Область техники изобретения
Изобретение относится к комбинированной терапии для лечения опухолей и метастаз опухолей, которая предусматривает введение антиангиогенных средств и фактора некроза опухоли альфа (TNFα) или молекулы, обладающей биологической активностью TNFα, необязательно совместно с другими цитотоксическими средствами, такими как гамма-интерферон (IFNγ), или химиотерапевтическими средствами, такими как цисплатин, или ингибиторами ErbB рецептора, такими как антитела к EGFR. Способ и фармацевтические композиции, содержащие указанные средства, могут обладать синергетически усиленным ингибирующим действием на пролиферацию опухолевых клеток каждого индивидуального терапевтического средства, обеспечивая более эффективное лечение по сравнению с введением каждого индивидуального компонента по отдельности.
Предпосылки создания изобретения:
Ангиогенез, который также называется неоваскуляризацией, представляет собой процесс васкуляризации ткани, который характеризуется вовлечением в процесс роста новых развивающихся кровеносных сосудов в ткани. Этот процесс опосредуется инфильтрацией эндотелиальных клеток и гладкомышечных клеток. Полагают, что процесс осуществляется по одному из трех путей: (1) сосуды могут отрастать из ранее существовавших сосудов; (2) развитие сосудов de novo может осуществляться из клеток-предшественников (образование и развитие сосудов); или (3) существующие небольшие сосуды могут увеличиваться в диаметре (Blood и др., 1990, Bioch. Biophys. Acta 1032, 89. Известно, что эндотелиальные клетки кровеносных сосудов содержат по крайней мере пять RGD-зависимых интегринов, включая рецептор витронектина (αvβ3 или αvβ5), рецепторы коллагена типа I и IV, рецептор ламинина, рецептор фибронектина/ламинина/коллагена и рецептор фибронектина (Davis и др., 1993, J. Cell. Biochem. 51, 206). Известно также, что гладкомышечные клетки содержат по крайней мере шесть RGD-зависимых интегринов, включая αvβ3 и αvβ5.
Ангиогенез представляет собой важный процесс в неонатальном росте, но также играет существенную роль при заживлении ран и в патогенезе разнообразных клинически важных заболеваний, включая воспаление ткани, артрит, псориаз, злокачественные новообразования, диабетическую ретинопатию, дегенерацию желтого пятна и другие неоваскуолярные глазные заболевания. Эти заболевания, связанные с ангиогенезом, также называют ангиогенными заболеваниями (Folkman и др., 1987, Science 235, 442).
Ингибирование адгезии клеток in vitro с применением моноклональных антител, иммуноспецифических к различным α и β субъединицами интегрина, происходит с вовлечением рецептора витронектина αvβ3 в адгезию клеток разных типов, включая эндотелиальные клетки микрососудов (Davis и др., 1993, J. Cell. Biol. 51, 206).
Интегрины представляют собой класс клеточных рецепторов, известных тем, что они связывают белки внеклеточного матрикса и таким образом опосредуют взаимодействия между клетками и между клетками и внеклеточным матриксом, в результате чего происходит адгезия клеток. Рецепторы интегрина представляют собой семейство белков, имеющие общие структурные характеристики, а именно нековалентный гетеродимерный гликопротеиновый комплекс, образуемый α и β субъединицами. Для рецептора витронектина, названного так в связи с тем, что он предпочтительно первоначально связывается с витронектином, сейчас известно, что к нему относятся три разных интегрина, обозначенные как αvβ1, αvβ3 и αvβ5. αvβ1 связывает фибронектин и витронектин. αvβ3 связывает многие лиганды, включая фибрин, фибриноген, ламинин, тромбоспондин, витронектин и фактор Виллебранда. αvβ5 связывает витронектин. Очевидно, что имеются разные интегрины с разными биологическими функциями, так же как разные интегрины и субъединицы имеют совместную биологическую специфичность. Один важный участок распознавания в лиганде для многих интегринов представляет собой трипептидную последовательность аргинин-глицин-аспарагиновая кислота (RGD). RGD была обнаружена во всех лигандах, указанных выше, для интегриновых рецепторов витронектина.
Этот RGD участок распознавания может имитироваться линейными и циклическими (поли)пептидами, которые включают RGD последовательность. Такие RGD пептиды известны как ингибиторы или антагонисты, соответственно, функции интегрина. Представляется важным отметить, что, тем не менее, в зависимости от последовательности и структуры RGD пептида, специфичность ингибирующего воздействия может видоизменятся в зависимости от специфичности целевых интегринов. Разнообразные RGD полипетиды интегринов разной специфичности описаны, например, Cheresh, и др., 1989, Cell 58, 945, Aumailley и др., 1991, FEBS Letts. 291, 50, и в многочисленных заявках на патент и патентах (например, патенты US 4,517,686, 4,578,079, 4,589,881, 4,614,517, 4,661,111, 4,792,525; ЕР 0770622).
Образование новых кровеносных сосудов, или ангиогенез, играет ключевую роль в росте злокачественных новообразований, и поэтому значительный интерес вызывает разработка веществ, которые ингибируют ангиогенез (см., например, Holmgren и др., 1995, Nature Medicine 1, 149; Folkman, 1995, Nature Medicine 1, 27; O′Reilly и др., 1994, Cell 79, 315). Применение антагонистов интегрина αvβ3 для ингибирования ангиогенеза известно в методах ингибирования роста солидных опухолей путем снижения кровоснабжения солидных опухолей (см., например, патенты US 5,753,230 и US 5,766,591, в которых описано применение антагонистов αvβ3, таких как синтетические полипептиды, моноклональные антитела и миметики αvβ3, которые связывают рецептор αvβ3 и ингибируют ангиогенез). Способы и композиции для ингибирования αvβ5-опосредованного ангиогенеза тканей с применением антагонистов рецептора витронектина αvβ5 описаны в WO 97/45447.
Ангиогенез характеризуется инвазией, миграцией и пролиферацией эндотелиальных клеток, этот процесс зависит от взаимодействия клеток с компонентами внеклеточного матрикса. В этой ситуации, рецепторы интегрина между клеткой и матриксом опосредуют распространение и миграцию клеток. Было показано, что рецепторы адгезии эндотелия интегрина αvβ3 играют ключевую роль, являясь специфической сосудистой мишенью при антиангиогенном лечении (Brooks и др., 1994, Science 264, 569; Friedlander и др., 1995, Science 270). Участие интегринов αvβ3 сосудов в ангиогенезе было продемонстрировано на нескольких моделях in vivo, в которых развитие новых кровеносных сосудов в трансплантированных человеческих опухолях полностью ингибировалось или при системном введении пептидных антагонистов интегрина αvβ3 и αvβ5, как указано выше, или, альтернативно, при помощи антитела LM609 к αvβ3 (Brooks и др., 1994, Cell 79, 1157; АТСС НВ 9537). Это антитело блокирует рецептор интегрина αvβ3, активация которого естественными лигандами ингибирует апоптоз пролиферирующих ангиогенных сосудистых клеток и таким образом нарушает формирование вновь развивающихся кровеносных сосудов, что является существенным аспектом для пролиферации опухолей. Тем не менее, в последнее время было обнаружено, что клетки меланомы способны образовывать паутиноподобные структуры кровеносных сосудов даже при отсутствии эндотелиальных клеток (Barinaga 1999, Science 285, 14), вследствие этого опухоли могут быть толерантными к некоторым антиангиогенным лекарственным средствам, которые являются эффективными только в присутствии эндотелиальной ткани.
Разнообразные молекулы, которые стимулируют пролиферацию, миграцию и агрегацию эндотелия, включают VEGF, Ang1 и bFGF, и представляют собой естественные факторы выживания. VEGF (фактор роста эндотелия сосудов) был идентифицирован в качестве селективного ангиогенного фактора роста, который может стимулировать митоз эндотелиальных клеток. Предполагают, что VEGF, в частности, является основным медиатором ангиогенеза в первичных опухолях и при ишемических глазных заболеваниях. VEGF представляет собой гомодимер (MB: 46000) и является специфическим ангиогенным фактором эндотелиальных клеток (Ferrara и др., 1992, Endocrin. Rev., 13, 18) и фактором проницаемости сосудов (Senger и др., 1986, Cancer Res., 465629), который с высоким сродством связывается с мембраносвязаными рецепторами с тирозинкиназной активностью (Jakeman и др., 1992, J.Clin. Invest., 89, 244). Материал, полученный при биопсии опухолей человека, вызывает усиление экспрессии мРНК VEGF в злокачественных клетках и мРНК рецептора VEGF в смежных эндотелиальных клетках. Максимальная экспрессия VEGF обнаруживается в участках опухоли, смежных с участками некрозов кровеносных сосудов (см. обзор Thomas и др., 1996, J.Biol. Chem. 271(2), 603; Folkman, 1995, Nature Medicine 1, 27). В заявке WO 97/45447 описано вовлечение интегрина αvβ5 в процесс образования новых сосудов, который, в частности, индуцируется VEGF, EGF и TGF-α, и обнаруживается, что антагонист αvβ5 может ингибировать ангиогенез, вызванный VEGF. Для эффективной противоопухолевой терапии также может использоваться в качестве мишени рецептор VEGF для ингибирования ангиогенеза, применяя моноклональные антитела (Witte и др., 1998, Cancer Metastasis Rev. 17(2), 155). Известно, что MAb DC-101 ингибирует ангиогенез клеток опухоли.
Тирозинкиназы представляют собой класс ферментов, которые катализируют перенос конечного фосфата аденозинтрифосфата на остатки тирозина в белковых субстратах. Полагают, что тирозинкиназы при помощи субстратного фосфорилирования играют решающую роль в передаче сигнала при разнообразных клеточных функциях. Несмотря на то что точно механизмы передачи сигнала все еще неясны, для тирозинкиназ было показано, что они являются важными факторами, которые принимают участие в пролиферации клеток, канцерогенезе и дифференциации клеток.
Тирозинкиназы могут быть разделены на тирозинкиназы рецепторного типа и нерецепторного типа. Оба типа тирозинкиназ - рецепторного и нерецеторного типа - вовлечены в пути передачи сигналов в клетках, приводящие к многим патогенным состояниям, включая злокачественные новообразования, псориаз и гипериммунные ответы. Многие тирозинкиназы вовлечены в рост клеток, а также в ангиогенез.
Тирозинкиназы рецепторного типа имеют внеклеточную, трансмембранную и внутриклеточную части, в то время как тирозинкиназы нерецепторного типа являются полностью внутриклеточными. Тирозинкиназы, связанные с рецептором, представляют собой трансмембранные белки, которые включают внеклеточный лиганд-связывающий домен, трансмембранную последовательность и цитоплазматический домен тирозинкиназы. Тирозинкиназы рецепторного типа включают большое количество трансмембранных рецепторов с разнообразной биологической активностью. В действительности, были идентифицированы разные подсемейства тирозинкиназ рецепторного типа. Такие тирозинкиназы включают рецепторы фактора роста фибробластов (FGF), рецепторы фактора роста эпидермиса (EGF) семейства основного класса ErbB и рецепторы фактора роста производных тромбоцитов (PDGF). Также сюда включают рецепторы фактора роста нервов (NGF), рецепторы зависимого от головного мозга нейротрофического фактора (BDNF) и рецепторы нейротрофина-3 (NT-3) и рецепторы нейротрофина-4 (NT-4).
Одно подсемейство тирозинкиназы рецепторного типа, обозначенное как подсемейство HER или ErbB, включает EGFR (ErbB1), HER2 (ErbB2 или p185neu), HER3 (ErbB3) и HER4 (ErbB4 или tyro2). Лиганды этого подсемейства рецепторов включают фактор роста эпителия (EGF), TGF-α, амфирегулин, НВ-EGF, бетацеллюлин и херегулин. Подсемейство PDGF включает семейство FLK, которое включает рецептор внутреннего домена киназы (KDR).
EGFR, кодируемый геном erbB1, вовлечен и является одной из причин возникновения злокачественных опухолей у людей. В частности, усиленная экспрессия EGFR наблюдается при раке молочной железы, мочевого пузыря, легких, головы, шеи и желудка, а также при глиобластомах. Усиленная экспрессия рецептора EGFR часто связана с усиленной выработкой лиганда EGFR, трансформирующего фактора роста альфа (TGF-α), в подобных опухолевых клетках происходит путем активации рецептора при помощи аутокринной стимуляции (Baselga и Mendelsohn, Pharmac. Ther. 64:127-154 (1994)). Рецептор EGF представляет собой трансмембранный гликопротеин, который имеет молекулярный вес 170,000, и был обнаружен во многих типах эпителиальных клеток. Он активируется по крайней мере тремя лигандами, EGF, TGF-α (трансформирующий фактор роста альфа) и амфирегулином. Было показано, что и фактор роста эпидермиса (EGF), и трансформирующий фактор роста альфа (TGF-α) связываются с рецептором EGF и приводят к пролиферации клеток и росту опухолей. Эти факторы роста не связываются с HER2 (Ulrich и Schlesinger, 1990, Cell 61, 203). В противоположность некоторым семействам факторов роста, которые идуцируют димеризацию рецептора в силу их димерной природы (например, PDGF), мономерные факторы роста, такие EGF, включают два связывающих участка к их рецепторам, и, следовательно, могут перекрестно связывать два соседних рецептора EGF (Lemmon и др., 1997, EMBO J. 16, 281). Димеризация рецептора является необходимой для стимуляции присущей каталитической активности и для аутофосфорилирования рецепторов факторов роста. Следует отметить, что рецепторы тирозинкиназы белков (PTKs) способны подвергаться как гомо-, так и гетеродимеризации.
Было показано, что антитела к рецептору EGF, хотя блокируют связывание EGF и TGF-α с рецептором, способны ингибировать пролиферацию опухолевых клеток. Принимая во внимание эти полученные данные, были выявлены множество мышиных и крысиных моноклональных антител к рецептору EGF и изучена их способность ингибировать рост опухолевых клеток in vitro и in vivo (Modjtahedi и Dean, 1994, J.Oncology 4, 277). Гуманизированное моноклинальное антитело 425 (hMAb 425, US 5.558,864; ЕР 0531472) и химерное моноклональное антитело 225 (cMAb 225, US 4,943,533 и ЕР 0359282), оба к рецептору EGF, выявляют эффективность при клинических исследованиях. Было показано, что антитело С225 ингибирует EGF-опосредованный рост опухолевых клеток in vitro и ингибирует образование человеческих опухолей in vivo у бесшерстых мышей. Кроме того, антитела обнаруживают, прежде всего, синергетический эффект с определенными химиотерапевтическими веществами (например, доксорубицином, адриамицином, таксолом и цисплатином) и уничтожают человеческие опухоли in vivo на моделях мышиных ксенотрансплантантов. Ye и др. (1999, Oncogene 18, 731) показали, что человеческие раковые клетки яичника могут успешно поддаваться лечению комбинацией обоих cMab 225 и гуманизированного MAb 4D5 к рецептору HER2.
Другой представитель семейства ErbB, HER2 (ErbB2 или р185nеu) был первоначально идентифицирован в качестве продукта трансформации гена из нейробластом крыс, подвергнутых химиотерапии. Активированная форма neu протоонкогена возникает вследствие точечной мутации (валина на глутаминовую кислоту) в трансмембранном участке кодуруемого белка. Амплификация человеческого гомолога neu наблюдается при раке молочной железы и яичников и коррелирует с неблагоприятным прогнозом (Slamon и др., Science, 235: 177-182 (1987); Slamon и др., Science, 244:707-712 (1989); US 4,968,603). ErbB2 (HER2) имеет молекулярный вес приблизительно 185,000, и обладает значительной гомологией к рецептору EGF (HER1), несмотря на то, что специфический лиганд для HER2 до сих пор еще не был окончательно идентифицирован.
Антитело 4D5 к рецептору HER2 дополнительно обнаруживает чувствительность к цитотоксическим действиям TNFα в клеточных линиях опухолей молочной железы, которые сверхэкспрессируют ErbB2 (патент US 5,677,171). Рекомбинантная гуманизированная модификация мышиного антитела 4D5 к ErbB2 (huMAb4D5-8, rhuMAb HER2 или HERCEPTIN®; US 5,821,337) обладает клинической активностью у пациентов с метастазами опухолей молочной железы, что сверхэкспрессируют ErbB2, которые предварительно получали интенсивное противораковое лечение (Baselga и др., J. Clin. Oncol. 14:737-744 (1996)). HERCEPTIN® был получен при испытаниях в 1998 для лечения пациентов с метастазами рака молочной железы, при этом опухоли сверхэкспрессируют белок ErbB2.
TNFα относится к большому семейству молекул, которое включает важные цитокины, такие как лиганд Fas, лиганд CD40, TRAIL, лимфотоксин и другие (Locksley и др., 2001, Cell 104:487-501). Кроме того, что он высвобождается из клеток разных типов, TNFα также существует в связанном с клеточной мембраной виде, в высокомолекулярной форме в клетках, и эта форма также является медиатором многих биологических действий. Полагают, что TNFα имеет некоторое значение в нормальном развитии и физиологии; однако он выявляет пагубные и деструктивные влияния во многих тканях при разных болезненных состояниях (Tracey и др., Ann. Rev. Med 1994; 45:491). Болезненные состояния, в которых TNFα играет значительную роль, включают септический шок, раковую кахексию, ревматоидный артрит и т.п.
Впервые TNFα человека был очищен в 1985 (см. Aggarwal и др., J.Biol. Chem. 1985, 260, 2345-2354). Вскоре после этого было завершено молекулярное клонирование кДНК TNF и клонирован локус TNF человека (Pennica и др., Nature 1984, 312, 124-729; Wang и др., Nature 1985, 313, 803-806). TNFα представляет собой тримерный полипептид с молекулярным весом 17 кДа, который главным образом вырабатывается макрофагами. Этот пептид вначале экспрессируется в виде трансмембранного белка с молекулярным весом 26 кДа, от которого отщепляется и высвобождается субъединица 17 кДа после протеолитического расщепления. TNFα вырабатывается, как правило, разнообразными клетками, например активированными макрофагами и фибробластами. Сообщалось, что TNFα индуцирует много разнообразных факторов. Также сообщалось, что TNFα также принимает участие, непосредственно или опосредованно, в разных заболеваниях, таких как инфекционные заболевания, аутоимунные заболевания, такие как системная красная волчанка (SLE) и артрит, СПИД, сепсис и некоторые другие типы инфекций.
TNFα и воспалительная ответная инфекция и повреждение ткани стимулирует каскад биохимических изменений, которые инициируют начало сложных реакций иммунной системы, которые совместно представляют собой воспалительную ответную реакцию. Развитие этой ответной реакции основывается, по крайней мере частично, на местном расширении сосудов или увеличении проницаемости сосудов и активации эндотелия кровеносных сосудов, что дает возможность белым кровяным тельцам эффективно циркулировать и мигрировать к поврежденному участку, и таким образом увеличивается вероятность их связывания с и разрушения любых антигенов. Полагают, что эндотелий кровеносных сосудов затем может активироваться или воспаляться. Как правило, воспаление представляет собой желательный иммунный ответ на разные непредвиденные воздействия, и по существу характеризуется быстрым началом и является непродолжительным (острое воспаление). Однако его длительная или неконтролируемая активность (хроническое воспаление) оказывает пагубное воздействие на организм и играет роль в патогенезе определенных иммунологических заболеваний, таких как септический шок, ревматоидный артрит, воспалительные заболевания кишечника и застойная сердечная недостаточность (см. "TNF и суперсемейство рецепторов TNF" в "Цитокины и рецепторы цитокинов", под ред. Bona и Revillard, Harvard Academic Publishers, Амстердам 2000, стр.118-148).
TNFα, также как и многие другие цитокины, секретируется макрофагами вскоре после инициации воспалительной ответной реакции и индуцирует коагуляцию, повышает проницаемость кровеносных сосудов и активирует экспрессию адгезионных молекул в клетках эндотелия сосудов.
TNFα никогда не является полностью полезным либо полностью вредным для хозяина. Таким образом, TNFα является эффективным модулятором функции эндотелиальных клеток. В зависимости от состояния сосудов он вызывает воспаление путем индуцирования активации эндотелиальных клеток и выживания или он вызывает некроз ткани путем индуцирования апопроза эндотелиальных клеток и разрушения сосудов (Pober, J.S., Pathol Biol (Paris) 46, 159-163 (1998); Aggarwal, & Natarajan, Eur. Cytokine Netw. 7, 93-124 (1996)). Многие внутриклеточные пути передачи сигналов опосредуются этими двумя различными ответными реакциями (Wallach и др., Annual Review of Immunology 17, 331-367 (1999)), но передача внеклеточных сигналов, определяющая, какое из двух воздействий TNFα осуществляется - выживание или гибель, являются непонятными.
Предпочтительно соотношение между его выработкой и регуляцией является гарантией, что хозяин может эффективно реагировать на поражение микроорганизмами без риска для состояния своего здоровья. Являясь медиатором воспаления, TNFα помогает организму в его борьбе против бактериальных инфекций и повреждении тканей при помощи повторной иммунизации соответствующего иммунного ответа. Тем не менее, его избыточная выработка приводит к хроническому воспалению, обладает пагубными воздействиями на организм и играет важную роль в патогенезе отдельных заболеваний.
IFNγ представляет собой эффективный усилитель TNFα (Dealtry и др., Eur Immunol, 17, 689-693, (1987)). Если TNFα вызывает апоптоз клеток, активация NF-κВ, транскрипционного фактора, который содействует выживанию клеток, может подавлять апоптоз, который индуцируется TNFα (Van Antwerp и др., Science 274, 787-789 (1996)).
TNFα принимает участие в передаче разнообразных клеточных сигналов, которые приводят к клеточным ответам, как, например, пролиферация, активация, дифференциация, но также и запрограммированная гибель клеток. Передачу клеточных сигналов к TNF(можно подразделить на ранние ответы, такие как активация киназ, фосфатаз, липаз, протеаз и факторов транскрипции, и поздние ответы, и таким образом более непрямые ответные реакции, такие как нарушение электрон-транспортной цепи в митохондриях, выработка радикалов, выработка оксида и высвобождение различных веществ. Многие ранние ответные реакции клеток, такие как восстановление домена гибели, содержащего переходные белки, активация NFKB или активация каспазы, также инициируются при помощи связывания других представителей семейства лигандов TNF с их соответствующими рецепторами. Соответственно, молекулы, подобные лимфотоксину, лиганду Fas или TRAIL могут взаимодействовать с избытком TNF (Grell и Clauss, I.c.).
