Группа изобретений относится к генной и тканевой инженерии и может быть использована в биотехнологии, медицине и сельском хозяйстве.
В настоящее время многие практические задачи, связанные с изменением свойств живого организма, могут быть решены методом генетической модификации. Такую модификацию клеток-мишеней можно осуществлять путем обеспечения транспорта экзогенного генетического материала через клеточную мембрану внутрь цитоплазмы клеток с последующей экспрессией закодированных в генетическом материале белков или с последующим взаимодействием генетического материала с внутриклеточными биополимерами с целью направленного изменения метаболизма модифицированных клеток.
При этом общей проблемой является доставка генетического материала к определенным тканям, органам, преодоление плазматической мембраны целевых клеток и транспорт к определенной мишени внутри клетки. Следует отметить, что поскольку любой генетический материал - потенциальный мутаген, очень важно предотвратить его попадание в другие, не нуждающиеся в трансформации, клетки организма.
Поэтому актуальной остается задача поиска пептидных векторов, которые способны обеспечить избирательный транспорт генетического материала в цитоплазму клеток-мишеней, а также последующий его транспорт внутрь их ядер, защитив по возможности генетический материал от внутриклеточной деградации. Особый практический интерес представляют быстрорастущие клетки млекопитающих - эпителиальные, эмбриональные и опухолевые. Большинство из них несет на поверхности рецепторы к специфическому белку, эпидермальному фактору роста (ЭФР). Поэтому перспективными в плане трансформации указанных клеток могут быть векторы-переносчики генетического материала, созданные на основе пептидных последовательностей, взаимодействующих с рецепторами к ЭФР.
Уровень технического состояния.
Наиболее простым в конструктивном отношении примером пептидного вектора для генетической трансформации клетки, несущей рецепторы к ЭФР, является комплекс, в котором последовательность эпидермального фактора роста, обеспечивающая специфическое взаимодействие с клетками-мишенями, ковалентно связана с выполняющим роль участка связывания нуклеиновой кислоты полилизином [1]. С помощью этого вектора осуществляют направленную доставку генетического материала в клетку млекопитающего, экспрессирующую на своей поверхности рецепторы к ЭФР. Однако описанный вектор и, соответственно, способ генетической модификации клетки с его использованием не предусматривают активного транспорта олигонуклеотидов в клеточное ядро, что может сказываться на сохранности экзогенного генетического материала внутри клетки.
Известен пептидный вектор, включающий последовательность TGF-альфа, специфически взаимодействующую с рецепторами к ЭФР; домен дрожжевого транскрипционного фактора GAL4 в качестве ДНК-связывающей области и домен транслокации экзотоксина A Pseudomonas для защиты вводимого в клетку генетического материала от разрушения в эндосомах [2]. Частично решая проблему сохранности в цитоплазме экзогенного материала, предлагаемый пептидный вектор имеет большой размер и включает два чужеродных для клетки белка, что при использовании его in vivo будет иметь следствием возникновение иммунного ответа. Кроме того, этот вектор, как и рассмотренный выше, применяется для генетической модификации клетки-мишени только в виде комплекса с поли-L-лизином, который обладает неспецифической токсичностью. Таким образом, использование данного вектора in vivo представляется неперспективным.
С учетом указанных недостатков закономерной представляется тенденция усовершенствования пептидных векторов к интересующим клеткам-мишеням, которая предусматривает использование особых пептидных последовательностей, совмещающих в себе способность связывать олигонуклеотиды и обеспечивать их специфический транспорт через ядерную мембрану, так называемых “сигналов ядерной локализации” (NLS). Примером такого пептида может служить сигнальная последовательность Т-антигена SV-40. Показана принципиальная возможность генетической модификации клетки позвоночных путем направленного транспорта в ядро экзогенного генетического материала, предварительно введенного в цитоплазму клетки с помощью микроинъекции [3]. Однако многочисленные исследования в этой области показали, что транспортная эффективность вектора, использующего NLS как для связывания, так и для проведения олигонуклеотидов в ядро, является сомнительной или, во всяком случае, неочевидной a priori, поскольку достоверно известно, что “проводящие” свойства NLS резко ослабевают в составе комплекса с генетическим материалом. Для того чтобы такие векторные системы обладали приемлемой эффективностью, соотношение вектора к олигонуклеотиду должно, по некоторым оценкам, быть выше 100:1 [4], что имеет следствием не только нежелательную “нагрузку” клетки инородным белком, но и возникновение пептид-пептидных взаимодействий, которые также снижают “транспортную” активность вектора.
Morris et al. ([5] - ближайший аналог) предложен способ генетической модификации фибробластов человека, предусматривающий использование двухкомпонентного пептидного вектора MPG, обеспечивающего направленный транспорт генетического материала в клетку-мишень и далее в ее ядро и состоящего из сигнала ядерной локализации большого Т-антигена SV-40 и пептида gp41 HIV-1. К основным недостаткам данного вектора и способа с его применением следует отнести низкую избирательность по отношению к клеткам-мишеням и недостаточную эффективность транспорта генетического материала в ядро, приемлемый уровень которого (по указанным выше причинам) достигается только при соотношении MPG-пептид: олигонуклеотид не менее 20:1.
В связи с этим основной задачей настоящего изобретения было создание простого по своей конструкции, эффективно действующего пептидного вектора, пригодного (не токсичного и не вызывающего иммунного ответа) для направленной генетической модификации in vivo клеток, несущих на своей поверхности рецепторы к ЭФР. Кроме того, учитывая, что химический синтез полипептидов далеко не всегда оправдывает себя в экономическом отношении, а также то, что при этом остается нерешенной задача постсинтетического фолдинга, в задачу настоящей работы входила разработка генно-инженерного способа получения значительных количеств такого вектора с правильно сформированной конформацией. При этом за основу был взят известный прием экспрессии чужеродного белка в Е. coli в виде гибридного продукта, в котором он слит с остатком модифицированного тиоредоксина (Glu30His, Gln62His) [6]. Дополнительные остатки His позволяют выделять слитый белок метало-хелат-аффинной хроматографией.
Раскрытие изобретения.
На основании известных результатов разработок в данной области исследований можно было предполагать, что решение задачи создания эффективного двухкомпонентного пептидного вектора, пригодного для генетической модификации in vivo клеток, активно экспрессирующих рецепторы к ЭФР, может быть найдено на пути поиска оптимальных для комбинирования (в плане природы аминокислотной последовательности и ее размера) последовательностей на основе NLS и последовательностей, обеспечивающих специфичное взаимодействие с рецепторами клетки-мишени, сочетание которых обладало бы минимальным уровнем неспецифических взаимодействий.
Такая комбинация компонентов пептидного вектора была найдена и представляла последовательно связанные между собой модифицированную последовательность сигнала Т-антигена SV-40, состоящую из 11 аминокислотных остатков, и аминокислотную последовательность эпидермального фактора роста человека из 53 аминокислотных остатков. В результате был получен новый пептидный вектор (из 64 аминокислот) с SEQ ID №1, получивший название PGE-k, который при осуществлении генетической трансформации обеспечивает целенаправленную доставку генетического материала к клеткам, несущим рецепторы к ЭФР, и транспорт его через клеточную и ядерную мембраны при молярном соотношении с олигонуклеотидом 3-10:1. Таким образом, настоящее изобретение обеспечивает повышение избирательности и эффективности способа генетической модификации рассматриваемых клеток-мишеней (технический результат).
Для получения нового пептидного вектора разработан способ, предусматривающий экспрессию в Е. coli химерной нуклеотидной последовательности (SEQ ID №5), кодирующей слитый белок (SEQ ID №4), состоящий из модифицированного тиоредоксина, PGE-k и объединяющей их линкерной последовательности, специфически расщепляемой энтерокиназой, а также необходимые для его осуществления рекомбинантная плазмида (pGEK) и рекомбинантный штамм Е. coli (BL21(DE3)/pLysS/pGEK, депонированный в ВКПМ под номером В-8389).
Таким образом, настоящее изобретение включает следующие объекты.
Первым объектом изобретения является пептидный вектор PGE-k, обеспечивающий избирательный транспорт экзогенного генетического материала в клетку млекопитающего, несущую наружные рецепторы к эпидермальному фактору роста, и последующее поступление его в ядро указанной клетки, с аминокислотной последовательностью SEQ ID №1, состоящей из производного аминокислотной последовательности эпидермального фактора роста человека (SEQ ID №2), которая является участком связывания с наружными рецепторами названной клетки, и последовательности сигнального пептида Т-антигена SV-40 (SEQ ID №3), которая отвечает за связывание экзогенного генетического материала и преодоление им ядерной оболочки.
Второй объект изобретения относится к нуклеотидной последовательности, кодирующей слитый белок, который состоит из мутантной формы тиоредоксина (Glu30His, Gln62His), линкерной последовательности -(Gly-Ser)2-Gly-(Asp)4-Lys-, специфически расщепляемой энтерокиназой, и последовательности пептидного вектора PGE-k и характеризуется аминокислотной последовательностью SEQ ID №4. Предпочтительно эта нуклеотидная последовательность представлена SEQ ID №5.
