Изобретение относится к медицине, а именно к химико-фармацевтической промышленности, и касается способа получения из растительного сырья вещества, обладающего иммуностимулирующей, противовирусной и антибактериальной активностью; вещества, полученного этим способом, и фармацевтической композиции на его основе.
Известен способ получения биологически активного полисахарида путем измельчения растительного сырья, в качестве которого используют ростки клубней картофеля, его экстрагирования кипящей водой, с последующим выдерживанием при 20oС в течение 16-18 часов, сепарацией экстракта, выдержкой его в течение не менее 20 дней, фракционированием его по молекулярному весу и выделением вещества с молекулярным весом более 10000 Дальтон, с последующим концентрированием и сушкой (Ru 2108800 C1, А 61 К 35/78, 20.04.1998). Вещество, полученное этим способом, обладает противовирусной и антибактериальной активностью и используется в качестве лекарственного препарата как наружного, так и внутреннего применения. Известен также способ получения из растительного сырья полисахаридов, обладающих иммуностимулирующим действием, путем обработки растительного сырья водным раствором формалина, выдерживания в подкисленной воде, с последующей экстракцией пектиновых полисахаридов водным раствором оксалата аммония, обработкой экстракта известным способом и лиофильной сушкой целевого продукта. В качестве растительного сырья используют пресноводные цветковые растения, например, любой вид ряски Lemna spp., предварительно измельченную свежую надземную часть высших растений, например, смолевки обыкновенной Oberna behen(L). (Ru 2149642, А 61 К 35/78, 27.05.2000). Ближайшим известным решением к заявленному является способ получения вещества, обладающего иммуностимулирующей, противовирусной и антибактериальной активностью, вещество, полученное этим способом, названное гамма-плантом (γ-PL), и фармацевтическая композиция на его основе (Л.А. Чекановская, А.В. Генералов, Выделение и характеристика биологически активного препарата "Гамма-планта" из ростков картофеля Solanum tuberosum", Хим.-фарм. ж-л, т.34, 3, 2000, с.51-56). Известный способ получения гамма-планта включает измельчение растительного сырья, в качестве которого используют ростки картофеля, экстракцию кипящей водой, центрифугирование водного экстракта, концентрирование, осаждение ацетоном, очистку и сушку целевого продукта. Очистка согласно известному способу включает диализ, гельфильтрацию на сорбенте АсА44, ионообменную хроматографию и на последнем этапе очистки жидкостную хроматографию высокого давления. Вещество, полученное описанным выше способом, представляет собой гликопротеин с молекулярной массой 70 КД, состоящий из углеводной и белковой частей в %-ном массовом соотношении 90:10. При этом углеводная часть гамма-планта на 70% состоит из глюкозы, на 3,7%-арабинозы, на 2,08%-ксилозы, 6,84%-галактозы, 0,58-маннозы, 1%-аминосахаров, 5,1%-уроновых кислот. Фармацевтическая композиция на основе гамма-планта содержит соответствующие наполнители или носители, позволяющие получать различные лекарственные формы. Однако эти препараты проявляют недостаточно высокую иммуностимулирующую активность, что обусловлено, по-видимому, балластными примесями, в т.ч. крахмалом, а также иной структурой активного компонента.
Задачей изобретения является получение нового вещества растительного происхождения из доступного сырья и препарата, обладающего более высокой иммуностимулирующей, противовирусной и антибактериальной активностью, разработка технологичного эффективного способа его получения и приготовление фармацевтической композиции на его основе, обладающей высоким терапевтическим действием.
Поставленная задача решается предлагаемым способом получения вещества, обладающего иммуностимулирующей, противовирусной и антибактериальной активностью, путем экстракции водой измельченного растительного сырья, в качестве которого используют растения семейства Dioscoreaceae, Plantaginaceae, Solanaceae, центрифугирования водного экстракта, его концентрирования, осаждения 96-%-ным этиловым спиртом в присутствии хлористого натрия, повторного переосаждения полученного осадка солевым или кислым агентом с последующей обработкой выделенного кислого пептидогликана-сырца раствором щелочи или насыщенным раствором соли щелочного металла, очисткой целевого продукта с помощью гельпроникающей хроматографии и сушкой.
