Изобретение относится к биотехнологии, в частности к генетической инженерии, и представляет собой рекомбинантную плазмидную ДНК, обуславливающую синтез и секрецию эпидермального фактора роста человека (чЭФР) в клетках дрожжей и штамм дрожжей Saccharomyces cerevisiae - продуцент чЭФР.
Изобретение может найти применение в микробиологической промышленности и медицине, в частности для создания новых противоожоговых и ранозаживляющих средств, для диагностики злокачественных заболеваний ассоциированных с гиперпродукцией рецептора чЭФР, а также в создании новых методов адресной доставки тех или иных терапевтических средств в клетки-мишени, специфически экспрессирующие рецептор чЭФР.
ЧЭФР - гормон пептидной природы, продуцируемый разнообразными тканями и содержащийся практически во всех биологических жидкостях организма человека. ЧЭФР синтезируется в виде предшественника длиной 1217 аминокислот и подвергается дальнейшему проетеолитическому процессингу с образованием зрелого белка с характерным размером в 53 и с тремя внутренними дисульфидными связями [1] . ЧЭФР действует на ткани, содержащие соответствующий рецептор на своей поверхности и стимулирует их рост и деление. Наиболее мощным митогеном in vivo и in vitro чЭФР является для эпителиальных и соединительно-тканных клеток. Детальное изучение спектра влияний ЭФР указывает на возможные пути его биотехнологического применения в качестве ранозаживляющего, цитопротективного, противоязвенного препарата и др. [2-7].
К настоящему времени известны рекомбинантные плазмидные ДНК, обеспечивающие экспрессию чЭФР в клетках бактерий Escherichia coli, B.subtilis и дрожжей S. cerevisiae и Pichia pastoris [8-11]. Использование секреторного пути микроорганизмов для получения биологически активных пептидов млекопитающих по сравнению с другими способами их биосинтеза (например, в виде гибридных белков) обладает рядом преимуществ, а именно:
- в процессе секреции происходит правильное созревание и сворачивание рекомбинантного белка;
- секретируемый белок накапливается в значительном количестве во внеклеточном пространстве рекомбинантных штаммом и защищен от действия внутриклеточных протеаз, что значительно облегчает его последующее выделение и очистку.
Преимущество дрожжей-сахаромицетов как штаммов-продуцентов секретируемых биологически активных пептидов млекопитающих состоит в том, что эти микроорганизмы безопасны для человека, свободны от эндотоксинов и аллергенов, присутствующих, например, в составе клеточных стенок бактерий Escherichia coli. Полученные препараты рекомбинантного ЧЭФР могут использоваться для научных, терапевтических и косметологических целей, а также в качестве компонентов питательных сред для культур животных клеток.
Целью предлагаемого изобретения является получение рекомбинантного штамма дрожжей, продуцирующего зрелый и биологически активный чЭФР с высоким выходом в секретируемой форме. Поставленная цель достигается тем, что сконструирована рекомбинантная плазмида YEp40AB, обеспечивающая синтез и секрецию чЭФР в клетках дрожжей.
Для достижения поставленной цели предлагается также штамм Saccharomyces cerevisiae 20B12 ВКМ CR-349D, содержащий рекомбинантную плазмидную ДНК YEp40AB.
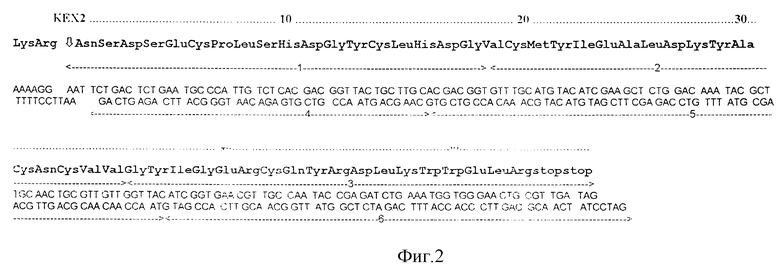
Сущность рекомбинантной плазмиды YEp40AB, чья генетическая карта представлена на фиг. 1, состоит в том, что она содержит синтетический ген ЧЭФР, встроенный в состав экспрессионного дрожжевого вектора YEpGPD/SEC, который в свою очередь содержит репликон плазмиды pUC18, репликон 2m ДНК дрожжей, промотор GPD1 гена дрожжей, сигнальную последовательность секреции и процессинга дрожжевого феромона альфа-фактора, терминатор гена PGK дрожжей, дрожжевой ген TRP1.
