Настоящее изобретение относится к нуклеиновым последовательностям, содержащим их векторам и их терапевтическим применениям, особенно в генной терапии. Более конкретно, настоящее изобретение относится к нуклеиновым последовательностям, которые включают ген, кодирующий внутриклеточный связывающий протеин (PIL), и их применению в генной терапии, в случае необходимости включенными в соответствующие векторы экспрессии.
Генная терапия состоит в исправлении дефекта или аномальности (мутация, аберрантная экспрессия и т. д. ) путем введения генетической информации в клетку или пораженный орган. Эта генетическая информация может быть введена либо ин витро в извлеченную из органа клетку, причем модифицированную клетку затем снова вводят в организм, либо непосредственно ин виво в соответствующую ткань. В этом отношении различные методы трансфекции и переноса генов описываются в литературе (см. Roemer и Friedman, Eur. J. Biochem., 208 (1992) 211). Вплоть до настоящего времени предлагаемые в уровне техники способы (подходы) генной терапии состоят в переносе генов, кодирующих активные полипептиды, которые принимают участие в генетических заболеваниях (гормоны, факторы роста и т.д.), антисмысловых генов или антигенных пептидов для реализации вакцин. Настоящее изобретение относится к новому способу (подходу) генной терапии, состоящему в переносе и экспрессии в целевой клетке (или ткани) внутриклеточного пептида, способного взаимодействовать с составляющими клеток и таким образом интерферировать с клеточными функциями. Настоящее изобретение преимущественно базируется на демонстрировании того, что можно экспрессировать ин виво модифицированные антитела, которые остаются во внутриклеточном пространстве и которые могут контролировать некоторые клеточные функции. Изобретение также базируется на демонстрировании того, что можно клонировать последовательности ДНК (дезоксирибонуклеиновой кислоты), кодирующие такие внутриклеточные антитела в векторах, особенно вирусных, для применения в генной терапии.
Использование антител в терапии человека, вообще, позволяет выявлять и нейтрализовать циркулирующие и/или локализованные биологические комплексы на поверхности клеток, вызывая серию явлений, управляемых иммунной системой, которая приводит к их устранению. Однако, во многих случаях, как раковые заболевания или заболевания, вызываемые, например, вирусами, этот способ бесполезен, так как антиген, ответственный за дерегуляцию ожидаемых клеток, недоступен вводимым антителам. В настоящем изобретении предлагается новый, особенно предпочтительный, терапевтический способ (подход), состоящий в непрерывном и внутриклеточном продуцировании антител или терапевтических агентов, связывание которых с их эпитопом уменьшает и/или аннулирует дерегуляцию.
Возможность экспрессировать рекомбинантным образом антитела уже описана в литературе. Так, в европейском патенте EР 99994 описывается экспрессия ин витро и очистка переменных участков тяжелых или легких цепей антител. Точно также, в патенте США 4946778 описывается экспрессия ин витро последовательностей ДНК, кодирующих модифицированные антитела, образованные переменными участками тяжелых и легких цепей антител, связываемых линкерами. Однако описанные в этом патенте антитела неактивны и обычно синтезируются в виде нерастворимых соединений(тел)-включений. Следовательно, антитела должны быть очищены, затем подвергнуты химическим обработкам (денатурация, ренатурация и т.д.) для восстановления активности. В настоящем изобретении демонстрируется возможность использования таких последовательностей ДНК для экспрессии прямо ин виво активных внутриклеточных антител. Таким образом, в настоящем изобретении демонстрируется возможность использования таких последовательностей ДНК, кодирующих внутриклеточные антитела, под контролем участков, позволяющих осуществлять их экспрессию в клетках млекопитающих, для генной терапии, особенно в случае человека. Этот новый способ, следовательно, позволяет подходить к составляющим клеток, недоступным при помощи классических методов вакцинации. Более того, этот способ не заключает в себе проявление иммунного ответа, а воздействует внутриклеточно.
Первый предмет изобретения, следовательно, заключается в последовательности нуклеиновых кислот, включающей ген, который кодирует внутриклеточный связывающий протеин (PIL) под контролем участков, позволяющих осуществлять его экспрессию в клетках млекопитающих.
Изобретение также относится к векторам, содержащим последовательность нуклеиновых кислот, такую, как описанная выше. Преимущественно векторы изобретения - вирусного происхождения, такие, как ретровирусы, аденовирусы, аденоассоциированные вирусы, вирусы герпеса, вирусы осповакцины, вирусы герпеса человека и т.д.
Изобретение относится также к применению этих последовательностей нуклеиновых кислот или этих векторов для приготовления фармацевтических композиций, предназначенных для хирургического и/или терапевтического лечения организма человека или животного. Оно также относится к любой фармацевтической композиции, включающей вектор, особенно вирусный вектор, или к последовательности нуклеиновых кислот, таких, как указанные выше.
В смысле настоящего изобретения термин "внутриклеточный связывающий протеин" (PIL) обозначает любой протеин или фрагмент протеина, способный распознавать составляющую клетки, в которой он экспрессирован, и взаимодействовать с ней селективным и родственным образом. Речь может идти о химических ковалентных или нековалентных взаимодействиях. Взаимодействие с клеточной составляющей (протеины, липиды, аминокислоты, иРНК, тРНК, рРНК, ДНК и т.д.) позволяет воздействовать на клеточную функцию, в которой содержится вышеуказанная составляющая, и таким образом контролировать (стимулировать, ингибировать, замедлять) эту функцию.
Предпочтительно PIL согласно изобретению образованы молекулами, происходящими от антител или обладающими связывающими свойствами, сравнимыми с таковыми антител. В особенности речь идет о протеинах, обладающих селективностью и сродством, достаточными для возможности протекания взаимодействия ин виво, оказывающим нейтрализующее воздействие на антиген. Эти молекулы в нижеследующем тексте обозначаются как внутриклеточные антитела в соответствии с их свойствами и их локализацией.
Антитела, молекулы надсемейства иммуноглобулинов, синтезируются естественно (главным образом лимфоцитами В) в форме секретированных (выделенных) протеинов. Они, следовательно, высвобождаются во внеклеточных пространствах (кровеносная система), где они проявляют свою активность (распознавание и связывание не самих антигенов). В настоящее время показано, что можно экспрессировать ин виво модифицированные гены, кодирующие внутриклеточные антитела, не затрагивая свойства специфичности и сродства антител. Последовательности нуклеиновых кислот согласно изобретению, которые кодируют внутриклеточные антитела, следовательно, включают ген антитела, модифицированный таким образом, что антитело не секретируется. В особенности ген антитела обычно модифицируют путем делеции или мутации последовательностей, ответственных за его секрецию. PIL согласно изобретению особенно могут быть образованы фрагментами антител, например, фрагментами Fab или F(ab)'2, которые содержат домены связи антигена. Использование этого типа внутриклеточного антитела, однако, заключает экспрессию последовательности нуклеиновых кислот, включающей несколько генов, кодирующих соответственно тяжелые и легкие участки этих фрагментов, и также заключает то, что эти цепи точно соединяются ин виво. На этом основании особенно предпочтительная форма внутриклеточных антител, используемых в рамках изобретения, образована пептидом, соответствующим сайту связывания переменного участка легкой цепи антитела, связанным за счет пептидного линкера с пептидом, соответствующим сайту связывания вариабельного участка тяжелой цепи антитела. Использование этого типа внутриклеточного антитела, обозначаемого ScFv, представляет интерес, так как эти антитела экспрессируются единственным геном. Конструкция последовательностей нуклеиновых кислот, кодирующих такие, модифицированные согласно изобретению антитела, иллюстрируется в примерах.
