Настоящее изобретение относится к конструкциям ДНК и растениям, их включающим. В частности, оно относится к промоторным последовательностям для экспрессии генов, которые придают растениям гербицидную устойчивость.
Недавние успехи в биотехнологии растений привели к созданию трансгенных растений, устойчивых к применяемым гербицидам. Гербицидная толерантность достигается использованием ряда различных трансгенных методов. Одним из хорошо документированных примеров является использование бактериального ксенобиотического детоксицирующего гена фосфинотрицинацетилтрансферазы (PAT) из Streptomyces hydroscopicus. Мутантные гены растительного происхождения, например измененный генный сайт - мишень, кодирующий ацетолактатсинтазу (ALS) из Arabidopsis, успешно использовались для образования трансгенных растений, устойчивых к применяемым гербицидам. Гены PAT и ALS были экспрессированы под контролем сильного конститутивного промотора.
Мы предлагаем систему, где гены, придающие гербицидную толерантность, экспрессировались бы индуцируемым способом, в зависимости от применения конкретного активирующего химического реагента. Этот подход обладает множеством выгод для фермера, включая нижеследующие:
1. Индуцируемый контроль за гербицидной толерантностью снизил бы любой отрицательный риск, связанный с высокими уровнями конститутивной экспрессии генов гербицидной устойчивости. Это может представлять отдельную проблему на ранних стадиях роста, когда высокие уровни трансгенного продукта могут непосредственно мешать нормальному развитию. Альтернативно, высокие уровни экспрессии генов гербицидной устойчивости могут вызывать метаболическое истощение ресурсов растения.
2. Экспрессия генов гербицидной устойчивости индуцируемым способом позволяет использовать определенный гербицид, о котором идет речь, для контроля за самосевами, если активирующий химический реагент исключили во время обработки.
3. Применение индуцируемого промотора для управления генами устойчивости к гербицидам будет снижать соответствующую опасность устойчивости сорных видов, становящихся большой проблемой. Если гены устойчивости попали в сорные виды из близких сельхозкультур, контроль можно было бы осуществлять с помощью соответствующего гербицида, в отсутствие индуцирующего химического реагента. Это особенно важно, если ген толерантности подкрепляет устойчивость при тотальном вегетативном контроле, гербицида, который использовали бы (без индуцирующего химического реагента) перед высевом соответствующей сельхозкультуры и, по возможности, после сбора урожая этой сельхозкультуры.
4. Например, можно представить себе, что гербицидная устойчивость у таких зерновых как пшеница, могущая скрещиваться с сорными дикими овсами, или что гербицидная устойчивость у масличной культуры рапса или канны может быть перенесена в дикие крестоцветные, придавая таким образом гербицидную устойчивость этим причиняющим беспокойство сорнякам. Еще одним примером является то, что индуцируемая экспрессия гербицидной устойчивости у сахарной свеклы будет снижать опасность дичания сахарной свеклы, что тоже становится проблемой.
Известно несколько систем регуляции гена (генные переключатели), используемых для придания индуцируемой гербицидной устойчивости у растений. Много таких генных переключателей описано в обзоре у Gatz (Current Opinion in Biotechnology (1996) 7, 168-172) и включает такие системы, как ген-переключатель тетрациклинового репрессора, система Lac-репрессора, индуцируемые медью системы, такие, которые основаны на АСЕ 1, промоторы, индуцируемые салициловой кислотой, включающие PR-la-систему и систему, основанную на стероидных гормонах, таких как глюкокортикоидная, прогестероновая и эстрогенная рецепторные системы. Модификации глюкокортикоидной рецепторной системы, которые включают GAL-4-связывающий домен дрожжей и VP-16-активатор, описаны у Aoyama с соавт. The Plant Cell, (1995) 7, 1773-1785 и предусматривают создание аналогичных систем, основанных, например, более на стероидных гормонах насекомых, чем на стероидных гормонах млекопитающих. Действительно, недавно описана система, основанная на экдизоновом рецепторе Hеliothis virescens. Известны также системы генной регуляции бензолсульфонамида (Hershey с соaвт. Plant Mol. Biol., 17, 679-690 (1991), которые основаны на alcR-белке из Asper-gillus nidulans и промоторах глутатион-S-трансферазы.
Известно также несколько генов, которые придают гербицидную устойчивость, например один из гербицидов, для которого описаны гены устойчивости и который используется необычайно широко, представляет собой N-фосфонометилглицин (глифосат), а его приемлемые для сельского хозяйства соли включают изопропиламиновые, триметилсульфониевые, натриевые, калиевые и аммониевые соли.
Первая цель настоящего изобретения заключается в создании химически индуцируемой генной экспрессионной кассеты, включающей индуцируемый промотор, присоединенный путем сшивки к гену-мишени, который придает устойчивость к гербициду.
Можно использовать любой ген гербицидной устойчивости, но гены, которые устойчивы к N-фосфонометилглицину или его солям или производным, представляются особенно предпочтительными.
Для придания гербицидной устойчивости может быть использовано несколько индуцируемых промоторов, и они включают любой из вышеперечисленных промоторов.
Однако для использования в этой области особенно полезен генный переключатель, основанный на регуляторном белке alc R из Aspergillus nidulans, который активирует экспрессию генов под alcA-промотором в присутствии некоторых спиртов и кетонов. Эта система описана в нашей Международной патентной публикации W093/21334, которая включена здесь путем ссылки.
Описана также система активации гена alcA/alcR из грибов Aspergillus nidulans. Путь утилизации этанола у А. nidulans ответствен за распад спиртов и альдегидов. Было установлено, что в метаболическом пути этанола участвуют три гена. Было показано, что гены аlсА и alcR тесно расположены в группе сцепления VII, a aIdA картирован в группе сцепления VIII (Pateman JH с соавт. , 1984, Proc. Soc. Lond, B217:243-264; Sealy-Lewis HM и Lockington RA, 1984, Curr. Genet, 8:253-259). Ген alcA кодирует ADHI у A. nidulans ADHI, a aldA кодирует AldDH, этот второй фермент ответствен за утилизацию этанола. Экспрессия генов alcA и aldА индуцируется этанолом и некоторыми другими индукторами (Creaser EH с соавт, 1984, Biochemical J, 255:449-454) с помощью активатора транскрипции alcR. Ген alcR и коиндуктор ответственны за экспрессию alcA и aldA, так как ряд мутаций и делеций в alcR приводят к плейотропной утрате ADHI и aldDH (Felenbok В с соавт, 1988, Gene, 73:385-396; Pateman с соавт. , 1984; Sealy-Lewis & Lockington, 1984). Белок ALCR активирует экспрессию a-ZcA путем связывания с тремя специфическими участками в промоторе alcA (Klumberg Р с соавт., 1992, J. Biol. Chem, 267:21146-21153).
Ген alcR был клонирован (Lockington с соавт., 1985, Gene, 33:137-149) и секвенирован (Felenbok с соавт., 1988). Экспрессия гена alcR является индуцируемой, ауторегулируемой и подвержена репрессии глюкозой, опосредованной с помощью репрессора CREA (Bailey С и Arst HN, 1975, Eur. J. Biochem, 51: 573-577; Lockington RA с соавт., 1987, Mol. Microbiology, 1:275-281; Dowzer CEA и Kelly JM, 1989, Curr. Genet, 15:457-459; Dowzer CEA и Kelly JM, 1991, Mol. Cell. Biol, 11:5701-5709). Регуляторный белок ALCR содержит 6 цистеинов вблизи своего N-конца, координированных в цинковый двухядерный кластер (Klumberg Р. с соавт., 1991, FEBS Letts, 280:11-16). Этот кластер связан с высококонсервативными ДНК-связывающими доменами, обнаруженными в факторах транскрипции других аскомицет. Показано, что транскрипционные факторы GAL4 и LAC9 имеют двухядерные комплексы, которые обладают структурой типа клеверного листа, содержащие два атома Zn(II) (Pan Т и Coleman JE, 1990, Biochemistry, 29: 3023-3029; Halvorsen YDC с соавт., 1990, J. Biol.Chem, 265:13283-13289). Структура ALCR сходна с этим типом, за исключением наличия асимметричной петли из 16 остатков между Cys-3 и Cys-4. ALCR положительно активирует собственную экспрессию путем связывания с двумя специфическими участками в своей промоторной области (Kulmberg P. с соавт., 1992, Molec. Cell. Biol, 12:1932-1939).
Регуляция тремя генами, alcR, а1сА и aldA, вовлеченными в путь утилизации этанола, осуществляется на уровне транскрипции (Lockington с соавт., 1987; Gwynne D с соавт. , 1987, Gene, 51:205-216; Picket с соавт., 1987, Gene, 51:217-226).
Имеются две другие алкогольдегидрогеназы, представленные у A.nidulans. ADHII присутствует в мицелии, выращенном в неиндуцируемой среде, и репрессируемая в присутствии этанола. ADHII, кодируемая alcB, также находится под контролем alcR (Sealy-Lewis & Lockington, 1984). Третья алкогольдегидрогеназа также была клонирована путем комплементации с adh-штаммом S. cerevisiae. Этот ген alcA картирован в группе сцепления VII, но не сцеплен с а1сА и alcR. Этот ген, alcC, кодирует ADHIII и утилизирует этанол очень слабо (McKnight GL с соавт., 1985, EMBO J, 4:2094-2099). Показано, что ADHIII вовлечена в выживаемость А. nidulans в периоды анаэробного стресса. Экспрессия аlсС не репрессируется в присутствии глюкозы, означая, что он может не находиться под контролем alcR (Roland LJ и Stromer JN, 1986, Mol. Cell. Biol, 6: 3368-3372).
Итак, A. nidulans экспрессирует фермент алкогольдегидрогеназу I (ADHI), кодируемый геном а1сА только тогда, когда он растет в присутствии различных спиртов и кетонов.
