Область техники, к которой относится изобретение
Изобретение относится к молекулярной биологии, микробиологии и медицине и обеспечивает способ определения генетических детерминант резистентности возбудителя туберкулеза - микобактерий туберкулезного комплекса (Mycobacterium tuberculosis, М. bovis, М. bovis BCG, М. africanum, М. microti, М. canettii, М. caprae, М. pinnipedii, М. mungi) к новейшим противотуберкулезным препаратам, включая бедаквилин и линезолид, в клиническом образце на дифференцирующем олигонуклеотидном микрочипе.
Уровень техники
В соответствии с приказом Министерства здравоохранения Российской Федерации №951 от 29.12.2014 г. пациентов с лекарственно-устойчивыми формами туберкулеза необходимо переводить на IV и V режимы химиотерапии, включающие бедаквилин (BDQ) и линезолид (LZN). Эффективность применения данных препаратов зависит как от общего состояния пациента, так и от фенотипических и генетических характеристик изолята М. tuberculosis, являющегося причиной болезни. Недавние исследования изолятов от больных, получающих терапию бедаквилином и линезолидом, позволил определить следующие состояния возбудителя туберкулеза (ТБ) - предсуществующая резистентность к бедаквилину у пациентов с лекарственно-устойчивыми формами ТБ и резистентность к бедаквилину и/или линезолиду, приобретаемая больными в ходе лечения (Zimenkov D, Nosova Е, Kulagina Е, Antonova О, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline- and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906), что подтверждает необходимость разработки молекулярно-генетического теста для оценки эффективности действия данных препаратов.
Бедаквилин (торговое название «Сиртуро», кодовые названия ТМС207 и R207910), диарилхинолон, полученный группой Koen Andries из Janssen Pharmaceutica, является первым за последние 40 лет противотуберкулезным препаратом с новым механизмом действия (Palomino, J.C. & Martin, A. TMC207 becomes bedaquiline, a new anti-TB drug. 2013. Future Microbiology. 8: 1071-80). В декабре 2012 года Food and Drug Administration (FDA, США) одобрено применение бедаквилина для комбинированной терапии туберкулеза с множественной лекарственной резистентностью (МЛУ) у взрослых.
Бедаквилин ингибирует микобактериальную АТФ-синтазу, препятствуя образованию АТФ в клетке и нарушению внутриклеточного метаболизма. Бедаквилин не влияет на активность АТФ-синтазы млекопитающих. Описано два вероятных механизма формирования резистентности М. tuberculosis к бедаквилину. Первый механизм связан с мутациями в гене atpE, кодирующем трансмембранную С-субъединицу АТФ-синтазы. Такой механизм был выявлен в штаммах с резистентностью к бедаквилину, полученной in vitro (Andries, K., Verhasselt, P., Guillemont, J., Gohlmann, H.W., Neefs, J.M., Winkler, H., Van Gestel, J., Timmerman, P., Zhu, M., Lee, E., Williams, P., de Chaffoy, D., Huitric, E., Hoffner, S., Cambau, E., Truffot-Pernot, C., Lounis, N. & Jarlier, V. A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis. 2005. Science. 307: 223-7; Segala, E., Sougakoff, W., Nevejans-Chauffour, A., Jarlier, V. & Petrella, S. New mutations in the mycobacterial ATP synthase: new insights into the binding of the diarylquinoline TMC207 to the ATP synthase C-ring structure. 2012. Antimicrobial Agents and Chemotherapy. 56: 2326-34). Секвенирование МЛУ-изолятов с селектированной in vitro резистентностью к бедаквилину позволило выявить следующие замены в гене atpE: N28V, D32V, E61D, А63Р, I66M, N28Q, L59V, E61N, А63Р и I66M, находящиеся в области взаимодействия бедаквилина и С-субъединицы. Однако только треть устойчивых штаммов обладала мутациями в гене atpE, остальные изоляты, по видимому, обладали детерминантами резистентности в иных локусах.
Секвенирование изолятов с искусственно полученной резистентностью (Hartkoorn, R.C., Uplekar, S. & Cole, S.T. Cross-resistance between clofazimine and bedaquiline through upregulation of MmpL5 in Mycobacterium tuberculosis. 2014. Antimicrobial Agents and Chemotherapy. 58, 2979-81.) и изолята от пациента с приобретенной резистентностью (Andries, K., Villellas, С., Coeck, N., Thys, K., Gevers, Т., Vranckx, L., Lounis, N., de Jong, B.C. & Koul, A. Acquired Resistance of Mycobacterium tuberculosis to Bedaquiline. 2014. PLoS One. 9: e102135) выявило мутации в локусе Rv0678, кодирующем транскрипционный репрессор ряда генов, включающих в себя mmpL5 и mmpS5, чьи продукты формируют эффлюксный насос, обеспечивающий транспорт наружу таких препаратов как клофазимин и эконазол. Так как любая инактивация репрессора приводит к повышению экспрессии эффлюксного комплекса, то спектр возможных мутаций в рамке считывания Rv0678 необычайно широк и включает в себя как мутации, приводящие к аминокислотным заменам в ключевых регионах белка, так и всевозможные нонсенс-мутации, ответственные за сбой рамки считывания и отсутствие полноценного белка-репрессора в клетке.
Результаты недавних исследований показали возможную ассоциацию мутаций в локусе pepQ с резистентностью к бедаквилину и клофазимину (Almeida D, Ioerger Т, Tyagi S et al. Mutations in pepQ confer low-level resistance to bedaquiline and clofazimine in Mycobacterium tuberculosis. 2016. Antimicrobial Agents and Chemotherapy. 60: 4590-9), однако в исследовании российской популяции бедаквилин-резистентных изолятов последовательность данного локуса была отнесена к дикому типу (Zimenkov D, Nosova Е, Kulagina Е, Antonova О, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline- and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906).
Линезолид (Zyvox, Pfizer), представитель класса оксазолидинонов, был одобрен FDA в 2000 г. для лечения инфекций, вызванных грамположительными бактериями, а также показал бактериостатическую активность против М. tuberculosis с множественной и широкой (ШЛУ) лекарственной резистентностью как in vitro, так и на животных моделях (Zhang, Z., Pang, Y., Wang, Y., Liu, C. & Zhao, Y. Beijing genotype of Mycobacterium tuberculosis is significantly associated with linezolid resistance in multidrug-resistant and extensively drug-resistant tuberculosis in China. 2014. International Journal of Antimicrobial Agents 43: 231-5). Исследования эффективности применения линезолида для лечения МЛУ-ТБ показали хорошую переносимость препарата и обосновали введение линезолида для терапии резистентных форм ТБ (Lee, М., Lee, J., Carroll, M.W., Choi, H., Min, S., Song, Т., Via, L.E., Goldfeder, L.C., Kang, E., Jin, В., Park, H., Kwak, H., Kim, H., Jeon, H.S., Jeong, I., Joh, J.S., Chen, R.Y., Olivier, K.N., Shaw, P.A., Follmann, D., Song, S.D., Lee, J.K., Lee, D., Kim, C.T., Dartois, V., Park, S.K., Cho, S.N. & Barry, C.E., 3rd. Linezolid for treatment of chronic extensively drug-resistant tuberculosis. 2012. New England Journal of Medicine. 367: 1508-18).
Считается, что линезолид, как и остальные оксазолидиноны, препятствует сборке рибосомы на этапе присоединения формилметиониновой тРНК, и таким образом блокирует инициацию трансляции (Shinabarger, D. Mechanism of action of the oxazolidinone antibacterial agents. 1999. Expert Opinion on Investigational Drugs. 8: 1195-202). В настоящее время установлено, что механизм действия линезолида похож на механизм действия других антибиотиков, связывающихся с пептидил-трансферазным центром рибосомы. Сравнение кристаллических структур комплекса 50S субъединицы с разными антибиотиками показало, что линезолид частично делит сайт связывания с другими антибиотиками, которые ингибируют синтез белка. Хлорамфеникол, анизомицин, клиндамицин, тиамулин и стрептомицин А имеют сайты связывания в А-сайте пептидил-трансферазного центра, перекрывающиеся с сайтом связывания линезолида.
Генетические детерминанты резистентности, выявленные в линезолид-устойчивых изолятах возбудителя туберкулеза, были обнаружены генах 23S рРНК и rplC фибосомный белок L3) (Richter, Е., Rusch-Gerdes, S. & Hillemann, D. First linezolid-resistant clinical isolates of Mycobacterium tuberculosis. 2007. Antimicrobial Agents and Chemotherapy. 51: 1534-6; Beckert, P., Hillemann, D., Kohl, T.A., Kalinowski, J., Richter, E., Niemann, S. & Feuerriegel, S. rplC T460C identified as a dominant mutation in linezolid-resistant Mycobacterium tuberculosis strains. 2012. Antimicrobial Agents and Chemotherapy. 56: 2743-5). В российской популяции линезолид-устойчивых изолятов идентифицированы замены C154R в различных вариантах в гене rplC, впервые описаны мутации g2294a и g2814t в гене 23S рРНК (Zimenkov D, Nosova Е, Kulagina Е, Antonova О, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline- and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906).
Таким образом, для первичного скрининга изолятов от больных туберкулезом, планируемых к терапии бедаквилином и линезолидом, актуальным представляется анализ генетических детерминант резистентности в локусах Rv0678, atpE, 23S рРНК, rplC. Принимая во внимание тот факт, что мутации в гене Rv0678 могут возникать спонтанно в любом положении открытой рамки считывания, и, кроме того, в резистентных к BDQ клинических изолятах выявлены гетеропопуляции микобактерий - смеси штаммов с разными мутациями в данном локусе, наиболее подходящим способом идентификации таких детерминант резистентности могли бы стать технологии секвенирования нового поколения (NGS). Однако успех внедрения технологий NGS во многом зависит от миниатюризации платформ, примером которых может быть система MinION (Votintseva А.А., Bradley P., Pankhurst L., Del Ojo Elias C., Loose M., Nilgiriwala K., Chatterjee A., Smith E.G., Sanderson N., Walker Т.M., Morgan M.R., Wyllie D.H., Walker A.S., Peto Т., Crook D.W. and Iqbal Z. Same-Day Diagnostic and Surveillance Data for Tuberculosis via Whole-Genome Sequencing of Direct Respiratory Samples. 2017. Journal of clinical microbiology. 55(5): 1285-1298), усовершенствования этапа пробоподготовки образца и выделения НК, упрощения и ускорения процедуры анализа в условиях потоковых тестов клинико-диагностической лаборатории (Khodakov D., Wang С. and Zhang D.Y. Diagnostics based on nucleic acid sequence variant profiling: PCR, hybridization, and NGS approaches. 2016. Advanced drug delivery reviews. 105(Pt A): 3-19.). Вместе с тем, полное отсутствие отечественных разработок, как в области приборной базы, так и в наборах реагентов, обеспечивающих цикл полногеномного/таргетного анализа клинического материала, не позволяет пока снизить цену до приемлемого уровня. Таким образом, в данной области существует острая потребность в разработке способа выявления генетических детерминант резистентности возбудителя туберкулеза к бедаквилину и линезолиду, который бы выгодно отличался от известных из уровня техники решений простотой проведения анализа, высокими специфичностью и информативностью в отношении количества определяемых детерминант резистентности, а также невысокой стоимостью.