Кроме того, последовательности нуклеиновых кислот, кодирующие внутриклеточные антитела согласно изобретению, также могут быть модифицированы химическим путем, ферментативным путем или генным путем с целью генерирования внутриклеточных стабилизированных антител и/или внутриклеточных многофункциональных антител, и/или уменьшенного размера, и/или для того, чтобы благоприятствовать их локализации в таком внутриклеточном пространстве. Так, последовательности нуклеиновых кислот изобретения могут включать последовательности, кодирующие пептиды ядерной локализации (NLS). В особенности, можно подвергать слиянию последовательности изобретения с последовательностью, кодирующей NLS вируса SV40, пептидная последовательность которого следующая: МРКККRК (Kalderon и др., Cell, 39 (1984) 499).
Как указано выше, последовательности нуклеиновых кислот согласно изобретению включают последовательности, позволяющие осуществлять экспрессию гена или генов, кодирующих PIL, в клетках млекопитающих. Обычно гены PIL, следовательно, находятся под контролем промоторных участков транскрипции и трансляции, функциональных в клетке млекопитающего, в которой достигнута экспрессия. Речь может идти о гомологичных по отношению к вышеуказанной клетке последовательностях, т.е. о последовательностях, естественно ответственных за экспрессию генов в вышеуказанной клетке. Речь также может идти о последовательностях различного происхождения, т.е. последовательностях, ответственных за экспрессию протеинов в других типах клеток; последовательностях, ответственных за экспрессию антител в природных условиях; вирусных последовательностях экспрессии, например, присутствующих в векторе, в который включены последовательности согласно изобретению, или еще синтетических или полусинтетических последовательностях.
В случае применения для человека в литературе описаны многочисленные функциональные промоторы, такие, как, например, вирусные промоторы CMV, SV40, Ela, MLP, LTR и т.д. Клеточные промоторы, такие, как, например, промотор гена виллина, представляют интерес, так как позволяют осуществлять специфическую тканевую экспрессию (ограниченную в кишечнике в случае виллина).
Кроме того, последовательности экспрессии также могут быть модифицированы, например, для их адаптации к экспрессии в особый тип вектора или клетки, для уменьшения их размера, для повышения активности их промотора транскрипции, для генерирования индуцируемых промоторов, улучшения их уровня регуляции или еще для изменения природы их регуляции. Такие модификации можно осуществлять, например, путем мутагенеза ин витро, путем введения дополнительных элементов контроля или синтетических последовательностей, или путем делеций или замещений оригинальных элементов контроля. Особенно предпочтительным может быть использование специфических тканевых промоторов, чтобы осуществлять экспрессию PIL только в один тип ткани.
Кроме того, когда последовательность нуклеиновых кислот не содержит последовательности экспрессии, то она может быть включена в вектор экспрессии, ниже такой последовательности.
Для получения вектора согласно изобретению, во-первых, следует идентифицировать клеточную функцию, например, принимающую участие или ответственную за патологию, на которую хотят воздействовать. Затем следует идентифицировать соответствующую клеточную составляющую, принимающую участие в этой функции, после чего нужно определить, какой подобный PIL наиболее адаптирован к этой составляющей (антитела, производные и т.д.), в зависимости от его локализации, его роли, его природы и т.д. Выбрав PIL, соответствующая последовательность нуклеиновых кислот может быть получена методами молекулярной биологии (химический синтез, клонирование, ферментативная модификация и т.д. ) и вставлена в соответствующий вектор согласно описанной в примерах методологии.
Другой предмет изобретения относится к фармацевтическим композициям, содержащим по крайней мере одну последовательность нуклеиновых кислот или вектор, такие, как описанные выше.
Последовательности согласно изобретению могут быть использованы такими, какие есть, например, после введения человеку или животному, для индуцирования внутриклеточной экспрессии PIL с целью затрагивания определенной клеточной функции. В особенности они могут быть введены в форме "голой" ДНК согласно способу, описанному в заявке ВОИС 90/11092. Их также можно вводить в закомплексованной форме, например, с DEAE-декстраном (Раgаnо и др., J. Virol. , 1 (1967), 891), с ядерными протеинами (Kaneda и др., Science, 243 (1989) 375), с липидами (Falgner и др., PNAS 84 (1987) 7413), в форме липосом (Praley и др., J. Biol. Chem. 255 (1980) 10431) и т.д.
В предпочтительном варианте реализации изобретения нуклеиновые последовательности, такие, как указанные выше, включают в вектор. Применение таких векторов позволяет на деле благоприятствовать пенетрации в клетки, повышать резистентность к ферментам и увеличивать стабильность и внутриклеточные уровни экспрессии. Векторы согласно изобретению могут быть как плазмидными, так и вирусными. Однако предпочитают использовать вирусный вектор.
В предпочтительном варианте реализации, следовательно, изобретение относится к последовательностям нуклеиновых кислот, таким, как указанные выше, включенным в вирусный вектор. Изобретение также относится к любому рекомбинантному вирусу, включающему встроенную в его геном по крайней мере одну последoвательность нуклеиновых кислот, кодирующую PIL.
Как указано выше, различные вирусы могут быть использованы в качестве векторов для переноса и экспрессии ин витро генов согласно изобретению. В качестве примера можно назвать ретровирусы (RSV, HMS, MMS и т.д.), вирус герпеса человека, аденоассоциированные вирусы, аденовирусы, вирус осповакцины и т.д.
Предпочтительно рекомбинантным вирусом согласно изобретению является дефектный вирус. Термин "дефектный вирус" обозначает вирус, неспособный реплицироваться в целевой клетке. Обычно геном дефектных вирусов, используемых в рамках настоящего изобретения, следовательно, лишен по крайней мере последовательностей, необходимых для репликации вышеуказанного вируса в зараженной клетке. Эти участки могут быть либо удалены (полностью или частично), либо сделаны нефункциональными, либо заменены другими последовательностями и особенно нуклеиновыми последовательностями изобретения. Предпочтительно дефектный вирус тем не менее сохраняет последовательности своего генома, которые необходимы для инкапсуляции вирусных частиц.
Дефектные рекомбинантные вирусы, производные ретровирусов, аденоассоциированных вирусов, вируса человеческого гepпеса и адоновирусов, уже описаны в литературе (Roemer и Friedmann, Eur. J. Biochem. 208 (1992) 211; Dobson и др. , Neuron, 5 (1990) 353; Chiocca и др., New. Biol. 2 (1990) 739; Miyano-hara и др.. New Biol. 4 (1992) 238; ВОИС 91/18088; Akli и др., Nature Genetics 3 (1993) 224; Stratford-Perricauder и др., Human Gene Therapy, 1 (1990) 241; европейский патент ЕР 185573; Levrero и др., Gene, 101 (1991) 195; европейский патент ЕР 243204).