Плазмида YEp40AB состоит из следующих элементов:
- фрагмента EcoRI "тупой"-HindIII размером 2,6 тпн плазмиды pUC18
- фрагмента EcoRI "тупой"-PstI размером 2,0 тпн 2 m ДНК дрожжей
- фрагмента EcoRI "тупой"-PstI размером 0,9 тпн несущего ген TRPI дрожжей
- фрагмента HindIII "тупой" EcoRI размером 1,25 тпн, содержащего промотор гена GPD1 дрожжей, слитый с препросегментом гена MFa1 дрожжей;
- фрагмента ClaI-HindIII размером 0,3 тпн, содержащего терминатор гена PGK1 дрожжей
- фрагмента EcoRI-ClaI размером 0,15 тпн, содержащего ген ЧЭФР.
Молекулярный вес плазмиды YEp40AB составляет 4,5 Мд, что соответствует размеру в 7,2 тпн.
Число копий полученной плазмиды составляет 10-15 на клетку дрожжей.
Плазмидная ДНК YEp40AB содержит уникальные места для расщепления рестриктазами AatII, BglII, ClaI, EcoRI, по два сайта для рестриктаз PvuII, XbaI, положения которых указаны в табл. 1. Этот факт был установлен с помощью рестрикционного анализа.
Плазмида YEp40AB реплицируется в клетках E.coli за счет репликона pUC18. В клетках дрожжей репликацию плазмиды обеспечивает репликон 2 mcm ДНК дрожжей. Синтез ЧЭФР в клетках дрожжей осуществляется под контролем конститутивного промотора GPD1 дрожжей и сигналов процессинга и секреции гена MFα1 и достигается при культивировании трансформированного штамма на обычных селективных и полуселективных средах.
Плазмида YEp40AB содержит гены:
- модифицированный синтетический ген чЭФР, который кодирует синтез этого белка в дрожжевой клетке. Этот ген, введенный между местами расщепления EcoRI и ClaI, имеет нуклеотидную последовательность, показанную на фиг. 2. Эта последовательность кодирует зрелый чЭФР. Перед этим геном находится синтетическая последовательность LysArg, представляющая собой сайт специфического процессинга для дрожжевой эндопротеазы КЕХ2, входящая в состав препросегмента гена MFa1 дрожжей.
- bla-ген, обеспечивающий синтез b-лактамазы и влияющий на устойчивость штаммов E.coli, несущих плазмиду, к ампициллину
- TRPI-ген, обеспечивающий прототрофность по триптофану трансформантов реципиентного штамма дрожжей, несущего плазмиду.
Образуемый продукт представляет собой зрелый ЧЭФР, содержащий аминокислотных остатков.
Митотическая стабильность плазмиды YEpGPD1/EGF в штамме S.cerevisiae 20B12 при росте в селективных условиях составляет 90%.
Штамм-продуцент чЭФР получают трансформацией клеток дрожжей Saccharomyces cerevisiae 20B12 [a/a trp1 pep4-3] предложенной рекомбинантной плазмидной ДНК. Выбор штамма обусловлен тем, что он несет единственную ауксотрофную мутацию по гену trp1-289, позволяющую проводить отбор трансформантов плазмидой YEp40AB по маркеру trp1, а также тем, что он дефектен по синтезу вакуолярных протеаз, что существенно влияет на выход синтезируемых гетерологичных белков.
Полученный после селекции трансформированный клон 20B12/YEp40AB способен при культивировании продуцировать чЭФР. Уровень синтеза чЭФР в сконструированном штамме составляет 10 мг/л при титре культуры 5•108 кл/мл, что следует из данных определения чЭФР в супернатантах культуральной среды штамма-продуцента с помощью иммуноферментного анализа.