Интегрин-опосредованная адгезия к внеклеточному матриксу (ЕСМ) является существенной для выживания большинства клеток, включая эндотелиальные клетки. Например, интегрин сосудов αvβ3 содействует пролиферации и выживанию ангиогенных эндотелиальных клеток и антагонисты αvβ3 индуцируют апоптоз ангиогенных эндотелиальных клеток и подавляют ангиогенез (Brooks и др., Cell 79, 1157-1164 (1994). Некоторые биохимические реакции, связанные с интегрин-опосредованным клеточным выживанием, включая активацию PI 3-K/AKT (Khwaja и др., Embo Journal 16, 2783-2793 (1997)) и NF-κВ (Scatena и др., J.Cell Biol 141, 1083-1093 (1998)) были идентифицированы в качестве путей передачи сигналов. Кроме интегринов, молекулы адгезии между клетками РЕСАМ-1 и VE Кадхерин также способствуют выживанию эндотелиальных клеток (Bird и др. J Cell Sci 112, 1989-1997 (1999); Carmeliet и др. Cell 98, 147-157 (1999)).
TNF является цитотоксическим для некоторых линий опухолевых клеток, но большинство из них почти не поддаются влиянию в процессе роста. Поэтому является маловероятным, что противоопухолевые воздействия TNF в некоторых моделях на животных (Balkwill и др., Cancer Res. 46: 3990-3993 (1986)) обусловлены прямым действием цитокинов на опухолевые клетки. В некоторых исследованиях было показано, что опосредованные механизмы хозяина вовлечены в регрессию опухоли, которая инициируется TNF (Manda и др., Cancer Res. 47: 3707-3711 (1987)). Накопление данных свидетельствует о том, что геморрагический некроз опухолей, который вызывается TNF, инициируется в эндотелиальных клетках уровнем внутриопухолевых сосудов (Havell и др., J. Exp.Med. 167: 1967-1985 (1988)).
Результаты клинических исследований TNF у раковых больных являются в общем, неутешительными (обзор Haranaka, J.Biol. Response Mod. 7: 525-534 (1988)). Обычно противоопухолевые действия TNF ограничиваются значительными побочными эффектами. Одним из подходов к ограничению побочных действий TNF является получение TNF мутантов, которые обнаруживают каждый активности рецептора TNF 1-специфического типа или разнообразные фармакодинамические свойства (Brouckaert и др., Circ. Shock 43: 185-190 (1994); Eggermont, Anticancer Res. 18: 3899-3905 (1998); Lucas и др., Int. J.Cancer 15: 543-549 (2001)). В последнее время наблюдаются успехи в лечении пациентов, которые страдают от меланом или сарком конечностей. Существенные положительные результаты могут быть получены при помощи технологии изолированной перфузии. Чрезмерные дозы TNF вплоть до 4 мг применяются в комбинации с цитостатиками или IFN (Lienard и др., J.Clin. Oncol. 10:52-60 (1992)). Местные ответные реакции, включая сильное размягчение и покраснение опухоли, связанные с сильной воспалительной ответной реакцией, подобны к опосредованным TNF противоопухолевым воздействиям в мышиных системах.
Было показано, что такое лечение пациентов с метастатической меланомой конечностей избирательно разрушает сосудистую сеть опухоли, но не повреждает покоящиеся интактные сосуды. Это воздействие связано с супрессией, индуцированной TNF и IFNγ, интегриновой αVβ3-функции в эндотелиальных клетках in vitro и индукцией апоптоза эндотелиальных клеток in vivo (Ruegg и др., Nature Med 4, 408-414 (1998)). Эти результаты свидетельствуют о том, что TNF в сочетании с дополнительными терапевтическими средствами может являться очень клинически эффективным при лечении некоторых опухолей, при условии, что может контролироваться системная токсичность.
В настоящем изобретении описывается, что молекулы, которые принимают участие в ангиогенезе, такие как интегрины, при модулировании активности TNFα, могут быть использованы напрямую в клиническом применении TNFα в качестве противоракового средства. Совместное введение антиангиогенных средств с TNFα, предпочтительно антагонистов интегрина, может избирательно повышать чувствительность рецепторов ангиогенеза, связанных с эндотелиальными клетками в отношении апоптотической активности TNF, в результате чего происходит усиленное разрушение сосудов опухоли. Кроме того, эта комбинированная терапия может способствовать уменьшению доз TNF для уменьшения системных побочных эффектов TNF.
Сущность изобретения
Настоящее изобретение впервые описывает новую концепцию лечения опухолей, которая предусматривает введение особи средства, которое блокирует или ингибирует ангиогенез, совместно с TNFα, мутантами TNF или TNF-подобными молекулами. Необязательно композиция в соответствии с данным изобретением включает дополнительные терапевтически активные соединения, предпочтительно выбранные из группы, включающей цитотоксические средства, химиотерапевтические средства и ингибиторы или антагонисты рецепторов семейства тирозинкиназы ErbB, такие как описаны далее более подробно. Таким образом, изобретение относится к фармацевтическим композициям, содержащим, предпочтительно, антиангиогенные средства, антагонисты интегрина (рецептора) и TNFα, мутанты TNF или TNF-подобные молекулы в терапевтически эффективном количестве. В частности, изобретение относится к фармацевтическим композициям, содержащим линейный или циклический RGD-пептиды и TNFα необязательно совместно с IFNγ. Предпочтительная композиция в соответствии с изобретением включает циклический пептид цикло-(Arg-Gly-Asp-DPhe-NMe-Val), TNFα и IFNγ. В соответствии с настоящим изобретением указанные терапевтические активные средства также могут быть представлены в виде фармацевтического набора, включающего упаковку, содержащую одно или более антиангиогенных средств, TNFα, и, необязательно, одно или более цитотоксических /химиотерапевтических средств/ анти-ErbB средств в единичных упаковках или в отдельных контейнерах. Изобретение относится, в частности, к комбинированной терапии, предусматривающей применение и введение, соответственно, двух или более молекул, где по крайней мере одна молекула обладает ингибиторной активностью по отношению к ангиогенезу и другая представляет собой TNFα. Кроме того, изобретение относится к комбинированной терапии, предусматривающей введение только одной (слитой) молекулы, имеющей антиангиогенную активность и активность TNFα, необязательно совместно с одним или более цитотоксическим/химиотерапевтическим средствами. Например, слитый белок, состоящий по существу из цикло-(Arg-Gly-Asp-DPhe-NMe-Val), который непосредственно присоединен к TNFα, или при помощи линкерной молекулы, может вводиться пациенту. Другим примером является антитело к интегрину, такое как LM609, как описано ниже, которое присоединено на С-конце его Fc-участка к TNFα. Дополнительным примером является биспецифическое антитело, присоединенное к TNFα, в котором одна часть является специфичной к рецептору интегрина или рецептору VEGF и другая часть специфичная к рецептору EGF.
В большинстве случаев введение может сопровождаться лучевой терапией, при этом лучевая терапия может осуществляться в основном одновременно или перед или после введения лекарственного средства. Введение различных средств в комбинированной терапии в соответствии с изобретением также может осуществляться в основном одновременно или последовательно. Опухоли, которые на поверхностях своих клеток несут рецепторы, вовлеченные в развитие кровеносных сосудов опухоли, могут успешно лечиться комбинированной терапией в соответствии с настоящим изобретением.
Известно, что опухоли используют альтернативные пути для своего развития и роста. Если один путь блокирован, они часто обладают способностью переключаться на другой путь путем экспрессии и использования других рецепторов и других путей передачи сигналов. Поэтому фармацевтические композиции в соответствии с настоящим изобретением могут блокировать несколько таких возможных стратегий развития опухоли и, следовательно, обеспечивать различные преимущества. Комбинации в соответствии с настоящим изобретением полезны для лечения и предотвращения опухолей, опухолеподобных нарушений, неоплазий и метастаз опухолей, что далее описывается более подробно. Предпочтительно сочетания различных средств по настоящему изобретению вводятся в комбинации в низких дозах, при этом такие дозы являются ниже тех доз, которые обычно используются в медицине. Преимущество снижения доз соединений, композиций, средств и терапий по настоящему изобретению, что вводятся особи, состоит в уменьшении количества побочных эффектов, связанных с введением высоких доз. Например, при снижении дозы химиотерапевтического средства, такого как метотрексат, происходит уменьшение частоты и степени тошноты и рвоты по сравнению с наблюдаемыми при введении высших доз. При снижении количества побочных действий улучшается качество жизни раковых больных. Дополнительными преимуществами снижения количества побочных эффектов является улучшение соблюдения больным режима и схемы лечения, снижение числа госпитализаций, необходимых для лечения побочных эффектов, и уменьшение введения обезболивающих средств, необходимых для лечения боли, связанной с побочными действиями. Альтернативно, способы и комбинации по настоящему изобретению могут также обеспечивать максимальную терапевтическую эффективность при высших дозах.
Комбинации в соответствии с изобретением выявляют неожиданный синергетический эффект. При введении комбинации лекарственных средств действительно наблюдается сокращение и распад опухоли при клинических исследованиях, в то время как не наблюдается существенного неблагоприятного взаимодействия лекарственных средств.
Более подробно, изобретение относится к:
- фармацевтической композиции, содержащей в терапевтически эффективном количестве по крайней мере (i) одно антиангиогенное средство и (ii) фактор некроза опухоли альфа (TNFα) или молекулу, обладающую биологической активностью TNFα, необязательно совместно с фармацевтически приемлемым носителем, наполнителем или растворителем;
- соответствующей фармацевтической композиции, в которой указанное антиангиогенное средство представляет собой ингибитор/антагонист интегрина (рецептора) или ингибитор/антагонист VEGF (рецептора);
- соответствующей фармацевтической композиции, в которой указанный ингибитор/антагонист рецептора интегрина представляет собой линейный или циклический пептид, который содержит RGD;
- соответствующей фармацевтической композиции, в которой указанный пептид, который содержит RGD, представляет собой цикло-(Arg-Gly-Asp-DPhe-NMeVal);
- соответствующей фармацевтической композиции, в которой указанное антиангиогенное средство представляет собой антитело или его иммунотерапевтическиактивный фрагмент, которое связывается с рецептором интегрина или рецептором VEGF;
- соответствующей фармацевтической композиции, в которой указанное антиангиогенное средство и TNFα связаны вместе с образованием одной слитой молекулы;
- соответствующей фармацевтической композиции, которая дополнительно содержит по крайней мере одно цитотоксическое и/или химиотерапевтическое средство;
- соответствующей фармацевтической композиции, в которой указанное цитотоксическое средство представляет собой гамма-интерферон (IFNγ) и/или другой эффективный цитокин;
- соответствующей фармацевтической композиции, в которой указанное химиотерапевтическое соединение выбирают из группы, включающей цисплатин, доксорубицин, гемцитабин, доцетаксел, паклитаксел (таксол), блеомицин;
- соответствующей фармацевтической композиции, которая дополнительно содержит ингибитор или антагонист рецептора ErbB семейства тирозинкиназы;
- соответствующей фармацевтической композиции, в которой указанный ингибитор представляет собой антитело к EGFR, антитело к HER2 или его иммунотерапевтическиактивный фрагмент;
- фармацевтическому набору, который включает упаковку, содержащую (i) по крайней мере одно антиангиогенное средство, предпочтительно ингибитор/антагонист рецептора интегрина, (ii) TNFα и необязательно (iii) дополнительное цитотоксическое и/или химиотерапевтическое средство;
- соответствующему предпочтительному фармацевтическому набору, содержащему
(i) цикло(Arg-Gly-Asp-DPhe-NMeVal), (ii) TNFα и (iii) IFNγ и необязательно (iii) дополнительное цитотоксическое и/или химиотерапевтическое средство и/или ингибитор или антагонист рецептора ErbB семейства тирозинкиназы;
- соответствующему фармацевтическому набору, в котором указанные фармацевтически активные средства находятся в отдельных контейнерах в указанной упаковке;
- применению фармацевтической композиции, как указано выше и в пунктах формулы изобретения, для приготовления лекарственного средства или композиции лекарственных средства для лечения опухолей и метастаз опухолей; и
- способу лечения опухолей или метастаз опухолей у особи, который предусматривает введение указанной особи одновременно или последовательно терапевтически эффективных фармацевтических композиций, как указано выше.
Подробное описание изобретение
Если не указанно другое, термины и фразы, применяемые в данном изобретении, имеют значения и определения, указанные ниже. Кроме того, эти определения и значения описывают изобретение более подробно, включая предпочтительные варианты осуществления.
"Биологические молекулы" включают естественные или синтетические молекулы, имеющие, как правило, молекулярный вес больше чем приблизительно 300, и предпочтительно представляют собой поли- и олигосахариды, олиго- и полипептиды, белки, пептиды, поли- и олигонуклеотиды, а также гликозилированные производные липидов. Как правило, биологические молекулы включают иммунотерапевтические средства, прежде всего антитела или их фрагменты, или функциональные производные этих антител или фрагментов, включая слитые белки.
"Рецептор" или "рецепторная молекула" представляет собой растворимый или мембранно-связанный/ассоциированный протеин или гликопротеин, содержащий один или более доменов, с которыми связываются лиганды с образованием комплекса рецептор-лиганд. При связывании с лигандом, которым может быть агонист или антагонист, происходит активация или инактивация рецептора и может инициироваться или блокироваться путь передачи сигнала.
Под "лигандом" или "лигандом рецептора" подразумевается естественное или синтетическое соединение, которое связывается с молекулой рецептора с образованием комплекса рецептор-лиганд. Под понятие лиганд подпадают агонисты, антагонисты и соединения с частичным агонистичным/антагонистичным действием. В соответствии с характерными особенностями данного изобретения этот термин включает, прежде всего, TNF-подобные лиганды.
Понятие "TNFα" как применяется в настоящем изобретении, включает, если не указано иное, все виды молекул TNF и молекул, обладающих биологической активностью TNFα, включая естественные и синтетические, пептидные или непептидные мутанты TNF, варианты или TNF-подобные лиганды. Предпочтительно понятие включает естественный пептидный TNFα.
"Агонист" или "агонист рецептора" представляет собой естественное или синтетическое соединение, которое связывается с рецептором с образованием комплекса рецептор-агонист, при этом происходит активирование указанного рецептора и комплекса рецептор-агонист, соответственно, инициализируется путь передачи сигнала либо другой биологический процесс.
Под "антагонистом" или "антагонистом рецептора" подразумевается естественное или синтетическое соединение, которое обладает противоположным к агонисту биологическим действием. Антагонист связывается с рецептором и блокирует активацию рецептора агонистом путем конкуренции с агонистом за рецептор. Антагонист определяется путем его способности блокировать действие агониста. Антагонист рецептора также может представлять собой антитело или его иммунотерапевтически эффективный фрагмент. Предпочтительные антагонисты в соответствии с настоящим изобретением приведены и обсуждаются ниже.
Понятие "терапевтически эффективное" или "терапевтически эффективное количество" относится к количеству лекарственного средства, эффективного для лечения заболевания или расстройства у млекопитающего. В случае злокачественного заболевания, терапевтически эффективное количество лекарственного средства может уменьшать число раковых клеток; уменьшать размер опухоли; ингибировать (например, несколько замедлять распространение и предпочтительно останавливать) инфильтрацию раковых клеток в периферические органы; ингибировать (например, несколько замедлять распространение и предпочтительно останавливать) метастазирование опухолей; ингибировать, несколько замедлять, рост опухоли; и/или ослаблять распространение одного или более симптомов, связанных с раком. В этом отношении лекарственное средство может предотвращать рост и/или уничтожать существующие раковые клетки, оно может быть цитостатическим и/или цитотоксическим. При терапии рака эффективность, например, может определяться путем измерения времени прогрессирования заболевания (ТТР) и/или измерения ответной реакции (RR).
Понятие "иммунотерапевтически эффективное" относится к биологическим молекулам, которые вызывают иммунный ответ у млекопитающего. Более конкретно, понятие относится к молекулам, которые могут распознавать и связывать антиген. Типично, антитела, фрагменты антител и слитые белки антитела, которые включают их антигенсвязывающие участки (комплементарные определяющие участки, CDRs), являются иммунотерапевтическиэффективными.
Понятие "пролекарство" как применяется в настоящем изобретении, относится к предшественнику или производной форме фармацевтически активного вещества, который является менее цитотоксическом по отношению к клеткам опухоли по сравнению с исходным лекарственным средством и способен активироваться под влиянием ферментов или превращаться в более активную исходную форму (см., например, "Пролекарства в химиотерапии рака", Biochemical Society Transactions, 14, стр.375-382, 615-ое собрание, Белфаст (1986)).
"Антиангиогенное средство" относится к естественному или синтетическому соединению, которое блокирует или в некоторой степени препятствует развитию кровеносных сосудов. Антиангиогенная молекула может, например, быть биологически активной молекулой, которая связывается и блокирует ангиогенный фактор роста или рецептор фактора роста. Предпочтительная антиангиогенная молекула при этом связывается с рецептором, предпочтительно рецептором интегрина или рецептором VEGF. Понятие в соответствии с изобретением также включает Пролекарства указанного антиангиогенного средства.
Существует много молекул, которые имеют разную структуру и исходный участок, который определяет антиангиогенные свойства. Более предпочтительные классы средств, которые ингибируют или блокируют ангиогенез, которые являются подходящими в соответствии с настоящим изобретением, представляют собой, например:
(i) антимитотические средства, такие как фторурацил, митомицин-С, таксол;
(ii) метаболиты эстрогена, такие как 2-метоксиэстрадиол;
(iii) ингибиторы матриксной металлопротеиназы (ММР), которые ингибируют цинкметаллопротеиназы (металлопротеазы) (например, бетимастат, ВВ 16, TIMPs, миноциклин, GM6001, или описанные в "Ингибирование матриксных металлопротеиназ: терапевтические применения" (Golub, Ежегодник Нью-йоркской Академии наук, том.878а; Грюневолд, Zucker (ред.), 1999);
(iv) антиангиогенные многофункциональные средства и факторы, такие как IFNα (патенты US 4,530,901; 4,503,035; 5,231,176); фрагменты ангиостатина и плазминогена (например, kringle 1-4, kringle 5, kringle 1-3 (O′Reilly, M.S. и др., Cell (Кембридж, Mass.) 79(2): 315-328, 1994; Сао и др., J.Biol. Chem. 271: 29461-29467, 1996; Сао и др., J. Biol. Chem 272: 22924-22928, 1997); эндостатин (O′Reilly, M.S. и др.. Cell 88(2), 277, 1997 и WO 97/15666), тромбоспондин (TSP-1; Frazier, 1991, Curr Opin Cell Biol 3(5):792); тромбоцитарный фактор 4 (PF4);
(v) ингибиторы активатора плазминогена/урокиназы;
(vi) антагонисты рецептора урокиназы;
(vii) гепариназы;
(viii) аналоги фумагиллина, такие как TNP-470;
(ix) ингибиторы тирозинкиназы, такие как SUI 01 (многие из выше- и нижеупомянутых антагонистов рецептора ErbB (антагонисты EGFR/HER2) также являются ингибиторами тирозинкиназы, и могут демонстрировать, следовательно, блокирующую активность по отношению к рецептору EGF, что приводит к ингибированию роста опухоли, также как и антиангиогенную активность, в результате чего происходит ингибирование развития кровеносных сосудов и эндотелиальных клеток, соответственно);
(х) сурамин и аналоги сурамина;
(xi) ангиостатические стероиды;
(xii) антагонисты VEGF и bFGF;
(xiii) антагонисты рецептора VEGF, такие как антитела к рецептору VEGF (DC-101);
(xiv) flk-1 и flt-1 антагонисты;
(xv) ингибиторы циклооксигеназы-11 такие как COX-II;
(xvi) антагонисты интегрина и антагонисты рецептора интегрина, такие как αv антагонисты и антагонисты αv рецептора, например антитела к рецептору αv и RGD пептиды. Предпочтительными в соответствии с настоящим изобретением являются антагонисты интегрина (рецептора).
Понятие "антагонисты /ингибиторы интегрина" или "антагонисты/ ингибиторы рецептора интегрина" относится к естественной или синтетической молекуле, которая блокирует и ингибирует рецептор интегрина. В некоторых случаях, понятие включает антагонисты к лигандам указанных рецепторов интегрина (такие как для αvβ3: витронектин, фибрин, фибриноген, фактор Виллебранда, тромбоспондин, ламинин; для αvβ5: витронектин; для αvβ1: фибронектин и витронектин; для αvβ6: фибронектин).
Предпочтительными в соответствии с изобретением являются антагонисты к рецепторам интегрина. Антагонисты интегрина (рецептора) могут представлять собой естественные или синтетические пептиды, непептиды, пептидомиметики, иммуноглобулины, такие как антитела или их функциональные фрагменты, или иммуноконъюгаты (слитые белки). Предпочтительными в соответствии с изобретением ингибиторами интегрина являются такие, которые связываются с рецептором αv интегринов (например αvβ3, αvβ5, αvβ6 и подклассов).
Предпочтительными ингибиторами интегрина являются αv антагонисты, и, в частности, αvβ3 антагонисты. Предпочтительными αv антагонистами в соответствии с изобретением являются RGD пептиды, пептидомиметики (непептидные) антагонисты и антитела к интегриновому рецептору, такие как антитела, блокирующие αv рецепторы.
Типичные неиммунологические антагонисты αvβ3 описаны в исследованиях в патентах US 5,753,230 и US 5,766,591. Предпочтительные антагонисты представляют собой линейные и циклические пептиды, которые содержат RGD. Циклические пептиды являются, как правило, более стабильными и обладают увеличенным периодом полужизни в сыворотке. Наиболее предпочтительным антагонистам интегрина по изобретению, тем не менее, является цикло-(Arg-Gly-Asp-DPhe-NMeVal) (EMD 121974, Cilengitide®, Merck KgaA, Германия; ЕР 0770622), который является эффективным блокатором рецепторов интегрина αvβ3, αvβ1, αvβ6, αvβ5, αIIbβ3.
