представитель обратных бетаинов, алкил 2-гидрокси-3-триметиламмониопропил фосфат (C16-AHTMAP: CAS® N. 99485-86-6), был описан Kurosaki, T. et al., Chem. Soc. Japan 11:1297-1301 (1990) и был получен в дар от KAO Chemical (Japan). Оксид амина, Ammonyx MO (оксид миристилдиметиламина: CAS® N. 3332-27-2) был получен как образец от Stepan Company, Northfield, IL. Использовавшийся c-алкилбетаин (CAS® N.96-55-9) и c-цетилбетаин (CAS® N. 95-56-0) был получен в качестве образца от R. T. Vanderbilt, Norwalk, CT.
Исследовалось несколько групп обратных бетаинов, можно было ожидать, что несколько образцов алкил 2-гидрокси-3-триметиламмониопропилфосфатов (Cn-AHTMAP), исходно описанных Kurosaki, T. et al., Chem. Soc. Japan 11: 1297-1301 (1990), могли дать эффект в методах по данному изобретению. К ним относились C12 (CAS® N. 99485-91-3), C14 (CAS® N. 132630-63-8), C16 (CAS® N. 99485-86-6), C18 (CAS® N. 99485-87-7). Zimmer, R.E. et al. (патент США 3560393) описал синтез нескольких интересных обратных бетаинов, которые все можно было обоснованно считать потенциально активными в методах по данному изобретению. Сюда входят вещества, которые объединяют катион аммония (β = -N⊕(CH3)3) с фосфинатанионом (γ = -PO
(β = -N⊕(CH3)3) с фосфонатанионом (γ = -PO
Варианты N,N-оксидов диметиламина, от которых обоснованно предполагалось получить активность в методах по данному изобретению, включают: C12 (CAS® N. 1643-20-5), C14 (CAS® N. 3332-27-2), C16 (CAS® N. 7128-91-8), C18 (CAS® N. 2571-88-2) и C22 (CAS® N.26483-35-2). Примером оксида амина, имеющего амидопропильную часть, может быть C18:1амидо (CAS® N. 14351-50-9. Это примеры веществ, у которых R1 получены от природных масел: кокамидопропил (CAS® N. 68155-09-9), бабассуамидопропил (CAS® N. 124046-26-0). Таким образом, "оксидоподобные" ("AO-подобные") вещества, показанные на табл. 3, могли быть определены как структуры, у которых γ = O⊖, R4 представляет ковалентную связь β-O, и которые включают все возможные комбинации R1, α, β, R2 и R3, что следует из табл. 2 и как было показано в данной работе.
Варианты c-алкилбетаинов (γ = COO⊖, β = -N⊕(CH3)3, R4=-CH-), от которых обоснованно предполагалось получить активность в методах по данному изобретению, включают: C10 (CAS® N. 96-55-9), C12 (CAS® N. 686-83-9), C14 (CAS® N. 16545-85-0), C16 (CAS® N. 95-56-0) и C18 (CAS® N. 686-84-0). Следовательно, "c-алкилбетаиноподобные" вещества ("cAB-подобные"), как показано в табл. 3, могли бы быть идентифицированы как такие вещества, у которых R1 присоединен к R4 и которые включают все возможные комбинации α, β, γ, R1, R2, R3 и R4, что следует из табл. 2 и как было показано в данной работе.
В качестве примера имидазольного бетаина может быть приведено вещество, у которого R4 происходит из масла кокосовых орехов (CAS® N. 68334-21-4). Следовательно, все вещества "подобные имидазольному бетаину" (ImB подобные), как можно было бы обоснованно предполагать, могут проявлять активность, необходимую для способов по данному изобретению, и могут, таким образом, быть определены как такие структуры, у которых β, по определению, имидазольного действия, и которые включают все возможные комбинации α, γ, R1, R2, R3 и R4, что следует из табл. 2 и как было показано в данной работе.
Следовательно, как "бетаиноподобные", в данной работе обозначаются все вещества, структуры которых описаны в табл. 2 и 3, включая, например, CB-подобные, SB-подобные, HSB-подобные, PB-подобные, StB-подобные, PhB-подобные, SoB-подобные, RevB-подобные, AO-подобные, cAB- подобные и ImB-подобные, которые проявляют SB-18-подобную активность. Следует отметить, что как в случае с Gallot, B. et al, J. Colloid Interface Sci. 121:514-521 (1988), модификации по положению γ, которые не приводят к изменению SB-18-подобной активности веществ, могут тоже быть включены в определяемую группу. Далее, включение нескольких зарядов в состав одной молекулы (например, двух или более, в числе которых по меньшей мере два заряда противоположных знаков), могло дать дипольный момент таким образом, что действие было сопоставимо с действием бетаина, как описано в данной работе, что проявляется как SB-18-подобная активность, которую также можно рассматривать как бетаиноподобную.
Применение бетаиноподобных детергентов
Применение бетаиноподобных свойств зависит от длины алкильной цепи, комбинации зарядов и структуры мостика. Длинные алкильные цепи, такие, как C16-C20, предпочтительнее из-за их способности разрушать тяжистые структуры и их очевидной способности более эффективно удаляться. Однако, пониженная растворимость таких длинноцепочечных алкилов может препятствовать эффективному применению таких бетаинов в способах по данному изобретению. Фосфобетаины, карбоксибетаины или сульфобетаины предпочтительнее из-за повышенной растворимости, обеспечиваемой их анионами, однако, карбоксибетаины и сульфобетаины более легко синтезируемы. Карбоксибетаины имеют характеристики более общие, присущие ионным детергентам, напротив, сульфобетаины проявляют свойства, более напоминающие свойства неионных детергентов. Мостик в виде прямой цепи является предпочтительным и пропильная группа является наиболее предпочтительной структурой мостика, потому что она (а) позволяет осуществлять "всаливание" детергента и (б) имеет наименьшую бактериостатическую активность. Метиленовый мостик наименее желателен, потому что он проявляет свойство высаливания. Метиленовая связь предпочтительна; однако, хотя амидопропильные связи не мешают проявлению активности всеми SB-18-подобными детергентами, они явно повышают неионные свойства. Таким образом, выбор SB-18-подобного детергента для использования в способах по данному изобретению может быть различным в зависимости от целей применения или системы, используемой для определения, или от желаемой бактериостатической активности. Например, если объектом исследования являются микроорганизмы, которые не группируются между собой, например М. paratuberculosis, эти свойства ассоциируются с диспергированием и ионная характеристика не является существенной. При таких обстоятельствах могут использоваться бетаины с более короткими цепями и меньшим проявлением ионных свойств. Напротив, если задача состоит в определении микроорганизмов, образующих при росте группы клеток, например М. tuberculosis, существенными являются ионный статус и более длинные цепи. Поскольку длина цепи и длина мостика являются важными детерминантами бактериостатической активности, бетаиноподобные структуры могут быть изменены для того, чтобы облегчить определение способом амплификации. Например, несмотря на то, что в приводимых в данной работе примерах используется ПЦР как предпочтительный метод определения, как обсуждалось выше, способы по данному изобретению могут облегчить определение также с применением других методов амплификации. Можно ожидать, что бетаин, который действует оптимально при применении ПЦР, может использоваться (но не обязательно, это будет оптимально) для других способов, например для лигазной цепной реакции (ligase chain reaction - LCR), амплификации по типу замещения цепи (strand displacement amplification - SDA) или системы амплификации на базе транскрипции (NASBA, 3SR или LAT), например. Далее приведено руководство по применению с целью разъяснить аспекты бетаиноподобных структур и их функции для того, чтобы проинструктировать специалистов в плане возможных модификаций SB-18-подобных детергентов, что может помочь в выборе оптимального бетаина для конкретного применения. Согласно приведенным выше ограничениям можно ожидать, что C18-карбоксипропилбетаин ( N. 78195-27-4) обеспечит идеальное сочетание растворимости, длины цепи и структуры мостика для широкого круга применений с использованием ПЦР, и поэтому методы по данному изобретению, использующие данное вещество, являются весьма высокопредпочитаемым вариантом воплощения изобретения. CAS® N 78195-27-4 может быть синтезировано с использованием технологий, например таких, как описаны в заявке N. 79100920[54- 100920] (Япония) и в патенте 81251394 (Япония).
N. 78195-27-4) обеспечит идеальное сочетание растворимости, длины цепи и структуры мостика для широкого круга применений с использованием ПЦР, и поэтому методы по данному изобретению, использующие данное вещество, являются весьма высокопредпочитаемым вариантом воплощения изобретения. CAS® N 78195-27-4 может быть синтезировано с использованием технологий, например таких, как описаны в заявке N. 79100920[54- 100920] (Япония) и в патенте 81251394 (Япония).
SB-18 в концентрациях ниже 2 мкМ не диспергирует бактерии; концентрации от 2 мкМ до 2 mM были наиболее эффективными, вероятно, как соответствующие критической концентрации мицеллообразования (critical micellar concentration (CMC)); SB-18 в концентрациях намного выше 2 mM были трудны для применения в виде раствора из-за их нерастворимости. Специалисту понятно, что эти рассмотрения будут изменяться в зависимости от того, какой именно детергент применяется. Например, с более короткой цепью детергенты будут нуждаться в более высоких концентрациях для превышения CMC, но температуры Краффта будут достигнуты при более низкой концентрации. Следовательно, концентрации в от 10 до 1000 крат выше CMC детергента, который может быть использован, и соотношение температуры исследования и температуры Краффта SB-подобного детергента должны быть тщательно откоррелированы. При использовании SB-18 необходимо применять среду с электролитами (10 mM - 100 mM соли) и нагревать раствор, например, до температуры от 37 до 42oC, но не ниже 37oC для того, чтобы сохранять детергент в мицеллярной форме. Варьируя SB-18-подобный детергент, который применяется в методах по данному изобретению, как обсуждается далее, можно устранить потребность в поддержании в теплом состоянии SB-18-подобного детергента. Например, при использовании C18- карбоксипропилбетаина (CAS® N. 78195-27-4) в концентрации, при которой не лизируются клетки (как правило ниже 2 mM), отпадает необходимость соблюдать жесткие температурные рамки для SB-18.
Как правило, температуры выше 50oC способствуют лизису. Следовательно, поскольку способы по данному изобретению создают преимущества в плане повышения эффективности выделения путем центрифугирования, только если бактериальные клетки остаются целыми, следует стараться избегать температур выше 50oC.
pH промывного раствора, содержащего детергент, следует поддерживать в диапазоне выше 5, но ниже 9. Ниже pH 5 SB-18-пoдoбныe дeтepгeнты могут начать вести себя несвойственным образом, а при pH выше 9 SB-18-подобные детергенты могут проявиться как анионные детергенты и будут высаливаться. Однако, любое значение pH, которое позволяет SB-18-подобному детергенту проявлять SB-18-подобную активность, является приемлемым. Время промывки с использованием такого детергента должно быть не менее 60 мин, чтобы произошло аккумулирование детергента. Инкубирования вплоть до 18 часов дают незначительное улучшение результатов по сравнению с 60-минутной инкубацией.
Бетаины как детергенты
Как правило, принято выделять три класса детергентов: ионные, включая как катионные, так и анионные; неионные, и амфотерные (Kirk-Othmer Encyclopedia of Chemical Technology, 3rd ed., vol. 22, John Wiley & Sons, New York (1983), p.332-387). Специфический класс детергентов, к которому бы принадлежали бетаины, единым образом не выделен. Согласно формальному подходу на основе зарядов некоторые специалисты классифицируют бетаины как "амфотерные", однако, Laughlin, R.G. Langmuir 7:842-847 (1991) приводит аргументы о том, что эти молекулы являются подклассом неионных детергентов. Очевидно, что бетаины обладают определенными специфическими характеристиками, что ставит их в категорию, отличную как от неионных, так и от ионных детергентов. Бетаины ведут себя в некоторых отношениях как ионные детергенты, и в других аспектах как неионные детергенты. Является установленным фактом, что в зависимости от структуры одна и та же молекула может проявлять как ионные детергентные свойства в соответствии с определенными условиями, так и неионные детергентные свойства в других условиях.
Детергенты как правило описывают на основании их физических характеристик и фазового поведения в рамках зависимости температуры (ось X) от концентрации (ось Y). В самых простых понятиях можно выделить три фазы применительно к чистому детергенту: мономерная, твердая и мицеллярная фазы. Каждый детергент может быть описан на основании системы параметров, при которых данная фаза существует. Изменения параметров системы вызывают переходы между фазами, что затем используется для дальнейшей характеристики поведения. Данная общая информация взята из: Shinoda, K., Nakagawa, T., Tamamushi, B. и Isemura, T. eds. Colloidal Surfactants, Academic Press, New York (1963); Schick, M. J. ed. Surfactant Series, vol. 1: Nonionic Surfactants, Marcel Dekker, New York (1967); Jungermann, E. ed., Surfactant Science Series, vol. 4: Cationic Surfactants, Marcel Dekker, New York (1970): Bluestein, B.R. и Hilton, C. L, eds., Surfactant Science Series, vol. 12: Amphoteric Surfactants, Marcel Dekker, New York (1982); Gloxhuber, C. и Kunstler, K. eds., Anionic Surfactants, Marcel Dekker, New York (1992).
Например, все детергенты в некоторой степени растворимы в водных растворах. При низких концентрациях существуют только мономеры. При возрастании концентрации если допустить, что температура достаточно высокая, происходит фазовый переход и начинают формироваться мицеллы. При этих условиях и в большинстве случаев дальнейшее увеличение концентрации способствует только увеличению концентраций мицелл. Концентрация, при которой происходит этот переход, называется "критической мицеллярной концентрацией" (CMC).
Так долго, как система находится выше критической температуры, CMC фактически является постоянной концентрацией для всех значений температуры. Следовательно, в целях данного обсуждения, CMC может наблюдаться как параллельная описанной выше оси X линия. Если система находится ниже этой критической температуры, наблюдается подвижный переход между мономерной фазой и твердой фазой, независимо от концентрации. Когда температура повышается, при увеличении концентрации выше CMC наблюдается фазовый переход между твердой и мицеллярной фазами. Эта температура, известная как температура Краффта (Krafft, F. et al., Ber. 28:2566-2573 (1895)), обычно упоминается как точка плавления гидратированных кристаллов, и совершенно так же не зависит от концентрации. Следовательно, для цели данного обсуждения, температура Краффта может быть представлена как линия, парллельная оси Y.
Точка, в которой CMC и температура Краффта пересекаются, известна как точка Краффта. Существенным в рамках данного изображения является характеристика размера (мицеллярный молекулярный вес и агрегирующее число) и формы сформировавшихся мицелл. Беря за основу эти рамки (температура, концентрация и система растворителей, включая различные электролиты и концентрации электролитов), за последнее столетие определили уникальные фазовое и мицеллярное поведение различных химических структур. Последующее обсуждение концентрируется на вопросах, связанных с клиническим аспектом (например, водные растворы), и подробно останавливается на таких характеристиках, которые, предположительно, важны для проявления свойств SB-18 и SB-18-подобных детергентов в целом и включены как руководство для специалистов, стремящихся оптимизировать бетаиноподобный детергент по данной заявке, например, при определении методом амплификации, но не ПЦР.
Поскольку CMC детергентов уменьшается по мере увеличения длины алкильной цепи, CMC неионных детергентов как правило на два порядка ниже, чем у ионных гомологов с той же длиной цепи. Это означает, что мономер неионного гомолога менее растворим. Однако, это различие строго зависит от вида и числа неионных частей молекулы. Например, Hsiao, L. et al., J. Phys. Chem. 60: 657-660 (1956) показали, что CMC полиоксиэтиленфениловых простых эфиров возрастает по мере увеличения числа полиоксиэтиленовых фрагментов в молекуле. Tokiwa, F. et al., Bull. Chem. Soc. Japan 35:1737-1739 (1954) и Schick, M.J. et al. , J. Phys. Chem. 66:1326-1333 (1962) показали, что мицеллярный вес и агрегирующее число возрастают вместе с увеличением числа единиц полиоксиэтилена. С другой стороны ионные детергенты показали лишь незначительные различия CMC у различных ионных образцов. Shinoda (Shinoda, K., Colloidal Surfactants, Academic Press, New York (1963) pp. 54-55) для того, чтобы проиллюстрировать эти незначительные различия, обобщил данные по нескольким различным ионным гомологам.
Напротив, CMC бетаинов зависит от дистанции, разделяющей заряды, также, как и от природы специфичных ионных групп. Например, Lattes, A. et al., Surfactants Soln. 11:127-1139 (1991) показали, что CMC серии карбоксибетаинов в воде возрастает по мере продвижения длины мостика от метила к этилу, пропилу. Weers, J. G. et al. , Langmuir 7:854-867 (1991) опровергают эти данные, показывая обратную тендецию. Weers, J.G. et al., Langmuir 7:854-867 (1991) далее показали, что: (i) CMC серии сульфобетаинов сначала возрастает по мере продвижения от пропила к бутилу, а затем снижается по мере продвижения от бутила к гексилу, и (ii) величины CMC карбоксибетаинов ниже, чем аналогичные значения у сульфобетаинов. Интересно, что по этому вопросу из результатов Nandakumar T. N. et al. , J. Oil Tech. Assoc. India 11:31-34 (1979) следует вывод, противоположный выводу Weers, J.G. et al., Langmuir 7: 854-867 (1991): величины CMC карбоксибетаинов выше, чем соответствующие величины у сульфобетаинов. Анализ зависимости CMC от ионной части молекулы может быть сделан путем сравнения величин CMC у C16- карбоксиэтилбетаина (8,3•10-5 М: Lattes, A. et al. , Surfactants Soln. 11:127-1139(1991)) с C16-фосфоэтилбетаином (4•10-5 М: Tsubone, K. et al., J. Am. Oil Chem. Soc. 67: 149-153 (1990)) и с C16-сульфоэтилбетаином (нерастворим: Weers, J.G. et al. Langmuir 7:854-867 (1991); Parris, N. et al., J. Am. Oil Chem. Soc. 53: 97- 100 (1976) сообщают о точке Краффта у C16-сульфоэтилбетаина >90oC).
Очевидно, что при сравнении с ионными детергентами для бетаинов выясняется более полное соотношение поверхностной активности с числом ионных групп. В действительности, противоречивые результаты можно объяснить чрезвычайной чувствительностью данных молекул к условиям эксперимента.
Изменение CMC в зависимости от длины цепи было описано с помощью приводимого ниже уравнения (Kevens, Н.В., et al., J. Am. Oil Chem. Soc. 30:74-80 (1953)), в котором A - константа для гомологического ряда и В - вторая константа, используемая для демонстрации зависимости CMC от длины цепи (m) для того же ряда:
LgC = A-Bm
Shinoda (Shinoda, K.B: Colloidal Surfactants, Academic Press, New York (1963) pp. 42-44) вывел значения B для различных детергентов. В целом, значения B ионных детергентов находятся на уровне Lg2 (например, 0,3) и Lg3 (например, 0,5) для неионных детергентов. Beckett, A.H. et al., J. Pharm. Pharmacol. 15: 422-431 (1963); Molyneux, P. et al., Trans. Faraday Soc. 61: 1043-1052 (1965); и Lattes, A. et al., Surfactants Soln. 11:127-1139 (1991) сообщают, что значение B для гомологического ряда карбоксибетаинов приблизительно составляет Lg 3. Weers, J.G. et al., Langmuir 7:854-867 (1991) определяют величину В для сульфопропилбетаинового ряда как 0,48.
Очевидно, что бетаины проявляют себя как неионные детергенты, что означает, что CMC показывает более строгую зависимость от длины алкильной цепи, чем у соответствующих ионных гомологов. Например, CMC бетаинов снижается логарифмически при добавлении каждых последующих двух атомов углерода (это четко показано в результатах исследования Nandakumar T.N. et al., J. Oil Tech. Assoc. India 11:31-34 (1979), а сравнение между ионными детергентами и бетаинами показано графически в работе Herrmann, K.W. J. Colloid Inter. Sci. 22: 352-359 (1966)). Однако, как обсуждалось выше, эта тенденция в некоторой степени нарушается в зависимости от длины мостика и от числа ионных групп. Например, гомологический ряд алкилгидроксифосфоэтилбетаина в публикации Tsubone, K. et al., J. Am. Oil Chem. Soc. 67:394-399 (1990) и гомологический ряд алкилфосфоэтилбетаина в публикации Tsubone, K. et al., J. Am. Oil Chem. Soc. 67:149-153 (1990) не показали такую степень зависимости. По данным этих авторов величина В составляет 0,1025 и 0,025, соответственно, что выражает даже меньшую зависимость от длины алкильной цепи, чем у их ионных гомологов. Такое отклонение фосфобетаинов отмечено Tsubone et al., J. Am. Oil Chem. Soc. 67:149-153 (1990). Эти результаты означают, что существует зависимость между кислото-щелочными свойствами и создаваемым дипольным моментом. Кроме того, показано общее соотношение между свойством, структурой и составом.
Ответы на изменения температуры возможно являются наиболее отличительными свойствами ионных и неионных детергентов. Например, повышение температуры вызывает повышение CMC у ионных детергентов и уменьшение у неионных детергентов. В работе Kuriyama, K. Kolloid-Z 180:55-64 (1961) сравниваются додецилсульфат натрия и метоксиполиоксиэтилендециловый эфир с целью четко продемонстрировать подобную дихотомию. Структуры, свойственные бетаинам, являются pH зависимыми. Tsubone et al. показали, что гидроксиалкилфосфобетаины (Tsubone, K. et al., J. Am. Oil Chem. Soc.67:394-399 (1990)) и фосфоэтилбетаины (Tsubone, K. et al., J. Am. Oil Chem. Soc.67:394-399 (1990)) действуют как ионные детергенты при pH 11,0, и как неионные детергенты при pH 6,0. CMC при pH 2,0 не изменяется под действием температуры. Следовательно, может получиться, что при низких pH некоторые бетаины поведут себя необычным образом, при высоких значениях pH все бетаины действуют как анионные детергенты и, интересно, что при средних значениях pH, при наличии цвиттериона, бетаины проявляют неионную структуру: CMC снижается при увеличении температуры.
На основании вышесказанного следует, что изменения мицеллярного веса и агрегационного числа в зависимости от температуры являются предсказуемыми. Мицеллярный вес и агрегационное число ионных детергентов медленно уменьшается по мере увеличения температуры (Nakagawa, T. et al., в: Colloidal Surfactants, Academic Press, New York (1963) pp. 123-126). С другой стороны, мицеллярный вес и агрегационное число неионных детергентов постепенно увеличивается экспоненциально по мере увеличения температуры (см.: Balmbra, R.R. et al. , Trans. Faraday Soc. 58:1661-1667 (1962) и Kuriyama, K. Kolloid-Z 180: 55-64 (1961)). Такое поведение является характерным свойством неионных детергентов и проявляется в явлении, известном как точка мутности, или как высшая критическая температура растворения.
Точка мутности фиксируется как начало появления мутности и предполагается, что она вызывается дегидратацией гидрофильной части молекул неионного детергента, что приводит к снижению растворимости. Следует отметить, что точка мутности, в отличие от температуры Краффта, не четко определяется температурой. В сущности, мицеллы все еще присутствуют; однако, вес мицелл и агрегатное число понемногу возрастают с возрастанием температуры, и, если температура продолжает повышаться, со временем происходит фазовое разделение. Nilson, P. et al., J. Phys. Chem. 88:6357-6362 (1984) показали аналогичное явление на примере сульфатобетаинов. Те же авторы показали, что сульфобетаины проявляют аналогичное свойство в гораздо меньшей степени, тогда как карбоксибетаины таким свойством не обладали. По существу Bhatia, A. et al. Colloid and Sufraces 69:277-292 (1993) показали, что мицеллярный вес и агрегатное число C18-карбоксиметилбетаинов уменьшается по мере увеличения температуры, что является характерной ответной реакцией ионных детергентов. Эти результаты означают, что ионные/неионные свойства бетаинов могут быть выбраны путем изменения нужным образом зарядов (например, дипольного момента).
Также эти детергенты различаются по изменению CMC в ответ на присутствие электролитов. Например, повышение концентрации электролитов вызывает лишь слабое уменьшение CMC неионных детергентов (Shinoda, K. et al., Bull. Chem. Soc. Japan 34:237-241 (1961)). Напротив, ионные детергенты показывают заметное уменьшение CMC при схожем изменении концентрации электролитов (Schwuger, V.M.J. Kolloid-Z. 233:979-985 (1969)). В результате происходит "высаливание" ионных детергентов. Иначе говоря, в то время как растворимость гегенионов повышается, растворимость за счет алкильной части молекулы понижается. Следовательно, температуру следует повышать, чтобы подобрать требуемый нагрев смеси алкильных цепей. На это явление также ссылаются как на поднятие точки Краффта (см.: Tartar, H.V. et al., J. Phys. Chem. 43:1173-1179 (1940); Tartar, H. V. et al., J. Am. Chem. Soc. 61:539-544 (1939); Nakayama, H. et al. , Bull. Chem. Soc. Japan 40:1797-1799 (1967); и Tsujii, K. et al., J. Phys. Chem. 84:2287-2291 (1980), приведены в качестве примеров).
Наиболее часто употребимые неионные детергенты имеют точки Краффта ниже 0oC, и, следовательно, не проявляют таких свойств. Если редуцируется гидрофильная часть молекулы, некоторые неионные детергенты несмотря на это действительно показывают более низкое значение пороговой растворимости. Schott, H. et al. , J. Pharm. Sci. 65:979-981 (1976) показали, что Brij 56 (оксид цетилполиоксиэтилена (C16E10): CAS® N.9004-95-9) и Brij 76 (оксид стеарилполиоксиэтилена (C18E10): CAS® N.9005- 00-9) могут быть высалены. Однако, можно отметить, что высаливание, наблюдавшееся Schott, H. et al., J. Pharm. Sci. 65: 979-981 (1976), гораздо менее выражено, чем высаливание, наблюдавшееся Tartar, H.V. et al., J. Phys. Chem. 43:1173-1179 (1940). Способность к высаливанию неионных детергентов имеет следующую тенденцию: SCN⊖ > I⊖ > NO
Способность понижать точку мутности имеет обратную тенденцию (Schott, H. et al., J. Pharm. Sci. 64:658-664 (1975)). Schott, H. et al., J. Pharm. Sci. 65: 979-981 (1976) фактически показали, что некоторые нитраты натрия и калия могут вызвать частичное совпадение точки мутности и температуры Краффта (Brij 56: оксид цетилполиоксиэтилена (C16E10), например), вызывая таким образом образование "аморфного геля". Возможно, поведение бетаинов в присутствии электролитов проявляет наиболее серьезное различие между этим классом детергентов и другими. Tsujii, K. et al., J. Phys. Chem. 82:1610-1614 (1978) показали, что C16- и C18-сульфопропилбетаины могут быть "всалены" при добавлении электролитов. Это означает, что точка Краффта существенно снижается при добавлении электролитов (например, точка плавления гидратированных кристаллов понижается). Tsujii, K. et al. , Yukagaku 30:495-499 (1981) позднее показали, что это явление было в прямой корреляции с длиной мостика: если длина мостика была более 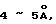 бетаин проявлял свойства по типу "всаливания", в то время, как при длине мостика короче
бетаин проявлял свойства по типу "всаливания", в то время, как при длине мостика короче 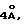 он проявлял свойства по типу "высаливания". Schott, H. et al., J. Pharm. Sci. 64:658-664 (1975) показал, что некоторые неионные детергенты могут быть всалены с учетом повышения точки мутности. Например, точка мутности Brij 96 (олеил полиоксиэтилен (C18:1E10): CAS® N.9004-98-2) в двухмолярном растворе HCl повышается при температуре 11,4oC. Однако, эти различия в температуре являются незначительными при сравнении с аналогичными величинами у бетаинов. Например, Tsujii, K. et al., J. Phys. Chem. 82:1610-1614 (1978) показали, что точка Краффта для SB-18 в чистой воде равна 73,4oC при концентрации детергента 2 mM и составляет величину 33,6oC в 1 М NaCl ( Δ 39,8oC). Tsujii, K. et al. , Yukagaku 30:495- 499 (1981) выдвигают гипотезу, что явление всаливания зависит от координации ионов по цвиттерионным головным группам и что существует минимальная перспектива, позволяющая использовать их продуктивно.
он проявлял свойства по типу "высаливания". Schott, H. et al., J. Pharm. Sci. 64:658-664 (1975) показал, что некоторые неионные детергенты могут быть всалены с учетом повышения точки мутности. Например, точка мутности Brij 96 (олеил полиоксиэтилен (C18:1E10): CAS® N.9004-98-2) в двухмолярном растворе HCl повышается при температуре 11,4oC. Однако, эти различия в температуре являются незначительными при сравнении с аналогичными величинами у бетаинов. Например, Tsujii, K. et al., J. Phys. Chem. 82:1610-1614 (1978) показали, что точка Краффта для SB-18 в чистой воде равна 73,4oC при концентрации детергента 2 mM и составляет величину 33,6oC в 1 М NaCl ( Δ 39,8oC). Tsujii, K. et al. , Yukagaku 30:495- 499 (1981) выдвигают гипотезу, что явление всаливания зависит от координации ионов по цвиттерионным головным группам и что существует минимальная перспектива, позволяющая использовать их продуктивно.
Степень, с которой бетаины высаливаются (для тех, которые относятся к типу высаливающихся), проявляется более разительно, чем способность к высаливанию у неионных детергентов, но в меньшей степени, чем наблюдается у гомологичных ионных детергентов (например, сравните данные Tsujii, K. et al., J. Phys. Chem. 82:1610-1614 (1978) с данными Tsujii, K. et al., Yukagaku 30: 495-499 (1981)). Способность бетаинов к всаливанию в присутствии различных солей придерживается классического "лиотропного ряда" (Tsujii, K. et al., J. Phys. Chem. 82: 1610-1614 (1978)). Следует сказать, что определенные ионы более эффективны при рассеивании точки Краффта по сравнению с другими ионами. Например, Tsujii, K. et al., J. Phys. Chem. 82:1610-1614 (1978) показали, что способность SB-18 к всаливанию следует тенденции: SCN⊖ > I⊖ > NO
Изменения мицеллярного веса и агрегатного числа в ответ на изменения концентрации электролитов может быть предсказано на основании способности к всаливанию и высаливанию. Мицеллярный вес и агрегатное число ионных детергентов возрастают по мере увеличения концентрации электролита (Shinoda, K., в: Colloidal Surfactants, Academic Press, New York (1963) pp.20-21, обзор многочисленных публикаций по этому вопросу). Becher, P., в: Surfactant Science Series, vol. 4: Nonionic Detergents, Marcel Dekker, New York (1967) pp. 500-504) всесторонне рассматривает воздействие электролитов на мицеллярный вес и агрегатное число неионных детергентов и приходит к выводу, что за исключением нескольких особых случаев, данные параметры по большей части не затрагиваются изменениями в концентрации электролитов. Однако, как известно, электролиты подавляют величину точки мутности неионных детергентов (Nakagawa, T. , et al. , Colloidal Surfactants, Academic press, New York (1963) pp. 129-135). Интересно отметить, что воздействие электролитов на детергенты обоих типов схоже, но противоположно: электролиты поднимают температуру Краффта у ионных детергентов (например, поднятие точки плавления кристаллов) и снижают точку мутности неионных детергентов (например, понижение температуры, при которой происходит фазовое разделение). Bhatia, A. et al., Colloids and Surfaces 69:277-292 (1993) показали, что мицеллярный вес и агрегатное число C18-карбоксиметилбетаина возрастает по мере увеличения концентрации Na⊕, однако, это происходит не так ярко выраженно, как у ионных детергентов.
Shinoda, K. Colloidal Surfactants, Academic Press, New York (1963) pp. 76-78 рассматривает ряд работ, показывающих, что как ионные, так и неионные детергенты, как правило, pH независимы. Следует сказать, что при равной ионной силе фазовое поведение этих детергентов равное при разных значениях pH (например, любое изменение, касающееся pH, может в целом быть отнесено к изменениям ионной силы, как обсуждалось выше).
Однако, свойства бетаинов чрезвычайно зависят от pH. Nandakumar T.N. et al., J. Oil Tech. Assoc. India 11:31-34 (1979) показывают, что несколько параметров, связанных с поверхностной активностью карбоксиметилбетаинов и сульфопропилбетаинов, зависят от pH. Tsubone et al. показали, что гидроксиалкилфосфобетаины (Tsubone et al., J. Am. Oil Chem. Soc. 67:394-399 (1990) и фосфобетаины (Tsubone, K. et al., J. Am. Oil Chem. Soc. 67:394-399 (1990)) также проявляют противоположные свойства при разных значениях pH. Однако следует понимать, что даже с теми детергентами, которые всаливаются, сочетание высокого значения pH (например, бетаины проявляются как ионные детергенты) и высокой концентрации соли приводит к высаливанию бетаина.
Tsubone, K. et al., J. Am. Oil Chem. Soc. 67: 149-153 (1990) показали, что пониженное значение pKa Cn-фосфоэтилбетаина попадает в диапазон 4,7-4,9, а повышенное - в диапазон 8,8 - 9,8. Weers, J.C. et al., Langmuir 7:854-867 (1991) показывают пониженные значения pK2 C12-карбоксибетаинов, имеющих разные длины мостиков. При возрастании длины мостика, пониженное значение pK2 возрастает. Таким образом, система pH является важным фактором, который надо учитывать при работе с бетаинами. Вкратце, зависимость от pH является проявлением комбинации зарядов, находящихся в бетаине, длины мостика и структуры. Использование бетаинов в методах по данному изобретению в жестко-щелочных условиях невозможно (например, бетаины выпадают в осадок при pH 14), хотя использование бетаинов в методах по данному изобретения в жестко-кислых условиях, не проявило таких же ограничений.
Существует еще один момент в отношении структуры мостика и его модификаций, связанный с использованием этих молекул. Parris, N. et al., J. Am. Oil Chem. Soc. 53: 60-63 (1976) сравнивают температуры Краффта сульфопропилбетаинов с соответственными гидроксипропилсульфобетаинами и показывают значительное повышение точки Краффта при этой модификации. Ветвление мостика с образованием "изо"-форм имеет даже более значительный эффект (Parris, N. et al. , J. Am. Oil Chem. Soc. 53:60- 63(1976)). Таким образом, выбор структуры мостика и системы электролита может влиять на использование этих молекул далее в методах по данному изобретению.
Уникальной чертой бетаинов является наличие дипольного момента, созданного структурным расположением противоположных зарядов (Laughlin, R. G. Langmuil 7: 842-847(1991). Полагают, что это расположение обеспечивает способность координировать структуру вода/ион так, что это приводит к нескольким привлекательным свойствам водной фазы. Упомянутое расположение может принимать различные формы. В табл. 3 приведены разнообразные структуры, явно бетаин-подобные, и в обсуждении табл. 3 перечисляет множество молекул этой категории. Оно включает "обратные бетаины", детергенты, из категории оксидоподобных аминов, c-алкильные бетаины и имидазольные производные бетаинов. Можно было бы предсказать, что, имея сильный дипольный момент, созданный бетаиноподобными структурами, свойственными этим молекулам, обратные бетаины, оксиды амина, c-алкильные бетаины и амидазольные производные бетаинов будут иметь похожую применимость в методах по данному изобретению. Свойства этих молекул обсуждаются ниже в связи с описанными здесь свойствами бетаинов.
Zimmer, R.E. (U.S. 3,560,393) описывает синтез большого числа разных обратных бетаинов, включающих фосфатные, фосфонатные, фосфинатные, аммонийные и сульфонийные функциональные группы. Kurosaki, T. et al., Chem.Soc. Japan 11:1297-1301 (1990) характеризуют обратные бетаины по типу: алкил 2-гидрокси-3- триметиламмонийпропил фосфаты (Cn-AHTMAP). Наблюдалось, что соединения Cn-AHTMAP имеют значения CMC, похожие на таковые соответствующих бетаинов, и B-значение 0,368, предполагая, что зависимость CMC от длины алкильной цепи ближе к таковой ионных детергентов, чем неионных детергентов. Следует, однако, повторить, что B-значения фосфобетаиновых серий по Tsubone et al. (Tsubone, K. et al., J. Am. Oil Chem. Soc. 67: 149-153 (1990) и Tsubone, K. et al., J. Am. Oil Chem. Soc. 67: 149-153 (1990)) кажутся даже более ионными по характеру, чем у соответствующих ионных детергентов. К сожалению, не существует данных относительно зависимости обратных бетаинов от pH.
Оксиды аминов (AO) очень похожи по структуре зарядов на бетаины; однако, мостик минимизирован до ковалентной связи N-О. Tsujii, K. et al., Yukagaki 30: 495-499 (1981), анализируя отношение межмостикового расстояния и способности к всаливанию/высаливанию, включили C18-AO в свое исследование. Эти авторы показали, что способность C18-AO аналогична способности C18- карбоксиметилбетаина; однако, точка Краффта повышена. Сравнение поведения при высаливании C18-AO (Tsujii, K. et al., Yukagaki 30:495-499 (1981)) с поведением при высаливании хлорида N,N,N- триметилоктадециламмония (Tsujii, K. et al., J.Phys.Chem. 82:1610- 1614 (1978)) показывает, что первый ведет себя более похоже на бетаины с коротким мостиком, чем на соответственный катионный детергент. Можно предполагать, что все важные характеристики дипольного момента бетаиноподобных структур функционируют хотя минимально в оксидах амина. Дальнейшая работа по оксидам аминов Fumikatsu, T. et al., J. Phys. Chem. 70:3437-3441 (1966) показала, что фазовое поведение детергентов этого класса зависит от pH, и Hermann, K.W. J. Colloid. Inter.Sci. 22:352-359 (1966) показал, что значения CMC, B-значения и мицеллярные свойства более близко свойственны бетаинами, чем соответствующим ионным детергентам. В материалах Hermann, K. W. J. Colloid.Inter.Sci. 22:352-359 (1966) даже предполагается, что оксиды аминов более неионные, чем соответствующие бетаины. McCutcheon's, Volume 1: Emulsifiers & Detergents, North American Edition, MC Publishing, Glen Rock, NJ, стр. 20, приводит оксиды аминов как отдельную категорию детергентов.
Большой объем информации относительно свойств c-алкилбетаинов представлен в серии статей Tori и соавторов (Tori, K. et al., Kolloid-Z Z. Polymere 187: 44-51 (1963); Tori, K. et al., Kolloid-Z Z. Polymere 188:47-52 (1963); Tori, K. et al., Kolloid-Z Z. Polymere 189:50-55 (1963); Tori, K. et al. , Kolloid-Z Z. Polymere 191:42- 48 (1963): Tori, K. et al., Kolloid-Z Z. Polymere 191: 48-52 (1963). Здесь опять видно, что фазовое поведение этих молекул похоже на n- алкилбетаины, обсуждавшиеся в табл. 2. Например, поведение при высаливании, как ожидается, имитирует таковое бетаинов с коротким мостиком, где высаливание не так значительно, как это видно для ионных гомологов. (Tori, K. et al., Kolloid-Z Z. Polymere 189:50- 55 (1963)). Дополнительно заметим, что CMC c-алкилбетаинов уменьшается с возрастанием температуры, что является отличительным признаком неионных детергентов. Далее, Molyneux, P. et al. , Trans. Faraday Soc. 61:1043-1052 (1965) изложили их данные по алкилбетаинам - зависимость CMC от длины алкильной цепи-, с аналогичными данными по n-алкилбетаинам Beckett, A.H., et al., J.Pharm.Pharmacol. 15: 422-431 (1963) в сравнении с данными по c-алкилбетаинам Tori, K. et al., Kolloid-Z Z. Polymere 191: 48-52 (1963), и показали, что B-значения n-алкилбетаинов и c- алкилбетаинов, как можно было ожидать, идентичны. Хотя популярность c-алкилбетаинов не достигает тех же размеров, как у n-алкилбетаинов, они коммерчески доступны: разнообразие, наблюдаемое в классе c-алкилбетаинов - это только часть класса n- алкилбетаинов.
Способность представить детергент в функциональной форме (т.е. в мицеллярной форме) в клиническом опыте (т.е. водных, содержащих электролиты в растворах) кажется важным аспектом функционирования бетаинов в методах изобретения. Таким образом, может оказаться, что выбор заряда (т.е. COO⊖ против SO
Очевидно следует заключение, что уникальной чертой бетаинов является то, что более длинные алкильные цепи могут быть использованы в клиническом опыте, так как имеют более привлекательные характеристики растворимости.
Бетаины как растворяющие агенты
Растворение - это действие по переведению в раствор молекул, которые нерастворимы или недостаточно растворимы в водных средах (Nakagawa, T. In: Nonionic Detergents, (1967) стр. 558-603). Теоретически, растворение происходит по одному из трех вариантов. Первый, растворяемое вещество может быть изолировано во внутренней части мицеллы (т.е. соединения неполярны). Второе, амфипатические молекулы, которые и полярны и неполярны до определенной степени, могут ассоциироваться с мицеллой таким же образом, как молекулы других детергентов (т.е. неполярная часть изолируется в ядре мицеллы, тогда как полярная часть ассоциирована с поверхностью мицеллы). Третий механизм действует для тех соединений, которые нерастворимы как в органических средах, так и в воде. Диметилфталат является классическим примером (McBain, J. W. et al., J.Am.Chem.Soc. 70:3838-3840 (1948). В этой модели предполагается, что молекулы адсорбируются на поверхности мицеллы. Таким образом, интуитивно предполагается, что поведение при растворении основывается на природе как детергента, так и растворяемого вещества.
McBain, J. W. et al., J. Phys. Chem. 55:655-662 (1951) сравнивали растворение n-гексана, циклогексана, циклогексена и бензола лауратом калия, гидрохлоридом додециламина и Triton X-100 и заключили, что ионные детергенты сильнее всего растворяют циклогексан, тогда как Triton X-100 сильнее всего растворяет бензол. Ионные и неионные детергенты, как ожидалось, вели себя по-разному при анализе на функциональном уровне.
Weers, J.C. et al., Langmuir 7:854-867 (1991) предоставили единственное исследование контролируемого растворения, сравнивающее анионный (SDS: CAS® N. 151-21-3), катионный (бромид C12-триметиламмония: CAS® N. 1119-94-4) и неионный детергент (C12-этиленоксид (C12E6)): CAS® N. 3055-96-7) с C12-карбоксиметилбетаином (CAS® N. 683-10-3) и C12сульфопропилбетаином (CAS® N. 14933-08-5). "Oil Blue A" был использован в качестве растворителя. Эти авторы показали, что способность к растворению возрастает в порядке: анионный <катионный<карбоксибетаин<сульфобетаин<неионный. На основании вышеприведенного обсуждения работы McBain, J.W. et al., J.Phys.Chem. 55:655-662 (1951) и того, что Oil Blue A является расплавленным полициклическим углеводородом (производным антрацена: 1,4-бис((1-метилэтил)амино)-9,10-антраценодионом (CAS® N. 14233-37-5), можно было ожидать такой иерархии. Интересно, что сульфобетаин растворяет больше Oil Blue A, чем карбоксибетаин. Это согласуется с вышеприведенным обсуждением, описывающим поведение карбоксибетаинов относительно сульфобетаинов: карбоксибетаины тяготеют к ионному поведению при сравнении с сульфобетаинами; или, с другой стороны, сульфобетаины тяготеют к неионным детергентам при сравнении с карбоксибетаинами.
Длина алкильной цепи непосредственно соотносится со способностью к растворению. C18-детергенты будут иметь великолепные растворяющие характеристики относительно C12-детергентов, даже C12-детергентов в других классах: Weers, J.C. et al., Langmuir 7:854-867 (1991) также показали, что "максимальная концентрация добавок" возрастает экспоненциально с возрастанием длины алкильной цепи. Например, если C12-этиленоксид (C12E6) растворяет немного больше Oil Blue A, чем C12-сульфопропилбетаин, C14-сульфопропилбетаин растворяет в 4 - 5 раз больше Oil Blue A, чем C12-этиленоксид (C12E6)) (Weers, J.C. et al., Langmuir 7:854-867 (1991).
Неотъемлема в рамках изучения феномена растворения концепция детергентности. Оптимальная детергентность требует сложной смеси поверхностно-активных свойств (Harris, J.C. In: Surfactant Science Series, vol. 1: Nonionic Detergents, (1964), стр. 683-732 рассматривают эти свойства). Решающим аспектом детергентности является представление о том, что если что-то растворено, оно должно поддерживаться в дисперсной форме.
Бетаины имеют прекрасные дисперсионные характеристики (Weit, J.K. et al. , J. Am. Oil Chem. Soc. 53:757-761 (1976)). Fernley, G.W. et al., J.Am.Oil Chem. Soc. 55: 98-103 (1978) рассмотрели использование бетаинов как детерзивных агентов и сравнили их с алкил-сульфонатными детергентами. Однако, растворяемые вещества в методах изобретения являются липидами и липопротеинами, ассоциированными с клеточной стенкой микобактерий.
Структура клеточной стенки микобактерий (см. McNeil, M.R. et al., Res. Microbiol. 142:451-463 (1991) для всестороннего обзора), состоящая в первую очередь из липидов, липопротеинов и миколовых кислот, имеет большую толщину и гидрофобна. Что касается миколовой кислоты, каждая миколовая кислота имеет несколько цепей приблизительно C14-20 (Σ = C76-C80) и единица клеточной стенки выделена как димиколат (Noll, H. et al., Biochim. Biophys. Acta 20: 299-309 (1956)). Обусловленный структурой миколовых кислот второй метод солюбилизации, как можно ожидать, будет действенным (т.е. неполярная часть отделена в ядре мицеллы, тогда как полярная часть ассоциирована с поверхностью мицеллы). Основываясь на вышеприведенной дискуссии, можно предсказать, что ионные детергенты будут лучшими солюбилизаторами миколовых кислот, чем неионные детергенты. Таким образом, периферически ассоциированные компоненты клеточной стенки должны быть очищены и поддерживаться в растворе для того, чтобы диспергировать М. tuberculosis. Young, D.B. et al., Res. Microbiol. 142: 55-65 (1991) показал, что Triton X- 100 (CAS® N. 9002-93-1) может удалять липопротеины из М. tuberculosis, однако фиг.3 предполагает, что этот детергент не может диспергировать бактерии. Таким образом, можно ожидать, что длинноцепочечные бетаины будут лучшими солюбилизаторами миколовых кислот, чем Triton Х-100. Обусловленное огромным размером и сложной структурой миколовых кислот поддержание их в растворе, как можно ожидать, будет проблематичным. Другими словами, SB-12, в дополнение к тому, что он относительно слабый солюбилизатор, сравнительно с SB-18, может оказаться неспособным поддерживать миколовые кислоты в дисперсном состоянии. Например, Bhatia, A. et al. , Colloids and Surfaces 69:277-292 (1993) сравнивали в деталях мицеллярную структуру C12, C16 и C18-карбоксиметилбетаинов. Гидродинамический радиус мицеллы C12-бетаина 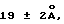 тогда как структура мицеллы обоих C16 и C18-бетаинов палочкообразная и зависит от концентрации поверхностно-активного вещества и электролита: устойчивая длина мицеллы C16-бетаина
тогда как структура мицеллы обоих C16 и C18-бетаинов палочкообразная и зависит от концентрации поверхностно-активного вещества и электролита: устойчивая длина мицеллы C16-бетаина 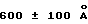 в 1,0 М NaCl при 25oC, тогда как постоянная длина мицеллы C18-бетаина
в 1,0 М NaCl при 25oC, тогда как постоянная длина мицеллы C18-бетаина 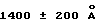 в 0,01 М NaCl при 43oC. Учитывая, что протяженность цепи C12-бетаина
в 0,01 М NaCl при 43oC. Учитывая, что протяженность цепи C12-бетаина 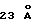 (Bhatia, A. et al., Colloids and Surfaces 69:277-292 (1993) и далее учитывая, что природа миколовых кислот гидрофобная, вероятность, что более короткая цепь поверхностно-активных веществ просто не сможет поддерживать солюбилизированную миколовую кислоту в растворе, может объяснить неспособность более коротких детергентов SB-серий диспергировать М. tuberculosis.
(Bhatia, A. et al., Colloids and Surfaces 69:277-292 (1993) и далее учитывая, что природа миколовых кислот гидрофобная, вероятность, что более короткая цепь поверхностно-активных веществ просто не сможет поддерживать солюбилизированную миколовую кислоту в растворе, может объяснить неспособность более коротких детергентов SB-серий диспергировать М. tuberculosis.
Краткая характеристика бетаинов, как детергентов
Бетаины имеют уникальные характеристики, которые помещают их в отдельный класс, но этот класс, кажется, находится где-то между ионными и неионными категориями. Поведение их очень сложное и зависит от структуры мостика и типа цвиттериона. В некоторых случаях бетаины ведут себя как ионные детергенты, и в других, бетаины ведут себя как неионные детергенты. Таким образом, выбор аниона и длины мостиковой группы имеет глубокое влияние на их использование.
Например, все характеристики растворимости непосредственно связаны с гидрофильностью головной группы. Гидрофильность головной группы в свою очередь зависит от аниона. Так как гидрофильность различных анионов сопровождается прогрессией SO
действительно уменьшается с увеличением температуры; что сродни ионному поведению. Однако, в отношении алкильной структуры все детергенты, как и ожидалось, ведут себя одинаково. Например, включение структурных изменений в алкильную цепь, будь то одна или несколько двойных связей, полярных групп, таких как гидроксилы, простые эфиры, амиды или сложные эфиры, или другие модификации, такие как включение циклопропановых колец, могут производить умеренные изменения в характере. Обусловленный широкой вариабельностью структуры, в сочетании с известными физическими характеристиками бетаино-подобных структур, подбор бетаина с идеальными характеристиками для использования в методах данного изобретения становится возможным. Например, карбоксибетаины, несмотря на то, что обладают негативным качеством - более высокой степенью зависимости от pH, дают более растворимые бетаины и могут, таким образом, позволить использование более длинных алкильных цепей. Дополнительно, карбоксибетаины, с тенденцией к ионному поведению, как можно ожидать, будут лучшими солюбилизаторами миколовых кислот среди бетаинов. Мостик должен быть длиннее, чем 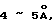 следовательно, пропильная группа будет минимальной. Hodge, D.J. et al, Langmuir 7:878-884 (1991) докладывал о C20-карбоксигексилбетаине с точкой Краффта 20oC. Ожидается, что C18-карбоксипропилбетаин (CAS® N. 78195-27-4) обладает оптимальными характеристиками.
следовательно, пропильная группа будет минимальной. Hodge, D.J. et al, Langmuir 7:878-884 (1991) докладывал о C20-карбоксигексилбетаине с точкой Краффта 20oC. Ожидается, что C18-карбоксипропилбетаин (CAS® N. 78195-27-4) обладает оптимальными характеристиками.
Суммируя, можно сказать, что длинноцепочечные алкилы требуются для дисперсии М. tuberculosis, и эти молекулы должны функционировать в контексте физиологически уместных условий (т.е. концентрация электролита приблизительно 100-150 мМ). Здесь лежит видимое преимущество бетаинов над ионными детергентами в методах по данному изобретению: бетаины всаливаются, а ионные детергенты высаливаются. Объяснение преимущества бетаинов перед неионными детергентами более спекулятивное, предположение в том, что бетаины в основном лучше солюбилизируют миколовые кислоты и другие липиды, вовлеченные в образование тяжей, чем неионные детергенты.
Хотя эти соображения действительно представляют объяснение, почему SB-18 был единственным детергентом, функционирующим в опыте по агрегированию, они не объясняют полностью бимодальное действие SB-18: а именно, почему SB-18 также улучшает сбор клеток М. avium (для которых агрегирование не является проблемой). Это обсуждается ниже.
Плавучесть микобактерий
Синтез липидов микобактериями - очень сложный и разнообразный предмет, со многими особенностями, которые видоспецифичны (по этому вопросу см. Bloch, K Adv. Enzymol. 45: 1-84 (1977): Minnikin, D.E. in: The Biology of Mycobacteria, C. Ratledge et al. eds., vol. 1 Academic Press, New York, 1982, стр. 95-184: Ratledge C. in: The Biology of Mycobacteria, C. Ratledge et al. eds., vol. 1 Academic Press, New York, 1982, стр. 53-92: Takayama, K. et al., in: The Mycobacteria a source book, part A, G.P. Kubika et al., eds. , Marcel Dekker, Inc. , New York, 1984, стр. 315-344). В целом клеточная стенка состоит на 60% из липидов, из которых приблизительно 50% находится в форме миколовых кислот. (Joklik, W.K. et al., Zinsser Microbiology, 20th edition, Appelton & Lange, Norwalk, CT, 1992, стр. 81 и 499).
Миколовая кислота придает уникальный характер этому классу микроорганизмов (Ratledge C. in: The Biology of Mycobacteria, С. Ratledge et al. eds. , vol. 1 Academic Press, New York, 1982, стр. 53-92 и особенно страницы 65-84 для детального описания синтеза миколовой кислоты).
Результат синтеза миколовой кислоты in vivo состоит в том, что образуются значительные количества CO2. Например, синтез остатка миколовой кислоты, содержащий 80 атомов углерода по пути малонил CoA, дает приблизительно 80 молекул CO2.
Существует несколько важных моментов. Во-первых, in vivo, микобактериальная клеточная стенка толще, чем ее копия in vitro (Rastogi, N. et al., Antimicrob. Agents Chemother. 20% 666-677 (1981)). Во-вторых, почти 18% сухого веса микобактериальной клетки (выросшей в отсутствие олеиновой кислоты, Tween 80 или BSA) составляет липид (Etinson, M.W., et al., Am. Rev. Resp. Dis. 104: 717-727 (1971)) и in vivo приблизительно 60% клеточной стенки - это комплексный липид и 50% от него - миколовая кислота (Joklik, W.K. et al. , Zinsser Microbiology, 20th edition, Appelton & Lange, Norwalk, CT, 1992, стр. 81, 499 и 503); следовательно микобактерии выделяют большое количество CO2 во время синтеза клеточной стенки. По существу, непосредственным результатом роста в обогащенных средах, содержащих олеиновую кислоту, Tween 80 и BSA, может быть снижение синтеза липидов и, следовательно, значительное уменьшение образования CO2. В итоге, является установленным, что необработанные клетки обладают плавучестью в поле центрифугирования, что означает, что любая теория плавучести, содержащая положение о включениях газа, уничтожает классические представления о "газовых вакуолях" как возможной причине (Walsby, A.E. Bact.Rev. 36:1-32 (1972). Это приводит исследователя к заключению, что микроорганизмы могут просто быть дегазированы для уменьшения или уничтожения плавучести. Например, как описано ниже, небольшое нагревание микобактерий (точка плавления фактора, способствующего агрегированию клеток - melting point (MP) - 43-45oC (Noll, H. et al., Biochim. Biophys. Acta 20: 299-309 (1956)), под вакуумом, делает возможным удаление достаточного количества задержанного CO2 для того, чтобы повлиять на плавучесть.
