Настоящее изобретение относится к новым слитым белкам, которые состоят из ассоциируемого с опухолью таргетирующего элемента, предпочтительно моноклонального антитела или его фрагмента, распознающего молекулу, которая предпочтительно экспрессируется на опухолевых клетках человека, такую как рецептор эпидермального фактора роста (EGFR) и биологически активный лиганд, выбираемый из группы хемокинных белков, предпочтительно из семейства C-X-C. Образующиеся в результате слитые белки могут использоваться для доставки биологически активного лиганда к определенной клетке-мишени или ткани. Новые иммуноконъюгаты могут применяться при лечении опухолей.
Предпосылки создания изобретения
Различные терапевтические концепции используются для лечения раковых больных. Прежде проводили клинические испытания с моноклональными антителами, которые распознают, конкретно или предпочтительно, молекулы на поверхности клетки, экспрессированные на раковых клетках. Целью такого подхода является индукция антителозависимой клеточно-опосредованной цитотоксичности /ADCC/ или комплемент-опосредованной цитотоксичности для уничтожения опухолевых клеток. Другим подходом является цитокинопосредуемая активация иммунного ответа. Индуцированная цитокином противоопухолевая активность может быть опосредована
1/ непосредственным цитотоксическим/цитостатическим действием цитокина на рост опухоли;
2/ опухолеантигенными неспецифическими механизмами, такими как ЛАК-активность или моноцит/гранулоцит - опосредованная цитотоксичность;
3/ опухолеантигенными специфическими иммунными реакциями, опосредуемыми CD4- и CD8 - позитивными Т-клетками.
В такой ситуации у животных-моделей наблюдают системный иммунитет против опухоли.
Однако, цитотоксичность высоких доз цитокинов и недостаточное присутствие in.situ ведут к концепции прицельной антиопухолевой терапии. Принцип прицельной противоопухолевой терапии основывается на физическом сцеплении молекулы-мишени, такой как моноклональное антитело, специфическое для опухолеспецифического антигена, с биологически активной эффекторной молекулой. Доставка эффекторных молекул молекулой-мишенью должна увеличить концентрацию цитокина в опухоли и уменьшить максимальную требуемую дозу. На животных-моделях показано, что in situ присутствие цитокина либо в результате внутриопухолевой инъекции, либо в результате выделения из рассеченных опухолевых клеток может привести к регрессии опухоли /см. обзор в Colombo and Forni, Immunology Today 15: 48-51, 1994/. В таких системах цитокины не ослабляют пролиферацию опухоли, но способны активировать быструю и мощную противоопухолевую реакцию. Следовательно, физическая комбинация эффекторной молекулы и нацеливающего элемента предоставляет средства уменьшения периферического присутствия и усиления наличия внутри опухоли биологически активного лиганда. Кроме того, отдельные опухолевые клетки или микрометастазы также могут быть мишенями для таких молекул.
Биологически активный лиганд для направляемого антителом таргетинга /targeting - конъюгирование лекарственного препарата с антителом, обеспечивающее направленную доставку препарата к ткани или органу-мишени/ должен индуцировать разрушение клетки-мишени либо непосредственно, либо путем создания окружающей среды, летальной для клетки-мишени. Это может быть достигнуто с помощью цитокинов, таких как IL-1, IL-2, IL-4, IL - 6, IL - 7, IL - 10, IL - 13, IFN /интерфероны/, TNF и КСФ-ы. Показано, что эти цитокины добиваются противоопухолевого действия либо непосредственно, либо косвенно - путем активации механизмов защитных сил организма /Mire-Sluis TIBTECH 11: 74-77, 1993; Colombo et al. , Cancer Res.52: 4853-4857, 1992; Thomas & Balkwill, Prarmac. Ther. 52: 307-330, 1991/.
Однако, большинство из этих цитокинов активирует эффекторные клетки, но не проявляет или проявляет очень слабую хемотаксическую активность для них, так что противоопухолевая активность может быть слабой в отсутствие подходящего количества эффекторных клеток в опухолевой ткани.
Однако, хемокины являются хемотаксическими для многих эффекторных клеток и, таким образом, будут усиливать их присутствие в месте опухоли, и, кроме того, они индуцируют ряд функций эффекторных клеток /см., например, Miller and Krangel (1992) "Biology and Biochemistry of the Chemokines: A. Novel Family of Chemotactic and Inflammatory Cytokines", Critical Reviews in Immunology 12, 17/.
IL - 8, MIP 2α /также известный как GRO- β/ и MIP 2 β /GPO- γ/ являются членами суперсемейства хемокинов C-X-C /также известного как суперсемейство малых цитокинов или интеркрины/. Они действуют как хемотаксические факторы и активируют функции эффекторных клеток, и, следовательно, могут представлять оптимальные эффекторные молекулы. Это семейство C-X-C хемокинов является группой недавно охарактеризованных малых белков /8-10 кд/, которые показывают 20-50% гомологию в аминокислотной последовательности и обладают хемотаксической и провоспалительной активностью. IL-8 имеет хорошо определяемую трехмерную структуру /Clore et al., Biochemistry 29: 1689-1696, 1990/ и общий с некоторыми CXC хемокинами N-концевой основной ELR-мотив. Ожидается, что вследствие гомологии последовательностей среди CXC хемокинов вполне подобной будет трехмерная структура. Это уже показано для MCAF/MCP-1 /Cronenborn & Clore Prot. Eng. 4, 263-269, 1991/.
IL-8 в растворе образует стабильные димеры /Clore et al, Biochemistry 29: 1689-1696, 1990/, и возможно, что слитый белок F (ab') IL-8 димеризуется путем взаимодействия двух мономеров IL-8 с образованием двухвалентного иммуноконъюгата. Это будет усиливать взаимодействие слитого белка с антигеном.
Члены группы C-X-C, которые до сих пор описаны, действуют, главным образом, на нейтрофильные гранулоциты. Гены локализованы на хромосоме 4. Членами этой группы являются PF4, основной белок тромбоцита, hlP 10, IL-8, MIP 2α и MIP 2 β.
Действие этих белков на нейтрофилы проявляется в хемотаксической активности, дегрануляции и респираторном всплеске /burst/ ISherry and Cerami, Current opinion in Immunology 3: 56-60, 1991; Oppenheim et al., Annu. Rev. Immunol. 9: 617-648, 1991; Miller and Krangel, Critical Reviewsin Immunology 12: 17-46, 1992; Clark-Lewis et al., J.Biol. Chem., 266: 23128-23134, 1991/.
Члены близкого родственного семейства C-C хемокинов действуют, главным образом, на моноциты. Гены локализованы на хромосоме 17. Такими белками являются LD78, Act-2, MCAF, 1309 и RANTES. Эти молекулы проявляют сильную хемотаксическую активность на моноцитах /Matsushima et al., Chem.Immunol. 51: 236-265, 1992; Oppenheim et al., Annu. Rev. Immunol. 9: 617-648, 1991/.
Фактор эпидермального роста /EGF/ представляет собой полипептидный гормон, который является митогенным для эпидермальных и эпителиальных клеток. Когда EQF взаимодействует с чувствительными клетками, он связывается с мембранными рецепторами /EGFR/. EGFR представляет собой трансмембранный гликопротеин порядка 170 кд и является генным продуктом с-erb-B протоонкогена.