Подходящие пептидные антагонисты, а также пептидомиметические (непептидные) антагонисты αvβ3/αvβ5/αvβ6 рецептора интегрина описаны в научной и патентной литературе. Например, на них ссылаются Hoekstra и Poulter, 1998, Curr. Med. Chem. 5, 195; WO 95/32710; WO 95/37655; WO 97/01540; WO 97/37655; WO 97/45137; WO 97/41844; WO 98/08840; WO 98/18460; WO 98/18461; WO 98/25892; WO 98/31359; WO 98/30542; WO 99/15506; WO 99/15507; WO 99/31061; WO 00/06169; ЕР 0853084; ЕР 0854140; ЕР 0854145; US 5,780,426; и US 6,048,861. Заявки на патенты, в которых описаны бензазепины, а также родственные бензодиазепины и бензоциклогептены, которые являются антагонистами рецептора интегрина αvβ3, которые также являются пригодными для применения в соответствии с настоящим изобретением, включают WO 96/00574, WO 96/00730, WO 96/06087, WO 96/26190, WO 97/24119, WO 97/24122, WO 97/24124, WO 98/15278, WO 99/05107, WO 99/06049, WO 99/15170, WO 99/15178, WO 97/34865, WO 97/01540, WO 98/30542, WO 99/11626 и WO 99/15508. Другие антагонисты рецептора интегрина, характеризующиеся ограниченными изменениями структуры основного кольца, описаны в WO 98/08840; WO 99/30709; WO 99/30713; WO 99/31099; WO 00/09503; US 5,919,792; US 5,925,655; US 5,981,546; и US 6,017,926. В US 6,048,861 и WO 00/72801 описан ряд производных нонановых кислот, которые являются сильными антагонистами рецептора интегрина αvβ3. Другие химически небольшие молекулы антагонистов интегрина (главным образом антагонистов витронектина) описаны в WO 00/38665. Для других антагонистов рецептора αvβ3 показано, что они являются эффективными ингибиторами ангиогенеза. Например, синтетические антагонисты рецептора, такие как (S)-10,11-дигидро-3-[3-(пиридин-2-иламино)-1-пропилокси]-5Н-дибензо[а, d]циклогептен-10-уксусная кислота (известная как SB-265123) исследовалась в разных модельных системах на млекопитающих. (Keenan и др., 1998, Bioorg. Med. Chem. Lett. 8(22), 3171; Ward и др., 1999, Drug Metab. Dispos. 27(11), 1232). Исследования для идентификации антагонистов интегрина, подходящих для применения в качестве антагониста, описаны, например Smith и др., 1990, J.Biol. Chem. 265, 12267, и в цитируемой патентной литературе. Антитела к рецептору интегрина также хорошо известны. Подходящие моноклональные антитела к интегрину (например, αvβ3, αvβ5, αvβ6) могут быть модифицированы для усовершенствования их антигенсвязывающего фрагмента, включая F(ab)2, Fab, и конструированный Fv или одноцепочечное антитело. Подходящее и предпочтительно применяемое моноклональное антитело к рецептору интегрина αvβ3 было идентифицировано как LM609 (Brooks и др., 1994, Cell 79, 1157; АТСС НВ 9537). Эффективное специфическое антитело к αvβ5, P1F6, описано в WO 97/45447, и оно также является предпочтительным в соответствии с данным изобретением. Также приемлемым селективным αvβ6 антителом является MAb 14D9.F8 (WO 99/37683, DSM ACC2331, Merck KGaA, Германия), а также MAb 17.E6 (ЕР 0719859, DSM ACC2160, Merck KGaA), которые селективны к αv-цепи рецепторов интегрина. Другим подходящим антителом к интегрину является коммерчески доступный Vitraxin®.
"Ангиогенный фактор роста или рецептор фактора роста" представляет собой фактор или рецептор, который при его активации способствует росту и развитию кровеносных сосудов. Типично к этой группе относятся фактор роста эндотелия сосудов (VEGF) и его рецептор.
Понятие "антитело" или "иммуноглобулин" здесь применяется в широком смысле и, в частности, относится к интактным моноклональным антителам, поликлональным антителам, многоспецифическим антителам (например, биспецифическим антителам), которые образуются по крайней мере из двух интактных антител, и к фрагментам антитела, при условии, что они проявляют необходимую биологическую активность. Как правило, понятие включает гетероантитела, которые состоят из двух или более антител или их фрагментов с разной связывающей специфичностью, соединенных вместе.
В зависимости от аминокислотной последовательности их постоянных участков, интактные антитела могут быть отнесены к разным "классам антител (иммуноглобулинов)". Существует пять основных классов интактных антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них в дальнейшем могут быть разделены на "подклассы" (изотипы), например, IgG1, lgG2, IgG3, lgG4, IgA и lgA2. Постоянные домены тяжелой цепи, которые соответствуют разным классам антител, называются α, δ, ε, γ и μ соответственно. Предпочтительно основной класс антител в соответствии с изобретением представляет собой IgG, более подробно IgG1 и lgG2.
Антитела обычно представляют собой гликопротеины, которые имеют молекулярный вес приблизительно 150,000 и состоят из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как число дисульфидных связей отличается в тяжелых цепях иммуноглобулинов разных изотипов. Каждая тяжелая и легкая цепь также регулярно соединены в пространстве междуцепочечными дисульфидными мостиками. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым расположено несколько постоянных доменов. Каждая легкая цепь имеет вариабельный домен (VL) на одном конце и постоянный домен на ее другом конце. Постоянный домен легкой цепи совмещается с первым постоянным доменом тяжелой цепи, и вариабельный домен легкой цепи совмещается с вариабельным доменом тяжелой цепи. Полагают, что специфические аминокислотные остатки образуют поверхность между вариабельными доменами легкой и тяжелой цепей. "Легкие цепи" антител из многих видов позвоночных могут быть отнесены к одному из двух явно отличных типов, которые называются каппа (κ) и лямбда (λ), на основании аминокислотных последовательностей их постоянных доменов.
Понятие "моноклональное антитело", как здесь применяется, относится к антителу, полученному из популяции по существу однородных антител, например, индивидуальные антитела, составляющие популяцию, являются идентичными, за исключением возможных мутаций, встречающихся в естественных условиях, которые могут присутствовать в единичном количестве. Моноклональные антитела являются высокоспецифичными к отдельному антигенному участку. Кроме того, в отличие от препаратов поликлональных антител, которые включают различные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело связывается с отдельной детерминатной в антигене. В связи с их специфичностью моноклональные антитела обладают преимуществами, так как могут быть синтезированы без загрязнения другими антителами. Способы получения моноклональных антител, включая гибридомный метод, описаны Kohler and Milstein (1975, Nature 256, 495) и в "Технологии моноклональных антител, получение и характеристика гибридом грызунов и людей" (1985, под ред. Burdon и др., Лабораторные технологии в биохимии и молекулярной биологии, том 13, научное издательство Элсевиер, Амстердам), или могут быть получены при помощи хорошо известных методов рекомбинантной ДНК (см., например, патент US 4,816,567). Моноклональные антитела также могут быть изолированы из библиотек фаговых антител, применяя, например, методику, описанную в Clackson и др., Nature, 352:624-628 (1991) и Marks и др., J. Mol. Biol., 222:58, 1-597 (1991).
Под понятием "химерное антитело" подразумеваются антитела, в которых часть тяжелой и/или легкой цепи является идентичной или гомологичной к соответствующим последовательностям антитела, извлеченным из отдельного вида или принадлежащим отдельному антителу класса или подкласса, в то время как остаток цепи(ей) является идентичным или гомологичным к соответствующим последовательностям антитела, извлеченным из другого вида или принадлежащим антителу другого класса или подкласса, а также фрагменты таких антител, при условии, что они выявляют необходимую биологическую активность (например: патент US 4,816,567; Morrison и др., Proc. Nat. Acad. Sci. USA, 81:6851-6855 (1984)). Способы получения химерных и гуманизированных антител также известны в данной области техники. Например, такие способы получения химерных антител описаны в патентах на имя Boss (Celltech) и Cabilly (Genentech) (патенты US 4,816,397; US 4,816,567).
"Гуманизированные антитела" представляют собой разновидность нечеловеческих (например, грызунов) химерных антител, которые включают минимальную последовательность, извлеченную из нечеловеческого иммуноглобулина. Главным образом, гуманизированные антитела представляют собой человеческие иммуноглобулины (реципиентное антитело), в котором остатки гипервариабельного участка (CDRs) реципиента заменены остатками гипервариабельного участка нечеловеческого вида (донорное антитело), такого как мышь, крыса, кролик или нечеловеческая обезьяна, которые имеет необходимую специфичность, аффинность и функциональную активность. В некоторых случаях, каркасный участок (FR) остатков человеческого иммуноглобулина заменяется соответствующими остатками нечеловеческого происхождения. Кроме того, гуманизированные антитела могут включать остатки, которые не обнаруживаются в реципиентном антителе или в донорном антителе. Эти модификации осуществляются для облегчения дальнейшей очистки антитела. В основном, гуманизированное антитело может содержать по существу все, по крайней мере один, и типично два вариабельных домена, в которых все или по существу все гипервариабельные петли соответствуют таковым нечеловеческого иммуноглобулина и все или по существу все FRs являются таковыми последовательности человеческого иммуноглобулина. Гуманизированное антитело необязательно также может содержать по крайней мере часть постоянного участка (Fc) иммуноглобулина, типично эта часть является частью иммуноглобулина человека. Способы получения гуманизированных антител описаны, например. Winter (US 5,225,539) и Boss (Celltech, US 4,816,397).
Понятие "вариабельный" или "FR" относится к тому обстоятельству, что определенные части вариабельных доменов в значительной степени отличаются по своей последовательности в антителах и используются для связывания и специфичности каждого индивидуального антитела с его индивидуальным антигеном. Однако вариабельность распределена неравномерно на всем протяжении вариабельных доменов антител. Она сконцентрирована в трех сегментах, которые называются гипервариабельными участками вариабельных доменов как в легкой цепи, так и в тяжелой цепи. Наиболее высококонсервативные участки вариабельных доменов называются каркасными участками (FRs). Вариабельные домены естественных тяжелых и легких цепей каждый включает четыре FRs (FR1-FR4), которые в значительной степени имеют β-складчатую конфигурацию, соединенные с тремя гипервариабельными участками, которые образуют связующие петли, и в некоторых случаях частично образуют β-складчатую структуру. Гипервариабельные участки в каждой цепи удерживаются вместе непосредственно близко при помощи FRs и с гипервариабельными участками другой цепи способствуют образованию антигенсвязывающего участка антител (см. Kabat и др., Sequences of Proteins of Immunological Interest, 5-е изд., Public Health Service, National Institutes of Health, Батезда, MD. (1991)). Постоянные домены непосредственно не вовлечены в связывание антитела с антигеном, но выполняют разные эффекторные функции, такие как участие антитела в антителозависимой клеточной токсичности (ADCC).
Понятие "гипервариабельный участок" или "CDR" при использовании в изобретении, относится к аминокислотным остатками антитела, которые ответственны за связывание антигена. Гипервариабельный участок, как правило, включает аминокислотные остатки с "комплементарного детерминирующего участка" или "CDR" (например, остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в легкой цепи вариабельного домена и 31-35 (H1), 50-65 (Н2) и 95-102 (Н3) в тяжелой цепи вариабельного домена; и/или такие остатки с "гипервариабельной петли" (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в легкой цепи вариабельного домена и 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) в тяжелой цепи вариабельного домена; Chothia и Lesk J. Mol. Biol. 196:901-917 (1987)).
"Каркасный участок" или "FR" остатки представляют собой такие остатки вариабельного домена, которые являются отличными от остатков гипервариабельного участка, как определяется в настоящем изобретении.
"Фрагменты антитела" включают часть интактного антитела, предпочтительно содержащую его антигенсвязывающий или вариабельный участок. Примеры фрагментов антитела включают Fab, Fab′, F(ab′)2, Fv и Fc фрагменты, дитела, линейные антитела, одноцепочечные молекулы антител; и многоспецифичные антитела, образованные с фрагмента(ов) антител. "Интактное" антитело представляет собой антитело, содержащее антигенсвязывающий вариабельный участок, а также постоянный домен (CL) легкой цепи и постоянные домены тяжелой цепи СН1, СН2 и СH3. Предпочтительно, инактное антитело выполняет одну или несколько эффекторных функций.
При расщеплении антитела папаином образуются два идентичных антигенсвязывающих фрагмента, которые называются "Fab" фрагментами, каждый из них включает один антигенсвязывающий участок и CL и СН1 участок, и остаток "Fc" фрагмента, чье название отображает его способность к быстрой кристаллизации.
"Fc" участок антител, как правило, включает СН2, СН3 и петлевой участок IgG1 или lgG2 основного класса антител. Петлевой участок представляет собой группу с приблизительно 15 аминокислотных остатков, которая соединяет СН1 участок с СН2-СН3 участками.
При обработке пепсином образуется "F(ab′)2" фрагмент, который имеет два антигенсвязывающих участка и сохраняет способность к перекрестному связыванию антигена. "Fv" представляет собой минимальный фрагмент антитела, который содержит полный участок распознавания антигена и участок связывания антигена. Этот участок состоит из димера одной тяжелой цепи и одного вариабельного домена легкой цепи, которые плотно нековалентно присоединены. В этой конфигурации три гипервариабельных участка (CDRs) каждого вариабельного домена взаимодействуют с образованием антигенсвязывающего участка на поверхности VH-VL димера. Совместно, шесть гипервариабельных участков определяют антигенсвязывающую специфичность антитела. Тем не менее, даже один вариабельный домен (или половина Fv, которая включает только три гипервариабельных участка, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя с меньшим сродством по сравнению с целым ангигенсвязывающим участком. Fab фрагмент также включает постоянный домен легкой цепи и первый постоянный домен (СН1) тяжелой цепи. "Fab'" фрагменты отличаются от Fab фрагментов путем прибавления нескольких остатков к карбоксильному концу домена тяжелой цепи СН1, включая один или более цистеинов с петлевого участка антитела. F(ab′)2 фрагменты антитела необязательно образуют пары Fab(фрагментов при наличии цистеинов в петле между ними. Другие химические связывания фрагментов антитела также известны (см., например, Hermanson, Технология биоконъюгатов. Academic Press, 1996; US 4,342,566). "Одноцепочечные Fv" или "scFv" фрагменты антитела включают V, и V, домены антител, в которых эти домены присутствуют в виде единичной полипептидной цепи. Предпочтительно Fv полипептид также включает полипептидный линкер между VH и VL доменами, который дает возможность scFv приобретать необходимую для связывания антигена структуру. Одноцепочечные FV антитела известны, например, из Plückthun (The Pharmacology of Monoclonal Antibodies, том 113, под ред. Rosenburg и Moore, Springer-Verlag. Нью Йорк. стр.269-315 (1994)), WO 93/16185; US 5,571,894; US 5,587,458; Huston и др. (1988, Proc. Natl. Acad. Sci. 85, 5879) или Skerra и Plueckthun (1988, Science 240,1038).
Понятие "дитела" относится к небольшим фрагментам антитела с двумя антигенсвязывающими участками, эти фрагменты включают вариабельный тяжелый домен (V,) связанный с вариабельным легким доменом (V,) в идентичных полипептидных цепях (V, - V,). При помощи линкера, который является очень коротким для образования пар между двумя доменами идентичных цепей, домены образуют пары с комплементарными доменами другой цепи и образуют два антигенсвязывающих участка. Более подробно дитела описаны, например, в ЕР 404,097; WO 93/11161.
"Биспецифические антитела" представляют собой отдельные, дивалентные антитела (или их иммунотерапевтическиэффективные фрагменты), которые имеют два различных специфичных участка связывания антигена. Например, первый участок связывания антигена представляет собой участок к рецептору ангиогенеза (например, интегрину или VEGF рецептору), тогда как второй участок связывания антигена представляет собой участок к рецептору ErbB (например, EGFR или HER2). Биспецифические антитела могут быть получены при помощи химических методов (см., например, Kranz и др. (1981) Proc. Natl. Acad. Sci. USA 78,5807), при помощи "полидомной" технологии (см. патент US 4,474,893) или при помощи технологии рекомбинантной ДНК, которые все известны per se. Дополнительные методы описаны в WO 91/00360, WO 92/05793 и WO 96/04305. Биспецифические антитела также могут быть получены из одной цепи антител (см., например, Huston и др. (1988) Proc. Natl. Acad. Sci. 85, 5879; Skerra и Plueckthun (1988) Science 240, 1038). Аналоги вариабельных участков антитела могут быть получены в виде единичной полипептидной цепи. Для образования биспецифического связывающего агента отдельные цепи антител могут быть связаны вместе при помощи химических методов или методов генетической инженерии, известных в данной области техники. Также представляется возможным получать биспецифические антитела в соответствии с данным изобретением при помощи использования последовательностей лейциновой застежки. Применяемые последовательности являются производными участков лейциновой застежки факторов транскрипции Fos и Jun (Landschulz и др., 1988, Science 240, 1759; см. обзор Maniatis и Abel, 1989, Nature 341, 24). Лейциновые застежки представляют собой специфические аминокислотные последовательности длиной приблизительно 20-40 остатков, в которых лейцин обычно встречается в каждом седьмом остатке. Такая последовательность застежки образует алифатическую α-спираль с остатками лейцина, расположенными в гидрофобном участке димера. Соответствующие пептиды для лейциновых застежек протеинов Fos и Jun предпочтительно образуют гетеродимеры (O′Shea и др., 1989, Science 245, 646). Биспецифические антитела, которые содержат застежки, и способы их получения также описаны в WO 92/10209 и WO 93/11162. Биспецифическое антитело в соответствии с изобретением может представлять собой антитело к рецептору VEGF и рецептору αVβ3, как обсуждалось выше относительно антител, обладающих единичной специфичностью.
Понятие "иммуноконъюгат" относится к антителу или иммуноглобулину, соответственно, или его иммунологически эффективному фрагменту, которое соединено ковалентной связью с иммунологически неактивной молекулой. Предпочтительно эта присоединенная часть представляет собой пептид или протеин, который может быть гликозилирован. Указанная молекула, которая не представляет собой антитело, может присоединятся к С-концу постоянных тяжелых цепей антитела или к N-концам вариабельных легких и/или тяжелых цепей. Присоединенные части могут быть связаны посредством линкерной молекулы, которая представляет собой, как правило, полипептид, который включает 3-15 аминокислотных остатков. Иммуноконъюгаты в соответствии с изобретением предпочтительно включают слитые протеины, содержащие иммуноглобулин или его иммунотерапевтически эффективный фрагмент к ангиогенному рецептору, предпочтительно к интегрину или рецептору VEGF и TNFα или слитый протеин, содержащий по существу TNFα и IFNγ или другой приемлемый цитокин, который присоединен его N-концом к С-концу указанного иммуноглобулина, предпочтительно в его Fc части.
Понятие "слитый протеин" относится к естественной или синтетической молекуле, включающей один или более иммунотерапевтически неэффективных (не антител) белков или пептидов, обладающих различной специфичностью, которые связаны вместе необязательно при помощи линкерной молекулы. Слитый протеин в соответствии с изобретением может представлять собой молекулы, состоящие из, например, цикло-(Arg-Gly-Asp-DPhe-NMeVal), сконденсированного с TNFα и/или IFNγ.
"Гетероантитела" представляют собой два или более антител или связывающих фрагмента антител, которые связаны вместе, каждый из которых обладает разной связывающей специфичностью. Гетероантитела могут быть получены при помощи совместного конъюгирования двух или более антител или фрагментов антител. Предпочтительно гетероантитела включают перекрестно связанные Fab/Fab' фрагменты. Для конъюгирования антител могут применятся разные связующие или поперечно-сшивающие агенты. Примерами являются белок А, карбоимид, М-сукцинимидил-3-ацетил-тиоацетат (SATA) и N-сукцинимидил-3-(2-пиридилдитио) пропионат (SPDP) (см., например, Karpovsky и др. (1984) J.EXP. Med. 160, 1686; Liu и др. (1985) Proc. Natl. Acad. Sci. USA 82, 8648). Другие подобные методы описаны Paulus, Behring Inst. Mitt., №.78, 118 (1985); Brennan и др. (1985) Science 30 m:81 или Glennie и др. (1987) J. Immunol. 139, 2367. Другим способом является применение α-фенилендималеимида (αPDM) для связывания трех Fab' фрагментов (WO 91/03493). Многоспецифичные антитела в контексте данного изобретения также являются приемлемыми и могут быть получены, например, в соответствии с исследованием WO 94/13804 и WO 98/50431.
"Эффекторные функции" антитела относятся к таким биологическим активностям, которые определяются Fc участком (естественной последовательностью Fc участка или аминокислотной последовательностью различных Fc участков) антитела. Примеры эффекторных функций антитела включают цитотоксичность, зависимую от комплемента, связывание Fc с рецептором, антителозависимую цитотоксичность, опосредованную клетками (ADCC), фагоцитоз; ингибирующую регуляцию рецепторов поверхности клеток (например, рецептора β-клеток), и т.д.
Понятие "ADCC" (антителозависимая цитотоксичность, опосредованная клетками) относится к опосредованной клетками реакции, в которой неспецифические цитотоксические клетки, которые экспрессируют рецепторы Fc (FcR) (например, естественные киллерные (NK) клетки, нейтрофилы и макрофаги), распознают связанное антитело на клетке-мишени и впоследствии вызывают лизис клетки-мишени. Первичные клетки, опосредующие ADCC, NK клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Для оценки ADCC активности молекул, которые представляют интерес, могут применятся исследования ADCC in vitro, такие как описанные в уровне техники (патенты US 5,500,362; US 5,821,337). Подходящие эффекторные клетки для таких исследований включают мононуклеарные клетки периферической крови (РВМС) и естественные киллерные (NK) клетки.
"Эффекторные клетки человека" представляют собой лейкоциты, которые экспрессируют один или несколько FcRs и выполняют эффекторные функции. Предпочтительно, клетки экспрессируют по крайней мере FcγRIII и выполняют ADCC эффекторные функции. Примерами лейкоцитов человека, которые опосредуют ADCC, являются мононуклеарные клетки периферической крови (РВМС), естественные киллерные (NK) клетки, моноциты, цитотоксические Т-клетки и нейтрофилы.
Понятия "Fc рецептор" или "FcR" применяются для описания рецептора, который связывается с Fc участком антитела. Предпочтительные FcR представляют собой естественную последовательность человеческого FcR. Кроме того, предпочтительный FcR представляет собой такой, который связывает IgG антитело (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII, и FcγRIII, включая аллельные варианты и альтернативно усиленные формы этих рецепторов. Обзоры относительно FcRs приведены, например, в Ravetch и Kinet, Annu. Rev. Immunol 9:457-92 (1991).