Особенно предпочтительно использовать нуклеиновые последовательности изобретения в форме, включенной в рекомбинантный дефектный аденовирус.
В действительности существуют различные серотипы аденовирусов, структура и свойства которых изменяются немного, но которые непатогенны для человека, и особенно иммунонеугнетенных субъектов. Кроме того, эти вирусы не интегрируются в геном клеток, которые они инфицируют, и могут включать значительные фрагменты экзогенной ДНК. Из различных серотипов предпочитают использовать в рамках настоящего изобретения аденовирусы типа 2 или 5 (Ad 2 или Ad 5). В случае аденовирусов Ad5 необходимые для репликации последовательности представляют собой участки Е1А и Е1B.
Кроме того, незначительный размер генов, кодирующих внутриклеточные антитела согласно изобретению, позволяет предпочтительным образом включать одновременно, в один и тот же вектор, несколько генов, кодирующих внутриклеточные антитела, направленные против различных участков одной или нескольких клеточных целевых составляющих. Особый вариант реализации изобретения, следовательно, состоит в векторе, особенно вирусном, включающем по крайней мере две последовательности нуклеиновых кислот, кодирующих внутриклеточные связывающие протеины, направленные против различных эпитопов.
Дефектные рекомбинантные вирусы согласно изобретению могут быть получены путем гомологичной рекомбинации между дефектным вирусом и плазмидой, несущей, между прочим, последовательность нуклеиновых кислот, такую, как указанная выше (Levrero и др., Gene, 101 (1991) 195; Graham., EMBO J. 3 (12) (1984) 2917). Гомологичная рекомбинация производится после котрансфекции вышеуказанными вирусом и плазмидой соответствующей линии клеток. Используемая линия клеток предпочтительно должна быть (I) трансформируемая вышеуказанными элементами и (II) содержать последовательности, способные дополнять часть генома дефектного вируса, предпочтительно в интегрированной форме, для того чтобы избежать опасностей рекомбинации. В качестве примера линии, используемой для получения дефектных рекомбинантных аденовирусов, можно указать линию почки эмбриона человека 293 (Graham и др., J. Gen. Virol. 36 (1977) 59), которая содержит особенно, интегрированной в ее геном, левую часть генома аденовируса Ad5 (12%). В качестве примера линии, используемой для получения дефектных рекомбинантных ретровирусов, можно указать линию CRIP(Danos и Mulligan, PNAS, 85 (1988) 6460).
Затем вирусы, которые размножились, извлекают и очищают согласно классическим способам молекулярной биологии.
Настоящее изобретение, следовательно, также относится к фармацевтической композиции, включающей по крайней мере один дефектный рекомбинантный вирус, такой, как указанный выше.
Фармацевтические композиции согласно изобретению могут быть сформулированы для введения топически, орально, парентерально, через нос, внутривенно, внутримышечно, подкожно, внутриглазным путем и т.д.
Предпочтительно фармацевтические композиции содержат фармацевтически приемлемые эксципиенты для вводимой путем инъекции формулировки. Речь может идти в особенности о солевых (мононатрийфосфат, динатрийфосфат, хлорид натрия, хлорид калия, хлорид кальция или хлорид магния и т.д. или смеси таких солей), стерильных, изотонических растворах или о сухих композициях, особенно лиофилизированных, которые путем добавления, в зависимости от случая, стерилизованной воды или физиологической сыворотки позволяют получать растворы для инъекций.
Используемые для введения дозы нуклеиновых кислот (последовательность или вектор) могут быть адаптированы в зависимости от различных параметров, и особенно в зависимости от используемого способа введения, соответствующей патологии, экспрессируемого гена или еще от продолжительности искомого лечения. Обычно, что касается рекомбинантных вирусов согласно изобретению, их формулируют и вводят в виде доз 104-1014 pfu/мл и предпочтительно 106-1010 pfu/мл. Термин "pfu" ("бляшкообразующая единица") соответствует инфекционной способности раствора вируса и определяется путем инфекции соответствующей культуры клеток и измеряется, обычно спустя 48 часов, числом бляшек инфицированных клеток. Способы определения титра pfu вирусного раствора хорошо освещены в литературе.
Предметом изобретения также является любая рекомбинантная клетка, содержащая последовательность нуклеиновых кислот, такую, как определенная выше.
Последовательности согласно изобретению, в случае необходимости включенные в векторы, и содержащие их фармацевтические композиции могут быть использованы для лечения многочисленных патологий. Так, они могут быть использованы для переноса и экспрессии генов ин виво в любой тип ткани. Лечение, кроме того, может быть целенаправленным в зависимости от излечиваемой патологии (перенос на уровне индивидуальной ткани особенно может быть определен выбором вектора, а экспрессия - выбором особого промотора). Последовательности или векторы согласно изобретению предпочтительно используют для продуцирования у человека или животного, ин виво и внутриклеточно, протеинов, способных специфически воздействовать на различные клеточные функции, такие, как клеточная пролиферация, синтез метаболитов, синтез протеинов, репликация и/или транскрипция ДНК и т.д. Таким образом, настоящее изобретение позволяет специфически, локально и эффективно лечить многочисленные дисфункции клеток (естественного) происхождения или возникающие в результате различных патологий, и в особенности раковые заболевания, вирусные или бактериальные заболевания, или, вообще, любую патологию, в которой может быть идентифицирован клеточный медиатор.
Применение для лечения патологий, связанных с пролиферацией клеток
В особенно предпочтительном варианте реализации последовательности нуклеиновых кислот согласно изобретению включают гены, кодирующие PIL, способные взаимодействовать и интерферировать с активностью факторов, принимающих участие в пролиферации клеток. Пролиферация клеток возбуждается множеством факторов, таких, как мембранные рецепторы (протеины G), онкогены, ферменты (протеины киназ, фарнезилтрансфераз, фосфолипаз и т.д.), нуклеозиды (АТР, АМР, GDP, GTP и т.д.), факторы активации [факторы обмена гуанозинов (GRF, GAP и т.д.), транскрипционные факторы и т.д.] и т.д. Внутриклеточная экспрессия PIL согласно изобретению, способных связывать и нейтрализовать такие факторы, позволяет контролировать процесс пролиферации клеток. Это представляет интерес в особенности в той ситуации, где пролиферация клеток не поддается естественным механизмам регуляции, приводя, например, к появлению опухолей. Многочисленные факторы (продукты онкогенных генов и факторы, принимающие участие в сигнализации эффекта этих продуктов) на деле ассоциируются с этими явлениями дерегуляции пролиферации клеток. Так, 90% аденокарцином поджелудочной железы содержат мутированный онкоген Ki-ras на двенадцатом кодоне (Almoguera и сотр., Cell, 53 (1988) 549). Точно также, наличие мутированного rаs-гена обращает на себя внимание в аденокарциномах ободочной кишки и раковых заболеваниях щитовидной железы (50%) или в карциномах легких и миелоидных лейкемиях (30%; Bos J.L., Cancer Res., 49, (1989) 4682). На сегодняшний день идентифицированы другие многочисленные онкогены (mус, fos, jun, ras, myb, erb и т.д.), мутированные формы которых представляются ответственными эа де~ регуляцию пролиферации клеток.