Полученный штамм дрожжей, несущий плазмиду YEp40AB, характеризуется следующими общими признаками:
Морфологические признаки. Клетки имеют форму от круглой до овальной, при делении почкующиеся.
Культуральные признаки.
Клетки хорошо растут на обычно используемых питательных средах. Время генерации около 90 мин в жидкой YNB-среде. На 2-2,5% питательном агаре "Difco" образуются круглые, гладкие, белые колонии с ровными краями. При выращивания на жидких YNB- и YEPD-средах образуется интенсивная ровная мутность.
Физиолого-биохимические признаки.
Оптимальная температура культивирования - от 28 до 30oC, оптимум pH - 6,6. В качестве источника углерода служит главным образом глюкоза, в некоторых случаях - этанол. Источником азота служат органические соединения (в виде пептона, аминокислот).
Способ получения рекомбинантной плазмиды YEp40AB иллюстрируется следующими примерами:
Пример 1. Конструирование промежуточной плазмиды pUC9/hEGFdBam
Плазмида pUC9/hEGF представляет собой вектор pUC9, содержащий синтетический ген эпидермального фактора роста человека [12], клонированный между сайтами EcoRI и BamHI полилинкера pUC9.
2 мкг ДНК плазмиды pUC9/hEGF линеаризовали с помощью рестриктазы BarnHI путем инкубирования в течение 120 мин при 37oC в высокосолевом буфере (100 mM NaCl, 50 mM Tris-HCl pH 7.5, 1 mM DTE, 10 mM MgCl2) с 10 единицами фермента. В реакционную смесь затем добавляли 2 мкл раствора, содержащего все четыре дезоксинуклеотидтрифосфата в концентрации по 2 mM каждого и 2 единицы Кленовского фрагмента ДНК-полимеразы I и продолжали инкубацию еще 30 мин при комнатной температуре.
Линеаризованную и "затупленную" по BamHI сайту плазмидную ДНК pUC9/hEGF освобождали от присутствующих белков и очищали с помощью набора GeneClean II (Bio101 Inc.) по инструкции фирмы-изготовителя.
100 нг полученного препарата в объеме 20 мкл лигировали в смеси состава 50 mM Tris-HCl pH 7.5, 10 mM MgCl2, 10 mM DTT, 1 mM ATP с добавлением T4-лигазы (2,5 единицы) в течение 3 часов при 16 градусах С.
200 мкл компетентных клеток штамма E.coli JM110 приготовленных стандартным способом [13] трансформировали 10 мкл лигазной смеси и трансформанты отбирали на чашках с питательным агаром, содержащим 50 мкг/мл ампициллина.
Плазмидную ДНК из полученных клонов выделяли с помощью щелочного метода [13] . Все плазмидные ДНК проверяли рестрикционным анализом на появление дополнительного сайта для рестриктазы ClaI, образуемого за счет "заполнения" сайта BamHI, как указано ниже:
1 мкг плазмидной ДНК инкубировали 2 часа при 37oC в 1х низкосолевом буфере с 2 единицами рестриктазы ClaI и полученные препараты анализировали в 1% агарозном геле. Те плазмиды, которые образовывали 3 тпн фрагмент ДНК, соответствовали правильной конструкции. Эта плазмида была обозначена как pUC/hEGFdBam.
Пример 2 Конструирование рекомбинантной плaзмиды YEp40AB для экспрессии и секреции чЭФР в клетках дрожжей
Конструирование осуществляли в соответствии со схемой, представленной на фиг. 1.
Плазмида YEpGPD/SEC [14] представляет собой вектор для экспрессии и секреции продуктов чужеродных генов в дрожжах.
