Настоящее изобретение относится к репликативно-дефектным ретровирусным векторам, кодирующим ген цитохрома Р450, которые способны конвертировать безвредные противораковые пролекарства в цитотоксические метаболиты. Кроме того, настоящее изобретение относится к применению таких ретровирусных векторов для получения лекарственных средств для лечения различных раковых заболеваний или удаления опухолей.
Уровень техники
Противораковые лекарственные средства, применяемые для лечения опухолей, в большинстве случаев применяются системно и распространяются по всему организму пациента. Высокие системные дозы таких лекарственных средств, необходимые для лечения рака, сочетаются с неприятными для пациента побочными действиями.
При попытке обойти эту проблему применяют пролекарства противораковых средств, которые должны претерпевать метаболизм или активироваться в организме прежде, чем становятся цитотоксичными. К несчастью, опухоли человека, содержащие соответствующие высокие уровни активирующих ферментов, являются редкими. Основной областью активации пролекарств является печень, и чтобы гарантировать, что опухоль в отдаленной области получает достаточную дозу активированного лекарственного средства, количество активированного пролекарства, принесенного в печень, должно быть достаточно высоким, и это обстоятельство снова приводит к токсическим побочным действиям на пациента.
Один из подходов, с помощью которых эти проблемы высокой общей концентрации активированных лекарственных средств можно обойти, может состоять в обеспечении средств активации пролекарства непосредственно или вблизи области опухоли. При таком подходе потребуется, чтобы опухолевые клетки или клетки в области опухоли, генетически трансформировались для продуцирования большого количества ферментов, необходимых для метаболизма противораковых пролекарств. Ретровирусные векторы идеально подходят для устойчивой доставки генов к клеткам, так как ретровирус способен интегрировать форму ДНК своего генома в геном клетки-хозяина, и, таким образом, все дочерние клетки инфицированной клетки будут нести ретровтирусный вектор, несущий терапевтический ген. Другое преимущество состоит в том, что ретровирусы инфицируют только делящиеся клетки, и следовательно, они являются идеальными носителями, доставляющими гены для опухолевых клеток.
Уже испытано много цитотоксических генов, переносимых ретровирусными векторами. Эти гены кодируют ферменты, конвертирующие вещества, являющиеся фармакодинамически и токсикологически инертными даже при высоких уровнях доз, но которые in vivo могут быть превращены в высокоактивные метаболиты (Connors, Т.А. (1995), Gene Therapy, 2:702-709).
Обнаружено, что соответствующим образом созданные для противораковой химиотерапии пролекарства являются эффективными при лечении у животных опухолей с высоким содержанием активирующего фермента (Connors, Т. and Whisson, М. (1966), Nature, 210: 866-867, and Cobb, L. et al. (1969), Biochemical Pharmacology, 18: 1519-1527). Однако клинические результаты являются неутешительными, так как обнаружено, что злокачественные опухоли человека, содержащие соответственно высокие уровни активирующих ферментов, являются редкими (Connors, Т. (1986), Xenobiotica, 16:975-988). Лечение пролекарствами, активируемыми вирусноуправляемыми ферментами (VDEPT), и более общее лечение пролекарствами, активируемыми генуправляемыми ферментами (GDEPT), родственны в том, что такое лечение также способствует разрушению опухолевых клеток посредством опухолеспецифической активации пролекарства. Однако в этом случае ген, кодирующий фермент, или специфически направляется на злокачественные клетки, или находится под контролем специфического промотора.
До настоящего времени больше всего усилий в области лечения пролекарствами концентрируется на применении в качестве гена-самоубийцы тимидинкиназы вируса человеческого простого герпеса (HSV-tk). Хотя фермент HSV-tk в сочетании с пролекарством ганцикловиром (GCV) рекомендован в качестве хорошей системы для GDEPT (Culver, К. et al., (1992), Science, 256: 1550-1552, Ram, Z. et al., (1993), Cancer Research, 53:83-88, и Chen, S., Shine, H. et al., (1994), Proc. Natl. Acad. Sci., 91:3054-3057), существует ряд теоретических соображений, приводящих к мысли, что это не самое лучшее сочетание. Во-первых, она является S-фазовоспецифическим средством; не действующим на клетки в состоянии покоя. Это происходит потому, что монофосфат GCV является неустойчивым и для получения токсического действия должен присутствовать, когда клетки входят в S-фазу. HSV-tk фосфорилирует GCV до формы монофосфата (реакция, которая не может быть осуществлена ферментами млекопитающих), который затем фосфорилируется клеточными ферментами до формы трифостата и включается в ДНК. Во-вторых, активное лекарственное средство является трифосфатом, и нельзя ожидать, что оно будет свободно диффундировать и вызывать эффект свидетеля (bystander effect). Однако свидетельствующий эффект наблюдается как in vitro, так и in vivo, хотя оказывается, что включается метаболическая кооперация, и в последнем случае некоторая доля действия может быть косвенным действием с участием иммунного компонента (Bi, W. , Parysek, L. et al., (1993), Human Gene Therapy, 4:725-731, Vile, R. and Hart, I. (1993), Cancer Research, 53: 3860-3864, и Freeman, S., Abboud, С. et al., (1993), Cancer Research, 53:5274-5283). Один из недостатков состоит в том, что эффект свидетеля зависит от контакта клетка-клетка. Это может происходить из-за наличия щелевых контактов, образованных за счет тесного контакта между трансдуцированными и окружающими клетками, способными переносить фосфорилированный ганцикловир.
Недавно сделано сообщение об интересных результатах, полученных с клетками, трансфектированными геном, кодирующим форму 2В1 крысиного цитохрома Р450, и обработанными затем циклофосфамидом (Chen, S., Shine, Н. et al., (1994), Proc. Natl. Acad. Sci., 91:3054-3057).
Цели изобретения
Целью настоящего изобретения являются новые репликативно-дефектные ретровирусные векторы, несущие ген, кодирующий цитохром Р450 под транскрипционным контролем специфического для клетки-мишени регуляторного элемента или промотора, или индуцируемого рентгеновским излучением промотора, которые можно трансфектировать в линии упаковывающих клеток для продуцирования рекомбинантных ретровирусных частиц, полезных для генной терапии различных злокачественных опухолей.