Индукция передается через регуляторный белок, кодируемый геном alcR и конститутивно экспрессированный. При наличии индуктора (спирта или кетона) этот регуляторный белок активирует экспрессию гена alcA. В присутствии индуктора этот регуляторный белок стимулирует также собственную экспрессию. Это означает, что высокие уровни фермента ADHI получают в индуцируемых условиях (т.е. при наличии спирта или кетона). И наоборот, этот ген аlсА и его продукт, ADHI, не экспрессируются в отсутствие индуктора. Экспрессия аlсА и образование соответствующего фермента репрессируются также в присутствии глюкозы.
Таким образом, промотор гена alcA представляет собой индуцируемый промотор, активированный регуляторным белком alcR в присутствии индуктора (т.е. с помощью сочетания белок/спирт или белок/кетон). Гены alcR и alcA (включая соответствующие промоторы) были клонированы и секвенированы (Lockington RA. с соавт. , 1985, Gene, 33:137-149; Felenbok В. с соавт., 1988, Gene, 73: 385-396; Gwynne с соавт., 1987, Gene, 51:205-216).
Гены алкогольдегидрогеназы (adh) были исследованы у некоторых растительных видов. У кукурузы и других зерновых они переключаются анаэробными условиями. Промоторная область генов adh кукурузы содержит 300 п.н. регуляторный элемент, необходимый для экспрессии в анаэробных условиях. Однако регуляторный белок, эквивалентный alcR, не обнаружен в каком-либо растении. Следовательно, данный тип генной регуляторной системы, alcR/alcA, неизвестен для растений. Конститутивная экспрессия alcR в растительных клетках не приводит к активации эндогенной adh-активности.
В соответствии со второй целью настоящего изобретения создали химически индуцируемую генную экспрессионную кассету растения, включающую первый промотор, присоединенный путем сшивки к регуляторной последовательности alcR, которая кодирует регуляторный белок alcR, и индуцируемый промотор, присоединенный путем сшивки к гену-мишени, который придает гербицидную устойчивость, отличающуюся тем, что индуцируемый промотор активируется регуляторным белком в присутствии эффективного экзогенного индуктора, в силу чего этот применяемый индуктор вызывает экспрессию этого гена-мишени.
Индуцируемый промотор предпочтительно получали из промотора гена аlсА, или же из alcR, aldA или alcR-индуцируемых генов.
Мы обнаружили, что переключатель alcA/alcR особенно подходит для приведения в действие генов толерантности к гербициду, по крайней мере, по следующим причинам.
1. Переключатель alcA/alcR был создан, чтобы стимулировать высокие уровни экспрессии гена. Кроме того, данный регуляторный белок alcR предпочтительно находится под управлением такого сильного конститутивного промотора, как полиубиквитин. Можно достигнуть высоких уровней индуцированной трансгенной экспрессии, которые сравнимы с экспрессией под таким сильным конститутивным промотором, как 35 CaMV.
2. Если генный переключатель используется в ситуации, когда соответствующий активирующий химический реагент применяется одновременно с соответствующим гербицидом, необходимо быстрое повышение уровней гена гербицидной устойчивости. Фигура 1 показывает экспрессию генного маркера (CAT) во времени, после применения индуцирующего химического агента. Это исследование показывает быстрое увеличение (2 часа) САТ-экспрессии после лиственного применения индуцирующего химического агента. Быстрая ранняя динамика индукции является причиной экспрессии регуляторного белка конститутивным способом, поэтому время запаздывания не наблюдается, пока имеет место синтез факторов транскрипции. Кроме того, мы выбрали простую двухкомпонентную систему, которая не зависит от комплексной сигнальной системы трансдукции.
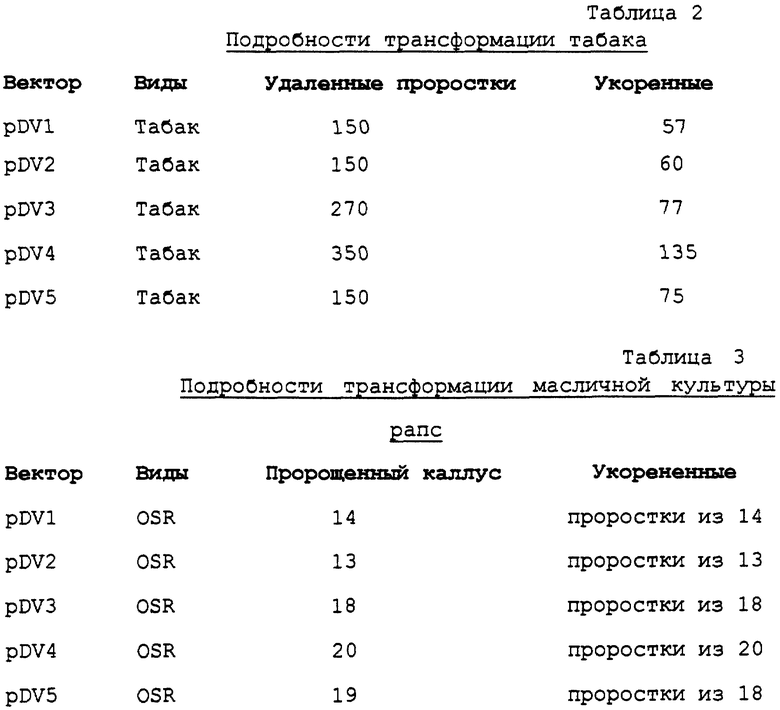
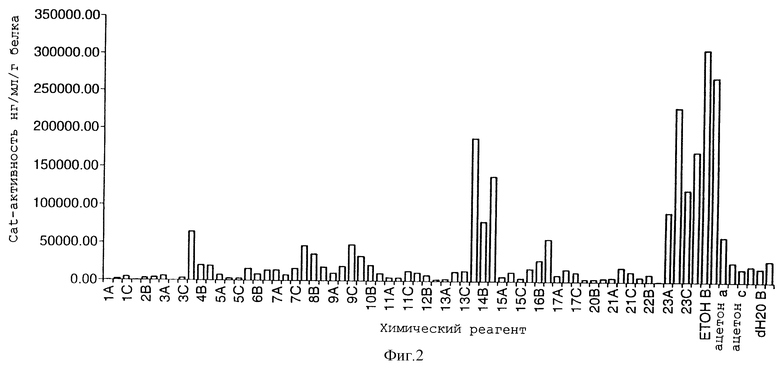
3. Мы протестировали специфичность системы alcA/alcR с по мощью ряда растворителей, используемых в агрономической практике. Гидропонная система для проращивания семян обнаружила, что этанол, бутан-2-ол и циклогексанон, все, дают высокие уровни индуцированной экспрессии репортерного гена (фигура 2). По сравнению со спиртами и кетонами, перечисленными в таблице 1, которые используются в агрономической практике в виде применяемого опрыскивания листьев, только этанол давал высокие уровни индуцированной активности репортерного гена (фигура 3).
Это представляется важным, так как неправильная индукция трансгенов не встречается при случайном выдерживании с растворителями данного списка. Этанол не является обычным компонентом агрохимических составов и поэтому соответствующую обработку опрыскиванием можно рассматривать в качестве специфического индуктора генного переключателя alcA/R для ситуации полевых условий.
4. Ряд биотических и абиотических стрессов, например патогенная инфекция, жара, холод, засуха, поранение, затопление, все они оказываются неспособными индуцировать alcA/alcR-переключение. Кроме того, ряд химических обработок нерастворителями, например салициловой кислотой, этиленом, абсцизовой кислотой, ауксином, гибберилиновой кислотой, различными агрохимикатами, все, не способны индуцировать alcA/alcR-систему.
Первый промотор может быть конститутивным или тканеспецифичным, запрограммированным на развитие или даже индуцируемым. Регуляторная последовательность, ген alcR, получена из Aspergillus nidulans и кодирует регуляторный белок alcR.
Индуцируемый промотор представляет собой предпочтительно промотор гена alcA, получаемый из Aspergillus nidulans, или "химерный" промотор, полученный из регуляторных последовательностей промотора, alcA и сердцевинной области промотора из генного промотора, который действует в растительных клетках (включая любой растительный генный промотор). Этот alcA-промотор или родственный "химерный" промотор активируется с помощью регуляторного белка alcR при применении спиртового или кетонового индуктора.
Индуцируемый промотор можно также получить из промотора гена aldAK, промотора гена аlсВ и промотора гена alcC, получаемых из Aspergillus nidilans.
Индуктор может быть любым эффективным химическим реагентом (таким как спирт или кетон). Подходящие химические реагенты для использования с alcA/alcR-полученной кассетой включают реагенты, которые перечислены у Creaser с соавт.(1984, Biochem J, 225, 449-454), такие как бутан-2-он (этилметилкетон), циклогексанон, ацетон, бутан-2-ол, 3-оксомасляная кислота, пропан-2-ол, этанол.
Генная экспрессионная кассета, ответственная за применяемый экзогенный химический индуктор, дающий возможность внешней активации экспрессии гена-мишени, регулируется с помощью данной кассеты. Указанная экспрессионная кассета является высокорегулируемой и пригодна для общего использования в растениях.
Две части указанной экспрессионной кассеты могут находиться в одном и том же конструкте или в отдельных конструктах. Первая часть включает соответствующую регуляторную кДНК или генную последовательность, субклонированную в экспрессионном векторе, с действующим растительным промотором, приводящим в действие его экспрессию. Вторая часть включает, по крайней мере, часть индуцируемого промотора, который контролирует экспрессию нижележащего гена-мишени. При наличии подходящего индуктора регуляторный белок, полученный с помощью первой части указанной кассеты, будет активировать экспрессию гена-мишени путем стимуляции индуцируемого промотора во второй части данной кассеты.
На практике данный конструкт или данные конструкты, включающие эту экспрессионную кассету настоящего изобретения, инсерцировали в растение путем трансформации. Экспрессия генов-мишеней в соответствующем конструкте, находящегося под контролем химически переключаемого промотора настоящего изобретения, может быть затем активирована в результате применения химического индуктора на соответствующее растение.