Экспрессия PIL, способных связывать эти клеточные факторы (предпочтительно их онкогенную форму) и, следовательно, замедлять или ингибировать их воздействия, дает возможность нового терапевтического подхода к раковым заболеваниям.
В особенно представляющем интерес варианте настоящее изобретение относится к векторам, содержащим последовательности нуклеиновых кислот, включающих ген, который кодирует внутриклеточное антитело, способное взаимодействовать с продуктом экспрессии онкогена или с фактором, принимающим участие в пути сигнализации онкогена.
Из целевых онкогенов в смысле изобретения можно назвать онкогены ras, fos, jun, myc, myb и erb.
Из факторов, принимающих участие в пути сигнализации онкогена, особенно можно назвать мембранные рецепторы, которые могут быть выявлены на уровне их внутриклеточных доменов [протеины G, киназы (пример, тирозинкиназа), фосфорилазы, фарнезилтрансферазы], факторы обмена нуклеозидов (факторы GAP, GRF и т.д.) и т.д.
Преимущественно настоящее изобретение относится к векторам, особенно вирусного происхождения, содержащим последовательность нуклеиновых кислот, кодирующую внутриклеточные антитела, направленные против онкогена или фактора, принимающего участие в пути сигнализации онкогена, образованную пептидом, соответствующим сайту связывания вариабельного участка легкой цепи антитела, связанным за счет пептидного линкера с пептидом, соответствующим сайту связывания вариабельного участка тяжелой цепи антитела.
Преимущественно изобретение относится к дефектным рекомбинантным вирусам, экспрессирующим внутриклеточное антитело, направленное против фактора пути ras-зависимой сигнализации.
Как это показывают примеры, экспрессия внутриклеточных антител anti-p21 (продукт экспрессии ras-гена), анти-GAP или anti-p53 позволяет ревертировать трансформирующий раковую клетку фенотип.
Применение для лечения вирусных патологий
Нуклеиновые последовательности согласно изобретению могут еще представлять собой последовательности, кодирующие внутриклеточные антитела, способные взаимодействовать с патогенным вирусом (ВИЧ, вирус папилломы и т. д.) в инфекционном цикле.
Преимущественно, в случае вируса ВИЧ, антивирусные агенты, имеющиеся в распоряжении на сегодняшний день или выдвигаемые на передний план в протоколах клинических испытаний, не позволяют блокировать вирус в его размножении, но в меру задерживают прогрессирование заболевания. Один из основных доводов в этом отношении заключается в появлении штаммов, резистентных к этим антивирусным агентам. Расширение эффективной вакцинации также наталкивается на многочисленные препятствия: генетическая изменяемость ВИЧ не позволяет точно определять антигенную стабильную структуру для иммунологического, гуморального или целлюлярного ответа против различных существующих штаммов.
Как в случае антивирусных агентов, где вирус резистентен к воздействию селекции за счет смещения точечных мутаций, в опытах по вакцинации, проводимых до настоящего времени, ВИЧ, по-видимому, не поддается иммунной (immunitaire) системе.
Настоящее изобретение состоит в новом подходе к лечению инфекции ВИЧ, заключающемся в блокирования вируса во время его репликативного цикла путем экспрессии PIL, и особенно внутриклеточных антител. Настоящее изобретение в особенности позволяет избавиться от проблемы генетической изменяемости вируса в противоположность антивирусным и при использовании вакцин способам. В случае классической вакцинации проявляющиеся иммунные ответы в принципе возникают против изменяемых участков. Напротив, использование внутриклеточных антител согласно изобретению позволяет выбрать эпитоп, не только сохранившийся, но и также необходимый, с функцией вирусного протеина. Предпочтительно внутриклеточное антитело направлено против эпктопа достаточного размера для препятствования резистентности и адаптируемости вируса за счет смещения точечных мутаций. Кроме того, незначительная величина кодирующего внутриклеточное антитело согласно изобретению гена позволяет предпочтительным образом иметь несколько участков (одного или нескольких протеинов) из одного и того же вектора генной терапии.
Два основных протеина регуляции вируса, tat и rev, представляют собой цели первой важности для осуществления настоящего изобретения. В самом деле, эти два протеина необходимы для вирусной репликации. Более того, экспрессия антисмысловых информационных рибонуклеиновых кислот или рибозимов (ribozymes), нацеленная на информационную РНК tat или rev, противодействует вирусной репликации. Механизм воздействия этих протеинов относительно хорошо подтвержден документальными данными: "tat" представляет собой фактор трансактивации транскрипции, тогда как "rev" обеспечивает транзицию (переход) между ранней и поздней фазами репликативного цикла. Эти два протеина подвергают испытанию на их активность путем фиксации на вирусных информационных РНК, и относительно хорошо устанавливают пределы необходимого для этой фиксации пептидного участка, необходимого для их функции. Кроме того, показано, что суперэкспрессия их места фиксации на РНК (участок ТАR для "tat" и RRE для "rev") ингибирует их функцию в вирусной экспрессии.
В этих условиях особенно предпочтительный вариант изобретения состоит в использовании последовательности ДНК, кодирующей внутриклеточное антитело, направленное против протеинов "tat" и "rev" и способной нейтрализовать их активность. Предпочтительно такие антитела направлены против участка "tat" или "rev", ответственного за их фиксацию на РНК.
Внутриклеточные антитела согласно изобретению, направленные против этих эпитопов, могут быть получены согласно описанной в примерах методологии. В особенности, что касается антитела anti-tat, то оно может быть получено путем генетической модификации из моноклонального антитела Т-В(12) (Hybridolab).
Другой целевой протеин, важный в репликации ВИЧ, функция которого легко может быть ингибирована внутриклеточным антителом, представляет собой протеин нуклеокапсида NCP7. В самом деле, этот протеин играет важную роль в ретротранскрипции (ранняя фаза), в интеграции и поздней, но тем не менее важной фазе: инксапсидации. Эта многофункциональность выражается в том, что он выполняет структурную, ферментативно активную роль. Он описывается как гибридаза, которая позволяет осуществлять комплексацию вирусных нуклеиновых кислот: РНК/ДНК и ДНК/ДНК во время ретротранскрипции, также как РНК/РНК и РНК/Lys3-тРНК на уровне инкапсидации. NCP7 является необходимым для продуцирования инфекционных вирусов.
Цитоплазматическая экспрессия путем генной терапии внутриклеточных антител, направленных против NCP7, ингибирующих его функцию в димеризации вирусных РНК и в инкапсидации, составляет другой особый вариант осуществления изобретения.
Внутриклеточные антитела согласно изобретению, направленные против нейтрализующих эпитопов этого протеина, могут быть получены согласно описанной в примерах методологии.
