Изобретение относится к новым анти-ЭФРР антителам и фрагментам антител, предпочтительно одноцепочечным Fv (оцFv), которые могут быть получены из фаг-антитело библиотек, сконструированных из клеток иммунизированных млекопитающих, предпочтительно мыши. Фрагменты антител, выделенные из фаг-антитело библиотек, могут быть сконструированы с целью создания частично гуманизированной полной молекулы антитела. Подобные химерные анти-ЭФРР антитела содержат постоянные области иммуноглобулинов человека и могут быть использованы так же, как и их фрагменты, в качестве средств диагностики и терапии опухолей человека.
Кроме того, в изобретении показано, что применение фаг-антитело библиотек является альтернативным и более гибким способом выделения антител из иммунизированных млекопитающих по сравнению со стандартной гибридомной технологией.
Изобретение относится также к лекарственным формам, содержащим указанные антитела и их фрагменты и предназначенные для лечения таких опухолей, как меланома, глиома или карцинома. Указанные антитела или их фрагменты могут быть также использованы в диагностических целях для обнаружения и оценки упомянутых опухолей in vitro и in vivo.
В описании применяются некоторые технические термины, определение которых приводится ниже.
"КОи" (каркасные области) означает четыре субрайона из вариабельных областей в тяжелой или легкой цепи, поддерживающих три ОКО.
"ОКО" (определяющие комплементарность области) относится к трем субрайонам вариабельных областей тяжелой или легкой цепи, включающих гипервариабельные последовательности и образующих петельные структуры, которые в первую очередь ответственны за прямой контакт с антигеном.
Термин "химерные" или гуманизированные антитела означает антитела, содержащие постоянные области, источником которых является человек, и вариабельные области (в том числе ОКО), происходящие из отличного от человека источника, например мыши.
Термин "гуманизированные" или полностью гуманизированные антитела относится к антителам, содержащим постоянные области КОи, источником которых служит человек, при этом ОКО происходит из отличного от человека источника.
"ЭФР" и "ЭФРР" означает эпидермальный фактор роста и его рецептор.
"ПЦР" означает полимеразноцепную реакцию.
"оцFv" означает одноцепочечный Fv, являющийся фрагментом антитела.
"VL" означает вариабельную область легкой цепи.
"VK" означает вариабельную область легкой каппа-цепи.
"VH" означает вариабельную область тяжелой цепи.
ФБС означает фосфатный буферный солевой раствор.
ФТС означает фетальная телячья сыворотка.
ССРХ означает сбалансированный солевой раствор Хенка.
ФИТЦ означает флюоресцеинизотиоцианат.
СКК означает смешанная клеточная культура.
Эпидермальный фактор роста (ЭФР) является полипептидным гормоном, характеризующимся митогенностью по отношению к эпидермальным и эпителиальным клеткам. При взаимодействии ЭФР с чувствительными клетками происходит связывание с мембранными рецепторами (ЭФРР). ЭФРР является трансмембранным гликопротеином примерно в 170 кД и генным продуктом протоонкогена c-erb-B.
MAb 425 является мышиными моноклональными антителами, созданными против хорошо известной клеточной линии A431 карциномы человека (ATCC CRL 1555) и связывающимися с полипептидным эпитопом внешнего домена ЭФРР человека с подавлением связывания ЭФР. MAb 425 (ATCC HB 9629), как найдено, передают клеточную цитотоксичность in vitro и подавляют рост клеток опухоли, происходящих из клеточных линий эпидермоидной и колоректальной карциномы in vitro (Kodeck и др., Cancer Res. 1987, 47, 3692). Гуманизированные и химерные варианты MAb 425 раскрыты в патенте WO 92/15683.
В последние несколько лет появились описания способов, которыми могут быть получены функциональные фрагменты антитела в эукариотных клетках-хозяевах, например E.coli (Skerra и Pliickthun, Science 1988, 240, 1038; Better и др., Science 1988, 240, 1041). Такие фрагменты включают Fv фрагмент и Fab фрагмент, причем Fv представляет особый интерес. Одноцепочечные Fv (в которых VL и VH цепи связаны друг с другом) описаны также в работах: Bird и др. Science 1988, 242, 423; Huston и др., Proc. Natl. Acad. Sci. США 1988, 85, 5879.
Технология с применением фаг-антитело библиотек является альтернативной гибридомной технологии выделения антител из иммунизированных животных. Гибридомная технология состоит в иммортализации клеток, продуцирующих антитела. Фаг-антитело технология заключается в иммортализации генов, кодирующих антитела (Winter G. и Milstein C., Nature, 1991, 349, 293). В фаг-антитело технологии гены вариабельной области легкой цепи (VL) и гены вариабельной области тяжелой цепи (VH) антитела подвергают ПЦР-амплификации, вариабельные области неупорядоченно соединяют и экспрессируют в виде фрагментов антитела на поверхности фаговых частиц, после чего библиотеки фаговых антител отбирают с выявлением антител, связывающихся с представляющими интерес антигенами.
Гибридомная технология оказалась чрезвычайно успешной в выделении мышиных моноклональных антител при условии возможности создания сильной иммунной реакции в селезенке животного. К примеру, мышиные MAb к рецептору эпидермального фактора роста (ЭФРР) человека выделены из селезенки мыши, иммунизированной внутрибрюшинно клетками A431 опухоли человека (Murthy и др., Arch. Biochem. Biophys. 1987, 252, 549). Потенциальное преимущество фаг-антитело технологии над гибридомной технологией заключается в возможности использования в качестве исходного продукта фактически любого источника экспрессирующих антитело клеток и возможности быстрого отбора большого числа различных антител. Другое преимущество фаг-антитело технологии состоит в том, что гены, кодирующие вариабельные области представляющих интерес антител, оказываются уже клонированными и непосредственно доступными для дальнейшей обработки методами генной инженерии.
Имеется сообщение о превращении противостолбнячного токсоидного Fab фрагмента, выделенного из фаг-антитело библиотеки, в полную молекулу антитела (Bender и др., Hum. Antibod. Hybridomas. 1983, 4, 74).
Последние десять лет в качестве альтернативы активной иммунизации с целью создания моноклональных антител (mAb) к широкому спектру антигенов из как человечьих, так и мышиных систем применяется иммунизация in vitro (например, Vaux D. J. T. , Helenius A., Melliman I., Nature 1988, 336, 36; Gathuru J. K. и др., J. Immunol. Methods, 1991, 137, 95; Bornebaek C.A.K., Immunol. Today, 1988, 9, 355). Преимущества такого подхода заключаются в том, что требуются очень небольшие количества антигена, и в том, что способ применим в создании человечьих гибридом. Однако образование IgM антител с плохим сродством и трудность иммортализации человечьих лимфоцитов после in vitro иммунизации - это постоянные проблемы, связанные с данной технологией.
Новый путь получения антител состоит в ПЦР амплификации репертуара генов вариабельных областей тяжелой (VH) и легкой (VL) цепей, которые затем неупорядоченно рекомбинируют и зкспрессируют в виде библиотек фагового отображения (7-9), Гены вариабельной области антитела клонируют и сливают с минорным белком оболочки (ген 3) в виде одноцепочного Fv фрагмента (оцFv) (10). Фаговая частица отражает на своей поверхности фрагмент антитела, которое может быть отобрано пеннингом с использованием способности антитела к связыванию. Данная технология обладает тем преимуществом, что неупорядоченная рокомбинация V-генов может привести к образованию новых пар оснований с неизвестной специфичностью и сродством, которые могут быть отобраны естественными процессами. Более того, такой подход делает возможным применение простых или in vitro иммунизированных лимфоцитов, источником которых является мышь или человек.
Предшествующие попытки получения к ЭФРР иммунизацией in vitro мышиных B-клеток и применением гибридомной технологии привели к антителам, характеризующимся низким сродством и перекрестной реакционностью. Для преодоления указанных недостатков использована комбинация иммунизации in vitro с последующим проведением ПЦР клонирования.
Таким образом, цель изобретения заключается в создании антител и фрагментов антител, обладающих высоким сродством к ЭФР-рецепторам и которые могут быть получены перспективной технологией, раскрываемой выше и ниже.
В настоящем изобретении проведено сравнение мышиных анти-ЭФРР антител, выделенных из трех различных фаг-антитело библиотек, с мышиными MAb (425), выделенными применением стандартной гибридомной технологии (Murthy и др., Arch. Biochem. Biophys. 1987, 252, 549; Kettleboroug и др., Protein Eng. 1991, 4, 773). Библиотеки приготовлены не только из селезенки иммунизированной мыши, но также из обезвоженных лимфатических узлов иммунизированной мыши и из in vitro иммунизированных мышиных клеток. Два одноцепочных Fv (оцFv), выделенные из библиотек, обработаны методами генной инженерии с созданием химерных полных молекул антитела, в которых мышиные вариабельные области соединены с человечьими постоянными областями.
Если более подробно, то настоящее изобретение относится к анти-ЭФРР одноцепочным Fv, получаемых из фаг-антитело библиотек, сконструированных из клеток, предпочтительно из клеток селезенки или обезвоженных лимфатических узлов иммунизированного млекопитающего, предпочтительно мыши или из in vitro иммунизированных клеток. В принципе, изобретение не ограничено только оцFv, но изобретением охватываются также и другие анти-ЭФРР фрагменты антитела, например: Fab или F(ab')2.
Некоторые оцFv по изобретению характеризуются четко определенными ДНК и аминокислотными последовательностями. Таким образом, другая цель изобретения заключается в создании одноцепочного Fv фрагмента, в котором вариабельные области тяжелой и легкой цепей состоят из ДНК и/или аминокислотной последовательности, выбранной из одной из последовательностей тяжелой и легкой цепей, представленных Последовательностями NN 1 - 32, приведенными в конце описания, предпочтительно последовательностями на фиг. 5 - 8.
Вследствие того что во многих случаях для диагностических и терапевтических целей могут быть использованы только полностью функциональные полные антитела, для изобретателей было интересно связать переменные области из одноцепочных Fv с постоянными областями иммуноглобулинов человека с образованием полных частично или полностью гуманизированных анти-ЭФРР антител.
Таким образом, еще одна цель изобретения состоит в создании полного анти-ЭФРР антитела, сконструированного из ДНК последовательностей, происходящих из фрагментов антитела согласно выше- и нижеприведенному определениям или определению, приведенному в формуле изобретения, и из ДНК последовательностей, происходящих из постоянных областей иммуноглобулинов человека, при этом в рекомендуемом воплощении изобретения тяжелая цепь состоит из аминокислотной последовательности гамма-1 цели, а легкая цепь состоит из аминокислотной последовательности каппа-цепи.
Согласно настоящему изобретению анти-ЭФРР оцFv выделяют применением технологии с использованием фаг-антитело библиотек. Таким образом, изобретение относится к способу получения анти-ЭФРР одноцепочного Fv, включающему следующие стадии:
(I) выделение РНК из клеток иммунизированного млекопитающего, предпочтительно мышиных клеток;
(II) синтез первой нити кДНК;
(III) амплификация VH и VK генов в кДНК из иммунизированных клеток;
(IV) клонирование указанных генов вместе с приемлемыми сайтами рестрикции в фагемидный вектор;
(V) трансформация клеток прокариотов лигирующими смесями;
(VI) отбор фаговых библиотек для фаговых антител, направленных на ЭФРР, применением очищенного ЭФРР, и
(VII) продуцирование целевого одноцепочного Fv в прокариотических клетках-хозяевах, предпочтительно E.coli.
