Изобретение относится к новым вирусным векторам, их получению и их использованию в генной терапии. Оно относится также к содержащим вышеуказанные вирусные векторы фармацевтическим композициям. Более конкретно, данное изобретение относится к рекомбинантным аденовирусам как векторам для генной терапии.
Генная терапия заключается в том, чтобы коррегировать дефицит или дефект (мутация, ошибочная экспрессия и т.д.) путем введения генетической информации в пораженные клетки или орган. Эта генетическая информация может вводиться или in vitro в клетку, извлеченную из органа, и затем измененная клетка снова вводится в организм или непосредственно in vivo в соответствующую ткань. Во втором случае существуют различные методики, среди которых различные методики трансфекции с применением комплексов ДНК и ДЭАЭ-декстрана (Pagano et al. , J. Virol. I (1967) 891), ДНК и нуклеопротеинов (Kaneda et al. , Science 243 (1989) 375), ДНК и липидов (Felgner et al., PNAS 84 (1987) 7413), использованием липосом (Fraley et al., J. Biol. Chem. 255 (1980) 10431) и т.д. Совсем недавно появилось использование вируса в качестве вектора для переноса генов в качестве обещающей альтернативы этим физическим методикам трансфекции. В этом отношении были испытаны различные вирусы на их способность инфицировать определенные клеточные популяции. В частности, испытаны ретровирусы (RSV, HMS, MMS и т.д.), вирус HSV, аденоассоциированные вирусы и аденовирусы.
Среди этих вирусов аденовирусы проявляют некоторые интересные свойства для использования в генной терапии. А именно, они имеют достаточно широкий круг хозяев, способны инфицировать клетки в состоянии покоя, не включаются в генном инфицированной клетки и на сегодняшний день не связаны со значительной патологией у людей.
Аденовирусы являются вирусами с двунитевой линейной ДНК размером около 36 кb. Их геном состоит конкретно из одного обратного повтора (ITR) c последовательностью инкапсидации на его конце из ранних и поздних геном (Cf фиг. 1). Главными ранними генами являются гены Е1 (E1a и E1b), Е2, Е3 и Е4. Главными поздними генами являются гены с L1 по L5.
Принимая во внимание свойства аденовирусов, упомянутые выше, они уже использовались для переноса генов in vivo. С этой целью были получены различные векторы, производные аденовирусов, включающие различные гены (β-gal, ОТС, α-1AT, цитокинов и т.д.). В каждой из этих конструкций аденовирус был модифицирован так, чтобы сделать его неспособным к репликации в инфицированной клетке. Таким образом, конструкции, описанные в предшествующем прототипе, являются аденовирусами, лишенными областей Е1 (E1a и/или Е1b) и в случае необходимости и Е3, на место которых встраиваются последовательности гетерологичной ДНК (Levrero et al., Gene 101 (1991) 195; Gosh-Choudrury et al., Gene 50 (1986) 161). Однако векторы, описанные в прототипе, имеют много недостатков, которые ограничивают их применение в генной терапии. В частности все эти векторы включают много вирусных генов, экспрессия которых in vivo нежелательна в режимах генной терапии. К тому же в эти векторы не могут быть включены фрагменты ДНК очень большого размера, что может быть обходимо для некоторых целей.
Данное изобретение позволяет устранить эти недостатки.
Данное изобретение описывает рекомбинанты аденовируса для генной терапии, способные эффективно переносить ДНК (до 30 кb) in vivо, стабильно экспрессировать на высоких уровнях эту ДНК in vivо, при полном ограничении риска продукции вирусных протеинов, трансмиссии вируса, патогенности и т.д. В частности, было обнаружено, что возможно значительно уменьшить размер генома аденовируса без помехи образованию инкапсидированной вирусной частицы. Это удивительно для размера, который наблюдался в случае другого вируса, например ретровируса, так как некоторые последовательности, распределенные по всей длине генома, необходимы для эффективной инкапсидации вирусных частиц. Вот почему реализация векторов, обладающих значительными внутренними делециями была сильно ограничена. Данное изобретение показывает также, что угнетение основных вирусных генов не мешает никак образованию вирусной частицы как таковой. К тому же рекомбинанты аденовирусов, полученные таким образом, сохраняют, несмотря на значительные модификации их геномной структуры, их полезные свойства сильной инфекционной способности, стабильности in vivо и т.д.
Векторы этого изобретения особенно выгодны, так как они дают возможность включения желательных последовательностей ДНК очень большого размера. Так возможно встроить ген, по длине превосходящий 30 кb. Это особенно интересно для некоторых патологий, для лечения которых необходима коэкспрессия многих генов или экспрессия очень больших генов. Так, например, в случае мышечной дистрофии до настоящего времени было невозможно перенести соответствующую ДНК нативного гена, ответственного за эту патологию (ген дистрофии) из-за его большого размера (14 кb).
Векторы этого изобретения также очень полезны потому, что они имеют очень мало функциональных вирусных областей, и поэтому отрицательные действия, связанные с использованием вируса в качестве векторов при генной терапии, такие как иммуногенность, патогенность, трансмиссия, репликация, рекомбинация и т.д., сильно снижены и даже устранены.
Данное изобретение представляет также, в частности, вирусные векторы, приспособленные для переноса и экспрессии in vivo желательных последовательностей ДНК.
Первый объект данного изобретения касается, таким образом, дефектного рекомбинантного аденовируса, содержащего
- последовательности ITR;
- последовательность, делающую возможной инкапсидацию;
- гетерологическую ДНК последовательность, в котором
- ген E1 является нефункциональным и
- по крайней мере, один из генов Е2, Е4, L1-L5 является нефункциональным.
С точки зрения данного изобретения термин "дефектный аденовирус" обозначает аденовирус, неспособный автономно реплицироваться в клетке-мишени. Обычно геном дефектных аденовирусов по этому изобретению, таким образом, лишен, по крайней мере, последовательностей, необходимых для репликации указанного вируса в инфицированной клетке. Эти области могут или удаляться (полностью или частично), или превращаться в нефункциональные, или заменяться на другие последовательности, а именно на последовательности гетерологичной ДНК.
Повторяющиеся инвертированные последовательности (ITR) представляют начало репликации аденовируса. Они располагаются на концах 3' и 5' вирусного генома (Сf фиг. 1), откуда они могут быть легко выделены по классическим методикам молекулярной биологии, известным специалистам. Нуклеотидная последовательность ITR аденовирусов человека (в частности, серотипов Ad2 и Ad5) описана в литературе, так же как и аденовиусов собаки (а именно, СAV1 и CAV2). Что касается аденовируса Ad5, например, левая последовательность ITR соответствует области, содержащей нуклеотиды генома с 1 по 103.
Последовательность инкапсидации (также называемая последовательностью Psi) необходима для инкапсидации вирусной ДНК. Эта область, таким образом, должна быть представлена, чтобы дать возможность получения дефектных рекомбинантных аденовирусов по этому изобретению.
Последовательность инкапсидации локализуется в геноме аденовируса между левой ITR (5') и геном E1 (Сf фиг. 1). Она может быть выделена или искусственно синтезирована по классическим методикам молекулярной биологии. Нуклеотидная последовательность последовательности инкапсидации аденовирусов человека (в частности, серотипов Ad2 и Ad5) описана в литературе, так же как и аденовирусов собаки (а именно, GAV1 и GAV2). В отношении аденовируса Ad5, например, последовательность инкапсидации соответствует области, включающей нуклеотиды генома со 194 по 358.