Можно использовать любой метод трансформации, подходящий для целевого растения или растительных клеток, в том числе инфицирование с помощью Agrobacterium tumefaciens, содержащей рекомбинантные Ti-плазмиды, электропорацию, микроинъецирование клеток и протопластов, трансформацию микропулями и трансформацию пыльцевой трубки. Полученные трансформированные клетки можно затем в соответствующих случаях регенерировать в целые растения, в которых этот новый ядерный материал стабильно инкорпорирован в данный геном. Этим путем могут быть получены трансформированные однодольные и двудольные растения.
Примеры генетически модифицированных растений, которые можно получить, включают полевые сельскохозяйственные культуры, зерновые, фрукты и овощи, такие как канна, подсолнечник, табак, сахарная свекла, хлопок, соя, кукуруза, пшеница, ячмень, рис, сорго, томаты, манго, персики, яблоки, груши, клубнику, бахчевые, картошку, морковь, салат, капусту, лук.
В настоящем изобретении, кроме того, создали растительную клетку, содержащую генную экспрессионную кассету, в соответствии с настоящим изобретением. Эта генная экспрессионная кассета может быть стабильно инкорпорирована в данный геном растения путем трансформации. В настоящем изобретении создали также растительную ткань или растение, включающее такие клетки и растения или семена, полученные из них.
В настоящем изобретении, кроме того, разработали способ для контроля экспрессии гена растения, включающий трансформирование растительной клетки с помощью химически индуцируемой генной экспрессионной кассеты растения, которая обладает первым промотором, присоединенным путем сшивки с регуляторной последовательностью alcR, которая кодирует регуляторный белок аlсА, и индуцируемым промотором, присоединенным путем сшивки к гену-мишени, который придает гербицидную устойчивость, отличающийся тем, что указанный индуцируемый промотор активирован с помощью регуляторного белка в присутствии эффективного экзогенного индуктора, в силу чего применяемый указанный индуктор вызывает экспрессию данного гена-мишени.
Эта стратегия индуцируемой экспрессии гербицидной устойчивости может быть осуществлена с помощью предварительного опрыскивания химическим активатором или в случае медленно действующих гербицидов, например N-фосфонометил-глицина (обычно известного в качестве глифосата), причем соответствующий химический индуктор может быть добавлен в смеси одновременно с данным гербицидом.
