Настоящее изобретение относится к способу получения трансгенных растений, имеющих модифицированный характер распределения фруктана по сравнению с нетрансформированными растениями; к ДНК-конструкциям для продуцирования трансгенных растений или растительных тканей; к трансгенным растениям или растительным тканям, имеющим модифицированный характер распределения по сравнению с нетрансформированными растениями; к фруктанам, выделенным из растений или тканей; к использованию трансгенных растений, тканей и фруктанов в различных целях; к семенам, полученным от трансгенных растений.
Растения продуцируют множество углеводов, употребляемых в пищу или используемых в промышленных целях. Примерами важных углеводов являются крахмал, целлюлоза и сахароза. Эти соединения используются человеком либо в их натуральном виде, либо в модифицированной форме, для употребления в пищу и в промышленном производстве. Культурные растения, продуцирующие указанные углеводы, были получены традиционными методами селекции растений. Помимо вышеуказанных хорошо известных углеводов, в растениях могут присутствовать и многие другие растительные углеводы, обладающие ценными свойствами.
Растительные углеводы могут быть разделены на две группы в зависимости от той роли, которую они играют в растениях. К первой группе относятся структурные углеводы, которые составляют, в основном, часть внеклеточного матрикса. Наиболее известным углеводом, принадлежащим к этой группе, является целлюлоза. Ко второй группе относятся неструктурные запасные углеводы, которые могут служить в качестве долговременных или кратковременных (неустойчивых) резервных углеводных отложений. Примерами таких углеводов могут служить крахмал, сахароза и фруктаны.
Фруктан представляет собой полимер, состоящий главным образом из повторяющихся фруктозных звеньев. Фруктаны в основном содержатся в тех видах растений, которые обычно не используются в рациональных фермерских хозяйствах. Фруктаны присутствуют в однодольных (например, Poaceae, Libiaceae) и двудольных (например, Compositae) растениях (Pollock & Cairus, 1991, Ann. Rev. Plant Physiol. Plant Mol. Biol. 42, 77-101; Hendry 1987, New Phytol. 106, 201 - 216).
Было высказано предположение, что, помимо своей роли как запасных растительных углеводов, фруктаны имеют и другие функции. Такими функциями могут быть, например, сообщение растению толерантности к сухому и холодному климату (индуцированная холодом потеря влаги (Pontis 1989, J. Plant Physiol. 134, 148 - 150; Gonzales et al., 1990, New Phytol. 115, 319-323)).
Однако фруктаны, продуцируемые растениями, обычно имеют ограниченную функциональность. По этим причинам фруктаны пока еще не находят широкого применения в пищевой промышленности и других производствах, несмотря на их общепризнанную потенциальную коммерческую ценность (Fuchs 1991. Biochem. Soc. Transact. 19, 555-560, A. Fuchs, ed. Inulin and Inulin-containing Crops, Elsevier, 1993), особенно по сравнению с другими углеводами.
Одна из главных проблем, не позволяющих широко использовать растительные фруктаны в промышленном производстве, заключается в ограниченном диапазоне видов растений, накапливающих фруктаны в значительных количествах. Во многих ценных сельскохозяйственных культурах, к которым, например, относятся сахарная свекла и картофель, фруктаны либо отсутствуют, либо содержатся в очень незначительных количествах. Растения, способные накапливать фруктаны до определенного уровня, часто обладают неблагоприятными агрономическими свойствами. Примером такого растения является Helianthus tuberosus.
Другой проблемой является ограниченная функциональность продуцируемых фруктанов. Указанная функциональность inter alia определяется длиной цепи фруктана, которая в растениях редко превышает так называемую "степень полимеризации" (DP) моносахаридов, равную 100. Но обычно в природе встречаются более мелкие фруктаны, степень полимеризации которых резко убывает до сбора или после сбора урожая и/или при его хранении. Низкая степень полимеризации (DP) фруктанов сильно ограничивает его утилизацию в последующих процессах и может приводить к снижению урожая.
В настоящее время установлено, что некоторые растения могут быть трансформированы в целях продуцирования фруктанов в соответствующих растениях-хозяевах.
Таким образом, настоящее изобретение относится к способу получения трансгенных растений, имеющих модифицированный характер распределения фруктанов по сравнению с нетрансформированными растениями, причем указанный способ включает в себя следующие стадии:
a) получение ДНК-конструкций, содержащих один или несколько генов фруктозилтрансферазы, или их модифицированных вариантов, надлежащим образом связанных с последовательностями промотора и терминатора, которые являются активными в растениях;
b) трансформацию растительной клетки с помощью указанных конструкций;
c) регенерацию трансгенных растений из трансформированной растительной клетки.
Гены фруктозилтрансферазы кодируют ферменты, которые катализируют образование фруктозо-фруктозных связей. В Международной патентной заявке WO 89/12386 для модифицирования состава растворимых твердых компонентов в растительных клетках было предложено продуцировать трансгенные растения, содержащие чужеродный ген, кодирующий фермент, способный к полимеризации углеводов, например таких, как различные сахарозы. В заявке WO 89/12386 раскрывается inter alia использование декстрансахарозы. Этот фермент использует сахарозу в качестве источника для полимеризации глюкозных звеньев. Однако настоящее изобретение относится к полимеризации фруктозных звеньев.
Авторами настоящего изобретения было установлено, что благодаря присутствию микробной 5'-нетранслируемой области гена фруктозилтрансферазы Sac B, в которой рамочный кодон ATG находится перед инициирующим кодоном ATG, невозможно получить приемлемые уровни экспрессии этого гена. Таким образом, в соответствии с другим вариантом настоящего изобретения было установлено, что соответствующая модификация 5'-нетранслируемой области гена фруктозилтрансферазы способствует повышению уровня экспрессии гена фруктозилтрансферазы в растительной клетке. Подходящей модификацией можно считать любую модификацию, которая не оказывает неблагоприятного воздействия на уровень экспрессии гена фруктозилтрансферазы. Фактически это означает, что любая последовательность, негативно влияющая на уровень экспрессии указанного гена, должна быть удалена. При этом предпочтительно, чтобы из указанного микробного гена была полностью удалена 5'-нетранслируемая область до кодона ATG.
Термин "характер распределения фруктана", используемый в настоящем описании, означает распределение фруктана в различных растительных тканях и в различных отделах растительной клетки, таких как цитозоль, вакуоль, апопласт и т.п. Характер распределения фруктана в растениях, не продуцирующих фруктан в нетрансформированном виде, может быть изменен путем введения гена, кодирующего фруктозилтрансферазу. Распределение фруктана в растении может быть также изменено путем переориентирования пути метаболизма в растениях или растительных клетках на определенное растение или определенные отделы растительных клеток.
Сахароза, которая является субстратом для фруктозилтрансфераз, представляет собой углевод, обнаруживаемый в нескольких различных участках клетки. Сахароза синтезируется в цитоплазме и значительное количество этого углевода обнаруживается в цитозоле, в вакуоли и во внеклеточном пространстве (апопласте).
Поскольку биохимические процессы в растительных клетках часто ограничиваются одним или несколькими отделами клетки, то предпочтительно стимулировать накопление продуктов введенных генов в конкретном отделе клетки. Поэтому на фруктозилтрансферазах, продуцируемых в трансгенных растениях, должны присутствовать последовательности, обозначающие направленную доставку этих ферментов в конкретный отдел клетки и являющиеся соответственно специфичными для данного отдела клетки. Были идентифицированы и проанализированы специфичные аминокислотные последовательности, обеспечивающие доставку фруктозилтрансферазы в различные участки клетки. ДНК-последовательности, кодирующие указанные последовательности специфических аминокислотных участков, могут быть сцеплены с нужными генами таким образом, чтобы после экспрессии такого химерного гена продуцировался белок, в котором целевая информация распознавалась и использовалась клеткой для доставки в соответствующие области этой клетки (Oakes et al. EPO 0486683, WO 91/19808, Symons et al. Bio/Technology 8, 217-221 (1990), Pen J. et al. Bio/Technology 10, 292-296 (1990)).
Поэтому в предпочтительном варианте осуществления настоящего изобретения полигенный экспрессирующий кластер содержит также последовательность, обеспечивающую направленную доставку фруктозилтрансферазы в один или несколько конкретных отделов растения или растительной клетки. Такими последовательностями - "поставщиками" могут быть любые ДНК-последовательности, способные обеспечивать целевую доставку фруктозилтрансферазы в один или несколько конкретных отделов растения или растительной клетки. Примерами таких последовательностей - "поставщиков" являются ответственные за доставку сигнальная и вакуолярная последовательность гена карбоксипептидазы Y (срy), или сигнальная и апопластическая последовательность гена белка, ассоциированного с патогенезом (pr-s). Таким образом, настоящее изобретение относится к способу, где путем добавления ответственной за доставку последовательности к ДНК-конструкции могут быть продуцированы трансгенные растения, в которых фруктозилтрансфераза направляется в один или несколько специфических отделов клетки, в результате чего обеспечивается нужное распределение фруктана в растении или в клетках растения.
Часто оказывается необходимым регулировать не только место, но также и время экспрессии вводимых генов. Например, предпочтительно ограничивать экспрессию вводимой ферментной активности конкретными участками растений, а именно теми органами растений, урожаи которых обычно собирают фермеры, например, такими органами растений как клубни, плоды или семена. Более того, часто оказывается желательным инициировать экспрессию в этих органах на определенной стадии развития. Это особенно предпочтительно в тех случаях, если экспрессия вводимых генов мешает нормальному развитию этих органов растения.