Раскрытие изобретения
В результате проведенных обширных научных исследований, анализа баз данных нуклеотидных последовательностей NCBI http://www.ncbi.nlm.nih.gov/genome/genomes/166?subset=, Welcome Trust Sanger institute http://www.sanger.ac.uk/resources/downloads/bacteria/mycobacterium.html, TBDReaMDB (www.tbdreamdb.com), результатов собственных геномных исследований (Zimenkov D, Nosova E, Kulagina E, Antonova O, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline- and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906) авторы настоящего изобретения обнаружили, что задача разработки способа определения генетических детерминант резистентности возбудителя туберкулеза к бедаквилину и линезолиду может быть успешно решена путем использования олигонуклеотидных микрочипов (биочипов), содержащих олигонуклеотидные зонды, последовательности которых специфичны к последовательности гена Rv0678, а также к мутантным вариантам генов atpE, 23SpPHK, rplC.
Технология олигонуклеотидных микрочипов низкой плотности успешно зарекомендовала себя для идентификации лекарственно-устойчивых изолятов возбудителя туберкулеза (Gryadunov D., Mikhailovich V., Lapa S., Roudinskii N., Donnikov M.,  S., Markova O.,
S., Markova O.,  A., Chernousova L., Skotnikova O., Moroz A., Zasedatelev A. and Mirzabekov A. Evaluation of hybridisation on oligonucleotide microarrays for analysis of drug-resistant Mycobacterium tuberculosis. 2005. Clinical microbiology and infection. 11(7): 531-539; Gryadunov D., Dementieva E., Mikhailovich V., Nasedkina Т., Rubina A., Savvateeva E., Fesenko E., Chudinov A., Zimenkov D., Kolchinsky A. and Zasedatelev A. Gel-based microarrays in clinical diagnostics in Russia. 2011. Expert review of molecular diagnostics. 11(8): 839-853; Zimenkov D.V., Antonova O.V., A.V., Isaeva Y.D., Krylova L.Y., Popov S.A., Zasedatelev A.S., Mikhailovich V.M. and Gryadunov D.A. Detection of second-line drug resistance in Mycobacterium tuberculosis using oligonucleotide microarrays. 2013. BMC infectious diseases. 13: 240; Zimenkov D.V., Kulagina E.V., Antonova O.V., Zhuravlev V.Y. and Gryadunov D.A. Simultaneous drug resistance detection and genotyping of Mycobacterium tuberculosis using a low-density hydrogel microarray. 2016. Journal of antimicrobial chemotherapy. 71(6): 1520-1531. Патенты РФ 2562866, 2376387).
A., Chernousova L., Skotnikova O., Moroz A., Zasedatelev A. and Mirzabekov A. Evaluation of hybridisation on oligonucleotide microarrays for analysis of drug-resistant Mycobacterium tuberculosis. 2005. Clinical microbiology and infection. 11(7): 531-539; Gryadunov D., Dementieva E., Mikhailovich V., Nasedkina Т., Rubina A., Savvateeva E., Fesenko E., Chudinov A., Zimenkov D., Kolchinsky A. and Zasedatelev A. Gel-based microarrays in clinical diagnostics in Russia. 2011. Expert review of molecular diagnostics. 11(8): 839-853; Zimenkov D.V., Antonova O.V., A.V., Isaeva Y.D., Krylova L.Y., Popov S.A., Zasedatelev A.S., Mikhailovich V.M. and Gryadunov D.A. Detection of second-line drug resistance in Mycobacterium tuberculosis using oligonucleotide microarrays. 2013. BMC infectious diseases. 13: 240; Zimenkov D.V., Kulagina E.V., Antonova O.V., Zhuravlev V.Y. and Gryadunov D.A. Simultaneous drug resistance detection and genotyping of Mycobacterium tuberculosis using a low-density hydrogel microarray. 2016. Journal of antimicrobial chemotherapy. 71(6): 1520-1531. Патенты РФ 2562866, 2376387).
Заявляемый в настоящем изобретении способ выгодно отличается от известных из уровня техники методов возможностью анализа ДНК возбудителя, выделенной из клинического материала, с целью установления наличия/отсутствия точечных мутаций, инсерций, делеций заранее неизвестной локализации в последовательности гена Rv0678, а также идентификации 15 мутаций в генах atpE, 23S рРНК, rplC, ассоциированных с резистентностью к бедаквилину и линезолиду. Метод не требует дорогостоящего оборудования и высококвалифицированного персонала. Данные, полученные с помощью заявленного способа, могут быть использованы для включения бедаквилина и/или линезолида в IV и V режимы терапии конкретного пациента, мониторинга эффективности лечения и отмены данных препаратов в силу приобретения резистентности.
В своем первом аспекте данное изобретение обеспечивает способ определения генетических детерминант резистентности возбудителя туберкулеза к бедаквилину и линезолиду. Способ основан на мультиплексной ПЦР с получением флуоресцентно-меченных фрагментов микобактериального генома с последующей гибридизацией этих фрагментов на биологическом микрочипе, содержащем набор олигонуклеотидов. Способ предусматривает следующие стадии:
а) мультиплексную амплификацию генов Rv0678, atpE, 23S рРНК, rplC с одновременным флуоресцентным маркированием при использовании геномной ДНК микобактерий туберкулезного комплекса в качестве матрицы и набора праймеров, последовательности которых представлены SEQ ID NO: 1-12, с получением флуоресцентно-меченных фрагментов микобактериального генома.
б) обеспечение олигонуклеотидного микрочипа для установления наличия/отсутствия точечных мутаций, инсерций, делеций заранее неизвестной локализации в последовательности гена Rv0678, а также идентификации мутаций в генах atpE, 23S рРНК, rplC, ассоциированных с резистентностью к бедаквилину и линезолиду, представляющего собой подложку, содержащую множество дискретных элементов, в каждом из которых иммобилизован уникальный олигонуклеотидный зонд, имеющий последовательность, комплементарную последовательности одноцепочечного фрагмента, полученного на стадии (а), и выбранную из группы, включающей: а) соответствующие последовательности гена Rv0678 дикого типа таким образом, что совпадение последовательности 5'-концевой части каждого следующего зонда составляет не менее 75% с последовательностью предыдущего, при этом зонды длиной 20 нуклеотидов каждый полностью перекрывают последовательность кодирующей области гена Rv0678; б) соответствующие последовательности фрагмента гена atpE дикого типа; в) соответствующие последовательности фрагмента мутантного варианта гена atpE, приводящего к резистентности микроорганизма к бедаквилину; г) соответствующие последовательности фрагмента гена rplC дикого типа; д) соответствующие последовательности фрагмента мутантного варианта гена rplC, приводящего к резистентности микроорганизма к линезолиду; е) соответствующие последовательности фрагмента гена 23S рРНК дикого типа; ж) соответствующие последовательности фрагмента мутантного варианта гена 23S рРНК, приводящего к резистентности микроорганизмов к линезолиду; при этом последовательности иммобилизованных олигонуклеотидных зондов представлены SEQ ID NO: 13-140;
(в) - гибридизацию амплифицированных флуоресцентно-меченных продуктов, полученных на стадии (а), на олигонуклеотидном микрочипе с образованием дуплексов с иммобилизованными зондами в условиях, обеспечивающих разрешение в один нуклеотид между образующимися в результате гибридизации совершенными и несовершенными дуплексами;
(г) регистрацию результатов гибридизации на олигонуклеотидном микрочипе, проведенной на стадии (в) с использованием анализатора флуоресценции и программного обеспечения, что позволяет использовать программную обработку интенсивностей сигналов с последующей интерпретацией результатов, которую проводят в два этапа: на первом этапе анализируют сигналы в группах элементов, содержащих зонды, соответствующие последовательностям генов atpE, 23S рРНК, rplC; на втором этапе анализируют гибридизационный профиль, включающий интенсивности сигналов элементов, содержащих зонды, соответствующие последовательности гена Rv0678.
В одном из своих воплощений способ характеризуется тем, что амплификацию фрагментов микобактериального генома проводят, используя образец ДНК, выделенный из мокроты, экссудата, бронхо-альвеолярного лаважа или промывных вод бронхов, или образец ДНК, выделенный из клинического изолята микобактерий.
В следующем воплощении способ характеризуется тем, что на стадии (а) для флуоресцентного маркирования получаемых ПЦР-фрагментов используют флуоресцентно меченый дезоксиуридинтрифосфат.
В следующем воплощении способ характеризуется тем, что в каждой серии экспериментов на стадии (а) в отдельной пробирке проводят амплификацию и флуоресцентного маркирование образца микобактериальной ДНК штамма дикого типа H37Rv с последующей гибридизацией полученных флуоресцентно-меченных ПЦР-фрагментов на отдельном микрочипе на стадии (в) и регистрацией результатов гибридизации на стадии (г) с получением эталонного гибридизационного профиля, включающего интенсивности сигналов элементов, содержащих зонды, соответствующие последовательности гена Rv0678 дикого типа.
В еще одном из воплощений способ характеризуется тем, что на стадии (г) на первом этапе интерпретации анализируют сигналы групп элементов, содержащих зонды, специфичные к одному из генов atpE, 23S рРНК, rplC, таким образом, что если максимальный сигнал в группе зарегистрирован в элементе, содержащем зонд дикого типа, то считают, что по данной аминокислотной/нуклеотидной позиции данного гена изучаемый образец ДНК возбудителя туберкулеза мутаций не имеет, в противном случае, если максимальный сигнал в группе зарегистрирован в элементе, содержащем зонд, последовательность которого соответствует мутантному варианту одного из генов, то считают, что по данной аминокислотной/нуклеотидной позиции данного гена изучаемый образец ДНК возбудителя туберкулеза имеет мутацию, приводящую к резистентности к бедаквилину или линезолиду.
В еще одном из своих воплощений способ характеризуется тем, что на стадии (г) на втором этапе интерпретации результатов эталонный гибридизационный профиль нормируют на гибридизационные профили анализируемых образцов ДНК с неизвестной последовательностью гена Rv0678 с получением нормированных профилей, различающихся для последовательностей ДНК дикого типа и мутантных вариантов гена Rv0678, тем самым относя последовательность гена Rv0678 к дикому типу, либо к содержащей точечную мутацию, делецию, инсерцию, приводящую к резистентности к бедаквилину.
Другие аспекты настоящего изобретения будут ясны из прилагаемых фигур, подробного описания и формулы изобретения.