Понятие "цитокин" представляет собой общий термин белков, которые высвобождаются одной популяцией клеток и воздействуют на другую клетку в качестве межклеточных медиаторов. Примерами таких цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. Цитокинами также являются гормоны роста, такие как человеческий гормон роста, N-метионильный человеческий гормон роста и гормон роста крупного рогатого скота; паратиреоидный гормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (FSH), тиреотропный гормон (TSH) и лютеинизирующий гормон (LH); фактор роста гепатоцитов; фактор роста фибробластов; пролактин; лактоген плаценты; мышиный гонадотропин-ассоциированный пептид; ингибин; активин; фактор роста эндотелия сосудов (VEGF); интегрин; тромбопоэтин (ТРО); факторы роста нервов, такие как NGFβ; фактор роста тромбоцитов; трансформирующие факторы роста (TGFs), такие как TGFα и TGFβ; эритропоэтин (ЕРО); интерфероны, такие как IFNα, IFNβ и IFNγ; колониестимулирующие факторы, такие как M-CSP, GM-CSF и G-CSF; интерлейкины, такие как IL-1, IL-1a, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12; и TNFα или TNFβ. Предпочтительные цитокины в соответствии с изобретением представляют собой интерфероны и TNFα.
Понятие "цитотоксическое средство", как применятся здесь, относится к веществу, которое ингибирует или предотвращает функцию клеток и/или вызывает разрушение клеток. Понятие также включает радиоактивные изотопы, химиотерапевтические средства и токсины, такие как ферментативно активные токсины бактерий, грибов, растительных или животных организмов или их фрагменты. Понятие также может включать представителей семейства цитокинов, предпочтительно IFNγ.
Понятие "химиотерапевтическое средство" или "противоопухолевое средство" включает химические средства, которые обладают противоопухолевыми действиями, например, предотвращают развитие, созревание или распространение опухолевых клеток, непосредственно воздействуя на клетки опухоли, например, путем цитостатических или цитотоксических действий, и непрямо воздействуя при помощи механизмов, таких как модификация биологического ответа. Приемлемые химиотерапевтические средства в соответствии с изобретением предпочтительно представляют собой естественные или синтетические химические соединения, но также и биологические молекулы, такие как белки, полипептиды, и т.п. Существует много противоопухолевых средств, которые являются коммерчески доступными, исследованных в клинических условиях и в доклинических условиях, которые могут быть включены в данное изобретение для лечения опухолей/новообразований при комбинированной терапии с TNFα и антиангиогенными средствами, как указывается выше, необязательно с другими средствами, такими как антагонисты EGF рецептора. Следует обратить внимание на то, что химиотерапевтические средства могут вводиться необязательно совместно с вышеуказанной комбинацией лекарственных средств. Примерами химиотерапевтических средств являются алкилирующие агенты, например азотистые иприты, этилениминовые соединения, алкилсульфонаты и другие соединения с алкилирующим действием, такие как нитрозомочевины, цисплатин и дакарбазин; антиметаболиты, например, фолиевая кислота, антагонисты пурина или пиримидина; ингибиторы митоза, например алкалоиды барвинка и производные подофиллотоксина; цитотоксические антибиотики и производные камптотецина. Предпочтительные химиотерапевтические средства или химиотерапия включают амифостин (этиол), цисплатин, дакарбазин (DTIC), дактиномицин, мехлорэтамин HCl (азотистый иприт), стрептозоцин, циклофосфамид, каррнустин (BCNU), ломустин (CCNU), доксорубицин (адриамицин), липодоксорубицин (доксил), гемцитабин (гемзар), даунорубицин, липодаунорубицин (дауноксом), прокарбазин, митомицин, цитарабин, этопозид, метотрексат, 5-фторурацил (5-FU), винбластин, винкристин, блеомицин, паклитаксел (таксол), доцетаксел (таксотер), альдеслейкин, аспарагиназа, бисульфан, карбоплатин, кладрибин, камптотецин, СРТ-11,10-гидрокси-7-этил-камптотецин (SN38), дакарбазин, флоксуридин, флударабин, гидросимочевина, ифосфамид, идарубицин, месна, альфа-интерферон, бета-интерферон, иринотекан, митоксантрон, топотекан, лейпролид, мегестрол, мельфалан, меркапропурин, пликамицин, митотан, пегаспаргас, пентостатин, пипоброман, пликамицин, стрептозоцин, тамоксифен, тенипозид, тестолактон, тиогуанин, тиотепа, 5-[бис-(2-хлорэтил)амино]урацил, винорелбин, хлорамбуцил и их сочетания.
Наиболее предпочтительными химиотерапевтическими средствами в соответствии с изобретением являются цисплатин, гемцитабин, доксорубицин, паклитаксел (таксол) и блеомицин.
Понятия "рак" и "опухоль" относятся или описывают физиологические состояния у млекопитающих, которые обычно характеризуются нерегулируемым клеточным ростом. При помощи фармацевтических композиций в соответствии с настоящим изобретением можно предотвращать развитие опухолей, при этом могут лечиться опухоли молочной железы, сердца, легких, тонкой кишки, толстой кишки, селезенки, почек, мочевого пузыря, головы и шеи, яичников, предстательной железы, головного мозга, поджелудочной железы, кожи, костей, костного мозга, крови, тимуса, матки, яичек, шейки матки и печени. Более специфично опухоли выбирают из группы, включающей аденому, ангиосаркому, астроцитому, эпителиальную карциному, герминому, глиобластому, глиому, гамартому, гемангиоэндотелиому, гемангиосаркому, гематому, гепатобластому, лейкоз, лимфому, медуллобластому, меланому, нейробластому, остеосаркому, ретинобластому, рабдомиосаркому, саркому и тератому. Более подробно, опухоль выбирают из группы, включающей акральную лентигинозную меланому, старческий кератоз, аденокарциному, аденокистозную карциному, аденомы, аденосаркому, аденосквамозную карциному, опухоли астроцитов, карциному бартолиновых желез, базально-клеточный рак, альвеоляно-клеточный рак, капиллярную, карциноидную опухоль, карциному, карциносаркому, кавернозную, холангио-карциному, хондросаркому, папиллому/саркому хориоидного сплетения, почечно-клеточный рак, цистаденому, опухоль ентодермальной полости, гиперплазию эндометрия, эндометриальную саркому, аденокарциному эндометрия, эпендимальную, эпителиоидную, саркому Эвинга, фибропластинчатую, фокальную узелковую гиперплазию, ульцерогенную аденому поджелудочной железы, опухоли половых клеток, глиобластому, глюкагоному, гемангибластому, гемангиоэндотелиому, гемангиомы, аденому печени, аденоматоз печени, печеночно-клеточный рак, инсулиному, интаэпителиальную неоплазию, преинваз ионную сквамозно-клеточную неоплазию, инвазивную сквамозно-клеточную карциному, крупноклеточную карциному, лейомиосаркому, ограниченную злокачественную меланому, злокачественную меланому, злокачественные мезотелиальные опухоли, медуллобластому, медуллоэпителиому, меланому, менингеальную, мезотелиальную, метастатическую карциному, слизеобразующий плоскоклеточный рак, нейробластому, нейроэпителиальную аденокарциному узелковую меланому, овсяновидно-клеточную карциному, олигодендроглиальную, остеосаркому, полипептид поджелудочной железы, папиллярную серозную аденокарциному, пинеалоцитные, гипофизарные опухоли, плазмацитому, ложную саркому, бластому легких, рак почки, ретинобластому, рабдомиосаркому, саркому, серозную карциному, мелкоклеточный рак, мягкотканевые карциномы, опухоль, секретирующую соматостатин, сквамозную карциному, плоскоклеточный рак, субмезотелиальную, поверхностную распространяющуюся меланому, недифференцированную карциному, увеальную меланому, сосочковую карциному, випому, хорошо дифференцированную карциному и аденосаркому почки (опухоль Вильмса).
"ErbB рецептор" представляет собой рецептор тирозинкиназы белков, который принадлежит к семейству рецепторов ErbB и включает рецепторы EGFR(ErbB1), ErbB2, ErbB3 и ErbB4 и другие представители этого семейства, которые будут идентифицированы в будущем. Рецептор ErbB в большинстве случаев может включать внеклеточный домен, который может связывать лиганд ErbB; липофильный трансмембранный домен; консервативный внутриклеточный домен тирозинкиназы; и карбоксиконцевой сигнальный домен, в котором находится несколько остатков тирозина, которые могут быть фосфорилированы. Рецептор ErbB может представлять собой "естественную последовательность" ErbB рецептора или "вариант его аминокислотной последовательности". Предпочтительно рецептор ErbB представляет собой естественную последовательность рецептора ErbB человека. ErbB1 относится к генам, кодирующим белковый продукт EGFR. Более предпочтительным является EGF рецептор (HER1). Выражения "ErbB1" и "HER1" взаимозаменяемо используются в настоящем изобретении и относятся к белку HER1 человека. Выражения "ErbB2" и "HER2" взаимозаменяемо используются в настоящем изобретении и относятся к белку HER2 человека. Предпочтительными в соответствии с данным изобретением являются ErbB1 рецепторы (EGFR).
"Лиганд ErbB" представляет собой полипептид, который связывается с и/или активирует ErbB рецептор. ErbB лиганды, которые связывают EGFR, включают EGF, TGF-α, амфирегулин, бетацеллюлин, HB-EGF и эпирегулин.
Понятие "антагонист/ингибитор рецептора ErbB" относится к естественной или синтетической молекуле, которая связывает и блокирует или ингибирует ErbB рецептор. Таким образом, при помощи блокирования рецептора антагонист препятствует связыванию ErbB лиганда (агониста) и активированию комплекса агониста/лиганда с рецептором. ErbB антагонисты могут быть к HER1 (EGFR) или HER2. Предпочтительными антагонистами по изобретению являются антагонисты к EGF рецептору (EGFR, HER1). Антагонист ErbB рецептора может представлять собой антитело или его иммунотерапевтически эффективный фрагмент или неиммунобиологические молекулы, такие как пептид, полипептидный белок. Также сюда входят химические молекулы, тем не менее, предпочтительными антагонистами в соответствии с изобретением являются антитела к EGFR и антитела к HER2. Предпочтительными антителами по изобретению являются антитела к Неr1 и антитела к Неr2, более предпочтительно антитела к Herl. Предпочтительные антитела к Her1 представляют собой MAb 425, предпочтительно гуманизированное MAb 425 (hMAb 425, US 5,558,864; ЕР 0531472) и химерное MAb 225 (cMAb 225, US 4,943,533 и ЕР 0359 282). Наиболее предпочтительным является моноклональное антитело h425, которое обладает высокой эффективностью при лекарственной монотерапии в сочетании с уменьшенными нежелательными и побочными эффектами. Наиболее предпочтительным антителом к HER2 является HERCEPTIN®, выпускаемый Genentech/Roche. Эффективные антагонисты EGF рецептора в соответствии с изобретением также могут включать естественные или синтетические химические соединения. В качестве примеров предпочтительных молекул этой категории можно привести органические соединения, металлоорганические соединения, соли органических и металлорганических соединений.
Примерами антагонистов HER2 рецептора являются: гетероарильные соединения, замещенные стирилом (US 5,656,655); бис- моно, и/или бициклические арильные гетероарильные, карбоциклические и гетерокарбоциклические соединения (US 5,646,153); трициклические пиримидиновые соединения (US 5,679,683); производные хиназолина, обладающие ингибиторной активностью по отношению к рецептору тирозинкиназы (US 5,616,582); гетероарилэтендиильные или гетероарилэтендииларильные соединения (US 5,196,446); соединение, обозначенное как 6-(2,6-дихлорфенил)-2-(4-(2-диэтил-аминоэтокси) фениламино)-8-метил-8Н-пиридо(2,3)-5-пиримидин-7-он (Panek, и др., 1997, J.Pharmacol. Exp. Therap. 283,1433) ингибирующие семейства рецепторов EGFR, PDGFR и FGFR.
"Лучевая терапия": Опухоли, которые могут поддаваться лечению фармацевтическими композициями в соответствии с изобретением, могут дополнительно подвергаться лечению при помощи облучения или радиоактивных лекарственных средств. Источник радиации может быть наружным или внутренним для пациента, который поддается лечению. Если источник является наружным для пациента, то это такое лечение известно как наружная лучевая терапия (EBRT). Если источник радиации является внутренним для пациента, то такое лечение называется брахитерапия (ВТ). Некоторые характерные радиоактивные атомы, которые могут применятся, включают радий, цезий-137, и иридий-192, америций-241 и золото-198, кобальт-57; медь-67; технеций-99; йодид-123; йодид-131; и индий-111. Также можно метить средства в соответствии с изобретением радиоактивными изотопами. Сегодня лучевая терапия представляет собой стандартное лечение для контроля неудалимых или неоперабельных опухолей и/или метастаз опухолей. Улучшенные результаты наблюдаются при сочетании лучевой терапии с химиотерапией. Лучевая терапия основывается на принципе, что радиация в высоких дозах в зонах облучения вызывает гибель репродуктивных клеток как опухолевых, так и нормальных тканей. Полученная доза радиации в основном определяется на основе поглощенной дозы (рад), времени и фракционирования, и может быть определена онкологом. Количество ионизирующей радиации, полученной пациентом, зависит от разнообразных соображений, однако двумя наиболее важными соображениями являются локализация опухоли по отношению к другим критическим структурам или органам тела, и протяженность распространения опухоли. Предпочтительным курсом лечения для пациента, который подвергается лучевой терапии, является лечение на протяжении периода 5-6 недель в тотальной дозе 50-60 Гр, которой облучается пациент в виде единичных дневных фракций по 1,8-2,0 Гр, 5 дней в неделю. Гр представляет собой сокращение от Грей и относится к дозе в 100 рад. В предпочтительном варианте осуществления изобретения предусматривается совместное воздействие, когда опухоли у человека подвергаются лечению антагонистом ангиогенеза и TNFα/IFNγ и облучением. Другими словами, ингибирование роста опухоли при воздействии указанными соединениями усиливается при комбинации с действием радиации и/или химиотерапевтическими средствами. Лучевая терапия необязательно может применятся в соответствии с изобретением. Это рекомендуется и является предпочтительным в случаях, в которых пациенту не может быть введено достаточного количества средств в соответствии с изобретением.
"Фармацевтическое лечение": Способ по изобретению включает разнообразные воздействия практического воплощения изобретения поэтапно. Например, средства в соответствии с изобретением могут вводиться одновременно, последовательно или отдельно. Кроме того, средства могут вводиться раздельно в течении интервала времени приблизительно 3 недели между введениями, например, от в основном введения непосредственно после введения первого средства до приблизительно 3 недель после введения первого средства. Способ можно осуществлять после хирургического вмешательства. Альтернативно хирургическое вмешательство можно осуществлять на протяжении интервала между введением первого активного средства и второго активного средства. Примером этого метода является комбинация настоящего способа с хирургическим удалением опухоли. Лечение в соответствии со способом обычно включает введение терапевтических композиций путем одного или нескольких циклов введения. Например, если осуществляется одновременное введение, терапевтическая композиция, которая включает оба средства, вводится на протяжении периода от приблизительно 2 дней до приблизительно 3 недель за один цикл. После этого цикл лечения может повторятся, если в этом есть необходимость, учитывая состояние пациента. Подобным образом, если осуществляется последовательное введение, время введения каждого индивидуального терапевтического вещества устанавливается на основании идентичного периода времени. Интервал между циклами может изменятся от 0 до 2 месяцев.
Средства по настоящему изобретению могут вводиться парентерально путем инъекции или постепенной инфузии на протяжении периода времени. Несмотря на то что ткани, подвергающейся лечению, обычно можно достичь при системном для организма введении, и более того, наиболее часто при лечении используется внутривенное введение терапевтических препаратов, другие ткани и доставка также рассматриваются, при вероятности того, что ткани-мишени содержат целевые молекулы. Таким образом, средства по настоящему изобретению могут вводиться внутриглазно, внутривенно, внутрибрюшинно, внутримышечно, подкожно, внутриполостно, черезкожно, путем ортотопической инъекции или инфузии, и также могут доставляться при помощи перистальтики. Терапевтические композиции содержат, например, антагонист интегрина по данному изобретению и обычно вводятся внутривенно, например, в виде инъекции единичной дозы. Терапевтические композиции по настоящему изобретению содержат физиологически толерантный носитель совместно с соответствующим средством, как описывается здесь, растворенным или диспергированым в нем в качестве активного компонента. Как применяется в данном изобретении, понятие "фармацевтически приемлемый" относится к композициям, носителям, растворителям и реактивам, которые представляют вещества, допускающие их введение млекопитающему без проявления нежелательных физиологических действий, таких как тошнота, головокружение, желудочные расстройства и подобные. Приготовление фармацевтической композиции, которая содержит активные компоненты в растворенном или диспергированом виде, понятно в данной области техники и не должно ограничиваться на основании состава. Обычно такие композиции приготавливают в виде инъецируемых, жидких растворов или суспензий, однако твердые формы, подходящие для растворов или суспензий, могут также быть приготовлены в виде раствора до использования. Составы также могут быть эмульгированы. Активный компонент может быть смешан с наполнителями, которые являются фармацевтически приемлемыми и совместимые с активным компонентом и в количествах, подходящих для применения в терапевтических методах, описанных здесь. Подходящие наполнители представляют собой, например, воду, физиологический раствор, декстрозу, глицерин, этанол и подобные и их сочетания. Кроме того, при необходимости, композиция может включать незначительные количества вспомогательных веществ, таких как смачивающие или эмульгирующие вещества, буферные вещества и подобные, которые усиливают эффективность активного компонента. Терапевтические композиции согласно настоящему изобретению могут включать фармацевтически приемлемые соли их компонентов. Фармацевтически приемлемые соли включают кислото-аддитивные соли (образованные свободной аминогруппой полипептида), которые образуются с неорганическими кислотами, такими как, например, соляная или фосфорная кислота, или такими органическими кислотами, как уксусная, винная, миндальная и подобные. Соли, образованные свободной карбоксильной группой, также могут являться производными неорганических оснований, таких как, например, гидроокиси натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, 2-этиламиноэтанол, гистидин, прокаин и т.п. Более предпочтительными являются соли с HCl, если в составе применяются циклические полипептидные антагонисты αv. Физиологически толерантные носители являются хорошо известными в данной области техники. Примерами жидких носителей являются стерильные водные растворы, которые не содержат других веществ, кроме активных компонентов и воды, или содержат буфер, такой как натрий-фосфатный при физиологическом значении рН, физиологический раствор или оба, такой как фосфатный буфер и физиологический раствор. Кроме того, дополнительно, водные носители могут включать более чем одну буферную соль, а также соли, такие как хлориды натрия и калия, декстрозу, полиэтиленгликоль и другие растворенные вещества. Жидкие композиции также могут включать жидкие фазы в дополнение к и исключая воду. Примерами таких дополнительных жидких фаз являются глицерин, растительные масла, такие как хлопковое масло, и водно-масляные эмульсии.
Обычно терапевтически эффективное количество иммунотерапевтического средства, например, в виде антитела, блокирующего рецептор интегрина или фрагмента антитела или конъюгата антитела или антитела, блокирующего VEGF рецептор, фрагмента или конъюгата, представляет собой такое количество, которого при введении в физиологически толератной композиции достаточно для достижения концентрации в плазме от приблизительно 0,01 микрограмма (мкг) на миллилитр (мл) до приблизительно 100 мкг/мл, предпочтительно от приблизительно 1 мкг/мл до приблизительно 5 мкг/мл и обычно приблизительно 5 мкг/мл. Дозировка может быть назначена по-другому, от приблизительно 0,1 мг/кг до приблизительно 300 мг/кг, предпочтительно от приблизительно 0,2 мг/кг до приблизительно 200 мг/кг, более предпочтительно от приблизительно 0,5 мг/кг до приблизительно 20 мг/кг, в виде одной или нескольких ежедневных доз на протяжении одного или нескольких дней. Если иммунотерапевтическое средство представлено в виде фрагмента моноклонального антитела или конъюгата, то количество может без труда устанавливаться на основании веса фрагмента/конъюгата, которое соответствует весу целого антитела. Предпочтительная молярная концентрация в плазме составляет от приблизительно 2 микромолей (мкМ) до приблизительно 5 миллимолей (мМ) и предпочтительно приблизительно от 100 мкМ до 1 мМ антагониста антитела. Терапевтически эффективное количество средства в соответствии с настоящим изобретением, которое представляет собой неиммуный терапевтический пептид или протеинный полипептид (например, TNFα, IFNγ) или другую биологическую молекулу с подобными размерами, обычно представляет собой такое количество полипептида, которого при введении в физиологически толератной композиции достаточно для достижения концентрации в плазме от приблизительно 0,1 микрограмма (мкг) на миллилитр (мл) до приблизительно 200 мкг/мл, предпочтительно от приблизительно 1 мкг/мл до приблизительно 150 мкг/мл. На основании полипептида, имеющего вес приблизительно 500 грамм на моль, предпочтительная молярная концентрация в плазме составляет от приблизительно 2 микромолей (мкМ) до приблизительно 5 миллимолей (мМ) и предпочтительно приблизительно 100 мкМ - 1 мМ антагониста полипептида. Обычная дозировка активного средства, которое предпочтительно представляет собой химический антагонист или (химическое) химиотерапевтическое средство в соответствии с изобретением (ни иммунотерапевтическое средство, ни неиммунотерапевтический пептид/протеин) представляет собой 10-1000 мг, предпочтительно приблизительно 20-200 мг, и более предпочтительно 50-100 мг на килограмм веса тела в день.
Фармацевтические композиции по изобретению могут включать лечение особи средствами, которые уменьшают или дают возможность избежать побочных действий, связанных с комбинированной терапией в соответствии с настоящим изобретением ("дополнительная терапия"), включая, но не ограничиваясь, такие средства, например, которые уменьшают токсическое действие противораковых лекарственных средств, например, ингибиторы резорбции кости, кардиопротекторные средства. Указанные дополнительные средства предотвращают или уменьшают степень тошноты и рвоты, обусловленных химиотерапией, лучевой терапией или операцией, или уменьшают процент инфекций, связанных с введением миелосупрессивных противораковых лекарственных средств. Дополнительные средства хорошо известны в данной области техники.