Фруктозилтрансферазы настоящего изобретения используют сахарозу в качестве субстрата для синтеза высокомолекулярных фруктанов. Многие микроорганизмы содержат фруктозилтрансферазы, обладающие способностью продуцировать фруктаны (часто называемые леванами) из сахарозы (A.Fuchs 1959, Thesis, University of Leiden, Han 1989 Adv. Appl. Microbiol. 35, 171-194). Эти ферменты могут переносить фруктозные звенья от сахарозы к фруктановой акцепторной молекуле для продуцирования высокомолекулярных фруктанов. Поскольку указанные акцепторные молекулы изначально происходят от сахарозы, они могут еще содержать концевую молекулу глюкозы. Реакция протекает по следующей схеме (см., например, Han 1989 Adv. Appl. Мicrobiol., 35, 171-194):
n(G-F) ---> G-(F)n + n - IG
G-F = сахароза, G = глюкоза, F = фруктоза.
В зависимости от конкретного вида фруктозилтрансферазы гликозидная связь, соединяющая фруктозные звенья, может быть (2-1)- или (2-6)-типа. В микроорганизмах, в одной и той же молекуле фруктана могут присутствовать связи обоих типов. Функциональность молекулы фруктана зависит от типа (2-1)- или (2-6)-связей скелета молекулы, от степени ее разветвления и полимеризации.
Многие микроорганизмы, такие как бактерии, дрожжи, грибы и т.п., содержат фруктозилтрансферазы, способные синтезировать фруктаны с высокой степенью полимеризации (DP) из сахарозы. Обычно накопление фруктанов происходит во внеклеточном пространстве.
Примерами бактерий, продуцирующих фруктозилтрансферазу, являются две бактерии Bacillus subtillis (Steinmenz et al., 1985 Mol. Gen. Genet. 200, 220-228) и Streptococcus mutant (Shirosa & kuramitsu 1988 J.Bactetiol. 170, 810-816), B. subtilis структурный ген sac B кодирует левансахаразу, т.е. фруктозилтрансферазу. Этот фермент способен превращать сахарозу во фруктозный полимер со степенью полимеризации, которая может легко превышать 10000. В этом фруктане, продуцируемом левансахаразой, в основном присутствуют связи (2-6)-типа с экстенсивным (2-1)-ветвлением.
В другом микроорганизме, а именно в S. mutant, ген ftf кодирует фруктозилтрансферазу, которая также продуцирует фруктаны с очень высокой степенью полимеризации. В этом фруктане фруктозные единицы в основном связаны между собой связями (2-1)-типа с (2-6)-ветвлением (Rosell & Birkhed 1974 Acta Chem. Scand. B28, 589)
Бактериальные фруктозилтрансферазы имеют относительно низкое значение Km (константа Михаэлиса) для сахарозы, составляющее приблизительно 20 мМ. Концентрации сахарозы в большинстве растений значительно выше, а поэтому эти ферменты должны быть активными в растениях. Другим важным свойством бактериальных фруктозилтрансфераз является их способность синтезировать леваны при низких температурах вплоть до 0oC (Tanaka и др., 1978 Aoric. Biol. Chem. 42, 323-326). Растения часто находятся в условиях подобных температур и в этих условиях ферменты должны быть еще вполне активными.
Выбор предпочтительного структурного гена (или генов) фруктозилтрансферазы зависит от требований, предъявляемых к степени полимеризации и к степени ветвления фруктанов.
Настоящее изобретение не ограничивается микробными фруктазилтрансферазами и для осуществления его целей могут быть также использованы фруктозилтрансферазы, происходящие от растений или любых других прокариотических или эукариотических источников.
Так, например, биосинтез и разложение фруктанов в растениях были исследованы на Helianthus tuberosus (топинамбур) и несколько травах (Pollock & Cairns 1991, Ann. Rev. Plant Physiol. Plant Mol. Biol. 42, 77-101; Pollock 1986, New Phytol. 104, 1 - 24). В растениях различных семейств синтезируются различные типы фруктанов, как линейные, так и разветвленные, имеющие гликозидные (2-1)- и (2-6)-связи между фруктозными молекулами. В растениях фруктаны накапливаются в вакуолях. Субстратом для биосинтеза фруктанов является сахароза, однако сродство растительных ферментов к сахарозе значительно ниже (приблизительно Km 100), чем в случае бактериальных ферментов. Растительные ферменты, участвующие в метаболизме фруктана, являются весьма толерантными к условиям низких температур (Pontis 1989, J. Plant Physiol. 134, 148-150; Gonzales et al., 1990 New Phytol. 115, 319-323), а поэтому эти ферменты могут быть с успехом использованы для вполне определенных целей.
Почти все растения обладают способностью к синтезу сахарозы, который происходит в цитоплазме. Сахароза представляет собой важнейшую транспортную форму углеводов, которая переносится от ее источника (ткань, "экспортирующая" чистый углевод) к "потребителю" (ткань, "импортирующая" чистый углевод) с помощью сосудистой системы. Этот перенос часто представляет собой транспорт сахарозы через внеклеточное пространство - апопласт. Кроме того, сахароза может быть использована растениями в качестве запасного углерода. В этом случае сахароза обычно аккумулируется в вакуолях растительных клеток, например в корнях сахарной свеклы. Из этого следует, что основными участками локализации сахарозы в растительных клетках является цитозаль и вакуоль. Кроме того, значительные количества сахарозы присутствуют во внеклеточном пространстве (апопласте) многих растений.
Предпочтительные последовательности, ответственные за доставку, должны направлять фруктозилтранферазу к тем отделам клетки, где находится субстрат, а именно к вакуоли, цитозоли и апопласту соответственно. Предпочтительным отделом для значительных накоплений фруктана является, например, вакуоль. Однако для других целей (например, для сообщения растению засухоустойчивости и холодоустойчивости) предпочтительными отделами для накопления фруктана могут оказаться цитозоль или апопласт.
Тот факт, что во многих растениях накопление сахарозы происходит в вакуолях, означает, что указанный участок клетки является идеальным для биосинтеза фруктана микробными или растительными ферментами.
Поэтому в предпочтительном варианте своего осуществления настоящее изобретение относится к способу вакуолярной экспрессии микробных, растительных или других фруктозилтрансфераз в трансгенных растениях. Этот способ предусматривает сшивание ДНК-последовательностей, кодирующих сигнал специфической экспрессии, сигнал вакуолярной локализации, и микробные, растительные или другие трансферазы.
Важными участками для накопления фруктана могут быть также и два других компартмента, в которых присутствует сахароза, а именно цитозоль и апопласт. Это может быть достигнуто путем направленной доставки микробной, растительной или какой-либо другой фруктозилтрансферазы в цитозоль или апопласт. Для осуществления цитозольной экспрессии не требуется введения в клетку специальной информации; для этого вполне достаточно экспрессии зрелых ферментов. Что касается апопластической экспрессии фруктозилтрансферазы, то для ее осуществления необходимо, чтобы к ферменту, осуществляющему целевую доставку в апопласт, была добавлена специфическая последовательность, обеспечивающая эту доставку.
Для осуществления экспрессии модифицированного гена фруктозилтрансферазы в растениях, к ДНК-конструкции необходимо добавить промоторные сигналы. Такие промоторы экспрессии могут быть специфичными к определенному типу клеток либо они могут быть активными в различных типах клеток широкого ряда. Кроме того, время экспрессии может быть определено, например, путем использования промоторов с временной регуляцией. Обычно, для экспрессии гена в растениях используют промотор 35S вируса мозаики цветной капусты, который является активным во многих типах клеток растений независимо от стадии их развития.
В зависимости от цели использования и от выбранного растения-мишени, предпочтительным промотором может быть тканеспецифический или конститутивный, сильный или слабый промотор. Для продуцирования фруктана в органах запасания, предпочтительным промотором является сильный промотор, специфичный для ткани-акцептора (т.е. накапливающей ткани).
Для еще большего увеличения уровней транскрипции, этот промотор может быть модифицирован так, чтобы он содержал дуплекс энхансеров.
Трансляция мРНК может быть улучшена путем добавления трансляционного энхансера, такого, как трансляционный энхансер РНК4 вируса мозаики люцерны, который должен присутствовать в транскрибированной 5'-нетранслируемой области.
Для правильной терминации транскрипции, к ДНК-конструкции должна быть добавлена последовательность растениеспецифического терминатора. Примером такой последовательности может служить терминатор гена нопалинсинтазы.
Настоящее изобретение не ограничивается природными фруктозилтрансферазами, и с таким же успехом может быть осуществлено с использованием их модифицированных вариантов. Модификации могут влиять на активность фруктозилтрансфераз, например, таким образом, что в результате может быть изменена степень полимеризации или структура продуцируемого фруктана.
Индуцируемая аккумуляция фруктанов в трансгенных растениях в соответствии с вышеописанным способом осуществляется с учетом экстракции фруктанов из этих растений в целях производства указанных фруктанов. В этих растениях фруктаны могут накапливаться в тех органах, которые обычно собирают при снятии урожаев, например в таких, как корни, корнеплоды, листья, стебли, клубни, плоды и семена.
Генетически модифицированные культурные растения, содержащие вышеупомянутую конструкцию, кодирующую фруктозилтрансферазу, являются эффективными продуцентами высококачественного углеводного полимера, используемого человеком.