Неожиданно было обнаружено, что настоящий способ, предусматривающий осуществление указанной выше последовательности операций, позволяет получить новое вещество, обладающее сильно выраженной иммуностимулирующей, противовирусной и антибактериальной активностью и представляющее собой водорастворимый кислый пептидогликан с молекулярной массой 1200-40000 КД, имеющий массовое соотношение между глюкозой и уроновыми кислотами, равное 1: 2-4, и фармацевтическую композицию на его основе, содержащую фармацевтически приемлемый носитель или наполнитель.
Предложенный способ предусматривает использование листьев, стеблей, корней, клубней и проростков растения, т.е. разных частей растения в любой стадии его развития или зрелости.
При переосаждении в качестве солевого агента используют цетилтриметиламмонийбромид или хлористый кальций или в качестве кислого агента - органическую или неорганическую кислоту или кислую неорганическую соль.
Проведение такого переосаждения позволило выделить кислый пептидогликан-сырец, отделив его от многочисленных сопутствующих примесей полисахаридов и белков.
При очистке целевого продукта с помощью гельпроникающей хроматографии в качестве хроматографирующего агента используются TSK HW-75F, сефароза 2В или 4В CL.
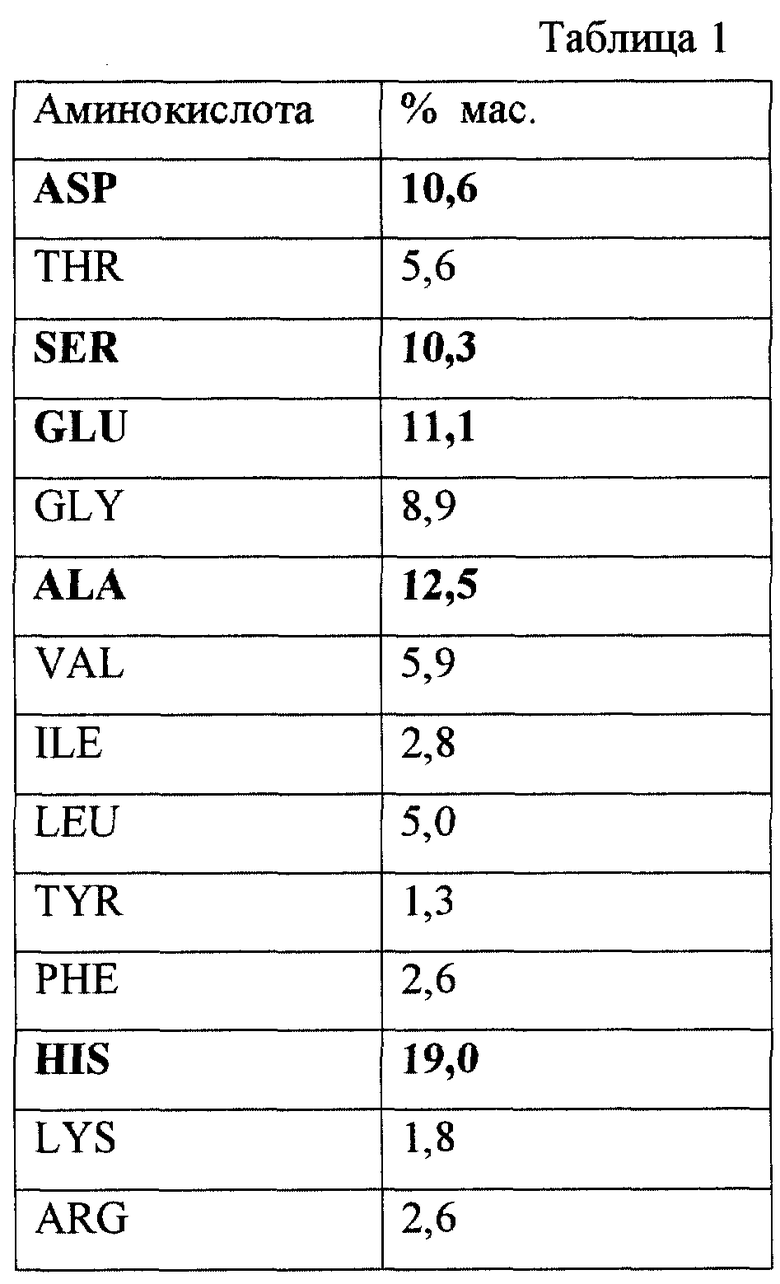
Вещество, полученное настоящим способом, характеризуется тем, что пептидная часть молекулы кислого пептидогликана составляет 13±3 мас.%.