Краткий перечень фигур
Для более ясного понимания сущности заявленного изобретения, а также для демонстрации его характерных черт и преимуществ далее приводятся ссылки на фигуры чертежей, на которых:
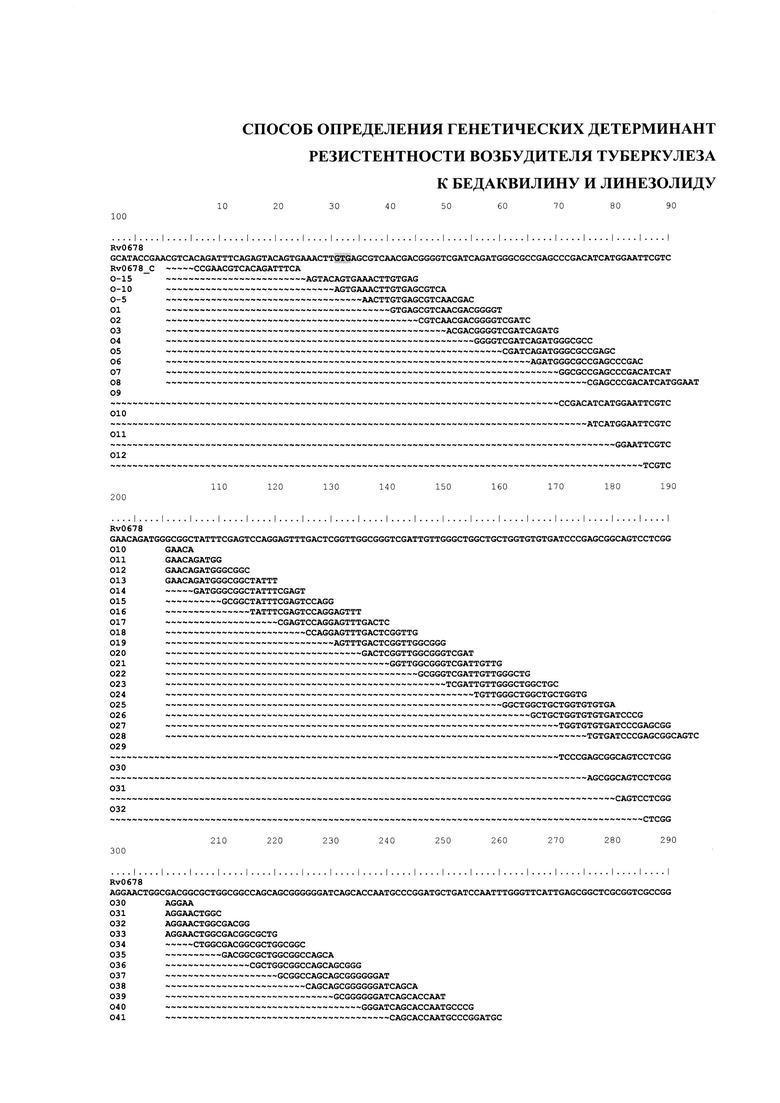
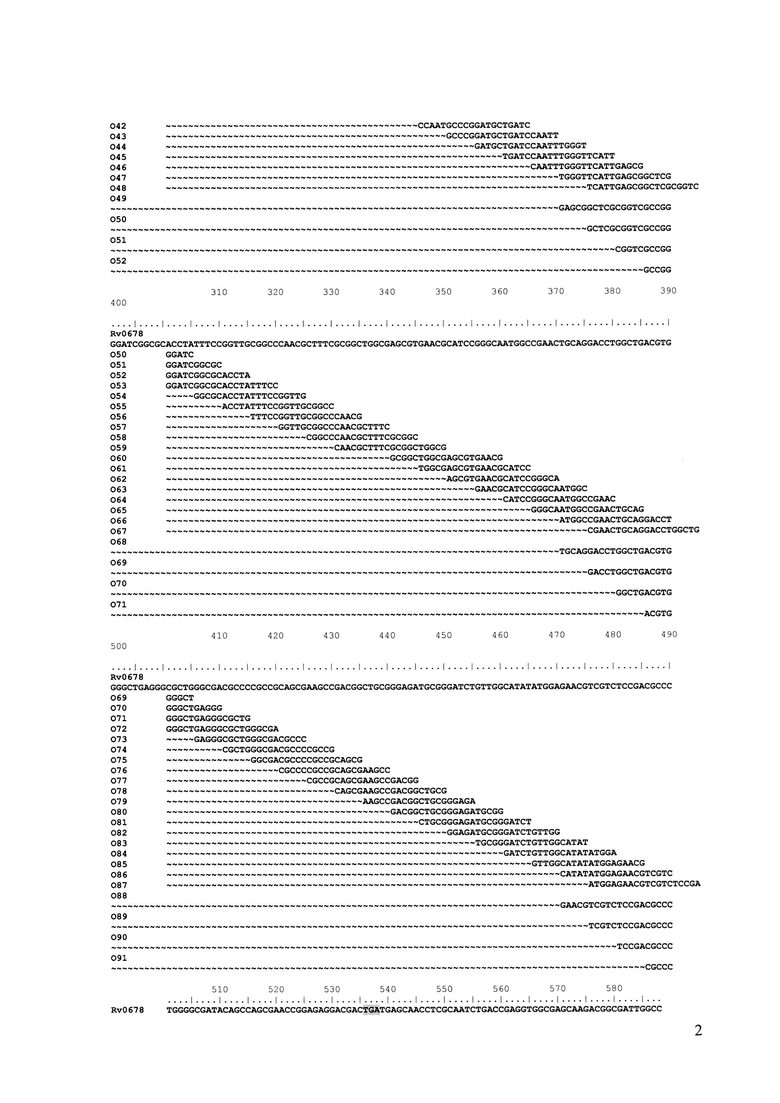
Фиг. 1. Представляет выравнивание последовательностей олигонуклеотидов длиной 20 оснований, перекрывающих последовательность дикого типа гена Rv0678. Выделены старт- и стоп-кодоны кодирующей области гена. Олигонуклеотид Rv0678_c предназначен для контроля полноразмерной амплификации гена Rv0678.
Фиг. 2. Представляет схему размещения дискриминирующих олигонуклеотидов на биочипе.
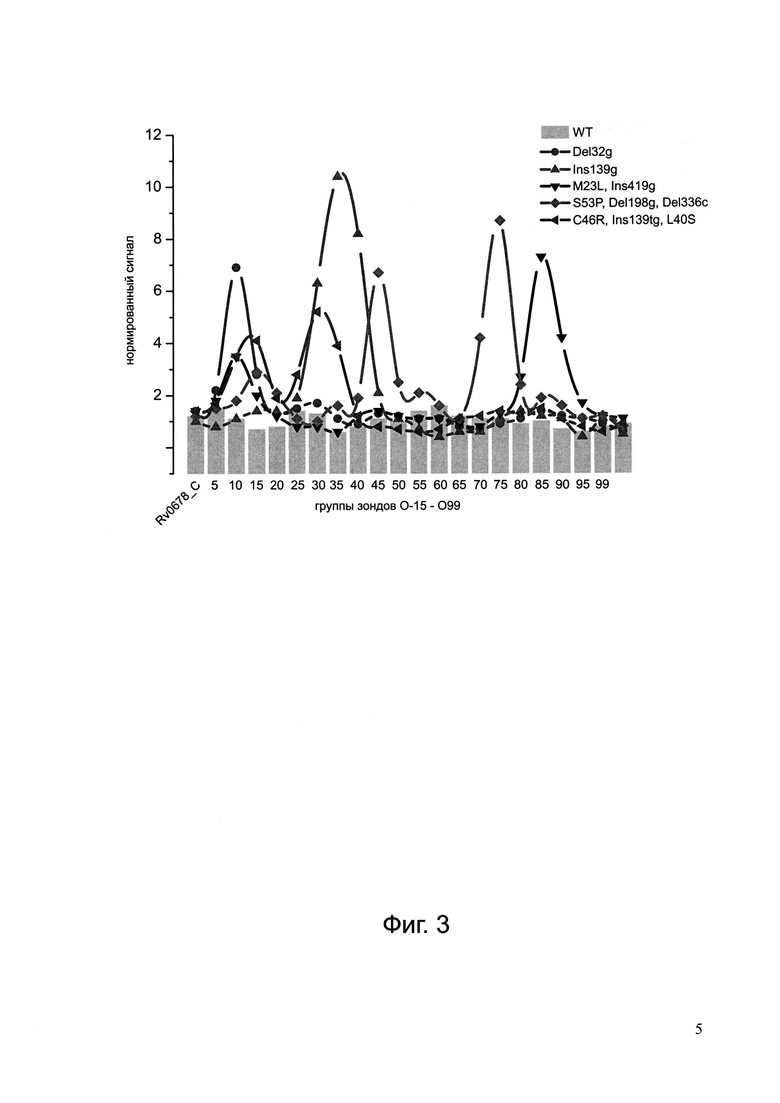
Фиг. 3. Нормированные гибридизационные профили, полученные при анализе последовательности гена Rv0678 в образцах ДНК с различными мутациями в данном гене. Нормированный гибридизационный профиль образца ДНК с последовательностью дикого типа по гену Rv0678 показан серыми столбцами.
Фиг. 4. Флуоресцентная картина гибридизации биочипа и результаты интерпретации при анализе ДНК, выделенной из клинического образца, содержащего штамм М. tuberculosis, чувствительный к бедаквилину и линезолиду.
Фиг. 5. Флуоресцентная картина гибридизации биочипа и результаты интерпретации при анализе ДНК, выделенной из клинического образца, содержащего резистентный к бедаквилину штамм М. tuberculosis, обладающий мутацией Del32g в гене Rv0678.
Фиг. 6. Флуоресцентная картина гибридизации биочипа и результаты интерпретации при анализе ДНК, выделенной из клинического образца, содержащего резистентный к бедаквилину штамм М. tuberculosis, обладающий, мутациями S53P, Del198g, Del336c в гене Rv0678.
Фиг. 7. Флуоресцентная картина гибридизации биочипа и результаты интерпретации при анализе ДНК, выделенной из клинического образца, содержащего резистентный к бедаквилину штамм М. tuberculosis, обладающий мутациями M23L и ins419g в гене Rv0678 и A63V в гене atpE.
Фиг. 8. Флуоресцентная картина гибридизации биочипа и результаты интерпретации при анализе ДНК, выделенной из клинического образца, содержащего резистентный к бедаквилину и линезолиду штамм М. tuberculosis, обладающий мутацией ins139g в гене Rv0678 и мутацией t460c в гене rplC.
Фиг. 9. Флуоресцентная картина гибридизации биочипа и результаты интерпретации при анализе ДНК, выделенной из клинического изолята, содержащего резистентный к линезолиду штамм М. tuberculosis, обладающий мутациями g2294a в гене 23S рРНК и t460c в гене rplC.
Осуществление изобретения
В заявленном способе предложено использование мультиплексной амплификации и флуоресцентного маркирования фрагментов микобактериального генома с получением флуоресцентно-меченных ПЦР-продуктов, при этом в качестве матрицы может быть использована геномная ДНК, выделенная из клинического респираторного материала, такого как мокрота, промывные воды или аспират бронхов, бронхоальвеолярный лаваж, экссудат, а также геномная ДНК, выделенная из клинических изолятов - культур клеток микобактерий, выращенных в жидкой или на твердой средах. Заявленный способ также предусматривает использование оригинального олигонуклеотидного микрочипа с иммобилизованными специфическими олигонуклеотидными зондами, процедуры гибридизации, регистрации и интерпретации результатов.