Настоящее изобретение относится к способу получения растений, обладающих новыми биосинтетическими свойствами, которые позволяют этим растениям продуцировать и аккумулировать фруктан. Эти растения имеют улучшенные свойства благодаря изменению соотношения "источник-потребитель" и выхода, либо они имеют улучшенные свойства в условиях абиотического и биотического стрессов. Иллюстрирующими, но не ограничивающими примерами таких стрессов могут служить засуха, световое излучение, температура, болезни и вредители. Указанными улучшенными свойствами растения в нормальных или стрессовых условиях является высокое содержание сухих веществ, улучшенный вкус или лучшая сохраняемость, более высокие питательные свойства и т.п. Указанные растения могут быть использованы в качестве сырья для производства фруктана.
В соответствии с настоящим изобретением может быть использован один ген фруктозилтрансферазы или комбинация генов фруктозилтрансферазы прокариотического или эукариотического происхождения. Эти гены могут кодировать ферменты для биосинтеза фруктанов широкого ряда.
Настоящее изобретение, кроме того, относится к семенам, черенкам, клубням, луковицам или другим частям трансгенных растений, которые могут быть использованы для непрерывного продуцирования последующих генераций указанных растений.
Фруктаны, продуцируемые с использованием трансгенных растений настоящего изобретения, могут быть использованы в пищевой промышленности и в других производствах. Так, например, указанные фруктаны могут быть использованы для изготовления пищевых продуктов, употребляемых человеком и животными, а именно для изготовления фруктозных сиропов, а также для изготовления химических или пластиковых материалов в обычной или модифицированной форме.
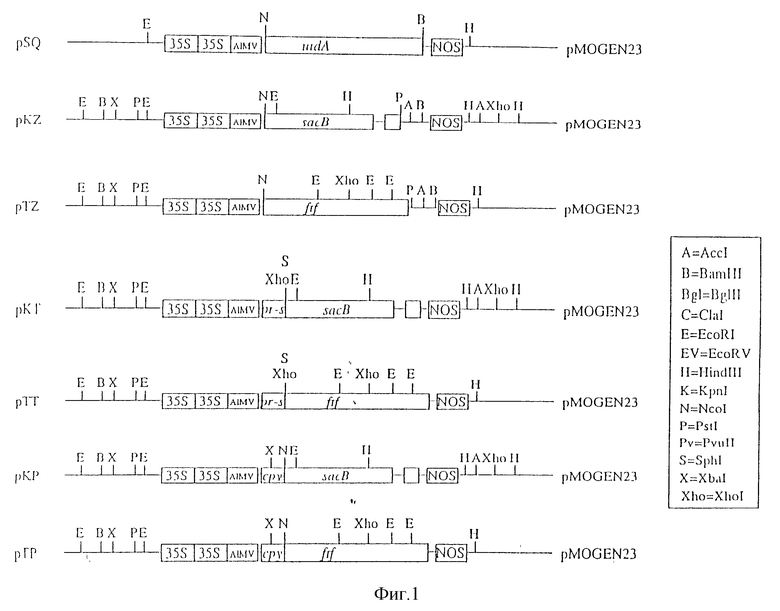
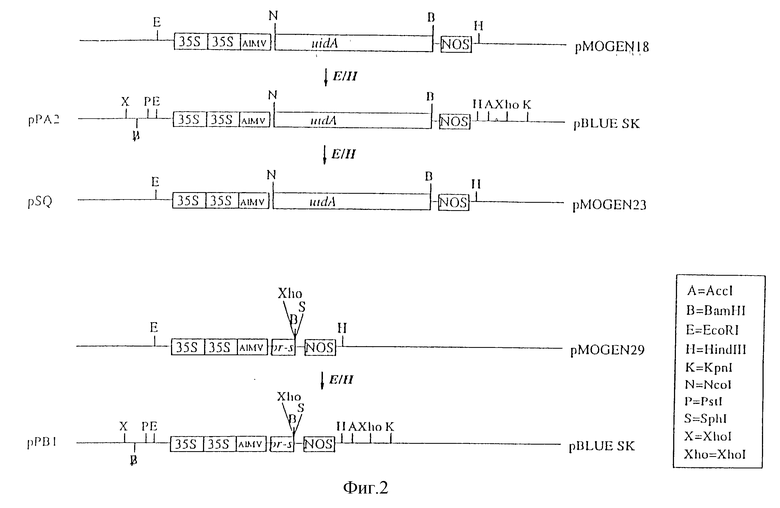
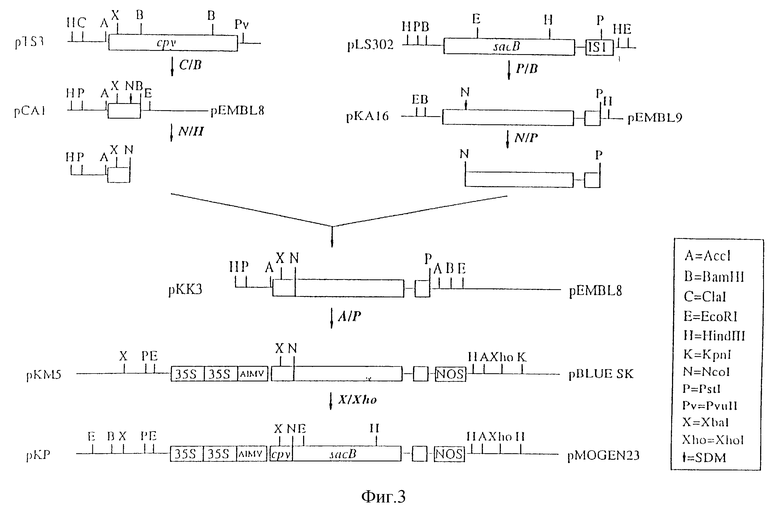
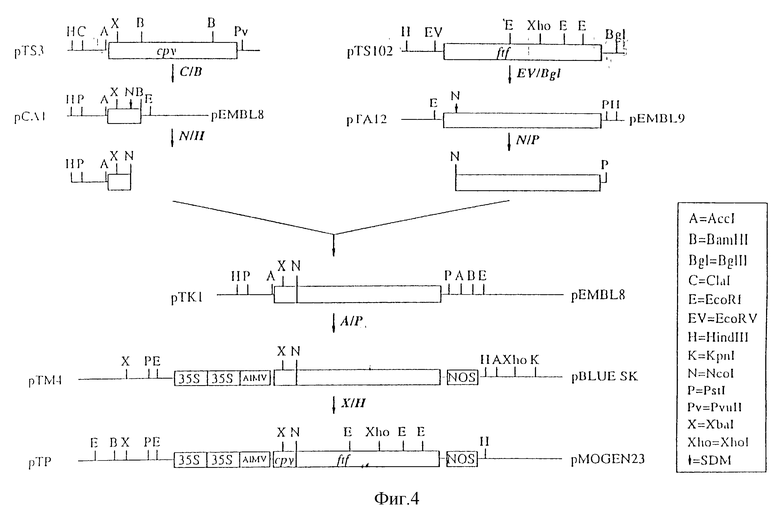
Настоящее изобретение иллюстрируется нижеследующими рисунками, где:
На фиг. 1 схематично показаны ДНК-конструкции;
На фиг. 2 показано конструирование плазмид рРА2 и рРВ1;
На фиг. 3 показано конструирование 35S-cpy-sacB-Nos в бинарном векторе (pKP);
На фиг. 4 показано конструирование 35S-cpy-ftf-Nos в бинарном векторе (pTP);
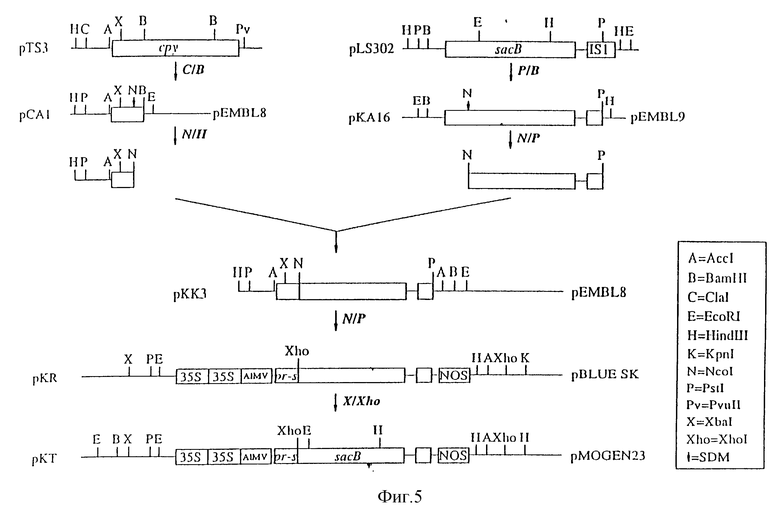
На фиг. 5 показано конструирование 35S-pr-s-sacB-Nos в бинарном векторе (pKT);
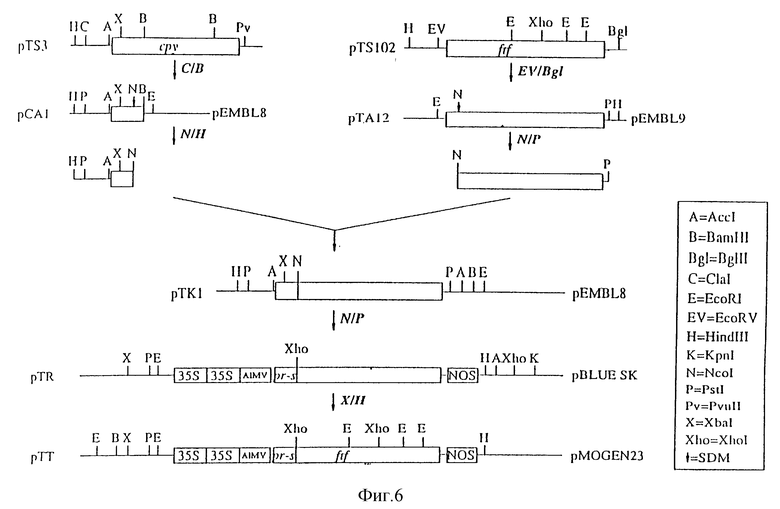
На фиг. 6 показано конструирование 35S-pr-s-ftf-Nos в бинарном векторе (pTT);
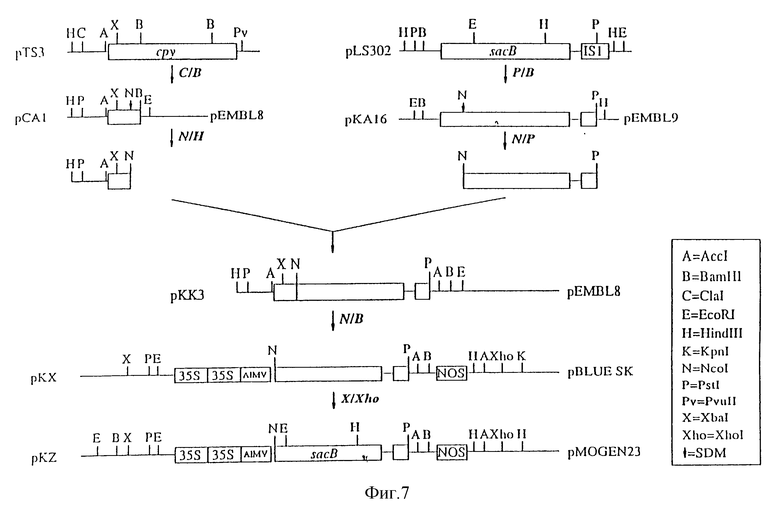
На фиг. 7 показано конструирование 35S-SacB-Nos в бинарном векторе (pKZ);
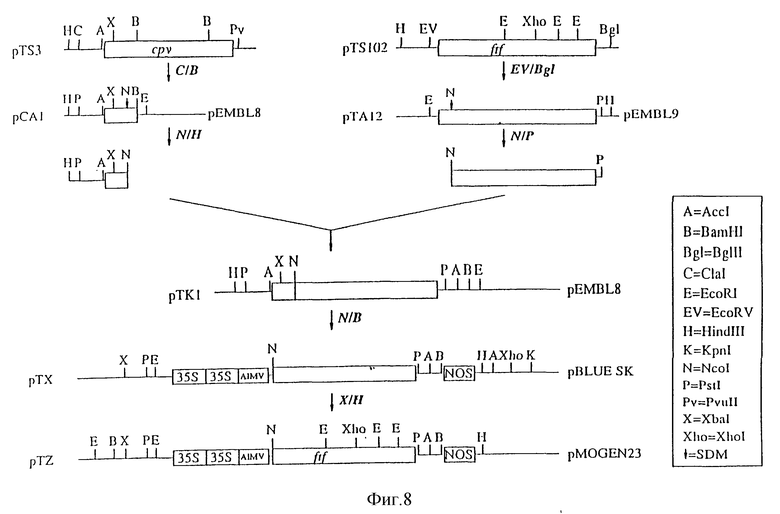
на фиг. 8 показано конструирование 35S-ftf-Nos в бинарном векторе (pTZ); и
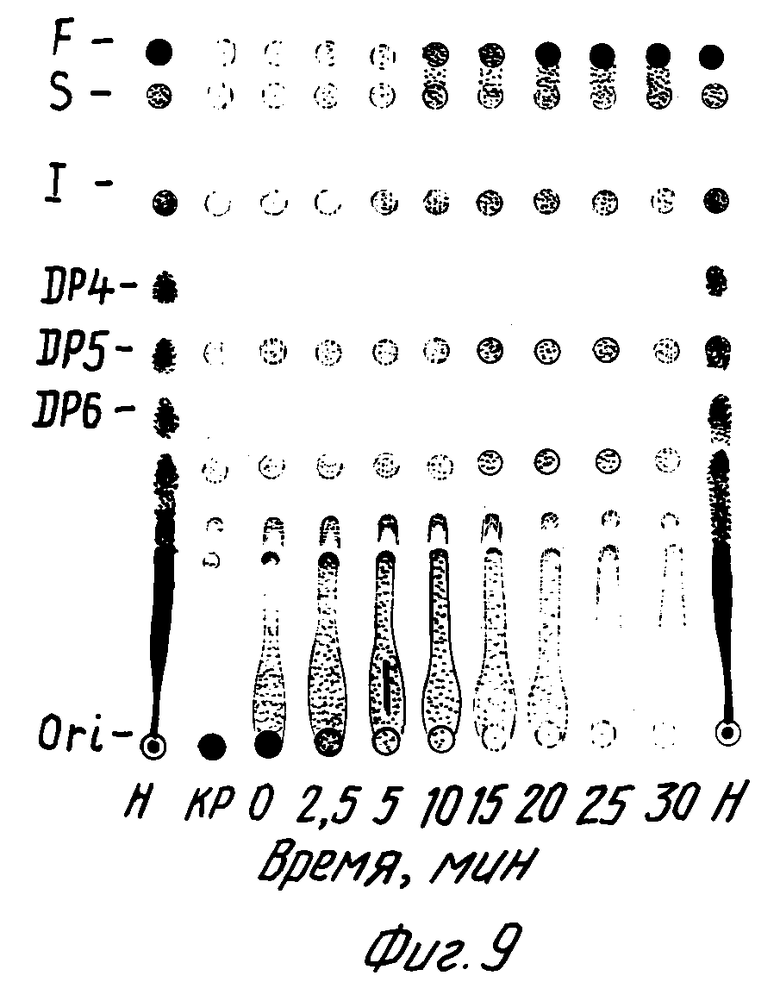
