Изобретение относится к электрохимии ферментов, в особенности электрохимии выделенных гидроксилаз.
Гидроксилазы известны, как катализаторы окисления различных субстратов. Например, метанмонооксигеназа является эффективным катализатором окисления метана молекулярным кислородом для получения метанола в качестве единственного продукта.
Исследования структуры и механизма электрохимии гидроксилаз описаны в литературе.
Известны две формы фермента метанмонооксигеназы, растворимая (sMMO) и в виде частиц (корпускулярная) (pMMO). Растворимый фермент, продуцируемый Methylococcus capsulatus (Bath), состоит из гидроксилаз (Mr 250.5 кДа), редуктазы (Mr 38.5 кДа) и регуляторного компонента белка В (Mr 15.9 кДа), каждый из которых необходим для активности этого фермента. Гидроксилаза состоит из двух протомеров с расположением a2b2g2, и кристаллическая структура этого компонента была определена рентгеновским исследованием. sMMO Methylosinus trichosporium OB3b имеет очень похожий состав и также хорошо описана. Активным сайтом у sMMO является центр с двумя атомами железа, связанными гидроксигруппой у неактивного фермента, находящийся в субъединице гидроксилазы. Восстанавливающие эквиваленты передаются от НАДФ активному центру через Fe2S2 и ФАД центры в редуктазе. Белок В не содержит ионов металлов или кофакторов, и детали его регуляторной роли неясны. У неактивного фермента атомы железа находятся в полностью окисленном состоянии Fe(III)Fe(III). Существует также два других состояния окисления, легко достигаемые в двухатомном железном кластере, а именно со смешанной валентностью Fe(III)Fe(II) и полностью восстановленное Fe(II)Fe(II). Именно Fe(II)Fe(II) форма гидроксилазы вступает в реакцию с кислородом активирует его в холе ферментативного превращения.
Окислительно-восстановительные потенциалы двухатомных железных центров у M. capsulatus (Bath) и M.trichosporum OB3b были измерены в трех независимых исследованиях. Значения Eo' для Fe(III)Fe(III)/Fe(III)Fe(II) и Fe(III)Fe(II)/Fe(II)Fe(II) были предметом дебатов. Окислительно-восстановительные свойства гидроксилазного компонента растворимой метанооксигеназы от двух различных микроорганизмов были подробно изучены. В предыдущих исследованиях использовали окислительно-восстановительное титрование с индикатором и спектроскопические методы для определения концентрации восстановленных образцов. В этих исследованиях использовали непрямое титрование гидроксилазных двухатомных железных центров окислительно-восстановительными медиаторами. Концентрации смешанно-валентных и полностью восстановленных гидроксилаз определяли при помощи ЭПР или сочетанием ЭПР со спектроскопией по Мессбауэру при очень низких температурах (4.2 - 18 K).
В области электрохимии белков было обнаружено, что тиол- или дисульфидсодержащие органические молекулы являются особенно хорошими модификаторами, поскольку они хемосорбируются золочением, образуя стабильный слой, покрывавший поверхность электрода. Подходящие аминокислоты также могут хемосорбироваться тем же способом и тем самым содействовать электрохимическим белковым превращениям на поверхности электрода. Было исследовано использование цистеинсодержащих пептидов, которые также содержат функциональные аминокислоты (т. е. аргинин, лизин, гистидин), в качестве активаторов электрохимических белковых реакций.
Однако в отличие от электрохимии белков осуществление непосредственных электрохимических мероприятий на окислительно-восстановительных ферментах без помощи медиаторов не является тривиальной задачей. Основная трудность состоит в том, что часто окислительно-восстановительные центры расположены глубоко в массе белка, далеко от его поверхности, так, что путь, который необходимо проделать электронам до электрода, может быть достаточно большим, что снижает скорость переноса до крайне малой величины. Также поскольку большинство окислительно-восстановительных ферментов значительно больше и структурно менее прочные по сравнению с другими белками, то они более подвержены деформации и потере активности на поверхности электрода.
Электрохимия гидроксилаз в присутствии медиатора усложняется из-за возможности взаимодействия между гидроксилазой и медиатором, при этом медиатор может препятствовать определению концентрации образцов при температурах, отличающихся от температур проведения окислительно-восстановительных реакций.
Задача данного изобретения состоит в том, чтобы избежать вышеперечисленных недостатков.
Согласно одному из объектов данного изобретения предлагается способ переноса электронов между электродом и ферментом в электрохимическом процессе, предусматривающий вызывание прилипания фермента к электроду.
Предпочтительно перенос является прямым (непосредственным).