Количество пептида определялось методом Лоури с использованием бычьего сывороточного альбумина в качестве стандарта (О.Н. Lowry, N.J. Rosenbrough et al., J. Biol. Chem., (1951), 193, 265-275).
Пептидная часть кислого пептидогликана состоит из следующих аминокислот, представленных в табл. 1.
Из данных табл. 1 следует, что количественно превалирует содержание гистидина (HIS), аланина (ALA), глутаминовой кислоты (GLU), аспарагиновой кислоты (ASP) и серина (SER).
В полисахаридную часть молекулы входят галактуроновая кислота, глюкоза, галактоза, манноза, арабиноза, рамноза, при следующем их массовом содержании: галактуроновой кислоты 18±6%, глюкозы 9±3%, галактозы 5,5±2%, маннозы 0,7±0,25%, арабинозы 3,8±1,3%, рамнозы 1,9±0,9%.
Количество кислых сахаров определяют по цветной реакции с 3,5-диметилфенолом в концентрированной серной кислоте (A.I. Usov, M.I. Bilan, N.G. Klochkova, Botanica Marina, 1995, 38, 43-51).
Анализ уроновых кислот методом ГЖХ в виде их триметилсилильных производных (P. Albersheim, Methods Enzimol., 1987, 118, 3-40) показал в основном наличие галактуроновой кислоты.
Количество нейтральных сахаров определяют методом ГЖХ в виде их ацетатов полиолов (P.Albersheim, Methods Enzimol., 1987, 118, 3-40).
Нижеследующие примеры иллюстрируют настоящее изобретение.
Пример 1.
Растительные клетки корневой части диоскории семейства Dioscoreaceae (D. cuacasia Lipsky) выращивались в культуральной среде. По окончании инкубации клеточную массу отделяют фильтрованием, а надосадочную жидкость концентрируют, диализуют против дистиллированной воды и лиофильно сушат.
К 5 г сухой смеси, полученной из культуральной жидкости, прибавляют 500 мл дистиллированной воды. Экстракцию проводят при комнатной температуре в течение 3 часов. Нерастворившийся осадок отделяют центрифугированием. Маточник концентрируют до объема 200 мл, добавляют 200 мг хлористого натрия, растворяют его и приливают 600 мл 96% этилового спирта. Осадок выделяют центрифугированием при 4000 об/мин в течение 30 мин. Осадок промывают 50 мл 96% этилового спирта, центрифугируют и сушат на воздухе. Затем проводят повторное переосаждение полученного осадка кислым агентом - органической кислотой. Для этого 1 г сухого полупродукта растворяют в 200 мл дистиллированной воды при перемешивании при комнатной температуре в течение 1 часа. К раствору прибавляют 5 мл 10% раствора трихлоруксусной кислоты. Выпавший осадок, представляющий собой кислый пептидогликан-сырец, отделяют центрифугированием. Промывают водой. Затем прибавляют 50 мл дистиллированной воды и при перемешивании прикапывают 25% раствор аммиака до полного растворения осадка.
Полученный раствор наносят на колонку TSK HW-75F. Колонку элюируют водой. Собирают первый высокомолекулярный пик с молекулярной массой от 1200 до 40000 КД. Раствор концентрируют на роторном испарителе и сушат лиофильно. Выход кислого пептидогликана составляет 39 мг. Массовое соотношение между глюкозой и уроновыми кислотами составляет 1:3.
Количество пептида в образце составляет 11,2%.
Пример 2.
100 г свежесобранной надземной части (стебли) диоскории семейства Dioscoreaceae (D. cuacasia Lipsky) измельчают, добавляют 500 мл дистиллированной воды и экстрагируют при нагревании 30-40oС в течение 5 часов.