Существуют различные серотипы аденовирусов, и, следовательно, структура и свойства несколько варьируют. Тем не менее эти вирусы демонстрируют сравнимое генетическое строение, и сведения, опубликованные по данному вопросу, могут быть легко воспроизведены специалистом для всех типов аденовирусов.
Аденовирусы этого изобретения могут происходить от человека, животного или быть смешанной природы (человеческой и от животных).
Что касается аденовирусов человеческого происхождения предпочтительно использовать те, которые по классификации относятся к группе С. Более предпочтительно из различных серотипов человеческих аденовирусов в рамках данного изобретения использовать аденовирусы типов 2 или 5 (Ad2 или Ad5).
Как показано выше, аденовирусы этого изобретения могут также иметь происхождение от животных или включать последовательности из аденовирусов животного происхождения. Заявитель на самом деле показывает, что аденовирусы животного происхождения способны инфицировать с большой эффективностью человеческие клетки и что они не способны размножаться в человеческих клетках, в которых они испытывались (Cf заявка FR 9305954). Заявитель также показывает, что аденовирус животного происхождения совсем не транскомплементарен аденовирусу человеческого происхождения, так что полностью устраняется риск рекомбинации и размножения in vivо в присутствии человеческого аденовируса, что могло бы привести к образованию инфицирующей частицы. Использование аденовируса или участков аденовирусов животного происхождения, таким образом, особенно выгодно, так как риск, присущий использованию вируса в качестве вектора в генной терапии к тому же очень мал.
Аденовирус животного происхождения, используемый в рамках данного изобретения, может быть собачьим, бычьим, мышиным (например, Mav1, Beard et al. , Virology, 75 (1990) 81), овечьим, свиным, птичьим или еще и обезьяньим (например, SAV). Более конкретно, из птичьих аденовирусов можно упомянуть серотипы с 1 по 10, доступные из АТСС, как, например, штаммы Phelps (АТСС VR-432), Fontes (АТСС VR-280), P7-A (АТСС VR-827), IBH-2A (АТСС VR-828), J2-A (АТСС VR-829), T8-A (АТСС VR-830), K-11 (АТСС VR-921) и еще штаммы для сравнения (референс-штаммы) АТСС VR-831 по 835. Из бычьих аденовирусов можно использовать различные известные серотипы и особенно те, которые находятся в АТСС (типы с 1 по 8) со штаммами сравнения АТСС VR-313, 314, 639-642, 768 и 769. Можно также назвать мышиные аденосирусы FL (АТСС VR-550) и Е20308 (АТСС VR-528), овечий аденовирус типа 5 (АТСС VR-1343) или типа 6 (АТСС VR-1340), свиной аденовирус 5359 или обезьяньи аденовирусы, особенно те, которые являются референс-аденовирусами в АТСС под номерами VR-591-594, 941-943, 195-203 и т.д.
Предпочтительно из различных аденовирусов животного происхождения в рамках этого изобретения используются аденовирусы или участки аденовирусов собак и особенно все штаммы аденовирусов САV2 (штамм Манхэттен или А26/61 (АТСС VR-800), например). Собачьи аденовирусы были объектом многочисленных структурных исследований. Таким образом, были описаны полные карты растрикции аденовирусов САV1 и САV2 в предшествующей работе (Spibey et al., J. Ien. Virol. 70 (1989) 165), и гены E1a, E3, так же как последовательности ITR были клонированы и секвенированы (смотрите, в частности, Spibey et al., Virus Res. 14 (1989) 241; Linne, Virus Res. 23 (1992) 119, WO 91/11525).
Как указано ранее, аденовирусы данного изобретения включают одну гетерологичную последовательность ДНК. Последовательностью гетерологичной ДНК называется вся последовательность ДНК, вводимая в рекомбинантный вирус, который ее переносит и/или экспрессия которой достигается в клетке-мишени.
В частности, последовательность гетерологичной ДНК может содержать один или множество терапевтических генов и/или один или множество генов, кодирующих антигенные пептиды.
Терапевтическими генами, которые могут таким образом переноситься, являются все гены, транскрипция и, в известных случаях, трансляция которых в клетке-мишени производит продукты, имеющие терапевтическое действие.
Можно говорить, в частности, о генах, кодирующих белковые продукты, имеющие терапевтическое действие. Белковым продуктом, кодируемым таким образом, может быть белок, пептид, аминокислота и т.д. Этот белковый продукт может быть гомологичным клетке-мишени (то есть продуктом, который в норме экспрессируется в клетке-мишени, когда в ней не присутствует никакая патология). В этом случае экспрессия белка дает возможность, например, покрыть недостаточную экспрессию в клетке или экспрессию неактивного или слабоактивного белка в соответствии с модификацией и, кроме того, дает возможность суперпродукции указанного белка. Терапевтический ген может также кодировать мутант клеточного белка, имеющего повышенную устойчивость, измененную активность и т. д. Белковый продукт может также быть гетерологичным клетке-мишени. В этом случае экспрессируемый белок может, например, дополнять или привносить в клетку недостающую активность, давая ей возможность бороться с патологией.
Среди терапевтических продуктов по данному изобретению можно привести, в частности, ферменты, производные из крови, гормоны, лимфокины, интерлейкины, интерфероны, TNF и т.д. (FR 9203120), факторы роста, нейротрансмиттеры и их предшественники или ферменты для синтеза, трофические факторы: BDNF, CNTF, NGF, IGF, GMF, AFGF, bFCF, NT3, NT5 и т.д.; аполипопротеины: Аро АI, Аро АIV, АроЕ и т.д. (FR 9305125), дистрофин или минидистрофин (FR 9111947), гены-супрессоры опухолей: р53, Rb Pap1A, DCC, k-rev и т.д. (FR 9304745), гены, кодирующие факторы, участвующие в коагуляции: факторы VII, VIII, IX и т.д.
Терапевтический ген может также быть геном или антисмысловой последовательностью, экспрессия которых в клетке-мишени дает возможность контролировать экспрессию генов или транскрипцию клеточной мРНК. C этих последовательностей, например, в клетке-мишени могут транскрибироваться РНК, комплементарная клеточной мРНК, и блокировать таким образом их трансляцию в белок, соответствующая методика описана в патенте ЕР 140308.
Как показано выше, последовательность гетерологичной ДНК может также включить один или множество генов, кодирующих антигенный пептид, способный генерировать у человека иммунный ответ. В этом особом случае осуществления изобретение дает возможность реализации таким образом вакцин, позволяющих иммунизировать человека, особенно против микроорганизмов или вирусов. Можно, в частности, говорить об антигенных пептидах, специфичных для вируса Эпштейна-Барра, вируса ВИЧ, вируса гепатита В (ЕР 185573), вируса псевдобешенства и специфичных для опухолей (ЕР 259212).