На фиг. 9 показан пример частичного кислотного гидролиза фруктана, выделенного из трансгенного растения табака КР. Эта картина частичного гидролиза идентична картине гидролиза фруктанов, продуцируемых Bacillus subtilis. На фиг. 9 были приняты следующие обозначения: F -фруктоза; S - сахароза (дисахарид); I - I-кестоза (трисахарид); DP4, DP5 и DP6 -тетра-, пента- и гексасахариды, соответственно; и H - экстракт из клубней Helianthus tuberosus (земляная груша), используемый в качестве стандарта.
Настоящее изобретение проиллюстрировано нижеследующими примерами, которые, однако, не следует рассматривать как некое ограничение объема изобретения.
Пример 1.
Экспрессия sacB-гена в вакуоли
1. Отбор генов для использования
Для генерирования фруктан-продуцирующих трансгенных растений проводили отбор генов, кодирующих белки, способные продуцировать фруктаны. В результате для использования в настоящем изобретении был выбран один из этих генов, а именно ген sacB Bacillus subbilis, кодирующий левансахаразу (Steinmetz M. и др., Mol. Gen. Genet. 200 : 220 - 228 (1985)). Этот фермент продуцирует, в основном, разветвленные фруктаны со (2-6)-связями в присутствии сахарозы.
Поскольку в растительных клетках сахароза накапливается в вакуолях, то вакуоли являются предпочтительным местом продуцирования фруктана. Для направленной доставки левансахаразы в вакуоль была использована ответственная за такую доставку область карбоксипептидазы Y (Valls L.A. и др., Cell 48, 887-897 (1987)).
2. Конструирование 35S-cpy-sacB-Nos в бинарном векторе (рКР)
Плазмида pLS 302 была описана Steinmeftz и др. (см. выше). Ген левансахзаразы (sacB) в виде BarHI-PstI - фрагмента клонировали из этой плазмиды в область множественного клонирования плазмиды рEMBL9 (Donte L. et. al., Nucleic Acids Res., 11, 1645 - 1655 (1983)) в соответствующие Bam HI и PstI - сайты, в результате чего получали плазмиду pKA16. Общая методика клонирования ДНК описана в литературе (см., Sambrook J. et al., Cold Spring Harbour, NY, Cold spring Harbour Laboratory (1989)).
Для создания Ncol - сайта возле сайта процессинга гена левансахаразы в положении нуклеотида 550 (Steinmentz M. и др. см. выше) осуществляли сайт-направленный мутагенез как описано Kramer W. и др. (Nucleic Acids, Res, 12, 9441-9456 (1984) с использованием следующего олигонуклеотида 5'-GCAACTCAAGCCATGGCGAAAGAAACG-3', в результате чего получали плазмиду pK 22.
В положении аминокислоты-2 по отношению к сайту процессинга (положение нуклеотида 544), фенилаланин заменялся на метионин. Ncol-Pst1 - фрагмент, в котором присутствует последовательность, кодирующая зрелый белок левансахаразы, был использован для последующего клонирования.
Плазмида pTs3, содержащая ген карбоксипептидазы Y(cpy), была описана Valls L.A. и др., см выше.
Первую часть cpy - гена клонировали из этой плазмиды в виде ClaI (-695) и BamHI (462)-фрагмента в область множественного клонирования плазмиды pEMBL 8 (Dente L. и др., см., выше) в соответствующие Acc 1 - Bam H1-сайты, в результате чего получали плазмиду pCAI.