Кроме того, цель изобретения заключается в раскрытии способа получения полного анти-ЭФРР антитела путем клонирования ДНК, кодирующей переменные области фрагментов анти-ЭФРР антитела, продуцируемых, как указано выше или по определению, приведенному в формуле изобретения, в, по меньшей мере, один эукариотический экспрессионный вектор, содержащий геномную ДНК, кодирующую постоянные области иммуноглобулинов человека, трансформацией эукариотических клеток указанным вектором(ами) с последующей экспрессией и выделением антитела.
Анти-ЭФРР оцFv и прежде всего полные анти-ЭФРР антитела могут быть использованы в диагностике и терапии опухолей человека. Таким образом, изобретение относится к лекарственным формам, содержащим анти-ЭДРР одноцепочный Fv или полное анти-ЭФРР антитело по вышеприведенному определению или по определению, приведенному в формуле изобретения.
Результаты и преимущества настоящего изобретения могут быть суммированы следующим образом.
Из фаг-антитело библиотек выделены новые мышиные анти-ЭФРР антитела. Новые антитела представлены по меньшей мере четырьмя различными VH подгруппами и четырьмя различными VK подгруппами (Kabat и др., "Представляющие иммунологический интерес последовательности белков", 5-ое изд., Деп. здоровья и службы человека США, Бетседа, 1991). Полученные антитела показали образование пар оснований и последовательностей отличных от мышиных MAb, выделенных применением гибридомной технологии. Мышиное MAb 425 имеет VH 2b и VK 4 пары, которые не обнаруживаются в фаговых антителах.
VH для оцFv L 3 11 D обладает наиболее высоким процентом идентичности с 425 VH (84,9%). Большая часть отличий приходится на ОКО. VK для оцFv S4 2D обладает наиболее высоким процентом идентичности с 425 VK (83,2%). И в этом случае большая часть отличий падает на ОКО, в частности ОКОЗ. В данном изобретении из фаг-антитело библиотек выделены разнообразные новые анти-ЭФРР антитела, и все эти антитела отличаются от 425 MAb по меньшей мере двумя оцFv фрагментами, распознающими расположенный на ЭФРР эпитоп, отличный от эпитопа, распознаваемого 425 MAb. В этом отличие от имеющегося сообщения, согласно которому антитела, выделенные из комбинированных библиотек, как утверждается, очень похожи на антитела, выделенные гибридомной технологией (Caton и Koprowski, Proc. Natl. Acad. Sci. США 1990, 87, 6450).
Из трех фаг-антитело библиотек наилучшей библиотекой с точки зрения числа стадий отбора, необходимых для получения антител с высоким сродством, и с точки зрения различий выделенных и обладающих высоким сродством антител, является библиотека, образованная из обезвоженного лимфатического узла. Лимфатические узлы выбраны при построении фаг-антитело библиотек в качестве источника РНК по двум причинам. Во-первых, предшествующей работой показано, что B-клетки, продуцирующие IgG антитела, с высоким сродством получают в большем количестве из подколенных лимфатических узлов после иммунизации через подушечку на лапке, чем из селезенки после иммунизации через брюшину (Venn и Dresser, J. Immunol. Methods 1987, 102, 95). Во-вторых, считается, что обезвоженные лимфатические узлы являются хорошим источником для выделения противоопухолевых антител человека. Таким образом, выделение мышиных анти-ЭФРР антител из подколенного лимфатического узла иммунизированной через подушечку на лапке мыши является моделью для выделения анти-ЭФРР антител человека из подмышечных лимфатических узлов больного раком молочной железы. Показана возможность получения библиотек хорошего размера из небольших количеств материала лимфатических узлов с последующим выделением из библиотек, обладающих высоким сродством антител.
Хотя из всех трех фаг-антитело библиотек были выделены мышиные анти-ЭФРР антитела, совсем не очевидно, что любое из вновь выделенных антител будет отличаться более высоким сродством по сравнению с мышиным 425 MAb, выделенным с помощью гибридомной технологии. Первоначальные анализы показывают, что происходящие из фаг-антитела оцFv оказываются с ЭФРР лучше, чем оцFv, сконструированные из 425 MAb (фиг. 2). В других экспериментах с молекулами полного химерного антитела одно из химерных антител (S4 2D) показало сродство к ЭФРР, равное сродству химерного 425 антитела. Второе химерное антитело (L3 11D) обладает в четыре раза более низким сродством по сравнению с химерным 425 антителом (фиг. 4). Данные по связыванию, полученные с применением оцFv, столь противоречивы, вероятно, вследствие присутствия в препаратах оцFv смесей мономеров и димеров (Criffiths и др., EMBO J. 1993, 12, 725). Напротив, трудно ожидать, что химерные IgG антитела будут образовывать димеры, и, как показано, химерные L 3 11D и S4 2D антитела, имеющие именно тот размер, который следует ожидать для двухвалентных, мономерных химерных IgG антител. Однако анализ аффинно очищенных 425, L 3 11D и S4 2D оцFv показал, что указанные препараты оцFv содержат мономерные, димерные и иные полимерные формы. Кроме того, относительные количества мономерных и полимерных форм меняется для каждого оцFv 425 оцFv имеет самый низкий процент димерных форм. Предположительно, димерные и особенно более крупные полимерные формы отличаются более сильным связыванием с очищенным ЭФРР по сравнению с мономерной формой. По-видимому, 425 оцFv характеризуются более слабой тенденцией к димеризации, чем вновь выделенные оцFv.
Хотя экспрессия фрагментов антител на поверхности фаговых частиц создает основу эффективного способа для быстрого отбора антител с целевой специфичностью, тем не менее ни фаговые антитела, ни сами фрагменты антител (оцFv или Fab) вряд ли станут целевыми конечными продуктами. К тому же показано, что мышиные оцFv, выделенные из фаговых библиотек, могут быть легко превращены в молекулы полного антитела. В этом случае мышиные вариабельные области соединяют с постоянными областями человека, в результате чего создаются частично гуманизированные химерные антитела.
Эти результаты показывают возможность применения фаг-антитело технологии для получения анти-ЭФРР фрагментов антитела от иммунизированной мыши. Из фрагментов антитела могут быть сконструированы молекулы полного антитела с необходимыми постоянными областями. В отдельных случаях для получения моноклональных антител от мыши в качестве выбранного способа может быть использована и гибридомная технология. Если при этом оказывается доступным в высшей степени иммуногенный антиген и, если могут быть получены несколько адекватных клеточных линий, продуцирующих одно или несколько противоантигенных антител, в этом случае, видимо, нет причин обращаться к фаг-антитело технологии. Однако, если в создании обладающих высоким сродством антител преимущества дает особая методика иммунизации, например, инъекцией в подушечку лапки или, если необходимо большое количество антител против разнообразных эпитопов на антигене, или, если необходимы антитела, направленные против очень точного и возможно менее иммуногенного эпитопа, в этом случае выбранным способом может служить фаг-антитело технология. Кроме того, если предстоит дополнительная обработка методами генной инженерии антител, тогда преимущества фаг-антитело технологии проявляется в том, что гены оказываются уже клонированными.
Настоящий подход, сочетающий in vitro иммунизацию конкретным антигеном с технологией ПЦР-клонирования, позволяет создавать оцFv фрагменты, реагирующие с ЭФРР и не вступающие в перекрестные реакции с другими антигенами. Методика иммунизации, о которой сообщается здесь, зависит от состояния антигена, который не растворим, но является препаратом мембранного везикула, и от самой культуральной среды, в которой отсутствует ФТС. Об обоих методиках сообщается, как о средстве повышения эффективности иммунизации in vitro за счет доступности антигена для представляющих антиген клеток (например, Brans P. и др., J. Immunol. Methods, 1987, 98, 11).
Результаты, полученные применением СКК, согласуются с известными работами (например: Borrebaek C. A. K. и Moller S.A., J. Immunol., 1986, 136, 3710; Moller S.A. и Borrebaeck C.A.K. в Borrebaek C.A.K. (ред.) "Иммунизация in vitro в гибридомной технологии". Эльзевир Сайенс Паблишерс Б.В., Амстердам 1988, стр. 3), в которых для усовершенствования процесса иммунизации in vitro в качестве источника лимфокинов предлагается применение надосадочной жидкости от СКК. Препараты мембранных везикул следует рассматривать как полиантиген, поскольку в таких везикулах присутствуют многие различные антигенные детерминанты. По этой причине кажется вероятным, что они будут создавать определенный уровень поликлональной активации. Все это нами исключено, поскольку специфичная анти-ЭФРР реакция абсолютно отлична от реакции, полученной после применения стандартного поликлонального активатора.
Вместо иммортализации B-клеток после иммунизации in vitro была применена молекулярная стратегия иммортализации VH и VL генов антитела. Указанные фрагменты моноклонального антитела были экспрессированы и продуцированы в бактерии. Система фагового отображения является эффективным методом выделения фрагментов антитела, направленных против специфичных антигенов. Присутствие терминирующего кодона между фрагментом антитела и g3p белком оболочки допускает переключение между поверхностным отображением и секрецией в виде растворимого оцFv фрагмента применением супрессорных несупрессорных штаммов (Hogoenboom и др., Nucl. Acids Res. 1991, 19, 4133).
Вследствие увеличения специфичной реакции и уровня мРНК в стимулированных in vitro антигеном B-клетках способ иммунизации in vitro вносит свой вклад в выделение фрагментов антитела с высокой специфичностью к антигену. После двух циклов отбора 100% клонов оказались положительными для связывания ЭФРР. Напротив, клоны, полученные способами in vivo иммунизации, были на 100% положительными после четырех циклов отбора (Kettleborough и др., патент EP 94/104160 и Eur. J. Immunol. 1994, 24, 952).
Применение библиотек фагового отображения из простых генов антитела могло бы позволить получать фрагменты специфичного антитела без иммунизации или после иммунизации in vitro, т.е. простым, быстрым и экономичным путем.
Биологические материалы и общие методы
Микроорганизмы, клеточные линии, плазмиды, фагемиды, промоторы, маркеры устойчивости, точки инициации репликации или иные фрагменты векторов, упоминаемые в данной заявке, являются коммерчески или в целом каким-то иным путем доступны. При условии, что в заявке не приводится никакой другой информации, все они использованы в виде примеров и несущественны по изобретению, и могут быть заменены другими приемлемыми инструментами и биологическими материалами соответственно.
Для клонирования оцFv и для продуцирования оцFv белков рекомендуются бактериальные хозяева. Примерами таких хозяев является E.coli и Bacillus.
Для продуцирования полных анти-ЭФРР антител по изобретению рекомендуются такие эукариотичные хозяева, как например: COS, CHO или дрожжи.
Методики, имеющие существенное значение для изобретения, подробно рассматриваются в описании. Другие, не раскрываемые подробно методики, соответствуют известным стандартным методам, хорошо известным специалистам или более подробно раскрываемым в цитируемых ссылках, заявках на патенты и справочной литературе.