Мышиное моноклональное антитело MAb 425 действует против клеточной линии карциномы A431 человека /ATCC CRL 1555/, и найдено, что оно связывается с полипептидным эпитопом на наружной области EGER. Найдено, что оно ингибирует связывание EGF и служит посредником опухолевой цитотоксичности in vitro и подавляет рост опухолевых клеток эпидермальной и колоректальной, происходящих из карциномы, клеточных линий in vitro /Rodeck et al., 1987, Cancer Res, 47, 3692/. Очеловеченная и химерная версии MAb 425 известны из заявки ВОИС 92/15683.
Таким образом, целью настоящего изобретения является создание антител или их фрагментов, содержащих эпитоп, направленный на антиген EGFR на поверхности опухолевой клетки, и биологически активный лиганд, обладающий высокой хемотаксической активностью к их эффекторным клеткам, и, таким образом, приводящим к низкотоксичной прицельной противоопухолевой терапии. Иммуноконъюгаты представляют собой, следовательно, улучшенный вариант аналогичных иммуноконъюгатов цитокин-антитело, которые подобны по эффективности в отношении их способности вызывать лизис опухоли, но не в отношении возможности эффективно притягивать эффекторные клетки к определенному месту вследствие их хемотаксических свойств.
Краткое изложение сущности изобретения
Изобретение относится к слитым белкам, которые объединяют часть моноклонального антитела - как минимум, сайт узнавания антитела, или полное моноклональное антитело, распознающее эпитом EGFR, с биологически активным лигандом, выбираемым из группы хемокинов, предпочтительно - из семейства C-X-C, в особенности - из IL-8. Конструкции, кодирующие эти слитые белки, генерируют методами технологии рекомбинантных ДНК. Слитые белки содержат вариабельную область тяжелой цепи антитела и CH1-домен константной области /CH1-конъюгаты, Fab - фрагмент/ и соответствующую легкую цепь, или вариабельную область тяжелой цепи антитела и CH1 - и CH2-домен константной области, или вариабельную область тяжелой цепи антитела и CH1-, CH2 и CH3-домен константной области, в каждом случае слитые с биологически активным лигандом. Посредством коэкспрессии с соответствующей легкой цепью может быть генерирован слитый белок, который нацеливается в антигенсекретирующие клетки и доставляет активный лиганд к определенному месту в организме.
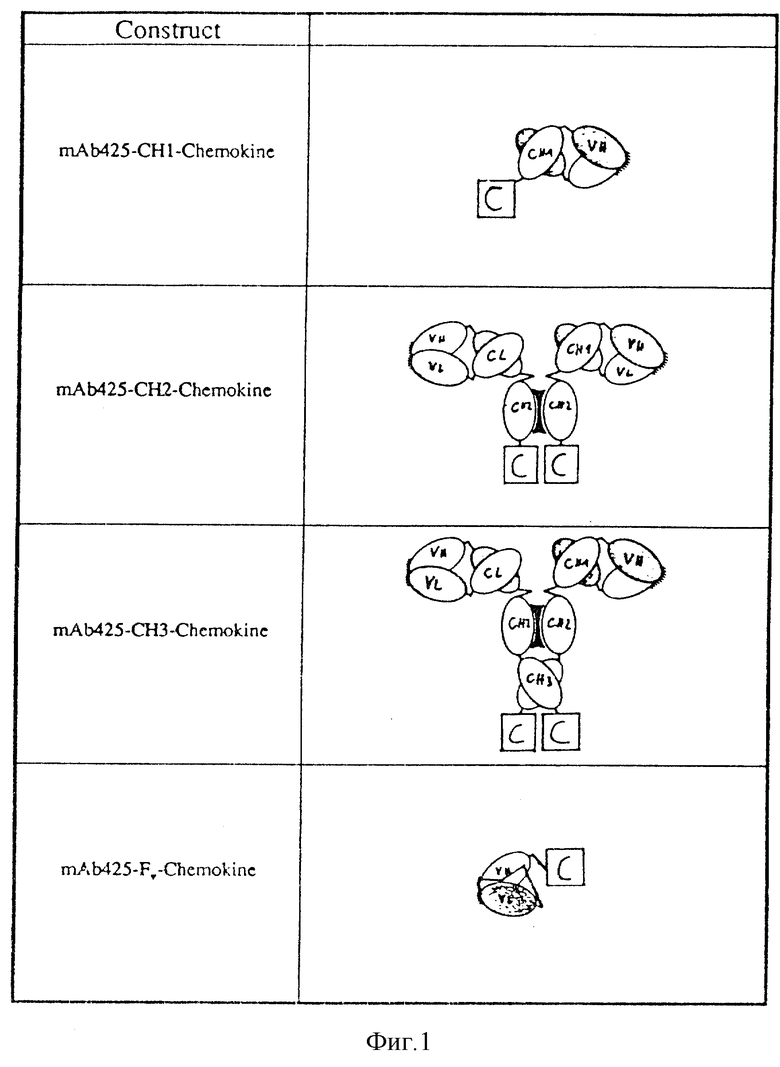
Аналогично, другой иммуноконъюгат может быть получен путем слияния хемокина с C-концом Fv-фрагмента антитела. В этом случае тяжелая и легкая цепи экспрессируются в одном полипептиде, где оба элемента комбинируются соответствующей линкерной последовательностью, чтобы обеспечить должную упорядоченность антигенсвязывающего центра антитела. Различные конструкции приводятся на фиг. 1.
Экспрессия иммуноконъюгатов дает в результате новые молекулы, которые сочетают две функции; во-первых, они попадают в антигенсекретирующие клетки /EGFR/, и во-вторых, они доставляют активный лиганд к определенному месту в организме. Эти лиганды являются сильными хемоатрактантами и активирующими молекулами и дают в результате инфильтрацию эффекторных клеток в месте опухоли и могут вызвать последующее разрушение опухоли.
С помощью иммуноконъюгатов, соответствующих настоящему изобретению, могут быть определены такие опухоли, как меланома, глиома и карцинома, и они могут успешно излечиваться при отсутствии общего заметного токсического действия.
Таким образом, целью настоящего изобретения является предложить иммуноконъюгат, содержащий моноклональное антитело или его фрагмент, направленные на опухолевую клетку, секретирующую эпитоп рецептора фактора эпидермального роста /EGFR/, и белковый лиганд-хемокин, который сливается с упомянутым антителом или фрагментом антитела.
Существуют различные группы хемокинов, такие как хемокины C-X-C и хемокины C-C.
Так, в предпочтительном варианте осуществления настоящего изобретения выбирают белок-хемокин из семейства C-X-C.
В пределах семейства C-X-C предпочтительным вариантом для настоящего изобретения является IL-8.
Следовательно, целью настоящего изобретения является предложить иммуноконъюгат, в котором белок-хемокин выбирают из семейства C-X-C, и он, предпочтительно представляет собой интерлейкин 8 /IL-8/.