Согласно другому объекту изобретения предлагается способ электрохимии фермента, предусматривающий прямую электрохимию, проводимую в отсутствие медиаторов.
В варианте выполнения изобретения фермент является монооксигеназой.
В другом варианте изобретения фермент является Р450 или модифицированным Р450.
В еще одном варианте изобретения фермент является метанмонооксигеназой.
В еще одном выполнении изобретения фермент является гидроксилазой.
Гидроксилаза может быть растворимой метилмонооксигеназой.
Фермент может содержать двухатомный железный активный сайт или альтернативно фермент может включать порфиринсодержащий активный сайт.
Предпочтительно электрохимию проводят при использовании модифицированного золотого электрода.
Согласно дополнительному объекту изобретения предлагается способ модифицирования электрода для подготовки электрода к использованию в электрохимии фермента при отсутствии медиаторов, который предусматривает обработку электрода белком.
Предпочтительным ферментом является монооксигеназа.
Подходящий фермент имеет двухатомный железный или порфиринсодержащий активный сайт.
Обычно пептид включает гексапептид.
Предпочтительно гексапептид содержит Цис и Лиз радикалы.
Подходящим гексапептидом является Лиз-Цис-Тре-Цис-Цис-Ала.
Обычно обработка включает вольтаметрические циклы работы электрода в растворе гексапептида, избегая при этом восстановительного очищения электрода.
Согласно еще одному объекту изобретения предлагается способ окисления субстрата молекулярным кислородом в присутствии фермента, предусматривающий прямую электрохимию, проводимую в отсутствие медиаторов.
В варианте выполнения изобретения фермент является монооксигеназой.
В другом варианте изобретения фермент имеет двухатомный железный или порфиринсодержащий активный сайт.
Субстрат может быть метаном, который окисляют до метанола, или субстрат может быть камфарой, которую окисляют до 5-экзо-гидроксикамфары, или же субстрат может содержать полициклические ароматические углеводороды, которые окисляют до 5-экзо-гидроксикамфары.
Гидроксилазу предпочтительно получают из цитохрома Р450cam.
Обычно цистеин удаляют из реакционной среды.
Мы обнаружили, что электрод, модифицированный пептидами, обеспечивает более благоприятную поверхность для фиксации фермента.
Мы полагаем, что для того, чтобы происходил электронный перенос между гидроксилазным компонентом M.capsulatus (Bath) sMMO и электродом, предпочтительна положительно заряженная поверхность: e-аминогруппа лизинового радикала в предпочтительном гексапептиде предположительно выполняет эту роль. Электрохимическая модификация электрода этим конкретным пентилом обеспечивает стабильный монослой и более однородное покрытие поверхности по сравнению с электродом, модифицированным хемосорбцией (т.е. простым погружением электрода в раствор пептида).
Электрод может быть модифицированным золотым электродом, например золотым электродом, модифицированным обработкой гексапептидом Лиз-Цис-Тре-Цис-Цис-Ала. Обработка может проводиться циклическим действием электрода в растворе гексапептида, избегая восстановительного очищения электрода.
Влияние белка B и белка B' на электрохимию раствора гидролазы очень важно. ЭПР спектроскопия смешанного валентного состояния показала, что белок В, связываясь с гидроксилазой, изменяет окружение двухатомного железного сайта. В каждом из этих предварительных исследований также наблюдалось, что они вызывают сдвиг потенциалов двухатомного железного сайта.