Обработка бактерий комплекса MAC
Для некоторых микроорганизмов, таких как бактерии MAC комплекса, необходима дальнейшая стадия обработки, инкубирование в течение достаточного отрезка времени при повышенной температуре, такой как 45oC, в вакууме под давлением приблизительно 600 мм рт.ст. Нет смысла говорить о минимальном времени экспозиции образцов, содержащих MAC микроорганизмы, в промывном растворе, содержащем детергент, с целью дисперсии, так как эффект наступает почти немедленно, но как правило желательно проводить промывку в течение 60 мин для того, чтобы позволить детергенту аккумулироваться в клетках так, чтобы компенсировать плавучесть. Полагают, что экспозиция образца в промывном растворе больше 60 мин дает ограниченное преимущество. Время для такой стадии вакуумного дегазирования должно быть по меньшей мере 20 мин и 60 мин предпочтительно. Инкубация до нескольких часов приводит только к незначительным улучшениям по сравнению с инкубацией на протяжении 60 мин, если используются SB-18-подобные детергенты. Однако, инкубация в несколько часов позволяет использовать большинство детергентов. Экспозиция MAC микроорганизмов в содержащем детергенты промывном растворе может быть одновременной со стадией дегазирования, однако могут быть практические соображения по безопасности при использовании этого воплощения. Температура стадии дегазирования должна быть между 30oC и 50oC, предпочтительно 40-42oC. Температура ниже 30oC, возможно, не усилит смягчение восковой оболочки MAC микроорганизмов, температура выше 50oC может усилить лизис организма. Так как жизнеспособность есть первое преимущество описанных здесь методов, и далее, так как методы дают преимущество в отношении улучшения эффективности выделения центрифугированием только в том случае, если бактериальные клетки интактны, температура при обработке не должна превышать 50oC для большинства описанных здесь микроорганизмов, но температура выше 50oC может быть использована в том случае, если она не вызывает лизиса некоторых специфических определяемых видов. Это может быть, однако, использовано для получения желаемого результата. Например, температуру можно выбрать так, чтобы лизировать известный определенный микроорганизм при определенных условиях обработки, но не лизировать другой вид микроорганизма в тех же самых определенных условиях обработки. В таком примере методы изобретения можно селективно обогащать для нелизирующихся микроорганизмов.
Не желая вдаваться в последующие объяснения, предполагается, что различия, наблюдаемые между микроорганизмами MTB-комплекса и МАС-комплекса, происходят из-за их относительных характеристик роста. В организмах MAC комплекса, которые растут преимущественно отдельными клетками, SB-18-подобные детергенты имеют три эффекта: (1) эффект нарушения основного поверхностного натяжения, наблюдаемый с большинством детергентов, (2) эффект дисперсии, наблюдаемый с организмами MTB комплекса (хотя и менее обязывающий) и (3) способность транспортироваться через мембрану и аккумулироваться в клетке. Weir, M.P. et al., Am. Rev. Res. Dis. 106:450-457 (1972) показали, что микобактерии очень эффективны в активном аккумулировании липидов, например, олеиновой кислоты. Schaefer, W.B. et al., J.Bacteriol. 90:1438-1447 (1965) показали, что аккумуляция липидов наблюдается в течение двух часов и ярко выраженные липоидные включения формируются в течение 24 часов. Последовательность аккумуляции липидов, в противоположность высказывавшимся ранее предположениям (Silverstolpe, L. Nord. Med. 40/48: 2220-2222 (1948); Davis, В. D. et al. Microbiology, Harper & Row, New York (1973), стр.847), может значительно изменять частичный специфический объем клетки таким образом, что это оказывается достаточным для частичного противодействия естественной плавучести.
Не желая вдаваться в последующие объяснения, предполагается, что стадия вакуумного дегазирования эффективно удаляет задержанный газ, а именно CO2, образованный в результате естественного метаболизма микроорганизма, но задержанный до некоторой степени в его наружней воскообразной клеточной стенке. Последняя теория основана на наблюдениях, что ограниченные, но различимые улучшения в выделении наблюдались в отсутствие добавленного детергента, и что любой детергент может быть использован, если микроорганизмы значительно дегазированы. Преодоление плавучести путем сочетания детергентов с дегазированием было рассмотрено для дальнейшего улучшения концентрирования клеток методами, связанными с силой тяжести, например, центрифугированием, как здесь обсуждалось.
Факт, что детергенты, иные чем SB-18-подобные, палочковидные или октадецил-подобные, могут быть использованы, когда микроорганизмы значительно дегазированы, предполагает, что естественная плавучесть организмов MAC комплекса скорее всего происходит от задержанных газов. Не задерживаясь на этом объяснении, предполагается, что раз естественная плавучесть организмов MAC комплекса полностью уничтожена, поверхностное натяжение остается единственным фактором, способствующим флотации. Так как большинство детергентов снимают поверхностное натяжение, много дополнительных детергентов можно использовать для улучшения выделения центрифугированием. Длительная инкубация под вакуумом может, однако, снизить жизнеспособность микроорганизмов.
Вакуумное давление в 600 мм рт.ст. удобно благодаря коммерческой доступности экономичных вакуумных насосов этого уровня. Однако, специалисты могут использовать любое давление или средства, которые достаточны для достижения желаемого результата по определению чувствительности микроорганизмов к детергентам, содержащим октадецил-подобные компоненты. После стадии промывки детергентом, таким как SB-18, и после, если необходимо, стадии дегазирования, микроорганизмы в образце собираются методами, известными в этой области, как например, центрифугирование, и лизируются, если нужно, для дальнейшего анализа, такого как определение нуклеиновых кислот с помощью амплификации. Не все методы определения требуют лизиса микроорганизмов. Например, определение методом высева или определение мембранных антигенов в иммунном методе не требует лизиса. Поскольку методы по данному изобретению не вызывают лизиса, микроорганизмы, имеющие структуры миколовых кислот в наружной мембране, и, особенно, микобактерии могут быть определены любой методикой, такой как амплификация или высев после обработки любым из детергентов, предлагаемых в данном изобретении.
Аккумулирование детергентов
Не вдаваясь подробно в последующие объяснения, аккумуляция детергентов объясняет один аспект SB-18-подобной активности.
Для проверки обоснования гипотезы дегазирования установлено, что культивирование микобактерий в присутствии Tween 80 (CAS® N. 9005-65-6)/олеиновой кислоты (CAS® N. 112-80-1) вызывает погруженный рост (Dubos, R.J., Exp. Biol. Med 58:361-362 (1945) и рост микобактерий в присутствии Tween 80/олеиновой кислоты вызывает образование липоидных включений (Schaefer, W. B. et al., J.Bacteriol. 90:1438-1447 (1965)), можно заключить, что, в противоположность имевшимся прежде мнениям (Silverstolpe, L. Nord. Med. 40/48: 2220-2222 (1948); Davis, B.D. et al. Microbiology, Harper & Row, New York (1973), стр.847), образование липоидных включений связано с погруженным ростом и, возможно, даже вызывает его. Также следует сделать заключение, что объяснение источника естественной плавучести этих микроорганизмов не было до сих пор осознано. Полагают, что бетаины, аналогично Tween 80/олеиновой кислоте, проникают в клетку таким образом, что они изолируются внутри клетки, и делая это, изменяют частичный специфический объем клетки, частично уменьшая естественную плавучесть этих организмов, таким образом повышая выделение путем центрифугирования. Факт, что бетаины долго использовались как компоненты коммерческих антибактериальных препаратов в составах детергентов
(Gomi, T. , JP 8895298 A2: JP 6395298), антикоррозионных препаратов (Nomcova, J. et al., CS 202494 B), средств для чистки зубов. (Oshino, K. et al., JP 92134025 A2; JP 04134025) предполагают, что они действительно проникают в микробные клетки. Факт, что клеточная стенка Mycobacterium chelonei 1000 до 10000 раз менее проницаема, чем у E.coli (Jarlier, V. et al., J.Bacteriol. 172:1418-1423 (1990), предполагает, что какой-то аспект природы бетаина вызывает то, что эти соединения изолируются предпочтительным образом.
Для того, чтобы молекула проникла в микробную клетку, она должна сначала пройти через клеточную стенку (наружная мембрана) и затем через клеточную мембрану (внутренняя мембрана). Проницаемость клеточных стенок обсуждалась Nikaido, H. et al., Microbiol.Rev. 49:1-32 (1985) и проницаемость микобактериальных клеток обсуждалась Connell, N.D. et al., In: Tuberculosis: Pathogenesis, Protection and Control, B.R. Bloom, ed., American Society for Microbiology, Washington, D. C. (1994) стр. 333-352. В основном для молекулы существует три пути преодоления наружной мембраны. Первый - "гидрофильный путь", обычно вовлекающий порин; второй - "гидрофобный путь", обычно включающий диффузию через гидрофобную область; и третий обозначается как "самостимулирующийся путь", или разрушение клеточной стенки агентом для обеспечения входа самой молекулы (суммировано из Hancock, R.E.W. Ann.Rev.Microbiol. 38:237-264 (1984)). Транспорт через клеточную стенку, обусловленный природой носителя, является самым обычным средством преодоления внутренней мембраны. Существует два основных класса транспорта, обусловленного носителем: "облегченная диффузия" и "активный транспорт". Оба включают использование специфичных белковых комплексов, имеющих определенный уровень субстратной специфичности, характеристики Михаэлиса-Ментена. Некоторые являются энергетически зависимыми, тогда как другие зависят от электрохимического градиента. Таким образом, клеточная стенка микобактерий представляет собой выдающийся пример барьера, который должен обеспечивать некоторый механизм обмена молекулами с внеклеточной средой.
В контексте методов по данному изобретению очевидно, что нужно иметь в виду обсуждаемые временные рамки. Например, большинство исследований, которые изучали бактерицидную или бактериостатическую активность определенного соединения, выращивали бактерии в присутствии указанной молекулы в течение продолжительного периода времени (например, дней). Через этот период времени отмечался рост и оценивалась эффективность. В методах данного изобретения обсуждаемые временные рамки ограничены несколькими часами. Таким образом, маловероятно, что такие механизмы, как гидрофобный путь или самостимулирующийся путь, были вовлечены.
Таким образом, поступление бетаинов через наружную мембрану должно использовать гидрофильный путь (т.е. порины) в первую очередь. Порины - "... белки, которые образуют неспецифические, открытые, наполненные водой каналы, позволяющие небольшим молекулам диффундировать через мембрану." (цитировано по: Connell, N.D. et al., In: Tuberculosis: Pathogenesis, Protection and Control, B.R. Bloom, ed., American Society for Microbiology, Washington, D.C. (1994) стр. 336. Trias, J.V. et al. Science 258:1479-1481 (1992) докладывали, что порин М. c...nae является 59 KD-белком, который образует пору с диаметром приблизительно 2,2 нм, и пределом исключения между 2000 и 3000 дальтон. Молекулярная масса SB-18 (CAS® N. 13177-41-8) 419,7 дальтон. Дополнительно, Tsubone, K., J. Am. Oil Chem.Soc. 67:149-153 (1990) и Tsubone, K., J. Am. Oil Chem.Soc. 67:394-399 (1990) изучали физические параметры нескольких N-алкил-фосфоэтилбетаинов и соответствующих 2- гидроксиалкил-фосфоэтилбетаинов, и показали, что площадь, занимаемая каждой молекулой для C16-бетаинов, составляет 0,475 нм2 и 0,761 нм2, соответственно. Таким образом, диаметр поперечного сечения головной группы бетаина приблизительно 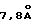 и
и 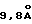 , соответственно. Bhatia, A.et al., Colloids Surf. 69:277-292 (1993) принимают объем головной группы C18-карбоксибетаина (CAS® N. 820-66-6) как
, соответственно. Bhatia, A.et al., Colloids Surf. 69:277-292 (1993) принимают объем головной группы C18-карбоксибетаина (CAS® N. 820-66-6) как 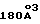 , что дает диаметр в
, что дает диаметр в 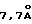 для этой головной группы. Для сравнения, диаметр головной группы SDS (CAS® N. 151-21-3) - порядка
для этой головной группы. Для сравнения, диаметр головной группы SDS (CAS® N. 151-21-3) - порядка 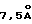 (Shinoda, K.J. , J. Phys.Chem. 59:432-435 (1955). Неионные детергенты, Tween 80 (Олеил полиоксиэтилен (n= 20) сорбитан: CAS® N. 9005-65-6), например, должны иметь значительно большие диаметры головной группы, это может быть пространственной помехой легкому проникновению через порин. В поддержку этого, Wayne, LG. et al., Am. Rev. Resp. Dis. 90:588-597 (1964) показали, что только некоторые микобактерии способны гидролизовать Tween, и рост тех, которые не способны гидролизовать Tween, не стимулируется добавлением этого детергента. Предполагают, что головная группа должна быть удалена перед ассимиляцией.
(Shinoda, K.J. , J. Phys.Chem. 59:432-435 (1955). Неионные детергенты, Tween 80 (Олеил полиоксиэтилен (n= 20) сорбитан: CAS® N. 9005-65-6), например, должны иметь значительно большие диаметры головной группы, это может быть пространственной помехой легкому проникновению через порин. В поддержку этого, Wayne, LG. et al., Am. Rev. Resp. Dis. 90:588-597 (1964) показали, что только некоторые микобактерии способны гидролизовать Tween, и рост тех, которые не способны гидролизовать Tween, не стимулируется добавлением этого детергента. Предполагают, что головная группа должна быть удалена перед ассимиляцией.
В данной работе показано использование некоторых неионных детергентов для улучшения выделения. Например, если головная группа Tween 80 слишком велика для того, чтобы пройти через порин, другие детергенты, такие как линейные полиоксиэтиленовые эфиры жирных кислот (напр. "Brij"-соединения), применяющиеся в методах по данному изобретению, как полагают, способны проникать в клетки и это позволяет им функционировать в методах данного изобретения. Hsiao, L. et al., J.Phys.Chem. 60:657-660 (1956) показал, что пространство на каждую молекулу полиоксиэтиленфениловых эфиров того же порядка, как и для бетаинов (то есть C9-, с, в среднем, 9,5 единицами POE, будет иметь и площадь 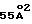 ). Здесь показано, что Brij 96 (олеилполиоксиэтиленовый эфир (C18:1E10): (CAS® N. 9004-98-2), который относительно неэффективен при диспергировании групп клеток М. tuberculosis, имеет аналогичную SB-18 активность в отношении преодоления плавучести. Не намереваясь задерживаться на следующем объяснении, отметим, что неионные детергенты, такие как Brij 96, функционируют в методах данного изобретения благодаря их гомологичной химической форме. То есть, Brij 96 похож на детергенты, подобные SB-18, в том, что у него такой же "палочкообразный" характер и у него нет стерических препятствий для проникновения в клетку. Таким образом, "палочкообразность", как она здесь называется, относится к неионным детергентам, проявляющим подобное SB-18 действие в отсутствие дегазирования, в методах по данному изобретению. Палочкообразные детергенты будут иметь похожие "осевые пропорции". Осевая пропорция, как определено McGraw-Hill Dictionary of Scientific and Technical Terms, 5th ed., Parker, S. P. ed. McGraw-Hill, Washington, D,C, (1994) стр. 168 - это "отношение большей оси к меньшей оси...". Например, Bhatia, A.et al., Colloids Surf. 69: 277-292 (1993) принимает протяженность длины цепи (т. е. большую ось) C12-карбоксилметилбетаина как
). Здесь показано, что Brij 96 (олеилполиоксиэтиленовый эфир (C18:1E10): (CAS® N. 9004-98-2), который относительно неэффективен при диспергировании групп клеток М. tuberculosis, имеет аналогичную SB-18 активность в отношении преодоления плавучести. Не намереваясь задерживаться на следующем объяснении, отметим, что неионные детергенты, такие как Brij 96, функционируют в методах данного изобретения благодаря их гомологичной химической форме. То есть, Brij 96 похож на детергенты, подобные SB-18, в том, что у него такой же "палочкообразный" характер и у него нет стерических препятствий для проникновения в клетку. Таким образом, "палочкообразность", как она здесь называется, относится к неионным детергентам, проявляющим подобное SB-18 действие в отсутствие дегазирования, в методах по данному изобретению. Палочкообразные детергенты будут иметь похожие "осевые пропорции". Осевая пропорция, как определено McGraw-Hill Dictionary of Scientific and Technical Terms, 5th ed., Parker, S. P. ed. McGraw-Hill, Washington, D,C, (1994) стр. 168 - это "отношение большей оси к меньшей оси...". Например, Bhatia, A.et al., Colloids Surf. 69: 277-292 (1993) принимает протяженность длины цепи (т. е. большую ось) C12-карбоксилметилбетаина как 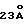 , и объем головной группы карбоксилметилбетаина как приблизительного
, и объем головной группы карбоксилметилбетаина как приблизительного 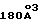 . Считая головную группу сферой, диаметр головной группы (т.е. меньшая ось) будет приблизительно
. Считая головную группу сферой, диаметр головной группы (т.е. меньшая ось) будет приблизительно 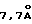 . Большая ось, по определению, будет перпендикулярна меньшей оси. Таким образом, осевое соотношение C12-карбоксилметилбетаина (CAS® N. 683-10-3) будет приблизительно 3 (23 : 7,7=2,99). Tsubone, K., J. Am. Oil Chem. Soc. 67:394-399 (1990) докладывал, что для 2-гидроксиалкилфосфоэтилбетаинов площадь, занятая головной группой - 0,761 нм2. Меньшая ось этих бетаинов будет
. Большая ось, по определению, будет перпендикулярна меньшей оси. Таким образом, осевое соотношение C12-карбоксилметилбетаина (CAS® N. 683-10-3) будет приблизительно 3 (23 : 7,7=2,99). Tsubone, K., J. Am. Oil Chem. Soc. 67:394-399 (1990) докладывал, что для 2-гидроксиалкилфосфоэтилбетаинов площадь, занятая головной группой - 0,761 нм2. Меньшая ось этих бетаинов будет 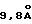 , давая значение осевого соотношения для C12-бетаина (CAS® N. 124591-53-3) в 2,3. Таким образом, можно ожидать, что осевое соотношение бетаинов находится в диапазоне от 1,5 для детергентов, у которых большая ось (R1) состоит из 8 атомов углерода, до приблизительно 6 для тех бетаинов, у которых гидрофобная область достигает 22 атомов углерода. Палочкообразные детергенты будут включать в себя все электрически нейтральные детергенты с длиной алкильной цепи 8-22 атомов углерода, предпочтительнее 12-20 атомов углерода и, наиболее предпочтительно, - 16-18 атомов углерода, что обнаруживает активность, аналогичную SB-18 в отсутствие дегазирования.
, давая значение осевого соотношения для C12-бетаина (CAS® N. 124591-53-3) в 2,3. Таким образом, можно ожидать, что осевое соотношение бетаинов находится в диапазоне от 1,5 для детергентов, у которых большая ось (R1) состоит из 8 атомов углерода, до приблизительно 6 для тех бетаинов, у которых гидрофобная область достигает 22 атомов углерода. Палочкообразные детергенты будут включать в себя все электрически нейтральные детергенты с длиной алкильной цепи 8-22 атомов углерода, предпочтительнее 12-20 атомов углерода и, наиболее предпочтительно, - 16-18 атомов углерода, что обнаруживает активность, аналогичную SB-18 в отсутствие дегазирования.
Октадецилоподобные детергенты
Было открыто, что определенные детергенты, неэффективные в диспергировании скоплений MTB, могут далее улучшать концентрирование MAC бактерий центрифугированием, если организмы дегазированы. Например, после обсуждавшейся выше вакуумной обработки может быть использовано много дополнительных детергентов, особенно те, которые подобны октадецилам по структуре. То есть, SB-16 и SB-18 становятся более эффективными в улучшении сбора клеток после дегазирования. Например, бромид триметилоктадецил-аммония (TMA-18: (CAS® N. 1120-02-1) и хлорид бензилдиметилоктадециламмония (BenzDMA-18), как теперь стало очевидно, имеют некоторый уровень эффективности в улучшении сбора клеток методами данного изобретения после того, как выполнено дегазирование. Неионные, октадециловые детергенты, такие как Tween 80 (CAS® N. 9005-65-6) и Span 80 (CAS® N. 1338-43-8) представляли тот же самый феномен после дегазирования. Когда задержанный газ удален, октадецил-подобные молекулы улучшают сбор путем изменения частичного специфичного объема клетки, как обсуждалось выше.
Таким образом, "октадецилоподобное" вещество, как это здесь называется, представляет собой молекулу детергента, обладающую частью, похожей на октадецил, аналогичную SB-18-подобной октадециловой части и не требующую присутствия функции цвиттериона для того, чтобы быть эффективной. Октадецилоподобные детергенты полезны в методах по данному изобретению, когда применяются к MAC комплексу микроорганизмов, и включают SB-18-подобные детергенты. Не все октадецил-подобные детергенты являются SB-18-подобными детергентами, но все SB-18-подобные детергенты являются октадецил- подобными детергентами. SB-18-подобные детергенты полезны в методах по данному изобретению, когда применяются к комплексу MTB (т.е., где дегазирование не требуется).
Неспособность анионных детергентов (напр. додецил сульфата натрия (SDS): CAS® N. 151-21-3) или октадецил сульфата натрия (SOS): CAS® N. 1120-04-3) или катионных солей алкилтриметиламмония (напр. TMA-12 (CAS® N. 1119-94-4) и TMA-18 (CAS® N. 120-02-1) работать таким же образом, как SB-18, объясняется следующим образом. Точка Краффта у длинноцепочечных ионных детергентов такова, что TMA-18 и SOS находятся в микрокристаллической форме и, таким образом недоступны для быстрого поглощения. Это может быть подтверждено проверкой методами Weir, M.P. et al., Amer. Rev. Res. Dis. 106:450-457 (1972): BSA был использован для эмульгирования олеиновой кислоты (CAS® N. 112-80-1) перед использованием. Эмульгирование позволяет представить олеиновую кислоту в форме, приемлемой для поглощения (т.е. в растворе). Объяснение исключения анионных детергентов, находящихся в растворе, состоит в том, что микобактериальные порины катион-селективны. Иными словами, канал имеет несколько пространственно расположенных анионных аминокислот у начала канала порина, что исключает анионные соединения (Trias, J.V. et al., Science 258:1479-1481 (1992); и Trias, J.V. et al., J.Biol.Chem. 268:6234-6240 (1993)). Jarlier, V. et al., J.Bacteriol. 172:1418-1423 (1990) показали, что цефалоспорины используют гидрофильный путь, чтобы проникнуть в клетку и далее, анионные цефалоспорины диффундируют в клетку значительно медленнее, чем те, у которых "нет чистого заряда". Препятствием, задерживающим движение катионных детергентов через наружную мембрану, является то, что порин по Trias, J.V. et al. , J. Biol.Chem. 268:6234-6240 (1993) зависит от напряжения. Аккумуляция катионных детергентов просто закроет канал.
Проникнув через внешнюю мембрану, бетаины пересекают внутреннюю мембрану путем активного транспорта. Schaefer, W.B. et al., J.Bacteriol. 90:1438-1447 (1965) изучали M. kansasii и Weir, M.P. et al., Amer. Rev. Res. Dis. 106: 450-457 (1972) изучали M. smegmatis и показали, что поглощение олеата быстрое и энергозависимое. Дополнительно, Weir, M.P. et al., Amer. Rev. Res. Dis. 106:450-457 (1972) показали, что вовлеченный механизм до определенного уровня стереоспецифичен. McCarthy, C.Infect.Immun. 4:199-204 (1971) изучал M. avium и докладывал, что 78 нмоль пальмитиновой кислоты (CAS® N. 57-10-3) могут быть удалены 1 миллиграммом клеток за 30 минут. Существуют, однако, специфические видовые свойства. Schaefer, W. B. et al., J.Bacteriol. 90: 1438-1447 (1965) заявляли, что MTB-комплекс организмов "...не демонстрирует тех же самых поразительных изменений в морфологии и мутности среды на средах с Tween 80 и олеиновой кислотой." Cella, J.A. et al., J.Am.Chem.Soc. 74: 2061-20-62 (1952) заключали, что C16-C18 четвертичные соли аммония вызывают наилучшую бактерицидную активность, и Tsubone, K., J.Pharm.Sci. 80:441-444 (1991) ясно показывает, что C16-фосфоэтилбетаин (CAS® N. 126712-88-7) имеет идеальную длину для указанной активности (как докладывалось, гомолог C18-мутный в условиях опыта). Оба автора показали, что соединения с короткой цепью являются относительно менее эффективными антимикробными веществами. Tsubone, K. , J. Pharm. Sci. 80:441-444 (1991) также показали значительную видовую зависимость от активности. Следовательно, что переносчик липидов демонстрирует значительно более высокую специфическую активность в отношении более длинных цепей липидов, и эта активность демонстрирует широкий спектр вариаций среди видов. Тем не менее, нет значительных препятствий, удерживающих бетаины от проникновения в клетку. Преимущество бетаинов в методах данного исследования состоит в том, что они идеальным образом структурно подходят клетке.
Если описанный путь вовлечен в SB-18-подобную активность, то могут существовать также дополнительные факторы, зависящие от pH и температуры. Например, McCarthy, C. Infect.Immun. 4:199-204 (1971) показал, что переносчик липидов М. avium зависит как от pH, так и от температуры: транспорт оптимален при 38oC и pH 6,5. Поскольку изменение pH и понижение температуры во время процесса могут иметь негативные последствия в отношении химической и фазовой структур SB-18, соответственно, эти параметры могут также нарушать способность аккумулировать SB-18-подобные детергенты. Например, Tsujii, K. et al., J.Phys.Chem. 82:1610-1614 (1978) показали, что SB-18 необходимо держать в 10 мМ NaCl выше 37,5oC для поддержания мицеллярной структуры. Tsubone, K. et al., J. Am. Oil Chem. Soc. 67:149-153 (1990) показали, что более низкий pKa Cn-фосфоэтилбетаина попадает в диапазон 4,7-4,9 и более высокий pKa - в диапазон 8,8-9,8. Weers, J.G. et al., Langmuir 7:854-867 (1991) определили более низкое значение pKa для широкого диапазона C12-карбоксибетаинов и показали его зависимость от структуры мостиковой группы. Сохранение условий опыта в пределах этих диапазонов pH обеспечивается функционированием бетаина в качестве детергента (т.е. мицеллярная форма поддерживается, допуская таким образом солюбилизацию миколовых кислот и липидов, связанных с тяжами микобактерий). Эксперименты, использующие мягкие кислотные буферы (напр. pH 5,7), не препятствовали существенным образом концентрированию клеток М. tuberculosis. Этот результат, однако, может быть видоспецифичным. Таким образом, для получения максимальной выгоды от использования бетаинов в методах по данному изобретению, для определения широкого разнообразия организмов, pH надо поддерживать выше 5,0, но не на много больше, чем 8,0, однако любое значение pH, позволяющее SB-18-подобному детергенту проявлять активность SB-18-подобного детергента, приемлемо.
Причина, по которой короткоцепочечные детергенты демонстрируют меньшую активность в описанных здесь опытах, может в действительности более основываться на факте, что существует связь между длиной алкильной цепи и антимикробной активностью. Наиболее интересные находки Tsubone, K.,J.Pharm.Sci. 80: 441-444 (1991) - это то, что из девяти проверенных организмов (как грибов, так и бактерий) на Propionobacterium acnes и Streptococcus mutans группа фосфобетаинов влияла значительно более драматично. Из девяти организмов, P. acnes наиболее близок к микобактериям. Установлено, что последовательность прямого праймера (TBv2-119 [SEQ ID N.:1:]) полностью гомологична P. acnes в этом участке в генной последовательности 16S рРНК. В заключение можно сказать, что или бетаины эффективно изолированы, или структуры чрезвычайно сильно действующие, или и то и другое. Имея теперь в основном описанное изобретение, то же самое можно легче понять через ссылки на следующие примеры, которые предлагаются в качестве иллюстрации, и не предполагается, что они ограничивают настоящее изобретение.
ПРИМЕРЫ
Пример 1
Материалы и Методы
I. Приготовление промывного буфера SB-18
Промывной раствор SB-18 (на 500 мл)
A. Добавьте 25 мл солей 20X SB-18 (см. ниже) к приблизительно 475 мл H2O (или 5 мл на 100 мл промывного буфера) и автоклавируйте (20 мин текучим паром).
B. Удалите колбу из автоклава и немедленно добавьте 5 мл 100X SB-18 (или 1 мл 100X SB-18 на 100 мл промывного буфера). Смешайте круговыми движениями и поместите на 40oC.
C. Храните колбы при 40oC до использования. (Колбы могут храниться несколько дней перед использованием).
D. Дополнительно: Непосредственно перед использованием согрейте 1,25 М DTT до комнатной температуры и добавьте 2 мл 1,25 М DTT на 500 мл SB-18-промывного раствора (окончательная концентрация: 5 мМ или 400 мкл 1,25 М DTT на 100 мл промывного буфера). Хорошо смешайте круговыми движениями. Важно, чтобы DTT не был холодным при добавлении.
Примечание: не используйте при наличии осадка.
E. Используйте как описано в примерах ниже.
F. Выбросите оставшееся содержимое после использования.
На любой стадии этой процедуры при наличии осадка не используйте промывной буфер. Дополнительно, растворы следует держать теплыми постоянно перед и во время использования. Температура Краффта для SB-18 в этих условиях приблизительно 37oC (Tsujii, K. et al., J.Phys.Chem. 82:1610- 1614 (1978)). Таким образом, раствор можно хранить выше этой температуры, чтобы избежать кристаллизации и быть уверенным в образовании стабильных мицелл.
II. Компоненты промывного буфера SB-18
По этой прописи делается 10 литров промывного буфера. Этого достаточно для обработки около 400 образцов по 25 мл каждый. В примерах в дальнейшем используют буфер, описанный ниже, однако, как указано в примерах 9 и 12, этот буфер может быть модифицирован, чтобы учесть специфику используемого SB-18-подобного детергента (прим. 9; фиг. 10) также, как и специфику требуемого способа обработки (прим. 12: табл. 11).
(A) Соли для промывного раствора 20X SB-18 (см. табл. 14).
H2O до 200 мл и смешать.
Простерилизуйте фильтрованием и разлейте на две порции по 100 мл. Храните при комнатной температуре. Этот раствор можно делать еженедельно или ежемесячно и хранить.
*Примечание: Использование аминокислот, таких как фенилаланин, не обязательно. Однако, аминокислоты следует автоклавировать только при pH 7,0. Таким образом, если аминокислоты использованы, pH у 20X основного раствора следует уменьшить до pH 7,0.
(B) Основной раствор 100X SB-18
(SB-18 удобно добавить, растворив первоначально в смеси 1:1 изопропанол: вода).
A. Перенесите 50 мл раствора изопропанол:вода (1:1, см. ниже) в градуированный цилиндр.
B. Взвесьте 3,358 г SB-18 (N-октадецил-N,N-диметил-3-аммоний-1- пропан сульфонат; По каталогу Sigma N. O 8004), поместите в цилиндр и осторожно перемешивайте. Позвольте раствору осесть в течение около 20 мин и осторожно перемешивайте примерно каждые 5 мин.
C. Добавьте еще раствора изопропанол:вода до примерно 75 мл и осторожно перемешайте. Когда SB-18 растворится (всего примерно 30 мин), доведите окончательный объем до 100 мл и перемешайте переворачиванием.
D. Перенесите раствор в стерильную пластиковую бутылочку Nalgene.
E. Этот раствор может храниться при комнатной температуре месяцами.
(C) 1,25 DTT
Растворите 7,71 г дитиотриэтол (154,2 г/моль) в 40 мл воды. Быстро разлейте на порции по 2 мл. Храните при -20oC.
III. Реагенты промывочного буфера SB-18
(A) 1 М NaHPO4 (см. табл. 15).
Дистиллированной воды до 1 литра.
Стерилизовать фильтрацией, разлить на аликвоты по 250 мл. Хранить при комнатной температуре.
Примечание: 1 М NaHPO4-буфер может со временем образовать осадок при комнатной температуре. Лучше сделать большой объем разведенного, 20X для продолжительного хранения.
*Примечание: pH 8,0 предпочтителен, если в сочетании с SB-18 не были использованы аминокислоты, при использовании аминокислот pH 7,0 предпочтительней.
(В) 5 М NaCl
NaCl (58,44 г/моль) - 292,2 г
Добавить H2O до 1 литра. Простерилизовать фильтрацией, разлить на аликвоты по 100 мл. Хранить при комнатной температуре.
(C) 1:1; изопропанол:вода
Смешайте 50 мл изопропанола с 50 мл воды.
IV. 2X Литический буфер
10X Taq Буфер - 8 мл
0,45% Tween 20 - 180 мкл
0,45% NP-40 - 180 мкл
200 мкг/мл протеиназы K - 160 мкл (50 мг/мл)
Вода до 40 мл.
(A) 10X Taq Буфер см. табл. 16).
Долить водой до 100 мл. Простерилизовать фильтрованием и разлить на порции по 9 мл. Хранить при -20oC.
(B) Протеиназа K (50 мг/мл)
Протеиназа K: 500 мг в 10 мл воды.
Разлить на порции по 175 мкл в пробирки для микроцентрифугирования объемом 0,5 мл. Хранить при -20oC.
(C) 1 М Трис-HCl pH 8,9
Трис основание (121,1 г/моль) - 9,472 г
Трис-HCl (157,6 г/моль) - 3,438 г
Водой долить до 100 мл. Стерилизация фильтрованием. Аликвоты: каждая по 25 мл. Хранить при 4oC.
(D) 4 М KCl
KCl (74.55 г/моль) - 29,82 г
Добавить воды до 100 мл. Простерилизовать фильтрованием. Аликвоты: каждая по 15 мл. Хранить при комнатной температуре.
V. Источники, использованные для воплощений, приведенных в качестве примеров (см. табл. 17).
VI. Вода
Для больших объемов, включая промывной раствор SB-18, была использована Milli-Q очищенная вода.
Для меньших объемов была использована вода Gibco/BRL:
1 литр - 15230-022
100 мл - 15230-014
"Меньшие" объемы включали те, что требовались для приготовления:
a. 50X соли для промывного раствора SB-18:
1 М NaHPO4 и 5 М NaCl
b. 100 K SB-18 основной раствор:
Изопропанол:вода (1:1)
c. 1,25 М DTT
d. 2X Литический буфер:
10X Taq Буфер, 1 М Трис-HCl pH 8,9, 4 М KCl и протеиназа K (50 мг/мл)
VII. Обработка клинического материала для высева
Все клинические материалы были направлены в нашу лабораторию для обычного высева и обработаны по методу N-ацетил-L- цистеин/NaOH (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 31-46). Бактериальные осадки были ресуспендированы в 1 мл стерильной воды и выделены порции для посева на среду BACTEC 12B (Becton- Dickinson, Towson, MD), для инкубации на Lowenstein-Jensen-агар (Becton-Dickinson, Towson, MD) и для анализа мазка (Kent, P.T. et al., "Public Health Mycobacteriology, " в A Guide for the Level III Laboratory, U.S. Department of Health and Human Service, Center for Disease Control, (1985) pp. 57-70). Оставшийся осадок был или заморожен при -20oC, или далее обработан для ПЦР, как описано ниже. Все культуры, положительные на BACTEC, были идентифицированы или методом Gen-Probe (GenProbe, San Diego, CA), или стандартными биохимическими анализами (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S. Department of Health and Human Service, Center for Disease Control, (1985) pp. 71-157).
VIII. Прямая амплификация осадка.
Таблица 4: 200 мкл осадка были непосредственно помещены в пробирку для микроцентрифугирования с закручивающейся крышкой, содержащую 200 мкл 2X литического буфера (40 мМ Трис-HCl [pH 8,3], 100 мМ KCl, 0,45% Tween 20, 0,45% NP-40 и 200 мкг/мл протеиназы K). Материал инкубировали при 60oC в течение 60 мин и затем кипятили 15 минут. Аликвота в 50 мкл была затем отобрана для амплификации путем ПЦР.
IX. Промывание осадка
Осадки промывались различными растворами, как описано ниже.
Таблица 5: было три следующих условия промывания: (i) Промывание водой: 25 мл стерильной воды добавляли к осадку, сильно перемешивали и затем немедленно подвергали центрифугированию при 3500 g в течение 20 мин при 4oC.
(ii) Промывание дитиотриэтолом (DTT): 25 мл 10 мМ KHPO4 (калий фосфатный буфер, смесь KHPO4 и K2HPO4) pH 8,0 и 5 мМ DTT были добавлены к осадку, перемешаны и инкубированы при комнатной температуре в течение 20 минут, затем подвергнуты центрифугированию при 7410 g в течение 20 мин при 4oC.
(iii) Промывание Tween 20/NP-40: 25 мл 10 мМ KHPO4 (pH 8,0) и 5 мМ DTT с 0,05% Tween 20 и 0,05% NP-40 были добавлены к осадку, перемешаны и инкубированы при комнатной температуре в течение 20 минут при встряхивании, затем подвергнуты центрифугированию при 7410 g в течение 20 мин при 4oC. Во всех случаях супернатант был слит и было добавлено 200 мкл стерильной воды для ресуспендирования осадка. Двести мкл были затем перенесены в пробирки для микроцентрифугирования с закручивающимися крышками, содержащими 200 мкл 2X литического буфера. Материалы инкубировали при 60oC 60 минут и затем кипятили 15 минут. Все пробирки были подвергнуты вращению в течение 1 мин при 12000 g перед отбором аликвоты в 50 мкл для амплификации.
Таблица 6: 25 мл 10 мМ NaHPO4 (pH 7,0), 45 мМ NaCl, 5 мМ DTT и также 200 мкМ или 2 мМ N-октадецил-N,N-диметил-3-аммоний-1-пропан сульфоната (SB-18 [Sigma Кат, N. 08004]) были добавлены к осадку, перемешаны и инкубированы при 37oC в течение 60 минут при встряхивании (140 об/мин), затем подвергнуты центрифугированию при 7410 g в течение 20 мин при 37oC. Супернатант слит и добавлено 200 мкл стерильной воды для ресуспендирования осадка. Двести мкл были затем перенесены в пробирки для микроцентрифугирования с закручивающимися крышками, содержащими 200 мкл 2X литического буфера. Материалы инкубировали при 60oC 60 минут и затем кипятили 30 минут. Все пробирки были подвергнуты вращению в течение 1 мин при 12000 g перед отбором аликвоты в 50 мкл для амплификации.
Таблица 9: 25 мл 10 мМ NaHPO4 (pH 7,0), 15 мМ NaCl, 5 мМ DTT и 2 мМ SB-18 были добавлены к осадку, перемешаны и инкубированы при 37oC в течение 60 минут при встряхивании (140 об/мин). Крышки были затем удалены и пробирки помещены в вакуумную печь, предварительно разогретую до 40oC. Вакуум 600 мм рт. ст. применяли в течение 60 минут. Все пробирки были удалены из вакуумной печи, крышки надеты и пробирки подвергнуты центрифугированию при 7410 g в течение 20 мин при 37oC. Супернатант был слит и осадки ресуспендированы с добавлением 200 мкл стерильной воды. Двести мкл были затем перенесены в пробирки для микроцентрифугирования с закручивающимися крышками, содержащими 200 мкл 2X литического буфера. Материалы инкубировали при 60oC 60 минут и затем кипятили 15 минут. Все пробирки были подвергнуты вращению в течение 1 мин при 12000 g перед отбором аликвоты в 50 мкл для амплификации. Для окончательной оценки все образцы были амплифицированы в повторе.
X. Система ПЦР
Разработанная система ПЦР уникальна и основывается на последовательности генов 16S рРНК, как опубликовано Rogall, T. et al., Int. J. Syst. Bacteriol. 40: 323-330 (1990). Праймеры были подобраны так, чтобы иметь возможность обеспечить оптимальную амплификацию следующих групп/видов микобактерий: М. tuberculosis (ТБ комплекс: [MTB]), М. avium - М. intracellulare, М. paratuberculosis (MAC комплекс), М. kansasii и М. marinum. Прямой праймер был создан в отношении нуклеотидов 119-144 (согласно номенклатуре Rogall, T. et al. , Int. J. Syst. Bacteriol. 40:323-330 (1990)) второй вариабельной (V2) области последовательности генов 16S рРНК этих микобактерий. Последовательность прямого праймера (TBv2-119): 5'-AAACTG GGT CTA ATA CCG GAT AGG A-3' [SEQ ID N. :1:]. Обратный праймер был создан в отношении нуклеотидов 431-453 третьей вариабельной области (V3). Последовательность обратного праймера (TBv3-453): 5'-CCA CCT ACC GTC AAT CCG AGA-3' [SEQ ID N.:2:]. Продукт амплификации был приблизительно 335 пар оснований (в зависимости от амплифицированного вида). Родоспецифический зонд был создан в отношении центральной части продукта амплификации и является общим для всех микобактерий. Его последовательность: 5'-GCG GGC iCA TCC CAC ACC GC-3' [SEQ ID N.:3:]. Видоспецифический зонд для MTB был создан в отношении особой части продукта амплификации и он специфичен для организмов ТБ комплекса. Его последовательность: 5'-GAC CAC GGG ATG CAT GTC TTG TG-3' [SEQ ID N.:4:].
Метод амплификации использует 9600 Thermal Cycler (Perkin Elmer, Norwalk, CT) и включает урацил-ДНК-гликозилазы (UDG [Life Technologies, Inc. Gaitherburg, MD]) схему стерилизации во всех реакциях (Longo, V.C. et al., Gene 93:125-128 (1990)). Каждые 100 мкл реакционной смеси содержали конечную концентрацию 200 мкМ dATP/dGTP и 400 мкМ dUTP (dU-dNTP: Boerhringer, Indianapolis, IN), 3,0 мМ MgCl2, 25 пмолей каждого праймера с 2,5 единицами Taq ДНК-полимеразы (Perkin Elmer, Norwalk, CT). Конечная концентрация буфера составляла 50 мМ Трис-HCl pH 8,9, 10 мМ KCl и 0,225% каждого Tween 20/NP-40. AmpliWax (Perkin Elmer, Norwalk, CT) было также включено для оптимизации процесса. Реакционные смеси были следующими: 25 мкл (содержащих 2,5 единицы Taq, 0,25 единицы UDG, dU-dNTP's и 25 пмолей TBv2-119 в 1X буфере) помещены в пробирку для центрифугирования на 0,2 мл с бусами AmpliWax, нагреты до 80oC в течение 5 мин и затем охлаждены. 25 мкл (содержащих MgCl2, 0,75 единиц UDG и 25 пмолей TBv3-453 в 1X буфере) были затем нанесены слоем на AmpliWax. Материал был добавлен в объеме 50 мкл в 1X-ПЦР буфере. Циклируемый профиль был: 45oC 5 мин, 94oC 7 мин, затем 40 циклов 94oC по 20 сек, 61oC 1 сек и 72oC 10 сек, при последующим выдерживании в реакционной зоне при 72oC. После термального циклирования реакционные смеси были заморожены при -20oC.
Положительный контроль амплификации был клонирован с использованием CloneAmp System (Life Technologies, Inc. Gaitherburg, MD). Праймеры TBv2-119 и TBv3-453 были модифицированы согласно производителю. Колонии Mycobacterium tuberculosis ATCC # 27294 (ATCC, Rockville, MD), выращенные в пробирках со скошенным агаром, помещали в
200 мкл ПЦР-литического буфера и кипятили 30 минут. Аликвоту в 10 мкл отбирали и обрабатывали для амплификации, как описано выше, за исключением того, что вместо 200 мкМ dTTP было использовано 400 мкМ dUTP, и фермент UDG был исключен из реакции. Продукт был клонирован в клетках DH5α согласно методике производителя (Life Technologies, Inc. Gaitherburg, MD). Плазмида (pOL332) была очищена Applied Biotechnologies, Inc. (Ellicot City, MD) и определена последовательность с использованием набора для циклического сиквинирования (Life Technologies, Inc. Gaitherburg, MD) для подтверждения идентичности. Очищенная плазмида была оценена количественно A260 считываниями и затем разбавлена для использования в качестве положительного контроля амплификации. Все действия по амплификации включали двойные реакции, содержащие 20 и 100 копий этой плазмиды. Дополнительно, двойные отрицательные контроли были также амплифицированы на каждом этапе. Эта плазмида также использовалась как контроль гибридизации в каждом блоте.
XI. Определение
Амплифицированные образцы были удалены из -20oC и приготовлены для блоттинга добавлением 100 мкл 2X-денатурированного раствора (1М NaOH/2М NaCl). Реакционные пробирки были затем помещены обратно в термальный циклер и нагреты до 60oC в течение 15 мин. Все зонды были помечены на 5'-конце с использованием γ- [32P]ATP и полинуклеотидкиназы (Life Technologies, Inc. Gaitherburg, MD) согласно указаниям производителя. Зонд был очищен с использованием колонки Chromaspin (...ontech, Palo alto, CA). Образцы были подвергнуты дот блоттингу на Nytran Plus (Schleicher & Schuell, Keene, NH) и обработаны зондом согласно рекомендациям производителя. Коротко, фильтр был обожжен при 80oC в течение часа и затем был прегибридизирован 3 часа при 42oC в 10X Denhardt'a, 6X SSPE, 1% SDS и 100 мкг/мл денатурированной ДНК спермы сельди. Фильтр затем был помещен в 6X SSPE и 3% SDS с 5'- 32P-меченым зондом и инкубировался в течение ночи или при 61oC (родоспецифичный зонд [SEQ ID N.:3:] ) или при 65oC (TB- специфичный зонд [SEQ ID N.:4:]). Фильтр был трижды промыт при комнатной температуре в 6X SSPE каждый раз 10 мин, и затем однократно или при 61oC 3 минуты в 1X SSPE для родоспецифичного зонда [SEQ ID N.:3:] , или при 65oC 3 минуты в IX SSPE для TB- специфичного зонда [SEQ ID N.:4:]. Затем фильтр был подвергнут авторадиографии. Фильтры были очищены кипячением в 0,1% SDS в течение 15 мин и заново обработаны зондом при необходимости.
XII. Оптимизация ПЦР
Положительный контроль был использован как средство оптимизировать систему. Концентрация MgCl2, концентрация нуклеотидов, различные профили амплификации и температуры обжигания были проверены для того, чтобы оптимизировать систему. Продукты ПЦР были проанализированы электрофорезом в агарозном геле с окраской бромистым этидием, а также дот блоттингом и гибридизацией. Условия амплификации, описанные выше, могут обычно амплифицировать 20 копий до определяемого уровня. Мы заметили феномен, уже описанный Carmody, M. W. et al., Biotechniques 15:692-699 (1993). Собственно, количество продукта dU-ПЦР, видимого на геле, больше, чем количество, оцениваемое гибридизацией (при сравнении известных количеств блоттированной dT-плазмиды). Мы пытались использовать "горячий старт UDG", как описано Loewy, Z.G. et al. , J.Clin.Micro. 32:135-138 (1994), но не наблюдали того же усиления сигнала.
XIII. Организация дот блотов
Как описано выше, продукт ПЦР денатурирован и ковалентно иммобилизован на нейлоновой мембране в двух измерениях. Создание порядка в двух измерениях выполнено с использованием дот-блот множащего аппарата ("The Convertible", Life Technologies, Inc. Gaitherburg, MD), который представляет собой стандартный формат 96 лунок (то есть 8 лунок на 12 лунок).
Нижний ряд на фиг. 2A и 16B, или два нижних ряда на фиг. 3, 4A, 5, 6, 7, 7A, 8, 9, 9A, 9B, 9C, 9D, 10, 11A, 12 и 13, или колонка 1 на фиг. 14 представляют результаты контролей гибридизации и амплификации. Контроли гибридизации - это лунки, обозначенные "108, 109, 1010" и поставленные в повторе в том же ряду на фиг. 2A, 16A и в той же колонке на фиг. 3, 4A, 5, 6, 7, 7A, 8, 9, 9A, 9B, 9C, 9D, 10, 11A, 12, 13 и 14. Указанное число копий плазмиды pOL332 было использовано как контроли гибридизации и их можно сравнивать от блота к блоту для определения экспозиции.
Таким же образом, контроли амплификации состоят из дублированных реакций, содержащих "0, 20 и 100" стартовых копий плазмиды pOL332, и могут быть использованы в рамках эксперимента для определения эффективности данного прохождения амплификации. Контроли амплификации также находятся в повторе на том же ряду на фиг. 2A, 16A и в той же колонке на фиг. 3, 4A, 5, 6, 7, 7A, 8, 9, 9A, 9B, 9C, 9D, 10, 11A, 12, 13 и 14.
B большинстве экспериментов используются мультиплицированные репликаты, которые считаются идентичными при начале эксперимента. Эти образцы репликатов помечены "a", "b", "c", "d" и т.д. и амплифицированы, по меньше мере, в повторе, как указано скобками, охватывающими отметку. Каждая экспериментальная лунка показана по меньшей мере в повторе, что отмечено скобками. Несколько экспериментов просто сравнивали "условия опыта", тогда как многие из экспериментов использовали несколько внутренних контролей: "отрицательный контроль опыта", "положительный контроль опыта" и "контроль ввода опыта". Контроль ввода опыта (также известный как "прямой ввод" или "прямая аликвота") представляет максимальное число копий, взятых в данном эксперименте (т. е. дот максимальной интенсивности). Сравнение этих серий с контролями испытания или проверкой условий представляет действенность процесса. Отрицательный контроль испытания (вода) собственно представляет низкую эффективность выделения, и положительный контроль испытания (2 мМ SB-18) представляет высокую эффективность выделения. Сравнение контролей: положительного, отрицательного и ввода испытания с различными проверками условий позволяют сделать заключение о том, улучшают ли модификации эффективность выделения.
XIV. Разрешение расхождения результатов.
В эмпирической работе все образцы, оказавшиеся положительными при высеве, но отрицательными по ПЦР, были проверены на ингибиторы. В этих случаях 1000 копий положительного контроля была введена в выпадающие образцы, чтобы проверить ингибирование амплификации. При оценке было несколько случаев, в которых дубликаты амплификации одного и того же образца давали разные результаты (т.е. один был положительным и один - отрицательным). Эти образцы были снова проверены в повторе и результаты разрешались аналогично тому, как описано Jackson et al., J.Clin.Microbiol. 31:3123-3128(1993): если хотя бы одна из второго комплекта реакций была положительной, образец считался бы положительным.
XV. Специфичность ПЦР
Для проверки специфичности системы с праймерами и зондами, описанными выше, три категории микроорганизмов были проверены на их способность быть амплифицированными. Три категории были основаны на предварительной группировке следующим образом: (i) те, которые должны амплифицироваться оптимально, (ii) те, которые могут амплифицироваться, но с уменьшенной действенностью из-за небольших ошибок спаривания оснований в последовательности праймера, и (iii) те, которые не должны амплифицироваться из-за отсутствия последовательностей в праймерах. Следующие штаммы Американской Коллекции Типовых Культур (ATCC, Rockville, MD) были использованы для проверки первой категории: М. tuberculosis (ATCC #25294), М. avium (ATCC #25291), М. intracellulare (ATCC # 13950), М. kansanii (ATCC # 12478), М. paratuberculosis (ATCC # 19698), М. marinum (ATCC #927), М. szulgai (ATCC #35799), М. gastri (ATCC # 15754). Следующие штаммы ATCC были использованы для проверки второй категории: М. xenopi (ATCC # 19250), M. gorgonae (ATCC # 14470), М. malmoense (ATCC # 29571), М. terra (ATCC # 15755), М. nonchromogenicum (ATCC # 19530). Следующие штаммы ATCC были использованы для проверки третьей категории: М. fortuitum (ATCC #6841) представлял быстрорастущие микобактерии, которые не должны амплифицироваться; неродственные, но похожие организмы, включающие Propionibacterium acnes (ATCC #6919), Corynebacterium xerosis (ATCC # 373) и Rhodococcus equi (ATCC #6939); другие микроорганизмы, найденные в значительных количествах в мокроте и бронхиальных материалах (Murray, P. R. et al., in: Manual of Clinical Microbiology, 5th edition, A. Balows et al. eds., Am.Soc.Microbiol., Washington, DC, 1991, pp. 488-490) были представлены Prevotella melaninogenica (ATCC # 25845), Peptostreptococcus magnus (ATCC # 15794). Все микобактерии росли на пробирках со скошенным агаром перед использованием. Все другие микроорганизмы были выращены, как предлагает ATCC.
Продукты ПЦР анализировали как гель-электрофорезом на агарозном геле с окраской бромистым этидием, так и дот блоттингом и гибридизацией. Все микроорганизмы первой категории амплифицировались с равной эффективностью. Микроорганизмы второй категории распались на две подгруппы, основанные на комплементарности к праймерам. M. gorgonae (1 ошибка спаривания оснований в прямом праймере и 2 ошибки спаривания оснований в обратном праймере) амплифицировалась только при высоком числе копий. М. malmoense (1 ошибка спаривания оснований только в прямом праймере) амплифицировалась с меньшей эффективностью. Во второй подгруппе М. terra, М. nonchromogenicum и М. xenopi каждая имела большое количество ошибок спаривания оснований и не амплифицировалась. Ни один из микроорганизмов третьей категории не амплифицировался даже при высоком числе копий.
Пример 2
Непосредственное кипячение образцов в ПЦР буфере было недостаточным
Сначала, для лучшего выделения микобактерий в форме, поддающейся амплификации, была сделана попытка непосредственного кипячения осадка, как описано Victor et al., J. Clin. Micro. 30:1514-1517 (1992). 76 замороженных осадков NALC/NaOH были непосредственно помещены в ПЦР-буфер и кипятились (табл. 4). Как описано выше, для прямой амплификации осадка, 200 мкл осадка были непосредственно перенесены в пробирку для микроцентрифугирования с закручивающейся крышкой, содержащую 200 мкл 2X литического буфера (40 мМ Трис-HCl [pH 8,3] , 100 мМ KCl, 0,45% Tween 20, 0,45% NP-40 и 200 мкг/мл протеиназы K). Осадки инкубировались при 60oC 60 мин и затем кипятились в течение 15 мин. Аликвоту в 50 мкл отбирали затем для амплификации ПЦР.
Таблица 4: Корреляция ПЦР с высевом, когда осадок NALC/NaOH амплифицирован непосредственно
Показаны результаты прямой ПЦР амплификации осадков NALC/NaOH. Осадок был удален перед амплификацией и выращен в культуральных колбах BACTEC 12B, как описано выше. Все положительные культуры МТБ или MAC были идентифицированы с использованием методики Gen-Probe (GenProbe, San Diego, CA). Все другие были идентифицированы биохимическим анализом (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U. S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 71-157).
Как показано в табл. 4, только 1 из 7 образцов, положительных при высеве, был определен амплификацией (14,3%), когда осадок был использован непосредственно. В каждом случае было возможно амплифицировать организм из культуры, предполагая, что праймерная пара функциональна в этих изолятах. Как было определено, каждый ложноотрицательный ответ был результатом ингибирования ПЦР из-за экстракции NALC/NaOH.
Пример 3
Эффекты ингибирования амплификации
Наша лаборатория обычно использует 3% NaOH для обработки клинических материалов (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 31-46). Концентрация NaOH (т.е. OH⊖) в 3% основном растворе составляет 0,75 мМ при pH 14. Общая концентрация Na⊕ в 3% растворе (включая 1,45% цитрат натрия) составляет 0,8 М. Были выполнены эксперименты, показанные на фиг. 1, для того, чтобы проверить потенциальное ингибирующее влияние на ПЦР, вызванное концентрацией соли (т.е. Na⊕) или pH (т.е. OH⊖ концентрацией). 5 мл NALC/NaOH (5% NALC, 1,45% цитрата натрия и 3% NaOH) были разбавлены в 10 раз 45 мл воды. Конечная концентрация растворенных компонентов составляла 3,1 мМ NALC, 5,6 мМ цитрата натрия и 75 мМ NaOH. Различные количества разбавленного NALC/NaOH были затем помещены в пробирки для микроцентрифугирования на 1,5 мл, содержащие 200 мкл 2X литического буфера. Приблизительно 1000 копий положительного контроля плазмиды pOL332 были помещены в каждую смесь и все объемы были доведены водой до конечного объема 400 мкл. Все пробирки инкубировали при 95oC в течение 15 мин и затем 50 мкл каждой было амплифицировано в повторе. Результаты показаны на фиг. 1 (см. секцию XIII в примере 1 для объяснения формата данных). Процент показывает количество NALC/NaOH в последней пробирке с реакцией амплификации. Например, 25% будут означать, что 25 мкл раствора NALC/NaOH (3,1 мМ NALC, 5,6 мМ цитрата натрия и 75 мМ NaOH) были помещены в реакционную смесь для амплификации в конечном объеме 100 мкл. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 также были подвергнуты блоттингу.