Для создания Ncol-сайта возле сайта процессинга, расположенного ниже вакуолярной последовательности доставки гена cpy в положении 330 (Valls L.A. и др. , см. выше), осуществляли сайт-направленный мутагенез, описанный Kramer W. и др. (см. выше) с использованием следующего олигонуклеотида: 5'-CGTGTCAACAAGACCATGGACCCTAA-3'. Полученная в результате плазмида pCB50 содержала первую часть гена cpy с Ncol - сайтом у сайта процессинга. В аминокислотном положении +2, по отношению к сайту процессинга в нуклеотидном положении 334, изолейцин заменялся на треонин. В аминокислотном положении +3 по отношению к сайту процессинга в нуклеотидном положении 337, лезин заменялся метионином. Hind 111 - Ncol фрагмент, в котором присутствует последовательность, кодирующая часть карбоксипептидазы Y, ответственную за целевую доставку в вакуоль, использовали для последующего клонирования.
Ответственную за доставку в вакуоль последовательность гена cpy сшивали со зрелой последовательностью гена левансахаразы, используя при этом трехточечное лигирование (Sambrook J. и др., см. выше).
Нижеследующие фрагменты были использованы в эквимолярных концентрациях, а именно: Hind 111 - Ncol - фрагмент гена cpy (кодирующего последовательность, осуществляющую доставку в вакуоль); Ncol- Pst1- фрагмент гена sacB; и фрагмент, содержащий вектор pEMBLB 8, переваренный ферментами Hind 111 - Pst1. Полученная в результате плазмида pKK3 кодирует конструкцию, полученную путем лигирования с сохранением рамки считывания гена карбоксипептидазы и гена Y-левансахаразы. Правильная рамка считывания гибридного гена была подтверждена анализом последовательности.
Плазмида pMOG18, которая содержит растительный промотор с энхансерным дуплексом и последовательности, усиливающие трансляцию мРНК, описана Symons et al., Bio/Technology 8, 217-221 (1990)). Эта плазмида содержит конструкцию "35S-промотор/uidA-ген/ Nos-терминатор". Вектор p Bluescript 11 sk (Stаratagene, San Diege, CA), из которого был удален внутренний Bam H1-сайт путем Bam H1-рестрикции, затупления липких концов фрагментом Кленова, и последующего лигирования, был использован для дальнейшего клонирования. 35S-uidA-Nos-фрагмент получали путем переваривания плазмиды pMOG18 ферментами EcоR1 и Hind III, а затем клонировали в Bam H1 - pBluescript в соответствующие EcoRI - Hind III - сайты, в результате чего получали плазмиду pPA2. Эту плазмиду переваривали ферментами Ncol и Bam H1 в целях удаления гена uid A. Полученный вектор обрабатывали s1-нуклеазой для удаления выступающих концов, а затем дефосфорилировали. Из плазмиды pKK3 путем переваривания ферментами Bcc1 и Pst 1 выделяли фрагмент, содержащий гибридный ген cpy-sacB. Указанные сайты затупляли фрагментом Кленова. После этого два фрагмента лигировали в полученную плазмиду pKM5. Эта плазмида содержала конструкцию "35S-промотор/ cpy-sacB-ген/Noc-терминатор".
Плазмиду pKM5 сначала переваривали ферментом Xho1, а затем частично переваривали ферментом Xba 1. Фрагмент, содержащий полную конструкцию (35S-cpy-sacB-Nos), клонировали в Xba-1 и Xho-1-сайт pMOG23 (Symons и др., см. выше), производной бинарного растительного вектора (Bevan M., Nucl. Acids Res, 12: 8711-8721), в результате чего получали плазмиду pKP.
3. Трансформация pKP в растения табака
Плазмиду pKP конъюгировали в Agrobacterium Dumefaciens LB 4404 (Hoe kema A. ex al, Nature 308, 179 - 180 (1983)) при тройном спаривании с использованием хелперной плазмиды pPK2013 (Lam S.T. et al., Plasmid 13, 200-204 (1985)). Эту конструкцию вводили в Nicotiana tabacum var. Petit Havanna (SPI) при помощи метода трансформации с использованием листовых дисков. Регенерированные растения, именуемые KP-растениями, отбирали на резистентность к канамицину и культивировали на MS-среде (Murashige & Skoog Phusiol. Plant 15, 473-497 (1962)), содержащей глюкозу вместо сахарозы. Затем растения выращивали на почве в теплице и анализировали.
4. Анализ KP-растений
Растения выращивали в теплице, после чего листовой материал обрезали и измельчали в пробирке Эппендорфа. После центрифугирования (2 мин., 16000 об/мин. ), 1 мкл супернатанта наносили каплями на ТСХ-пластины (Cairus A.J. & Pollock C. Y. New Phytol 109, 399 - 405 (1988)). Затем три раза проявляли в смеси ацетона и воды (85:15) и обрабатывали путем опрыскивания мочевиной как описано в Wise C.S. et al, Analitical Chemictry 27, 33-36 (1955)). В этом методе предпочтительно окрашиваются фруктоза и фруктозосодержащие полимеры.
Фруктозные полимеры не были обнаружены ни в нетрансформированных растениях, ни в растениях, трансформированных неродственными конструкциями.
Скриннинг трансформантов с использованием вышеуказанного метода обнаружил экстенсивную аккумуляцию фруктанов в этих растениях. Уровни экспрессии фруктанов варьировались между отдельными трансформированными растениями, содержащими одну и ту же конструкцию, как это обычно бывает в экспериментах с растениями-трансформантами. Такие отклонения в уровнях экспрессии зависят, главным образом, от сайта геномной интеграции (эффект положения).
Аккумуляцию фруктанов в этих растениях, кроме того, исследовали путем выделения больших количеств фруктана (Livingstone 111 DA, Plant Physiol. 92, 767-769 (1990). Этот фруктан анализировали путем определения его размера на ЖХБР-колонке (жидкостная хроматография быстрого разрешения) (Phаrmacia) с Superosa 6HP 10/30, в результате чего фруктаны были обнаружены в свободном объеме колонки, что указывало на степень полимеризации > 25000 фруктозных звеньев. Фруктан, продуцированный левансахаразой Bacillus subtilis, аналогичным образом элюируется в свободном объеме Superose-колонки.
Частичная деградация посредством кислотного гидролиза (см. фиг. 9) показала характерную картину распределения продуктов гидролиза. Очищенные растительные и бактериальные фруктаны обнаруживали идентичную картину деградации на ТСХ. Полный кислотный гидролиз и последующий ВЭЖХ-анализ на Aminex HPX87C-колонке (Biorad) при 85oC с использованием воды в качестве элюента показали, что указанный фруктан состоит из фруктозы.
Очищенные фруктаны были также проанализированы с помощью протонного ЯМР. Фруктан, выделенный из трансгенных растений, не обнаруживал каких-либо различий в характере пиков по сравнению с фруктаном, синтезированным левансахаразой Basillus subtilis.
Мы не обнаружили какой-либо фруктан-гидролизирующей активности в нескольких видах растений (табак, сахарная свекла, томаты, картофель). При инкубировании экстрактов растительного белка с фруктаном в течение продолжительного периода времени в фосфатно-цитратном буфере pH 5,0 высвобождения фруктозы не наблюдалось.
После физиологического старения растения фруктаны остаются в трансгенных растениях, что подтверждает их стабильность, проиллюстрированную с помощью белковых экстрактов. Накопление фруктанов происходит во всех частях растения, что соответствует неспецифической активности промотора 35S в трансгенных растениях.
Трансгенные растения давали нормальные семена. Прорастание семян проходило идентично прорастанию семян нетрансформированных растений, а содержащаяся в трансгенных растениях конструкция стабильно наследовалась последующей генерацией, которая продуцировала фруктаны аналогичным образом.
Пример 2
Экспрессия гена ftf в вакуоли
1. Отбор генов
Для генерирования фруктанпродуцирующих растений проводили отбор генов, кодирующих белки, способные продуцировать фруктаны. Для этого был использован другой ген, а именно ген ftf Streptococcus mutant, кодирующий фруктозил-трансферазу (Shirosa T. & Kuramitsu H. K. , Y.Bacteriol 170, 810-816 (1988)). Этот фермент продуцирует, главным образом, разветвленные фруктаны со (2-1)-связями в присутствии сахарозы. Поскольку в растительных клетках сахароза может накапливаться в вакуолях, то вакуоли являются предпочтительным местом продуцирования фруктанов. Поэтому был выбран ген, кодирующий белок, способный к самодоставке в вакуоль, а именно карбоксипептидазу Y (Valls L. A. et al., см. выше). Сигнальную последовательность и последовательность, ответственную за доставку в вакуоль, от указанного гена использовали как описано в примере 1.
2. Конструирование 35S-cpy-ftf-Nos в бинарном векторе (pTP)
Плазмида pTS 102 была описана Shiroza T. и Kuramitsu H.K. (см. выше). Ген фруктозилтрансферазы (ftf) клонировали из этой плазмиды в виде EcoR-Bg III-фрагмента в область множественного клонирования pEMBL9 (Dente L. и др., см. выше) в соответствующие Sma I - Bam HI-сайты, в в результате чего получали плазмиду pTA12.
Для создания Ncol-сайта возле области процессинга гена ftf (положение нуклеотида 783 (Shiroza T. & Kuramitsu H.K., см. выше) осуществляли сайт-направленный мутагенез, как описано Kramer W. и др. (см. выше) с использованием следующего олигонуклеотида:
5'-GGCTCTCTTCTGTTCCATGGCAGATGAACC-3'
в результате чего получали плазмиду pTD2.
В положении аминокислоты +1 (положение нуклеотида 783) по отношению к области процессинга, глутамин заменялся на метионин. Ncol-PstI-фрагмент, в котором присутствует последовательность, кодирующая зрелый белок фруктозилтрансферазы, использовали для последующего клонирования.