Краткие пояснения к диаграммам
Фиг. 1 - аминокислотные последовательности оцFv, выделенных из фаг-антитело библиотек. (A) оцFv из библиотеки лимфатического узла. (B) оцFv из библиотеки селезенки. Указаны определяющие комплементарность области (ОКО) и каркасные области (КО).
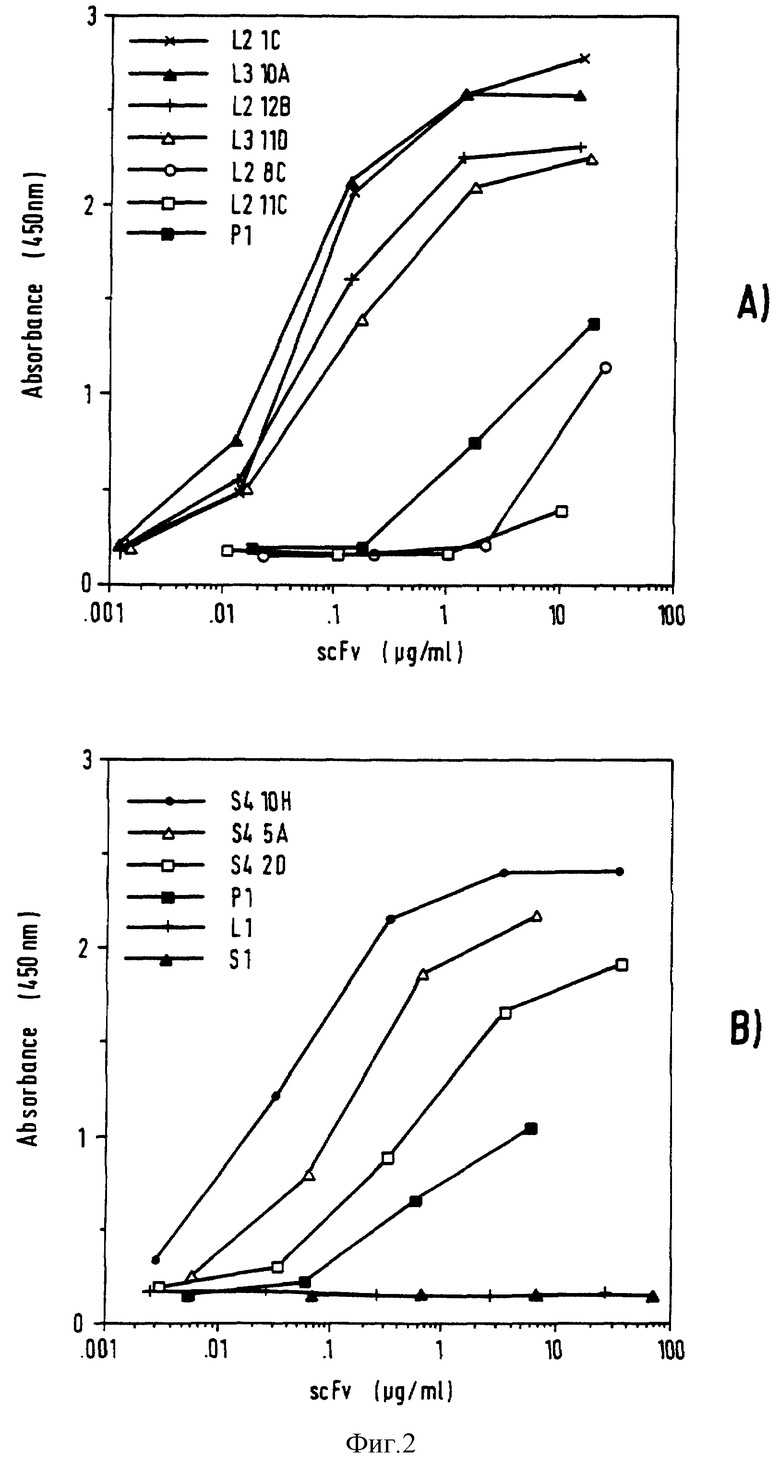
Фиг. 2 - связывание оцFv с ЭФРР. Определяют концентрации оцFv в бактериальных надосадочных жидкостях и оцFv анализируют методом ФИСА (ELISA) на связывание с очищенным ЭФРР. (A) оцFv из библиотеки лимфатического узла. (B) оцFv из селезенки библиотеки. P1 (положительный контроль) относится к оцFv происходящим из 425 MAb. L1 и S1 (отрицательный контроль) относится к несвязывающимся оцFv из предварительно отобранных библиотек лимфатического узла и библиотек селезенки.
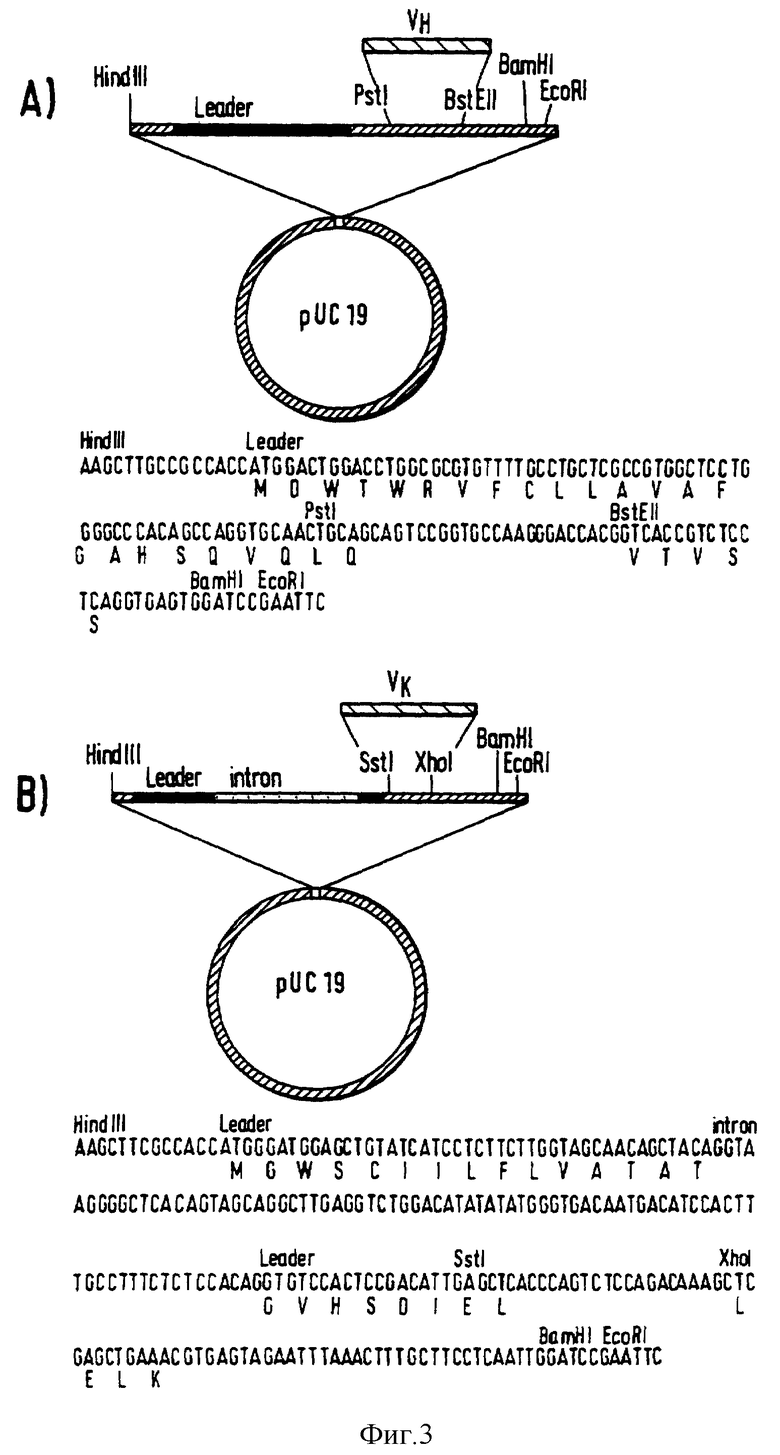
Фиг. 3 - промежуточные вектора, применяемые для реконструкции вариабельных областей для экспрессии в клетках млекопитающего. (A) VH вектор. (B) VL вектор.
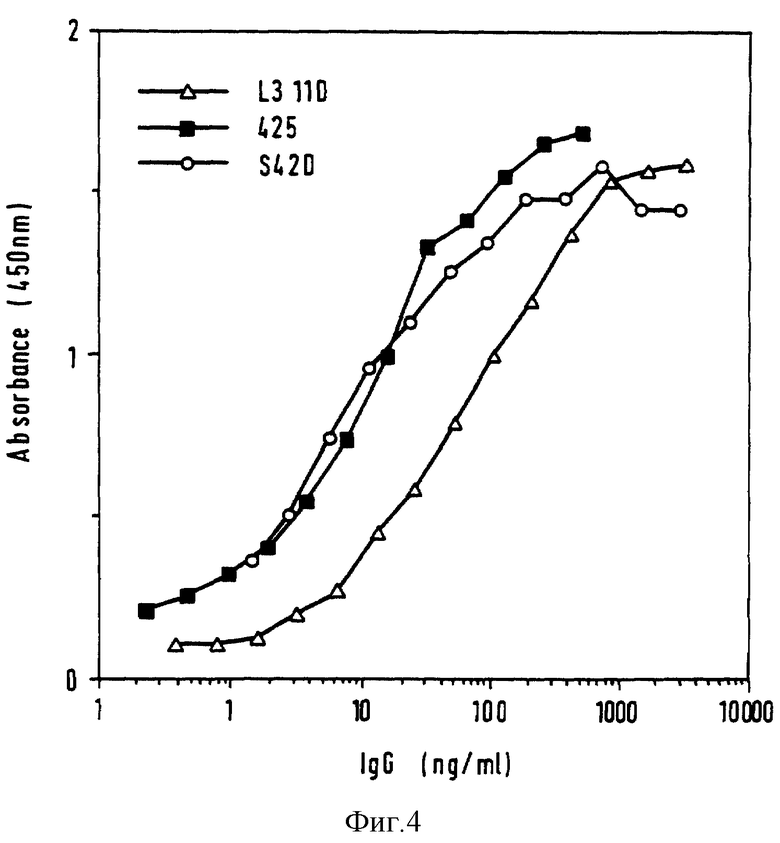
Фиг. 4 - связывание химерных полных антител с ЭФРР. Методом ФИСА определяют концентрации антител в надосадочных жидкостях от COS клеток и антитела испытывают методом ФИСА на связывание с очищенным ЭФРР.
Фиг. 5 - ДНК и аминокислотная последовательность оцFv N L211C. (A) Легкая цепь. (B) Тяжелая цепь.
Положение аминокислот:
(A) ФР.-1: 1-23, ОКО-1: 24-34,
ФР.-2: 25-49, OКO-2: 50-56,
ФР.-3: 57-88, ОКО-3: 89-97,
ФР.-4: 98-109.