В основном, последовательность гетерологичной ДНК включает как последовательности, обеспечивающие экспрессию терапевтического гена, так и/или ген, кодирующий антигенный петид в инфицируемой клетке. Можно упомянуть последовательности, которые естественно ответственны за экспрессию рассматриваемого гена, когда эти последовательности способны функционировать в инфицируемой клетке. Можно также сказать о последовательностях различного происхождения (ответственных за экспрессию других белков или полностью синтетических). А именно, можно упомянуть промоторные последовательности генов эукареиотов или вирусов. Например, можно говорить о промоторных последовательностях генома той клетки, которую хотят инфицировать. Также можно сказать о промоторных последовательностях генома вируса, включая используемый аденовирус. В этом отношении можно привести в качестве примера промоторы генов Е1А, MLP, CMV, RSV и т.д. Кроме того, эти последовательности экспрессии могут модифицироваться путем добавления последовательностей активации, регуляции и т.д. С другой стороны, когда встраиваемый ген не содержит последовательностей экспрессии, он может помещаться в геном дефектного вируса после такой последовательности.
С другой стороны, последовательность гетерологичной ДНК может также содержать в обратном направлении от терапевтического гена сигнальную последовательность, обеспечивающую секрецию терапевтического продукта из клетки-мишени. Эта сигнальная последовательность может быть природной сигнальной последовательностью терапевтического продукта, но могут быть также названы совсем другие функциональные сигнальные последовательности или искусственная сигнальная последовательность.
Как показано здесь ранее, векторы этого изобретения имеют, по крайней мере, один нефункциональный ген из Е2, Е4, L1-L5. Рассматриваемый вирусный ген можно сделать нефункциональным с помощью всех известных специалистам методик, а именно путем супрессии, замещения, делеции или добавления одного или многих оснований в него или рассматриваемые гены. Такие модификации могут быть получены in vitro (на выделенной ДНК) или in situ, например, или с помощью методов генной инженерии, или путем обработки мутагенами.
В качестве мутагенных средств можно привести, например, физические средства, такие как жесткое облучение (рентгеновское, гамма, ультрафиолетовое и т. д.), или химические средства, способные воздействовать на различные функциональные группы оснований ДНК, например алкилирующие средства (этиленметансульфонат ЭМС), N-метил-N'-нитро-N-нитрозогуанидин, N-нитрохинолеин-1-оксид (NXO), интеркалирующие средства и т.д.
Под делецией в данном изобретении подразумевается любая супрессия рассматриваемого гена. А именно, можно говорить о всей или части области, кодирующей указанный ген, и/или о всей или части промоторной области транскрипции указанного гена. Супрессия может быть выполнена путем расщепления с помощью соответствующих ферментов растрикции, затем лигирования, в соответствии с классическими методами молекулярной биологии, так как показано в примерах.
Генетические модификации могут также осуществляться путем разрушения генов, например, по методу, первоначально описанному Rothstein (Meth. Enzymol. , 101 (1983) 202). В данном случае вся кодирующая последовательность или ее часть предпочтительно разрушается, чтобы сделать возможной замену путем гомологичной рекомбинации геномной последовательности на нефункциональную последовательность или мутантную.
Там, где указанные генетические модификации могут локализоваться, это - в части, кодирующей рассматриваемый ген, или вне кодирующей области, например в областях, ответственных за экспрессию и/или регуляцию транскрипции указанных генов. Нефункциональный характер указанных генов может, таким образом, проявляться продукцией неактивного белка вследствие структурных или конформационных модификаций, отсутствием продукции, продукцией белка, имеющего измененную активность или продукцией природного белка на сниженном уровне или в соответствии с желаемым способом регуляции.
С другой стороны, некоторые изменения, такие как точечные мутации, по природе способны исправляться или быть ослаблены с помощью клеточных механизмов. Такие генетические изменения представляют в таком случае ограниченный интерес для производства. Таким образом, особенно предпочтительно, чтобы свойство нефункциональности было бы совершенно стабильным сегрегационно и/или необратимым.
По преимуществу ген становится нефункциональным в результате частичной или полной делеции.
Предпочтительно дефектные рекомбинантные аденовирусы этого изобретения лишены поздних генов аденовируса.
Наиболее преимущественный способ этого изобретения состоит в получении рекомбинантного дефектного аденовируса, включающего
- последовательности ITR,
- последовательность, обеспечивающую инкапсидацию,
- последовательность гетерологичной ДНК и
- область, несущую ген или часть гена Е2.
Другой особенно выгодный способ этого изобретения состоит в получении дефектного рекомбинантного аденовируса, содержащего
- последовательности ITR,
- последовательность, обеспечивающую инкапсидацию,
- последовательность гетерологичной ДНК и
- область, несущую ген или часть гена Е4.
Всегда, особенно при преимущественном способе, векторы этого изобретения обладают, кроме того, функциональным геном Е3 под контролем гетерологичного промотора. Наиболее предпочтительно, когда векторы обладают частью гена Е3, обеспечивающей возможность экспрессии белка gp 19.
Дефектные рекомбинантные аденовирусы по этому изобретению могут быть получены различными способами.
Первый способ состоит в трансфекции ДНК в дефектный рекомбинантный вирус, полученный in vitro (или путем лигирования, или через плазмидную форму) в компетентной клеточной линии, то есть несущую при культивировании все функции, необходимые для комплементации дефектного вируса. Эти функции предпочтительно интегрированы в геном клетки, что позволяет избежать риска рекомбинации и придает повышенную стабильность клеточной линии. Получение таких клеточных линий описано в примерах.
Второй подход состоит в котранфекции соответствующей клеточной линии ДНК дефектного рекомбинантного вируса, полученного in vitro (или путем лигирования, или в форме плазмиды) и ДНК хелперного вируса. По этому методу не нужно располагать компетентной клеточной линией, способной восполнять дефектные функции рекомбинантного аденовируса. Часть этих функций на деле восполняется хелперным вирусом. Этот хелперный вирус должен сам быть дефектным, и клеточная линия при культивировании несет необходимые функции для его дополнения.
Получение дефектных рекомбинантных вирусов этого изобретения по этому методу также показано в примерах.
Среди клеточных линий, используемых в рамках этого второго подхода, можно, в особенности, назвать линию клеток почки эмбриона человека 293, клеток КВ, клеток Неla, MDCK, GHK и т.д. (Сf примеры).
Затем векторы, которые размножены, выделяют, очищают и амплифицируют по классическим методикам молекулярной биологии.
Данное изобретение, таким образом, имеет отношение также и к клеточным линиям, инфицируемым аденовирусом, подразумевая интегрирование в их геном функции, необходимой для дополнения дефектного рекомбинантного аденовируса, такого как описанный ранее. В частности, оно касается клеточных линий, включающих интегрированные в их геном области Е1 и Е2 (а именно, области, кодирующей белок 72К), и/или Е4, и/или ген рецептора для глюкокортикоидов. Предпочтительно эти линии получены для части линии 293 или gmDBP6.
Данное изобретение относится также ко всей фармацевтической композиции, содержащей один или несколько дефектных рекомбинантных аденовирусов, таких, которые описаны выше. Фармацевтические композиции этого изобретения могут быть сформированы для введения путем местного применения, перорального, парентерального, интраназального, внутривенного, внутримышечного, подкожного, внутриглазного, трансдермального введения и т.д.
Предпочтительно фармацевтическая композиция содержит фармацевтически приемлемые носители для инъекционных форм. Можно, в частности, назвать растворы солей (мононатриевый фосфат, динатриевый фосфат, хлорид натрия, калия, кальция или магния и т.д. или смеси этих солей), стерильные, изотонические или высушенные композиции, особенно лиофилизированные, которым по потребности путем добавления стерилизованной воды или физиологической сыворотки придают вид (состав) инъекционных растворов.