Сущность изобретения
Настоящее изобретение также включает, среди прочего, следующие аспекты, по отдельности или в сочетании:
репликативно-дефектный ретровирусный вектор, несущий ген цитохрома Р450 под транскрипционным контролем специфического для клетки-мишени регуляторного элемента или промотора, или промотора, индуцируемого рентгеновским излучением;
описанный выше репликативно-дефектный ретровирусный вектор, содержащий область 5'-LTR структуры U3-R-U5; одну или несколько последовательностей, выбранных среди кодирующих и некодирующих последовательностей; и область 3'-LTR, содержащую полностью или частично делегированную область U3, при этом указанная делегированная область U3 заменяется полилинкерной последовательностью, содержащей специфический для клетки-мишени регуляторный элемент или промотор или индуцируемый рентгеновским излучением промотор, с последующей областью R и U5, отличающийся тем, что по меньшей мере одна из кодирующих последовательностей кодирует цитохром Р450;
описанный выше репликативно-дефектный ретровирусный вектор, при этом специфический для клетки-мишени регуляторный элемент или промотор выбирают среди одного или нескольких элементов группы, состоящей из специфических регуляторных элементов и промоторов WAP, MMTV, b-лактоглобулина и казеина, специфических регуляторных элементов и промоторов поджелудочной железы, включая карбоангидразу II и b-глюкокиназные регуляторные элементы и промоторы, лимфоцитных специфических регуляторных элементов и промоторов, включая иммуноглобулин и лимфоцитные специфические регуляторные элементы MMTV, и специфических регуляторных элементов и промоторов MMTV, обеспечивающих чувствительность к глюкокортикоидным гормонам или направляющих экспрессию в молочной железе;
описанный выше репликативно-дефектный ретровирусный вектор, при этом указанные области LTR выбирают среди по меньшей мере одного элемента из группы, состоящей из LTR, MLV, MMTV, MSV, SIV, HIV, HTLV, FIV, FeLV, BLV и MPMV;
описанный выше репликативно-дефектный ретровирусный вектор, при этом указанный ретровирусный вектор имеет основой BAG-вектор или вектор pLXSN;
описанный выше репликативно-дефектный ретровирусный вектор, при этом указанный ретровирусный вектор представляет pLX2Bl, полученный так, как описано в примере 1;
описанный выше репликативно-дефектный ретровирусный вектор, при этом указанный ретровирусный вектор представляет pc3/2Bl, полученный так, как описано в примере 2;
описанный выше репликативно-дефектный ретровирусный вектор, при этом ретровирусные последовательности, участвующие в интеграции ретровируса, изменены или по меньшей мере частично делегированы;
описанный выше репликативно-дефектный ретровирусный вектор, при этом указанные регуляторные элементы или промоторы контролируются трансдействующими факторами;
линию упаковывающих клеток, трансфектированную описанным выше репликативно-дефектным ретровирусным вектором; причем указанная линия упаковывающих клеток дает убежище по меньшей мере одной ретровирусной или рекомбинантной ретровирусной конструкции, кодирующей белки, необходимые для упаковки указанного ретровирусного вектора;
описанную выше линию упаковывающих клеток, при этом линия упаковывающих клеток происходит от грызуна, собаки, кошки или человека и гистологически совместима с человеческой тканью;
описанную выше линию упаковывающих клеток, при этом линию упаковывающих клеток выбирают из группы, состоящей из psi-2, psi-Crypt, psi-AM, GP+E-86, PA317 и GP+envAM-12;
рекомбинантную ретровирусную частицу, полученную путем культивирования при подходящих условиях описанной выше линии упаковывающих клеток, необязательно, с последующим выделением полученной рекомбинантной ретровирусной частицы;
фармацевтическую композицию, содержащую описанную выше рекомбинантную ретровирусную частицу или описанную выше линию упаковывающих клеток;
описанную выше линию упаковывающих клеток, инкапсулированную в пористую оболочку, проницаемую для рекомбинантных ретровирусных частиц, продуцированных указанной выше линией упаковывающих клеток;
описанную выше линию упаковывающих клеток, инкапсулированную в комплексный материал, образованный сульфатом целлюлозы и полиметилдиаллиламмонием;
способ удаления опухолевых клеток, включающий введение нуждающемуся в этом субъекту терапевтически эффективного количества описанной выше рекомбинантной вирусной частицы, описанной выше линии упаковывающих клеток или описанной выше инкапсулированной линии упаковывающих клеток либо одновременно, либо с временным интервалом с противораковым пролекарством, которое может быть активировано цитохромом Р450;
описанный выше способ в случае, когда опухолевые клетки являются клетками опухоли молочной железы или опухоли поджелудочной железы;
описанный выше способ, отличающийся тем, что рекомбинантную ретровирусную частицу, линию упаковывающих клеток или описанную выше инкапсулированную линию упаковывающих клеток вводят посредством инъекции или имплантации в опухоль или в область опухоли;
применение описанной выше рекомбинантной ретровирусной частицы, описанной выше линии упаковывающих клеток или описанной выше инкапсулированной линии упаковывающих клеток для получения фармацевтической композиции, полезной для удаления опухолевых клеток;
ретровирусный провирус, интегрированный в геном человека, несущий ген цитохрома Р450 под транскрипционным контролем специфического для клетки-мишени регуляторного элемента или промотора или индуцируемого рентгеновским излучением промотора; и
человеческую клетку, содержащую ген цитохрома Р450 под транскрипционным контролем специфического для клетки-мишени регуляторного элемента или промотора или индуцируемого рентгеновским излучением промотора.
Подробное описание изобретения
Цитохромы Р450 образуют обширную группу монооксигеназ, катализирующих окисление широкого ряда субстратов. Они продуцируются некоторыми бактериями, дрожжами и высшими организмами, в которых играют некую роль при детоксификации ксенобиотиков, в реакциях биоактивирования и при метаболизме различных эндогенных соединений.
Цитохром Р450 катализирует гидроксилирование традиционно используемых противораковых пролекарств циклофосфамида (СРА) и ифосфамида до их активных токсических форм. Обычно экспрессия гена эндогенного цитохрома Р450 у пациента ограничивается в печени, и противоопухолевое действие системно применяемых СРА зависит от последующего системного распространения из печени токсических лекарственных метаболитов. Это приводит к проблемам токсичности, так как активированное лекарственное средство поражает не только опухоль, но поражает также другие здоровые ткани пациента, такие как костный мозг и почки.
Терапевтический подход, при котором ген цитохрома Р450 вводится непосредственно в опухолевые клетки и сверхэкспрессируется в этих клетках, является обходным решением этой проблемы. Токсические метаболиты, продуцированные в трансдуцированных опухолевых клетках, поражают окружающие нетрансдуцированные опухолевые клетки в зависимости от концентрационного градиента. Дополнительным преимуществом системы цитохром Р-450/СРА является отсутствие зависимости цитотоксического действия на окружающие клетки от репликации клеток. Это происходит потому, что один из активных генерированных метаболитов служит причиной межуровневых связей вне зависимости от фазы клеточного цикла. Позднее, в процессе синтеза ДНК, эти межуровневые связи приводят к гибели клеток.
Ретровирусные векторы являются обычно наиболее используемыми носителями для переноса генов для клинических испытаний для получения надежных результатов. Большинство таких испытаний, однако, проводятся ex vivo, когда клетки пациента выделяют, модифицируют в культуре, а затем снова вводят пациенту.
В случае лечения злокачественных опухолей обычно возможно выделить клетки пациента (или опухолевые клетки, или здоровые клетки), инфицировать их in vitro рекомбинантной ретровирусной частицей, несущей ген, кодирующий цитохром Р450, а затем вернуть их пациенту в место вблизи опухоли. Такой подход, однако, исключительно трудоемок, поскольку клетки каждого пациента должны быть выделены, культивированы, трансдуцированы генной конструкцией и успешно возвращены без заражения случайными веществами. Стоимость и время, предполагаемые при таком подходе, ограничивают его практическую пригодность.