(B) ФР.-1: 1-30, ОКО-1: 31-35,
ФР.-2: 36-49, ОКО-2: 50-66,
ФР.-3: 67-98, ОКО-3: 99-108,
ФР.-4: 109-119.
Фиг. 6 - ДНК и аминокислотная последовательность оцFv N L2 12B. (A) Легкая цепь. (B) Тяжелая цепь.
Положение аминокислот:
(A) ФР.-1: 1-23, ОКО-1: 24-38,
ФР.-2: 39-49, ОКО-2: 50-56,
ФР.-3: 57-88, ОКО-3: 89-97,
ФР.-4: 98-109.
(B) ФР.-1: 1-30, ОКО-1: 31-35,
ФР.-2: 36-49, ОКО-2: 50-66,
ФР.-3: 67-98, ОКО-3: 99-108,
ФР.-4: 109-119.
Фиг. 7 - ДНК и аминокислотная последовательность оцFv N L3 11D. (A) Легкая цепь. (B) Тяжелая цепь.
Положения аминокислот во ФР. и ОКО соответствуют положениям, приведенным для фиг. 6.
Фиг. 8 - ДНК и аминокислотная последовательность оцFv N S4 2D.
(A) Легкая цепь. (B) Тяжелая цепь.
Положения аминокислот:
(A) ФР.-1: 1-23, ОКО-1: 24-35,
ФР.-2: 36-50, ОКО-2: 51-57,
ФР.-3: 58-89, ОКО-3: 90-98,
ФР.-4: 99-110.
(B) ФР.-1: 1-30, ОКО-1: 31-35,
ФР.-2: 36-49, ОКО-2: 50-66,
ФР.-3: 67-98, ОКО-3: 99-107,
ФР.-4: 108-118.
Приведенные на фиг. 5-8 последовательности указаны также в Перечне последовательностей, являющихся частью описания данного изобретения.
Подробное описание изобретения
(1) Конструирование и скрининг фаг-антитело библиотек
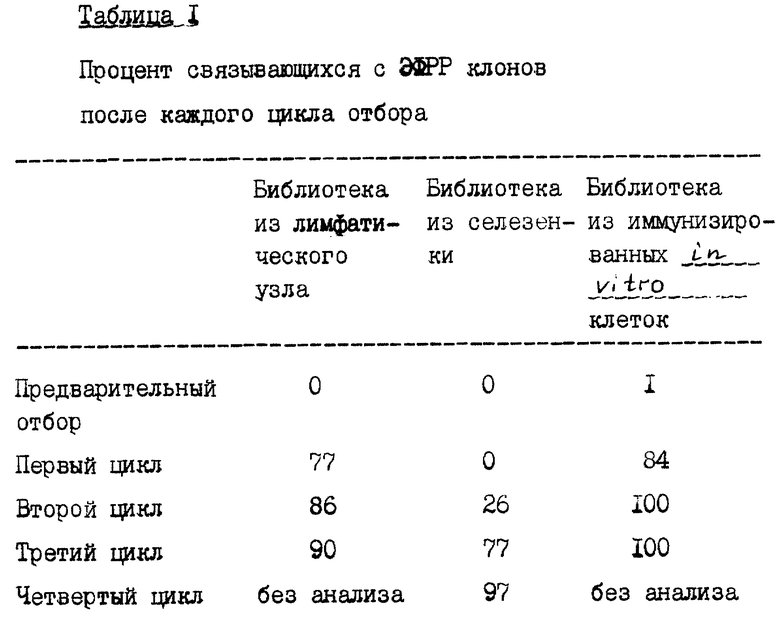
Сконструированы три фаг-антитело библиотеки: одна из селезенки мыши, иммунизированной клеточной линией A431 карциномы человека (8,8•105 элементов), одна из подколенного лимфатического узла мыши, иммунизированной в подушечку лапки очищенным ЭФРР (6,5•106 элементов) и одна из мышиных лимфоцитов, иммунизированных in vitro A431 везикулами (1,1•105 элементов) (подробности конструирования A431 везикулов и иммунизации in vitro приведены в Примерах 1, 2). Перед отбором по меньшей мере 46 клонов из каждой библиотеки анализируют методом BstNI фингерпринтинга (Clackson и др., Nature 1991, 352, 624) с целью определения отличий в наборах детерминант. Наблюдается широкий спектр типов переваривания. Также перед отбором оцFv из 96 клонов из каждой библиотеки оценивают методом ФИСА на связывание с ЭФРР. Ни один оцFv из библиотеки селезенки и лимфатического узла не связывается с ЭФРР. Один из оцFv из иммунизированной in vitro библиотеки связывается с ЭФРР, После одного цикла отбора с применением покрытых ЭФРР иммунопробирок наблюдается явное обогащение оцFv, связывающимися с ЭФРР, в случае библиотеки лимфатического и в случае библиотеки из иммунизированных in vitro клеток. Дня обнаружения каких-либо оцFv, связывающихся с ЭФРР, из библиотеки селезенки требуется второй цикл отбора. После третьего цикла отбора большая часть оцFv из библиотеки лимфатического узла и библиотеки из иммунизированных in vitro клеток оказываются положительными на связывание с ЭФРР. После четвертого цикла отбора в случае библиотеки селезенки большая часть оцFv оказываются положительными на связывание с ЭФРР (табл. 1).
(2) Анализ последовательности связывающихся с ЭФРР клонов
После каждого цикла отбора оцFv вставки из связывающихся с ЭФРР клонов анализируют методом BstNI фингерпринтинга (Clackson и др., Nature 1991, 352, 624). В результате становится очевидным, что происходит обогащение для определенных типов переваривания. Клоны с разными BstNI фингерпринтами выбраны после второго и третьего циклов отбора в случае библиотеки из лимфатического узла и после третьего и четвертого циклов отбора в случае библиотеки из селезенки для проведения ДНК-секвенирования VH и VL генов. Клоны с более поздних циклов отбора анализируются по той причине, что, как ожидается, на поздних этапах отбора появляются антитела, обладающие более высоким сродством (Clackson и др., Nature 1991, 352, 624).
Шестнадцать клонов из библиотеки лимфатического узла подвергнуты секвенированию с получением шести различных оцFv (фиг. 1). Пять из них являются парными уникальным VH и VL. Шестой клон является вариантом ранее обнаруженной VH с шестью изменениями в аминокислотах, пять из которых приходятся на каркасную область (ФР.-1). Два из таких изменений могут быть отнесены за счет применения вырожденного VHIBACK-SFI праймера (Hoogenboom и др., Nucl. Acids Res. 1991, 19, 4133). Прочие могут быть результатом ошибок в ПЦР. VH разделены на две подгруппы: VH2b и VH3d, в то время как VK охватываются четырьмя подгруппами: VK3, VK4, VK5 и VK6 (Kabat и др. "Представляющие иммунологический интерес последовательности белков", 5-ое изд. Деп. Здоровья и службы человека США, Бетесда 1991). Десять индивидуальных клонов из библиотеки селезенки подвергнуты секвенированию, в результате выявлены четыре разных оцFv. Три из них являются парными уникальным VH и VK, в то время как четвертый был аналогичен одной из предшествующих пар, но только с двумя аминокислотными отличиями в VН, одно из которых обнаруживается в определяющей комплементарность области (ОКО-2), и двумя отличиями в аминокислотах в VK. Подразделение на подгруппы выявило VH из подгрупп VH2a, VH2c и VH3d и VK из подгрупп VK3 и VK4. Сравнение оцFv из библиотек лимфатического узла и селезенки выявило всего лишь один оцFv, являющимся общим для обеих библиотек (оцFv L3 10A/оцFv S4 10H) (фиг. 1). При анализе методом ФИСА указанный клон, видимо, прочно связывается с ЭФРР. Хотя и принимались особые меры, чтобы исключить всякое перекрестное загрязнение между библиотеками, тем не менее трудно исключить небольшое загрязнение в случае клона, прочно связывающего ЭФРР. Однако, с учетом инбредной природы линии Balb/c мышей возможно, что в двух различных библиотеках возникают одинаковые оцFv.
(3) Анализ сродства и специфичности связывания с ЭФРР
На основе хорошего связывания с антигеном и отличий в ДНК последовательностях для дополнительного анализа отобрано несколько оцFv, происходящих из библиотек лимфатического узла и селезенки. Отобранные оцFv анализируют методом ФИСА на связывание с очищенным ЭФРР, связывание с отличными от ЭФРР антигенами и связывание с клеточными линиями опухолей, экспрессирующими и не экспрессирующими ЭФРР. В качестве положительного контроля готовят оцFv из мышиных 425 MAb (P1), в качестве отрицательного из фаговых антител, выделенных из библиотек лимфатического узла и селезенки для проведения отбора (L1 и S1 соответственно). Концентрацию оцFv определяют сравнением разбавлений подлежащего анализу оцFv с разбавлениями очищенного оцFv известной концентрации в Вестерн-блотинге.
оцFv анализируют методом ФИСА на связывание с очищенным ЭФРР, и полученные результаты откладывают на графике (фиг. 2). Появляется возможность оценить оцFv с точки зрения их связывания с ЭФРР. Полученная оценка воспроизводима в разных опытах. К наиболее прочно связывающимся с ЭФРР относятся следующие оцFv: L2 1C и L3 10A из библиотеки лимфатического узла и S4 10H из селезеночной библиотеки. Как указано ранее, оцFv L3 10A и S4 10H имеют одинаковые ДНК последовательности. оцFv (S4 5A), очень похожий на оцFv S4 10H, с двумя изменениями в аминокислотах в VH и двумя - в VK получает более низкую, чем у S4 10H, оценку. Напротив, различия, наблюдаемые между L2 12B и L3 11D, видимо, не оказывают значительного влияния на связывание. Из выделенных оцFv только два (L2 8C и L2 11C), видимо, связываются хуже, чем оцFv 425.
Методом ФИСА анализируют оцFv на связывание с нанесенными на пластик набором неродственных белков (овальбумин, лизолим куриного яйца, цитохром Ц, глицеральдегид-3-фосфатдегидрогеназа, СВА альбумин и БСА). Ни один из оцFv не дал сигнал выше фона.
Методом ФИСА анализируют оцFv на связывание с тремя клеточными линиями опухоли. Клеточные линии A431 и MDA MB 468 являются ЭФРР-несущими клетками опухоли, выделенными из вульвы и молочной железы соответственно. Клеточная линия SK-MEL-23 является ганглиозиднесущей клеточной линией меланомы, которая включена в качестве отрицательного контроля. Из десяти проанализированных оцFv только четыре связываются как с очищенным ЭФРР, так и с ЭФРР-несущими клетками опухоли (L2 12B, L3 11D, L2 11C и S4 2D, фиг. 5-8). Никакого связывания с SK-MEL-23 клетками не обнаружено. Имеется несколько возможных объяснений этому неожиданному результату. Одно из них заключается в том, что ЭФРР, используемый для иммунизации, отбора и ФИСА, является секретируемым родственным ЭФРР белком (Weber и др., Science 1984, 224, 294). Этот белок имеет у C-окончания 17 дополнительных аминокислот (Giinther и др., J. Biol. Chem., 1990, 265, 22082). Методом ФИСА анализируют оцFv на связывание с пептидом из таких 17 аминокислот и никакого связывания не наблюдают. Возможно, что секретируемый родственный ЭФРР белок и ЭФРР на поверхности клетки опухоли имеют отличия в конформации и гликозилирования.
