[0001] Изобретение относится к слитым белкам интерлейкина-7 (IL-7), способам их получения и к их применению. Слитые белки включают иммуноглобулиновую часть, слитую непосредственно или опосредованно с IL-7, которая может быть модифицирована в специфических положениях по сравнению с IL-7 дикого типа для того, чтобы улучшить биологические и фармацевтические свойства. Белки в соответствии с изобретением являются особенно полезными при лечении расстройств, сопровождающихся иммунодефицитами, и, в частности, заболеваний, которые вовлекают дефициты Т-клеток.
Предпосылки изобретения
[0002] Разнообразные расстройства и способы лечения вовлекают дефицит иммунных клеток. Например, инфекция ВИЧ приводит к потере CD4+ Т-клеток, в то время как способы лечения, такие как химиотерапия и радиационная терапия, обычно приводят к потере широкого разнообразия клеток крови. Были сделаны попытки обеспечить специфические белковые препараты, которые могут пополнить специфические типы иммунных клеток, теряемых в результате заболевания или терапии. Например, в раковой химиотерапии эритропоэтин используется для пополнения красных клеток крови, колониестимулирующий фактор гранулоцитов (G-CSF) используется для пополнения гранулоцитов, а колониестимулирующий фактор гранулоцитов и макрофагов (GM-CSF) используется для пополнения гранулоцитов и макрофагов. Эти белковые лекарственные средства несмотря на то, что являются полезными, имеют относительно короткий период полураспада в сыворотке крови, так что пополнение иммунных клеток часто является неэффективным. Более того, в клиническом применение в настоящее время не существует никакого специфического лечения для специфической стимуляции развития Т- или В-клеток, даже несмотря на то, что потеря этих клеток в результате заболевания или после определенных миелоабляционных способов лечения известна как явление, которое является особенно вредным для здоровья пациента. Таким образом, существует потребность в области техники, которая заключается в развитии стимуляторов и восстановителей иммунной системы, в частности лимфоцитов, которые имеют продолжительный период полураспада в сыворотке крови.
Короткое изложение сущности изобретения
[0003] Настоящее изобретение направлено на слитые белки интерлейкина-7 (IL-7), которые имеют улучшенные биологические свойства по сравнению с соответствующими белками IL-7 дикого типа. Более того, настоящее изобретение основывается, частично, на обнаружении того факта, что слитые белки IL-7, обладающие особенными структурными свойствами, имеют улучшенные биологические свойства по сравнению с рекомбинантным IL-7 дикого типа.
[0004] Соответственно, в одном аспекте изобретение характеризует белок слияния, который включает одну часть, содержащую цепь иммуноглобулина (Ig), и вторую часть, содержащую интерлейкин-7 (IL-7), где слитый белок IL-7 имеет повышенную биологическую активность, такую как длительный период полураспада в сыворотке крови или улучшение выживания или роста иммунных клеток по сравнению с IL-7 дикого типа.
[0005] В одном воплощении изобретение характеризует белок слияния, который включает одну часть, содержащую цепь иммуноглобулина (Ig), и вторую часть, содержащую IL-7, где аминокислотные остатки в положениях 70 и 91 IL-7 являются гликозилированными, а аминокислотный остаток в положении 116 IL-7 является негликозилированным. Повсюду в этом документе аминокислотные положения IL-7 относятся к соответствующим положениям в последовательности зрелого человеческого IL-7. В одном воплощении аминокислотный остаток в положении 116 IL-7 представляет собой аспарагин. В другом воплощении аминокислотный остаток в положении 116 IL-7 является измененным таким образом, что он не служит сайтом гликозилирования. В одном воплощении остаток IL-7 включает дисульфидную связь между Cys2 и Cys92, Cys34 и Cys129, и Cys47 и Cys141 IL-7.
[0006] В другом воплощении изобретение предусматривает слитый белок, содержащий одну часть, которая включает цепь Ig, и вторую часть, включающую IL-7, где IL-7 включает дисульфидную связь между Cys2 и Cys92, Cys34 и Cys129, и Cys47 и Cys141 IL-7. В одном воплощении аминокислотный остаток в положении 116 IL-7 является негликозилированным. В другом воплощении аминокислотный остаток в положении 116 IL-7 представляет собой аспарагин или является измененным таким образом, что он не служит сайтом гликозилирования. В другом воплощении аминокислотные остатки в положениях 70 и 91 IL-7 являются гликозилированными.
[0007] Цепь Ig обычно представляет собой интактное антитело или его часть, такую как участок Fс. Цепь Ig слитого белка IL-7 может иметь происхождение от любого известного изотипа Ig и может включать, по крайней мере, часть одного или более константных доменов. Например, константный домен может быть выбран из группы, которая состоит из участка СН1, шарнирного участка антитела, участка СН2 и участка СН3. В одном воплощении остаток Ig включает шарнирный участок, участок СН2 и участок СН3. Цепь Ig необязательно является связанной с частью IL-7 с помощью линкера.
[0008] Остатки Ig одного изотипа антитела, такие как IgG1 или IgG2, и гибридные участки Ig разрешаются в данном изобретении. Например, в одном воплощении остаток Ig включает шарнирный участок, который имеет происхождение от одного изотипа (то есть IgG2), и участок СН, полученный от другого изотипа (то есть IgG1). Цепь Ig, включающая часть Fc IgG1, может преимущественно быть модифицирована для включения мутаций Asn297Gln и Тyr296Аlа. Кроме того, цепь Ig, включающая часть Fc IgG2, может быть преимущественно модифицирована для включения мутаций Asn297Gln и Phe296Ala.
[0009] IL-7 часть слитого белка IL-7, описанного выше, может включать зрелую часть IL-7 части. В одном воплощении IL-7 часть может дополнительно включать делецию, такую как внутренняя делеция. В одном примере IL-7 может включать делецию восемнадцати аминокислот от аминокислоты 96 до 114 SEQ ID NO: 1.
[0010] В других воплощениях изобретение включает очищенные нуклеиновые кислоты, кодирующие белки слияния IL-7, описанные выше, и культивируемые хозяйские клетки, включающие эти нуклеиновые кислоты.
[0011] В другом аспекте изобретение включает способ получения слитого белка IL-7, включающего экспрессию в хозяйской клетке нуклеиновой кислоты, описанной выше, и выделение слитого белка.
[0012] В другом аспекте изобретение включает композицию, такую как фармацевтическая композиция, содержащая слитый белок, описанный выше.
[0013] В другом аспекте изобретение включает способ лечения пациента путем введения слитых белков Fc-IL-7.
Краткое описание чертежей
[0014] Фигура 1 показывает аминокислотную последовательность человеческого IL-7 (SEQ ID NO:1). Сигнальная последовательность представлена жирным шрифтом. Восемнадцать аминокислот, которые могут быть делетированы из IL-7 последовательности, представлены жирным шрифтом и курсивом.
[0015] Фигура 2 показывает аминокислотную последовательность коровьего IL-7 (SEQ ID NO:2). Сигнальная последовательность представлена жирным шрифтом.
[0016] Фигура 3 показывает аминокислотную последовательность бараньего IL-7 (SEQ ID NO:3). Сигнальная последовательность представлена жирным шрифтом.
[0017] Фигура 4 показывает аминокислотную последовательность зрелого человеческого Fcγ1-IL-7 (SEQ ID NO:4).
[0018] Фигура 5 показывает аминокислотную последовательность зрелого человеческого Fcγ2(h) (FN>AQ)-IL-7 (SEQ ID NO:5).
[0019] Фигура 6 показывает аминокислотную последовательность зрелого человеческого Fcγ1 (линкер 1)-IL-7 (SEQ ID NO:6).
[0020] Фигура 7 показывает аминокислотную последовательность зрелого человеческого Fcγ1 (YN>AQ) (линкер2)-1Е-7 (SEQ ID NO:7).
[0021] Фигура 8 показывает аминокислотную последовательность зрелого человеческого Fcγ1 (YN>AQ,d) (линкер2)-IL-7 (SEQ ID NO:8).
[0022] Фигура 9 представляет собой последовательность нуклеиновой кислоты для участка Fc человеческого Fcγ1-IL-7 (SEQ ID NO:22).
[0023] Фигура 10 представляет собой последовательность нуклеиновой кислоты для участка Fc человеческого Fcγ1 (YN>AQ)-IL-7 (SEQ ID NO:21).
[0024] Фигура 11 представляет собой последовательность нуклеиновой кислоты для участка Fc человеческого Fcγ2(h)-IL-7 (SEQ ID NO:20).
[0025] Фигура 12 представляет собой последовательность нуклеиновой кислоты для участка Fc человеческого Fcγ2(h) (FN>AQ)-IL-7 (SEQ ID NO:19).
[0026] Фигура 13 представляет собой графическое изображение фармакокинетического профиля рекомбинантного человеческого IL-7 (незаштрихованные квадраты) и слитого белка Fcγ2(h) (FN>AQ)-IL-7 (незаштрихованные ромбы) Примера 7. Концентрации в сыворотке введенных слитых белков IL-7 (в нг/мл) измеряли в течение времени (в часах).
[0027] Фигура 14 представляет собой графическое изображение восстановления В-клеток у облученных мышей с пересаженным костным мозгом и обработанных с помощью рекомбинантного человеческого IL-7 (незаштрихованные символы), человеческого Fc-IL-7 (заштрихованные символы) или ФБР (X). Белки вводили через день (квадраты) и один раз в неделю (треугольники). Пунктирная линия представляет концентрацию В-клеток у донорной мыши.
[0028] Фигура 15 представляет собой графическое изображение восстановления Т-клеток у облученных мышей с пересаженным костным мозгом и обработанных с помощью рекомбинантного человеческого IL-7 (незаштрихованные символы), человеческого Fc-IL-7 (заштрихованные символы) или ФБР (X). Белки вводили через день (квадраты) и один раз в неделю (треугольники). Пунктирная линия представляет концентрацию Т-клеток у донорной мыши.
[0029] Фигура 16 представляет собой точечную схему, иллюстрирующую популяции лимфоцитов образцов крови (верхний ряд) и селезенки (нижний ряд) у облученных мышей с пересаженным костным мозгом и обработанных с помощью huFcγ2(h) (FN>AQ)-IL-7 (первые две колонки), необработанных контролей (последняя колонка). Первая колонка представляет восстановленные эндогенные лимфоциты (CD45.2+), а вторая колонка представляет восстановленные донорные лимфоциты (CD45.1+). Т-лимфоциты определяли как CD3 позитивные клетки, показанные в расположенном нижнем правом квадранте. В-лимфоциты определяли как В220 позитивные клетки, показанные в расположенном верхнем левом квадранте.
Подробное описание изобретения
[0030] Изобретение обеспечивает слитые белки IL-7, которые обладают улучшенной биологической активностью по сравнению с белками IL-7 дикого типа. В частности, изобретение обеспечивает слитые белки IL-7, которые включают иммуноглобулиновую часть (Ig). Эти Ig-IL-7 слитые белки обладают улучшенной биологической активностью, такой как более продолжительный период полураспада в сыворотке крови по сравнению с белками IL-7 дикого типа, что делает их приемлемыми для применения при лечении состояний, сопровождающихся дефицитом иммунных клеток, таких как дефициты лимфоцитов.
[0031] Изобретение также частично основывается на открытии того факта, что слитые белки IL-7, которые обладают особыми структурными характеристиками, также имеют улучшенные биологические свойства. Несмотря на то что аминокислотная последовательность IL-7 млекопитающих является хорошо известной, информация о структуре белков IL-7, имеющих происхождение от эукариотических клеток, в том числе, например, каким образом происходит сборка белков и каким является влияние его предсказанных N-связанных сайтов гликозилирования на его биологическую структуру, является слабо изученным. Например, человеческий белок IL-7 имеет цистеин в положениях 2, 34, 47, 92, 129 и 141 зрелого белка и три потенциальных N-связанных сайта гликозилирования в положениях аспарагин (Asn)70, Asn91 и Asn116. Однако точная структура IL-7, синтезированного при культивировании эукариотических клеток, является неизвестной.
[0032] Настоящее изобретение включает слитые белки IL-7, которые имеют особые структурные формы и повышенную биологическую активность. Например, слитые белки IL-7, имеющие дисульфидную модель связывания Cys2-Cys92, Cys34-Cys129 и Cys47-141, являются более активными in vivo, чем рекомбинантный белок IL-7 дикого типа.