С другой стороны, можно представить себе аллогенный подход, когда один тип клеток инфицируется рекомбинантной ретровирусной частицей, несущей ген, кодирующий Р450, а затем используется для лечения многих разных пациентов. Такой подход гораздо проще осуществить при предположении, что можно преодолеть проблемы иммунного отторжения. Однако большинство опухолей на подходят для генной терапии ex vivo.
В идеале ген, кодирующий цитохром Р450, должен быть введен в опухолевые клетки или в клетки вблизи опухоли in vivo.
Доставка генов in vivo ставит множество новых проблем. Прежде всего и что важнее всего, следует обратиться к вопросам безопасности. Основное беспокойство в случае возможной генной терапии in vivo как с точки зрения безопасности, так и с практической точки зрения, является нацеливание экспрессии. Ясно, что терапевтические гены, переносимые векторами, не должны экспрессироваться во всех тканях без разбора, а только в требуемой клетке-мишени. Это особенно важно, когда переносимые гены являются таким пролекарством, активирующим гены, созданным для удаления специфических опухолевых клеток. Удаление других клеток, не являющихся мишенями, очевидно, будет весьма нежелательным.
По существу, случайная интеграция провирусной формы ретровирусного генома в геном инфицированной клетки является серьезной этической проблемой, поскольку такая случайная интеграция может привести к активации протоонкогенов и, таким образом, привести к развитию новой раковой опухоли. Большая часть исследователей обычно соглашается, что вероятность репликации дефектных ретровирусов, таких как все ретровирусы, используемые сегодня, при интеграции в или вблизи клеточного гена, участвующего в управлении клеточной пролиферацией, исчезающе мала. Однако обычно также предполагается, что бурная экспансия популяции репликативнокомпетентных ретровирусов из единичного события заражения будет в итоге обеспечивать достаточное число событий интеграции, чтобы придать такой фенотипической интеграции весьма реальную вероятность.
Системы с ретровирусными векторами оптимизируют для минимизации таких изменений присутствующего репликативнокомпетентного вируса. Однако документальные данные ясно подтверждают, что рекомбинационные события между компонентами ретровирусной векторной системы могут привести к генерации потенциально патогенного репликативнокомпетентного вируса, и для минимизации этого риска рекомбинации созданы многие поколения векторных систем (Salmons, В. and Gunzburg, W.H. (1993), Human Gene Therapy, 4(2):129-41).
Ретровирусные векторные системы состоят из двух компонентов.
1) Сам ретровирусный вектор является модифицированным ретровирусом (векторной плазмидой), в котором гены, кодирующие вирусные белки, заменены терапевтическими генами. Так как замена генов, кодирующих вирусные белки, эффективно ослабляет вирус, его необходимо "спасти" с помощью второго компонента в системе, который предоставляет недостающие вирусные белки для модифицированного ретровируса.
2) Вторым компонентом является клеточная линия, продуцирующая большое количество вирусных белков, однако не обладающая способностью продуцировать репликативнокомпетентный вирус. Такая клеточная линия известна как линия упаковывающих клеток и состоит из линии клеток, трансфектированных одной или несколькими плазмидами, несущими гены, дающие возможность для упаковки модифицированного ретровирусного вектора.
Чтобы генерировать рекомбинантную ретровирусную частицу, ретровирусный вектор трансфектируют в линию упаковывающих клеток. При этих условиях модифицированный ретровирусный геном, включающий вставленный терапевтический ген, транскрибируется из ретровирусного вектора и упаковывается в модифицированные ретровирусные частицы. Эти рекомбинантные ретровирусные частицы затем используют для инфицирования опухолевых клеток, во время которого векторный геном и любой цитотоксический ген интегрируются в ДНК клетки-мишени. Клетка, инфицированная такой рекомбинантной вирусной частицей, не может продуцировать новый векторный вирус, так как в таких клетках отсутствуют вирусные белки, но ДНК вектора, несущего терапевтический ген, интегрирована в клеточную ДНК и теперь может экспрессироваться в инфицированной клетке.
Ранее описан ряд ретровирусных векторных систем, которые должны давать возможность нацеленной доставки переносимых цитотоксических генов (Salmons, В. and Gunzburg, W.H. (1993), Human Gene Therapy, 4(2):129-41). Большинство таких подходов включает или ограничение события инфицирования предварительно определенными типами клеток, или использование гетерологичных промоторов для направления экспрессии связанных гетерологичных терапевтических генов в определенные типы опухолевых клеток. Используются гетерологичные промоторы, которые должны "загонять" экспрессию связанных генов только в тип клеток, в которых этот промотор обычно активен и/или дополнительно поддается контролю. Такие промоторы в сочетании с терапевтически геном предварительно вставляют в тело ретровирусных векторов вместо генов gag, pol или env.
Ретровирусные длинные концевые повторы (LTR), фланкирующие эти гены, несут ретровирусный промотор, который вообще неспецифичен в том смысле, что он может "загонять" экспрессию во многие различные типы клеток (Majors, J. (1990), в "Retroviruses - Strategies of replication (Swanstrom, R. and Vogt, P. K. , Eds.): Springer-Verlag, Berlin: 49-52). Сообщается о промоторной интерференции между промотором LTR и гетерологичными внутренними промоторами, такими как тканеспецифические промоторы, описанные выше. Кроме того, известно, что ретровирусные LTR укрывают сильные энхансеры, которые могут либо независимо, либо вместе с ретровирусным промотором влиять на экспрессию клеточных генов вблизи места интеграции ретровируса. Известно, что этот механизм вносит вклад в онкогенность животных (van Lohuizen, M. and Berns, A. (1990), Biochem. Biophys. Acta, 1032:213-235). Эти два наблюдения поддержали разработку самоинактивирующихся векторов (SIN), в которых ретровирусные промоторы функционально инактивируются в клетке-мишени (WO 94/29437). К другим модификациям этих векторов относятся инсерция кассет промоторных генов в области LTR для создания двухкопийных векторов (WO 89/11539). Однако в обоих этих векторах гетерологичные промоторы, вставленные или в тело вектора, или в область LTR, непосредственно связываются с терапевтическим геном.
Ранее описанный упомянутый выше вектор SIN, несущий делегированный 3'LTR (WO 94/29437), используется в дополнение к гетерологичному промотору, такому как промотор цитомегаловируса (CMV), вместо ретровирусного промотора 5'LTR (5'LTR без U3) для ограничения экспрессии векторной конструкции в линии упаковывающих клеток. В 3'LTR (WO 94/29437) также включен сигнал гетерологичного полиаденилирования.
Целью настоящего изобретения является конструкция безопасного ретровирусного вектора, которая укрывает ген цитохрома Р450 как терапевтическую основу. Этот новый вектор несет гетерологичный определяющий, индуцибельный или тканеспецифический промотор и/или регуляторные элементы в 3'LTR, который после инфекции становится дуплицированным и транслоцированным к 5'LTR в клетке-мишени. Таким образом, в инфицированной клетке введенный промотор регулирует экспрессию гена цитохрома Р450, который вставлен в тело вектора. Этот вектор не претерпевает самоинактивации, но вместо этого обменивает промотор, давая начало вектору с названием РrоСоn для векторов промоторной конверсии. Основы и преимущества системы РrоСоn подробнее описываются в WO 96/07748.