Для получения "вектора" 10 мкг ДНК плазмиды YEpGPD/SEC инкубировали 2 часа при 37oC в высокосолевом буфере с 10 единицами EcoRI в объеме 20 мкл, полноту гидролиза контролировали электрофорезом в 1% агарозном геле. Затем смесь прогревали 15 мин при 70oC для инактивации рестриктазы и разбавляли в 5 раз 1х низкосолевым буфером. Далее добавляли 10 единиц рестриктазы ClaI и продолжали инкубацию еще 2 часа при 37 градусах.
Смесь депротеинизировали с помощью набора Gene CleanII и ДНК растворяли в 20 мкл ТЕ-буфера.
Для получения "фрагмента" 30 мкг плазмиды PUC9/hEGFdBam гидролизовали последовательно рестриктазами EcoRI и ClaI так же, как указано выше, пропорционально увеличив в три раза начальный объем инкубационной смеси. После депротеинизации с помощью набора Gene CleanII ДНК растворяли в 40 мкл ТЕ-буфера и разделяли с помощью электрофореза в 2,5% "Low-Melting" агарозном геле. Из геля выделяли фрагмент размером 160 пн, содержащий модифицированный ген hEGF.
0,2 мкг EcoRI-ClaI вектора YEpGPD/SEC лигировали с 0,1 мкг EcoRI - ClaI фрагмента модифицированного гена hEGF и трансформировали клетки штамма E. coli JM110, как описано в примере 1. Полученные клоны анализировали на правильность конструкции с помощью рестриктного анализа. Те клоны, которые содержали уникальный сайт BglII (внутренний сайт для гена hEGF), и при расщеплении с помощью других рестриктаз, образовывали характерные фрагменты, как указано в таблице, соответствовали правильной конструкции. Отобранная в результате плазмида, содержащая ген hEGF векторе YEpGPD/SEC, была обозначена как Yep40AB.
Пример 3. Получение штамма дрожжей 20B12, содержащего плазмиду Yep40AB
Клетки реципиентного штамма S.cerevisiae 20B12 [a trpl-189 pep4-3] трансформировали плазмидной ДНК YEpGPD/hEGF по методу Ито и др. [13]. Колонию дрожжей с пластинки агара переносили в 50 мл жидкой YEPD-среды и инкубировали в течение ночи при 30oC до плотности ОД660 1,0-2,0. Культуру центрифугировали и осадок промывали буфером следующего состава: 10 mM Tris-HCl, pH 7,5, 0,1 mM EDTA (TE-буфер). Промытые клетки ресуспендировали в 50 мл ТЕ-буфера, содержащего 0,1 М LiAc и инкубировали 2 часа при 30oC. После центрифугирования клетки суспендировали в 0,5 мл LiAc/ТЕ-буфера, добавляли 10-20 мкг плазмидной ДНК Yep40AB и смесь инкубировали 30 мин при 30oC. Затем добавляли 1 мл 50% PEG4000, перемешивали и инкубировали 1 час при 30oC. Затем следовал тепловой шок - 5 мин при 42oC. Клетки центрифугировали и промывали TE-буфером. Промытые клетки ресуспендировали в 500 мкл воды и высевали по 100 мкл на чашки Петри с селективным агаром (0,67% "Yeast Nitrogen Base". Difco, 2% глюкозы, 2% агара). Трансформанты появлялись через 2-4 дня инкубации при 30oC.
Пример 4. Тестирование чЭФР в супернатанах культуральной среды рекомбинантных штаммов.
Для анализа экспрессии производных чЭФР в трансформантах штамма 20B12 параллельно выращивали клоны 20B12/YEpGPD/hEGF, и в качестве отрицательного контроля - штамм 20B12/YEpGPD/SEC в 1 мл минимальной YNB-среды. После культивирования в течение 12 часов при 30oC по 0,5 ml суспензии клеток из девяти параллельных культур переносили в 10 мл YNB-среды с 0,5% казаминокислот и продолжали инкубирование в течение 48 часов. Супернатант культуральной среды получали после осаждения клеток.