[0033] Более того, изобретение обеспечивает форму слитого белка IL-7, в которой два или три потенциальных N-связанных сайта гликозилирования IL-7 являются гликозилированными. В частности, Asn70 и Asn91 зрелого белка являются гликозилированными, в то время как предсказанный N-связанный сайт гликозилирования в положении Asn116 IL-7 не является гликозилированным. Такой слитый белок IL-7 является более активным in vivo, чем рекомбинантный IL-7 дикого типа.
[0034] Изобретение также включает слитые белки IL-7, в которых остаток IL-7 содержит делецию и которые сохраняют сравнимую активность по сравнению с соответствующими немодифицированными слитыми белками IL-7. Например, изобретение обеспечивает форму Ig-IL-7, в которой остаток IL-7 содержит делецию восемнадцати внутренних аминокислот, соответствующую последовательности VKGRKPAALGEAQPTKSL (SEQ ID NO:9).
Слитые белки интерлейкина-7
[0035] Обычно IL-7 часть белка сливают с белком носителя. В одном воплощении белок носителя располагается на N-терминальном конце слитого белка, а белок IL-7 располагается на С-терминальном конце. В другом воплощении слитый белок IL-7 располагается на N-терминальном конце слитого белка, а белок носителя располагается на С-терминальном конце.
[0036] Как используется в данной заявке, термин «интерлейкин-7» или «IL-7» означает полипептиды IL-7 и их производные и аналоги, которые имеют существенную идентичность аминокислотной последовательности с зрелым человеческим IL-7 дикого типа и существенно эквивалентую биологическую активность, например, в стандартных биоанализах или анализах связывающей аффинности рецептора IL-7. Например, IL-7 относится к аминокислотной последовательности рекомбинантного или нерекомбинантного полипептида, который имеет аминокислотную последовательность i) нативного или существующего в природе аллельного варианта полипептида IL-7, ii) биологически активного фрагмента полипептида IL-7, iii) биологически активного полипептидного аналога полипептида IL-7, или iv) биологически активного варианта полипептида IL-7. Полипептиды IL-7 в соответствии с изобретением могут быть получены из любых видов, например человека, коровы или овцы. Последовательность нуклеиновой кислоты и аминокислотная последовательность IL-7 являются хорошо известными в области техники. Например, аминокислотная последовательность человеческого IL-7 имеет депозитный номер в Genbank NM 000880 (SEQ ID NO:1), она показана на Фигуре 1; аминокислотная последовательность мышиного IL-7 имеет депозитный номер в Genbank NM 008371; аминокислотная последовательность IL-7 крысы имеет депозитный номер в Genbank AF 367210; аминокислотная последовательность IL-7 коровы имеет депозитный номер в Genbank NM 173924 (SEQ ID NO:2), она показана на Фигуре 2; а IL-7 аминокислотная последовательность овцы имеет депозитный номер в Genbank 1110089 (SEQ ID NO: 3), она показана на Фигуре 3. Сигнальная последовательность для полипептида каждого вида выделена жирным шрифтом на каждой из Фигур и обычно не включается в случае, если IL-7 часть слита своей С-терминальной частью с белком носителя.
[0037] «Вариант» белка IL-7 определяется как аминокислотная последовательность, которая изменена в одной или более аминокислотах. Вариант имеет «консервативные» изменения в том случае, если заменяющая аминокислота обладает подобными структурными или химическими свойствами, например замена лейцина изолейцином. Более редко вариант может иметь «неконсервативные» изменения, например замену глицина триптофаном. Подобные минорные вариации могут также включать делеции аминокислот или инсерции, или и то, и другое. Руководство по поводу определения, какая и как много аминокислотных остатков могут быть заменены, встроены или делетированы без устранения биологической активности, могут быть найдены при использовании компьютерных программ, которые являются хорошо известными в области техники, например, программного обеспечения для молекулярного моделирования или для получения выравнивания. Варианты IL-7 белков, входящие в объем данного изобретения, включают белки IL-7, которые сохраняют активность IL-7. Полипептиды IL-7, которые включают добавления, замены или делеции, также включаются в данное изобретение до тех пор, пока они сохраняют существенно эквивалентную биологическую активность IL-7. Например, укороченные варианты IL-7, которые сохраняют сравнимую биологическую активность, что и форма полной длины белка IL-7, включаются в данное изобретение. Активность белка IL-7 может быть измерена при использовании in vitro анализов клеточной пролиферации, таких как описано в Примере 6 выше. Активность вариантов IL-7 в соответствии с изобретением поддерживает биологическую активность, по крайней мере, 10%, 20%, 40%, 60%, но более предпочтительно 80%, 90%, 95% и даже более предпочтительно 99% по отношению с IL-7 дикого типа.
[0038] Варианты белков IL-7 также включают полипептиды, которые обладают, по крайней мере, приблизительно 70%, 75%, 80%, 85%, 90%, 92%, 95%, 96%, 97%, 98%, 99% или более идентичностью последовательности с последовательностью IL-7 дикого типа. Для определения процента идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот эти последовательности выравниваются для целей оптимального выравнивания (например, в первую аминокислотную последовательность или последовательность нуклеиновой кислоты могут быть введены пробелы для оптимального выравнивания со второй аминокислотной последовательностью или последовательность нуклеиновой кислоты). Процент идентичности между двумя последовательностями представляет собой функцию номера идентичных положений, общих для последовательностей (то есть % гомологии = число идентичных положений/общее число положений·100 раз). Определение процента гомологии между двумя последовательностями может быть осуществлено при использовании математического алгоритма. Предпочтительный неограничивающий пример математического алгоритма, используемого для сравнения двух последовательностей, представляет собой алгоритм Karlin и Altschul (1990) Proc. Natl. Acad. Sci. USA 87: 2264-68, модифицированного в соответствии с Karlin и Altschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-77. Такой алгоритм введен в программы NBLAST и XBLAST Altschul, и др., (1990) J. Mol. Biol. 215: 403-10. BLAST нуклеотидный поиск может быть осуществлен с помощью программы NBLAST, шкала = 100, разрядность = 12. BLAST белковый поиск может быть осуществлен с помощью программы XBLAST, шкала = 50, разрядность = 3. Для получения выравнивания, содержащего пробелы, для целей сравнения может быть использована программа Gapped BLAST, как описано у Altschul и др,. (1997) Nucleic Acids Research 25(17): 3389-3402. При использовании программ BLAST и Gapped BLAST можно использовать параметры по умолчанию соответствующих программ (например, XBLAST и NBLAST).
[0039] Потенциальные Т-клеточные или В-клеточные эпитопы в остатке IL-7 могут быть удалены или модифицированы в Fc-IL-7 слитые белки в соответствии с изобретением. Примеры остатков IL-7 с устраненной иммуногенностью раскрыты в во временной заявке США под названием «Варианты IL-7 со сниженной иммуногенностью» (Номер дела патентного поверенного LEX-035PR), которая была подана в Ведомство США по патентам и товарным знакам 9 декабря 2004 года.
Белок носителя
[0040] Белок носителя может представлять собой любой остаток, ковалентно слитый с белком IL-7. В одном воплощении белок носителя представляет собой альбумин, например человеческий сывороточный альбумин. В другом воплощении белок носителя представляет собой остаток иммуноголобулина (Ig), такой как тяжелая цепь Ig. Цепь Ig может быть получена из IgA, IgD, IgE, IgG или IgM. В соответствии с изобретением остаток Ig может образовывать интактное антитело и может нацеливать слитый белок IL-7 на специфические целевые сайты в организме. Слитые белки, использующие нацеливание антител, являются известными для специалиста в данной области. В другом воплощении остаток Ig носителя дополнительно включает легкую цепь Ig.
[0041] В одном воплощении остаток Ig включает участок Fc. Как используется в данной заявке, термин «Fc часть» охватывает домены, которые имеют происхождение от константного участка иммуноглобулина, предпочтительно человеческого иммуноглобулина, в том числе фрагмента, аналога, варианта, мутанта или производной константного участка. Приемлемые иммуноглобулины включают IgG1, IgG2, IgG3, IgG4 и другие классы. Константный участок иммуноглобулина определяется как природный или синтетически полученный полипептид, гомологичный иммуноглобулину С-терминального участка, он может включать домен СН1, шарнирный участок, СН2 домен, СН3 домен или СН4 домен, отдельно или в любой комбинации. В данном изобретении часть Fc обычно включает СН2 домен. Например, Fc часть может включать шарнир-СН2-СН3. Альтернативно, Fc часть может включать весь или часть шарнирного участка, СН2 домен и/или СН3 домен и/или СН4 домен.
[0042] Константный участок иммуноглобулина является ответственным за множество важных функций антитела, в том числе связывание с рецептором Fc (FcR) и фиксацию комплемента. Существует пять основных классов константных участков тяжелой цепи, которые классифицируются как IgA., IgG, IgD, IgE и IgM. Например, IgG подразделяется на четыре подкласса: 1, 2, 3 и 4, которые также известны как IgG1, IgG2, IgG3 и IgG4 соответственно.
[0043] Молекулы IgG взаимодействуют с многочисленными классами клеточных рецепторов, в том числе тремя классами рецепторов Fсγ (FcγR), специфичных для IgG класса антител, а именно FcγRI, FcγRII и FcγRIII. Важные последовательности для связывания IgG с рецепторами FcγR, как было сообщено, расположены в доменах СН2 и СН3. Период полураспада антитела в сыворотке крови находится под влиянием способности этого антитела связываться с рецептором Fc (FcR). Подобно этому, период полураспада слитых белков иммуноглобулина также находится под влиянием способности связываться с такими рецепторами (Gillies и др., (1999) Cancer Res. 59: 2159-66). По сравнению с таковыми IgG1, СН2 и СН3, домены IgG2 и IgG4 имеют биохимически невыявляемую или пониженную связывающую аффинность к рецепторам Fc. Было показано, что слитые белки иммуноглобулина, содержащие СН2 и СН3 домены IgG2 или IgG4, имеют более длительные периоды полураспада по сравнению с соответствующими слитыми белками, содержащими СН2 и СН3 домены IgG1 (Патент США №5,541,087; Lo и др., (1998) Protein Engineering, 11: 495-500). В соответствии с определенными воплощениями данного изобретения предпочтительные СН2 и СН3 домены получены из изотипа антитела со сниженными связывающей аффиностью антитела и эффекторными функциями, таких как, например, IgG2 или IgG4. Более предпочтительно СН2 и СН3 домены являются полученными из IgG2.
[0044] Шарнирный участок обычно располагается С-терминально по отношению к СН1 домену константного участка тяжелой цепи. В изотипах IgG дисульфидные связи обычно возникают внутри этого шарнирного участка, что позволяет образовывать тетрамерную молекулу антитела. В этом участке доминирующими являются пролины, серины и треонины. В рамках настоящего изобретения шарнирный участок является обычно, по крайней мере, гомологичным природному участку иммуноглобулина, который включает остатки цистеина для образования дисульфидных связей, связывающих два остатка Fc. Характерные последовательности шарнирных участков для иммуноглобулинов человека и мыши являются известными в области техники и могут быть найдены у Borrebaeck, ред., (1992) Antibody Engineering. A Practical Guide. W.H. Freeman и Со. Приемлемые шарнирные участки для настоящего изобретения могут быть получены из IgG1, IgG2, IgG3, IgG4 и других классов иммуноглобулинов.
[0045] Шарнирный участок IgG1 содержит три цистеина, второй и третий из которых являются вовлеченными в дисульфидные связи между двумя тяжелыми цепями иммуноглобулина. Эти одинаковые два цистеина позволяют осуществлять эффективное и стойкое дисульфидное связывание части Fc. Таким образом, предпочтительный шарнирный участок в соответствии с настоящим изобретением получен из IgG1, более предпочтительно из человеческого IgG1, в котором первый цистеин является предпочтительно мутированным до другой аминокислоты, предпочтительно серина.
[0046] Шарнирный участок изотипа IgG2 содержит четыре дисульфидные связи, которые имеют тенденцию к усилению олигомеризации и возможному неправильному дисульфидному связыванию при секреции в рекомбинантных системах. Приемлемые шарнирные участки могут быть получены из шарнирного участка IgG2; при этом каждый из двух первых цистеинов является предпочтительно мутированными до другой аминокислоты.
[0047] Шарнирный участок IgG4 известен как такой, который формирует неэффективные внутрицепочечные дисульфидные связи. Однако приемлемый шарнирный участок для настоящего изобретения может быть получен из шарнирного участка IgG4, предпочтительно содержащего мутацию, которая способствует правильному формированию дисульфидных связей между остатками тяжелых цепей (Angal и др., (1993) Mol. Immunol., 30: 105-8).