Результаты, представленные на фиг. 1, предполагают, что разведенный раствор NALC/NaOH должен быть разведен в дальнейшем более чем в 10 раз в конечной смеси для амплификации для того, чтобы ПЦР функционировала действенно. Таким образом, как добавление соли, так и OH⊖ должно быть менее чем примерно 5 мМ для эффективного процесса амплификации. Так как ПЦР более чувствительна к изменениям в pH (относительно слабого повышения в отношении соли), причина ингибирования - в изменениях pH. Учитывая вариабельность консистенции материала, невозможно удалить всю NALC/NaOH простым декантированием. Эти данные подтверждают, что необходима дальнейшая обработка образца, если NALC/NaOH использовалась на первой стадии обработки материала. Это заключение далее поддерживается данными Noordhoek, G.T. et al., J. Clin. Micro. 32:277- 284 (1994): две лаборатории, имевшие худшие результаты в этом исследовании, не очищали NALC/NaOH-осадки.
Пример 4
Влияние дополнительных вторичных стадий промывания водой и DTT
I. Промывание водой
Для уменьшения потерь на каждый образец и увеличения действенности, когда требуется обработать большое число образцов, была проверена применимость вторичной стадии промывки (2o-Промывка) для дальнейшего избавления от ингибиторов, таких как NALC/NaOH (табл. 5).
Условия амплификации, как описано выше в Материалах и Методах.
Сначала была использована простая промывка водой для промывания осадка. К осадку добавили 25 мл стерильной воды, перемешали и затем немедленно подвергли центрифугированию при 3500 g в течение 20 мин при 4oC. Супернатант слит и 200 мкл стерильной воды добавили для ресуспендирования осадка. Затем 200 мкл были перенесены в пробирки для микроцентрифугирования с завинчивающейся крышкой, содержащие 200 мкл 2X литического буфера. Материал инкубировали при 60oC 60 мин и затем кипятили 15 мин. Все пробирки были подвергнуты вращению при 12000 g перед взятием 50 мкл для амплификации.
После включения этой стадии промывки водой корреляция улучшилась до 33% (табл. 5: 7 ПЦР-положительных из 21 положительных при высеве [n=407]). Как сказано выше, возможно амплифицировать микроорганизм из высева: однако, в противоположность сказанному выше, ингибирование не является первичной проблемой, так как некоторые материалы были отрицательны и при повторной амплификации. Более того, выделение М. avium тоже является значительной проблемой. Вообще, когда корреляции были низкими, эти результаты совпадали с большинством исследований, обсужденных в табл. 1. Из 33 методологий, показывающих корреляцию менее 100% в табл. 1, 21 включают какую-либо стадию буферного обмена. Оставшиеся 12 не включают стадию буферного обмена и 9 ссылаются на ингибиторы, как на проблему.
II. Промывка DTT
Shawar, R.M. et al., J.Clin. Micro. 31:61-65 (1993) и Hermans, P.W.M.et al. , J. Clin. Micro. 28:1204-1213 (1990) показали, что мокрота подавляет ПЦР. По-видимому, кислые мукополисахариды являются ингибиторами (Ochert, A. S. et al., PCR Methods and Application 3:365-368 (1994). С другой стороны, вязкая природа мокроты может также влиять на действенность амплификации. Buck, G.E. et al., J. Clin. Micro. 30:1331-1334 (1992) специально ссылается на вязкость как на фактор, влияющий на ложноотрицательные результаты. Некоторые экстремальные проявления этого условия, условия, которое прямо пропорционально стадии заболевания, наблюдались в ранних исследованиях. Таким образом, в попытке преодолеть проблему, композиция 2o-промывки была изменена так, чтобы разжижить мокроту.
Для лучшего разжижения мокроты, 10 мМ KHPO4, pH 8,0 и 5 мМ дитиотриэтола (DTT) были включены в стадию промывки, аналогично методу Hirsch, S.R. et al. , J. Lab. Clin. Med. 74:346- 353 (1969) и RCF была поднята во время стадии центрифугирования, как рекомендовано Ratnam, S. et al., J.Clin. Microbiol. 23: 582-585 (1986) и Rickman, T.W. et al., J.Clin. Microbiol. 11:618-620 (1980). 25 мл 10 мМ KHPO4, pH 8,0 и 5 мМ DTT были добавлены к осадку, перемешаны, инкубированы при комнатной температуре 20 мин и затем центрифугированы при 7410 g 20 мин при 4oC. Во всех случаях, супернатант был слит и добавлено 200 мкл стерильной воды для ресуспендирования осадка. 200 мкл были перенесены в пробирки для микроцентрифугирования с закручивающимися крышками, содержащими 200 мкл 2X литического буфера. Материалы инкубировались при 60oC 60 мин и затем их кипятили 15 мин. Все пробирки были подвергнуты вращению при 12000 g перед взятием 50 мкл для амплификации.
Как показано в табл. 5, 175 случайно выбранных материалов были промыты в этих условиях. Консистенция образцов улучшилась значительно и корреляция тоже улучшилась, на этот раз до 60% (6 ПЦР-положительных из 10 положительных при высеве). Как прежде, с одним исключением (см. примечание C в табл. 5), было возможно амплифицировать микроорганизм из высева (или из пробирки со скошенным агаром), и только один ложноотрицательный результат можно отнести за счет ингибирования. Однако, "статистические выпадения" снова наблюдались среди репликантов того же самого образца.
Хотя промывка осадка показала снижение ингибирования, истинно ложноотрицательные результаты не были полностью обусловлены ингибированием: при таких условиях ошибка метода доминирует над проблемой ложноотрицательных результатов.
Таблица 5: Корреляция ПЦР и высевов при использовании различных условий промывания.
Показаны результаты различных условий опытов при промывке осадков и корреляция ПЦР и высевов. Все материалы были обработаны NALC/NaOH по методике Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center tor Disease Control, (1985) pp. 31-46).
Осадок был удален перед промыванием и выращен в культуральных колбах BACTEC 12B, как описано выше. Все положительные культуры МТБ, MAC или М. kansasii были идентифицированы с использованием методики Gen-Probe (GenProbe, San Diego, CA). Все другие были идентифицированы биохимическим анализом (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 71-157). Графа Условия представляет дальнейшую обработку образцов промыванием. Были использованы три различных вторичных условия промывки: (i) вода, (ii) 10 мМ KHPO4, pH 8,0 и 5 мМ DTT и (iii) 10 мМ KHPO4, pH 8,0 и 5 мМ DTT, содержащий 0,05% Tween 20 и 0,05% NP-40, как описано в Материалах и Методах.
Пример 5
Разделение микобактерий в процессе центрифугирования
Во время описанных выше экспериментов был исследован вопрос, не могут ли микобактерий быть потеряны во время стадии промывания. Фиг. 2 описывает экспериментальную разработку опыта по стимулированию обработки (далее называемому "опыт по обработке"). Mycobacterium tuberculosis, выросшая в пробирках со скошенным агаром, была использована как источник в этих экспериментах.
Небольшая масса клеток была сначала перенесена в 10 мл 10 мМ KHPO4 (pH 8,0) и перемешивалась для образования исходной бактериальной суспензии. Исходная бактериальная суспензия затем была разбавлена в 2•103 раз и 20 мкл перенесли в 15 мл 2o- промывного буфера. Этот перенос повторялся еще 5 раз до общего числа образцов, равного 6. В то же самое время 6 аликвот по 20 мкл были непосредственно взяты из разведения 2•103 в 6 аликвот, содержащих по 380 мкл литического буфера для прямой амплификации. Эти последние аликвоты (далее называемые "прямыми", "прямого ввода" или "взятыми непосредственно" контролями) служили контролями для общего введения мишени в данном эксперименте. Пробирки, содержащие промывной буфер, были затем подвергнуты центрифугированию (7410 g при 4oC 20 мин). После центрифугирования аликвоты супернатанта (200 мкл) были помещены в 200 мкл 2X литического буфера. Супернатант был вылит, осадок ресуспендирован в 200 мкл воды и 200 мкл ресуспендированного осадка были смешаны с 200 мкл 2X литического буфера. Аликвоты в 50 мкл каждой серии были подвергнуты ПЦР, в четырех повторностях.
Ввод в этом эксперименте оценивался приблизительно в 10000 копий. Таким образом, прямая аликвота может представлять амплификацию приблизительно 1000 копий. Если организмы лизированы или гомогенно распределены в супернатанте, реакционная смесь амплификации будет содержать примерно 20 копий. Амплификация осадка представляет предположительную эффективность выделения центрифугированием. Результаты показаны на фиг. 2A (см. секцию XIII в примере 1 для объяснения формата данных).
На фиг. 2A продукты ПЦР были денатурированы, блоттированы, обработаны зондами и подвергнуты авторадиографии, как описано в Материалах и Методах. Шесть различных "материалов" представлены как a, b, c, d, e и f и должны быть идентичными. Четыре повторности амплификаций фракции супернатанта и осадка для данного материала даны на одной линии (1, 2, 3 и 4). Четыре повторности амплификаций аликвот прямого ввода, соответственно данному материалу, показаны на одной и той же линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы. Результаты (фиг. 2A) показывают, что по некоторым осадкам они отрицательны, по некоторым фракциям супернатанта - положительны, и обе фракции демонстрируют ошибки опыта.
Одним возможным объяснением для приведенных выше результатов было то, что микобактерии лизированы. Осмотически лизинг этих микроорганизмов необычен. Thacore, H. et al., Rev. Infect. Dis. 3:960-972 (1981) и Sato, H. et al. , Can.J.Microbiol. 12:255-261 (1966) докладывали, что требуются исключительные условия для получения сферопластов и "теней" у MTB. С другой стороны, Thacore, H. et al. Proc. Soc. Exptl. Biol. Med. 114:43-47 (1963) показали, что клетки, растущие в присутствии ЭДТА, приклеиваются к стенкам сосудов. Это, однако, легко преодолеть добавлением лизоцима к среде. Точное объяснение, по-видимому, в разделении микобактерии из-за плавучести. Наблюдения фиг. 2A не удивительны в свете результатов Klein, G.C. et al., Am.J. Clin. Pathol. 22:581-585 (1952): как осадок, так и супернатант были положительны при высеве в 88,8% и 82,4% всех материалов, центрифугированных при 2000 об/мин и 3000 об/мин соответственно; и осадок был отрицательным при высеве, тогда как супернатант был положительным при высеве в 2,7% и 2,2% всех материалов, центрифугированных при 2000 об/мин и 3000 об/мин соответственно.
Пример 6
Влияние клеточной стенки на выделение микобактерии.
Dubos, R. J. et al., J.Exp.Med. 83:409-423 (1946) заключают, что погруженный рост в присутствии Tween 80 (CAS® N. 9005-65-6) обусловлен "намачиванием" поверхности клетки. Если поверхностное натяжение ответственно за выпадающие результаты и поверхностное натяжение можно преодолеть добавлением неионных детергентов, кажется логичным, что эти реагенты смогут улучшить корреляцию с высевами. Однако, как показано ниже, дело не в этом.
Young, D.B. et al., Res. Microbiol. 142:55-65 (1991) продемонстрировали, что периферически ассоциированные липопротеины могут быть удалены инкубацией в неионном детергенте Triton X-100 (CAS® N. 9002-93-1). Неионные детергенты Tween 20 (CAS® N. 9005-64-5) и NP-40 (CAS® N. 127087-87-0), которые обычно используются как добавки к ПЦР, были, таким образом, включены во вторичную стадию промывания в попытке повлиять на поверхностное натяжение (или прилипаемость) микобактериальных клеток (табл. 5).
Осадки, обработанные NALC/NaOH, были промыты Tween20/NP-40 промывным раствором: 25 мл 10 мМ KHPO4 (pH 8,0) и 5 мМ DTT с 0,05% Tween 20 и 0,05% NP-40 были добавлены к осадку, перемешаны, инкубированы при комнатной температуре 20 мин при встряхивании и затем подвергнуты центрифугированию при 7410 g в течение 20 мин при 4oC. Во всех случаях пробирки были декантированы и добавлены 200 мкл стерильной воды для ресуспендирования осадка. 200 мкл были перенесены в пробирки для микроцентрифугирования, содержащие 200 мкл 2X литического буфера. Материалы инкубировали при 60oC 60 минут и затем кипятили 15 минут. Все пробирки были подвергнуты вращению при 12000 g перед взятием 50 мкл для амплификации.
Как показано в табл. 5, из 359 амплифицированных материалов 26 были положительными при высеве как для MTB-комплекса (5 материалов), так и MAC-комплекса (21 материал). MTB была амплифицирована только из 4 материалов (80%) и MAC была амплифицирована из 9 материалов (43%). Опять, отсутствие ингибирования было показано во всех, кроме одного из ложноотрицательных результатах; микроорганизмы могут быть амплифицированы из культуры и наблюдалось, что в ложноотрицательных результатах доминировали статистические выпадения.
Таким образом, суммируя результаты промывок, показанные в табл. 5, можно видеть, что была достигнута корреляция в 80% с высевом у MTB (хотя присутствовало относительно небольшое число изолятов MTB [n=10]). Результаты, касающиеся способности изолировать бактерии MAC-комплекса этим способом, поразительно отличались: корреляция с высевом была 37% [n=46]). Это расхождение также отражено в материалах табл. 1.
Для последующего подтверждения этих наблюдений был повторен эксперимент по обработке, показанный на фиг. 2, с использованием М. tuberculosis в присутствии 0,1% Triton X-100 (CAS® N. 9002-93-1). Фиг. 3 показывает результаты этого эксперимента. Опыт по обработке, описанный на фиг. 2, был использован в этом эксперименте за исключением того, что 2o-промывной буфер содержал воду или 10 мМ Трис-HCl pH 8,0 с добавлением 0,1% Triton X-100 и 5 мМ DTT. Кроме того, были взяты только 4 аликвоты и пробирки инкубировали при комнатной температуре в течение 20 мин при встряхивании. Двойные амплификации четырех аликвот, представленные как a, b, c и d, должны быть идентичны. Двойные амплификации прямых аликвот показаны на соответственно обозначенной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Результаты, показанные на фиг. 3, согласуются с приведенными выше данными (табл. 5): не следует ожидать, что неионные детергенты будут улучшать эффективность выделения. Собственно, наблюдались большие вариации, которые могут быть вызваны группированием клеток. Отметьте сверхэкспозицию этого блота (фиг. 3) относительно других блотов. Было необходимо рассмотреть эти результаты.
Заключение, связанное с этими результатами, состоит в том, что если должны быть предприняты дополнительные шаги для дальнейшего освобождения образцов от ингибиторов, простое "намачивание" клеток не решает эту проблему полностью. Это далее доказывается фактом, что из пяти исследований, показанных в табл. 1, те, которые использовали неионные детергенты для промывки осадков перед амплификацией (Clarridge, J. E. et al., J.Clin.Micro. 31: 2049-2056 (1993); Irula, J. V. et al., J.Clin.Micro. 31:1811-1814 (1993); Kolk, A.H.J. et al., J.Clin.Micro. 30:2567-2575 (1992); Shawar, R.M. et al., J. Clin. Micro. 31: 61-65 (1993) и Sritharan, V. et al., Mol.Cell Probes 5: 385-395 (1991)) не достигли 100% корреляции с высевом. Robinson, L. et al., J. Clin. Micro. 27:84-91 (1941) докладывали годами раньше, что агенты, которые смягчают поверхностное натяжение, были слабыми в усилении выделения центрифугированием. Если, в противоположность предположению Dubos (Dubos, R.J. et al. , J.Exp.Biol.Med. 58:361-362 (1945); Dubos, R.J. et al., J.Exp.Med. 83:409-423 (1946), Tween 80, олеиновая кислота и BSA влияют на пленочный рост in vivo, что противоположно компенсированию поверхностного натяжения, тогда следует ожидать, что группы клеток будут присутствовать в эксперименте, описанном здесь (фиг. 3). Основываясь на представленных здесь результатах (табл. 5) и на уровне техники (табл. 1), не получено преимуществ при включении этих неионных детергентов в промывной буфер.
Пример 7
Агрегация и дисперсия микобактерий.
Когда периферически ассоциированные липидные компоненты наверняка удалены Triton X-100 в вышеуказанном эксперименте (фиг.3), клетки все еще агрегированы. В качестве следующего шага были предприняты попытки улучшить выделение дисперсией. Фиг.4 детализирует экспериментальный замысел опыта (далее обозначенный как "опыт по агрегации"), который был предпринят для оценки способности различных детергентов диспергировать группы клеток микобактерий. Этот метод основывается на факте, что агрегация обостряет ошибки опыта: множественные аликвоты дадут похожие результаты амплификации, только если группы клеток будут диспергированы. Более того, каждая аликвота одного и того же образца будет иметь равное число микроорганизмов, только если они гомогенно распределены по раствору. Можно ожидать, что ситуация обострится при низком числе копий. Теоретически, дисперсия групп клеток должна увеличивать вероятность точного диагноза, даже при относительно низком числе копий, потому что все аликвоты будут иметь более высокую вероятность попадания в мишень.
Осуществление опыта, задуманного для оценки способности детергентов диспергировать микобактерии (фиг. 4), нуждалось в М. tuberculosis, выросшей в пробирках со скошенным агаром в качестве источника для этого эксперимента. Небольшая масса клеток была помещена сначала в 5 мл 10 мМ KHPO4 (pH 8,0) и перемешана для образования бактериального препарата. Препарат был затем разбавлен в 2•103 раз переносом 10 мкл в 20 мл 2o-промывной буфер. Различные аспекты 2o-промывного буфера менялись, включая детергент и его концентрации, использованный буфер, так же как и ионную силу и pH буфера, время инкубации, температуру инкубации и наличие или отсутствие взбалтывания. После инкубации 20 мкл были перенесены в 25 мл нормализующего буфера (10 мМ KHPO4 pH 8,0, 0,05% Tween 20, 0,05% NP-40 и 5 мМ DTT). Этот перенос был повторен трижды до образования четырех образцов. В то же самое время 4 аликвоты в 20 мкл были непосредственно удалены из 2•103-кратного разведения в четыре различные пробирки для микроцентрифугирования, содержащие 380 мкл литического буфера для прямой амплификации. Эти последние аликвоты (называемые далее "прямыми", "прямого ввода" или "взятыми непосредственно" контролями) служили контролями для общего введения в данном эксперименте. Пробирки, содержащие нормализующий буфер, были подвергнуты центрифугированию при 7410 g в течение 20 мин при 4oC. Во всех случаях супернатант слит и добавлено 200 мкл стерильной воды для ресуспендирования осадка. Немедленно были отобраны аликвоты в 200 мкл и помещены в 200 мкл 2X лизисного буфера. Все пробирки были подготовлены для ПЦР как описано в Материалах и Методах. Аликвоты в 50 мкл от каждой серии были подвергнуты ПЦР в повторе, для того, чтобы компенсировать разведение и нормализовать ввод.
Для получения данных, показанных на фиг.4A, продукты ПЦР из каждого опыта были денатурированы, блоттированы, обработаны зондами и подвергнуты авторадиографии, как описано в Материалах и Методах (см. выше). Дублированные амплификации четырех различных "материалов" для каждого условия представлены как a, b, c и d и должны быть идентичными. Двойные повторности амплификаций прямых аликвот, для каждого условия показаны на соответственно отмеченной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
На фиг.4A показан характерный результат сравнения отсутствия детергента (т. е. воды), 0,1% Tween 80 в 10 мМ Трис-HCl pH 8,0 и 2 мМ N-октадецил-N,N-диметил-3-аммоний-1-пропан сульфонат (SB-18: CAS® N. 13177-41-8) в 10 мМ NaHPO4, 15 мМ NaCl. Все пробирки инкубировали при 37oC 60 минут при встряхивании (140 об/мин) перед тем, как брать аликвоту в нормализующий буфер. Из проверенных соединений единственным детергентом, постоянно демонстрирующим значительную активность в этом опыте, был сульфопропилбетаин, SB-18. SB-16 (CAS® N. 2281-11-0) показывал некоторую дисперсию активности: SB-16 функционировал, хотя непостоянно. Другие детергенты и другие детергенты в сульфопропилбетаиновой серии, но с длиной менее чем 16 алкил-углеродов (SB-10 (CAS® N. 15163-36-7), SB-12 (CAS® N. 14933-08-5), SB-14 (CAS® N. 14933-09-6) были слабыми в дезагрегировании групп клеток MTB. Дисперсия групп клеток MTB увеличивает вероятность того, что даже при низком числе копий, во всех аликвотах будет находиться мишень.
Попытки подтвердить теорию дисперсии анализом кислотоустойчивых мазков не были успешными. Видимо, включение любого детергента значительно ухудшает способность клеток фиксироваться на предметном стекле нагреванием.
Пример 8
SB-18 способствует выделению микобактерий центрифугированием
Приведенные выше результаты были расширены путем использования опыта по обработке, описанного на фиг. 2. Наблюдалась прямая корреляция между действенностью детергента в разделении групп клеток и выделением в опыте по обработке. Фиг.5 показывает репрезентативный результат с использованием как M. avium, так и М. tuberculosis в присутствии и отсутствие 2 мМ SB-18. Схема, описанная на фиг. 2, сопровождалась экспериментом, показанным на фиг. 5. 2o-промывной буфер содержал 10 мМ NaHPO4 pH 7,0, 15 мМ NaCl, 2 мМ SB-18, 2 мМ фенилаланина и 5 мМ DTT и сравнивался с водой. Дополнительно, пробирки инкубировали при 37oC в течение 60 мин при встряхивании (140 об/мин). Дублированные амплификации четырех аликвот представлены как a, b, c и d и должны быть идентичными. Двойные повторности амплификаций прямых аликвот, соответственно данным видам, показаны на соответственно отмеченной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Этот блот был выбран, так как характерное для М. tuberculosis слипание, собирание в группы и характерное для М. avium более диффузное распространение явно различимы. Дополнительно, в отношении слипания, отмеченного в этом блоте, наблюдалось, что фенилаланин, повышая сбор организмов MAC, уменьшает действенность дисперсионной активности SB-18. Несмотря на это, значительные улучшения в выделении обычно наблюдались с обоими организмами.
Табл. 6 описывает результаты обработки осадков NALC/NaOH промывочным буфером с добавлением 2 мМ SB-18.
Таблица 6: Корреляция ПЦР и результатов высева при включении в промывной буфер SB-18
Результаты включения SB-18 в 2o-промывной буфер представлены, как описано в Материалах и Методах. Состав буфера, использованного для промывки осадка в этих сериях образцов, был 10 мМ NaHPO4 (pH 7,0), 45 мМ NaCl, 5 мМ DTT и 200 мкМ или 2 мМ SB-18. Осадок был удален перед промыванием и выращен в BACTEC 12B культуральных колбах. Все положительные высевы MTB, MAC и М. kansasii были идентифицированы с использованием методики Gen-Probe (GenProbe, San Diego, CA). Все другие микроорганизмы были идентифицированы биохимическим анализом (Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 71-157). Осадки были промыты в течение 60 мин при 37oC при встряхивании (140 об/мин). После промывки осадка образцы были подвергнуты центрифугированию и далее - ПЦР, как описано в Материалах и Методах.
Из 754 образцов, промытых буфером, содержащим SB-18, 23 образца были положительными при высеве для М. avium, 5 были положительными при высеве для М. tuberculosis и 1 была положительна при высеве для М. kansasii. Корреляция к высеву была следующей: М. avium, 65%: М.tuberculosis, 100% и М. kansasii, 100%. Так как реальное количество положительных при высеве материалов было низким, ясно, что наблюдалось различимое улучшение общей корреляции. Из восьми образцов М. avium, давших ложноотрицательные ответы при предварительном просмотре, разрешение разногласий показало, что 5 - из-за низкого числа копий (множественные амплификации были и положительные, и отрицательные), один, как было подтверждено, был результатом присутствия ингибиторов, и два были "необъясненными" (множественные амплификации все были отрицательными, но присутствие ингибиторов исключается). Дополнительно, начала проявляться корреляция с образцами М. gordonae, положительными при высеве. Тот факт, что положительные амплификации М. gordonae наблюдались in vitro только тогда, когда было представлено высокое число копий, рассматривался как дальнейшее свидетельство того, что общая эффективность выделения значительно улучшилась.
Когда способность изолировать М. avium и М.tuberculosis для экспериментальных целей из клинического материала и из пробирок со скошенным агаром была снова улучшена, появилось несколько наблюдений из работы in vitro, которые нуждаются в обсуждении. Во-первых, из-за разницы в вводе от эксперимента к эксперименту, было очень трудно получить значащие результаты. При вводе с очень низким числом копий наблюдались драматические ошибки с теми детергентами, которые не влияли на агрегирование MTB, интерпретация была невозможной. При вводе с высоким числом копий не удавалось наблюдать значащие различия, так как амплификация всех реакционных смесей проходила в линейной фазе. Требуется тщательное балансирование между этими двумя крайностями и оказывается трудно это исполнить. A priori, оценено, что требуется от 100 до 1000 копий, чтобы получить значащие результаты. Менее 100 копий образуют драматические ошибки в центрифугированных осадках, и более 1000 копий дают относительно гомогенные растворы, которые гарантированно содержат клетки, независимо от природы обработки раствора. Второе наблюдение относится к ожидаемым относительным различиям. Например, при использовании промывания простой водой или неионным детергентом, наблюдается 80% корреляция с высевом для MTB (табл. 5). Предположение состоит в том, что только краевые случайные улучшения (приблизительно 20%) требуются для достижения 100% корреляции. Для М. avium, с другой стороны, потребуется более чем двойное увеличение эффективности, чтобы достигнуть 100% корреляции. В свете первого наблюдения, отмеченного только что, эта точка зрения еще более усложняет накопление значимых данных. Несмотря на это, были получены значительные улучшения в описанных условиях и предполагается, что потери были значительно больше, чем первоначально предполагалось. "Необъясненные" результаты для MAC- микроорганизмов могут быть только улучшены призывом снизить количественные потери во время проведения анализа. Третье наблюдение состоит в том, что различия в способности изолировать М. avium и М.tuberculosis продолжаются: эти два организма, похоже, ведут себя по-разному, как in vivo, так и in vitro, с MTB, демонстрирующим гораздо более значительные улучшения в результате добавления SB-18 (фиг.5). Далее, факт, что MAC-микроорганизмы растут отдельными клетками, предполагает, что механизмы действия SB-18 функционируют не только благодаря дисперсии.
Попытки улучшить опыт сфокусировались на включении добавок. Так как смешивание детергентов кажется непродуктивным, и в соответствии с открытиями Linfield, W.M. et al., J.Am.Oil Chem.Soc. 40:114-117 (1963), включение аминокислот в буфер улучшает качество выделения умеренно, но ухудшает способность диспергировать MTB. Может быть, что аминокислоты просто активно изолируются, тем самым понижая естественную плавучесть микобактерий до некоторого уровня. Несмотря на это, все эмпирические данные предполагают, что какой-то вводимый материал все еще потерян.
Пример 9
Использование бетаинов в выделении микобактерий
SB-18 (CAS® N. 13177-41-8) и SB-16 (CAS® N. 2281-1-0) (хотя и в меньшей степени) были единственными детергентами, проверенными выше, на функционирование в опыте по агрегированию (фиг.4A показывает результаты применения SB-18). Далее, результаты, приведенные на фиг.5, предполагают, что механизм, по которому функционирует SB-18, не ограничивается только дисперсией. Таким образом, были предприняты серии экспериментов в попытке объяснить черты SB-18, которые способствуют улучшениям в определении (фиг. 6-9A).
Результаты, показанные на фиг. 6, основаны на модификации опыта, показанного на фиг.2 и используют несколько ионных гомологов SB-18: октадецилсульфат натрия (SOS: CAS® N. 1120-04-3), бромид триметилоктадециламмония (TMA-18: CAS® N. 1120-02-1) и 3-[(3- хлорамидопропил)-диметиламмоний]-1-пропан сульфонат (CHAPS: CAS® N. 75621-03-3). SOS и TMA-18 являются анионными и катионными октадецил детергентами и CHAPS цвиттерионная желчная соль с частью молекулы - (N,N-диметиламмоний) пропан сульфонат. Все промывные буферы содержали 10 мМ NaHPO4 pH 8,0 и 15 мМ NaCl, за исключением водного контроля. Для каждой серии были подготовлены пробирки в четырех повторностях и к ним было добавлено 2 мМ SB-18, или 200 мкМ SOS, или 2 мМ TMA-18 или 2 мМ CHAPS. Раствор SOS концентрацией 2 мМ образует белый осадок, ингибирующий ПЦР-реакцию. Поэтому был использован раствор 200 мкМ. Все пробирки инокулировали 20 мкл исходной бактериальной суспензии М. tuberculosis и инкубировали при 37oC 60 минут при встряхивании перед центрифугированием. Дублированные амплификации четырех повторностей представлены как a, b, c и d и должны быть идентичными. Дублированные амплификации прямых аликвот показаны на соответственно отмеченной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Результаты показывают, что SB-18 является единственным детергентом среди проверенных серий, функционирующим эффективно. Повторные эксперименты привели к следующим заключениям: (a) SOS, кроме того, что очень дорог, при использованных условиях выпадает в осадок, (b) хотя TMA-18 и CHAPS проявляются лучше, чем SOS или водный контроль, однако не дают сигнала, наблюдаемого обычно с SB-18.
Результаты, показанные в фиг. 7 и 7A, основаны на модификации опыта, показанного на фиг.2, и используют SB-серии детергентов: SB-12, SB-14, SB-16 и SB-18 как с Mycobacterium tuberculosis, так и с Mycobacterium avium, соответственно. Эти гомологи отличаются только длиной алкильной цепи. Все 2o-промывные буферы содержали 10 мМ NaHPO4 pH 8,0, 15 мМ NaCl и 5 мМ DTT, за исключением водного контроля. Для каждой серии были подготовлены пробирки в четырех повторностях и к ним были добавлены до конечной концентрации 2 мМ соответствующие детергенты. Все пробирки инокулировали или 20 мкл исходной бактериальной суспензии М. tuberculosis (фиг. 7), или 20 мкл исходной бактериальной суспензии М. avium (фиг.7A). Все пробирки инкубировали при 37oC 60 минут при встряхивании (140 об/мин) перед центрифугированием. Дублированные амплификации четырех повторностей для каждого детергента представлены как a, b, c и d и должны быть идентичными. Дублированные амплификации прямых аликвот показаны на соответственно помеченной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Как показано на фиг. 7, оказалось, что SB-18 и SB-16 дают результаты как одинаковой интенсивности, так и однородного распределения. С другой стороны, SB-14, показав исключительные надежды в улучшении выделения М. tuberculosis центрифугированием, продемонстрировал результаты, которые характерны для неоднородного распределения, вызванного слипанием клеток. SB-12, явно выступавший лучше, чем вода, вел себя не так хорошо, как гомологи с более высокой длиной алкильной цепи. В отношении результатов фиг. 7A, можно сказать, что все детергенты в этих сериях функционировали относительно одинаково в пределах улучшения выделения М. avium. Эти результаты были довольно типичными за исключением того, что при использовании М. tuberculosis, SB-16 в основном давал менее однородные результаты, чем SB-19.
После завершения экспериментов, показанных в фиг.3, стало ясно, что слипание клеток все еще является основной заботой. Как обсуждалось в Примере 7, линия проведенных исследований была нацелена на уменьшение этого феномена. Использование SB-18 в методах по данному изобретению
было оригинально открыто в процессе случайного поиска детергентов, и было идентифицировано на основании его способности диспергировать группы клеток М. tuberculosis. Факт, что все SB-серии детергентов улучшали выделение М. tuberculosis и М. avium, в сочетании с фактом, что М. avium растет отдельными клетками (т.е. не требуется дисперсии), предполагает, что детергенты SB-серии имеют что-то общее, позволяющее им функционировать в методах по данному изобретению. Иначе говоря: в дополнение к SB-гомологам с длинной цепью, способным диспергировать скопления клеток, все SB-18-подобные детергенты способны, некоторым еще неопределенным образом, улучшать выделение этого класса микроорганизмов. Таким образом, данные предполагают двойной механизм действия SB-18. Изобретатель показывает дальше, как эта двойная способность SB-18 распространяется дальше на другие молекулы.
SB-18-серии детергентов, также известные как сульфопропилбетаины, являются частью большого, широкого класса цвиттерионных детергентов, известных как бетаины. SB-18-серии цвиттерионных детергентов доступны благодаря торговле (напр. Aldrich, Milwaukee, WI; CalBiochem., La Jolla.CA; Fluka, Ronkonkoma, NY, Sigma, St. Lois). Существует большое число производителей гомологичных структур и четыре различных образца были сначала получены от трех различных производителей: Корпорация Henkel (Hoboken, NJ) предоставила "Velvetex AB-45" и "Velvetex BK-35"; Химическая Компания Inolex (Philadelphia, PA) предоставила "Lexaine® C" и химическая Корпорация Goldschmidt (Hopewell, VA) предоставила "TEGO® Betain L 5351". "Velvetex AB-45" (CAS® N. 68424-94-2) является коко-карбоксилметилбетаином (R1 - масло кокосового ореха, α =-CH2-, R2, R3 - -CH3 и R4 - -CH2-, бета= N⊕ и γ = -COO⊖: см. Таблицу 2 для структурных кодов.); "Velvetex BK-35" (CAS® N. 61789-40-0) и "TEGO® Betain L 5351" (CAS® N. "представляющий частную собственность"), являются все коко-амодопропил карбоксиметилбетаинами (α =-C(O)NHC3H6-). Кокосовое масло имеет сложный состав, однако использование этого масла в синтезе бетаинов приводит к тому, что R1 состоит из следующей смеси алкильных цепей: 45,4% лауриновой (C12), 18,0% имиристиновой (C14), 10,5% пальмитиновой (C16), 2,3% стеариновой (C18), 0,4% арахидиновой (C20), 7,4% олеиновой (C18:1) и 5,4% других (состав масла по CRC Handbook of Chemistry and Physics, 55th ed., CRC Press, Cleveland. OH (1974) стр. D-192-193).
Результаты, показанные на фиг.8, основаны на модификации опыта фиг.2 и используют коммерчески доступные препараты бетаинов, описанные выше: Velvetex AB-45, Velvetex BK-35, Lexaine® C и TEGO® Betain L 5351. Два мМ SB-18 были использованы в качестве положительного контроля, и вода была использована как отрицательный контроль. Все 2o-промывные буферы содержали 10 мМ NaHPO4, pH 8,0 и 10 мМ NaCl, за исключением водного контроля. Пробирки в четырех повторностях были приготовлены для каждой из серий и доведены до конечной концентрации 10 мМ соответствующим коко-бетаиновым детергентом. Все пробирки были инокулированы исходной бактериальной суспензией М. tuberculosis. Все пробирки инкубировали при 37oC 60 минут при встряхивании (140 об/мин) перед центрифугированием. Дублированные амплификации четырех повторностей для каждой серии детергентов представлены как a, b, c и d и должны быть идентичными. Дублированные амплификации прямых аликвот показаны на соответственно помеченной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Ясно, что эти коммерческие препараты показывают некоторый уровень "SB-18-подобной активности" в отношении улучшения способности выделения этих организмов центрифугированием. В соответствии с составом алькильной части молекулы и результатами, представленными в таблице 7, можно a priori ожидать, что эти препараты будут и действительно ведут себя промежуточным образом между SB-12, SB-14 и SB-16. Например, они улучшают выделение, но имеют ограниченную способность в отношении диспергирования этих организмов. Следует заметить, что так как три из реагентов имели идентичные структуры (Velvetex BK-35, Lexaine® C и TEGO® Betain L 5351), следовало ожидать, что они будут функционировать идентично, и так оно и было. (Заметим, что два из них имели разные CAS® Номера). Факт, что все три действительно функционировали с равной эффективностью, позволяет предположить, что действие независимо от производителя. Дополнительно отметим, что, несмотря на то, что препараты содержали примеси, они функционировали в этом опыте.
Дополнительные источники бетаин-подобных молекул проверены ниже. Происхождение этих молекул представлено так, чтобы результаты проверки широкого разнообразия гомологичных структур можно было лучше обсудить.
Происхождение бетаинов
Бетаины являются цвиттерионными детергентными молекулами, которые по существу являются центром положительного заряда, отделенного от центра отрицательного заряда. Алкил может быть связан или с катионом, или с анионом, или с мостиком, разделяющим заряды. Таблица 2 дает общую структуру наиболее распространенных классов бетаинов и обсуждение таблицы 2 дает множество примеров. Таблица 3 дает несколько структурных вариаций на тему бетаинов и обсуждение таблицы 3 дает несколько примеров этих подклассов. Катион, в подавляющем большинстве случаев, является четвертичным азотом. Анион демонстрирует значительно более высокую степень разнообразия. Например, аммонийные версии карбоксибетаинов, фосфобетаинов и сульфобетаинов коммерчески легко доступны.
В дополнение к упомянутым выше источникам, в издании 1994 "McCutcheon's" (McCutcheon's, Volume 1: Emulsifiers & Detergents, North American Edition, MC Publiching Glen Rock, NJ, стр. 290-291) перечислено 93 коммерчески доступных "производных бетаинов". Сегодня существует огромная масса научной информации, технической экспертизы и возможностей производств, имеющих отношение к бетаинам.
Представленные здесь результаты показывают, что комбинация структурных и поведенческих идиосинкразий улучшает действие бетаинов в методах настоящего изобретения. Обусловленное разнообразием и уникальным характером, проявляемыми этим классом цвиттерионных детергентов (табл. 2 и 3), большое разнообразие бетаинов, представляющее широкий спектр структурных вариантов, было проверено для того, чтобы определить способность этих гомологов улучшать выделение Mycobacterium tuberculosis, табл. 7 суммирует структуры, источники и CAS® номера бетаинов, использованных в фиг. 9-9D.
Исследование изменений в структуре бетаинов
Сравнение фиг. 6, 7, 7A и 8 и обсуждение структуры бетаинов показало, что нет уникальной комбинации сульфоната, четвертичного диметиламина и октадецильной цепи, которая способствует SB-18-подобной активности. С другой стороны обнаружилось, что анион отделен от катиона и оба ковалентно присоединены к одной и той же молекуле. Целью эксперимента, показанного на фиг.9, было исследование роли дипольного момента, образующегося в результате сосуществования противоположных зарядов, в SB-18-подобных молекулах. Результаты, представленные на фиг.9, основываются на модификации обработки результатов анализа, представленных на фиг.2, и включают различные бетаины с различными зарядами и структурами. Два соединения были получены из коммерческих источников, а три были синтезированы. SB-18 контроль (CAS® N. 13177-41-8) - C18-сульфопропилбетаин, приобретенный от Sigma, St. Louis, MO. SB-18 содержит четвертичный диметиламин с сульфонатной группой (-SO
Как показано на фиг.9, оказалось, что все тестируемые детергенты в этом анализе работают одинаково хорошо. Основываясь на представленных аргументах и на том факте, что функции обратимых бетаинов аналогичны SB-18, показано, что все бетаино-подобные молекулы проявляют SB-18-подобную активность. Основой для этой группы, как предполагается, является непосредственное сосуществование противоположных зарядов, в результате чего образуется электрически нейтральная молекула с большим дипольным моментом. Результат такой комбинации лучше всего представлен в обзоре Laughlin R.G.Langmuir 7:842-847 (1991): "Цвиттерионные функциональные группы, обладающие наибольшей полярностью, определенной для неионного класса гидрофильных групп". Тот факт, что различные ионные группы могут быть взаимозаменяемо применены в описанном здесь анализе, определяет степень полярности головной группы как характеристику указанных детергентов. Например, степень полярности, созданная полиоксиэтиленовой группой электрически нейтральной и имеющей примерно такие же молекулярные размеры, как, скажем аммонийпропансульфонатная группа SB-18, сравнительно незначительна. Представлены примеры для сравнения CMC и точек Краффта Brij 76 (CAS® N-9005-00-9) и SB-18 с октадецилсульфонатом натрия. Brij 76 (стеарил-этилен оксид (C18E10)) обладает CMC примерно 3•10-5 М (определено: Schott, H. et al. , J.Phar.Sci.64:6558-664 (1975) на основе олеилового эквивалента), точка Краффта - 45,5oC и точка помутнения 64oC (Schott. H. et al., J.Phar.Sci.65:979-981 (1976)). SB-18 обладает CMC примерно 3•10-6 (Nandakumar T. N. et al., J. Oil Tech. Assoc. India 11:31-34 (1979)), точка Краффта - 89oC в чистой воде и 37,5oC в 10 ммоль NaCl (Tsujii.K. et al. J.Phys.Chem. 82:1610-1614 (1978)). Критическая температура октадецилсульфоната натрия - 57oC при концентрации 7,5•10-4 (Tartar.H.V. et al., J.Am.Chem.Soc. 61:539-544 (1939)). SB-18 менее растворим (т.е. обладает самым низким CMC), но имеет самую низкую температуру Краффта в присутствии соли (т. е. самую низкую температуру плавления гидратированного кристалла). Ясно, что связь между CMC, температурой Краффта и полярностью является сложной, однако оказалось, что способность цвиттериона структурировать воду в присутствии электролитов является ключом действия. Поэтому с учетом перечисленных в табл. 2 и 3 катионов и анионов рассматривается большое количество ионов, которые могут объединиться, образовывая функциональные бетаины в методах данного изобретения.
Исследования бетаиновой мостиковой структуры
Результаты, представленные на фиг.9, предполагают, что важной характеристикой структуры бетаина является присутствие противоположных зарядов. Эти данные вместе с важностью бетаиновой мостиковой структуры также предполагают, что как только комбинация мостиковой структуры и алькильной цепи позволяет образовывать мицеллы в процессе анализа, бетаин будет проявлять SB-18-подобную активность. Рассмотрение структур на фиг.9 показывает, что Detain PB имеет метильный мостик, сульфатоэтилбетаин и фосфоэтилбетаин - этильный мостик, SB-18-пропильный мостик, а C16-AHTMAP - оксипропильный мостик. Результаты, представленные на фиг. 9A, показывают дополнительные мостиковые структуры и зарядные комбинации.
Они основаны на модификации обработки результатов анализа, представленных на фиг.2, и рассматривают различные бетаины, которые варьируются на основе мостиковой структуры отдельно от зарядов. Два соединения были получены из коммерческих источников, а три были синтезированы. Ammonix MO (CAS® N. 3332-27-2) - оксид C14-диметиламина (мостиком является N-связь: N⊕-O⊖ был получен как образец от Stepan Company, Northfield, IL. Следует отметить, что Ammonix MO может подразумевать другую вариацию структуры дипольного момента, обсуждаемую на фиг. 9. Darvan NS - c-алкилбетаин - см. табл. 3 (алкил присоединен к мостику) был получен как образец от R.T.Vanderbilt, Norwalk.CT. Производитель утверждает, что это соединение является смесью c-децил-бетаина (CAS® N. 96-55-9) и c-цетил-бетаина (CAS® N. 96-56-0). Пропорции каждого не представлены. Три синтеза были проведены по договору с Ecochem.Inc.of Chaska.MN (все химические характеристики были получены Ecochem.).
Первый синтетический бетаин, C18-карбоксиэтилбетаин (CAS® N.30612-73-8) содержит этильный мостик (R4 = -CH2CH2) и был синтезирован по методике Weers, J. G. et al., Langmuir 7:854-867 (1991). Второй синтетический бетаин C18-сульфобутилбетаин (CAS® N. 22313-73-1) содержит бутиловый мостик (R4 = -CH2CH2CH2-) и был синтезирован по методике Parris N. et al., J. Am. Oil Chem. Soc.53:97-100 (1976). Третий синтетический бетаин, C16-оксипропилсульфобетаин (CAS® N.7425-12-9) содержит оксипропильный мостик (R4 = -CH2CH(OH)CH2-) и был синтезирован по методике Parris N. et al., J. Am. Oil Chem. Soc. 53: 60-63 (1976). Два ммоль SB-18 применяли в качестве положительного контроля, а воду - в качестве отрицательного. Все буферы для промывания содержали 10 ммоль NaHPO4 (натрийфосфатный буфер, смесь NaH2PO4 и Na2HPO4), pH 8,0 и 15 ммоль NaCl, за исключением водного контроля. Четыре пробирки для репликаций были получены для каждой серии и были разбавлены подходящим детергентом до конечной концентрации 2 ммоль, за исключением Darvan NS, который применялся в концентрации 10 ммоль. Все пробирки были инокулированы 20 мкл исходной бактериальной суспензией М.Tuberculosis, а затем инкубированы при 37oC в течение 60 минут при перемешивании (140 об/мин) перед центрифугированием. Повторные амплификации четырех репликантов для каждой серии детергентов представлены как a, b, c и d, и могли быть идентичны. Повторные амплификации ряда аликвот показаны определенной линией. Повторные контроли 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий также были блоттированы.
За исключением C16гидрооксипропилсульфобетаина, все тестируемые соединения действовали в методах изобретения. А именно, все тестируемые соединения проявляли некоторую SB-18-подобную активность. Интересно, что Darvan NS в комбинации с C10 и C16 бетаинами действовал незначительно. Поскольку пропорции каждого неизвестны, Darvan NS экстремально разбавлен, даже больше, чем коко-производные, поэтому предполагается, что он содержит очень высокую концентрацию C10-производных. Невзирая на это, c-алкил проявлял некоторую степень SB-18-подобной активности. Дополнительные эксперименты с оксидом амина показали, что эта структура обладает отличной эффективностью, хотя постоянно наблюдалось образование комков. К сожалению, увеличение длины цепи в AO в клинических условиях не может быть допустимо. Например, температура Краффта для C18-AO составляет примерно 43oC и относилась к типу высаливания (Tsujii. K. et al., Yukagaku 30:495-499 (1981)). Кроме того, как предсказано на фиг. 9, оксиды амина дают другой пример структурной вариабельности дипольного момента. Как C18-карбоксиэтилбетаин, так и C18-сульфобутилбетаин, в основном, ведут себя аналогично SB-18: обработка различных экспериментов, применяющих эти два бетаина, обычно дают довольно единообразные результаты.
Неспособность C16-гидрооксипропилсульфобетаина способствовать выделению бактерий при центрифугировании может быть объяснена тем фактом, что детергент осаждается при добавлении к NaHPO4. На основании информации, касающейся взаимодействия бетаиновой мостиковой структуры с различными ионами, квалифицированный специалист может модифицировать систематические параметры и таким образом C16-оксипропилсульфобетаин может быть применен.
Например, если расстояние между зарядами 4 ≈ 5 ангстрем было минимальным расстоянием, позволяющим всаливание (пропильное расстояние будет составлять примерно 5,5 ангстрем (Tsujii K. et al. Yukagaku 30:495-499 (1981)), и способность к всаливанию описывается следующей тенденцией: SCN⊖ > I⊖ > NO
Исходя из этих данных, можно согласиться с Laughlin, R.G.Langmuir 7: 842-847 (1991) в том, что эффективность бетаинов большей частью более зависит от структуры зарядов, чем от мостиковой структуры. Другими словами, как только мостик способствует образованию мицелл при системных условиях, может наблюдаться сосуществование зарядов на той же молекуле, что способствует применению указанных детергентов в методах данного изобретения. Поэтому важность структуры мостика выразится, в первую очередь, в возможности продуктивного присутствия данного детергента в контексте условий анализа. Например, если мостики способствуют образованию мицелл при указанных в анализе температуре, электролите и его концентрации, то бетаин будет проявлять SB-18-подобную активность.
Исследование влияния алкила и алкильной связи на функцию бетаина
Внимательное исследование применяемых в эксперименте структур, представленных на фиг. 9 и 9A, показало, что в методах данного изобретения был применен комплексный ряд алкильных структур. Например, неразветвленные C10, C14, C16, C18 цепи углеводородов были основой гидрофобных доменов (R1) в этих примерах. Кроме того, в сульфатобетаине применяется амидопропильная связь для ковалентного связывания алкила с четвертичным азотом. Подразумевается, что бетаиновая функция меньше зависит от структуры алкила. На основании этого можно утверждать, что как только комбинация зарядов, мостиковой структуры и алкильной структуры способствует образованию мицелл в контексте данного анализа, бетаин будет проявлять SB-18-подобную активность. Для подтверждения этого замечания дальнейшими примерами было протестировано несколько дополнительных бетаинов. Результаты, представленные на фиг. 9B, основываются на модификации обработки результатов анализа, представленных на фиг. 2, и используют различные бетаины с различной алкильной структурой, строением и связью с четвертичным азотом. Все пять соединений были получены из коммерческих источников. Velvetex AB-45 (CAS® N.68424-94-2) применяет масло коксового ореха в качестве источника алкильной цепи (состав масла кокосового ореха дан ниже) и был получен как образец от Henkel Corporation, Emery Group Cospha, Hoboken, NJ. Алкильная связь Velvetex AB-45 является простым удлинением алкильной цепи. Миратаин ASC (CAS® N. 108797-84-8) содержит 3-бутокси-2-гидроксил в качестве группы, соединяющей алкил и четвертичный азот. Он был получен как образец от Rhone-Poulenc, Surfactant & Specialty Division, Cranberry, NJ. В информации на его CAS® N его структура обозначена, как "1-пропанаминий, 3-бутокси-2-окси-(2-окси-3- сульфопропил)-диметил-, внутренняя соль". Плотность этой жидкости предполагает, что алкил, приблизительно додецил, присоединен к бутоксилу, хотя точная структура неизвестна. Schercotaine IAB (CAS® N.6179-44-8) содержит амидопропил, соединяющий алкил с четвертичным азотом, и был получен как образец от Scher Chemical, Inc. , Clifton, NJ. Производитель утверждает, что алкил является изостеарилом, отмечая, что молекула может быть разветвлена, не указывая каким образом. В информации на его CAS® N, однако, обозначено, что алкил является октадецилом. В Velvetex OLB-50 (CAS® N.871-37-4) применяется олеил (C18:1) в качестве алкильной цепи (т.е. вводится двойная связь) и был образец получен от Henkel Corporation, Emery Group Cospha, Hoboken, NJ. Incronam B-40 содержит необычно длинную алкильную цепь (C22) и был получен как образец от Croda, Inc. , Parsippany, NJ. Для него не был дан CAS® N, однако, аналогичный продукт был продан из Соединенных Штатов тем же производителем с тем же торговым знаком, обозначающим его активный ингредиент как бетанамид. Его CAS® N был 84082-44-0.
В информации на его CAS®N обозначено, что его алкильный состав является смесью C8-C22. Оба были проданы как пасты, отмечая преобладание исключительно длинноцепочечных алкилов. Основной раствор детергента Incronam B-40 может быть получен при растворении порции пасты в смеси изоамиловый спирт: вода в соотношении 10:1. 2 ммоль SB-18 были положительным контролем, а вода - отрицательным. Все буферы для промывания содержали 10 ммоль NaHPO4, pH 8,0 и 15 ммоль NaCl, за исключением водного контроля. Четыре пробирки для репликаций были получены для каждой серии и были разбавлены подходящим детергентом до конечной концентрации 2 ммоль, за исключением Incronam B-40, который применялся в концентрации 200 мкмоль. Все пробирки были инокулированы 20 мкл исходной бактериальной суспензией М.tuberculosis, а затем инкубированы при 37oC в течение 60 минут при перемешивании (140 об/мин) перед центрифугированием. Повторные амплификации четырех репликантов для каждой серии детергентов представлены как a, b, c и d, и должны быть идентичны. Повторные амплификации ряда аликвот показаны определенной линией. Повторные контроли 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108 109 и 1010 копий также были блоттированы.
Опять же, все указанные бетаины, за исключением Incronam B-40, проявили SB-18-подобную активность. Velvetex AB-45 дал немного более единообразные результаты в этих экспериментах, чем представленные на фиг. 8. Миратаин ASC, содержащий цепь неизвестной длины, был экстремально разбавлен и проявил неспособность диспергировать микобактерии. Эти результаты, происходящие от слипания в комочки, были типичными, как и для бетаинов с более короткой цепью. Schercotaine IAB и Velvetex OLB-50 являются октадецильными детергентами, первый содержит насыщенную цепь, а последний - ненасыщенную. Оба указанных бетаина дали результаты, которые обычно получаются с SB-18: обработка указанными бетаинами давала довольно единообразные результаты, хотя наблюдалось немного больше вариаций с неочищенными образцами. Очевиден вывод, что как только алкил способствует образованию мицелл при системных условиях, будет наблюдаться сосуществование зарядов в одной и той же молекуле, что способствует применению указанных детергентов в методах данного изобретения. Incronam B-40, аналогично C16- оксипропилсульфобетаину (фиг.9A), осаждается при добавлении к NaHPO4, и поэтому не ожидается, что он проявит ту же степень эффективности. Основываясь на информации, касающейся взаимодействия бетаиновой мостиковой структуры с различными электролитами, следует отметить, что Incronam B-40 представляет более сложную проблему, чем C16-оксипропилсульфобетаин (фиг. 9A). Например, если расстояние между зарядами 4 - 5 ангстрем было минимальным расстоянием, позволяющим высаливание (метильное расстояние будет составлять примерно 3,1 ангстрем (Tsujii K. et al. Yukagaku 30: 495-499 (1981)), и способность к высаливанию описывается следующей тенденцией: SCN⊖ > I⊖ > NO
Бетаины, полученные из природных масел
Подавляющее большинство коммерческих бетаинов получены из натуральных масел, главным образом из масла коксового ореха. Исходя из результатов фиг. 8, 9, 9A и 9B, можно сказать, что эти бетаины также должны действовать в методах данного изобретения. Представленные на фиг. 9C и 9D результаты получены при использовании несколько различных бетаинов, алкилы которых получены исключительно из природных масел. Эти структурные отличия основываются на длине, строении алкила, его смеси, связи с четвертичным азотом, мостиковых структурах и анионном заместителе. В сущности, указанные бетаины дают уникальную возможность дальнейшего исследования связи между различными аспектами строения бетаинов. Каждый эксперимент сначала представлен, затем обсуждается на обоих фиг.