Другие вирусные составляющие также могут составлять объект генной терапии согласно изобретению, а именно место фиксации молекулы CD4 (пример: гликопротеин оболочки (gр120/41), участок мультимеризации оболочки, место расщепления gр120/gр41, протеаза, интеграза и вообще любой вирусный протеин. Эти домены обычно недоступны, когда оболочка находится в естественной (нативной) форме. В связи с этим они очень мало иммуногенны и инициирующая их вакцинация невозможна. Настоящее изобретение позволяет выявить как цель эти вирусные составляющие и таким образом обладает очень высоким терапевтическим потенциалом. Кроме того, как указано выше, небольшая величина генов, кодирующих внутриклеточное антитело, согласно изобретению позволяет предпочтительным образом экспрессировать одновременно, в тот же самый вектор, несколько внутриклеточных антител, направленных против различных участков одной или нескольких из этих составляющих.
Нуклеиновые последовательности согласно изобретению также могут представлять собой последовательности, кодирующие PIL, которые способны взаимодействовать и интерферировать с активностью факторов, принимающих участие в синтезе метаболитов, в синтезе протеинов или еще в репликации и/или транскрипции ДНК.
Настоящее изобретение более полно описывается с помощью нижеследующих примеров, которые нужно рассматривать как иллюстративные и не ограничивающие объема охраны изобретения.
Подписи под чертежом.
(A): карта рестрикции ScFv-ras.
(B): карта рестрикции ScFv-Gap.
(C): нейтрализующее воздействие транзиторной экспрессии внутриклеточного антитела изобретения на трансформирующую способность онкогена "ras" или "Нег2".
Ру - контрольные клетки;
ScFv - трансфектированные одной плазмидой psv2.ScFv. ras клетки;
VAL - клетки, трансфектированные вектором, экспрессирующим онкогенный ras, Ha-ras Val12;
VAL+ScFv - клетки, котрансфектированные плазмидой psv2. ScFv. ras и Ha-ras Val12;
NER2 - клетки, трансфектированные экспрессирующим онкогенный Неr2 вектором;
NER2+ScFv - клетки, котрансфектированные плазмидой psv2. ScFv. ras и Неr2.
Общие методы молекулярной биологии
Классически используемые в молекулярной биологии методы, такие, как препаративные экстракции плазмидной ДНК, центрифугирование плазмидной ДНК по градиенту хлорида цезия, электрофорез на агарозогелях или акриламидогелях, очистка фрагментов ДНК путем электроэлюирования, экстракция протеинов фенолом или смесью фенола с хлороформом, преципитация (осаждение) ДНК в солевой среде этанолом или изопропанолом, трансформация в Escherichia coli и т.д., хорошо известны специалисту и широко описаны в литературе [Maniatis Т. и др. , "Molecular Cloning, a Laboratory Manual", Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1982; Ausubel F.M. и др. (eds.) "Current Protocols in Molecular Biology", John Wiley and Sons. New York, 1987].
Плазмиды типа pBR322, pUC и фаги серии М13 имеются в продаже (Bethesda Research Laboratories).
Для лигатур фрагменты ДНК могут быть разделены по их величине путем электрофореза в гелях агарозы или акриламида, проэкстрагированы фенолом или смесью фенола с хлороформом, осаждены в этаноле, затем инкубированы в присутствии ДНК-ли-газы фага Т4 (Biolabs) согласно рекомендациям поставщика.
Заполнение выступающих 5'-концов может быть осуществлено с помощью фрагмента Кленова ДНК-полимеразы I Е. coli (Biolabs) согласно инструкциям изготовителя. Деструкцию выступающих 3'-концов осуществляют в присутствии ДНК-полимеразы фага Т4 (Biolabs), используемой согласно рекомендациям изготовителя, Деструкцию выступающих 5'-концов осуществляют путем осторожной обработки с помощью нуклеазы S1.
Направленный мутагенез ин витро с помощью синтетических олигодезоксинуклеотидов может быть осуществлен по методу, разработанному Taylor и др. [Nucleic Acids Res. , 13 (1985) 8749 - 8764], используя набор, выпускаемый фирмой Amersham.
Ферментативная амплификация фрагментов ДНК по так называемому способу PCR [Polymerase - catalyzed Chain Reaction (катализируемая полимеразой цепная реакция), Saiki R.K. и др., Science, 230 (1985) 1350 - 1354; Mullis К.В. и Faloona Р.A., Meth. Enzym., 155 (1987) 335 - 350] может быть осуществлена при использовании "концевой циклической ДНК" (Perkin Elmer Cetus) согласно указаниям изготовителя.
Проверка (контроль) нуклеотидяых последовательностей может быть осуществлена по методу, разработанному Sanger и др. [Proc. Natl. Acad. Sci., USA, 74 (1977) 5463 - 5467], используя набор, выпускаемый фирмой Amersham.
ПРИМЕР 1. Клонирование и экспрессия последовательности ДНК, клонирующей внутриклеточное антитело anti-ras.
Этот пример описывает клонирование и экспрессию последовательности нуклеиновых кислот, кодирующей внутриклеточный связывающий протеин, воспроизводящий свойства моноклонального антитела Y13-259. Антитело Y13-259 направлено против протеинов Ras (ATCC CRL 1742) (J. Virol. 43, 294 - 304, 1982) и представляет собой антитело, нейтрализующее функцию онкогенных протеинов Ras, сразу инъецируемое в клетки [Smith и сотр., (1986), Nature, 320, 540 - 543; Кung и сотр., Ехр. Cell Res. (1986), 162, 363 - 371].
1.1. Получение последовательности ДНК.
ДНК-последовательность, кодирующую внутриклеточное антитело (фрагмент ScFv), получают по способу, описанному в патенте США 4946778. Эту последовательность затем, под контролем функционального промотора, вводят в клетки млекопитающих.
PHK-poly-A изолируют из культуры клеток гибридомы, которая выделяет антитело Y13-259, согласно способу, описанному Chirguin S.H. и др. [Biochemistry, 18, 5294 (1979)]. Эти РНК используют для реакции реверсия - транскрипция с помощью праймеров, образованных случайными гексануклеотидами. Полученные кДНК служат матрицей двух реакций PCR:
- одна, предназначенная для амплификации переменного фрагмента тяжелой цепи (VH) Y13-259 с помощью специфических праймеров мышиных VH;
- вторая, позволяющая получать фрагмент VL, используя смесь 10 праймеров, производимых от мышиных последовательностей.