Эта стратегия может быть принята для любого сочетания гербицида, корреспондирующего с геном, придающим устойчивость. Например, генный переключатель alcA/alcR может быть использован с:
1. Геном глутатион-S-трансферазы (GST-27) кукурузы (смотрите нашу Международную патентную публикацию W090/08826), который придает устойчивость к хлорацетанилидным гербицидам, таким как ацетохлор, метолахлор и алахлор.
2. Фосфинотрицинацетилтрансферазой (PAT), которая придает устойчивость к данному гербициду, обычно известному в качестве глуфозината.
3. Мутантными генами ацетолактатсинтазы кукурузы (смотрите нашу Международную патентную публикацию W090/14000) и с другими генами, которые придают устойчивость к сульфонилмочевине и имидазолинонам.
4. Генами, которые придают устойчивость к глифосату. Такие гены включают ген глифосатоксиредуктазы (GOX) (смотрите Международную патентную публикацию W092/00377 от имени Monsanto Company); генами, которые кодируют синтазу 5-енолпирувил-3-фосфошикимовой кислоты (EPSPS), включающей EPSPS Класса I и Класса II, генами, которые кодируют мутантную EPSPS, и генами, которые кодируют слитые пептиды EPSPS, такие, которые включают транзитный пептид хлоропласта и EPSPS (смотрите, например, ЕР 218571, ЕР 293358, W091/04323, W092/04449 и W092/06201 от имени Monsanto Company); и генами, которые участвуют в экспрессии СРЛиазы (CPLyase).
Кроме того, мы опишем предпочтительные признаки и варианты осуществления настоящего изобретения в нижеприведенных примерах, не ограничивающимися ими, и со ссылками на соответствующие иллюстрации, в которых:
Фигура 1 иллюстрирует экспрессию маркерного гена (CAT) во времени после применения индуцирующего химического реагента;
Фигура 2 иллюстрирует соответствующие уровни экспрессии индуцированного репортерного гена в корнях, смачиваемых рядом растворителей;
Фигура 3 иллюстрирует соответствующие уровни активности индуцированного репортерного гена при опрыскивании листвы соответствующими химическими реагентами, перечисленными в Таблице 1;
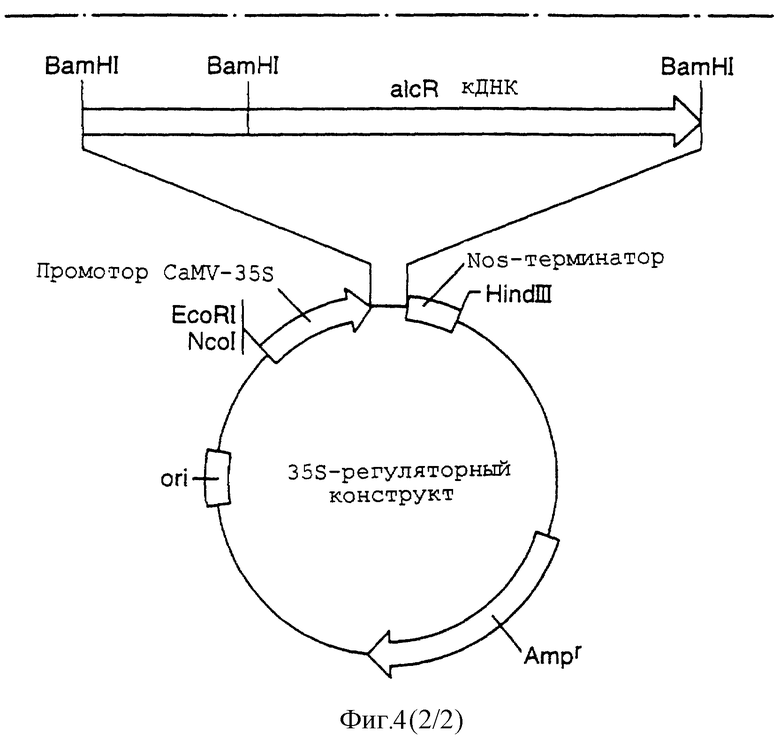
Фигура 4 иллюстрирует образование 35S-регуляторного конструкта в результате лигирования кДНК alcR no pJRl.
Фигура 5 иллюстрирует образование репортерного конструкта;
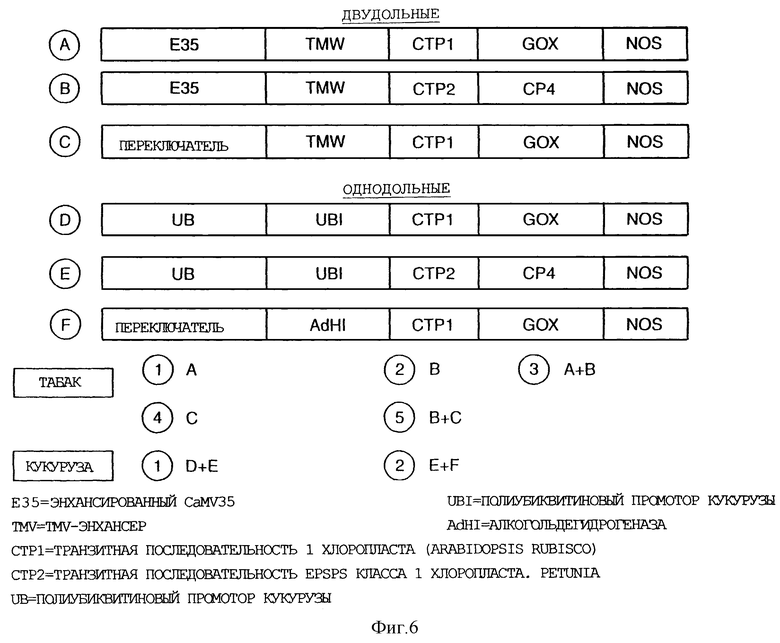
Фигура 6 представляет собой сводку кассет и конкретных конструктов, трансформирующих растение;
Фигура 7 A,B,C иллюстрирует транзитную последовательность 1 хлоропласта из Arabidopsis RUBISCO (СРТ 1) (варианты);