Антитела, которые могут использоваться в соответствии с настоящим изобретением, представляют собой либо целые антитела, либо их фрагменты. Подходящими фрагментами являются фрагменты антител Fv, Fab или F(ab') 2/CH1 - фрагменты антител - в соответствии со стилем настоящей заявки/, CH3 - и CH2 - фрагменты антител /фиг. 1/. Предпочтительными вариантами осуществления изобретения являются фрагменты Fv, CH1 -, CH2 - и CH3 - фрагменты антител.
Целью настоящего изобретения, следовательно, является предложить иммуноконъюгат, в котором антитело представляет собой Fab - фрагмент или F(ab') 2 - фрагмент, состоящий, по существу, из вариабельной области тяжелой цепи антитела, CH1 - домена константной области и соответствующей легкой цепи /конъюгат антитело-CH1/, другой иммуноконъюгат, в котором антитело представляет собой фрагмент антитела, состоящий, по существу, из вариабельной области тяжелой цепи антитела, CH1 - и CH2 - доменов константной областей соответствующей легкой цепи /конъюгат антитело- CH2/, еще один иммуноконъюгат, в котором антитело представляет собой полное антитело, состоящее, по существу, из вариабельной области тяжелой цепи антитела, CH1-, CH2- и CH3-доменов константной области и соответствующей легкой цепи /конъюгат антитело - CH3), и, наконец, еще один иммуноконъюгат, в котором антитело состоит, по существу, из вариабельной области тяжелой цепи антитела, соответствующей легкой цепи и полипептидной последовательности, которая связывает легкую и тяжелую цепи /конъюгат антитело - Fv/.
Иммуноконъюгаты по настоящему изобретению могут включать, необязательно, сайт рестрикции между антителом /фрагментом/ и белком-хемокином, который создает возможность вводить в него, например, специфический линкерный пептид, для того, чтобы обеспечить оптимальное связывание конъюгата с эпитопом- мишенью. Соответствующие линкерные пептиды и способы их введения хорошо известны в технике и описываются ниже. В соответствии с изобретением выбирают сайт рестрикции, который является единственным в отдельной ДНК-конструкции. Предпочтительными сайтами рестрикции являются Ncol и Bcll.
Таким образом, еще одной целью настоящего изобретения является предложить иммуноконъюгат, содержащий аминокислоту /последовательность/, которая может быть выведена к сайту рестрикции ДНК между антителом или фрагментом антитела и биологически активным лигандом, причем упомянутый сайт является единственным в полной слитой конструкции.
Еще одной целью настоящего изобретения является предложить иммуноконъюгат, содержащий линкерный пептид между антителом или фрагментом антитела и биологически активным лигандом.
В принципе, подходящими являются все антитела, которые нацеливаются на рецепторы EGF на поверхности опухолевых клеток. Однако, предпочтительным вариантом осуществления изобретения является моноклональное антитело 425.
Кроме того, целью настоящего изобретения является предложить иммуноконъюгат, в котором антитело или фрагмент антитела происходят от мышиного, очеловеченного или химерного MAb 425 и которое выбирают, предпочтительно, из группы, состоящей из (MAb 425)-CH1-1L8, (MAb 425)-CH2-(Ncol)-1L8, (Mab425)-CH2-(BclI)- IL8, (Mab425)-Fv-IL8, (MAb425)-CH3-IL8.
Кроме того, целью настоящего изобретения является предложить способ изготовления иммуноконъюгата, определение которому дается выше и ниже, и в формуле изобретения, путем слияния ДНК- последовательностей, кодирующих антитело или фрагмент антитела, и биологически активного лиганда друг с другом на однонитевой ДНК с помощью олигонуклеотида, который является комплементарным к нужной слитой ДНК-последовательности, помещения получающейся в результате конструкции в вектор экспрессии, который трансформируется в организм-хозяина, культивирования клеток-хозяев в питательном растворе и экспрессии слитого белка.
Иммуноконъюгаты настоящего изобретения являются подходящими для применения при лечении.
Таким образом, целью настоящего изобретения является предложить фармацевтическую композицию, содержащую по крайней мере один из конъюгатов, описанных выше, ниже и в формуле изобретения, и физиологически приемлемый носитель.
Настоящее изобретение имеет целью генерировать слитый белок, состоящий из моноклонального антитела, как прицеливающегося /нацеливающегося/ элемента, и хемокина IL-8 как эффекторной молекулы с хемотаксическими и активирующими свойствами.
В первую очередь можно показать, что IL-8 и другие хемокины /такие как MIP-ы/ сохраняют свою биологическую активность, когда N-конец блокируется дополнительными аминокислотами, такими как составляющая антитела. Следовательно, такая молекула будет пригодна для прицельной противоопухолевой терапии в том отношении, что эффекторные клетки направляются к EGF - рецепторным положительным опухолевым клеткам и активируются in situ.
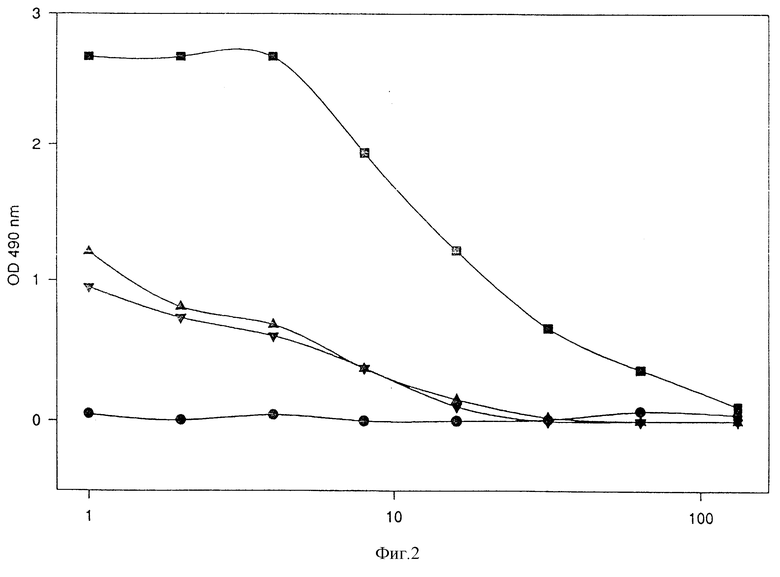
Можно показать, что кДНК, кодирующая тяжелую цепь MAb 425, и белок IL-8 могут быть слиты методами молекулярной биологии, и белки могут быть экспрессированы в подходящих системах экспрессии, и что способность к связыванию EGF-рецепторов сохраняется в слитых белках /фиг. 2/.
Главными клетками-мишенями для IL-8 являются нейтрофильные гранулоциты, которые имеют три биологические функции:
- хемотактическое перемещение вдоль хемотактического градиента;
- выделение накопленных гранул предварительно продуцированными протеолитическими ферментами;
- прямое продуцирование супероксиданионов /респираторный всплеск/.
Соответственно, исследуют слитые белки (MAb 425)/Ncol/ IL-8 и (MAb-425)/BClI/IL-8 на их хемотаксическую активность, индукцию выделения MPO и индукцию выделения супероксида.