Иммунотерапевтические средства в соответствии с изобретением могут дополнительно вводиться со вспомогательными средствами, аналогичными BCG и стимуляторами иммунной системы. Кроме того, композиции могут включать иммунотерапевтические средства или химиотерапевтические средства, содержащие цитотоксически эффективные радиоактивно меченные изотопы, или другие цитотоксические средства, такие как цитотоксические пептиды (например цитокины) или цитотоксические лекарственные средства и подобные. Понятие "фармацевтический набор" для лечения опухолей или метастаз опухолей относится к упаковке, и, как правило, инструкциям для применения реагентов в способах лечения опухолей и метастаз опухолей. Реагент в наборе согласно данному изобретению обычно представленный терапевтической композицией, как указано в изобретении, и следовательно, может быть представлен разнообразными формами, приемлемыми для распределения в наборе. Такие формы могут включать жидкость, порошок, таблетки, суспензии и аналогичные составы для предусмотренных антагониста и/или слитого протеина согласно данному изобретению. Реагенты могут располагаться в отдельных контейнерах, подходящих для отдельного введения в соответствии с настоящими методами, или альтернативно могут располагаться в композиции в единичном контейнере в упаковке. Упаковка может включать достаточное количество для одной или нескольких дозировок регентов в соответствии с способами лечения, описанными здесь. Набор по данному изобретению также включает "инструкцию по применению" веществ, содержащихся в упаковке.
Описание фигур
Фигура 1. Образование HUVEC сфероидов и выживание не требует связывания интегрина. (а) Блокирование анти-VE-кадхерина (75) mAb или Са2+-уменьшение (EDTA, EDTA/Ca2+) ингибирует образование HUVEC сфероидов, в то время как блокирование mAbs против интегрина α1(Lia1/2), α5(SAM-1), αVβ3 (LM609) и РЕСАМ-1 (10D9) или RGD-пептида не обладает таким действием. (b) Жизнеспособность. HUVEC, извлекаемый из культур сфероидов (О) или фибронектина (•), обладает подобными жизнеспособными профилями.
Фигура 2. Интегринзависимая адгезия защищает HUVEC от TNF-индуцированного апаптоза. (a) YoPro-1 поглощение: воздействие TNF (Т) и TNF/IFNγ (T1) не вызывает YoPro-1 окрашивания в связанных с фибронектином HUVEC, в то время как вызывает сильное YoPro-1 окрашивание в HUVEC сфероидах, что подавляется ингибиторами каспазы BOG и ZVAD. TNF±IFNγ (T1). С, необработанные культуры. (b) Демонстрация активации каспазы-3 и высвобождения PARP (показано стрелкой) при помощи вестерн-блоттинга в обработанных TNE/IFNα (T1) сфероидах, но не в связанных с фибронектином HUVEC. С, необработанные культуры, (с, d) Кривые жизнеспособности HUVEC, подданных воздействию TNF  TNF/IFNγ
TNF/IFNγ  или контрольной сыворотки (О). (е) Жизнеспособность HUVEC, культивированных на иммобилизованных антителах (imAbs) к α1
или контрольной сыворотки (О). (е) Жизнеспособность HUVEC, культивированных на иммобилизованных антителах (imAbs) к α1  αvβ3
αvβ3 и α4 интегринам (О/I) при отсутствии (незакрашенные символы) или в присутствии (закрашенные символы) TNF/IFNγ.
и α4 интегринам (О/I) при отсутствии (незакрашенные символы) или в присутствии (закрашенные символы) TNF/IFNγ.
Фигура 3. Индуцированная TNF активация NF-κВ не требует связывания интегрина (а) Вестерн-блоттинг и (b) анализ изменения электрофоретической подвижности (EMSA) демонстрируют параллельную кинетику I-кВ фосфорилирования (Pi-κВ), I-κВ деградации (I-κВ) и NF-κВ ядерной транслокации (EMSA) в связанных с фибронектином HUVEC и сфероидах, подвергнутых воздействию TNF/IFNγ. (с) Проточный цитометрический анализ свидетельствует об идентичной индукции экспрессии ICAM-1 поверхностями клеток на фибронектине и в культурах сфероидов HUVEC, подданных воздействию TNF (----) или TNF/IFNγ―). (...) необработанные клетки.
Экспрессия РЕСАМ-1 показана в качестве контроля.
Фигура 4. Активация Akt является существенной для выживания HUVEC и требует связывания интегрина. (а) Выявление фосфорилированного (Pi-Akt) и общего Akt (Akt) в связанных с фибронектином и сфероидных HUVEC культурах стимулируется TNF/IFNγ на протяжении указанного времени. (b) Левая панель: ингибиторы PI-3 киназы вортманнин (W) и LY294002 (LY) повышают чувствительность связанных с фибронектином HUVEC к TNF (Т) и TNF/IFNγ(Т1)-индуцированному апоптозу. Мертвые клетки визуализировались путем окрашивания YoPro-1. Правая панель: кривые выживания связанных с фибронектином HUVEC, подвергнутых воздействию LY294002  TNF/IFNγ (Δ), или LY294002/TNF/IFNγ
TNF/IFNγ (Δ), или LY294002/TNF/IFNγ  . (О) необработанные культуры. (с) Значительная активность PI-3 киназы (р110*) и Akt (Aktmp), но не дикого типа Akt (Aktwt) или контрольной плазмиды (pBS), содействовала выживанию сфероидов, подданных воздействию TNF
. (О) необработанные культуры. (с) Значительная активность PI-3 киназы (р110*) и Akt (Aktmp), но не дикого типа Akt (Aktwt) или контрольной плазмиды (pBS), содействовала выживанию сфероидов, подданных воздействию TNF  или TNF/IFN (
или TNF/IFN ( ). (О) необработанные культуры. Вставки представляют собой проточный цитометрический анализ EGFP флюоресценции трансфектированных клеток (% положительных клеток), (d) HUVEC электропорированы с контрольной плазмидой (pBS) или конститутивно активными Akt (Akt*) и инфицированы AdΔNI-κB или AdLacZ, культивировались в качестве связанного с фибронектином монослоя или сфероидов в отсутствие (С) или в присутствии TNF (Т) или TNF/IFNγ (T1). Апоптотические клетки определяли путем окрашивания YoPro-1. Жизнеспособные связанные с фибронектином клетки окрашивались кристаллическим фиолетовым, (е) HUVEC электропорировали с контрольной плазмидой (незакрашенные символы) или pAktmp (закрашенные символы) и инфицировали AdANI-κВ
). (О) необработанные культуры. Вставки представляют собой проточный цитометрический анализ EGFP флюоресценции трансфектированных клеток (% положительных клеток), (d) HUVEC электропорированы с контрольной плазмидой (pBS) или конститутивно активными Akt (Akt*) и инфицированы AdΔNI-κB или AdLacZ, культивировались в качестве связанного с фибронектином монослоя или сфероидов в отсутствие (С) или в присутствии TNF (Т) или TNF/IFNγ (T1). Апоптотические клетки определяли путем окрашивания YoPro-1. Жизнеспособные связанные с фибронектином клетки окрашивались кристаллическим фиолетовым, (е) HUVEC электропорировали с контрольной плазмидой (незакрашенные символы) или pAktmp (закрашенные символы) и инфицировали AdANI-κВ  или AdLacZ
или AdLacZ  , и культивировали на фибронектине в присутствии разных концентраций TNF и живые связанные клетки определялись путем измерения оптической плотности (О.П.) окрашенных кристаллическим фиолетовым лунок. (f) Анализировали проточной цитометрией экспрессию ICAM-1 в необработанных HUVEC (...), или HUVEC, обработанных TNF (-----) и TNF/LY294002 (―) (левая панель), также как HUVEC, инфицированных AdΔNI-кВ (средняя панель) или AdLacZ и подвергнутых воздействию TNF (------) и TNF/IFNγ (―).
, и культивировали на фибронектине в присутствии разных концентраций TNF и живые связанные клетки определялись путем измерения оптической плотности (О.П.) окрашенных кристаллическим фиолетовым лунок. (f) Анализировали проточной цитометрией экспрессию ICAM-1 в необработанных HUVEC (...), или HUVEC, обработанных TNF (-----) и TNF/LY294002 (―) (левая панель), также как HUVEC, инфицированных AdΔNI-кВ (средняя панель) или AdLacZ и подвергнутых воздействию TNF (------) и TNF/IFNγ (―).
Фигура 5. (а-с) Анализ при помощи вестерн-блоттинга Pi-Akt, MDM2, р53, Pi-FKHR/FRKHL1 (а), и Pi-MEK, Pi-p38 и Pi-JNK и Pi-ERK в фибронектиновых и сфероидных HUVEC культурах, подвергнутых воздействию TNF/IFNγ на протяжении указанного времени. Сумма Akt, FKHR, MEK, p38, ERK и JNK белка свидетельствует об одинаковом количестве общего белка. Сфероидные культуры характеризуются недостаточностью фосфорилирования Akt и FKHR/FKRL1, повышенными уровнями р53 и усиленным фосфорилированием MEK, p38, ERK и JNK в ответ на действие TNF/IFNγ по сравнению со связанными с фибронектином клетками.
Фигура 6. Уменьшение связывания интегрина усиливает цитотоксичность TNF in vitro. (a) HUVEC культивировали на фибронектине или PLL на протяжении 16 часов при отсутствии (С) или в присутствии TNF (Т) или TNF/IFNγ (T1). Апоптоз и выживание адгезированных клеток определяли окрашиванием с YoPro-1 и кристаллическим фиолетовым, соответственно. (b) EMD121974 разрушали αVβ3-опосредованную адгезию HUVEC на желатине  , но не α5β1 компонент α5β1/αVβ3-опосредованной адгезии к фибронектину
, но не α5β1 компонент α5β1/αVβ3-опосредованной адгезии к фибронектину  . Контрольный пептид EMD135981 был неэффективным (незакрашенные символы). (с) HUVEC культивировали на фибронектине при отсутствии (С) или в присутствии TNF/IFNγ (T1), EMD121974 и EMD135981, как указано. Клетки, подвергнутые апоптозу, и адгезированные клетки определяли путем окрашивания с YoPro-1 и контрастной микроскопией соответственно. (d) Кривые выживания HUVBC эксперимента представленные на панеле с. Без пептида
. Контрольный пептид EMD135981 был неэффективным (незакрашенные символы). (с) HUVEC культивировали на фибронектине при отсутствии (С) или в присутствии TNF/IFNγ (T1), EMD121974 и EMD135981, как указано. Клетки, подвергнутые апоптозу, и адгезированные клетки определяли путем окрашивания с YoPro-1 и контрастной микроскопией соответственно. (d) Кривые выживания HUVBC эксперимента представленные на панеле с. Без пептида  ; EMD121974 ; EMD135981
; EMD121974 ; EMD135981  . Необработанные культуры - незакрашенные символы. Культуры, обработанные TNF/IFNγ, - закрашенные символы. (е) Кривые выживания HUVEC, электропорированных с Aktmp (незакрашенные символы) или pBS (закрашенные символы), и культивированных на фибронектине и обрабобтанных TNF/IFN γ без или в присутствии пептидов EMD121974 или EMD135981 . Aktmp предотвращает гибель клеток, вызванную комбинированным воздействием EMD121974 и TNF/IFN.
. Необработанные культуры - незакрашенные символы. Культуры, обработанные TNF/IFNγ, - закрашенные символы. (е) Кривые выживания HUVEC, электропорированных с Aktmp (незакрашенные символы) или pBS (закрашенные символы), и культивированных на фибронектине и обрабобтанных TNF/IFN γ без или в присутствии пептидов EMD121974 или EMD135981 . Aktmp предотвращает гибель клеток, вызванную комбинированным воздействием EMD121974 и TNF/IFN.
Фигура 7. Уменьшение связывания интегрина усиливает цитотоксичность TNF in vivo. BN крысы, которые имеют BN-175 изогенную саркому мягких тканей, обрабатывались EMD121974  , TNF
, TNF  или EMD121974/TNF при помощи техники ILP. (О) ложно обработанные крысы. Рост опухолей определяли на протяжении 6 дней после ILP. Результаты представлены в виде значений объема опухоли ± с.к.о. (n=6). Небольшие фрагменты изогенной саркомы мягких тканей BN-175 имплантировали в правую заднюю лапу самцов крыс BN, и обработку начинали после достижения опухоли размеров в диаметре 12-14 мм (Manusama и др., Oncol. Rep.6, 173-177. (1999)). Затем бедренную артерию и вену канюлизировали силастиковой трубкой и коллатерали перевязывали жгутом. Перфузию осуществляли на протяжении 30 мин с 5 мл Heamaccel® (2,4 мл/мин), при которых лекарственные средства вводили в виде болюсов (EMD121974, 500 мкг, конечная концентрация в перфузате 170 МКМ; TNF, 50 мкг). Перфузат насыщали кислородом и ногу выдерживали при 38-39°С в теплом матраце. Крысам, которым вводили EMD121974 при помощи перфузии, также системно вводили пептид 2 часа до и 3 часа после ILP (100 мг/кг внутрибрюшинно). Диаметр опухоли измеряли в двух направлениях при помощи циркуля и объем опухоли (V) рассчитывали по формуле (V=0,4)(А2ХВ, где В представляет собой наибольший диаметр и А представляет собой диаметр, перпендикулярный к В). На группу обрабатывали 6 крыс. Местные и общие побочные эффекты оценивали, как описано (Manusama и др., Oncol. Rep.6, 173-177. (1999)).
или EMD121974/TNF при помощи техники ILP. (О) ложно обработанные крысы. Рост опухолей определяли на протяжении 6 дней после ILP. Результаты представлены в виде значений объема опухоли ± с.к.о. (n=6). Небольшие фрагменты изогенной саркомы мягких тканей BN-175 имплантировали в правую заднюю лапу самцов крыс BN, и обработку начинали после достижения опухоли размеров в диаметре 12-14 мм (Manusama и др., Oncol. Rep.6, 173-177. (1999)). Затем бедренную артерию и вену канюлизировали силастиковой трубкой и коллатерали перевязывали жгутом. Перфузию осуществляли на протяжении 30 мин с 5 мл Heamaccel® (2,4 мл/мин), при которых лекарственные средства вводили в виде болюсов (EMD121974, 500 мкг, конечная концентрация в перфузате 170 МКМ; TNF, 50 мкг). Перфузат насыщали кислородом и ногу выдерживали при 38-39°С в теплом матраце. Крысам, которым вводили EMD121974 при помощи перфузии, также системно вводили пептид 2 часа до и 3 часа после ILP (100 мг/кг внутрибрюшинно). Диаметр опухоли измеряли в двух направлениях при помощи циркуля и объем опухоли (V) рассчитывали по формуле (V=0,4)(А2ХВ, где В представляет собой наибольший диаметр и А представляет собой диаметр, перпендикулярный к В). На группу обрабатывали 6 крыс. Местные и общие побочные эффекты оценивали, как описано (Manusama и др., Oncol. Rep.6, 173-177. (1999)).
Фигура 8. Уменьшение связывания интегрина усиливает цитотоксичность, вызванную TNF, TRAIL и FasL, in vitro. HUVEC культивировали на протяжении ночи на покрытых фибронектином микротитрационных планшетах при отсутствии (контроль) или в присутствии EMD121974 (300 мкМ), TNF (200 нг/мл), FasL (200 нг/мл), TRAIL (200 нг/мл), LIGHT (200 нг/мл) и IFN γ (330 нг/мл), как указано. Жизнеспособность определяли при помощи MST анализа.
Более подробно изобретение может быть описано при помощи следующих примеров:
Пример 1: Интегринзависимая адгезия эндотелиальных клеток в отношении TNFα-индуцированного апоптоза.
HUVEC образование сфероидов и выживание не требует интегринов
Для изучения влияния связывания интегрина на TNF-индуцированный апоптоз мы определяли условия, при которых эндотелиальные клетки могут культивироваться без интегринзависимой адгезии. Суспензия единичных клеток эндотелия быстро погибает вследствие анойкиса (апоптоза, индуцированного отделением клеток) (Meredith и др., Mol. Biol Cell. 4, 953-961 (1993)), что является препятствием для дальнейшего анализа. Однако при посеве клеток эндотелия пупочной вены человека (HUVEC) при высокой плотности (1,0×106 клеток/мл) в лунках, покрытых БСА, образуются многоклеточные сфероиды в течение 2-4 часов и могут сохраняться на протяжении 24 часов в зависимости от VE-кадхерина и независимо от какого-либо определяемого влияния со стороны интегринов. Ингибирование VE-кадхериновой активности при помощи блокирования моноклональным антителом (mAb), или путем уменьшения Са-Mg2+ при помощи EDTA, блокирует образование сфероидов, тогда как ингибирующий mAbs в отношении к α2, α3, α5, α6, β1, αVβ3 или αVβ5 интегринам, блокирующие пептиды на основе RGD и блокирующий в отношении РЕСАМ-1 mAb, сами или в комбинации, не оказывают влияния на HUVEC сфероиды (Фиг.1a, и данные не показаны).
Для определения действия сфероидной культуры на жизнеспособность клеток сфероиды и связанные с фибронектином HUVEC собирали в интервале между 6 и 72 часами выращивания культуры на планшете, серийно разводили и в дальнейшем культивировали на протяжении последующих 48 часов перед тем, как определяли относительное число клеток. Сдвиг влево или сглаживание кривой растворения свидетельствует о снижении жизнеспособности. Через 6, 12, 16 и 24 часа после выращивания культуры на планшете жизнеспособность HUVEC, восстановленных из сфероидных культур, была сопоставима с такой у связанных с фибронектином культур, но через 36 часов она прогрессивно снижалась (Фиг.1b через 16 часов, и данные не показаны). Взятые вместе, эти результаты свидетельствуют о том, что HUVEC может образовывать сфероиды, которые являются жизнеспособными на протяжении 24 часов при отсутствии интегринзависимой адгезии.
Пример 2: Адгезия к фибронектину защищает HUVEC от TNF-индуцированного апоптоза.
Для изучения того факта, модулируют ли интегрины TNF-опосредованный апоптоз, мы культивировали HUVEC на фибронектине (интегринзависимая адгезия) или в качестве сфероидов (интегриннезаивисимая адгезия) при отсутствии или в присутствии TNF (200 нг/мл) и IFNγ (330 нг/мл), усилителя цитотоксичности TNF (Dealtry и др., Eur. J. Immunol. 17, 689-693 (1987)). Воздействие TNF±IFNγ на монослои HUVEC на фибронектине ("фибронектинсвязанные HUVEC") не вызывает повышения апоптоза, о чем свидетельствует отсутствие поглощения YoPro-1 (Idziorek и др., J. Immunol. Methods 185, 249-258 (1995)), связывание поверхностями клеток аннексина V, активация каспазы-3 и разложение ее субстрата PARP (Фиг.2а, 2b и данные не показаны). В отличие от этого, обработка сфероидов TNF±IFNγ вызывает усиление поглощения YoPro-1 (усиление подавляется ингибиторами каспазы BOG, Z-VAD, IETD и DVED), фрагментацию ДНК, активацию каспазы-3 и разложение PARP (Фиг.2а, 2b и данные не показаны). Для исследования воздействия TNF±IFNγ на выживание клеток мы определяли жизнеспособность необработанных и обработанных культур. Действие TNF±IFNγ на фибронектинсвязанные HUVEC не влияло на жизнеспособность клеток (Фиг.2с). Обработка сфероидов TNF вызывала гибель более 80% клеток и комбинированная обработка TNF/IFNγ вызывала гибель всех клеток (Фиг.2d). Обработка только одним IFNγ не являлась цитотоксической (данные не показаны). HUVEC прилипают к иммобилизированному фибронектину при помощи αVβ3 и α5β1 интегринов (Rüegg и др., Nature Med. 4, 408-414 (1998)). Для изучения индивидуального вклада этих интегринов в выживание клеток на фибронектине мы культивировали HUVEC на иммобилизированных на пластике mAbs (imAbs) к αVβ3, α1, α5 и α4 интегринам. Иммобилизированные анти-αvβ3, анти-α5 и анти-α1 mAbs защищают HUVEC от TNF-индуцированной гибели, в то время как анти-α4 mAbs такого действия не обнаруживают (Фиг.2е и данные не показаны). Исходя из этих результатов нами был сделан вывод, что αVβ3 и αVβ1 интегрин-опосредованная адгезия подавляет TNF-индуцированный апоптоз, и при его отсутствии повышается чувствительность HUVEC к TNF и каспаз-опосредованному апопозу.
Пример 2: Интегринзависимая передача сигналов защищает эндотелиальные клетки от TNFα-индуцированного апоптоза.
TNFиндуцированная активация NF-κВ не требует связывания интегрина
Активация ядерного фактора-κВ (NF-κВ) способствует выживанию клеток, подвергнутых воздействию TNF (Beg & Baltimore, Science 274, 782-784; Van Antwerp и др., Science 274, 787-789 (1996)). Так как адгезия клеток при помощи интегринов активирует NF-κВ (Scatena и др., J, Cell Biol. 141, 1083-1093 (1998)), мы изучали, является ли чувствительность сфероидов к индуцированному TNF апоптозу обусловленной недостатком активации NF-κВ. Активация NF-κВ оценивалась путем измерения I-κВ фосфорилирования и деградации, NF-κВ ядерной транслокации и экспрессии поверхностями клеток ICAM-1, NF-κВ-индуцируемым геном (Collins и др., Faseb J. 9, 899-909. (1995)), в сфероидах и фибронектинсвязанных HUVEC культурах, подданных воздействию TNF±IFNγ. Мы не наблюдали существенных отличий в I-κВ фосфорилировании и деградации, NF-κВ ядерной транслокации или экспрессии ICAM-1 (Фиг.3а-с), что свидетельствует о том, что TNF-идуцированный апоптоз в культурах HUVEC сфероидов не связан с ослабленной активацией NF-κВ.
Пример 3: Активация обусловлена связыванием интегрина и является существенной для выживания клеток.