Фигура 8 показывает последовательность плазмиды pMJBl;
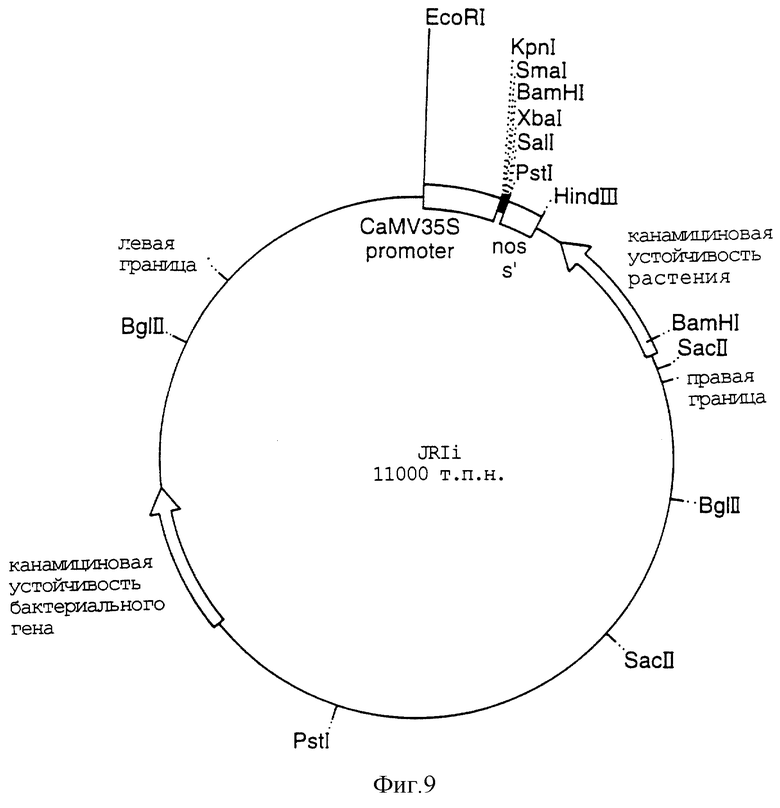
Фигура 9 представляет собой карту плазмиды pJRIi;
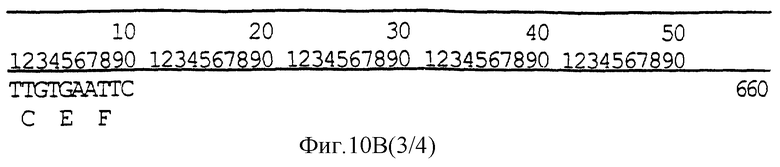
Фигура 10 (A, B, C) иллюстрирует транзитную последовательность СТР2 хлоропласта из гена EPSPS класса I из Petunia hybrida (варианты);
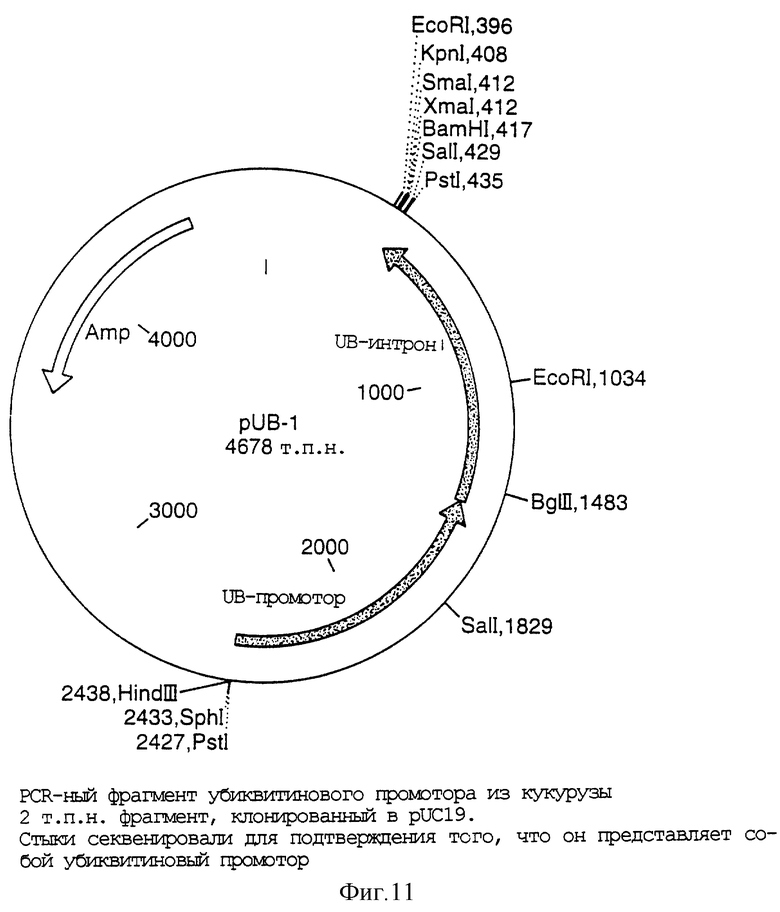
Фигура 11 представляет собой карту плазмиды pUB-1;
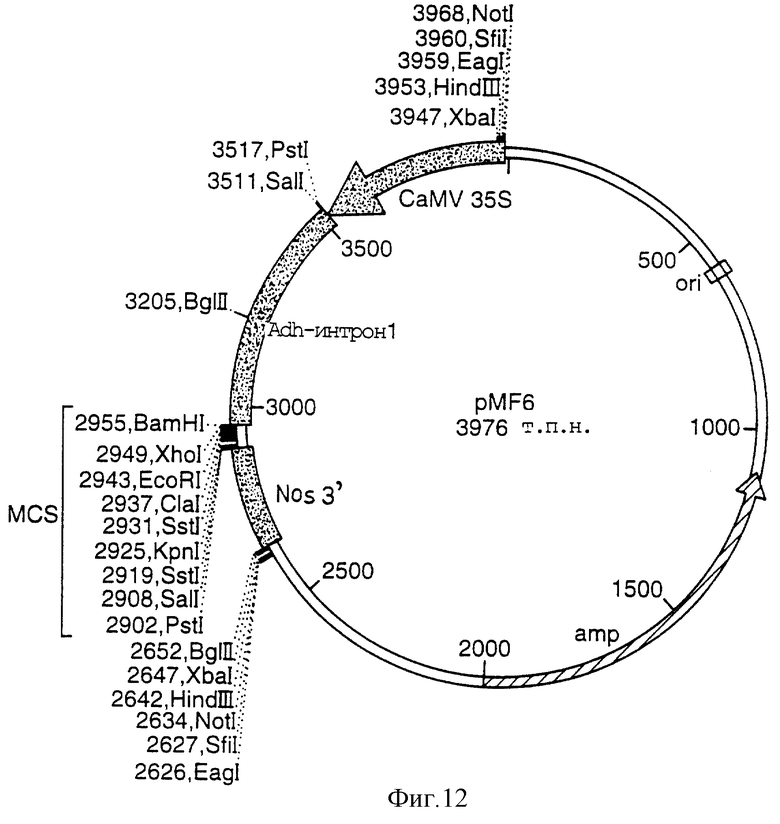
Фигура 12 представляет собой карту плазмиды pMF6;
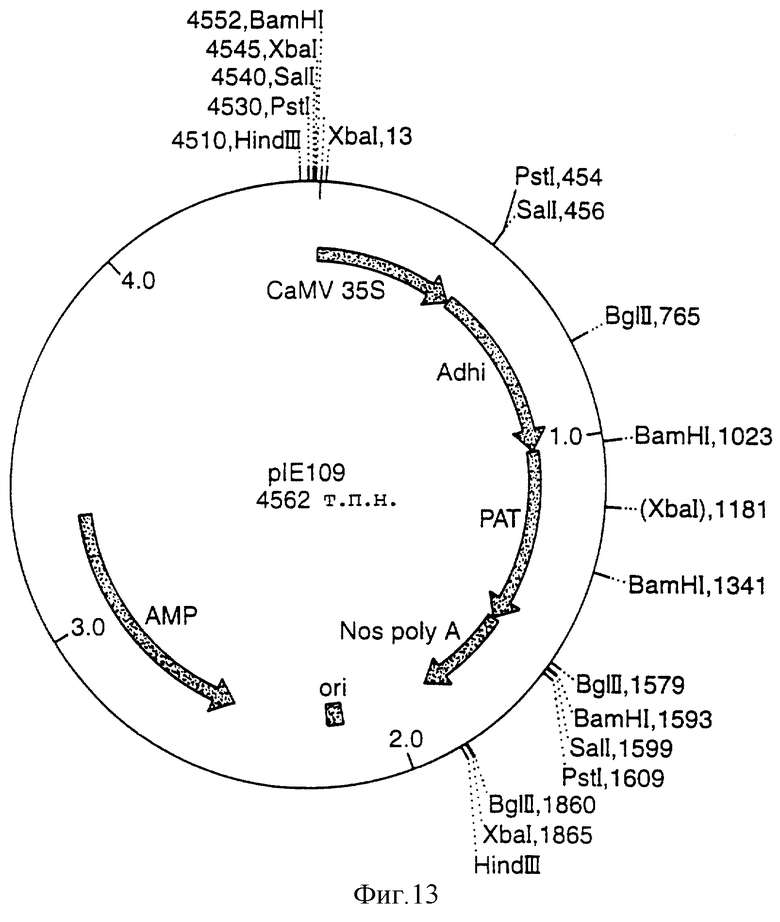
Фигура 13 представляет собой карту плазмиды рIЕ109, в которой величины выражены в парах нуклеотидов (не в масштабе) и используемыми нижеприведенными сокращениями:
ADHi - Алкогольдегидрогеназа из кукурузы;
РАТ - Фосфинотрицинацетилтрансфераза (Basta-устойчивый ген);
АМР - Ген устойчивости к ампициллину;
СаMV-35S - Промoтор 35S вируса мoзаики цветной капусты;
nos Poly A - Область poly А нопалинсинтазы;
ori - ColE1-ориджин репликации из pUC
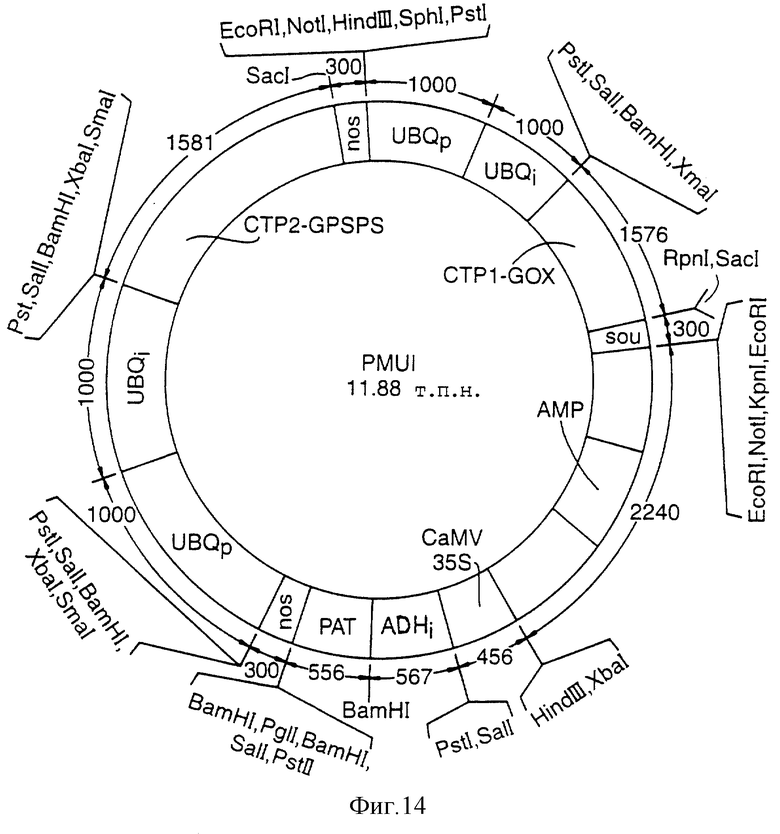
Фигура 14 представляет собой карту плазмиды pMVI, в которой величины выражены в парах нуклеотидов (не в масштабе), а сокращения представлены так же, как и на Фигуре 13, с нижеследующими дополнительными сокращениями:
UBQp - убиквитиновый промотор кукурузы
UBQi - убиквитиновый интрон кукурузы
nos - 3'-терминатор нопалинсинтазы
CZPI GOX - транзитный пептид хлоропласта - последовательность глифосатоксидазы
CZP2 GPSPS - транзитный пептид хлоропласта - последовательность EPSPS-синтетазы
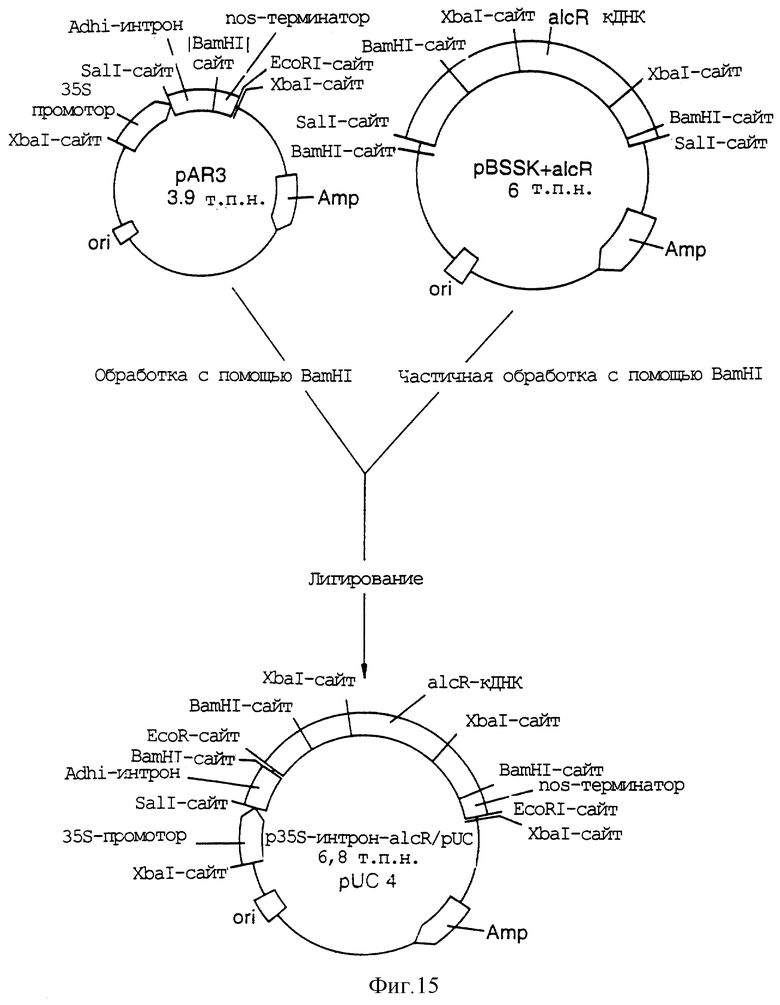
Фигура 15 показывает получение плазмиды pUC4 путем лигирования рАr3 и pBSSK;
Фигура 16 представляет собой карту плазмиды pMV2, в которой величины выражены в парах нуклеотидов (не приведено к масштабу), а сокращения представлены так же, как на Фигуре 14, с нижеследующими дополнительными сокращениями:
А1сА - аlсА-промотор Aspergillus nidulans
AlcR - alcR-промотор Aspergillus nidulans
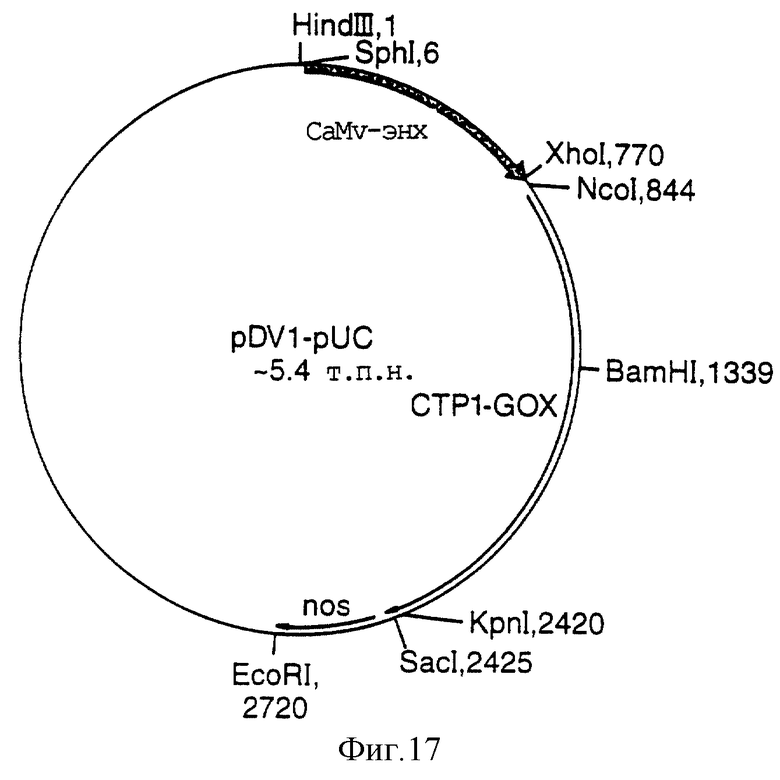
Фигура 17 представляет собой карту плазмиды pDVl-pUC;
Фигура 18 представляет собой карту плазмиды pDV2-pUC;
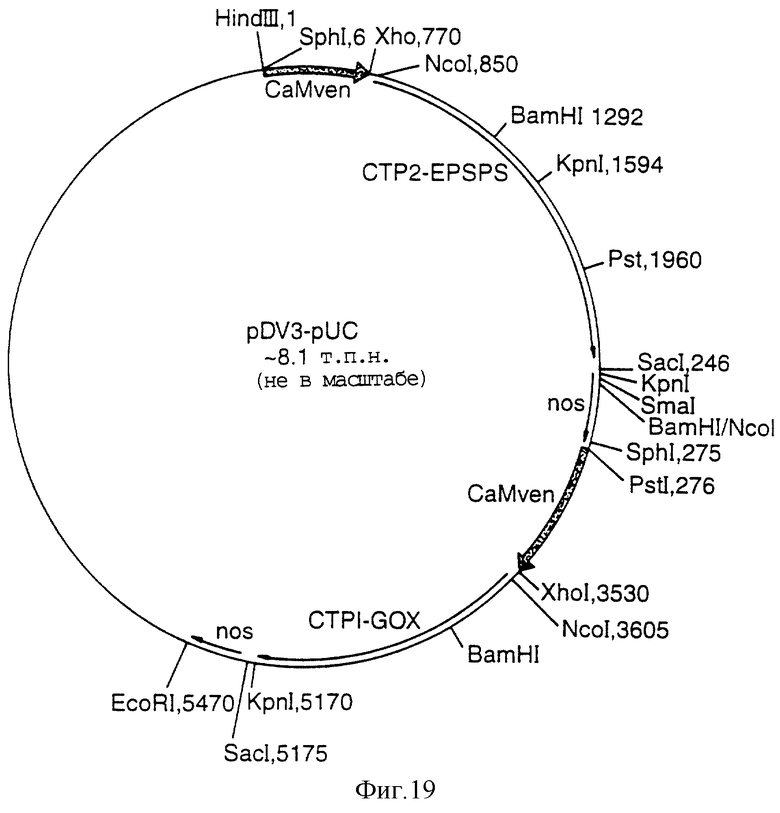
Фигура 19 представляет собой карту плазмиды pDV3-Bin;
Фигура 20 представляет собой карту плазмиды pDV4-Bin;
Фигура 21 представляет собой карту плазмиды pDV5-Bin; и
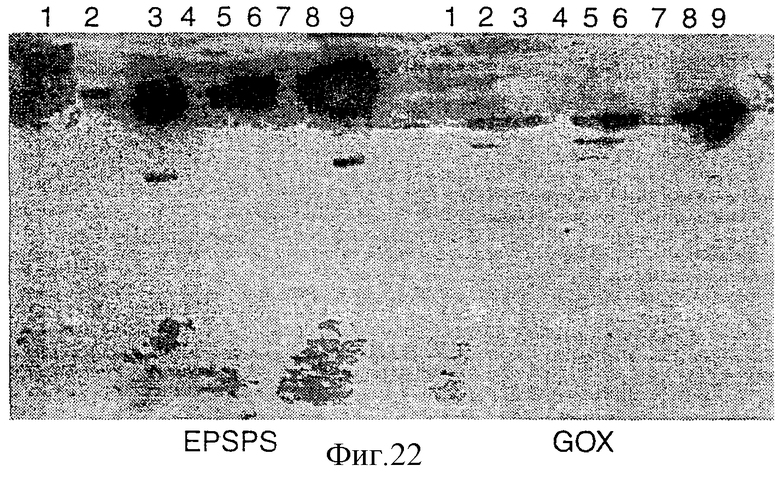
Фигура 22 представляет собой вестерн-блот, показывающий экспрессию EPSPS и GOX в трансформантах.