[0048] В соответствии с настоящим изобретением часть Fc может содержать СН2 и/или СН3 и/или СН4 домены и шарнирный участок, который имеет происхождение от различных изотипов антитела, то есть является гибридной Fc частью. Например, в одном воплощении, Fc часть содержит СН2 и/или СН3 домены, полученные из IgG2 или IgG4, и мутантный шарнирный участок, полученный из IgG1. Как используется в данной заявке, Fcγ2(h) относится к воплощению, в котором шарнирный участок имеет происхождение от IgG1, a остальные константные домены происходят из IgG2. Альтернативно, мутантный шарнирный участок, полученный из другого подкласса IgG, используется в гибридной части Fc. Например, может использоваться мутантная форма шарнира IgG4, которая позволяет осуществлять эффективное дисульфидное связывание между двумя тяжелыми цепями. Мутантный шарнирный участок может также иметь происхождение от шарнирного участка IgG2, в котором каждый из двух первых цистеинов является мутированным до другой аминокислоты. Такие гибридные части Fc способствуют экспрессии на высоком уровне и улучшают правильную сборку Fc-IL-7 слитых белков. Сборка таких гибридных Fc частей является известной в области техники и была описана в опубликованной патентной заявке США №2003-0044423.
[0049] В некоторых воплощениях Fc часть содержит модификации аминокислот такие, которые обычно удлиняют период полураспада Fc слитого белка в сыворотке крови. Такие аминокислотные модификации включают мутации, существенно снижающие или устраняющие Fc связывание с рецептором или активность в отношении фиксации комплемента. Например, сайт гликозилирования в пределах Fc части тяжелой цепи иммуноглобулина может быть удален. В IgG1 сайт гликозилирования представляет собой Asn297 в пределах аминокислотной последовательности Gln-Tyr-Asn-Ser (SEQ ID NO:30). В других изотипах иммуноглобулина сайт гликозилирования соответствует Asn297 IgG1. Например, в IgG2 и IgG4 сайт гликозилирования представляет собой аспарагин в пределах аминокислотной последовательности Gln-Phe-Asn-Ser (SEQ ID NO:29). В соответствии с этим мутация Asn297 IgG1 удаляет сайт гликозилирования в Fc части, имеющей происхождение от IgG1. В одном воплощении Asn297 является замененным на Gln. В другом воплощении тирозин в пределах аминокислотной последовательности Gln-Tyr-Asn-Ser (SEQ ID NO:30) является дополнительно мутированным для устранения потенциального эпитопа, не принадлежащего Т-клеткам, полученного в результате мутации аспарагина. Как используется в данной заявке, эпитоп Т-клеток представляет собой полипептидную последовательность в белке, которая взаимодействует с или связывается с молекулами МНС класса II. Например, аминокислотная последовательность Gln-Tyr-Asn-Ser (SEQ ID NO:30) в пределах IgG1 тяжелой цепи может быть заменена на аминокислотную последовательность Gln-Ala-Gln-Ser (SEQ ID NO:28). Подобно этому, мутация аспарагина в пределах аминокислотной последовательности Gln-Phe-Asn-Ser (SEQ ID NO:29) в IgG2 или IgG4 устраняет сайт гликозилирования в Fc части, имеющей происхождение от тяжелой цепи IgG2 или IgG4. В одном воплощении аспарагин заменяется глутамином. В других воплощениях фенилаланин в пределах аминокислотной последовательности Gln-Phe-Asn-Ser (SEQ ID NO:29) является дополнительно мутированным для устранения потенциального эпитопа, не принадлежащего Т-клеткам, полученного в результате мутации аспарагина. Например, аминокислотная последовательность Gln-Phe-Asn-Ser (SEQ ID NO: 29) в пределах тяжелой цепи IgG2 или IgG4 может быть заменена аминокислотной последовательностью Gln-Ala-Gln-Ser (SEQ ID NO: 28).
[0050] Как показали наблюдения, изменение аминокислот поблизости соединения части Fc и не-Fc части может значительно повысить период полураспада в сыворотке крови слитого белка Fc (опубликованная патентная заявка США №2002-0147311). Соответственно, участок соединения слитого белка Fc-IL-7 или IL-7-Fc в соответствии с настоящим изобретением может также содержать изменения, которые соответствуют природным последовательностям тяжелой цепи иммуноглобулина и IL-7 и предпочтительно лежат в пределах приблизительно 10 аминокислот точки соединения. Эти аминокислотные изменения могут вызывать повышение гидрофобности путем, например, изменения С-терминального лизина Fc части на гидрофобную аминокислоту, такую как аланин или лейцин. Еще в одном воплощении в соответствии с изобретением С-терминальный лизин и предшествующий глицин части Fc являются делетированными.
[0051] В других воплощениях Fc часть содержит изменения аминокислот сегмента Leu-Ser-Leu-Ser поблизости С-терминального конца Fc части тяжелой цепи иммуноглобулина. Аминокислотные замены сегмента Leu-Ser-Leu-Ser (SEQ ID NO:27) устраняют потенциальные соединительные эпитопы Т-клеток. В одном воплощении аминокислотная последовательность Leu-Ser-Leu-Ser (SEQ ID NO:27) поблизости С-терминального конца Fc части заменяется на аминокислотную последовательность Ala-Thr-Ala-Thr (SEQ ID NO:26). В других воплощениях аминокислоты в пределах сегмента Leu-Ser-Leu-Ser (SEQ ID NO:27) заменяются на другие аминокислоты, такие как глицин или пролин. Подробно способы осуществления аминокислотных замен сегмента Leu-Ser-Leu-Ser (SEQ ID NO:27) поблизости С-терминального конца IgG1, IgG2, IgG3, IgG4 или других классов молекул иммуноглобулина, а также другие примеры модификаций для изменения соединительных эпитопов Т-клеток, были описаны в опубликованной заявке США №2003-0166877.
Спейсер
[0052] В одном воплощении спейсерный или линкерный пептид является встроенным между белком носителя и слитым белком IL-7. Например, спейсер размещается непосредственно С-терминально по отношению к последней аминокислоте константного участка Ig. Спейсерный или линкерный пептид является предпочтительно незаряженным и более предпочтительно неполярным или гидрофобным. Длина спейсерного или линкерного пептида предпочтительно составляет от 1 до приблизительно 100 аминокислот, более предпочтительно от 1 до приблизительно 50 аминокислот, или от 1 до приблизительно 25 аминокислот, и даже более предпочтительно от 1 до приблизительно 15 аминокислот, и даже более предпочтительно менее чем 10 аминокислот. В одном воплощении спейсер содержит последовательность (G4S)n, где n является меньшим чем 5. В предпочтительном воплощении спейсер содержит последовательность G4SG4 (SEQ ID NO: 25). Еще в одном воплощении спейсер содержит фрагмент, который узнается как N-связанный сайт гликозилирования. Еще в одном воплощении спейсер содержит фрагмент, который узнается сайт-специфическим агентом расщепления. В альтернативном воплощении изобретения белок носителя и слитый белок IL-7 разделены синтетическим спейсером, например, PNA спейсером, который предпочтительно является незаряженным и более предпочтительно неполярным или гидрофобным.
Получение слитых белков IL-7
[0053] Неограничивающие способы для синтеза полезных воплощений в соответствии с изобретением описаны в Примерах данной заявки, как и анализы, полезные для определения in vitro свойств, фармакокинетики и in vivo активности на животных моделях.
[0054] Слитые белки IL-7 в соответствии с изобретением могут быть получены при использовании рекомбинантных экспрессионных векторов, известных в области техники. Термин «экспрессионные векторы» относится к способной к репликации конструкции ДНК, которая кодирует желаемый слитый белок IL-7 и которая содержит транскрипционную единицу, включающую объединение (1) генетического(их) элемента(ов), который(ые) играет (играют) регуляторную роль в генной экспрессии, например, промоторов, операторов или энхансеров, оперативно связанного(ых) с (2) последовательностью ДНК, кодирующей желаемый слитый белок IL-7, который транскрибируется в мРНК и транслируется в белок, и (3) приемлемые последовательности инициации транскрипции и трансляции, а также терминации транскрипции и трансляции. Выбор промотора или других регуляторных элементов обычно варьирует в зависимости от намеченной хозяйской клетки. Предпочтительный экспрессионный вектор в соответствии с изобретением представляет собой Fc экспрессионный вектор, полученный из PdCs-huFc экспрессионного вектора, описанного у Lo и др., Protein Engineering (1998) 11: 495.
[0055] В предпочтительном примере нуклеиновая кислота, кодирующая слитый белок IL-7, трансфецируется в хозяйскую клетку при использовании методик рекомбинантной ДНК. В контексте настоящего изобретения чужеродная ДНК включает последовательность, кодирующую белки в соответствии с изобретением. Приемлемые хозяйские клетки включают прокариотические, дрожжевые и высшие эукариотические клетки. Предпочтительные хозяйские клетки представляют собой эукариотические клетки.
[0056] Рекомбинантные слитые белки IL-7 могут быть экспрессированы в дрожжевых хозяевах, предпочтительно из видов Saccharomyces, таких как S. cerevisiae. Могут также использоваться дрожжи других родов, таких как Pichia или Kluyveromyces. Дрожжевые векторы будут обычно содержать источник репликации из дрожжевой плазмиды или автономно реплицирующуюся последовательность (ARS), промотор, ДНК, кодирующую слитый белок IL-7, последовательности для полиаденилирования и терминации транскрипции, а также гены для селекции. Приемлемые промоторные последовательности в дрожжевых векторах включают промоторы для металлотионеина, 3-фосфоглицераткиназы или других гликолитических ферментов, таких как энолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкоза-4-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозафосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
[0057] Различные системы культур клеток млекопитающих или насекомых могут использоваться для экспрессии рекомбинантного белка. Бакуловирусные системы для получения белков в клетках насекомых являются хорошо известными в области техники. Примеры приемлемых хозяйских клеток млекопитающих включают клетки NS/0, L клетки, С127, 3Т3, клетки яичника китайского хомячка (СНО), линии клеток HeLa и ВНК. Дополнительные приемлемые хозяйские клетки млекопитающих включают клетки CV-1 (ATCC CCL70) и клетки COS-7, которые получены из почки обезьян. Другая приемлемая линия клеток почки обезьян, CV-1/EBNA, была получена путем трансфекции клеточной линии CV-1 с помощью гена, кодирующего ядерный антиген-1 вируса Эпштейна-Барр (EBNA-1) и с помощью вектора, содержащего регуляторные последовательности цитомегаловируса (CMV) (McMahan и др., (1991) EMBO J. 10: 2821). Ген EBNA-1 позволяет осуществлять эписомальную репликацию экспрессионных векторов, таких как HAV-EO или pDC406, которые содержат EBV источник репликации.
[0058] Экспрессионные векторы млекопитающих могут включать нетранскрибируемые элементы, такие как источник репликации, приемлемый промотор и энхансер, связанные с геном, который подвергают репликации, и другие 5' или 3' фланкирующие нетранскрибируемые последовательности, а также 5' или 3' нетранслируемые последовательности, такие как необходимые сайты связывания рибосом, сайты полиаденилирования, донорные и акцепторные сайты сплайсинга и последовательности терминации транскрипции. Обычно используемые промоторы и энхансеры являются такими, которые получены из вируса полиомы, аденовируса 2, обезьяньего вируса 40 (SV40) и цитомегаловируса человека. ДНК последовательности, полученные из вирусного генома SV40, например, SV40 источник, ранний и поздний промоторы, энхансеры, сплайсер и сайты полиаденилирования, могут использоваться для получения других генетических элементов, необходимых для экспрессии гетерологичной последовательности ДНК.
[0059] Для секреции слитого белка IL-7 из хозяйской клетки экспрессионный вектор включает ДНК, кодирующую сигнальный и лидерный пептиды. В настоящем изобретении можно использовать ДНК, кодирующую нативную сигнальную последовательность IL-7, или альтернативно, можно использовать ДНК, кодирующую гетерологичную сигнальную последовательность, такую как сигнальная последовательность из другого интерлейкина или из молекулы секретируемого Ig.
[0060] Настоящее изобретение также обеспечивает способ для получения рекомбинантных белков в соответствии с настоящим изобретением, включающий культивирование хозяйской клетки, трансформированной с помощью экспрессионного вектора, включающего последовательность ДНК, которая кодирует слитый белок IL-7, в условиях, благоприятствующих экспрессии. Желаемый белок потом очищают из культуральной среды или клеточных экстрактов. Например, супернатанты из экспрессионных систем, которые секретируют рекомбинантный белок в культуральную среду могут быть сначала сконцентрированы при использовании коммерчески доступных фильтров для концентрирования белка, например, ультрафильтрационного элемента Amicon или Millipore Pellicon. После этапа концентрирования для концентрата можно использовать приемлемый матрикс для очистки, который известен в области техники. Например, Fc-IL-7 слитые белки обычно поглощают при использовании матрикса, связанного с протеином А.