Определение содержания белка со свойствами чЭФР проводили методом иммуноферментного анализа с использованием поли- и моноклональных антител. На 96-луночные иммунологические пластины адсорбировали по 50 нг поликлональных кроличьих антител против чЭФР в течение 8 часов при 4oC. Все места неспецифического связывания белка блокировали 0,5% обезжиренным сухим молоком в буфере состава 0,15 М NaCl, 10 mM NaPi pH 7,4, 0,1% Tween 20 (PBS/Tween) в течение 30 мин. Затем супернатанты (в различных разведениях) переносили на пластины. В качестве положительного контроля использовали рекомбинантный hEGF фирмы "Serva". Пластины инкубировали в течение 2 часов при 37oC. После многократной промывки добавляли анти-hEGF mAb. После промывки инкубировали пластины с козлиными антимышиными антителами, коньюгированными с пероксидазой хрена (разведение 1:500, 2 часа при 37oC. Связанная пероксидаза выявлялась путем окрашивания в 0,1 М цитрат-натрийфосфатном буфере pH 5,0, содержащем 0,16% ортофенилендиамин и 0,08% H2O2. Через 15 мин реакцию останавливали добавлением 50 мкл 3М серной кислоты и определяли интенсивность поглощения при длине волны 492 нм в приборе "Multiscan" (фирма Flow). Данные, полученные для различных образцов, усредняли и сопоставляли с контрольными образцами чЭФР фирмы "Serva". Определенный таким образом уровень экспрессии чЭФР в штамме 20B12/Yep40AB составлял 10 мг/л культуральной среды при плотности 5•108 кл/мл.
Пример 5. Выделение чЭФР из супернатантов культуральной среды.
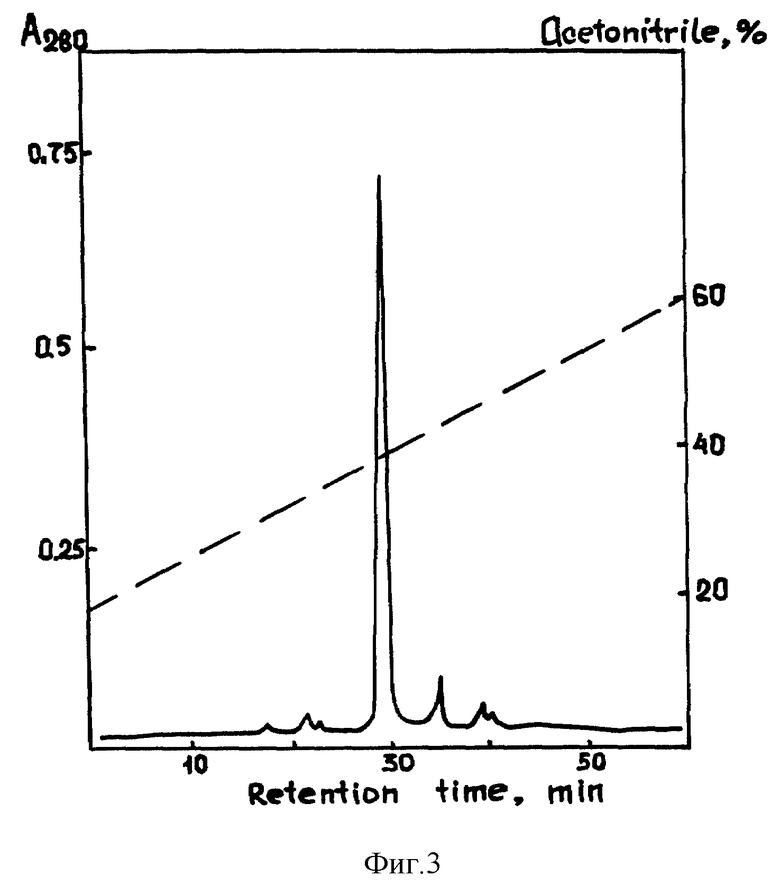
Культуральную среду, полученную после культивирования рекомбинантного штамма 20B12/Yep40AB в течение 48 часов в объеме 1 л среды YNB-CAA, центрифугировали (12 000 об/мин, 20 мин) и супернатант наносили на колонку (2х4 см) с Davisil C18 (90-130 мкм), уравновешенную 10% ацетонитрилом (2 объема). Колонку промывали 2 объемами 20% ацетонитрила с 0,1% ТФА. Рекомбинантный чЭФР элюировали 2 объемами 60% ацетонитрила, содержащего 0,1% ТФА при скорости потока 5 мл/мин. Элюат лиофилизировали с помощью Speed-Vac ("Savant") и проводили заключительную стадию очистки с помощью обращенно-фазовой высокоэффективной жидкостной хроматографии. Для этого препарат после лиофилизации растворяли в 0,1% ТФА, центрифугировали для отделения нерастворимых примесей и наносили на колонку mBondapak C18 (0,78 х 30 cm) фирмы "Waters". ЧЭФР элюировали с помощью линейного градиента концентрации ацетонитрила (0-60%) в 0,1% ТФА /фиг. 3/.