Третий объект изобретения - это рекомбинантная плазмида pGEK для экспрессии слитого белка, включающего пептидный вектор PGE-k, которая характеризуется тем, что имеет размер 3691 п.о. и состоит из
- NdeI/AatII фрагмента плазмиды pGEMEX-1, имеющего размер 2347 п.о. и содержащего участок инициации репликации (Ori) и ген устойчивости к ампициллину (Bla);
- AatII/HindIII фрагмента плазмиды pGEMEX-1, имеющего размер 755 п.о. и содержащего второй участок инициации репликации (f1Ori), и
- нуклеотидной последовательности SEQ ID №5.
Четвертым объектом заявленной группы является штамм Escherichia coli ВКПМ В-8389, содержащий плазмиду pGEK и являющийся продуцентом слитого белка, включающего пептидный вектор PGE-k.
Пятый объект изобретения относится к способу получения пептидного вектора, предусматривающему:
- культивирование рекомбинантного штамма Е. coli ВКПМ В-8389 в условиях, обеспечивающих экспрессию включающего пептидный вектор слитого белка,
- гидролиз полученного слитого белка энтерокиназой или другой протеазой, распознающей включенную в него линкерную последовательность и специфически расщепляющей пептидную связь, следующую за указанным линкером, и
- выделение целевого продукта.
Шестым объектом заявленной группы является способ генетической модификации клетки млекопитающего, несущей наружные рецепторы к эпидермальному фактору роста, предусматривающий контактирование названной клетки с экзогенным генетическим материалом в присутствии пептидного вектора PGE-k.
Описание чертежей.
На Фиг.1 представлена физическая и генетическая карта новой рекомбинантной плазмиды pGEK.
Обозначения: Bla - ген устойчивости к ампициллину, f1ori- участок начала репликации фага f1, T7p, T7t - промотор и терминатор полимеразы Т7, SP6p - промотор полимеразы SP6.
На Фиг.2 представлена нуклеотидная последовательность генов тиоредоксина (Glu30His, Gln62His), линкера и эпидермального фактора роста, проклонированных в плазмиду pGEMEX-1 по сайтам рестрикции Ndel (905)* и BamHI (42)*, где
 - фрагменты последовательности pGEMEX-1 (Promega);
- фрагменты последовательности pGEMEX-1 (Promega);
 - последовательность гена тиоредоксина (Glu30His. Gln62His);
- последовательность гена тиоредоксина (Glu30His. Gln62His);
 - последовательность гена линкера состава -(Gly-Ser)2-Gly-(Asp)4-Lys-
- последовательность гена линкера состава -(Gly-Ser)2-Gly-(Asp)4-Lys-
 - последовательность гена человеческого эпидермального фактора роста.
- последовательность гена человеческого эпидермального фактора роста.
* расположение сайта рестрикции по каталогу Promega.
На Фиг.3 представлена схема конструирования плазмиды pGEMEX-1/Thio-hEGF.
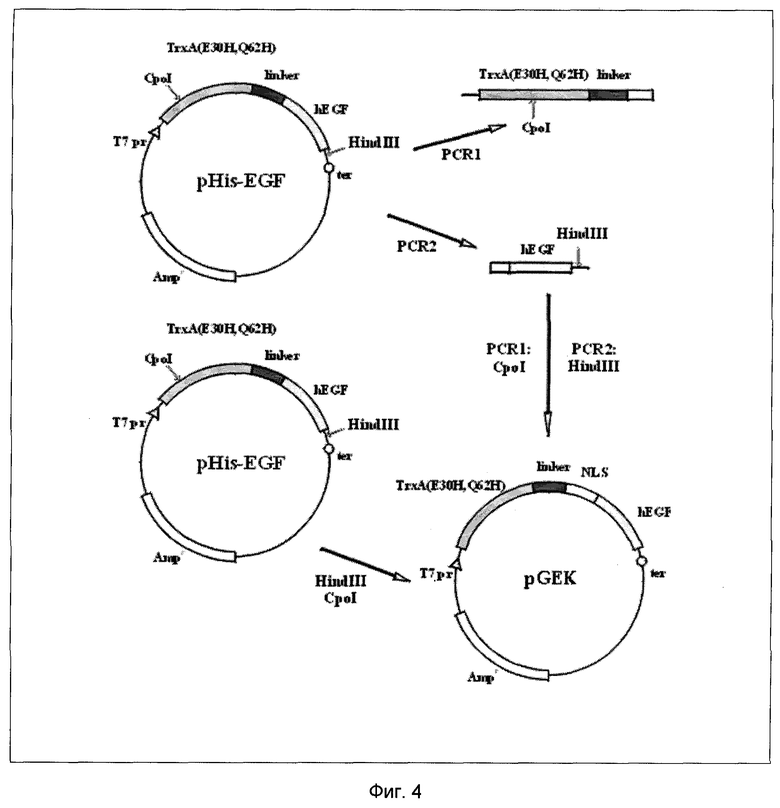
На Фиг.4 представлена схема конструирования плазмиды pGEK.
На Фиг.5 представлена схема включения в последовательность гена гибридного белка формулы SEQ ID №4 гена участка связывания с транспортируемым экзогенным генетическим материалом формулы SEQ ID № 2.
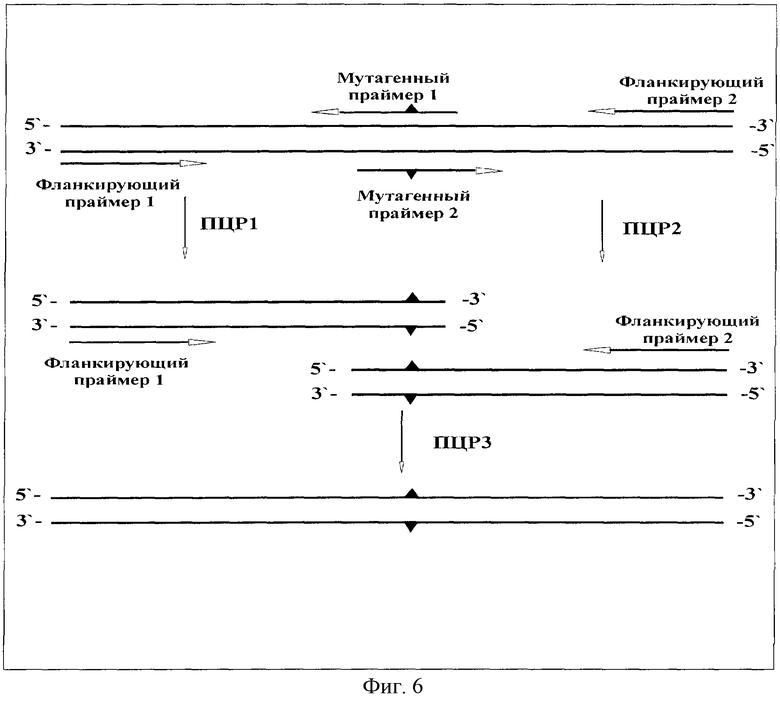
На Фиг.6 представлена схема введения точечных мутаций при помощи полимеразной цепной реакции.
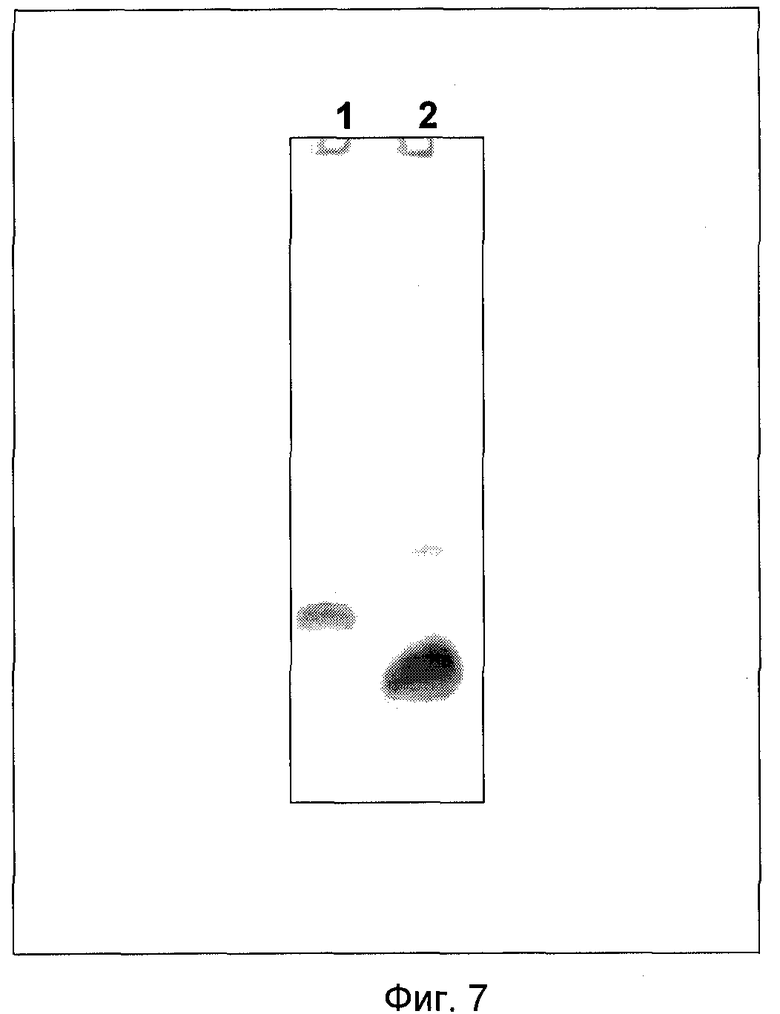
На Фиг.7 представлены результаты электрофоретического фракционирования в 15% Трицин-ПААГ очищенного нового пептидного вектора PGE-k формулы SEQ ID №1 в сравнении с рекомбинантным пептидом, человеческим эпидермальным фактором роста, где
1 - очищенный новый пептидный вектор формулы SEQ ID №1,
2 - контроль: рекомбинантный пептид, человеческий эпидермальный фактор роста.
