Настоящее изобретение относится к новому гену, обозначенному Grb 3-3, к его вариантам и к их применениям, например, в генной противораковой терапии.
Различные гены, называемые онкогенами и супрессорными генами, вовлечены в контроль клеточного деления. Среди них ras-гены и их продукты, обычно обозначаемые протеинами p21, играют ключевую роль в пролиферации клеток во всех эукариотных организмах, где они были исследованы. Так было показано, что некоторые специфические модификации этих протеинов лишены своего нормального контроля и становятся онкогенными. Так, большое число человеческих опухолей было ассоциировано с наличием модифицированных ras-генов. Кроме того, гиперэкспрессия этих протеинов p21 может привести к нарушению пролиферации клеток. Следовательно, понимание точной роли этих протеинов p21 в клетках, способа их функционирования и их характеристик является основной целью для понимания и подхода к терапии канцерогенеза.
Различные факторы, причастные к пути ras-зависимой сигнализации, были идентифицированы. Среди них фигурирует ген Grb 2, который кодирует протеин 23-25 кДа, имеющий структуру SH3-SH2-SH3 /Lowensteinetal, Cell 70 /1992/ 431; Matuoka et al., PNAS 89 /1992/ 9015/. Возможно продукт гена Grb 2 взаимодействует с протеинами, фосфорилированными по тирозину в его SH2 области и с фактором обмена GDP класса SOS в его SH3 области /Egah et al., Nature 363/ 1993/ 45/. Таким образом он является одним из компонентов трансформирующей активности продукта ras-гена. Настоящее изобретение вытекает из обнаружения, клонирования и характеристики изоформы гена Grb 2, обозначенной Grb 3-3, имеющей делецию в области SH2. Этот ген определен во взрослых тканях: соответствующая мРНК находится под единой полосой 1,5 kb и транслируется в протеин 19 кДа. Из-за наличия делеции в области SH2 продукт гена Grb 3-3 не способен взаимодействовать с фосфорилированными по тирозину протеинами /фосфорилированный EGF рецептор/, но он сохраняет способность взаимодействовать с областями, богатыми пролином, протеинов SOS. Следовательно, благодаря наличию делеции продукт гена Grb 3-3 способен препятствовать клеточным воздействиям продукта гена Grb 2. Перенос этого гена in vivo или его вариантов, включая антисмысловые последовательности, позволяет, следовательно, препятствовать процессам пролиферации, дифференциации и/или смерти клеток.
Итак, первый объект изобретения относится к нуклеотидной последовательности, включающей весь или часть гена Grb 3-3,последовательность SEQ ID N 1 (последовательности 1-9 приведены в конце описания).
Другой объект изобретения относится к нуклеотидной последовательности, происходящей из SEQ ID N 1 и способной ингибировать по крайней мере частично экспрессию протеина Grb 2 или Grb 3-3. В частности, изобретение относится к антисмысловым последовательностям, экспрессия которых в клетке-мишени позволяет контролировать транскрипцию клеточной мРНК. Такие последовательности могут быть, например, транскрибированы в клетке-мишени в РНК, комплементарные клеточные мРНК Grb 2 или Grb 3-3, и блокировать таким образом их трансляцию в протеин согласно методике, описанной в европейском патенте EP 140 308. Такие последовательности могут быть составлены полностью или частично из нуклеотидной последовательности SEQ ID N 1, транскрибированной в обратной ориентации.
Как указывалось выше, Grb 2 является по крайней мере бифункциональным протеином, имеющим в его области SH2 специфические последовательности, фосфорилированные по тирозину, и в двух областях SH3 факторы обмена семейства SOS. Grb 3-3, потерявший свою способность ассоциироваться с протеинами, фосфорилированными по тирозину, может, следовательно, образовать только комплекс с протеинами SOS. Следовательно, Grb 3-3 может препятствовать образованию комплекса Grb 2-SOS на рецепторах аутофосфорилированных факторов роста или на ассоциированных протеинах, также фосфорилированных по тирозину, таких как SHC или IRSI. способен блокировать это комплексообразование. Он способен блокировать митогенные пути и вызвать смерть клетки. Заявитель действительно показал, что протеин Grb 3-3 проявляет свое действие во время некоторых физиологических процессов, например при созревании тимуса у крыс. Заявитель также показал, что Grb 3-3 способен вызвать смерть клеток при апоптозе различных типов клеток. Эти очень ценные свойства могут быть доказаны /i/ путем инъекции рекомбинантного протеина в фибробласты ЗТЗ и /ii/ путем переноса последовательности, кодирующей Grb 3-3, в клетки ЗТЗ /пример 4/. Следовательно, Grb 3-3 способен вызывать клеточную смерть живых клеток, таких как иммортализованные /бессмертные/, раковые или зародышевые клетки. Как показано в примерах, Grb 2 способен противодействовать действиям Grb 3-3.