Результаты, представленные на фиг.9C, основаны на модификации обработки результатов анализа, представленных на фиг.2, и касаются различных бетаинов, которые получены исключительно из природных масел. Все пять соединений были получены из коммерческих источников. TEGO Бетаин L5351 (CAS® N. "Собственность") - карбоксиметилбетаин, который использует масло коксового ореха в качестве источника алкильной цепи в комбинации с амидопропильной связью. TEGO Бетаин L5351 был получен как образец от Henkel Corporation, Emery Group Cospha, Hoboken, NJ. Масло кокосового ореха, выделенное из Cocos nucifera, является сложной смесью жирных кислот следующего состава: 45,4% лауриновой (C12), 18,0% миристиновой (C14), 10,5% пальмитиновой (C16), 2,3% стеариновой (C18), 0,4% арахидоновой (C20), 7,4% олеиновой (C18:1) и 5,4% других кислот (состав масла взят из CRC Handbook of Chemistry and Physics, 55th ed. CRC Press, Cleveland, OH (1974) pp.D-192-193). Crosultaine C-50 (CAS® N.68139-30-0) - сульфобетаин, который использует оксипропильный мостик и масло коксового ореха в качестве источника алкильной цепи в комбинации с амидопропильной связью. Crosultaine C-50 получен как образец от Croda, Inc., Parsippany, NJ. Incronam BA-30 (CAS® N не обозначен) - карбоксиметилбетаин, который применяет масло ореха пальмы как основу гидрофобного домена, в комбинации с амидопропильной связью. Он был получен как образец от Croda, Inc. , Parsippany, NJ. Масло пальмового ореха, выделенное из Attalea funifera, является сложной смесью жирных кислот следующего состава: 44,1% лауриновой (C12), 15,4% миристиновой (C14), 8,5% пальмитиновой (C16), 2,7% стеариновой (C18), 0,2% арахидоновой (C20), 16,1% олеиновой (C18:1), 1,4% линолевой (C18:2) и 11,6% других (состав масла взят из CRC Handbook of Chemistry and Physics, 55th ed. CRC Press, Cleveland, OH (1974) pp.D-192-193). Rewoteric AM-R40 (CAS® N.71850-81-2) - карбоксиметилбетаин, который применяет касторовое масло как источник алкильной цепи в комбинации с амидопропильной связью. Он был получен как образец от Sherex Chemical Company, Dublin, OH. Касторовое масло, выделенное из Ricinus communis, является сложной смесью жирных кислот, но содержит главным образом рицинолевую (87%), олеиновую (7,4%) и линолевую (3,1%) кислоты (состав масла взят из CRC Handbook of Chemistry and Physics, 55th ed. CRC Press, Cleveland, OH (1974) pp.D-192-193). Рицинолевая кислота или 12-окси-9-октадеценовая кислота (цис) содержит ненасыщенную C18-цепь и представлена формулой: C6H13CH(OH)CH2CH= CH(CH2)7COOH. Линолевая кислота или 9,12-октадекадиеновая кислота также содержит ненасыщенную C18-цепь и представлена формулой: CH3(CH2)4CH=CHCH2CH= CH(CH2)7COOH.
Schercotain WOAB (CAS® N. не обозначен) предположительно является карбоксиметилбетаином, который использует масло зародышей пшеницы в качестве источника алкильной цепи в сочетании с амидопропиловой связью. Schercotaine WOAB был получен в качестве образца из Scher Chemicals, Inc., Clifton, NJ. Масло зародышей пшеницы из Triticum aesticum является сложной смесью жирных кислот, но преимущественно состоит из линолевой кислоты (52,3%), олеиновой кислоты (28,1%) и линоленовой кислоты (3,6%). Баланс приблизительно 16% составляет смесь лауриновой, миристиновой, пальмитиновой, стеариновой и арахидиновой кислот (состав масла по CRC Handbook of Chemistry and Physics, 55th ed., CRC Press, Cleveland. OH (1974) стр. D-192-193). Линоленовая кислота или 9,12,15-октадекантриеноевая кислота (цис,цис,цис), также как ненасыщенная C18-цепь, имеет структуру: CH3[CH2CH==CH]3(CH2)7COOH. 2 мМ SB-18 были использованы в качестве положительного контроля, и вода была использована в качестве отрицательного контроля. Все промывные буферы содержали 10 мМ NaHPO4, pH 8,0 и 15 мМ NaCl, за исключением водного контроля. Для каждой серии были приготовлены четыре повторности пробирок. Incronam BA-30, Rewoteric AM-R40 и Schercotain WOAB были добавлены к соответственному промывному буферу до конечной концентрации 2 мМ, и TEGO Betain L 5351 и Crosultaine C-50 были приведены к конечной концентрации 10 мМ. Ни один из этих детергентов не преципитировал при добавлении натрий фосфатного буфера. Все пробирки инокулировали 20 мкл бактериальной исходной суспензии М. tuberculosis. Все пробирки инкубировали при 37oC 60 минут при встряхивании (140 об/мин) перед центрифугированием. Дублированные амплификации четырех повторностей для каждой серии детергентов представлены как a, b, c и d и должны быть идентичными. Двойные повторности амплификаций прямых аликвот показаны на соответственно отмеченной линии. Повторные контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Результаты, показанные на фиг. 9D, основаны на модификации метода обработки, показанного на фиг. 2, и используют несколько бетаинов, которые также происходят исключительно из натуральных масел. Различие этих структур основано на длине алкила, структуре алкила, смесях алкилов, связью с четвертичным азотом, структурами мостика и анионной частью молекулы. Все пять молекул получены из коммерческих источников. Chembetaine S (CAS® N. не приведен) является карбоксиметилбетаином, который использует соевое масло в качестве источника алкильной цепи в сочетании с амидопропиловой связью. Chembetaine S был получен в качестве образца из Chemron Corporation, Paso Robles, CA. Соевое масло, полученное из Glycine soja, является сложной смесью жирных кислот, но оно преимущественно состоит из линолевой кислоты (50,7%), олеиновой кислоты (28,9%) и пальмитиновой кислоты (9,8%). Дополнительные компоненты включают: линоленовую (6,5%), стеариновую (2,4%) и арахидиновую (0,9%) кислоту (состав масла по CRC Handbook of Chemistry and Physics, 55th ed. , CRC Press, Cleveland. OH (1974) стр. D-192- 193). Hetaine CLA (CAS®N. не приведен) - это бетаин неизвестной структуры, который использует canola масло как источник алкильной цепи, в сочетании с амидопропиловой связью. Hetaine CLA был получен как образец из Heterene, Inc., Paterson, NJ. Состав "масла canola" наверняка не известен. Crosultaine T-30 (CAS® N. не приведен) является сульфобетаином, который имеет гидроксипропиловый мостик и сало как источник для алкильной цепи в сочетании с амидопропиловой связью. Crosultaine T-30 был получен как образец от Croda, Ic., Parsippany, NJ. Сало обычно происходит из говядины (Bovis taurus) или баранины (Ovis aries). Сало, происходящее из говядины, имеет следующий состав: 49,6% олеиновой кислоты, 27,4% пальмитиновой кислоты, 14,1% стеариновой кислоты, 6,3% миристиновой кислоты и 2,5% линолевой кислоты. Сало, происходящее из баранины, имеет следующий состав: 36,0% олеиновой кислоты, 24,6% пальмитиновой кислоты, 30,5% стеариновой кислоты, 4,6% миристиновой кислоты и 4,3% линолевой кислоты (состав масла по CRC Handbook of Chemistry and Physics, 55th ed., CRC Press, Cleveland. OH (1974) стр. D-192-193). Источник сала, использованный производителем Crosultaine T-30, не известен. Rewoteric TEG (CAS® N. 70750-46-8) это бис-гидроксиэтилглиценат, имеющий сало как источник для алкильной цепи. Rewoteric TEG был получен как образец от Sherex Chemical Company, Dublin, OH. Источник сала, использованный для производства Rewoteric TEG, не известен. Crosultaine E-30 (CAS® N. не приведен) является сульфобетаином, который имеет гидроксипропиловый мостик и рапсовое масло как источник для алкильной цепи в сочетании с амидопропиловой связью. Crosultaine E-30 был получен как образец от Croda, Ic., Parsippany, NJ. Рапсовое масло, полученное из Brassica campestris, является сложной смесью жирных кислот, но оно состоит преимущественно из эруковой кислоты (50%), олеиновой (32%) и линолевой кислоты (15%) (состав масла по CRC Handbook of Chemistry and Physics, 55th ed. , CRC Press, Cleveland. OH (1974) стр. D-192-193). Эруковая кислота, или цис-13-докосеноевая кислота, имеет ненасыщенную C20 цепь со структурой: CH3(CH2)7CH= =CH(CH2)11COOH.
Сначала Hetaine CLA, Crosultaine T-30 и Crosultaine E-30 все преципитировали при добавлении NaHPO4 (конечная концентрация 2 мМ детергента). Таким образом, условия были изменены и вместо этого был использован 50 мМ Трис-HCl pH 8,0. Два мМ SB-18 было использовано в качестве положительного контроля (в обычном буфере 10 мМ NaHPO4, pH 8,0, 15 мМ NaCl) и вода была использована в качестве отрицательного контроля. Детергентные промывные буферы содержали 50 мМ Трис-HCl pH 8,0, за исключением водного контроля. Для каждой серии были приготовлены четыре повторности пробирок. Chembetaine S и Rewoteric TEG были добавлены к соответствующим промывным буферам до конечной концентрации 2 мМ, и Hetaine CLA, Crosultaine T-30 и Crosultaine E-30 были приведены к конечной концентрации 200 мкМ. В Трис-буфере Hetaine CLA и Crosultaine E-30 оказались мутными, тогда как Crosultaine T-30 оставался прозрачным. Crosultaine T-30, однако, преципитировал перед окончательным центрифугированием в одном из экспериментов (не показан). Все пробирки инокулировали 20 мкл бактериального препарата М. tuberculosis. Все пробирки инкубировали при 37oC 60 минут при встряхивании (140 об/мин) перед центрифугированием. Дуплицированные амплификации четырех повторностей для каждой серии детергентов представлены как a, b, c и d и должны быть идентичными. Двойные повторности амплификаций прямых аликвот показаны на соответственно отмеченной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Бетаины, представленные на фиг. 9C и 9D, были отобраны для того, чтобы проверить влияние смесей алкильных частей молекул, сложных алкильных структур, различных комбинаций зарядов и различных связей в отношении эффективности в методах данного изобретения. Как заключает фиг.9B, модификации алкильной структуры имеют очень небольшое влияние в отношении методов данного изобретения, так как эти структуры образуют мицеллы при параметрах, используемых системой. Другими словами, если комбинация зарядов, структуры мостиковой группы и алкила такова, что образование мицелл возможно, тогда бетаины будут повышать сбор микобактерий центрифугированием. Сало, кокосовое, бабассу, соевое, рапсовое масла и масло зачатков пшеницы дают широкий спектр сложных смесей насыщенных и ненасыщенных алькильных цепей и касторовое масло используется как гидроксилированный алкил. Исключая Rewoteric TEG, гидрофобные части молекул связаны с четвертичным азотом через амидопропиловую группу и в качестве группы, эти структуры используются в сочетании как с карбоксиметил-, так и гидроксипропилсульфо - частью молекулы. На основании информации, касающейся влияния структуры мостиков на поведение бетаинов, можно сказать, что метиленовые мостики и гидроксипропиловые мостики уменьшают способность к использованию более длинных алкильных цепей в описанном здесь опыте. Эта уменьшенная растворимость, однако, преодолевается в некоторой степени использованием амидопропиловой связи.
Как TEGO Betain L 5351, так и Crosultaine C-50 показывали более однородные результаты в этом эксперименте, чем обычно наблюдалось (фиг. 9C). Incronam BA-30, другое масло, состоящее преимущественно из коротких цепей, давало результаты амплификации, более характерные для слипания групп клеток, но все еще демонстрировало SB-18-подобную активность в отношении улучшения сбора в общем (фиг. 9C). Результаты, показанные здесь с Rewoteric TEG и Schercotain WOAB, были весьма типичными: здесь также наблюдался результат амплификации, вызванный слипанием (фиг.9C). Другой детергент, который был первично октадециловым и который обычно показывал более сильное слипание, чем ожидалось, был Chembetaine S. A priori можно ожидать, что октадециловые детергенты будут вести себя похоже. Например, SB-18, C18- карбоксиэтилбетаин и C18-сульфобутилбетаин (фиг. 9F), Velvetex OLB-50 и Schercotain IAB (фиг. 9B), Rewoteric AM-R40 и Schercotain WOAB (фиг.9C), и Chembetaine S, Crosultaine T-30, Rewoteric TEG и Crosultaine E-30 (фиг.9D), все состояли главным образом из насыщенных и ненасыщенных октадецил-подобных цепей. Хотя они действуют лучше, чем более короткие цепи в повышении эффективности собирания материала, они отличаются друг от друга в облегчении группирования бактерий. Возможно, возрастание сложности детергентов, при использовании смесей цепей различной длины, или включении двойных связей или других функциональных групп, таких как гидроксильные группы, уменьшает способность растворять миколовые кислоты и липиды, ассоциированные со связыванием. С другой стороны, многие из этих коммерческих препаратов неочищены и, возможно, этот фактор влияет на общую действенность.
Возможно, наиболее интересными из комбинаций, результаты испытаний которых приведены на фиг. 9C и 9D, были те, которые представлены Hetaine CLA, Crosultaine T-30 и Crosultaine E-30. Если под "маслом canola" производитель Hetaine CLA подразумевает кукурузное масло, то кукурузное масло, происходящее из Zea mays, является сложной смесью жирных кислот, но состоит преимущественно из олеиновой (49,6%), линоловой (34,3%) и пальмитиновой (10,2%) кислот (состав масла по CRC Handbook of Chemistry and Physics, 55th ed., CRC Press, Cleveland. OH (1974) стр. D-192-193). Факт, что Velvetex OLB-50 (олеилкарбоксиметилбетаин) функционировал без проблем, предполагает, что т. к. смесь алкилов Hetaine CLA очень вязкая, то она представляет собой что-то другое и возможно включает группы, которые подчеркивают неионный характер (т.е. гидроксипропилсульфобетаин). Факт, что Crosultaine T-30 имеет небольшие проблемы с растворимостью, тогда как Rewoteric TEG функционирует без инцидентов (т. е. они оба используют сало), подтверждает природу анионной растворимости обоих (-COO⊖ против -SO
Наиболее интересный аспект фиг.9D то, что растворы казались мутными (т. е. наблюдалась точка мутности) как с Hetaine CLA, так и с Crosultaine E-30. В противоположность C16- гидроксипропилсульфобетаину (фиг.9A) оба этих бетаина функционировали в методах данного изобретения. Такие результаты не парадоксальны. Напротив, считается, что момент преципитации вызывается дегидрированием гидрофильной части неионного детергента, и поэтому точка мутности, в отличие от температуры Краффта, не является хорошо определяемой температурой, мицеллы могут существовать, когда наблюдается мутность. В основном, в начале помутнения мицеллярный вес и число агрегирования постепенно возрастает с возрастанием температуры, и если температура растет далее, в конечном счете наступает фазовое разделение. (Nakagawa.T. et al., In: Colloidal Surfactants, Academic Press, New York (1963) стр. 121-135). Таким образом, так как Hetaine CLA и Crosultaine E-30 не доходят до фазового разделения, можно ожидать, что они будут функционировать в опыте по обработке.
Nilsson, P. et al., J.Phys.Chem. 88:6357-6362 (1984) наблюдал точки преципитации в сульфобетаинах и Faulkner, P. G. et al., Langmuir 5:924-926 (1989) описывал изменения, которые имеют место при гидроксилировании мостика у сульфобетаинов. Интересным аспектом этих результатов является то, что один бетаин, C16-гидроксипропилсульфобетаин, выступал как ионный детергент и был растворен в фосфатном буфере: тогда как другой, но очень близко родственный бетаин, Crosultaine E-30 (C20:1-, C18:1- и C18:2-амидопропил гидроксипропилсульфобетаин), вел себя как неионный детергент и стал мутным (сточки зрения этого обсуждения, структура Hetaine CLA неизвестна.) Дополнительно предлагается несколько очень интересных аспектов структуры бетаина, касающихся взаимодействия амидопропиловой связи, гидроксипропилового мостика и длины алкильной цепи, эти наблюдения предполагают получить возможность иметь предсказуемые результаты при добавлении электролитов. Например, растворимость C16-гидроксипропилсульфобетаина следует улучшать с помощью другого иона (напр. иодид калия (KJ) вместо NaHPO4), благодаря эффекту повышения точки Краффта (Tsujii, K. et al., J.Phys. Chem. 82:1610-1614 (1978); Tsujii, K. et al. , Yukagaki 30:495-499 (1981)), тогда как можно ожидать, что добавление того же самого иона усилит точку преципитации Hetaine CLA и Crosultaine E-30 (Schott, H. et al., J.Pharm.Sci. 64:658-664 (1975); Schott, H. et al., J. Pharm. Sci. 65: 979-981 (1976)). Собственно говоря, как предсказано, добавление 100 мМ KJ вызвало очищение раствора C16- гидроксипропилсульфобетаина, но вызвало ухудшение в мутности Hetaine CLA и Crosultaine E-30. Таким образом, использование двух последних потребует минимизации концентрации солей. Кажется, что функционирование амидопропила усиливает неионный характер. Например, амидопропиловая часть молекулы увеличивает общую головную группу путем увеличения гидрофильной части детергента, аналогично возрастанию числа полиоксиэтиленовых групп, как обсуждалось Hsiao, I. et al., J.Phys.Chem. 60: 657-660 (1956).
Одной из главных проблем использования коммерческих образцов, представленных здесь, является их относительная чистота. Например, SB-18 (Sigma, St. Lois, MO) предложен в качестве очищенной внутренней соли, которая, как утверждается, 98% чистоты. Incronam B-40, Hetaine CLA, Crosultaine T-30 и Crosultaine E-30 все предложены в качестве грубых смесей продуктов реакции. Процент активного материала, заявленный производителем: 44-48%, 38-42%, 30-34%, соответственно. Дополнительно, содержание соли (NaCl) в каждом заявлено: 4,2-4,6%, 5%, 2,4-4,0% и 2,5-4,0%, соответственно. Попытка образовать 2 мМ раствор любого из этих детергентов будет также образовывать раствор 3-4 мМ NaCl, что в некоторых случаях создаст раствор, который невозможно применить.
Фиг. 10 проверяет различные параметры опыта, модифицированные для улучшения применения C16-гидроксипропилсульфобетаина, Incronam B-40, Hetaine CLA, Crosultaine T-30 и Crosultaine E-30. Условия опыта были модифицированы следующим образом:
C16-гидроксипропилсульфобетаин- и Crosultaine T-30-буфер состоял из 10 мМ Трис-HCl pH 8,0 и 50 мМ KJ. Incronam B-40-, Hetaine CLA- и Crosultaine E-30- буфер был просто 200 мкМ Трис-HCl pH 8,0. Каждый детергент имел незамутненный раствор при 200 мкМ. Каждый буфер был также доведен до 37oC. Контроль SB-18 использовал 10 мМ Трис-HCl pH 8,0 и 50 мМ KJ буфер. Для каждой серии были приготовлены четыре повторности пробирок. Все пробирки инокулировали 20 мкл бактериального препарата М. tuberculosis. Все пробирки инкубировали при 37oC 60 минут при встряхивании (140 об/мин) перед центрифугированием. Дуплицированные амплификации четырех повторностей для каждой серии детергентов представлены как a, b, c и d и должны быть идентичными. Двойные повторности амплификаций прямых аликвот показаны на соответственно отмеченной линии. Контроли копий 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также блоттированы.
Хотя Hetaine CLA, Crosultaine T-30 и Crosultaine E-30, как было показано, функционировали в оригинальных условиях (фиг.9D), Incronam B-40 и C16-гидроксипропилсульфобетаин были несколько хуже с точки зрения функционирования. Этот эксперимент показал, что основываясь на физической природе бетаинов, могут быть сделаны предсказуемые модификации условий опыта, для предотвращения применения соединения, которое не будет функционировать. Дополнительно этот эксперимент показал важность функционального проявления детергента (т.е. в мицеллярной фазе).
Заключение
Хотя алкильные цепи C12-C16 позволяют улучшить выделение всех микобактерий, C18-C22 алкильные цепи также способствуют разрушению связывания микобактерий. Диметил, четвертичный азот, в комбинации с карбоксильным анионом (-COO⊖) дают наиболее высокий уровень растворимости экономически эффективным образом. Наиболее предпочтительная структура мостиковой группы это прямая алкильная цепь (т.е. без модификаций), с минимум тремя атомами углерода (т. е. пропиловый мостик). Хотя влияние амидопропиловой связи трудно распознать, для простоты, примем, что связь будет нуждаться только в расширении гидрофобной части. C16-карбоксипропилбетаин (CAS® N. 78195-27-4) сочетает оптимальные характеристики растворимости, структуры мостиковой группы и ионного характера, оставаясь относительно простым для синтеза. (JP Appl. N. 79100920[54-100920]; JP 8125139).
Для C16-карбоксипропилбетаина (CAS® N. 78195-27-4), использованного здесь, для приготовления соединения была использована следующая процедура (обсужденная с Lafayette D. Foland в Clorox Technical Center, Pleasanton, CA):
(1) Добавьте 1 эквивалент (%,1 моль или 29,7 г) N,N-диметилоктадециламина (297 г/моль: CAS® N. 124-28-7) к 1,1 эквивалентов (0,11 молей или 19.9 г) этиловому эфиру броммасляной кислоты (195 г/моль: CAS® N.2969-81-5) под аргоном для образования точного раствора.
(2) Поместите флакон в масляную баню при 50-60oC и помешивайте раствор (под аргоном) в течение ночи. Образуются крупные белые куски. [Отметим: это очень экзотермическая реакция. При проведении это реакции важно следить за температурой, чтобы промежуточные продукты не сгорели].
(3) Добавьте 600 мл изопропилового спирта прямо в продукты реакции и немедленно добавьте во флакон 150 г активированной анионообменной смолы AG 1-XB (Bio-Rad N. 140-1422) или аналогичную анионообменную смолу, такую как Amberlite IRA-400OH (Sigma IRA 440C) с гидроксидом-противоионом и перемешивайте суспензию в течение ночи. [Отметим: активируйте (или реактивируйте) анионообменную смолу смешиванием 150 г смолы с 1 л 5% NaOH. Перед использованием промойте смолу на колонке 5 литрами дистиллированной воды. Количество смолы может значительно уменьшиться и это может быть определено рутинными методами.]
(4) Удалите анионообменную смолу фильтрацией
(5) Удалите растворитель с использованием Rotovap и поместите окончательную твердую фракцию на ночь в линию с высоким вакуумом.
(6) Рекристаллизуйте материал дважды следующим образом:
- Поместите высушенный бетаин в 400 мл эфира и доведите до кипения. Титруйте метанол во флакон до момента, когда материал растворится. Остудите флакон до комнатной температуры (белые кристаллы начнут формироваться почти немедленно). Поместите флакон при 0-4oC на ночь. Удалите кристаллизованный материал фильтрацией и поместите в линию с высоким вакуумом на ночь. [Отметим: Материал освобождается из анионообменной смолы довольно чистым, и если 95% чистоты приемлемы, тогда будет необходима только одна рекристаллизация (или, возможно, без рекристаллизации). Заметим также, что двухпротонный, неполярный растворитель может быть заменой эфира.
Эта процедура дает приблизительно 20 г очищенного C18-карбоксипропилбетаина (приблизительно 50% выход). N,N- диметилоктадециламин доступен из AZKO, Inc. (N. Armeen DM 18D), American Tokyo Kasei, lnc. (N. D1609) и Pfaltz & Bauer, Inc. (N. D42680) и Fluka (N. 16540). Заметим, что этиловый эфир бромомасляной кислоты может быть использован вместо метилового эфира иодомасляной кислоты, использованного в патенте Kazuo (JP 8125139); аналогично, Amberlite IRA-400OH анионообменная смола от Sigma может быть использована вместо AG 1-XB от Bio-Rad. Промежуточный продукт, имеющий 412 г/моль, будет иметь 2,4 ммоль/г. Amberlite-смола имеет емкость 4 meq/г согласно каталогу Sigma. Для уменьшения концентрации соли единственной рекристаллизации вполне достаточно.
Заметим, что C18-карбоксипропилбетаин (CAS® N.78195-27-4) чрезвычайно бактериостатичен и концентрации этого соединения больше 2 мМ в опытах по данному изобретению могут привести к нежелательному лизису бактерий.
Важными аспектами условий опыта, таким образом, являются температура, pH, ионная сила раствора и специфические электролиты. Например, pH выше 5,0, но ниже 9,0 будет абсолютно необходим для многих бетаинов для поддержания цвиттериона. Температура процесса должна поддерживаться выше температуры Краффта для специфических SB-18-подобных детергентов. Например, температура опыта для SB-18 должна держаться 37oC и выше. (Tsujii, K. et al., J.Phys. Chem. 82:1610-1614 (1978), тогда как температура опыта для C18-карбоксипропилбетаина (CAS® N. 78195-27-4) должна поддерживаться только 25oC (Tsujii, K. et al., Yukagaki 30:495-499 (1981)). Ионная сила зависит от использованого иона. Например, для того, чтобы уменьшить засоленность хлоридом натрия, требуется более высокая концентрация, чем при применении такой соли, как иодид калия.
Когда обсуждается выбор ПЦР-амплификации в качестве метода определения, хлорид калия, хотя и не идеальный, удовлетворителен и дает необходимый эффект при концентрациях, полезных относительно условий амплификации.
Результаты показывают, что возможно логическое комбинирование структуры бетаина и выбор замысла опыта, и могут быть оптимизированы по желанию специалиста. SB-18 является наиболее эффективным бетаином, который в настоящее время легко доступен с коммерческой точки зрения в очищенной форме, хотя, как отмечалось раньше, бетаины, такие как C18-карбоксипропилбетаин (CAS® N. 78195-27-4) легко синтезируются в лаборатории или путем синтеза по заказу коммерческим поставщиком.
Собранные данные фиг. 6, 7, 8, 9 до 9D и фиг. 10 показывают: (1) что SB-18 уникален среди перечисленных гомологичных ионных детергентов в том, что он улучшает дисперсию микобактерий и сбор их центрифугированием (фиг.6); (2) что все сульфопропилбетаиновые серии имеют некоторый уровень эффективности в улучшении сбора обоих классов микобактерий (фиг.7 и 7A); (3) что большое разнообразие гомологичных "бетаиноподобных" детергентов проявляют "SB-18-подобную активность" в методах по данному изобретению, и что эти характеристики соответствуют предсказанному поведению (фиг.8, 9, 9A, 9B, 9C, 9D); и (4) что изменение параметров системы, основанное на понимании поведения этого класса молекул, может применяться для того, чтобы позволить бетаинам, которые не будут иным образом функционировать в методах данного изобретения, проявлять SB-18-подобную активность (фиг. 10).
В заключение, полярность дипольного момента, созданного разделением зарядов, вызывает функциональное применение молекулы детергента в клиническом опыте. Более важно, в отношении дисперсии связанных микобактерий, то, что созданный дипольный момент позволяет более длинным цепям алкильных частей молекулы быть использованными в водных, содержащих электролиты растворах. Последствием этого является способность использовать детергенты, которые имеют более высокие способности к растворению при таких условиях реакции, которые обычны для клинической лаборатории. Однако, как было прежде замечено, способность диспергировать эти организмы не объясняет полностью способность молекул с более короткими цепями улучшать сбор организмов, которые не образуют групп клеток. Другая половина ответа будет лучше понята из следующего Примера.
Пример 10
Дегазирование М. tuberculosis и М. avium
Спорно, что гидрофобная природа клеточной стенки этого класса микроорганизмов впрямую ответственна за пленочный рост. Многочисленные публикации ссылаются на Dubos, R.J. Exp.Biol.Med. 58:3611-362 (1945) и утверждают, что Tween 80 позволяет "быстрый", "диффузный" или "погруженный рост" как результат "смачивания". Результаты фиг.4A и табл. 6, однако, предполагают, что вне зависимости от того, связано ли поверхностное натяжение и слипание непосредственно с поверхностным, пленочным ростом, факт в том, что механизмы, благодаря которым при использовании Tween 80 наблюдается этот феномен, отличны от SB-18. Так как М. avium растет более диффузным образом и демонстрирует минимальное слипание, можно ожидать, что на MTB будут более драматично влиять эксперименты, описанные на фиг.5. Смысл в том, что Tween 80 должен действовать in vivo, вызывая погруженный рост. Несмотря на это, кажется логичным, что в отсутствие поверхностного натяжения существование погруженного роста и плавучести должно быть некоторым образом близко связано.
Silverstolpe, L. Nord.Med. 40/48:2220-2222 (1948) определил, что пределы удельного веса туберкулезных бацилл от 1,07 до 0,79, со значением, немного меньшим, чем 1,00. Silverstolpe, L Nord.Med. 40/48:2220-2222 (1948) заключает (цитата на языке оригинала). Литературно переведено: "Относительно низкий удельный вес бактерий можно объяснить содержанием их жиров." Идея в том, что действительное количество липидов в клеточной стенке отвечает за низкий удельный вес, как это принято в технике. В популярном микробиологическом тексте (1973) авторы утверждали: "... поразительное обилие липидов в клеточной стенке (60% от сухого веса) отвечает за гидрофобный характер микроорганизмов, что ведет к слипанию их друг с другом при росте на жидких средах и флотации на поверхности." (Davis, B.D. et al., Microbiology, Harper & Row, New York, 1973, стр.847). Однако, согласно данному изобретению, не только содержание липидов ответственно за низкий удельный вес микобактерий.
Согласно направлению исследований, можно предположить, что парциальный удельный объем липидов в клетке компенсирует парциальный удельный объем остальных компонентов, чтобы позволить флотацию. Это дает возможность предположить, что, в отсутствие поверхностного натяжения, дополнительный липидный компонент в клеточной стенке М. avium непосредственно ответственен за увеличивающуюся трудность выделения в процессе центрифугирования (табл. 1). Stinson, M.W. et al., Am.Rev.Resp.Dis. 104:717-727 (1971) показали, что при оценке по глицерину, липидная оболочка составляет 17,7% от сухого веса клетки, тогда как в присутствии Tween 80 содержание липидов увеличивается до 34,1%.
Так как Tween 80 предотвращает рост в погруженной культуре, эта гипотеза не может быть рациональным объяснением плавучести: содержание липидов возрастает почти в два раза, однако клетки, как это не парадоксально, остаются погруженными.
Schaefer, W. B. et al., J.Bacteriol. 90:1438-1447 (1965) показывали это несоответствие годами раньше при опубликовании драматичных микрофотографий, показывающих образование липоидных тел в микобактериях в присутствии как олеин-BSA (бычий сывороточный альбумин), так и Tween 80-BSA содержащих сред. Weir, M. P. et al., Amer. Rev. Res. Dis. 106:450-457 (1972) и McCarthy, C. Infect. Immun. 4:199-204 (1971) характеризовали также быстрый подъем олеиновой кислоты в условиях культивирования. Представляется, что данные, необходимые для поддержания гипотезы о связи между плавучестью и общим содержанием липидов, отсутствуют. Из сравнения этих открытий можно прийти к объективному заключению, что, возможно, аккумуляция липидов способствует в некоторой степени погруженному росту.
Сделанные выше комментарии приводят к предположению, что погруженный рост в присутствии Tween 80 (CAS® N.9005-65-6) должен быть результатом некоторых иных изменений в структуре клетки. Доклад Yamori, S. et al., Microbiol. Immunol. 35:921-926 (1991) подтверждает, что при добавлении OADC (олеиновой кислоты, BSA, дектрозы, каталазы и NaCl) к среде, содержащей Tween 80, имели место значительные изменения клеточной стенки. Эти авторы заключают, что чувствительность к препаратам изменяется в результате изменения клеточной стенки (напр. , клеточная стенка становится более проницаемой). Дополнительно, Rastogi, N. et al., Antimicrob. Agents Chemother. 20:666-677 (1981) показали десятью годами раньше, что клеточная стенка значительно утоньшается в пассажах при культивировании, и что далее, в обогащенной среде, содержащей олеиновую кислоту, BSA и Tween 80, биохимические характеристики также изменяются. Рассмотренные вместе, эти наблюдения подразумевают, что если поверхностное натяжение может быть вовлечено в пленочный рост, ни поверхностное натяжение, ни состав липидов не могут полностью объяснить "плавучести" или "низко-специфичной тяжести". Вместо этого изменения в клеточной стенке должны быть ответственны за погруженный рост. Кроме того, должны быть и другие возможности, например, (i) процесс, при котором эти липиды включаются в стенку, (ii) структуры, образующиеся при инкорпорации, или (iii) комбинация этих возможностей.
В основном, существует два пути, при которых липид может быть ответственен за поверхностный рост. Первый, по предположению Dubos, R.J. Exp. Biol. Med. 58:361- 362 (1945), это поверхностное натяжение. Вторая теория, которая происходит из работы Silverstolpe, L Nord.Med. 40/48:2220-2222 (1948), основывается на "частичном специфичном липидном объеме" этих микроорганизмов.
Как здесь описано, согласно изобретению, полагают, что наиболее эффективным путем для микобактерий, как облигатных аэробов, для поддержания плавучести в отсутствие поверхностного натяжения или частичных специфичных липидных объемов, является задерживание газов (напр., CO2), образующихся как побочный продукт при построении клеточной стенки. Приблизительно 80 молекул CO2 образуются на каждый остаток миколовой кислоты, синтезированной по пути малонил CoA. Образование CO2 микроорганизмом, близко родственным микобактериям, Propionobacterium shermanii, образует "глаза" (т.е. дыры) в швейцарском сыре (Sherman. J. M. J.Bacteriol. 6:379-392 (1921)). На молекулярном уровне клеточная стенка может просто иметь макромолекулярные карманы с задержанным в них CO2 в переплетенной сети отвержденных миколовых кислот и гликолипидов. Так как температура плавления связывающего фактора (6,6'-димиколата α,α-D-трегалозы) составляет 43-45oC (Noll, H. et al., Biochim. Biophys. Acta 20: 299-309 (1956)), восковидная консистенция клеточной стенки может действовать как ловушка для газа в макромолекулярных карманах. Полагают, что помещение буфера под вакуум приводит к высвобождению растворенного CO2 из буфера так, что создается градиент концентрации. Это, в свою очередь, позволяет CO2, задержанному в клеточной стенке, раствориться в буфере. Предполагается, что этот процесс довольно неэффективен и является еще менее эффективными для разрушения тяжей.
На фиг. 11 показана схема окончательной конфигурации общего протокола процесса. На фиг. 11A показаны результаты дегазирования в присутствии и отсутствие SB-18 как для М. tuberculosis, так и для М. avium.
Для данных, представленных на фиг. 11A, схема, показанная на фиг.2, модифицирована, как описано на фиг. 11. Промывной буфер содержит воду или 10 мМ NaHPO4, pH 7,0, 15 мМ NaCl, 2 мМ SB-18, 2 мМ фенилаланина и 5 мМ DTT. Дополнительно, пробирки были сначала инкубированы при 37oC в течение 60 минут при встряхивании (140 об/мин) и затем или непосредственно центрифугировались, или помещались под вакуум (600 мм рт.ст.) на 60 мин при 40oC перед центрифугированием. Перед дегазированием крышки были удалены. В этом эксперименте были проверены как М. tuberculosis, так и М. avium. Дубликаты амплификаций четырех аликвот представлены как a, b, c и d, должны быть идентичными. Дубликаты амплификаций прямых аликвот, соответственно данным видам, показаны на соответственно помеченой линии. Повторные контроли 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также подвергнуты блоттингу.
Ясно, что как поверхностное натяжение, так и плавучесть играют значительную роль в объяснении поведения этих микроорганизмов в процессе амплификации. Даже в отсутствие каких-либо добавленных детергентов наблюдался как правило больший сбор клеток, когда организмы подвергались вакуумной обработке (см. выделение MAC в воде с вакуумным дегазированием без добавления детергентов. ) Ряд экспериментов привел к заключению, что как поверхностное натяжение, так и задержание газа оказывают влияние как для М. tuberculosis, так и для М. avium, доминирующей чертой для М. tuberculosis является поверхностное натяжение, и доминантной характеристикой для М. avium является задержанный газ. То есть, если SB-18 улучшает выделение обоих классов микобактерий, на MTB сильнее влияет детергент, тогда как на MAC дегазирование оказывает большее действие, чем на MTB. Дополнительно, дегазирование MTB в этих условиях, возможно, более эффективно благодаря тому, что SB-18 диспергирует эти микроорганизмы.
Если гипотеза относительно задержанного газа правильна, продолжительное дегазирование должно позволять любому детергенту улучшать выделение М. tuberculosis. Например, так как проблемы, окружающие ложноотрицательные результаты MTB, включают агрегирование, поверхностное натяжение и плавучесть, то если плавучесть может быть компенсирована до некоторой степени дегазированием, тогда остаются только агрегирование и поверхностное натяжение. Так как большинство детергентов, данных в подходящей концентрации, могут преодолеть поверхностное натяжение, остается только погрешность измерения. Фиг. 12 это показывает.
Для данных, представленных на фиг. 12, схема, показанная на фиг. 2, снова использована, как описано на фиг. 11. Промывной буфер содержит воду или 0,1% Тритона X-100 (CAS® N.9002-93-1) в 10 мМ Трис-HCl, pH 8,0 или 10 мМ NaHPO4, pH 7,0, 15 мМ NaCl, 2 мМ SB-18 (CAS® N. 13177-41-8) и 5 мМ DTT. Дополнительно, были подготовлены три комплекта пробирок для каждого из условий. После инокуляции, все пробирки инкубировались сначала при 37oC в течение 60 мин при встряхивании (140 об/мин). Один комплект пробирок центрифугировали непосредственно, тогда как остальные два комплекта для каждого из трех условий подвергались вакуумной обработке (600 мм рт.ст.). Один комплект пробирок был помещен под вакуум только на 60 мин при 40oC перед центрифугированием, и последний комплект пробирок был помещен под вакуум на 6 часов при 40oC перед центрифугированием (перед дегазированием крышки были удалены). В этом эксперименте была исследована только М. tuberculosis. Дубликаты амплификаций четырех аликвот представлены как a, b, c и d, должны быть идентичными. Дубликаты амплификаций прямых аликвот, соответственно данным видам, показаны на соответственно помеченной линии. Повторные контроли 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также подвергнуты блоттингу.
Ясно, что продолжительное дегазирование улучшает способность выделения этих микроорганизмов центрифугированием. При этих условиях включение любого детергента, не нарушающее тяжей, смягчает поверхностное натяжение до уровня, при котором увеличивается выделение бактерий путем центрифугирования. Следует заметить, что дегазирование, как ожидается, более эффективно в присутствии тех детергентов, которые нарушают образование связывающего вещества.
Другие детергенты с SB-18-подобной активностью
Следующие исследования проверяли, проявляют ли себя другие детергенты, аналогичные бетаинам, по отношению к клетке таким образом, что они изолируются внутри клетки и в процессе этого изменяют частичный специфичный объем клетки для того, чтобы частично нейтрализовать естественную плавучесть этих организмов, таким образом повышая выделение путем центрифугирования.
Например, предполагая, что прохождение бетаинов через наружную мембрану использует гидрофильный путь (Connell, N.D. et al., In: Tuberculosis: Pathogenesis, Protection and Control, B.R. Bloom, ed., American Sosiety for Microbiology, Washington, D.C. (1994) pp. 333-352), и прохождение через внутреннюю мембрану использует липидный транспорт (Schaefer, W.B. et al., J. Bacteriol. 90:1438-1447 (1965); Weir, M.P. et al., Amer. Rev. Res. Dis. 106: 450-457 (1972) и McCarthy, C. Infect.lmmun. 4:199-204 (1971)) можно предварительно использовать класс неионных детергентов с похожими характеристиками, например, приблизительно-октадециловый детергент, что минимизирует объем головной группы. Если головная группа Tween 80 слишком объемна для того, чтобы пройти через пору, другие детергенты, такие как линейные полиоксиэтиленовые эфиры жирных кислот (напр., соединения "Brij"), могут проявлять себя таким образом, который допускает прохождение в клетки, что позволяет функционирование в рамках методов данного изобретения.
Hsiao, L. et al., J. Phys. Chem. 60:657-660 (1956) показали, что пространство для молекулы полиоксиэтиленфениловых эфиров того же порядка как и для бетаинов (напр. C9 - с в среднем 9,5 POE единиц будет занимать пространство в 55 A2). На фиг. 13 сравнивается SB-18 с Brij 96 (олеилполиоксиэтиленовый эфир(C18:1E10)) и Tween 80 (олеилполиоксиэтиленовый сорбитан (n=20)) в процессе анализа.
Эксперимент на фиг. 13 следует за анализом, представленным на фиг.2, и использует 2 мМ SB-18, 1% Brij 96 (CAS® N.9004-98-2) и 1% Tween 80 (CAS® N. 9005-65-6) в 10 мМ NaHPO4, pH 8,0, 15 мМ NaCl. В качестве отрицательного контроля использовалась вода. Пробирки в четырех повторностях были приготовлены для каждой серии. Все пробирки инокулировали 20 мкл бактериальной культуры М. tuberculosis и затем инкубировали при 37oC в течение 60 мин при встряхивании (140 об/мин) перед центрифугированием. Дубликаты амплификаций четырех аликвот, представленные как a, b, c и d, должны быть идентичными. Дубликаты амплификаций прямых аликвот, соответственно данным видам, показаны на соответственно помеченной линии. Повторные контроли 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также подвергнуты блоттингу.
Применение Brij 96 в многочисленных экспериментах, подобных этому, представлено на фиг. 13 и показывает сравнимые результаты. Brij 96 демонстрирует активность, аналогичную SB-18 в отношении компенсации плавучести; однако, как ожидалось, Brij 96 имеет ограниченную способность в отношении дисперсии М. tuberculosis. Несмотря на это, результаты, показанные здесь по Tween 80 (фиг. 13), согласуются с результатами на фиг.4A: Tween 80 не только улучшает выделение, но и диспергирует эти микроорганизмы. В поддержку гипотезы, что Brij 96 может проникать в клетку, Dubos, R.J. et al., J.Exptl.Med. 83:409-423 (1946) показали, что гомологи Brij 96 могут быть использованы для стимулирования роста в культуре. К сожалению, Dubos показал, что эти соединения, как наблюдалось, образуют преципитаты в присутствии сыворотки и других протеинов. Полезность преципитирующих белков с использованием линейных полимеров полиоксиэтилена уже известна в течение некоторого времени (см. Yamamoto, K.R. et al., Virology 40:734 (1970), в качестве одного из примеров). Это ограничивало бы полезность этих детергентов для обработки клинического материала.
Дальнейшие проблемы с использованием этих неионных детергентов заключаются в том, что они имеют очень узкий диапазон, в пределах которого могут функционировать. Например, на основании наших наблюдений, предпочтительны более длинноцепочечные алкилы, более длинные цепи алкилов имеют пониженную растворимость. Brij 96 имеет точку Краффта ниже 2oC и точку мутности при 54oC, тогда как Brij 76 (стеарилполиоксиэтиленовый эфир (C18E10): (CAS® N. 9005-00-9), имеет точку Краффта приблизительно 46oC и точку мутности при 64oC (Schott, H. et al., J. Pharm. Sci. 64:658-664 (1975); Schott.H. et al., J. Pharm. Sci. 65: 979-981 (1976). Brij 72 (стеарилполиоксиэтиленовый эфир (C18E2): (CAS® N. 9005-00-9) и Brij 92 (олеилполиоксиэтиленовый эфир (C18:1E2) (CAS® N. 9004-98-2) оба нерастворимы при условиях опыта. Возрастание растворимости требует возрастания числа единиц POE. Если число единиц POE возрастает, также возрастает пространство головной группы (Hsiao, L. et al. J.Phys.Chem. 60:657-660 (1956)).
Дополнительно, в пределах этого узкого диапазона, неионные детергенты ведут себя необычно. Например, Schott. H. et al., J.Pharm.Sci. 65:979-981 (1976) показали, что некоторые соли - нитраты натрия и калия могут вызывать перекрывание точки мутности и температуры Краффта Brij 56 (цетилполиоксиэтиленовый эфир (C68E10): (CAS® N.9004-95-9), вызывая таким образом образование "аморфного геля". Brij 56 имеет точку Краффта 32,5oC и точку мутности при 67oC (Schott. H. et al., J.Pharm.Sci. 65:979-981 (1976)). Снова преимущества бетаинов связаны с фактом, что длинноцепочечные алкилы могут быть надежно использованы в клинических условиях для диспергирования любого связывания и компенсирования плавучести.
Данные на фиг. 14 демонстрируют, что определенные классы детергентов имеют более высокую склонность к облегчению сбора путем центрифугирования, если микроорганизмы подвергнуты ограниченному дегазированию под вакуумом. На фиг. 14 показан представительный результат с использованием М. avium, когда далее следует модифицированная версия опыта, показанная на фиг. 11. Способы промывки с использованием как воды, так и 2 мМ SB-18 (10 мМ NaHPO4, pH 7,0, 15 мМ NaCl, 2 мМ SB-18 и 5 мМ DTT) сравнивали с несколькими различными детергентами в том же натрий-фосфатном буфере (10 мМ NaHPO4, pH 7,0, 15 мМ NaCl, 2 мМ SB-18, 2 мМ фенилаланина и 5 мМ DTT). Две серии детергентов четвертичного аммония были использованы для проверки как химической структуры, так и длины цепи алкила. В первой серии детергенты были длинноцепочечными алкил-триметиламмониевыми солями. Они включали (a) бромид додецилтриметиламмония (TMA-12: (CAS® N. 1119-94-4), смешанный бромид алкил-триметиламмония (mTMA: mTMA - это в основном тетрадецил-гомолог (C14) с присутствием некоторого количества C12- и C16- гомологов и (c) бромид октадецилтриметиламмония (TMA-18: (CAS® N. 1120-02-1). Во второй использованной серии детергенты были солями длинноцепочечного алкил-бензилдиметиламмония. Они включали: (d) хлорид бензалкония (BenzAlk (CAS® N. 8001-54-5)): BenzAlk в основном является гомологом C12, с присутствием некоторого количества C14 и C16); (e) хлорид бензилдиметилтетрадециламмония (BenzDMA-14: (CAS® N. 139-08-2) и (f) хлорид бензилдиметилстеарил аммония (BenzDMA-18). Каждый был обработан таким же образом, как было описано для SB-18 (Пример 1), и добавлен буфер до конечной концентрации 2 мМ. Пробирки были сначала инкубированы при 37oC в течение 60 минут при встряхивании (140 об/мин) и затем инкубировались в течение 60 мин при 40oC под вакуумом (600 мм рт. ст.). Дубликаты амплификаций четырех аликвот, представленные как a, b, c и d, должны быть идентичными. Дубликаты амплификаций прямых аликвот, соответственно данным видам, показаны на соответственно помеченной линии. Повторные контроли 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также подвергнуты блоттингу.
В соответствии с представленными здесь данными, хотя все детергенты, как наблюдалось, проявляли дополнительную эффективность, приблизительно-октадециловые детергенты, которые были способны к более активному изолированию, были значительно лучше в повышении выделения клеток центрифугированием, если дегазирование компенсировало плавучесть. Похожие эксперименты показывали, что неионные, октадециловые детергенты Tween 80 и Span 80, не способные прежде ни улучшать сбор организмов, не подвергнутых дегазированию, ни дезагрегировать микобактериальные клетки, давали результаты, аналогичные наблюдаемым на фиг. 14 (известно, что Tween 80 содержит малые количества гидролизованного детергента, и Span 80 просто является сорбитановым эфиром стеариновой кислоты).
Следовательно, дегазирование микобактерий открывает класс "приблизительно-октадециловых" детергентов (также называемых "приблизительно-октадецилподобными" детергентами), который включает SB-18 подобные детергенты, и который может теперь использоваться как SB-18-подобные детергенты для более эффективного выделения микроорганизмов. Так как микроорганизмы комплекса MAC растут сначала отдельными клетками, детергенты, использованные в эксперименте (фиг. 14), легко нарушали поверхностное натяжение и диспергировали клетки. Полагают, что эти детергенты, благодаря их прибилизительно-октадециловому характеру, более эффективно переносятся через мембрану и аккумулируются в клетке, как обсуждалось выше.
Таким образом, более длинные алкильные цепи важны по двум причинам. Во-первых, более длинные цепи облегчают дисперсию, и во-вторых, активнее осуществляют процесс изоляции. Детергенты с длинной цепью, менее растворимые, требуют специальных условий для функционирования. Например, ионные детергенты требуют минимизации электролитов и неионные детергенты требуют более крупных головных групп. Клинический материал требует работы в контексте растворов, содержащих электролиты, и активный транспорт детергентов в бактериальные клетки требует максимального уменьшения головной группы. Здесь и видны преимущества бетаинов: они функционируют более эффективно в растворах, содержащих электролиты, и у них минимизирован объем головной группы. Итак, бетаины предлагают уникальную возможность усилить быструю диагностику микобактериозов, так как длинноцепочечные алкилы могут быть надежно использованы в клинических условиях для диспергирования тяжей и скоплений клеток и компенсации плавучести.
Дополнительные черты, относящиеся к предназначению бетаинов для описанных здесь методов, предложены в работе Tsubone, K. et al., J.Pharm.Sci. 80: 441-444 (1991). Эти авторы показывают, что, как было отмечено, действенность антимикробной активности зависит от длины алкильной цепи, длины мостиковой группы и длины R2 и R3 (R2 и R3, как показано в табл. 2). Например, C16 в большинстве случаев представляется идеальной длиной цепи (C18-фосфоэтилбетаин очевидно преципитирует), метиловые группы в R2 и R3 позициях представляются выражающими наиболее высокий уровень антимикробной активности благодаря стерическим эффектам, и мостиковая группа C2 демонстрирует наиболее высокую антимикробную активность, тогда как C3 показывает минимум активности. Возрастание длины мостиковой группы свыше C3 показывает устойчивое возрастание антимикробной активности. В более поздней статье авторы показывают, что эти феномены кореллируют со способностью хелатировать двухвалентные катионы (Tsubone, K. et al. , J. Pharm.Sci. 80:1051-1054 (1991)).
В заключение, так как природа, состав и пространственные отношения заряженных частей делают бетаины уникальными, и контактная комбинация ионов приводит к изменению бетаинов, вариабельность внутри семейства; то основываясь на приведенном выше обсуждении, можно конструцировать бетаины с желаемыми характеристиками. Например, более длинная алкильная цепь может обеспечивать более эффективное изолирование и смягчать проблемы со связыванием. Применение в качестве мостика пропиловой группы уменьшает до минимума бактериостатическую активность. Группы R2 и R3 следует минимизировать до метиловых групп для избежания стерического препятствия. Несмотря на это, представление о минимизации бактериостатической активности с помощью пропилового мостика (напр. SB-18 является сульфопропилбетаином) немедленно поднимает вопрос о жизнеспособности.
Жизнеспособность микобактерий, обработанных SB-18
В отношении М. tuberculosis этот опыт дает жизнеспособную культуру. В табл. 8 показаны данные по необработанной культуре BACTEC в сравнении с освобождением 14CO2 у необработанных микобактерий (A и B); клетки, инкубированные в ПЦР лизисном буфере без дополнительной обработки (C и D); клетки, инкубированные в лизисном буфере и обработанные при 60oC в течение часа и затем обработанные при 95oC в течение 15 мин (E и F); и клетки, обработанные SB-18 (G и Н).
Поскольку опыт с SB-18 дает жизнеспособные микроорганизмы и продолжительное дегазирование усиливает действенность любого детергента в улучшении сбора центрифугированием; в связи с тем, что микобактерии являются облигатными аэробами, продолжительное дегазирование, как можно ожидать, подвергнет риску жизнеспособность.
В табл. 9 показаны результаты комбинирования обработки SB-18 с дегазированием NALC/NaOH-осадков.
Таблица 9: Корреляция ПЦР и результатов высева с использованием SB-18 в промывном буфере и вакуумного дегазирования промытых осадков.
Представлены результаты включения стадии вакуумного дегазирования в дополнение к добавлению SB-18 к 2o-Промывному буферу. Все препараты были сначала обработаны NALC/NaOH (Kent, P.T. et al., "Public Health Mycobacteriology" в A Guide for the level III laboratory, U.S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46). Осадок был сначала удален промыванием и инкубировался в BACTEC 12B культуральных колбах. Все положительные культуры MTB, MAC и М. kansasii были идентифицированы с помощью методики Gen-Probe (Gen-Probe, San Diego, CA). Все остальные микроорганизмы были идентифицированы с помощью биохимического анализа (Kent, P. T. et al., "Public Health Mycobacteriology" в A Guide for the level III laboratory, U. S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 71-157). Состав буфера, использованного для промывки осадков NALC/NaOH в этих сериях образцов, был 10 мМ NaHPO4, pH 8,0, 15 мМ DTT, 2 мМ SB-18. Осадки промывали в течение 60 мин при 37oC при встряхивании (140 об/мин). После промывания осадка образцы были помещены под вакуум 600 мм рт. ст. при 40oC на 60 мин. Образцы были затем подвергнуты центрифугированию (5000 g в течение 20 мин при 37oC) и далее подвергнуты ПЦР, как описано в Материалах и Методах.
Из 504 образцов, промытых SB-18 при 37oC и дегазированных под вакуумом 600 мм рт.ст. при 40oC и амплифицированных в повторе, 13 образцов дали положительный результат высева для комплекса MAC, 4 дали положительный результат высева для комплекса MTB и 4 дали положительный результат высева для М. kansasii. Анализ результатов показывает, что 9 из образцов, давших положительный результат при высеве для MAC, были также положительны при ПЦР (70%); 3 из образцов, давших положительный результат при высеве для MTB, были также положительны при ПЦР (75%); и все препараты М. kansasii, давшие положительный результат при высеве, были ПЦР-положительны (100%). Три из образцов MAC дали ложный результат из-за содержания ингибиторов и один был необъяснен. Образец MTB дал ложный результат, продемонстрировав выпадающие статистические характеристики вне разрешения (низкое число копий). Три из образцов MAC, положительных при высеве и ПЦР, продемонстрировали характеристики, согласующиеся с статистическими выпадениями (т.к. один из дубликатов амплификации был положительным и один отрицательным). Те образцы, которые, как наблюдалось, представляли постоянно повторяющуюся погрешность, требуют расширения срока инкубации в культуре для определения этим методом.
Верным выводом является то, что неполные результаты не являются следствием проблем, связанных с методикой SB-18/дегазирование. Напротив, все данные, представленные выше, в сочетании с фактом, что образцы были сначала обработаны NALC/NaOH, показывают, что чистота образцов в значительной степени включает в себя предварительную обработку по методике SB-18/дегазация. Тот факт, что препараты MAC все еще представляют статистические выпадения и необъясненные результаты, предполагает далее, что несовершенные результаты будут преодолены, как только современные методы будут использоваться как средство обработки образцов на начальной стадии.
Замечания по безопасности
Так как крышки пробирок с препаратами должны быть удалены перед вакуумным дегазированием, может образоваться большое количество аэрозоля при несчастном случае. Таким образом, были приняты следующие правила для того, чтобы убедиться в безопасности персонала. Во-первых, все данные были получены с использованием последовательно промывок детергентом и методологии вакуумного дегазирования. Это было сделано исключительно по соображениям безопасности: так как стадия дегазирования требует, чтобы крышки были удалены, а стадия промывки детергентом требует перемешивания, выполнение этих двух стадий одновременно может привести к тому, что крышки будут отделены и персонал подвергнется воздействию препаратов. Во- вторых, аппарат, показанный на фиг. 15, помещен в вытяжной шкаф с ламинарным потоком. Работающему на аппарате следует убедиться, что поток воздуха в вытяжном шкафу с ламинарным потоком не прерывается вакуумной печью. В-третьих, весь воздух, поступающий из вакуумной печи, проходит через два комплекта фильтров для удаления частиц пыли. Первый фильтр представляет собой фильтр шприцевого типа 0,2μ (Gelman, Ann Arbor, Michigan), который подходит по размеру к вакуумной линии. Этот фильтр прикреплен к вакуумной печи через короткий отрезок вакуумной трубки. Используются шланговые зажимы для его иммобилизации, так что возможность случайного отсоединения исключается. Шприцевый фильтр заменяется ежемесячно, автоклавируется и помещается на место. Второй фильтр 0,3μ HEPA- CAP (Whatman, Clifton, NJ) и он также подходит к вакуумной линии. Этот фильтр соединяется со шприцевым фильтром без шланговых зажимов, так что он может быть удален, обеззаражен и храниться, если это необходимо. Фильтр HEAP-CAP заменяется раз в квартал. Наконец, вакуумная печь обеззараживается по тому же расписанию вместе с вытяжным шкафом с ламинарным потоком.