[0061] «Изолированный» или «очищенный» слитый белок IL-7 или его биологически активная часть означает, что он является существенно свободным от клеточного материала или других загрязняющих белков клетки или ткани, из которых этот слитый белок IL-7 получен, или что он является существенно свободным от химических предшественников или других химических соединений, если он получен с помощью химического синтеза. Выражение «существенно свободный от клеточного материала» включает препараты слитого белка IL-7, в которых белок отделен от компонентов клетки, из которых он изолирован или получен рекомбинантным путем. В одном воплощении выражение «существенно свободный от клеточного материала» включает препараты слитого белка IL-7, которые содержат менее чем 30% (по сухому веществу) неслитого белка IL-7 (которые также называются в данной заявке как «загрязняющий белок»), более предпочтительно менее чем приблизительно 20% белка, отличного от слитого белка IL-7, еще более предпочтительно менее чем приблизительно 10% белка, отличного от слитого белка IL-7, и наиболее предпочтительно менее чем приблизительно 5% белка, отличного от слитого белка IL-7. Когда слитый белок IL-7 или его биологически активная часть очищается из рекомбинантного источника, он также предпочтительно является существенно свободным от культуральной среды, то есть культуральная среда представляет менее чем приблизительно 20%, более предпочтительно менее чем приблизительно 10%, и наиболее предпочтительно менее чем приблизительно 5% от объема белкового препарата.
[0062] Термин «существенно чистый слитый белок Ig-IL-7» относится к препарату, в котором Ig-слитый белок IL-7 составляет, по крайней мере, 60%, 70%, 80%, 90%, 95% или 99% белков препарата. В одном из своих воплощений изобретение включает существенно чистые препараты слитых белков Ig-IL-7, имеющие дисульфидную связь между Cys2 и Cys92, Cys34 и Cys129, Cys47 и Cys141. В другом своем воплощении изобретение характеризует существенно чистые препараты слитых белков Ig-IL-7, в которых Asn116 не является гликозилированным, a Asn70 и Asn91 являются гликозилированными.
Способы лечения при использовании белков Fc-IL-7
[0063] Слитые белки IL-7 в соответствии с изобретением являются полезными при лечении иммунодефицитов и в ускорении восстановления иммунной системы, которое возникает, например, после заболеваний или лечений, которые являются иммуносупрессивными по своей природе. Например, слитые белки IL-7 могут использоваться для лечения инфекционных патогенов, расстройств иммунной системы и для усиления роста (в том числе пролиферации) специфических типов иммунных клеток. Кроме того, слитые белки IL-7 могут использоваться при лечении различных форм рака, такого как рак мочевого пузыря, рак легкого, рак мозга, рак молочной железы, рак кожи и рак предстательной железы. В одном воплощении является полезным лечить пациентов, которые подверглись одному или более циклам химиотерапии, с помощью слитых белков IL-7, как описано выше, для того, чтобы способствовать восстановлению иммунной системы. Альтернативно, слитые белки IL-7 являются полезными в адоптивных трансплантациях Т-клеток. Например, слитые белки IL-7 могут вводиться для размножения и выживаемости трансплантированных Т-клеток или для размножения популяции изолированных Т-клеток ex vivo. Альтернативно, является также полезным вводить слитые белки IL-7, описанные выше, пациентам с ВИЧ, а также пожилым пациентам, подвергнутым трансплантации, или другим пациентам с супрессированной функцией иммунной системы.
Введение
[0064] Слитые белки IL-7 в соответствии с изобретением могут быть введены в фармацевтическую композицию, приемлемую для введения. Такие композиции обычно включают слитый белок IL-7 и фармацевтически приемлемый носитель. Как используется в данной заявке, выражение «фармацевтически приемлемый носитель» предназначено для включения любого или всех растворителей, дисперсионной среды, покрытий, антибактериальных и противогрибковых агентов, изотонических агентов и агентов, замедляющих поглощение, и тому подобных, совместимых с фармацевтическим введением. Применение таких сред и агентов для фармацевтически активных веществ является хорошо известным в области техники.
[0065] Фармацевтическая композиция в соответствии с изобретением рецептируется таким образом, чтобы быть совместимой с путем введения. Примеры способов введения включают парентеральное, например, внутривенное, интрадермальное, подкожное, пероральное (например, путем ингаляции), трансдермальное (местное), введение через слизистую оболочку или ректальное введение. Растворы или суспензии, используемые для парентарального, интрадермального или подкожного применения, могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, жирные масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота; буферы, такие как ацетатный, цитратный или фосфатный, и агенты для доводки тоничности, такие как хлорид натрия или декстроза. Значение рН может доводиться с помощью кислот или оснований, таких как хлористоводородная кислота или гидроокись натрия. Препарат для парентерального введения может быть заключен в ампулы, одноразовые шприцы или флаконы, содержащие много доз, изготовленные из стекла или пластика.
[0066] Лекарственные средства, которые содержат слитые белки IL-7 в соответствии с изобретением, могут иметь концентрацию от 0,01 до 100% (мас./мас.), хотя их количество варьирует в зависимости от дозированной формы лекарственных средств.
[0067] Вводимая доза зависит от веса тела пациентов, тяжести заболевания и мнения врача. Однако обычно является целесообразным вводить приблизительно от 0,01 до приблизительно 10 мг/кг веса тела в день, предпочтительно от приблизительно 0,02 до приблизительно 2 мг/кг, и более предпочтительно от приблизительно 0,5 мг/кг в случае инъекции. Доза может вводиться один раз или несколько раз в день в соответствии с тяжестью заболевания и мнением врача.
[0068] Композиции в соответствии с изобретением являются полезными, когда вводятся с одним или более другими терапевтическими агентами, например, молекулой, которая является известной как таковая, которая полезна для пополнения клеток крови. Например, такая молекула может представлять собой эритропоэтин, который является известным как агент, способствующий пополнению клеток крови, G-CSF, который используется для пополнения нейтрофилов или GM-CSF, который используется для пополнения гранулоцитов и макрофагов.
ПРИМЕР 1
Клонирование вариантов (hu) Fc-IL-7 и huFc-IL-7 человека
[0069] Нуклеиновую кислоту, кодирующую зрелую форму человеческого IL-7 (то есть ту, в которой отсутствует ее N-терминальная сигнальная последовательность) подвергали амплификации с помощью полимеразной цепной реакции (ПЦР), используя прямые и обратные праймеры, которые встраивали рестрикционные сайты для Sma I и Xho I соответственно. Амплифицированный продукт ПЦР клонировали в вектор pCRII (Invitrogen, Carlsbad, CA), а его последовательность проверяли. Аминокислотная последовательность зрелого IL-7 показана в виде SEQ ID NO: 1. Sma I/Xho I переваренные фрагменты IL-7 переносили в обработанный таким же образом экспрессионный вектор, полученный из pdCs-huFc, что приводило к получению химерной последовательности между huFc и IL-7 с IL-7, размещенным в рамке считывания, непосредственно ниже последовательности, кодирующей домен СН3 Fc (смотри Lo и др., Protein Engineering (1998) 11: 495).
[0070] Серии экспрессионных векторов получали из вектора pdCs-huFc, кодирующего фрагмент Fc, который обычно включает шарнирный участок, СН2 домен, и СН3 домен Ig и который был сконструирован для включения специфических изменений с участок Fc. Таким образом, путем перемещения взад-вперед фрагмента IL-7 между этими векторами, были получены серии huFc-слитых белков IL-7, которые отличаются от своего скелета Fc. Для созданий различных скелетов приемлемые мутации сначала вводили в последовательность Fc с помощью способов, известных в области техники. Поскольку участок Fc вектора, полученного из pdCs-huFc, является фланкированным Af1II рестрикционным сайтом и SmaI рестрикционным сайтом, то путем проведения ПЦР нуклеиновой кислоты приемлемым образом модифицированного скелета и при использовании праймеров, которые встраивают рестрикционные сайты для Af1II и SmaI соответственно, полученный таким образом фрагмент нуклеиновой кислоты, кодирующий Fc, может быть потом заменен в векторе, полученном из pdCs-huFc, как фрагмент Af1II - SmaI. Af1II последовательность CTTAAGC (SEQ ID NO:24) находится выше начала последовательности Fc GAGCCCAAA (SEQ ID NO:23), которая представляет собой начало шарнирного участка, как показано на Фигуре 20. Сайт SmaI CCCGGGT (SEQ ID NO:17) находится на конце домена СН3, как показано с помощью подчеркивания нуклеиновых кислот на Фигуре 12, и кодирует Pro-Gly аминокислоты, предшествующие остатку аланина, полученному в результате мутации с изменением лизина на аланин в конце участка СН3.
[0071] Например, huFcγ1-IL-7 конструируют так, чтобы он содержал шарнирный участок, СН2 и СН3 домены, полученные из подкласса IgG1. В контексте Fc слитого белка IgGγ1 шарнирный участок в дополнение содержит мутацию, предусматривающую замену первого цистеина на серин. Последовательность кодируемого слитого белка показана на Фигуре 4 (SEQ ID NO:4), в то время как последовательность, представленная в SEQ ID NO:22, кодирует зрелый huFcγ1 скелет вектора.
[0072] В дополнение были получены Fcγ1-слитые белки IL-7, которые включают дипептидную мутацию YN на AQ для устранения сайта гликозилирования на Fc (соответствует N297 в IgGγ1), а также потенциального иммуногенного эпитопа Т-клеток в соответствии со способами, описанными выше. Последовательность скелета зрелого Fc для huFcγ1 (YN>AQ) раскрыта в SEQ ID NO:21. Замену аланином и глицином тирозина и аспарагина осуществляли с помощью первого введения мутаций в скелет Fc с помощью подхода перекрывания ПЦР. Два перекрывающихся комплементарных мутагенных праймера использовали для получения двух фрагментов ПЦР, которые использовали в качестве матрицы во втором цикле амплификации для получения одного фрагмента, содержащего приемлемые замены кодона. Мутагенный праймер в смысловом направлении представлял собой 5'-AGCAGGCCCAGAGCACGTACCGTGTGGT-3' (мутация подчеркнута) (SEQ ID NO:36). Комплементарная цепь представляла собой 5'-GTACGTGCTCTGGGCCTGCTCCTCCCGC-3' (SEQ ID NO:37). Фланкирующий направленный вперед праймер представлял собой 5'-CTCTCTGCAGAGCCCAAATCT-3' (SEQ ID NO:38), он также содержал сайт PstI. В антисмысловом направлении фланкирующий обратный праймер представлял собой 5'-CAGGGTGTACACCTGTGGTTC-3' (SEQ ID NO:33), он также содержал сайт BsrGI. После амплификации последовательность проверяли с помощью стандартных способов и подвергали рестрикции с помощью BsrGI и PstI. Полученным фрагментом потом заменяли немутантный участок Fc.
[0073] huFcγ2(b) (FN>AQ)-IL-7 также конструировали при использовании описанных ранее методик. Этот слитый белок включал измененный шарнирный участок, который имеет происхождение от подкласса IgGγ1, в то время как СН2 и СНЗ домены были получены из подкласса IgGγ2. В дополнение, дипептидная мутация FN на AQ была включена для устранения сайта гликозилирования на Fc (соответствует N297 в IgGγ1), а также для устранения потенциального иммуногенного эпитопа Т-клеток. Последовательность кодируемого слитого белка показана на Фигуре 5 (SEQ ID NO:5). Последовательность скелета зрелого Fc huFcγ2(h) (FN>AQ) представлена последовательностью SEQ ID NO:19.
[0074] В дополнение, были получены Fc-слитые белки IL-7, которые включают последовательность гибкого линкера между остатком Fc и остатком IL-7. Например, встраивали линкерный полипептид с последовательностью GGGGSGGGGSGGGGS (линкер1, SEQ ID NO: 34). Для получения huFcγ1 (линкер 1)-IL-7 синтетический олигонуклеотидный дуплекс последовательности 5'-GGGTGCAGGGGGCGGGGGCAGCGGGGGCGGAGGATCCGGCGGGGGCТС-3' (SEQ ID NO:18) встраивали путем лигирования тупых концов в уникальный сайт SmaI экспрессионного вектора pdCs-huFc-IL-7 и проверяли ориентацию дуплекса. Прямой праймер конструировали таким образом, чтобы аминокислотные остатки Pro - Gly кодировались кодонами, охватывающими сайт SmaI (СCCGGGT) (SEQ ID NO:17), а результирующий остаток Ala (полученный в результате замены кодируемого лизина на аланин) СН3 участка оставляли. Аминокислотная последовательность кодируемого слитого белка показана на Фигуре 6 (SEQ ID NO:6).