Кроме того, исследование экспрессии Grb 3-3, проведенное во время заражения лимфоцитарных клеток вирусом HIV, позволило показать, что массированное продуцирование вируса, наблюдаемое через 7 дней после заражения, коррелирует с гиперэкспрессией мРНК Grb 3-3 зараженными клетками /пример 5/. Этот эксперимент показывает, что удаление или противодействие клеточным эффектам Grb 3-3 также может позволить сохранить жизнь зараженным клеткам, а именно зараженным VIH, и также позволить лимфоцитам T4 продолжать выполнять роль иммунной защиты. В этом отношении изобретение также относится к применению соединений, способных устранить или препятствовать по крайней мере частично клеточным эффектам Grb 3-3, для приготовления фармацевтической композиции, предназначенной для лечения СПИДа. Более конкретно используемыми соединениями могут быть:
- генетические антисмысловые последовательности такие, как определено выше,
- специфические олигонуклеотидные Grb 3-3, модифицированные или нет для лучшей стабильности или биодоступности /фосфоротиоаты, вставки и т.п./. Предпочтительно речь идет об олигонуклеотидах, содержащих локализованную кодирующую последовательность между N-концевой SH3 областью и остаточной SH2 областью.
- любая последовательность, перенос которой в зараженные клетки вызывает гиперэкспрессию Grb 2.
Нуклеотидные последовательности согласно изобретению могут быть использованы как таковые, например, после инъекции человеку или животному, чтобы вызвать защиту или лечить раковые заболевания. В частности, они могут быть инъецированы в форме голой ДНК по методике, описанной в заявке WO 90/11092/. Они также могут быть введены в форме комплексов, например, с DEAE-декстраном /Pagano et al., J.Virol.I /1967/ 891/, с ядерными протеинами /Kaheda et al., Science 243 /1989/ 375/, с липидами /Felgner et al., PNAS 84 /1987/ 7413/, в форме липосом /Fraley et al., J.Biol.Chem.255 /1980/ 10431/ и т.д.
Предпочтительно нуклеотидные последовательности согласно изобретению являются частью вектора. Применение такого вектора позволяет в действительности улучшить введение нуклеиновой кислоты в обрабатываемую клетку, а также повысить ее стабильность в указанных клетках, что позволяет получить длительный терапевтический эффект. Кроме того, можно вводить несколько последовательностей нуклеиновой кислоты в одном и том же векторе, что также повышает эффективность лечения.
Используемый вектор может быть различного происхождения, лишь бы он был способен трансформировать клетки животных, предпочтительно человеческие опухолевые клетки. В предпочтительном варианте осуществления изобретения используют вирусный вектор, который может быть выбран среди аденовирусов, ретровирусов, аденоассоциированных вирусов /AAV/, вируса герпеса, цитомегаловируса /CMV/, вируса осповакцины и т.п. Векторы, происходящие из аденовирусов, ретровирусов или AAV, включающие гетерологические последовательности нуклеиновых кислот, были описаны в литературе [Akli et al., Nature Genetics 3 /1993/ 224; Stratford-Perricandet et al., Huwan Gene Therapy I /1990/ 241; EP 185573, Levrero et.al., Gene 101 /1991/ 195; Le Gal la Salla et al., Science 259 /1993/ 988; Roemer et Friedmann, Eur.J.Biochem.208 /1992/ 211; Dobson et al., Neuron 5 /1990/ 353; Chiocca et al., NewBiol. 2 /1990/ 739; Miyanohara et al., New Biol. 4 /1992/ 238; WO 91 /18088].
Следовательно, настоящее изобретение также относится к любому рекомбинантному вирусу, содержащему вставленную в его геном нуклеотидную последовательность, такую, как описано выше.
Целесообразно, чтобы рекомбинантный вирус согласно изобретению был дефектным вирусом. Термин "дефектный вирус" означает вирус, неспособный реплицироваться в клетке-мишени. Обычно, следовательно, геном дефектных вирусов, используемых в рамках настоящего изобретения, лишен по крайней мере последовательностей, необходимых для репликации указанного вируса в зараженной клетке. Эти области или могут быть удалены /полностью или частично/, или сделаны нефункциональными, или замещены другими последовательностями и, например, нуклеиновой кислотой изобретения. Предпочтительно, чтобы дефектный вирус тем не менее сохранил последовательности своего генома, которые необходимы для инкапсидации вирусных частиц.
Особенно выгодно использовать нуклеиновые последовательности изобретения в форме включения в аденовирус, в AAV или в рекомбинантный дефектный ретровирус.
Что касается аденовирусов, существуют различные серотипы, структура и свойства которых мало меняются, но которые не являются патогенными для человека, а именно не угнетающими иммунную систему. Кроме того, эти вирусы не интегрируются геномом клеток, которые они заражают, и могут включать значительные фрагменты экзогенной ДНК. Среди различных серотипов предпочтительно использовать в рамках настоящего изобретения аденовирусы типа 2 или 5 /Ad2 или Ad5/. В случае аденовируса Ad5 последовательности, необходимые для репликации, представляют собой области EIA или EIB.
Рекомбинантные дефектные вирусы изобретения могут быть получены путем гомологичной рекомбинации между дефектный вирусом и плазмидой, несущей наряду с другими нуклеотидную последовательность, такую, как определено выше /Levrero et al. , Gene 101 /1991/ 195; Graham, EMBO J. 3/12/ /1984/ 2917/. Гомологичная рекомбинация происходит после ko-трансфекции указанного вируса и плазмиды в соответствующую клеточную линию. Используемая клеточная линия предпочтительно должна быть /i/ трансформируемой указанными элементами и /ii/ сдержать последовательности, способные дополнять часть генома дефектного вируса, предпочтительно в интегрированной форме, чтобы избежать опасностей рекомбинации. В качестве примера линии, используемой для получения рекомбинантных дефектных аденовирусов, можно провести линию человеческой зародышевой почки 293/ Crahamet al., J. Gel.Virol. 36/1977/ 59/, которая содержит, например, интегрированную в ее геном левую часть генома аденовируса Ad5 /12%/. В качестве примера линии, используемой для получения рекомбинантных дефектных ретровирусов, можно привести линию CRIP /Danos et Mulligan, PNAS85 /1988/ 6460/.