Затем анализировалась активация Akt/РКВ, активирование киназы белков при помощи TNF, что способствует выживанию эндотелиальных клеток (Madge & Pober, J.Biol. Chem. 275, 15458-15465. (2000)). Основное Akt фосфорилирование в фибронектин-связанных HUVEC повышалось при воздействии TNF/IFNγ, что согласуется с конститутивной и TNF-индуцированной Akt активацией. В отличие от этого, не наблюдалось фосфорилирования Akt в необработанных сфероидах и воздействие TNF/IFN вызывало только слабое фосфорилирование (Фиг.4а). Для оценки значимости активации Akt для выживания HUVEC мы обрабатывали фибронектинсвязанные клетки вортманнином и LY294002, двумя фармакологическими ингибиторами фосфоинозитид-3 (PI-3) киназы, активатора начальных процессов активации Akt (Kandel, & Hay, Exp. Cell Res. 253, 210-229. (1999)). Мы также экспрессировали конститутивно активную форму Akt (Aktmp) и каталитическую субъединицу PI-3 киназы (р110*) в сфероидах. Обработка вортманнином и LY294002 вызывает усиление апоптоза и снижает выживание фибронектинсвязанных клеток при воздействии TNF±IFNγ (Fig. 4b), в то время как Aktmp и р110*, но не дикий тип Akt (Aktwt) или контрольная плазмида (pBS), защищает сфероиды от апоптоза, индуцируемого TNF±IFNγ (Фиг.4с). На основе этих результатов нами был сделан вывод о том, что активация Akt является существенной для выживания HUVEC, подвергнутых воздействию TNF±IFNγ, и что как основная, так и TNF-индуцировання активация Akt зависит от связывания интегрина.
вызывало только слабое фосфорилирование (Фиг.4а). Для оценки значимости активации Akt для выживания HUVEC мы обрабатывали фибронектинсвязанные клетки вортманнином и LY294002, двумя фармакологическими ингибиторами фосфоинозитид-3 (PI-3) киназы, активатора начальных процессов активации Akt (Kandel, & Hay, Exp. Cell Res. 253, 210-229. (1999)). Мы также экспрессировали конститутивно активную форму Akt (Aktmp) и каталитическую субъединицу PI-3 киназы (р110*) в сфероидах. Обработка вортманнином и LY294002 вызывает усиление апоптоза и снижает выживание фибронектинсвязанных клеток при воздействии TNF±IFNγ (Fig. 4b), в то время как Aktmp и р110*, но не дикий тип Akt (Aktwt) или контрольная плазмида (pBS), защищает сфероиды от апоптоза, индуцируемого TNF±IFNγ (Фиг.4с). На основе этих результатов нами был сделан вывод о том, что активация Akt является существенной для выживания HUVEC, подвергнутых воздействию TNF±IFNγ, и что как основная, так и TNF-индуцировання активация Akt зависит от связывания интегрина.
Пример 4: Выживание обработанных TNF HUVEC требует активации Akt и NF-κВ.
Aktmp подавляет TNF-индуцированный апоптоз сфероидов в присутствии активной NF-κВ. Мы также изучали, является ли активация обеих путей передачи сигнала NF-κВ и Akt необходимой для выживания, или достаточно активации исключительно Akt. Мы блокировали активацию NF-B в клетках, которые экспрессируют конститутивно активный Akt (Aktmp) путем инфицирования HUVEC аденовирусом, который экспрессирует неразлагаемый 1-κВ (AdΔNI-κВ - который предотвращает IkB-NFκB диссоциацию (Brown и др., Science 267, 1485-1488. (1995)). AdΔNI-κВ повышает чувствительность фибронектин-связанных HUVEC к TMF±IFNγ-индуцированному апоптозу и на это не влияет Aktmp. Контрольная электропорация (pBS) или заражение аденовирусом (AdLacZ) также не оказывает влияния. AdΔNI-κВ также повышает чувствительность сфероидов, которые сверхэкспрессируют Aktmp, к TNF±IFNγ-индуцированному апоптозу (Фиг.4d). Для определения того факта, может ли Akt защищать от воздействия TNF в низких дозах в HUVEC при отсутствии NF-κВ активации, адгезированные монослои wt и HUVEC, которые сверхэкспрессируют Aktmp, инфицировались AdΔNI-κВ и подвергались воздействию TNF (0,33-100 нг/мл). AdΔNI-κВ повышал чувствительность HUVEC к апоптозу (TNF>0,1 нг/мл), но Aktmp не оказывал защитного воздействия на такие HUVECS даже при таких низких дозах TNF (Фиг.4е). Более того, LY294002 и вортманнин не ингибировали индуцированную TNF экспрессию ICAM-1, что указывает на то, что NF-κВ активация в HUVEC не требует передачи сигналов при помощи Akt (Фиг.4f и данные не показаны), и согласуется с индукцией ICAM-1 в сфероидах (см. фигуру 3с). В отличие от этого, инфицирование HUVEC AdΔNI-κВ подавляет экспрессию ICAM-1 в ответ на действие TNF±IFNγ (Фиг.4f). Взятые вместе, эти результаты свидетельствуют о том, что выживание HUVEC при воздействии TNF±IFN γ требует совместной активации Akt и NF-κВ.
Пример 5: Связывание интегрина способствует активации FKHR и MDM2 и подавляет фосфорилирование МЕК, р38 и JNK
Антиапоптотическое действие Akt первоначально приписывалось его фосфорилированию и ингибированию каспазы-9 и Bad (Datta и др., Genes Dev. 13, 2905-2927. (1999)). Сейчас, однако, показано, что Akt-зависимое выживание связано с фосфорилированием и ингибированием "forkhead" транскрипционных факторов (FKHR/FKHRL1) (Datta и др., Genes Dev. 13, 2905-2927. (1999); Brunet и др., Cell 96, 857-868. (1999)) и MDM2, р53 деградацией (Mayo & Donner, Proc. Natl. Acad. Sci. USA 98, 11598-11603. (2001)), и подавлением активации протеинкиназ ERK, р38 и JNK (Rommel и др., Science 286, 1738-1741. (1999); Gratton и др., J. Biol. Chem. 276, 30359-30365. (2001); Park и др., J. Biol. Chem. 277, 2573-2578. (2002); Madge & Pober, J. Biol. Chem. 275, 15458-15465. (2000)). Мы исследовали, является ли недостаточность связывания интегрина и Akt передача сигналов связаны с перестройкой этих путей передачи сигналов. Нами определялись уровни MDM2, р53 и фосфорилирования FKHR/FRKHL1, МЕК, р38 и -JNK в связанных HUVEC и сфероидах, подданных воздействию TNF/IFNγ. Такие сфероиды обладали недостаточным фосфорилированием FKHR/FKRL1, пониженными уровнями MDM2 и накоплением р53 по сравнению с фибронектинсвязанными клетками (Фиг.5а). Кроме того, сфероиды имели повышенные основное и TNF/IFNγ-индуцированное фосфорилирование МЕК, р3 8 и JNK (Фиг.5b).
Эти результаты согласуются с ролью Akt в поддерживании выживания путем ингибирования FKHR/FKHRL1, путем снижения уровней р53 и путем подавления фосфорилирования МЕК, р38 и JNK.
Пример 6: Ингибирование интегринзависимой адгезии при помощи соединений с небольшими молекулами повышает чувствительность эндотелиальных клеток к TNFα-индуцированному апоптозу in vitro и in vivo.
Снижение связывания интегрина повышает чувствительность связанных HUVEC к апоптозу, индуцированному TNF.
Повышение чувствительности к TNF в условиях снижения связывания интегрина не является однозначным для сфероидов: культивирование HUVEC на поли-L-лизине (PLL), субстрате, который способствует интегринзависимой адгезии (Bershadsky и др., Curr. Biol. 6, 1279-1289. (1996)) сохраняет выживание на PLL, и добавление TNF±IFN γ вызывает массивную гибель (Фиг.6а), эта гибель предотвращается экспрессией Aktmp (не показано). Кроме того, мы селективно ингибировали интегрин αVβ3 в HUVEC на фибронектине при помощи EMD121974 ((циклический (Arg-Gly-Asp-D-Phe-[N-Me]-Val), который является антагонистическим циклопептидом αVβ3/αVβ5) (Dechantsreiter и др., J. Med. Chem. 42, 3033-3040. (1999)), что не оказывает влияния на  компонент α5β1/αVβ3-зависимой адгезии на фибронектине (Фиг.6b). Тогда как ни TNF/IFNγ, ни EMD121974 отдельно не влияли на выживание, сочетанное воздействие TNF/IFNγ и EMD121974 (но не неингибиторного контрольного пептида EMD135981) повышало апоптоз и отделение (Фиг.6с), и уменьшало выживание (Фиг.6d). Экспрессия Aktmp защищала фибронектинсвязанные HUVEC от апоптоза, индуцированного TNF, IFNγ и EMD121974 (Фиг.6е).
компонент α5β1/αVβ3-зависимой адгезии на фибронектине (Фиг.6b). Тогда как ни TNF/IFNγ, ни EMD121974 отдельно не влияли на выживание, сочетанное воздействие TNF/IFNγ и EMD121974 (но не неингибиторного контрольного пептида EMD135981) повышало апоптоз и отделение (Фиг.6с), и уменьшало выживание (Фиг.6d). Экспрессия Aktmp защищала фибронектинсвязанные HUVEC от апоптоза, индуцированного TNF, IFNγ и EMD121974 (Фиг.6е).
Пример 7: Уменьшение связывания интегрина повышает чувствительность связанных HUVEC к апоптозу, индуцированному различными лигандами гибели семейства TNF-лигандов.
Повышение чувствительности к проапоптотическим действиям передачи сигналов при помощи рецепторов гибели при уменьшении связывания интегрина не ограничивается TNF, но также наблюдается при воздействии TRAIL и FasL в присутствии EMD121974 на HUVEC, культивируемых на фибронектине. LIGHT, лиганд, который связывается с рецепторами, уменьшая участок гибели, не обнаруживает синергизма с αVβ5-блокированием (Фиг.7).
Пример 8: EMD121974 повышает чувствительность, установленную для опухолей, к противоопухолевым действиям TNF
Ангиогенные эндотелиальные клетки экспрессируют αVβ3 интегрин и связывание αVβ3 способствует выживанию эндотелиальных клеток (Brooks и др., Cell 79, 1157-1164 (1994); Brooks и др., Science 264, 569-571 (1994)). Сведения, что EMD121974 повышает чувствительность эндотелиальных клеток к апоптозу, индуцируемому TNF in vitro, свидетельствуют о том, что это соединение может усиливать противоопухолевое действие TNF. Для изучения этой гипотезы мы подвергали лечению крыс, которые имеют изогенную саркому мягких тканей BN175, которая является чрезвычайно агрессивной и васкуляризированной опухолью, резистентной к цитотоксическому действию TNF in vitro и in vivo (Manusama и др., Oncol. Rep.6, 173-177. (1999)). Мы применяли методику изолированной перфузии конечности (ILP) для введения TNF, EMD121974 или их сочетания к конечности с опухолью. Лечение при помощи TNF или пептида отдельно не оказывало влияния на рост опухоли. В противоположность этому, комбинированное введение TNF и EMD121974 вызывало полную регрессию опухоли у 50% животных и в общем существенное уменьшение роста опухоли (Фиг.8). При этом у животных, подданных воздействию EMD121974/TNF, не наблюдалось местной и общей токсичности, что свидетельствует о том, что EMD121974 селективно повышает чувствительность опухолей в отношении цитотоксичности TNF. Так как опухолевые клетки BN175 являются нечувствительными к TNF и не экспрессируют активный αVβ3 интегрин, о чем свидельствует их слабая адгезия к фибриногену даже в присутствии высоких концентраций Mn2+, и их низкая чувствительность к селективным ингибиторам αVβ3, таким как EMD 121974 (неопубликованное исследование), нами был сделан вывод о том, что эффективное синергетическое противоопухолевое действие наиболее вероятно связано с разрушением сосудистой сети опухоли.
Взятые вместе с нашими полученными in vitro данными, они являются существенным подтверждением значимости интегрина αVβ3 по сравнению с αVβ1 в этой системе для контроля выживания эндотелия.
Пример 9: Культивирование HUVEC и электропорация
HUVEC получали и культивировали, как описано ранее (Ruegg и др., Nature Med 4, 408-414 (1998)) и применяли между пассажами 3 и 7. Полная сыворотка представляла собой: M199 (Life technologies, Базель, Швейцария), 10% PCS (сыворотки плода коровы) (Seromed, Берлин, Германия), 12 мкг/мл экстракта головного мозга крупного рогатого скота (Clonetics-Bio Whittaker, Walkersville, MD, США), 10 нг/мл человеческого рекомбинантного EGF (фактора роста эпидермиса) (Peprotech, London, UK), 25 ед./мл гепарина, 1 мкг/мл гидрокортизона (Sigma Chemie), 2 мМ L-глутамина, 100 мкг/мл стрептомицина и 100 ед./мл пенициллина (Life Technologies). Для электропорации HUVEC ресуспендировали в полной сыворотке, инкубировали на льду на протяжении 5 мин с ДНК (20 мкг специфической плазмиды и 5 мкг pEGFP-C1) и электропорировали с генератором импульсов для генетической трансформации (Biorad, Глаттбругг, Швейцария). Электропорированные HUVEC культивировали на протяжении 48 часов перед использованием. Приблизительно 80% клеток экспрессировали EGFP 40 часов после электропорации.
Пример 10: Образование сфероидов
HUVEC собирали в результате расщепления трипсином, ресуспендировали в полной сыворотке при 1,0×106 клеток/мл и высевали 1 мл/лунку в 12 лунковые планшеты нетканевой культуры (Evergreen Scientific, Лос Анжелос, СА, США) предварительно покрытые 1% БСА. Для изучения агрегации 200 мкл клеточной суспензии высеивали в микролунки ELISA планшетов, покрытые 1% БСА (Maxisorp II, NUNC, Росклид, Дания) отдельно или в присутствии mAbs (10 мкг/мл), EDTA (5 мМ) или Са /EDTA (10/5 мМ). Образование сфероидов оценивали через 6 часов и 16 часов. Микроснимки делали при помощи микроскопа Televal 31 (Carl Zeiss AG, Цюрих, Швейцария).
Пример 11: Морфологический анализ сфероидов
Для оценки морфологии сфероиды вставляли в Epon (Fluka Chemie) и толстые срезы окрашивали 1% метиленом/азуровым синим. Для иммунологического окрашивания замороженные срезы сфероидов фиксировали в 4% (Fluka Chemis, Буче, Швейцария) формальдегиде. После блокирования 1% БСА срезы последовательно инкубировали на протяжении 1 часа с праймером mAb (20 мкг/мл) и циан3-меченной GaM антисывороткой (West Grove, PA, США). Для реакции TUNEL замороженные срезы сфероидов фиксировали в 4% параформальдегиде и обрабатывали как описано в (Ruegg и др., I.c.). Подсчитывали окрашивание сфероидов с йодидом пропидия для общего содержания ДНК. Срезы наблюдали в епифлуоресцентном микроскопе (Axioskop, Carl Zeiss AG), оборудованном CCD камерой, (Photonic Science, Милхем, UK) или при помощи лазерного конфокального микроскопа (LSM 410, Carl Zeiss AG). Индекс апоптоза определяли путем вычисления соотношения между зелеными (TUNEL окрашенные фрагменты ДНК) и красными (окрашенная йодидом пропидия общая ДНК) пикселями.
Число исследованных сфероидов на условие составляло: С, 31; Т, 21; Т1, 12. Для определения апоптических клеток в культуре краситель ДНК YoPro-1 (250 нМ) добавляли к целой культуре или к собранным подвижным клеткам (Delhase, М., Li, N. & Karin, M. Kinase regulation in inflammatory response. Nature 406, 367-368. (2000)). Культуры наблюдали в инвертированном флуоресцентном микроскопе (Leica DM IRB, Хербругг, Швейцария). Для электронной микроскопии сфероиды фиксировали 2,5% глутаральдегидом в 100 мМ какодилатного буфера и в последующем фиксировали в 1% OsO4. Клетки обезвоживали этанолом и вставляли в Epon. Сверхтонкие срезы исследовали, используя трансмиссионный электронный микроскоп Philips CM10.
Пример 12: Выживание клеток и пролиферация.
Для выживания HUVEC сфероиды на планшете 1×106 клеток/мл в покрытых 1% БСА 24 мм лунках, или адгезированные клетки на планшете 4×105 клеток в покрытых фибронектином 3 мкг/мл 35 мм лунках планшетов нетканевых культур (Evergreen Scientific), подвергали стимуляции с TNFα (200 нг/мл=104 ед./мл)± IFNγ (330 нг/мл=104 ед./мл). Ингибиторы киназы или EMD пептиды добавляли 1 час или 4 часа перед стимуляцией, соответственно при следующих концентрациях: вортманнин, 100 нМ; LY294002, 20 мкМ; EMD пептиды, 300 мкМ. Через 16 часов культуры, клетки собирали при помощи диссоциации (5 мин при 20°С для сфероидов) с 5 мМ EDTA или 1х трипсина (для адгезированных культур), промывали, ресуспендировали в полной сыворотке до 4×105 клеток/мл, аликвота до 100 мкл/лунку в микротитровальных планшетах тканевых культур (Falcon, Becton Dickinson) и титровали в 1:2 этапах в трех повторах. Относительное число клеток оценивали через 48 часов путем измерения превращения МТТ на протяжении последних 4 часов в культуре. Результаты поданы в виде значений О.П. при 540 нм (Packard Spectra Count, Мериден, СТ, США) и представлены в виде значений при трех повторах на лунку±с.о.
Пример 13: Исследования отделения клеток
Maxisorp II ELISA планшеты покрывали 1 мкг/лунку фибронектина или 0,5% желатина на протяжении ночи при 4°С в фосфатно-солевом буферном растворе PBS. Покрытые лунки промывали и блокировали с 1% БСА на протяжении 2 часов при 37°С и промывали перед использованием. Добавляли 2×104 кл/лунку в основной сыворотке без FCS и быстро осаждали путем центрифугирования (40×g). Клеткам давали возможность адгезировать на протяжении 2 часов при 37°С перед прибавлением пептидов в разных концентрациях. Через 2 часа клетки промывали теплым фосфатно-солевым буферным раствором и прикрепленные клетки фиксировали в 2% параформальдегиде, окрашивали 0,5% кристаллическим фиолетовым (Sigma Chemie) и измеряли О.П. при 620 нм (Packard Spectra Count). Результаты поданы в виде значений О.П. и представлены в виде значений при двух повторах на лунку ± с.о. специфической адгезии (=адгезия на ЕСМ белке минус адгезия на БСА).
Пример 14: Проточная цитометрия
Непрямое иммунологическое окрашивание HUVEC и экспрессию EGFP выполняли согласно стандартному протоколу (Ruegg и др., I.c.). Погибшие клетки исключали при помощи окрашивания йодидом пропидия. Все образцы анализировали при помощи FACScan II® и Cell Quest® компьютерных программ (Becton Dickinson, Mountain View CA, США).
Пример 15: Исследование изменения электрофоретической подвижности (EMSA)
Ядерные экстракты HUVEC (1×10 клеток на условие) приготавливали как описано (Cai и др., J.Biot Chem 272, 96-101. (1997)) и инкубировали с синтетическим двунитевым 31-мерным олигонуклетотидом, содержащим κВ последовательности HIV человека с длинным концевым повтором, меченным на конце [γ-32 Р]АТФ, используя Т4 киназу. Связывание NF-κВ с меченным Р олигонуклетидом определяли путем PAGE и авторадиографии.
Пример 16: Вестерн-блоттинг
50 мкл супернатанта лизата клеток (1×106 в 250 мкл 2× Laemmli буфера) растворяли при помощи 7,5%-12,5% SDS-PAGE и переносили при помощи влажного промокания (Bio Pad) на Immobilon-P мембраны (Millipore, Волкетсвилл, Швейцария). Мембраны последовательно инкубировали в 5% сухом молоке на протяжении 1 часа, с исходным антителом на протяжении ночи при 4°С, и с HRP-меченным GaM (Dako, Зуг, Швейцария) на протяжении 1 часа. Для определения использовали систему ECL (Amersham-Pharmacia Biotech). Для повторного зондирования мембраны очищали в 2% SDS, 50 мМ Трис и 100 мМ ВМЕ на протяжении 30 мин при 50°С.