Дозы вируса, используемые для инъекции, могут быть адаптированы в зависимости от различных параметров и особенно в зависимости от используемого способа введения, затрагиваемой патологии, гена, который нужно экспрессировать, и от продолжительности искомого лечения. Обычно рекомбинантные аденовирусы по этому изобретению готовятся и вводятся в дозах, содержащих от 104 до 1014 бое/мл и предпочтительно от 106 до 1010 бое/мл. Термин бое ("бляшкообразующая единица") соответствует инфицирующей способности раствора вируса и определяется по инфицированию соответствующей клеточной культуры и подсчету, обычно через 5 дней, числа бляшек инфицированных клеток. Методики определения титра бое вирусного раствора хорошо представлены в литературе.
В соответствии со встроенной последовательностью гетерологичной ДНК аденовирусы этого изобретения могут использоваться для лечения или предотвращения многих патологий, включая генетические болезни (дистрофию, цистофиброз и т.д.), нейродегенеративные болезни (Альцгеймера, Паркинсона, ALS и т. д.), раковых заболеваний, патологий, связанных с расстройствами коагуляции или с дислипопротеинемиями, патологиями, связанными с вирусными инфекциями (гепатиты, SIDA и т.д.), и т.д.
Далее изобретение будет более полно описано с помощью примеров, которые следуют далее и которые должны рассматриваться как иллюстративные, а не ограничивающие.
Описание чертежей
Фиг. 1: генетическое строение аденовируса Аd5. Полная последовательность Аd5 доступна на основе базы данных и позволяет специалисту выбрать или создать любой сайт рестрикции и, таким образом, выделить всю область из генома.
Фиг. 2: карта рестрикции аденовируса САV2 штамм Манхэттен (по Spibey et al., упомянутой выше).
Фиг. 3: создание дефектных вирусов этого изобретения путем лигирования.
Фиг. 4: конструкция рекомбинантного вируса, несущего ген Е4.
Фиг. 5: конструкция рекомбинантного вируса, несущего ген Е2.
Фиг. 6: конструкция и общий вид плазмиды рРY32 и pPY31.
Фиг. 7: общий вид плазмиды рРY55.
Фиг. 8: общий вид плазмиды р2.
Фиг. 9: общий вид промежуточной плазмиды, используемой для построения плазмиды рITRL5-E4.
Фиг. 10: общий вид плазмиды рITRL5-E4.
Общие методики молекулярной биологии
Классические методы, используемые в молекулярной биологии, такие как препаративная экстракция плазмидной ДНК, центрифугирование плазмидной ДНК в градиенте хлорида цезия, электрофорез в агарозном или акриламидном геле, очистка фрагментов ДНК путем электроэлюции, экстракция белков фенолом или фенолом-хлороформом, осаждение ДНК в солевой среде этанолом или изопропанолом, трансформация Е. соli и т.д., хорошо известны специалистам и широко описаны в литературе (Maniatis T. et al., "Molecular Cloning, a Laboratory Manual", Cold Spring Harbor Laboratory, Cold Sprung Harbor. N.Y., 1982; Ausubel F. M. et al. (eds), "Current Protocols in Molecular Biology", John Wiley and Sons, New York, 1987).
Плазмиды типа рВR322, рUC и фаги серии М13 закуплены (Веthesda Research Laboratories).
Для лигирования фрагмены ДНК могут быть разделены по их размеру путем электрофореза в агарозном или акриламидном геле, экстрагированы фенолом или смесью фенола/хлороформа, осаждены этанолом и затем инкубированы в присутствии ДНК-лигазы фага Т4 (Biolabs) по рекомендациям поставщика. Заполнение выступающих концов 5' может быть осуществлено с помощью фрагмента Кленова ДНК по спецификации производителя. Устранение выступающих концов 5' выполняется в присутствии ДНК-полимеразы фага Т4 (Biolabs), используемой по рекомендациям производителя. Устранение выступающих концов 5' выполняется путем осторожной обработки нуклеазой SI.
Направленный мутагенез in vitro с помощью синтетических олигодезоксинуклеотидов может выполняться по методу, разработанному Taylor et al. (Nucleic Acids Res. 13 (1985) 8749-8764), с использованием набора фирмы Amersham.
Ферментативная амплификация фрагментов ДНК с помощью метода РСR (Polymerase-catalyzed Chain Reaction, Saiki R.K. et al., Science, 230 (1985) 1350-1354; Mullis K.B. et Faloona F.A., Meth. Enzym. 155 (1987) 335-350) может осуществляться при использовании "DNA thermal cycler" (Perkin Elmer Cetus) по описанию изготовителя.
Определение нуклеотидных последовательностей может выполняться по методу, разработанному Sanger et al. (Proc. Natl. Acad. Sci. USA, 74 (1977) 5463-5467) с использованием набора фирмы Amersham.
Используемые клеточные линии
В примерах, которые следуют ниже, используются или могут использоваться следующие клеточные линии.
- Линия 293 клеток почки эмбриона человека (Graham et. al., J. Gen. Virol. 36 (1977) 59). Эта линия содержит, конкретно, встроенную в ее геном левую часть человеческого аденовируса Аd5 (12%).
- Линия клеток КВ человека: выделенная из эпидермальной карциномы человека эта линия доступна из АТСС (N хран. ССL17), так же как доступны и условия ее культивирования.
- Линия клеток Hela человека: полученная из карциномы человеческого эпителия эта линия доступна в АТСС (N хран. ССL2), также доступны и условия ее культивирования.
- Линия клеток МDСК собаки: Условия культуры клеток MDCK были описаны, в частности, Macatney et al., Science 44 (1988) 9.
- Линия клеток gmDBP6 (Brough et al., Virology 190 (1992) 624). Эта линия образована клетками Hela, несущими ген аденовируса Е2 под контролем LTR MMTV.
Пример 1.
Этот пример показывает возможность рекомбинантного аденовируса, лишенного основных вирусных генов. Для этого была создана серия мутантов с делецией в аденовирусе путем лигирования in vitro, и каждый из этих мутантов был контрансфицирован с вирусом-хелпером в клетки КВ. Эти клетки не позволяют размножаться вирусу, дефектному по Е1, причем транскомплементация направлена на область Е1.
Были получены различные мутанты с делецией на основе аденовируса Аd5 путем расщепления и затем лигирования in vitro. Для этого по методике, описанной Zipp et al. (J. Virol. 63 (1989) 5133), выделена вирусная ДНК из Аd5, подвергнута расщеплению в присутствии различных ферментов рестрикции (Сf фиг. 3), затем продукт расщепления лигировали в присутствии Т4 ДНК-лигазы. Размер различных делеций затем контролировали в 0,8% агарозном геле с ДСН (SDS). Эти мутанты затем картировали (Сf фиг.3). Эти различные мутанты несут следующие области:
mt1: лигирования между фрагментами Аd5 - 0-20642 (SauI) и (SauI) 33797-35935,
mt2: лигирования между фрагментами Аd5 - 0-19549 (NdeI) и (NdeI) 31089-35935,
mt3: лигирования между фрагментами Аd5 - 0-10754 (AatII) и (AatII) 25915-35935,
mt4: лигирования между фрагментами Аd5 - 0-11311 (MluI) и (MluI) 24392-35935,
mt5: лигирования между фрагментами Аd5 - 0-9462 (SaII) и (XhoI) 29791-35935,
mt6: лигирования между фрагментами Аd5 - 0-5788 (ХhoI) и (ХhoI) 29791-35935,
mt7: лигирования между фрагментами Аd5 - 0-3665 (SphI) и (SphI) 31224-35935.