Затем размножившиеся вирусы собирают и очищают по классическим методикам молекулярной биологии.
Объектом настоящего изобретения также является фармацевтическая композиция, содержащая по крайней мере один рекомбинантный вирус или одну нуклеотидную последовательность, такие, как определено выше.
Фармацевтические композиции изобретения могут быть сформулированы с учетом пути введения как для местного введения оральные, парентеральные, интраназальные, внутривенные, внутримышечные, подкожные, внутриглазные и т.п.
Предпочтительно фармацевтические композиции содержат фармацевтически приемлемые носители для рецептуры для инъекций, необязательно непосредственно в обрабатываемую опухоль. В частности, можно привести солевые растворы /мононатрийфосфат, динатрий фосфат, хлорид натрия, калия, кальция или магния, и т.п., или смеси этих солей) стерильные, изотонические, или сухие композиции, например лиофилизированные, которые при добавлении в зависимости от ситуации стерилизованной воды или физиологической сыворотки позволяют получить растворы для инъекций.
Дозы нуклеиновых кислот /последовательности или векторы/, используемые для введения, могут быть адаптированы в зависимости от различных параметров, например, в зависимости от используемого способа введения, конкретной патологии, действующей нуклеиновой кислоты, или еще от длительности желаемого лечения. Обычно, что касается рекомбинантных вирусов согласно изобретению, их формулируют и вводят в форме доз, содержащих между 104 и 1014 pfu/мл, предпочтительно 106 - 1010 pfu/мл. Термин pfu /"бляшкообразующая единица"/ соответствует заражающей мощности раствора вируса и определяется путем заражения соответствующей клеточной культуры и измеряется обычно через 48 часов по числу бляшек зараженных клеток. Методики определения титра pfu раствора вируса хорошо документированы в литературе.
Такие фармацевтические композиции могут быть использованы для людей для лечения и/или профилактики рака. В частности, продукты изобретения способны модулировать активность ras-протеинов, они позволяют включаться в процессы развития раков, а именно они могут ингибировать активность онкогенов, трансформирующая активность которых проходит через функциональное p21-GAP взаимодействие. Действительно, многочисленные раки ассоциируются с наличием онкогенных ras-протеинов. Среди раков, наиболее часто содержащих мутированные ras-гены, можно привести, например, аденокарциномы поджелудочной железы, 90% которых имеют мутированный Ki-ras-онкоген в двенадцатом кодоне /Almoguera et coll., Cell 53/ 1988/ 549/, аденокарциномы ободочной кишки и раки щитовидной железы /50%/ или карциномы легких и миелоидные лейкемии /30%/, Bos, J.L. Cancer Res. 49 /1989/ 4682/. Более обычно композиции согласно изобретению могут быть использованы для лечения любого типа патологий, в которых наблюдается аномальная пролиферация клеток, путем индукции апоптоза, а также любых патологий, характеризующихся смертью клеток при апоптозе /СПИД, хорея Хантингтона, Паркинсон/, с помощью соединений, блокирующих эффекты Grb 3-3 /например, антисмысловые/.
Настоящее изобретение будет более полно описано с помощью примеров, приведенных ниже, которые должны рассматриваться как иллюстрирующие, но не ограничивающие.
Подписи к фигурам:
Фиг. 1 : Схематическая иллюстрация структурных областей -Grb 2 и Grb 3-3.
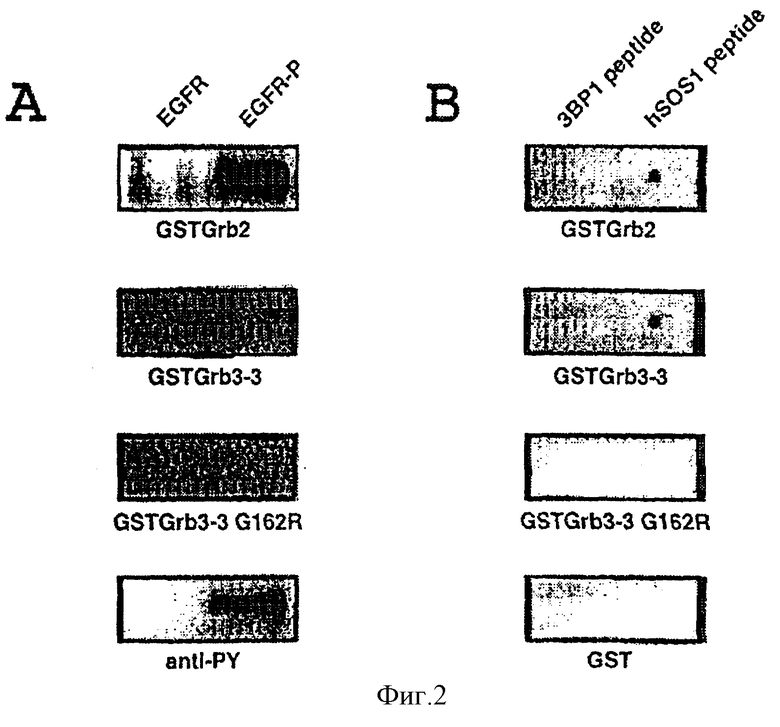
Фиг. 2: Изучение связи Grb 3-3 и рецептором EFG /фиг.2,a/ и с пептидами, богатыми пролином /фиг.2,в/.
