Это изобретение было сделано при правительственной поддержке, предоставленной Национальным институтом здравоохранения на основании субсидий N PO1 HL-46528 и PO1 HL-45168. В этой связи правительство имеет определенные права на это изобретение.
Область техники, к которой относится изобретение
Предметом настоящего изобретения являются способы увеличения эффективности трансдукции клеток вирусами и, в частности, способы усиления опосредованного вирусом переноса генов в клетки с помощью фибронектина и/или фрагментов фибронектина.
Известный уровень техники
Прогресс в области понимания молекулярной основы многих болезней человека, а также совершенствования технологии переноса генов сделал возможным разработку процедуры соматической генотерапии для лечения серьезных генетических заболеваний. В настоящее время с помощью генотерапии можно лечить заболевания, вызванные дефектом или недостаточностью фермента или другого белка, требующие точного регулирования уровня этих компонентов, и заболевания костного мозга человека.
Например, одним заболеванием, которое можно лечить с помощью генотерапии, является аденозиндезаминазная (ADA) недостаточность, которая вызывает тяжелую форму комбинированного иммунодефицита. У больных, страдающих аденозиндезаминазной недостаточностью, в клетках костного мозга содержится мало фермента или он вообще отсутствует. Однако аденозиндезаминазную недостаточность можно лечить путем трансплантации совместимого костного мозга. ADA-нормальные клетки характеризуются избирательным действием по сравнению с пораженными клетками и обычно восстанавливают костный мозг больного.
Клетки костного мозга являются хорошей мишенью для соматической генотерапии, так как ткань костного мозга легко использовать для выполнения in vitro анализов и она содержит репопулирующие клетки. Альтернативно было продемонстрировано, что пуповинная кровь содержит большое количество первичных клеток-предшественников. Успешный перенос генов в кроветворные стволовые клетки, долгоживущие репопулирующие клетки, позволяет вылечивать разные заболевания благодаря получению потомства этих клеток.
Перенос генов и долговременная экспрессия генов в репопулирующих стволовых клетках была достигнута в моделях, полученных рядом ученых в результате исследований, проведенных на мышах. Однако при выполнении in vivo экспериментов на более крупных животных, таких как собаки и приматы, успех оказался весьма ограниченным, что связано главным образом с низкой эффективностью заражения первичных кроветворных стволовых клеток. Применение текущей технологии переноса генов далее осложнено при лечении людей несколькими факторами, которые включают низкие количества стволовых клеток, имеющихся в костном мозге взрослого человека, отсутствие приемлемых методов очистки таких клеток и небольшую долю таких первичных клеток в клеточном цикле.
При выполнении экспериментов как на мышах, так и крупных животных, включающих исследование клеток костного мозга, было установлено, что наиболее успешные процедуры включали сокультивирование клеток-мишеней с линиями ретровирусных клеток-продуцентов. Кроме того, в основе большинства утвержденных Управлением по контролю за продуктами и лекарствами экспериментов по переносу генов у людей лежат рекомбинантные ретровирусные векторы, предназначенные для трансдукции генов. Рекомбинантные ретровирусные векторы весьма желательны для генотерапии, так как они эффективно переносят, точно и стабильно интегрируют экзогенную ДНК в клеточную ДНК. Эти векторы содержат экзогенную ДНК, служащую для переноса генов, и затем модифицируются с целью устранения вирусного патогенеза. Благодаря такой модификации продуцирование вирусов достигается в результате использования ретровирусных упаковочных клеток. Однако для клинической генотерапии более желательна безклеточная трансдукция в связи с вопросами биобезопасности и контроля качества. К сожалению, эффективный перенос генов в такие кроветворные клетки, как стволовые клетки, оказывается невозможным без сокультивирования с вирусопродуцирующими клетками.
Совсем недавно было установлено, что эффективность переноса генов можно увеличить путем оказания воздействия на клетки-мишени со стороны стромальных клеток во время инфицирования. Стромальные клетки являются основным компонентом кроветворного микроокружения. Кроветворное микроокружение состоит из организованной сети макрофагов, стромальных клеток, клеток эндотелия, адипоцитов и сложного экстрацеллюлярного матрикса, составленного из целого ряда определенных адгезионных молекул. Молекулы сложного экстрацеллюлярного матрикса, такие как ламинин, коллаген, тромбоспондин, протеогликаны, гликозаминогликаны и фибронектин, образуют связывающие центры как для кроветворных клеток, так и для факторов роста. До сих пор не ясен механизм, лежащий в основе такого стимулирующего действия стромальных клеток на ретровирусную инфекцию, но уже известно, что физиологическая регуляция пролиферации и дифференциации кроветворных клеток происходит тогда, когда эти клетки непосредственно контактируют с клетками кроветворного микроокружения.
Эффективный перенос генов в долгоживущие репопулирующие кроветворные стволовые клетки и другие клетки остается проблематичным, препятствуя широкому применению в настоящее время методов переноса генов для лечения заболеваний, связанных с кроветворным процессом, и других болезней. Существует насущная необходимость в методах эффективного переноса генетического материала в клетки млекопитающих без опасных последствий и ограничений, присущих используемым ранее методам. Настоящее изобретение направлено на удовлетворение этих потребностей.
Краткое изложение сущности изобретения
Одним предпочтительным вариантом осуществления настоящего изобретения предусматривается способ увеличения частоты трансдукции кроветворных клеток посредством ретровирусного вектора. Этот способ включает инфицирование жизнеспособных кроветворных клеток рекомбинантным ретровирусным вектором с дефектом репликации в присутствии по существу чистого фибронектина и/или его фрагментов, эффективно увеличивающих частоту клеточной трансдукции ретровирусом. Фибронектин и/или его фрагменты можно получить из натуральных материалов или синтезировать искусственным путем (например, создать генетически с помощью методов рекомбинантного или химического синтеза) либо получить из комбинации натуральных и синтетических материалов. Кроме того, совершенно очевидно, что полипептид или полипептиды фибронектина, используемые в этом изобретении, могут включать мутации, образующие натуральную последовательность аминокислот фибронектина, которая тем не менее представляет функциональные полипептиды, обладающие адгезионными свойствами, необходимыми для достижения повышенной трансдукции в соответствии с этим изобретением.
Другим предпочтительным вариантом осуществления настоящего изобретения предусматривается способ получения трансдуцированных кроветворных клеток, который включает инфицирование жизнеспособных кроветворных клеток рекомбинантным ретровирусом с дефектом репликации, содержащим экзогенную ДНК, в присутствии иммобилизованного фибронектина, иммобилизованных фрагментов фибронектина или их иммобилизованной смеси в количествах, достаточных для увеличения частоты клеточной трансдукции ретровирусом.
Еще одним предпочтительным вариантом осуществления настоящего изобретения предусматривается усовершенствованный способ трансплантации клеток. Этот способ включает стадии получения жизнеспособных кроветворных клеток у животного-донора; инфицирование кроветворных клеток вектором рекомбинантного ретровируса с целью получения трансдуцированных жизнеспособных кроветворных клеток, причем инфицирование производится в присутствии фибронектина и/или его фрагмента в иммобилизованной форме, способствующей увеличению частоты трансдукции; и введение трансдуцированных жизнеспособных кроветворных клеток животному-реципиенту в виде клеточного трансплантата. В одном предпочтительном случае инфицированные клетки можно ввести аутологическому донору.
Еще одним предпочтительным вариантом осуществления настоящего изобретения предусматривается способ получения трансдуцировнных клеток пуповинной крови с целью их трансплантации. Этот способ включает инфицирование кроветворных клеток, взятых из пуповинной крови, рекомбинантным ретровирусным вектором с дефектом репликации в присутствии достаточного количества иммобилизованного фибронектина и/или его фрагментов с целью увеличения частоты трансдукции кроветворных клеток под действием ретровирусного вектора. Настоящее изобретение включает также популяции жизнеспособных трансдуцированных клеток из пуповинной крови, полученных в соответствии с таким способом, и способы трансплантации клеток, которые включают введение животному популяций трансдуцированных клеток в качестве клеточного трансплантата.
В соответствии с особыми аспектами вышеуказанных вариантов осуществления настоящего изобретения используемый фибронектин или его фрагмент должен содержать первую аминокислотную последовательность, которая обеспечивает связывающую ретровирус активность области связывания гепарина П фибронектина, и вторую аминокислотную последовательность, которая обеспечивает связывающую клетки активность области CS-1 фибронектина. Одновременное использование этих двух областей связывания фибронектина обеспечило весьма значительное увеличение трансдукции клеток-мишеней под действием ретровируса.
Еще одним предпочтительным вариантом осуществления настоящего изобретения предусматривается способ создания генно-инженерной конструкции с целью увеличения частоты трансдукции заранее определенной клетки-мишени под действием ретровирусного вектора. Этот способ включает стадию ковалентного связывания лиганда, который связывает клетку-мишень с полипептидом, содержащим аминокислотную последовательность, которая обеспечивает связывающую ретровирус активность области связывания гепарина П фибронектина. Настоящее изобретение включает также способы использования этих генно-инженерных конструкций для увеличения частоты трансдукции заранее определенных клеток-мишеней посредством ретровирусного вектора и процедуры трансплантации с использованием трансдуцированных клеток.
Еще одним предпочтительным вариантом осуществления настоящего изобретения предусматривается способ локализации количества вируса, включающий культивирование среды, содержащей вирус в присутствии необходимого имобилизованного количества фибронектина или фрагментов фибронектина, обеспечивающего связывание вирусом области связывания гепарина П фибронектина с целью локализации количества вируса.
Другими предпочтительными вариантами осуществления настоящего изобретения предусматриваются культуры трансдуцированных жизнеспособных кроветворных и других клеток, содержащих чистый и/или иммобилизованный фибронектин или его фрагменты, а также наборы для опосредованного вирусом переноса ДНК в клетки, как это далее рассматривается в описании изобретения.
Целью настоящего изобретения является создание способов эффективного инфицирования ретровирусом клеток млекопитающих.
Другой целью настоящего изобретения является создание способов переноса генов с помощью ретровирусных векторов, которые не требуют сокультивирования.
Еще одной целью настоящего изобретения является создание усовершенствованных способов и культур клеток для аутологической и/или аллогенной трансплантации клеток.
Эти и другие цели, преимущества и отличительные особенности настоящего изобретения станут совершенно очевидными из приводимого ниже описания изобретения.
Описание чертежей
Фиг. 1 - схематическое изображение молекулы фибронектина, включающей фрагменты химотрипсина.
Фиг. 2 - изображение эффективности инфицирования коммитированных клеток-предшественников человека в присутствии фрагментов фибронектина с использованием вектора TKNEO, как это далее описывается в примере 1, см. ниже.
Фиг. 3 - сравнение эффективности инфицирования различных коммитированных кроветворных клеток-предшественников человека в присутствии фрагментов фибронектина с использованием вектора TKNEO, как это далее описывается в примере 1, см. ниже.
Фиг. 4 - сравнение наличия аденозиндезаминазы человека у мышей, трансплантированной вместе с клетками костного мозга, которые были трансдуцированы посредством (i) сокультивирования (столбцы 2-4), (ii) инфицирования надостадочной жидкости в присутствии иммобилизованных фрагментов фибронектина (столбцы 5 - 7) и инфицирования надосадочной жидкости на основе бычьего сывороточного альбумина (столбцы 8 - 10), как это далее описывается в примере 7, см. ниже. Контрольные пробы для аденозиндезаминазы человека показаны в столбцах 1 и 12, а для аденозиндезаминазы мышей - в столбце 11.
Фиг. 5 - изображение связывания ретровируса с фрагментами фибронектина, как это далее описывается в примере 8, см. ниже.
Фиг. 6 - изображение того, что связывание ретровируса с фрагментами фибронектина зависит от дозы, как это далее описывается в примере 8, см. ниже.
Фиг. 7 - схематическое изображение, иллюстрирующее разные фрагменты рекомбинантного фибронектина, используемого в примерах 9 - 11, см. ниже.
Фиг. 8 - изображение связывания ретровируса с разными фрагментами фибронектина, включая несколько рекомбинантных фрагментов, как это описывается в примере 9, см. ниже.
Фиг. 9 - изображение того, как гепарин блокирует связывание ретровируса с фрагментами фибронектина, как это описывается в примере 9, см. ниже.
Фиг. 10 - изображение эффективности инфицирования ретровирусом кроветворных клеток мышей в присутствии разных фрагментов фибронектина, как это далее описывается в примере 10, см. ниже.
Фиг. 11 - сравнение наличия аденазиндезаминазы человека у мышей, которым были трансплантированы клетки костного мозга, трансдуцированные посредством (i) сокультивирования, (ii) инфицирования надосадочной жидкости с использованием разных фрагментов фибронектина и (iii) инфицирования надосадочной жидкости с использованием бычьего сывороточного альбумина, как это описывается в примере 11, см. ниже.