Смесь отжимают с помощью механического пресса. Водный экстракт диализуют и концентрируют до объема 100 мл путем упаривания на роторном испарителе. К концентрату прибавляют хлористый натрий из расчета 100 мг на 100 мл и 3 объема 96% этилового спирта. Осадок выделяют центрифугированием при 4000 об/мин в течение 30 мин.
Осадок промывают 30 мл 96% этилового спирта, центрифугируют и сушат на воздухе. Затем проводят повторное переосаждение полученного осадка кислым агентом - неорганической кислотой. Для этого 1 г сухого полупродукта растворяют в 200 мл дистиллированной воды при перемешивании при комнатной температуре в течение 1 часа. Нерастворившийся осадок отбрасывают, а к маточнику прибавляют 5 мл конц. НСl. Выпавший осадок отделяют центрифугированием. Промывают водой. Затем прибавляют 50 мл дистиллированной воды и при перемешивании прикапывают 25% раствор аммиака до полного растворения осадка.
Полученный раствор наносят на колонку с TSK HW-75F. Колонку элюируют водой. Собирают первый высокомолекулярный пик с молекулярной массой от 1200 до 40000 КД. Раствор концентрируют на роторном испарителе и сушат лиофильно. Выход кислого пептидогликана составляет 16 мг. Массовое соотношение между глюкозой и уроновыми кислотами составляет 1:2,2.
Количество пептида в образце составляет 14,7%.
Пример 3.
100 г свежесобранных листьев подорожника семейства Plantaginaceae (P. major L.) измельчают, добавляют 500 мл дистиллированной воды и экстрагируют при нагревании 30-40oС в течение 4 часов.
Смесь отжимают с помощью механического пресса. Водный экстракт диализуют и концентрируют до объема 100 мл путем упаривания на роторном испарителе. К концентрату прибавляют хлористый натрий из расчета 100 мг на 100 мл и 3 объема 96% этилового спирта. Осадок выделяют центрифугированием при 4000 об/мин в течение 30 мин.
Осадок промывают 30 мл 96% этилового спирта, центрифугируют и сушат на воздухе. Затем проводят повторное переосаждение полученного осадка кислым агентом - неорганической кислотой. Для этого 1 г сухого полупродукта растворяют в 200 мл дистиллированной воды при перемешивании при комнатной температуре в течение 1 часа. Нерастворившийся осадок отбрасывают, а к маточнику прибавляют 5 мл конц. НСl. Выпавший осадок отделяют центрифугированием. Промывают водой. Затем прибавляют 50 мл дистиллированной воды и при перемешивании прикапывают 25% раствор аммиака до полного растворения осадка.
Полученный раствор наносят на колонку TSK HW-75F. Колонку элюируют водой. Собирают первый высокомолекулярный пик с молекулярной массой от 1200 до 40000 КД. Раствор концентрируют на роторном испарителе и сушат лиофильно. Выход кислого пептидогликана составляет 16 мг. Массовое соотношение между глюкозой и уроновыми кислотами составляет 1:2,7.
Количество пептида в образце составляет 13,8%.
Пример 4.
Аналогичен примеру 3, но для высаживания кислого пептидогликана вместо 5 мл конц. HCl используют 200 мл насыщенного раствора кислой неорганической аммонийной соли МН4NО3 или (NH4)2SO4. Выход конечного продукта составляет 14 мг. Массовое соотношение между глюкозой и уроновыми кислотами составляет 1: 2,4.
Количество пептида в образце составляет 15,8%.
Пример 5.
5 кг проростков картофеля семейства Solanaceae (S. tuberosum) измельчают, прибавляют 10 л воды и экстрагируют при комнатной температуре при перемешивании в течение 2 часов. Смесь отжимают с помощью механического пресса. Водный экстракт диализуют и концентрируют до объема 1 л путем ультрафильтрации на фильтре 10 КД.
К концентрату прибавляют хлористый натрий из расчета 100 мг на 100 мл и 3 объема 96% этилового спирта. Осадок выделяют центрифугированием при 4000 об/мин в течение 30 мин.