[0075] Дополнительные Fc-слитые белки IL-7 конструировали так, чтобы они включали линкерный полипептид с последовательностью GGGGSGGGG (линкер2, SEQ ID NO:25). Для получения huFcγ1 (YN>AQ) (линкер2)-IL-7 амплифицированный продукт ПЦР, полученный из пары праймеров 5'-CCCGGGCGCCGGCGGTGGAGGATCAGGTGGTGGCGGTGATTGTGATATTGAAGGTAAAGATG-3' (содержащий кодируемую линкерную последовательность, SEQ ID NO: 15) и 5'-ATCATGTCTGGATCCCTCGA-3' (SEQ ID NO:14) на приемлемой матричной плазмиде pdCs-Fc-IL-7, клонировали в вектор pCRII (Invitrogen, Carlsbad, СА), а его последовательность проверяли. Переваренный фрагмент Хmа I/Xho I, кодирующий линкер2/IL-7, потом трансфецировали в обработанный таким же образом экспрессионный вектор, полученный из pdCs-huFc. Вектор модифицировали так, чтобы он содержал скелет зрелого Fc huFcγ1 (YN>AQ) с последовательностью SEQ ID NO:21. Аминокислотная последовательность кодируемого слитого белка показана на Фигуре 7 (SEQ ID NO:7).
[0076] Подобно этому huFcγ1 (YN>AQ,d) (линкер2)-IL-7 получали при использовании пары праймеров 5'-CCCGGGCGGTGGAGGATCAGGTGGTGGCGGTGATTGTGATATTGAAGGTAAAGATG-3' (SEQ ID NO: 16) и 5'-ATCATGTCTGGATCCCTCGA-3' (SEQ ID NO:12). huFcγ1 (YN>AQ,d) (линкер2)-IL-7 отличается от предыдущего слитого белка huFcγ1 (YN>AQ) (линкер2)-IL-7 тем, что он не содержит двух терминальных аминокислотных остатков Fc части слитого белка. В частности, предпочтительнее, чем терминация последовательностью …ATATPGA (SEQ ID NO:11), является завершение Fc части последовательностью …AT ATP (SEQ ID NO:10). Аминокислотная последовательность кодируемого слитого белка показана на Фигуре 8 (SEQ ID NO:8).
ПРИМЕР 2
Трансфекция и экспрессия Fc-IL-7 слитых белков
[0077] Использовали электропорацию для введения ДНК, кодирующей слитые белки IL-7, описанные выше, в клеточную линию мышиной миеломы NS/0. Для осуществления электропорации клетки NS/0 выращивали в модифицированной Дюльбекко среде Игла с добавлением 10% инактивированной прогреванием фетальной сыворотки теленка, 2 мМ глутамина и пенициллина/стрептомицина. Приблизительно 5×106 клеток один раз промывали с помощью фосфатно-буферного раствора (ФБР) и ресуспендировали в 0,5 мл ФБР. 10 мкг линеаризованной плазмидной ДНК для huFcγ1-IL-7 потом инкубировали с клетками в кювете Gene Pulser (0,4 см электродная щель, BioRad) на льду в течение 10 мин. Электропорацию осуществляли при использовании Gene Pulser (BioRad, Hercules, CA) с параметрами 0,25 В и 500 мкФ. Клетки оставляли для восстановления в течение 10 мин на льду, после чего их ресуспендировали в ростовой среде и высаживали на планшеты на 96 ячеек.
[0078] Отбирали стабильно трансфецированные клоны по их росту в присутствии 100 нМ метотрексата (МТХ), который прибавляли к ростовой среде через два дня после трансфекции. Клетки подпитывали каждые три дня от двух до трех раз, при этом МТХ-резистентные клоны появлялись через 2-3 недели. Супернатанты, полученные из клонов, анализировали с помощью Supernatants анти-Fc ELISA для идентификации клонов, которые вырабатывают большие количества слитых белков IL-7. Клоны, имеющие высокие показатели продукции, изолировали и размножали в ростовой среде, содержащей 100 нМ МТХ. Обычно использовали ростовую среду, не содержащую сыворотки, такую как среда H-SFM или CD (Life Technologies).
ПРИМЕР 3
Биохимический анализ huFc-IL-7 слитых белков
[0079] Использовали обычную характеристику с помощью SDS-ПАГЭ для оценки целостности слитых белков. Исследовали различия между вариантами huFc-IL-7, huFcγ1-IL-7, huFcγ2(h) (FN>AQ)-IL-7, huFcγ1 (линкер 1)-IL-7, huFcγ1 (YN>AQ) (линкер2)-IL-7 и huFcγ1 (YN>AQ,d)-IL-7. Слитые белки huFc-IL-7, экспрессированные клетками NS/0, собирали на шариках Протеин А сефарозы (Repligen, Needham, MA) из среды тканевой культуры, в которую они секретировались, вымывали с помощью кипячения в буфере для белковых образцов при наличии или при отсутствии восстанавливающего агента, такого как β-меркаптоэтанол. Образцы разделяли с помощью SDS-ПАГЭ и полоски белков визуализировали при использовании окрашивания Кумасси. С помощью SDS-ПАГЭ исследуемые huFc-слитые белки IL-7 были обычно хорошо выражены, они присутствовали существенно в виде одной полосы в геле, было также обнаружено, что в образцах huFc-IL-7 вариантов, включающих линкер, вторичные полосы, которые могут представлять собой укороченный материал, были в значительной мере уменьшенными.
[0080] Очищенные huPc-слитые белки IL-7 также подвергали анализу с помощью гель-фильтрации (SEC) для оценки степени агрегации вариантов huFc-IL-7. Кратко, супернатанты клеточных культур загружали в предварительно уравновешенную Протеин А сефарозную колонку с высокой скоростью прокачки, колонку дважды хорошо промывали в физиологическом буфере (таком как 100 мМ фосфат натрия, 150 мМ NaCl при нейтральном значении рН), и полоску белка элюировали при значении рН приблизительно 2,5-3 в том же солевом буфере, что описан ранее. Фракции сразу же нейтрализовали.
[0081] Было обнаружено, что для каждого из проанализированных слитых белков, по крайней мере, 50% продукта было мономерным, а в общем случае более чем 65% продукта. Термин «мономерный», как используется в данной заявке, относится к неагрегированным белкам. Понятно, что белки, содержащие Fc часть, обычно образуют комплекс, связанный дисульфидной связью, которые обычно включают две полипептидные цепи (разве только две части Fc не присутствуют в одном и том же полипептиде) и могут считаться «единичным димером». Термин «мономерный» не предназначен для того, чтобы исключить такие связанные дисульфидной связью виды, но только для того, чтобы указать на то, что такие белки не являются агрегированными. Для получения фактически мономерного препарата huFc-слитого белка IL-7 (приблизительно 98%) элюат, полученный в результате очистки с помощью сефарозы-Протеина А, загружали в колонку для препаративной SEC (Superdex) и собирали фракцию мономерного пика. Обычно концентрация восстановленного белка составляла приблизительно 1 мг/мл. Если это было необходимым, то образец концентрировали, например, с помощью центрифужного диализа (например, VivaSpin) с отсечением молекулярного веса 10-30 кДа.
Дисульфидное связывание
[0082] IL-7 содержит шесть остатков Cys, которые могут образовывать дисульфидные связи, в положениях Cys2, Cys34, Cys47, Cys92, Cys129 и Cys141 последовательности зрелого белка IL-7. Сборку huFcγl-IL-7 анализировали с помощью определения модели дисульфидных связей, присутствующих в остатке IL-7 слитого белка. Кратко, пептидные карты huFcγl-IL-7 получали из трипсинизированного материала и анализировали на присутствие модели пептидных фрагментов. Белок huFcγ1-IL-7 подвергали трипсинизации либо в нативной форме, либо после восстановления и алкилирования. Для подсчета пептидных фрагментов, которые могут быть гликозилированными, образцы нативного и денатурированного белков дополнительно обрабатывали с помощью PNGазы F для удаления цепей гликозила перед пептическим перевариванием. Пептидные фрагменты фракционировали с помощью жидкостной хроматографии высокого разрешения (ВЭЖХ), и их массу определяли с помощью масс-спектроскопии.
[0083] В отношении Fcγ1-IL-7 пептидный фрагмент, содержащий дисульфидную связь Cys47-Cys141 («3-6»), может быть предсказан как такой, который имеет массу 1447,6, в то время как пептидный фрагмент, содержащий пептидную связь Cys2-Cys141 («1-6»), будет предсказан как такой, который имеет массу 1426,6. Подобно этому, пептидный фрагмент, содержащий дисульфидную связь Cys34-Cys129 («2-5»), может быть предсказан как такой, который имеет массу 2738,3. Действительно, пептидные фрагменты с массой 1447,6 («3-6») и 2738,3 («2-5») были идентифицированы в образцах, полученных из нативного белка Fc-IL-7 независимо от того, были ли образцы обработаны с помощью PNGазы F или нет, но не в образцах восстановленного Fc-IL-7. В противовес этому, пептидный фрагмент, имеющий массу 1426,6 («1-6»), не был обнаружен ни в одном образце. Таким образом, Fcγ1-IL-7 содержал дисульфидные связи Cys47-Cys141 и Cys34-Cys129, но не Cys2-Cys141. Было отмечено, что пептидный фрагмент предсказанной массы 2439,2, соответствующей фрагменту, содержащему Cys2-Cys92 («1-4»), был идентифицирован только в образце из нативного слитого белка, обработанного PNGазой F. Фактически Cys92 лежал внутри трипептидного фрагмента Asn91 Cys92 Thr93, показывая, что Asn91 был гликозилирован в huFcγ1-IL-7. Таким образом, модель дисульфидного связывания huFcγ1-IL-7 была в соответствии с Cys2-Cys92, Cys34-Cys129 и Cys47-Cys141. Эта экспериментально определенная конфигурация дисульфидных связей Fc-IL-7 находится в противоречии с экспериментально определенной конфигурацией, показанной для полученного с помощью бактерий и подвергнутого повторной сборке IL-7 (Cosenza и др., (1997) JBC 272:32995).
N-связанные сайты гликозилирования
[0084] Человеческий IL-7 содержит три потенциальных сайта гликозилирования в положениях Asn70, Asn91 и Asn116 последовательности зрелого белка IL-7. Пептидные карты huFcγ1-IL-7 (восстановленный/алкилированный) анализировали на присутствие сигнатурных пептидных фрагментов. При гликозилировании эти сигнатурные фрагменты будут выявляться только в образцах, обработанных с помощью PNGазы F. Массы 1489,7, 1719,9 и 718,3 могли бы быть предсказаны для триптических пептидных фрагментов, содержащих немодифицированные остатки для Asn70, Asn91 и Asn116 соответственно.
[0085] Действительно, пептидные фрагменты с массами 1489,7 и 1719,9 были идентифицированы в образцах, которые были обработаны с помощью PNGазы F, однако отсутствовали в необработанном образце, что свидетельствовало о том, что Asn70 (содержащийся в последовательности 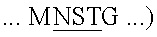 (SEQ ID NO:31) и Asn91 (содержащийся в последовательности
(SEQ ID NO:31) и Asn91 (содержащийся в последовательности  (SEQ ID NO:32) были действительно гликозилированными. Неожиданно триптический фрагмент с массой 718,3, соответствующий SLEENK (SEQ ID NO:35), идентифицировали в обоих образцах, обработанных PNGазой F, и в необработанном образце, что свидетельствовало о том, что Asn116 не был гликозилированным. Этот факт был неожиданным, поскольку Asn116 в последовательности IL-7 человека
(SEQ ID NO:32) были действительно гликозилированными. Неожиданно триптический фрагмент с массой 718,3, соответствующий SLEENK (SEQ ID NO:35), идентифицировали в обоих образцах, обработанных PNGазой F, и в необработанном образце, что свидетельствовало о том, что Asn116 не был гликозилированным. Этот факт был неожиданным, поскольку Asn116 в последовательности IL-7 человека 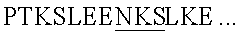 (смотри SEQ ID NO:1) является предсказанным как N-связанный сайт гликозилирования. Предполагаемый NKS сайт гликозилирования сохраняется у овцы, коровы, а также у человека.