Так как промоторная конверсия не приводит к самоинактивации, ретровирусный вектор в клетке-мишени будет транскрипционно активным, кроме того, LTR будут содержать в значительной степени гетерологические промотер/энхансерные последовательности клетки-мишени. Это будет снижать вероятность того, что интегрированный вектор в клетке-мишени будет подвергаться той же инактивации в течение длительного времени, какая описана для традиционных векторов (Xu, L., Yee, J.K. et al., (1989), Virology, 171:331-341), а также будет снижать возможность рекомбинации с эндогенными ретровирусными последовательностями для генерации потенциально патогенного репликативнокомпетентного вируса, увеличивая безопасность системы.
Согласно изобретению 5'LTR ретровирусной векторной конструкции не модифицируется, и экспрессия вирусного вектора ограничивается в упаковывающих клетках нормальным ретровирусным промотором U3. Имеется возможность для обычного ретровирусного полиаденелирования, и в 3'LTR не включаются никакие сигналы гетерологичного полиаденелирования. Это важно для разработки способов генной терапии in vivo, так как нормальная физиологическая регуляция вируса через нормальный вирусный промотор и, вероятно, включающая также обычный вирусный контроль полиаденелирования, будет преобладать длительное время in vivo, в то время как упаковывающие клетки продуцируют рекомбинантный вирус.
Для достижения вышеуказанной и других целей изобретение предоставляет ретровирусный вектор, претерпевающий промоторную конверсию содержащей 5'LTR области структуры U3-R-U5; одну или несколько кодирующих последовательностей, выбранных из группы генов, известных как гены цитохрома Р450; и 3'LTR область, содержащую полностью или частично делегированную область U3, при этом указанная делегированная область U3 заменяется гегерологичным промотором, за которым следует область R и U5.
Указанный промотор может быть или конститутивным как промежуточный ранний промотор/энхансер цитомегаловируса (CMV), или стимулируемым, например, глюкокортикоидными гормонами (например, промотор MMTV), или специфическим для клетки-мишени.
Регуляторные элементы, специфические для клетки-мишени, и промоторы выбирают среди одного или нескольких элементов любого гена, но в данном варианте воплощения изобретения они могут быть из числа промоторов, включая карбоангидразу II и регуляторные элементы и промоторы β-глюкокиназы, лимфоцитных специфических регуляторных элементов и промоторов, включая карбоангидразу II и регуляторные элементы и промоторы β-глюкокиназы, лимфоцитных специфических регуляторных элементов кислых белков молочной сыворотки (WAP), вируса опухоли молочной железы мыши (MMTV), β-лактоглобулиновых и казеиновых специфических регуляторных элементов и промоторов, специфических регуляторных элементов и промоторов поджелудочной железы, включая иммуноглобулиновые и MMTV лимфоцитные специфические регуляторные элементы и промоторы и MMTV специфические регуляторные элементы и промоторы, придающие чувствительность к глюкокортикоидным гормонам или направляющие экспрессию в молочную железу. К другим промоторам относятся, например, промоторы CD4, CD34 и IL2. Указанные регуляторные элементы и промоторы регулируют предпочтительно экспрессию указанного ретровирусного вектора.
Оказывается, что область промотора WAP, требуемая для опосредования специфичности молочной железы, представляет рестрикционный фрагмент XhoI/Xbal в 320 п.о. (от -413 до -93) (Kolb, A.F., Günzburg, W.H., Albang, R., Brem, G., Erfle, V. , and Salmons, B. (1995), Biochem. Biophys. Res. Commun., 217, 1045-1052). Кроме того, некоторые эксперименты показывают, что фрагмент промотора MMTV PstI в 0,6 т.п.о. (Salmons, В., Corner, В., Calberg Baca, C.M., and Ponta, H. (1985), Virology, 144:101-114) может играть некую роль в регуляции специфичности экспрессии в молочной железе, демонстрируемой MMTV (Kolb, A. F., Günzburg, W.H., Albang, R., Brem, G., Erfle, V., and Salmons, B. (1995), Biochem. Biophys. Res. Commun., 217, 1045-1052).
Области LTR выбирают среди, по меньшей мере, одного элемента из группы, состоящей из LTR вируса мышиного лейкоза (MLV), вируса опухоли молочной железы мыши (MMTV), вируса саркомы мышей (MSV), вируса обезьяньего иммунодефицита (SIV), вируса иммунодефицита человека (ВИЧ), вируса человеческого Т-клеточного лейкоза (HLTV), вируса иммунодефицита кошек (FIV), вируса лейкоза кошек (FELV), бычьего вируса лейкоза (BLV) и обезьяньего вируса Мейсона-Пфайзера (MPMV).
Ретровирусный вектор имеет своей основой предпочтительно или вектор LXSN (Miller, A.D., and Rosman, G.J. (1989), Biotechniques, 7:980-990), или вектор pBAG (Price, J., Turner, D. et al., (1987), Proc. Natl. Acad. Sci. USA, 84:156-160), или гибрид этих обоих векторов.
Кодирующая последовательность терапевтического гена может представлять ген любого цитохрома Р450, но наиболее предпочтительной является форма 2В1 крысиного цитохрома Р450, по определению Fuji-Kuriyama, Y., Mizukami, Y. et al., (1982), Proc. Natl. Acad. Sci. USA, 79:2793-2797.
В другом варианте воплощения изобретения дается система с ретровирусным вектором, содержащая описанный выше ретровирусный вектор в качестве первого компонента и линию упаковывающих клеток, укрывающую по меньшей мере одну ретровирусную или рекомбинантную ретровирусную конструкцию, кодирующую белки, требуемые для указанного ретровирусного вектора, который упаковывается.
Линию упаковывающих клеток выбирают предпочтительно среди элементов группы, состоящей из ψ-2, ψ-Crypt, ψ-AM, GP+E-86, PA317 и GP+envAM-12, или любую из линий, супертрансфектированных рекомбинантными конструкциями, создающих возможность для экспрессии поверхностных белков от других содержащих оболочки вирусов.
Изобретение включает также мРНК, происходящую от ретровирусного вектора по изобретению.
В линии упаковывающих клеток экспрессия ретровирусного вектора регулируется обычным неселективным ретровирусным промотором, который содержится в области U3. Однако как только вектор входит в клетку-мишень, происходит промоторная конверсия и ген Р450 экспрессируется из тканеспецифического или индуцибельного выбранного промотора, вставленного в вектор РrоСоn. В действительности не только можно включать в систему любой тканеспецифический промотор, обеспечивающий селективное нацеливание широкого ряда различных клеточных типов, но, кроме того, после события конверсии структура и свойства ретровирусного вектора более не походят на структуру и свойства вируса. Это, конечно, имеет исключительно важное значение с точки зрения безопасности.