Из плазмиды pCB50 (описанный в примере 1), для последующего клонирования использовали Hind III-Ncol-фрагмент, в котором присутствует последовательность, кодирующая часть белка карбоксипептидазы Y, ответственную за доставку в вакуоль.
Последовательность CpV-гена, ответственную за доставку в вакуоль, легировали со зрелой последовательностью гена фруктозилтрансферазы, используя технику трехточечного лигирования (Sambrook J. и др., см. выше). Нижеследующие фрагменты были использованы в эквимолярных концентрациях: Hind III-Ncol-фрагмент cpv-гена (кодирующего последовательность, ответственную за доставку в вакуоль); Ncol-PstI-фрагмент ftf-гена; и фрагмент, содержащий вектор pEMB 8, который был переварен ферментами Hind III и PstI. Полученная в результате плазмида pTKI кодирует гибридную конструкцию, образованную путем лигирования с сохранением рамки считывания гена карбоксипептидазы Y и фруктозилтрансферазы. Правильность рамки считывания гибридного гена была подтверждена анализом последовательности.
Плазмиду pPA2, описанную в примере 1, переваривали ферментами Ncol и Bam HI для удаления гена uidA. Полученный вектор обрабатывали SI-нуклеазой для удаления выступающих концов, а затем дефосфорилировали. Из плазмиды pTKI путем переваривания ферментами AccI и PstI выделяли фрагмент, содержащий cpv-ftf-гибрид. AccI- и PstI-сайты затупляли фрагментом Кленова. Полученные фрагменты лигировали и получали плазмиду pTM4, которая содержала конструкцию 35S-промотор/ cpv/ftf-гибридный ген/ Nos-терминатор.
Плазмиду pTM4 сначала переваривали ферментом XhoI, а затем частично переваривали ферментом XbaI. Фрагмент, содержащий полную конструкцию (35S-cpy-ftf-Nos), клонировали в XbaI и Hind III-сайт плазмиды pMOG23 (Symons и др. , см. выше), представляющей собой производное растительного бинарного вектора pBIN19 (Bevan M. , см. выше), в результате чего получали плазмиду pTP.
Трансгенные растения, именуемые TP-растениями, которые содержат вышеописанную конструкцию, генерировали как описано в примере 1.
3. Анализ TP-растений
Скриннинг трансформантов с использованием TCX-метода, описанного в примере 1, выявил аккумуляцию фруктана с нормальными отклонениями, обусловленными эффектом положения.
Аккумуляцию фруктанов в этих растениях, кроме того, исследовали путем выделения больших количеств фруктана (Livingstone III DA., см. выше). Этот фруктан анализировали путем определения его размера на ЖХБР-колонке с Superose 6HP 10/30 (Pharmacia), в результате чего фруктаны были обнаружены в свободном объеме колонки, что указывало на степень полимеризации > 25000 фруктозных звеньев. Аналогичным образом, фруктан, продуцируемый фруктозилтрансферазой Streptococcus mutans, элюировался в свободном объеме Superose-колонки. Частичная деградация посредством кислотного гидролиза показала характерную картину распределения продуктов гидролиза. Очищенные растительные и бактерильные фруктаны обнаруживали идентичную картину деградации на ТСХ. Полный кислотный гидролиз и последующий ВЭЖХ-анализ на Aminex HPX87C-колонке при 85oC с использованием воды в качестве элюента показали, что указанный фруктан состоит из фруктозы.
В этом эксперименте, как и в примере 1, растения, даже после их физиологического старения содержали фруктаны.
Семена также оказались нормальными, т.е., фертильными, а способность к продуцированию фруктана наследовалась последующей генерацией.
Пример 3.
Экспрессия гена sacB в апопласте
1. Отбор генов
Для генерирования фруктанпродуцирующих растений проводили отбор генов, кодирующих белки, способные продуцировать фруктаны. Для этого был использован один из этих генов, а именно ген sacB, кодирующий левансахаразу (Steinmetz M и др., см. выше), и описанный в примере 1.
Поскольку в растениях табака сахароза транспортируется во флоэму через апопласт (межклеточное пространство), то апопласт также является предпочтительным местом накопления фруктанов. Поэтому был выбран ген, кодирующий белок, способный к самодоставке в апопласт, а именно ген pr-s, кодирующий белок s, ассоциированный с патогенезом (Coruelissen B.J.C. и др., Nature 321, 531-532 (1986). От этого гена использовали последовательность "экспорт"-сигнала.
2. Конструирование 35S/pr-s-sacB/Nos в бинарном векторе (pKT)
Плазмида pMOG29 была описана Pen J. и др. (Bio/Technology 10, 292-296 (1992)). Ген (pr-s) белка, связанного с патогенезом (Cornelissen B.J.C. и др. , см. выше; Van kan S.A.L. и др. Plant Mol. Biol. 12, 153-155 (1989)), имеет сигнальную последовательность, благоприятствующую целевой доставке белков во внеклеточное пространство (апопласт) (Pen J. и др., см. выше). Плазмида pMOG29 содержит конструкцию "35S-промотор/pr-s последовательность /Nos-терминатор". Для последующего клонирования использовали pBluescript 11 SK (Stratagene), из которого был удален внутренний Nam HI-сайт путем разрезания ферментом Bam H1 с последующим затуплением липких концов фрагментом Кленова и лигированием.
Конструкцию "35S-промотор/pr-s-последовательность/Nos-терминатор", полученную путем EcoRI- и Hind III- переваривания плазмиды pMOG29, клонировали в указанный pBluescript и соответствующий EcoRI и Hind III-сайт, в результате чего получали плазмиду pPBI. Эту плазмиду переваривали ферментом Bam HI, а липкие концы затупляли фрагментом Кленова. Из плазмиды pKK3 (пример 1) выделяли ген acB путем переваривания ферментами Ncol и PstI. Ncol- и PstI-сайты затупляли фрагментом Кленова. После лигирования по тупым концам получали плазмиду pKP. Плазмида pKP содержит встроенный с сохранением рамки считывания гибридный pr-s-ген левансахаразы. Лигирование с сохранением рамки считывания было подтверждено анализом последовательности. Указанная конструкция кодирует аминокислотную последовательность:
MNFLKSFPFYAFLCFGQYFVAVTHARAS,
за которой следует последовательность белка левансахаразы, начиная с метионина, аланина и остатка зрелой последовательностью левансахаразы (Steinmetz M. и др., см выше). Эту конструкцию переваривали ферментом XbaI, а затем частично переваривали ферментом XhoI. Фрагмент, содержащий полную конструкцию (35 S/pr-s-sacB/Nos), клонировали в XbaI- и XhoI-сайты плазмиды pMOG23 (Symons и др. , см. выше), представляющей собой производное растительного бинарного вектора pBIN19 (Bevan M., см. выше), в результате чего получали плазмиду pKT.
Трансгенные растения, именуемые KT-растениями и содержащие вышеописанную конструкцию, генерировали, как описано в примере 1.
3. Анализ KT-растений
Скриннинг трансформантов с использованием вышеуказанного ТСХ-метода, описанного в примере 1, выявил аккумуляцию фруктана в этих растениях с нормальными отклонениями, обусловленными эффектом положения.
Аккумуляцию фруктанов в этих растениях, кроме того, исследовали путем выделения фруктана в более крупных количествах (Hivingstone 111 D.A. см. выше). Результаты, полученные с помощью хроматографии на суперозе и полного кислотного гидролиза с последующим ЖХБР-анализом, показали, что вышеописанный фруктан является высокомолекулярным соединением и состоит из фруктозы.
В этом эксперименте, как и в примере 1, растения, после их физиологического старения, содержали фруктаны. Аналогичным образом семена этих растений были фертильными, а способность к продуцированию фруктана наследовалась последующей генерацией.
Пример 4.
Экспрессия гена ftf в апопласте
1. Отбор генов
Для генерирования фруктанпродуцирующих растений проводили отбор генов, кодирующих белки, способные продуцировать фруктаны. Для этого был использован другой ген, а именно ген ftf, кодирующий фруктозилтрансферазу (Shiroza T. & Kuramitsu H.K. и др., см. выше), и описанный в примере 2. Поскольку в растениях табака, сахароза транспортируется во флоэму через апопласт, то апопласт также является предпочтительным
местом накопления фруктанов. Поэтому был выбран ген, кодирующий белок, способный к самодоставке в апопласт, а именно ген pr-s (Cornelissen B.J.C. и др. , см. выше), кодирующий белок S, ассоциированный с патогенезом. От этого гена использовали сигнальную последовательность.
2. Конструирование 35S/pr-s-ftf/Nos в бинарном векторе (pTT)
Плазмиду pPPBI (пример 3) переваривали ферментом Bam HI, а липкие концы затупляли фрагментом Кленова. Из плазмиды pTKI (пример 2) выделяли ген ftf путем переваривания ферментами Ncol и PstI. Указанные рестрикционные сайты затупляли фрагментом Кленова и после лигирования по тупым концам получали плазмиду pTP. Плазмида pTP содержит встроенную с сохранением рамки считывания гибридную конструкцию pr-s-ftf. Лигирование с сохранением рамки считывания было подтверждено анализом последовательности. Указанная конструкция кодирует аминокислотную последовательность:
MNFLKSFPFYAFLCFGQYEVAVTHARAS
и последовательность белка фруктозилтрансферазы, начинающуюся с метионина зрелой фруктозилтрансферазы (Shiroza T., см. выше).