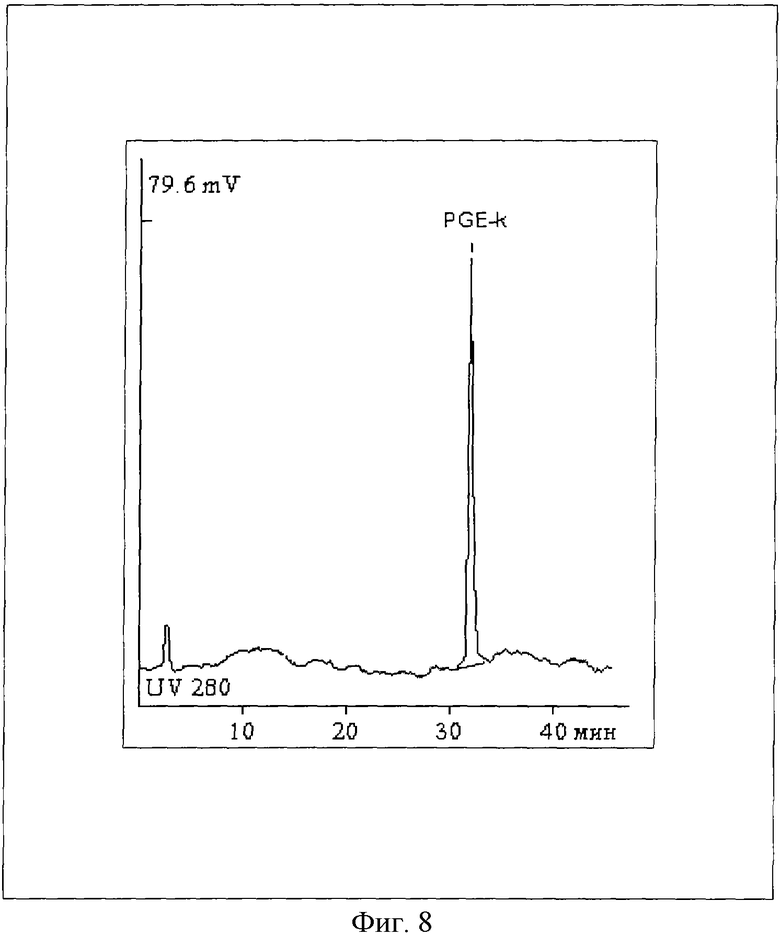
На Фиг.8 представлен ВЭЖХ-анализ очищенного нового пептидного вектора формулы SEQ ID № 1. Колонка Диасорб С16Т, 4х250 мм (Элсика, Россия), линейный градиент MeCN (от 5% до 95%, 45 мин) в 0,01% водной CF3COOH.
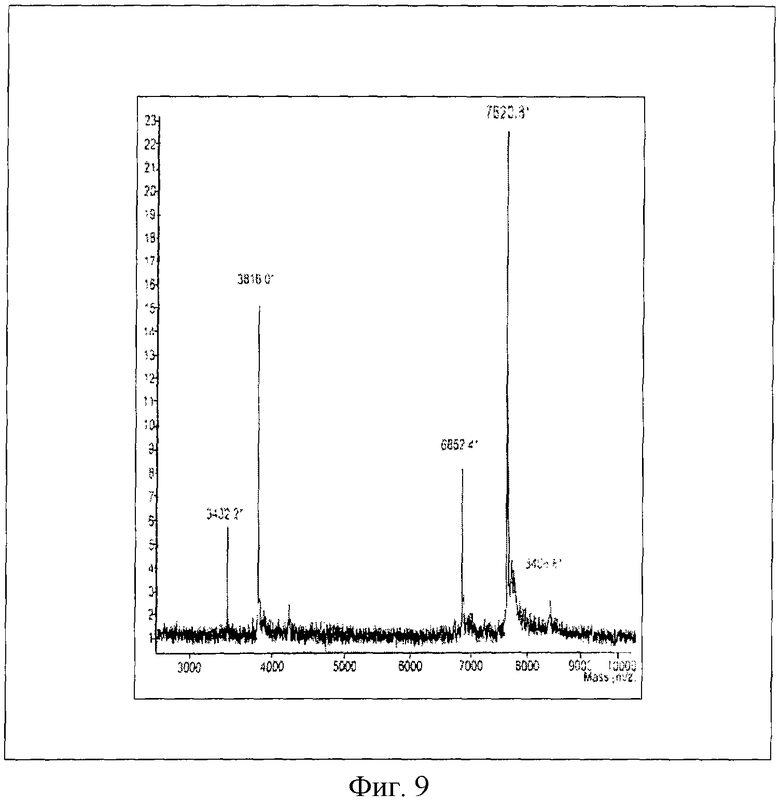
На Фиг.9 представлен масс-спектр (MALDI) нового пептидного вектора формулы SEQ ID № 1.
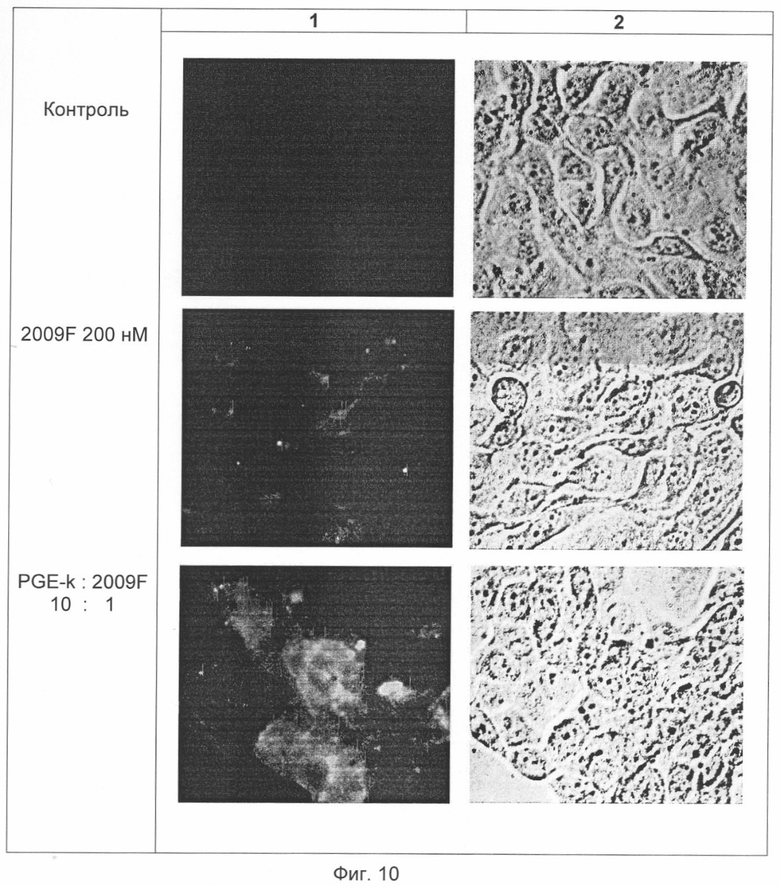
На Фиг.10 представлены микрофотографии (1 - флюоресцентные, 2 - инверсионные) опухолевых клеток линии А-431 через 18 часов инкубации с 5'-FITC-меченным фосфотиоатным олигонуклеотидом 2009F и со смесью его и нового пептидного вектора формулы SEQ ID №1 в молярном соотношении 1:10.
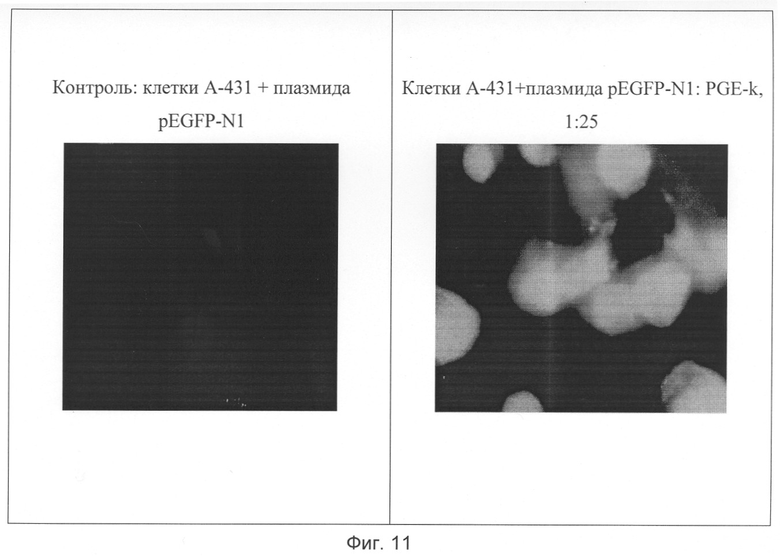
На Фиг.11 представлены флюоресцентные микрофотографии опухолевых клеток А-431 на пятый день инкубации с плазмидой pEGFP-N1 и со смесью ее и нового пептидного вектора формулы SEQ ID №1 в молярном соотношении 1:25.