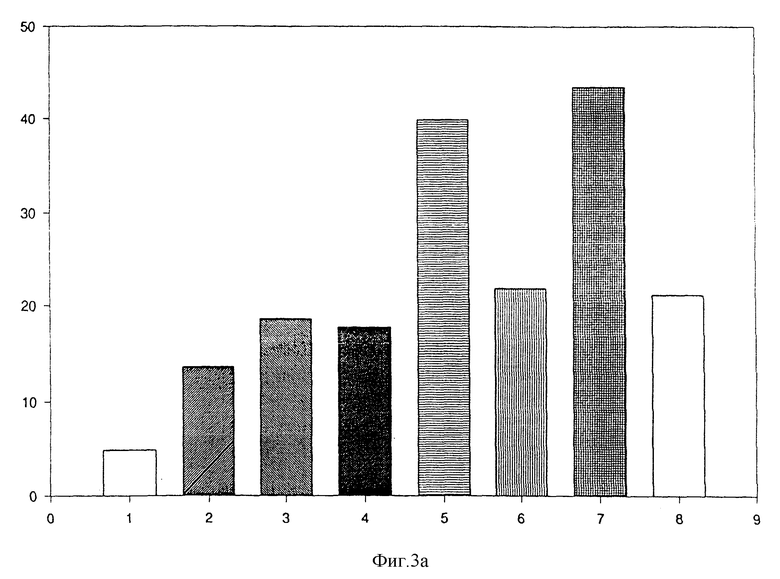
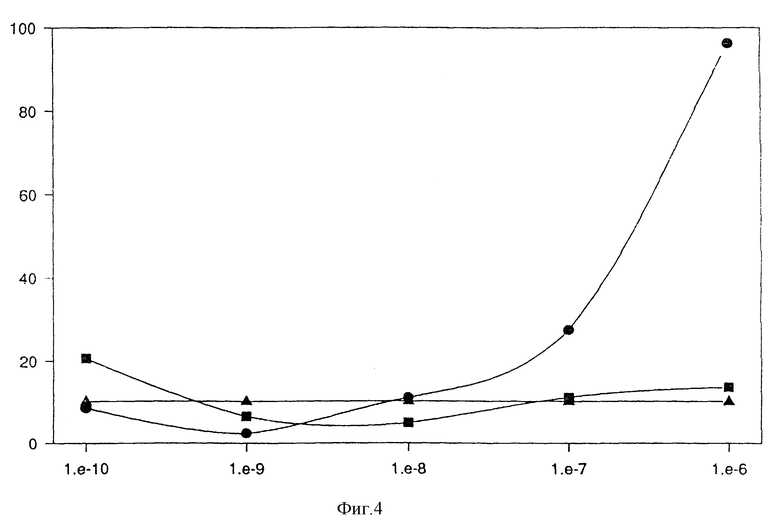
Кроме того, можно показать, что слитые белки, соответствующие изобретению /например, (MAb 425)/ Ncol/IL-8 и (MAb 425)/BclI/IL-8/, вызывают хемотаксическую активность, индукцию выделения МРО и выделения супероксида. Результаты, приведенные на фиг. 3а, показывают, что оба слитых белка являются хемотаксическими для нейтрофилов человека в области рекомбинантного IL-8 /фиг. 3/. Кроме слитых белков (MAb 425) /Ncol/IL-8 и (MAb 425)/BclI/IL-8, которые должны монтировать двухвалентную форму из одновалентной, создают слитый белок F (ab')-IL-8, который экспрессируют в Е.coli и очищают. Фиг. 4 показывает, что слитый белок F(ab')-IL-8 является хемотаксическим для нейтрофилов человека, когда сравнивается с (MAb 425) F (ab'), экспрессированным в E.coli, и, соответственно, очищенным.
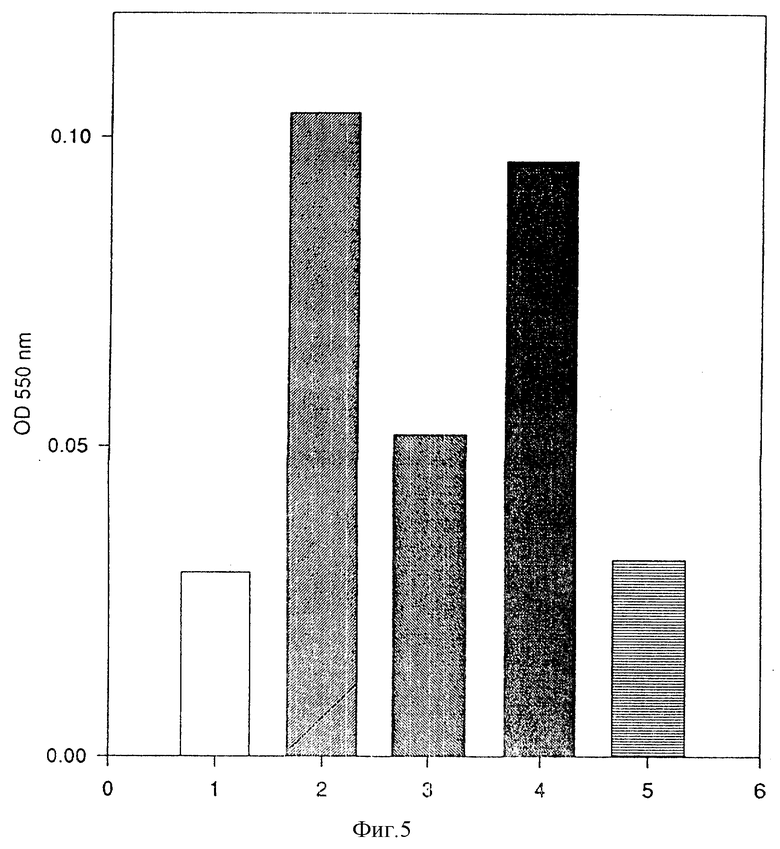
Слитый белок (MAb 425)/IL-8 обладает более сильной способностью к секреции супероксида по сравнению со свободным IL-8 /фиг. 5/, слитый белок (MAb 425)/Ncol/IL-8 менее активен, но величины более высокие, чем контрольные значения /фиг. 5/. Один MAb 425 не показывает активности. Все испытания осуществляют, используя цитохалазин В в качестве энхансера, который один не обнаруживает активности /данные не приводятся/.
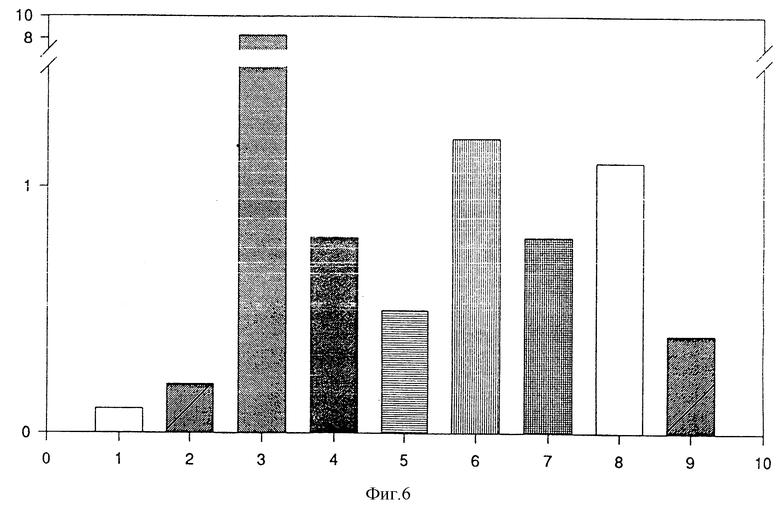
Оба слитых белка индуцируют секрецию миелопероксидазы /MPO/, но слитый белок (MAb 425) /Ncol/IL-8 более активен, чем слитый белок (MAb 425)/IL-8 /фиг. 6/. Все данные вычисляют в соответствии с данными для клеток, лизированных тритоном, которые принимают за 100% содержание фермента. Все данные получают, используя в качестве энхансера цитохалазин В.