Для дальнейших исследований связывания с клетками опухоли три оцFv (L2 11A, L3 11D и S4 2D) очищают и анализируют на связывание с A431 клетками опухоли проточной цитометрией. В качестве положительного контроля применяют 425 оцFv. Из трех испытанных оцFv только L3 11D и S4 2D связываются с A431 клетками. Эти два оцFv проявляют связывание по своему характеру, аналогичное оцFv 425.
Очищенные оцFv, приготовленные из двух отдельных образцов, связывающихся как с ЭФРР, так и с ЭФРР-несущими клетками опухоли (L3 11D и S4 2D) испытывают в анализах на конкурентное связывание с мышиными 425 MAb. В то время как очищенный оцFv 425 способен препятствовать связыванию 425 MAb с ЭФРР в данном интервале концентраций, оцFv L3 11D и S4 2D не препятствуют связыванию мышиного 425 MAb с ЭФРР в тех же концентрациях. Эти два оцFv, видимо, распознают эпитоп на ЭФРР, отличающийся от эпитопа, распознаваемого мышиным 425 MAb.
(4) Химерные полные антитела, происходящие из оцFv
Два оцFv (L3 11D и S4 2D) отобраны для превращения в молекулу полного антитела. ДНК, кодирующие мышиные VH и VK, клонируют в промежуточные вектора, содержащие ДНК последовательности, кодирующие лидерные последовательности иммуноглобулина и сигналы сплайс-донора (фиг. 3). Регулировка положения участков клонирования в VH промежуточном векторе означает замену первого остатка в VH (аспаргиновой кислоты на глутаминовую кислоту). Из промежуточных векторов ДНК фрагменты, содержащие VH и VK, уже присоединенные к лидерным последовательностям и сплайс-донора, клонируют в клеточные экспрессионные вектора млекопитающего, содержащие ДНК, кодирующие либо постоянную гамма-1-область человека, либо постоянную каппа-область человека (Maeda и др. , Hum. Antibod. Hybridomas 1991, 2, 124). Для каждого химерного антитела COS клетки трансфектируют одновременно векторами экспрессии тяжелой цепи и легкой цепи. В качестве положительного контроля, клетки также одновременно трансфектируют векторами экспрессии тяжелой и легкой цепей, кодирующими химерное 425 антитело (Kelleborough и др., Protein Eng. 1991, 4, 773). От клеток отделяют среду, которую анализируют методом ФИСА с целью определения концентрации присутствующего антитела и способности антитела связываться с ЭФРР (фиг. 4). При сравнении концентрации, необходимой для достижения полумаксимума связывания с антигеном, химерное S4 2D антитело связывается с ЭФРР на уровне, равном связыванию для химерного 425 антитела. Однако химерное L3 11D антитело связывается с ЭФРР примерно в четыре раза хуже, чем химерное 425 антитело. Сродство химерного 425 антитела (Kettleborough и др., Protein Eng. 1991, 4, 773), определенное анализом с конкурентным связыванием, равняется 1,9 и 108 М-1. Полученные результаты довольно неожиданны, поскольку предшествующие данные анализа оцFv указывают на то, что и оцFv S4 2D, и оцFv L3 11D связываются с ЭФРР лучше, чем оцFv 425 (фиг. 2). Очищенные белком A образцы химерных L3 11D и S4 2D антител анализируют методом НДС-ПАГЭ в восстановительных и невосстановительных условиях. Кроме того, химерные L3 11D и S4 2D антитела анализируют проточной цитометрией на связывание с A431 и SK-MEL-23 клетками. Оба химерных антитела хорошо связываются с ЭФРР-экспрессирующими A431 клетками и не связываются с ЭФРР-отрицательными SK-MEL-23 клетками.
(5) Применение в терапии и диагностике
Фрагменты антител и полные антитела по изобретению могут быть введены больному человеку в лечебных целях.
Таким образом, цель изобретения состоит в создании лекарственной формы, содержащей в качестве активного компонента по меньшей мере одно антитело или один фрагмент антитела по вышеприведенному определению и определению, представленному в формуле изобретения, в смеси с одним или несколькими фармацевтически приемлемыми носителями, наполнителями или разбавителями.
Как правило, антитело данного изобретения инъецируют внутривенно или парентерально. Обычно дозировочные интервалы вводимых фрагментов антител достаточно велики для создания необходимого подавляющего опухоль или рассасывающего опухоль эффекта. Дозировка зависит от возраста, состояния, пола и тяжести заболевания больного и может меняться от 0,1 мг/кг до 200 мг/кг, предпочтительно от 0,1 мг/кг до 100 мг/кг/доза в одной или нескольких дозах, вводимых ежедневно в течение одного или нескольких дней.
Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примеры неводных растворителей включают: пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло и инъецируемые органические сложные эфиры, такие как этилолеат и другие, известные специалистам растворители, приемлемые для таких целей. Антитела данного изобретения могут быть использованы в лекарственной форме, содержащей физиологически приемлемый носитель. Примеры таких приемлемых носителей включают: солевой раствор, ФБС, раствор Ринджера или раствор Ринджера с лактозой. В лекарственных формах могут также присутствовать консерванты и другие добавки, например: антибиотики, антиокислители и хелатобразующие агенты.
Для поддержки цитотоксичности антитела (фрагмента) оно может быть также конъюгировано известными методами с цитокинами, например ИЛ-2.
Лекарственные формы настоящего изобретения пригодны для лечения всех видов опухолей, в том числе: меланом, глиом и карцином, а также опухолей в системе кровообращения и твердых опухолей.
Для диагностических целей антитело может быть коньюгировано, например, с непроницаемым для излучения красителем или может быть помечен радиоактивным изотопом. Рекомендуемым способом метки является иодогенный способ. В диагностических целях антитело рекомендуют вводить в виде F(ab')2 или оцFv фрагментов. При этом достигаются лучшие результаты, поскольку нет необходимости в вычитании фона.
Пример 1. A431 везикулы
Препараты отделенных мембранных везикул получают по ранее описанной методике (Cohen и др., J.Biol Chem. 1982, 257, 1523; Yeaton и др., J.Biol. Chem. 1983, 258, 9254) с некоторыми модификациями. Конфлюентные колбы, содержащие A431 клетки, промывают ФБС, содержащим кальций и магний. Добавляют гипотонический ФВС и колбы встряхивают 15 минут. Затем клетки промывают везикуляционным буфером (100 мМ NaCl, 50 мМ Na2HPO4, 5 мМ KCl, 0,5 мМ MgCl2, pH 8,5). После добавления везикулярного буфера колбы перемешивают при комнатной температуре и при 37oC. Затем буфер декантируют через металлическое сито в охлаждаемые льдом пробирки на 60 мл и центрифугируют при 150•g при 4oC. Осадок отбрасывают, а надосадочную жидкость ультрацентрифугируют 90 минут при 39000 об/мин. Конечный осадок вновь суспендируют в Гепес буфере (pH 7,4). Для анализа ЭФРР из везикул образцы осаждают 9 объемами этанола, вновь суспендируют в 0,08 М Трис (pH 6,8) и затем проводят НДС-ПАГЭ с MAb 425 в качестве стандарта.
Содержание белка в препаратах определяют количественно модифицированным методом Кумасси Плюс с применением БСА в качестве стандарта при 595 нм. Для анализа ЭФРР из везикул образцы осаждают 9 объемами этанола (примерно сутки при 4oC). Осадок вновь суспендируют в Трис (0,08 М, pH 6,8) и затем проводят НДС-ПАГЭ (5%-ый упаковывающий гель, 1 ч, 35 мА; 10%-ый проходящий гель, 2,5 ч, 40 мА), Образцы и стандарт готовят в двух экземплярах. Один из них окрашивают Кумасси голубым, а другой наносят в виде пятен на нитроцеллюлозные листы (12 B, 16 ч при 4oC) и обрабатывают мышиным mab 425 (анти-ЭФРР) и антимышиным IgG-антителом, конъюгированным с щелочной фоcфатазой.
Для in vitro иммунизации применяют три среды. Среда-1 (С1), среда-2 (С2) и смешанная тимоцитовая культуральная среда или смешанная клеточная культура (СКК). С1 состоит из HLI (Вертрекс Лабораториз, США) с добавкой 50 мМ 2-меркаптоэтанола и 2 мМ L-глутамина (Гибко). С2 состоит из HLI с добавкой 50 мМ 2-меркаптоэтанола, 40 Е/мл ИЛ-2 (Гензим), 20 мг/мл вспомогательного пептида (Сигма), 2 мМ L-глутамина, 100 Е/мл пенициллина (Гибко), 100 мг/мл стрептомицина (Гибко), 4% и 20% ФТС (Биолоджикал Индастриз). СКК приготовлена по методике Vaux (1). Если вкратце, то суспензии клеток вилочковых желез трехнедельных мышей линий Balb/c и C57/BL-1 готовят продавливанием вилочковых желез через стерильное сито на 50 меш. Клеточную суспензию отделяют, промывают дважды HBSS и число живых клеток определяют методом исключения с трипановым голубым. Тимоциты затем культивируют при плотности 2,5•106 тимоцитов каждой линии на мл в HLI среде, содержащей 4% ФТС, 2 мМ L-глутамина, 100 Е/мл пенициллина и 100 мг/мл стрептомицина. Спустя 48 часов надосадочную жидкость отделяют, фильтруют через 0,22 мм фильтр и хранят при -70oC.
Суспензию спленоцитов от неиммунизированных мышей линии Balb/c в возрасте восьми недель получают способом, приведенном для получения суспензии тимоцитов. Жизнеспособность клеток определяют методом исключения с трипановым голубым.
Пример 2. In vitro иммунизация и скрининг
Для иммунизации in vitro применяют три среды. Среда-1 (С1), cреда-2 (С2) и смешанная тимоцитовая культурная среда или смешанная клеточная культура (СКК), С1 состоит из HLI (Вентрекс Лабораториз, США) с добавкой 50 мМ 2-меркаптоэтанола и 2 мМ L-глутамина (Гибко), С2 состоит из HLI с добавкой 50 мМ 2-меркаптоэтанола, 40 Е/мл ИЛ-2 (Гензим), 20 мг/мл вспомогательного пептида (Сигма), 2 мМ L-глутамина, 100 Е/мл пенициллина (Гибко), 100 мг/мл стрептомицина (Гибко), 4% или 20% ФТС (Биолоджикал Индастриз). СКК готовят по методике Vaux (1). Если вкратце, то суспензии тимусов трехнедельных мышей линий Balb/c и C57/BL-1 получают продавливанием вилочковых желез через стерильное сито на 50 меш. Клеточную суспензию отделяют, промывают дважды HBSS и число живых клеток определяют методом исключения с трипановым голубым. Тимоциты затем культивируют при плотности 2,5•106 каждой линии на мл HLI среды, содержащей 4% ФТС, 2 мМ L-глутамина, 100 Е/мл пенициллина и 100 мг/мл стрептомицина. Спустя 48 часов надосадочную жидкость отделяют, фильтруют через фильтр на 0,22 мм и хранят при -70oC.