ПРИМЕРЫ
Для генов, придающих устойчивость к глифосату, мы вы брали пояснение на примере генного переключателя alcA/alcR. Данный переключатель использовали для приведения в действие индуцируемой экспрессии глифосатоксидазы (GOX) в растениях. Переключаемую GOX экспрессировали одну или в сочетании с конститутивной экспрессией 5-енол-пирувилшикимат-3-фосфата (EPSPS) CP4. Конструкты оптимизировали для экспрессии в однодольных и двудольных видах сельскохозяйственных культур.
ПРИМЕР 1
Получение alcR-регуляторного конструкта.
Опубликование геномной ДНК-последовательности alcR позволяет выделить образец кДНК alcR.
кДНК alcR клонировали по экспрессионным векторам pJR1(pUC). pJR1 содержат промотор 35S вируса мозаики цветной капусты. Этот промотор является конститутивным растительным промотором и непрерывно экспрессирует соответствующий регуляторный белок. Сигнал полиаденилирования nos находится в экспрессионном векторе.
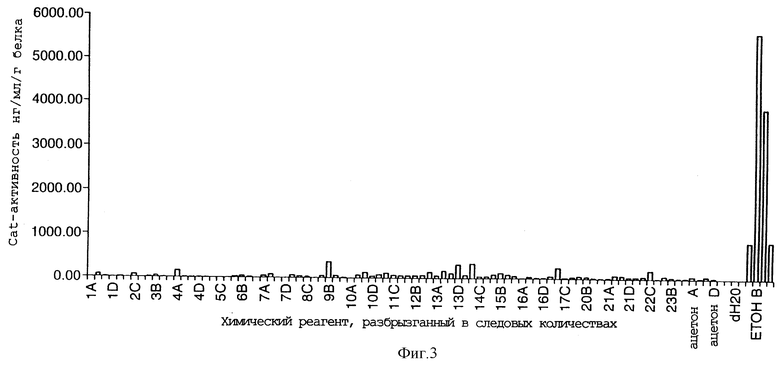
Фигура 4 иллюстрирует образование 35S-регуляторного конструкта путем лигирования alcR-кДНК по pJRl. Частично рестрицированный с помощью BamHI клон кДНК alcR подвергали электрофорезу в агарозном геле и вырезали и выделяли очисткой 2,6 т.п.н. фрагмент. Выделенный фрагмент затем лигировали по pJRl-вектору, который был рестрицирован с помощью ВатНI, и обрабатывали фосфатазой для предотвращения рециркуляризации. Ген alcR был таким образом помещен под контроль промотора CaMV-35S и 3'-сигнал полиаденилирования nos в этом "35S-аlсР"-конструкте.
ПРИМЕР 2
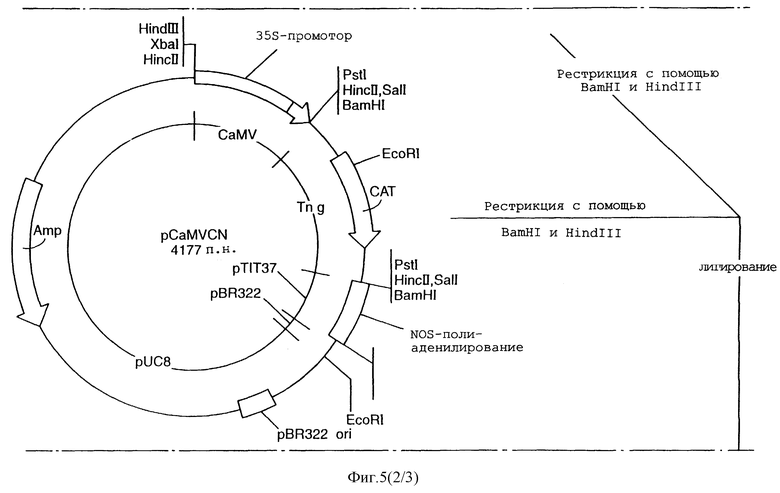
Получение репортерного конструкта alcA-CAT, содержащего химерный промотор.
Плазмида pCaMVCN содержит бактериальную хлорамфениколтрансферазу (CAT) репортерного гена между 35S-промотором и терминатором транскрипции nos (конструкт "35S-CAT").
Промотор аlсА субклонировали в вектор pCaMVCN для получения конструкта "alcA-CAT". Слияние части промотора alcA и части 35S-промотора создает химерный промотор, который позволяет экспрессироваться генам под его контролем.
Фигура 5 иллюстрирует получение репортерного конструкта. Промотор alcA и 35S-промотор обладают идентичными ТАТА-боксами, которые были использованы для сшивки двух промоторов воедино, с использованием методики рекомбинантной PCR: 246 п. н. участок из промотора аlсА и 5'-конца гена CAT из pCaMVCN (содержащего часть сердцевинной области -70 35S-промотора) были отдельно амплифицированы, а затем вместе сплайсированы с использованием PCR. Полученный рекомбинантный фрагмент подвергли затем рестрикционной обработке с помощью BamHI и HindIII. Вектор pCaMVCN частично обрабатывали с помощью ВаmHI и HindIII, затем подвергали электрофорезу так, чтобы можно было бы выделить правильный фрагмент и лигировать с данным рекомбинантным фрагментом.
Лигированные смеси были трансформированы в Е. coli и наслоены на богатую агаровую среду. Из полученных колоний минипрепаративно выделяли плазмидную ДНК, а рекомбинантные клоны извлекали с помощью электрофореза по размеру и рестрикционного картирования. Лигированные стыки секвенировали, чтобы проверить правильность извлеченных рекомбинантов.
ПРИМЕР 3
Конструкты, устойчивые к глифосату
На Фигуре 6 показана сводка полученных кассет и конкретных конструктов, трансформирующих растение.
Вектор 1 двудольных
Вектор 1 представляет собой конститутивную контрольную плазмиду, содержащую ген глифосатоксидазы (GOX), слитый с транзитной последовательностью 1 хлоропласта из Arabidopsis RUBISCO (СРТ 1) (Фигура 7), приводимую в действие с помощью энхансированного 35S-промотора CaMV (ES) и трансляционной энхансерной последовательности TMV-омега (TMV). Вектор 1 использует терминатор нопалинсинтазы (nos). Синтетический GOX-ген с добавлением СТР 1 синтезировали по информации из патентной публикации W092/00377 с добавлением Ncol-сайта на трансляционном старте ATG, а Крn I - на 5'-конце. Внутренние сайты Sph I и Nco I-сайт делегировали во время синтеза, обычно, без аминокислотного изменения. Синтезированную последовательность СТР 1 GOX выделяли в виде фрагмента Nco I Крп I и лигировали с использованием стандартных методик молекулярного клонирования в разрезанную NcoI KpnI pMJB 1 плазмиду, основанную на pIBT 211, содержащую 35-промотор CaMV, с дуплицированным энхансером, сшитым с энхансерной последовательностью трансляции вируса табачной мозаики, замещающей 5'-нетранслируемую лидерную последовательность вируса гравировки табака и терминированной сигналом poly(А) нопалин-синтазы (nos) (Фигура 8).
Кассету, содержащую энхансированную 35 CaMV TMV-последовательность СТР 1 GOX и nos-терминатор (двудольный вектор 1 pUC Фигуры 17), выделяли в виде фрагмента HindIII EcoRI и лигировали в разрезанную HindIII EcoRI pJRIi, основанную на растительном трансформирующем векторе Bin 19 (Фигура 9).
Вектор 2 двудольных
Синтетический СР4-ген EPSPS, слитый с транзитной последовательностью СТР2 хлоропласта (Фигура 10) из гена EPSPS класса I из Petunia hybrida, синтезировали по данным из патента WO 92/04449 с помощью Ncol на ATG инициации трансляции. Внутренний Sph I-сайт был молчащим в СР4-гене EPSPS, обычно, без аминокислотного изменения.
Фрагмент, содержащий синтетический СТР 2 СР4 EPSPS выделяли в виде фрагмента NcoI Sac I и лигировали по pMJBI. Фрагмент, содержащий 35-промотор CaMV с дуплицированным энхансером, транзитным пептидом СТР 2 последовательности TMV-омега, EPSPS и терминатором nos, выделяли в виде фрагмента EcoRI Hind III (вектор 2 pUC двудольных, Фигуры 18) и клонировали по pJRIi, чтобы получить вектор 2 pUC двудольных (Фигура 18).
При секвенировании стыков двудольного вектора 2 идентифицировали дополнительную последовательность, вставленную между SасI-сайтом и началом nos-терминатора. Она имела следующий вид:
5' AGG CTG CTT GAT GAG CTC GGT ACC CGG GGA TCC ATG GAG CCG ААТ 3'
Вектор 3 двудольных
Контрольный вектор с генами EPSPS и GOX конструировали путем разрезания двудольного вектора с помощью EcoRI и инсерцирования линкера ΔEcoRI Sph I ΔEcoRI. Последовательность этого линкера приведена ниже:
5' ААТ TAG GGG CAT GCC CCT 3'
Полученный вектор резали с помощью Sph I, чтобы освободить кассету В, которая была клонирована по SрhI-сайту в двудольном векторе 1), 5' с CaMV-промотором. Затем из двудольного вектора 3-pUC (Фигура 19) вырезали кассеты 1) и 2) в виде фрагмента HindIII и EcoRl и клонировали по pJRIi.