Ранее показано, что для связывания рецептора и сигнальной трансдукции требуется N-концевая часть молекулы IL-8 с хорошо сохранившимся мотивом E-L-R. Трехмерная структура IL-8 представляет собой гомодимер, в котором оба N-окончания находятся в доступной конфигурации. Возможно, что биологическая активность IL-8 отменяется, когда N -концы блокируются дополнительными аминокислотами. Поэтому между двумя кДНК вводят сайты рестрикции (Ncol/BclI), чтобы ввести линкерные пептиды, и посредством этого восстановить доступность N -окончаний.
Другие подходы к созданию слитых белков, такие как химическая связь двух элементов, ведут скорее к неопределенным структурам, которые могут изменяться от партии к партии. Кроме того, химическое сочетание может разрушить вторичную структуру лиганда или создать ситуацию, когда большинство белков являются неактивными в отношении связывания рецептора вследствие недоступности. Напротив, подход по настоящему изобретению генерирует слитые белки определенной структуры, которые могут быть экспрессированы с воспроизводимым качеством почти без ограничения.
Суммируя сказанное, слитые протеины по настоящему изобретению обладают следующими свойствами:
- связываются с позитивными клетками EGFR,
- вызывают хемотаксическую активность,
- индуцируют секрецию МРО и супероксида,
- индуцируют лизис опухоли in situ.
Следовательно, иммуноконъюгаты, соответствующие настоящему изобретению, являются подходящими для лечения опухолей.
Краткое описание таблиц и чертежей
Табл. 1
Таблица 1 показывает последовательность праймеров, используемых для PCR (полимеразно/цепьевой реакции) при генерации либо Ncol, либо BclI-сайта при создании слитых белков для эукариотной экспрессии.
Фиг. 1
Модели иммуноконъюгатов антитело-цитокин. С=цитокин; VH = вариабельная область тяжелой цепи; VL = вариабельная область легкой цепи; CH = константная область тяжелой цепи; CL= константная область легкой цепи.
Фиг. 2
Демонстрация MAb 425 в супернатантах с трансфекцией COS -7 ELISA анти-EGF-R;
квадраты: супернатант (MAb 425)-CH3;
треугольники: супернатант (MAb 425)-CH2-(Ncol) - IL -8;
перевернутые треугольники: супернатант (MAb425)-CH2-(BclI)-IL -8;
кружки: супернатант pHCMV;
горизонтальная ось: разбавление супернатантов;
вертикальная ось: оптическая плотность при 490 нм.
Фиг. 3а
Индукция хемотаксиса супернатантами с трансфекцией COS -7:
1 колонка: контрольный DMEM/S;
2 колонка: неразбавленный супернатант pHCMV;
3 колонка: супернатант (MAb 425)-CH3, разбавленный 1:14 /в соответствии с результатами ELISA EGF-r/;
4 колонка: супернатант (MAb 425)-CH3, разбавленный 1:28 /в соответствии с результатами ELISA EGF-r/;
5 колонка: неразбавленный супернатант (MAb 425)-CH2-(BclI)- -IL-8/1.76•10-10 моль/л*/;
6 колонка: супернатант (MAb 425)-CH2-(BclI- 1L-8, разбавленный 1:2;
7 колонка: неразбавленный супернатант (MAb 425)-CH2-(Ncol)- IL-8/2,0 • 1010 моль/л*/;
8 колонка: супернатант (MAb 425)-CH2-(Ncol)-IL-8, разбавленный 1:2, концентрацию IL-8 определяют ELlSA /Amersham/;
вертикальная ось: число клеток в поле подсчета.
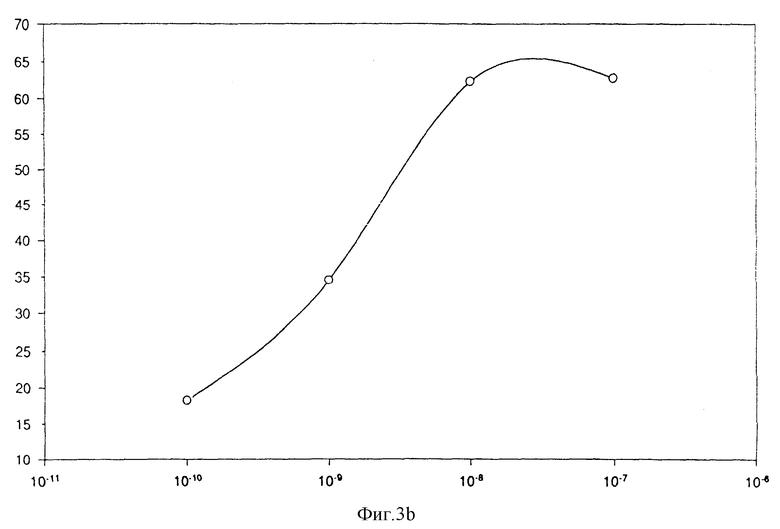
Фиг. 3b
Индукция хемотаксиса очищенным IL-8:
вертикальная ось: число клеток в поле подсчета;
горизонтальная ось: концентрация IL-8 /моль/л/.
Фиг. 4
Индукция хемотаксиса (MAb 425)-CH1-IL-8 /E.coli/:
кружки: (MAb 425)-CH1-IL-8, экспрессированный в Е.coli;
квадраты: (MAb 425)- F (ab'), экспрессированный в E.coli;
треугольники: контрольный, Дульбекко с BSA /бычий сыворотчный альбумин/;
вертикальная ось: число клеток в поле подсчета;
горизонтальная ось: концентрация (моль/л).
Фиг. 5
Индукция секреции супероксида супернатантами с трансфекцией COS -7:
1 колонка: нестимулированные клетки;
2 колонка: IL-8, 10-7 M;
3 колонка: неразбавленный супернатант (MAb 425)-CH2-(Ncol)- IL-8 /2,0 • 1010 моль /л*/;
4 колонка: неразбавленный супернатант (MAb 425)-CH2-(BclI)- IL-8/1,76 • 10-10 моль/л*/;
5 колонка: неразбавленный супернатант (MAb 425)-CH3/О моль/л*
х концентрацию IL-8 определяют ELISA /Amersham/
вертикальная ось: оптическая плотность при 550 нм.