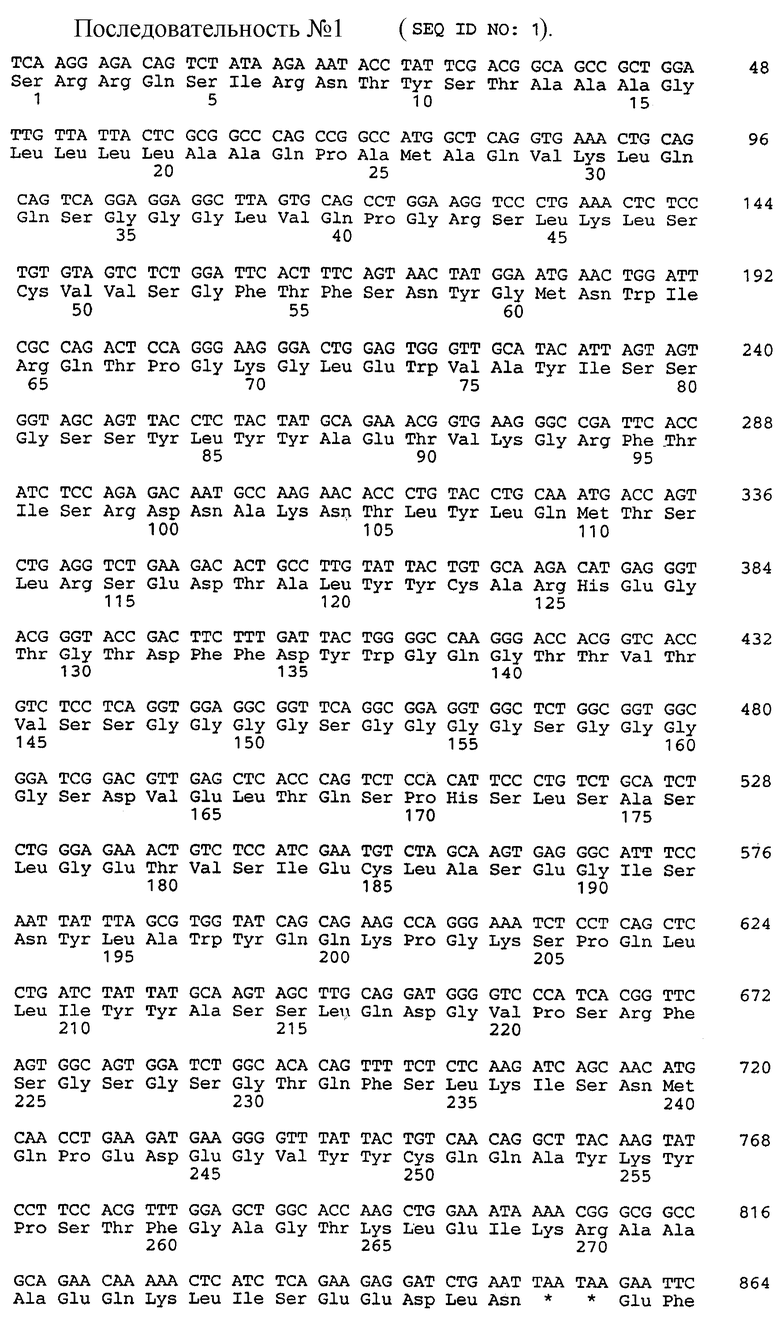
Два фрагмента длиной 340 п.о. и 325 п.о. получают таким образом и затем объединяют благодаря линкеру, который позволяет осуществлять точное расположение кДНК VH в виде 5' этой VL. Этот линкер кодирует 15 аминокислот, образованных тремя повторениями звена (Gly)4Ser (Orlandi R. и др., Proc. Natl. Acad. Sci., USA, 86, 3833-3837 (1979)]. Последовательность внутриклеточного антитела представлена на SEQ ID 1 (остатки 28-270). Эта последовательность придает достаточно степеней свободы для слияния VH-VL, чтобы могло произойти их объединение в параллельной ориентации и обеспечить корректное сродство для антигена.
Слитую последовательность нуклеиновых кислот VH-линкер-VL затем вставляют в фагомиду (un phagemide), который делает возможной экспрессию внутриклеточного антитела (фрагмент ScFv) на поверхность фага М13 (см. чертеж, А). Эта экспрессия позволяет легко осуществлять идентификацию и селекцию внутриклеточных антител, которые точно распознают антиген.
1.2. Функциональная оценка модифицированного внутриклеточного антитела.
ДНК-последовательность, которая кодирует внутриклеточное модифицированное anti-Ras антитело (VН-линкер-VL), выделяется из фагомиды путем рестрикции, затем ее вставляют в вектор sv2 под контролем энхансерной пары раннего промотора SV40 (Schweighoffer и др., Science, 256, 825-827, 1992), чтобы исследовать ее способность антагонизировать воздействия онкогевного Ras. Таким образом полученную плазмиду обозначают как psv2. ScFv. ras. Функциональную оценку осуществляют согласно нескольким тестам:
а) Транзиторная трансфекция в клетки млекопитающих
Для оценки путем транзиторной трансфекции в клетки млекопитающих плазмиду psv2. ScFv. ras котрансфектируют в клетки NIH 3Т3 согласно протоколу, описанному Schweighoffer и др. [Science, 256, 825-827 (1992)], с помощью вектора, который позволяет осуществлять экспрессию гена Ha-ras Val12. Состояние активации пути изучаемой сигнализации регистрируют путем измерения ферментной активности, получаемой от гена-носителя "хлорамфеникол-ацетил-трансферазы (CAT)", находящегося под контролем промотора, содержащего нуклеотидные элементы, отвечающие в виде "транс" воздействию Ras (RRE), также котрансфектированные: эти элементы RRE образованы гибридным ТК-энхансерным промотором полиомы (Wasylyk и сотр., ЕМВО, J., 7, 2475. 1988).
Полученные результаты представлены на чертеже, С. Анализ достигаемых активностей CAT показывает способность модифицированного внутриклеточного антитела, полученного из антитела Y13-259, антагонизировать активность онкогенного Ras.
Плазмида psv2.ScFv.ras, котрансфектированная плазмидой, допускающей экспрессию онкогена, наделенного активностью тирозинкиназы, онкогена НЕR2 (человеческий эпидермальный фактор роста типа II), также блокирует его активность в тесте с плазмидой "CAT" (см. чертеж).
б) Образование центров трансформированных клеток
Канцерогенные (раковые) клетки обладают свойством образовывать центры (очаги) трансформации и особенно фибробласты NIH 3Т3, экспрессирующие онкогенный Ras (Barlat и сотр., Оnсоgеnе (1993), В, 215-218).
Клетки NIH 3T3 культивируют, как в предыдущем тесте, в среде Dulbecco, модифицированной Eagle (DMEM), содержащей 10% эмбриональной телячьей сыворотки, при 37oС при влажности окружающей среды, содержаний 5% СО2. Эти клетки затем котрансфецируют с онкогенным Ras; Ha-ras Val12, плазмида psv2.ScFv. ras (см. п. а) выше, и 10-кратным избытком резистентного к неомицину гена, путем метода трансфекиии катионными липидами (Schweighoffer и сотр., Science, 256, 825-827, 1992). Одинаковое полное количество ДНК трансфецируют в каждой чашке.
Спустя 24 часа после трансфекции, трансфецированные клетки, происходящие из каждой чайки Петри диаметром 100 мм, разделяют в соотношении 1-10 и культивируют в той же самой среде, но в присутствии G418 (GIBCO/BRL) в количестве 0,4 мг на 1 мл среды. Число центров трансформации, полученное на 1 мкг трансфецированной ДНК, подсчитывают после 14 дней культивирования.
Полученные результаты представлены в таблице (см. в конце текста). Они представляют собой среднее из четырех независимых опытов.
Полученные результаты ясно показывают, что внутриклеточное anti-ras антитело очень сильно уменьшает трансформирующую способность онкогснного rаs-гена.
Кроме того, при рассмотрении полученных в п. а) результатов экспрессия этого фрагмента ScFv антитела Y13-I59 также должна противодействовать трансформации за счет других онкогенов, как HЕR1, HER2, облегчая активацию клеточных протеинов Ras.