Пример 11
Метод SB-18-дегазирование превосходит процесс NALC/NaOH
Схема на фиг.2 была модифицирована так, чтобы было сделано прямое сравнение путем как высева, так и амплификации между необработанной М. tuberculosis, М. tuberculosis, обработанной с использованием SB-18 и дегазирования, и М. tuberculosis, обработанной NALC/NaOH. На фиг. 16 представлены необходимые модификации. Таблица 10 представляет необработанные данные по подсчету 14CO2, высвобождающегося из культур BACTEC 12B (Beckton Dickinson, Sparks, MD) для каждого из трех условий. Эти данные представлены на фиг.16A, и фиг. 16B представляет результаты амплификации, когда аликвоты обработанных образцов, тех же, что были использованы для инокуляции в высевах, амплифицированы ПЦР.
В этом эксперименте следует описание фиг. 16. Клетки М. tuberculosis были собраны из пробирок со скошенным агаром и суспендированы в воде. Суспензия далее была разбавлена для образования бактериального препарата, который далее использовался для инокулирования всех образцов. Два комплекта пробирок, каждый состоящий из 6 репликатов, были приготовлены для каждого этапа процесса. Рабочий раствор SB-18 состоял из 15 мМ NaHPO4, pH 8,0, 15 мМ NaCl, 2 мМ SB-18 и 5 мМ DTT. Двадцать мкл суспензии клеток были добавлены к каждой пробирке SB-18 и затем все были сначала инкубированы при 37oC в течение 60 мин при встряхивании (140 об/мин) и затем были помещены под вакуум (600 мм рт. ст.) на 60 мин при 40oC (перед дегазированием крышки были удалены). Пробирки были подвергнуты центрифугированию (5000 g в течение 20 мин при 37oC) и декантированы. Пять мл NALC/NaOH (Kent, P.T. et al., "Public Health Mycobacteriology" в A Guide for the level III laboratory, U.S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46) были помещены в каждый репликат этого комплекта и было добавлено 20 мкл бактериального материала. Пробирки с NALC/NaOH инкубировались 15 мин при комнатной температуре, разбавлены 45 мл стерильной воды, подвергнуты центрифугированию (2000 g в течение 20 мин при 4oC) и затем декантированы. Все осадки были ресуспендированы в 300 мкл стерильной воды. Были приготовлены шесть образцов положительного контроля для слежения за жизнеспособностью и силой максимального амплификационного сигнала. Эти контроли были приготовлены простым смешиванием 280 мкл стерильной воды с 20 мкл бактериального препарата. Все образцы были затем разделены для анализа как методом высева, так и амплификацией. Это было выполнено следующим образом: дублированные аликвоты по 25 мкл были использованы для инокулирования культур BACTEC 12B и аликвота в 200 мкл этого образца была добавлена к 200 мкл 2X лизисного буфера. Колбы с культурами проверялись обычным образом на высвобождение 14CO2 на счетчике BACTEC 460TB. Амплификацию выполняли как описано в Материалах и Методах. Дубликаты амплификаций шести образцов из каждого комплекта обозначены как a, b, c и d и должны быть идентичными (фиг. 16B). Повторные контроли 0, 20 и 100 копий были амплифицированы одновременно. Контроли гибридизации 108, 109 и 1010 копий были также подвергнуты блоттингу.
Следует сделать два основных замечания и несколько менее существенных наблюдений, касающихся этих данных. Первое основное замечание: результаты всех обработанных SB-18 высевов оказались положительными, и второе основное замечание: ни один высев из NALC/NaOH не оказался положительным. Известно, что NALC/NaOH подвергает риску жизнеспособность микобактерий (Krasnov, I. et al. , Am.J.Clin.Path. 45:352-355 (1966); Kubica.J.P.W. et al., Am.Rev.Resir. Dis. 87: 775-779 (1963)), результаты амплификации подтверждают (фиг. 16B), что действительно имели место большие потери микроорганизмов. В некоторых случаях эти потери количественны. Так как клинические данные представлены в табл. 9, использующей осадки, обработанные NALC/NaOH, должно быть ясно, что уровень, до которого повреждены образцы перед промыванием SB-18 и дегазированием, не является тривиальным.
Первое менее важное наблюдение то, что положительные контроли требуют почти две недели для перехода порогового значения (15 cpm). Сравнение этого с данными табл. 8, в которой положительные контроли клеток, обработанных SB-18, были положительны на 5 день, показывает чрезвычайно низкое число копий, использованных в этих экспериментах. Второе, данные в табл. 10 предполагают, что клетки, обработанные SB-18, запаздывают относительно контрольных клеток примерно на неделю. Этому есть три объяснения: (a) как обсуждалось в примере 10, предполагается, что SB-18 имеет некоторую бактериостатическую активность; (b) положительные контроли представляют истинное, максимальное потребление, тогда как потери, без сомнения, имеют место с обработанными клетками (при низком числе копий этот эффект обостряется); и, наконец (c) декантированные пробирки далее не обрабатывались (т.е. объем образца после добавления воды был во всех случаях больше 300 мкл), приводя к эффекту разбавления. Если степень, в которой детергент бактериостатичен, не может быть определена, как это и есть в действительности, ясно, что SB-18 не бактерициден. В отношении вышеуказанного "c", этот эксперимент был повторен и супернатант был перенесен в пробирки для микроцентрифугирования и обрабатывался далее для того, чтобы свести к минимуму как бактериостатический эффект, так и эффект разбавления. В этом эксперименте результаты SB-18 и контроля были неразличимы и два из 12 высевов NALC/NaOH были положительными.
Невозможно избежать заключения, что корни недостатка с приведенными выше диагнозами инфекций М. tuberculosis лежат в методах обработки, используемых в настоящее время в технике. Small, P.M. et al., New Engl.J.Med. 330:1703-1709 (1994) и Allard, D. et al., New Engl.J.Med. 330:1710-1716 (1994) изменили мышление медицинского общества (Hamburg, М.A. et al., New Engl.J.Med. 330: 1750-1751 (1994) опубликованием длины фрагмента рестрикции в исследованиях полиморфизма (RFLP) и предположением, что более трети инфекций ТБ в Соединенных Штатах были переданы в последнее время. Прежде оценивалось, что 90% инфекций латентны и возникают из неактивных очагов. Данные, представленные в этих примерах (и особенно пример 11), подразумевают в значительной степени, что из-за сочетания медленного роста, плавучести, агрегации, неправильных методов обработки и отсутствия надежных и чувствительных способов диагностики истинный процент случаев, возникающих из латентных инфекций или недавних трансмиссий, еще не установлен.
Из двух первичных источников ложно-отрицательных результатов, ингибитора и низкого числа копий, методы изобретения лучше разрешают последний. Детергент до некоторого уровня разрушает и понижает плавучесть, дегазирование еще понижает плавучесть. В результате микроорганизмы, имеющие эти характеристики, могут теперь быть более эффективно собраны для определения. Далее, так как методы данного изобретения усиливают способность собирать и экстрагировать низкое число микроорганизмов, имеющихся в образце, методы данного изобретения таким образом усиливают действенность способов определения, когда необходимо работать с низким числом копий.
Примеры 12 и 13 представлены как дополнительные примеры того, как использование SB-12 подобных детергентов может применяться на практике для биологических или других препаратов. Определено, что клинические лаборатории обязаны следовать рекомендованным опубликованным правилам (Kent, P.T. et al. , "Public Health Mycobacteriology" в A Guide for the level III laboratory, U. S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46), дан пример 12. Пример 13 с точки зрения результатов, представленных в нем, предлагает, чтобы современные методологии были бы оставлены для того, чтобы технологии, основанные на амплификации, надежно использовались в клинических лабораториях.
Таблица 10. Сравнение обработки SB-18 с обработкой NALC/NaOH
Представлены результаты высевов эксперимента по обработке, описанные на фиг. 16. Цифры, представляющие подсчет освобожденного 14CO2 для данного образца, определены на счетчике BACTEC 460TB (Becton Dickinson, Sparks, MD). Каждый образец проверяли регулярно в течение периода в 8 недель ("Days in Culture"). Три комплекта культур с шестью репликами каждый были инициированы: "Ввод", представляющий группу положительного контроля (образцы A-F); "SB- 18", представляющий М. tuberculosis, обработанную с помощью методов по данному изобретению, как показано на фиг. 11 (образцы G-L); и "NALC", представляющий M. tuberculosis, обработанную с помощью рекомендованного метода (Kent, P.T. et al., "Public Health Mycobacteriology" в A Guide for the level III laboratory, U.S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46) (образцы M-R). Каждый образец культивировали в двойной повторности (обозначенные 1 или 2). Значение выше 15 считалось положительным. Значения выше 999 выходят за шкалу и отмечались как "+". Данные для данного дня усредненные и приведены на фиг.16A.
Пример 12
Способ подготовки для амплификации микобактерий из клинических препаратов, которые предварительно были обработаны стандартными методами
Этот пример дан с точки зрения того, что клинические лаборатории должны следовать рекомендованным опубликованным правилам (Kent, P. T. et al., "Public Health Mycobacteriology" в A Guide for the level III laboratory, U. S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46).
Следующая методика разрешает упомянутые выше проблемы и полезна специально для подготовки микобактерий к амплификации для осадков, которые образованы любым из стандартных методов из клинических материалов (включая, например, мокроту, цереброспинальную жидкость (CSF) и мочу). Следующая методика полезна для подготовки микобактерий, включая, но не ограничиваясь, комплекс М. tuberculosis, комплекс М. avium и М. kansasii. Как видно из табл. 11, pH промывного буфера необходимо тщательно контролировать так, чтобы подобная SB-18 активность могла быть получена в методах по данному изобретению.
1. Микобактерии сначала экстрагируются из клинического материала с помощью Методики NALC/NaOH или любой методики, рекомендованной Kent (Kent, P.T. et al., "Public Health Mycobacteriology" в A Guide for the level III laboratory, U.S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46).
2. Осадок удаляется для высева и микроскопирования мазка.
3. Осадок может быть далее осветлен или очищен перед следующей стадией в случае необходимости. Такое осветление может проводиться с помощью методик, известных в технике для исследуемых жидкостей тела, включая, например, центрифугирование, гель-хроматографию и фильтрацию.
(a) Проведение стадий 4, 5 или 6 в зависимости от применения.
(b) Стадии 8-15 описаны для применения совместно с ПЦР. Эти стадии могут быть модифицированы так, чтобы результирующий обработанный осадок мог быть использован совместно с любым из описанных выше методов.
4. Промывка детергентом SB-18:
(i) Для оставшегося осадка доведите объем до приблизительно 25 мл, используя вторичный промывочный буфер SB-18. Отметим: вторичный промывочный буфер следует наливать в ту же самую коническую пробирку объемом 50 мл, которую использовали для обработки первоначального материала (т.е. не меняйте пробирки). (Хотя обычно используются конические пробирки объемом 50 мл, специалист может, конечно, использовать соответствующий контейнер любого размера, подходящий для желаемого процесса.)
(ii) Сильно перемешайте для того, чтобы ресуспендировать слипшийся осадок, и инкубируйте при 37oC в течение 60 мин (140 об/мин или достаточно сильно, чтобы разбить культуру и диспергировать ее в жидкости) сразу перед следующей стадией. Температура 37oC облегчает дисперсию и необходима для того, чтобы удержать детергент от преципитации.
(iii) Переходите к стадии 7
5. Вакуумное дегазирование:
(i) Оставшийся осадок довести до объема 25 мл водой или любым подходящим буфером. Отметим: воду следует наливать в ту же самую коническую пробирку объемом 50 мл, которую использовали для обработки первоначального материала (т.е. не меняйте пробирки).
(ii) Инкубируйте в течение 60 мин при 40oC под вакуумом 600 мм рт.ст. (крышки с пробирок для этой стадии следует удалить).
(iii) Переходите к стадии 7
6. Промывка детергентом SB-18 и вакуумное дегазирование:
(i) Оставшийся осадок довести до объема 25 мл водой или любым подходящим буфером. Отметим: воду следует наливать в ту же самую коническую пробирку объемом 50 мл, которую использовали для обработки первоначального материала (т.е. не меняйте пробирки).
(ii) Сильно перемешайте для того, чтобы ресуспендировать слипшийся осадок, и инкубируйте при 37oC в течение 60 мин (140 об/мин или достаточно сильно, чтобы разбить культуру и диспергировать ее в жидкости) сразу перед следующей стадией. Температура 37oC облегчает дисперсию и необходима для того, чтобы удержать детергент от преципитации.