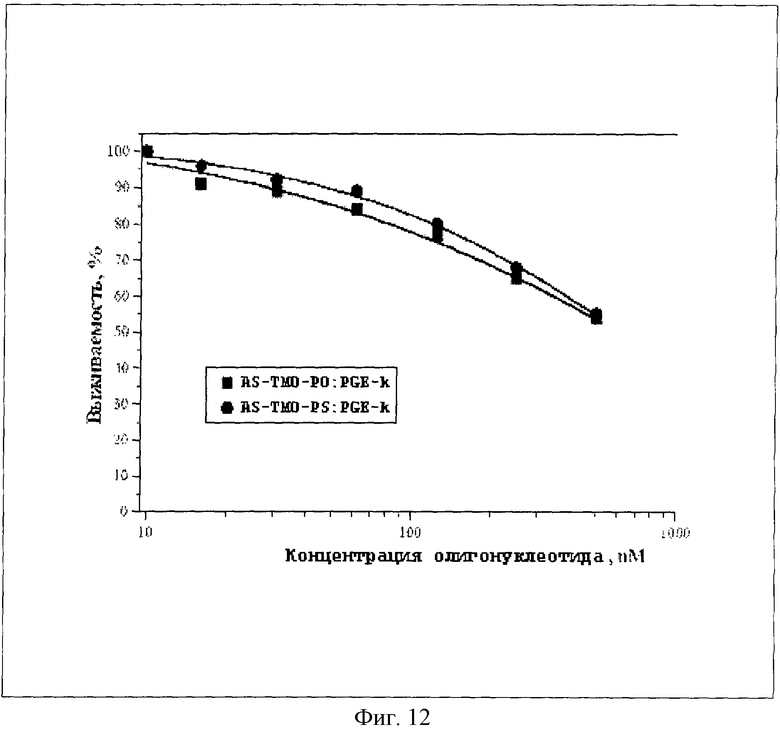
На Фиг.12 показано влияние смесей нового пептидного вектора формулы SEQ ID № 1 с олигонуклеотидами AS-TMO-PO и AS-TMO-PS на выживаемость клеток карциномы молочной железы человека линии MCF-7 (молярное соотношение белок-олигонуклеотид 5:1).
В Прил. 1. приведен список заявляемых олигодезоксирибонуклеотидных и аминокислотных последовательностей.
В Табл. 1 приведен список синтезированных олигонуклеотидов.
В Табл. 2 показано влияние вектора PGE-k на доставку флюоресцеин-меченного антисенс-тиоолигонуклеотида в опухолевые клетки.
В Табл. 3. показано влияние вектора PGE-k на доставку репортерной плазмиды в различные опухолевые клетки.
В Табл. 4. показано изменение цитостатической активности антисенса AS-ТМО и его тио-аналога в отношении культуры опухолевых клеток под действием липофектина и вектора PGE-k.
Ниже приведены примеры осуществления изобретения.
Пример 1 описывает химический синтез праймеров и антисенс-олигонуклеотидов, в том числе модифицированных, а также метод ферментативного 5'-фосфорилирования фрагментов ДНК.
Примеры 2-5 описывают клонирование гена тиоредоксина и получение плазмиды pBluescript II/Thio, содержащей этот ген. Источником хромосомной ДНК служил штамм Escherichia coli TG1.
Клонирование проводили при помощи метода ПЦР на основании известной нуклеотидной последовательности. Для амплификации гена использовали пару праймеров Thio5 и Тhiо3 (примечание: в работе использовались синтетические олигодезоксирибонуклеотиды, далее именуемые "олигонуклеотиды"; см. Табл. 1), фланкирующих кодирующую часть гена тиоредоксина с 5'- и 3’’-концов соответственно. Нуклеотидная последовательность праймеров включала рестрикционные сайты, необходимые для последующего клонирования в экспрессионном векторе. Thio5 содержал сайт рестрикции Ndel, а Тhio3 - сайт Nael.
Фрагменты ДНК, полученные в ходе ПЦР, фракционировали в агарозном геле, выделяли из агарозы, фосфорилировали с помощью полинуклеотид-киназы фага Т4, субклонировали в состав вектора pBluescript II SK (-) и получили плазмиду pBluescript II/Thio.
В этом и остальных примерах выделенные из клонов плазмиды секвенировались в пределах вставки по методу Сенгера.
Пример 6 описывает, согласно Фиг.3, получение плазмиды pGEMEX-1/hEGF, содержащей ген эпидермального фактора роста субклонированием этого гена в коммерческую плазмиду, pGEMEX-1, по сайтам рестрикции EcoRI и BamHI.
Пример 7 описывает, согласно Фиг.3, конструирование плазмиды pGEMEX-1 /Thio, содержащей ген тиоредоксина, переклонированием гена тиоредоксина из плазмиды pBluescript II/Thio в плазмиду pGEMEX-1 по сайтам рестрикции Ndel и BamHI.
Пример 8 описывает, согласно Фиг.3, получение и клонирование плазмиды pGEMEX-1/Thio-hEGF, содержащей ген слитого белка формулы SEQ ID № 4. Введение линкера было необходимо для направленного гидролиза энтерокиназой слитого белка формулы SEQ ID № 4 по сайту (Asp)4-Lys-Lys.
Пример 9 описывает введение точечных мутаций, Glu30His и Gln62His, в ген тиоредоксина.
Известно, что замена двух остатков в тиоредоксине Е. coli (Giu30→His и Gln62→His) позволяет выделять его и его производные металлохелатной аффинной хроматографией за счет связывания остатков гистидина с иммобилизованными ионами Ni2+ или Cu2+. Введение каждой из мутации включало в себя три ПЦР по общей схеме (Фиг.6). В результате получена плазмида pHis-EGF, содержащая ген указанного слитого белка.
Пример 10 описывает введение в состав плазмиды pHis-EGF фрагмента, кодирующего участок связывания генетического материала формулы SEQ ID №2 согласно Фиг.4, 5. Фрагмент вводился при помощи двух ПЦР с участием олигонуклеотидов С LINKER и N-EGF. В результате получена рекомбинантная плазмида pGEK.
Примеры 11, 12 описывают получение и выращивание штамма Escherichia coli В-8389 ВКПМ, а также выделение слитого белка формулы SEQ ID №4 с использованием металло-хелатной и ионообменной хроматографии. Фракции анализировались гель-электрофорезом.
Пример 13 описывает выделение и очистку пептидного вектора формулы SEQ ID №1. Слитый белок формулы SEQ ID №4 подвергали расщеплению каталитической субъединицей энтерокиназы по сайту (Asp)4-Lys-Lys. Полученный пептидный вектор PGE-k формулы SEQ ID №1 очищали ионообменной хроматографией. Продукт охарактеризован УФ- и масс-спектрами (Фиг.9), электрофорезом в ПААГ и ВЭЖХ (Фиг.7, 8). Аминокислотным анализом подтверждена последовательность десяти N-концевых аминокислот (NH2-Lys-Lys-Lys-Lys-Arg-Lys-Val-Glu-Asp-Pro-).
В Примерах 14-16 приведено описание опытов по определению влияния пептидного вектора PGE-k на доставку в культуры опухолевых клеток плазмидной ДНК (Пример 15) и олигонуклеотидов-антисенсов (Примеры 14 и 16).
В экспериментах использовались культуры клеток, характеризующиеся различным количеством поверхностных ЭФР-рецепторов: HeLa (клетки карциномы шейки матки человека), MCF-7 (клетки аденокарциномы молочной железы человека), А431 (клетки эпидермоидной карциномы вульвы человека), KB (клетки человеческой карциномы ротовой полости), К562 (клетки миелобластной лейкемии человека).
В качестве отрицательного контроля использовались клетки К562, поверхность которых практически не несет ЭФР-рецепторы, как и поверхность соматических клеток в норме. Как положительный контроль рассматривали результаты доставки тех же генетических конструкций в клетки липофекцией.
Универсальность пептидного вектора формулы SEQ ID №1 по отношению к структуре доставляемых молекул ДНК подтверждали в экспериментах с различными олигонуклеотидами, в том числе с модифицированными, и с плазмидной ДНК.
Примеры 14 и 16 описывают опыты по эффективной доставке в клетки-мишени олигомеров F2009 (антисенс к гену Вс12 ингибитора апоптоза), AS-TMO-РО (антисенса к РНК-компоненту теломеразы) и его фосфоротиоатного аналога AS-TMO-PS, содержащего вместо связей O-Р(O)-O- связи O-P(S)-O-.
Аналогичные данные были получены и для олигонуклеотидов другой структуры, например антисенсов к генам С-мус, mdr1 и др.
В Примере 14 приведены экспериментальные данные по определению влияния пептидного вектора формулы SEQ ID №1 на эффективность доставки в опухолевые клетки А431 и К562 фосфоротиоатного антисенс-олигонуклеотида 2009F, несущего 5'-флюоресцеиновую метку (FITC) (см. Фиг.10 и Табл. 2). Свойства фосфоротиоатных олигонуклеотидов, в частности высокая внутриклеточная стабильность, позволяет надежно соотнести регистрируемую флюоресценцию с внутриклеточным расположением именно олигомера, а не продуктов его гидролиза. Из данных Табл. 2 следует, что использование пептидного вектора PGE-k приводило к резкому, более чем в 5 раз, увеличению эффективности доставки 2009F в клетки А431, обогащенные ЭФР-рецепторами. В то же время, данный пептидный вектор не только не способствовал, а даже препятствовал попаданию 2009F в нецелевые рецептородефицитные клетки К562, защищая их от действия 2009F, что подтверждает селективность PGE-k.