Разумеется, специалист, на основании описанных в настоящей заявке результатов, может воспроизводить изобретение с последовательностями нуклеиновых кислот, кодирующими внутриклеточные антитела (такие, как фрагменты ScFv), направленные против других клеточных составляющих. Их можно получать либо из известных антител, направленных против клеточных составляющих, либо путем идентификации нейтрализующего антигена, иммунизации посредством этого антигена или его предпочтительного эпитопа, затем получения внутриклеточного антитела из полученных антитела, его мРНК или гибридомы. Также могут быть использованы другие составляющие, принимающие участие в процессах трансформации клеток. Например, другие последовательности ДНК, кодирующие внутриклеточные антитела anti-ras связи, могут быть получены по той же самой методологии из гибридом М38, M8, М90 и М30 (АТСС НВ 9158), антитела которых направлены соответственно против остатков 1-23, 24-69, 90-106, 107-130 и 131-152 протеина Ha-ras. Более того, как указано выше, векторы, несущие одновременно несколько последовательностей, кодирующих различные внутриклеточные антитела, предпочтительно могут быть получены для придания лучшей нейтрализующей активности.
ПРИМЕР 2. Клонирование и экспрессия последовательности ДНК, кодирующей внутриклеточное антитело anti-GAP.
В этом примере описывается клонирование и экспрессия последовательности нуклеиновых кислот, кодирующей внутриклеточный связывающий протеин, репродуцирующий свойства моноклонального антитела, направленного против протеина GAP.
Протеин GAP (активирующий GTPase протеин) принимает участие в ras-зависимом пути сигнализации. Он взаимодействует с протеинами ras каталитически и с увеличенной в 100-200 раз скоростью гидролиза GTP, измеряемой ин витро для нормального протеина р21. В различных исследовательских работах показано, что каталитический домен этого протеина примерно из 1044 аминокислот расположен на карбоксиконцевом участке (остатки 702-1044) и этот участок является ответственным за взаимодействие протеина GAP с протеинами ras (см. ВОИС 91/02749).
В настоящее время показано, что моноклональное антитело, направленное против так называемого "SH3" домена протеина GAP, нейтрализует функции онкогенных протеинов Ras в зиготе Xenope (Duchesne и сотр., Science, 259, 525-528, 1993).
Согласно описанной в п.1.1 методологии можно синтезировать последовательность ДНК, кодирующую внутриклеточное антитело (фрагмент ScFv), соответствующую этому антителу (SEQ ID 2, остатки 11-250, см. чертеж, В). Такая последовательность, включенная в вектор, может позволять ингибирование трансформирующей способности онкогенного ras-гена в опухолевых клетках.
Кроме того, заявитель также более точно идентифицировал распознаваемые этим антителом эпитопы. Эти эпитопы затем были синтезированы искусственно, и их можно использовать для генерирования новых нейтрализующих антител, которые могут служить для осуществления изобретения.
а) Более точная идентификация
Идентификацию реализуют по методу "идентификации антигенных детерминант секвенциального типа в пределах белкового антигена". Этот метод базируется на том принципе, что данное антитело может реагировать с пептидами из 5-15 аминокислот. В силу этого идентификация последовательных эпитопов может быть получена путем приготовления полного набора регенерируемых пептидов, из 5-15 аминокислот, соответствующих полной последовательности рассматриваемого антигена. Этот метод используют для определения функциональных эпитопов домена "SH3" GAP. Для этого, всю совокупность этого домена исследуют путем последовательных (регенераций) восстановлении, путем синтеза докапелтида из любых двух аминокислот.
Синтез регенерируемых пептидов
35 пептидов, составляющих всю совокупность фрагмента (см. чертеж), синтезируют химическим путем. Синтез осуществляют по 2 раза, на двух независимых носителях, по методу Fmoс/трет-бутил на твердой фазе (набор Cambridge Research Biochemicals).
Выделение функциональных эпитопов
Функциональные эпитопы, распознаваемые антителом Ас200, обнаруживают путем ферментного иммуносорбентного анализа (тест ELISA) с помощью кроличьего аити-мышиного (lapin anti-souris) антитела, соединенного с пероксидазой. Хромогенный субстрат используемого фермента представляет собой амино-ди-(3-этилбензотиазодинсульфонат) (ABTS).
Полученные результаты показывают, что распознаваемые этим антителом эпитопы следующие: PVEDRRRVRAI; EISF; EDGWM.
Эти эпитопы могут быть использованы специалистом согласно классическим способам для генерирования антител, нейтрализующих воздействия ras. Эти антитела или гибридомы, подуцирующие их, затем используют для генерирования последовательностей нуклеиновых кислот и векторов изобретения согласно вышеописанной методологии.
ПРИМЕР 3. Получение последовательности нуклеиновых кислот, кодирующей внутриклеточное антитело anti-Ki-ras, из иРНК, выделенных из селезенок мышей, иммунизированных пептидами, происходящими из гиперпеременных частей Ki-Ras (2А и 2В).
В этом примере описывается получение последовательностей нуклеиновых кислот, кодирующих внутриклеточные антитела (такие, как фрагменты ScFv) согласно изобретению путем идентификации нейтрализующего антигена, иммунизации посредством этого антигена или его предпочтительного эпитопа, затем получения последовательности нуклеиновых кислот из полученных антитела, его мРНК или гибридомы.
Этот пример демонстрирует возможность применения настоящего изобретения к любому антигену или желательному эпитопу, даже когда не располагают никаким моноклональным антителом, направленным против вышеуказанного антигена или эпитопа.
Используемым в этом примере антигеном является протеин Ki-Ras. Более конкретно, используемыми для иммунизации антигенами являются пептиды из 25 и 24 аминокислот, соответствующие концам следующих протеинов Ki-Ras 2A и 2В.
Пептид 2A: QYRLKKISKEEKTPGCVKIKKCIIM.
Пептид 2В: KYREKNSKDGKKKKKKSKTKCIIM.
После иммунизации мышей этими пептидами, согласно классическим методам иммунологии, извлекают селезенки и кДНК получают из мРНК, кДНК, кодирующие переменные участки, затем клонируют, что приводит к составу банка фагов, экспрессирующих ScPv, соответствующие совокупности реестра используемых мышей. Внутриклеточные антитела (фрагменты ScFv), распознающие пептиды 2A и 2В, затем идентифицируют и выделяют путем последовательных стадий селекции по сродству на колонке и на микротитрационном планшете.
Эти ScFv затем испытывают на функциональность согласно описанной в примере 1 методике.
Изложенная в этом примере стратегия позволяет предпочтительным образом селекционировать внутриклеточные антитела, специфические к онкогенным Ki-Ras, которые, следовательно, не затрагивают других протоонкогенных Ras. Селективность таких "инструментов", следовательно, не только клеточная (вследствие прерывания путей трансдукиии в трансформированные за счет Ras клетки), но и также молекулярная.
ПРИМЕР 4. Получение последовательности нуклеиновых кислот, кодирующей внутриклеточное антитело, направленное против мутированных протеинов р53 (анти-р53).