(iii) Инкубируйте в течение 60 мин при 40oC под вакуумом 600 мм рт.ст. (крышки с пробирок для этой стадии следует удалить).
(iv) Переходите к стадии 7.
7. Вращайте пробирку 20 минут при 37oC. Отметим: в примерах была использована клиническая центрифуга IEC Model PR7000M. Использовали максимальную скорость оборотов/g, установленную для этого ротора, 5200 об/мин /7410g.
8. Пока пробирки вращаются, налейте 200 мкл 2X литического буфера в соответствующие номера помеченных пробирок для микроцентрифугирования с резьбой на 1,5 мл (оставьте пробирки при 4oC).
9. После центрифугирования декантируйте супернатант, тщательно стараясь не сместить осадок. Декантируйте полностью, насколько это возможно.
10. Добавьте к осадку 200 мкл воды.
11. Ресуспендируйте осадок путем сильного перемешивания.
12. Перенесите 200 мкл препарата в соответствующим образом помеченные пробирки для микроцентрифугирования с резьбой на 1,5 мл, содержащие 200 мкл 2X литического буфера (из вышеупомянутой стадии 8).
13. Сильно перемешивайте и инкубируйте пробирки при 60oC в течение 60 мин.
14. Сильно перемешивайте и инкубируйте пробирки при 95oC в течение 30 мин.
15. Сильно перемешивайте и немедленно выполняйте ПЦР, или храните при - 20oC. Пробирки следует подвергнуть стадии быстрого центрифугирования непосредственно перед взятием аликвоты для амплификации.
Замечания по мониторингу pH в SB-18 Промывном буфере
Примеры 10 и 11 показывают, что pH промывного буфера надо отслеживать так, чтобы была достигнута максимальная активность SB-18, табл. 11 исследует pH двух различных SB-18 промывных буферов при добавлении раствора NALC/NaOH. Если следовать методам примера 12, промывной раствор должен иметь более высокую буферную силу, как 50 мМ Трис-HCl.
Таблица 11: Добавление NALC/NaOH к SB-18-Буферу
Таблица ниже показывает pH промывного буфера SB-18 при добавлении различных количеств раствора NALC/NaOH. Были использованы три различных концентрации раствора NALC/NaOH (2%, 3% или 4%). Они представляют наиболее часто встречающиеся концентрации NALC/NaOH, используемые в клинической лаборатории для обработки материалов для определения микобактерий (Kent, P.T. et al. , "Public Health Mycobacteriology" в A Guide for the level III laboratory, U.S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46). Эти соотношения относятся к проценту NaOH раствора, добавленного к фактическому материалу. Во время обработки фактического образца и после короткой инкубации для обеззараживания материала, этот раствор далее разводится или водой, или солевым фосфатным буфером (PBS) перед центрифугированием. В целях этого эксперимента и в порядке имитирования клинической обработки, 5 мл каждого из растворов NALC/NaOH добавляли в конические пробирки объемом 50 мл и немедленно разбавляли 45 мл воды. Различные количества (1 мл, 2 мл, 3 мл, 4 мл или 5 мл) разведенных растворов NALC/NaOH были затем добавлены к 25 мл (A) - 10 мМ NaHPO4, pH 8,0, 15 мМ NaCl или (B) - 50 мМ Трис-HCl, pH 8,0, 50 мМ KCl, и был определен pH.
Пример 13
Методика приготовления микобактерий для высева, мазка или амплификации с использованием SB-18 или дегазирования в качестве принципиальной стадии исследования.
Этот пример дан в свете результатов, представленных здесь. При этом предлагается, чтобы современные методологии были оставлены и в клинической лаборатории надежно использовались методики, основанные на амплификации.
Последующая процедура разрешает упомянутые выше проблемы и применима специально для немедленного выделения микобактерий для высева, мазка и амплификации из образцов, которые не были обработаны каким-либо стандартным методом, более того, образцы обработаны непосредственно методом по данному изобретению. Эта методика может использоваться для выделения микобактерий из клинических материалов (включая, например, мокроту, цереброспинальную жидкость (CSF) и мочу) и применима специально для приготовления микобактерий, включая, но не ограничиваясь, комплекс М. tuberculosis, комплекс М. avium и М. kansasii. Эта методика может быть использована для выделения микобактерий из полужидких материалов (включая, но не ограничиваясь М. paratuberculosis из коровьего навоза, М. avium из птичьего помета или других микобактерий из почвы). Дополнительно, другие микобактерии (включая, но не ограничиваясь комплексом М. avium и М. paratuberculosis) могут быть изолированы из биологических жидкостей, таких как молоко или цельная кровь. Эта методика может быть использована для выделения микобактерий, включая, но не ограничиваясь М. gordonae из источника в окружающей среде, такого как вода. Эта методика может быть использована для выделения микобактерий из любого экзотического источника, такого как чешуя рыб и рептилий, образцы кожи амфибий или другие образцы ткани, человеческие или другие (включая, но не ограничиваясь, комплекс М. tuberculosis (MTB), комплекс М. avium (MAC), М. marinum, М. fortuitum, М. chelonae).
ПРИМЕЧАНИЯ:
(a) Стадии 1-4 описывают, как использовать образец, который не был сначала обработан по какой-либо стандартной методике, более того, образец обрабатывается по методу данного изобретения.
(b) Стадии 11-14 описаны для применения совместно с ПЦР. Эти стадии могут быть модифицированы так, чтобы результирующий обработанный осадок мог быть использован совместно с любым из описанных выше методов.
(c) Начните процедуру со стадий 1, 2, 3 или 4 в зависимости от применения.
(d) Любой образец, требующий осветления или очищения, или и того и другого перед началом этой обработки или перед любой стадией может быть обработан с помощью методик, известных в технике, включая, например, центрифугирование, гель-хроматографию, и/или ионообменную хроматографию, и/или фильтрацию. Описанный метод осветления и/или очищения остается на усмотрение специалиста и будет зависеть от применения. Стадия 4 включена, чтобы служить этому примером.
1. Промывка детергентом SB-18:
(i) К материалу (1-2 г или 1-2 мл) добавьте приблизительно 25 мл промывного буфера SB-18. Отметим: промывной буфер следует наливать в коническую пробирку объемом 50 мл с оригинальным материалом.
(ii) Сильно перемешайте для того, чтобы суспендировать образец, и инкубируйте при 37oC в течение 60 мин (140 об/мин или достаточно сильно, чтобы разбить культуру и диспергировать ее в жидкости). Температура 37oC облегчает дисперсию и необходима для того, чтобы удержать детергент от преципитации.
(iii) Переходите к стадии 5
2. Вакуумное дегазирование:
(i) Суспендируйте примерно 1-2 г образца или 1-2 мл жидкого образца в 6 мл стерильной воды или любого другого подходящего буфера, в конической пробирке объемом 50 мл. (Образцы воды следует брать с поверхности).
(ii) Инкубируйте в течение 60 мин при 40oC под вакуумом примерно 600 мм рт.ст. (крышки с пробирок для этой стадии следует удалить).
(iii) Переходите к стадии 5
3. Промывка детергентом SB-18 и вакуумное дегазирование:
(i) К материалу (1-2 г или 1-2 мл) добавьте приблизительно 25 мл промывного буфера SB-18. Отметим: промывной буфер следует наливать в коническую пробирку объемом 50 мл с оригинальным материалом.
(ii) Сильно перемешайте для того, чтобы суспендировать образец, и инкубируйте при 37oC в течение 60 мин (140 об/мин или достаточно сильно, чтобы разбить культуру и диспергировать ее в жидкости). Температура 37oC облегчает дисперсию и необходима для того, чтобы удержать детергент от преципитации.
(iii) Инкубируйте в течение 60 мин при 40oC под вакуумом примерно 600 мм рт.ст. (крышки с пробирок для этой стадии следует удалить).
(iv) Переходите к стадии 5
4. Промывка детергентом SB-18 с последующим очищением и/или осветлением (Вакуумное дегазирование необязательно).
(i) Суспендируте примерно 1-2 г образца или 1-2 мл жидкого образца в 6 мл стерильной воды или любого другого подходящего буфера, в конической пробирке объемом 50 мл.
(ii) Сильно перемешайте образец и оставьте стоять при комнатной температуре на 10 минут.
(iii) Приготовьте пробирки с 1,25X SB-18 Промывным Буфером и храните при 37oC.
(iv) Осветлите образец центрифугированием при 2700 об/мин (2000 g) при 25oC в течение 10 мин или с любой другой силой, необходимой для осветления образца без нарушения дальнейшей обработки микобактерий в супернатанте (эта стадия необязательна, если в образце нет видимых включений).
(v) Удалите из центрифуги и оставьте стоять на 10 мин при комнатной температуре.
(vi) Тщательно декантируйте большую часть супернатанта каждого образца непосредственно через колонку G-50 (Pharmacia, Piscataway, NJ) в соответственно помеченные конические пробирки объемом 50 мл, содержащие промывной буфер 1,25X SB-18. Удалите желаемый объем, но оставьте часть, наиболее близкую к осадку. Постарайтесь перенести как можно меньше осадка.
(vii) Сильно перемешайте содержимое и затем инкубируйте при 37oC в течение 60 мин при встряхивании (140 об/мин).
(viii) Удалите крышки и инкубируйте в течение 60 мин при 40oC под вакуумом, приблизительно 600 мм рт.ст. (Эта стадия необязательна).
(ix) Переходите к стадии 5
5. Вращайте пробирку 20 минут при 37oC. Отметим: в примерах была использована клиническая центрифуга IEC Model PR7000M. Использовали максимальную скорость оборотов/g, установленную для этого ротора, 5200 об/мин /7410 g.
6. Пока пробирки вращаются, налейте 200 мкл 2X литического буфера в соответствующие номера помеченных пробирок для микроцентрифугирования с резьбой на 1,5 мл (оставьте пробирки при 4oC).
7. После центрифугирования декантируйте супернатант, тщательно стараясь не сместить осадок. Декантируйте полностью, насколько это возможно.
8. Добавьте к осадку 200 мкл воды.
9. Ресуспендируйте осадок путем сильного перемешивания.
10. Отберите аликвоту в 100 мкл для высева, если требуется. Обычно остается от 100 до 200 мкл супернатанта на дне пробирки. Добавление более чем 200 мкл воды к осадку обычно не обязательно: будет достаточно жидкости для высева, мазка и амплификации. Отметим: для анализа мазка детергент должен быть удален, насколько это возможно.
11. Перенесите 200 мкл препарата в соответствующим образом помеченные пробирки для микроцентрифугирования с резьбой на 1,5 мл, содержащие 200 мкл 2X Лизисного Буфера (из вышеупомянутой стадии 6).
12. Сильно перемешивайте и инкубируйте пробирки при 60oC в течение 60 мин.
13. Сильно перемешивайте и инкубируйте пробирки при 95oC в течение 30 мин.
14. Сильно перемешивайте и немедленно выполняйте ПЦР или храните при -20oC. Пробирки следует подвергнуть стадии быстрого центрифугирования непосредственно перед взятием аликвоты для амплификации.
Сравнение клинических образцов, обработанных NALC/NaOH и SB-18
В табл. 12 сравниваются тринадцать образцов, которые были обработаны как NALC/NaOH, так и SB-18. Использованные материалы представляли собой образцы, отброшенные клинической лабораторией, часть которых была обработана NALC/NaOH. Начальный объем этих образцов доходил до максимального рекомендуемого объема, предложенного Kent, P.T et al., "Public Health Mycobacteriology" в A Guide for the level III laboratory, U.S. Department of Health and Human Service, Centers for Disease Control (1985), стр. 31-46. Таким образом, был использован остаток оригинального материала, который был отброшен с последующей инокуляцией культуры осадками NALC/NaOH. Обычно для обработки SB-18 остается меньше 1 мл. Культуры 1, 10, 12 и 13 при обработке обоими методами были положительны при высеве. Однако, образец SB-18 из культуры 1 не содержал кислотоустойчивого материала. Как было показано позже, рост в этой культуре проявляется за счет штамма Pseudomonas. Все культуры 10, 12 и 13 содержали кислотоустойчивый материал при обработке обоими методами. Осадки SB-18 были амплифицированы и образцы 4, 10, 12 и 13 были ПЦР-положительными. Образец 1 содержал ингибиторы, и образец 4 был получен от пациента, диагностированного в прошлом инфекцией М. tuberculosis. Допускается, что этот пациент проходил лекарственную терапию. Образец 4 при микроскопии мазка дал отрицательный результат. Результаты в табл. 12, хотя и очень ограниченные, показывают, что образцы могут быть обработаны SB-18 и амплифицированы для того, чтобы оценить наличие микобактериальной ДНК. Дополнительно, SB-18 имеет значительную бактерицидную и бактериостатическую активность. Однако, Pseudomonas, который также довольно непроницаемый объект (Jarlier, V. et al., J. Bacteriol. 172%1418-1423 (1990), возможно, имеет отношение к высевам обработанных SB-18 образцов. Следует заметить, что образцы 10, 12 и 13, обработанные SB-18, которые предоставляли значительно меньше материала, быстрее давали образцы, положительные при высеве.
Таблица 12: Обработка клинического материала непосредственно SB-18.