Эту конструкцию переваривали ферментами XbaI и Hind 111. Фрагмент, содержащий полную конструкцию (35S/pr-s-ftf/Nos), клонировали в XbaI Hind 111-сайты плазмиды pMOG23 (Symons и др., см. выше), представляющей собой производное растительного бинарного вектора pBIN19 (Bevan M. см. выше), в результате чего получали плазмиду pTT.
Трансгенные растения, именуемые TT-растениями и содержащие вышеописанную конструкцию, генерировали как описано в примере 1.
3. Анализ TT-растений
Скриннинг трансформантов с использованием TCX-метода описанного в примере 1, выявил аккумуляцию фруктана с нормальными отклонениями, обусловленными эффектом положения.
Аккумуляцию фруктанов в этих растениях, кроме того, исследовали путем выделения больших количеств фруктана (Livingstone III D.A. см. выше). Результаты, полученные с помощью хроматографии на суперозе и полного кислотного гидролиза с последующим ЖХБР-анализом, показали, что указанный фруктан является высокомолекулярным соединением и состоит из фруктозы.
Как и в примере 1, растения после их физиологического старения содержали фруктаны. Аналогичным образом, семена этих растений были фертильными, а способность к продуцированию фруктана наследовалась последующей генерацией.
Пример 5
Экспрессия sacB в цитоплазме
1. Отбор генов.
Для генерирования фруктанпродуцирующих растений проводили отбор генов, кодирующих белки, способные продуцировать фруктаны. Для этого был выбран один из генов, а именно ген acB, кодирующий левансахаразу (Steinmetz M. и др., см. выше) и описанный в примере 1.
Поскольку в растительных клетках сахароза синтезируется в цитоплазме, то цитоплазма также является предпочтительным местом накопления фруктанов. Так как ядерные белки продуцируются в цитоплазме, то в последовательности-переносчике нет необходимости.
2. Конструирование 35S-sacB-Noc в бинарном векторе (pKz)
Плазмиду pPA2 (пример 1) переваривали ферментами Ncol и Bam HI и векторсодержащий фрагмент выделяли. Из плазмиды pKK3 (пример 1) выделяли ген sacB в виде Ncol-Bam HI-фрагмента. Оба эти фрагмента лигировали, в результате чего получали плазмиду pKX. Эта плазмида кодирует конструкцию зрелой левансахаразы. Правильность полученной конструкции была подтверждена анализом последовательности.
Эту конструкцию переваривали ферментами XbaI и XhoI. Фрагмент, содержащий полную конструкцию (35S-sacB-Noc), клонировали в XbaI- и XhoI-сайты плазмиды pMOG23 (Symons и др., см. выше), представляющей собой производное растительного бинарного вектора pBIN19 (Bevan M. см. выше), в результате чего получали плазмиду pKz.
Трансгенные растения, именуемые KT-растениями и содержащие вышеописанную конструкцию, генерировали как описано в примере 1.
3. Анализ KT-растений
Скриннинг трансформантов с использованием TCX-метода, описанного в примере 1, выявил аккумуляцию фруктана с нормальными отклонениями, обусловленными эффектом положения.
Аккумуляцию фруктанов в этих растениях, кроме того, исследовали путем выделения больших количеств фруктана (Livingstone 111 D.A. см. выше). Результаты, полученные с помощью хроматографии на суперозе и полного кислотного гидролиза с последующим ЖХБР-анализом, показали, что указанный фруктан является высокомолекулярным соединением и состоит из фруктозы.
Как и в примере 1, растения даже после их физиологического старения содержали фруктаны. Аналогично семена этих растений были фертильными, а способность к продуцированию фруктана наследовалась последующей генерацией.
Пример 6
Экспрессия гена ftf в цитоплазме
1. Отбор генов
Для генерирования фруктанпродуцирующих растений проводили отбор генов, кодирующих белки, способные продуцировать фруктаны. Для этого был использован другой ген, а именно ген ftf, кодирующий фруктозилтрансферазу (Shiraza T. & Kuramitsu H.K., см. выше) и описанный в примере 2. Поскольку в растительных клетках сахароза синтезируется в цитоплазме, то цитоплазма является предпочтительным местом накопления фруктанов. Так как ядерные белки продуцируются в цитоплазме, то в последовательности-переносчике нет необходимости.
2. Конструирование 35S-ftf-Nos в бинарном векторе (pTz)
Плазмиду pPA2 (пример 1) переваривали ферментами Ncol и Bam HI и векторсодержащий фрагмент выделяли. Из плазмиды pTK1 (пример 2) выделяли ген ftf в виде Ncol-Bam HI-фрагмента. Затем оба фрагмента лигировали и получали плазмиду pTX. Эта плазмида кодирует конструкцию зрелой фруктозилтрансферазы. Правильность полученной конструкции была подтверждена анализом последовательности.
Эту конструкцию переваривали ферментами Xba I и Hind III. Фрагмент, содержащий полную конструкцию (35S-ftf-Nos), клонировали в VbaI и Hind 111-сайты плазмиды pMOG23 (Symons и др., см. выше), представляющей собой производное растительного бинарного вектора pBIN19 (Bevan M. см. выше), в результате чего получали плазмиду pTz.
Трансгенные растения, именуемые Tz-растениями и содержащие вышеописанную конструкцию, генерировали как описано в примере 1.
Анализ Tz-растений
Скриннинг трансформантов с использованием TCX-метода, описанного в примере 1, показал аккумуляцию фруктана с нормальными отклонениями, обусловленными эффектом положения.
Аккумуляцию фруктанов в этих растениях, кроме того, исследовали путем выделения больших количеств фруктана (Livingstone 111 D.A. см. выше). Результаты, полученные с помощью хроматографии на суперозе и полного кислотного гидролиза с последующим ЖХБР-анализом, показали, что указанный фруктан является высокомолекулярным соединением и состоит из фруктозы.
Как и в примере 1, даже после физиологического старения растения содержали фруктаны. Аналогично семена этих растений были фертильными, а способность к продуцированию фруктана наследовалась последующим поколением.
Пример 7
Приобретенные свойства трансгенных растений табака, несущих конструкции фруктозилтрансферазы
1. Общие свойства
Все трансгенные линии растений табака, несущие различные конструкции фруктозилтрансферазы и полученные как описано в примерах 1 - 6, аккумулируют молекулы фруктана. Уровни аккумуляции фруктана отличаются для отдельных трансформантов, что, очевидно, обусловлено влиянием сайта интеграции трансгена на генную экспрессию (эффект положения), и, кроме того, они отличаются для шести различных использованных конструкций. С помощью гибридизационного Саузерн-анализа (Sambrook и др., см. выше) определяли число ДНК-копий, интегрированных в отдельные геномы растений. Число интегрированных копий варьировалось от 1 до 8, но большинство растений содержало лишь одну или две копии данной конструкции. Степень полимеризации фруктанов составляла до 20000 фруктозных звеньев или более. Фруктаны накапливались во всех исследуемых органах, включая листья, стебли, корни и семена. Это свойство наследовалось потомством в соответствии с концепцией Менделя.
Идентичность фруктанов подтверждалась анализами, проводимыми путем определения размеров фруктанов, а также с помощью гидролиза и протонной ЯМР-спектроскопии, как было описано выше. Кроме того, были проведены исследования, в которых антитела, направленные против фруктофуранозильных связей (HaII B. и др., 1990, Mol. Immunology 27: 351 - 361; Hall B и др., 1992, The Journal Immunology 149: 1605 - 1612), реагировали с фруктанами, продуцированными в трансгенных растениях. Для этих исследований фруктаны наносили пятнами на нитроцеллюлозную фильтровальную бумагу и иммобилизовали путем 15-минутной обработки при 120oC. Фильтры инкубировали со специфическим мышиным антителом (Hall и др., см. выше), и связывание антител с фруктаном обнаруживали с помощью щелочной фосфатазы, конъюгированной со вторым козьим антителом против неспецифических частей мышиного антитела.
Фруктаны присутствуют в растениях на протяжении всего их жизненного цикла. После синтезирования фруктан остается стабильным. Если и существует метаболизм фруктана, то его уровень очень невысок. Фруктан обнаруживается даже в листьях растений на стадии увядания, причем в количестве, сравнимом с его количеством в зрелых листьях. Содержание фруктана в трансгенных растениях табака составляет порядка 3 - 8% по сухой массе зрелых листьев. При этом с помощью ЖХБР-хроматографии, описанной выше, было установлено, что уровни других растворимых углеводов (глюкозы, сахарозы и фруктозы) сравнимы с их уровнями в нетрансформированных растениях дикого типа.
2. Характеризация фенотипа
В условиях нормального роста не наблюдалось какого-либо различия в росте или морфологии между нетрансформированными контрольными растениями табака и фруктан-аккумулирующими трансгенными растениями, несущими ftf-ген S.mutant в трех различных конфигурациях, упомянутых в примерах 2, 4 и 6 (TP, Tz и TT-линии). Это справедливо также для растений, несущих ген sacB B.subtilis, присоединенный к гену cpy, несущему сигнал доставки (KP-растения, пример 1). Растения, экспрессирующие sacB -ген B.subtilis в цитоплазме или апопласте (Kz, KT, примеры 3 и 5), обнаруживали некротические повреждения зрелых листьев и стеблей. Строгость фенотипа коррелирует с уровнем аккумулированных фруктанов. Указанные растения способны к продуцированию фертильных семян.
Пример 8
Повышенная продуктивность трансгенных растений табака, несущих фруктозилтрансферазные конструкции, в условиях стресса.
Продуктивность в условиях стресса характерных фруктан-аккумулирующих растений табака, несущих cpy-sacB-конструкцию (KP, пример 1), сравнивали с продуктивностью нетрансформированных контрольных растений.