Осадок промывают 300 мл 96% этилового спирта, центрифугируют и сушат на воздухе. Затем проводят повторное переосаждение полученного осадка солевым агентом - хлористым кальцием. Для этого 10 г сухого полупродукта растворяют в 1 л дистиллированной воды при перемешивании при комнатной температуре в течение 1 часа. Нерастворившийся осадок отбрасывают, а к маточнику прибавляют 150 мл 5% раствора хлористого кальция. Выпавший осадок, представляющий собой кислый пептидогликан-сырец, отделяют центрифугированием. Затем переводят его в растворимую форму путем добавления насыщенного раствора хлористого натрия и выдерживают смесь при перемешивании и нагревании при 50oС до полного растворения. Полученный раствор наносят на колонку TSK HW-75F. Колонку элюируют водой, собирают первый высокомолекулярный пик с молекулярной массой от 1200 до 40000 КД. Раствор концентрируют на роторном испарителе и сушат лиофильно. Выход кислого пептидогликана составляет 170 мг. Массовое соотношение между глюкозой и уроновыми кислотами составляет 1:4.
Пример 6.
Аналогичен примеру 4, но в качестве органов растения используют клубни картофеля и в качестве солевого агента при проведении повторного переосаждения осадка используют 9% раствор цетилтриметиламмонийбромида. Выход кислого пептидогликана составляет 14 мг. Массовое соотношение между глюкозой и уроновыми кислотами составляет 1:3,5. Количество пептида в образце составляет 16%.
Пример 7. Иммуностимулирующая активность, проявляющаяся в активации синтеза антител.
Введение кислого пептидогликана (КПГ) одновременно с чужеродным антигеном приводит к значительному усилению интенсивности продукции антител, специфичных к введенному антигену. В экспериментах использовали мышей СВА, С57В1/6 (CBAxC57B1/6)F1 и BALB/c. В качестве антигенов для иммунизации мышей применяли бычий сывороточный альбумин (БСА), яичный альбумин (ЯА) или бараньи эритроциты (БЭ). Эритроциты дефибринированной крови барана трижды отмывали, центрифугируя в 50-кратном объеме раствора Хэнкса, затем ресуспендировали в том же растворе. Мышам вводили внутрибрюшинно 2 млн. БЭ. Иммунизацию 200 мкг БСА проводили внутрибрюшинно, повторную иммунизацию той же дозой БСА выполняли спустя 4 недели. ЯА вводили внутрибрюшинно в дозе 50 мкг, повторные иммунизации ЯА проводили через 2 и 4 недели после первой иммунизации.
Соответствующую навеску КПГ растворяли в растворе Хэнкса или физиологическом растворе NaCl за 2-3 часа до использования. КПГ вводили мышам в дозах от 1 до 1000 мкг. В случае иммунизации мышей БЭ, БСА или ЯА одновременно с антигеном вводили КПГ.
Уровень иммунного ответа у мышей, иммунизированных БЭ, определяли по количеству антителообразующих клеток (АОК), выявляемых в селезенке методом Ерне через 4-5 сут после иммунизации. Содержание антител, специфичных к ЯА или БСА, в сыворотке крови мышей определяли методом твердофазного иммуноферментного анализа (ИФА). Изотип антител, специфичных к БСА, определяли методом ИФА с помощью кроличьих антител, специфичных к IgM, IgG1, IgG2a, IgG2b или IgG3 мыши.
Совместное введение КПГ с гетерологичным антигеном экспериментальным мышам приводило к иммунной реакции, значительно более интенсивной, чем при введении только антигена. Так, инъекция КПГ вместе с субоптимальной иммуногенной дозой БЭ приводила к 10-кратному усилению продукции антителопродуцентов, специфичных к БЭ.
После иммунизации БСА в сочетании с КПГ уровень (ИФА-титр) специфических к БСА антител в сыворотке крови мышей достигал 1:20000, что было намного выше уровня специфических антител при иммунизации только БСА (ИФА-титр не более 1: 500). При введении КПГ существенно увеличивалась не только интенсивность, но и продолжительность вторичной иммунной реакции на БСА. При этом доминировали изотипы антител IgG1, IgG2a и IgG3. Продукция IgM и IgG2b, специфичных к БСА, повышалась под влиянием КПГ в меньшей степени.
Адъювантный эффект отчетливо зависел от дозы КПГ. При вторичной реакции на БСА оптимальной иммуностимулирующей дозой оказалась доза 1 мкг КПГ, при первичной иммунной реакции на гетерологичные эритроциты оптимальными были дозы 10 мкг КПГ или более.