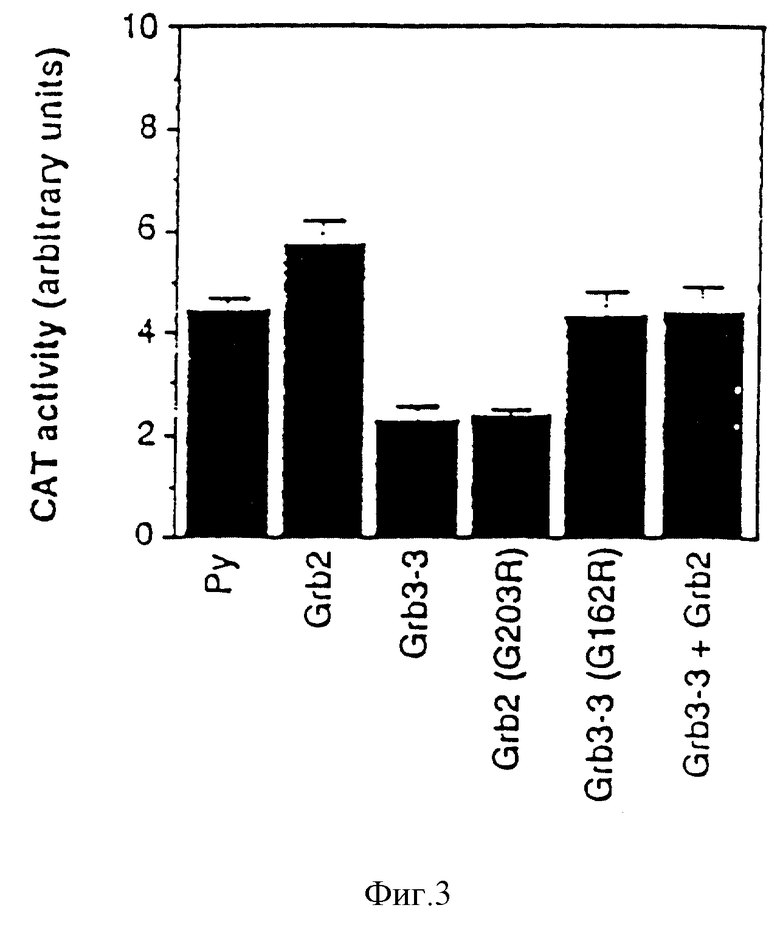
Фиг. 3: Действие Grb 3-3 на трансактивацию через ras-RRE- производное гена-усилителя полинома вируса.
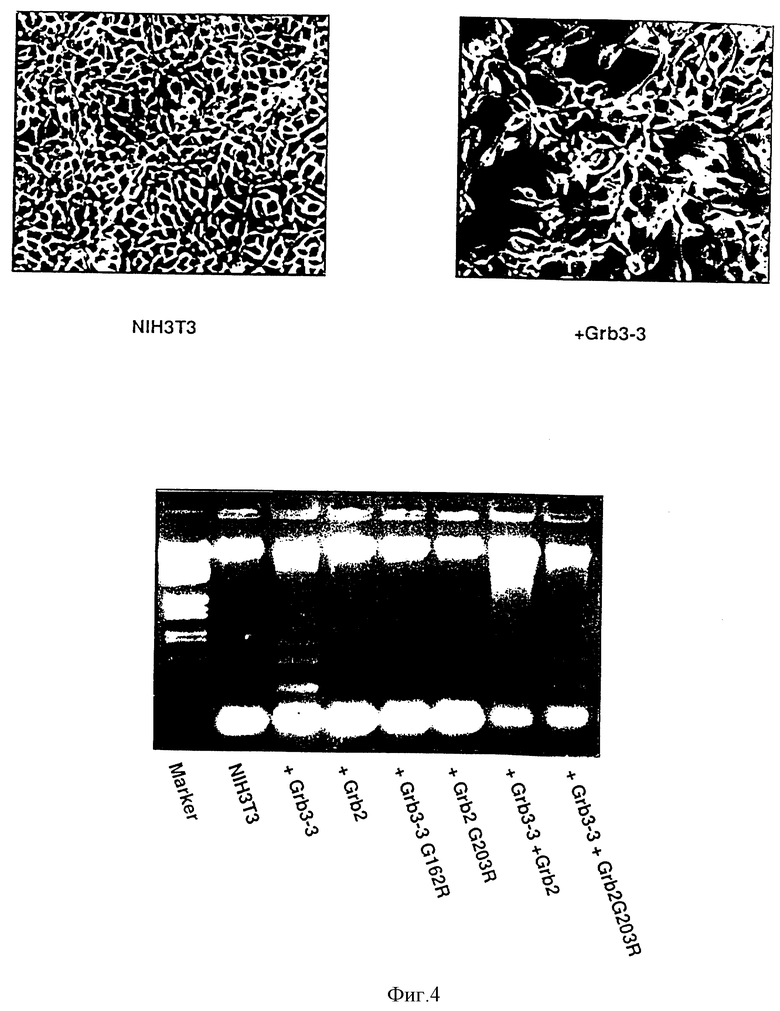
Фиг. 4: Выявление смерти клетки, вызванное Grb 3-3 на фибробластах ЗТЗ.