По данным SDS-электрофореза в 15% ПААГ чистота полученного препарата составляла более 98%. /фиг. 4/.
Литература
1. Scott J, Patterson S, Rall L, Bell GI, Crawford R, Penschow J, Niall H, Coghlan J The structure and biosynthesis of epidermal growth factor precursor. J Cell Sci Suppl 1985; 3:19-28
2. Schultz G, Khaw PT, Oxford K, MaCauley S, Van Setten G, Chegini N Growth factors and ocular wound healing. Eye 1994; 8(Pt 2): 184-187
3. Greenhalgh DG. The role of growth factors in wound healing. J Trauma. 1996 Jul 1; 41(1): 159-167.
4. Schultz G, et al. EGF and TGF-alpha in wound healing and repair. J Cell Biochem. 1991 Apr 1; 45(4): 346-352.
5. Cristiano RJ, et al. Epidermal growth factor mediated DNA delivery into lung cancer cells via the epidermal growth factor receptor. Cancer Gene Ther. 1996 Jan 1; 3(1): 4-10.
6. Jinno H, et al. Epidermal growth factor receptor-dependent cytotoxic effect by an EGF-ribonuclease conjugate on human cancer cell lines-a trial for less immunogenic chimeric toxin. Cancer Chemother Pharmacol. 1996 Jan 1; 38(4): 303-308.
7. Ohno К, et al. Multi-drug delivery system using streptavidin-transforming growth factoralpha chimeric protein. DNA Cell Biol. 1996 May 1; 15(5): 401-406.
8. Morioka-Fujimoto К, et al. Modified enterotoxin signal sequences increase secretion level of the recombinant human epidermal growth factor in Escherichia coli. J Biol Chem. 1991 Jan 25; 266(3): 1728-1732.
9. Yamagata H, Nakahama К., Suzuki Y, Kakinuma A, Tsukagoshi N, Udaka S Use of Bacillus brevis for efficient synthesis and secretion of human epidermal growth factor. Proc Natl Acad Sci USA 1989 May; 86(10): 3589-3593
10. Urdea MS, Merryweather JP, Mullenbach GT, Coit D, Heberlein U, Valenzuela P, Barr PJ Chemical synthesis of a gene for human epidermal growth factor urogastrone and its expression in yeast. Proc Natl Acad Sci USA 1983 Dec; 80(24): 7461-7465
11. Clare JJ, Romanes MA, Payment FB, Rowedder JE, Smith MA, Payne MM. Sreekrishna K, Henwood CA Production of mouse epidermal growth factor in yeast: high-level secretion using Pichia pastoris strains containing multiple gene copies. Gene 1991 Sep 15; 105(2): 205-212
12. Батчикова Н.В., Птицын Л.Р., Альтман И.Б., Степанов А.И., Ажаев А.Л. Химический синтез и клонирование гена эпидермального фактора роста человека Биоорган. Химия, 1988 т. 14, N 5 стр. 621-630.
13. Ausubel F.M., Brent R.G., Kingston R.E., Moore D.D., Seidman J.G., Smith J.A., Struhl K.A. Current Protocols in Molecular Biology. Massachusets General Hospital and Harvard Medical School, John Willey & Sons Inc., 1994.