Принципиальная схема анализа микобактериальной ДНК с целью определения генетических детерминант резистентности возбудителя туберкулеза к бедаквилину и линезолиду с помощью олигонуклеотидного микрочипа.
Клинический образец подвергали деконтаминации и лизису клеток с целью обеспечения доступа к геномной ДНК. Одним из пригодных способов является разжижение в щелочных условиях в присутствии N-ацетил-L-цистеина и кипячение с детергентом для обеспечения доступа к ДНК и деконтаминации образца (Kent, Р.Т., and G.P. Kubica. Public health mycobacteriology. A guide for level III laboratory. 1985 Centers for Disease Control and Prevention, Atlanta, GA). Для этих целей также могут быть использованы иные способы, известные специалистам в данной области, такие как разрушение клеток с помощью ультразвука (Padilla Е, Gonzalez V, Manterola JM, et al. Evaluation of two different cell lysis methods for releasing mycobacterial nucleic acids in the INNO-LiPA mycobacteria test. 2003. Diagnostic Microbiology and Infectious Diseases. 46(1): 19-23), лизис при помощи гуанидина тиоцианата - саркозина (Chakravorty S, Tyagi JS. Novel use of guanidinium isothiocyanate in the isolation of Mycobacterium tuberculosis DNA from clinical material. 2001. FEMS Microbiology Letters. 205(1): 113-7.) и т.д. Очистку геномной ДНК после проведения лизиса проводили с использованием автоматических роботизированных станций, например Freedom EVO® Clinical (Tecan Group Ltd., Германия), или коммерческих наборов, таких, например, как «ПРОБА-НК» (ООО «НПО «ДНК-Технология», Регистрационное удостоверение Росздравнадзора № ФСР 2008/02938), «Реагент в пробирках для выделения ДНК из биопроб с целью последующего анализа методом полимеразной цепной реакции (ДНК-ЭКСПРЕСС)» (ООО «НПФ «Литех», Регистрационное удостоверение Росздравнадзора ФСР 2007/00362), Комплект реагентов для выделения РНК/ДНК из клинического материала «РИБО-преп» по ТУ 9398-071-01897593-2008 (ФБУН ЦНИИ Эпидемиологии Роспотребнадзора, Регистрационное удостоверение Росздравнадзора ФСР 2008/03147).
Полученный препарат микобактериальной геномной ДНК использовали в качестве матрицы в мультиплексной ПЦР. В ходе амплификации происходила наработка фрагментов генов Rv0678, atpE, 23SрРНК, rplC.
Праймеры для проведения мультиплексной ПЦР выбирали таким образом, чтобы они фланкировали кодирующую область гена Rv0678, а также сегменты генов atpE, 23S рРНК, rplC, где находятся наиболее часто встречающиеся мутации, ассоциированные с резистентностью микроорганизма к бедаквилину или линезолиду. Используя специализированное программное обеспечение, например Oligo v. 6.3 (Molecular Biology Insights Inc., США) или Fast PCR (http://www.biocenter.helsinki.fi/bi/Programs/fastpcr.htm) или другие коммерчески доступные программы, или программы, свободно доступные в сети Internet, рассчитывали температуры плавления праймеров и, варьируя их длину, добивались того, чтобы разброс температур отжига праймеров внутри набора не превышал 3-4°C. При подборе праймеров избегали таких последовательностей, которые способны формировать вторичные структуры типа шпильки с высокими температурами плавления, а также последовательностей, образующих между собой дуплексы, состоящие более чем из трех-пяти нуклеотидов. Каждый выбранный праймер должен обладать уникальной специфичностью в отношении анализируемого участка последовательности генома микобактерий туберкулезного комплекса. Специфичность праймеров проверяли с помощью программного обеспечения, использующего поиск в базах нуклеотидных последовательностей по алгоритму BLAST (например, www.ncbi.nlm.nih.gov/BLAST).
Для обеспечения эффективной амплификации с одновременным введением флуоресцентной метки во все вышеперечисленные сегменты микобактериального генома в едином реакционном объеме использовали праймеры следующей конструкции. Последовательность каждого добавляемого в реакционную смесь праймера состояла из двух частей - 3'-специфичной, т.е. комплементарной последовательности фрагмента генома М. tuberculosis, и 5'-универсальной (адаптерной), различавшейся для прямых и обратных праймеров. Помимо таких праймеров, содержащих и специфические, и адаптерные последовательности, в реакционную смесь добавляли два праймера, последовательности которых комплементарны последовательностям адаптерной части составных праймеров (SEQ ID NO: 1, 2). Данные адаптерные праймеры присутствовали в реакционной смеси в различных концентрациях, с целью наработки преимущественно одноцепочечных фрагментов, последовательности которых комплементарны последовательностям олигонуклеотидов, иммобилизованных на биочипе.
Расчетные температуры плавления специфичных и адаптерных последовательностей выбирали равными 70°C и 55°C, соответственно, а профиль амплификации включал две стадии по 30 циклов каждая, с температурами отжига 64°C на первой, и 50°C на второй стадии.
Таким образом, в ходе ПЦР в едином реакционном объеме на первой стадии за счет гибридизации и элонгации составных праймеров при использовании геномной ДНК в качестве матрицы происходила наработка двуцепочечньгх ПЦР-продуктов, содержащих на концах последовательности, специфичные к адаптерным праймерам, а затем, в ходе второй стадии, полученные ПЦР-продукты служили матрицей для наработки одноцепочечных фрагментов при использовании адаптерных праймеров с меньшей температурой отжига (Zimenkov D.V., Kulagina E.V., Antonova O.V., Zhuravlev V.Y. and Gryadunov D.A. Simultaneous drug resistance detection and genotyping of Mycobacterium tuberculosis using a low-density hydrogel microarray. 2016. Journal of antimicrobial chemotherapy. 71(6): 1520-1531).
Флуоресцентное маркирование исследуемых фрагментов микобактериального генома проводили, добавляя в ПЦР-смесь флуоресцентный субстрат - конъюгат дезоксиуридинтрифосфата и красителя индодикарбоцианинового ряда, с длиной волны возбуждения, равной (640±5) нм и длиной волны флуоресценции, равной (665±5) нм. В ходе ПЦР данный субстрат встраивался Taq-ДНК-полимеразой в растущую цепь ДНК, обеспечивая на выходе флуоресцентно-меченные ПЦР-продукты, анализируемые далее посредством гибридизации на биочипе (Kuznetsova V.E., Spitsyn М.А., Shershov V.E., Guseinov Т.О., Fesenko E.E., Lapa S.A., Ikonnikova A.Y., Avdonina M.A., Nasedkina T.V., Zasedatelev A.S. and Chudinov A.V. Novel fluorescently labelled nucleotides: synthesis, spectral properties and application in polymerase chain reaction. 2016. Mendeleev Communications. 26(2): 95-98).
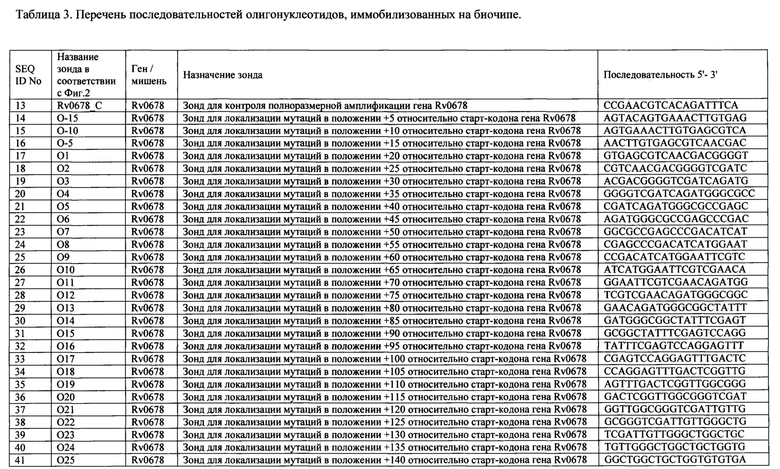
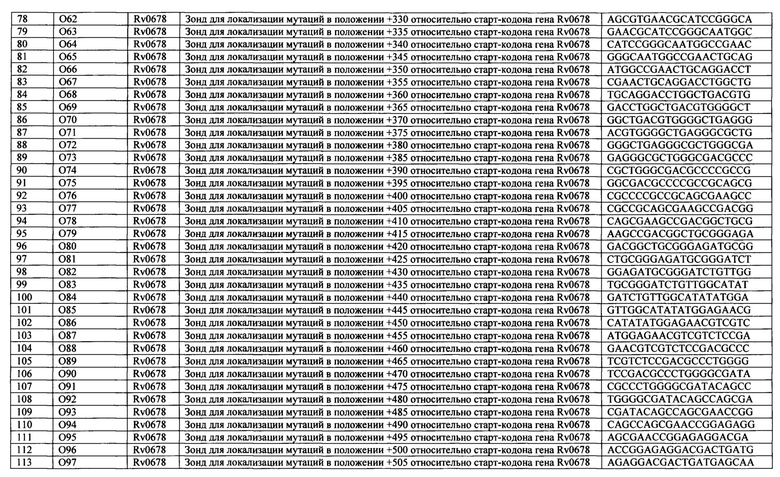
Выбор дискриминирующих олигонуклеотидов осуществляли согласно двум различным алгоритмам в зависимости от анализируемой мишени. Для гена Rv0678, в котором в любом положении кодирующей области спонтанно возникают точечные мутации, делеций, инсерций, приводящие к резистентности к бедаквилину, конструировали олигонуклеотиды длиной 20 оснований каждый, соответствующие последовательности дикого типа, последовательно перекрывающие весь ген Rv0678 таким образом, что совпадение последовательности 5'-концевой части каждого следующего зонда составляет не менее 75% с последовательностью предыдущего (Фиг. 1). Такой набор олигонуклеотидов (SEQ ID NO: 13-115) позволил определить наличие/отсутствие детерминант неустановленной локализации по всей длине гена с большей точностью, чем описанные ранее модели «рамочных» микрочипов (Volokhov DV, Chizhikov VE, Denkin S, Zhang Y. Molecular detection of drug-resistant Mycobacterium tuberculosis with a scanning-frame oligonucleotide microarray. 2009. Methods in Molecular Biology. 465:395-41; Akama T, Suzuki K, Tanigawa K, Kawashima A, Wu H, Nakata N, Osana Y, Sakakibara Y, Ishii N. Whole-genome tiling array analysis of Mycobacterium leprae RNA reveals high expression of pseudogenes and noncoding regions. 2009. Journal of Bacteriology. 191(10):3321-7.), включая возможность идентификации сочетанных мутаций и смесей ДНК дикого типа и/или мутантных вариантов.