Каждый из мутантов, полученных выше, был контрансфицирован с вирусной ДНК Ad. RSV. Bgal (Stratford-Perricaudet et al., J. Clin. Invest. 90 (1992) 626) в клетки КB в присутствии фосфата кальция. Клетки собирали через 8 дней после трансфекции и супернатанты культуры собирали, затем амплифицировали в клетках КВ до получения накоплений 50 чашек для каждой трансфекции. Сначала из каждого образца была выделена эписомная ДНК и разделена в градиенте хлорида цезия. В каждом случае наблюдались две четкие полосы вируса, которые выделяли и анализировали. Более тяжелая соответствовала вирусной ДНК Ad. RSV. Bgal и более легкая - ДНК рекомбинантного вируса, произведенная путем лигирования (фиг. 3). Титр, полученный для этой последней, составляет около 108 бое/мл.
Вторая серия мутантов с делецией в аденовирусе создавалась путем лигирования in vitro по той же самой методологии. Эти другие мутанты несут следующие области:
mt8: лигирования между фрагментами 0-4623 (Apa1) Ad RSV. Bgal и (Apa1) 31909-35935 Ad5,
mt9: лигирования между фрагментами 0-10178 (BgIII) Ad RSV. Bgal и (BamHI) 21562-35935 Ad5.
Эти мутанты, несущие ген LacZ под контролем промотора LTR вируса RSV, затем коинфицировали в клетки 293 в присутствии вирусной ДНК Н21808 (Weinberg et al. , J. Virol. 57 (1986) 833), которая лишена области Е4. По этой второй методике транскомплементация направлена на Е4, а не на Е1.
Эта методика позволяет так же, как описано выше, получать вирусные рекомбинанты, обладающие только вирусным геном, но не областью Е4.
Пример 2.
В этом примере описывается получение дефектных рекомбинантных аденовирусов по этому изобретению путем котрансфекции ДНК рекомбинантного вируса, включенной в плазмиду, с вирусом-хелпером.
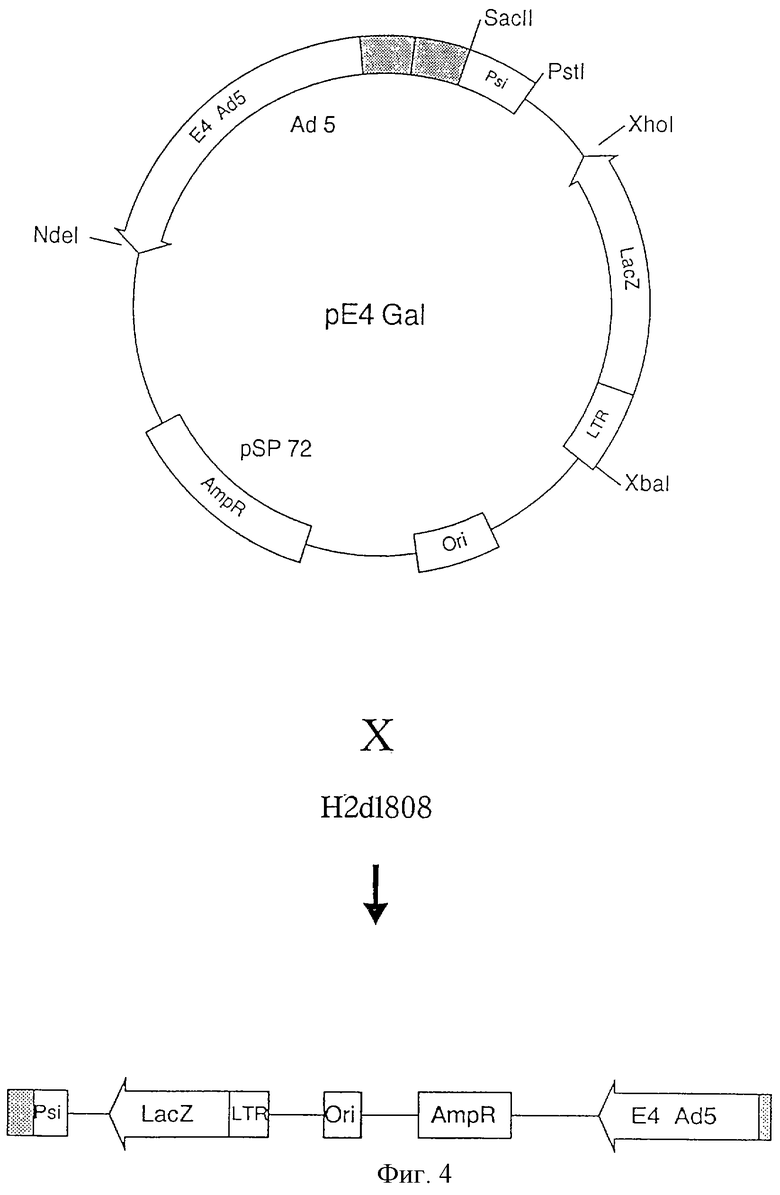
Для этого была сконструирована плазмида, несущая примыкающие ITR Ad5, последовательность инкапсуляции, ген Е4 под контролем соответствующего промотора и в качестве гетерологичного гена ген LacZ под контролем промотора LTR вируса RSV (фиг.4). Эта плазмида, обозначенная рЕ4Gаl была получена путем клонирования и лигирования следующих фрагментов (см. фиг.4):
- фрагмента НindIII-SacII, происходящего из плазмиды рFG144 (Graham et al. , EMBO J. 8 (1989) 2077). Этот фрагмент несет последовательности ITR Ad5 от головы к хвосту и последовательность инкапсидации: фрагмент HindIII (34920)-SacII (352);
- фрагмента Аd5, содержащегося между сайтами SacII (расположенном на уровне пары оснований 3827) и PstI (расположенном на уровне пары оснований 4245);
- фрагмента рSP 72 (Promega), содержащегося между сайтами PstI (pb 32) и Sa1I (pb 34);
- фрагмента XhoI-XbaI плазмиды рAdLTRGalIX, описанного в Stratford-Perricaudet et al. , (JCI 90 (1992) 626). Этот фрагмент несет ген LacZ под контролем LTP вируса RSV;
- фрагмента XbaI (pb40)-NdeI (pb 2379) плазмиды pSP 72;
- фрагмента NdeI (pb 31079) - HindIII (pb 34930) Ad5. Этот фрагмент, локализованный в правом конце генома Ad5, содержит область Е4 под контролем соответствующего ему промотора. Он был клонирован на уровне сайтов NdeI (2379) плазмиды pSP 72 и HindIII первого фрагмента.
Эта плазмида была получена путем клонирования различных фрагментов в указанных областях плазмиды рSP 72. Понятно, что эквивалентные фрагменты могут быть получены специалистами исходя из других источников.
Плазмида рЕ4Gal затем контрансфицируется с ДНК вируса Н2dl 808 в клетки 293 в присутствии фосфата кальция. Затем получают рекомбинантный вирус, как описано в примере 1. Этот вирус несет в качестве единственного вирусного гена ген Е4 аденовируса Ad5 (фиг. 4).