Вектор двудольных 4
Индуцируемый GOX-вектор конструировали путем вырезания САТ-гена из "palcCAT" в виде фрагмента PstI. Векторную полосу, содержащую alcA-промотор и nos-терминатор, выделяли очисткой на геле и использовали для лигирования с линкером PstI-XhoI-KpnI-PstI, последовательность которого имеет следующий вид:
5' GCC ACT CGA GCT AGG TAC CCT GCA 3'
Ее ориентирование подтвердили анализом последовательности. TMV-омега и CTPI GOX-последовательность из двудольного вектора 1) выделили в виде фрагмента XhoI KpnI и клонировали по alcA nos-вектору, содержащему линкер XhoI-KpnI-PstI. Nos-кассету alcA TMV CTP1 GOX вырезали в виде фрагмента Hindlll и клонировали по растительному трансформирующему вектору "p35S-alc R", содержащему nos-терминатор кДНК alcR под контролем 35 CaMV-промотора, с образованием двудольного вектора 4 (Фигура 20).
Вектор двудольных 5
Двудольный вектор 5 (Фигура 22), содержащий индуцируемый ген GOX и конститутивный ген EPSPS, получали с использованием следующей стратегии клонирования. Двудольный вектор 2 (pDV2-pUC) модифицировали путем клонирования в линкере ΔEcoRI-HindIII-ΔEcoRI по ScoRI-сайту, чтобы можно было вырезать CaMV еN-СТР2-ЕРSРS-поs-кассету в виде фрагмента HindIII. Затем этот фрагмент лигировали в pDV4-Bin, разрезанный HindIII. Рекомбинанты, содержащие все три кассеты, т.е. 35S-AlcR, CaMVen-CTP2-EPSPS и AlcA-CTPl-GOX-nos отбирали путем гибридизации с помощью радиоактивно меченных олигонуклеотидов. Подтверждение ориентирования осуществляли путем секвенирования по всем границам.
Векторы однодольных
Вектор 1: Кассета D
Линкер EcoRI-NotI-EcoRI (5' AATTCATTTGCGGCCGCAAATG3') встраивали в двудольный вектор pDVl. Эту плазмиду резали с помощью NcoI и 5'-выступ достраивали с помощью фрагмента Кленова ДНК-полимеразы I. Затем линеаризованный вектор резали с помощью NotI и полученный тупой/NotI-фрагмент, содержащий СТР1 GOX и nos-терминатор, лигировали по обработанному SmaI/NotI вектору pPUBl (Фигура 12), содержащему полиубиквитиновый промотор, полиубиквитиновый интрон с KpnI-NotI-KpnI-линкерной (5'CATTTGCGGCCGCAAATGGTAC3') вставкой. HindIII-NotI-HindIII-линкер (5'AGCTTGCAGCGGCCGCTGCA3') инсерцировали в получаемый конструкт.
Вектор 1: Кассета Е
EcoRI-NotI-EcoRI-линкер (5'AATTCATTTGCGGCCGCAAATG3') встраивали в двудольный вектор pDV2. Эту плазмиду разрезали с помощью NcoI и 5'-выступ достраивали с помощью фрагмента Кленова ДНК-полимеразы I. Полученный линейный вектор разрезали затем с помощью NotI и полученный тупой/NotI-фрагмент, содержащий СТР2 EPSPS и nos-терминатор лигировали по обработанному SmaI/NotI вектору pPUBl, содержащему полиубиквитиновый промотор, полиубиквитиновый интрон с KpnI-NotI-KpnI-линкерной (5'CATTTGCGGCCGCAAATGGT AC3') вставкой, чтобы создать плазмиду 1. Селективная маркерная кассета PAT (промотор CaMV-35S, AdhI-интрон, ген фосфинотрицинацетилтрансферазы (PAT), nos-терминатор) вырезали из рIЕ108 (Фигура 14) и клонировали по HindIII-сайту в плазмиде 1 для получения однодольной кассеты Е. Диагностическую рестрикционную обработку использовали для подтверждения того, что селективная маркерная кассета встроена 5'-3' в той же ориентации, что и кассета СТР2 EPSPS.
Фрагмент, содержащий полиубиквитиновый промотор, полиубиквитиновый интрон, СТР1 GOX и nos-терминатор, вырезали из кассеты D с помощью NotI и лигировали по NotI-кассете Е для образования однодольного вектора 1 (Фигура 14). Рестрикционную обработку использовали для подтверждения того, что две кассеты были инсерцированы в одной и той же ориентации.
Селективную маркерную кассету (промотор-СаМV-35, AdhI-интрон, ген фосфинотрицинацетилтрансферазы (PAT), nos) вырезали из рIЕ108 и клонировали по Hind III-сайту в 5), чтобы получить однодольную кассету Е.
Вектор 1
Фрагмент, содержащий полиубиквитиновый промотор, полиубиквитиновый интрон GOX и nos вырезали из кассеты D с помощью NotI и клонировали по NotI-резанной кассете Е с образованием однодольного вектора 1.
Вектор 2: Кассета F
EcoRI-фрагмент из pUC4 (Фигура 15), содержащий последовательности кДНК alcR и nos-терминатора, достраивали с помощью фрагмента Кленова ДНК-полимеразы I по тупому концу, лигировали по pUBl с помощью КрnI-NotI-KpnI-линкерной вставки и ориентировали после рестрикционного анализа. Селективную маркерную кассету PAT инсерцировали в HindIII-сайт после вырезания из рIЕ108 и ориентировали с помощью рестрикционного анализа, чтобы создать вектор 1. Вышеуказанную плазмиду 1, содержащую полиубиквитиновый промотор, полиубиквитиновый интрон, СТР2 EPSPS и nos-терминатор, разрезали с помощью HindIII и ΔHindIII-NotI-HindIII-линкера:
5'AGCTCGCAGCGGCCGCTGCA3'
5'GCGTCGCCGGCGACGTTCGA3'
встраивали и ориентировали путем секвенирования для создания вектора 2.
ClaI-NcoI-ClaI-линкер (5'CGATGCAGCCATGGCTGCAT3') встраивали в pMF6 (Фигура 13) для получения вектора 3. NcoI/KpnI-фрагмент, содержащий СТР1 GOX, вырезали из pDVl и встраивали в NcoI/KpnI резанный вектор 3 для создания вектора 4. SalI-фрагмент, содержащий кукурузный AdhI-интрон, СТР1 GOX, вырезали из вектора 4 и лигировали в SalI-резанную pUC2, содержащую alcA-промотор и nos-терминатор, и ориентировали с помощью секвенирования для создания вектора 5. HindIII-фрагмент из вектора 5, содержащий alcA-промотор, кукурузный Adhl-интрон, СТР1 GOX и nos-терминатор, лигировали в HindIII-разрезанный вектор 2 и ориентировали путем рестрикционной обработки. NotI-фрагмент из данного полученного конструкта, содержащего полиубиквитиновый промотор, полиубиквитиновый интрон, СТР2 EPSPS, nos-терминатор, alcA-промотор, кукурузный Adhl-интрон, СТР1 GOX и nos-терминатор, лигировали по Notl-разрезанному вектору 1 и ориентировали путем рестрикционного анализа для создания однодольного вектора 2 (Фигура 16).
ПРИМЕР 4
Трансформация растений
Плазмиды для трансформации двудольных переносили в Agrobacterium tumefaciens LBA4404, используя метод замораживания-оттаивания, описанный у Holsters с соавт., 1978.
Трансформанты табака получали с помощью метода листовых дисков, описанного у Bevan 1984. Проростки регенерировали на среде, содержащей 100 мг/л канамицина. После укоренения растения переносили в теплицу и выращивали в условиях 16 ч свет/8 ч темноты.
Трансформации масличного растения paпс (Brassica napus cv westar) осуществляли с использованием метода семядольных черешков, описанного у Moloney с соавт., 1989. Селекцию трансформированного материала осуществляли на канамицине (15 мг/л). Укоренившиеся проростки сразу же переносили на почву, основанную на компосте, и выращивали до стадии зрелости в контролируемых тепличных условиях (16 ч день 20oС днем, 15oС ночью 60% RH).
Трансформацию кукурузы осуществляли с использованием метода бомбардировки частицами, как описано у Klein с соавт., 1988. Селекцию осуществляли на биолофосе, 1 мг/л.
Трансформацию сахарной свеклы осуществляли с использованием метода сохраняемых клеточных протопластов, смотрите нашу Международную патентную публикацию W095/10178.
Результаты, описывающие детали получения трансгенных растений, представлены в Таблицах 2 и 3.
ПРИМЕР 5
Анализ трансгенных растений
Полимеразная цепная реакция (PCR)
Геномную ДНК для PCR-анализа получали в соответствии с методом, описанным у Edwards с соавт., 1992. PCR осуществляли с использованием условий, описанных у Jepson с соавт., Plant Molecular Biology Reporter, 9(2), 131-138 (1991). Для каждой интродуцированной кассеты создавали наборы праймеров.
Растения анализировали с использованием следующих сочетаний олигонуклеотидов: -
pDVl TMV1+GOX 1, GOX3+nos1
pDV2 TMV1+EPSPS, EPSPS3+nos1
pDV3 EPSPS3+GOX1
pDV4 35S+AlcRl, AlcA2+GOX1
pDV5 35S+AlcRl, AlcA2+GOX1, TMV1+EPSPS1
Олигонуклеотидные последовательности приведены ниже:-
TMV1 5'CTCGAGTATTTCAACAATTACCAAC
GOX1 5'AATCAAGGTAACCTTGAATCCA
GOX3 5'ACCACCAACGGTGTTCTTGCTGTTGA
NOS1 5'GCATTACATGTTAATTATTACATGCTT
EPSPS1 5'GTGATACGAGTTTCGCTAGCGAGAC
EPSPS3 5'TACCTTGCGTGGACCAAAGACTCC
35S 5'GTCAACATGGTGGAGCACG
AlcR1 5'GTCCGCACGGAGAG-CCCAAACGA
Селекция на глифосате
Кривые гибели для табака сорт Samsun и рапса сорт Westar в глифосате
Оба вида испытывали в ряду концентраций глифосата путем инсерции, в случае табака - 5-6 мм стеблевого сегмента, несущего листовой узел, и в случае растущей верхушки плюс два листа, масличной культуры рапс, в МS-среду, содержащей глифосат в концентрации 0, 0,0055, 0,011, 0,0275, 0,055 и 0,01 мМ изопропиламиновой соли глифосата. Полученные результаты оценивали через две недели роста, как показано в Таблице 4.