1. Стресс, вызываемый засухой
Стресс, вызываемый засухой, индуцировали в серии растений путем добавления к растениям, выращенным на вермикулите, полиэтиленгликоля со средней молекулярной массой 10000 Дальтон (ПЭГ 10000). Диапазон уровней указанного стресса индуцировали с использованием ПЭГ-растворов, содержащих до 20% ПЭГ 10000. Кроме того, определяли массу свежей ткани, массу сухой ткани растений, и темпа роста (см/день). Фруктан-аккумулирующие KP-растения, подвергнутые стрессу, вызываемому засухой, росли быстрее и давали значительно более высокие выходы, чем обычные нетрансформированные растения в аналогичных условиях стресса. По сравнению с нетрансформированными растениями, подверженными стрессовым условиям, трансгенные KP-растения имели, в типичном случае, повышенную массу в свежем состоянии, составляющую до 19%, и сухую массу, составляющую до 32%.
2. Стрессовые условия, вызываемые световым освещением
Для характерных трансгенных KP-растений и нетрансформированных растений, содержащихся в субоптимальных условиях освещения, проводили сравнения показателей выхода. KP-растения, выращенные в условиях низкого освещения (5000 люкс), непосредственно сравнивали с контрольными (нетрансформированными) растениями. Фруктан-аккумулирующие KP-растения росли быстрее и давали значительно более высокие выходы, чем нетрансформированные растения. Массы KP-растений в свежем виде и в сухом виде на 18% превышали соответствующие массы нетрансформированных растений. KP-растения имели больше листьев, чем нетрансформированные растения.
Высокие интенсивности излучения, особенно, в сочетании с субоптимальными условиями культивирования, также вызывали глубокий стресс у большинства видов растений. Производительность фруктан-аккумулирующих растений в этих условиях также превышает производительность нетрансформированных растений в аналогичных условиях. Таким образом, фруктаны играют роль дополнительных запасов углеводов и тем самым защищают растение от повреждений, вызываемых световым стрессом.
3. Стресс, вызываемый холодом
Показатели роста трансгенных KP-растений и нетрансформированных растений сравнивали в условиях стресса, вызываемого холодом. При культивировании в контролируемых условиях при 12oC выходы KP-растений, определенные исходя из их сырой и сухой массы, на 19% превышали выходы нетрансформированных растений.
Стрессовые условия, указанные в пп. 1), 2) и 3), способствовали более чем 4-кратному увеличению уровня фруктана в трансгенных KP-растениях по сравнению с уровнями фруктазы в KP-растениях, не подвергавшихся стрессу. Накопление фруктана в растениях приводит к более высоким показателям их роста; причем степень накопления фруктана коррелирует со степенью усиления роста.
Более высокая продуктивность в стрессовых условиях не ограничивается только конкретными примерами 1), 2) и 3) и может иметь место в широком диапазоне неблагоприятных условий окружающей среды. Кроме того, повышенная производительность растений не ограничивается лишь усиленным темпом их роста или увеличением сырой или сухой массы; трансгенные растения могут иметь улучшенные свойства и в отношении других экономически важных аспектов их жизнедеятельности как до сбора, так и после сбора урожая.
Помимо улучшенных свойств в условиях абиотических стрессов, упомянутых выше, фруктан-аккумулирующие растения могут также обладать повышенной резистентностью к биологическим стрессам, например к заболеваниям и поражению вредителями (Farrar, b "Pests and pathogens: plant response to foliar attack", p. 107 - 127, Ed. P. G. Ayres, BIOS Publisher 1992). Накопление фруктана может приводить к метаболическим или структурным модификациям в данном растении и тем самым способствовать снижению его восприимчивости к заболеваниям и поражениям вредителями. Иллюстрирующими, но не ограничивающими примерами таких модификаций могут служить изменения в усвояемости, строении ткани, и вкусовых качеств растений, а также изменения уровней других первичных и вторичных метаболических продуктов растения.
Растения с естественно или искусственно модифицированными типами распределения углеводов могут быть предпочтительными объектами для работы по дальнейшему улучшению их свойств в стрессовых условиях, например, для еще большего увеличения накопления фруктана в их тканях, иллюстрирующими, но не ограничивающими примерами таких растений могут служить натуральные мутанты в отношении метаболизма крахмала и сахарозы, а также растения, в которых метаболизм крахмала и сахарозы был модифицирован с помощью молекулярно-генетической техники, описанной, напр. Sonnewald & Willmitzer в Plant Physiology 99, 1267-1270, 1992.
Пример 9
Общее применение описанной технологии
1. Конструкции
Для иллюстрации общей применимости используемой технологии были изготовлены cpy-sacB конструкции, описанные в примере 1. При этом могут быть использованы другие гены любого происхождения, кодирующие полипептиды, которые способны синтезировать фруктан, и которые состоят, по крайней мере, из двух связанных между собой фруктозных звеньев. Кроме того, эффективность вводимых конструкций, выражающаяся в способности трансгенных растений накапливать фруктан в соответствующих количествах, в определенной период времени, а также в соответствующих органах или тканях, может быть модифицирована путем использования регуляторных сигнальных последовательностей, иллюстрирующими, но не ограничивающими примерами которых могут служить конститутивные промоторы, органоспецифические промоторы, промоторы с временной регуляцией, сигналы полиаденилирования, трансляционные энхансеры, транскрипционные энхансеры и т.п. Кроме того, локализация полипептида в клетке может регулироваться использованием сигнальных элементов, ответственных за целевую доставку полипептида в тот или иной отдел клетки, например, сигнальных элементов любого происхождения, ответственных за направленную доставку в вакуоль или апопласт. После введения в растения любой комбинации из вышеуказанных элементов значительные количества фруктанов могут накапливаться в вегетативных органах растений, таких, как листья, корни и стебли, а также в репродуктивных органах, таких, как клубни, корнеплоды, плоды и семена.
2. Применение в различных видах растений
Для иллюстрации общей применимости используемой технологии, конструкцию cpy-sacB, описанную в примере 1, вводили в культурные растения, например картофель (Solanum tuberosum L), которые могут быть, но необязательно, трансформированы в соответствии с описанием Visser (Plant Culture Manual B5, 1-9, Kluwer Academic Publishers, 1991). Полученные трансгенные растения накапливали фруктаны в каждом органе на протяжении всего цикла своего развития. Уровни фруктана в листьях и клубнях могут составлять выше 9% по сырой массе. Кроме того, аналогичную конструкцию вводили в свеклу (Beta vulgaris L.), которая может быть, но необязательно, трансформирована в соответствии с описанием D'Halluim etal, Biotechnology 10, 309 - 314 (1992). Полученные трансгенные растения свеклы накапливали значительные количества фруктана со степенью полимеризации до 25000 или более в своих листьях и корнеплодах. Ту же самую cpy-sacB конструкцию вводили в брюкву (Brassica napis L.), которая может быть, но необязательно трансформирована в соответствии с описанием Block и др., (Plant Physiol. 91, 674 - 701 (1989)). Полученные трансгенные растения накапливали, например, в своих листьях и запасающих органах значительные количества фруктана со степенью полимеризации до 25000 или более.
Примерами других видов растений, которые могут быть модифицированы в целях сообщения им способности накапливать фруктаны, могут служить кукуруза (Lea may L. ), пшеница (Triticum aestivum L.) ячмень (Hordeum vulgare L.), рис (Oryza sativa L.), соя (Glucin max L.), горох (Pisum sativum L.), фасоль (Phascolus vulgaric L.), цикорий (Cichorium intybus L.), сахарный тростник (Saccharum officinarum L.), ямс съедобный (Dioscorea esculenta L.), маниок съедобный (кассава) (Manihot esculenta L.) и травы (например, Lolium spp. (плевел), Poa spp. (мятлик) и Festuca spp. (овсянница)).
Растения с естественно или искусственно модифицированными типами распределения углеводов могут быть предпочтительными объектами для модификации метаболизма фруктана, особенно, в отношении повышения уровней накопления фруктана. Иллюстрирующими, но не ограничивающими примерами таких растений являются натуральные мутанты, в которых имеются изменения в метаболизме крахмала и сахарозы, а также растения, в которых метаболизм крахмала и сахарозы был модифицирован с помощью молекулярно-генетической техники, описанной, например, Sonnewald & Willmitzer b "Plant Phusiology 99, 1267-1270, 1992.
Изобретение относится к сельскохозяйственной и пищевой промышленности. ДНК-конструкции гена фруктозилтрансферазы вводятся в геном растения и тем самым модифицируют характер распределения в них фруктанов. Трансгенные растения получают регенерацией из трансформированной растительной клетки. Полученные растения обладают повышенной продуктивностью по сравнению с нетрансформированными при выращивании в стрессовых условиях - при засухе, недостатке освещения, пониженных температурах - за счет увеличенного содержания фруктанов. ДНК-конструкции позволяют получать растения с фруктанами, имеющими более высокую степень полимеризации, чем обычно. Такие фруктаны могут быть более широко использованы в промышленном производстве. 7 с. и 15 з.п. ф-лы, 9 ил.
Приоритет по пунктам:
28.12.1992 по пп.1 - 19;
28.12.1993 по пп.20 - 22.
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |
| Биохимия растений.: Пер | |||
| с англ | |||
| под ред | |||
| Кретовича В.Л | |||
| - М.: Мир, 1968, с | |||
| Способ приготовления кирпичей для футеровки печей, служащих для получения сернистого натрия из серно-натриевой соли | 1921 |
|
SU154A1 |