Его геном имеет размер около 12 кb, такой, который дает возможность встройки гетерологичной ДНК весьма большого размера (до 20 кb). К тому же специалист может легко заменить ген LacZ на совершенно другой терапевтический ген из тех, которые упомянуты выше. С другой стороны этот вирус несет некоторые последовательности, происходящие из плазмиды рSP 72, которые могут исключаться с помощью классических методик молекулярной биологии, если необходимо.
Пример 3.
В этом примере описывается получение другого дефектного рекомбинантного аденовируса по этому изобретению путем коинфицирования с помощью вируса-хелпера, ДНК рекомбинантного вируса, включенного в плазмиду.
Для этого была создана плазмида, несущая примыкающие ITR Ad5, последовательность инкапсидации, ген Е2 Аd2 под контролем собственного промотора и, в качестве гетерологичного гена, ген LacZ под контролем промотора LTR вируса RSV (фиг. 5). Эта плазмида, обозначенная как рЕ2Ga I, была получена путем клонирования и лигирования следующих фрагментов (см. фиг.5):
- фрагмента HindIII-SacII, происходящего из плазмиды рFG144 (Graham et al. , EMBO J. 8 (1989) 2077). Этот фрагмент несет последовательности ITR Ad5 от головы к хвосту и последовательность инкапсидации: фрагмент HindIII (34920)-SacII (352);
- фрагмента Ad5, содержащегося между сайтами SacII (локализованного на уровне пары оснований 3827) и PstI (локализованного на уровне пары оснований 4245);
- фрагмента рSP 72 (Promega), содержащегося между сайтами PstI (pb 32) и Sa1I (pb 34);
- фрагмента XhoI - XbaI плазмиды рАdLTRGalIX, описанного Stratford - Perricaudet et al. (JCI 90 (1992) 626). Этот фрагмент несет ген LacZ под контролем LTR вируса RSV. Он был клонирован на уровне сайтов Sa1I (34) и XbaI плазмиды рSP 72.
- фрагмента pSP 72 (Promega), содержащегося между сайтами XbaI (pb 34) и BamHI (pb 46);
- фрагмента BamHI (pb 21606) - SmaI (pb 27339) Ad2. Этот фрагмент генома Ad2 содержит область Е2 под контролем его соответствующего промотора. Он был клонирован на уровне сайтов BamHI (46) и ЕcoRV плазмиды pSP 72;
- фрагмента ЕсоRV (рb 81)- HindIII (pb 16) плазмиды рPS 72.
Эта плазмида была получена путем клонирования различных фрагментов в указанные области плазмиды pSP 72. Понятно, что специалисты могут получить эквивалентные фрагменты, исходя из других источников.
Далее плазмида рЕ2GaI котрансфицировалась вместе с ДНК вируса Н2d1802, лишенной области Е2 (Rice et al. J. Virol. 56 (1985) 767), в клетки 293 в присутствии фосфата кальция. Рекомбинантный вирус затем получали, как описано в примере 1. Этот вирус несет в качестве единственного вирусного гена ген Е2 аденовируса Аd2 (фиг.5). Его геном имеет размер около 12 кb, такой, который дает возможность встройки гетерологичной ДНК весьма большого размера (до 20 кb). Таким образом, специалист может легко заменить ген LacZ на любой другой терапевтический ген, такой, который упомянут выше. В другой стороны, этот вирус несет некоторые последовательности, происходящие из промежуточной плазмиды, которые могут быть устранены с помощью классических методик молекулярной биологии, если необходимо.
Пример 4.
В этом примере описывается создание клеточных линий, комплементирующих области Е1, Е2 и/или Е4 аденовируса. Эти линии позволяют создавать конструкции рекомбинантных аденовирусов по этому изобретению, лишенных этих областей, не прибегая к использованию вируса-хелпера. Эти вирусы получены путем рекомбинации in vivo и могут включать важные гетерологичные последовательности.
В описанных клеточных линиях области Е2 и Е4, потенциально цитотоксичные, помещены под контроль индуцируемого промотора: LTR MMTV (Pharmacia), который индуцируется дексаметазоном, или нативным или в минимальной форме, описанной в PNAS 90 (1993) 5603; или подавляемой тетрациклином системой, описанной Gossen и Büjard (PNAS 89 (1992) 5547). Понятно, что могут использоваться другие промоторы, частности варианты LTR MMTV, несущие, например, гетерологичные области регуляции (а именно, "энхансерную" область). Линии этого изобретения были созданы путем трансфекции соответствующих клеток в присутствии фосфата кальция фрагментом ДНК, несущим указанные гены (области аденовируса и/или ген рецептора для глюкокортикоидов) под контролем промотора транскрипции и терминатора (сайта полиаденилирования). Терминатор может быть или природным терминатором трансфицируемого гена, или другим терминатором, как, например, терминатор раннего мессенджера вируса SV40. Предпочтительно фрагмент ДНК несет также ген, дающий возможность селекции трансформированных клеток, как, например, ген устойчивости к генетицину. Ген устойчивости может нести также фрагмент другой ДНК, котрансфицируемой с первой.
После трасфекции трасформированные клетки отбирали, и их ДНК анализировали для подтверждения интеграции фрагмента ДНК в геном.
Эта методика позволяет получить следующие клеточные линии.
1. Клетки 293, обладающие геном 72К области Е2 Ad5 под контролем LTR MMTV.
2. Клетки 293, обладающие геном 72К области Е2 Ad5 под контролем LTR MMTV и геном рецептора для глюкокортикоидов.
3. Клетки 293, обладающие геном 72К области Е2 Ad5 под контролем LTR MMTV и областью Е4 под контролем LTR MMTV.
4. Клетки 293, обладающие геном 72К области Е2 Ad5 под контролем LTR MMTV, областью Е4 под контролем LTR MMTV и геном рецептора для глюкокортикоидов.
5. Клетки 293, обладающие областью Е4 под контролем LTR MMTV.
6. Клетки 293, обладающие областью Е4 под контролем LTR MMTV и геном рецептора для глюкокортикоидов.
7. Клетки gmDBP6, обладающие областями Е1А и Е1В под контролем их соответствующего промотора.
8. Клетки gmDBP6, обладающие областями Е1А и Е1В под контролем их собственного протомотора и областью Е4 под контролем LTR MMTV.
Пример 5.
В этом примере описывается получение дефектных рекомбинантных аденовирусов по этому изобретению, геном которых лишен генов Е1, Е3 и Е4. По предпочтительному способу осуществления, проиллюстрированному в этом примере и в примере 3, геном рекомбинантных аденовирусов этого изобретения модифицирован так, чтобы гены Е1 и Е4 были бы, по крайней мере, нефункциональными. Такие аденовирусы обладают прежде всего способностью включения важных гетерологичных генов. С другой стороны, эти векторы имеют высокую безопасность, потому что делеция области Е4, которая участвует в регуляции экспрессии поздних генов, стабильности поздних ядерных РНК, прекращения экспрессии белков клетки-хозяина и эффективности репликации вирусной ДНК. Эти векторы обладают, таким образом, фоновым шумом транскрипции, и экспрессия вирусных генов сильно снижена. И, наконец, по особенно предпочтительному способу эти векторы могут производиться до титров, сравнимых с таковыми диких аденовирусов.