Сравнивались данные высевов, которые показали 13 клинических материалов, обработанных как NALC/NaOH, так и SB-18. Конкретно, методика NALC/NaOH была проведена по Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S. Department of Health and Human Service, Center for Disease Control, (1985) pp. 31-46 (3% окончательно) и методика SB-18 - как это описано выше с началом на стадии 3, и затем продолжена по стадиям 5 - 14. Осадки NALC/NaOH были проверены на кислотоустойчивый материал согласно методике Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 71-157. Использованные материалы представляли собой образцы, отброшенные клинической лабораторией, часть которых была обработана NALC/NaOH. Цифры, представляющие выход высвобождающегося из данного образца 14CO2, были получены на счетчике BACTEC 460TB (Bacton Dickinson, Sparks, MD). Значение более 15 считалось положительным. Положительные культуры были проверены на кислотоустойчивый материал согласно методике Kent, P.T. et al., "Public Health Mycobacteriology," в A Guide for the Level III Laboratory, U.S.Department of Health and Human Service, Center for Disease Control, (1985) pp. 71-157. Если было показано, что образец положителен по кислотоустойчивости, культура более не проверялась, обозначаясь "+". НД обозначает, что образцу не было 7 недель.
Пример 14
Способ количественного выделения комплекса М. avium из цельной крови.
(1) Добавьте 1 мл свежей цельной крови в 10 мл буфера, содержащего SB-18-подобный или приблизительно-октадециловый детергент, и сильно перемешайте растворы.
Отметим: цельная кровь должна быть взята в пробирку для сбора с ACD (цитратно-сахарозный буфер) (Becton-Dickinson) и использована в течение 24 часов. Содержимое пробирки для сбора должно быть тщательно перемешано после взятия.
Отметим: эта методика была оптимизирована с использованием CB-18 (CAS® N. 78195-27-4). Использованный буфер: 20 мМ Трис-HCl, pH 8,0, 2 мМ NaCl и 1 мМ CB-18. Наиболее высокие концентрации CB-18 (напр. 2 мМ) способствовали лизису этих микроорганизмов. Модификацию этого способа следует оптимизировать для использования SB-18-подобного или приблизительно-октадецилового детергента.
(2) Инкубируйте при 37oC в течение 60 мин при встряхивании (140 об/мин).
(3) Удалите крышки и поместите в вакуумную печь. Инкубируйте при 42oC в течение 60 мин под вакуумом 600 мм рт.ст.
(4) Закройте крышками и центрифугируйте при 3500 g 20 минут при 30oC.
Отметим: следует быть осторожным при центрифугировании пробирок объемом 15 мл. Во-первых, должны быть использованы подходящие роторные адаптеры. Во-вторых, не следует использовать полистироловые пробирки из-за возможности разрушения: следует использовать полипропиленовые пробирки.
(5) Декантируйте пробирки и добавьте 500 мкл промывного буфера, содержащего тот же самый SB-18-подобный или приблизительно-октадециловый детергент, и ресуспендируйте осадок. [Отметим: обычно образуется небольшой гелеобразный осадок. Не пытайтесь удалить из пробирки весь супернатант.]
(6) Перенесите пипеткой ресуспендированный осадок в пробирку для микроцентрифугирования с резьбой на 1,5 мл.
(7) Поместите пробирки в микроцентрифугу и вращайте 10 мин при комнатной температуре на максимальной скорости (температура этой стадии может изменяться в зависимости от применяемого в методе детергента).
(8) Удалите супернатант и добавьте 300 мкл стерильной воды к материалу.
(9) Ресуспендируйте осадок и перенесите 200 мкл в 2X лизисный буфер для амплификации. Обработайте материал для амплификации и определения, как описано в примере 1.
(10) Оставшийся осадок переносится для анализа в культуральную колбу BACTEC 12B (Becton-Dickinson).
Пример 15
Модифицированный метод обеззараживания щавелевой кислотой.
Возможное заражение Pseudomonas является самой большой проблемой при использовании метода примера 13. Если это требуется, Pseudomonas может быть элиминирован добавлением щавелевой кислоты в питательную среду или предварительной обработкой зараженной мокроты щавелевой кислотой. Метод такого обеззараживания следует далее. Этот метод модифицирует методику с щавелевой кислотой, описанную на стр. 43-44 Kent, P.T. et al., "Public Health Mycobacteriology, " в A Guide for the Level III Laboratory, U.S. Department of Health and Human Service, Center for Disease Control, (1985), стр. 31-46.
(1) Поместите от 0,5 до 2 мл необработанного материала или 2 мл зараженной культуры в коническую пробирку на 50 мл.
(2) Добавьте к материалу или зараженной культуре эквивалентный объем 5% щавелевой кислоты (CAS® N. 6153-56-6).
5% Щавелевая кислота: 5 г дигидрата щавелевой кислоты (126,02 г/моль) в 100 мл воды.
(3) Инкубируйте при комнатной температуре 15 минут.
(4) Добавьте одну десятую (1/10) объема "Нейтрализующего раствора NaOH" к смеси и немедленно сильно перемешайте. Например, если к материалу были добавлены 2 мл щавелевой кислоты, тогда "Нейтрализующего раствора NaOH" будет добавлено к материалу 200 мкл.
Нейтрализующий раствор NaOH: 31,76 граммов гидроокиси натрия (40 г/моль) в 100 мл воды (окончательная концентрация 7,94 М).
(5) Добавьте к материалу буфер, содержащий SB-18-подобный или приблизительно-октадециловый детергент до окончательного объема примерно 25 мл.
(6) Инкубируйте при 37oC 60 минут при встряхивании (140 об/мин).
(7) Удалите крышки и перенесите в вакуумную печь. Инкубируйте при 42oC 60 минут под вакуумом 600 мм рт.ст.
(8) Закройте крышками и центрифугируйте при 5000 g в течение 20 мин при 37oC.
(9) Декантируйте пробирки и добавьте к материалу 300 мкл стерильной воды.
(10) Ресуспендируйте осадок и перенесите 200 мкл в 2X лизисный буфер для амплификации. Обрабатывайте материал для амплификации и определения, как описано в примере 1.
(11) Оставшийся осадок переносится для анализа в культуральные колбы BACTEC 12B.
Пример 16.
Способ любого из примеров 12, 13, 14 или 15 за исключением того, что желаемый немикобактериальный микроорганизм, или желаемая группа микобактерий, или комплекс видов микобактерий, или комплекс микобактерий, такой как М. tuberculosis (MTB) комплекс, М. avium (MAC) комплекс, MAIS комплекс и комплекс М. fortuitum, также как быстрорастущие и медленнорастущие микобактерии, включая специфические и неспецифические фотохромогены, не-фотохромогены, скотохромогены, и в особенности М. africanum, М. asiaticum, М. avium, М. bovis (BCG), М. butyricum, М. chelonae, М. duvalii, М. flavescens, М. fortuitum, М. gastri, М. gorgonae, М. haemophilum, М. intracellularae, М. kansasii, М. leprae, М. lepraemurium, М. linda, М. lufu, М. marinum, М. malmoense, М. microti, М. mucoscum, М. nonchromogenicum, М. paratuberculosis. М. peregrinum, М. phlei, М. rhodochrous, М. scrofulaceum, М. shimoidei, М. simiae, М. smegmatis, M.szulgai, М. terrae, М. thermoresistable, М. triviale, М. tuberculesis, М. ulcerans, М. vaccae, М. xenopi определены путем определения их генетического материала или определением антигена, указывающего на присутствие этих микроорганизмов.
Пример 17
Способ, показанный в любом из примеров 12-16, за исключением того, что детергент является SB-18-подобным детергентом, как отмечено в табл. 2 и 3, иным, чем SB-18.
Пример 18.
Способ, показанный в любом из примеров 12-16, за исключением того, что детергент является приблизительно-октадециловым детергентом, иным, чем SB-18.
Пример 19.
Способ, показанный в любом из примеров 12-16, за исключением того, что по меньшей мере два различных октадецил-подобных детергента использованы в стадиях процесса.
Пример 20.
Список номеров®, химическое название и структура для номеров® соответствует представленному в табл. 13.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
(1) ОСНОВНАЯ ИНФОРМАЦИЯ
(I) ЗАЯВИТЕЛЬ: Конинг Клиникэл Лэборэтриз, Инк., г.Балтимор, штат Мэриленд, США
(II) НАЗВАНИЕ ИЗОБРЕТЕНИЯ: Способ приготовления образца для определения микроорганизмов (варианты) и набор для обработки образцов с целью определения микобактерий
(III) ЧИСЛО ПОСЛЕДОВАТЕЛЬНОСТЕЙ: 4
(IV) АДРЕС ДЛЯ КОРРЕСПОНДЕНЦИЙ:
(A) АДРЕСАТЫ: Sterne, Kessler, Goldstein & Fox
(B) УЛИЦА: 1100 New York Avenue
(C) ГОРОД: Вашингтон
(D) ШТАТ: Федеральный округ Колумбия
(E) СТРАНА: США
(F) КОД: 20005-3934
(V) ЧИТАЕМАЯ КОМПЬЮТЕРОМ ФОРМА:
(A) СРЕДСТВО ПЕРЕДАЧИ: Дискета
(B) КОМПЬЮТЕР: IBM-совместимый
(C) ОПЕРАТИВНАЯ СИСТЕМА: PS-DOS/MS-DOS
(D) ПРОГРАММНОЕ ОБЕСПЕЧЕНИЕ: Patentin Release #1.0, Version #1.25
(VI) ДАННЫЕ НАСТОЯЩЕЙ ЗАЯВКИ:
(A) НОМЕР ЗАЯВКИ:
(B) ДАТА ПРИОРИТЕТА
(C) КЛАССИФИКАЦИЯ:
(VII) ДАТА ПЕРВИЧНОЙ ЗАЯВКИ:
(A) НОМЕР ЗАЯВКИ: US 08/399,564
(B) ДАТА ПРИОРИТЕТА: 23.02.95
(C) КЛАССИФИКАЦИЯ:
(VII) ДАТА ПЕРВИЧНОЙ ЗАЯВКИ:
(A) НОМЕР ЗАЯВКИ: US 08/224,592
(B) ДАТА ПРИОРИТЕТА: 07.04.94
(C) КЛАССИФИКАЦИЯ:
(VII) ДАТА ПЕРВИЧНОЙ ЗАЯВКИ:
(A) НОМЕР ЗАЯВКИ: US 08/221,731
(B) ДАТА ПРИОРИТЕТА: 05.04.94
(C) КЛАССИФИКАЦИЯ:
(VIII) ПОВЕРЕННЫЙ/АГЕНТ
(A) ИМЯ: Cimbala Michele A.
(B) РЕГИСТРАЦИОННЫЙ НОМЕР: 33,851
(C) ССЫЛКА/НОМЕР ДОКУМЕНТА: 1453.0030003/MAC
(IX) ИНФОРМАЦИЯ ДЛЯ ТЕЛЕКОММУНИКАЦИЙ:
(A) ТЕЛЕФОН: (202) 371-2600
(B) ТЕЛЕФАКС: (202) 371-2540
(C) ТЕЛЕКС: 248636 SSK
(2) Информация о SEQ ID N: 1:
(i) Характеристики последовательности:
(A) Длина: 25 пар оснований
(B) Тип: нуклеиновая кислота
(C) Число нитей: две
(B) Топология: линейная
(xi) Описание последовательности: SEQ ID N: 1:
AAACTGGGTC TAATACCGGA TAGGA
(2) Информация о SEQ ID N:2:
(i) Характеристики последовательности:
(A) Длина: 21 пар оснований
(B) Тип: нуклеиновая кислота
(C) Число нитей: одна
(B) Топология: линейная
(xi) Описание последовательности: SEQ ID N:2:
CCACCTACCG TCAATCCGAG A
(2) Информация о SEQ ID N:3:
(i) Характеристики последовательности:
(A) Длина: 20 пар оснований
(B) Тип: нуклеиновая кислота
(C) Число нитей: одна
(B) Топология: линейная
(ix) Свойства:
(A) Имя/ключ: модифицированное основание
(B) Положение: 7
(D) Дополнительная информация: /основание, меченное меткой /Примечание = "Нуклеотид в положении 7 является инозином".
(xi) Описание последовательности: SEQ ID N:3:
GCGGGCNCAT CCCACACCGC
(2) Информация о SEQ ID N:4:
(i) Характеристики последовательности:
(A) Длина: 23 пар оснований
(B) Тип: нуклеиновая кислота
(C) Число нитей: две
(B) Топология: линейная
(xi) Описание последовательности: SEQ ID N:4:
GACCACGGGGA TGGATGTCTT GTG:
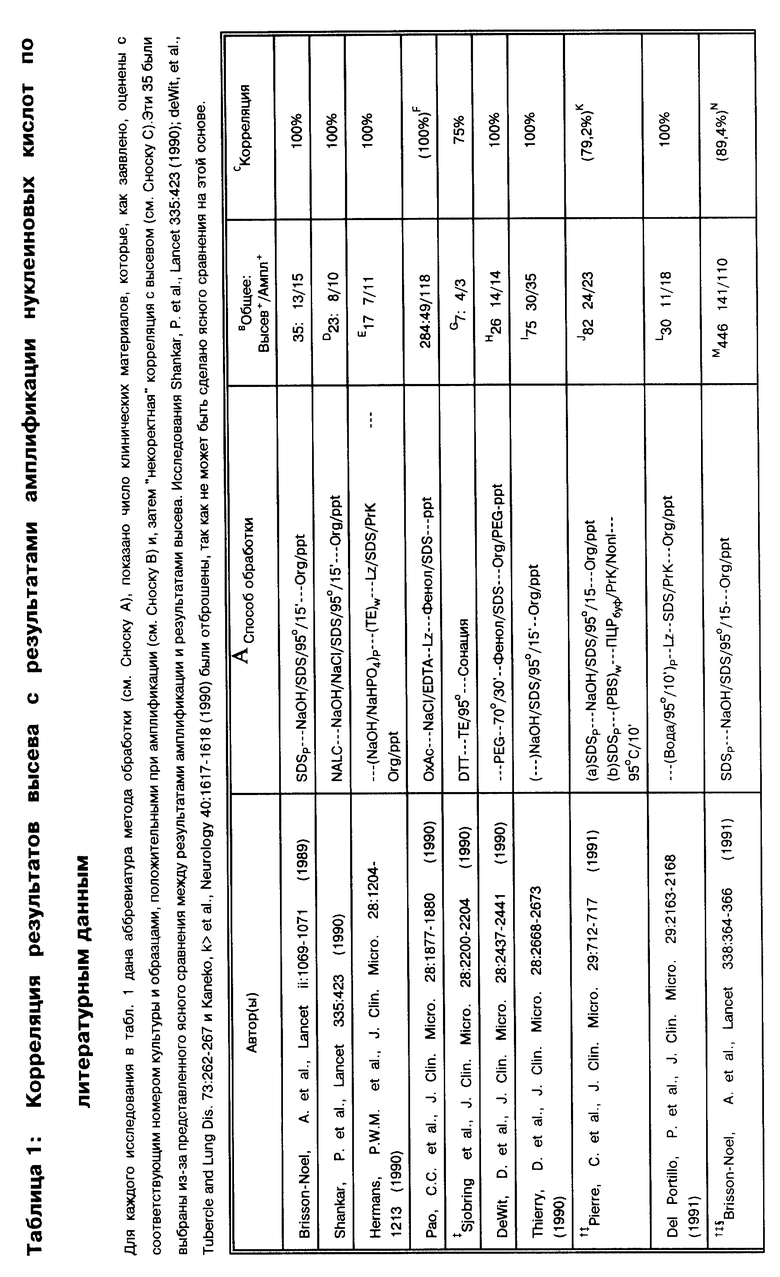
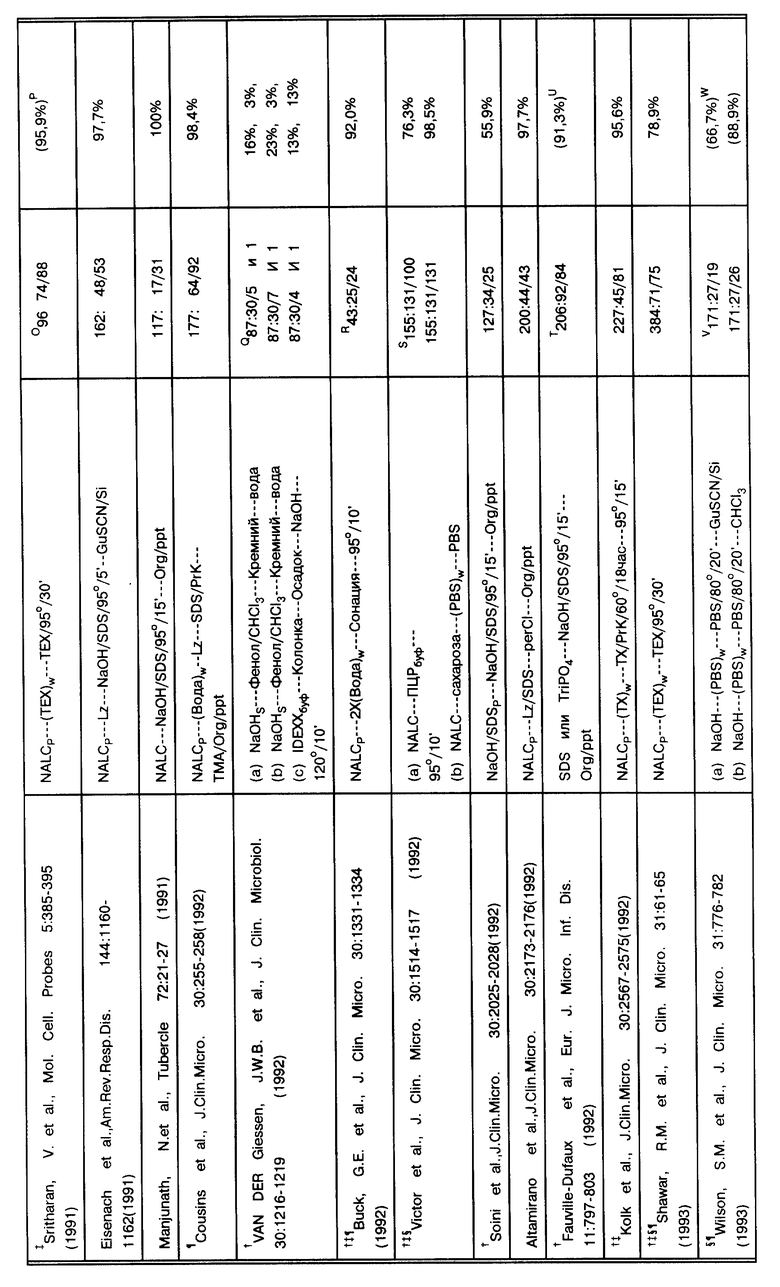
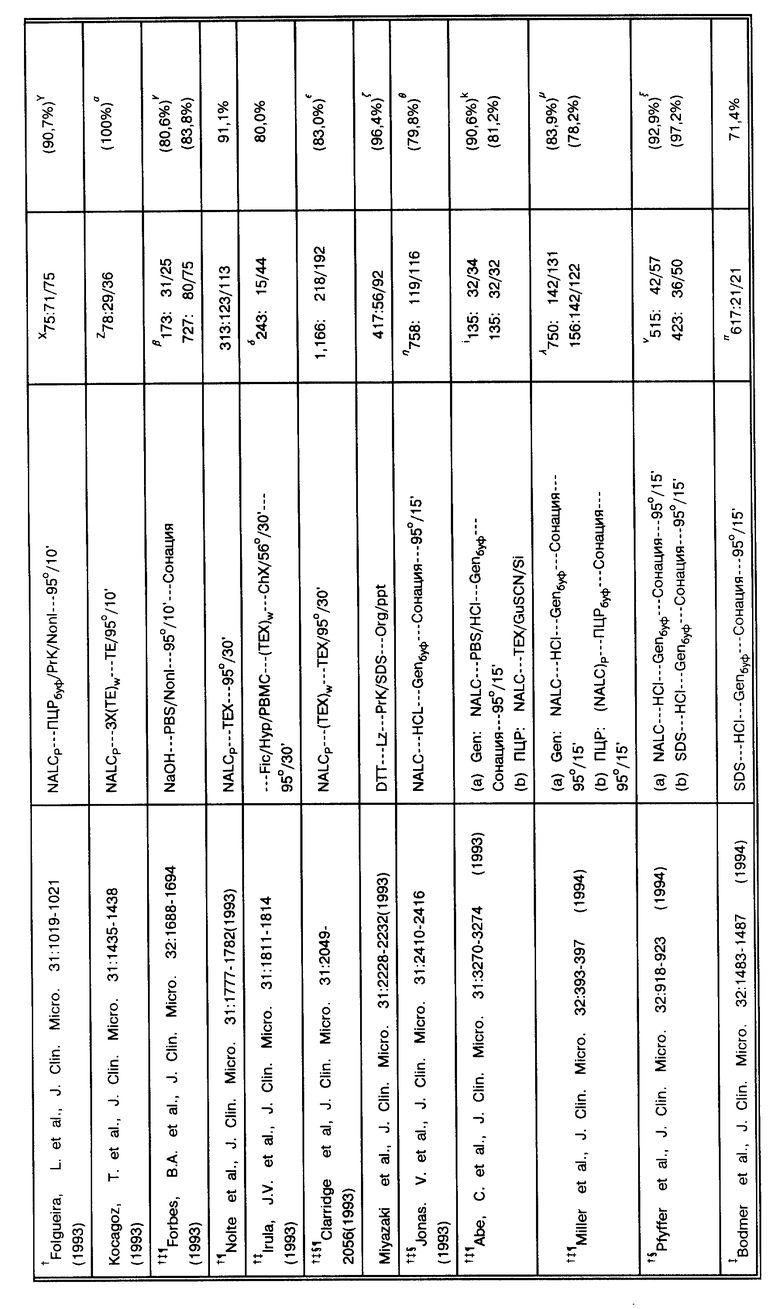
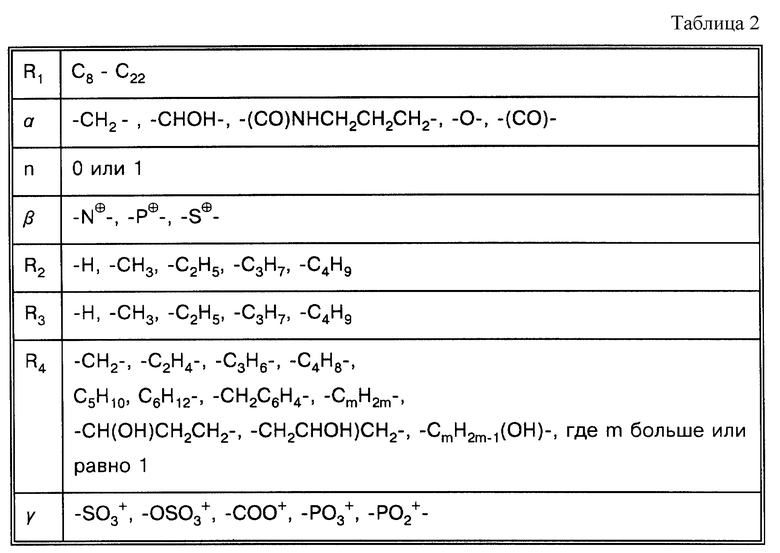
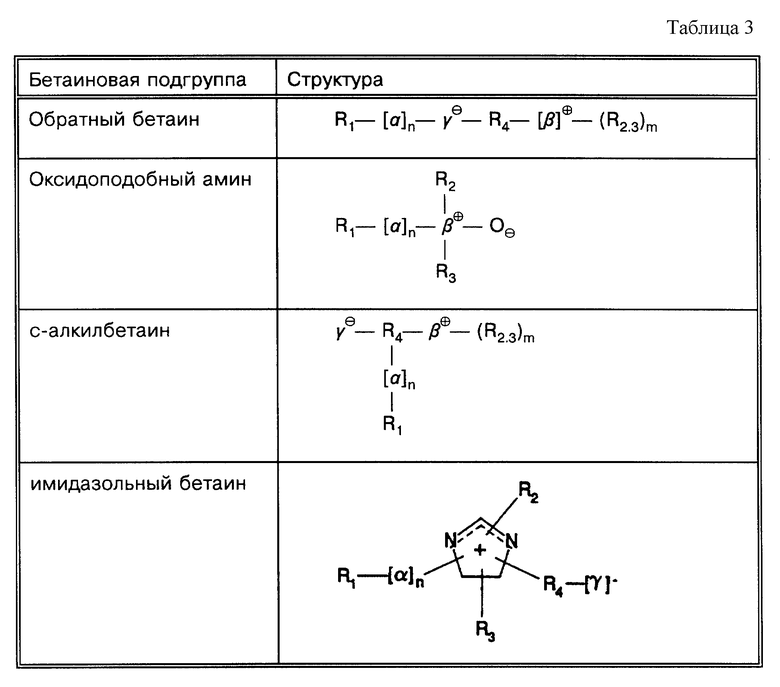
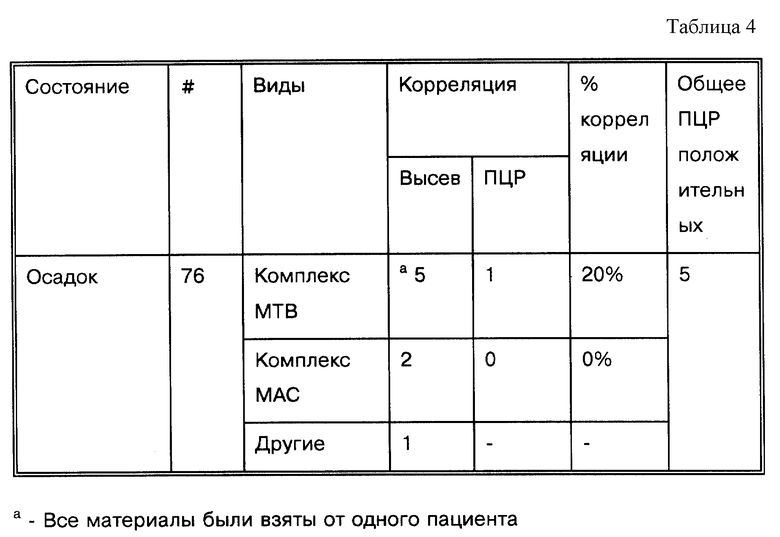
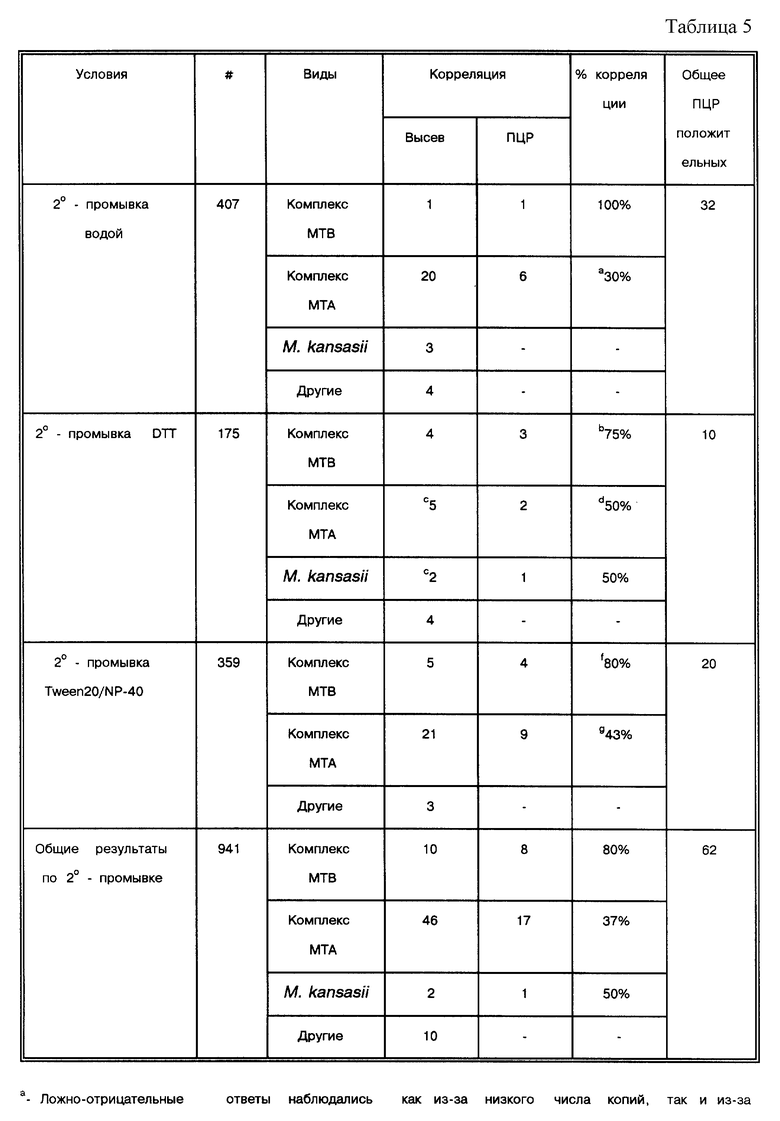

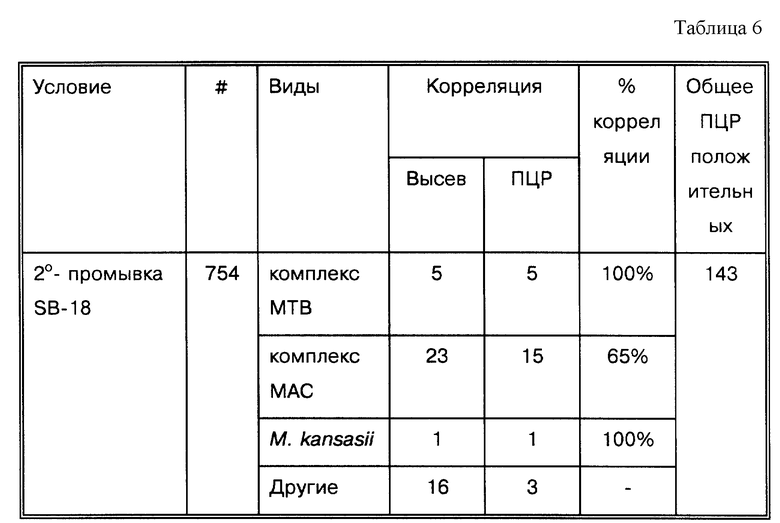
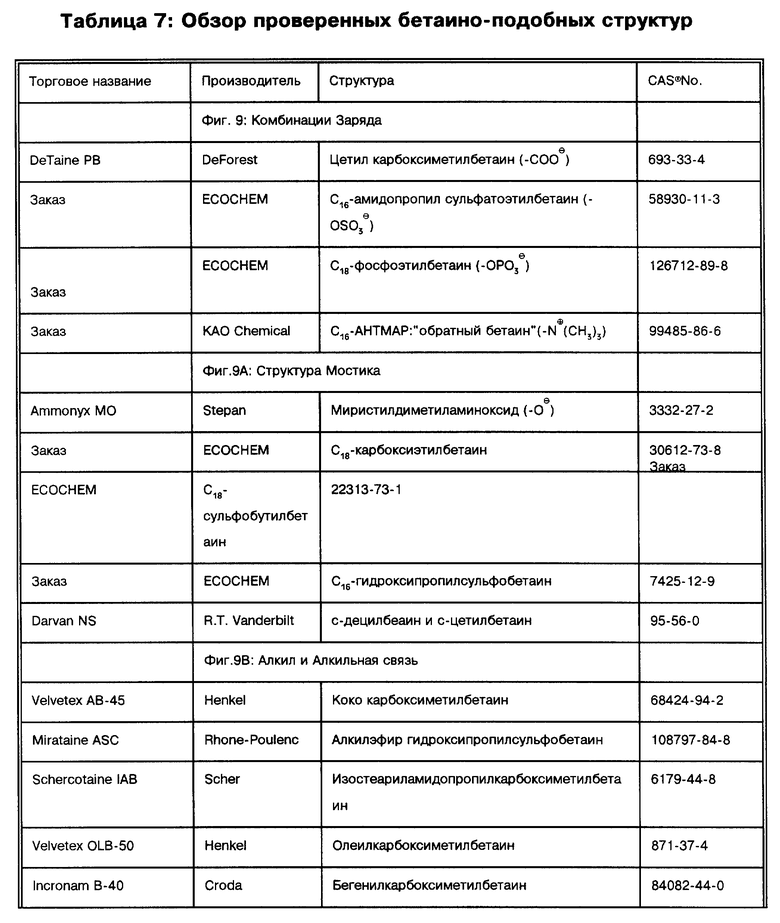
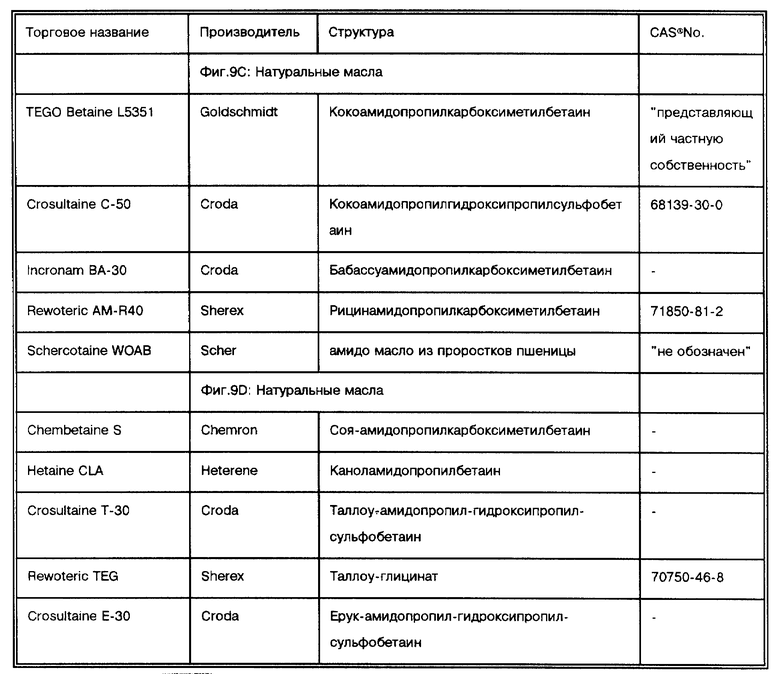
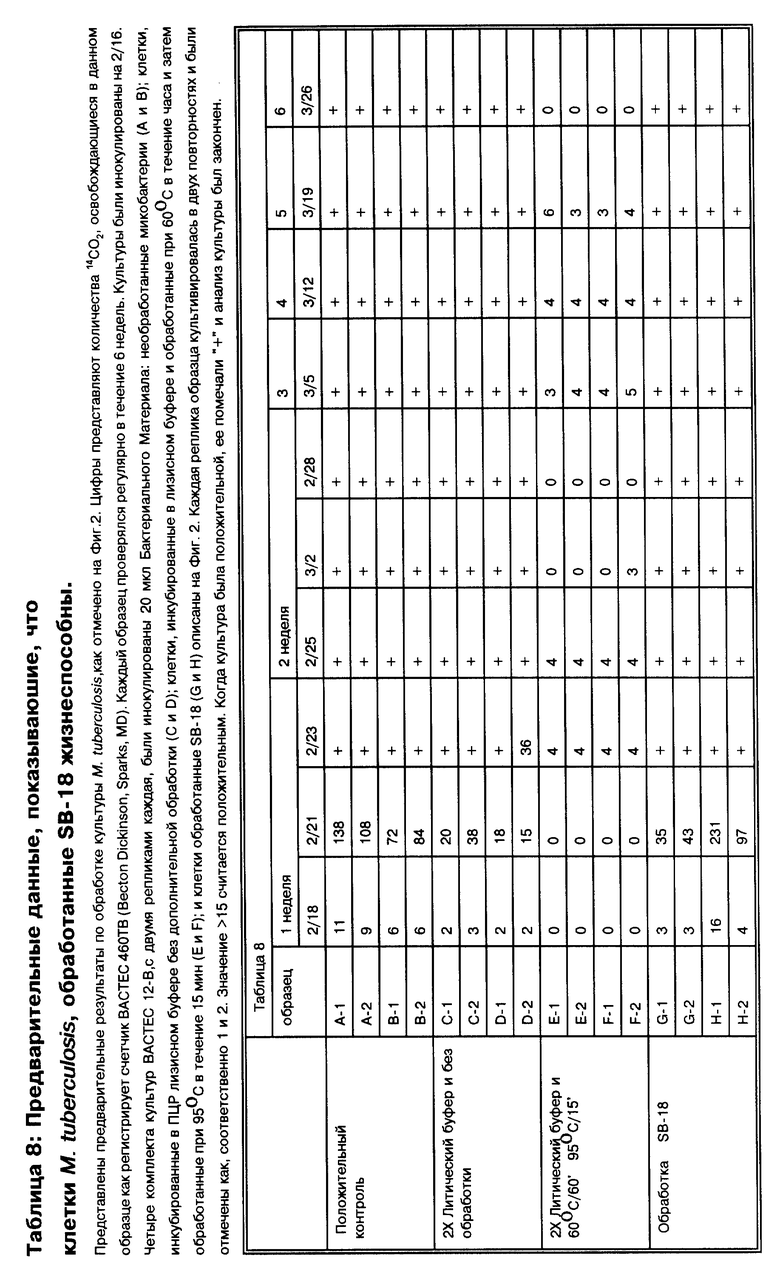
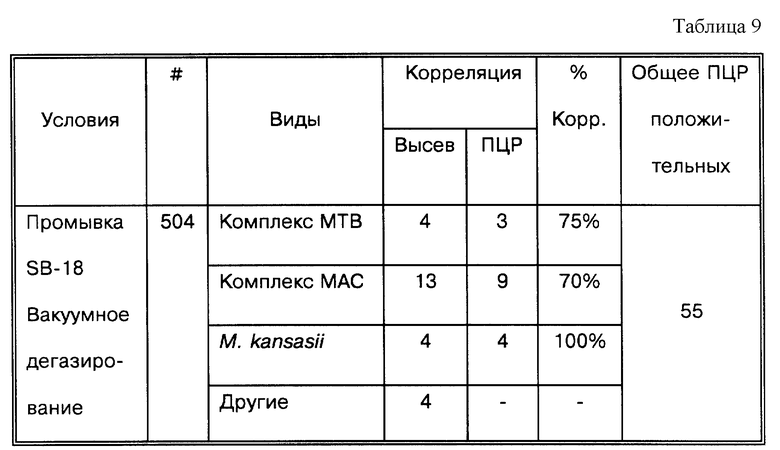
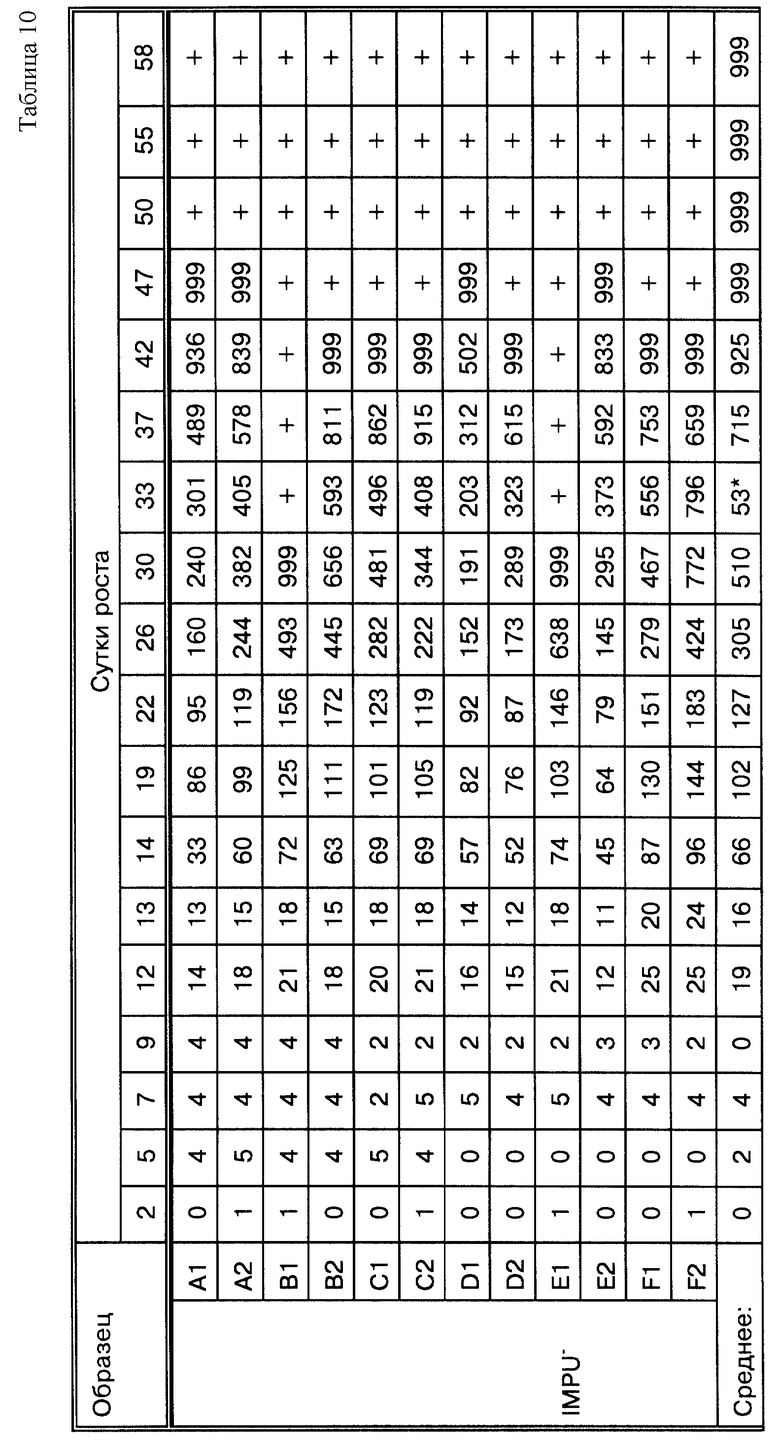
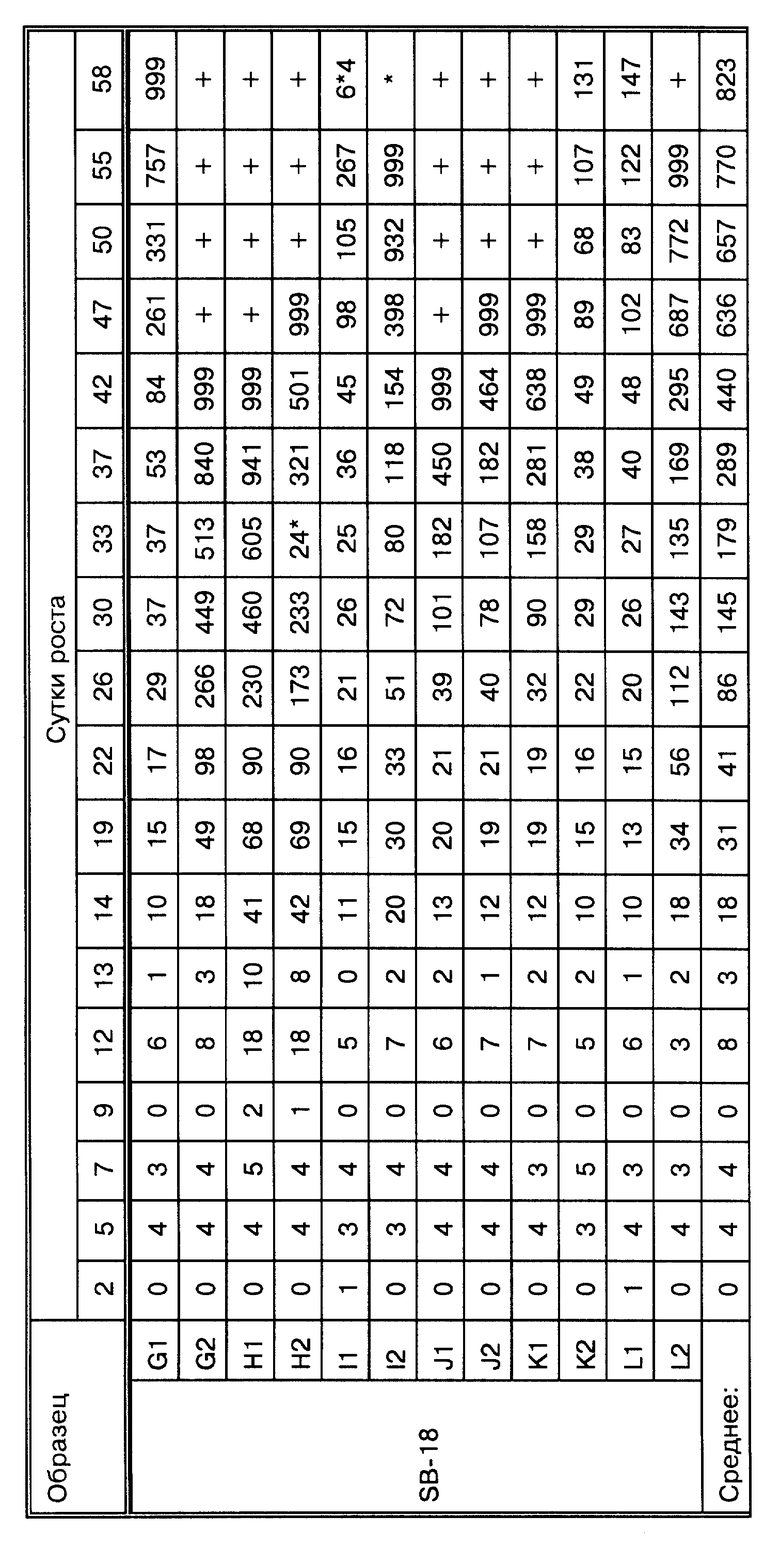
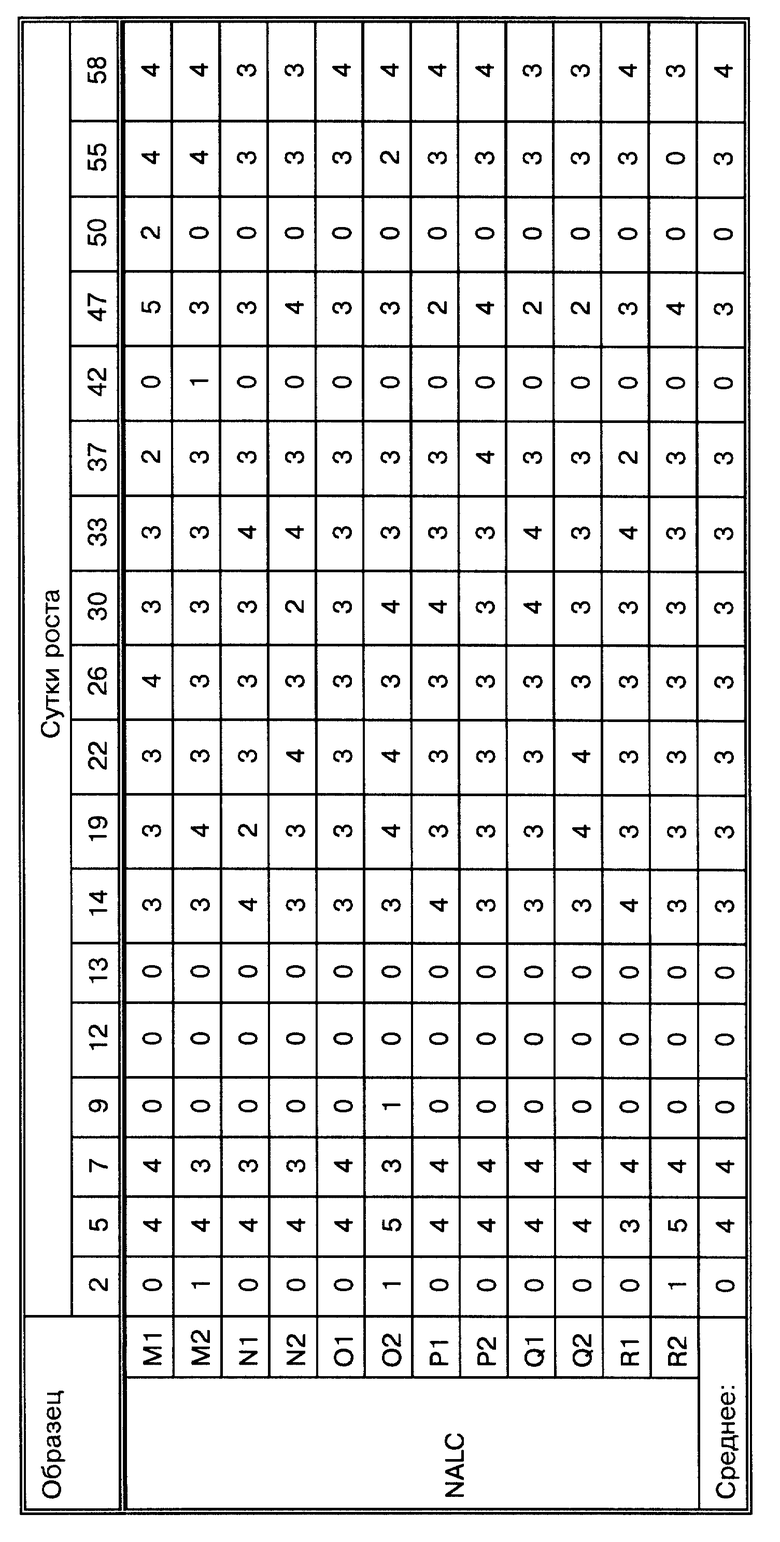
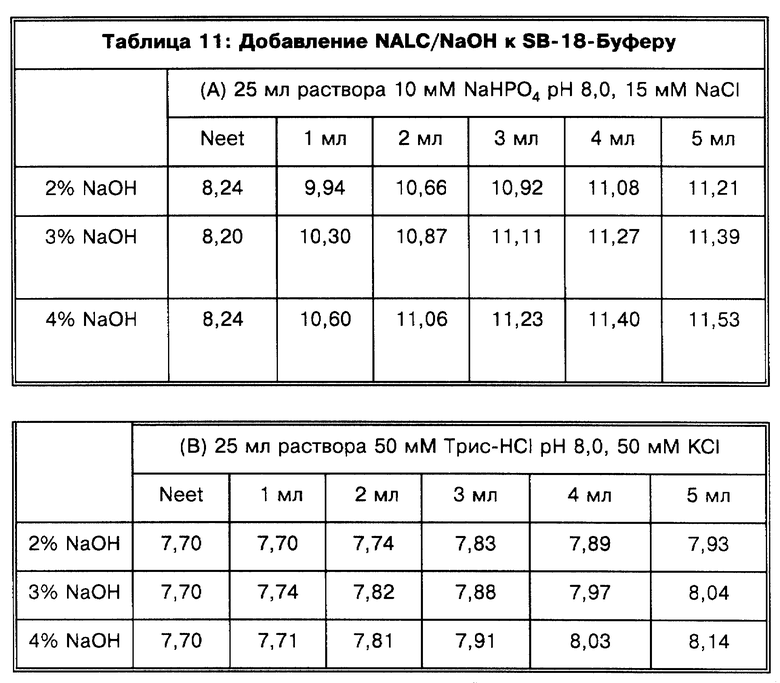
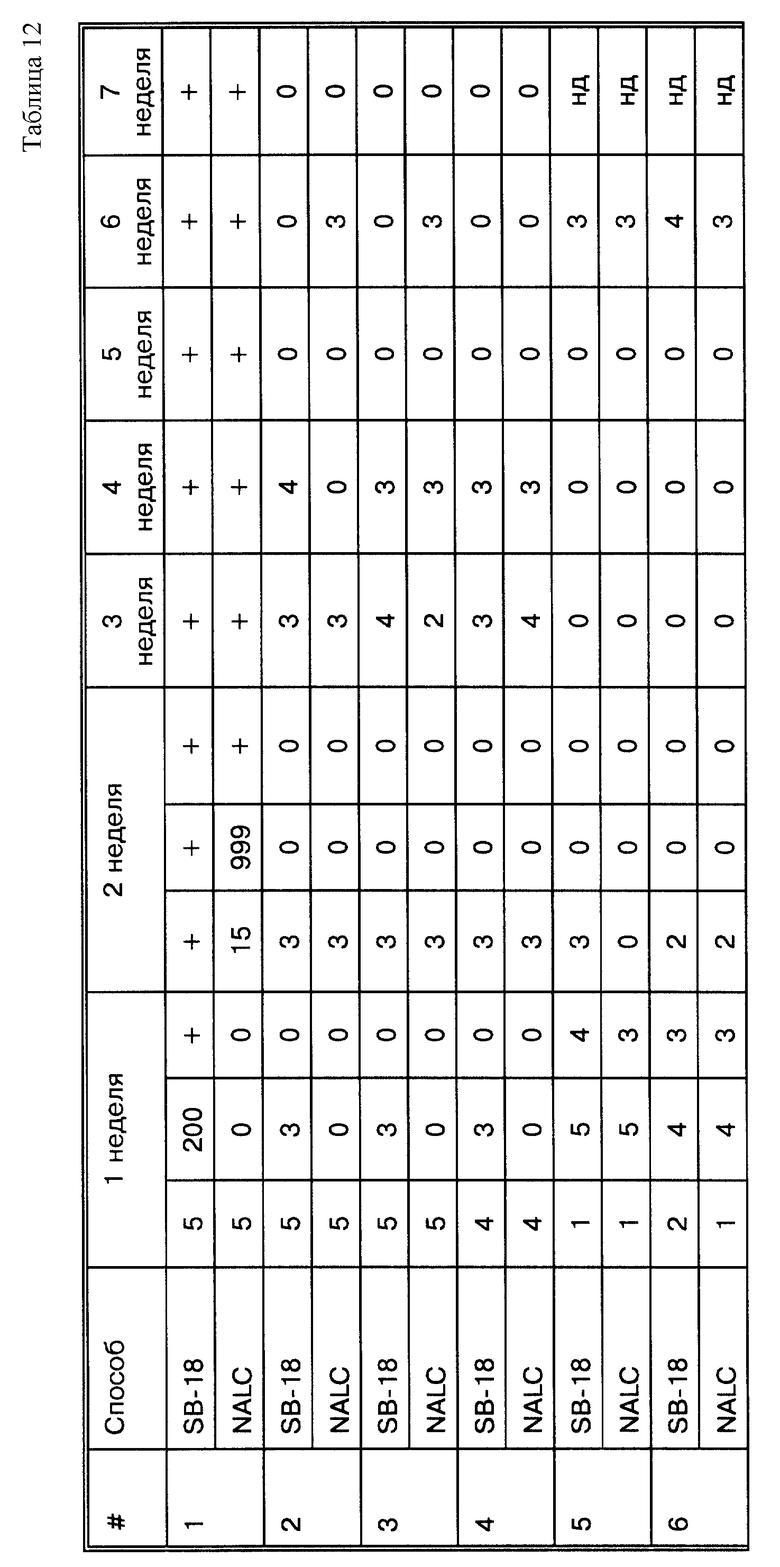
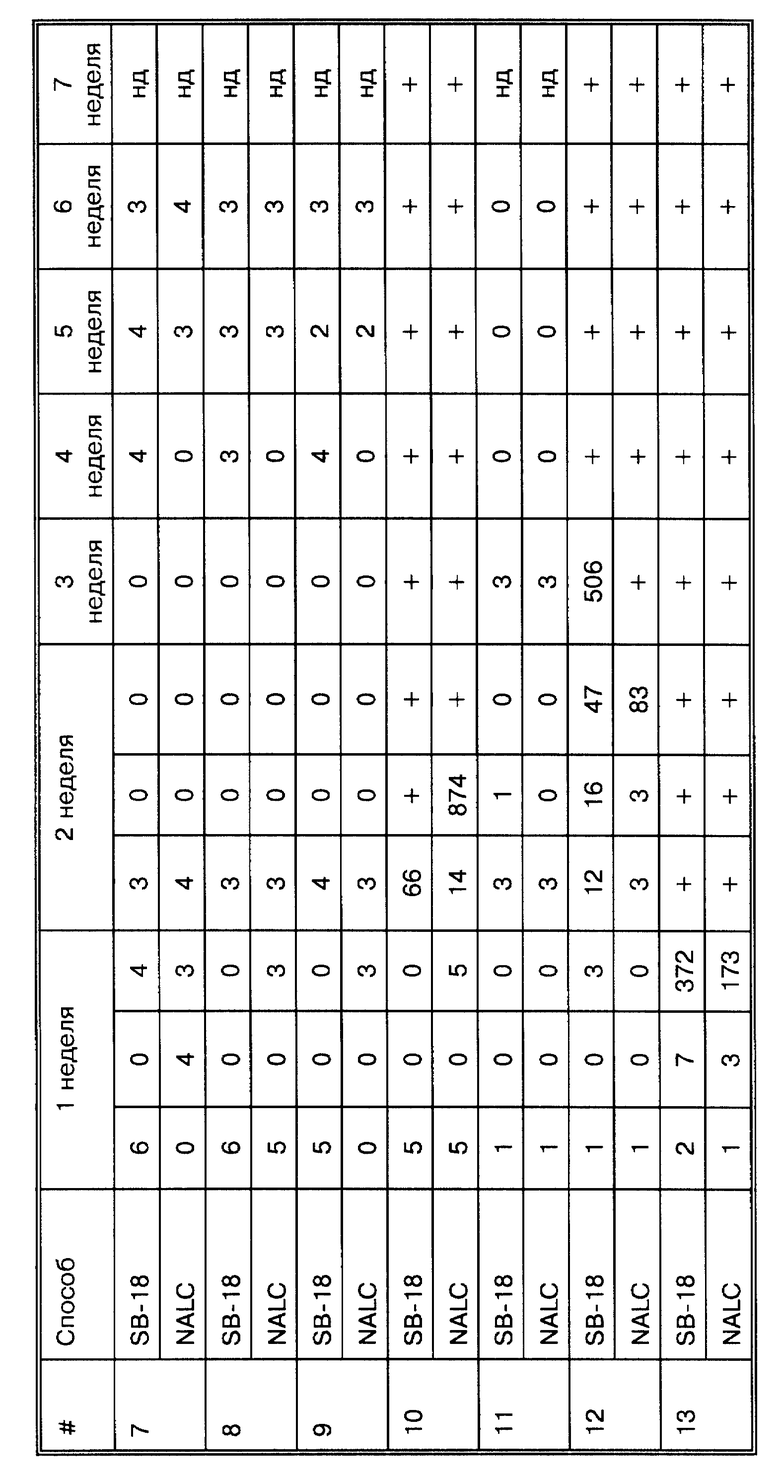

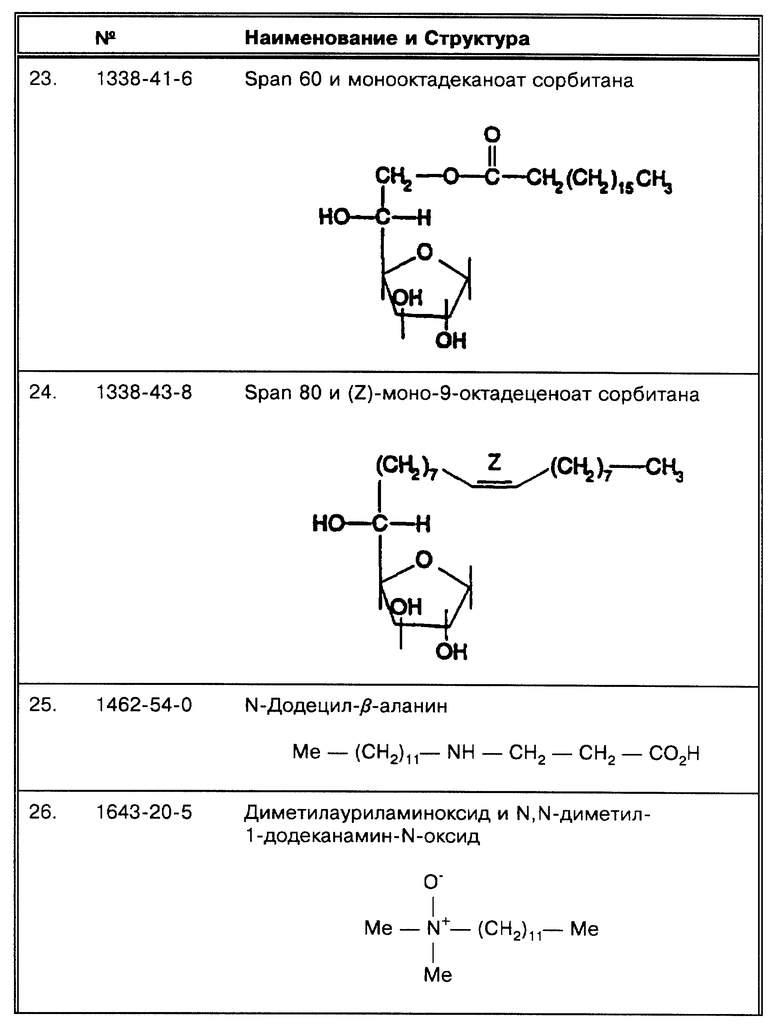
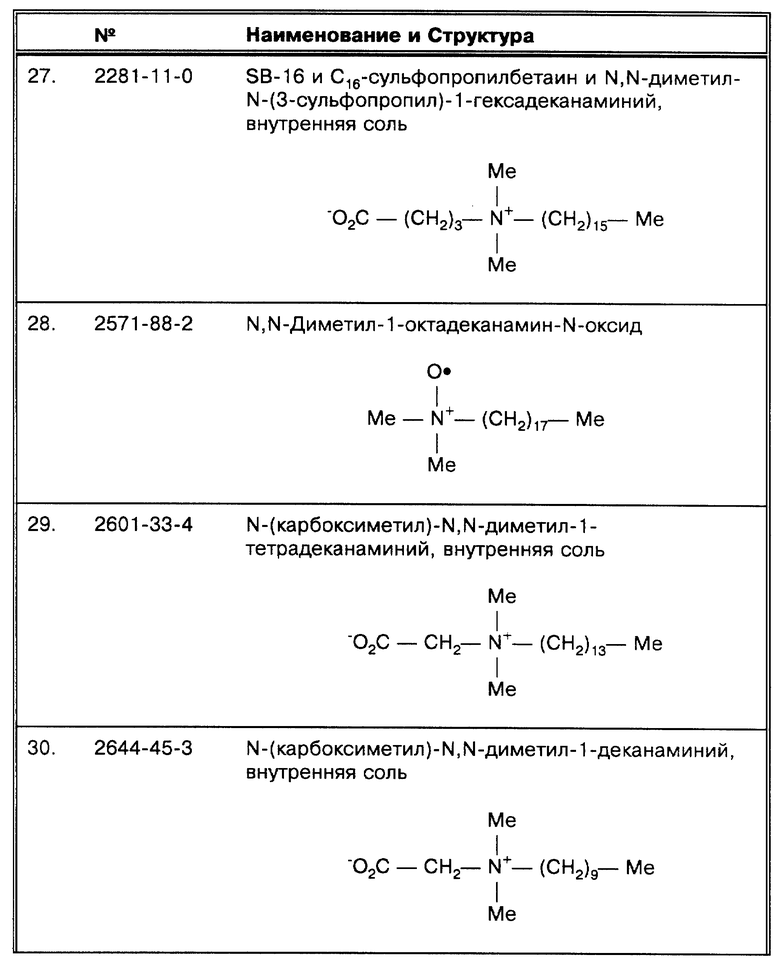
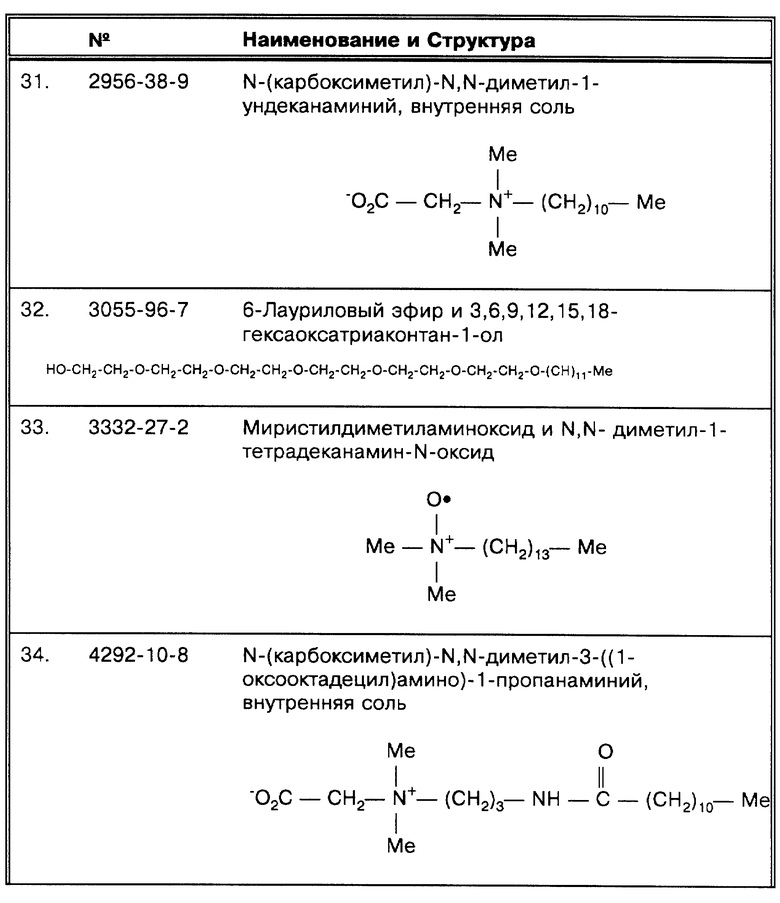
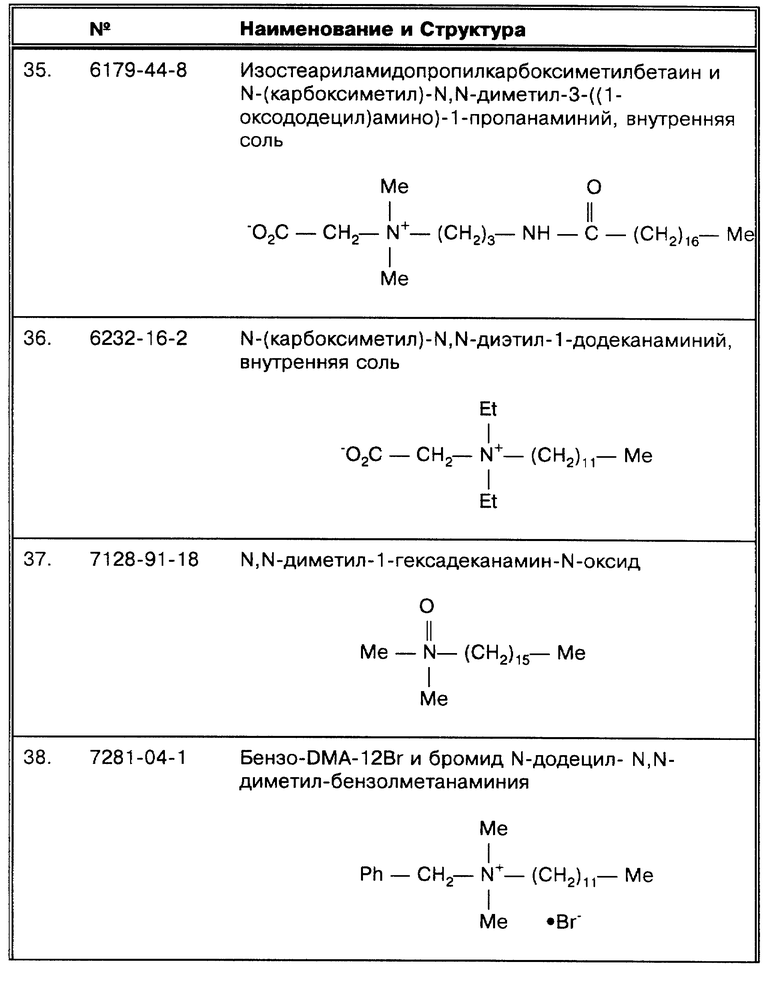
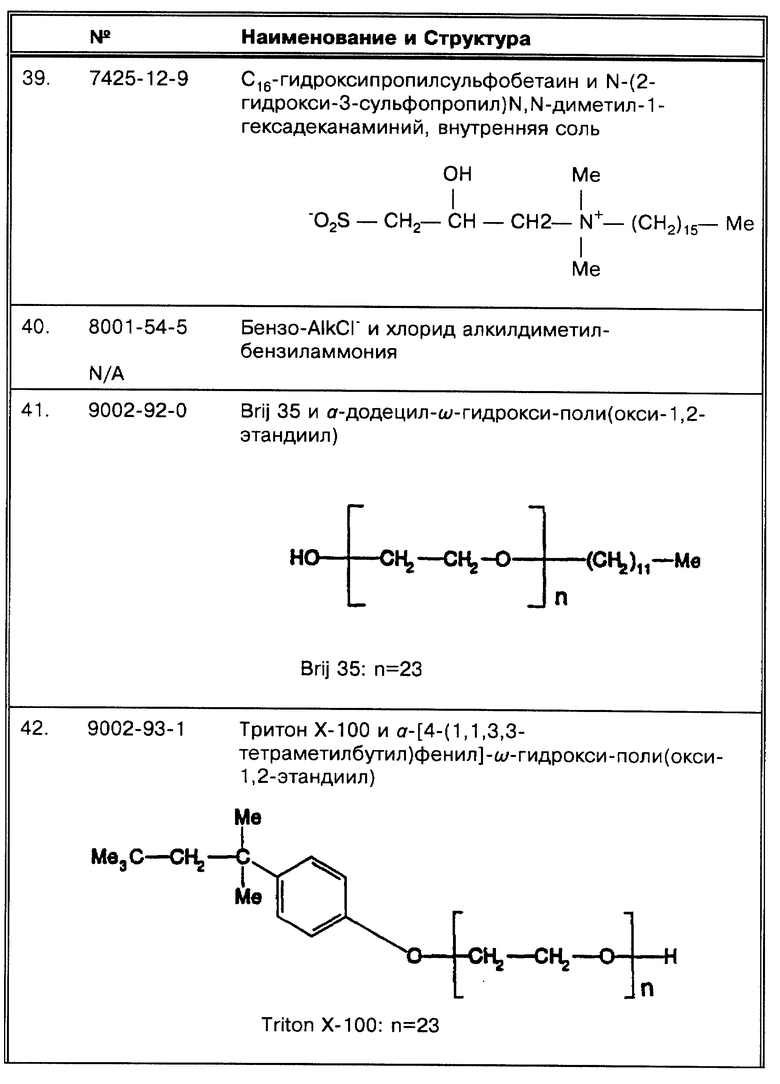
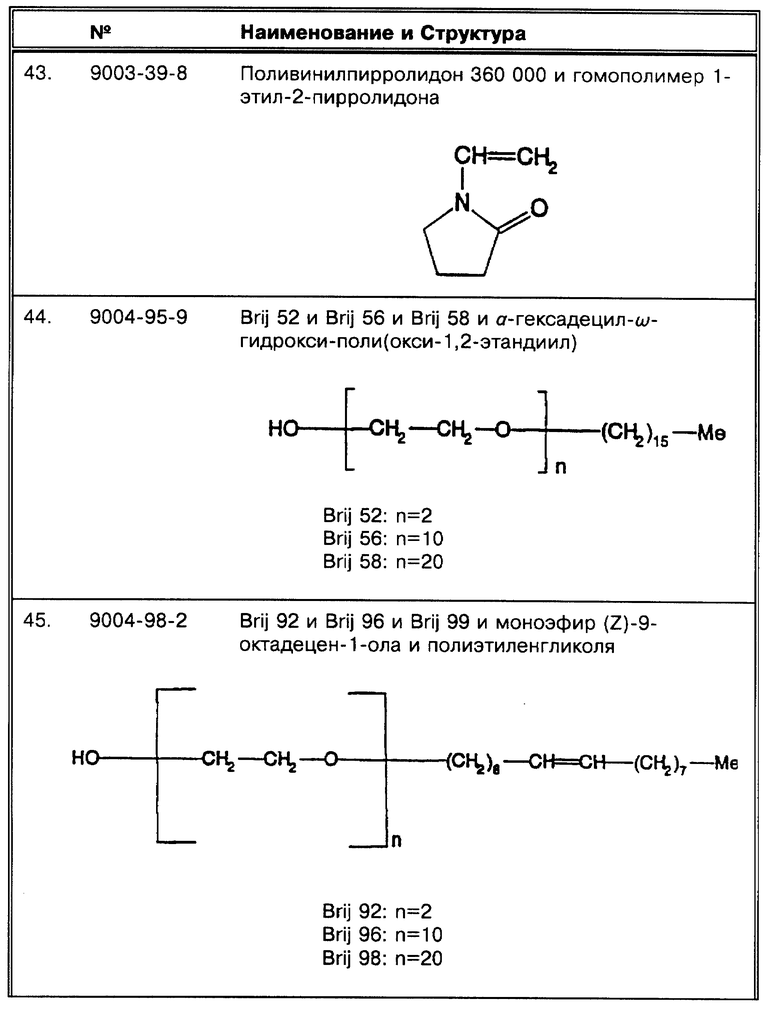

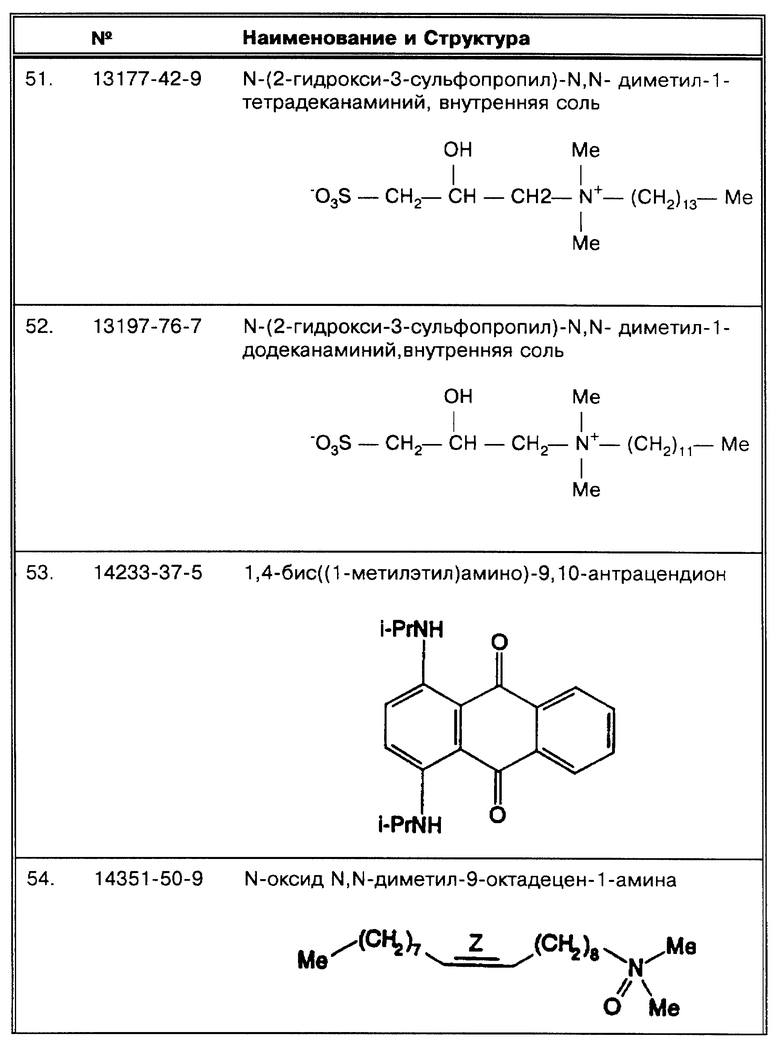
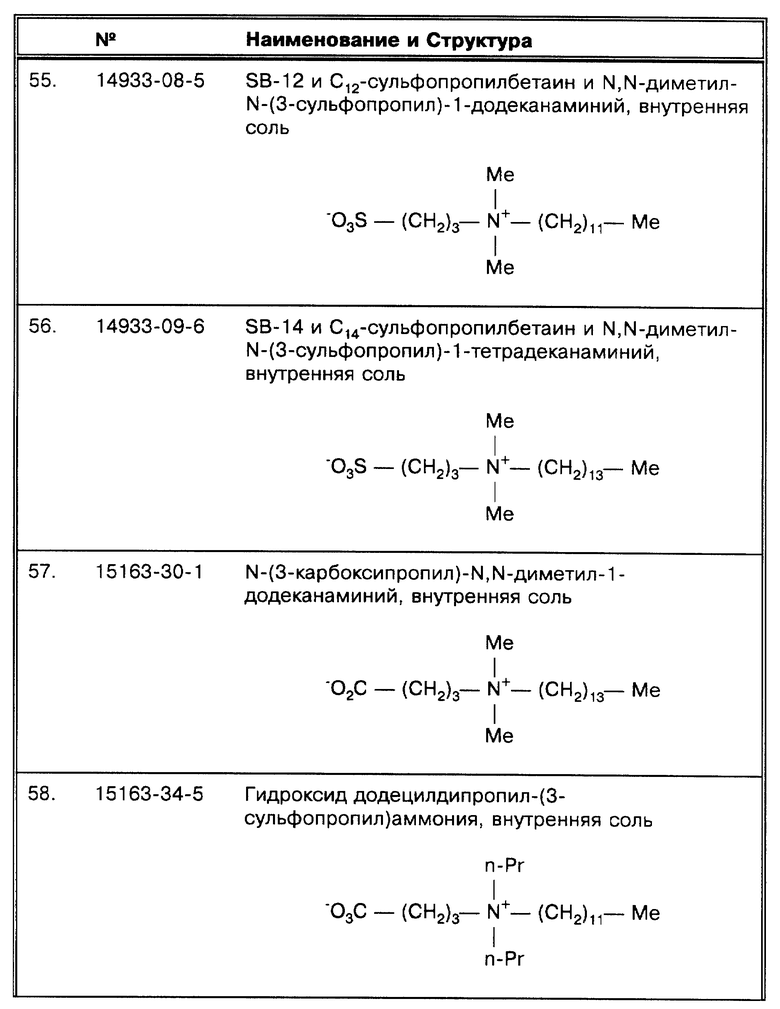
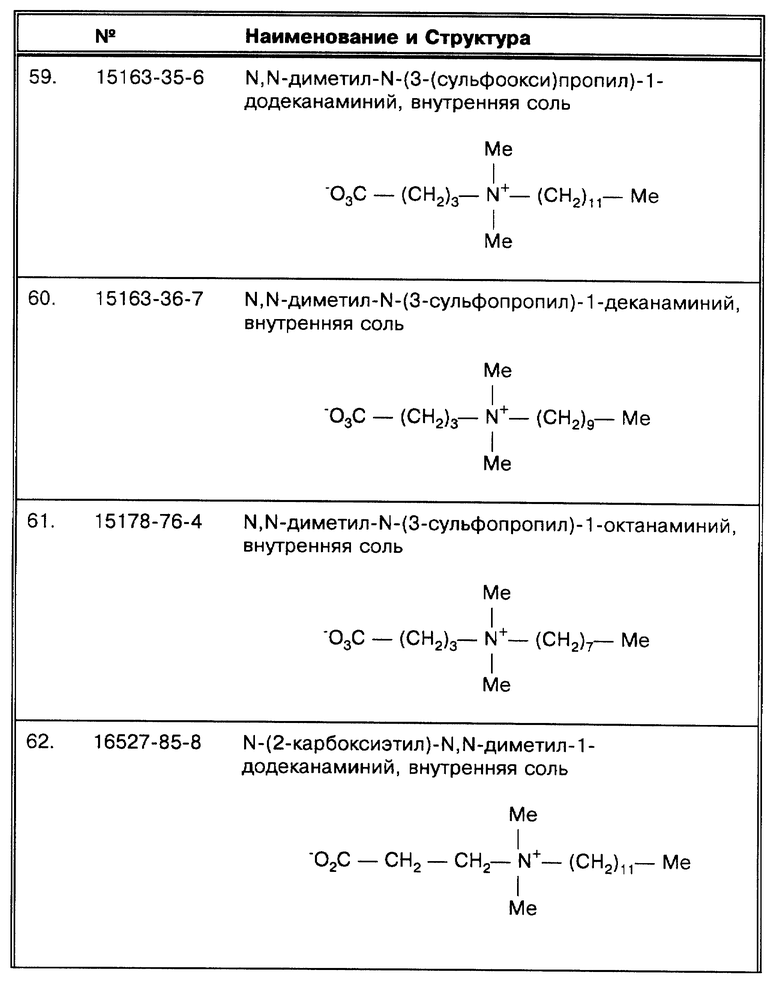

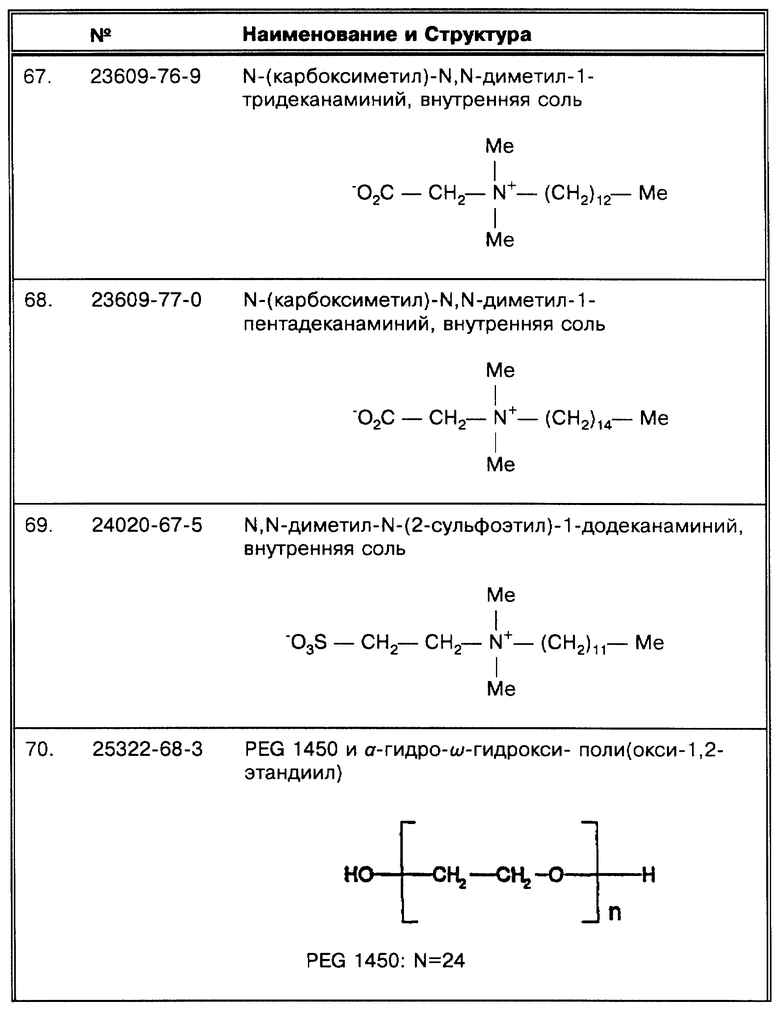
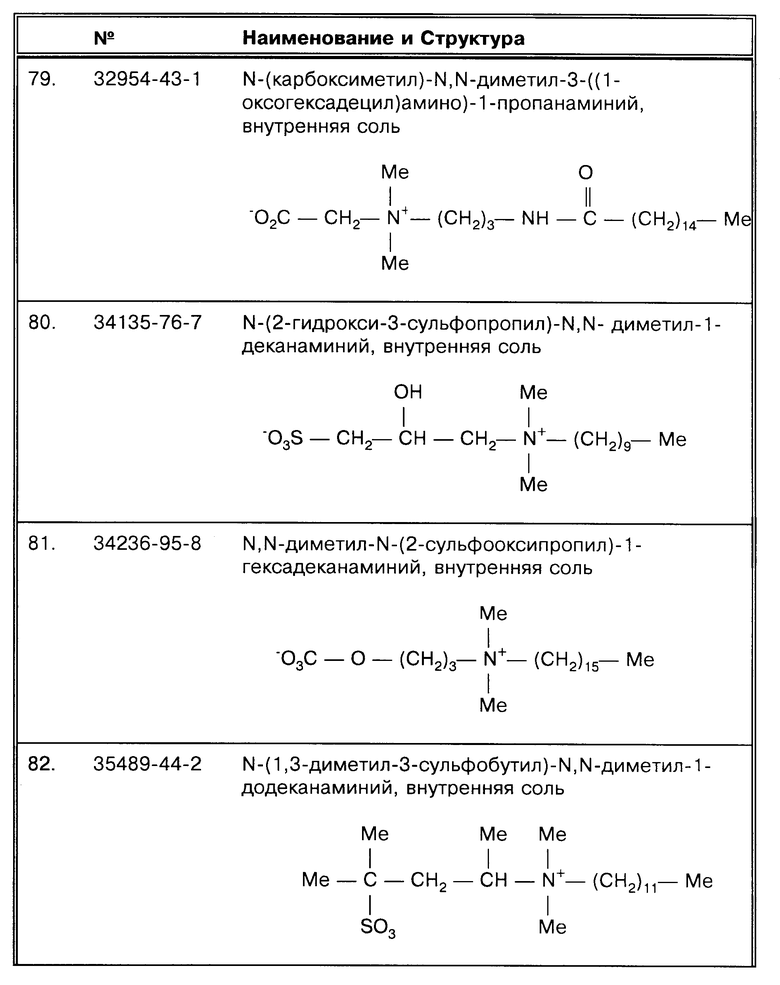
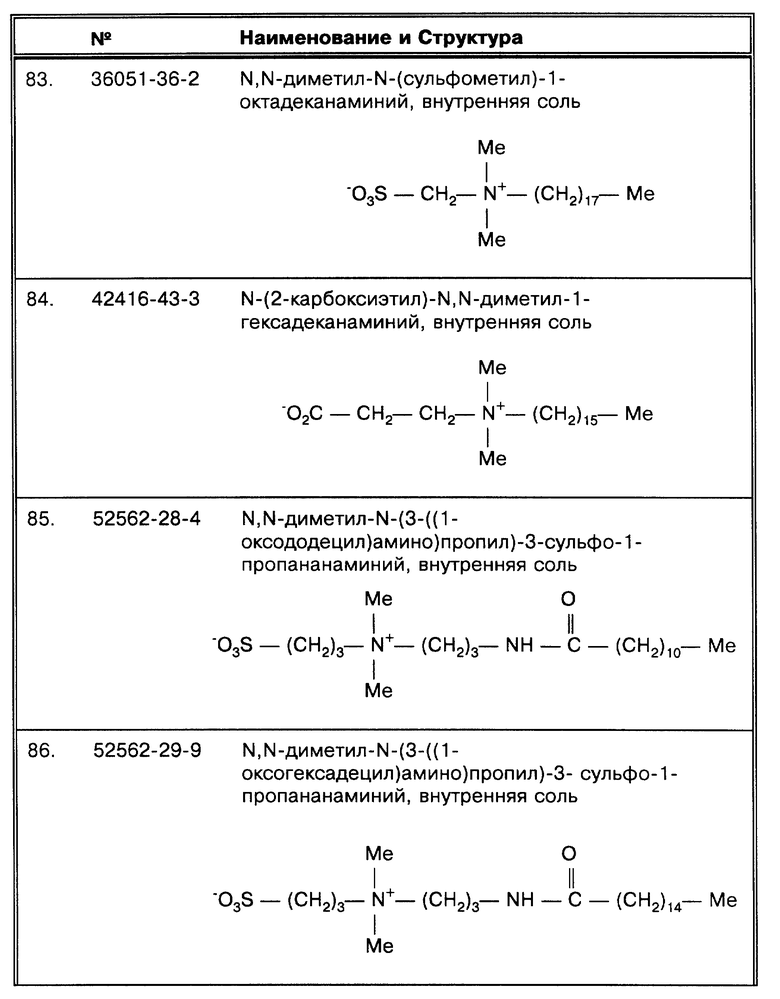
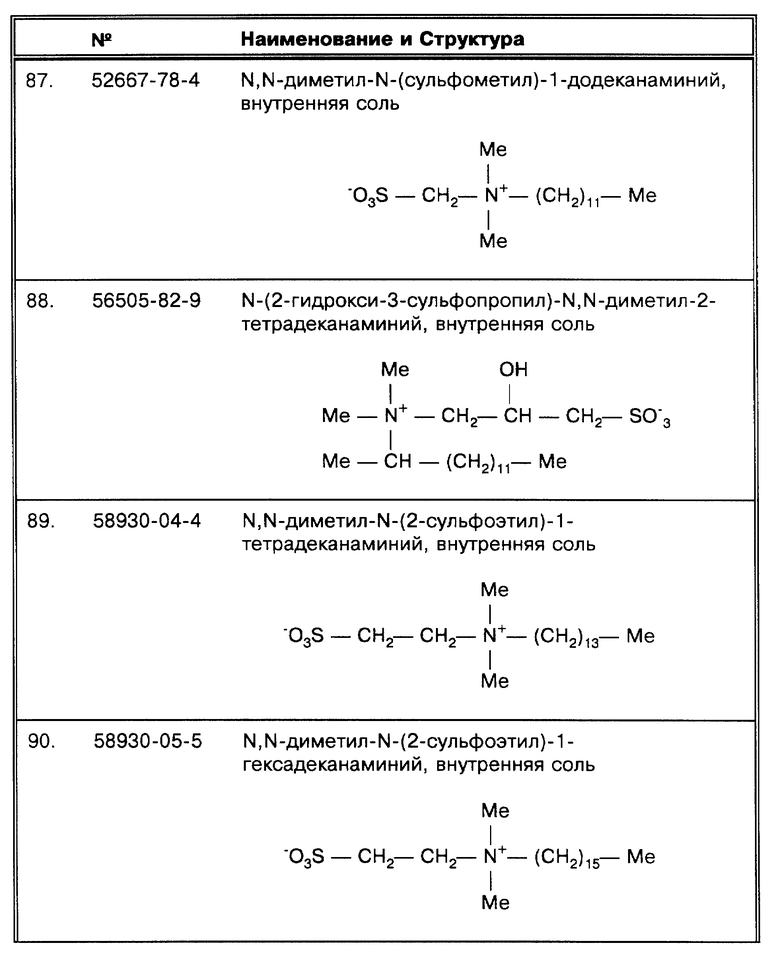
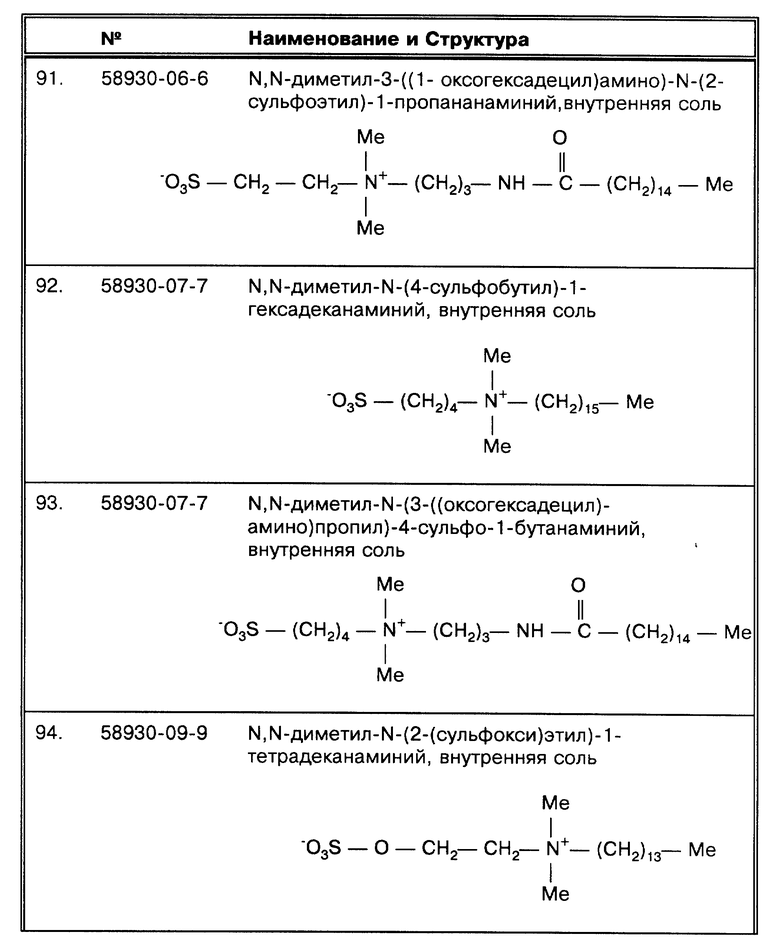
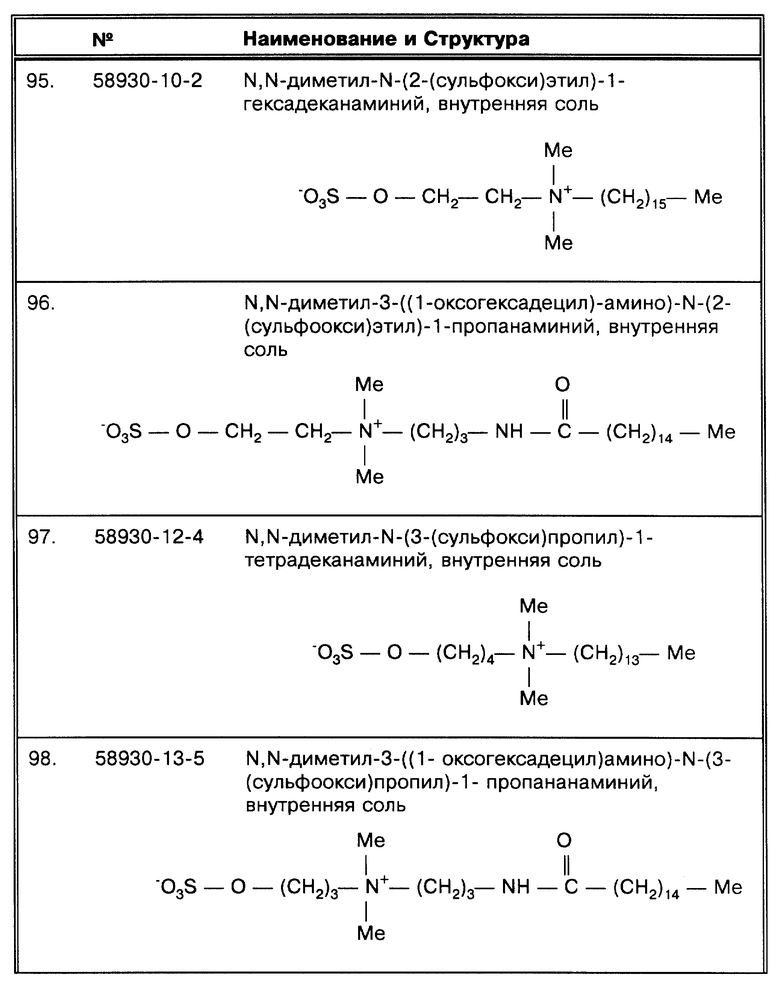
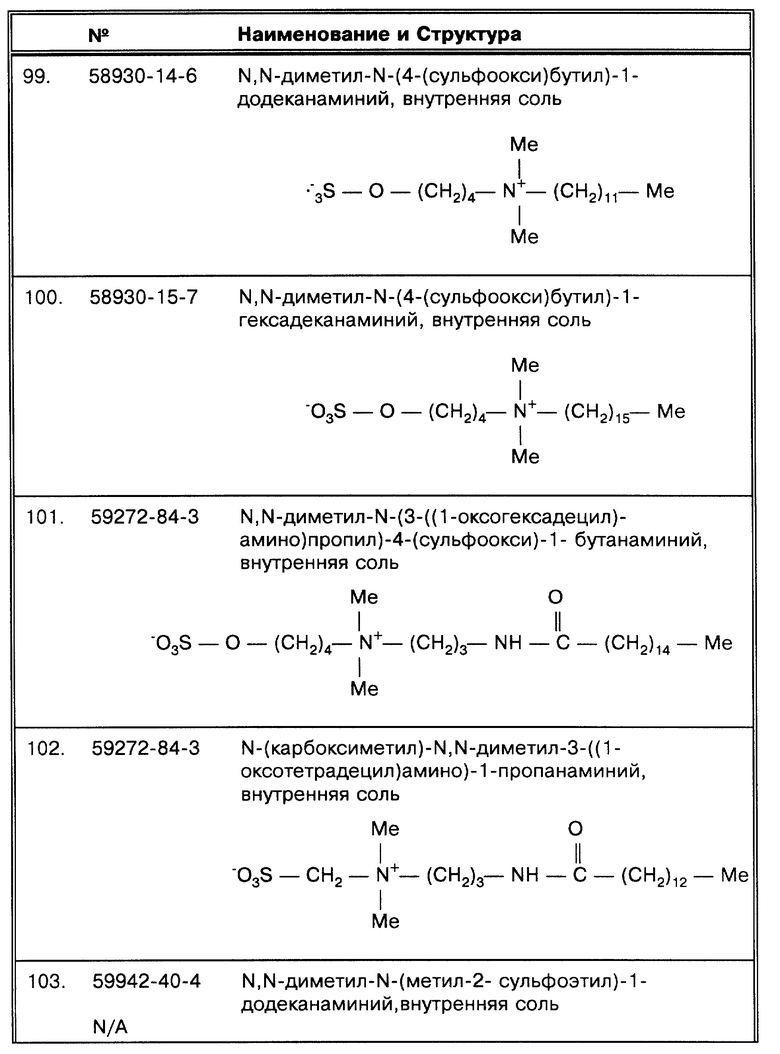

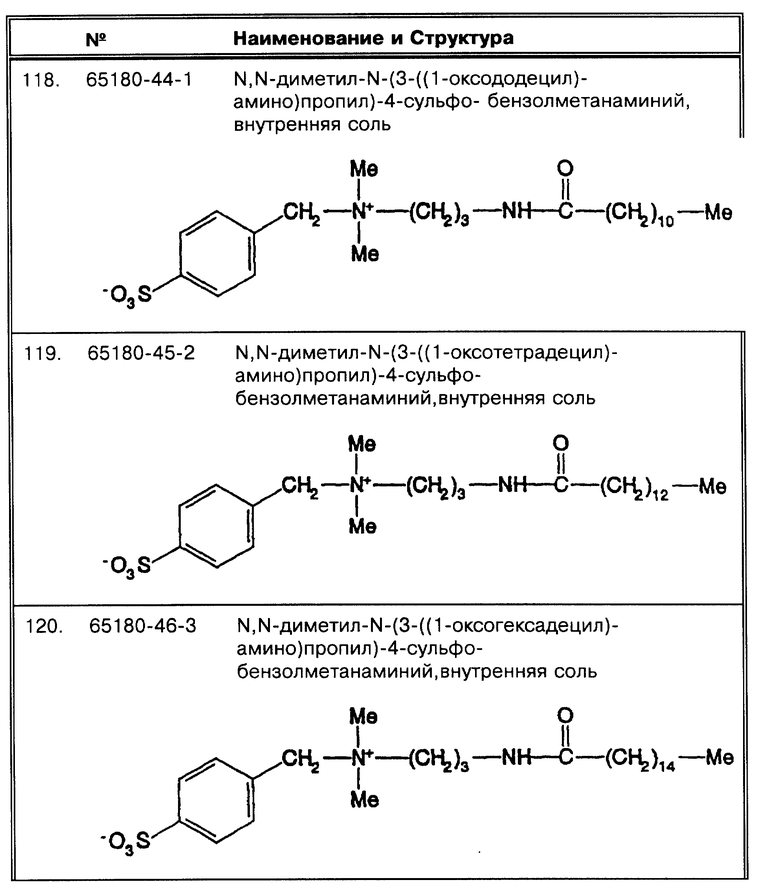
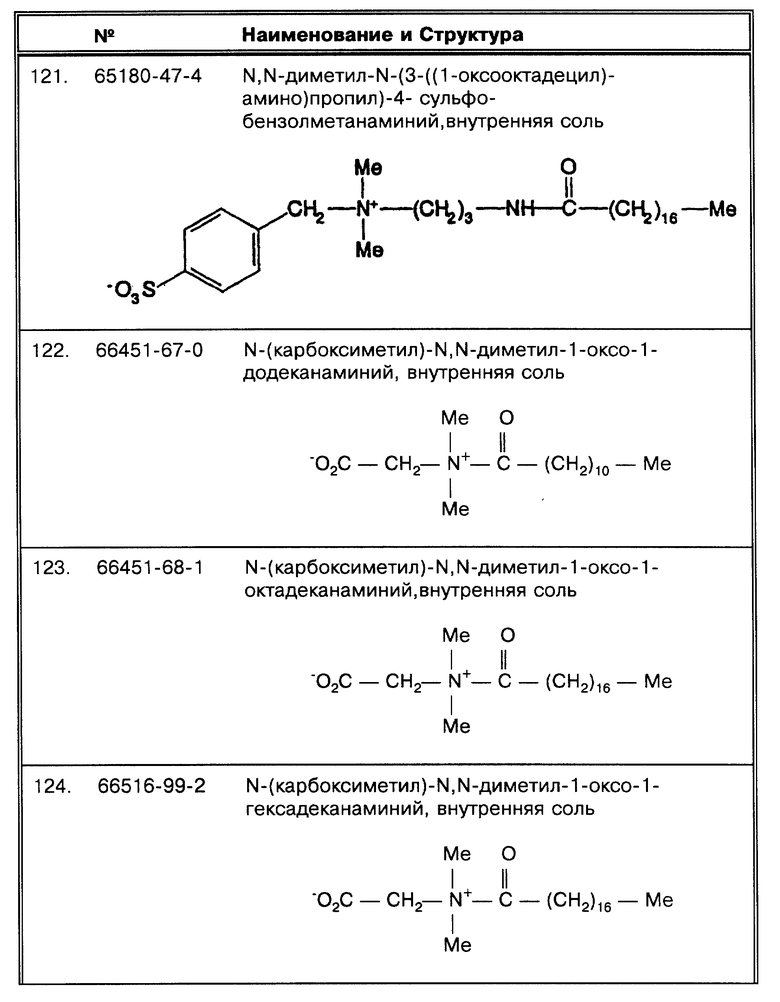

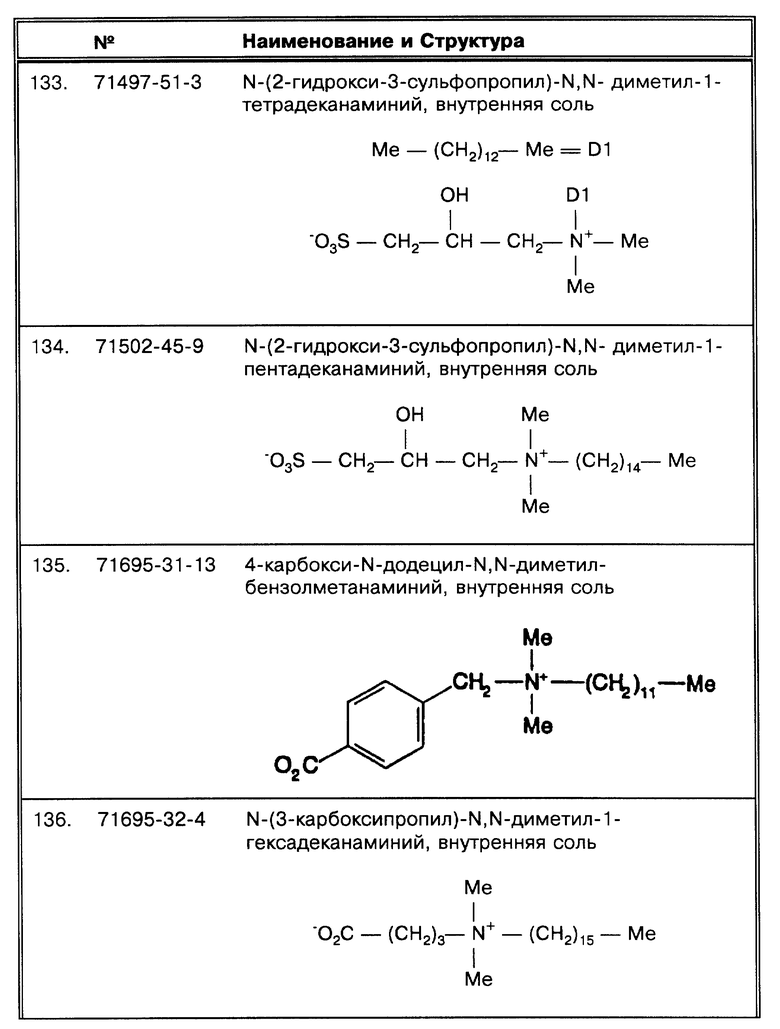
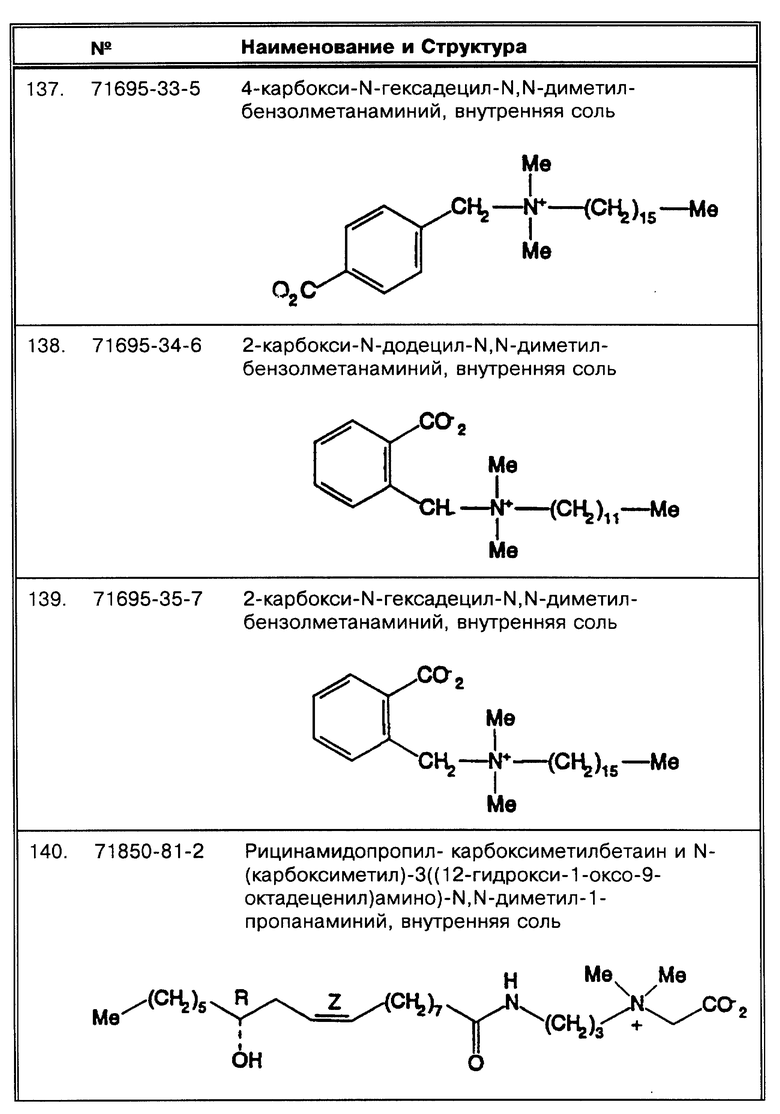
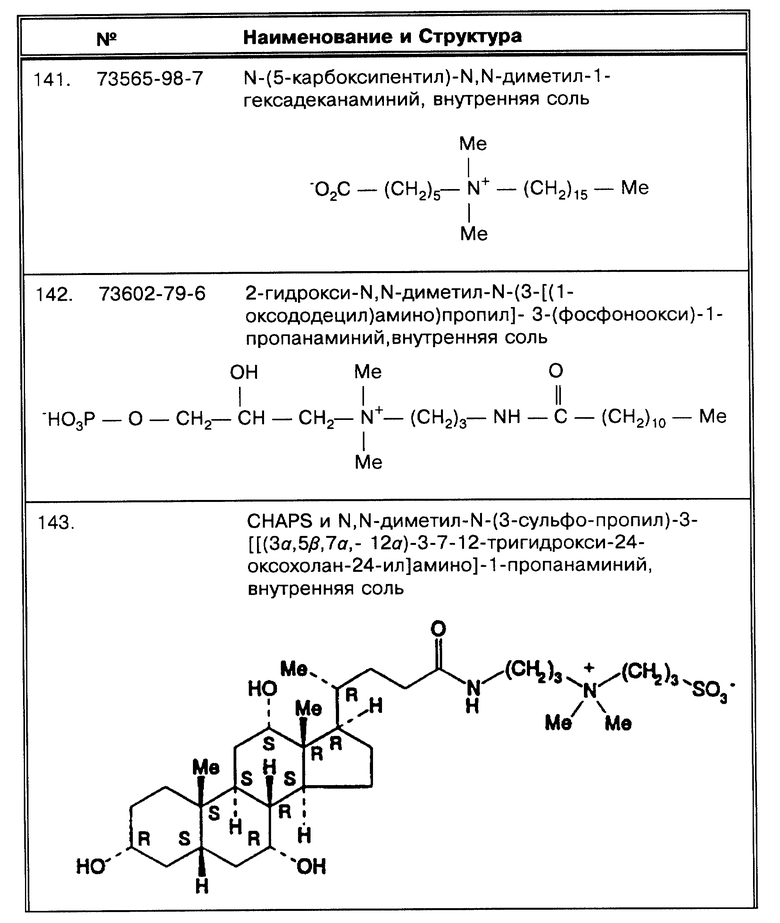
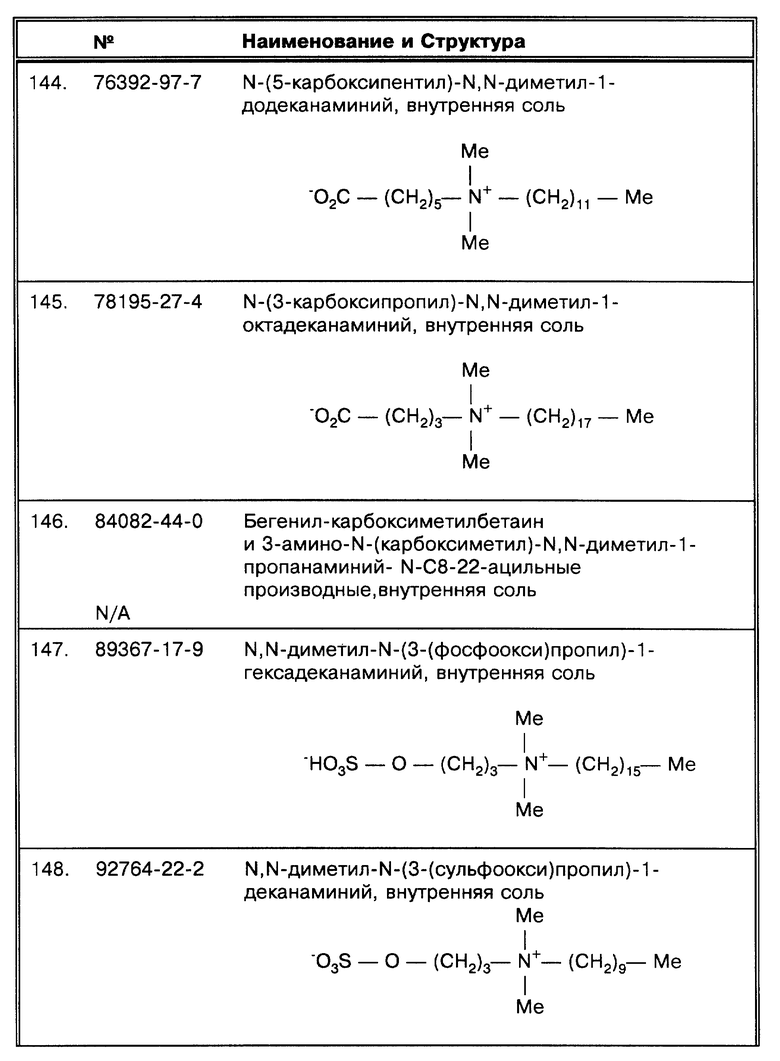
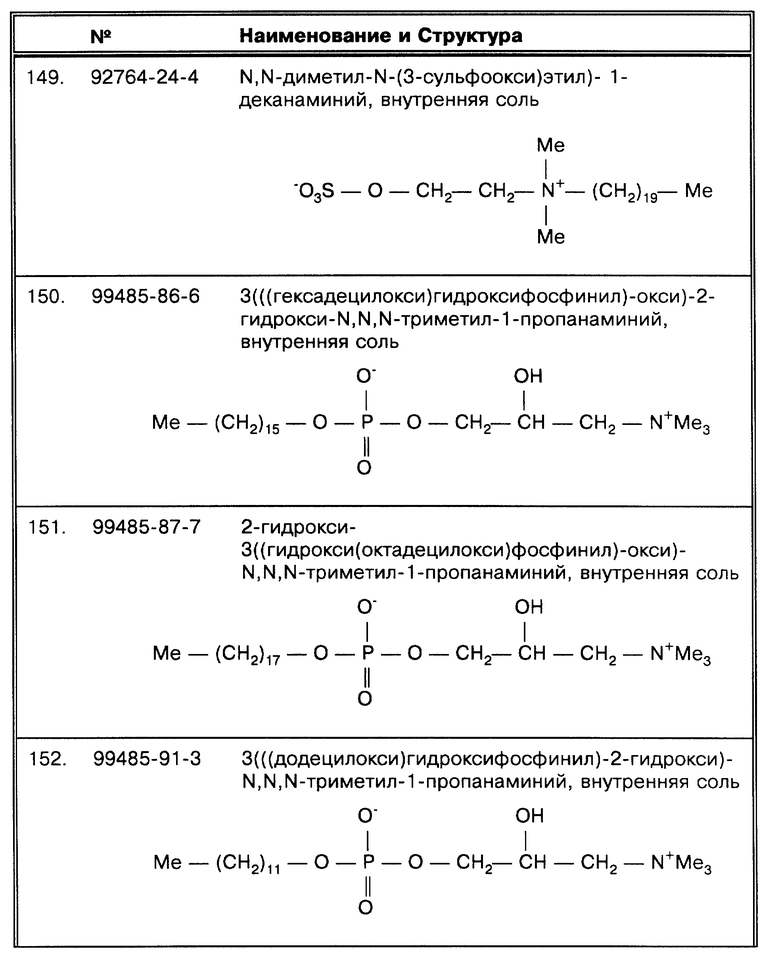
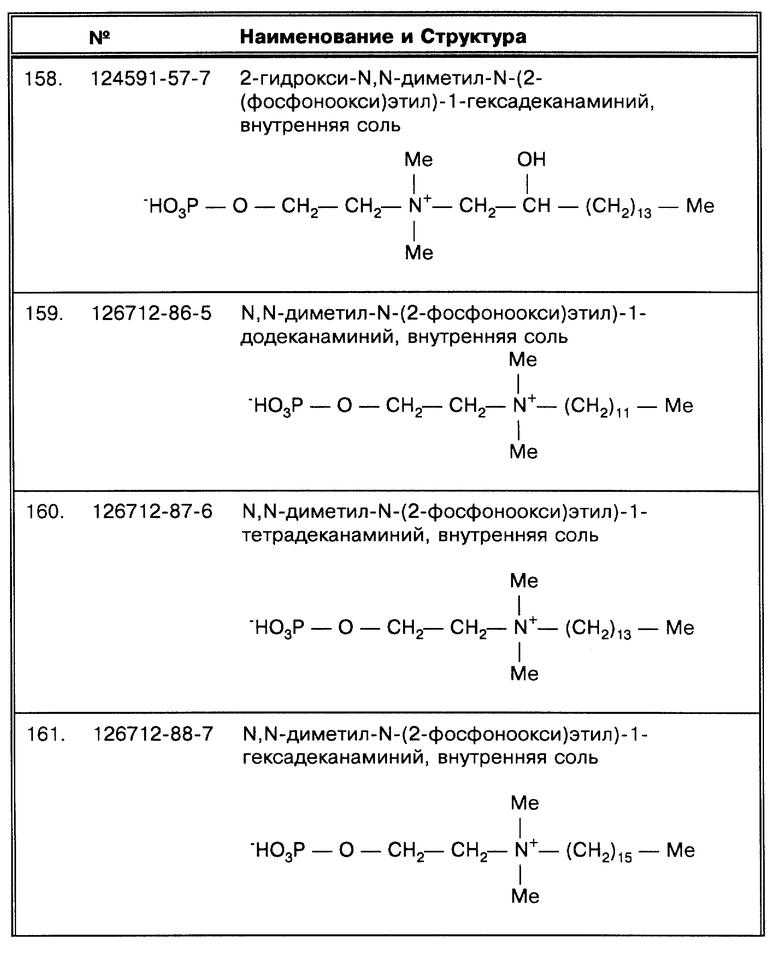
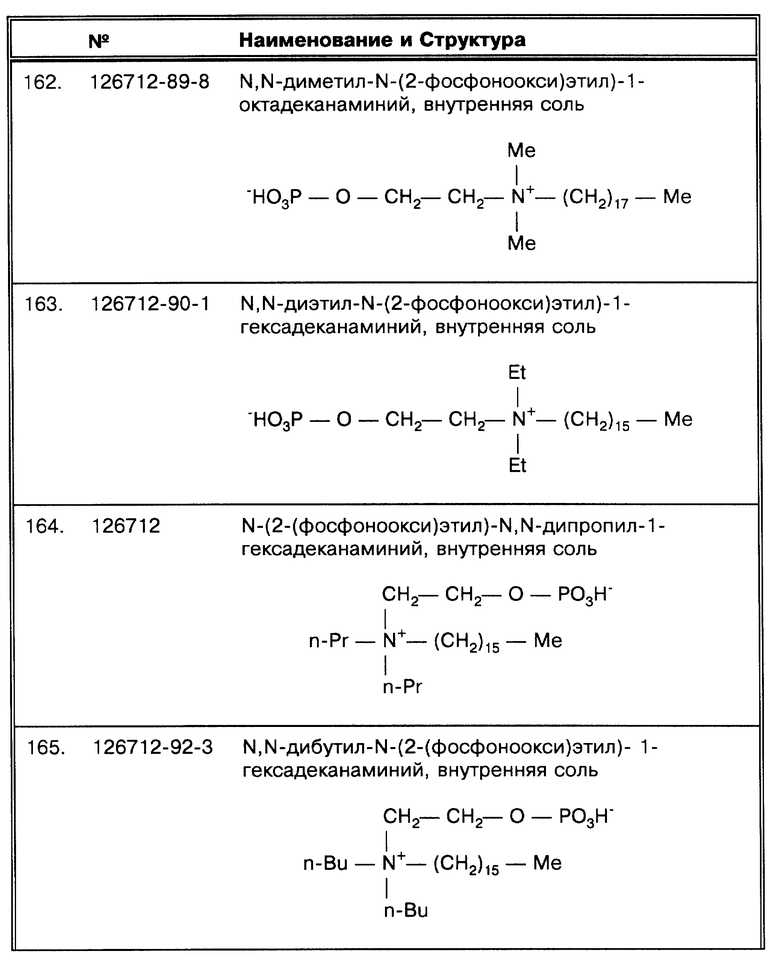
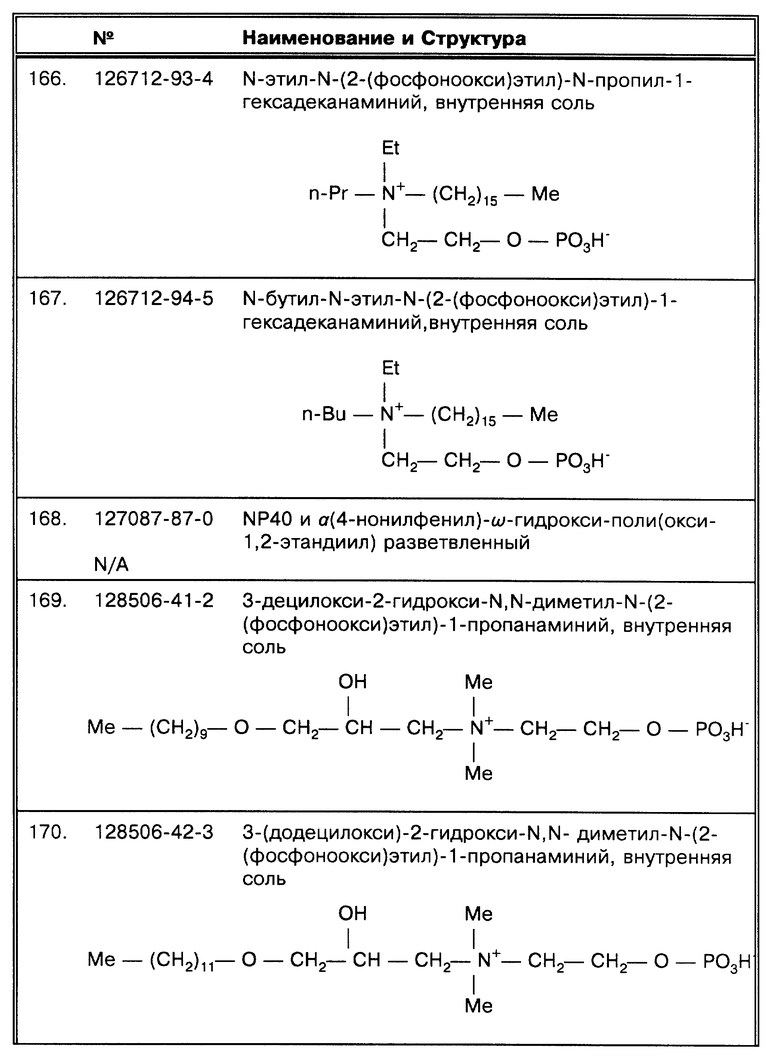
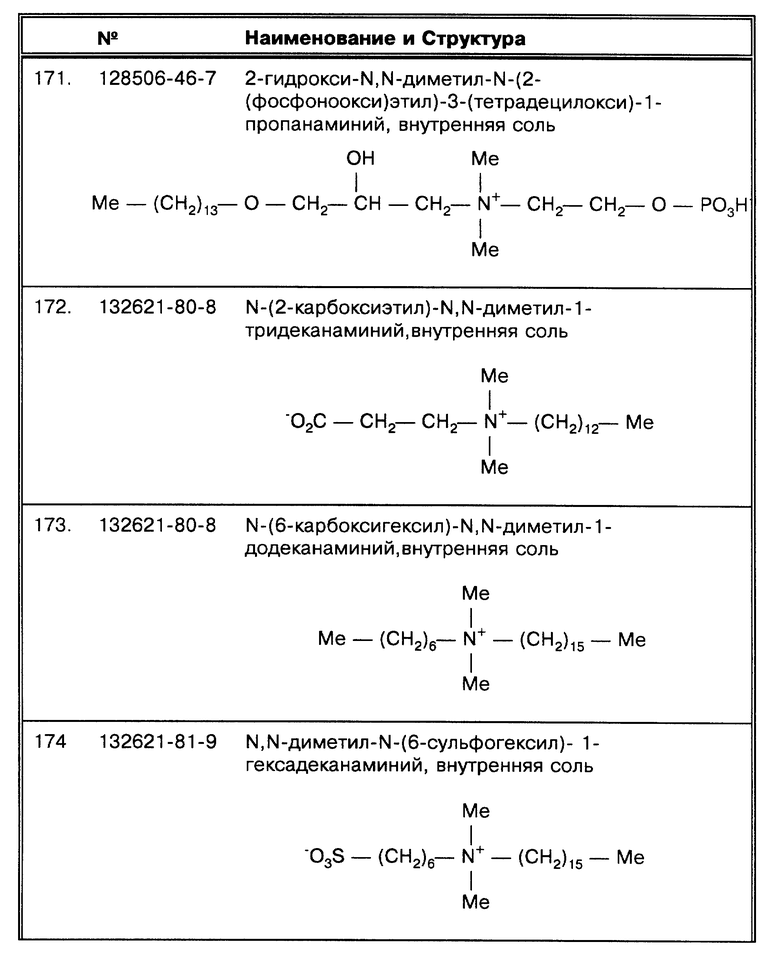
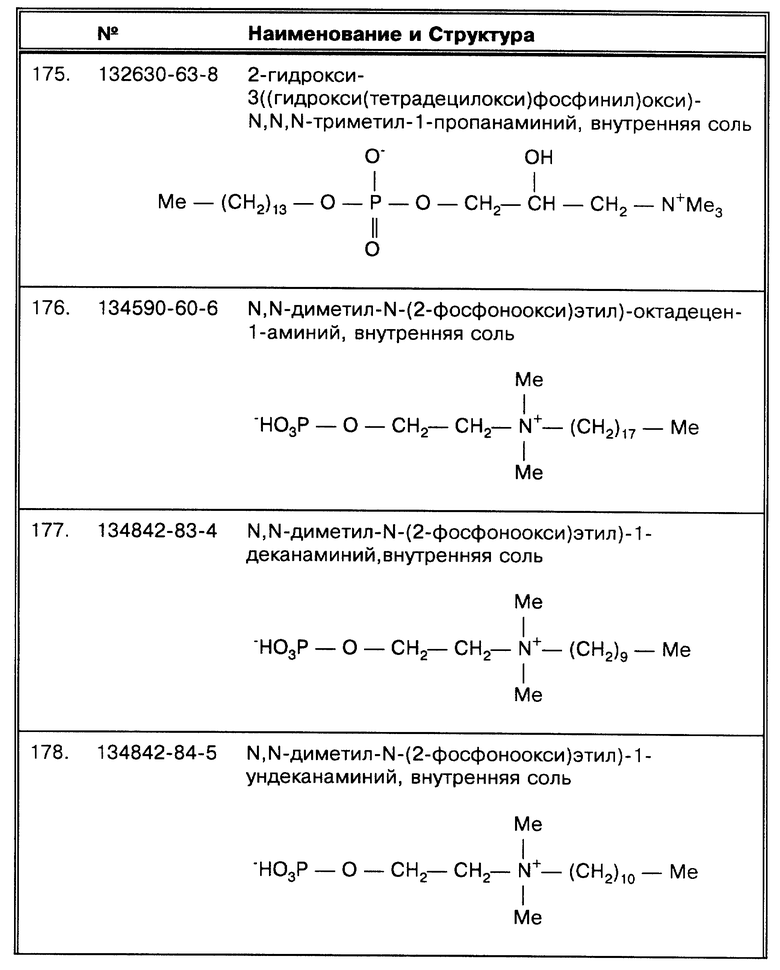
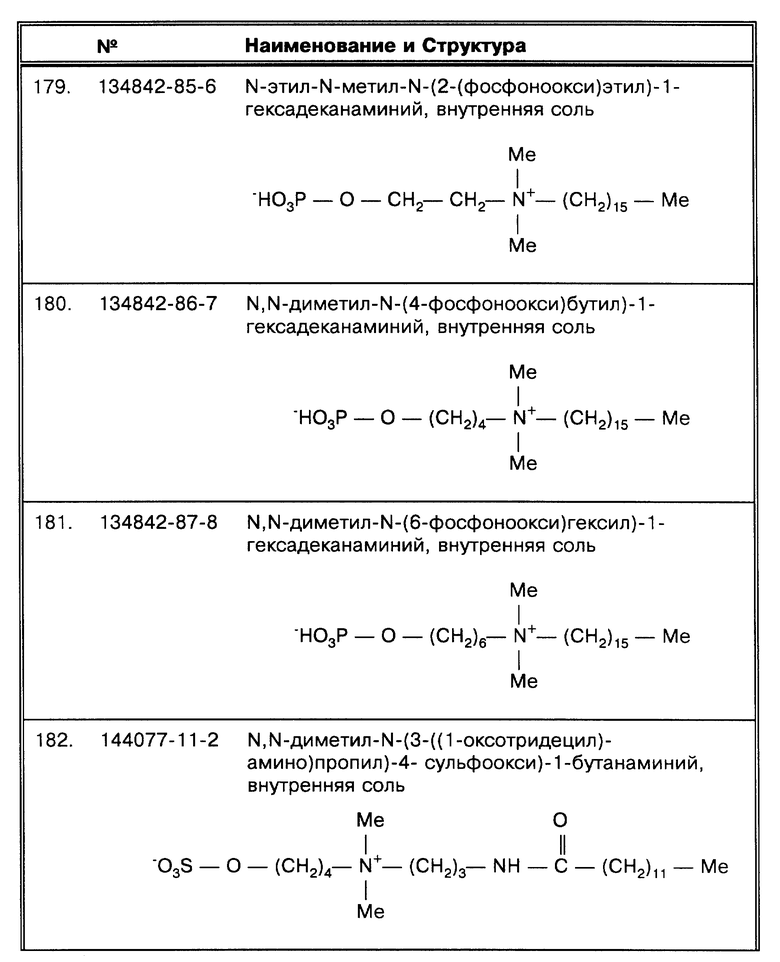
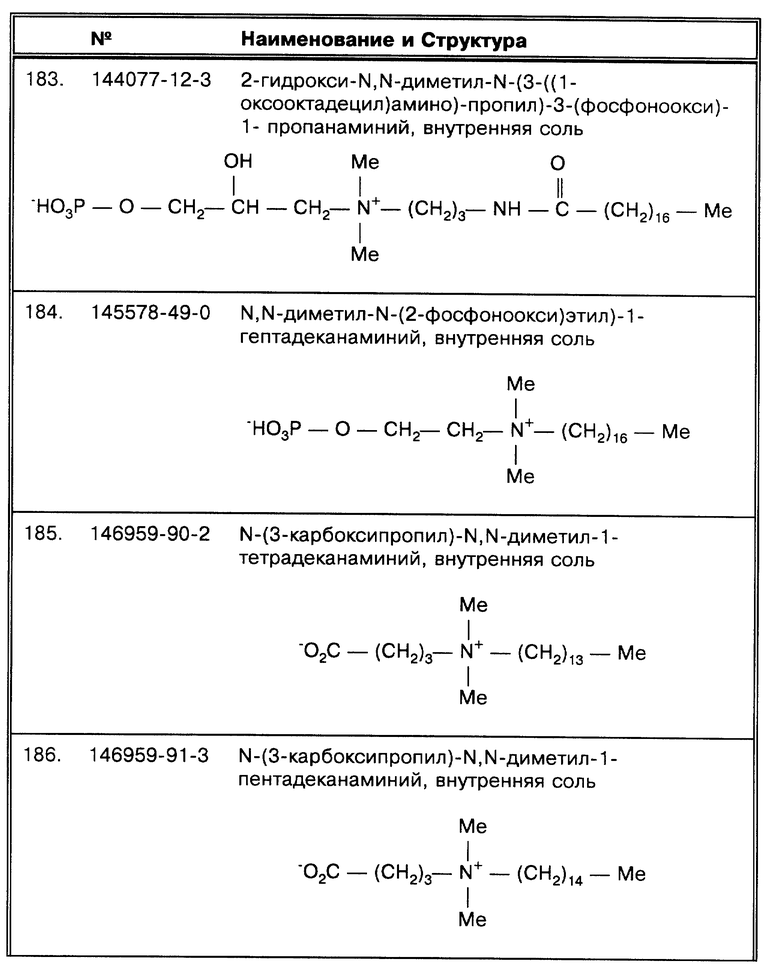
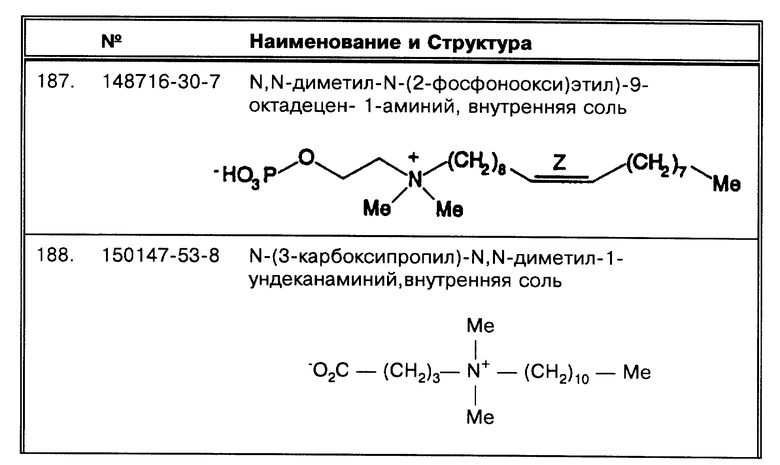
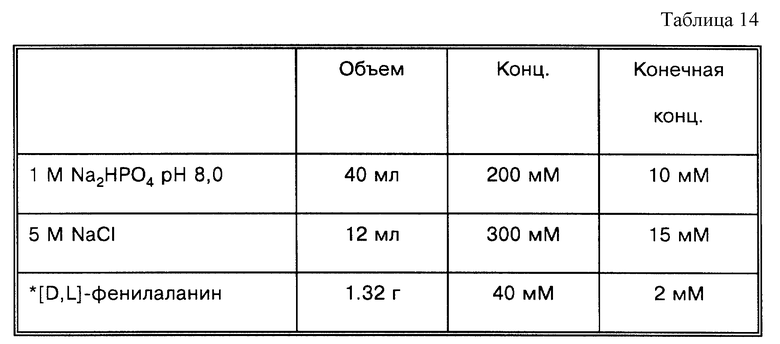

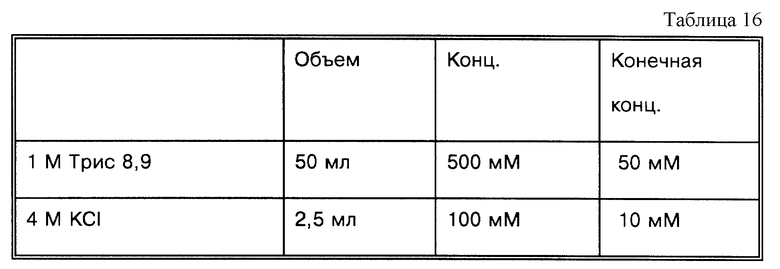
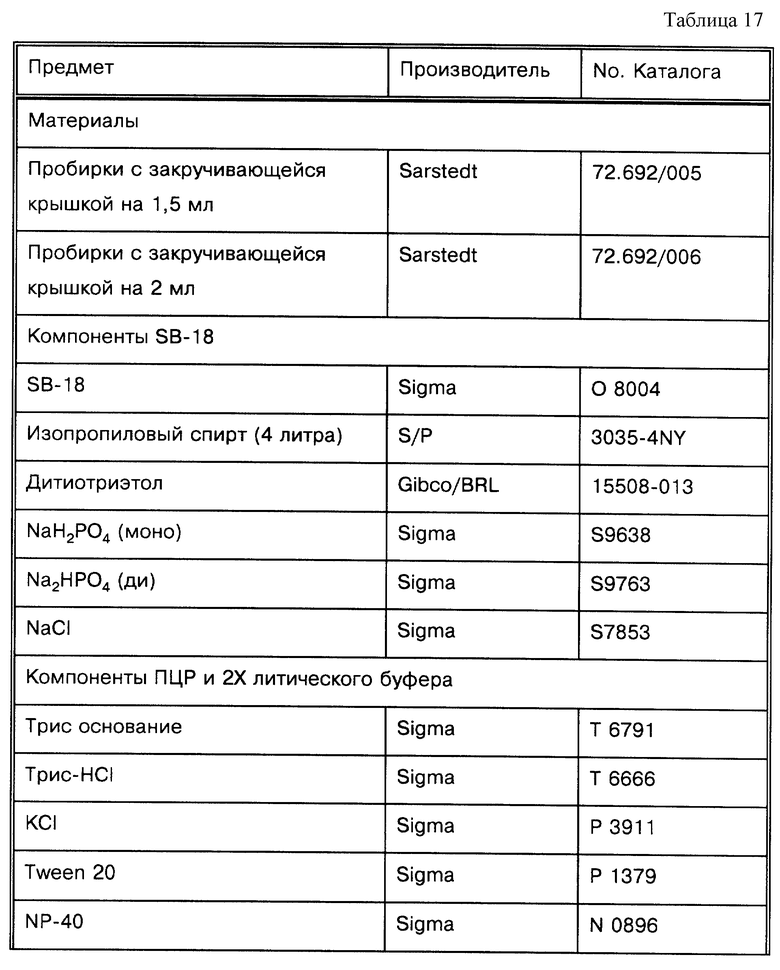
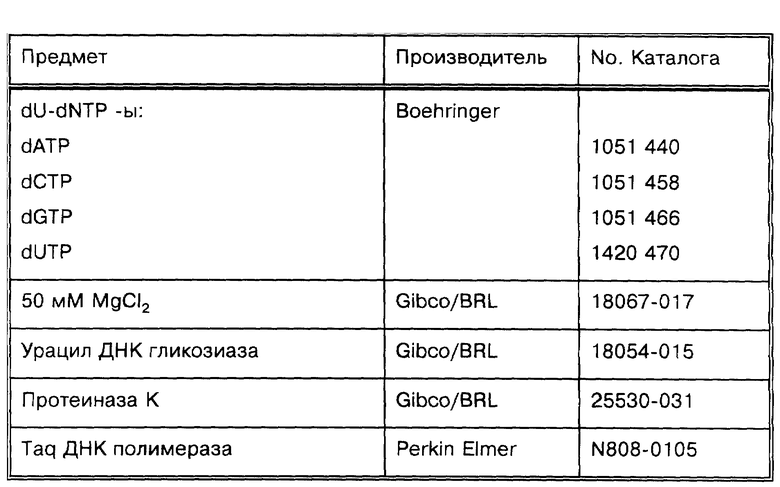
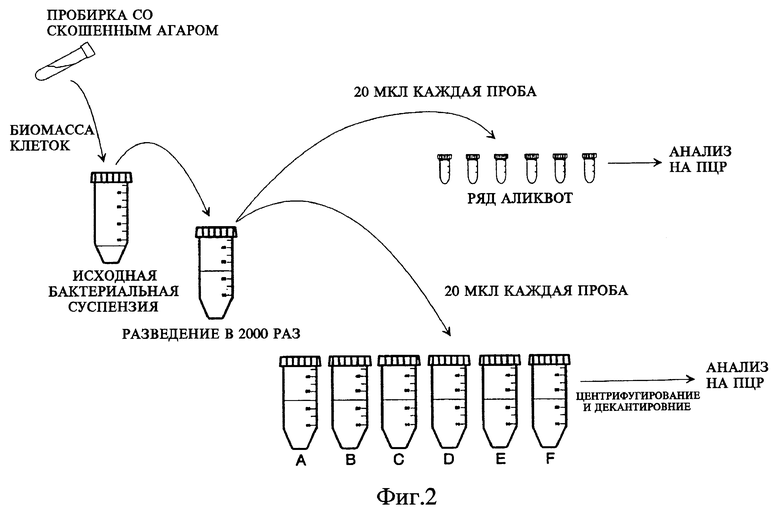
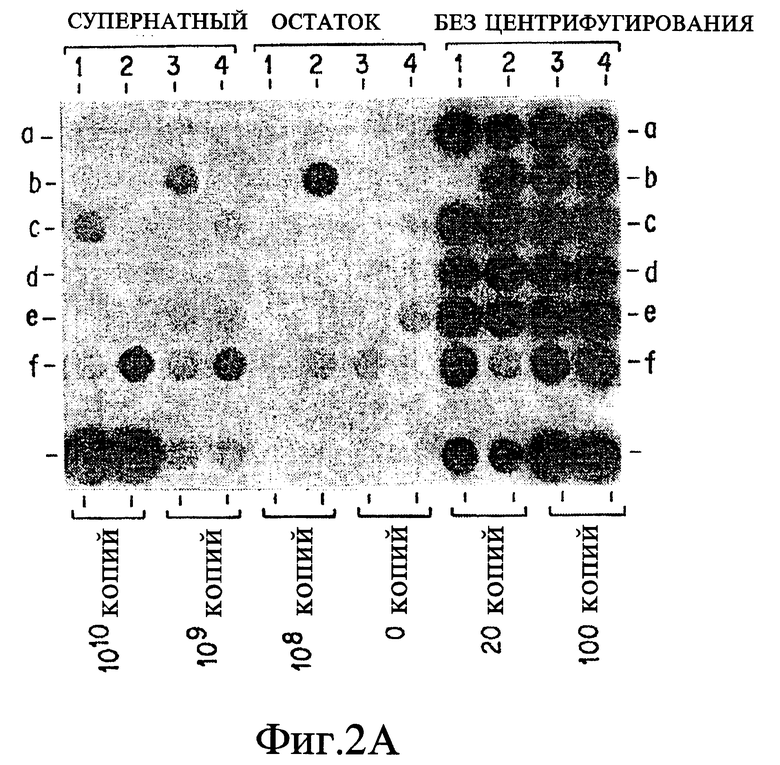
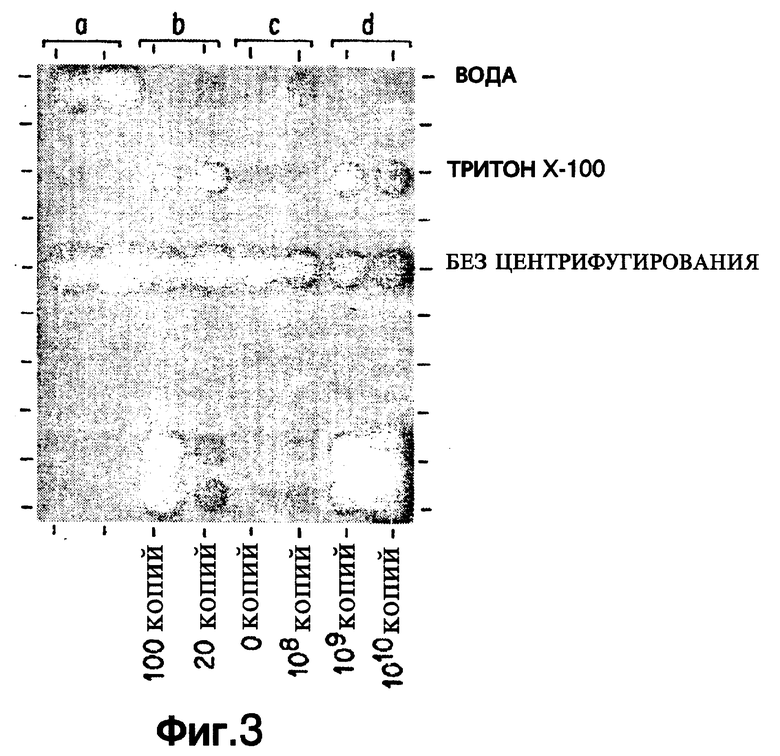
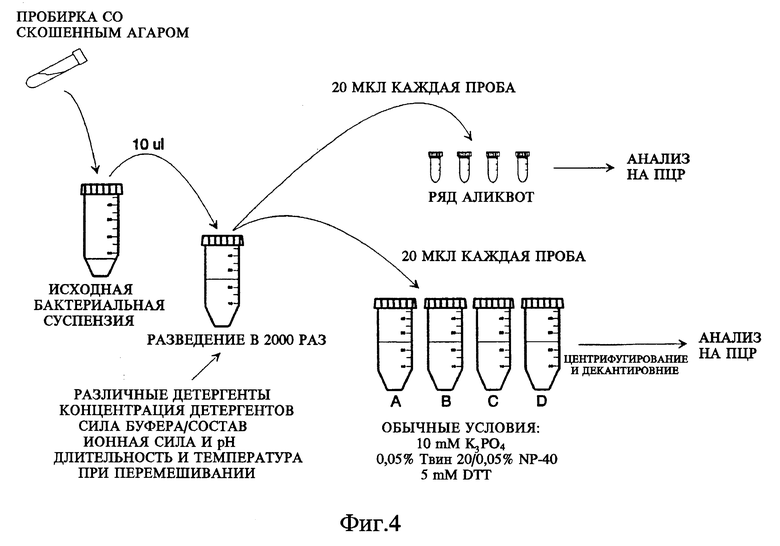
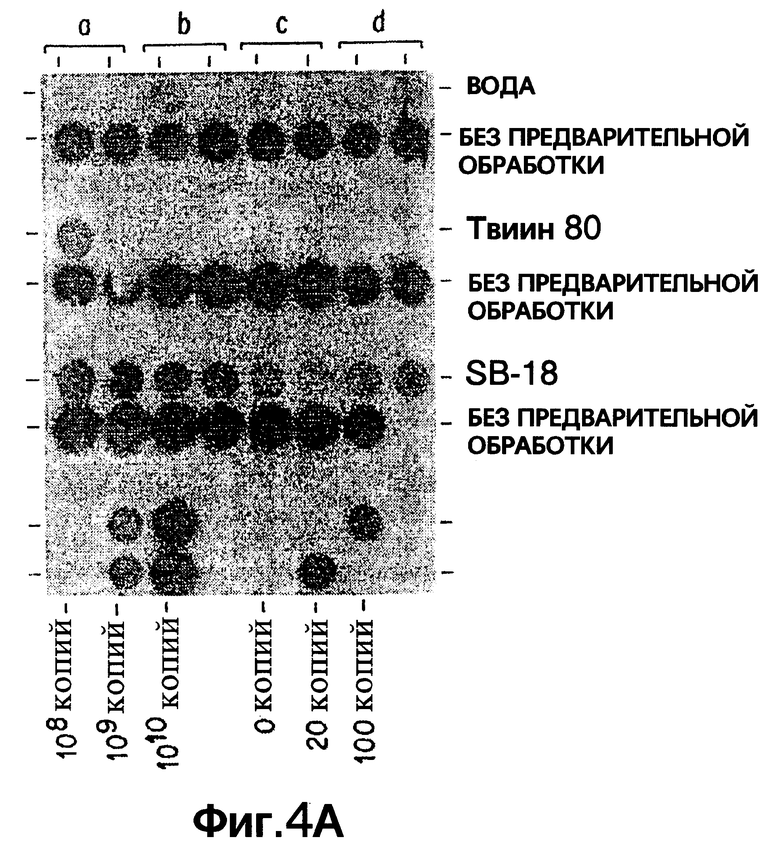
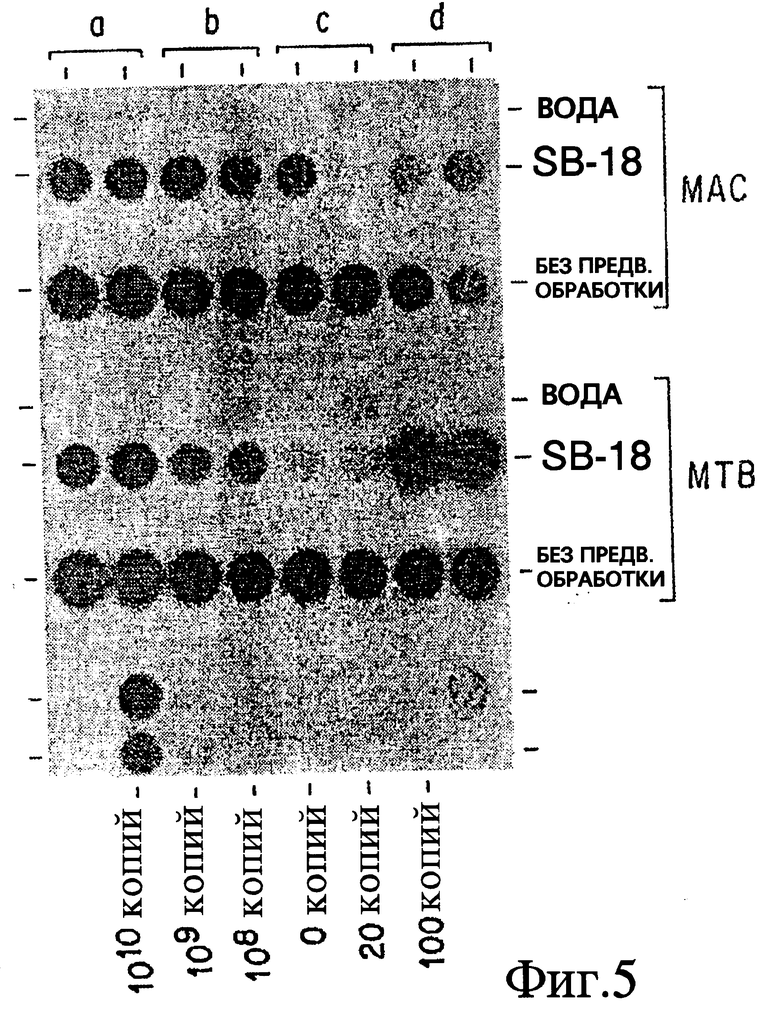
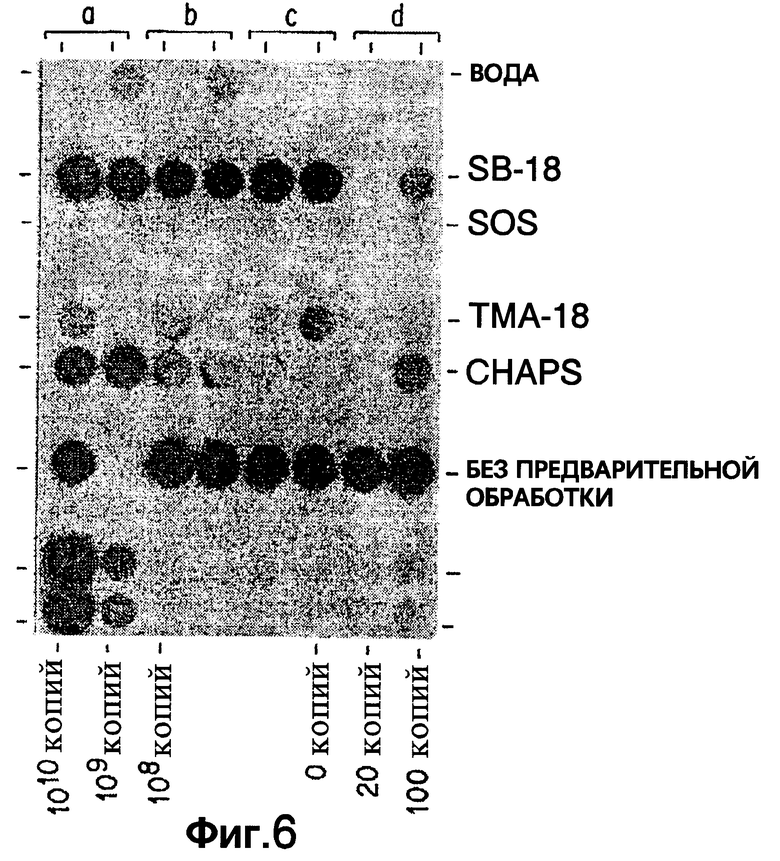
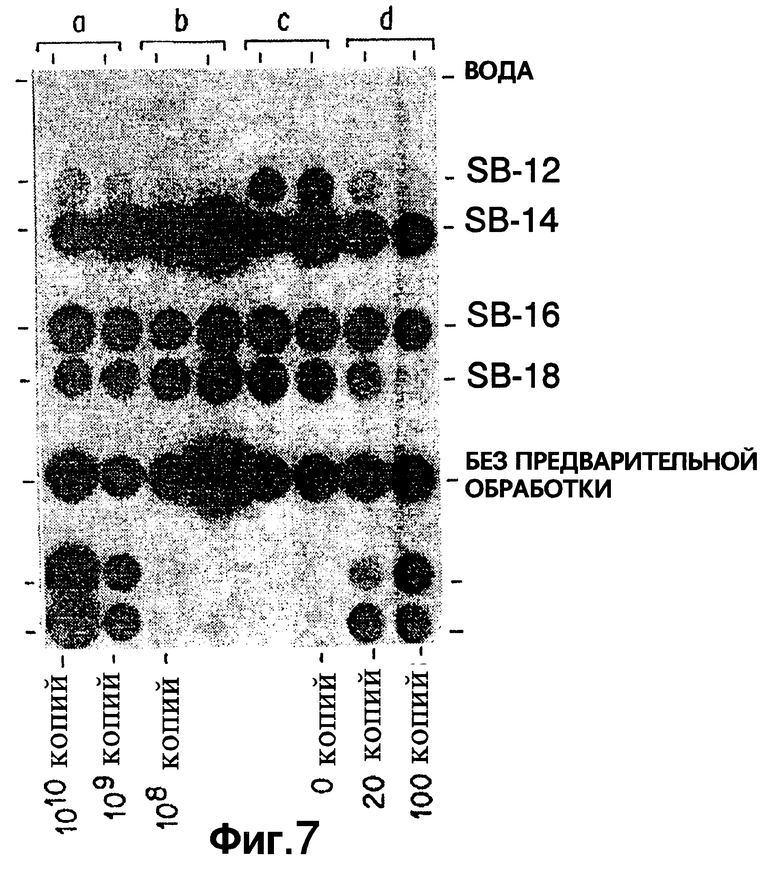
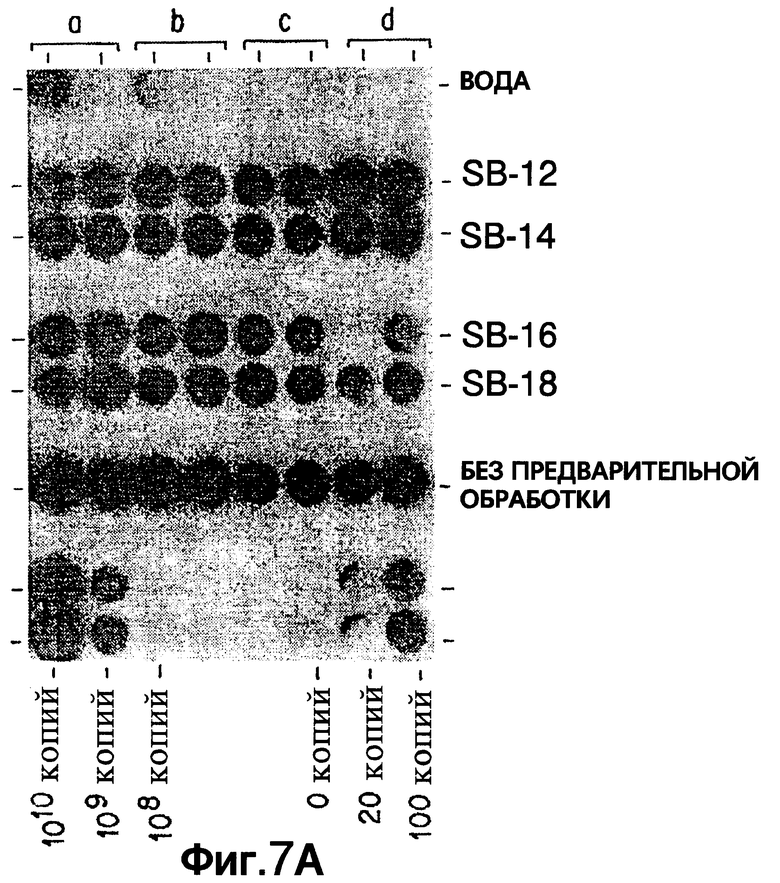
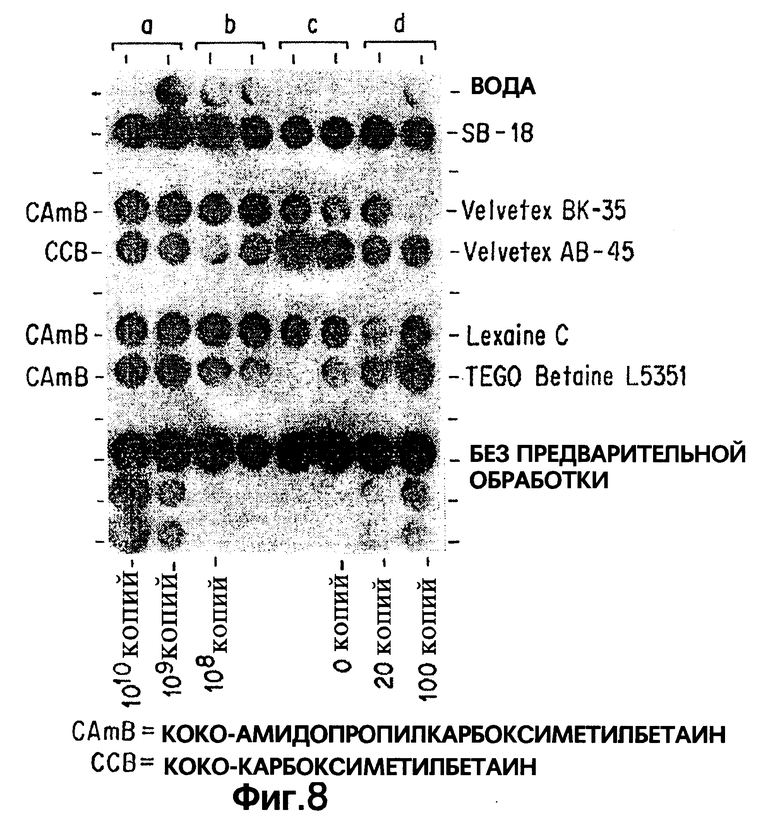
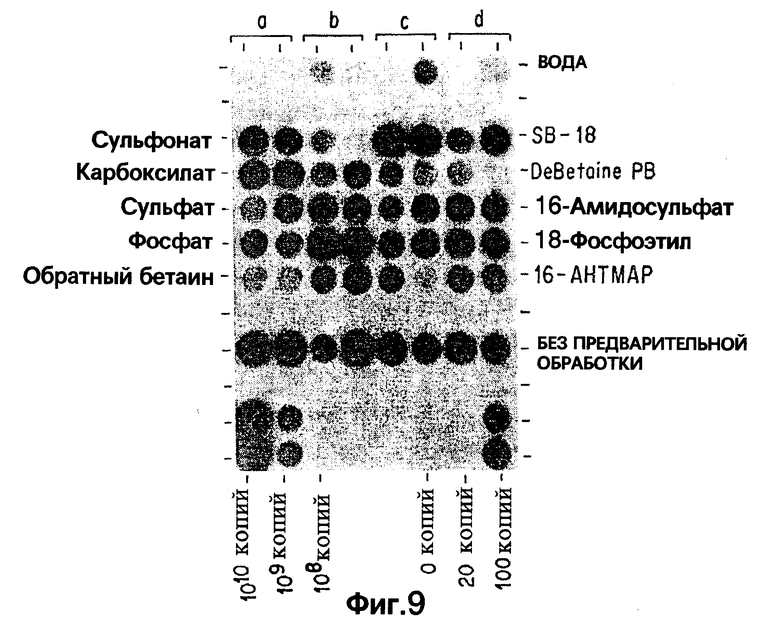
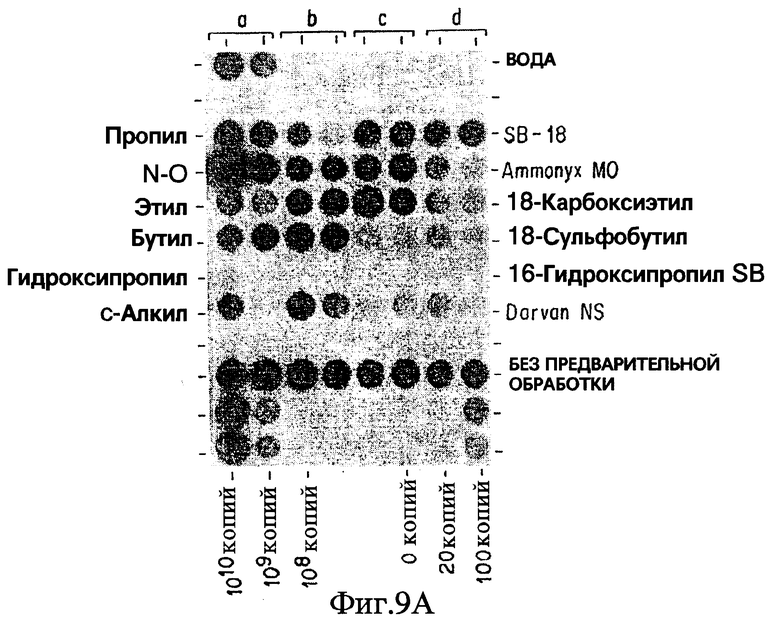
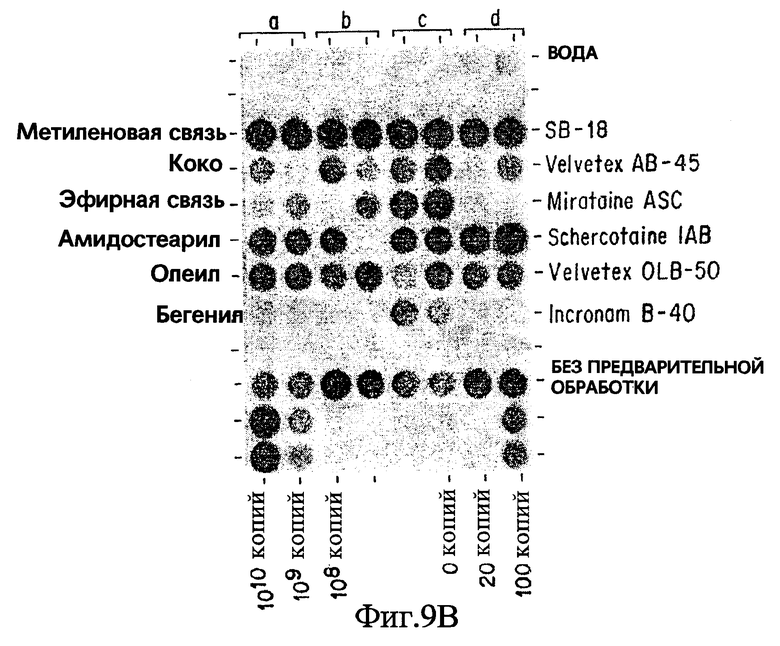
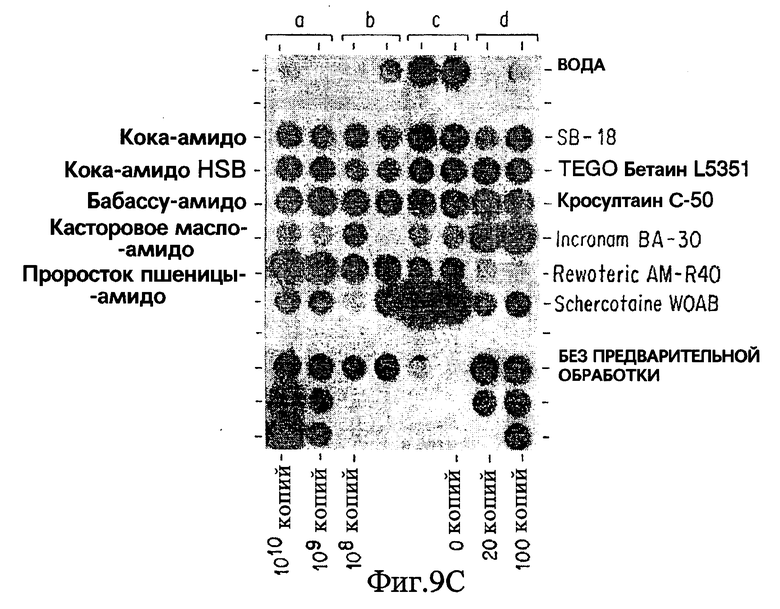
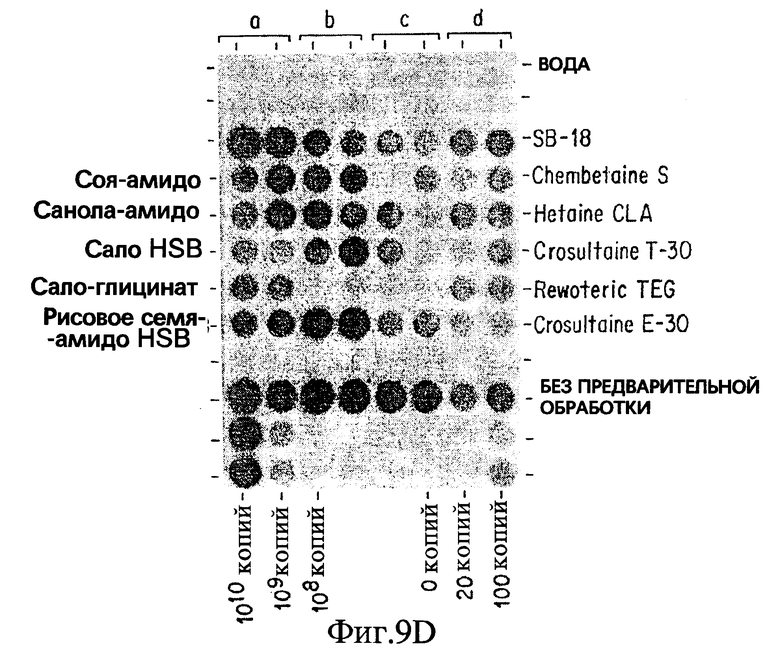