14. Сидорович B.E., Королев С.В., Карпычев И.В., Эльдаров М.А., Скрябин К. Г. Набор векторов для направленного сортинга гетерологичных белков в дрожжах Saccharomyces cerevisiae. Биотехнология, 1996, N 2, стр. 10-17.


Изобретение относится к биотехнологии и может быть использовано для получения рекомбинантного эпидермального фактора роста человека (ЧЭФР). Рекомбинантный штамм Saccharomyces cerevisiae - продуцент ЧЭФР получают путем трансформации рекомбинантной плазмидой YEp40AB, направляющей синтез рекомбинантного эпидермального фактора роста человека и его экспорт в среду культивирования. Штамм Saccharomyces cerevisiae депонирован в ВКМ под номером ВКМ CR-349D. Штамм способен продуцировать ЧЭФР в количестве 10 мг на литр культуры продуцента с плотностью 5 • 108 кл/мл. Рекомбинантный ЧЭФР по своим биологическим и физико-химическим свойствам соответствует природному ЧЭФР. Изобретение позволяет получить биологически активный ЧЭФР с высоким выходом в секретируемой форме. 4 ил., 1 табл.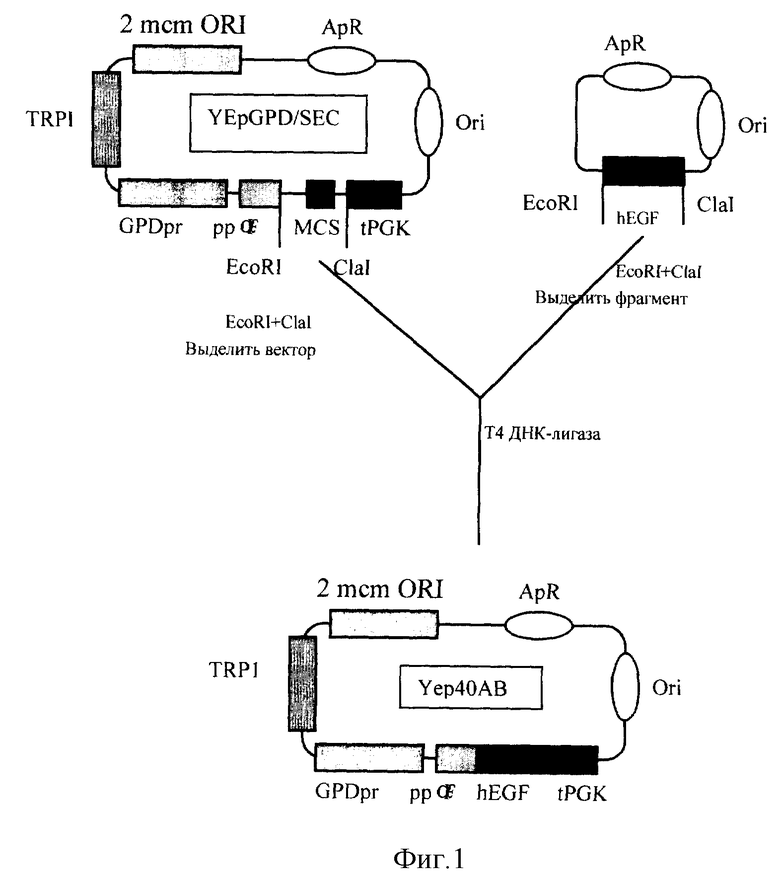
Штамм дрожжей Saccharomyces cerevisial 20 B 12, содержащий рекомбинантную плазмидную ДНК YEp 40 АВ, - ВКМ CR-349D - продуцент эпидермального фактора роста человека.
| Urdea M.S | |||
| et al | |||
| Proc.Natl.Acad | |||
| Sci USA, 1983, v.80, p.7461-7465 | |||
| Экономайзер | 0 |
|
SU94A1 |
| US 5162208, 10.11.92 | |||
| US 5521086, 28.05.96. | |||