Эти аденовирусы получали, начиная с плазмиды рPY55, несущей модифицированную правую часть генома аденовируса Ad5, или путем котрансфекции с холперной плазмидой (см. также примеры 1, 2 и 3), или с помощью дополняющей линии (пример 4).
5.1. Cоздание плазмиды рPY55.
а) Построение плазмиды рPY32
Фрагмент АvrII-BcII плазмиды РFG144 (F.L. Graham et al. EMBO J. 8 (1989) 2077-2085), соответствующей правому концу генома аденовируса Ad5, сначала был клонирован между сайтами XbaI и BamHI вектора рIC19H, начиная с контекста dam-. Это давало плазмиду рPY23. Интересной характеристикой плазмиды рPY23 является то, что сайт Sa1I, происходящий из мультисайта клонирования вектора рIC19Н, остается единственным и что он располагается около правого конца генома аденовируса Ad5. Фрагмент HaeIII-Sa1I плазмиды рPY23, который содержит правый конец генома аденовируса Ad5, начиная с сайта HaeIII, располагается в положении 35614 и затем клонируется между сайтами ECoRV и XhoI вектора рIС20Н, что дает плазмиду рPY29. Интересной характеристикой этой плазмиды является то, что сайты XbаI и ClaI, происходящие из мультисайта клонирования вектора рIС20Н, располагаются около соединения ЕсоRV/НаеIII, полученного в результате клонирования. Кроме того, это соединение модифицирует нуклеотидный контекст, непосредственно примыкающий к сайту СlаI, который теперь становится метилируемым в контексте dam+. Фрагмент XbaI (30470) - MaeII (32811) генома аденовируса Ad5 затем клонировали между сайтами XbaI и ClaI плазмиды рPY29, полученной, начиная с контекста dam-, что дает плазмиду рPY30. Фрагмент SStI плазмиды рPY30, который соответствует последовательности генома аденовируса Ad5 с сайта SStI в положении 30556 до правого конца, в заключение клонировали между сайтами SStI вектора рIС20Н, что дает плазмиду рPY31, карта рестрикции воставки которой, локализованной между сайтами HindIII, дана на фиг.6.
Плазмида рPY32 была получена после частичного расщепления плазмиды рPY31 с помощью BgIII с последующим полным расщеплением с помощью BamHI, а затем повторным лигированием. Плазмида рPY32 соответствует, таким образом, делеции генома аденовируса Ad5, расположенной между сайтом BamHI плазмиды рPY31 и сайтом BgIII, локализованном в положении 30818. Карта рестрикции фрагмента HindIII плазмиды pPY32 дана на фиг.6. Особенность плазмиды рPY32 состоит в том, что она обладает уникальными сайтами SaII и XbaI.
b) Построение плазмиды pPY47
Фрагмент BamHI (21562) - XbaI (28592) генома аденовируса Ad5 сначала клонировали между сайтами BamHI и XbaI вектора р1С19Н, полученного начиная с контекста dam-, что давало плазмиду рPY17. Эта плазмида содержит, таким образом, фрагмент HindIII (26328)-BglII (28133) генома аденовируса Ad5, который может быть клонирован между сайтами HindIII и BglII вектора p1C20Р, для получения плазмиды рPY34. Особенность этой плазмиды состоит в том, что сайт BamHI, происходящий из мультисайта клонирования, располагается непосредственно вблизи сайта HindIII (26328) генома аденовируса Ad5.
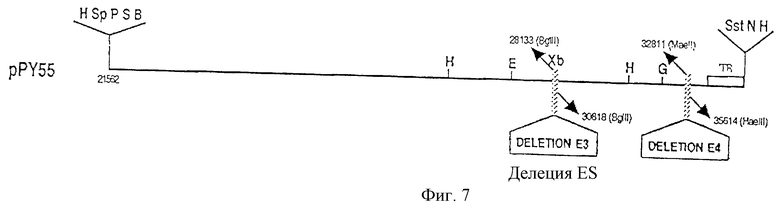
Фрагмент BamHI (21562)-HindIII (26328) генома аденовируса Ad5 происходящий из плазмиды рPY17, затем клонируется между сайтами BamHI и HindIII плазмиды рPY34, что дает плазмиду рPY39. Фрагмент BamHI - XbaI плазмиды рPY39, полученной, начиная с контекста dam-, содержащий часть генома аденовируса Ad5, заключающегося между сайтами BamHI (21562) и BglII (28133), затем клонировали между сайтами BamHI и XbaI вектора рIС19Н, полученного начиная с контекста dam-. Это дает плазмиду рPY47, интересной особенностью которой является то, что сайт Sa1I, происходящий из мультисайта клонирования, локализуется вблизи сайта HindIII (фиг.7).
с) Построение плазмиды рPY55
Фрагмент SaII - XbaI плазмиды рPY47, полученный, начиная с контекста dam-, и который содержит часть генома аденовируса Ad5, начиная с сайта BamHI (21562) до сайта BglII (28133), клонировали между сайтами SalI и XbaI плазмиды рPY32, что давало плазмиду рPY55. Эта плазмида непосредственно может использоваться для получения рекомбинантных аденовирусов, лишенных, по крайней мере, области Е3 (делеция между сайтами BglII, расположенными в положениях 28133 и 30818 генома аденовируса Ad5) и полной области Е4 (делеция между сайтами MaeII (32811) и НаеIII (35614) генома аденовируса Ad5 (фиг. 6, 7).
5.2. Получение аденовируса, содержащего, по крайней мере, одну делецию в области Е4 и предпочтительно, по крайней мере, в областях Е1 и Е4.
а) Получение путем котрансфекции с вирусом-хелпером Е4 клеток 293
Принцип, основанный на транскомплементации между "минивирусом" (вирус-хелпер), экспрессирующим область Е4, и рекомбинантным вирусом, лишенным, по крайней мере, Е3 и Е4. Эти вирусы были получены или путем лигирования in vitro, или после рекомбинации in vivo по следующей стратегии.
(I) ДНК вируса Ad-d1324 (Thimmappaya et al., Cell 31 (1982) 543) и плазмиду рPY55, расщепленные BamHI, сначала лигировали in vitro, затем котрансфицировали с плазмидой рЕАCaI (описанной в примере 2) в клетки 293.
(II) ДНК вируса Ad-d1324, расщепленную EcoRI и плазмиду рPY55, расщепленную BamHI котрансфицировали с плазмидой рЕ4GaI в клетки 293.
(III) ДНК аденовируса Ad5 и плазмиду рPY55, расщепленные BamHI, лигировали и затем котрансфицировали с плазмидой pE4GaI в клетки 293.
(IV) ДНК аденовируса Ad5, расщепленную EcoRI и плазмиду рPY55, расщепленную BamHI, котрансфицировали с рЕАGаI в клетки 293.
Стратегии (I) и (II) позволяют получить рекомбинантный вирус, лишенный областей E1, Е3 и Е4; стратегии (III) и (IV) позволяют получить рекомбинантный вирус, лишенный области Е1, но выражающий какой-нибудь трансген, может использоваться вместо ДНК вируса d1324 в соответствии со стратегиями (I) или (II), в целях получения рекомбинантного вируса, лишенного областей Е1, Е3 и Е4 и выражающий указанный трансген.
b) Получение с помощью клеточных линий, транскомплементирующих функции Е1 и Е4
Принцип, положенный здесь в основу, тот, что линия клеток, производная от линии, экспрессирующей области Е1, например линия 293, и также экспрессирующая, по крайней мере, открытые рамки считывания ORF6 и ORF6/7 области Е4 аденовируса Ad5 под контролем промотора, например индуцируемого, способна транскомплементировать сразу обе области Е1 и Е4 аденовируса Ad5. Такие линии описаны в примере 4.