Селекцию по трансформантам, толерантным к глифосату, осуществляли при концентрациях глифосата 0,01 и 0,05 мМ.
Конститутивно толерантные растения
Затем по данным, полученным для растений дикого типа, скринировали первичные трансформанты pDV1, 2 и 3 PCR+ve на MS-среде, содержащей глифосат в вышеуказанных концентрациях. Для табака это осуществляли путем инсерции трех или четырех частей стебля трансформанта в данную среду и использования нетрансформированного Samsun в качестве контроля. Оценка была основана на проведении полученного большинства. Растения, проявившие толерантность при повышенных концентрациях гербицида, росли в теплице до стадии зрелости для сбора семян.
Тест на расщепление
Семена стерилизовали в 10% хлорной извести в течение 10 мин. После нескольких промываний в стерильной воде 200 семян высадили в 1/2 MS-среду (2,3 г/л MS-соли, 1,5% сахарозы, 0,8% бактоагара, рН 5,9), содержащую 100 мг/л канамицина. Семена проращивали при 26oС с фотопериодом 16 часов/8 часов свет/темнота перед оценкой.
Вестерн-анализ
Получение антител
Белки GOX и EPSPS сверхэкпрессировали в Е. coli с использованием экспрессионной системы рЕТ. После индукции IPTG, GOX и EPSPS элюировали из клеточной массы, выращенной в качалочной склянке, и использовали для иммунизации кроликов (два животных на клон).
Приготовление тканевых экстрактов для иммуноблоттинга
120 мг ткани листьев плюс 60 мг PVPP и 500 мкл экстрагирующего буфера (50 мМ Трис-НСl рН 8, 1 мМ ЭДТА, 0,3 мМ DTT) измельчали в смесителе в течение нескольких минут. После гомогенизации полученный экстракт центрифугировали при 15000 об/мин в течение 15 мин. Полученный супернатант хранили при -80oС до востребования. Концентрации белка в экстракте измеряли по Брэдфорду.
SDS-PAGE и иммуноблоттинг
25 мкг белка фракционировали с помощью SDS-PAGE. Буфер для электрофореза содержал 14,4% (в/о) глицина, 1% (в/о) SDS и 3% (в/о) триса-основания. Белковые образцы вносили по Laemmli.
После SDS-PAGE белки электроблоттировали в течение ночи при 40 мА в нитроцеллюлозу (Hybond С, Amersham), используя прибор для электроблота от Biorad. Полученный фильтр окрашивали в 0,05% CPTS, растворенным в 12 мМ НС1. Блоты споласкивали в 12 мМ НС1 и деокрашивали в течение 5-10 мин в 0,5 NаНСО3 с последующей тщательной отмывкой при помощи Н2О. Затем фильтры фиксировали, иммунодетектировали в соответствии с Amersham ECL-набором. Непрямую иммунодетекцию осуществляли при разведении 1:10000 поликлонального кроличьего анти-GOX или анти-EPSPS в качестве первого антитела и при разведении 1: 1000 антикроличьего второго антитела, ассоциированного с пероксидазой хрена. Для удаления фона осуществляли дополнительную промывку в течение ночи. Детекцию осуществляли, используя набор ECL от Amershama, и полученные результаты показаны на Фигуре 21, в которой дорожка (1) представляет собой контроль, а остальные дорожки представляют трансформанты. Данный вестерн-анализ демонстрирует, что некоторые трансформанты обладают способностью экспрессировать GOX и EPSPS.
Конститутивно толерантные растения
Клеточные экстракты получали из каждого растения, толерантного к глифосату, и количество экспрессированного белка оценивали с помощью вестерн-анализа с использованием соответствующего антитела к соответствующему трансформанту. Растения, экспрессирующие очень высокие уровни GOX и EPSPS, тестировали при повышенных концентрациях глифосата по сравнению с уровнем экспрессии при гербицидной толерантности.
Индуцируемые толерантные растения
Для демонстрации индуцируемой толерантности к глифосату PCR-позитивные первичные трансформанты из трансформаций с помощью pDV4 и 5 сразу переносили в теплицу. Спустя две недели эти растения индуцировали с помощью смачивания корней этанолом (5%-ный раствор) и оставляли на 24 часа до осуществления вестерн-анализа, чтобы определить уровень экспрессии GOX после индукции. После периода времени, позволяющего растению вернуться в неиндуцированное состояние, такой вестерн-анализ повторяли, чтобы иметь возможность отобрать индуцируемые толерантные растения. Растения, которые показали наивысшие уровни экспрессии GOX, брали для дальнейшего анализа. Уровни GOX определяли с помощью вестерн-анализа на 6, 12, 18, 24, 36, 48 часов после этанольной обработки.
Высокоэкспрессирующие GOX-растения по pDV4 и по pDV5 использовали для опытов в теплице для демонстрации индуцируемой толерантности к глифосату. Растения индуцировали, используя ряд этанольных концентраций (1-15%) путем смачивания корней растений, выращенных в горшках. После GOX-индукции растения опрыскивали глифосатом. Контроли дикого типа и неиндуцированные растения также обрабатывали гербицидом.
Нозерн-анализ
Первичные трансформанты, содержащие двудольные векторы 2)., 3)., анализировали с помощью нозерн-блот-анализа, используя СТР EPSPS-пробу в виде NсоISасI-фрагмента. Первичные трансформанты, содержащие двудольные векторы 1). 3)., анализировали с помощью нозерн-блоттирования, используя CTPI GOX-пробу в виде NcoIKpnI-фрагмента. Аналогично, трансгенные злаковые линии, содержащие однодольные векторы 1). и 2)., анализировали с использованием СТР2 EPSPS-пробы.
Трансформанты, содержащие двудольный вектор 5). или однодольный вектор 2). , обрабатывали путем смачивания листьев 5%-ным этанолом для индукции GOX-уровней. РНК выделяли через 24 часа после обработки и подвергали нозерн-анализу с помощью CTPI GOX-пробы.
Первичные трансформанты, которые оказались PCR-позитивными по соответствующим кассетам и показали уровни транскрипции GOX и EPSPS, отбирали для дальнейшего анализа.
Анализ глифосатоксиредуктазы
Анализ глифосатоксиредуктазы осуществляли как описано у Kishore и Barry (WO 92/00377). Эти ограниченные, глифосатзависимые измерения потребления кислорода с использованием кислородного электрода обнаруживают образование глифосата в результате взаимодействия с 2,4-динитрофенилгидразином и определением гидрозона, с использованием HPLC или, предпочтительно, с использованием [3-14С]-глифосата в качестве субстрата, и обнаруживают образование радиоактивной аминометилфосфоновой кислоты посредством HPLC на анионобменной колонке.
Анализ EPSPS
Анализ активности синтазы 5-енолпирувилшикимат-3-фосфата (EPSPS) в растительных экстрактах осуществляли (1) в результате регистрации исчезновения фосфоенолпируватного субстрата (как описано у Rubin J.L., Gaines C.G. и Jensen R. A. , в Plant Physiol (1984 75, 839-845) или (2) в результате проведения опыта в обратном направлении и сопряжения с пируваткиназой и лактатдегидрогеназой (как описано у Mousdale D.M. и Coggins J.R. в Planta (1984) 160, 78-83 или (3) в результате использования 14(-меченного фосфоенолпирувата в качестве субстрата и обнаружения образования радиоактивного EPSPS с помощью HPLC на анионобменной колонке и выявления используемого радиоактивного детектора в потоке, как описано у Della-Ciolla с соавт. в Proc. Nat. Acad. Sci. (USA) (1986), 83, 6873-6877. Последний анализ применили в подтверждение того, что EPSPS-синтазная активность оказалась, как ожидали, относительно устойчивой к ингибированию глифосатом.















Изобретение относится к биотехнологии и может быть использовано в селекции растений. Химически индуцируемая генная экспрессионная кассета включает индуцируемый промотор и ген-мишень, придающий устойчивость растению к N-фосфонометилглицину и его производным. Введение химически индуцируемой генной экспрессионной кассеты в растительные клетки позволяет контролировать устойчивость растения к гербициду путем ее возникновения только в присутствии экзогенного химического индуктора, что обеспечивает избирательное уничтожение сорняков. 4 с. и 5 з.п. ф-лы, 22 ил., 4 табл.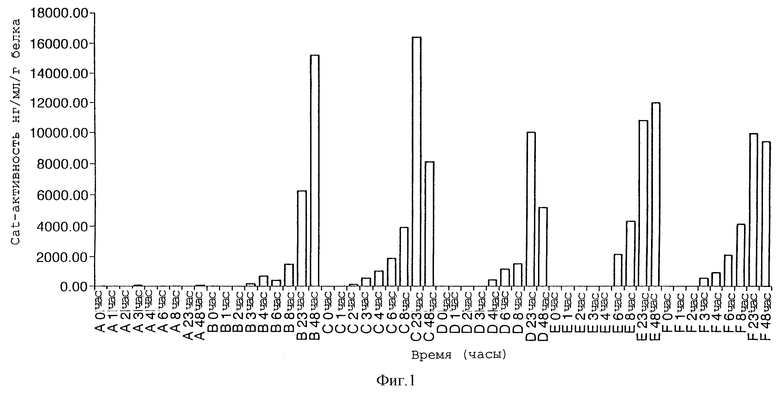
| СПОСОБ АВТОМАТИЧЕСКОГО РЕГУЛИРОВАНИЯ ПРОЦЕССА ОБЖИГА В ПЕЧАХ С КИПЯЩИМ СЛОЕМ | 0 |
|
SU388186A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| СПОСОБ ПОЛУЧЕНИЯ ТЕРМОРЕАКТИВНЫХ СМОЛ | 0 |
|
SU332104A1 |