Описание предпочтительных вариантов осуществления изобретения
Для получения более четкого представления о принципах настоящего изобретения ниже приводится описание некоторых вариантов его осуществления. Тем не менее, следует помнить, что они никоим образом не ограничивают объем изобретения, причем предполагается, что специалистам в этой области должны быть очевидны изменения, модификации и способы применения принципов по настоящему изобретению, приведенные здесь в качестве иллюстрации.
Как указывалось выше, настоящим изобретением предусматриваются способы увеличения частоты трансдукции жизнеспособных клеток под действием таких вирусов, как ретровирусы. Настоящим изобретением предусматриваются также способы эффективного переноса генов в жизнеспособные клетки с использованием рекомбинантных ретровирусных векторов, способы получения трансдуцированных клеток, а также способы и материалы для получения аутологичных и других клеточных трансплантатов.
Одной отличительной особенностью настоящего изобретения является открытие того, что фибронектин и его фрагменты, содержащие область адгезии клеток CS-1 фибронектина, значительно усиливают опосредованный ретровирусом перенос генов в такие клетки, как кроветворные клетки, например коммитировнные клетки-предшественники и стволовые кроветворные клетки или инициирующие клетки долгоживущей культуры, имеющие рецептор фибронектина и, таким образом, обладающие способностью связывания с фибронектином или его фрагментами. Весьма благоприятным является тот фактор, что увеличение эффективности не требует сокультивирования с продуцирующими вирус клетками. Другие отличительные особенности настоящего изобретения основываются на открытии связывающей вирусы области фибронектина, расположенной в области связывания гепарина П. Эту связывающую вирусы область можно использовать для локализации вирусных частиц во многих применениях, включающих, например, широкий диапазон генно-инженерных конструкций, предназначенных для доставки вируса к клетке-мишени.
Рекомбинантные вирусные векторы в соответствии с определенными предпочтительными аспектами настоящего изобретения содержат экзогенную ДНК и не являются патогенными, то есть у них имеется дефект репликации. Эти векторы эффективно переносят, точно и стабильно интегрируют экзогенную ДНК в ДНК клеток-хозяев, таких как клетки животных, в частности клетки млекопитающих. Например, в соответствии с настоящим изобретением нуклеотидную последовательность, включающую ряд оснований из кодирующей последовательности представляющего интерес гена, можно включить в рекомбинантный ретровирусный вектор с целью переноса гена под контролем соответствующего промотора. Таким промотором обычно является экзогенный промотор. В этой связи экзогенная ДНК может содержать ДНК, полученную натуральным или искусственным путем, и может состоять из частей, полученных из гетерологичных источников, причем эти части могут представлять собой натуральные или химические синтезированные молекулы и могут быть соединены посредством лигирования или других известных способов. Как указывалось выше, введенная нуклеотидная последовательность должна находиться под контролем промотора и считываться в прямом направлении. Говоря другими словами, последовательность промотора обычно находится в обратном направлении (то есть в 5' направлении) от кодирующей последовательности. Хорошо известно, что здесь могут присутствовать или отсутствовать другие регулирующие элементы (например, последовательности энхансеров), которые взаимодействуют с промотором и инициирующим кодоном транскрипции с целью получения транскрипции экзогенной кодирующей последовательности. Фраза "под контролем" подразумевает наличие таких элементов, которые необходимы для получения транскрипции введенного гена. Кроме того, рекомбинантная ДНК предпочтительно включает терминирующую последовательность, находящуюся в прямом направлении от введенной кодирующей последовательности.
Можно использовать ретровирусные векторы, которые включают экзогенную ДНК, представляющую селектирующий маркер или обладающую другим селективным преимуществом. Например, векторы могут содержать один или несколько экзогенных генов, которые являются резистентными к разным селективным агентам, включающим такие антибиотики, как неомицин. Репрезентативные векторы, которые можно использовать в этом изобретении, включают, например, вектор N/ZipTKNEO (TKNEO) (титр: 1•105 G418r колониеобразующих единиц/мл на клетки N1H 3N3), вектор ZipPGK-hADA и вектор ZipPGK-mADA, которые ранее описывались Морицем и др. (1993) в журнале J. Exp. Med. 178:529. В векторе TKNEO неофосфотрансферазные последовательности выражены в виде смысловой ориентации (по отношению к 5' длинному концевому повтору - LTR) через промотор тимидинкиназы вируса герпеса простого. Этот вектор содержит ген селекируемого маркера, который обеспечивает резистентность к неомицину, что облегчает идентификацию трансдуцированных клеток. В векторе ZipPGK- hADA кДНК аденозиндезаминазы человека ("hADA") выражается в смысловой ориентации по отношению к 5' длинному концевому повтору через промотор фосфоглицераткиназы человека (PGK). Он содержит только одну экспрессируемую генетическую последовательность и не имеет селектируемого маркера доминанты. Вектор ZipPG-K-mADA (PGK-mADA) идентичен вектору ZipPGK-hADA за исключением того, что кДНК аденозиндезаминазы человека заменена ДНК аденозиндезаминазы мышей (mADA). Эти и другие вирусные векторы и способы их получения хорошо известны и их применение в настоящем изобретении не может вызвать затруднений у специалистов в этой области.
Вирусные векторы, используемые в этом изобретении, обладают способностью связываться с аминокислотной последовательностью области связывания гепарина П фибронектина, в том числе и фибронектина человека. Поскольку настоящее изобретение не ограничено теоретическими выкладками, то можно предположить, что совместная локализация вируса и клетки-мишени посредством привязывания вируса и клетки к соответствующим функциональным областям способствует увеличению трансдукции клетки вирусом. В этом отношении способность вируса связываться с аминокислотной последовательностью области связывания гепарина П и, следовательно, обеспечивать эффективное достижение целей настоящего изобретения можно легко подтвердить с помощью стандартных процедур, аналогичных тем, которые описываются в приводимых ниже примерах 8 и 9. То есть эти анализы позволяют определить степень связывания вирусных частиц с иммобилизованными полипептидами, содержащими область связывания гепарина П с тем, чтобы противостоять вымыванию из иммобилизованной полипептидной матрицы. Коротко говоря, содержащую вирус надосадочную жидкость можно инкубировать в лунке с иммобилизованным полипептидом, включающим область связывания гепарина П фибронектина. Эту лунку затем интенсивно промывают буфером, представляющим собой физиологический раствор, после чего в этой лунке инкубируют клетки-мишени для вируса с целью определения в ней уровня инфицирующей активности. Определяют снижение инфицирующей активности или титра по сравнению с исходной содержащей вирус надосадочной жидкостью и сравнивают полученный показатель с аналогичной контрольной серией (полученной, например, при использовании лунки, покрытой бычьим сывороточным альбумином). Значительно более высокий титр в лунке, содержащей область гепарина П, по сравнению с контрольной лункой означает, что данный вирус пригоден для использования в соответствии с целями настоящего изобретения. Для облегчения процедуры скрининга вирусный вектор может содержать ген селектируемого маркера, как это описывалось выше.
Фрагменты фибронектина, предназначенные для использования в этом изобретении, могут иметь естественное или искусственное происхождение и могут быть получены из природных материалов с достаточно высокой степенью чистоты, например, так как это описывалось ранее Руослахти и др. (1981) в журнале J. Biol. Chem. 256: 7277; Пател и Лодиш (1986) в журнале J. Cell Biol. 102:449; и Бернарди и др. (1987) в журнале J. Cell Biol. 105:489. В связи с этим ссылка в этом изобретении на чистый фибронектин или его фрагменты означает, что они не содержат других белков, вместе с которыми обычно встречается фибронектин. Чистый фибронектин или его фрагменты, предназначенные для применения в этом изобретении, можно также получить рекомбинантным путем, например, так как это описывается в патенте США N 5198423, выданном 30 марта 1993 г. Тагучи и др. с передачей прав на патент Такара Шузо Компани, Лтд., Киото, Япония. В частности, рекомбинантные фрагменты, идентифицируемые в приводимых ниже примерах, как Н-271, Н-296, CH-271, CH-296 и C-CS1, а также способы их получения, детально описываются в противопоставленном патенте. Фрагмент C247, используемый в приводимых ниже примерах, был получен в соответствии с описанием, приведенным в патенте США N 5102988. Эти фрагменты или фрагменты, из которых они могут быть получены стандартными способами, получают путем культивирования клеток E.coli, зарегистрированных научно-исследовательским институтом ферментации агентства промышленной науки и технологии, Япония, под названиями PERM P-10721 (Н-296), FERM ВР-2799 (C-277, связанный с Н-271 с помощью метионина), FERM ВР-2800 (C-277, связанный с Н-296 с помощью метионина) и FERM ВР-2264 (Н-271) и описанных также в патенте США N 5198423. Кроме того, полезная информация об используемых в этом изобретении фрагментах фибронектина или исходных материалах для получения таких фрагментов содержится в работе Кимизука и др., опубликованной в журнале J. Biochem., 110, 284-291 (1991), в которой более подробно описываются вышеуказанные рекомбинантные фрагменты; в журнале EMBO J. 1755-1759 (1985), где описывается структура гена фибронектина человека; и в журнале Biocnemistry, 25, 4936-4941 (1986), где рассматривается область связывания гепарина П фибронектина человека. Было установлено, что фрагменты фибронектина, содержащие как область адгезии клеток CS-1, так и область связывания гепарина П, например, в виде 30 или 35 кд фрагмента (фибронектин 30/35) и в виде разных рекомбинантных фрагментов, рассматриваемых в приводимых ниже примерах, значительно увеличивают эффективность переноса генов в кроветворные клетки, и в настоящем изобретении предпочтение отдается именно им. Необходимо понять, что полипептид или полипептиды, относящиеся к фибронектину и используемые в этом изобретении, образуют аминокислотную последовательность, обеспечивая связывающую клетки активность области адгезии клеток CS-1 фибронектина, а также аминокислотную последовательность области связывания гепарина П фибронектина, которая обеспечивает связывание вируса. Вполне понятно, что необходимая активность связывания клеток и вирусов может быть обеспечена как нативными аминокислотными последовательностями этих функциональных областей фибронектина, так и аминокислотными последовательностями, которые отличаются от нативных последовательностей, но обладают аналогичной активностью связывания клеток и вирусов. Подобные аминокислотные последовательности характеризуются значительной гомологией в отношении соответствующих нативных последовательностей и могут включать такие последовательности, в которых аминокислоты были удалены, замещены и/или модифицированы, обеспечивая при этом получение аминокислотной последовательности с требуемыми характеристиками связывания клеток или вирусов.
Разработанные биотехнологии достигли такого уровня, при котором может быть легко произведена делеция, замещение, добавление или другая модификация аминокислот в функциональных областях. Полученные аминокислотные последовательности затем могут быть подвергнуты тестированию стандартными методами с целью определения требуемой активности связывания клеток или вирусов. Например, активность связывания вирусов мутанта или модифицированных форм области связывания гепарина П фибронектина можно тестировать так, как описывается ниже, в частности, в примерах 8 и 9, путем культивирования вирусов, промывки и анализов вирусных титров, что позволяет определить степень инфицирования по сравнению с контрольной пробой. С учетом приведенных здесь материалов анализы связывания будут представлять для специалистов в этой области обычную экспериментальную работу.
Связывание клеток с модифицированными или мутантными формами области адгезии клеток CS-1 фибронектина или с другими связывающими клетки полипептидами можно анализировать с помощью известных способов. Например, такие процедуры включают методы, описанные в журнале Nature 352: 438-441 (1991). Коротко говоря, пластиковые чашки покрывали связывающим клетки полипептидом и предназначенную для анализа популяцию клеток помещали в среду на время от 30 минут до 2 часов. После инкубации клетки, не связанные с белком, выделяли, считали и анализировали в отношении жизнеспособности. Клетки, связанные с полипептидом, также выделяли с помощью трипсина или буфера для диссоциации клеток (например, Gibco), считали и тестировали в отношении жизнеспособности. В некоторых случаях, например в случае колониеобразующих кроветворных клеток, такие клетки культивировали еще в течение 12-14 дней, после чего оценивали колониеобразующие характеристики клеток. Затем высчитывали процент адгезивных клеток и сравнивали этот результат с контрольной пробой, полученной при использовании бычьего сывороточного альбумина. Значительное связывание клеток-мишеней с анализированным полипептидом свидетельствует о том, что данная комбинация полипептида и клеток пригодна для достижения целей настоящего изобретения, и этот полипептид может быть присоединен к связывающему ретровирус фрагменту фибронектина с образованием конструкции по настоящему изобретению, предназначенной для увеличения инфицирования клеток-мишеней вирусным вектором.