С целью идентификации генетических детерминант установленной локализации в генах atpE, 23S рРНК, rplC для каждой позиции, для которой известны мутации, конструировали набор специфичных дискриминирующих олигонуклеотидов, способный выявлять известные варианты замен. Последовательности зондов для иммобилизации на биочипе подбирали с учетом размера и сложности анализируемой последовательности и, в частности, наличия повторов и протяженных гомополимерных последовательностей. Определяли длину дискриминирующих олигонуклеотидов, обеспечивающую их специфичность в отношении анализируемой последовательности. Используя программное обеспечение, например Oligo v. 6.3 (Molecular Biology Insights Inc., США), рассчитывали температуры плавления олигонуклеотидов и, варьируя их длину, добивались того, чтобы разброс температур плавления олигонуклеотидов составлял не более 2-3°C. Избегали таких олигонуклеотидов, которые способны формировать вторичные структуры типа шпильки с высокими температурами плавления. Положение определяемых вариабельных нуклеотидов и других нуклеотидных перестроек выбирали по возможности не далее 1-4 нуклеотида от середины соответствующего дискриминирующего олигонуклеотида.
Дискриминирующие олигонуклеотиды иммобилизовали в гелевых элементах, которые наносились на подложку формата предметного стекла в виде капель диаметром от 50 до 100 мкм с периодом 50-100 мкм, без использования специальных приспособлений, например, кварцевых масок. В качестве материала подложки использовали полимеры (полипропилен, полиэтилен, полибутилентерефталат, полиметакрилат, поликарбонат, полистирол), либо стекло. Совместная полимеризация олигонуклеотидов с основными компонентами геля обеспечивалась за счет действия ультрафиолетового излучения. В результате этой реакции иммобилизуемые молекулы ковалентно присоединялись к мономерам растущей полимерной цепи и равномерно распределялись во всем объеме каждой гелевой ячейки (Rubina AY,  SV, Dementieva EI et al. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production. 2004. Analytical Biochemistry. 325: 92-106).
SV, Dementieva EI et al. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production. 2004. Analytical Biochemistry. 325: 92-106).
ПЦР-продукты, полученные на стадии ПЦР, гибридизовали на дифференцирующем биочипе с иммобилизованными олигонуклеотидами, комплементарными последовательностям изучаемых на предмет наличия мутаций генов. Гибридизацию проводили в растворе, содержащем буферный компонент для поддержания рН, соль для создания ионной силы и хаотропный (дестабилизирующий водородные связи) агент, в герметичной гибридизационной камере при температуре, зависящей от температуры плавления иммобилизованных на микрочипе дискриминирующих олигонуклеотидов. В качестве дестабилизирующего водородные связи агента использовали, например, гуанидин тиоцианат, мочевину или формамид. Выбор оптимальной температуры гибридизации проводили с учетом удобства практического применения системы. Дискриминирующие олигонуклеотиды, заявленные в настоящем изобретении, обладали температурой плавления в интервале от 42 до 44°C, что позволило проводить гибридизацию при 37°C с использованием хаотропного агента. Температура 37°C удобна тем, что большинство клинических лабораторий оснащены термостатами, поддерживающими эту температуру.
Анализируемые фрагменты генов atpE, 23S рРНК, rplC образовывали совершенные гибридизационные дуплексы только с соответствующими (полностью комплементарными) олигонуклеотидами. Со всеми остальными олигонуклеотидами изучаемые фрагменты ДНК формировали несовершенный дуплекс. Дискриминацию совершенных и несовершенных дуплексов выполняли путем сравнения интенсивностей флуоресценции ячеек, в которых образовались дуплексы. Интенсивность сигнала в ячейке, в которой образовался совершенный гибридизационный дуплекс (Iсов) выше, чем в таковой, где образовался несовершенный дуплекс (Iнесов). Проведение гибридизации при оптимальных условиях (температура, подобранная концентрация хаотропного агента и ионная сила гибридизационного буфера) позволили добиться соотношения Iсов/Iнесов≥2 между двумя ячейками, содержащими зонды, принадлежащие одной группе, и различающиеся на один нуклеотид.
В каждой из групп ячеек, содержащих олигонуклеотиды, специфичные к одному из генов atpE, 23S рРНК, rplC, присутствовала ячейка, содержащая зонд дикого типа (обведена толстой линией на схеме размещения олигонуклеотидов, Фиг. 2) и ячейки, содержащие зонды на возможные мутации. Для каждой группы ячеек регистрировали максимальный сигнал, превосходящий остальные более чем в 2 раза (проверяли соотношение Iсов/Iнесов≥2). Если максимальный сигнал фиксировали в ячейке, соответствующей ДНК дикого типа (без мутаций, т.е., принадлежащей микроорганизму, чувствительному к лекарственному препарату), то считали, что по данной аминокислотной/нуклеотидной позиции (группе ячеек) изучаемый образец мутаций не имел. Если максимальный сигнал регистрировали в ячейке, соответствующей ДНК с мутацией (мутациями) (т.е., принадлежащей микроорганизму, устойчивому к лекарственному препарату), то считали, что по данной аминокислотной/нуклеотидной позиции (группе ячеек) изучаемый образец имел аминокислотную/нуклеотидную замену, приводящую к возникновению резистентности. Изучаемый образец ДНК признавали принадлежащим к чувствительному штамму микобактерий, если по каждой вариабельной аминокислотной/нуклеотидной позиции (группе ячеек) он был охарактеризован как относящийся к дикому типу. Изучаемый образец ДНК признавали принадлежащим к устойчивому штамму микобактерий, если, как минимум, по одной вариабельной аминокислотной/нуклеотидной позиции (группе ячеек) образец был охарактеризован как имеющий мутацию, приводящую к возникновению резистентности. Для таких образцов дополнительно выясняли тип препарата (бедаквилин/линезолид), к которому обнаружена устойчивость, установив группу, по которой образец отнесен к резистентному типу.
Интенсивность сигналов ячеек с дуплексами, образованными в результате гибридизации ПЦР-продукта, содержащего полноразмерный фрагмент кодирующей области гена Rv0678, и иммобилизованными олигонуклеотидами 'O-15'-'O99' (Фиг. 2), перекрывающими последовательность данного локуса, зависела от степени гомологии прогибридизовавшихся последовательностей, вторичных структур гибридизуемого ПЦР-продукта и иммобилизованного зонда, концентрации исходной ДНК-матрицы. В общем случае, регистрировали гибридизационный профиль в виде набора интенсивностей сигналов ячеек с дуплексами, образовавшимися при гибридизации последовательности гена Rv0678 анализируемого образца. Для оценки наличия/отсутствия точечных мутаций, делеций, или инсерций в данном профиле, в каждой серии исследуемых образцов проводили анализ образца микобактериальной ДНК штамма дикого типа H37Rv посредством постановки мультиплексной ПЦР в отдельной пробирке и гибридизации на биочипе с получением эталонного гибридизационного профиля, соответствующего ДНК дикого типа. Выполняя нормировку сигналов каждого элемента эталонного профиля на соответствующий сигнал элемента в профиле анализируемого образца устанавливали наличие/отсутствие мутаций определенной локализации с точностью, в среднем, до 20 нуклеотидов. В то время как нормированный гибридизационный профиль образца ДНК с последовательностью дикого типа по гену Rv0678 представлял собой набор сигналов со значениями 1,5±0,25 (серые столбцы на Фиг. 3), наличие мутаций(и) в последовательности данного гена приводило к падению сигнала в 2,5 и более раза в соответствующих элементах нормированного профиля, формируя пики - выбросы (Фиг. 3), тем самым свидетельствуя о присутствии детерминант(ы) резистентности к бедаквилину в последовательности гена Rv0678 анализируемого образца ДНК.
Данный алгоритм реализовывали в программном обеспечении анализатора флуоресценции биочипов, позволяющем проводить автоматическую регистрацию и интерпретацию результатов посредством захвата флуоресцентной картины гибридизации, вычисления сигнала в каждой ячейке, сравнения сигналов внутри групп и выдачи отчета об отсутствии/наличии детерминант резистентности к бедаквилину и/или линезолиду.
Полученные результаты могут быть использованы для включения бедаквилина и/или линезолида в режимы терапии туберкулеза конкретного пациента, мониторинга эффективности лечения и отмены данных препаратов в силу приобретения резистентности.
Далее изобретение будет проиллюстрировано примерами, которые предназначены для обеспечения лучшего понимания сущности заявленного изобретения, но не должны рассматриваться как ограничивающие данное изобретение.
Примеры
Пример 1. Обработка клинического образца/изолята и выделение ДНК. Проведение мультиплексной ПЦР и гибридизации на олигонуклеотидном микрочипе. Регистрация и интерпретация результатов гибридизации.
Обработка клинического образца и выделение ДНК.
1. Клинический респираторный образец (мокроту) смешивали в соотношении 1:1 по объему со свежеприготовленным 0,5% раствором N-ацетил-L-цистеина (NALC) в 2% NaOH. Образец тщательно перемешивали на вортексе и выдерживали при комнатной температуре в течение 20 мин. К образцу добавляли фосфатный буфер рН 6,8 в соотношении 1:5 по объему и центрифугировали в течение 30 мин при 3000 об/мин. При использовании спинномозговой жидкости проводили предварительное центрифугирование в течение 10 мин при 10000 об/мин.
2. Осадок клеток суспендировали в 1,5 мл ТЕ буфера (10 мМ Трис-НС1, 1 мМ ЭДТА), рН 8,0, и осаждали при 3000 об/мин в течение 30 мин. Отмывку повторяли еще раз.
3. К полученному осадку добавляли 30 мкл ТЕ буфера, рН 8,0, содержавшего 1% (об/об) Тритон Х-100, и выдерживали в сухом термостате при 95°C в течение 30 мин.
4. Образец центрифугировали при 10000 об/мин в течение 10 мин
5. Дальнейшую очистку ДНК проводили с использованием набора «ПРОБА-НК» (ООО «НПО «ДНК-Технология») согласно инструкции производителя. Полученный раствор ДНК в объем 50 мкл пригоден для проведения мультиплексной ПЦР.
Обработка клинического изолята и выделение ДНК.
Суспензию клеток клинического изолята, представляющего собой культуру, снятую с плотной или жидкой среды, промывали ТЕ-буфером (10 мМ Трис-HCl, 1 мМ ЭДТА, рН 8.0), осаждали центрифугированием при 10,000 g в течение 10 мин при 4°C и прогревали в ТЕ-буфере, содержавшем 1% Triton Х-100 при 95°C в течение 30 мин. Очистку микобактериальной ДНК из лизатов проводили с использованием набора «Проба-НК» («ДНК-Технология», Россия), либо на автоматизированной станции Freedom EVO® Clinical (Tecan Group Ltd., Германия).
Проведение мультиплексной ПЦР.
Для каждого анализируемого образца готовили стерильную пробирку вместимостью 0,2 мл для проведения ПЦР, маркируя их «N», где N - номер анализируемого образца. Добавляли одну пробирку 'K' для положительного контрольного образца.
Состав ПЦР-смеси в пробирках 'N' и 'К'
- 1X ПЦР-буфер для HS Taq ДНК полимеразы (ЗАО «Евроген», Россия»);
- 200 мкМ каждого dATP, dCTP, dGTP, dUTP (Евроген)
- 1 мкм флуоресцентного субстрата IMD515-dUTP (ООО «БИОЧИП-ИМБ», Россия)
- Смесь праймеров ПР. Последовательности праймеров и их концентрации в смеси представлены в Таблице 1.