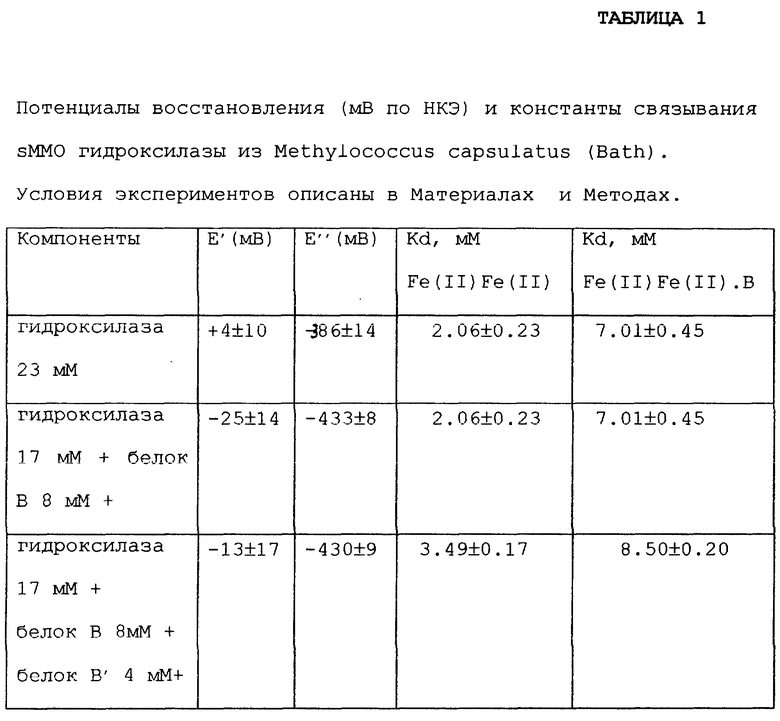
Известно, что образование комплекса "гидроксилаза - белок B" изменяет свойства двухатомного железного сайта, влияя на распределение продуктов и увеличение скоростей образования некоторых промежуточных продуктов в sMMO каталитическом цикле. Все эти эффекты могут быть вызваны способностью белка В вызывать конформационные изменения в гидроксилазе при связывании с ней, которые передаются активному сайту. Мы обнаружили, что на окислительно-восстановительные потенциалы гидроксилазы влияет присутствие белка В, что наблюдалось и в предыдущих исследованиях. Предпочтительно отрицательный сдвиг окислительно-восстановительных потенциалов, вызванный связыванием белка В с гидроксилазой, обусловлен изменением в первой координационной сфере атома(ов) железа или состояния протонирования гидроксильного мостика. Другие эффекты, такие как изменения доступности растворителя или структуры окружения активного сайта, вызываемые конформационными изменениями при связывании белка В, также могут являться причиной изменений потенциала. По нашим оценкам Kd для Fe(II)Fe(II) комплекса равна 2.06±0.23 М и 7.01±0.45 М для Fe(II)Fe(III) комплекса. Таким образом, уменьшение окислительно-восстановительных потенциалов может являться простым отражением снижения аффинности (сродства) между гидроксилазным и B-белковым компонентами в зависимости от степени окисления первого.
В ферментной системе M.capsulatus (Bath) две формы регуляторного белка В были стандартно очищены. Они представляют собой белок B полной длины и часть из двенадцати аминокислот белка B'. Исследования связывания при использовании поверхностного плазменного резонанса (SPR) показали, что белок B' также образует комплекс с полностью окисленной гидроксилазой, которая только лишь в три раза менее стабильна, чем соответствующий комплекс белка В. Наши исследования дают основания полагать, что белок B' связывается с тем же сайтом, что и белок B, но первый не способен вызывать такие же конформационные изменения в гидроксилазе, которые передавались бы на двухатомный железный сайт и которые изменяли бы окислительно-восстановительные потенциалы и другие свойства активного сайта. Образование усеченного белка В' может, таким образом, играть некоторую регуляторную роль в организме. При определенных условиях белок В расщепляется с образованием белка В' и у гидроксилазы могут наблюдаться конформационные изменения (и сопутствующие изменения свойств активного сайта). Таким образом клетка может сохранять небольшие ресурсы в условиях стресса. Мы обнаружили, что прямая электрохимия без введения медиаторов возможна с белком неэлектронного переноса, имеющим такие же размеры, как и гидроксилазный компонент растворимой метанмонооксигеназы.
Как отмечалось выше, процесс может представлять собой, например, окисление метана до метанола или окисление камфары до 5-экзо-гидроксикамфары и, в частности, окисление камфары до 5-экзо-гидроксикамфары при использовании гидроксилазы, полученной из цитохрома Р450cam. В последнем процессе, как нами обнаружено, желательно удалить цистеин из реакционной среды.
Гидроксилазы с порфириновым центром, которые характерны для бактерий Pseudomonas putida (выращенных на камфаре), являются идеальной системой для изучения регуляции электронного переноса между двумя окислительно-восстановительными центрами (разных) молекул белков.
Цитохром P450cam катализирует монооксигенирование D-(+)- камфары с получением 5-экзо-гидроксикамфары, для которой необходим внешний источник двух восстанавливающих эквивалентов. Эти электроны переносятся от НАДФ к железу гема-Р450cam посредством комбинированного действия ФАД-флавопротеина путидаредоксин редуктазы и железосерного белка путидаредоксина. Значительные изменения, наблюдаемые в оптическом спектре гема при связывании с камфарой, были объяснены спектроскопией по Мессбауэру и ЭПР исследованиями как вызываемые изменением спин-состояния железа в трехвалентном железном геме от малого спина в отсутствие субстрата до высокого спина у белка, связанного с субстратом. Кроме того, также отмечалось, что это изменение спин-состояния при связывании с камфарой также сопровождается изменением окислительно-восстановительного потенциала от -540 мВ до -414 мB (по НКЭ (насыщенный каломельный электрод)). Этот сдвиг восстановительного потенциала гема является основной характеристикой Р450cam каталитического цикла, позволяя протекать первому электронному переносу от путидаредоксина и таким образом инициируя каталитическое превращение.