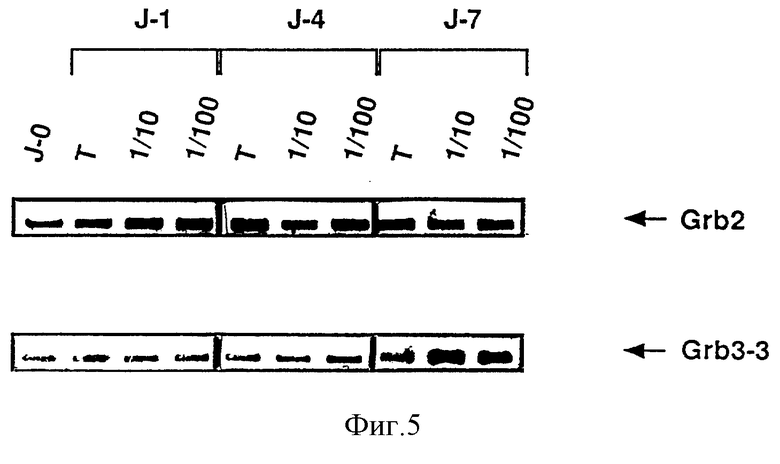
Фиг. 5: Выявление экспрессии Grb 3-3 в клетках, зараженных вирусом HIV.
Обычные методики молекулярной биологии.
Методики, обычно используемые в молекулярной биологии, такие как препаративные экстракции плазмидных ДНК, центрифугирование плазмидной ДНК в градиенте хлорида цезия, электрофорез в агарозном и акриламидном гелях, очистка фрагментов ДНК электроизоляцией, экстракция протеинов фенолом или фенолом-хлороформом, осаждение ДНК в солевой среде этанолом или изопропанолом, трансформация Esaherichia coli и т.п. хорошо известны специалистам в этой области и подробно описаны в литературе /Maniatis T. et al., "Molecular Cloning, а Laboratory Manual", Cold Spring Harbor Laboratory Cold Spring Harbor, N.Y. , 1982; Ausubel F.M. et al. /eds/, "Current Protocols in Molecular Biology", John Wiley and Sons, New York, 1987/.
Плазмиды типа pBR322, pLC и фаги серии M13 имеют коммерческое происхождение /Bethesda Research Laboratories/.
Для лигирования фрагменты ДНК могут быть разделены в соответствии с их размером электрофорезом на агарозном или акриламидном гелях, экстрагированы фенолом или смесью фенол/хлороформ, осаждены этанолом, потом инкубированы в присутствии ДНК лигазы фага T4 /Biolabs/ в соответствии с рекомендациями изготовителя.
Заполнение выступающих 5'- концов может быть проведено фрагментом ДНК полимеразы I Кленова Е. coli /Biolabs/ по описанию изготовителя. Деструкцию выступающих 3'- концов проводят в присутствии ДНК полимеразы фага T4 /Biolabs/, используемой в соответствии с рекомендациями изготовителя. Разрушение /деструкция/ выступающих 5'- концов проводят осторожной обработкой нуклеазой I.
Направленный мутагенез in vitro синтетическими олигонуклеотидами может быть проведен в соответствии с методикой, разработанной Taylor et al. /Nucleic Acids Res. 13 /1985/ 8749-8764/, используя набор, выпускаемый Amersham.
Ферментативная амплификация фрагментов ДНК по методике, называемой PCR /Полимеразная цепная реакция, Saiki R.K. et al., Science 230 /1985/ 1350-1354; Mullis K.B. et Faloona F.A. Meth.Ensym 155 /1987/ 335-350/, может быть проведена с использованием "ДНК термального циклера" /Perkin Elmer Cetus/ по описаниям изготовителя.
Проверка нуклеотидных последовательностей может быть проведена по методике, разработанной Sanger et al. /Proc. Natl.Acad. Sci.USA, 74/1977/ 5463-5467/ с использованием набора, выпускаемого Amersham.
Примеры
1. Выделение гена Crb 3-3
Ген выделяют скринингом полосы человеческой ДНК с помощью зонда, происходящего из последовательности гена Grb 2.
500000 рекомбинантных фагов Лямбда gt11, несущих фрагменты ДНК, выделенной из банка человеческой плаценты /Clontech/, были скринированы с помощью зонда, происходящего из последовательности гена Grb 2. Использованный зонд соответствует первым 8 аминокислотам протеина Grb 2 и имеет следующую последовательность:
ATGGAAGCCATCGCCAAATATGAC (SEQ ID N 2)
Таким образом было идентифицировано 10 положительных клонов. Вставка из 10 клонов была изолирована в виде фрагментов EcoRI, клонирована в плазмиде M13 p18 и секвенирована. Среди этих 10 клонов 9 имеет вставки, идентичные последовательности Grb 2. Один из них имеет вставку размером меньше, чем ген Grb 2, из-за делеции в области SH2 /фиг.1/. Анализ остальной последовательности показал полную идентичность с соответствующими областями Grb 2, включая некодирующие области 5' и 3'. Фаза открытого считывания этого клона кодирует протеин из 177 аминокислот /SEQ ID N 1/, содержащий 2 области SH3 по краям незавершенной SH2 области /фиг.1/. Аминокислоты, отсутствующие в области SH2 /остатки 60-100 в протеине Grb 2/, соответствуют остаткам, участвующим в связывании Grb 2 с пептидами, содержащими фосфорилированные тирозины.