Фиг. 6
Индукция секреции МРО супернатантами с трансфекцией COS-7:
1 колонка: нестимулированные клетки;
2 колонка: клетки, стимулированные цитохалазином В;
3 колонка: IL-8, 10-7 М;
4 колонка: неразбавленный супернатант (MAb 425)-CH3;
5 колонка: супернатант (MAb 425)-CH3, разбавленный 1:2;
6 колонка: неразбавленный супернатант (MAb 425)-CH2-(Ncol)-IL-8;
7 колонка: супернатант (MAb 425)-CH2-(Ncol)-IL-8, разбавленный 1:2;
8 колонка; неразбавленный супернатант (MAb 425)-CH2-(BclI)- IL-8;
9 колонка: супернатант (MAb 425)-CH2-IL-8, разбавленный 1:2;
вертикальная ось: активность в % MPO при сравнении с общим количеством МРО /100%/.
Подробное описание
Общие замечания
Все микроорганизмы, клеточные линии, плазмиды, промоторы, маркеры устойчивости, ориджины репликации, сайты рестрикции или другие фрагменты векторов, которые упоминаются в настоящей заявке, доступны коммерчески или, вообще, из других источников. При условии, что нет иных оговорок, они используются только в качестве примеров, и не существенны, в соответствии с изобретением, и могут быть замещены другими подходящими инструментами и биологическими материалами, соответственно.
Технические приемы, которые, в соответствии с изобретением, являются существенными, описываются подробно ниже. Другие технические приемы, которые подробно не описываются, являются стандартными методами, которые хорошо известны специалистам в этой области техники, или описываются подробно в цитируемых ссылках и патентных заявках и в известной литературе /например, "Antibodies, A Laboratory Manual", Harlow, Lane, Cold Spring Har bor, 1988/.
Моноклональные антитела
MAb 425 представляет собой мышиное моноклональное антитело IgG1, действующее против клеточной линии карциномы человека A 431 /АТСС CRL 1555/. MAb 425 связывается с полипептидным эпитопом поверхностного домена EGF- рецептора человека и конкурирует со связыванием EGF. Обнаружено, что MAb 425 посредничает в цитотоксичности опухоли in vitro и подавляет рост опухолевых клеток клеточных линий, происходящих из эпидермоидной и колоректальной карциномы, in vitro /Rodeck et al, Cancer. Res. 47: 3692, 1987/. Гуманизированная и химерная версии MAb 425 раскрываются в заявке ВОИС 92/15683.
Хемокины
Кодирующие хемокины кДНК либо закупаются в British Biotechnology Limited / человеческий IL-8 BBG, 44: Herrmann Biermann GmbH, Bad Nauheim FRG/, либо генерируются из РНК, выделенных из продуцирующей цитокин человеческой клеточной линии U 937 /АТСС CRL 1593/. Полную РНК из продуцирующих хемокин клеток выделяют c RNAZOl /WAK-Chemie, Германия/ в соответствии с рекомендациями изготовителя. Потом РНК транскрибируют в кДНК, и кодирующие хемокин последовательности усиливают PGR, используя подходящие праймеры, выведенные из опубликованных ДНК-последовательностей.
Векторы
pUC 19 является частью серии клонирующих векторов плазмиды E.coli с относительно высокой копийностью и содержит сегменты pBR322 и M13mp19. pUC 19 содержит индуцибельный бактериальный lac промотор-оператор с последующим сайтом множественного клонирования /Yanisch-Perron et al., Cene 33: 103-109, 1985/. Векторы pUC коммерчески доступны /напpимер, New England Biolabs/.
Фагемидные вeктopы pBluescipt KS/SK + KS/SK - получают из pUc 19. Векторы являются коммерчески доступными /Stratagene, Heidelberg/. Векторы прокариетной экспрессии основываются на векторе pSW1 /Ward et al, Nature 341: 544-546, 1989/, который является производным вектора pSW1/Ward et al., Nature 341: 544-546, 1989/, который является производным вектора pUC 19. pSW1 содержит последовательность, кодирующую лидерный пептид гена бактериального pelB из Erwina carotovora /Lei et al., J Bact, 169: 4379-4383, 1987/. Чужеродные ДНК могут быть введены в рамку позади лидерной последовательности для направления экспрессии белка в периплазму.
Вектор эукариотной экспрессии pHCMV /Gillies et al., Cell 33: 717, 1983/ содержит ориджин репликации обезьяньего вируса 40 /SV40/ и область промотора и энхансера цитомегаловируса человека. За областью промотора/энхансера следует сайт множественного клонирования для интродукции генов, которые экспрессируют. В этом векторе соединяют химерную форму вариабельной области тяжелой цепи MAb 425 и область c γ 1CH2, слитую с хемокином на конце CH2-домена, чтобы генерировать слитый белок с тяжелой цепью MAb 425. Цепь слитого 1g может быть собрана в иммуноконъюгат путем объединения ее с соответствующей легкой цепью с образованием области одновалентного связывания антигена, которая затем может быть ассоциирована с образованием двухвалентного иммуноконъюгата, специфического для антигена- мишени. Конструкции с тяжелой и легкой цепями могут размещаться в одном или в раздельных векторах.
Экспрессия слитых белков в эукариотных клетках
Конструирование векторов эукариотной экспрессии для экспрессии слитого белка (fab 425)-хемокин
Слияние MAb-425 и хемокинов по технологии PCR
Константную область человеческого c γ 1 вставляют в pUC в качестве фрагмента Bam H1/BamH1. Константная область c γ 1 содержит два SaclI-сайта: один располагается в 5'-интроне, 40 пар основ, в прямом направлении сайта 5'-BamHI, и другой располагается в прямом направлении сайта 5'-BamHI. 580 пар осн., и в обратном направлении начала CH3-домена, 140 пар основ. Второй SaclI-сайт является подходящим для дальнейшего субклонирования, и, таким образом, первый SaclI-сайт разрушается введением сайта SnaBI с адаптором. В этой конструкции / Δ SaclI c γ 1/ фрагменты в прямом направлении из сайта SaclI могут легко обмениваться.
Человеческий IL-8 вырезают из вектора pUC 18 /BglII /Eco R1/ и вставляют в pBluescript Sk+IStratagene GmbH, Heidelberg/ (Smal/EcoR1/, таким образом, что BgeII и Smal -сайт утрачиваются. Фрагмент SaclI/Xba1 клона Δ SaclI Cγ1 вставляют в pBlueskript SK+. Оба гена усиливают подходящими праймерами, используя технологию PCR:
для Δ Sacl1 c γ 1: праймер 3:концевая последовательность CH2-домена и Ncol-сайта;
праймер 5': праймер обратного секвенирования;
- для IL-8: праймер 3': праймер универсального секвенирования;
праймер 5': сайт Ncol и начальная последовательность IL-8.
Продукты разрезают и легируют SK+ SoclI/EcoRI. В получающейся в результате пептидной последовательности C-концевой лизин CH2-домена заменяют метионином, и N-концевой серии части IL -8 заменяют глицином.
Вновь генерированная последовательность в области соединения двух полипептидов имеет вид;
5' AAA GCC ATG GGT GCT 3'
Lys Ala Met GLy Ala
Cγ1CH2 <--- --->IL-8/2-72/.