Мы обнаружили, что ток катодного пика обнаруживает линейную зависимость от квадратного корня скорости сканирования (v) для связанного камфарой P450cam, показывая, что суммарный процесс является диффузионно управляемым.
Однако для фермента, не содержащего камфару, наблюдалось отклонение от линейной зависимости. Это может быть либо вызвано адсорбцией на поверхности электрода, или результатом того, что гетерогенный электронный перенос больше не контролируется диффузией. Первое маловероятно, поскольку глобальная структура связанного с камфарой и не содержащего камфары Р450cam сходна, и, таким образом, следует ожидать, что обе формы имеют схожую аффинность к поверхности электрода. Таким образом, это позволяет предположить, что скорость гетерогенного электронного переноса к форме, свободной от камфары, ниже, чем к форме, связанной с камфарой. Это может быть вызвано реорганизацией относительно высокого уровня реорганизацией, необходимой для получения пятикоординатной высокоспиновой формы двухвалентного железа из шестикоординатной низкоспиновой формы фермента, не содержащей камфары, что влечет за собой потерю лиганда воды, связанного с гемом. Это требование сходно с тем, которое налагается на миоглобин, который, как было показано, должен иметь высокую энергию реорганизации по сравнению с восстановлением цитохрома, который содержит шестикоординатные атомы железа гема одновременно в трехвалентном и двухвалентном состоянии.
В случае связанного с камфарой P450cam обе - трехвалентная и двухвалентная - формы фермента содержат пятикоординатный высокоспиновый атом железа гема, и барьер реорганизации как для гомогенного, так и гетерогенного электронного переноса должен быть низким.
Электрохимический ответ P450cam на отрицательно заряженный cpg электрод без покрытия точно подтверждает, что несмотря на общий отрицательный заряд белка при pH 7.4 (p1=4.55) специфическая структура положительно заряженной поверхности аминокислотных радикалов способствует взаимодействию между ферментом и поверхностью электрода таким образом, чтобы мог происходить гетерогенный электронный перенос. Компьютерное моделирование и исследования мутагенеза показывают, что основные радикалы Apr-72, Apr-112, Apr-364 и Лиз-344 на поверхности pf P450cam взаимодействуют с кислыми радикалами на поверхности путидаредоксина в комплексе, образованном из этих двух белков. Согласно кристаллической структуре высокого разрешения P450cam, эти поверхностные радикалы расположены на проксимальной (ближней) к гему стороне и в той области фермента, где гем наиболее близок (приблизительно 10  ) к его поверхности. Поскольку константа скорости электронного переноса обычно уменьшается при разделении донора и акцептора, разумно предположить, что эта область поверхности P450cam будет образовывать одновременно сайт связывания с путидаредоксином и область взаимодействия с электродом. Таким образом, это означает, что положительно заряженные радикалы также участвуют во взаимодействии между Р450cam и cpg-электродом без покрытия.
) к его поверхности. Поскольку константа скорости электронного переноса обычно уменьшается при разделении донора и акцептора, разумно предположить, что эта область поверхности P450cam будет образовывать одновременно сайт связывания с путидаредоксином и область взаимодействия с электродом. Таким образом, это означает, что положительно заряженные радикалы также участвуют во взаимодействии между Р450cam и cpg-электродом без покрытия.
Таким образом, графитовый электрод с гладкой кромкой может успешно заменить цепь белков физиологического электронного переноса при переносе первого электрона к цитохрому P450cam, причем прямая электрохимия является зависимой от субстрата.
Далее изобретение будет описано исключительно в качестве примера со ссылкой на сопровождающие иллюстрации, на которых:
Фиг. 1 - дифференциальная вольтамперограмма 23.6 мМ растворов гидроксилазы на Лиз-Цис-Тре-Цис-Цис-Ала модифицированном золотом электроде в 40 мМ MOPS, pH 7.0 (фоновый ток был вычтен).
Фиг. 2А и 2Б - диаграммы влияния увеличения концентрации белка B (1) или B' (2) на потенциалы электронного переноса первого электрона (а) и второго электрона (б). [Гидроксилаза] = 21 мМ, 40 мМ MOPS, pH 7.0. Электрод проверяли предварительно перед каждым измерением.