- 5 ед. HS Taq ДНК полимеразы (ЗАО «Евроген», Россия»);
В 30 мкл ПЦР-смесей вносили 3 мкл раствора ДНК, полученного в п. 5 процедуры обработки клинического образца и выделения ДНК. К 30 мкл ПЦР-смеси в пробирке 'K' добавляли 3 мкл раствора ДНК лабораторного штамма М. tuberculosis H37R.V.

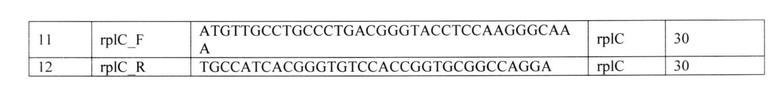
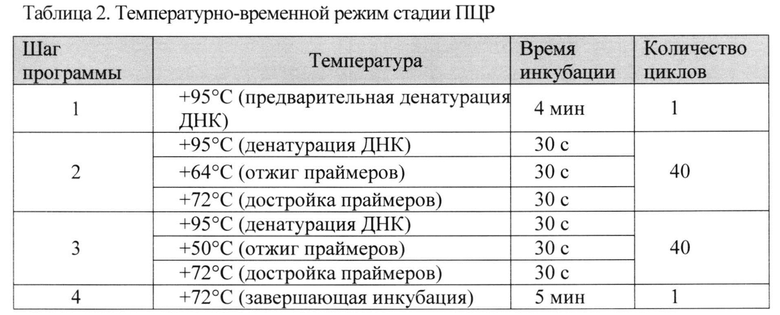
Мультиплексную ПЦР проводили на программируемом термостате S1000 (Bio-Rad, США) согласно температурному режиму, приведенному в Таблице 2.
Проведение гибридизации и отмывки на биочипе.
В пробирку объемом 0,5 мл вносили 10 мкл гибридизационного буфера (3М гуанидинтиоцианата, 150 мМ буфера HEPES рН 7,5, 15 мМ ЭДТА). Вносили 20 мкл реакционной смеси из пробирки 'N' после стадии ПЦР. Полученную смесь перемешивали на вортексе, капли собирали центрифугированием в течение 10 с при 1000 g.
30 мкл полученной гибридизационной смеси вносили в одно из отверстий реакционной камеры биочипа. Аналогичным образом проводили готовили гибридизационную смесь пробирки 'K' и вносили ее в камеру отдельного биочипа.
Гибридизацию проводили при 37°C в течение 6-12 часов. По окончании гибридизации биочип трижды промывали дистиллированной водой при 37°C, снимали гибридизационную камеру и высушивали биочип в потоке воздуха.
Регистрация и интерпретация результатов гибридизации.
Регистрацию флуоресцентного изображения микрочипа выполняли на универсальном аппаратно-программном комплексе (УАПК) (ООО «БИОЧИП-ИМБ». Россия) для анализа биологических микрочипов с использованием специализированного программного обеспечения 'ImageWare®' (ООО «БИОЧИП-ИМБ»). Интерпретацию результатов проводили согласно приведенному выше алгоритму, реализованному в модуле программного обеспечения для анализа флуоресцентных изображений микрочипов 'Imageware®'.
Пример 2. Олигонуклеотидный микрочип для идентификации генетических детерминант резистентности (точечных мутаций, инсерций, делеций) возбудителя туберкулеза к бедаквилину и линезолиду.
Синтез олигонуклеотидов проводили на автоматическом синтезаторе ABI-394 DNA/RNA synthesizer («Applied Biosystems», США) с использованием стандартного фосфорамидитного метода и очищали методом обращенно-фазовой ВЭЖХ (комплекс «Gilson», Франция). Олигонуклеотиды для иммобилизации в элементах микрочипа содержали спейсер со свободной аминогруппой, введенной в процессе синтеза с использованием 5'-Amino-Modifier С6 («Glen Research», США).
Иммобилизацию олигонуклеотидов на подложке биочипа в формате предметного стекла проводили, как было описано ранее ( SV, Chechetkin VR, Somova OG, Antonova OV, Moiseeva OV, Prokopenko DV, Yurasov RA, Gryadunov DA, Chudinov AV. Kinetic effects on signal normalization in oligonucleotide microchips with labeled immobilized probes. Journal of Biomolecular Structure and Dynamics. 2009 27(2):235-44).
Схема биочипа представлена на Фиг. 2. Биочип содержал 134 гелевых элемента, включая 128 ячейки с иммобилизованными олигонуклеотидами, 4 ячейки пустого геля с индексом '0', выполняющих роль отрицательного контроля и необходимых для вычисления фонового флуоресцентного сигнала и 2 ячейки с индексом «М», содержащие ковалентно связанный флуоресцентный краситель и использующиеся для автоматического вычисления интенсивности флуоресценции ячеек биочипа после гибридизации.
Биочип включал элементы с иммобилизованными зондами, в том числе:
а) олигонуклеотиды 'Rv0678_c', 'O-15'-'O99' для выявления наличия/отсутствия точечных мутаций, инсерций, делеций неустановленной локализации в кодирующей области гена Rv0678, ассоциированных с резистентностью к бедаквилину, последовательно перекрывающие весь ген Rv0678 таким образом, что совпадение последовательности 5'-концевой части каждого следующего зонда составляло не менее 75% с последовательностью предыдущего (перекрытие на 75%);
б) олигонуклеотиды для идентификации точечных мутаций определенной локализации в гене atpE (замены D28N/A/G, D32V, L59V, E61D/N, A63V/P, I66M), приводящих к резистентности к бедаквилину;
в) олигонуклеотиды для идентификации мутации в гене rplC (замена C154R в вариантах t460c/t460a+t462g), ассоциированной с резистентностью к линезолиду;
г) олигонуклеотиды для идентификации мутаций в гене 23 S рРНК (замены g2294a, g2814t), приводящие к резистентности к линезолиду.
Ячейки, содержащие олигонуклеотиды для выявления мутаций в генах atpE, rplC, 23S рРНК объединены в группы, соответствующие вариабельным аминокислотным остаткам или нуклеотидам в регуляторных областях генов. В каждой из групп присутствует элемент, содержащий олигонуклеотид, способный формировать совершенный гибридизационный дуплекс с ДНК, не имеющей мутаций (т.е. с ДНК дикого типа (WT)), в позициях, соответствующих следующим аминокислотным остаткам или нуклеотидам в регуляторных областях генов (такие ячейки обведены толстой линией на Фиг. 2). Остальные ячейки в группе содержат зонды, специфичные к различным мутациям, приводящим к замене аминокислоты или нуклеотида. Такие зонды формируют несовершенные гибридизационные дуплексы с ДНК дикого типа.
Перечень последовательностей олигонуклеотидов, иммобилизованных на биочипе, приведен в Таблице 3.


Пример 3. Анализ клинического образца, содержащего штамм М. tuberculosis, чувствительный к бедаквилину и линезолиду.
Клинический образец (мокрота), полученный от пациента, с подтвержденным микроскопией наличием кислотоустойчивых бактерий (БК+), был разделен на 2 части, одна из которых после деконтаминации (N-ацетил-L-цистеин и NaOH) и нейтрализации была передана для микробиологических исследований. Идентификацию микобактерий туберкулезного комплекса проводили по результатам биохимических тестов (Kent РТ, Kubica GP. Public health mycobacteriology. A guide for level III laboratory. Atlanta, GA: Centers for Disease Control and Prevention, 1985). Фенотипическую характеризацию изолята посредством определения минимальных ингибирующих концентраций, характеризующих устойчивость к бедаквилину и линезолиду, проводили в соответствии с опубликованными ранее протоколами (Torrea G, Coeck N, Desmaretz С, Van De Parre Т, Van Poucke T, Lounis N, de Jong ВС, Rigouts L. Bedaquiline susceptibility testing of Mycobacterium tuberculosis in an automated liquid culture system. 2015. Journal of Antimicrobial Chemotherapy. 70(8):2300-5; Zimenkov D, Nosova E, Kulagina E, Antonova O, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline-and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906). Значения критических концентраций по бедаквилину и линезолиду составили 0,06 мг/л и 1 мг/л, соответственно. Превышение данных значений свидетельствовало об резистентности изолята М. tuberculosis к данным противотуберкулезным препаратам. Вторую часть образца мокроты анализировали согласно изобретению, как описано в Примерах 1 и 2.
На Фиг. 4 представлен результат анализа ДНК, выделенной из данного клинического образца, и исследованной согласно изобретению.
Согласно алгоритму обработки результатов гибридизации анализ групп ячеек с зондами, специфичными к детерминантам резистентности в генах atpE, rplC, 23S рРНК, показал, что в каждой группе совершенный гибридизационный дуплекс зарегистрирован в ячейке, соответствующей дикому типу. Значения сигналов элементов биочипа в нормированном гибридизационном профиле (Фиг. 3, серые столбцы) по гену Rv0678 составляли в среднем 1,5±0,25, что соответствует дикому типу. Методом секвенирования гена Rv0678 у данного штамма подтверждена последовательность дикого типа.
Таким образом, ни в одном из исследуемых фрагментов генома М. tuberculosis мутации обнаружены не были. Значения минимальных ингибирующих концентраций (МИК) по бедаквилину и линезолиду составили 0,03 мг/л и 0,25 мг/л, соответственно, что характеризовало данный изолят как фенотипически чувствительный. Таким образом, полученный результат на биочипе об отсутствии детерминант резистентности к бедаквилину и линезолиду совпал с микробиологическими данными.
Пример 4. Анализ клинического образца, содержащего штамм М. tuberculosis, обладающий резистентностью к бедаквилину.
Клинический образец (экссудат), полученный от пациента, с подтвержденным микроскопией наличием кислотоустойчивых бактерий (БК+), был разделен на 2 части, одна из которых после деконтаминации (N-ацетил-L-цистеин и NaOH) и нейтрализации была передана для микробиологических исследований. Идентификацию микобактерий туберкулезного комплекса проводили по результатам биохимических тестов (Kent РТ, Kubica GP. Public health mycobacteriology. A guide for level III laboratory. Atlanta, GA: Centers for Disease Control and Prevention, 1985). Фенотипическую характеризацию изолята посредством определения минимальных ингибирующих концентраций, характеризующих устойчивость к бедаквилину и линезолиду, проводили в соответствии с опубликованными ранее протоколами (Torrea G, Coeck N, Desmaretz С, Van De Parre Т, Van Poucke T, Lounis N, de Jong ВС, Rigouts L. Bedaquiline susceptibility testing of Mycobacterium tuberculosis in an automated liquid culture system. 2015. Journal of Antimicrobial Chemotherapy. 70(8):2300-5; Zimenkov D, Nosova E, Kulagina E, Antonova O, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline-and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906). Значения критических концентраций по бедаквилину и линезолиду составили 0,06 мг/л и 1 мг/л, соответственно. Превышение данных значений свидетельствовало об резистентности изолята М. tuberculosis к данным противотуберкулезным препаратам. Вторую часть образца мокроты анализировали согласно изобретению, как описано в Примерах 1 и 2.
На Фиг. 5 представлен результат анализа ДНК, выделенной из данного клинического образца, и исследованной согласно изобретению.