2. Активность связывания протеина Grb 3-3
Как указывалось выше, протеин Grb 2 является медиатором взаимодействия межу рецепторами фосфорилированных факторов роста и факторами SOS. Это пример показывает, что протеин Grb 3-3 не способен взаимодействовать с фосфорилированным рецептором EGF, но он сохраняет свою способность взаимодействовать с пептидом, обогащенным пролином, происходящим из последовательности человеческого фактора SOSI.
Способность связывания Grb 3-3 была изучена с использованием протеинов слияния в Глутатион-S-Трансферазу /GST/, биотинилированных. Этот тип слияния позволяет быстро и эффективно очистить рекомбинантные продукты. Для этого последовательности изобретения были проявлены в штамме E.coli TGI в форме протеинов слияния с GST по методике, описанной Smith et Johnson /Gene 67 /1988/ 31/. Короче, гены Grb 2 и Grb 3-3сначала были модифицированы введением с обеих сторон стартового и стоп-кодонов сайта BamHI. Для этого фазы открытого считывания этих генов были амплифицированы PCR с помощью следующих олигонуклеотидов:
Олигонуклеотид I /5'/ /SEQ ID N 3/:
GAATTCGGATCCATGGAAGCCATCGCCAAATATGACTTC
Олгонуклеотид II /3'/ /SEQ ID N 4/:
GAATTCGGATCCTTAGACGTTCCGGTTCACGGGGGTGAC
Подчеркнутая часть соответствует созданному сайту BamHI, следующему за или предшествующему стартовому и стоп-кодонам.
Амплифицированные таким образом гены затем клонируют в форме фрагментов BamHI в вектор pGEX 2T /Pharmacia/, лианеризованный тем же самым ферментом, в 3' и в фазу кДНК, кодирующей GST. Полученные таким образом векторы затем используют для трансформации штамма E.coli TCI. Трансформированные таким образом клетки предварительно культивируют в течение ночи при 37oC, разбавляют 1/10 в среде LB, прибавляют IPTG, чтобы вызвать экспрессию /2 часа 25oC/, потом культивируют 21 час примерно при 25oC. Затем клетки лизируют и полученные слитые протеины очищают за счет сродства на колонке Агароза-GSH. Для этого бактериальный лизат инкубируют в присутствии геля /полученного и уравновешенного с буфером лизиса/ в течение 15 минут при 4oC. После 3 промывок Трис-HCl pH 7,4 протеины элюируют в присутствии буфера Трис-HCl pH 7,7 содержащего избыток GST. Супернатант собирают и центрифугируют.
Тот же протокол используют для получения мутанта Grb 2, в котором глицин 203 заменен аргинином /Grb 2G203R/, и мутант Grb3-3, в котором глицин 162 заменен аргинином /Grb 3-3G162R/. Мутант Grb 2G203R был описан как не имеющий активности в тесте на реинициацию синтеза ДНК /Lowenstein et al., выше/. Мутант Grb 3-3G162R несет ту же мутацию в том же положении и, следовательно, тоже должен быть неактивным.
Эти мутанты были получены мутагенезом с помощью PCR генов Grb 2 и Grb 3-3, используя в 5' олигонуклеотид 1, описанный выше, и в 3' олигонуклеотид III, приведенный ниже, в котором мутированный кодон подчеркнут:
Олигонуклеотид III /3'/ /SEQ ID N 5/:
GACGTTCCGGTTCACGGGGGTGACATAATTGCGGGGAAACATGCGGGTC
Амплифицированные таким образом фрагменты затем элюируют, реамплифицируют PCR с олигонуклеотидами I и II, потом клонируют в вектор pGEX 2T. Затем получают мутанты, как описано выше.
Затем протеины, слитые с GST /GST-Grb2, GST-Grb3-3, GST-Grb3-3,GI62R и GST/, биотинилируют по классическим методикам, известным специалистам (См. общие методики молекулярной биологии, а также Mayer et al., PNAS 88/ 1991/ 627/, и используют в качестве зондов для определения связывания с иммобилизованным фосфорилированным рецептором EGF /2.1/, потом с пептидом, происходящим из hSOSI /2.2/.
2.1. Связывание с фосфорилированным рецептором EGF
Протокол: используемый EGF рецептор очищают, исходя из клеток A431, путем иммобилизации на WGA-сефарозе по методике, описанной Duchesne et al. /Science 259 /1993/ 525/. Сначала стимулируют 2 мкг этого рецептора 1 мкМ EGF в течение 10 минут при 22oC, потом инкубируют с или без холодной АТР /10 мкМ/ в присутствии 2,5 мМ MnCl2 в буфере HNTC /20 мМ Hepes, 150 мM NaCl, 0,1% Тритон, 10% глицерина, pH 7,5/ при 4oC в течение 2 минут. Затем останавливают фосфорилирование рецептора добавлением буфера разложения. Потом образцы помещают на гель SDS-PAGE 4-20%, потом переносят на мембраны из поливинилидендифторида /PVDF/. Пятна затем инкубируют в присутствии различных слитых биотинилированных GST /2 мкг/мл/, потом проявляют с помощью стрептавадина, соединенного со щелочной фосфатазой /Promega/. Рецепторы EGF также подвергают иммуноблоттингу в присутствии антител анти-фосфотирозинов /анти-РУ/ для подтверждения, что рецепторы уже фосфорилированы.