В соответствии с наиболее характерными аспектами настоящего изобретения связывающий вирус полипептид, используемый для усиления трансдукции под действием ретровирусных векторов, должен включать (i) первую аминокислотную последовательность, которая соответствует Ala1690 - Thr1960 области связывания гепарина П фибронектина человека, которая представлена формулой (идентификационный номер последовательности 1 см. в конце описания).
или в достаточной мере подобную аминокислотную последовательность, обладающую способностью связывать ретровирус; и (ii) вторую аминокислотную последовательность, которая соответствует одной части области связывания IIICS фибронектина человека (область связывания клеток CS-1), которая представлена формулой (идентификационный номер последовательности 2, см. в конце описания):
или в достаточной мере подобную аминокислотную последовательность, обладающую способностью связывать кроветворные клетки, в частности первичные клетки-предшественники и/или долгоживующие репопулирующие (стволовые) клетки.
Как отмечалось выше, совершенно очевидно, что в объем настоящего изобретения входят модификации и/или мутации этих нативных последовательностей, если полученная аминокислотная последовательность в достаточной степени подобна нативной последовательности и обладает способностью связывать вирус (в случае области связывания гепарина П), а также способностью связывать клетки-мишени (в случае области CS-1).
Одним аспектом настоящего изобретения предусматривается метод соматической генотерапии, который включает in vitro клеточную терапию и последующую трансплантацию клеток-мишеней хозяину, который также известен как "прививка" хозяину трансдуцированных клеток-мишеней. Кроветворные или другие клетки могут быть взяты у человека или другого млекопитающего с помощью стандартных методов. Например, кроветворные клетки можно получить из костного мозга, крови периферической кровеносной системы человека-донора или пуповинной крови. Полученные кроветворные клетки могут необязательно быть подвергнуты обработке с помощью стандартных методов, чтобы обогатить их стволовыми клетками и/или первичными клетками-предшественниками. Кроветворные клетки затем можно инкубировать соответствующим образом, например, на планшетах с культурами тканей. Необязательно, на этом этапе несвязанные (adherent - negative) мононуклеарные клетки с низкой плотностью можно предварительно стимулировать до инфицирования ретровирусами. Предварительное стимулирование, известное в этой области и применяемое в этом изобретении, представляет собой процесс воздействия на клетки симулирующими рост факторами до инфицирования ретровирусами. Было доказано, что подобное предварительное стимулирование улучшает трансдукцию кроветворных клеток ретровирусами.
Вслед за предварительным стимулированием клетки можно собирать и инкубировать с фибронектином или его фрагментами, как описывается в этом изобретении, в результате чего увеличивается частота трансдукции клеток под действием ретровирусов. Клетки предпочтительно инкубируют с очищенным и/или нерастворимым, например иммобилизованным, фибронектином или его фрагментами. После этого клетки инфицируют рекомбинантным вирусом, например ретровирусом, содержащим ген, служащим для устранения недостаточности или дефекта фермента или другого белка в клетках, в присутствии фибронектина или его фрагмента в количестве, достаточном для увеличения чистоты трансдукции клеток под действием вируса. Полученные трансдуцированные кроветворные клетки можно затем ввести, например, внутривенно животному-реципиенту, предпочтительно аутологическому донору, но имеющему также аллогенные трансплантаты, что особенно верно при использовании в качестве трансплантата клеток пуповинной крови, как это описывается ниже.
Способы по настоящему изобретению можно использовать в области маркирования генов или генотерапии для лечения разных заболеваний, в том числе заболеваний костного мозга, включающих, например, разные формы рака и лейкоз, заболеваний, связанных с недостаточностью или дефектами белка, и для модификации кроветворных клеток с целью снижения их резистентности к другим терапевтическим методам, таким как химиотерапия. Типичными заболеваниями, для лечения которых можно использовать это изобретение, являются аденозиндезаминазная недостаточность, например вызванный ею тяжелый комбинированный иммунодефицит, острый миелолейкоз у детей, нейробластома, острый миелолейкоз у взрослых и острый лимфобластный лейкоз.
В соответствии с одним предпочтительным вариантом осуществления настоящего изобретения клетки, используемые для клеточной трансплантации, получают из пуповинной крови человека. Таким образом, пуповинную кровь можно собирать и обогащать жизнеспособными примитивными клетками-предшественниками и/или стволовыми клетками, например, путем получения популяции несвязанных мононуклеарных клеток с низкой плостностью. Такую популяцию затем при желании предварительно стимулируют и инкубируют в присутствии ретровирусного вектора и иммобилизованного и/или очищенного фибронектина или его фрагментов с целью увеличения эффективности трансдукции клеток под действием вектора. В этой связи было установлено, что трансдукция примитивных кроветворных и/или стволовых клеток, полученных из пуповинной крови, значительно увеличивается в присутствии фибронектина или его фрагментов, даже если фибронектин не образует часть комплексной внеклеточной матрицы в пуповинной крови и даже если примитивные клетки-предшественники и стволовые клетки из пуповинной крови имеют характеристики, отличные от показателей костного мозга. В частности, стволовые клетки пуповинной крови характеризовались как CD34+, HLA-DR+, а стволовые клетки из костного мозга характеризовались как CD34+, HLA-DR-. Сделанное авторами изобретения открытие того, что примитивные клетки-предшественники, полученные из пуповинной крови, эффективно трансдуцируются в присутствии фибронектина или его фрагментов, позволяет использовать удобный и значительно обогащенный стволовыми клетками источник кроветворных клеток. Кроме того, данные, свидетельствующие об успешной трансплантации многочисленным больным аллогенных трансплантатов пуповинной крови, обогащенной примитивными клетками-предшественниками и стволовыми клетками, делает пуповинную кровь предпочтительным источником получения кроветворных клеток. См. Кохли-Кумер и др., Brit. Haematol. 85:419-422 (1993); Броксмейер и др., Blood Cell 17:313-329 (1991); Глюкман и др., Br. J. Haematol. 45:557 (1980); Хидельберг; Sptinger - Verlag, стр. 60-68 (1968); Вагнер и др., Blood, 79: 1874-1881 (1992) и Вагнер и др., Blood, 82-86а (реферат).
Собранные трансдуцированные кроветворные или другие клетки можно при желании подвергнуть тестированию с целью определения эффективности трансдукции и экспрессии гена. Например, значительные улучшения опосредованного ретровирусом переноса генов, достигаемые в результате применения настоящего изобретения, демонстрируются приводимыми ниже примерами, в которых описываются некоторые тесты, свидетельствующие о высокой эффективности инфицирования и переноса генов ретровирусами в присутствии фибронектина или его фрагментов. В частности, кроветворные клетки мышей, инфицированные ретровирусом PGK-hADA, характеризуются высокими уровнями перенесенной кДНК аденозиндезаминазы. Аналогичным образом, колонии клеток-предшественников человека, инфицированные вирусом PGK-mADA, характеризовались уровнями аденозиндезаминазы мышей, которые в 10 раз превосходили уровни эндогенного белка аденозиндезаминазы человека. Поэтому, для того чтобы точно проанализировать эффективность переноса, колонии клеток-предшественников считали трансдуцированными только в том случае, если экспрессия перенесенной аденозиндезаминазы мышей равнялась или превышала уровни эндогенной аденозиндезаминазы человека. Высокие уровни экспрессии неогена, полученного из вектора TKNEO, были обнаружены посредством лекарственной устойчивости G418 при выполнении анализа на неофосфотрансферазную (продукт неогена) активность.
Как указывалось выше, способы по настоящему изобретению можно успешно выполнить без необходимости сокультивирования в присутствии ретровирусных клеток-продуцентов. Так, в соответствии с одним из аспектов этого изобретения опосредованный ретровирусом перенос генов может осуществляться при отсутствии каких-либо клеток кроме кроветворных клеток-мишеней или других клеток. Например, можно культивировать клетки-продуценты, содержащие плазмиду ретровирусного вектора, и собирать надосадочную жидкость. Содержащую ретровирусы надосадочную жидкость можно затем использовать для инфицирования кроветворных клеток в присутствии фибронектина и/или его фрагментов, которые предпочтительно являются иммобилизованными, то есть нанесенными на субстрат, на котором производится инфицирование, или контактирующими со средой, предназначенной для инфицирования. В этой связи любые клетки-продуценты, которые образуют ретровирусы с высоким титром и без вирусов-помощников, считаются пригодными для применения в этом изобретении. Они включают, например, упаковочные клетки, такие как Psi-2, C2, PA12, PA317 и GP+envAM12, а также многие другие известные линии упаковочных клеток.
В соответствии с другими отличительными особенностями настоящего изобретения сильное связывание вируса с аминокислотами в области связывания гепарина П фибронектина можно использовать для создания систем доставки, используемых для вирусной терапии в отношении клеток самых разных типов. Так, полипептид, включающий область связывания ретровируса из фибронектина, может быть ковалентно связан с любым лигандом, который придает этой конструкции специфичность в отношении клеток-мишеней. Такой подход делает ненужным необходимое ранее создание специальных линий ретровирусных клеток для каждой клетки-мишени (Kasahara N. , A, M. Dozy и G.W. Kan, Science, т. 266, стр. 1373-1376 (1994) и Valsesia-Wittmann, S., A. Drynda, G. Deleage, M. Auailley, J. M. Heard, O. Danos, G. Verdier и F.Z. Cosset, J. Virol. т. 68, стр. 4609 - 4619 (1994). Специфичность конструкции клеток-мишеней может быть достигнута благодаря применению лиганда, включающего, например,
1) белок, обеспечивающий адгезию клеток,
2) гормоны или цитокины,
3) моноклональные антитела, относящиеся к клеткам-мишеням,
4) углеводы, которые связывают клетки-мишени (Г. Ашвел и др., Annu, Rev. Biochem., т 51, стр. 531-554 (1982)),
5) метаболиты для клеток-мишеней или
6) функциональные полипептиды, связывающие клетки-мишени. Эффективность конструкции для доставки генов можно улучшить, включив в нее несколько областей связывания вируса гепарина П, в результате чего увеличивается количество вирусных частиц, доставляемых к клеткам-мишеням. Например, было установлено, что связывающая клетки область фибронектина человека, которая соответствует Pro1239 - Ser1515, как это описывается в патенте США N 5198423, обеспечивает связывание с клетками, в том числе с клетками ВНК и B16-F10 (Кимизука и др., J. Biochem., т. 110, стр. 285-291 (1991)). Кроме того, было установлено, что область гепарина П сама по себе обеспечивает связывание с фибробластами, клетками эндотелия и раковыми клетками. Эти полипептидные последовательности могут присоединяться к области связывания ретровируса из фибронектина и к заранее определенным клеткам-мишеням, предназначенным для инфицирования ретровирусом.
Типичные применения в системе кроветворных клеток включают также конструкцию эритропоэтина или G-CSF, присоединяемую к области связывания ретровируса фибронектина для доставки вируса соответственно к эритроидным или гранулоцитным клеткам- предшественникам. Другим применением в соответствии с настоящим изобретением является соединение области или областей связывания ретровируса с лигандом, который специфично или преимущественно осуществляет связывание с раковыми клетками. Например, было установлено, что на in vitro и даже in vivo рост раковых клеток молочной железы можно влиять с помощью веществ, соединяющихся с рецепторами на клетках-мишенях, подобных производным высвобождения лютеинизирующего гормона Эммонс Г. и др. Hum. Reprod., 9:1364- 1379 (1994), эстрогенам, Толчер А.В., Oncol. 8:39-43 (1994) или антиэстрогенам, Ховелл А. и др., Lancet,
345: 29-30 (1995), прогестогенам или антипрогестогенам, Клижн. Ф.Г., и др., приложение 9 к журналу Hum. Reprod, 1:181-189 (1994), Гриффитс К. и др. , Semin. Oncol. , 21:672-687 (1994), которые служат в качестве лигандов в конструкциях по настоящему изобретению, содержащих одну или несколько областей связывания вирусов из фибронектина. Как видно из нижеследующих примеров, воздействие на (раковые) клетки щитовидной железы может быть весьма специфическим благодаря использованию конструкций с джодидом, а на (раковые) клетки печени можно воздействовать с помощью конструкций, содержащих альфалипопротеины высокой плотности или их части. И, наконец, конструкции моноклональных антител и области связывания ретровирусов фибронектина позволяют целенаправленно воздействовать на любые клетки и органы, для которых предназначено антитело. Таким образом можно целенаправленно воздействовать на целый ряд клеток млекопитающих, делая упор на способность области связывания ретровируса фибронектина связывать и локализовать вирусные векторы.