Пример 15 описывает эксперимент по доставке в клетки HeLa и А431 плазмиды pEGFP-N1(Clontech, США), содержащей под ядерным промотором ген зеленого флюоресцирующего белка (см. Фиг.11, Табл. 3). Экспрессия генов в составе такой конструкции надежно подтверждает попадание плазмиды в клеточное ядро.
Доставка препарата плазмидной ДНК pEGFP-N1 в клетки культур HeLa, А431 и К562 липофекцией во всех случаях приводила к трансформации более 90% обработанных клеток, т.е. такое количество клеток экспрессировало репортерный белок. Инкубирование клеток этих культур со свободной плазмидой не приводило к образованию трансформантов. Выращивание клеточных культур в тех же условиях, но с добавлением смесей плазмиды pEGFP-N1 с пептидным вектором формулы SEQ ID №1, приводила к флюоресценции только тех клеток, которые несли поверхностные ЭФР-рецепторы (HeLa и А431). Данные Табл. 3 показывают, что количество трансформантов зависело от природы культуры клеток и молярного соотношения компонентов смеси и было максимальным для обогащенных ЭФР-рецепторами клеток культуры А431 и нулевым для рецептородефицитных К562.
В целом результаты надежно свидетельствуют об избирательной доставке плазмидной ДНК в ядра клеток, несущих наружные рецепторы к эпидермальному фактору роста.
Пример 16 описывает определение влияния пептидного вектора формулы SEQ ID №1 на цитостатическую активность олигонуклеотидов AS-TMO-PO и AS-TMO-PS в отношении культуры опухолевых клеток линии MCF-7. В рассмотренных пределах концентраций (до 1000 нМ) олигонуклеотиды не проявляли цитостатической активности. Принципиальное различие во влиянии на выживаемость клеток наблюдалось при доставке олигомеров внутрь клеток липофекцией. Фосфоротиоатный антисенс AS-TMO-PS обладал высокой токсичностью (IC50=109 нМ), а фосфодиэфирный AS-TMO-PO был нетоксичен.
Инкубирование клеток со смесями этих олигомеров с белком PGE-k в молярном соотношении олигонуклеотид - PGE-k 1:5 приводило к одинаковым значениям IC50, равным для обоих антисенсов 470 нМ (см. Фиг.12 и Табл. 4).
Приведенные данные иллюстрируют способность нового пептидного вектора не только доставлять олигонуклеотиды в клетки-мишени, но и защищать их от внутриклеточной деградации. Пептидный вектор формулы SEQ ID №1 позволяет с одинаковой эффективностью использовать и фосфоротиоатные олигонуклеотиды-антисенсы, и их природные аналоги.
Из данных Примеров 14-16 следует, что пептидный вектор PGE-k уступает в активности известному трансфицирующему средству, липофектину, примерно в 4 раза. Однако липофектин действует неселективно, независимо от типа клеток, и способен сам по себе проявлять неизбирательное цитотоксическое действие, нарушая барьерные свойства наружной клеточной мембраны. Поэтому новый пептидный вектор формулы SEQ ID № 1 имеет неоспоримое преимущество перед липофектином вследствие специфичности своего действия. Кроме того, применение нового пептидного вектора формулы SEQ ID № 1 впервые дает возможность использовать в практике генотерапии фосфодиэфирные антисенс-олигонуклеотиды вместо токсичных и существенно более дорогих тио-аналогов.
Пример 1. Синтез и фосфорилирование праймеров.
Олигодезоксирибонуклеотиды, в том числе праймеры (см. Табл. 1), были синтезированы твердофазным амидофосфитным методом на автоматическом синтезаторе ДНК АСМ 2У ("Биоссет", Россия) с использованием коммерческих защищенных амидофосфитных производных нуклеозидов и флюоресцеина (FITC), нуклеозид-полимеров и других реагентов (Glen Research, США). В синтезе фосфоротиоатного олигонуклеотида, AS-TMO-PS, вместо стандартного окислителя, содержащего йод и воду, использовали 0,05 М раствор 3Н-1,2-бензодитиол-3-он-1,1-диоксида (Glen Research, США) в ацетонитриле. Деблокирование и расщепление связи синтезированного олигомера с носителем осуществляли аммонолизом (28% аммиак, 5 часов, 50°С). 5'-Диметокситритил-производные всех олигонуклеотидов были очищены ион-парной обращенно-фазовой ВЭЖХ на колонке Диасорб С16Т, 4×250 мм (Элсика, Россия) в линейном градиенте MeCN (от 12,5% до 45%, 30 мин) в 0,1М водном NEt3-AcOH при 50°С, деблокированы 80% водной АсОН и выделены в виде литиевых солей. Синтезированные олигонуклеотиды характеризовали УФ-спектрофотометрией, масс-спектроскопией лазерной десорбции, анализировали методами ВЭЖХ и электрофореза в ПААГ.
5'-Фосфорилирование олигонуклеотидов проводили с помощью полинуклеотидкиназы фага Т4 в стандартных условиях. В 10 мкл реакционной смеси содержалось 20-40 пмолей олигонуклеотида, 1 мкл 10×PNK-буфера ("NEB", США): 500 мМ Tris-HCI, 100 мМ МgСl2, 50 мМ дитиотреитола, 1 мМ спермидина, 1 мМ EDTA, 2 мкл 5 мМ rАТР и 10 единиц полинуклеотидкиназы. Реакционную смесь выдерживали в течение 1 ч при 37°С, затем инактивировали фермент 30 мин при 70°С.
Пример 2. Выделение плазмидной ДНК из клеток штаммов Е. соli.
Выделение плазмидной ДНК из всех штаммов Е. соli проводили по модифицированному методу Birinboim & Doly. Клетки из 15 мл ночной культуры осаждали центрифугированием и осадок ресуспендировали в 1 мл буфера (25 мМ Tris-HCI, рН 8,0, глюкоза до 50 мМ, ЭДТА до 10 мМ, лизоцим до 4 мг/мл). Клеточную суспензию инкубировали 30 мин при комнатной температуре и добавляли 2 мл 0,2 М NaOH, содержащего 1% SDS. Смесь выдерживали в ледяной бане в течение 5 мин. Затем при 0°С добавляли 1,5 мл 3 М КОАс, рН 4,8, выдерживали 30 мин, осадок отделяли центрифугированием (30 мин, 12000 g). К надосадочной жидкости добавляли 1/3 объема 40% PEG 6000, выдерживали при 4°С 30 мин. Смесь центрифугировали (10 мин, 12000 g), осадок растворяли в 2 мл буферного раствора (20 мМ Tris-HCl, 1 мМ ЭДТА, рН 8,0), добавляли 4 мл насыщенного раствора NH4OAc, выдерживали 15 мин при 4°С. Осадок отфуговывали (15 мин, 12000 g), к супернатанту добавляли 4 мл изопропанола, выдерживали 15 мин при 4°С и повторяли центрифугирование. Осадок промывали 70% этанолом и сушили в вакууме.
Пример 3. Выделение хромосомной ДНК из клеток штаммов Е. соli.
1.5 мл ночной культуры клеток Escherichia coli TG1 центрифугировали (12000 g, 3 мин). Клетки ресуспендировали в 600 мкл буфера (20 мМ Tris-HCl, рН 8.0, 1 мМ ЭДТА, 0.5% ДНС, 100 мкг/мл протеиназы К), выдерживали 1 час при 37°С. К суспензии добавяли 100 мкл 5 М NaCl, перемешивали и затем добавляли 80 мкл 0.7 М NaCl, содержащего 10% цетилтриметиламмонийбромида. Суспензию перемешивали и выдерживали 1 час при 65°С. Последовательно экстрагировали равными объемами хлороформа и смеси фенол/хлороформ, уравновешенной в 0.2 М Tris. Водную фазу смешивали с 0.6 объема iPrOH. ДНК осаждали центрифугированием (10 с, 12000g), промывали 70% ЕtOН и высушивали в эксикаторе.
Пример 4. Условия проведения полимеразных цепных реакций (ПЦР).
4.1. Амплификации гена, кодирующего тиоредоксин.
Реакционная смесь содержала 10 нг хромосомной ДНК штамма Е. coli TG1 (см. Пример 3), 0.2 мМ каждого из dNTP, 1 мкМ каждого из праймеров, Thio5 и Thio3 и 1 единицу активности термостабильной полимеразы Vent в 50 мкл буфера (20 мМ Tris-HCl, рН 8.8, 2 мМ MgS04, 10 мМ КСl, 10 мМ (NH4)2SO4, 0.1% Тритон X 100). Сверху на реакционную смесь наслаивали 25 мкл минерального масла ("USB", США). Цикл амплификации включал в себя денатурацию ДНК (96°С, 1 мин), отжиг (60°С, 1 мин) и элонгацию (74°С, 30 с). Всего проводили 35 циклов реакции на амплификаторе фирмы "Techne" (Германия). Продукты ПЦР электрофоретически разделяли в 1.7% агарозном при напряженности электрического поля 12 V/см и выделяли фрагмент длиной 333 пары оснований.