Суспензию спленоцитов от неиммунизированных мышей в возрасте восьми недель линии Balb/c получают способом, приведенным для получения суспензии тимоцитов. Жизнеспособность клеток определяют методом исключения с трипановым голубым.
Суспензионную клеточную культуру из тимусов трехнедельных мышей линий Balb/c и C57/BL-1 получают продавливанием вилочковых желез через стерильное сито на 50 меш. Клеточную суспензию отделяют, промывают дважды HBSS и число жизнеспособных клеток определяют методом исключения с трипановым голубым. Тимоциты затем культивируют при плотности 2,5•106 тимоцитов каждой линии на мл HLI-среды, содержащей 4% ФТС, 2 мМ L-глутамина, 100 Е/мл пенициллина и 100 мг/мл стрептомицина. Спустя 48 часов надосадочную жидкость отделяют, фильтруют и оставляют на хранение. Суспензию спленоцитов от неиммунизированных мышей линии Balb/c в возрасте восьми недель получают способом, приведенным для получения суспензии тимоцитов. Жизнеспособность клеток определяют методом исключения с трипановым голубым.
Иммунизацию in vitro осуществляют в 6-луночных планшетах (Костар). Лунки, содержащие 107 спленоцитов в 3,5 мл С1-среды (состоит из HL-I-среды (Вентрекс Лабораториз, США) с добавкой 60 мкМ 2-меркаптоэтанола и 2 мМ L-глутамина (Гибко)), инкубируют (37oC, 5% CO2) с везикулами, несущими ЭФРР в необходимой концентрации. В контрольные лунки вносят везикулы из клеток, не экспрессирующих ЭФРР, или ФБС. Через несколько часов в каждую лунку вносят 3,5 мл С2-среды (состоит из HLI с добавкой 50 мкМ 2-меркаптоэтанола, 40 Е/мл ИЛ-2 (Гензим), 20 мкг/мл вспомогательного пептида (Сигма), 2 мМ L-глутамина; 100 Е/мл пенициллина (Гибко), 100 мкг/мл стрептомицина (Гибко)), содержащей 4% и 10% ФТС (Биолоджикал Индастриз). В некоторых опытах С2 заменена СКК-средой (смешанная тимоцитная среда культивирования или смешанная клеточная культура (Vaux и др. , Nature) 1988, 336, 36) с добавкой вспомогательного пептида (20 мкг/мл) и ИЛ-2 (40 Е/мл). (Необходимо отметить, что конечная концентрация ФТС, ИЛ-2 и вспомогательного пептида в культуре снижена на 50%). Клетки инкубируют в течение 72, 96, 120 или 144 ч в тех же условиях и наконец клетки анализируют на присутствие специфичного иммуноглобулина, или обрабатывают с целью выделения РНК.
Скрининг проводят с очищенными антигенами или фиксированными A431 клетками. Методика по существу соответствует ранее описанной (Carroll и др., Hybridboma 1990, 9, 81) с некоторыми модификациями. Если вкратце, то стерильные 96-луночные планшеты (Нунк, Максисорб) покрывают примерно на сутки очищенным ЭФРР (2,5 мкг/мл), CD3 ганглиозидом (2 мкг/мл) или РНКазой (10 мкг/мл) в ФБС. В случае применения в качестве антигена A431 клеток, клетки культивируют в 96-луночном планшете до конфлюэнтного состояния и фиксируют 0,1% глутаральдегида. Иммунизированные in vitro лимфоциты, промытые и вновь суспендированные в HLI среде с добавкой 2% ФТС и 2 мМ L-глутамина, вносят в каждую лунку в концентрации 5•105 клеток/мл и 1•105 клеток и инкубируют (37oC, 5% CO2) 48 ч. Для каждой группы делают шестнадцать дублей. Затем лимфоциты удаляют 5-кратным промыванием ФБС, содержащим 0,1% Твин-20. Специфичные иммуноглобулины обнаруживают с помощью меченных пероксидазой кроличьих антимышиных иммуноглобулинов (Дако) (1 час, 37oC). В качестве субстрата применяют 2,2'-азинобис(3-этилбензтиазолин-6-сульфониевая кислота)-диаммониевую соль (АБТС) (Сигма) в цитрат-фосфатном буфере.
Пример 3. Конструирование библиотеки
Конструируют три библиотеки: из РНК, полученной из селезенки мыши, иммунизированной внутрибрюшинно A431 клетками (Murthy и др., Arch. Biochem. Biophys 1987, 252, 549), из подколенного лимфатического узла мыши, иммунизированной в подушечку лапки очищенным ЭФРР, и из мышиных клеток, иммунизированных in vitro A431 везикулами. Синтезируют кДНК с первой цепочкой. Гены VH и VK ПЦР-амплифицируют и упорядочивают (Clackson и др., Nature 1991, 352, 624). С помощью ПЦР вводят NotI и SfiI сайты рестрикции и оцFv клонируют в фагемидный вектор pHEIN (Hogoenboom и др., NucI. Acids Res. 1991, 19, 4133). Лигированные смеси подвергают электропорации в клетки E.coli и образовавшиеся колонии соскабливают в среду с созданием общей библиотечной партии (Narks и др., J. Mol. Biol. 1991, 222, 581).
Пример 4. Скрининг библиотеки
Фаговые антитела выделяют из библиотеки с помощью M13K07 фага-помощника (Промега, Мэдисон, ВИ) (Marks и др., J. Mol. Biol. 1991, 222, 581). Иммунопробирки (Нунк, Лайф Сайенсиз, Великобритания) покрывают примерно на сутки 4 мл 2,5 мкг/мл ЭФРР в ФБС. После трех промывок ФБС пробирки инкубируют по меньшей мере 1 ч при 37oC в ФБС, содержащем 2% молочного порошка (ФБСМ). Фаг (1012-1013) вновь суспендируют в 4 мл ФБСМ и инкубируют 1 ч при комнатной температуре в покрытых ЭФРР пробирках. Пробирки промывают 20 раз ФБС с 0,1% Твин-20 и 20 раз ФБС. Связанный фаг элюируют после 10 мин инкубирования 1 мл 0,1 М раствора триэтиламина с перемешиванием в начале и конце. Элюированный фаг нейтрализуют добавлением 0,5 мл 1 М Трис-HCl (pH 7,5) и применяют для инфицирования TGI клеток E.coli в лаг-фазе. Инфицированные клетки наносят на пластины и индивидуальные колонки отбирают для маломасштабного индуцирования оцFv. Оставшиеся колонки соскабливают в среду и аликвоту используют для приготовления фага для следующего цикла скрининга.
Пример 5. Продуцирование и анализ оцFv
Растворимые оцFv продуцируют в E. coli HB2151 по известной методике (например, Kettleborough и др., см. выше). Концентрации оцFv в бактериальных надосадочных жидкостях определяют применением в качестве стандарта препаратов очищенного оцFv известной концентрации. Надосадочные жидкости фильтруют и добавляют азид натрия до концентрации 0,1%. Последовательные разбавления надосадочных жидкостей и стандарта наносят в виде пятен на фильтры Иммобилон-PVDF (Миллипор, Уотфорд, Великобритания) с применением 96-луночного коллектора. Фильтры обрабатывают, как при Вестерн блотинге (Towbin и др. , Proc. Natl. Acad. Sci. США 1979, 76, 4350). Детектирование оцFv осуществляют применением антитела (9E10), направленного на C-концевой tag (Munno и Pelham, Cell, 1986, 46, 291), с последующим применением конъюгированного пероксидазой козьего антимышиного IgG и IgM антитела (Джексон Иммуно Рисерч Лаб. Инк., Уэстр Грув, ПА). Продукты реакции проявляют с помощью ECL системы (Амерсхэм, Айлесбэри, Великобритания). Предварительно ополоснутые авторадиограммы сканируют на денситометре. Строят стандартную кривую, которую используют для определения концентраций оцFv в надосадочных жидкостях.
На покрытых ЭФРР пластинах (2,5 мкг/мл) проводят ФИСА со связыванием антигена. Содержащие оцFv надосадочные жидкости разбавляют ФБСМ и добавляют к пластинам. Связанные оцFv детектируют применением 9E10 антитела вышеописанным способом. Надосадочные жидкости, кроме того, анализируют на связывание с набором неродственных белков и пластиком. Пластины для ФИСА покрывают примерно на сутки овальбумином, лизозимом куриного яйца, цитохромом Ц, глицеральдегид-3-фосфат-дегидрогеназой, мышиным альбумином (СВА линия) и БСА в концентрации 100 мкг/мл. Неразбавленные надосадочные жидкости, содержащие 2% сухого молока, добавляют дублями к пластинам с покрытием и связанные оцFv детектируют вышеописанным способом.
ФИСА на связывание клеток проводят применением клеточных линий опухоли: A431 (ATCC CRL 1555), MDA MB 468 (ATCC HTB 132) и SK-MEL-23 (отрицательный контроль). Рост клеток до конфлюэнтного состояния проводят в обработанных поли-D-лизином 96-луночных планшетах для тканевых культур (Нунк). Клетки промывают МДСИ (ДМЭМ) и блокируют 2 ч при 37oC добавлением ФБС, содержащего 2,5% БСА. После всасывания в каждую лунку вносят надосадочные жидкости вместе с равным объемом 2 х УТ среды, содержащей 4% сухого молока, и инкубируют 1 ч при 4oC. Связанные оцFv детектируют вышеприведенным способом.
ФИСА на конкурентной основе проводят прединкубированием 10 мин ЭФРР-покрытых пластин для ФИСА с 50 мкл очищенного оцFv (100 мкг/мл). Затем добавляют мышиное MAb 425 (50 мкл) с получением концентраций от 3,13 до 200 нг/мл. После инкубирования и промывания связанное мышиное MAb 425 детектируют с помощью конъюгированного с пероксидазой козьего антимышиного IgG и IgM антитела.
Пример 6. ДНК анализ
Для BstNI фингерпринтинга вставки оцFv из индивидуальных клонов амплифицируют методом ПЦР и полученные продукту гидролизуют в присутствии BstNI (Clackson и др., Nature 1991, 352, 624). Последовательность ДНК определяют применением набора Секвеназа (Юнайтид Стейтс Биокемикал, Кливлэнд, ОГ).
Пример 7. Очистка оцFv
Бактериальные надосадочные жидкости осветляют и фильтруют используя фильтры на 0,2 мкм, перед загрузкой колонку на 1 мл с очищенным ЭФРР (5 мг), присоединенным к активированной цианогеномбромидом Сефарозе 4B (Фармация, Уппсула, Швеция). Колонку промывают 30 мл ФБС и затем 5 мл 0,2 М глицина (pH 5). Элюирование оцFv осуществляют 0,2 М глицин/HCl (pH 2,8). Элюат нейтрализуют 10-кратным количеством ФБС. Содержащие белок фракции объединяют и буфер заменяют ультрафильтрацией (Амикон, Стоунхауз, Великобритания) на ФБС, содержащий 1% БСА и 0,05% азида натрия.