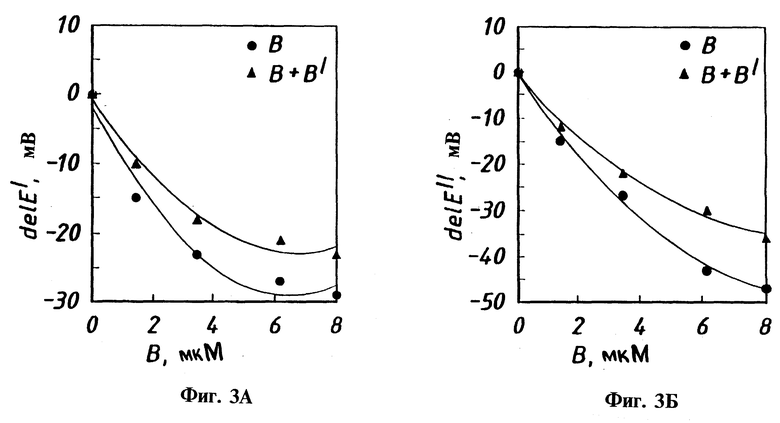
Фиг. 3А и 3Б - диаграммы влияния увеличения концентрации белка В на потенциалы электронного переноса первого электрона (а) и второго электрона (б) в присутствии белка В'. [Гидроксилаза] = 21 мМ, [B'] = 4 мМ, 40 мМ, pH 7.0.
Фиг. 4 - циклические вольтамперограммы цитохрома Р450cam.
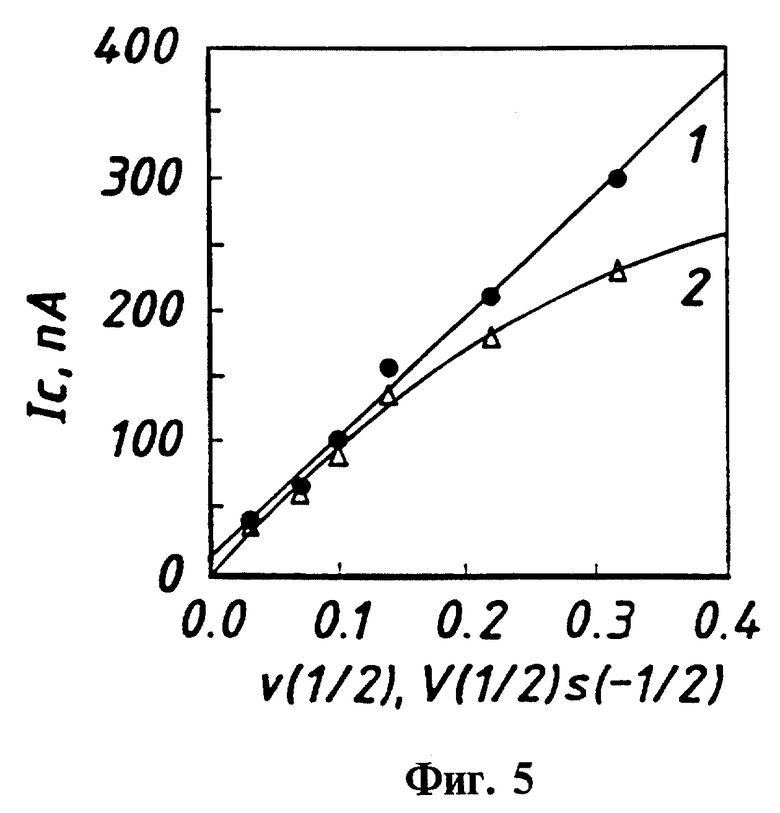
Фиг. 5 - зависимость тока катодного пика от скорости развертки потенциала для цитохрома Р450cam, связанного с камфарой и не содержащего камфары.
Примеры.
(1) Рост Фермента
Выращивание микроорганизма Methylococcus capsulatus (Bath) проводили, как описано в Pilkington, S.J. & Dalton, H. (1990) "Soluble Methane Monoxygenase from Methylococcus capsulatus (Bath)". Methods in Enzymol. 188, 181-190.
При очистке гидроксилазного и B-белкового компонентов были внесены следующие модификации в опубликованные методики: После начальной стадии ионного обмена, гидроксилазу пропускали через Superdex 200 гелевую инфильтрационную колонку. Окончательную очистку проводили элюированием чистой гидроксилазы из Mono Q ионообменной колонны с использованием 0-30%-ного градиента 1 M NaCl. Белок В был очищен сходным способом за исключением того, что гелевой фильтрационный средой была Superdex 75.