Рекомбинантный вирус, лишенный областей Е1, Е3 и Е4, может, таким образом, быть получен путем лигирования in vitro или путем рекомбинации in vivo по методикам, описанным здесь выше. Какая бы методика не использовалась для получения вирусов, лишенных, по крайней мере, области Е4, после трансфекции в эти использованные клетки наблюдался цитопатический эффект (показывающий продукцию рекомбинантных вирусов). Затем клетки собирали, разрушали путем трех циклов замораживания-размораживания в их супернатанте, затем центрифугировали при 4000 об/мин в течение 10 минут. Супернатант, полученный таким образом, затем амплифицировали на свежей клеточной культуре (клетки 293 по методикам а) и клетки 293, экспрессирующие область Е4, по методике b)). Затем вирус очищали от мембран и его ДНК анализировали по методу Hirt (упомянутому выше). Концентраты штамма вируса получали затем в градиенте плотности хлорида цезия.
Пример 6.
В этом примере описывается получение дефектных рекомбинантных аденовирусов, геном которых лишен генов Е1, Е3, L5 и Е4. Эти векторы особенно предпочтительны, потому что область L5 кодирует последовательность, которая является белком, чрезвычайно токсичным для клетки.
Эти аденовирусы получали, начиная с плазмиды р2, несущей модифицированную правую часть генома аденовируса А5, путем котрансфекции с различными хелперными плазмидами. Они могут быть также получены с помощью дополняющей линии.
6.1. Построение плазмиды р2.
Эта плазмида содержит всю правую область генома аденовируса Ad5, начиная с сайта BamHI (21562), из которой удален фрагмент, заключенный между сайтами XbaI (28592) и AvrII (35463), несущий гены Е3, L5 и Е4. Плазмида р2 была получена путем клонирования и лигирования следующих фрагментов в плазмиду рIС19R, расщепленную BamHI и дефосфорилированную (см. фиг.8):
- фрагмента генома аденовируса Ad5, содержащегося между сайтами BamHI (21562) и XbaI (28592), и
- правого конца генома аденовируса Ad5 (содержащего правую ITR), начиная с сайта AvrII (35463) по сайт BclI (соответствующий BamHI).
6.2. Построение хелперной плазмиды (рITRL5-Е4), несущей ген L5.
Хелперная плазмида рITR L5-Е4 привносит гены Е4 и L5. Она соответствует плазмиде рЕ4Ga1 описанной в примере 2, несущей, кроме того, область L5, кодирующую нить (последовательность) под контролем промотора МLР аденовируса Ad2. Плазмида рITR L5-Е4 была построена следующим образом (фиг.9 и 10).
Был синтезирован олигонуклеотид из 58 pb, содержащий в направлении 53-3' сайт HindIII, АТG нити и последовательность, кодирующую нить до сайта NdeI в положении 31089 генома аденовируса Ad5. Последовательность этого нуклеотида дана здесь ниже в ориентации 5'-3':
Сайты HindIII на 5' и Ndel на 3' подчеркнуты одной линией, АТC нити подчеркнута двойной линией.
Фрагмент SSpI - HindIII, содержащий последовательность промотора MLP, следующую за состоящим из трех частей лидером аденовируса Ad2 был выделен из плазмиды pMLР10 (Ballay et al., (1987) UCLA Symposia on molecular and cellular biology, New Series, vol.70, Robinson et al. (Eds) New York, 481). Этот фрагмент был встроен вместе с олигонуклеотидом из 58 pb, описанным здесь выше, между сайтами Ndel и ЕсоRV плазмиды pIC19R c получением промежуточной плазмиды (см. фиг.9).
Фрагмент SacII (затупленный) NdeI плазмиды pE4GaI (пример 2) затем вводили в промежуточную плазмиду между сайтами SSpI и NddeI с получением плазмиды рITRL5-Е4 (фиг.10).
6.3. Получение дефектных рекомбинантных аденовирусов, содержащих делецию в областях Е1, Е3, L5 и Е4.
а) Получение путем котрансфекции с вирусом-хелпером клеток 293.
Принцип основан на транскомплементации между "минивирусом" (вирус-хелпер), экспрессирующим область L5 или области Е4 и L5, и рекомбинантным вирусом, лишенным, по крайней мере, Е3, Е4 и L5.
Эти вирусы были получены или путем лигирования in vitro или после рекомбинации in vivo по следующим стратегиям.
(I) ДНК вируса Ad-d1324 (Thimmappaya et al., Cell 31 (1982) 543) и плазмиду р2, расщепленные BamHI, сначала лигировали in vitro, затем котрансфицировали с плазмидой-хелпером pITRL5-Е4 (пример 6.2) в клетки 293.
(II) ДНК вируса Ad-d1324, расщепленную ЕсоРI и плазмиду р2, расщепленную BamHI, котрансфицировали вместе с плазмидой рITRL-5-Е4 в клетки 293.
(III) ДНК аденовируса Ad5 и плазмиду р2, расщепленные BamHI, лигировали, затем котрансфицировали с плазмидой рITRL5-Е4 в клетки 293.
(IV) ДНК аденовируса Ad5, расщепленную ЕсоRI и плазмиду р2, расщепленную BamHI котрансфицировали с рITRL5-Е4 в клетки 293.
Стратегии (I) и (II) позволяют получать рекомбинантный аденовирус, лишенный областей Е1, Е3, L5 и Е4; стратегии (III) и (IV) позволяют получать рекомбинантный аденовирус, лишенный областей Е3, L5 и Е4. Разумеется, ДНК рекомбинантного вируса, лишенная области Е1, но выражающая какой-нибудь трансген, может использоваться вместо ДНК вируса Ad-d1324 по стратегиям (I) или (II) в целях получения рекомбинантного вируса, лишенного областей Е1, Е3, L5 и Е4, и экспрессирующего указанный трансген.
Методики, описанные здесь выше, могут также использоваться с вирусом-хелпером, несущим только область L5, при использовании клеточной линии, способной экспрессировать области Е1 и Е4 аденвируса, такой, которая описана в примере 4.
С другой стороны, также можно использовать дополняющую линию, способную экспрессировать области Е1, Е4 и L5, так, чтобы полностью освободиться от вируса-хелпера.
После трансфекции вирусные продукты извлекают, амплифицируют и очищают при условиях, описанных в примере 5.







Изобретение относится к новым вирусным векторам. Представлен дефектный рекомбинантный аденовирусный вектор, в котором ген Е1 и, по меньшей мере, один из генов Е2-Е4, L1-L5 являются нефункциональными. Предпочтительно вектор получают от аденовируса собак серотипа CAV1 или CAV2 штамма Manhattan или А26/61 (АТСС VR-800). Вектор включают в фармацевтическую композицию, пригодную для генной терапии. 2 с. и 28 з.п.ф-лы, 10 ил.
| US 4920211, 24.04.1990. |