В этом примере описывается получение последовательностей нуклеиновых кислот, кодирующих внутриклеточные антитела (такие, как фрагменты ScFv), направленные против мутированных протеинов р53. Эти внутриклеточные антитела получают из различных моноклональных антител, направленных против вышеуказанных, мутированных протеинов р53.
Кодирующий протеин р53 ген изменен в очень большом числе опухолевых клеток (Caron de Promentel и Soussi, Genes, 4, 1-15, 1992). Мутированный протеин р53 не имеет такой же конформации, что и исходный (дикий) р53 (Lane и Benchimol, Genes Dev., 4, 1-8, 1990). Это изменение конформации может быть определено с помощью моноклональных антител (Milner и Cook, Virology, 154, 21-30, 1986; Milner, Nature, 310, 143-145, 1984).
Антитела рАВ240 распознают мутированные формы протеинов Р53.
Внутриклеточная экспрессия фрагмента ScFv специфических антител мутированного р53 или протеинов, специфически взаимодействующих с этими мутированными протеинами р53, должна индуцировать благоприятный эффект в содержащих мутированный р53 опухолях.
ПРИМЕР 5. Клонирование и экспрессия последовательности ДНК, кодирующей внутриклеточное антитело, направленное против вируса папилломы.
В этом примере описывается клонирование и экспрессия ДНК-последовательности, кодирующей внутриклеточное антитело (фрагмент ScFv), направленное против вируса папилломы человека (HPV).
Используемым вирусным протеином является протеин Е6. Этот протеин продуцируется вирусом HPV16 и вирусом HPV18, которые ответственны за 90% раковых заболеваний головного мозга у женщины и идентифицированы в предканцерозных эпителиальных поражениях (Riou и др., Lancet, 335 (1990) 117). Продуцирование гена Е6 приводит к образованию опухолей при сильном уменьшении количества дикого р53, антионкогена, в положительных клетках HPV (Wrede и др., Mol. Carcinog. 4 (1991) 171). В случае других опухолей р53 ингибируется различными механизмами: мутация (см. пример 4) или ассоциация с такими протеинами, как MDM2.
Последовательность протеина Е6 описана в литературе (Haw ley-Nelson и др., EMBO J. 8 (1989) 3905;  и др., J. Virol., 63 (1989) 4417). Особые участки этого протеина могут быть идентифицированы путем "идентификации антигенных детерминант секвенциального типа в пределах белкового антигена" (см. пример 2), затем использованы для иммунизации мышей согласно методике, описанной в примере 3. ДНК-последовательность, кодирующая внутриклеточное антитело (фрагмент ScPv), направленное против протеина Е6 вируса лалилломы человека (HPV), затем получают по вышеописанной методологии. Функциональность этой последовательности доказывается после экспрессии ин виво путем измерения
и др., J. Virol., 63 (1989) 4417). Особые участки этого протеина могут быть идентифицированы путем "идентификации антигенных детерминант секвенциального типа в пределах белкового антигена" (см. пример 2), затем использованы для иммунизации мышей согласно методике, описанной в примере 3. ДНК-последовательность, кодирующая внутриклеточное антитело (фрагмент ScPv), направленное против протеина Е6 вируса лалилломы человека (HPV), затем получают по вышеописанной методологии. Функциональность этой последовательности доказывается после экспрессии ин виво путем измерения
- увеличения содержания дикого р53 в экспрессирующих Е6 клетках;
- морфологической реверсии трансформированных за счет HPV клеток;
- блокирования воздействий Е6 на трансактивацию р53 и
- ингибирования трансформации с помощью Е6 человеческих кератиноцитов и фибробластов.
ПРИМЕР 6. Получение последовательности нуклеиновых кислот, кодирующей внутриклеточное анти-ВИЧ антитело из мРНК, выделенных из селезенок мышей, иммунизированных с помощью пептидов, происходящих из активированных участков протеинов tat у rev и NCP7.
В этом примере описывается получение последовательностей нуклеиновых кислот, кодирующих внутриклеточные антитела согласно изобретению (таких, как фрагменты ScPv) путем идентификации нейтрализующего антигена, иммунизации посредством этого антигена или его предпочтительного эпитопа, затем получения последовательности нуклеиновых кислот на полученных антитела, его мРНК или гибридомы.
Этот пример демонстрирует еще возможность применения настоящего изобретения к любому антигену или желательному эпитопу, даже когда никакое моноклональное антитело, направленное против вышеуказанного антигена или эпитопа, но было описано в уровне техники.
Используемыми (целевыми) антигенами в этом примере являются протеины tat, rev и NCP7 вируса ВИЧ. Более конкретно, используемыми для иммунизации антигенами являются следующие пептиды из 6, 9 и 16 аминокислот, соответствующие участкам этих протеинов, ответственным за их взаимодействие с мРНК (для tat и rev) или димериэацию РНК (для NCP7).
Пептид tat: RKKRRQRRR,
Пептид rev: RQARRNRRRRWRERQR,
Пептид NCP7: RAPRKK.
После иммунизации мышей с помощью этиx пептидов, согласно классическим методам иммунологии, извлекают селезенки и из мРHК получают кДНК. кДHК, кодирующие переменные участки, затем клонируют, что приводит к составу банка фагов, экспрессирующих ScPv, соответствующему совокупности реестра используемых мышей. Внутриклеточные антитела (фрагменты ScFv), распознающие пептиды tat, rev и NCP7, затем идентифицируют и выделяют путем последовательных стадий селекции по сродству на колонке или на микротитрационном планшете.
Эти ScFv затем исследуют на функциональность по их способности блокировать репликативный цикл вируса ВИЧ.



Изобретение относится к генетической инженерии. Предложен фрагмент ДНК, кодирующий внутриклеточный связывающий полипептид. Нуклеотидная последовательность фрагмента и аминокислотная последовательность полипептида приведены в описании. На основе предложенного фрагмента получена фармкомпозиция для лечения рака. Изобретение позволяет получить лекарственное средство для проведения генной терапии. 2 c. и 4 з.п. ф-лы, 1 ил., 1 табл.
| US 4946778 А, 07.08.1990 | |||
| BIOOCA et al., Intracellular Expression of Anti-p21 ras Single Chain Fragments Inhibits Melotic Maturation of Xenopus Oocytes, Biochemical & Biophysical Research Commun | |||
| Способ утилизации отработанного щелока из бучильных котлов отбельных фабрик | 1923 |
|
SU197A1 |
| SULLENGER et al., Overexpression of TAR Sequences Renders Cells Resistant to Human Immunodeficiency Virus Replication, Cell, 63, 1990, р.601-608 | |||
| POSNER et al., An IgG Human Monoclonal Antibody That Reacts with HIV-1/GP120, Inhibits Virus Binding To Cells, and Neutralizes Infection, The Journal Immunology 146(12), 1991, p.4325-4332 | |||
| Уплотнение вращающейся печи | 1974 |
|
SU520499A1 |
| WO 9205250 A1, 02.04.1992 | |||
| WO 9402610 A1, 03.02.1994 | |||
| WO 9312232 A1, 24.06.1993 | |||
| Моноклональные антитела | |||
| /Под редакцией Р.Г.КЕННЕТА и др | |||
| - М.: Медицина, 1983, стр.175-189. | |||