Такую же процедуру осуществляют, используя праймеры /таблица/ для введения сайта BclI между двумя генами. Получающийся в результате слитый ген имеет сайт BclI между геном константной области c γ 1 и геном IL-8, кодируя полную последовательность CH2-домена, две дополнительных аминокислоты /валин, изолейцин/ и последовательность IL-8 без первых двух аминокислот /серин, аланин/.
Вновь генерированная последовательность в области соединения двух полипептидов имеет вид:
5' GCC AAA GTG ATC AAA CAAA 3'
Ala Lys Val Ile Lys Glu
c γ 1CH2 <--- ---> IL-8/3-72/.
Продукты PCR субклонируют в SK+, используя сайты рестрикции SaclI и EcoR1. Для эукариотной экспрессии эти слитые гены клонируют в вектор pHCMV.
Экспрессия иммуноконъюгатов в эукариотных клетках
Экспрессия иммуноконъюгатов в эукариотных клетках требует введения в клетки-хозяева векторной ДНК, содержащей тяжелую и легкую цепи. Описан ряд различных способов, таких как электропорация, использование диэтиламиноэтилдекстрана, фосфата кальция, липофектина или слияние протопластов. Может быть использован любой тип клетки-хозяина, при условии, что последовательности рекомбинантных ДНК, кодирующие иммуноконъюгат, транскрибируются должным образом в мРНК в этом типе клеток. Клетки-хозяева могут представлять собой клетки миеломы мыши, которые не продуцируют иммуноглобулин, такие как Sp2/O-AG14/ATCC CRL 1581/, P3X63Ag-8.53 /ATCC CRL 1580/, или клетки хомячка, такие как CHO-K1 /ATCC CCL 61/, или
CHO/DHFR-/ATCC CRL 9096/, или
ВНК-21 /ATCC CCL 10/. Для переходной экспрессии могут быть использованы COS-1 /ATCC CRL 1650/ или COS -7 /ATCC CRL 1651/.
Кратковременная экспрессия иммуноконъюгатов
Вектор экспрессии pHCMV содержит сайт инициации репликации обезьяньего вируса SV40. Линия клеток COS-7 является производной линии клеток обезьяны CV-1, которая трансформирована дефектным по сайту инициации репликации вирусом SV40. Следовательно, плазмиды, содержащие сайт инициации-репликации вируса SV40, будут амплифицированы, и будет улучшаться выработка иммуноконъюгатов. Супернатанты собирают спустя 72 часа, и определяют связывание EGF- рецептора и концентрацию хемокина методом EUSA.
Перманентная экспрессия иммуноконъюгатов
Векторы, содержащие рекомбинантные конструкции для экспрессии иммуноконъюгатов, вводят в подходящие клетки-хозяева. Конструкции с тяжелой и легкой цепью могут быть помещены в одном и том же или в раздельных векторах; в последнем случае оба вектора могут нести идентичный селекционный маркер, такой как устойчивость к неомицину /Neomycin resistance или дегидрофолатредуктаза /DHER/, или два различных селекционных маркера, которые отбирают для присутствия из обоих векторов. Селекция для маркера DHER может быть осуществлена только в линиях отрицательных клеток DHER, таких как CHO/DHFR. Смешанные популяции для экспрессии иммуноконъюгатов анализируют ELISA для специфического EGF-рецептора. Дальнейшую селекцию для положительных моноклональных антител осуществляют посредством клонирования при низкой плотности клеток.
Очистка иммуноконъюгатов (MAb 425)-хемокин
Иммуноконъюгаты MAb 425, продуцированные клеткой-хозяином, могут быть собраны и очищены любым подходящим способом, таким как аффинная хроматография, с использованием антигена-мишени, антицитокинных антител или антиидотипических антител /например, Harlow, Lane, цит./выше/.
В настоящей работе очистка достигается с помощью антиидиотипических антител, которые продуцируют из MAb 425 стандартными способами /например, Kostelny et al.(1992), J.Immunol, 148, 1547/.
Чтобы получить чистые иммуноконъюгаты Fv, штаммы E.coli, подходящие для экспрессии белка, трансформируют плазмидами экспрессии /см. ниже/. Клетки выращивают до OD578 =0.5, и индуцируют изопропил- β -D-тиогалактопиранозидом /IPTG/ /1 мМ/. Клетки выращивают в течение ночи, и собирают супернатанты и клетки. Супернатант помещают в колонку с антиидиотопическим антиMAb 425, подготовленную в соответствии с обычными методиками. Колонку промывают забуференным фосфатом 0.5 М NaCl, и связанные белки элюируют 100 мМ глицина с 0,5 М NaCl при pH 2,5. Элюат сразу же нейтрализуют трис. 2.5 М, pH 8,0. Фракции, содержащие (MAb 425)-CH1-IL-8, соединяют, концентрируют и диализуют против ЗФР.
Конструирование векторов прокариотной экспрессии для (Fаb 425)-хемокина, и экспрессия слитого белка Fv-хемокин
Фрагмент Fv строят по Glockshuber et al. /Biochemistry - 29: 1362-1367, 1990/. ДНК-последовательности, кодирующие легкую цепь и Fd-фрагмент тяжелой цепи фрагмента Fv, введены в сайт множественного клонирования вектора pSW1. Зрелая кодирующая последовательность легкой цепи, зрелая кодирующая последовательность тяжелой цепи и кодирующая последовательность Fv предшествуют лидерному пептиду бактериального pel B гена. Кодирующая последовательность тяжелой цепи содержит сайт Ncol /конец 3'/. Кодирующие хемокин кДНК модифицируют PCR, чтобы ввести сайты рестрикции Ncol (конец 5') и Notl /конец 3' /или EcoR1/для вставки Fv/. Гены хемокина вставляют в рамку непосредственно в CH1-домен тяжелой цепи фрагмента Fv. С другой стороны, линкерный пептид, такой как (Gly-Gly-Gly-Gly-Ser)x, где x может иметь значения от 1 до 4, может быть вставлен между CH1-доменом и геном хемокина. Такие линкеры и способы их получения известны из литературы /например, Curtis et al. 1991, Proc. Natl, Acad. Sci. U.S.A. 88, 5809/.
Эти векторы облегчают эффективную экспрессию функциональных слитых белков F(ab') (=CH1) и Fv - хемокин в E.coli. Слитые белки хемокина с легкой и тяжелой цепью располагаются на отдельной дицистронной матричной PHK, поставленной под контроль индуцибельного lac промотора /Skerra and Plucrthun, Science 242: 1038-1040, 1988/. Следовательно, экспрессия слитого белка Fab/Fv может быть индуцирована в соответствии с условиями культивирования. Трансляция обоих белков из дистронной матричной РНК благоприятствует синтезу равного количества слитого белка Fd - хемокин и легкой цепи, увеличивая, таким образом, шансы для правильной сборки функциональных слитых белков Fdb/Fv. Два полипептида секретируются в периплазму E.coli, где происходит укладка, образование дисульфидных связей и сборка слитого белка Fab 425CH1/Fv. Длительное культивирование бактерий ведет к частичной проницаемости наружной мембраны E. coli, создавая возможность для диффузии слитых белков в культуральную среду.