Эти векторные системы будут применяться для получения рекомбинантного вируса, который можно использовать для заражения опухолевых или здоровых клеток или in vitro, или in vivo.
Рекомбинантные вирусы, очищенные или сконцентрированные, можно сохранить, в первую очередь, путем добавления достаточного количества буфера для композиции в среду, содержащую рекомбинантный ретровирус, с целью образования водной суспензии. Буфер для композиции представляет водный раствор, в котором в воде содержатся сахарид, высокомолекулярная структурная добавка и буферный компонент. Водный раствор может также содержать одну или несколько аминокислот.
Рекомбинантный ретровирус можно также сохранить в очищенной форме. Конкретнее, перед добавлением буфера для композиции неочищенный рекомбинантный ретровирус, описанный выше, можно очистить посредством пропускания через фильтр, а затем сконцентрировать, например, с помощью системы для концентрирования с поперечным потоком (Filtron Technology Corp., Nortborough, Миннесота). В рамках одного из вариантов воплощения изобретения к концентрату добавляют ДНКазу для переваривания экзогенной ДНК. Затем перевар диафильтруют для удаления избытка компонентов среды и закрепляют ретровирус в более желательном забуференном растворе. Затем диафильтрат пропускают через гель-хроматографическую колонку с сефадексом S-500 и элюируют очищенный рекомбинантный вирус. К элюату добавляют достаточное количество буфера для композиции для достижения нужной конечной концентрации составляющих и для минимального разведения рекомбинантного ретровируса и затем водную суспензию хранят предпочтительно при -70oС или сразу же высушивают. Как отмечалось выше, буфер для композиции представляет водный раствор, в котором в воде содержатся сахарид, высокомолекулярная структурная добавка и буферный компонент. Водный раствор может также содержать одну или несколько аминокислот.
Неочищенный рекомбинантный вирус можно также очистить колоночной ионообменной хроматографией. Обычно сырой рекомбинантный вирус очищают посредством пропускания его через фильтр и загружают фильтрат в колонку, содержащую матрицу из целлюлозы с высокой степенью сульфирования. Рекомбинантный ретровирус элюируют из колонки в очищенной форме с использованием буфера с большим содержанием соли. Затем буфер с высоким содержанием соли заменяют на более желательный буфер посредством пропускания элюата через колонку для эксклюзионной хроматографии. Затем к очищенному рекомбинантному ретровирусу добавляют буфер для композиции, как описано выше, и водную суспензию или сразу же сушат или хранят предпочтительно при -70oС.
Водную суспензию неочищенной или очищенной формы можно высушить лиофилизацией или испарением при температуре окружающей среды. Конкретно, лиофилизация включает стадии охлаждения водной суспензии до температуры ниже температуры стеклования или ниже эвтектической температуры водной суспензии и удаления воды из охлажденной суспензии путем сублимации с образованием лиофилизованного ретровируса. Однажды лиофилизованный, рекомбинантный ретровирус стабилен и может храниться при 20-25oС, что подробнее обсуждается ниже.
По методу испарения воду из водной суспензии удаляют при температуре окружающей среды посредством испарения. Воду можно также удалять посредством распылительной сушки.
Водные растворы, используемые для композиции, как описано ранее, состоят из сахарида, высокомолекулярной структурной добавки, буферирующего компонента и воды. Раствор может также содержать одну или несколько аминокислот. Сочетание этих компонентов действует как сохраняющее активность рекомбинантного вируса после замораживания и лиофилизации или сушки посредством испарения.
Высокомолекулярная структурная добавка способствует предотвращению вирусной агрегации во время замораживания и обеспечивает структурную основу в лиофилизованном или высушенном состоянии. В контексте настоящего изобретения структурные добавки рассматриваются как "высокомолекулярные", если их молекулярная масса превышает 5000. Предпочтительной высокомолекулярной структурной добавкой является человеческий сывороточный альбумин. Аминокислоты, если они присутствуют, служат для лучшего сохранения вирусной инфективности после охлаждения и оттаивания водной суспензии. Кроме того, аминокислоты также служат для лучшего сохранения вирусной инфекционности во время сублимации охлажденной водной суспензии и также в лиофилизованном состоянии.
Буферный компонент придает раствору свойства буфера посредством поддержания относительно постоянного рН. Можно использовать ряд буферов в зависимости от желательного интервала рН, предпочтительно с рН от 7,0 до 7,8.
Водные растворы для композиции с рекомбинантными ретровирусами подробнее описываются в WO A2 96/121014.
Кроме того предпочтительно чтобы водный раствор содержал нейтральную соль, используемую для того, чтобы довести конечную композицию с рекомбинантным ретровирусом до соответствующей изоосмотической концентрации соли.
Лиофилизованные или обезвоженные ретровирусы можно восстановить с использованием различных веществ, но предпочтительно их восстанавливают с применением воды. В некоторых случаях можно также использовать разбавленные солевые растворы, которые придают конечной композиции изотоничность. Кроме того, может быть выгодным использование водных растворов, содержащих компоненты, усиливающие, как известно, активность восстановленного ретровируса. Такими компонентами являются цитокины, такие как IL-2, поликатионы, такие как протаминсульфат, или иные компоненты, усиливающие трансдукционную эффективность восстановленного ретровируса. Лиофилизованные или обезвоженные ретровирусы можно восстановить с помощью любого подходящего объема воды или восстанавливающих агентов, создающего возможность для существенной а предпочтительно полной, солюбилизации лиофилизованного или обезвоженного образца.
Рекомбинантные ретровирусные частицы можно вводить в самые разные места, в том числе, например, в такие области, как орган или местоположение опухоли. В рамках других вариантов воплощения изобретения рекомбинантный ретровирус можно вводить перорально, внутривенно, трансбуккально или под язык, интраперитонеально или подкожно. Суточная доза зависит от соответствующего способа введения, формы, в которой ее вводят, указания направления введения, субъекта, которому назначается введение, и массы тела данного субъекта, а также выбора и опыта лечащего врача.
Описанные здесь способы введения можно осуществить просто посредством прямого введения с применением иглы, катетера или соответствующего приспособления. В частности, при некоторых вариантах воплощения изобретения можно вводить непосредственно одну или несколько лекарственных форм.
В одном из вариантов воплощения изобретения упаковывающие клетки заключают в капсулу. Для эффективного лечения продуцирующие вирус клетки должны оставаться в живых в органах-мишенях длительное время после имплантации, и в течение этого времени вирус должен продуцироваться и выделяться из упаковывающих клеток. Таким образом, упаковывающие клетки, продуцирующие вирус, в действительности будут составлять небольшую фабрику по производству вирусов, размещенную в месте применения. Это создаст возможность эффективной доставки рекомбинантного вируса in vivo. С другой стороны, инфицированные здоровые клетки либо человеческого происхождения, либо клетки, происходящие от других видов, будут инкапсулироваться и имплантироваться, обеспечивая небольшую фабрику по конверсии пролекарства, которая может быть размещена вблизи или в массе опухоли.