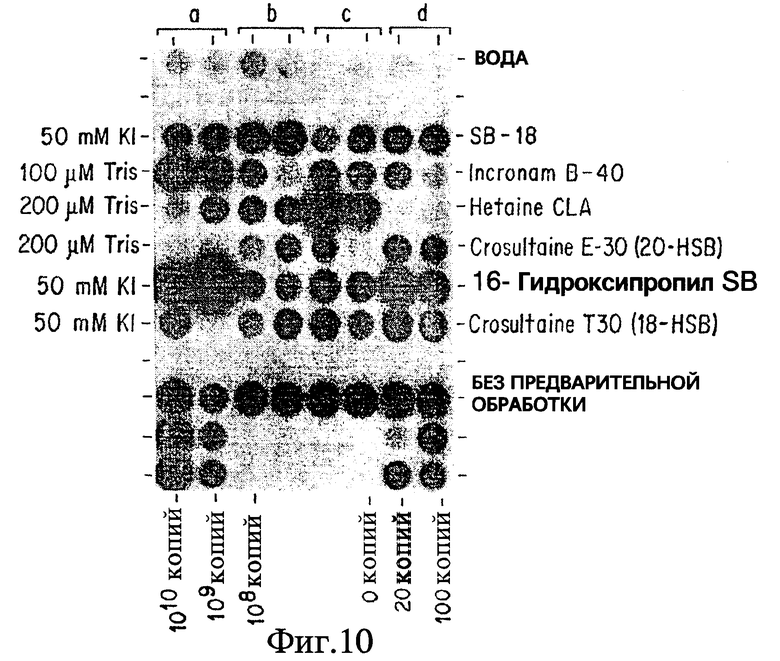
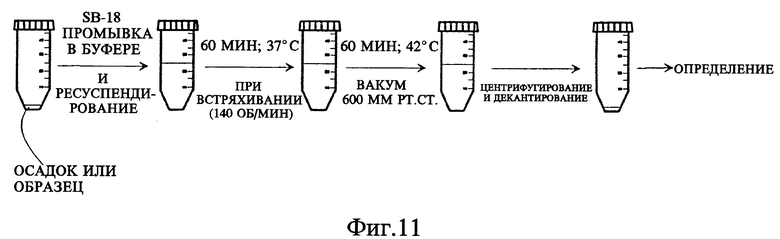
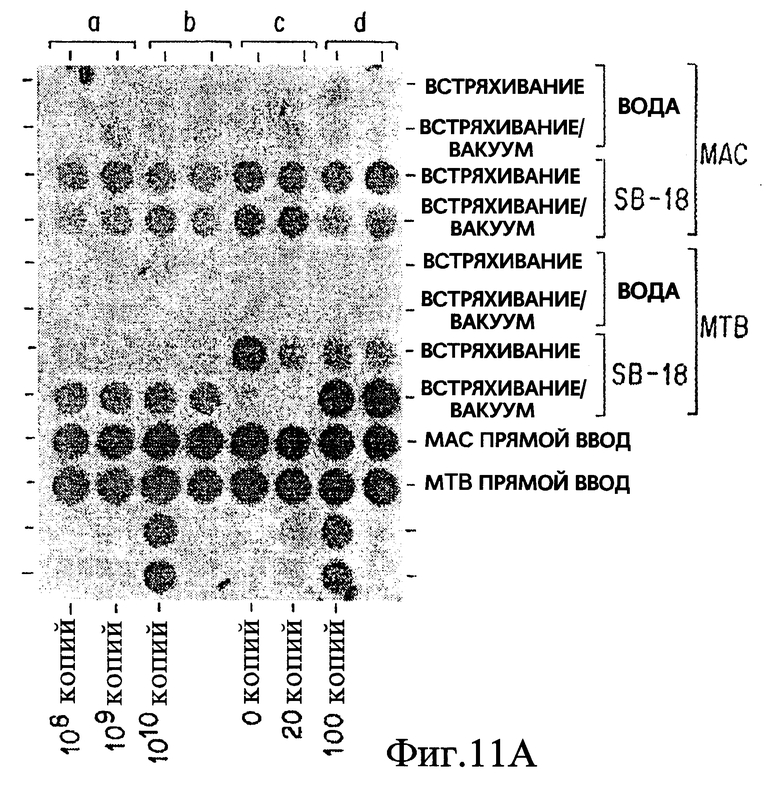
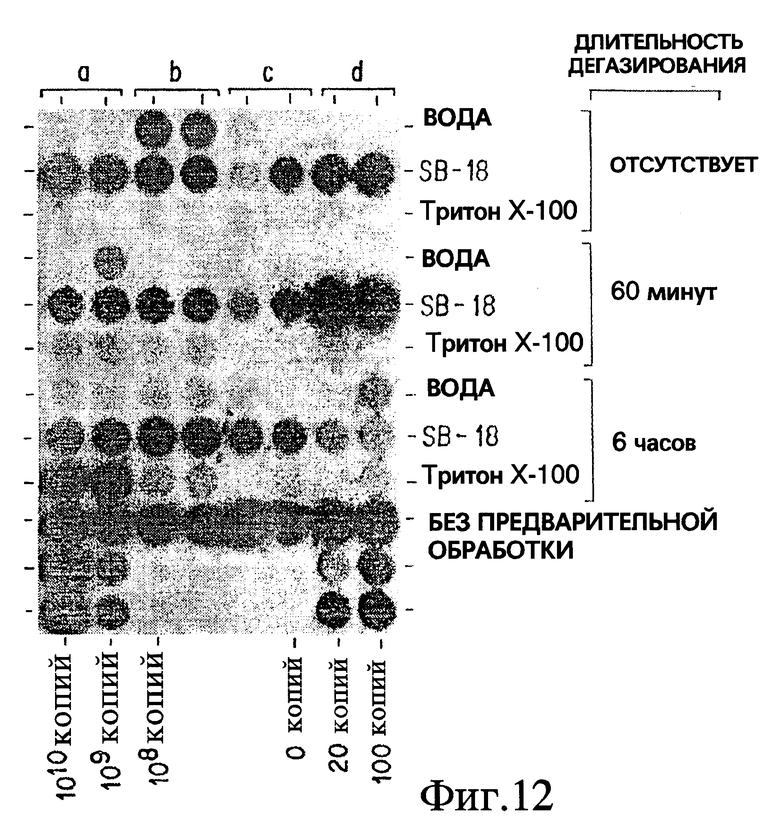
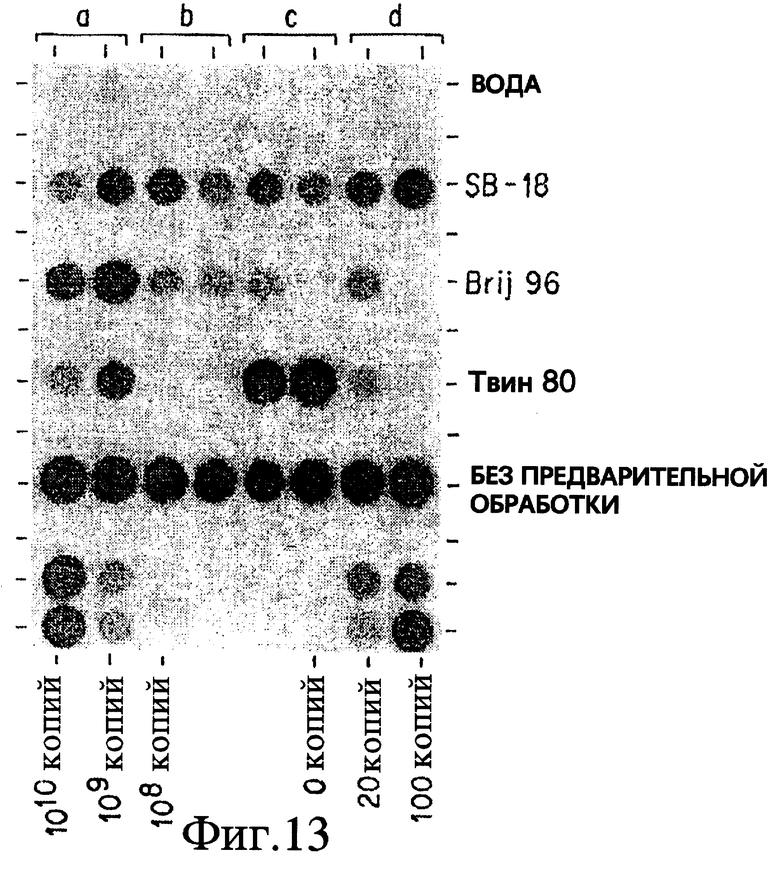
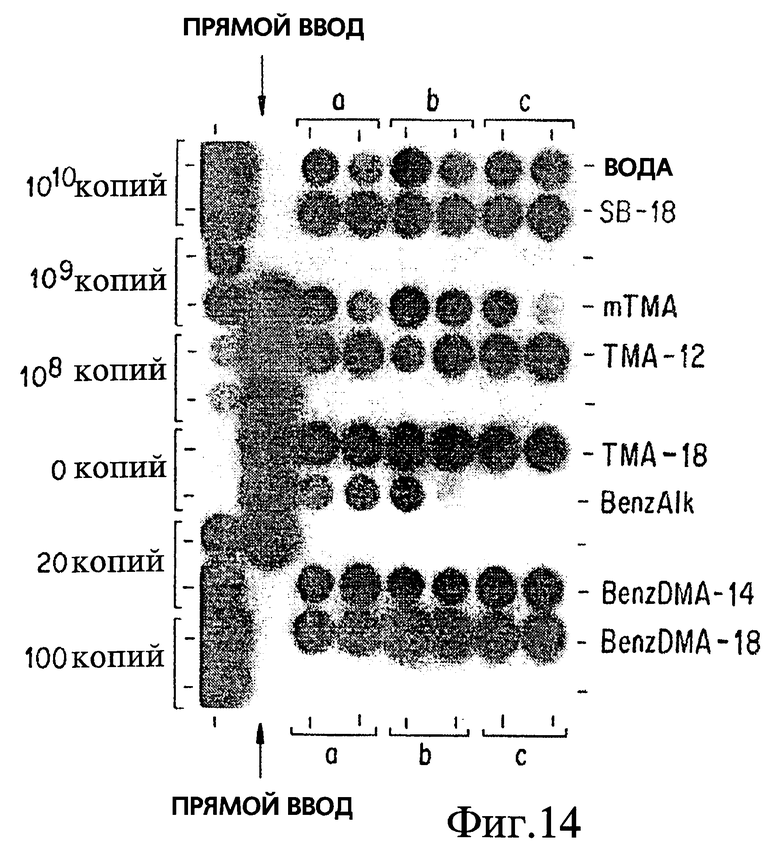
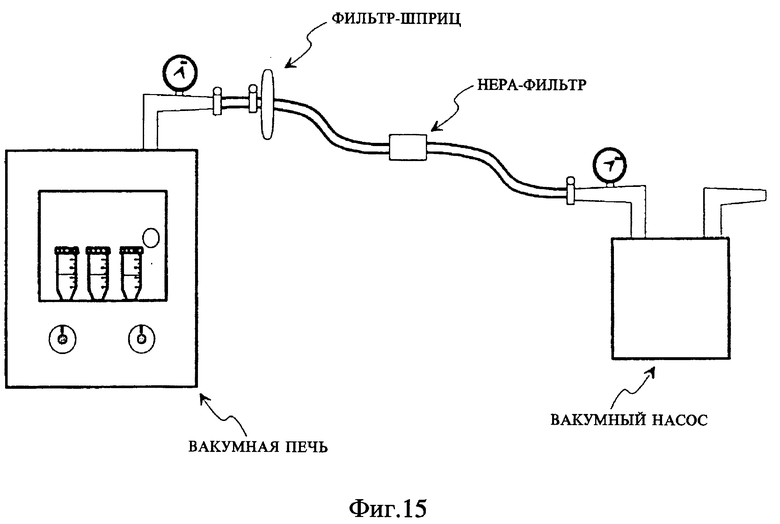
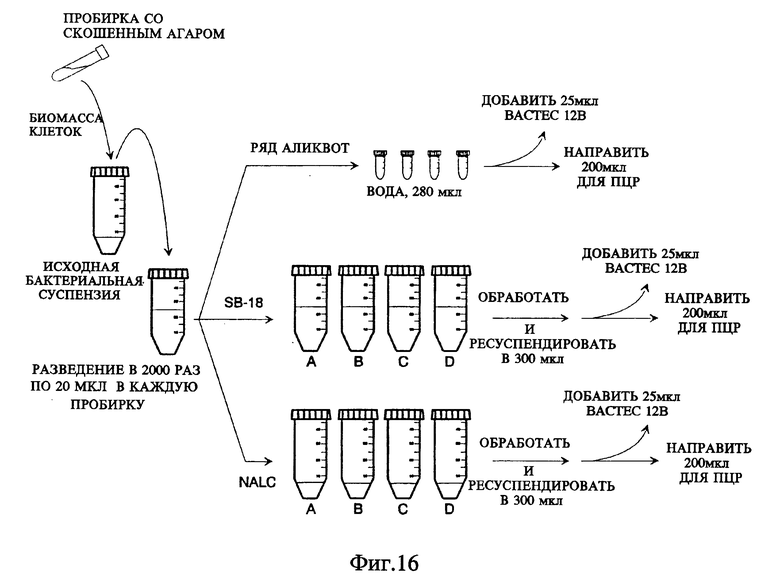
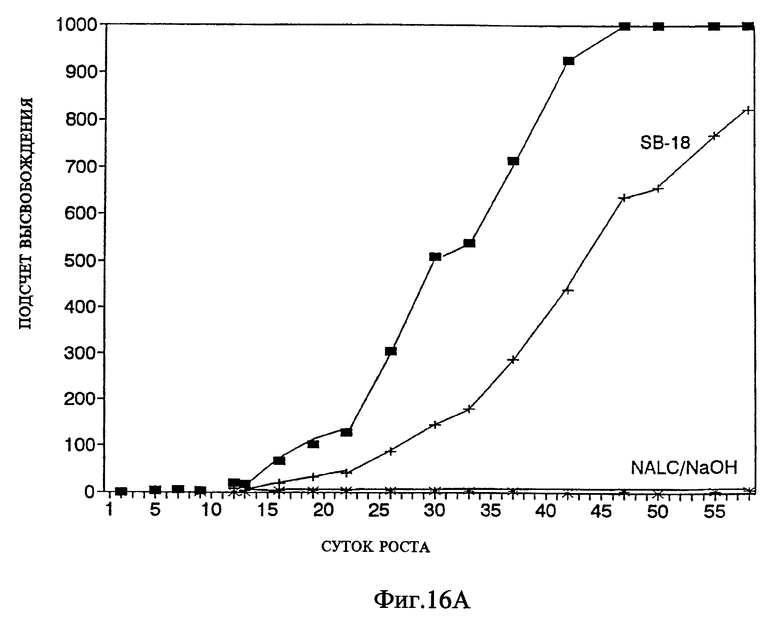
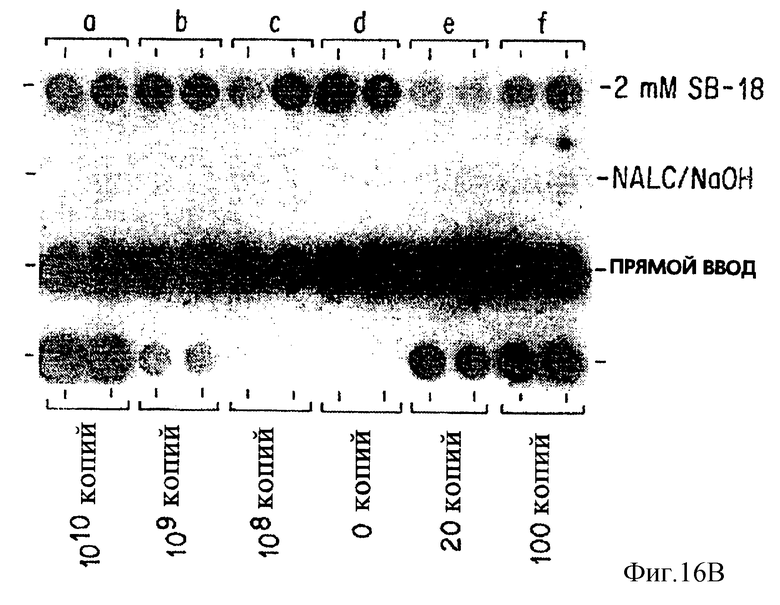
Изобретение относятся к микробиологии. При определении микроорганизма образец обрабатывают композицией, содержащей SB-18-подобный или палочковидный детергент. Определяемый микроорганизм должен содержать в наружной мембране структуры, подобные миколовой кислоте. Использование изобретения позволяет повысить точность определения микроорганизмов. 3 с. и 73 з.п.ф-лы, 16 ил., 17 табл.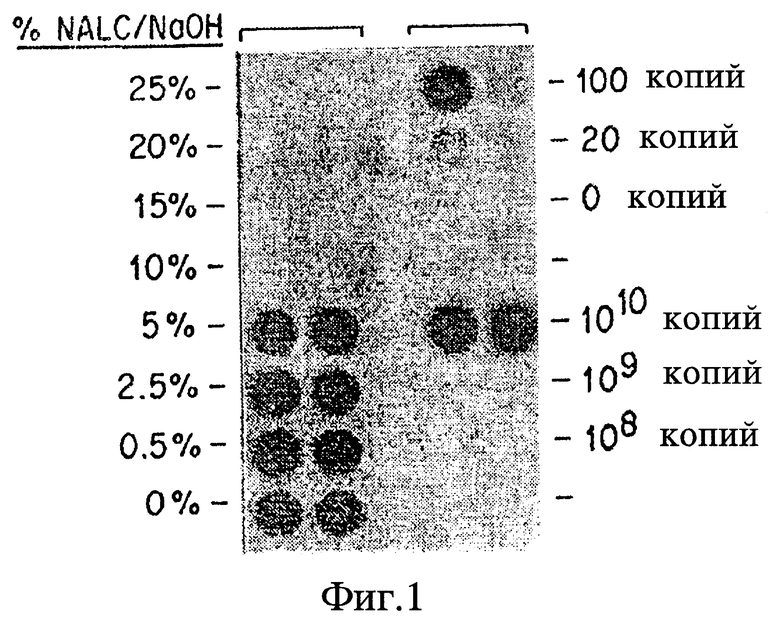
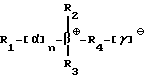
где R1 - C8-C22;
α- -CH2-, -CH(OH)-, -(CO)-NH-CH2CH2CH2-, -O- или -C(O)-;
n = 0 или 1;
β- -N⊕-, -P⊕- или -S⊕-;
R2- -H, -CH3, -C2H5, -C3H7 или C4H9;
R3- -H, -CH3, -C2H5, -C3H7 или C4H9;
R4 - -CH2-, -C2H4-, -C3H6-, -C4H8-, -C5H10-, -C6H12-, -CH2-C6H4-, -CmH2m-, -CH(OH)CH2CH2-, -CH2CH(OH)CH2- или -CmH2m-1(OH)-, где m ≥ 1;
γ- -SO
6. Способ по п.3, отличающийся тем, что в качестве детергента используют HSB-подобный детергент.
N,N-диметил-N-(сульфометил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(сульфометил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(сульфометил)-1-октадеканаминий, внутренняя соль,
N,N-диметил-N-(2-сульфоэтил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(2-сульфоэтил)-1-тетрадеканаминий, внутренняя соль,
N,N-диметил-N-(2-сульфоэтил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-3-((1-оксогексадецил)амино)-N-(2-сульфоэтил)-1-пропанаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-октанаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-деканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-тетрадеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-пентадеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-октадеканаминий, внутренняя соль,
гидроксид додецилдипропил(3-сульфопропил)аммония, внутренняя соль,
N, N-диметил-N-(3-(1-оксододецил)амино)-3-сульфо-1-пропанаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксогексадецил)амино)пропил)-3-сульфо-1-пропанаминий, внутренняя соль,
N,N-диметил-N-(метил-2-сульфоэтил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(метил-2-сульфоэтил)-1-тетрадеканаминий, внутренняя соль,
N,N-диметил-N-(метил-2-сульфоэтил)-1-гексадеканаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксогексадецил)амино)пропил)-2-сульфо-1-пропанаминий, внутренняя соль,
N,N-диметил-N-(4-сульфобутил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(4-сульфобутил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(4-сульфобутил)-1-октадеканаминий, внутренняя соль,
N-(1,3-диметил-3-сульфобутил)-N, N-диметил-1-додеканаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксогексадецил)амино)пропил)-4-сульфо-1-бутанаминий, внутренняя соль,
N,N-диметил-N-(6-сульфогексил)-1-гексадеканаминий, внутренняя соль,
N-додецил-N,N-диметил-4-сульфо-бензолметанаминий, внутренняя соль,
N,N-диметил-4-сульфо-N-тетрадецил-бензолметанаминий, внутренняя соль,
N-гексадецил-N,N-диметил-4-сульфо-бензолметанаминий, внутренняя соль,
N,N-диметил-N-октадецил-4-сульфо-бензолметанаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксододецил)амино)пропил)-4-сульфо-бензолметанаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксотетрадецил)амино)пропил)-4-сульфо-бензолметанаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксогексадецил)амино)пропил)-4-сульфо-бензолметанаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксооктадецил)амино)пропил)-4-сульфо-бензолметанаминий, внутренняя соль,
N-(2-гидрокси-3-сульфопропил)-N, N-диметил-1-деканаминий, внутренняя соль,
N-(2-гидрокси-3-сульфопропил)-N, N-диметил-1-додеканаминий, внутренняя соль,
N-(2-гидрокси-3-сульфопропил)-N,N-диметил-2-тетрадеканаминий, внутренняя соль,
N-(2-гидрокси-3-сульфопропил)-N,N-диметил-1-пентадеканаминий, внутренняя соль,
N-(2-гидрокси-3-сульфопропил)-N,N-диметил-1-гексадеканаминий, внутренняя соль,
N-(2-гидрокси-3-сульфопропил)-N, N-диметил-1-октадеканаминий, внутренняя соль,
2-гидрокси-N, N-диметил-N-(3-((1-оксододецил)амино)пропил)-3-сульфо-1-пропанаминий, внутренняя соль,
2-гидрокси-N, N-диметил-N-(3-((1-оксотетрадецил)амино)пропил)-3-сульфо-1-пропанаминий, внутренняя соль,
2-гидрокси-N, N-диметил-N-(3-((1-оксогексадецил)амино)пропил)-3-сульфо-1-пропанаминий, внутренняя соль,
2-гидрокси-N, N-диметил-N-(3-((1-оксооктадецил)амино)пропил)-3-сульфо-1-пропанаминий, внутренняя соль,
N-(2-гидрокси-3-сульфопропил)-N,N-диметил-1-тетрадеканаминий, внутренняя соль и
N-(2-гидрокси-3-сульфопропил)-N,N-диметил-1-тетрадеканаминий, внутренняя соль,
коко-амидопропилоксисульфобетаин,
алкиловый эфир гидроксипропилсульфобетаина,
таллоу-амидопропилгидроксипропилсульфобетаин,
эрукоамидопропилгидроксипропилсульфобетаин и каноламидопропилбетаин.
N,N-диметил-N-(2-сульфоэтил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(2-сульфоэтил)-1-тетрадеканаминий, внутренняя соль,
N,N-диметил-N-(2-сульфоэтил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-тетрадеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-пентадеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(3-сульфопропил)-1-октадеканаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксододецил)амино)пропил)-3-сульфо-1-пропанаминий, внутренняя соль,
N, N-диметил-N-(3-((1-оксогексадецил)амино)пропил)-3-сульфо-1-пропанаминий, внутренняя соль,
N,N-диметил-N-(4-сульфобутил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(4-сульфобутил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(4-сульфобутил)-1-октадеканаминий, внутренняя соль,
N, N-диметил-N-(3-((оксогексадецил)амино)пропил)-4-сульфо-1-бутанаминий, внутренняя соль.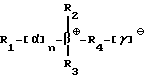
где R1 - C8-C22;
α- -CH2-, -CH(OH)-, -(CO)-NH-CH2CH2CH2-, -O- или -C(O)-;
n = 0 или 1;
β- -N⊕-, -P⊕- или -S⊕-;
R2- -H, -CH3, -C2H5, -C3H7 или C4H9;
R3- -H, -CH3, -C2H5, -C3H7 или C4H9;
R4 - -CH2-, -C2H4-, -C3H6-, -C4H8-, -C5H10-, -C6H12-, -CH2-C6H4-, -CmH2m-, -CH(OH)CH2CH2-, -CH2CH(OH)CH2- или -CmH2m-1(OH)-, где m ≥ 1;
γ- -COO⊖.
13. Способ по п.12, отличающийся тем, что указанный CB-подобный детергент выбран из группы следующих соединений:
N-(карбоксиметил)-N, N-диметил-1-гексадеканаминий, внутренняя соль, коко-карбоксиметилбетаин,
N-(карбоксиметил)-N,N-диметил-9-октадецен-1-аминий, внутренняя соль,
N-(карбоксиметил)-N,N-диметил-3-((1-оксооктадецил)амино)-1-пропанаминий, внутренняя соль,
N-C8-C22-ацильные производные 3-амино-N-(карбоксиметил)-N,N-диметил-1-пропанаминия, внутренняя соль,
N-(карбоксиметил)-3-((12-гидрокси-1-оксо-9-октадеценил)амино)N, N-диметил-1-пропанаминий, внутренняя соль,
коко-амидопропилкарбоксиметилбетаин
N-(2-карбоксиэтил)-N,N-диметил-1-додеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-тридеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-тетрадеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-гексадеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-октадеканаминий, внутренняя соль,
N-додецил-β-аланин,
N-(3-карбоксипропил)-N,N-диметил-1-ундеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-додеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-тетрадеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-пентадеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-гексадеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-октадеканаминий, внутренняя соль,
N-(4-карбоксибутил)-N,N-диметил-1-додеканаминий, внутренняя соль,
N-(5-карбоксипентил)-N,N-диметил-1-додеканаминий, внутренняя соль,
N-(5-карбоксипентил)-N,N-диметил-1-гексадеканаминий, внутренняя соль,
N-(6-карбоксигексил)-N,N-диметил-1-додеканаминий, внутренняя соль,
4-карбокси-N-додецил-N,N-диметил-бензолметанаминий, внутренняя соль,
2-карбокси-N-додецил-N,N-диметил-бензолметанаминий, внутренняя соль,
4-карбокси-N-гексадецил-N,N-диметил-бензолметанаминий, внутренняя соль,
2-карбокси-N-гексадецил-N,N-диметил-бензолметанаминий, внутренняя соль,
таллоу-глицинат (CAS® 70750-46-8), соя-амидопропилкарбоксиметилбетаин и бабассу-амидопропилкарбоксиметилбетаин.
N-(2-карбоксиэтил)-N,N-диметил-1-додеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-тридеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-тетрадеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-гексадеканаминий, внутренняя соль,
N-(2-карбоксиэтил)-N,N-диметил-1-октадеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-ундеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-додеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-тетрадеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-пентадеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-гексадеканаминий, внутренняя соль,
N-(3-карбоксипропил)-N,N-диметил-1-октадеканаминий, внутренняя соль,
N-(4-карбоксибутил)-N,N-диметил-1-додеканаминий, внутренняя соль.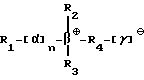
где R1 - C8-C22;
α- -CH2-, -CH(OH)-, -(CO)-NH-CH2CH2CH2-, -O- или -C(O)-;
n = 0 или 1;
β- -N⊕-, -P⊕- или -S⊕-;
R2- -H, -CH3, -C2H5, -C3H7 или C4H9;
R3- -H, -CH3, -C2H5, -C3H7 или C4H9;
R4 - -CH2-, -C2H4-, -C3H6-, -C4H8-, -C5H10-, -C6H12-, -CH2-C6H4-, -CmH2m-, -CH(OH)CH2CH2-, -CH2CH(OH)CH2- или -CmH2m-1(OH)-, где m ≥ 1;
γ- -PO
N,N-диметил-N-(2-(фосфоноокси)этил)-1-деканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-ундеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-тетрадеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-гептадеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-октадеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-октадецен-1-аминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-9-октадецен-1-аминий, внутренняя соль,
N,N-диэтил-N-(2-(фосфоноокси)этил)-1-гексадеканаминий, внутренняя соль,
N-(2-(фосфоноокси)этил)-N, N-дипропил-1-гексадеканаминий, внутренняя соль,
N,N-дибутил-N-(2-(фосфоноокси)этил)-1-гексадеканаминий, внутренняя соль,
N-этил-N-(2-(фосфоноокси)этил)-N-пропил-1-гексадеканаминий, внутренняя соль,
N-этил-N-метил-N-(2-(фосфоноокси)этил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(3-(фосфоноокси)пропил)-1-гексадеканаминий, внутренняя соль,
N, N-диметил-N-(4-(фосфоноокси)бутил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(6-(фосфоноокси)гексил)-1-гексадеканаминий, внутренняя соль,
2-гидрокси-N, N-диметил-N-(2-(фосфоноокси)этил)-1-додеканаминий, внутренняя соль,
2-гидрокси-N,N-диметил-N-(2-(фосфоноокси)этил)-1-тетрадеканаминий, внутренняя соль,
2-гидрокси-N,N-диметил-N-(2-(фосфоноокси)этил)-1-гексадеканаминий, внутренняя соль,
N-бутил-N-этил-N-(2-(фосфоноокси)этил)-1-гексадеканаминий, внутренняя соль,
2-гидрокси-N,N-диметил-N-(3-((1-оксододецил)амино)пропил)-3-(фосфоноокси)-1-пропанаминий, внутренняя соль,
2-гидрокси-N, N-диметил-N-(3-((1-оксооктадецил)амино)пропил)-3-(фосфоноокси)-1-пропанаминий, внутренняя соль,
3-(децилокси)-2-гидрокси-N, N-диметил-N-(2-(фосфоноокси)этил)-1-пропанаминий, внутренняя соль,
3-(додецилокси)-2-гидрокси-N, N-диметил-N-(2-(фосфоноокси)этил)-1-пропанаминий, внутренняя соль,
2-гидрокси-N, N-диметил-N-(2-(фосфоноокси)этил)-3-(тетрадецилокси)-1-пропанаминий, внутренняя соль.
N,N-диметил-N-(2-(фосфоноокси)этил)-1-додеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-тетрадеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-гексадеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-гептадеканаминий, внутренняя соль,
N,N-диметил-N-(2-(фосфоноокси)этил)-1-октадеканаминий, внутренняя соль,
N,N-диметил-N-(3-(фосфоноокси)пропил)-1-гексадеканаминий, внутренняя соль и
N, N-диметил-N-(4-(фосфоноокси)бутил)-1-гексадеканаминий, внутренняя соль.
25 Способ приготовления образца для определения, по крайней мере, одного вида микроорганизма, присутствующего в образце, предусматривающий обработку клинического материала или небиологической пробы с последующим определением, по крайней мере, одного вида микроорганизма, отличающийся тем, что указанный микроорганизм содержит в наружной мембране структуры, подобные миколовой кислоте, обработку осуществляют при пониженном давлении, достаточном для изменения плавучести, по крайней мере, одного вида указанного микроорганизма в данном образце, а определение микроорганизма производят по наблюдаемому изменению плавучести.
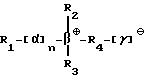
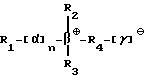
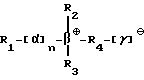
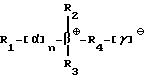
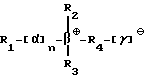
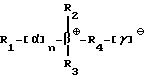
| Экономайзер | 0 |
|
SU94A1 |
| СПОСОБ ТРАНСПОРТИРОВКИ СЖИЖЕННОГО ПРИРОДНОГО ГАЗА | 1999 |
|
RU2228486C2 |
| Способ проведения гибридизационного анализа | 1989 |
|
SU1620938A1 |