Результаты: полученные результаты представлены на фиг.2,a. Они показывают, как и ожидалось, что протеин Grb 2 взаимодействует с рецептором EGF только в фосфорилированной форме. Затем они показывают, что протеин Grb 3-3 не связан с рецептором EGF, какой бы ни была его степень фосфорилирования.
2.2. Связывание пептида, производного от hSOSI
Протокол: синтезируют два пептида, обогащенных пролином:
Пептид hSOSI: GTPEVPVPPPVPPRRRPESA: Этот пептид соответствует остаткам 1143-1162 протеина hSOSI/Li et.al., Nature 363 /1993/ 83/, ответственного за взаимодействие между Grb 2 и hSOSI /SEQ ID N 6/.
Пептид 3BPI: PPPLPPLV: Этот пептид происходит из протеина 3BPI, который известен для эффективного связывания области SH3 Abl и Src /Cicchelli et al. , Science 257 /1992/ 803/ /SEQ ID N 7/.
Каждый из этих пептидов /1 Мкл, 10 мг/мл/ иммобилизуют на мембране из нитроцеллюлозы. Затем мембраны инкубируют в блокирующем буфере /Трис 20 мМ pH 7,6, 150 мМ NaCl, 0,1% Твин, 3% сывороточный альбумин/. Потом мембраны инкубируют всю ночь при 4oC в присутствии различных слитых биотинилированных GST /4 мкг/мл/, потом проявляют с помощью стрептавидина, соединенного со щелочной фосфатазой /Promeoa/.
Результаты: полученные результаты представлены на фиг.2,в. Они показывают, что Grb 3-3, как и Grb 2, способен связывать пептид hSOSI. Они кроме того показывают, что это взаимодействие является специфическим, потому что не наблюдается никакого связывания с пептидом 3BPI. Кроме того, результаты также показывают, что мутант Grb 3-3C162R не способен связывать пептид hSOSI, что подтверждает важность этого остатка и функциональную роль этого взаимодействия.
3. Активность протеина Grb 3-3
Этот пример показывает, что несмотря на его делецию в SH2 области протеин Grb 3-3 обладает функциональным эффектом.
Активность протеина Grb 3-3 была исследована путем определения его способности кооперироваться с ras для трансактивации промотора, обладающего ответом к ras (RRE), и управляющего экспрессией гена-репортера.
Использованный протокол описан, например, Schweighofter et al. Science 256 /1992/ 825. Короче, использованный промотор является синтетическим промотором, составленным из промотора гена тимидин каназы мыши и 4 элементов PEAL, повторяющихся, происходящих из усилителя полиомы /Wasylyk et al, EMBO J. 7 /1988/ 2475/: промотор Py-TK. Этот промотор управляет экспрессией гена-репортера, в случае бактериального гена хлорамфеникол ацетил трансферазы /CAT/: вектор Py-TK-CAT. Векторы экспрессии испытанных генов были сконструированы вставкой указанных генов в форме фрагментов BamHI в сайт BglII плазмиды pSV2. Этот сайт позволяет поместить гены под контроль раннего промотора SV40.
Клетки EP22 с 40% слияния были заражены 0,5 мкг вектора Py-TK-CAT, одного /Py/ или в присутствии вектора экспрессии, несущего под контролем раннего промотора SV40 ген: Grb 2, 2 мкг, Grb 3-3, 2 мкг, Grb 2/C203P/ 2 мкг, Grb 3-3/C162P/ 2 мкг, или Grb 3-32 мкг + Grb 2, 2 мкг. В каждом случае устанавливают суммарное количество ДНК 5 мкг с вектором экспрессии без вставки. Проводят заражение в присутствии липоспермина /Transfectam, IBF-Sepracor/. Клетки выдерживают 48 часов в культуре в среде ДМЕМ, дополненной 0,5% фетальной сыворотки теленка. Затем определяют активность CAT /трансактивацию PPE/, как описано Wasylyk et al., /PNAS85 /1988/ 7952/.
Полученные результаты представлены на фиг. 3. Они четко показывают, что экспрессия протеина Grb 3-3 препятствует эффектам активации рецептора фактора роста. Они также показывают, что избыток Grb 2 противодействует влияниям Grb 3-3 на ответ фактора роста.
4. Grb 3-3 вызывает апоптоз клеток.
Этот пример показывает прямое участие Grb 3-3 в клеточном апоптозе. Это свойство делает особенно целесообразными применения для лечения патологий, вызванных в результате пролиферации клеток /раки, рестеноз и т.п./.
Возникновение апоптоза клеток с помощью Grb 3-3 показано /i/ путем инъекции рекомбинантного протеина в фиброфласты ЗТЗ и /ii/ путем переноса последовательности, кодирующей Grb 3-3, в клетки ЗТЗ.