4.2. Остальные ПЦР.
Реакционные смеси содержали 100 нг плазмидной ДНК, 0,2 мМ каждого из dNTP, 1 мкМ каждого из праймеров и 1 единицу активности термостабильной полимеразы Vent в 50 мкл буфера (20 мМ Tris-HCl, рН 8.8, 2 мМ MgSO4, 10 мМ KCl, 10 мМ (NH4)2SO4, 0.1% Тритон X 100).
Пример 5. Клонирование гена тиоредоксина и получение плазмиды pBluescript ll/Thio.
Плазмиду pBluescript II SK- ("Stratogene", США), гидролизовали эндонуклеазой Smal. Гидролизат обрабатывали равным объемом фенол/хлороформ (1:1), к водной фазе добавляли 1/10 объема 3 М раствора AcONa, рН 6.0, и 3 объема этанола, выдерживали 30 мин при 4°С, центрифугировали при 12000 g 15 мин. Осадок промывали 70% этанолом, высушивали, растворяли в 40 мкл 10 мМ Tris-HCI, рН 8.0. К раствору добавляли 5 мкл 10х CIAP буфера (0.5 М Tris-HCI, 10 мМ MgCl2, 1 мМ ZnCl2, 10 мМ спермидина, рН 9.3), 5 мкл фосфатазы (0.1 ед./мкл) и инкубировали при 37°С в течение 30 мин. К реакционной смеси добавили 300 мкл стоп-буфера (10 мМ Tris-HCI, рН 7.5, 1 мМ ЕДТА, 200 мМ NaCl, 0.5% ДСН), затем обрабатывали раствор, осаждали и высушивали осадок, как описано выше для гидролизата в данном Примере. 50 нг полученного продукта смешивали с 5 нг продукта ПЦР длиной 333 пары оснований, полученного в Примере 4.1, и 5 единицами Т4 ДНК-лигазы в 10 мкл 1х буфера (40 мМ Tris-HCI, 10 мМ MgCl2, 10 мМ дитиотреитол, 0,5 мМ АТР, рН 7.8). После инкубации лигазной смеси (16°С в течение ночи) ею протрансформировали клетки штамма Е. соli XL-1 Blue (Stratogene, США) из расчета 10 нг плазмидной ДНК на 1,5×107 клеток (в 100 мкл) и высевали на чашки Петри с агаризированной средой, содержащей 100 мкг/мл ампициллина, инкубировали в течение ночи при 37°С. Инокулировали отдельными колониями по 3 мл среды YT (8 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl), содержащей 100 мкг/мл ампициллина, и инкубировали в течение ночи при 37°С и 260 об./мин на качалке "Certomat Н" ("В.Braun Melsungen", Германия). Выделенную из трансформированных клеток (как в Примере 2) плазмидную ДНК секвенировали в пределах вставки по методу Сенгера, используя для этого Т7 и обратный праймеры. Сконструированная плазмида получила название pBluescript ll/Thio.
Пример 6. Субклонирование гена hEGF в векторе pGEMEX-1.
Плазмиду pGEMEX-1 последовательно гидролизовали рестриктазами EcoRI и ВатHl, обрабатывая гидролизат, как в Примере 5. Полученный продукт дефосфорилировали и обрабатывали, как в Примере 5. 50 нг полученного продукта смешивали с 5 нг гена hEGF [7] и лигировали в условиях Примера 5. Лигазной смесью протрансформировали клетки штамма Е. соli XL-1 Blue в условиях Примера 5. Выращивание клеток, выделение полученной плазмиды pGEMEX-1/hEGF и подтверждение ее структуры также проводили по методике Примера 5.
Пример 7. Получением плазмиды pGEMEX-1/Thio.
Плазмиду pGEMEX-1 последовательно гидролизовали рестриктазами Aatll и BamHI, обрабатывая гидролизат, как в Примере 5 и выделяя фрагмент Aatll/ BamHI (pGEMEX-1).
Аналогично из той же плазмиды последовательным действием рестриктаз NdeI и AatII был получен и выделен фрагмент NdeI/AatII (pGEMEX-1).
Из плазмиды pBluescript II/Thio (Пример 5) действием рестриктаз NdeI и BamHI аналогично был получен фрагмент NdeII/BamHI (pBluescript II/Thio).
Дефосфорилирование полученных фрагментов и их совместное лигирование с образованием плазмиды pGEMEX-1/Thio проводили, как в Примере 5.
Пример 8. Получение плазмиды pGEMEX-1/Thio-hEGF
Плазмиду pGEMEX-1/hEGF (Пример 7) расщепляли рестриктазами AatII и EcoRI, а плазмиду pGEMEX-1/Thio - рестриктазами AatII и TurboNael. Фрагменты AatIIEcoRI (pGEMEX-1/hEGF) и AatII/TurboNael (pGEMEX-1/Thio) объединяли с линкером, дуплексом олигонуклеотидов SpE1 и SрЕ2, и лигировали, как в Примере 5. В условиях Примера 5 реакционной смесью трансформировали штамм Е. coli XL1 Blue и выделяли плазмиднуюДНК из трансформантов. Полученные плазмиды анализировали путем гидролиза рестриктазами TurboNael и EcoRI, затем ДНК отобранных клонов секвенировали методом Сэнгера. Результирующий вектор получил название pGEMEX-1/Thio-hEGF.
Пример 9. Получение плазмиды pHis-hEGF.
9.1.
В условиях Примера 4.2 (температура отжига праймеров 36°С) провели 20 циклов ПЦР на плазмиде pGEMEX-1/Thio-hEGF (Пример 8) с праймерами E30R и SP6. Продукты ПЦР электрофоретически разделяли, как в Примере 4.1, и выделили фрагмент длиной 487 пары оснований.
9.2.
ПЦР на плазмиде pGEMEX-1/Thio-hEGF с праймерами E30L и Т7 и разделение продуктов ПЦР проводили в условиях Примера 9.1. Выделили фрагмент длиной 178 пар оснований.
ПЦР 9.3.
Реакционная смесь содержала 40 нг продукта ПЦР 9.2 и 20 нг продукта ПЦР 9.1. Отжиг праймеров проводили при 28°С в течение 1 мин. Всего проводили 30 циклов ПЦР. Остальные условия проведения ПЦР - как в Примере 9.1. Обработку реакционной смеси и манипуляции с продуктом ПЦР проводили, как в Примере 5.
9.4. Введение мутаций Glu30His и Gln62His в ген тиоредоксина Е. соli.
Согласно схеме (Фиг.6) последовательно провели 2 мутации. В условиях Примера 9.1 проводили ПЦР на ДНК из Примера 9.3 с мутагенными праймерами Q62R и Q62L и фланкирующими праймерами Т7 и SP6. Продукт мутации, обработанный и выделенный, как в Примере 9.1, подвергли мутации Glu30His с другими мутагенными праймерами, E30L и E30R в условиях ПЦР и выделения продукта мутации Примера 9.1. Продукт последней ПЦР гидролизовали рестриктазами Ndel и HindIII, выделяли полученный фрагмент ДНК длиной 557 пар оснований и клонировали его в вектор pGEMEX-1, как описано в Примере 5. Полученная плазмида получила название pHis-hEGF.
Пример 10. Получение рекомбинантной плазмиды pGEK.
10.1.
Проводили 24 цикла ПЦР со смесью плазмиды pHis-hEGF (Пример 9.4), праймера Т7 и фосфорилированного C-LINKER'a в условиях Примера 4.2. Температурный режим: денатурация ДНК-1 мин при 96°С, отжиг праймера -1 мин при 49°С, синтез ДНК -1 мин при 74°С.
10.2.
При тех же условиях провели ПЦР со смесью плазмиды pHis-hEGF, праймера SP6 и фосфорилированного N-EGF.
10.3.
Продукты ПЦР 10.1 и 10.2 длиной 460 и 238 выделяли электрофорезом, как в Примере 4.1, и гидролизовали соответственно эндонуклеазами Сроl и Hindlll. Продукты гидролиза длиной 278 и 210 п.о. соответственно вновь выделяли электрофорезом. Плазмиду pHis-hEGF последовательно гидролизовали эндонуклеазами CpoI и HindIII и выделяли CpoI/HindIII (pHis-hEGF), как в Примере 5. CpoI/HindIII (pHis-hEGF) дефосфорилировали в условиях Примера 5. Дефосфорилированный CpoI/HindIII (pHis-hEGF) смешивали с гидролизованными продуктами ПЦР 10.1 и 10.2, лигировали, выделяли продукт лигирования, клонировали его в Е. соli XL-1 Blue, выращивали трансформанты и выделяли из них плазмидную ДНК, как в Примере 5. Отобранные клоны секвенировались в пределах вставки по методу Сенгера. Полученная экспрессионная конструкция получила название “рекомбинантной плазмиды pGEK”.
Пример 11. Трансформация штамма Escherichia coli BL21 (DE3)pLysS и выращивание нового штамма Escherichia соli В-8389 ВКПМ.