Согласно алгоритму обработки результатов гибридизации анализ групп ячеек с зондами, специфичными к детерминантам резистентности в генах atpE, rplC, 23S рРНК, показал, что в каждой группе совершенный гибридизационный дуплекс зарегистрирован в ячейке, соответствующей дикому типу. В нормированном гибридизационном профиле (Фиг. 3, профиль  ) по гену Rv0678 максимальное значение сигнала в элементе 'O9' составило 7,55, тем самым был выявлен пик в области 25-45 нуклеотида относительно старт-кодона, свидетельствующий о наличии мутации в данной области гена Rv0678. Методом секвенирования гена Rv0678 в данном штамме была выявлена делеция гуанина (g) в позиции 32 относительно старт-кодона.
) по гену Rv0678 максимальное значение сигнала в элементе 'O9' составило 7,55, тем самым был выявлен пик в области 25-45 нуклеотида относительно старт-кодона, свидетельствующий о наличии мутации в данной области гена Rv0678. Методом секвенирования гена Rv0678 в данном штамме была выявлена делеция гуанина (g) в позиции 32 относительно старт-кодона.
Микробиологический анализ показал, что минимальная ингибирующая концентрация данного изолята по бедаквилину составила 0,12 мг/л, превышая значение критической концентрации 0,06 мг/л, характеризующее устойчивое состояние штамма. Значение МИК изолята по линезолиду, равное 0,5 мг/л, соответствовало чувствительной форме. Таким образом, полученный на биочипе результат о наличии детерминанты резистентности к бедаквилину и дикому типу по линезолиду совпал с микробиологическими данными.
Пример 5. Анализ клинического образца, содержащего штамм М. tuberculosis, обладающий резистентностью к бедаквилину.
Клинический образец (смыв бронхов), полученный от пациента, с подтвержденным микроскопией наличием кислотоустойчивых бактерий (БК+), был разделен на 2 части, одна из которых после деконтаминации (N-ацетил-L-цистеин и NaOH) и нейтрализации была передана для микробиологических исследований. Идентификацию микобактерий туберкулезного комплекса проводили по результатам биохимических тестов (Kent РТ, Kubica GP. Public health mycobacteriology. A guide for level III laboratory. Atlanta, GA: Centers for Disease Control and Prevention, 1985). Фенотипическую характеризацию изолята посредством определения минимальных ингибирующих концентраций, характеризующих устойчивость к бедаквилину и линезолиду, проводили в соответствии с опубликованными ранее протоколами (Torrea G, Coeck N, Desmaretz С, Van De Parre Т, Van Poucke T, Lounis N, de Jong ВС, Rigouts L. Bedaquiline susceptibility testing of Mycobacterium tuberculosis in an automated liquid culture system. 2015. Journal of Antimicrobial Chemotherapy. 70(8):2300-5; Zimenkov D, Nosova E, Kulagina E, Antonova O, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline-and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906). Значения критических концентраций по бедаквилину и линезолиду составили 0,06 мг/л и 1 мг/л, соответственно. Превышение данных значений свидетельствовало об резистентности изолята М. tuberculosis к данным противотуберкулезным препаратам. Вторую часть образца мокроты анализировали согласно изобретению, как описано в Примерах 1 и 2.
На Фиг. 6 представлен результат анализа ДНК, выделенной из данного клинического образца, и исследованной согласно изобретению.
Согласно алгоритму обработки результатов гибридизации анализ групп ячеек с зондами, специфичными к детерминантам резистентности в генах atpE, rplC, 23S рРНК, показал, что в каждой группе совершенный гибридизационный дуплекс зарегистрирован в ячейке, соответствующей дикому типу. В нормированном гибридизационном профиле (Фиг. 3, профиль  ) по гену Rv0678 максимальное значение сигнала в элементе 'O13' составило 3,1, тем самым был выявлен пик в области 40-60 нуклеотида относительно старт-кодона, свидетельствующий о наличии мутации в данной области гена Rv0678. Аналогичным образом были выявлены пики в областях 185-205 и 325-345 относительно старт-кодона, также свидетельствующие о наличии мутаций. Методом секвенирования гена Rv0678 в данном штамме были выявлены замены S53P, делеция гуанина (g) в позиции 198 и делеция цитозина (с) в позиции 336.
) по гену Rv0678 максимальное значение сигнала в элементе 'O13' составило 3,1, тем самым был выявлен пик в области 40-60 нуклеотида относительно старт-кодона, свидетельствующий о наличии мутации в данной области гена Rv0678. Аналогичным образом были выявлены пики в областях 185-205 и 325-345 относительно старт-кодона, также свидетельствующие о наличии мутаций. Методом секвенирования гена Rv0678 в данном штамме были выявлены замены S53P, делеция гуанина (g) в позиции 198 и делеция цитозина (с) в позиции 336.
Микробиологический анализ показал, что МИК данного изолята по бедаквилину составила 0,25 мг/л, превышая значение критической концентрации 0,06 мг/л, характеризующее устойчивое состояние штамма. Значение МИК изолята по линезолиду, равное 0,25 мг/л, соответствовало чувствительной форме. Таким образом, полученный на биочипе результат о наличии детерминант резистентности к бедаквилину и дикому типу по линезолиду совпал с микробиологическими данными.
Пример 6. Анализ клинического образца, содержащего штамм М. tuberculosis, обладающий резистентностью к бедаквилину.
Образец мокроты, полученный от пациента, с подтвержденным микроскопией наличием кислотоустойчивых бактерий (БК+), был разделен на 2 части, одна из которых после деконтаминации (N-ацетил-L-цистеин и NaOH) и нейтрализации была передана для микробиологических исследований. Идентификацию микобактерий туберкулезного комплекса проводили по результатам биохимических тестов (Kent РТ, Kubica GP. Public health mycobacteriology. A guide for level III laboratory. Atlanta, GA: Centers for Disease Control and Prevention, 1985). Фенотипическую характеризацию изолята посредством определения минимальных ингибирующих концентраций, характеризующих устойчивость к бедаквилину и линезолиду, проводили в соответствии с опубликованными ранее протоколами (Torrea G, Coeck N, Desmaretz С, Van De Parre Т, Van Poucke T, Lounis N, de Jong ВС, Rigouts L. Bedaquiline susceptibility testing of Mycobacterium tuberculosis in an automated liquid culture system. 2015. Journal of Antimicrobial Chemotherapy. 70(8):2300-5; Zimenkov D, Nosova E, Kulagina E, Antonova O, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline-and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906). Значения критических концентраций по бедаквилину и линезолиду составили 0,06 мг/л и 1 мг/л, соответственно. Превышение данных значений свидетельствовало об резистентности изолята М. tuberculosis к данным противотуберкулезным препаратам. Вторую часть образца мокроты анализировали согласно изобретению, как описано в Примерах 1 и 2.
На Фиг. 7 представлен результат анализа ДНК, выделенной из данного клинического образца, и исследованной согласно изобретению.
Анализ групп ячеек с зондами, специфичными к детерминантам резистентности в генах atpE, rplC, 23S рРНК, показал, что в группе ячеек, соответствующих вариабельной аминокислоте 63 в гене atpE, совершенный гибридизационный дуплекс зарегистрирован в ячейке, соответствующей мутации A63V. Следовательно, ДНК штамма содержит мутацию, ассоциированную с резистентностью к бедаквилину. В группах ячеек с зондами, специфичными к детерминантам резистентности к линезолиду, совершенные гибридизационные дуплексы были зарегистрированы только в ячейках, соответствующих дикому типу.
В нормированном гибридизационном профиле (Фиг. 3, профиль по гену Rv0678 максимальное значение сигнала в элементе 'O7' составило 3,67, тем самым был выявлен пик в области 15-25 нуклеотида относительно старт-кодона, свидетельствующий о наличии мутации в данной области гена Rv0678. Аналогичным образом был выявлен пик в области 410-430 относительно старт-кодона, также свидетельствующий о наличии мутации. Методом секвенирования гена Rv0678 в данном штамме были выявлены замены M23L, инсерция гуанина (g) в позиции 419.
Микробиологический анализ показал, что МИК данного изолята по бедаквилину составила 1,0 мг/л, превышая значение критической концентрации 0,06 мг/л, характеризующее устойчивое состояние штамма. Значение МИК изолята по линезолиду, равное 0,25 мг/л, соответствовало чувствительной форме. Таким образом, полученный результат о наличии детерминант резистентности к бедаквилину и дикому типу по линезолиду совпал с микробиологическими данными.
Пример 7. Анализ клинического образца, содержащего штамм М. tuberculosis, обладающий резистентностью к бедаквилину и линезолиду.
Образец промывных вод бронхов, полученный от пациента, с подтвержденным микроскопией наличием кислотоустойчивых бактерий (БК+), был разделен на 2 части, одна из которых после деконтаминации (N-ацетил-L-цистеин и NaOH) и нейтрализации была передана для микробиологических исследований. Идентификацию микобактерий туберкулезного комплекса проводили по результатам биохимических тестов (Kent РТ, Kubica GP. Public health mycobacteriology. A guide for level III laboratory. Atlanta, GA: Centers for Disease Control and Prevention, 1985). Фенотипическую характеризацию изолята посредством определения минимальных ингибирующих концентраций, характеризующих устойчивость к бедаквилину и линезолиду, проводили в соответствии с опубликованными ранее протоколами (Torrea G, Coeck N, Desmaretz С, Van De Parre Т, Van Poucke T, Lounis N, de Jong ВС, Rigouts L. Bedaquiline susceptibility testing of Mycobacterium tuberculosis in an automated liquid culture system. 2015. Journal of Antimicrobial Chemotherapy. 70(8):2300-5; Zimenkov D, Nosova E, Kulagina E, Antonova O, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline-and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906). Значения критических концентраций по бедаквилину и линезолиду составили 0,06 мг/л и 1 мг/л, соответственно. Превышение данных значений свидетельствовало об резистентности изолята М. tuberculosis к данным противотуберкулезным препаратам. Вторую часть образца мокроты анализировали согласно изобретению, как описано в Примерах 1 и 2.
На Фиг. 8 представлен результат анализа ДНК, выделенной из данного клинического образца, и исследованной согласно изобретению.
Анализ групп ячеек с зондами, специфичными к детерминантам резистентности в генах atpE, rplC, 23S рРНК, показал, что в группе ячеек, соответствующих вариабельной позиции 460 в гене rplC, совершенный гибридизационный дуплекс зарегистрирован в ячейке, соответствующей мутации t460c (замена тимина на цитозин). Следовательно, ДНК штамма содержит мутацию, ассоциированную с резистентностью к линезолиду. В других группах ячеек с зондами, специфичными к детерминантам резистентности к бедаквилину или линезолиду, совершенные гибридизационные дуплексы были зарегистрированы только в ячейках, соответствующих дикому типу.
В нормированном гибридизационном профиле (Фиг. 3, профиль  ) по гену Rv0678 максимальное значение сигнала в элементе 'О30' составило 10,72, тем самым был выявлен пик в области 130-150 нуклеотида относительно старт-кодона, свидетельствующий о наличии мутации в данной области гена Rv0678. Методом секвенирования гена Rv0678 в данном штамме была выявлена инсерция гуанина (g) в позиции 139.