(II) Модификация электродов
Золотые электроды были модифицированы при циклической работе электродов при пониженных потенциалах в 5 мМ растворе гексапептида Лиз-Цис-Тре-Цис-Цис-Ала. При проведении цикла восстановления особое внимание уделялось тому, чтобы не превышать предела -0.85 В, поскольку восстановление протонов до водорода приводит к восстановительному очищению золотой поверхности. Было обнаружено, что десять циклов приводят к адекватной модификации. Гексапептид, используемый для модифицирования золотого электрода, был приобретен у Sigma Chemical Company, Poole, Dorset.
(III) Измерение окислительно-восстановительных потенциалов
В качестве метода измерения окислительно-восстановительных потенциалов гидроксилазных двухатомных железных центров была выбрана дифференциальная пульсационная вольтамперометрия (ДПВ).
ДПВ эксперименты с sMMO гидроксилазой дали две волны при 4±10 (волна I на фиг. 1) и -386±144 мВ (волна II на фиг. 1) при катодном сканировании. Пики были соотнесены с переносом соответственно первого и второго электронов к гироксилазному двухатомному железному центру.
Дифференциальная пульсационная вольтамперометрия (ДПВ) проводилась на стандартном стационарном поликристаллическом золотом дисковом (4 мм диаметр) электроде, помещенном в трехэлектродную, двухкамерную стеклянную ячейку небольшого объема (300-400 мл). Ненасыщенный каломельный электрод (НКЭ) (Radiometer К-401) служил в качестве электрода сравнения. Все потенциалы приведены здесь по отношению к НКЭ. Все эксперименты проводились в 40 мМ MOPS буфере при pH 7.0 при 23oC.
(IV) Влияние белков B и B'
Кроме того, определяли влияние белков B и B' на окислительно-восстановительные реакции.
Дополнительные эксперименты проводили при добавлении регуляторного белка B к раствору, используемому для дифференциальной пульсационной вольтамперометрии для определения его влияния на электрохимию гидроксилазы. В этих экспериментах белок В добавляли к гидроксилазе в минимальном соотношении 2:1 (В: гидроксилаза). Высокие соотношения В:гидроксилаза приводят к уменьшению обоих пиков тока до пренебрежимо малых величин, вероятно, из-за загрязнения электрода. Электрод был модифицирован до измерений каждой из различных концентраций белка В для обеспечения наиболее однородного, свежего покрытия. Как показано на фиг. 2, увеличение концентрации белка B приводит к сдвигу обоих первых и вторых потенциалов пиков волн в область более низких значений. Однако изменения каждого потенциала были различны. Общий сдвиг первого потенциала был 29 мВ, но 47 мВ для второго. Это обусловлено различными константами связывания белка В с двухатомным железным центром гидроксилазы в смешанном и полностью восстановленном состояниях.
Константы диссоциации. Kd=2.06±0.23 М и 7.01±0.45 М были определены для белка, связанного с Fe(III)Fe(II) и Fe(II)Fe (II) формами гидроксилазы соответственно, согласно способу, описанному Klotz, I.М. & Hunston, D.L. (1970) "Properties of Graphical Representation of Multiple Classes of Binding Sites", Biochem. 10, 3065-3069. Как можно видеть на фиг. 2, добавление чистого белка B', являющегося усеченной формой белка B, не оказала никакого влияния на электрохимию sMMO гидроксилазы. Однако в смеси B+B наблюдалось, что влияние белка на сдвиг потенциала становится менее выраженным, а также наблюдалось небольшое увеличение Kd для обоих восстановленных состояний белка. В присутствии белка B' для комплексов "гидроксилаза - белок B" были определены Kd=3.49±0.23 М для смешанного валентного и Kd=8.50±0.23 М для полностью окисленного состояния. Полученные данные свидетельствуют, что в B+B' смеси восстановление гидроксилазы становится до некоторой степени ограниченным. Потенциалы восстановления и константы связывания комплексов "гидроксилаза - белок B" при различных условиях исследования суммированы в таблице 1.