(смотри SEQ ID NO:1) является предсказанным как N-связанный сайт гликозилирования. Предполагаемый NKS сайт гликозилирования сохраняется у овцы, коровы, а также у человека.
[0086] Анализ моделей дисульфидного связывания и N-связанных сайтов гликозилирования, повторенный с образцами Fcγ1-(линкер 1)-IL-7 и Fcγ2h (FN>AQ)-IL-7, давал подобные результаты.
ПРИМЕР 4
Процедуры ELISA
[0087] Концентрации белковых продуктов в супернатантах МТХ-резистентных клонов и других аналитических образцах определяли с помощью анти-huFc ELISA, как описано подробно ниже. Планшеты ELISA покрывали с помощью аффинно-чистого козьего античеловеческого IgG (H+L) (Jackson Immuno Research Laboratories, West Grove, PA) при концентрации 5 мкг/мл в ФБР в количестве 100 мкл/ячейка в планшетах на 96 ячеек. Покрытые планшеты накрывали оберткой и инкубировали при температуре 4°С в течение ночи. Планшеты четыре раза промывали с помощью 0,05% Твина (Твина 20) в ФБР и блокировали с помощью 1% БСА/1% козья сыворотка в ФБР, 200 мкл/ячейка. После инкубации с блокирующим буфером при температуре 37°С в течение 2 часов планшеты четыре раза промывали с помощью 0,05% Твина в ФБР и окончательно высушивали. Исследуемые образцы разводили приемлемым образом в образце буфера (1% БСА/1% козья сыворотка/0,05% Твин в ФБР). Стандартную кривую получали при использовании химерного антитела (с человеческим Fc) известной концентрации. Для получения стандартной кривой готовили серийные разведения в образце буфера для получения стандартной кривой, при этом концентрации колебались в интервале от 125 нг/мл до 3,9 нг/мл. Разведенные образцы и стандарты прибавляли к планшету в количестве 100 мкл/ячейка и планшет инкубировали при температуре 37°С в течение 2 часов. После инкубации планшеты промывали 8 раз с помощью 0,05% Твина в ФБР. К каждой ячейке прибавляли 100 мкл вторичного античеловеческого IgG, конъюгированного с пероксидазой хрена, и разводили до приблизительно 1:120000 в буфере для образцов. Точное разведение вторичного антитела определяли для каждой группы HRP-конъюгированного античеловеческого IgG. После инкубации при температуре 37°С в течение 2 часов планшет промывали 8 раз с помощью 0,05% Твина в ФБР.
[0088] Раствор субстрата прибавляли к планшету в количестве 100 мкл/ячейка. Раствор готовили путем растворения 30 мг OPD (дигидрохлорид о-фенилендиамина) (OPD), (1 таблетка) в 15 мл буфера 0,025 М лимонной кислоты/0,05 М Na2HPO4, pH 5, который содержал 0,03% свежеприготовленной перекиси водорода. Окрашивание развивалось в течение приблизительно 30 минут при комнатной температуре в темноте. Реакцию останавливали путем прибавления 4 н. серной кислоты, 100 мкл/ячейка. Планшеты считывали с помощью устройства для считывания, которое было установлено как на 490, так и на 650 нм, а также было запрограммировано для вычитания фоновой оптической плотности (OD) при 650 нм из OD при 490 нм.
[0089] Концентрацию человеческого IL-7 в образцах сыворотки животных, подвергнутых лечению с помощью huFc-слитых белков IL-7 или рекомбинантного человеческого IL-7, определяли с помощью ELISA существенно так, как описано выше. Человеческий IL-7 собирали с помощью мышиного античеловеческого IL-7 антитела (R&D Systems, Minneapolis, MN) и определяли с помощью козьего античеловеческого IL-7 биотинилированного антитела (R&D Systems, Minneapolis, MN).
ПРИМЕР 5
Очистка huFc-IL-7 белков
[0090] Стандартную очистку слитых белков, содержащих Fc, осуществляли на основе сродства остатка Fc белка для Протеина А. Кратко, клетки NS/0, экспрессирующие приемлемый слитый белок, выращивали на среде для тканевой культуры и собирали супернатанты, содержащие экспрессированный белок, после чего загружали их в предварительно уравновешенную Протеин А сефарозную колонку с высокой скоростью прокачки. Колонку хорошо промывали в буфере (таком как 100 мМ фосфата натрия, 150 мМ NaCl при нейтральном значении рН). Связанный белок элюировали при низком значении рН (рН 2,5-3) в том же буфере, что описан ранее, и фракции сразу же нейтрализовали.
[0091] Для получения препарата неагрегированного huFc-слитого белка IL-7 (приблизительно 98% мономера) элюат загружали в колонку для препаративной SEC (Superdex) и собирали фракцию мономерного пика. Обычно концентрация восстановленного белка составляла приблизительно от 0,5 мг/мл до 2 мг/мл. Если это было необходимым, то образец концентрировали, например, с помощью центрифужного диализа (например, VivaSpin) с отсечением молекулярного веса 30 кДа.
ПРИМЕР 6
In vitro активность белков huFc-IL-7
[0092] Цитокиновую активность huFc-слитых белков IL-7 определяли в in vitro биоанализе клеточной пролиферации. Человеческие РВМС (мононуклеарные клетки периферической крови) активировали с помощью РНА-Р для получения клеток, которые являются чувствительными к IL-7. Пролиферацию измеряли в стандартном анализе встраивания тимидина. Кратко, РВМС сначала инкубировали в течение пяти дней с 10 мкг/мл РНА-Р, клетки промывали и потом инкубировали в среде с добавлением huFc-IL-7 слитых белков в сериях разведений в общей сложности в течение 48 часов. В течение заключительных 12 часов образцы подвергали обработке импульсами с помощью 0,3 мкКи [метил-3H]тимидина (Dupont-NEN-027). Потом клетки хорошо промывали, собирали и лизировали на стеклянных фильтрах. 3Н-тимидин, встроенный в ДНК, измеряли на сцинтилляционном счетчике. В качестве стандарта анализировали белок huIL-7 дикого типа, полученный от R&D Systems (Minneapolis, MN) или полученный от National Institute for Biological Standards and Control (NIBSC).
[0093] Значение ED50 клеточной пролиферации для huFc-слитых белков IL-7 получали путем вычерчивания кривой зависимости ответа от дозы в соответствии со стандартными методиками и определяли концентрацию белка, которая приводит к ответу, равному половине максимального. Оценивали слитые белки huFcγ1-IL-7, huFcγ2(h) (FN>AQ)-IL-7 и huFcγ1 (линкер1)-IL-7. Значения ED50 для слитых белков были довольно подобными друг другу, отличаясь в пределах трехкратного интервала друг от друга. Таким образом, было обнаружено, что эти изменения в остатке Fc имели слабое влияние на активность IL-7 слитого белка.
[0094] В дополнение к этому было обнаружено, что значения ED50 этих слитых белков были приблизительно в 3-10 раз выше, чем значение ED50, полученное для huIL-7, коммерчески доступного от R&D Systems. Поскольку этот коммерческий препарат был получен на бактериях и не был гликозилированным, то подвергали оценке ферментативно дегликозилированный белок huFcγ1-IL-7, полученный путем обработки с помощью PNGазы F. Было обнаружено, что он имеет активность, подобную таковой для необработанной формы. Без намерения связывать это с теорией, до некоторой степени пониженная активность слитых белков может достигаться не за счет гликозилирования остатка IL-7, а вместо этого за счет стерического эффекта, который является результатом ограниченного N-терминального конца остатка IL-7.
ПРИМЕР 7
Фармакокинетика белков huFc-IL-7
[0095] Оценивали фармакокинетические профили (РК) huFc-слитого белка IL-7 и рекомбинантного человеческого IL-7 (Peprotech, Rocky Hill, NJ), результаты представлены на Фигуре 13. Одну подкожную инъекцию эквимолярных количеств huFcγ 2(h) (FN>AQ)-IL-7 или рекомбинантного человеческого IL-7 (50 микрограмм) вводили группам C57BL6/J мышей. Образцы крови получали путем обескровливания с помощью инъекции в ретроорбитальную область (то есть в момент времени t=0 мин), через 30 мин, 1 час, 2 часа, 4 часа, 8 часов, 24, 48, 72, 96, 120 и 144 часов после инъекции. Образцы собирали в пробирки с гепарином для предотвращения сворачивания крови, а клетки удаляли центрифугированием при высокой скорости в микроцентрифуге Эппендорфа в течение 4 мин при 12500 g. Значения РК подсчитывали с помощью РК 2.0™ комплекта программного обеспечения (Summit Research Services, Montrose, CO).
[0096] Концентрацию вводимого IL-7 определяли в четырехкратных образцах плазмы в каждой точке времени с помощью анализа ELISA, специфического для человеческого IL-7. При этом было показано, что фармакокинетическое поведение huFc-IL-7 и рекомбинантного IL-7 значительно отличаются. Для рекомбинантного человеческого IL-7 максимальная концентрация (Сmax) составляла 23,5 нг/мл в момент времени 2,0 часа после инъекции (Тmax), в то время как для huFc-IL-7 Сmax составляла 1588,7 нг/мл через 24 часа после инъекции. В дополнение, в то время как рекомбинантный человеческий IL-7 поглощался быстрее, чем huFc-IL-7 (β-фаза полужизни составляла 0,9 часа против 12,4 часа), huFc-IL-7 элиминировался из циркуляции приблизительно в 9 раз медленнее во время β-фазы. Таким образом, в единицах AUC (площадь под кривой) как меры общего воздействия лекарственного средства мыши, которые получали huFc-IL-7, подвергались в 572 раза более высокому воздействию вводимого белка, чем мыши, получающие рекомбинантный человеческий IL-7. Эти данные демонстрируют значительное усовершенствование huFc-слитых белков IL-7 по отношению к свободному рекомбинантному IL-7 в отношении их РК. Было также обнаружено, что РК профили huFc-IL-7 слитых белков, таких как huFcγ1-IL-7 и huFcγ2(h) (FN>AQ)-IL-7, huFcγ1 (YN>AQ)(линкер2)-IL-7 и huFcγ1 (YN>AQ,d) (линкер2)-IL-7, которые вводили мышам путем внутривенной инъекции, были подобны друг другу.
ПРИМЕР 8
Эффективность huFc-IL-7 у лимфопенических мышей после трансплантации костного мозга (КМ)
[0097] Эффектиность huFc-слитых белков IL-7 по сравнению с рекомбинантным человеческим IL-7 оценивали in vivo. Например, huFcγ2(h) (FN>AQ)-IL-7 или рекомбинантный человеческий IL-7 (Peprotech, Rocky Hill, New Jersey) вводили лимфопеническим мышам после трансплантации костного мозга (КМ), истощенного по Т-клеткам, и оценивали восстановление популяции иммунных клеток.
[0098] По существу, реципиентных мышей подвергали летальному облучению перед трансплантацией КМ с помощью двух доз 600 сГр общего облучения тела при интервале 4 часа, клетки КМ, ресуспендированные в ФБР, вводили путем инфузии в хвостовые вены реципиентных мышей. Через регулярные интервалы, начиная от 5 дня, реципиентным мышам вводили подкожно эквимолярное количество huFc-IL-7 (7 мкг) или рекомбинантного человеческого IL-7 (2,5 мкг) (Peprotech, Rocky Hill, NJ). В ходе всего эксперимента брали образцы крови у реципиентных мышей и измеряли концентрации клеток лимфоцитов в образцах.
[0099] Для трансплантаций клетки КМ получали асептически из бедра и большой берцовой кости BL/6.SJL (Н2b, CD45.1) мышей (Jackson Labs, Bar Harbor, ME) и истощали препарат по Т-клеткам путем удаления магнитно-меченных Т-клеток с помощью колонок MACS® (Miltenyi Biotec, Auburn, CA). Степень истощения Т-клеток отслеживали с помощью FACS анализа при использовании флуоресцентно меченных антител против CD45, αβ-TCR (Т-клетки) и 7-аминоактиномицина D (7-AAD, апоптические клетки) (Calbiochem, X). 10×106 живых (7-ААD-негативных) клеток КМ (содержащих менее, чем 1% Т-клеток) использовали на одну реципиентную мышь. При изогенных трансплантациях КМ в качестве реципиентного мышиного штамма использовали мышей В6 (Н2b, CD45.2), а для аллогенных трансплантаций КМ выбирали мышей B6C3F1 (Н2b/k, CD45.2).