Продолжительность эффективности такого подхода зависит от (1) защиты клеток от иммунной системы хозяина, которая, естественно, будет удалять продуцирующие вирус или инфицированные клетки, в особенности если это клетки от другого вида, как это обычно бывает в случае клеток, продуцирующих ретровирусный вектор, и (2) выживания клеток in situ продолжительное время, которого может потребовать васкуляриэация.
Обнаружено, что длительного продуцирования векторного вируса из имплантированных упаковывающих клеток можно достичь с помощью соответствующего инкапсулирования перед имплантацией продуцирующих вирус упаковывающих клеток в микрокапсулы с полупроницаемыми оболочками. Кроме того, обнаружено, что такие капсулы хорошо приживаются в хозяине, васкуляризуются и не вызывают иммунной или воспалительной реакции хозяина. Эти находки вместе с полупроницаемостыо капсульных оболочек создают возможность длительной доставки ретровирусного вектора in vivo.
Разработана технология инкапсулирования для инкапсуляции упаковывающих клеток, продуцирующих вирус, и инфицированных вирусом или здоровых клеток в материалы на основе целлюлозы. С использованием этой технологии инкапсулируют до 1010, но предпочтительно 105-107 клеток в электролитный комплекс (например, из альгината и полилизина или предпочтительнее из сульфата целлюлозы и полидиметилдиаллиламмонийхлорида) или другие пористые структуры (такие как полиамиды, полисульфоны). Получающиеся в результате капсулы имеют различный диаметр от 0,01 до 5 мм, но предпочтительно 0,1-1 мм. Следовательно, можно изготовить капсулы, содержащие различное число клеток. Капсула является полупроницаемой, с достаточно большими порами, позволяющими вирусам или молекулам пролекарства проходить через них, но достаточно маленькими, чтобы предохранить клетки от доступа иммунной системы, причем таким образом существенно снижается иммунная реакция на эти клетки. Капсулы и инкапсулированные клетки культивируют в обычной среде для выращивания клеток (природа которой зависит от инкапсулируемой клеточной линии) в стандартных условиях влажности, температуры и содержания СO2.
После соответствующего периода культивирования (обычно не менее 1 часа и не более 30 суток) капсулы, содержащие клетки, можно имплантировать в различные области хирургическим путем, или непосредственно, или посредством инъекции с использованием шприца.
В различные моменты времени после имплантации инкапсулированных клеток хозяина можно лечить циклофосфамидом или ифосфамидом либо местно, либо системно. Клетки, инфицированные вирусом, экспрессирующим цитохромом Р450, будут превращать эти пролекарства в активные метаболиты, вызывающие алкилирование и перекресное связывание ДНК. Все клетки, несущие и экспрессирующие ген цитохрома Р450 (такие как инкапсулированные инфицированные клетки или инкапсулированные упаковывающие клетки) будут также катализировать эту конверсию. В одном из вариантов воплощения данного изобретения эти инкапсулированные инфицированные или упаковывающие клетки будут являться или медленно делящимися клетками, или клетками, обработанными митомицином С, небольшими дозами радиоактивного излучения или другим способом, с целью предотвращения репликации клеток и предохранения, таким образом, клеток от поражения их самих цитотоксическим действием пролекарств.
В дальнейшем изобретение будет поясняться приведенными примерами. Эти примеры, однако, никоим образом не предназначаются для ограничения объема настоящего изобретения, так как будут очевидны понятные модификации, и также еще и другие модификации и замены будут очевидны для специалиста в этой области техники.
Пример 1
В этом примере описывается конструирование ретровирусного экспрессирующего вектора для внутриопухолевой инфекции, содержащего ген крысиного цитохрома Р450 2В1.
Вектор экспрессии pLX2Bl создают путем лигирования фрагментов, полученных из плазмиды pLX125, и pSWl (Kedzie, K.M., Escobar, G.Y., Grimm, S.W., He, Y. A. , Pepperl, D.J., Regan, J.W., Stevens, J.C., and Halpert, J.R., (1991), J. Biol. Chem., 266 (33):2215-21). pLX125 получают так, как описано в РСТ ЕР 96/04447).
Плазмиду pLX125 линеаризируют HpaI и получающиеся в результате затупленные концы дефосфорилируют с использованием телячьей кишечной фосфатазы. ДНК очищают посредством разделения на 1% агарозном геле, иссечения и обработки с применением протокола Qiaquick (Qiagen). После осаждения этанолом ДНК ресуспендируют в воде.
Клонирующий вектор pSWl переваривают с SmaI и HindII и получают два фрагмента с затупленными концами. Переваренную смесь разделяют на 1% агарозном геле. Самый короткий фрагмент (1,5 т.п.о.), содержащий кДНК крысиного цитохрома Р450 2В1 (Fuji-Kuriyama, Y., Mizukami, Y. et al. (1982), Proc. Natl. Acad. Sci. USA, 79:2793-2797) иссекают и элюируют с использованием протокола экстракции ДНК Qiaquick, осаждают этанолом и ресуспендируют в воде.
Смешивают вместе 7,6 фмоль pLXl25 и 24 фмоль SmaI/HindII-фрагмента pSWl и лигируют в течение 3 суток при 12oС с использованием Т-4-лигазы (Boehringer). Лигазу инактивируют при 65oС в течение 10 мин и осаждают ДНК 10-кратным объемом бутанола. Осажденную ДНК ресуспендируют в воде и подвергают электропорации в бактерии DH10B (Gibco). Отбирают устойчивые к ампициллину колонии, получают ДНК и переваривают с SspBI/SalI, BamHI/SspBI, Pvul и BamHI. Конечную исправленную плазмиду обозначают pLX2Bl.
Липофекция
Накануне липофекции 3•105 ретровирусных упаковывающих клеток РА317 (Miller, A.D., and Buttimore, С. (1986), Mol. Cell. Biol., 6:2895-2902) высевают в 10-см чашки Петри или чашки для культивирования микроорганизмов. В день заражения 4 мкг pLX2Bl смешивают с 300 мкл бессывороточной среды. Параллельно смешивают 45 мкл липофектамина (Gibco BRL) с 300 мкл бессывороточной среды. Содержащий плазмиду раствор добавляют к смеси с липофектамином и инкубируют в течение 45 мин. Через 35 мин клетки промывают один раз 6 мл бессывороточной среды. К липофекционной смеси добавляют 2,4 мл бессывороточной среды и 1 мл получающейся смеси помещают на полученные клетки. Через 5 часов добавляют 3,5 мл модифицированной по способу Дульбекко среды Игла, содержащей 20% FCS. На следующий день клетки обрабатывают трипсином и разводят 1: 20 и высевают в 100-мм чашку. Через 24 часа среду заменяют на среду, содержащую аналог неомицина G418. Выделяют клеточные популяции и анализируют на экспрессию цитохрома Р450.
Супернатант от этих популяций используют для заражения клеток-мишеней СК. Фильтруют 1 мл вируссодержащего супернатанта от 5•106 клеток через фильтр 0,45 мкм и добавляют к 1•106 клеток-мишеней СК в присутствии 8 мкг/мл полибрена. Через 4 часа добавляют 1 мл модифицированной по способу Дульбекко среды Игла с 10% фетальной телячьей сыворотки.