Клетки штамма Е. соli BL21(DE3)pLysS (фирма "Novagen, Inc.", Cat#69388-3, Мэдисон, штат Нью Джерси, 53711, США) трансформировали плазмидой pGEK из расчета 10 нг pGEK на 1 - 5×107 клеток по методике Примера 5 с получением целевого нового штамма Escherichia coli BL2l(DE3)/pLysS/pGEK (№В-8389 ВКПМ). Затем культуру высевали на чашки Петри с агаризированной средой, содержащей 100 мкг/мл ампициллина и 34 мкг/мл хлорамфеникола, и инкубировали в течение ночи при 37°С. Отдельные колонии (около 100 шт.) собирали стеклянной палочкой и суспендировали в 100 мл среды ТВ (12 г/л триптона, 24 г/л дрожжевого экстракта, 0,04% глицерина, 1/10 объема фосфатного буфера (0,17 М КН2РO4, 0,72 М К2НРO4)). Культуру выращивали в присутствии 100 мкг/мл ампициллина и 34 мг/мл хлорамфеникола при 30°С и 260 об./мин. на качалке "Certomat Н" ("В.Braun Melsungen", Германия) до достижения культурой оптической плотности OD550=2.0 о.е. Далее добавили изопропил-β-D-тиогалактозид до конечной концентрации 0.05 мМ и продолжили инкубацию при 13°С и 260 об./мин 24 часа.
Новый штамм содержит 300-400 копий рекомбинантной плазмиды pGEK, это количество поддерживается стабильно в течение, по крайней мере, 8 генераций.
Штамм Е. соli BL2l(DE3)/pLysS/pGEK синтезирует слитый белок TrxA-EGF-NLS, формулы SEQ ID №4, в количестве 300 мг на 1 л культуральной жидкости при ее плотности, соответствующей 4×109 кл/мл.
Устойчивость: F-, ompT, hsdSB (rB-, mB-), dcm, gal, λ (DE3), plysED, CmR AmpR. Концентрация ампициллина - 100 мкг/мл, хлорамфеникола - 25 мкг/мл.
Штамм не является зоо- и фитопатогенным.
Пример 12. Выделение слитого белка TrxA-EGF-NLS формулы SEQ ID №4.
Полученную культуру клеток Escherichia coli B-8389 ВКПМ центрифугировали (2500 g, 1 час). Клетки ресуспендировали в 50 мл буфера: 100 мМ Tris-HCl+фенилметилсульфонилфторид до 0.1 мМ, рН 8.0 и обработали ультразвуком на дезинтеграторе УЗДН-2Т (7-10 раз по 30 с при t° не выше 10°С). Добавили равный объем 0.3 М NaCl и перемешали, центрифугировали при 9000 g 1 ч. Надосадочную жидкость нанесли на колонку (25×100 мм, Chelatlng-Sepharose (Ni2+) ("Pharmacia", Швеция)). Колонку отмыли буфером: 50 мМ Tris-HCl, рН 8.0, 0.15 М NaCl, затем буфером: 50 мМ Tris-HCl, рН 8.0,1 М NaCl, и вновь предыдущим буфером, каждый раз добиваясь отсутствия поглощения элюата при λ=280 нм. Полученный целевой продукт - слитый белок формулы SEQ ID № 4 - вымывали буфером: 50 мМ Tris-HCI, рН 8.0, 0.25 М NaCI, 0.1 М имидазол. Элюат диализовали (4°С, мембрана Spectra/Por 10000): дважды против 50 мМ Tris-HCl, рН 8.0, 8-12 ч, 1 раз против буфера: 50 мМ Tris-HCl, 1 мМ CuSO4, рН 8.0, 24-36 ч и дважды против 20 мМ Tris-HCl, рН 7.6, 8-12 ч. Диализат нанесли на колонку MonoQ HR10/10 ("Pharmacia", Швеция), уравновешенную последним буфером. Слитый белок формулы SEQ ID №4 вымывали линейным градиентом NaCI (0.1 М-0.5 М) в этом же буфере. Фракции анализировали гель-электрофорезом и объединили те из них, в которых содержалось наибольшее количество целевого слитого белка (молекулярная масса - 20 kDa).
Пример 13. Выделение и очистка пептидного вектора PGE-k формулы SEQ ID № 1.
К полученному препарату слитого белка формулы SEQ ID №4, обессоленному при помощи устройства Ultrafree-15 (Biomax-5K) (Millipore), добавили 1/10 объема энтерокиназного буфера: 0,5 М Tris-HCI, 0,01 М CaCl2,1% Твин 20, рН 8.0, и энтерокиназу EKmax ("Invitrogen", США) из расчета 1 ед. фермента на 3 мг гибридного белка, инкубировали 20 часов (37°С). Продукты гидролиза фракционировали на колонке MonoQ HR10/10 и анализировали (как описано в Примере 10). Выход чистого пептидного вектора PGE-k формулы SEQ ID № 1 составлял 1,8-2,2 мг. m/e [М.в.-Н] 7620,8 (рис. 9).
Пример 14. Определение влияния пептидного вектора PGE-k на доставку в культуры опухолевых клеток 5'-флюоресцеин-меченного тио-олигонуклеотида 2009F.
Опухолевые клетки HeLa, MCF-7, A431 и KB культивировали в пластиковых культуральных флаконах (Costar, Великобритания) в среде DMEM (Gibco, США), содержащей 10% эмбриональной бычьей сыворотки (Gibco) и 50 μg/ml гентамицина (Sigma, США) при 37°С в увлажненной атмосфере, содержащей 5% СO2. Клетки линии К562 культивировали в тех же условиях, но с использованием среды RPMI 1640 (Gibco, США). Клеточные культуры рассевали 2 раза в неделю и высевали в 12-луночные планшеты на покровные стекла или непосредственно в лунки за сутки до эксперимента.
Трансфекцию клеток A431 и К562 осуществляли растворами тио-5'-FIТС-олигонуклеотида 2009 F в присутствии различных PGE-k. Для приготовления трансфицирующих смесей в соотношениях компонентов, приведенных в Табл. 2, использовали раствор PGE-k (40 мкМ, в 50 мМ Tris-HCl, 0.17М NaCl, pH 8) и 50 мМ раствор 2009 F в воде. Во всех экспериментах концентрация по олигонуклеотиду при инкубировании составляла 200 нМ. Через 18 часов культуральную среду удаляли, стекла с клетками дважды промывали фосфатно-солевым буфером (PBS), клетки фиксировали 4% параформальдегидом в PBS, заключали в мовиол и исследовали флуоресценцию с помощью флуоресцентного микроскопа. Для проточной цитофлуориметрии (цитофлуориметр EPICS С, Coulter, Германия) клетки после окончания инкубации снимали с пластика с помощью раствора 0.05% трипсина в растворе Версена (Sigma, США), промывали PBS и фиксировали 2% параформальдегидом. Для возбуждения флуоресценции использовали аргоновый лазер (Coherent, США) с длиной волны 488 нм. Для микрофотографии клеток здесь и далее использовали инвертированный микроскоп Nikon (Япония) и флуоресцентный микроскоп Opton (Германия) (Фиг.10). Результаты опытов приведены в Табл. 2. Они показывают, что добавление пептидного вектора PGE-k к флюоресцеин-меченному тио-антисенсу в несколько раз увеличивает интенсивность флуоресценции рецептор-обогащенных клеток и, следовательно, увеличивает количество доставленного в клетки тио-антисенса. У рецептор-дефицитных клеток К562, напротив, при добавлении PGE-k интенсивность флуоресценции снижается.
Пример 15. Определение влияния пептидного вектора PGE-k на доставку в культуры опухолевых клеток плазмидной ДНК.
Трансфекцию культур клеток HeLa, А431и К562, подготовленных, как описано в Примере 14, осуществляли растворами препарата плазмиды pEGFP-N1 (Clontech) в воде в присутствии различных количеств PGE-k (Табл. 3). Перед трансфекцией клетки промывали средой DMEM, не содержащей сыворотку, добавляли 2 мл свежей DMEM и инкубировали в течение 2 часов. За 15 мин до трансфекции среду меняли на свежую и добавляли хлорокин (Sigma) в концентрации 25 мкМ.
Для приготовления трансфекционных смесей с соотношением компонентов, приведенных в Табл. 3, использовали растворы плазмиды pEGFP-N1 (78 нМ, ~ 220 мкг/мл) в воде и раствор пептидного вектора PGE-k (40 мкМ, в 50 мМ Tris-HCl 0.17M NaCl, pH8). Смеси не теряли трансфицирующих свойств, по крайней мере, в течение недели. Растворы добавляли к клеткам из расчета 1 мкг плазмиды/мл культуры на 24 часа, после чего среду меняли на стандартную среду DMEM и инкубировали 24-96 час. Затем исследовали флуоресценцию трансфицированных клеток, как в Примере 14. Трансфекцию клеток HeLa, A431 и К562 плазмидой с помощью липофектина (InvitroGen, США) проводили в соответствии с протоколом изготовителя. Экспериментальные данные, приведенные в Табл. 3, показывают, что свободная плазмида pEGFP-N1 не трансфицирует опухолевые клетки. В присутствии пептидного вектора формулы SEQ ID №1 происходит трансфекция только рецептор-обогащенных клеток HeLa и A431.
Пример 16. Определение влияния пептидного вектора PGE-k на цитостатическую активность олигонуклеотидов AS-TMO-PO и AS-TMO-PS в отношении опухолевых клеток.