Другой предпочтительный вариант осуществления настоящего изобретения включает получение конструкции, которую можно использовать для усиления вирусной трансдукции клетки-мишени. Связывающая вирусы аминокислотная последовательность области связывания гепарина П фибронектина присоединяется к лиганду, с которым связана клетка-мишень. Как указывалось выше, лигандом может быть, например, полипептид из фибронектина или другого белка (включая обеспечивающий адгезию клеток белок, в частности ламинин, коллаген, витронектин, остеопонтин или тромбоспондин), гормон, метаболит, антитело (включая моноклональные антитела) или любой другой лиганд, обладающий способностью связывания, предпочтительно специфического, с клеткой-мишенью. Полученную общую конструкцию можно использовать в иммобилизованной форме аналогично тому, как используются полипептиды фибронектина, специально рассматриваемые в приводимых ниже примерах.
Такие конструкции и подходы на основе целенаправленного воздействия на клетки могут in vitro применяться, как указывалось выше, а также в процессе in vivo целенаправленного переноса ретровируса с учетом различных факторов, таких как стабильность и специфичность конструкции и взаимодействие ретровирусной конструкции в физиологических условиях. Специфичность можно также улучшить путем изменения системы доставки с целью локализации доставки конструкции только к клеткам-мишеням, например, с помощью катеризации воротной вены для целенаправленного воздействия на клетки печени.
Опосредованный ретровирусом перенос ДНК можно осуществлять с помощью наборов, специально предназначенных для практического осуществления методов по настоящему изобретению. Поэтому еще одним аспектом настоящего изобретения предусматриваются наборы, которые включают определенное количество чистого полипептида или рассмотренной выше конструкции, которая усиливает трансдукцию клеток-мишеней под действием ретровирусов, вместе с искусственным субстратом, на котором можно производить инфицирование ретровирусом. Полипептид или другая конструкция могут быть предусмотрены отдельно или нанесены на искусственный субстрат. В случае процедур инфицирования, предназначенных для кроветворных клеток человека, наборы могут также включать факторы роста кроветворных клеток для предварительной симуляции клеток. Кроме того, эти наборы могут включать рекомбинантные ретровирусные векторы, представленные выше для осуществления трансдукции. Эти наборы должны включать стерильную упаковку, которая надежно изолирует различные компоненты друг от друга, не допуская их смешивания во время работы с этим набором. Например, для этой цели обычно используются формованные пластиковые изделия с несколькими отделениями, предназначенными для раздельного хранения компонентов, входящих в состав набора.
Для более полного и глубокого понимания сущности настоящего изобретения приводятся следующие типичные примеры. Совершенно очевидно, что эти примеры являются иллюстративными и никоим образом не ограничивают объем изобретения.
ПРИМЕР 1
Перенос генов в клетки костного мозга с использованием вектора TKNEO
1.1. Получение надосадочной жидкости с вирусом
Клетки-продуценты GP+Env AM 12 (см. Марковиц и др. (1988) Virology 167: 400), содержащие ретровирусный плазмидный вектор TKNEO, культивировали в среде Дульбекко, модифицированной по способу Исков (IMOM, Gibco, Гэтерсберг, шт. Мэриленд), которая содержала 10% околоплодной сыворотки теленка (Hyclone, Логан, шт. Юта), 100 единиц/мл пенициллина и 100 микрограмм/мл стрептомицина (P/S, предоставлены фирмой "Gibco"). Содержащую вирус надосадочную жидкость собирали в течение ночи путем добавления 10 мл среды Дульбекко, модифицированной по способу Исков, содержащей 20% околоплодной сыворотки теленка, на конфлюэнтных планшетах. Собранную среду фильтровали через 0,45-микронные фильтры (Gelman Sciences, Энн-Арбор, шт. Мичиган) и хранили до использования при температуре -80oC.
1.2 Получение фрагментов фибронектина
Фибронектин очищали от плазмы человека (Lifesource Гленвью, шт. Иллинойс), как описывалось ранее в работе Руослахти и др., Methods Enzymol. 82: 803-831 (1982), за исключением того, что колонку с желатином и агарозой промывали 1 М раствором мочевины до элюирования фибронектина 4 М раствором мочевины. Очищенный фибронектин подвергали медленному диализу при температуре 4oC с использованием 10 ммолей 3-(циклогексиламино)-1-пропансульфоновой кислоты, 150 ммолей NaCl, 2 ммолей CaCl2 с показателем pH 11,0 и хранили в виде аликвот при температуре -80oC. Область связывания клеток химотрипсина (CS-1) и фрагменты связывания гепарина П фибронектина очищали, как описывалось выше (Руослахти и др. (1982), там же, Патель и Лодиш, J., Cell Biol., 102, стр. 449-456 (1986) и Бернарди и др., J. Cell, Biol. 105, стр. 489-498 (1987). Три основных связывающих гепарин фрагмента (30 kD, 35kD) и 42kD) были получены в 1 М растворе элюата NaCl из колонки с гепарином и агарозой. Для дальнейшей очистки этих связывающих гепарин фрагментов 1 М раствор элюата NaCl подвергали диализу в течение ночи при температуре 4oC с использованием 10 ммолей трис-HCl, показатель pH 7, и пропускали через анионообменную колонку (2 мл быстрый поток диэтиламиноэтил-сефарозы (Farmacia Fine Chemicals, Упсала, Швеция)/мг белка), которую уравновешивали 10 ммолями трис-HCl, показатель pH 7,0. Фрагменты 30/35 kD собирали в виде несвязанной фракции, а фрагмент 42kD элюировали из колонки с помощью 100 ммолей NaCl. Из 500 мг фибронектина было получено примерно 26 мг фрагментов 30/35 kD) и 4 мг фрагмента 42kD. Фрагмент 42kD, а не фрагменты 30/35kD распознавались антителом в области связывания фибрина при определении с помощью метода вестерн-блотирования. Кроме того, фрагмент 42kD связывается с колонкой с иммобилизованным фибрином и сефарозой.
Для использования в процедуре инфицирования фрагменты фибронектина иммобилизовали на 35 или 100 мм чашках Петри (Falcon, Линкольн-Парк, ш. Нью-Джерси) при концентрации 75 пмолей/см2, как описывалось Пателем и Лодишем (1986), см. выше. На контрольные чашки аналогичным образом наносили 2% (не содержащий фибронектина) бычьего сывороточного альбумина (BSA, Boehringer Mannheim, Мангейм, Германия).
1.3. Процедура инфицирования ретровирусом
Пробы костного мозга, взятые у здоровых взрослых доноров, собирали в пробирки, содержащие стерильный, без консерванта гепаринсульфат натрия, в соответствии с процедурами, утвержденными Институционным советом медицинского колледжа университета шт. Индиана. Мононуклеарные клетки с низкой плотностью получали путем центрифугирования на Ficoll-Hypaque (плотность 1,077 г/мл. Pharmacia, Пискатавэй, шт. Нью-Джерси) в течение 45 минут при температуре 25oC. Пластичные связанные клетки удаляли из клеток костного мозга с низкой плотностью путем дополнительной инкубации на планшетах с культурой ткани в течение 4-16 часов при температуре 37oC с 5% CO2 в среде Дульлекко, модифицированной по способу Исков, с добавлением 2% околоплодной сыворотки теленка.
Несвязанные мононуклеарные клетки с низкой плотностью предварительно стимулировали до инфицирования ретровирусом, так как описывали Лускей и др. (1992), Blood 80-396, в течение 48 часов при температуре 37oC с 5% CO2 в среде Дульбекко, модифицированной по способу Исков, содержащей 20% околоплодной сыворотки теленка, 100 единиц/мл рекомбинантного интерлейкина 6 человека, 100 нг/мл фактора роста рекомбинантных стволовых клеток человека (предоставлены фирмой "Amges" Фаузенд-Оукс, шт. Калифорния) и смеси пенициллина/стрептомицина при плотности клеток 1 • 106 клеток/мл на чашках Петри. Предварительно стимулированные клетки собирали путем интенсивного пипетирования с целью удаления клеток, слабо прилипших к пластику.
Предварительно стимулированные клетки (5 • 105 клеток/мл) инкубировали в течение 6 часов на планшетах, на которые был нанесен бычий сывороточный альбумин (контрольные планшеты), фибронектин или его фрагменты (подвергнутые ультрафиолетовому облучению для достижения лучшего связывания белков с пластиковым планшетом), а затем инфицировали содержащей вирус надосадочной жидкостью в присутствии факторов роста (описанных выше) и 7,5 микрограмм/мл полибрена (Aldrich Chemical, Милуоки, шт. Висконсин). Содержащую вирус надосадочную жидкость заменяли (включая факторы роста и 5,0 микрограмм/мл полибрена) через 2 часа, после чего клетки инкубировали в течение еще 12-24 часов. При каждой смене среды вновь добавляли несвязанные клетки.
В соответствии с процедурой инфицирования несвязанные клетки сливали, а связанные кроветворные клетки собирали из культур с использованием буфера для диссоциации клеток (без фермента/на основе забуференного фосфатом физиологического раствора, Gibco) в соответствии с инструкциями изготовителя. В несвязанную фракцию вводили связанные клетки, дважды промывали и производили подсчет. Собранные клетки культивировали на метилцеллюлозном предшественнике для клоногенных анализов или на долгоживущих культурах костного мозга.
1.4. Долгоживущие культуры костного мозга.
Анализы инициирующих клеток долгоживущей культуры (стволовая клетка человека) выполняли в соответствии с ранее описанными методами при внесении незначительных изменений. Сатерленд и др. Blood, 74:1563 (1989). Вкратце этот метод можно описать следующим образом: 0,5-1•106 инфицированных клеток высевали в долгоживущие культуры костного мозга на конфлюэнтные, предварительно облученные (как описывалось выше) аллогенные фибробласты костного мозга человека в 5 мл среды Дульбекко, модифицированной по способу Исков, которая содержала 10% околоплодной сыворотки теленка, 10% сыворотки лошади (Sigma), смесь пенициллина/стрептомицина, 1 • 10-5 М раствор гидрокортизона (Upjohn, Каламазу, шт. Мичиган) и 320 мосмолей хлористого натрия на 6-луночных планшетах, заполненных культурой ткани (Costar Кеймбридж, шт. Массачусетс). Долгоживущую культуру костного мозга инкубировали при температуре 33oC в 5% CO2 и еженедельно удалили 50% среды и несвязанных клеток. Через пять недель инициирующие клетки долгоживущих культур уничтожали, используя буфер для диссоциации клеток для удаления связанных кроветворных клеток из фибробластов костного мозга. Несвязанные и связанные кроветворные клетки собирали и культивировали на метилцеллюлозе с целью получения колоний на основе инициирующих клеток долгоживущей культуры.
1.5. Клоногенные анализы на метилцеллюлозе
Анализы на метилцеллюлозе выполняли так, как описывалось Токсозом и др., Proc. Natl. Acad. Sci., США, т. 89, стр. 7350 (1992) с внесением незначительных изменений. Вкратце этот метод можно описать следующим образом: 2-5 • 104 инфицированных клеток костного мозга взрослого человека культивировали с 5 единицами/мл эритропоэтина (Epo, Amgen), 100 нг/мл фактора роста рекомбинантных стволовых клеток человека, 10 нг/мл рекомбинантного интерлейкина 3 человека (Genzyme, Кеймбридж, шт. Массачусетс) в 1 мл 2,4% метилцеллюлозы среды Дульбекко, модифицированной по способу Исков (Fluka, Ронконкома, шт. Нью-Йорк), которая содержала 25% околоплодной сыворотки теленка, 10% плазмы человека, 10-5 М раствора бетамеркаптоэтанола (Sigma) и смесь пенициллина/стрептомицина. Культуры инкубировали при температуре 37oC в атмосфере, состоящей из 5% CO2 и 95% воздуха, и на 13-й день под инвертированным микроскопом производили оценку колоний (> 50 клеток) на содержание гранулоцитов и макрофагов (гранулоцитарно-макрофагальная колониеобразующая единица), содержание миелоидных и эритроидных элементов (колониеобразующая единица смеси) или на содержание только эритроидных элементов (эритроидная бурстобразующая единица - БОЕэ).
1.6 Анализ инфицирования ретровирусом
Эффективность инфицирования вирусом TKNEO анализировали путем определения процента колоний на метилцеллюлозе, устойчивых к воздействию 1,5 мг/мл (сухой порошок, Gibco) G418 на 13-й день. В каждом эксперименте производили псевдозаражения путем инкубирования костного мозга на линии упаковочных клеток GP+EnvAM 12 без создания рекомбинантного вируса. Культура этих клеток, псевдозараженных 1,5 мг/мл G418, стабильно демонстрировала образование < 1% фоновых колоний.