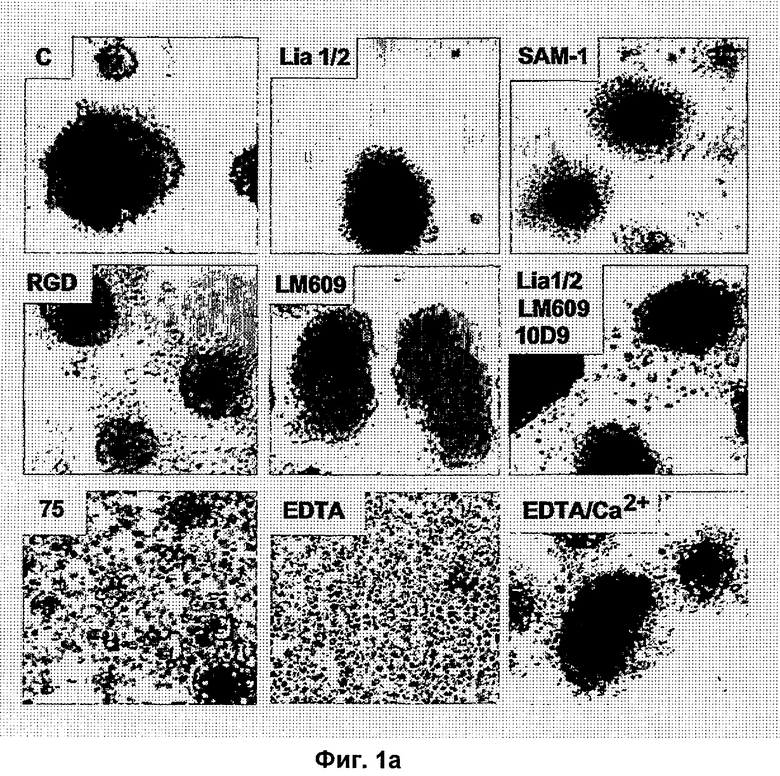
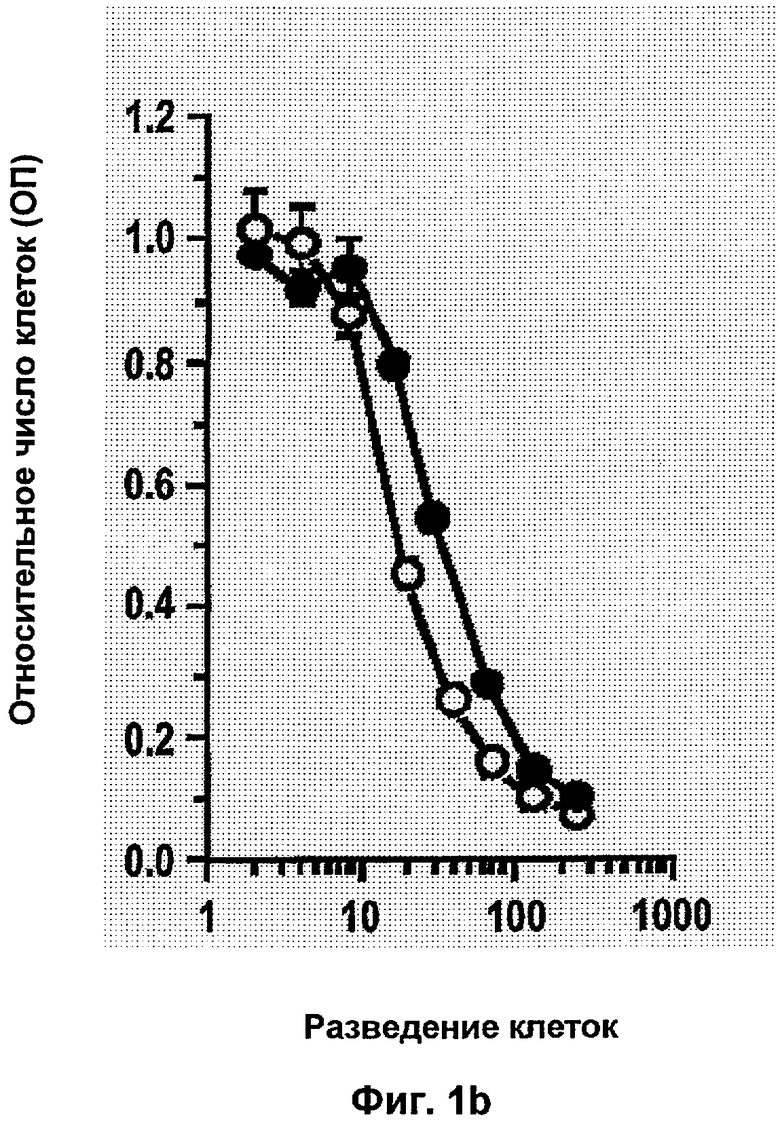

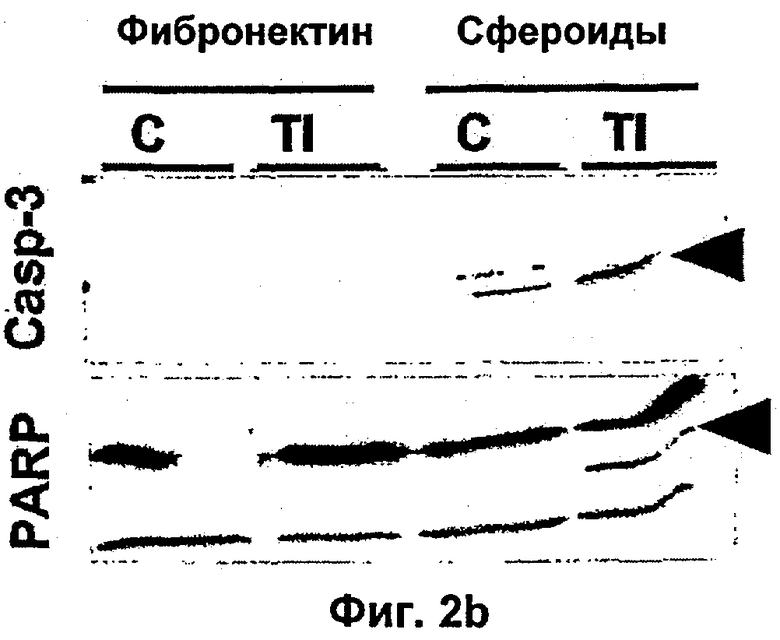
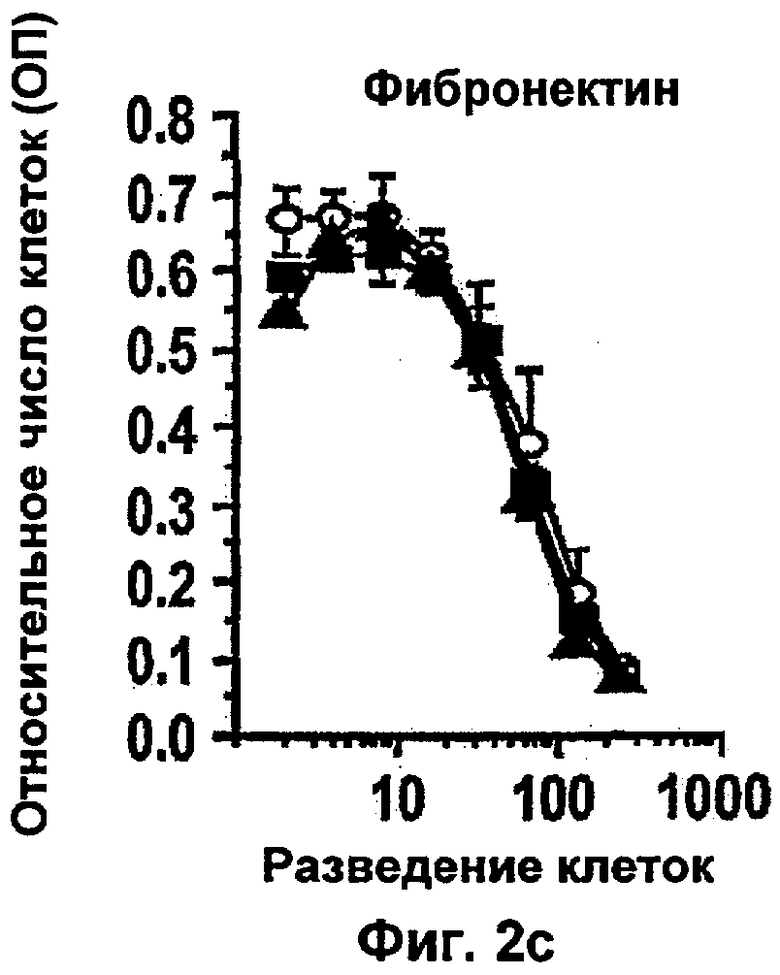
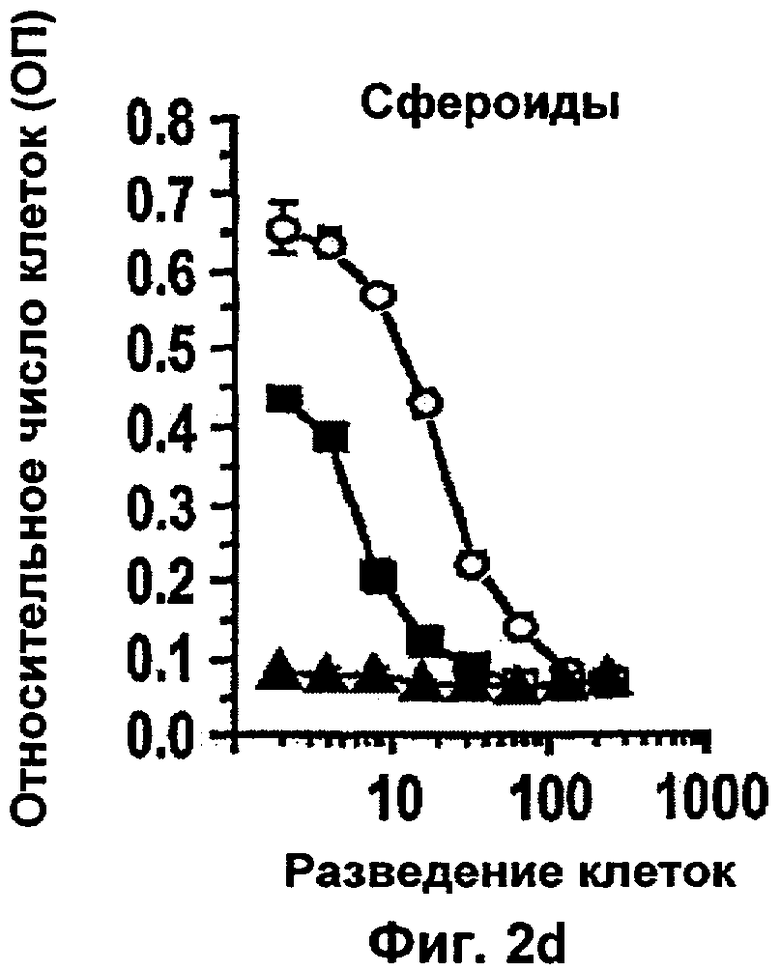
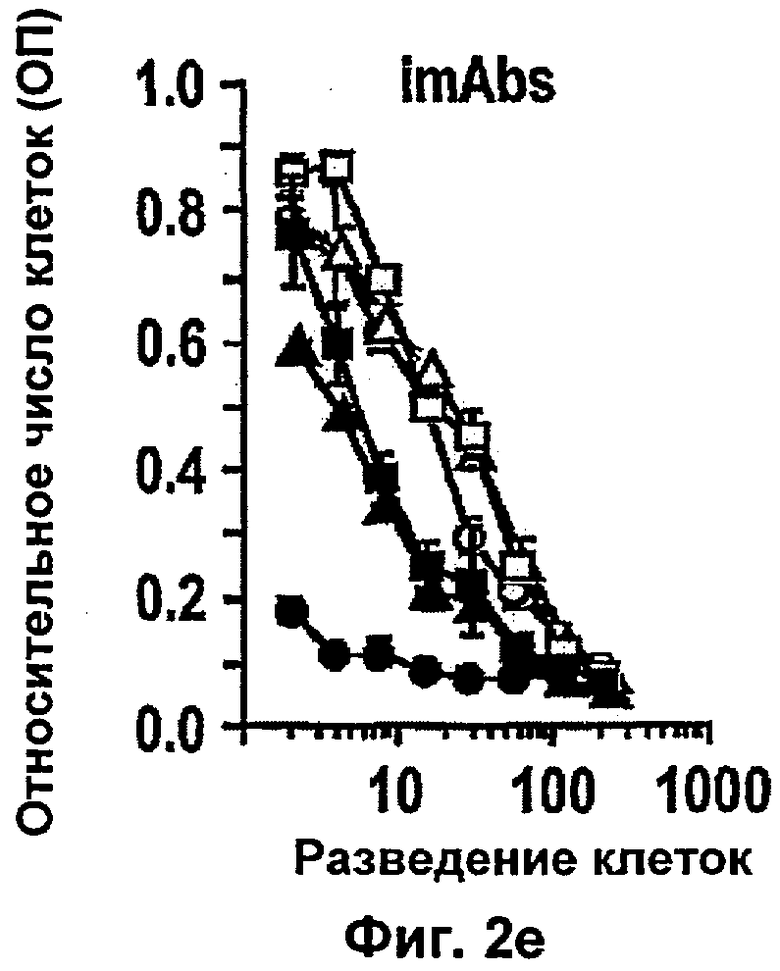
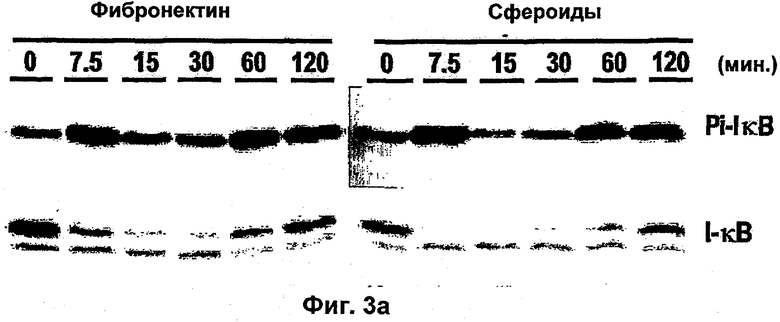
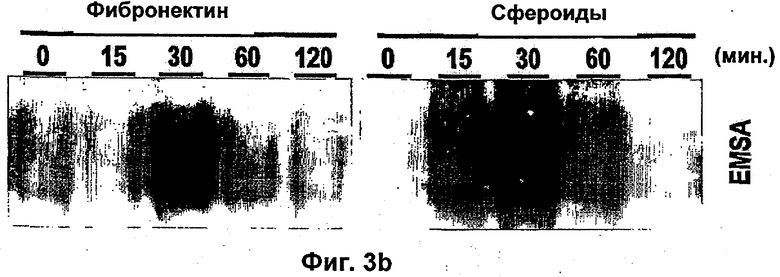
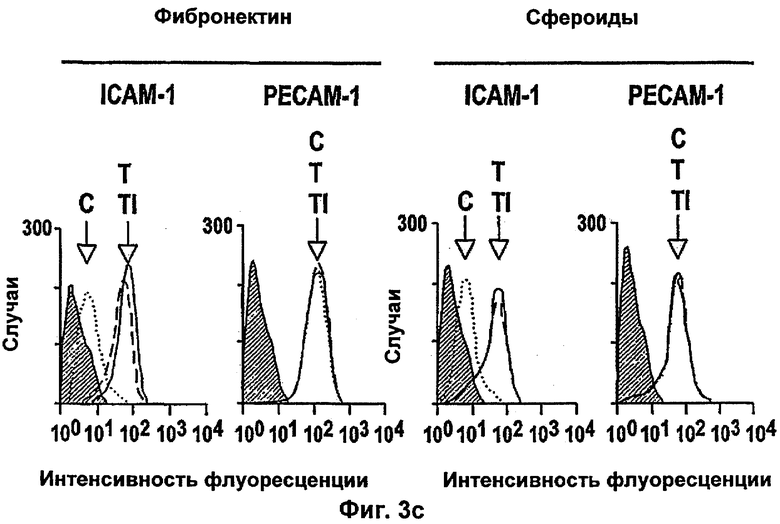

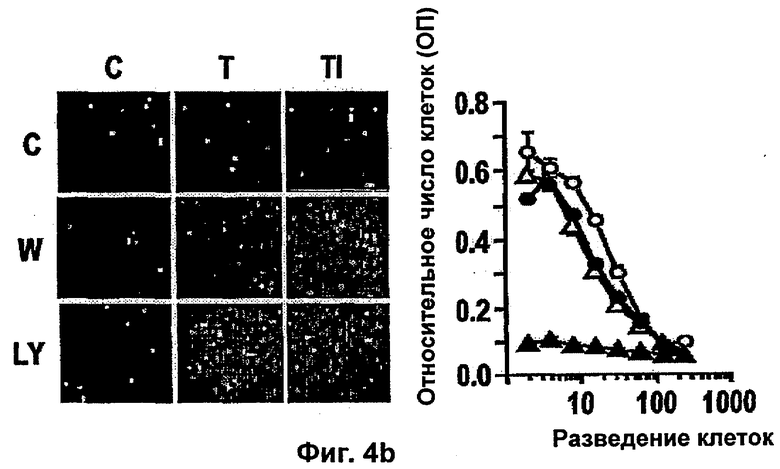
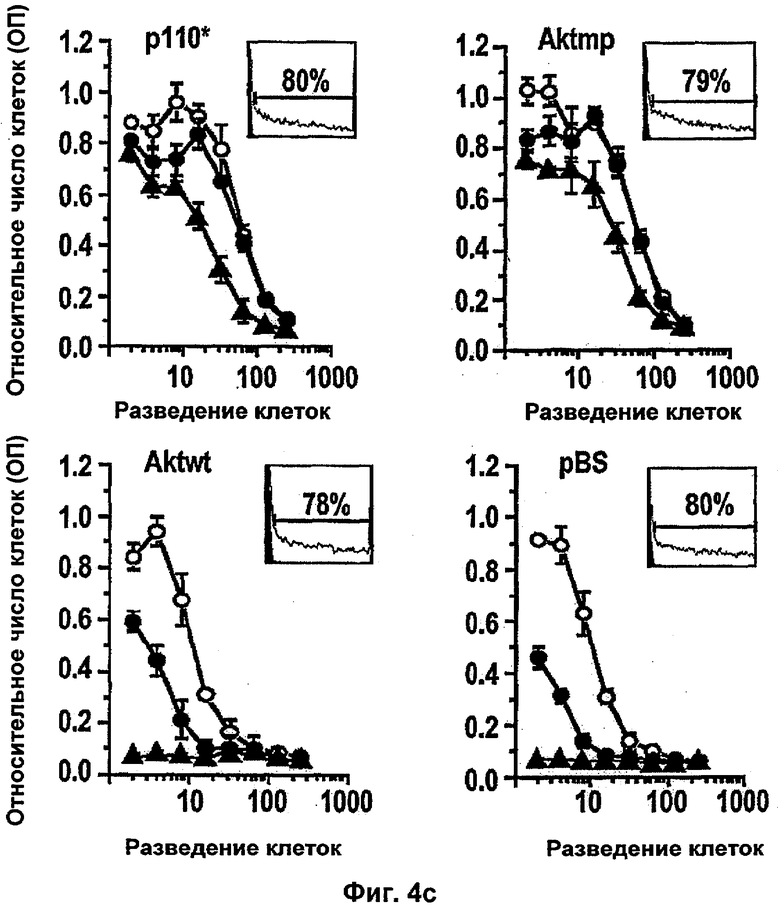
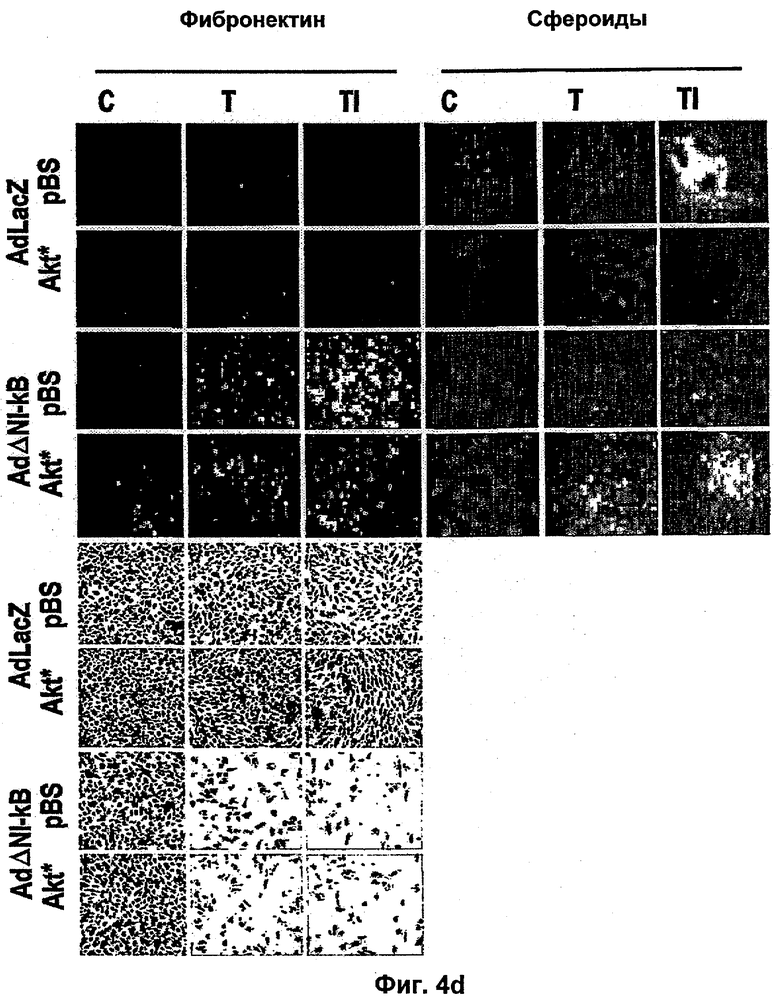
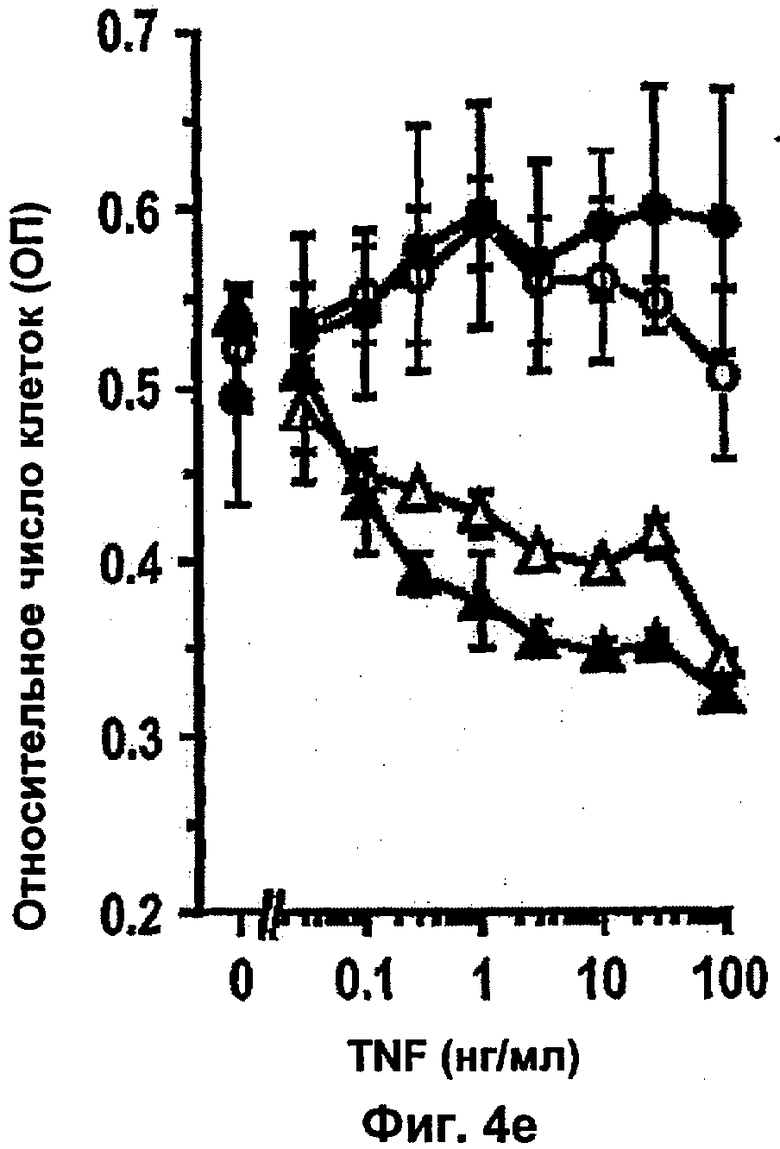
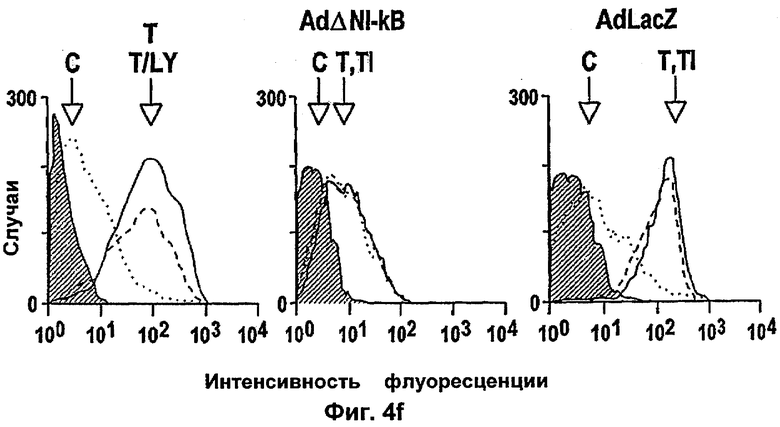
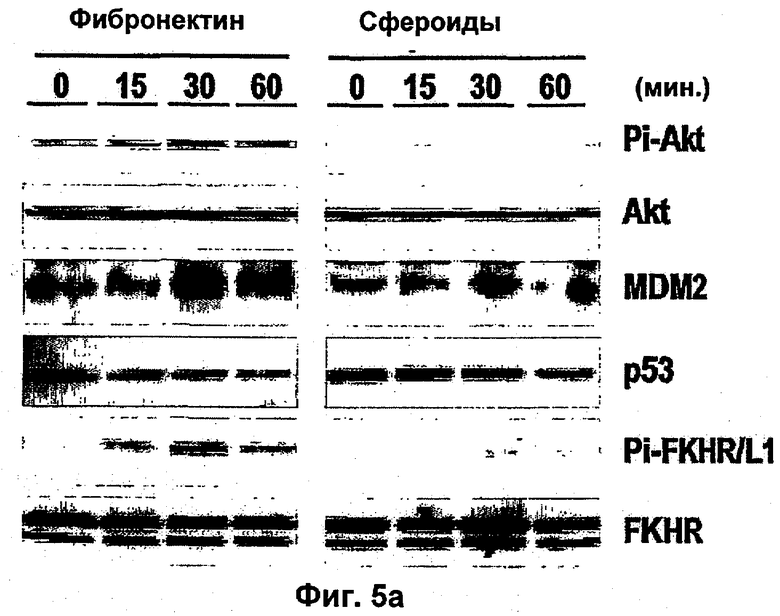
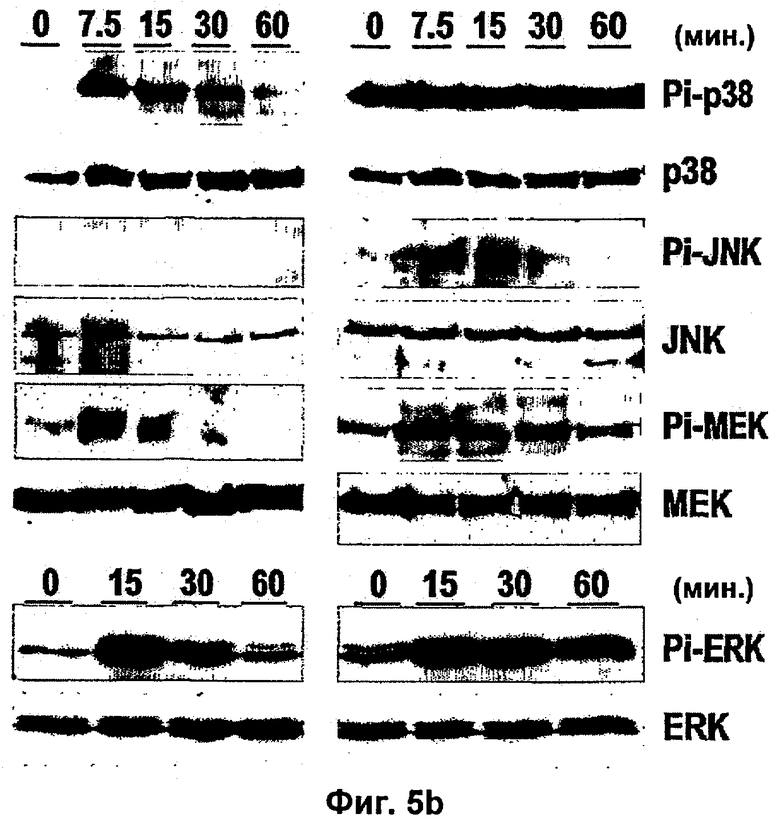
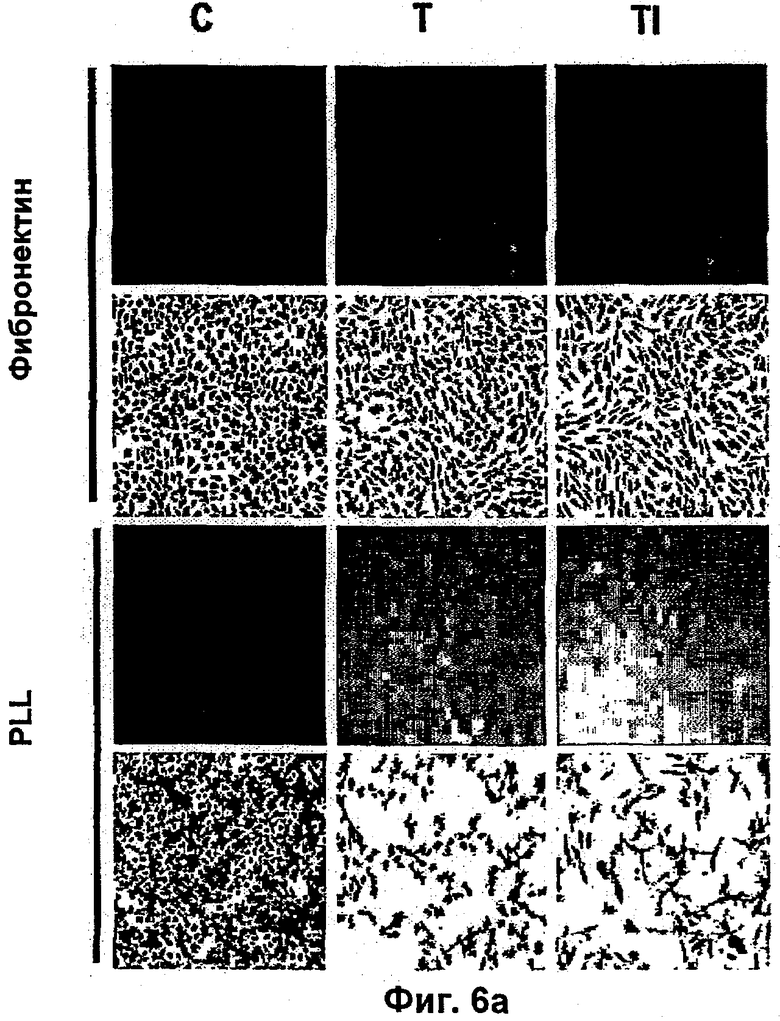
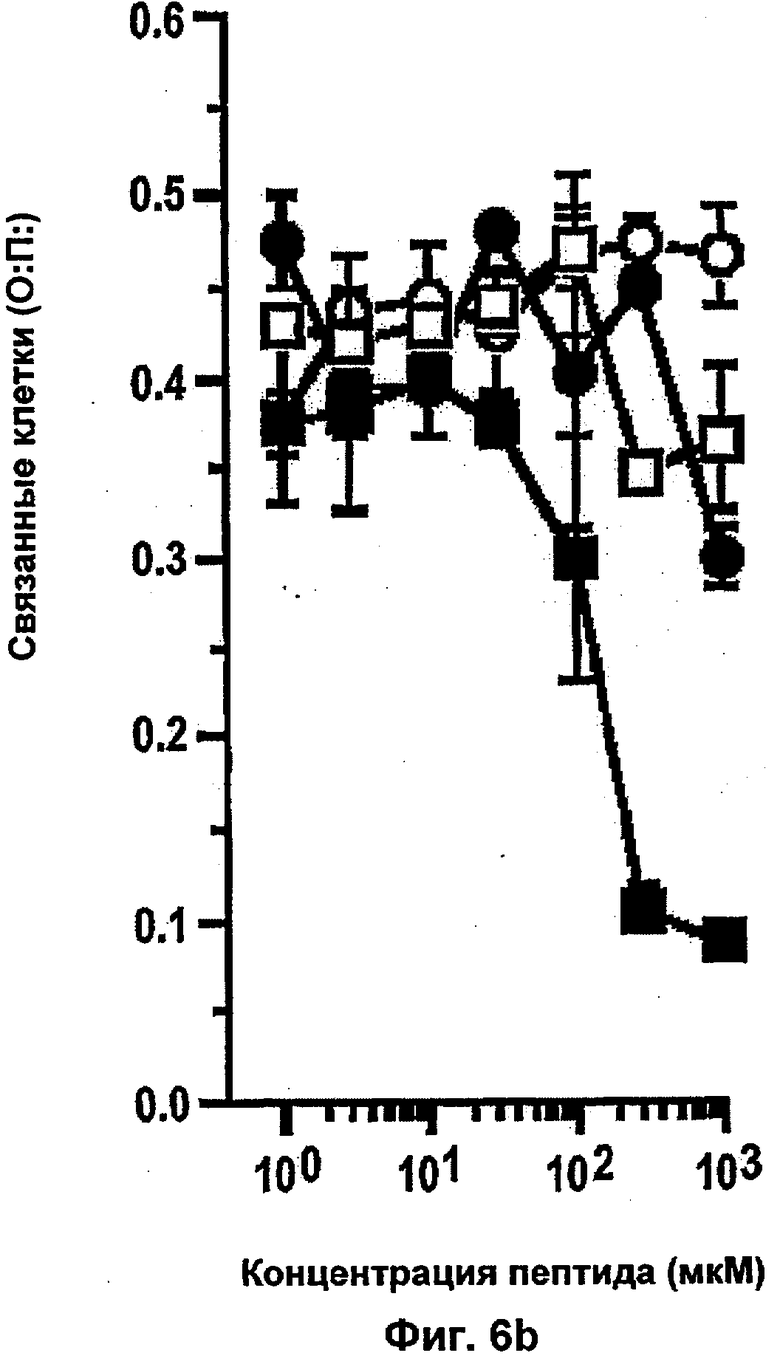
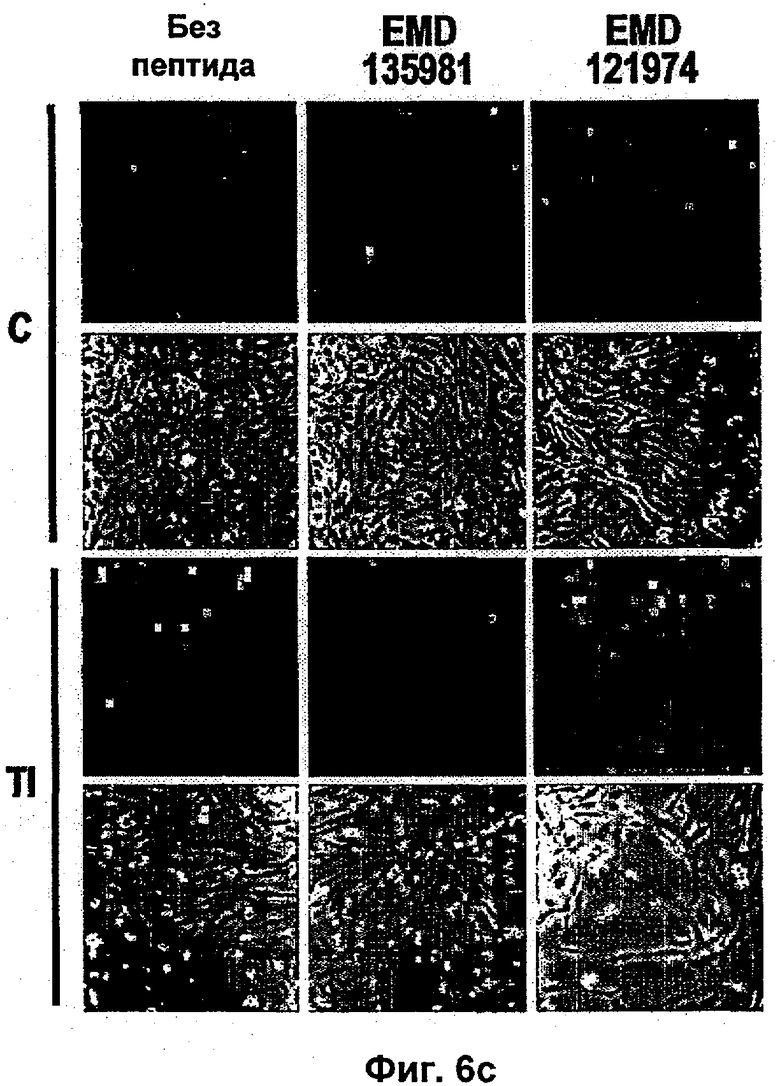
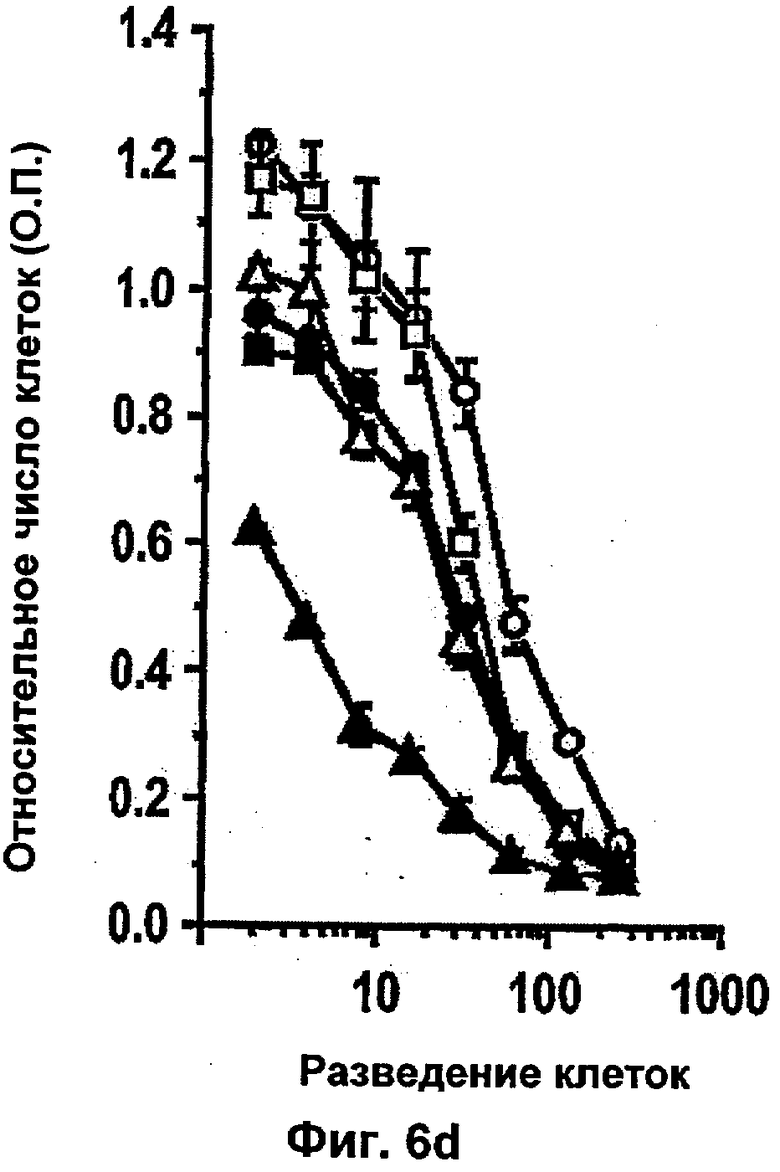

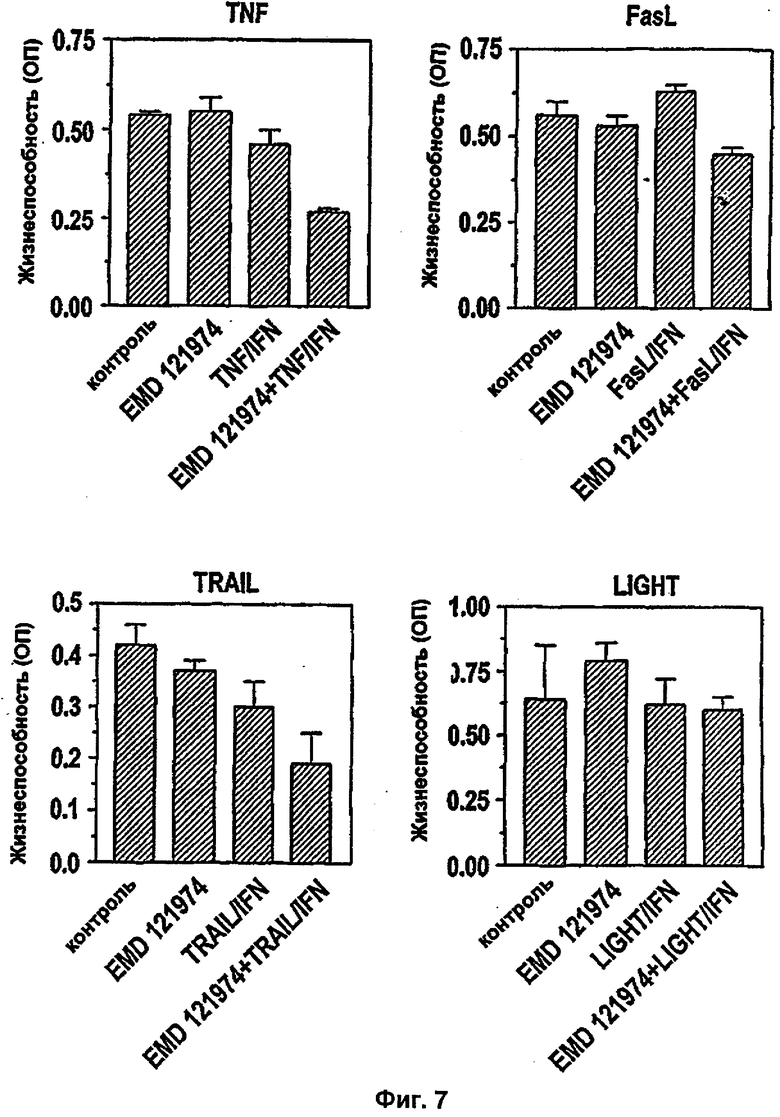
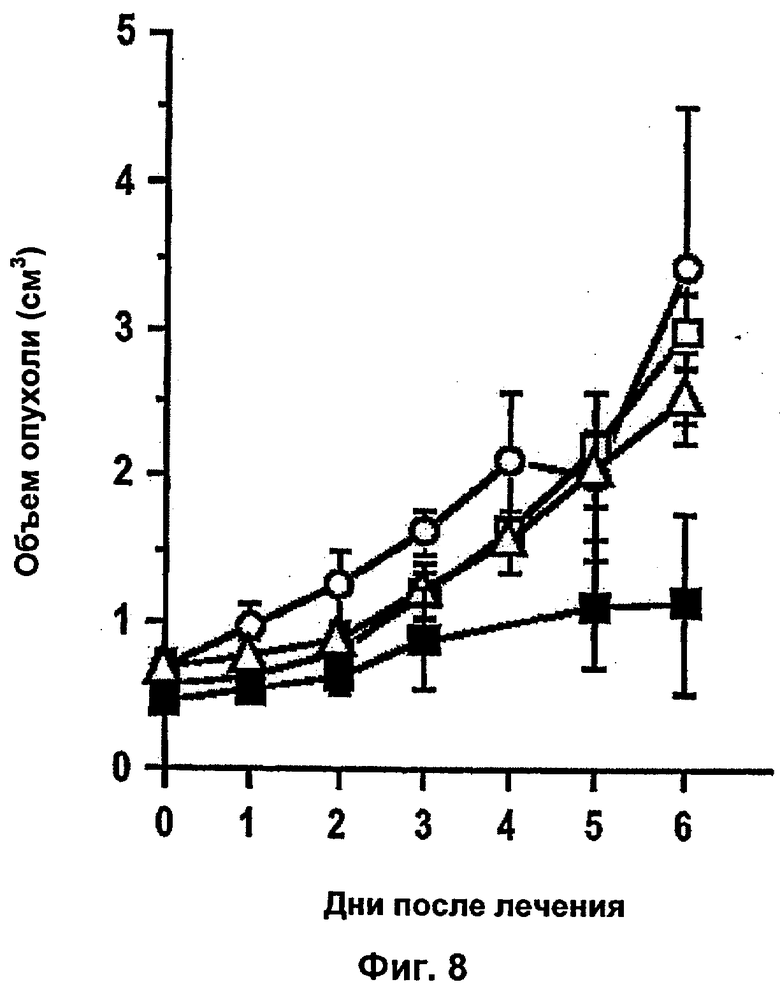