Пример 8. Анализ очищенных оцFv на КСВФ (FACS)
A431 клетки трипсинизируют и инкубируют в МДСИ, содержащей 10% ФТС. Клетки промывают дважды холодной МДСИ и фильтруют через сито на 45 мкм. Клетки (106) инкубируют 30 мин на льду в 50 мкл ФБС, 1% БСА с очищенными оцFv. После двух промывок холодным ФБС связанные оцFv детектируют применением 50 мкл ФИТЦ-конъюгированного 9E10 антитела (100 мкг/мл). После выдерживания 30 мин на льду клетки промывают один раз ФБС, фиксируют в ФБС, содержащем 1% формальдегида, и анализируют с помощью прибора ФАКСКАН (Бектон-Дикинсон, Коули, Великобритания).
Пример 9. Конструирование, анализ и экспрессия полных химерных антител
Использованием PstI и BstEII сайтов ДНК кодирующие VH отобранных оцFv субклонируют в промежуточный VH вектор, содержащий эукариотическую лидерную последовательность, происходящую из антитела HG3 CL человека (Rechavi и др., Proc. Natl. Acad. Sci США 1983, 80, 855) и сайт сплайс-донора (фиг. 3). ДНК, кодирующие VK, подготавливают для вставки в промежуточный VK вектор применением ПЦР праймеров с целью введения XhoI и SstI сайтов у 5'- и 3'-концов:
(VK Передний: 5'-CCG TTT CAG CTC GAG CTT GGT CCC-3', Vk Задний: 5'-GAC ATT GAG CTC ACC CAG TCT CCA-3').
SstI-XhoI фрагменты клонируют в промежуточный VK вектор, содержащий эукариотическую лидерную последовательность, происходящую из реконструированной легкой цепи CAMPATH-1 человека (Riechmann и др., Nature 1998, 332, 21) и сплайс-донора сайта (фиг. 3). ДНК, кодирующие вариабельные области плюс эукариотические фланкирующие области, клонируют в виде HindIII-BamI фрагментов в вектора экспрессии клеток млекопитающего, содержащие геномные ДНК, кодирующие постоянную гамма-1-область человека или постоянную каппа-область человека (Maeda и др., Hum. Antibod. Hybridomas 1991, 2, 124). Вектора экспрессии тяжелой и легкой цепей вводят электропорацией в COS клетки. Спустя 72 ч среду отделяют и химерные анти-ЭФРР антитела анализируют методом ФИСА (Kettleborough и др., Protein Eng. 1991, 4, 773).
Пример 10. Продуцирование оцFv, происходящих из in vitro иммунизированных клеток
Раскрываемые ниже способы являются слегка модифицированными вышеприведенными способами. Способы иммунизации, конструирования библиотеки и скрининга приведены в Примерах 1-4. Ниже более подробно раскрываются следующие стадии.
После скрининга первичной библиотеки и клонов, полученных в трех циклах пэннинга, отбирают несколько единичных устойчивых к ампициллину колоний. Фагемидную ДНК получают щелочным лизисом и применяют для трансфекции E.coli HB2151 (несупрессорный штамм) тепловым ударом. Колониями инокулируют 2хТУ-Amp-Glu и выращивают примерно сутки при 30oC. Для инокуляции 50 мл 2хТУ бульона, содержащего ампициллин (100 мг/мл) и 0,1% глюкозы, применяют в 5 мл, после чего клетки выращивают 1 ч со встряхиванием при 30oC (до лаг-фазы). Клетки собирают и экспрессию растворимого оцFv индуцируют добавлением изопропил- β -D-тиогалактопиранозида (ИПТГ) до конечной концентрации 1 мМ (De Bells D. и Schwartz I., Nucleic Acids Res. 1990, 18, 1311). Культуры выращивают примерно сутки при 80oC со встряхиванием. Отобранные надосадочные жидкости, содержащие оцFv, осветляют центрифугированием и фильтрованием через фильтры на 0,22 мм и анализируют. Бактериальные надосадочные жидкости анализируют на связывание с ЭФРР методом ФИСА по известной методике (Kettleborough и др., патент EP 94/104160 и Eur. J. Immunol. 1994, 24, 952). Специфичность отобранных оцFv фрагментов проверяют методом ФИСА с применением пластинок, покрытых различными белками, родственными и неродственными ЭФРР, а также других антигенов и пластика. Применяют следующие антигены: РНКазу, БСА, OVA, GD3 ганглиозид, витронектина рецептор (ВНР), гликопротеин тромбоцитов IIbIIIa (ГПIIbIIIa) и дисиалил-лакто-N-тетраозу (ДСЛНТ). Покрытие выдерживают примерно сутки при оптимальной для каждого антигена концентрации. Пластинки для ФИСА с покрытием блокируют 1 ч при 37oC добавлением 1,5% обезжиренного молока в ФБС (мас./об.). После промывания 100 мл, содержащих оцFv надосадочных жидкостей, вносят в лунки микротитрационного планшета и инкубируют 2 ч при 37oC. Связанный оцFv детектируют с помощью анти-c-myc антитела 9E10 (отработанная культурная среда от Myc 1-9E10.2 гибрида), и конъюгированного с щелочной фосфатазой кроличьего антимышиного антитела (Дако).
Для выявления способности оцFv связываться с клеточным ЭФРР с помощью КСВФ анализа и иммунофлюоресценции с нефиксированными клетками применяют три ЭФРР-несущие клеточные линии опухоли: A431, MDA MB 231 аденокарциномы молочной железы человека (ATCC HTB 26) и HT29 аденокарциномы оболочной кишки человека (ATCC HTB 38), а также неэкспрессирующую ЭФРР клеточную линию WM164. Для косвенного иммунофлюоресцентного анализа клетки наносят на пластинки Тарасаки (2•104 клеток/лунку) и культивируют 24 ч. Затем клетки инкубируют 90 мин при комнатной температуре с 20 мл сырой бактериальной надосадочной жидкости, содержащей жидкости, содержащей оцFv фрагменты. Инкубирование с первичным антителом (анти-c-myc) и вторичным антителом ведут 60 мин при комнатной температуре. Применяемое в качестве вторичного антитела ФИТЦ-конъюгированное кроличье антимышиное антитело (Дако) разбавляют в отношении 1:20.
Для КСВФ анализа 5•105 клеток промывают ФБС с 1% БСА и 0,1% азида натрия (ФБС-БСА) и инкубируют 20 мин при 4oC с 50 мл сырой бактериальной надосадочной жидкости. После двух промывок холодным ФБС-БСА связанный оцFv детектируют с помощью анти-c-myc антитела и ФИТЦ-конъюгированного козьего антимышиного антитела (Бектон-Дикинсон), разбавленных в отношении 1:25 в ФБС-БСА. Добавляют йодид пропидия (ЙП) до конечной концентрации 5 мг/мл. Анализ проточной цитометрией проводят на приборе ЭПИКС, Профиль 11, снабженным охлаждаемым воздухом аргоновым лазером. Для возбуждения используют линию 488 нм (15 мВ). Для отбора эмиссии ФИТЦ применяют пропускной фильтр для полосы 530 нм, а для отбора эмиссии ЙП применяют пропускной фильтр для полосы 625 нм. Живые клетки отбирают настройкой биткарты на переднее и боковое рассеивание и путем исключения ЙП-окрашенных клеток.
Различия в первичных и отобранных библиотеках определяют ПЦР амплификацией клонирования фрагментов (Giis'sow D., Clackson T., Nucleic Acids Res. 1989, 17, 4000) и анализом продуктов гидролиза в присутствии BstNI (8). Последовательности некоторых клонов определяют с помощью набора Секвеназа (ЮСБ) дидезокси-методом терминации цепи (Sanger и др., Proc. Natl. Acad. Sci. США 1977, 74, 5463).
Сырые бактериальные надосадочные жидкости подвергают НДС-ПАГЭ на 12,5%-ом геле. Вестерн блотинг осуществляют по существу по методике Towbin (Towbin и др., J. Proc. Natl. Acad. Sci. США 1979, 76, 4350). Белки переносят электроблотингом на Иммобилон-P (Миллипор) или нитроцеллюлозу (Био-Рад). Пятна блокируют ФБС, содержащим 2% обезжиренного молока (мас./об.). оцFv фрагменты детектируют применением анти-c-myc антитела (9E10), конъюгированного с пероксидазой антимышиного антитела (Джексон) и усиленной хемилюминесцентной системы (ECL, Амерсхэм).
Количественным анализом отделенных от мембраны везикул определяют общую концентрацию белка в 2,5 мг/мл, из которых только 10-14% соответствуют ЭФРР (Sato и др., J. Nat. Cancer Inst. 1989, 21, 1601; Yeaton R. и др., J. Biol. Chem. 1983, 258, 9254), т.е. 250-350 мг/мл. Электрофоретический анализ с применением НДС-ПАГЭ с последующим окрашиванием Кумасси голубым показал, что везикулы содержат довольно сложную смесь белков. Никакого разложения белков не обнаружено. Анализ Вестерн блотингом выявил, что в применяемых нами условиях эксперимента в препарате мембранного везикула присутствуют полные молекулы ЭФР рецептора.
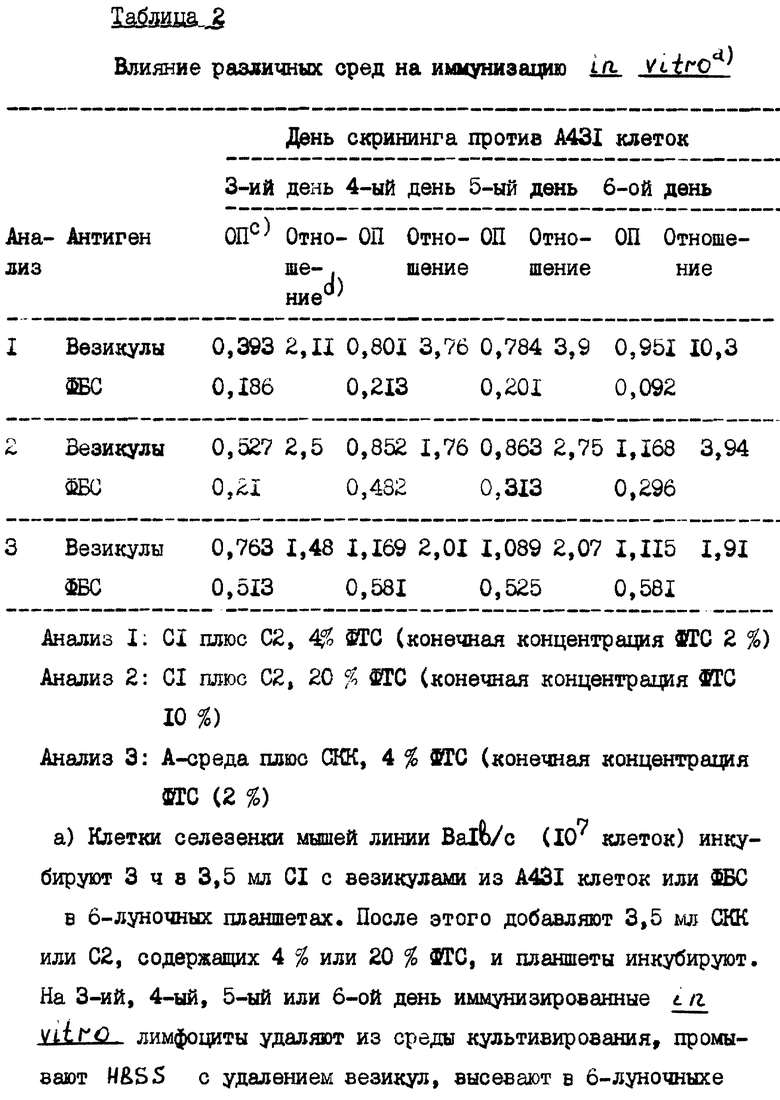
С целью выявления требований, предъявляемых к ФТС и лимфокинам, проводят сравнение СКК и С2, содержащих 20% или 4% ФТС. Везикулы, несущие ЭФРР или ФБС, применяют соответственно в качестве антигена и контроля. В шестилуночных планшетах инкубируют 3 ч спленоциты в С1-среде (бензосывороточная) с добавлением или без антигена. Затем добавляют С2 или СКК и спустя 72, 96, 120 или 144 ч проводят скрининг с применением фиксированных A431 клеток. Во всех опытах число выделенных жизнеспособных клеток колеблется в интервале 20-40%, что согласуется с опубликованными результатами (Gavilondo-Cowley J. и др., "Иммунизация in vitro в гибридомной технологии", Элзевир Сайенс Паблишерс Б. В., Амстердам 1988, стр. 131). Максимальная специфичная реакция в случае СКК наблюдалась на четвертый день, в то время как в случае С2, содержащей 4% или 20% ФТС (2% или 10% конечная концентрация) максимальная реакция запаздывала до шестого дня (табл. 2). Однако СКК и 10% ФТС инициируют неспецифичную реакцию, вероятно, за счет поликлональной активации, что можно видеть, если выразить результаты в виде отношения специфичной к неспецифичной реакции. Для проведения дополнительных анализов нами решено использовать С2-среду с добавкой 4% ФТС и 6 дней культивирования.