1.7 Эффективность переноса генов в коммитированные клетки-предшественники
Эффективность трансдукции сравнивали путем инфицирования клеток костного мозга при культивировании на чашках, покрытых фрагментами фибронектина 30/35 или бычьим сывороточным альбумином. При создании этих условий не было обнаружено различий в количестве колоний, полученных после инфицирования без выполнения отбора. На фиг. 2 показано процентное значение колоний G418 после инфицирования. Более высокий процент колоний G418r был отмечен на фрагментах фибронектина 30/35 для клеток-предшественников всех типов, в том числе и тех, которые были выделены из клеток-предшественников с ограниченной линией дифференцировки (БОЕЭ и КОЕ-ГМ) и множественной линией дифференцировки (КОЕ-смесь). Инфицирование всех коммитированных клеток-предшественников увеличилось в 9 раз на фрагментах фибронектина 30/35 по сравнению с бычьим сывороточным альбумином.
1.8. Эффективность переноса генов в инициирующие клетки долгоживущей культуры
Перенос генов в инициирующие клетки долгоживущей культуры оценивали с использованием вектора TKNEO. Перенос генов в колонии, полученных из инициирующих клеток долгоживущей культуры, был обнаружен только после инфицирования на фрагментах фибронектина 30/35 (16% колоний G418' по сравнению с 0% колоний G-418r при использовании фрагментов фибронектина 30/35 по сравнению с бычьим сывороточным альбумином).
1.9. Влияние специфичности фрагментов фибронектина 30/35 на эффективность инфицирования кроветворных клеток
Для определения специфичности более высокой эффективности переноса генов, наблюдаемой при использовании фрагментов фибронектина 30/35, производили инфицирование TKNEO на планшетах, покрытых бычьим сывороточным альбумином, фрагментами фибронектина 30/35, интактным фибронетином, 115 мд фрагмента фибронектина, имеющего центральную область связывания клеток, содержащую тетрапептидную последовательность RGDS и 42 кд C-терминального фрагмента фибронектина (42FN), характеризующегося наличием области связывания гепарина П, но не имеющего последовательности CS-1 (фиг. 1). Инфицирование на бычьем сывороточном альбумине позволило получить 3±1% БОЕэ G418r, 1±1% КОЕ-ГМ G418r и 0±0% КОЕ-смеси G418r. В центральной области связывания клеток не было отмечено значительного увеличения колоний G418r, в то время как несколько более высокое инфицирование БОЕэ (6,0 ± 1%) было отмечено на фибронектине 42 (фиг. 3). Однако интактный фибронектин способствовал увеличению переноса генов во все коммитированные клетки-предшественники. Процентное значение колоний G418r после инфицирования на интактном фибронектине было меньше, чем на фрагментах фибронектина 30/35 во всех линиях дифференцировки, включая БОЕэ (16±2 по сравнению с 24±4%), КОЕ-ГМ (5±2 по сравнению с 20±4%) и КОЕ-смесь (6±1 по сравнению с 9±1; соответственно интактный фибронектин по сравнению с фрагментами фибронектина 30/35).
ПРИМЕР 2
Перенос генов в клетки костного мозга с использованием вектора PGK-mADA
2.1. Общие процедуры
Надосадочную жидкость с вирусом PGK-mADA получали так же, как описывалось для вектора TKNEO в примере 1. Химотрипсиновые фрагменты фибронектина (фиг. 1) получали аналогично примеру 1, при этом выполняли процедуру инфицирования ретровирусом, описанную в примере 1. Анализы инициирующих клеток долгоживущей культуры (стволовые клетки человека) и анализы на метилцеллюлозе выполняли в соответствии с примером 1.
2.2. Анализ инфицирования ретровирусом
Эффективность инфицирования вектором PGK-mADA определяли с помощью анализа белка путем выполнения электрофореза изофермента аденозиндезаминазы. Анализ отдельных колоний клеток-предшественников выполняли в соответствии с описанием, приведенным Морицем (1993), Лимом и др. (1989) Proc. Natl. Acad. Sci. , США, т. 86, стр. 8892. Для повышения точности анализа эффективности переноса трансдуцированными считали только те колонии, которые демонстрировали образование аденозиндезаминазы мышей на таком же или более высоком уровне по сравнению с эндогенной аденозиндезаминазой человека. Для анализа пулов колоний их выделяли из культуры на метилцеллюлозе, собирали в 1,5 мл микропробирки (Rainin, Вобурн, шт. Массачусетс), промывали теплой средой и забуференным фосфатом физиологическим раствором, центрифугировали и хранили при температуре -20oC. Для анализа аденозиндезаминазы клетки лизировали в 5-микролитровом буфере для лизирования путем повторения циклов замораживания и оттаивания, после чего выполняли электрофорез изофермента так же, как описывалось выше.
2.3. Эффективность переноса генов в коммитированные клетки-предшественники
Эффективность трансдукции сравнивали путем инфицирования клеток костного мозга при культивировании на чашках, покрытых фрагментами фибронектина 30/35 или бычьим сывороточным альбумином. В этих условиях не было обнаружено различий в количестве колоний, полученных после инфицирования без выполнения отбора. Как показано в таблице 1, эффективность инфицирования всех коммитированных клеток-предшественников была значительно выше на фрагментах фибронектина 30/35, чем на бычьем сывороточном альбумине. Как и ожидалось в случае вектора с более высоким титром (~1•107 вирусов/мл), эффективность трансдукции коммитированных клеток-предшественников была очень высокой. Из таблицы 1 видно, что инфицирование костного мозга на фрагментах фибронектина 30/35 посредством PGK-mADA позволило достичь почти 100% трансдукции коммитированных клеток-предшественников в двух отдельных экспериментах.
2.4. Эффективность переноса генов в инициирующие клетки долгоживущей культуры
В четырех независимых экспериментах, выполненных с использованием вектора PGK-wADA, значительная часть колоний клеток-предшественников, выделенных через 5 недель (то есть колонии инициирующих клеток долгоживущей культуры), характеризовались экспрессией гена аденозиндезаминазы мышей. Экспрессия составляла от 2/12 (17%) до 6/6 (100%) анализированных колоний (таблица 2). Экспрессия введенного гена аденозиндезаминазы мышей превосходила или по крайней мере равнялась величине активности эндогенной аденозиндезаминазы человека во всех колониях, считавшихся положительными. Эффективность инфицирования для PGK-mADA была выше, чем для TKNEO. Как видно из таблицы 2, инфицирование костного мозга на фрагментах фибронектина 30/35 посредством PGK-mADA позволило получить почти 100% трансдукцию коммитированных клеток-предшественников в двух отдельных экспериментах.
2.5. Влияние специфичности фрагментов фибронектина 30/35 на эффективность инфицирования кроветворных клеток
Эффективность переноса генов в инициирующие клетки долгоживущей культуры была более высокой на фрагментах фибронектина 30/35. Из- за относительно небольшого размера этих вторичных колоний, образованных инициирующими клетками долгоживущей культуры, возможность выполнения анализа белка в отдельных колониях была ограничена. После инфицирования вектором PGK-mADA на бычьем сывороточном альбумине, интактном фибронектине и фрагменте фибронектина 42 экспрессия аденозиндезаминазы мышей в колониях, образованных инициирующими клетками долгоживущей культуры, составила 0/6, 0/4 и 0/3, в то время как аналогичный показатель для колоний, инфицированных на фрагментах фибронектина 30/35, составил 3/5. Кроме того, когда несколько колоний, образованных инициирующими клетками долгоживущей культуры, объединяли перед выполнением анализа в двух дополнительных экспериментах, экспрессия аденозиндезаминазы мышей была обнаружена только после инфицирования на фрагментах фибронектина 30/35 и в гораздо меньшей степени на интактном фибронектине, при этом она отсутствовала при использовании фрагмента фибронектина 42 или бычьего сывороточного альбумина.
ПРИМЕР 3
Перенос генов в клетки костного мозга с использованием вектора PGK-hADA
3.1. Общая процедура
Надосадочную жидкость с вирусом PGK-hADA получали так же, как описывалось для вектора TKNEO в примере 1. Химотрипсиновые фрагменты фибронектина (фиг. 1) получали в соответствии с описанием, приведенным в примере 1, при этом выполняли процедуру инфицирования ретровирусом по примеру 1. Анализы инициирующих клеток долгоживущей культуры и анализы на метилцеллюлозе выполняли аналогично примеру 1.
3.2. Анализ инфицирования ретровирусом
Для анализа пулов колоний их выделяли из культуры на метилцеллюлозе, собирали в 1,5 мл микропробирки (Raini, Вобурн, шт. Массачусетс), промывали теплой средой и забуференным фосфатом физиологическим раствором, центрифугировали и хранили при температуре -20oC. Для анализа аденазиндезаминазы клетки лизировали в 5 микролитрах буфера для лизирования путем повторения циклов замораживания и оттаивания, после чего выполняли электрофорез изофермента, как это описывалось выше.
ПРИМЕР 4
Перенос генов в клетки пуповинной крови с использованием вектора TKNEO
4.1. Общая процедура
Надосадочную жидкость с вирусом TKNEO и химотрипсиновые фрагменты фибронектина (фиг. 1) получали так же, как описывалось в примере 1. Выполняли процедуру инфицирования ретровирусом по примеру 1 за исключением того, что в пробирки, содержащие гепарин, собирали пуповинную кровь нормальных, доношенных новорожденных в соответствии с процедурами, утвержденными Институционным советом медицинского колледжа университета шт. Индиана, и использовали ее вместо клеток костного мозга. Анализы инициирующих клеток долгоживущей культуры (стволовые клетки человека) и анализы на метилцеллюлозе выполняли в соответствии с примером 1.
4.2. Эффективность переноса генов в коммитированные клетки-предшественники
Уровень инфицирования на фрагментах фибронектина 30/35 вырос более чем в четыре раза по сравнению с использованием бычьего сывороточного альбумина в трех отдельных экспериментах (таблица 3).
ПРИМЕР 5
Перенос генов в клетки пуповинной крови с использованием вектора PGK-mADA
5.1. Общая процедура
Надосадочную жидкость с вирусом PGK-mADA и химотрипсиновые фрагменты фибронектина (фиг. 1) получали так же, как описывалось в примере 1. Выполняли процедуру инфицирования ретровирусом по примеру 1 за исключением того, что в пробирки, содержащие гепарин, собирали пуповинную кровь нормальных, доношенных новорожденных в соответствии с процедурами, утвержденными Институционным советом медицинского колледжа шт. Индиана. Анализы инициирующих клеток долгоживущей культуры и анализы на метилцеллюлозе выполняли в соответствии с примером 1.
5.2. Эффективность переноса генов в инициирующие клетки долгоживущей культуры
При использовании вектора PGK-mADA с более высоким титром анализ колоний, образованных инициирующими клетками долгоживущей культуры, показал высокую экспрессию введенной кДНК аденозиндезаминазы мышей только на культурах, полученных из пуповинной крови, инфицированной надосадочной жидкостью на фрагментах фибронектина 30/35. Небольшая экспрессия аденозиндезаминазы мышей была обнаружена в колониях, образованных инициирующими клетками долгоживущей культуры, на контрольных чашках с бычьим сывороточным альбумином.
Результаты, приведенные в примерах 4 и 5, показывают, что повышенная эффективность инфицирования при использовании фрагментов фибронектина 30/35 может также быть достигнута при использовании клеток-предшественников пуповинной крови и стволовых клеток.
ПРИМЕР 6
Перенос генов в клетки пуповинной крови с использованием вектора PGK-hADA
Надосадочную жидкость с вирусом PGK-hADA и химотрипсиновые фрагменты фибронектина (фиг. 1) получали так же, как описывалось для вектора TKNEO в примере 1. Выполняли процедуру инфицирования ретровирусом по примеру 1 за исключением того, что в пробирки, содержащие гепарин, собирали пуповинную кровь нормальных, доношенных новорожденных в соответствии с процедурами, утвержденными Институционным советом медицинского колледжа университета шт. Индиана, и использовали ее вместо клеток костного мозга. Анализы инициирующих клеток долгоживущей культуры и анализы на метилцеллюлозе выполняли в соответствии с примером 1.
ПРИМЕРЫ 7-11
Ретровирусные векторы и линии клеток-предшественников для примеров 7-11
Для примеров 7-11 использовали две линии образующих ретровирус упаковочных клеток: экотропную линию клеток GP + E86 (Markowitz D., S. Goff и A. Bank, J. Virol., 62, стр. 1120-1124 (1988) и амфотропную линию клеток GP + env AM12 (Markowitz D., S. Goff и A. Bank, Virology, т. 167, стр. 400-406 (1988)). Использованные в описанных здесь исследованиях ретровирусные векторы и клоны клеток-продуцентов приведены в таблице 4.