На следующий день клетки обрабатывают трипсином, разводят и через 24 часа помещают в среду для селекции, дополнительно содержащую 400 мкг/мл G418. Через 2 недели выделяют колонии, устойчивые к G418, и испытывают на активность цитохрома Р450 2В1. Высевают 2•l04 клеток в 3-см чашку и подвергают действию ифосфамида в концентрации от 0 до 5 мМ. Наблюдают более высокую чувствительность клеток, инфицированных ретровирусом с цитохромом Р450 2В1, по сравнению с контрольными неинфицированными клетками.
Инкапсуляция
Полученные упаковывающие клетки, продуцирующие ретровирусный вектор, инкапсулируют так, как описано в WO 97/01357 в примере 2.
Имплантация
Полученные капсулы вводят посредством операции типа "ключ-скважина" вблизи или в трансплантированные или спонтанные опухоли мышей BALB/c или GR. В место каждой операции вставляют примерно 6 капсул диаметром 1 мм. Место операции закрывают 1 швом. Затем мышей обрабатывают циклофосфамидом или ифосфамидом местно путем прямой инъекции в опухоль 100 мкл СРА при концентрации 20 мг/мл, или системно при концентрации 130 мг СРА на кг массы тела, i.p., и 40-60 мг IFO на кг массы тела, i.p., на протяжении максимум 10 недель. В продолжение этого периода ежедневно контролируют размер и макроскопический вид опухоли. Затем мышей умерщвляют, удаляют ткань, содержащую вставленные капсулы, и готовят гистологические срезы для оптического и электронного микроскопа. Эти срезы ясно показывают хорошее приживление капсул, васкуляризацию и отсутствие признаков наличия лимфоцитов, указывающих на клеточную иммунную реакцию. Эти срезы также показывают отсутствие признаков гибели или некроза клеток в капсулах. Напротив, опухоли показывают некроз, и макроскопически имеется четкое уменьшение размера в процессе испытаний.
Пример 2
В этом примере описывается конструирование устойчивой клеточной линии, экспрессирующей в основном крысиный цитохром Р450 2В1.
Экспрессирующий вектор рс3/2В1 строят путем лигирования фрагментов, полученных из плазмиды pcDNA3 (Invitrogen), и pSWl (Kedzie, K.M., Escobar, G.Y. , Grimm, S.W., He, Y.A., Pepperl, D.J., Regan, J.W., Stevens, J.C., and Halpert, J.R. (1991), J. Biol. Chem., 266(33):2215-21).
Плазмиду pcDNA3 переваривают с XhoI/XbaI и получающиеся в результате фрагменты с "липкими" концами дефосфорилируют с использованием телячьей кишечной фосфатазы. ДНК векторного скелета очищают путем разделения на 1% агарозном геле, иссечения и обработки с применением протокола Qiaquick (Qiagen). После осаждения этанолом ДНК ресуспендируют в воде.
Клонирующий вектор pSWl переваривают с XhoI и XbaI, и получают два фрагмента. Смесь переваривания разделяют на 1% агарозном геле. Самый короткий фрагмент (1,5 т.п.о.), содержащий кДНК крысиного цитохрома Р450 2В1 (Fuji-Kuriyama, Y, Mizukami, Y. et al. (1982), Proc. Natl. Acad. Sci. USA, 79: 2793-2797) иссекают и элюируют с использованием протокола экстракции ДНК Qiaquick, осаждают этанолом и ресуспендируют в воде.
Смешивают вместе 8,3 фмоль скелета pcDNA3 и 24,8 фмоль XhoI/ XbaI-фрагмента Pswl, и лигируют в течение 1 суток при 12oС с использованием Т4-лигазы (Boehringer). Инактивируют лигазу при 65oС в течение 10 мин и осаждают ДНК 10-кратным объемом бутанола. Осажденную ДНК ресуспендируют в воде и подвергают электорпорации в бактерии DH10B (Gibco). Отбирают колонии, устойчивые к ампициллину, получают ДНК и переваривают с EcoRI, BamHI, EcoRV и XhoI. Конечную скорректированную плазмиду обозначают рс3/2В1.
Липофекция
Накануне липофекции 3•106 кошачьих почечных клеток высевают в 100-мм чашки. В день заражения 4 мкг рс3/2В1 смешивают со 100 мкл бессывороточной среды. Параллельно смешивают 15 мкл липофектамина со 100 мкл бессывороточной среды. Содержащий плазмиду раствор добавляют к смеси с липофектамином и инкубируют в течение 45 мин. Через 35 мин клетки промывают один раз 2 мл бессывороточной среды. К липофекционной смеси добавляют 800 мкл бессывороточной среды и 1 мл получающейся смеси помещают на полученные клетки. Через 6 часов добавляют 1 мл DMEM (Glutamax) с 10% FCS. На следующий день клетки обрабатывают трипсином, разводят 1:10 и высевают в 100-мм чашку. Через 24 часа среду заменяют на среду против неомицина. Через 14 суток выделяют устойчивые к неомицину клоны и испытывают на присутствие и активность вектора.
Экспрессирующие цитохром Р450 почечные клетки кошки показывают в десять раз большую чувствительность к ифосфамиду или циклофосфамиду по сравнению с клетками дикого типа.
Эффект свидетеля можно показать на кошачьих почечных клетках дикого типа, а также на клетках Rin5, происходящих из опухоли поджелудочной железы. Это демонстрируется путем сокультивирования клонов почечных клеток кошки с цитохромом Р450 либо с непродуцирующими кошачьими почечными клетками дикого типа либо с клетками Rin5, происходящими из опухоли поджелудочной железы крысы, и добавления ифосфамида. Не продуцирующие СК и Rin клетки уничтожаются токсичными метаболитами изофосфамида, которые продуцируются и выделяются кошачьими почечными клетками с цитохромом Р450 2В1. Титрационные исследования с использованием одного из клонов кошачьих почечных клеток с цитохромом Р450 2В1 показывают, что всего 0,25 мМ изофосфамида оказывает специфическое токсическое действие на эти клетки, а 1-2 мМ приводит к гибели всех клеток. С помощью анализа методом полимеразной цепной реакции клонов кошачьих почечных клеток с цитохромом Р450 2В1 показывают, что получена ДНК генной конструкции Р450 2В1. Другой биохимический анализ клонов с использованием ферментативного деалкилирования 7-пентоксиредоруфина цитохромом Р450 2В1 показывает, что генетически модифицированные клетки продуцируют цитохром Р450 2В1.
Капсулы, содержащие эти клетки, получают так, как описано в примере 1, и имплантируют их мышам вблизи местоположения опухоли. После обработки циклофосфамидом или ифосфамидом оценивают эффективность этой обработки так, как описано выше.
Ретровирусный экспрессирующий вектор, содержащий ген, кодирующий крысиный цитохром Р450-2В1, можно также получить так, как описано ниже в примере 4.