[00100] Концентрации клеток лимфоцитов (как представлено в Таблице) измеряли существенно так, как описано Brocklebank и Sparrow (Brocklebank и Sparrow (2001) Cytometry 46: 254). Кратко, флуоресцентные шарики (TruCOUNT™ Tubes, BD Biosciences, San Jose, CA) растворяли в 40 мкл ФБР, содержащего смесь специфических для лимфоцитов антител. Последовательно прибавляли 10 мкл крови с добавлением антикоагулянта, перемешивали и инкубировали в течение 30 минут в темноте и при комнатной температуре. Красные клетки крови подвергали лизису в 450 мкл литического раствора для красных клеток крови (BD Biosciences, San Jose, CA) и образцы анализировали с помощью проточной цитометрии (BD FACSCalibur™, BD Biosciences, San Jose, CA). Концентрацию определенной популяции лимфоцитов (например, В-клеток, Т-клеток или суммарных лейкоцитов) определяли путем создания обособленных окон вокруг лимфоцитов и флуоресцентных шариков и считывания количества событий внутри каждого окна. Количество лимфоцитов, ограниченных окном, на микролитр подсчитывали путем деления числа событий в районе лимфоцита, ограниченного окном, на число событий в районе шарика, ограниченного окном. Это число умножали на дробь, в знаменателе которой находится число шариков на пробирку TruCOUNT™ (обеспечивается поставщиком), а в числителе - объем образца, и в завершении умножали на кратность разведения.
[00101] В одном эксперименте восстановление лимфоцитов анализировали при параметрах изогенной трансплантации КМ при использовании материалов и способов, указанных выше. Реципиентных мышей инъецировали с помощью huFcγ2(h) (FN>AQ)-IL-7 при дозе 7 мкг (125 мкг IL-7/кг веса тела), и осуществляли измерение лимфоцитарных клеток так, как описано. Донорные лимфоциты определяли как CD45.1 позитивные клетки, в то время как эндогенные лимфоциты реципиентных мышей определяли как CD45.2 позитивные клетки. Клетки В-лимфоцитов и Т-лимфоцитов идентифицировали при использовании В220 и CD3 маркеров лимфоцитов соответственно. Было обнаружено, что на 49 день происходило повторное заселение донорными лимфоцитами (CD45.1 позитивные клетки) реципиентной мыши до уровней, сравнимых с необлученными контрольными мышами, в то время как эндогенные лимфоциты (CD45.2 позитивные клетки) не размножались в значительной степени. В дополнение к этому, обработка с помощью huFc-слитого белка IL-7 не вызывала значительной токсичности. Эти результаты демонстрируют эффективность слитого белка в размножении адоптивно перенесенных популяций лимфоцитов. Эти результаты представлены на Фигуре 16.
[00102] В другом эксперименте модель аллогенной трансплантации КМ, которая лучше имитирует клиническое осуществление трансплантации, использовали для сравнения huFc-слитого белка IL-7 с рекомбинантным человеческим IL-7, результаты показаны в Таблице. Опять-таки, использовали материалы и способы, описанные выше. huFcγ2(h) (FN>AQ)-IL-7 и человеческий IL-7 (эквивалент 125 мкг IL-7/кг веса тела) вводили либо через день (q2d), или один раз в неделю (q7d) от 5 дня до 56 дня после трансплантации. Обработанные ФБР донорные мыши и необлученные мыши, а также реципиентные мыши с пересаженным костным мозгом, обработанные ФБР, служили в качестве контролей.
[00103] У реципиентных мышей, обработанных слитым белком, донорные В-клетки (CD45.1+, В220+, CD19+) достигали уровней базовой линии (как определено при использовании концентрации В-клеток в крови у донорных контрольных мышей) через 14 или 16 дней после трансплантации в случае, когда осуществляли введение по схеме q2d или q7d соответственно. В противовес этому, у реципиентных мышей, обработанных с помощью рекомбинантного человеческого IL-7, ни один из режимов дозировки не давал эффекта, количество В-клеток у реципиентных мышей, обработанных ФБР и человеческим IL-7, требовало приблизительно одинакового времени для достижения уровней базовой линии, приблизительно 28 дней. В дополнение к ускорению восстановления В-клеток обработка huFc-IL-7 усиливала беспрерывное размножение В-клеток до 33 дня: huFc-IL-7, вводимый в соответствии с режимом q2d, приводил к 7-кратному увеличению, в то время как введение в соответствии с режимом q7d приводило к 2,5-кратному увеличению количества В-клеток по сравнению с контрольными мышами. После 33 дня количество В-клеток падало, но все же было приблизительно в 2 раза выше, чем у контрольных мышей. Также после 33 дня уровни введенных белков IL-7 падали в крови, что частично могло быть по причине образования нейтрализующих антител к слитому белку человека. Фигура 14 представляет результаты восстановления В-клеток у облученных мышей с трансплантированным костным мозгом, обработанных с помощью рекомбинантного человеческого IL-7 и huFc-IL-7.
[00104] Подобные результаты наблюдали в отношении Т-клеток, полученных от донора (CD45.1+, CD3+, TCRα+β+). Обработка с помощью huFc-слитого белка IL-7 приводила к ускоренному восстановлению Т-клеток, в то время как обработка при использовании рекомбинантного человеческого IL-7 не оказывала такого действия. Максимальные уровни Т-клеток достигались приблизительно на 49 день. Однако количества Т-клеток выше базовой линии (то есть концентрация Т-клеток в крови донорных мышей) достигались только при режиме дозировки q2d huFc-IL-7 слитого белка, достигая приблизительно 1,5-кратного увеличения количества Т-клеток по отношению к контрольным мышам. Фигура 15 представляет эти результаты восстановления Т-клеток у облученных мышей, которым трансплантировали костный мозг и которых подвергали лечению с помощью рекомбинантного человеческого IL-7 и huFc-IL-7.
[00105] Несмотря на кратковременное повышение количества донорных В-клеток и Т-клеток у реципиентных мышей при определенных условиях, ни одна из экспериментальных мышей не продемонстрировала никаких признаков заболеваемости в ходе эксперимента. Анализ внутренних органов на 55 день не выявил никаких патологический отклонений ни в печени, ни в почках, легких, селезенке, тимусе, лимфатических узлах, желудке, тонком кишечнике и толстом кишечнике. Таким образом, этот эксперимент по аллогенной трансплантации продемонстрировал, что huFc-слитый белок IL-7 в значительной степени превосходил в экспериментах in vivo рекомбинантный человеческий IL-7 в отношении восстановления лимфоцитов после создания миелоабляционных условий.
ПРИМЕР 9
Эффективность huFc-IL-7 при трансплантациях Т-клеток у лимфопенических мышей
[00106] Эффективность huFc-слитых белков IL-7 также оценивали на модели трансплантации Т-клеток. По сути, гомогенную (клональную) популяцию Т-клеток трансплантировали в иммунодефицитных, облученных мышей. При этом реципиентным мышам вводили huFc-IL-7 слитый белок и оценивали степень восстановления Т-клеток и, в конечном счете, функцию Т-клеток.
[00107] Для получения гомогенной популяции Т-клеток спленоциты брали у мышей Р14 TCR-tg/RAG (Charles River Laboratories, Wilmington, MA), которые не имели В-клеток. В дополнение к этому, Т-клетки этих мышей экспрессировали трансгенный рецептор Т-клеток (TCR), Р14, который является специфическим для вирусного эпитопа (gp33 LCMV).
[00108] Суспензии единичных клеток вводили внутривенно в хвостовые вены RAG Cγ+ иммунодефицитных мышей (Charles River Laboratories), которые были подвергнуты однократному облучению дозой 650 рад (сублетальная доза) за 4 часа перед осуществлением трансплантации. В выбранные дни, начиная от 2 дня, реципиентным мышам вводили 7 мкг слитого huFcγ2(h) (FN>AQ)-IL-7. Контрольной группе реципиентных мышей вводили ФБР. Степень восстановления Т-клеток в ответ на huFc-слитый белок IL-7 или ФБР определяли путем измерения присутствия Р14 Т-клеток (CD8+ Vβ8.1+Vα2+ клетки) в крови с помощью проточной цитометрии.
[00109] Было обнаружено, что на 35 день мыши, которым вводили huFc-слитый белок IL-7, имели 17-кратное повышение количества Т-клеток (35000 клеток/мкл) по сравнению с контрольными мышами (2000 клеток/мкл). Действительно, уровни восстановления Т-клеток превышали таковые, которые наблюдали у необработанных мышей Р14 TCR (23000 клеток/мкл). В дополнение к этому, у мышей, которых подвергли лечению huFc-IL-7, значительная фракция восстановленных Т-клеток обладала понижающей регуляцией IL-2Rα рецепторной субъединицы, CD25, на поверхности клеток. Таким образом, huFc-слитый белок был не только полезным в размножении трансплантированных Т-клеток, но также в дополнение к этому мог обусловливать тот факт, что перенесенные Т-клетки были чувствительными к цитокинам, таким как IL-2.
ПРИМЕР 10
Вспомогательная терапия huFc-IL-7 для пациентов с нарушением иммунитета
[00110] Рассматриваются многочисленные клинические назначения, при которых пациенты могут получить пользу от вспомогательной терапии с помощью huFc-IL-7. Например, получают развитие новые способы лечения для педиатрических пациентов со злокачественным заболеванием, таким как лимфобластная или миелоидная лейкемия, которых подвергают миелоабляционной терапии, такие пациенты подвергаются лечению с помощью аллогенной гематопоэтической трансплантации стволовых клеток для восстановления иммунной системы.
[00111] Для значительного повышения потенциала донорного пула для таких пациентов было обнаружено, что G-CSF мобилизованные стволовые клетки периферической крови (PBSC), полученные от подходящих неродственных доноров или гаплоидентичных доноров с неподходящими 1-3 HLA локусами, могут быть источником клеток при условии, что трансплантат является истощенным по Т-клеткам (смотри Handgretinger и др., (2001) Annals NY Acad. Sciences 938: 340-357). Такое истощение значительно снижало частоту возникновения острого отторжения трансплантата на основе явления «трансплантат против хозяина» (GvHD); однако предполагается, что благодаря низкой концентрации Т-клеток существовала значительная задержка в восстановлении иммунной системы. Для пациентов существовал высокий риск вирусных инфекций в течение, по крайней мере, первых 6 месяцев после трансплантации, а уровни Т-клеток не возвращались к нормальному в течение года (Handgretinger и др., (2001) Annals NY Acad. Sciences 938: 340-357; Lang и др., (2003) Blood 101: 1630-6). Таким образом, было бы желательно повысить темп восстановления Т-клеток и других иммунных клеток у таких пациентов.
[00112] Пациенты, которые получат выгоду от терапии с помощью huFc-IL-7, включают пациентов с детской лейкемией, такой как лимфобластная или миелоидная лейкемия. Дети, имеющие такое расстройство, будут сначала подвергаться миелоабляционной терапии, которая может основываться либо на химиотерапевтическом агенте бусулфане, либо на облучении всего организма в сочетании с химиотерапией. Например, в соответствии с диагнозом и возрастом пациента подвергают облучению всего тела (обычно 6 обработок по 2 Гр каждая), а также вводят кроличий антитимоцитарный глобулин (10 мг/кг ежедневно в течение 3 дней), этопозид (40 мг/кг) и циклофосфамид (120 мг/кг).
[00113] Для получения CD34 позитивных (pos) стволовых клеток для трансплантации стволовые клетки периферической крови (PBSC) гистосовместимого (аллогенного) донора мобилизуют с помощью ежедневной дозы 10 микрограмм/кг G-CSF в течение 6 дней и собирают их при использовании лейкафареза на 5 и 6 день. Обычно получают и трансплантируют приблизительно 20×106/кг CD34 позитивных стволовых клеток. CD34 позитивные стволовые клетки очищают от PBSC с помощью позитивного отбора при использовании анти-CD34 антитела в системе SuperMACS (магнитно-активированная сортировка клеток, Miltenyi Biotec) и разводят. Истощение Т-клеток обычно составляет 5 log, до приблизительно 10×103 клеток/кг. Агрегаты и другой дебрис исключают из трансплантата с помощью FACS сортировки. Клеточную суспензию вводят путем инфузии пациенту через центральный венозный катетер. Необязательно трансплантат может включать очищенные популяции других иммунных клеток, таких как гаплоидентичные NK клетки, DC, моноциты, а также CD34 негативные стволовые клетки.