Изобретение относится к области медицины, а именно к комбинированной терапии, использующей антиангиогенные средства и TNFα. Сущность изобретения включает фармацевтическую композицию для лечения эндотелиальных опухолей, содержащая в терапевтически эффективном количестве антиангиогенное средство, представляющее собой пептид, включающий последовательность RGD, а именно цикло (Arg-Gly-Asp-DPhe-NMeVal) и фактор некроза опухоли TNFα. Изобретение также включает набор, включающий фармацевтическую композицию, применение указанных фармацевтической композиции и набора для приготовления лекарственного средства и способ лечения. Преимущество изобретения заключается в создании средства эффективного для лечения эндотелиальных опухолей. 4 н. и 11 з.п. ф-лы, 8 ил.
(i) цикло (Arg-Gly-Asp-DPhe-NMeVal),
(ii) TNFα,
(iii) IFNγ.
(i) цикло(Arg-Gly- Asp-DPhe-NMeVal),
(ii) TNFα,
(iii) IFNγ.
(ii) TNFα.
(i) цикло(Arg-Gly-Asp-DPhe-NMeVal),
(ii) TNFα, и
(iii) IFNγ.
| EP 1077218 A, 21.02.1991 | |||
| Способ изготовления сотового заполнителя | 1978 |
|
SU770622A1 |
| LEJEUNE F.J | |||
| et al | |||
| Clinical applications of TNF-alpha in cancer, carent opinion in immunology, Carent Biology LTD, XX, 1998, v.10, № 5, pp.573-580. | |||