Все линии клеток культивировали в модифицированной по способу Дульбекко среде Игла (Gibco, Гранд-Айленд, шт. Нью-Йорк), содержащей 10% околоплодной сыворотки теленка (Hyclone, Логан, шт. Юта), 100 единиц/мл пенициллина и 100 мкг/мл стептомицина (Gibco), за исключением того, что клетки EAL2a выращивали в модифицированной по способу Дульбекко среде Игла F12 (Gibco) с 10% околоплодной сыворотки теленка. Содержащую вирус надосадочную жидкость собирали путем добавления 10 мл альфа-минимальной поддерживающей среды (Gibco) для клеток мышей или среды Дульбекко, модифицированной по способу Исков (Gibco), для клеток человека, каждая из которых содержала 10% околоплодной сыворотки теленка и смесь пенициллина/стрептомицина с целью слияния 10 см планшетов в течение ночи. Собранную среду фильтровали через 0,45-микронные фильтры (Gelman Scienses Энн-Арбор, шт. Мичиган) и хранили до использования при температуре -80oC.
ПРИМЕР 7
Трансдукция первичных кроветворных клеток мышей
7.1. Экспериментальная часть
Для проведения исследований с использованием клеток мышей костный мозг брали из бедренной и большеберцовой костей мышей C3H/HeJ в возрасте 6-8 недель через 2 дня после введения 150 мг/кг 5-фторурацила (SoloPack Laboratories, Фрэнклин-Парк, шт. Иллинойс) (Lim B., J.F. Aperley, S.H. Orhin и D. A. Williams, Proc. Natl. Acad. Sci, США, т. 86, стр. 8892-8896 (1989)). Клетки предварительно стимулировали при концентрации 5 • 105 клеток/мл в среде Дульбекко, модифицированной по способу Исков, которая содержала 20% околоплодной сыворотки теленка и смесь пенициллина/стрептомицина со 100 мг/мл фактора роста рекомбинантных стволовых клеток крысы (Amgen Фаузенд-Оакс, шт. Калифорния) и 100 единицами/мл рекомбинантного интерлейкина 6 человека (Pepro Tech Inc., Рок-Хил, г. Нью-Джерси) в течение 48 часов (Luskey B. D. , M. Rosenblatt, K. Zsebo и D.A. Williams, Blood, т. 80, стр. 396-402 (1992)). Затем сравнивали эффективность переноса генов вектором PGK-hADA, полученным с помощью клеток-продуцентов ЕРНА-5, с использованием трех разных процедур инфицирования:
1) инфицирование надосадочной жидкости;
2) инфицирование надосадочной жидкости на фрагментах фибронектина 30/35;
3) сокультирование на клетках-продуцентах ЕРНА-5. Поэтому 100 мм бактериологические чашки покрывали 2,5 мкг/см2 фрагментов фибронектина 30/35 (эквивалентно 75 ммолям/см2), растворенных в 5 мл забуференного фосфатом физиологического раствора (Gibco), после чего в течение 1 часа выдерживали при комнатной температуре и ультрафиолетовом освещении в чашках с открытой крышкой и еще в течение одного часа - в чашках с закрытой крышкой. После блокировки 2% бычьего сывороточного альбумина (Fraction V; Boehringer Mannheim, Индианаполис, шт. Индиана) в течение 30 минут при комнатной температуре чашки промывали один раз сбалансированным солевым раствором Хенкса, в который добавляли 2,5% (в объемном отношении) 1 М раствора гепеса (Gibco). Для инфицирования надосадочной жидкости чашки покрывали только бычьим сывороточным альбумином. 5 • 106 предварительно стимулированных клеток донора инкубировали вместе с 10 мл содержащей вирус надосадочной жидкости, полученной из клеток ЕРНА-5, в которую добавляли 100 единиц/мл рекомбинантного интерлейкина 6 человека, 100 нг/мл фактора роста рекомбинантных стволовых клеток человека и 7,5 мкг/мл полибрена. Несвязанные клетки собирали и вновь добавляли вместе со свежей надосадочной жидкостью с вирусом. Для сокультивирования клетки ЕРНА-5 в 4 мл среды инкубировали с 10 мкг/мл митомицина в течение 2 часов при температуре 37oC, промывали, обрабатывали трипсином и высевали на 100 мл чашки с культурой ткани при концентрации 3 • 106 клеток в 10 мл альфа-минимальной поддерживающей среды с 20% околоплодной сыворотки теленка и смеси пенициллина/стрептомицина. На следующий день в течение 48 часов добавляли 5 • 106 предварительно стимулированных клеток костного мозга со 100 единицами/мл рекомбинантного интерлейкина 6 человека, 100 нг/мл фактора роста рекомбинантных стволовых клеток крыс и 4 мкг/полибрена. В соответствии с процедурой инфицирования несвязанные клетки сливали, а адгезионные кроветворные клетки выделяли из культур с использованием буфера для диссоциации клеток (без фермента /на основе забуференного фосфатом физиологического раствора, Gibco), следуя инструкциям изготовителя. Связанные клетки добавляли к фракции несвязанных клеток, дважды промывали и суспендировали в 1 мл смеси сбалансированного солевого раствора Хенкса и Хепеса. Все клетки, полученные из 5 • 106 предварительно стимулированных клеток, инъецировали в хвостовые вены трех мышей-реципиентов, все тело которых подвергали облучению смертельной дозой (700 плюс 400 cGy, источник 137CS) (Luskey B. D. , M. Rosenblatt, K. Zsebo и D.A. Williams, Blood, т. 80 стр. 396-402 (1992)). Трансдукцию кроветворных стволовых клеток анализировали путем обследования мышей, прошедших восстановительную терапию, в отношении экспрессии введенной кДНК аденозиндезаминазы человека. Изоферментный анализ аденозиндезаминазы выполняли у трансплантированных мышей путем исследования клеток крови периферической кровеносной системы в присутствии белка аденозиндезаминазы человека с использованием ацетата целлюлозы при выполнении in situ ферментного анализа (Lim B., D.A. Williams и S.H. Orkin, Mol. Cell. Biol. , т. 7, стр. 3459-3465 (1987)). Исследование начинали проводить через 4 месяца после трансплантации и повторяли его ежемесячно.
7.2. Результаты
Восстановление долгоживущих клеток костного мозга у мышей при помощи генетически измененных кроветворных стволовых клеток обычно считается достаточным для определения эффективности трансдукции стволовых клеток через 4 месяца после трансплантации. Проведенный через 7 месяцев изоферментный анализ реципиентов трансдуцированного костного мозга показал, что:
1) экспрессия кДНК аденозиндезаминазы человека имела место при инфицировании сокультуры или надосадочной жидкости на фрагментах фибронектина 30/35 и отсутствовала в группе, трансплантированной после инфицирования надосадочной жидкости без фрагментов фибронектина;
2) уровни экспрессии были сравнимы для групп, инфицированных по методу сокультивирования и при использовании фрагментов фибронектина 30/35. Как показано на фиг. 4, столбцы 2 - 4, у трех мышей, которым был трансплантирован костный мозг, трансдуцированный сокультурой на клетках ЕРНА-5, была легко обнаружена аденозиндезаминаза человека. Аналогичные уровни аденозиндезаминазы человека были обнаружены у трех мышей, которым были трансплантированы кроветворные клетки, трансдуцированные путем инфицирования надосадочной жидкости с фрагментами фибронектина 30/35 (фиг. 4, столбцы 5-7). В отличие от этого аденозиндезаминаза человека не была обнаружена у трех мышей, которым были трансплантированы кроветворные клетки, трансдуцированные путем инфицирования надосадочной жидкости на бычьем сывороточном альбумине (фиг. 4, столбцы 8 - 10). Контрольные результаты для аденозиндезаминазы человека приведены в столбцах 1 и 12, а для аденозиндезаминазы мышей в столбце 11 фиг. 4. Зона, в которой приведены результаты для мышей, столбцы 2-10, свидетельствует о том, что были введены одинаковые количества белка. Эти данные показывают, что трансдукция долгоживущих восстанавливающих кроветворных стволовых клеток путем инфицирования надосадочной жидкости на фрагментах фибронектина 30/35 эквивалентна аналогичному показателю для сокультуры и значительно превосходит инфицирование надосадочной жидкости без фрагментов фибронектина 30/35.
ПРИМЕР 8
Механизм улучшения трансдукции путем связывания ретровирусных векторов с фрагментами фибронектина 30/35
8.1. Экспериментальная часть
Чтобы установить, является ли более высокая трансдукция результатом солокализации вируса и кроветворных клеток, мы анализировали частицы рекомбинантного ретровируса в отношении связывания с фрагментами фибронектина 30/35. Поэтому планшеты, покрытые фрагментами фибронектина 30/35, в течение 30 минут предварительно инкубировали с надосадочной жидкостью, содержащей вирус TKNEO, после чего интенсивно промывали. Вирусный титр надосадочной жидкости определяли с использованием клеток NIH/3T3 в соответствии со стандартными методами (Markowitz D. , S. Goff и A. Bank., J. Virol., 62, стр. 1120-1124 (1988)). Клетки 3T3 помещали при концентрации 1000 клеток/лунку на 6-луночный планшет с культурой ткани и выращивали в течение ночи. В каждую лунку, содержащую 7,5 мк/мл полибрена, последовательно добавляли растворы надосадочной жидкости с вирусом и инкубировали в течение 2,5 часов при температуре 37oC, после чего добавляли 2 мл среды. Через 24 часа эту среду заменяли средой, содержащей G418 (0,75 мг/мл, сухой порошок, Gibco), и инкубировали планшеты в течение 10-12 дней. Резистентные к G418 колонии (G418r) окрашивали через 10 - 12 дней и производили подсчет. Число колоний/лунку, умноженное на величину разбавлений содержащей вирус надосадочной жидкостью, использовали в качестве инфекционных частиц (КОЕ)/мл надосадочной жидкости. Мы оценивали/"титровали" количество ретровирусных частиц, оставшихся на 35 мм планшетах, покрытых фрагментами фибронектина 30/35 или бычьим сывороточным альбумином, после предварительной инкубации с содержащей вирус надосадочной жидкостью и интенсивного промывания путем добавления 1000 клеток NIH/3T3 на 35 мм бактериологическую чашку вместе с полибреном. Через двадцать четыре часа в культуры вводили среду, содержащую 0,75 мг/мл G418 (сухой порошок), после чего клетки продолжали инкубировать в течение 10-12 дней. После инкубации производили количественное определение наличия адгезионного вируса посредством определения числа резистентных к G418 колоний NIH-3T3.
Чтобы установить, зависит ли связывание вируса с фрагментами фибронектина 30/35 от дозы, повторяли вышеуказанные эксперименты с увеличением концентраций фрагментов фибронектина 30/35, наносимых на чашки. Поэтому 35 мм бактериологические чашки покрывали 1, 4, 10 и 20 мкг/см2 фрагментами фибронектина 30/35, как это описывалось выше. Разбавленный в соотношении 1:50 штамм вируса TKNEO, титр которого перед этим составил 1•104 инфекционных частиц/мл, в течение 30 минут инкубировали на планшетах, покрытых фрагментами фибронектина 30/35. После интенсивной промывки в каждую лунку добавляли 2000 клеток NIH-3T3. Отбор производили в соответствии с приведенным выше описанием, и через 10 дней после отбора производили подсчет резистентных к G418 колоний клеток NIH-3T3.
8.2. Результаты
На фиг. 5 приведены результаты одного из трех типичных экспериментов. При использовании содержащей TKNEO надосадочной жидкости титры ретровируса, измеряемые числом колоний G419r в клетках NIH/3T3, сократились более чем на 3 логарифма (с 4 • 103 до 0) на планшетах, покрытых бычьим сывороточным альбумином, в то время как на планшетах, покрытых фрагментами фибронектина 30/35, сокращение титра составило только 1 логарифм. Эти данные показывают, что происходит количественное связывание ретровируса с фрагментами фибронектина 30/35 и не происходит связывания с чашками, покрытыми бычьим сывороточным альбумином (контрольные чашки). На фиг. 6 показано, что большее число резистентных к G418 колоний было обнаружено тогда, когда содержащую вирус надосадочную жидкость инкубировали на планшетах, покрытых фрагментами фибронектина 30/35 с более высокими концентрациями. Поэтому связывание вируса с фрагментами фибронектина 30/35 зависит от дозы.