Пример 3
Сначала частично переваривают плазмиду pLX125 рестрикционным ферментом XhoI и получают вектор, который линеаризуют в позиции 3547. Эту линейную плазмиду затем переваривают с рестрикционным ферментом SspBI для удаления короткого фрагмента в пределах полилинкера pLX125. В препаративном геле правильно вырезанный векторный фрагмент выглядит как самая широкая полоса. С использованием протокола Quiaex (Qiagen) в этой полосе элюируют ДНК и очищают от геля.
Чтобы получить ген крысиного цитохрома Р450 2В1, клетки из линии клеток гепатомы крысы НТС лизируют раствором D (4 М гуанидийтиоцианата, 25 мМ цитрата натрия, рН 7, 0,5% N-лаурилсаркозиннатрия, 0,1 М 2-меркаптоэтанола), экстрагируют полную РНК, добавляя 1/15 объема 3 М ацетата натрия в таком же объеме насыщенного водой фенола и добавляют 1/5 объема хлороформа с изоамиловым спиртом (49: 1), всю смесь энергично перемешивают. После 15-минутной выдержки на льду экстракт центрифугируют 20 мин при 4oС при 10000 g. РНК в водной фазе осаждают одним объемом изопропанола в течение 30 мин при -20oС и центрифугируют при 10000 g при 4oС. Осадок промывают 70% этанолом и оставляют при комнатной температуре на 15 мин. После 5 мин центрифугирования при 4oС при 10000 g осадок сушат в вакуумной сушилке и снова растворяют в 0,5% растворе SDS (ДСН).
Экстрагированную ДНК обратно транскрибируют с использованием протокола для синтеза кДНК (Pharmacia). Получающуюся в результате кДНК используют в качестве матрицы для PCR. Затравки создают таким образом, чтобы они содержали сайт рестрикции SspBI (подчеркнут) в левосторонней затравке 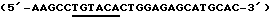 и сайт XhoI (подчеркнут) в правосторонней затравке
и сайт XhoI (подчеркнут) в правосторонней затравке  Обе затравки имеют дополнительные основания по 5'-концам для большей эффективности расщепления соответствующим рестрикционным ферментом. Продукт в 1562 п.о. переваривают с XhoI и SspBI, и получают три фрагмента.
Обе затравки имеют дополнительные основания по 5'-концам для большей эффективности расщепления соответствующим рестрикционным ферментом. Продукт в 1562 п.о. переваривают с XhoI и SspBI, и получают три фрагмента.
Самый длинный фрагмент (1545 п.о.), содержащий ген цитохрома Р450, лигируют с переваренной XhoI/SspBI плазмидой PLX125.
Устойчивую клеточную линию, которая экспрессирует крысиный цитохром Р450, форма 2В1, конструктивно можно получить также так, как описано ниже в примере 5.
Пример 4
Чтобы получить мРНК из крысиного цитохрома Р450, форма 2В1, умерщвляют четырехнедельных самок крыс, извлекают печень и сразу замораживают в жидком азоте. Замороженную печень помещают в стерилизованный фильтрацией буфер GTC (6M гуанидийизоцианата, 5 мМ цитрата натрия, 0,1 М 2-меркаптоэтанола, 0,5% N-лаурилсаркозилнатрия) и гомогенизируют при комнатной температуре. Для отделения РНК печеночный экстракт помещают на слой хлорида цезия (5,7 М хлорида цезия, 0,1 М ЭДТА) и центрифугируют на бакет-роторе при 20oС и 32000 об/мин в течение ночи. После полного удаления супернатанта осажденную РНК снова растворяют в охлажденном на льду 10 мМ трис, рН 7,5 и осаждают в течение ночи при -20oС 1/15 объема 3 М ацетата натрия и 2,5 объемами этанола. РНК центрифугируют в течение 40 мин при 8000 об/мин и 4oС и высушенный осадок ресуспендируют в стерильной воде.
Экстрагированную ДНК обратно транскрибируют с использованием протокола синтеза кДНК (Pharmacia). Получающуюся в результате кДНК используют в качестве матрицы для последующей PCR. Затравки создают таким образом, чтобы они содержали сайт рестрикции EcoRI в левосторонней затравке (5'-CGTGCGGAATTCGGCGGATTCAGCAT-3') и сайт EcoRVI в правосторонней затравке (5'-ATAACGGATATCACCTGGCTGCCTCA-3'). Обе затравки имеют дополнительные основания по 5'-концам для большей эффективности разрезающего фермента. Амплификат в 1588 п. о. переваривают с EcoRI и EcoRV и получают три фрагмента.
Самый длинный фрагмент (1572 п.о.), содержащий ген цитохрома Р450 2В1, лигируют с переваренной EcoRI/EcoRV плазмидой pcDNA3 (Invitrogen).
Пример 5
Конструирование вектора РrоСоn, содержащего ген крысиного цитохрома Р450 под контролем промотора WAP. Экстрагированную РНК, полученную в примере 4, обратно транскрибируют с использованием протокола синтеза кДНК (Pharmacia). Получающуюся в результате кДНК используют в качестве матрицы для PCR. Затравки создают таким образом, чтобы они содержали сайт рестрикции BamHI (подчеркнут) в левосторонней затравке (например, 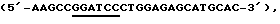 и сайт BamHI (подчеркнут) в правосторонней затравке (например, (5′-CGATTAGGATCCCTGCCTCA-3′). Обе затравки имеют дополнительные основания у 5'-концов для большей эффективности расщепления соответствующим рестрикционным ферментом. Продукт в 1562 п.о. переваривают с BamHI и полученный фрагмент, содержащий ген цитохрома Р450, лигируют с переваренной BamHI плазмидой pWAP.6 (см. заявку РСТ РСТ ЕР96 03922).
и сайт BamHI (подчеркнут) в правосторонней затравке (например, (5′-CGATTAGGATCCCTGCCTCA-3′). Обе затравки имеют дополнительные основания у 5'-концов для большей эффективности расщепления соответствующим рестрикционным ферментом. Продукт в 1562 п.о. переваривают с BamHI и полученный фрагмент, содержащий ген цитохрома Р450, лигируют с переваренной BamHI плазмидой pWAP.6 (см. заявку РСТ РСТ ЕР96 03922).
Изобретение относится к области медицины. Описывается реплекативно-дефектный ретровирусный вектор, несущий ген цитохрома Р450 под транскрипционным контролем регулярных элементов или промоторов, специфических для клетки-мишени, или промоторов, индуцируемых рентгеновским излучением. Изобретение позволяет расширить арсенал противоопухолевых лекарственных композиций. 4 c. и 5 з.п.ф-лы.
| CHEN S.H., CHINE Y.D | |||
| et al | |||
| Proc | |||
| Natl | |||
| Acad | |||
| Sci USA, 1994, Apr | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Gene therapy for brain tamors: regression of experimental gliomas by adenoviris-mediated gene transfer in vivo | |||
| КАТАЛИЗАТОР ДЛЯ ПОЛУЧЕНИЯ ФЕНОЛА И АЦЕТОНА | 1993 |
|
RU2039593C1 |
| US 4789550 A, 06.12.1988. | |||