Клетки аденокарциномы молочной железы человека линии MCF-7, подготовленные, как описано в Примере 14, обрабатывали водными растворами: олигонуклеотида AS-TMO-PO, олигонуклеотида AS-TMO-PS (тио-аналога AS-TMO-PO) или смесями этих олигонуклеотидов с новым пептидным вектором PGE-k в молярных соотношениях 1:5 в диапазоне концентраций 1×10-9-1×10-6 М по олигонуклеотиду. Клетки инкубировали 120 часов в условиях Примера 14. Их выживаемость определяли с помощью МТТ-теста и оценивали в процентах от соответствующего холостого контроля. Трансфекцию олигонуклеотидами с помощью липофектина (InvitroGen, США) проводили в соответствии с протоколом изготовителя. Значения показателя цитостатической активности IC50 для указанных олигонуклеотидов и их смесей с пептидным вектором PGE-k в отношении опухолевых клеток MCF-7 приведены в Табл. 4. Результаты показывают, что AS-TMO-PO не проявляет такой активности ни в свободном виде, ни в присутствии липофектина. В присутствии пептидного вектора формулы SEQ ID №1 оба антисенса проявляют примерно одинаковую цитостатическую активность.
Литература
1. Cristano R.J., Roth J.A. EGF-targeted nucleic acid delivery// Пат. США № WO 96/30536, выдан 03.10.1996.
2. Fominaya J., Uherek С., Wels W. A chimeric fusion protein containing transforming growth factor-alpha mediates gene transfer via binding to the EGF receptor// Gene Ther. 1998. V.5. № 4. Р.521-30.
3. Collas P., Alestrom P. Nuclear localization signals: a driving force for nuclear transport of plasmid DNA in zebrafish// Biochem. Cell. Biol. 1997. V.75. P.633-640.
4. Siebenkotten G., Christine R. Cellular transport system for the transfer of a nucleic acid through the nuclear envelope and methods thereof// Пат. США № WO 00/40742. Выдан 18.02.2003.
5. Morris M.C., Vidal P., Chaloin L, Heitz F., Divita G. A new peptide vector for efficient delivery of oligonucleotides into mammalian cells// Nucleic Acids Res. 1997. V.25. № 14. Р.2730-2736.
6. Lu Zh., DiBlasio-Smith E.A., Grant K.L., Warne N.W., LaVallie E.R., Collins-Racie L.A., Follettie M.T., Williamson M.J., McCoy J.M. Histidine Patch Thioredoxins. Mutant forms of thioredoxin with metal chelating affinity that provide for convenient purifications of thioredoxin fusion proteins//J. Biol. Chem. 1996. V.271. № 9. P.5059-5065.
7. Эльдаров М.А., Кагиянц С.М., Позмогова Г.Е., Луценко С.В., Северин Е.С., Кирпичников М.П., Скрябин К.Г. Штамм дрожжей S. cerevisiae BKM CR-349D - продуцент эпидермального фактора роста человека// Патент РФ 1999. №21505001 выдан 10.06.2000.
Приложение 1. Перечень олигодезоксирибонуклеотидных и аминокислотных последовательностей.
SEQ ID № 1 - аминокислотная последовательность заявляемого пептидного вектор PGE-k:
NH2-Lys-Lys-Lys-Lys-Arg-Lys-Val-Glu-Asp-Pro-Tyr-Asn-Ser-Asp-Ser-Glu-Cys-Pro-Leu-Ser-His-Asp-Gly-Tyr-Cys-Leu-His-Asp-Gly-Val-Cys-Met-Tyr-lle-Glu-Ala-Leu-Asp-Lys-Tyr-Ala-Cys-Asn-Cys-Val-Val-Gly-Tyr-lle-Gly-Glu-Arg-Cys-Gln-Tyr-Arg-Asp-Leu-Lys-Trp-Trp-Glu-Leu-Arg-COOH
SEQ ID № 2 - аминокислотная последовательность остатка пептидного человеческого эпидермального фактора роста:
-Asn-Ser-Asp-Ser-Glu-Cys-Pro-Leu-Ser-His-Asp-Gly-Tyr-Cys-Leu-His-Asp-Gly-Val-Cys-Met-Tyr-lle-Glu-Ala-Leu-Asp-Lys-Tyr-Ala-Cys-Asn-Cys-Val-Val-Gly-Tyr-lle-Gly-Glu-Arg-Cys-Gln-Tyr-Arg-Asp-Leu-Lys-Trp-Trp-Glu-Leu-Arg-COOH
SEQ ID № 3 - аминокислотная последовательность участка связывания с транспортируемым экзогенным генетическим материалом:
NH2-Lys-Lys-Lys-Lys-Arg-Lys-Val-Glu-Asp-Pro-Tyr-
SEQ ID №4 - аминокислотная последовательность слитого белка:
NH2-X-Y-Lys-Lys-Lys-Lys-Arg-Lys-Val-Glu-Asp-Pro-Tyr-Asn-Ser-Asp-Ser-Glu-Cys-Pro-Leu-Ser-His-Asp-Gly-Tyr-Cys-Leu-His-Asp-Gly-Val-Cys-Met-Tyr-lle-Glu-Ala-Leu-Asp-Lys-Tyr-Ala-Cys-Asn-Cys-Val-Val-Gly-Tyr-lle-Gly-Glu-Arg-Cys-Gln-Tyr-Arg-Asp-Leu-Lys-Trp-Trp-Glu-Leu-Arg-COOH, где "X" - остаток тиоредоксина с мутациями Glu30His, Gln62His, "Y" - линкер -(Gly-Ser)2-Gly-(Asp)4-Lys-
SEQ ID №5 - нуклеотидная последовательность, кодирующая слитый белок формулы SEQ ID № 4:
5'-ATGAGCGATAAAATTATTCACCTGACTGACGACAGTTTTGACACGGATGTACTCAAAGCGGACGGGGCGATCCTCGTCGATTTCTGGGCACACTGGTGCGGTCCGTGCAAAATGATCGCCCCGATTCTGGATGAAATCGCTGACGAATATCAGGGCAAACTGACCGTTGCAAAACTGAACATCGATCATAACCCTGGCACTGCGCCGAAATATGGCATCCGTGGTATCCCGACTCTGCTGCTGTTCAAAAACGGTGAAGTGGCGGCAACCAAAGTGGGTGCACTGTCTAAAGGTCAGTTGAAAGAGTTCCTCGACGCTAACCTGGCCGGCTCTGGTTCTGGTGATGACGATGACAAGAAAAAAAAGAAACGTAAAGTTGAGGATCCGTACAATTCTGACTCTGAATGCCCATTGTCTCACGACGGTTACTGCTTGCACGACGGTGTTTGCATGTACATCGAAGCTCTGGACAAATACGCTTGCAACTGCGTTGTTGGTTACATCGGTGAACGTTGCCAATACCGAGATCTGAAATGGTGGGAACTGCGTTGA-3'



Изобретение относится к генной и тканевой инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве. Создан простой в конструктивном отношении пептидный вектор (PGE-k), состоящий из полипептидной последовательности эпидермального фактора роста (ЭФР) и модифицированной последовательности сигнального пептида Т-антигена SV-40. Новый вектор PGE-k способен обеспечить избирательное поступление генетического материала в цитоплазму клетки-мишени, несущей наружные рецепторы к ЭФР, и последующий транспорт его через ядерную мембрану. Предложен способ получения пептидного вектора PGE-k, предусматривающий его экспрессию в виде слитого белка “мутантный тиоредоксин-линкер-вектор” и расщепление экспрессированного в E.coli продукта в области линкера с помощью специфической протеазы. Получен рекомбинантный штамм E.coli B-8389 ВКПМ-продуцент слитого белка, включающего PGE-k. Простота конструкции, эффективность действия, отсутствие токсичности и иммуногенности определяют пригодность данного вектора для направленной генетической модификации in vivo эпителиальных, эмбриональных и опухолевых клеток. 6 с. и 1 з.п. ф-лы, 12 ил., 4 табл.

- NdeI/AatII фрагмента плазмиды pGEMEX-1, имеющего размер 2347 п.о. и содержащего участок инициации репликации Ori и ген устойчивости к ампициллину Вlа;
- AatII/HindIII фрагмента плазмиды pGEMEX-1, имеющего размер 755 п.о. и содержащего flOri,
- нуклеотидной последовательности по п.3.
- культивирование трансгенного штамма по п.5 в условиях, обеспечивающих экспрессию включающего пептидный вектор слитого белка;
- выделение и последующий гидролиз слитого белка энтерокиназой или другой протеазой, распознающей включенную в него линкерную последовательность и специфически расщепляющей пептидную связь, следующую за указанным линкером, и
- выделение целевого продукта.
| СПОСОБ НАПРАВЛЕННОЙ ГЕНЕТИЧЕСКОЙ ТРАНСФОРМАЦИИ МОЛОЧНОЙ ЖЕЛЕЗЫ ЖИВОТНОГО И УСТРОЙСТВО ДЛЯ ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В МОЛОЧНЫЙ ПРОТОК МОЛОЧНОЙ ЖЕЛЕЗЫ ЖИВОТНОГО | 1993 |
|
RU2025487C1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| COLLASP, ALESTROMP., Cell BioL, 1997, 75(5), 633-640 | |||
| US 5646016, 10.12.1993. | |||