/i/ Инъекция рекомбинантного протеина
Рекомбинантный протеин Grb 3-3 был получен в форме слитого протеина с GST по протоколу, приведенному в примере 2. Слитый протеин затем обрабатывают тромбином /0,25%, Sigma/ для отделения части GST, потом очищают ионообменной хроматографией на колонке моноQ. Затем фракции, содержащие рекомбинантный протеин, концентрируют с помощью микроконцентраторов Microsep /Filtron/ в фосфатном буфере 20 мМ /pH 7/, содержащем 100 мМ NaCl. Полученный таким образом очищенный протеин инжектируют /1-3 мг/мл/ в культивируемые клетки ЗТЗ с помощью автоматического микроинжектора Эппендорфа. Затем клетки инкубируют при 34oC и фотографируют с регулярными интервалами, чтобы проследить морфологические превращения. Полученные результаты показывают, что через 5 часов после инъекции Grb 3-3 основная часть клеток умирает, тогда как инъекция в тех же условиях Grb 2 или мутанта Grb 3-3/C162R/ не оказывает никакого влияния на выживаемость клеток.
/ii/ Перенос последовательности, кодирующей рекомбинантный протеин.
Конструируют плазмиду, содержащую последовательность SEQ ID N 1, кодирующую протеин Grb 3-3 под контролем раннего промотора вируса SV40.
Фибропласты ЗТЗ с 40% слияния были заражены в присутствии липоспермина /Transfectam/, IBF-Sepracor/ 0,5 или 2 мг этой плазмиды экспрессии. Через 48 часов после заражения 50% этих клеток суспендируют в среде, а оставшиеся клетки, прилипшие к стенкам, имеют очень сильные морфологические изменения /фиг.4/. Анализ с помощью электрофореза на агарозном геле показал, кроме того, что клетки имеют конструкцию фрагментации олигонуклеосомальной ДНК, характерную для мертвых клеток /фиг.4/. Напротив, клетки, зараженные в тех же условиях плазмидой экспрессии Grb 2, Grb 3-3/C162R/, или Grb 2 /C203R/, сохраняют нормальную морфологию, являются всегда живыми и не имеют никакой фрагментации ДНК. Как показано на фиг. 4, соэкспрессия Grb 2 позволяет препятствовать действиям Grb 3-3.
Эти результаты четко показывают, следовательно, что Grb 3-3 представляют собой ген-убийцу, способный вызывать апоптоз клеток. Как указывалось выше, это свойство делает применения особенно выгодными для лечения патологий, возникающих в результате пролиферации клеток, таких как, например, раки, рестеноз и т.п.
5. Подтверждение экспрессии Grb 3-3 в лимфоцитах, зараженных вирусом HIV.
Этот пример показывает, что во время цикла заражения лимфоцитов T вирусом HIV относительное количество мРНК Grb 2 и Grb 3-3 модифицировано и что информационная РНК Grb 3-3 сверхпроявляется в момент обильной продукции вируса и смерти клеток.
Лимфоциты периферической крови заражают вирусом HIV-1 в двух разбавлениях /1/10 и 1/100/ в течение 1, 4 или 7 дней. Затем анализируют мРНК клеток путем обратной PCR с помощью олигонуклеотидов, специфических к Grb 2 и Grb 3-3, чтобы определить относительную пропорцию информационных мРНК Grb 2 и Grb 3-3. Использованные олигонуклеотиды, специфические к Grb3-3, являются следующими:
Олигонуклеотид IV /3'/:
ATGTTCCAAACGGATGTGGTTT (SEQ ID N 8)
Олигонуклеотид V /5'/:
ATAGAAATGAAACCACATCCGTTT (SEQ ID N 9)
Полученные результаты представлены на фиг. 5. Они четко показывают, что через 7 дней после заражения вирусом HI мРНК Grb 3-3 сверхпроявляется. Как показано определением протеина p24 и обратной транскриптазы вируса, день 7 также соответствует периоду, за который наблюдается обильная продукция вируса.




Изобретение относится к биотехнологии и может быть использовано для регуляции пролиферации клеток. Ген Grb 3-3 выделяют с помощью олигонуклеотидного зонда, происходящего из последовательности гена Grb 2. Ген Grb 3-3 представляет собой изоформу гена Grb 2, имеет делецию в области SH2 и способен вызывать апоптоз клеток. Антисмысловая последовательность к гену Grb 3-3 обладает способностью препятствовать вызываемым Grb 3-3 клеточным эффектом. Изобретение позволяет создавать фармацевтические композиции для лечения СПИДа. 3 с.п.ф-лы, 5 ил.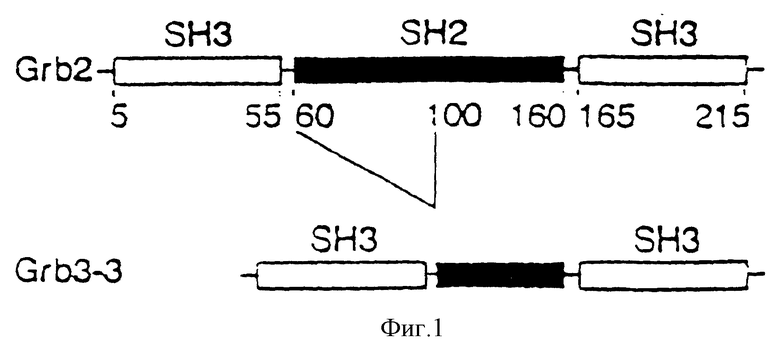
| Egan S.E | |||
| et al., Nature, 1993, v.363, p.45-51 | |||
| Lowenstein E.J | |||
| et al., Cell, 1992, v.70, p.431-442 | |||
| Duchesne M., et al., Science, 1993, v.259, p.525-528 | |||
| Suen K | |||
| Et al., Molekular and Cellular Biology, 1993, v.13, p.5500-5512. |