Связывающие свойства иммуноконъюгатов Mab 425
Связывающие свойства иммуноконъюгатов MAb 425 определяют ELISA для специфического EGF-рецептора. В нескольких словах, титрационные микропланшеты покрывают слоем очищенного EGF-peцептора в течение ночи при 4oC. Микропланшеты инкубируют с супернатантами, содержащими слитый белок, или с супернатантами, содержащими фрагменты неконъюгированного MAb. Микропланшеты промывают, чтобы удалить несвязанный материал, и определяют связанное с EGF-рецептором антитело посредством инкубации с козьим античеловеческим IgG и IgM /с тяжелой и легкой цепью/, конъюгированным с пероксидазой, в субстрате. Количество связанного с EGF-рецептором определенного белка определяют измерениями при 490 нм.
Биологическая активность иммуноконъюгатов (Mab 425)-IL-8
Изоляция эффекторных клеток
Для определения биологической активности нейтрофильные гранулоциты периферической крови человека изолируют из цельной крови здоровых доноров непосредственно перед определением, как описано ранее в Haslett et al. /Am. J. Pathol. 119: 101-110, 1985/. Отделяют плазму центрифугированием, эритроциты - осаждением с декстраном, и наконец, лимфоциты и лейкоциты отделяют центрифугированием в градиенте перколла. Выделенные нейтрофилы используют сразу же.
Определение хемотаксической активности
Определение хемотаксиса осуществляют в соответствии с Falk et al. /J. Immunol. Methods. 33: 239-247, 1980/. Коротко говоря, используют 48-ячеичную камеру Бойдена и 5- мкм-мембраны. Очищенные нейтрофилы ресуспендируют в модифицированной по способу Дульбекко среде Игла /DMEM/ /DMEM/; 1% пенициллина, 1% стрептомицина, 10% FCS /фетальная телячья сыворотка/, 2 мМ L- глутамина, 1 мМ пирувата Na, 10 мМ HEPES/ при концентрации 1 • 106 клеток/мл. Нижние части загружают супернатантами, содержащими слитый белок, или контрольными супернатантами, накрывают мембраной, и, в заключение, верхние части ячеек загружают суспензией клеток. После инкубации при 37oC в течение 30 минут мембраны удаляют и фиксируют в 2% глутардиальдегиде в течение 10 минут. Затем связанные с мембраной клетки покрашивают гематоксилином железа по Вейгерту /Weigert/ /сигма-диагностика/ в течение трех минут. Число клеток, связанных с мембраной, определяют с помощью микроскопа.
Определение способности индуцировать выделение фермента в нейтрофилах
Чтобы оценить способность иммуноконъюгатов на нейтрофилах, индуцировать выделение гранул, контролируют активность миелопероксидазы в супернатанте /Henson et al. , J. Immunol. 121: 851, 1978/. Испытания проводят в 96-луночных титрационных микропланшетах с 5 • 105 клетками на лунку. После инкубации /37oC/ со стимулами микропланшеты центрифугируют, и бесклеточный супернатант переносят в другой 96-луночный титрационный микропланшет. Бесклеточные супернатанты инкубируют с динизидином /в качестве субстрата/ и измеряют оптическую плотность при 492 нм. В качестве позитивного контроля используют FMLP с концентрацией 10-7. Для определения общего содержания фермента клетки без стимулов лизируют тритоном. Активность вычисляют как процент от общего содержания фермента /лизис/.
Определение способности выделять супероксид
Цитохром восстанавливают с О2-, и посредством этого изменяют его оптическую плотность. Изменение оптической плотности является ценным указателем при оценке супероксидной активности. Испытания проводят по Guthrie et al. /J. Exp. Med. 160: 1656-1671, 1984/ в 96-луночных титрационных микропланшетах с 5 • 105- клетками на лунку. После инкубации со стимулами и цитохромом с микропланшеты центрифугируют, и определяют оптическую плотность супернатантов при 550 нм.
Другие иммуноконъюгаты
В соответствии с приведенным выше описанием получают и исследуют иммуноконъюгаты против EGFR-CH1, -CH2-, CH3 и -Fv /с или без сайта рестрикции и линкера/, включающие в качестве хемокинной компоненты MIP-2 α и MIP-2β. Эти конструкции проявляют свойства, подобные свойствам производных IL-8.
Терапевтическое применение иммуноконъюгатов
Иммуноконъюгаты по настоящему изобретению могут вводиться пациентам-людям в лечебных целях. Следовательно, целью настоящего изобретения является предложить фармацевтическую формулировку, содержащую в качестве активного ингредиента по крайней мере один слитый белок, определение которому дается выше и в формуле изобретения, в сочетании с одним или несколькими фармацевтически приемлемыми и подходящими для него носителями, эксципиентами или разбавителями.
Как правило, иммуноконъюгаты настоящего изобретения будут вводиться внутривенной или парентеральной инъекцией. Обычно интервалы дозировки при введении иммуноконъюгатов являются достаточно широкими, чтобы получить нужный эффект подавления и разрушения опухоли. Дозировка будет зависеть от возраста, состояния, пола и тяжести заболевания у пациента и может изменяться от 0,1 мг/кг до 200 мг/кг, предпочтительно от 0,1 мг/кг до 100 мг/кг, в одной дозе при ежедневном введении одной или нескольких доз в течение одного или нескольких дней.
Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и приемлемые для инъекций сложные органические эфиры, такие как этилолеаты, и другие известные в технике растворители, подходящие для таких целей. Иммуноконъюгаты настоящего изобретения могут использоваться в композиции, содержащей физиологически приемлемый носитель. Примерами таких подходящих носителей является физиологический раствор, ЗФР, раствор Рингера или лактированный раствор Рингера. В фармацевтических формулировках могут также присутствовать консерванты и другие добавки, такие как антибиотики, антиоксиданты и хелатообразователи.
Фармацевтические формулировки настоящего изобретения являются подходящими для лечения всех разновидностей опухолей, включая меланомы, глиомы и карциномы, так же, как и для лечения кровяных опухолей и твердых опухолей.

Изобретение относится к медицине и касается иммуноконъюгатов, обладающих противоопухолевой активностью, и способа их получения. Сущность изобретения включает иммуноконъюгат, представляющий собой моноклональное антитело или его фрагмент, специфичное для молекулы рецептора EGF человека и представителя семейства хемокинов, а именно интерлейкина-8. Способ включает в себя слияние ДНК-последовательностей, кодирующих моноклональное антитело и фрагмент антитела, специфичного для молекулы рецептора EGF человека и биологически активного лиганда, получение комплементарной ДНК, включение ее в экспрессирующий вектор, трансформацию указанным вектором клеток хозяина и их культивирование в питательной среде в условиях, при которых экспрессируется указанный иммуноконъюгат. Полученные иммуноконьюгаты индуцируют цитотоксическую активность. 3 с. и 6 з.п.ф-лы, 6 ил., 1 табл.
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| 0 |
|
SU235805A1 | |
| Шланговое соединение | 0 |
|
SU88A1 |
| Огнетушитель | 0 |
|
SU91A1 |