ПРИМЕР 9
Связывание вируса с рекомбинантными фрагментами фибронектина
9.1. Экспериментальная часть
Кимизука и др. ранее сообщали о наличии экспрессии клонированных последовательностей ДНК фибронектина у E.coli (Kimizuka F., Y. Taguchi, Y. Ohdate, Y. Kawase, T. Shimojo, D. Hashino, I. Kato, K. Sekiguchi и K. Titani, J. Biochem., т. 110, стр. 284-291 (1991)). Клонированные и химерные пептиды содержат одну или несколько важных последовательностей в фибронектине, которые, как известно, участвуют в адгезии клеток (включая RGDS, CS-1 и участок связывания гепарина), см. фиг. 7. Чтобы проанализировать, может ли ретровирус связываться с этими рекомбинантными фрагментами фибронектина, анализы по образованию колоний клеток 3T3 повторяли на планшетах, покрытых рекомбинантными фрагментами C-274, H-271, H-296, CH-271, CH-296 и C-CS1, а также с использованием фрагментов фибронектина 30/35 в качестве положительной контрольной группы при выполнении двух разных разбавлений (1:10 и 1:100) замороженного штамма ретровируса TKNEO с 1•104 инфекционных частиц/мл. Фрагменты фибронектина использовали с концентрацией 120-130 пмолей/см2 (эквивалентно 4 мкг/см2 для C-274, Н-271, Н-296, C-CS1, фрагментов фибронектина 30/35 и 8 мкг/см2 для CH-271 и CH-296). На планшеты наносили покрытие, добавляли вирус, планшеты медленно промывали, на 24 часа вводили клетки NIH/3T3, затем их выращивали в селекционной среде в течение 10 дней, после чего колонии окрашивали и производили подсчет.
9.2. Результаты
На фиг. 8 показано, что число резистентных к G418 колоний (а следовательно и к адгезии вируса) увеличилось для фрагментов H-271, H-296, CH-271 и CH-296. Кроме того, из этой фигуры видно, что число связанных вирусов сравнимо для этих рекомбинантных фрагментов и фрагментов фибронектина 30/35, хотя в этой работе фрагмент Н-271 характеризовался самым высоким уровнем связывания вируса. Общими для всех этих 5 фрагментов фибронектина являются повторы 12-14 типа III, которые включают участок связывания гепарина с высоким сродством (Ruoslahti E., Ann. Rev. Biochem., т.57, стр. 375-413 (1988) и Kimizuka F. , Y. Taguchi, Y. Ohdate, Y. Kawase, T. Shimojo, K. Hashino, I. Kato, K. Sekiguchi и K. Titani, J. Biochem.,
т. 110, стр. 284-291 (1991)), вероятно, расположенный в повторе 13 (Kimisuka, F., Y. Taguchi, Y. Ohdate, Y. Kawase, T. Shimojo, K. Hashino, I. Kato, K. Sekiguchi и K. Titani, J. Biochem., т. 110, стр. 284-291 (1991)). Это позволяет предположить, что связывание вируса происходит на этом известном участке адгезии. Доказательства этого были получены в результате предварительной инкубации чашек, покрытых 4 мкг/см2 CH-271 с увеличивающимися концентрациями (10-1000 мкг/мл) сульфата гепарина, сильно заряженная молекула которого, как известно, ингибирует связывание клеток с участком связывания гепарина. Как видно из фиг. 9, число резистентных к G418 колоний сокращается вслед за предварительной инкубацией CH-271 с сульфатом гепарина в возрастающих концентрациях. Эти данные позволяют предположить, что связывание вируса с фибронектином опосредовано участком связывания гепарина с высоким сродством, расположенным рядом с участком CS-1 в карбоксиконцевых областях фибронектина.
ПРИМЕР 10
Трансдукция кроветворных клеток на рекомбинантных фрагментах фибронектина
10.1. Экспериментальная часть
Чтобы проанализировать, будет ли описанная выше более высокая трансдукция кроветворных клеток на фрагментах фибронектина 30/35 наблюдаться у рекомбинантных фрагментов фибронектина, мы оценивали эффективность трансдукции инфекций надосадочной жидкости in vitro путем анализов колониеобразующих клеток с высоким пролиферативным потенциалом. Используя векторы EAL2a, мы сравнивали разные рекомбинантные фрагменты фибронектина и фрагменты фибронектина 30/35 с инфицированием надосадочной жидкости на бычьем сывороточном альбумине в отношении влияния, оказываемого на трансдукцию кроветворных клеток, при использовании показателей роста резистентных к G418 колоний в качестве индикатора переноса генов. Кроме того, мы сравнивали способность вирусных частиц, уже связанных с фрагментами фибронектина (по сравнению с вирусами, находящимися в надосадочной жидкости), трансдуцировать кроветворные клетки. От 0,5 до 1 • 106 предварительно стимулированных клеток костного мозга инкубировали на 35 мм чашках Петри, покрытых фибронектином, в 1-2 мл содержащей вирус EAL2a надосадочной жидкости с факторами роста и 5 мкг/мл полибрена, как это описывалось выше. Для оценки трансдукции кроветворных клеток вирусом, связанным с фрагментом фибронектина, 35 мм чашки, покрытые фибронектином, инкубировали вместе с содержащей вирус надосадочной жидкостью и трижды промывали 2 мл забуференного фосфатом физиологического раствора. После этого добавляли от 0,5 до 1•106 предварительно стимулированных клеток костного мозга в 2 мл среды, дополненной факторами роста и полибреном. Через 22 часа клетки собирали и анализировали колониеобразующие клетки с высоким пролиферативным потенциалом с 1,5 мг/мл G418 и без него, как описывалось выше (Bradley T.R. и D. Metcalf, Aust. J., Exp. Biol. Med. Sci., т. 44, стр. 287-293 (1966)). Эти культуры инкубировали 14 дней в 7% CO2 при температуре 33oC, после чего высчитывали эффективность переноса генов в виде процентного значения резистентных к G418 колоний.
10.2 Результаты
Трансдукция первичных кроветворных клеток при инфицировании надосадочной жидкости (фиг. 10), была значительно выше, чем при инфицировании на бычьем сывороточном альбумине для всех фрагментов, которые включали участок связывания гепарина и, по крайней мере, еще один более активный участок адгезии клеток (сплошные столбцы). Эффективность трансдукции рекомбинантных фрагментов CH-271, H-296, CH-296 и C-CS1 была аналогична действию всех трех фрагментов фибронектина 30/35, которые включают как участок связывания гепарина, так и CS-1 участок. Во всех других случаях трансдукция сильно снижались. Эти данные показывают, что повышение трансдукции первичных кроветворных клеток, ранее продемонстрированное фрагментами фибронектина 30/35, может быть повторено рекомбинантными фрагментами фибронектина. Это далее свидетельствует о том, что вирус, непосредственно связанный с фибронектином, способен осуществлять генетическую трансдукцию кроветворных клеток. И, наконец, эти данные подтверждают, что наличие как CS-1 участка, так и участка связывания гепарина полезно для усиления трансдукции кроветворных (стволовых) клеток-предшественников.
ПРИМЕР 11
Восстановление долгоживущих клеток костного мозга мышей в результате трансдукции клеток мышей-доноров на рекомбинантных фрагментах фибронектина
11.1 Экспериментальная часть
Мы повторяли представленные выше in vitro исследования (начиная с примера 10) для примитивных кроветворных клеток-предшественников для сравнения инфицирования надосадочной жидкости на бычьем сывороточном альбумине с фрагментами фибронектина 30/35 и рекомбинантных фрагментов фибронектина с сокультурой, производя трансплантацию костного мозга для оценки влияния, оказываемого на восстановление кроветворных стволовых клеток. Мышам, получившим смертельную дозу радиации, инъецировали клетки донора, которые трансдуцировали вектором ЕРНА-5, содержащими кДНК аденозиндезаминазы человека. Через 1 месяц с помощью изоферментных анализов аденозиндезаминазы определяли эффективность переноса генов из крови периферической кровеносной системы.