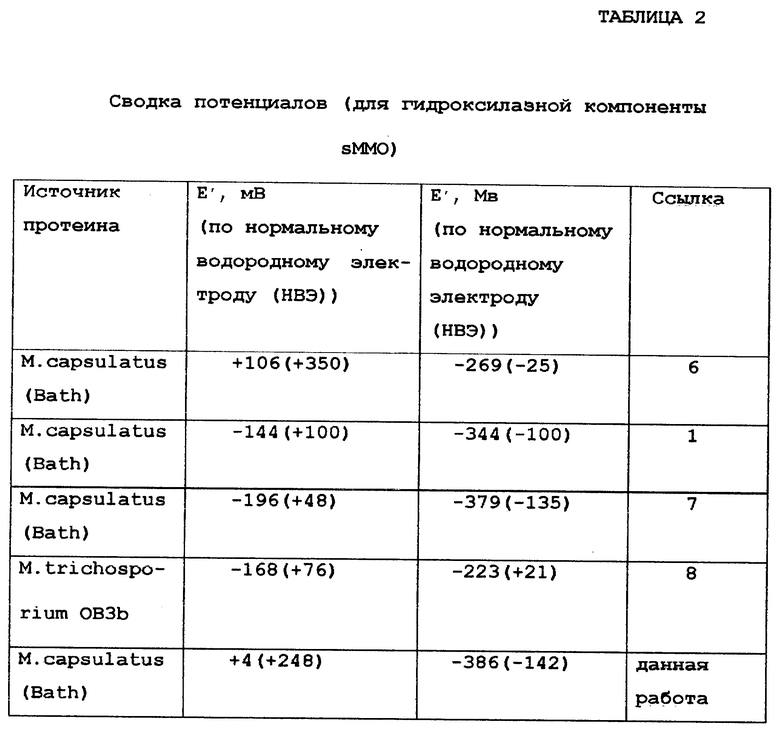
Предварительные исследования окислительно-восстановительных свойств гидроксилазной компоненты sMMO дали довольно большой диапазон окислительно-восстановительных потенциалов для каждой стадии переноса электрона. Два формальных окислительно-восстановительных потенциала для M.capsuiatus (Bath) были определены при использовании индикаторного окислительно-восстановительного титрования и ЭПР спектроскопии. Степень восстановления отслеживалась исключительно по определению значения спина в характерическом ЭПР спектре смешанного валентного состояния. Woodland et аl. сообщал о значениях E'= -106 мВ и E'' = -269 мВ, и значения, полученные Lui, были E' = -196 мВ и E'' = -379 мВ, и в самых последних измерениях было получено E' = -144 мВ и E''= -344 мВ.
В исследованиях M. trichosporium ОВ3b sMMO спектроскопия по Мессбауэру использовалась параллельно с ЭПР-спектроскопией для определения концентрации восстановленных образцов. Титрование M. trichosporium OB3b sMMO привело к формальным потенциалам E'= -168 мВ и E'' = -233 мВ.
Эти данные по окислительно-восстановительным потенциалам sMMO гидроксилазы суммированы в таблице 2. В этом исследовании значения окислительно-восстановительных потенциалов определены для индивидуальных переносов электронов при использовании прямого электрохимического метода с модифицированным золотым электродом. Восстановление полностью окисленной формы Fe(III)Fe(III) до смешанного валентного состояния Fe(II)Fe(III) наблюдается при 4±10, при этом потенциал образования полностью восстановленных Fe(II)Fe(II) образцов был определен как -368±10 мВ (E'). Эти значения отличаются от тех, которые были сообщены ранее, и, как отмечалось выше, отличаются друг от друга (таблица 2). До настоящего времени нет четкого объяснения для этих различий, но приготовление, чистота и активность белков, используемых в каждом исследовании, как впрочем и некоторые морфологические различия между M.richosporium OB3b и M.capsulatus (Batch) гидроксилаз, могут быть существенными. Однако гидроксилазные компоненты этих двух микроорганизмов имели практически одинаковые спектроскопические характеристики и очень сходную гомологию, а также сходные характеристики субстрата. Поэтому должны учитываться и другие возможные источники расхождения электрохимических результатов. Уже указывалось, что возможным источником ошибки является зависимость абсолютного определения значения ЭПР-спектра только для одного из трех окислительно-восстановительных состояний sMMO гидроксилазы как метода определения степени общего восстановления. Другим источником может быть сам метод измерения. Ранее многокомпонентные реакционные смеси составлялись в виде набора, содержащего окислительно-восстановительные красители (для калибровки оси потенциала), дитионит или восстановленный метилвиологен в качестве восстановителя и карбонат железа (III) в качестве окислителя. Взаимодействия, наблюдаемые между белком и этими компонентами, могут влиять на окислительно-восстановительные потенциалы и приводить к сообщениям о специфическом случае взаимодействия окислительно-восстановительного индикатора с гидроксилазой. Дополнительно может наблюдаться температурная зависимость, связанная с этими потенциалами.