Присутствие на поверхности везикул ЭФРР усиливает реакцию на антиген. В методике, аналогичной вышеприведенной, сравнивают везикулы из экспрессирующих и неэкспрессирующих клеточных линий. Лимфоциты культивируют 3 ч с везикулами с С1-среде. После этого добавляют С2, содержащую 4% ФТС. Спустя 6 дней лимфоциты из каждой группы культивируют 48 ч в 96-луночных планшетах, покрытых ЭФРР, фиксированными A431 клетками, РНКазой или CD3. Как и ожидалось, результаты проведенных анализов показали мультиспецифичность характера реакции (табл. 3). Реакционоспособность по отношению к ЭФРР явно повышается (на основании оптической плотности) в случае применения в качестве антигена экспрессирующих ЭФРР везикул.
Проанализировав полученные результаты, делают предположение, что антигензависимая реакция после иммунизации in vitro, в ходе которой образуется несколько объединений иммунизированных лимфоцитов против ЭФРР, пригодна для ПЦР-клонирования вариабельных областей.
После клонирования оцFv фрагментов, полученных в ходе иммунизации in vitro в pHENI фагемид получена библиотека из 1,1•105 клонов. Эта библиотека создана параллельно с еще двумя библиотеками, созданными in vivo иммунизацией. Конструирование таких фаговых библиотек описано ранее (Kettleborough и др., патент EP 94/104160 и Eur. J. Immunol. 1994, 24, 952).
Для отбора оцFv фрагментов, связывающихся с ЭФРР, фаг подергают пэннингу с применением ЭФРР-покрытых иммунопробирок. Элюируемый фаг применяют для повторного инфицирования SupE штамма E.coli. В общей сложности проведено три цикла отбора. В каждом цикле для определения фона параллельно анализируют пробирку без антигена. На иммунопробирку наносят 1,5•1010 фаговых частиц и элюированием получают 6,6•104 частиц из иммунопробирок, в то время как из фоновой популяции получают только 200 колоний. После третьего пэннинга наносят 1•1011 фагов, а элюируют 5,6•1010.
Для дополнительной характеристики оцFv фрагментов из фаговых популяций до отбора и после каждого цикла отбора отбирают 22 клона.
Отличия в библиотеках определяют анализом продуктов гидролиза в присутствии BstNI клонированных фрагментов. До отбора библиотеки, видимо, очень резко отличаются друг от друга. Фингерпринтинг связывающихся клонов, полученных после первого цикла отбора, указывает на присутствие нескольких групп с одинаковым характером рестрикции.
Клоны отбирают с различных циклов отбора на основе их типов сжигания. Определением ДНК последовательности выявлено присутствие разных последовательностей в большинстве отобранных клонов. Длина и состав определяющих комплементарность областей (ОКО) клонов 10D2, 5D3, 10E2, 1B3, 4B3 и 5E2 были различными. Большая часть вариаций наблюдается в ОКО из VH и VL последовательностей. Клоны 5D3 и 1E3, полученные после третьего цикла отбора, характеризуются прочным связыванием с ЭФРР при анализе методом ФИСА и проточной цитометрией и имеют одинаковые последовательности.
Растворимые оцFv фрагменты получены выращиванием несупрессорного штамма HB 2151 E.coli.
Для проверки продуцирования оцFv бактериальную среду от отдельных клонов анализируют электрофорезом на геле. Анализом Вестерн блотингом выявлена четкая полоса в районе 35000 кДа.
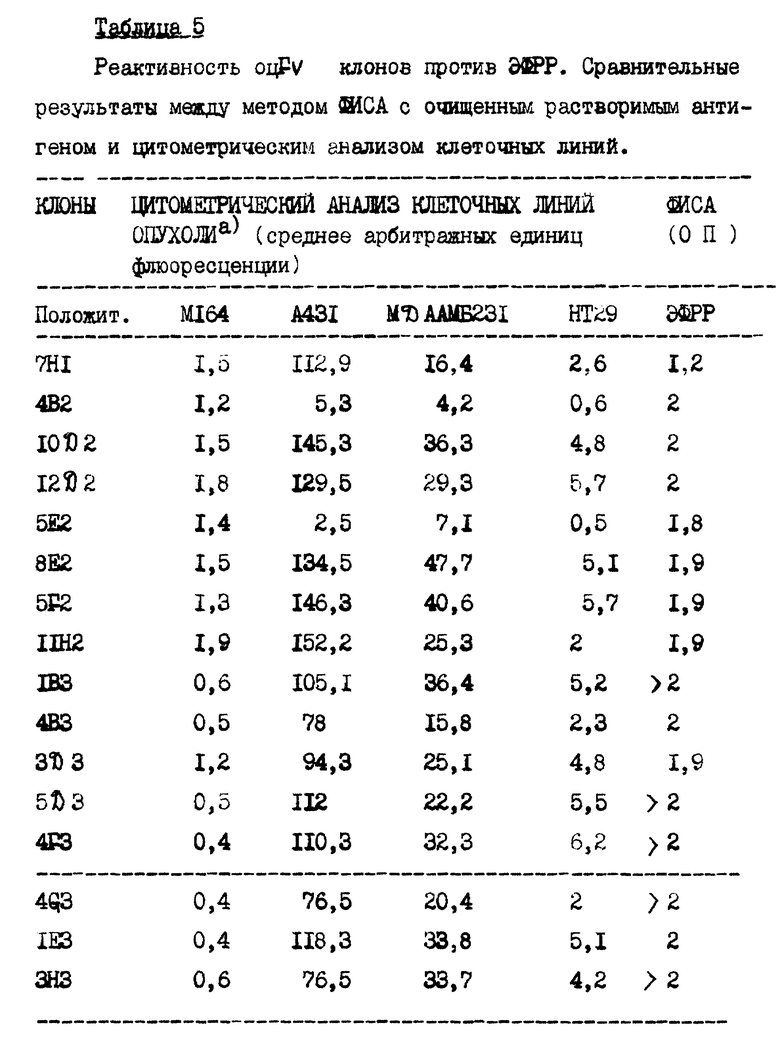
Клоны, обладающие активностью в связывании с ЭФРР, идентифицируют методом ФИСА. Для изучения перекрестной реактивности отобранных клонов проводят ФИСА анализы с применением различных антигенов. На пластинки для ФИСА в оптимальной концентрации наносят следующие антигены: ЭФРР, РНКазу, БСА, KLH, OVA, GD3 ганглиозид, рецептор витронектина, гликопротеин IIbIIIa тромбоцитов и дисиалил-лакто-N-тетраозу (табл. 4). Никакого связывания с отличными от ЭФРР антигенами не обнаружено. Кроме того, проведен анализ способности оцFv связываться с тремя ЭФРР-несущими клеточными линиями опухоли (эпидермоидная карцинома человека A431, аденокарцинома молочной железы человека MDA MB 231 и аденокарцинома ободочной кишки человека HT 29). В качестве отрицательного контроля применяют WM 164 меланомы человека, не экспрессирующей ЭФРР. Связывающиеся с клеточными линиями опухоли оцFv фрагменты анализируют косвенной иммунофлюоресценцией с использованием нефиксированных клеток, количественные определения проведены КСВФ анализа. Применение нефиксированных клеток гарантирует сохранение естественной конформации мембранных рецепторов. Положительные клоны показали четкую флюоресценцию в случае применения A431 клеток. Флюоресценция в случае других ЭФРР-несущих клеточных линий была слабой. Никакого связывания с отрицательными клеточными линиями не отмечено. Эти результаты подтверждены проточной цитометрией. Семнадцать положительных клонов и три отрицательных клона анализируют на связывание с A431, MDA MB 231 и HT 29 проточной цитометрией. В качестве отрицательной клеточной линии применяют WM 164. В качестве положительного контроля применяют оцFv 425 (P1 клон), а в качестве отрицательного контроля - вектор клонирования (HEN). Полученные результаты суммированы в табл. 5. Два клона (4B2 и 5E2) оказались положительными на связывание с ЭФРР согласно анализу методом ФИСА, но отрицательными на связывание с ЭФРР-экспрессирующими клеточными линиями опухоли.
























































Изобретение относится к новым анти-ЭФРР антителам и к их одноцепочечным Fv (оцFv) фрагментам, которые могут быть получены из фаг-антитело библиотек, сконструированных из клеток иммунизированного млекопитающего, предпочтительно мыши. Два одноцепочечных Fv, выделенных из фаг-антитело библиотек, обработаны методами генной инженерии с созданием молекул частично гуманизированного полного антитела. Такие химерные анти-ЭФРР антитела содержат постоянные области иммуноглобулинов человека и могут быть использованы так же, как и их одноцепочечные Fv, в качестве средств для диагностики и лечения опухолей человека. 5 с. и 4 з.п. ф-лы, 5 табл., 9 ил.
Приоритет по пунктам:
17.03.94 по пп.1 - 9 с последовательностями 1 - 16;
02.12.94 по пп.1 - 9 с последовательностями 17 - 32.
| WO 9516037 A1, 15.06.1995 | |||
| WO 9520045 A1, 27.07.1995. |