) по гену Rv0678 максимальное значение сигнала в элементе 'О30' составило 10,72, тем самым был выявлен пик в области 130-150 нуклеотида относительно старт-кодона, свидетельствующий о наличии мутации в данной области гена Rv0678. Методом секвенирования гена Rv0678 в данном штамме была выявлена инсерция гуанина (g) в позиции 139.
Микробиологический анализ показал, что МИК данного изолята по бедаквилину составила 0,12 мг/л, превышая значение критической концентрации 0,06 мг/л, характеризующее устойчивое состояние штамма. Значение МИК изолята по линезолиду составило 2 мг/л, превышая значение критической концентрации 1,0 мг/л, характеризующее устойчивое состояние штамма. Таким образом, полученный результат о наличии детерминант резистентности к бедаквилину и линезолиду совпал с микробиологическими данными.
Пример 8. Анализ клинического изолята, содержащего штамм М. tuberculosis, обладающий резистентностью к линезолиду.
Клинический изолят (культура клеток) был разделен на 2 части, одна из которых была использована для фенотипической характеризации штамма посредством определения минимальных ингибирующих концентраций, характеризующих устойчивость к бедаквилину и линезолиду (Torrea G, Coeck N, Desmaretz С, Van De Parre Т, Van Poucke T, Lounis N, de Jong ВС, Rigouts L. Bedaquiline susceptibility testing of Mycobacterium tuberculosis in an automated liquid culture system. 2015. Journal of Antimicrobial Chemotherapy. 70(8):2300-5; Zimenkov D, Nosova E, Kulagina E, Antonova O, Arslanbaeva L, Isakova A, Krylova L, Peretokina I, Makarova M, Safonova S, Borisov S, Gryadunov D. Examination of bedaquiline- and linezolid-resistant Mycobacterium tuberculosis isolates from the Moscow region. 2017. Journal of antimicrobial chemotherapy. 72 (7): 1901-1906). Значения критических концентраций по бедаквилину и линезолиду составили 0,06 мг/л и 1 мг/л, соответственно. Превышение данных значений свидетельствовало об резистентности изолята М. tuberculosis к данным противотуберкулезным препаратам. Вторую часть клинического изолята анализировали согласно изобретению, как описано в Примерах 1 и 2.
На Фиг. 9 представлен результат анализа ДНК, выделенной из данного клинического образца, и исследованной согласно изобретению.
Анализ групп ячеек с зондами, специфичными к детерминантам резистентности в генах atpE, rplC, 23S рРНК, показал, что в группе ячеек, соответствующих вариабельной позиции 460 в гене rplC, совершенный гибридизационный дуплекс зарегистрирован в ячейке, соответствующей мутации t460c (замена тимина на цитозин). В группе элементов, соответствующих вариабельной позиции 2294 в гене 23S рРНК, совершенный гибридизационный дуплекс зарегистрирован в ячейке, соответствующей мутации g2294a (замена гуанина на аденин). Следовательно, ДНК штамма содержит мутации, ассоциированные с резистентностью к линезолиду. В других группе ячеек с зондами, специфичными к гену atpE, совершенные гибридизационные дуплексы были зарегистрированы только в ячейках, соответствующих дикому типу.
Значения сигналов элементов биочипа в нормированном гибридизационном профиле по гену Rv0678 составляли в среднем 1,3±0,4, что соответствует дикому типу. Методом секвенирования гена Rv0678 у данного штамма подтверждена последовательность дикого типа.
Микробиологический анализ показал, что МИК данного изолята по бедаквилину составила 0,3 мг/л, характеризуя штамм как чувствительный. Значение МИК изолята по линезолиду составило 32 мг/л, превышая значение критической концентрации 1,0 мг/л, характеризующее устойчивое состояние штамма. Таким образом, полученный результат об отсутствии детерминант резистентности к бедаквилину и наличии мутаций, приводящих к резистентности к линезолиду, совпал с микробиологическими данными.
Таким образом, представленное изобретение позволяет в короткие сроки выявлять наличие/отсутствие детерминант резистентности М. tuberculosis к бедаквилину и линезолиду в клиническом материале, получаемом от больных туберкулезом. В силу высокой стоимости этих противотуберкулезных препаратов, выявления предсуществующей резистентности и формирования резистентности в процессе терапии полученные данные могут быть использованы для включения бедаквилина и/или линезолида в ГУ и V режимы терапии конкретного пациента, мониторинга эффективности лечения и отмены данных препаратов в случае формирования резистентности.
Хотя предпочтительные воплощения настоящего изобретения и их преимущества были подробно описаны выше, специалист в данной области сможет внести различные изменения, дополнения или, наоборот, что-то опустить, не выходя при этом за рамки данного изобретения, которые определяются прилагаемой ниже формулой изобретения.








Изобретение относится к области медицины, молекулярной биологии и микробиологии. Предложен способ определения генетических детерминант резистентности возбудителя туберкулеза к бедаквилину и линезолиду, включающий мультиплексную амплификацию генов Rv0678, atpE, 23S рРНК, rplC, обеспечение олигонуклеотидного микрочипа для установления наличия/отсутствия точечных мутаций, инсерций, делеций в указанных генах, гибридизацию амплифицированных флуоресцентно-меченных продуктов на олигонуклеотидном микрочипе и регистрацию результатов гибридизации. Изобретение позволяет установить наличие/отсутствие детерминант неизвестной заранее локализации в последовательности гена Rv0678, а также идентифицировать мутации в генах atpE, 23S рРНК, rplC, ассоциированных с устойчивостью к бедаквилину и линезолиду. 5 з.п. ф-лы, 9 ил., 3 табл., 8 пр.
1. Способ определения генетических детерминант резистентности возбудителя туберкулеза к бедаквилину и линезолиду, включающий:
(а) мультиплексную амплификацию генов Rv0678, atpE, 23S рРНК, rplC с одновременным флуоресцентным маркированием при использовании геномной ДНК микобактерий туберкулезного комплекса в качестве матрицы и набора праймеров, последовательности которых представлены SEQ ID NO: 1-12, с получением флуоресцентно-меченных фрагментов микобактериального генома;
(б) обеспечение олигонуклеотидного микрочипа для установления наличия/отсутствия точечных мутаций, инсерций, делеций заранее неизвестной локализации в последовательности гена Rv0678, а также идентификации мутаций в генах atpE, 23S рРНК, rplC, ассоциированных с резистентностью к бедаквилину и линезолиду, представляющего собой подложку, содержащую множество дискретных элементов, в каждом из которых иммобилизован уникальный олигонуклеотидный зонд, имеющий последовательность, комплементарную последовательности одноцепочечного фрагмента, полученного на стадии (а), и выбранную из группы, включающей: а) соответствующие последовательности гена Rv0678 дикого типа таким образом, что совпадение последовательности 5'-концевой части каждого следующего зонда составляет не менее 75% с последовательностью предыдущего, при этом зонды длиной 20 нуклеотидов каждый полностью перекрывают последовательность кодирующей области гена Rv0678; б) соответствующие последовательности фрагмента гена atpE дикого типа; в) соответствующие последовательности фрагмента мутантного варианта гена atpE, приводящего к резистентности микроорганизма к бедаквилину; г) соответствующие последовательности фрагмента гена rplC дикого типа; д) соответствующие последовательности фрагмента мутантного варианта гена rplC, приводящего к резистентности микроорганизма к линезолиду; е) соответствующие последовательности фрагмента гена 23S рРНК дикого типа; ж) соответствующие последовательности фрагмента мутантного варианта гена 23S рРНК, приводящего к резистентности микроорганизмов к линезолиду; при этом последовательности иммобилизованных олигонуклеотидных зондов представлены SEQ ID NO: 13-140;
(в) гибридизацию амплифицированных флуоресцентно-меченных продуктов, полученных на стадии (а), на олигонуклеотидном микрочипе с образованием дуплексов с иммобилизованными зондами в условиях, обеспечивающих разрешение в один нуклеотид между образующимися в результате гибридизации совершенными и несовершенными дуплексами;
(г) регистрацию результатов гибридизации на олигонуклеотидном микрочипе, проведенной на стадии (в), с использованием анализатора флуоресценции и программного обеспечения, что позволяет использовать программную обработку интенсивностей сигналов с последующей интерпретацией результатов, которую проводят в два этапа: на первом этапе анализируют сигналы в группах элементов, содержащих зонды, соответствующие последовательностям генов atpE, 23S рРНК, rplC; на втором этапе анализируют гибридизационный профиль, включающий интенсивности сигналов элементов, содержащих зонды, соответствующие последовательности гена Rv0678.
2. Способ по п. 1, в котором амплификацию фрагментов микобактериального генома проводят, используя образец ДНК, выделенный из мокроты, экссудата, бронхоальвеолярного лаважа или промывных вод бронхов, или образец ДНК, выделенный из клинического изолята микобактерий.
3. Способ по п. 1, характеризующийся тем, что на стадии (а) для флуоресцентного маркирования получаемых ПЦР-фрагментов используют флуоресцентно-меченный дезоксиуридинтрифосфат.
4. Способ по п. 1, характеризующийся тем, что в каждой серии экспериментов на стадии (а) в отдельной пробирке проводят амплификацию и флуоресцентное маркирование образца микобактериальной ДНК штамма дикого типа H37Rv с последующей гибридизацией полученных флуоресцентно-меченных ПЦР-фрагментов на отдельном микрочипе на стадии (в) и регистрацией результатов гибридизации на стадии (г) с получением эталонного гибридизационного профиля, включающего интенсивности сигналов элементов, содержащих зонды, соответствующие последовательности гена Rv0678 дикого типа.
5. Способ по п. 1, характеризующийся тем, что на стадии (г) на первом этапе интерпретации анализируют сигналы групп элементов, содержащих зонды, специфичные к одному из генов atpE, 23S рРНК, rplC, таким образом, что если максимальный сигнал в группе зарегистрирован в элементе, содержащем зонд дикого типа, то считают, что по данной аминокислотной/нуклеотидной позиции данного гена изучаемый образец ДНК возбудителя туберкулеза мутаций не имеет, в противном случае, если максимальный сигнал в группе зарегистрирован в элементе, содержащем зонд, последовательность которого соответствует мутантному варианту одного из генов, то считают, что по данной аминокислотной/нуклеотидной позиции данного гена изучаемый образец ДНК возбудителя туберкулеза имеет мутацию, приводящую к резистентности к бедаквилину или линезолиду.
6. Способ по п. 4, характеризующийся тем, что на стадии (г) на втором этапе интерпретации результатов эталонный гибридизационный профиль нормируют на гибридизационные профили анализируемых образцов ДНК с неизвестной последовательностью гена Rv0678 с получением нормированных профилей, различающихся для последовательностей ДНК дикого типа и мутантных вариантов гена Rv0678, тем самым относя последовательность гена Rv0678 к дикому типу, либо к содержащей точечную мутацию, делецию, инсерцию, приводящую к резистентности к бедаквилину.