[00114] Для оценки приживления трасплантата осуществляют абсолютный подсчет нейтрофилов. Приживление трансплантата считается успешным, если уровни нейтрофилов остаются выше 50 клеток/микролитр. За восстановлением иммунных клеток наблюдают с помощью FACS анализа, сначала еженедельно, а тогда, когда Т-клетки начинают восстанавливаться, каждые 3 месяца.
[00115] Для усиления восстановления иммунной системы пациента подвергают лечению с помощью huFc-слитого белка IL-7, такого как huFcγ2h (FN>AQ)-IL-7 или huFcγ2 (YN>AQ) (линкер2)-IL-7. Приблизительно через 3 недели после трансплантации (или после установления приживления трансплантата) пациент получает подкожное введение huFcγ2h (FN>AQ)-IL-7 или huFcγ2 (YN>AQ) (линкер2)-IL-7 в дозе приблизительно 1,5 мг/м2 (или дозу в интервале от 0,15 мг/м2 до 15 мг/м2) приблизительно два раза в неделю в течение 6-12 месяцев до тех пор, пока количество Т-клеток не достигнет 50% от нормальных уровней. При этом обнаруживается, что прогноз для пациента улучшается благодаря сниженному риску вирусной инфекции, что представляет собой одно из основных послетрансплантационных осложнений. Также было обнаружено, что это лечение не повышает значительно риск острого GvHD.
[00116] В дополнение к введению белка huFc-IL-7 другие способы лечения в оптимальном случае проводили с профилактической целью. Таковые включают, например, ацикловир, метронидазол, флуканазол и ко-тримоксазол. В течение первых трех месяцев пациенту можно еженедельно необязательно вводить иммуноглобулины, а также G-CSF.









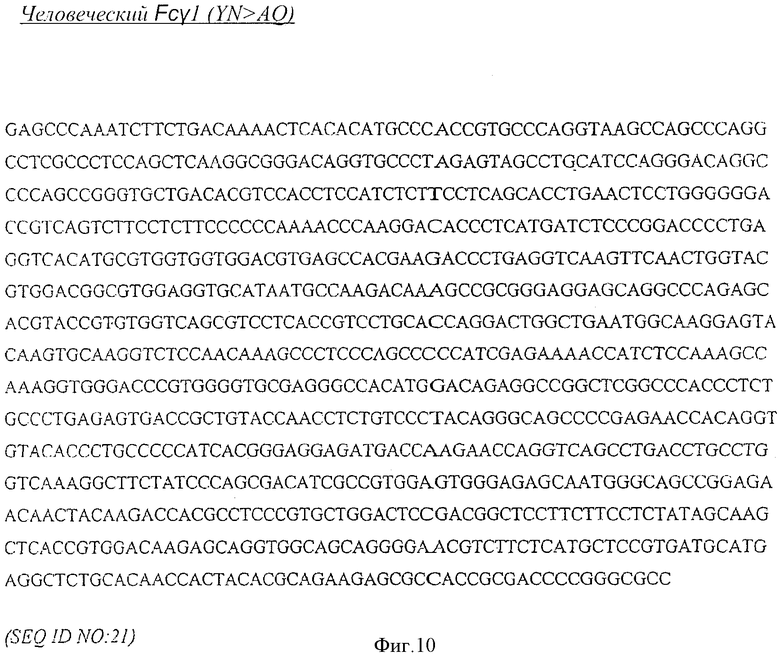


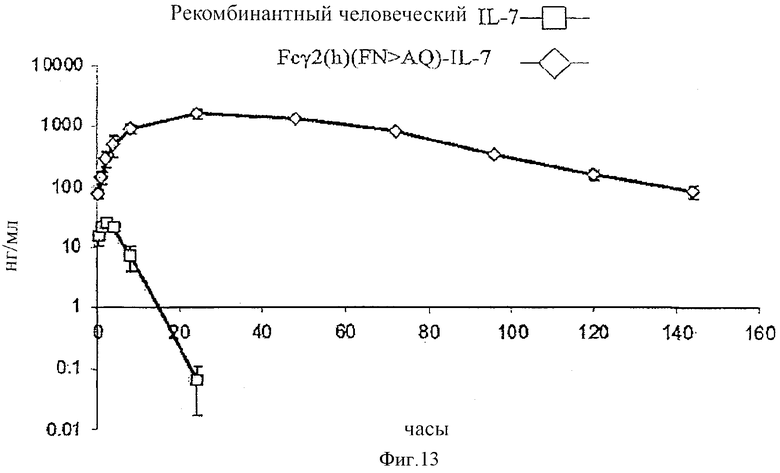
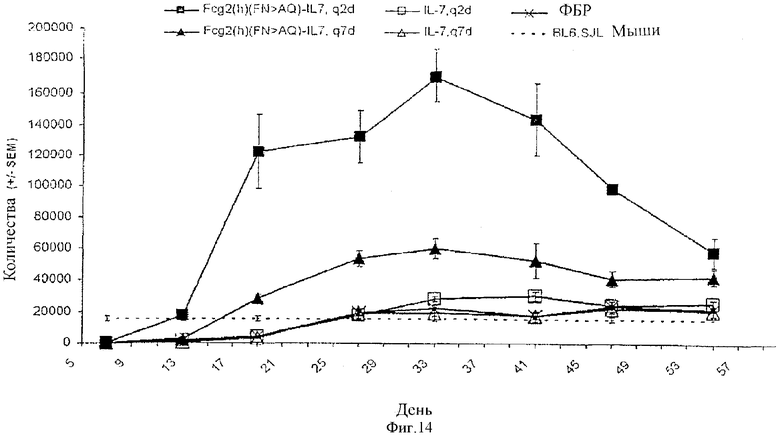
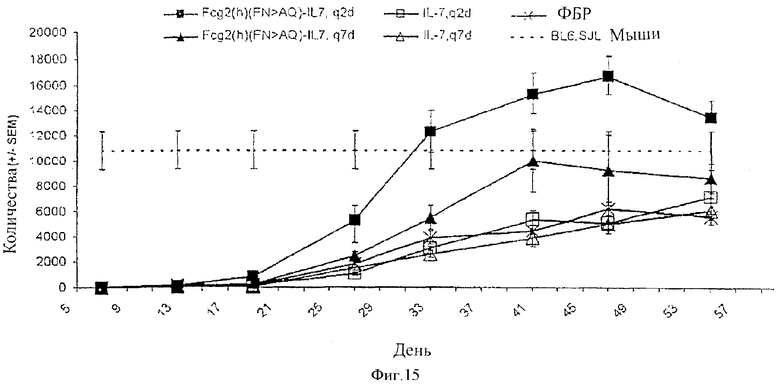
Изобретение относится к биотехнологии. Описан слитый белок, включающий цепочку иммуноглобулина и молекулу IL-7 или ее фрагмент, проявляющий активность, присущую IL-7, которая является модифицированной по сравнению с IL-7 дикого типа, где модификация в IL-7 представляет собой аминокислотные остатки в положениях 70 и 91, которые являются гликозилированными, а аминокислотный остаток в положении 116 является негликозилированным. Предложена фармацевтическая композиция, содержащая описанный белок. Представлен способ получения описанного слитого белка, включающий трансформацию клетки-хозяина с помощью ДНК, кодирующей описанный слитый белок IL-7, культивирование клетки-хозяина и сбор слитого белка IL-7. Изобретение может быть использовано при лечении расстройств, сопровождающихся иммунными дефицитами, и, в частности, заболеваний, которые вовлекают дефициты Т-клеток. 6 н. и 21 з.п. ф-лы, 16 ил., 1 табл.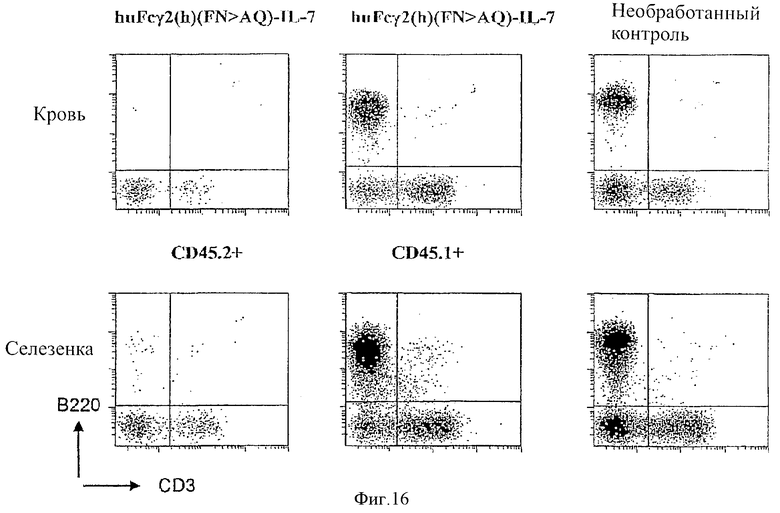
1. Слитый белок для лечения рака или иммунных расстройств, сопровождающихся иммунодефицитами, включающий цепочку иммуноглобулина и молекулу IL-7 или ее фрагмент, проявляющий активность, присущую IL-7, которая является модифицированной по сравнению с IL-7 дикого типа, связанные вместе непосредственно или посредством молекулы линкера, где модификация в IL-7 представляет собой аминокислотные остатки в положениях 70 и 91, которые являются гликозилированными, а аминокислотный остаток в положении 116 является негликозилированным.
2. Слитый белок по п.1, где в пределах молекулы IL-7 существуют дисульфидные связи между Cys2 и Cys92, Cys34 и Cys129 и Cys47 и Cys141.
3. Слитый белок по п.1, в котором аминокислотный остаток в положении 116 представляет собой аспарагин.
4. Слитый белок по п.1, в котором аминокислотный остаток в положении 116 является негликозилированным.
5. Слитый белок по п.1, в котором аминокислотный остаток в положении 116 является измененным так, что он не служит сайтом гликозилирования.
6. Слитый белок по п.1, в котором молекула IL-7 укорочена и включает делецию восемнадцати аминокислот от аминокислоты 96 до аминокислоты 114 SEQ ID NO:1.
7. Слитый белок по п.1, в котором молекула IL-7 представляет собой IL-7 человека.
8. Слитый белок по п.1, в котором молекула IL-7 представляет собой фрагмент зрелого IL-7.
9. Слитый белок по п.1, в котором цепочка иммуноглобулина включает по меньшей мере часть константного домена.
10. Слитый белок по п.9, в котором константный домен является выбранным из группы, состоящей из CH1, CH2 и СН3.
11. Слитый белок по п.9, в котором константный домен представляет собой константный домен IgG1.
12. Слитый белок по п.11, в котором Asn297 константного домена IgG1 является модифицированным.
13. Слитый белок по п.12, в котором модификация Asn297 представляет собой Asn297Gln.
14. Слитый белок по п.12, который дополнительно включает модификацию в Tyr296 константного домена IgGl.
15. Слитый белок по п.14, в котором модификация Tyr296 представляет собой Тyr296Аlа.
16. Слитый белок по п.9, в котором константный домен представляет собой константный домен IgG2.
17. Слитый белок по п.16, в котором цепочка IgG2 включает шарнир IgG1.
18. Слитый белок по п.17, в котором Asn297 константного домена IgG2 является модифицированным.
19. Слитый белок по п.18, в котором модификация Asn297 представляет собой Asn297Gln.
20. Слитый белок по п.18, дополнительно включающий модификацию в Phe296 константного домена IgG2.
21. Слитый белок по п.20, в котором модификация Phe296 представляет собой Phe296Ala.
22. Слитый белок по п.1 или 2, в котором молекула IL-7 слитого белка обладает повышенной биологической активностью по сравнению с IL-7 дикого типа.
23. Изолированная молекула ДНК, кодирующая слитый белок IL-7 по п.1 или 2.
24. Культивируемая клетка-хозяина, экспрессирующая слитый белок и включающая ДНК по п.23.
25. Способ получения слитого белка, включающий: трансформацию клетки-хозяина с помощью ДНК по п.23, культивирование клетки-хозяина и сбор слитого белка IL-7.
26. Фармацевтическая композиция, приемлемая для лечения рака или иммунных расстройств, сопровождающихся иммунодефицитами, включающая фармацевтически эффективное количество слитого белка IL- 7 по п.1 или 2 вместе с фармацевтически приемлемым носителем, разбавителем или наполнителем.
27. Применение слитого белка по п.1 или 2 для получения лекарственного средства для лечения рака и иммунных расстройств, сопровождающихся иммунодефицитами.
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| ИММУНОКОНЪЮГАТЫ, СПОСОБ ПОЛУЧЕНИЯ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1995 |
|
RU2157701C2 |