11.2. Результаты
На фиг. 11 отчетливо видно, что фрагмент фибронектина H-296, содержащий как участок связывания гепарина, так и CS-1 участок, показывает такие же результаты, что и фрагменты фибронектина 30/35 и сокультура. Фрагменты, которые не имеют этих участков, являются менее эффективными в отношении трансдукции трансплантируемых кроветворных клеток. Эти данные показывают, что солокализация примитивных кроветворных столбовых клеток и ретровируса, связанного с рекомбинантными фрагментами фибронектина, включающими как CS-1 участок, так и участок связывания гепарина, эффективно трансдуцируют трансплантируемые кроветворные клетки.
Хотя это изобретение было детально проиллюстрировано и рассмотрено в приводимом выше описании, его следует считать иллюстративными и не ограничивающим объем изобретения. Вполне понятно, что были представлены только предпочтительные варианты осуществления настоящего изобретения и весьма желательно, чтобы патентом были защищены все изменения и модификации, входящие в объем изобретения.
Все приведенные здесь публикации включены в совокупности в качестве противопоставленных материалов, что равнозначно их включению и полному изложению по отдельности.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ, ПОВЫШАЮЩИЕ ЧИСЛО КОПИЙ ВЕКТОРА (ЧКВ), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2744603C2 |
| ПОЛИПЕПТИДЫ, ПРОИСХОДЯЩИЕ ИЗ ТРИПТОФАНИЛ-ТРНК-СИНТЕТАЗЫ, И ИХ ПРИМЕНЕНИЕ ДЛЯ РЕГУЛЯЦИИ РАЗВИТИЯ КРОВЕНОСНЫХ СОСУДОВ | 2002 |
|
RU2297425C2 |
| СПОСОБЫ СТАБИЛЬНОЙ ТРАНСДУКЦИИ КЛЕТОК ВИРУСНЫМИ ВЕКТОРАМИ | 2001 |
|
RU2280074C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ПРИМЕНИМЫЕ ДЛЯ МОДУЛЯЦИИ АНГИОГЕНЕЗА С ИСПОЛЬЗОВАНИЕМ ПРОТЕИНКИНАЗЫ RAF И RAS | 2000 |
|
RU2257911C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2540490C2 |
| СПОСОБЫ И СОСТАВЫ ДЛЯ ПРОИЗВОДСТВА ЛИМФОЦИТОВ И ИХ РЕГУЛИРУЕМОГО УВЕЛИЧЕНИЯ | 2017 |
|
RU2755059C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНЕЙ МОЗГА | 2014 |
|
RU2664471C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2478400C2 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ, ФИБРОБЛАСТ, СПОСОБ ОТДЕЛЕНИЯ КЛЕТОК | 1991 |
|
RU2164240C2 |
| ТРАНСДУЦИРУЮЩИЕ ЦИТОХРОМ Р450 РЕТРОВИРУСНЫЕ ВЕКТОРЫ | 1997 |
|
RU2185821C2 |














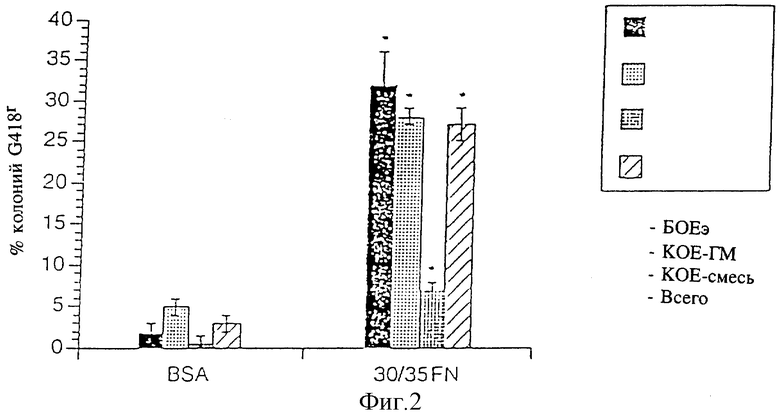
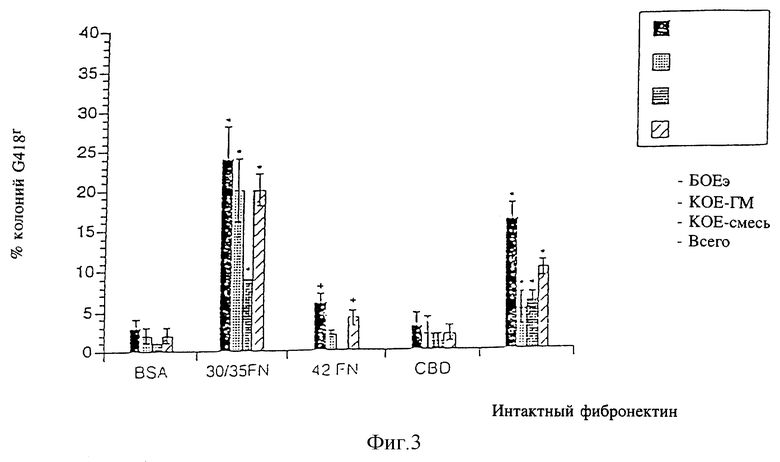

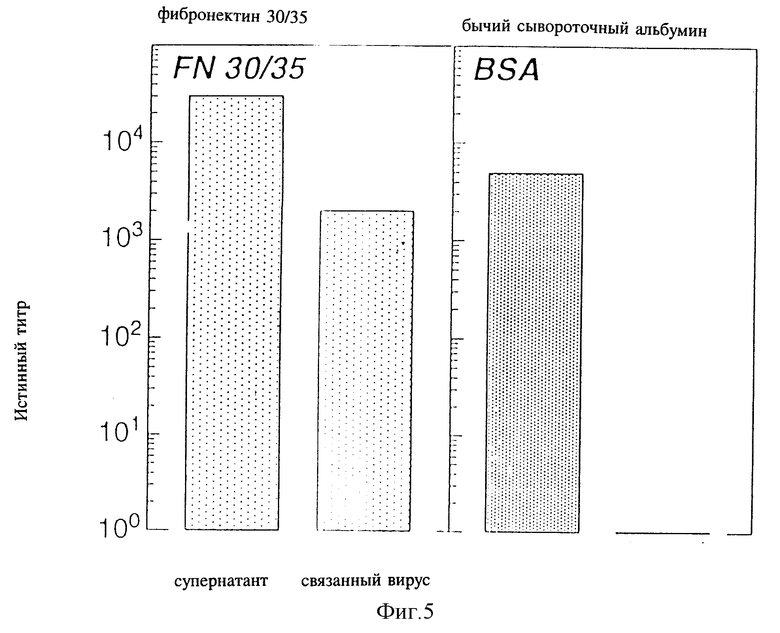
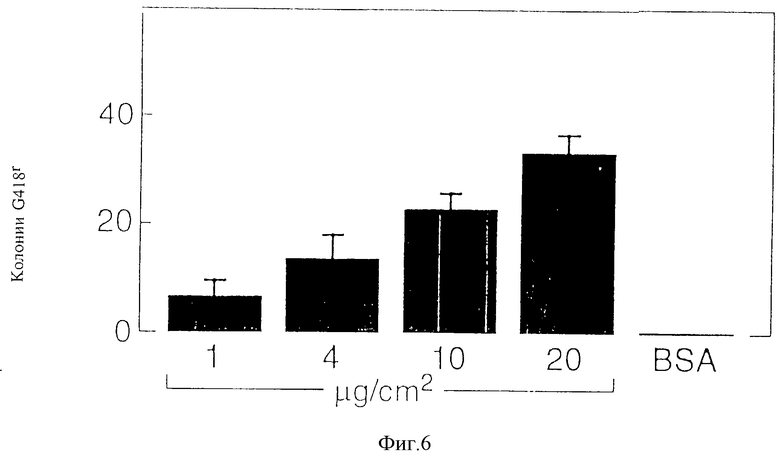
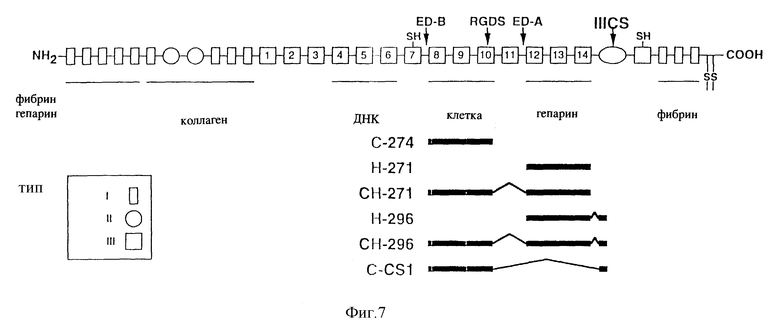
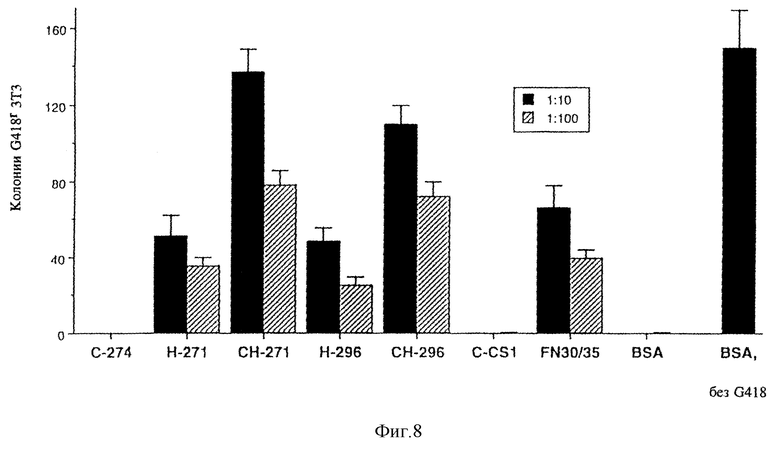


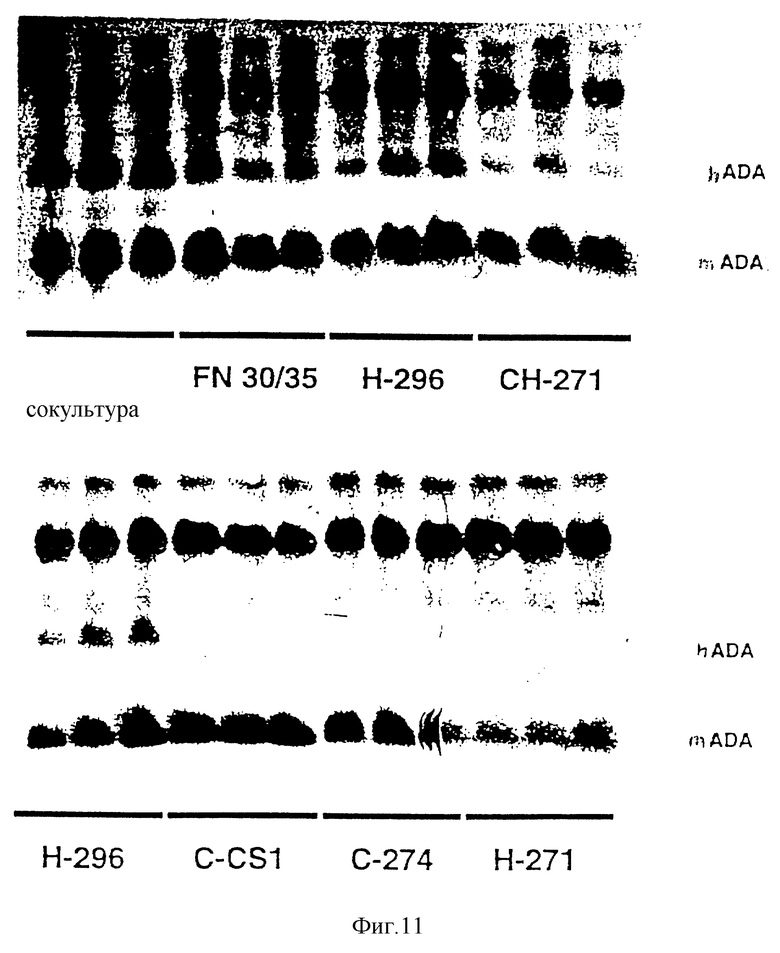
Изобретение относится к медицине и касается опосредованного вирусом усиленного переноса ДНК и способа повышения эффективности трансдукции кроветворных и других клеток под действием ретровируса. Способ включает инфицирование клеток в присутствии фибронектина или его фрагментов. Фибронектин и его фрагменты значительно усиливают опосредованный ретровирусом перенос генов в клетки, в частности кроветворные клетки, включая коммитированные клетки-предшественники и первичные кроветворные стволовые клетки. Настоящим изобретением предусматриваются также усовершенствованные способы соматической генотерапии на основе усиленного переноса генов, популяции кроветворных клеток и новые конструкции, предназначенные для усиления опосредованного ретровирусом переноса ДНК в клетки, и их использование. Преимущество изобретения заключается в разработке эффективного переноса генетического материала в клетки млекопитающих без опасных последствий и ограничений. 18 с. и 70 з.п. ф-лы, 4 табл., 11 ил.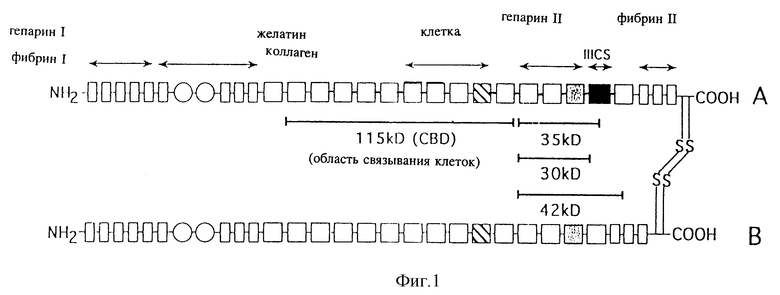
или по существу подобную аминокислотную последовательность, обладающую способностью связывать ретровирусы; так и (II) вторую аминокислотную последовательность, представленную формулой
Asp Gly Leu Pro Gln Leu Val Thr Leu Pro His Pro Asn Leu His Gly Pro Glu Iie Leu Asp Val Pro Ser Thr
или по существу подобную аминокислотную последовательность, обладающую способностью связывать примитивные кроветворные клетки.
или по существу подобную аминокислотную последовательность, обладающую способностью связывать ретровирусы;
и вторую аминокислотную последовательность, представленную формулой
Asp Gly Leu Pro Gln Leu Val Thr Leu Pro His Pro Asn Leu His Gly Pro Glu Ile Leu Asp Val Pro Ser Thr
или по существу подобную аминокислотную последовательность, обладающую способностью связывать примитивные кроветворные клетки.
или по существу подобную аминокислотную последовательность, обладающую способностью связывать ретровирусы; и
вторую аминокислотную последовательность, представленную формулой
Asp Gly Leu Pro Gln Leu Val Thr Leu Pro His Pro Asn Leu His Gly Pro Glu Ile Leu Asp Val Pro Ser Thr
или по существу подобную аминокислотную последовательность, обладающую способностью связывать примитивные кроветворные клетки.
или по существу подобную аминокислотную последовательность, обладающую способностью связывать ретровирусы; и вторую аминокислотную последовательность, представленную формулой Asp Gly Leu Pro Gln Leu Val Thr Leu Pro His Pro Asn Leu His Gly Pro Glu Ile Leu Asp Val Pro Ser Thr
или по существу подобную аминокислотную последовательность, обладающую способностью связывать примитивные кроветворные клетки.
или по существу подобную аминокислотную последовательность, обеспечивающую ретровирус-связываюшую активность домена гепарина-II фибронектина.
или по существу подобную аминокислотную последовательность, обеспечивающую ретровирус-связывающую активность.
или по существу подобную аминокислотную последовательность, обеспечивающую ретровирус-связывающую активность.
(а) по существу чистый рекомбинантный полипептид, содержащий (I) первую аминокислотную последовательность домена гепарина-II фибронектина человека, которая обеспечивает ретровирус-связывающую активность, и (II) вторую аминокислотную последовательность, которая обеспечивает связывающую клетки активность домена СS-1 фибронектина человека;
(b) искусственный субстрат, предназначенный для инкубации ретровирусного вектора в контакте с кроветворными клетками человека; и (с) факторы роста кроветворных клеток, предназначенные для предварительного стимулирования кроветворных клеток.
(d) рекомбинантный ретровирусный вектор, предназначенный для трансдукции кроветворных клеток человека.
или по существу подобную аминокислотную последовательность, обладающую ретровирус-связывающей активностью.
Asp Gly Leu Pro Gln Leu Val Thr Leu Pro His Pro Asn Leu His Gly Pro Glu Ile Leu Asp Val Pro Ser Thr
или по существу подобную аминокислотную последовательность, обеспечивающую способность связывания примитивных кроветворных клеток.
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| ПАВЛОВ А.Д., МОРЩАКОВА Е.Ф | |||
| Регуляция эритропоэза | |||
| -М.: Медицина, 1987, с | |||
| Спускная труба при плотине | 0 |
|
SU77A1 |
| PATEL et al | |||
| The fibronectin receptor of mamalian erytroid precursor cells, characterization ahd developmental regulation, J | |||
| of Cell | |||
| Biol | |||
| Пневматический водоподъемный аппарат-двигатель | 1917 |
|
SU1986A1 |
| Транспортер для перевозки товарных вагонов по трамвайным путям | 1919 |
|
SU102A1 |