ЭПР и измерения по Мессбауэру проводили при 4.2 - 18 K.
Таблица 1 иллюстрирует взаимоотношение между окислительно-восстановительными потенциалами и белком B, связанным с гидроксилазой, в зависимости от степени окисления двухатомного железного центра. Несмотря на различия M. capsulatus (Bath) и M.trichosporium OB3b (таблица 2), константы диссоциации для двух Fe(III)Fe(II). В комплексов сходны, т.е. Kd=2.06 мМ для первого и Kd= 1.7 мМ для последнего. Однако константы диссоциации для комплексов Fe(II)Fe(II). B значительно различаются, т.е. Kd=7.01 мМ для M.capsulatus (Bath) и Kd=500 мМ для OB3b.
Интересные эффекты наблюдаются в том случае, если в растворе для электрохимического восстановления гидроксилазы присутствует белок В'. Сам по себе белок В' не оказывает никакого влияния на окислительно-восстановительное поведение гидроксилазы. Однако в присутствии белка В' белок В становится менее прочно связанным и в результате наблюдается незначительно большая Kd, равная 3.49±0.17 М для Fe(II)Fe(III) формы и 8.5±0.20 М для Fe(II)Fe(II) комплекса. Это означает, что белок В' также связывается с гидроксилазой, но не образует прочного комплекса, что согласуется с данными SPR.
(V) Электрохимия
Прямая электрохимия sMMO гидроксилазных двухатомных железных центров без медиаторов была проведена при использовании модифицированных золотых электродов.
(VI) Использование цитохрома P450cam.
На фиг. 4 представлены циклические вольтамперограммы графитового электрода (1) с гладкой кромкой 15 М цитохрома pH 7.4 и (2) 18 М цитохрома P450cam в 40 мМ натрий фосфатном буфере, pH 7,4, содержащий 1 мМ D-(+)-камфары. Форма фермента, связанного с камфарой, показала ответ сходной формы (фиг. 4) при -390±10 мВ. Эти потенциалы и сдвиг в потенциале при связывании с камфарой на 136 мВ в сторону более положительных значений удовлетворительно согласуются с величинами, ранее определенными на основе потенциометрии, равными -540 мВ и -414 мВ соответственно.
На фиг. 5 представлена зависимость тока катодного пика от скорости сканирования потенциала для связанного с камфарой и не содержащего камфары цитохрома Р450cam. Условия те же, что и на фиг. 1, за исключением того, что температура составляла 18oC. Ток катодного пика проявляет линейную зависимость от квадратного корня скорости сканирования  для связанного с камфарой Р450cam, что свидетельствует о том, что суммарный процесс является диффузионно управляемым.
для связанного с камфарой Р450cam, что свидетельствует о том, что суммарный процесс является диффузионно управляемым.
(VII) Электрохимическое превращение камфары в 5-экзо-гидроксикамфару
После проведения электрохимии цитохрома P450cam фермент использовали для превращения камфары в 5-экзо-гидроксикамфару. Фермент удерживали на поверхности электрода при помощи мембраны Истмана, AQ420. Потенциал изменялся от ±0.2 B до -0.8 B с низкой скоростью сканирования при 50 мВ/с; молекулярный кислород пропускали через раствор, систему периодически исследовали на присутствие 5-экзо-гидроксикамфары. Было обнаружено, что образование 5-экзо-гидроксикамфары увеличивается до 100% после 2 часов.
Хотя данное описание содержит теоретические утверждения, касающиеся механизмов, по которым, как предполагается, протекают определенные описанные здесь процессы, понятно, что данное изобретение не ограничивается данными утверждениями.


Область использования - электрохимия ферментов. Способ переноса электронов между электродом и ферментом в электрохимическом процессе предусматривает вызывание адгезии фермента к электроду, причем перенос является прямым. Ферментом является метанмонооксигеназа, а электрохимию проводят в отсутствие медиаторов. Технический результат - упрощение электрохимического определения концентрации образцов. 3 с. и 14 з.п. ф-лы, 2 табл., 5 ил.
| Экономайзер | 0 |
|
SU94A1 |
| US 4541908 A, 17.09.1985 | |||
| Способ определения концентрации физиологически активных веществ | 1981 |
|
SU1035498A1 |
| WONG L.S., VILKER V.L | |||
| Direct electrochemistry of putidaredoxin at a modified golol electrode | |||
| Journal of electroanalytical chemistry, 1995, v.389, n | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Питательное приспособление к трепальным машинам для лубовых растений | 1922 |
|
SU201A1 |