Результаты влияния КПГ на продукцию антител к ЯА представлены в табл. 2. ЯА оказался высоко иммуногенным антигеном для мышей BALB/c, титр антител после введения 50 мкг ЯА без какого-либо адъюванта достигал 1:13000. Совместное введение КПГ с 50 мкг ЯА приводило к значительному повышению уровня антительного ответа (титр 1:22000). КПГ оказывал более сильное адъювантное действие, чем известный иммуноадъювант липопептид Pam3Cys-Ser-Lys4 (титр 1: 20000), но уступал адъюванту Фройнда (титр 1:33000).
Пример. 8. Иммуностимулирующая активность: активация тканевых макрофагов.
Тканевые макрофаги мыши, полученные из перитонеального экссудата, значительно активируются при их культивировании in vitro в присутствии КПГ. Активация макрофагов проявляется в изменении их размеров и формы, а также в изменении их метаболической и ферментативной активностей.
Клетки перитонеального экссудата (КПЭ) получали путем промывания брюшной полости мышей (CBAxC57B1/6)F1 3 мл среды 199. Полученные от 10-15 мышей КПЭ в концентрации 2-2,5•106 в 1 мл собирали в силиконизированные пробирки.
Продукция макрофагами окислительных радикалов
Суспензию клеток перитонеального экссудата разливали по 1 мл в пробирки хемилюминографа и инкубировали при температуре 37oС в увлажненной атмосфере 5% СО2 в воздухе в течение 2 часов. После инкубации неприлипшие клетки смывали, а прикрепившиеся к пробирке клетки 2 раза промывали средой 199. Затем в пробирки вносили по 1 мл полной среды RPMI-1640, содержащей КПГ в концентрациях от 0,2 до 50 мкг в 1 мл и инкубировали в указанных условиях еще 24 часа. После инкубации убирали надосадок, добавляли 0,5 мл буферного раствора (рН 7,2), приготовленного из раствора Хенкса (без фенолового красного), дополненного 5 mM глюкозы, 10 mМ HEPES-буфера, 0,62 mM люминола (Sigma Chemical Co), и оценивали уровень спонтанной и индуцированной зимозаном хемилюминесценции. Результаты по сумме трех экспериментов представлены в виде среднего числа импульсов в минуту на 1 млн. клеток.
Инкубация прилипающих КПЭ в присутствии КПГ в течение 24 часов не влияла на уровень их спонтанной хемилюминесценции, но значительно (на 50%) повышала способность клеток к продукции окислительных метаболитов в ответ на зимозан, представляющий собой компоненты клеточной стенки микроорганизмов. При этом увеличение хемилюминесцентной активности наступает уже при концентрации КПГ 0,2 мкг/мл, а максимальный эффект достигается при концентрации 5,5 мкг/мл, что хорошо коррелирует с концентрацией КПГ, приводящей к максимальной активации макрофагов, если судить по их морфологическим признакам.
Активность 5'-нуклеотидазы (5'-НТД) на поверхности макрофагов
Определение уровня фермента 5'-НТД является одним из высоко информативных способов оценки активации макрофагов. Установлено, что падение активности данного фермента под влиянием иммуномодуляторов характеризует наличие у них иммуноадъювантных свойств и коррелирует с высокой противоинфекционной активностью.
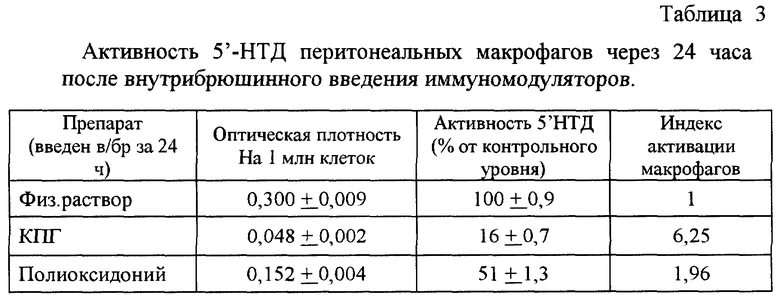
Для определения влияния КПГ на уровень 5'-НТД перитонеальных макрофагов мышам (CBAxC57B1/6)F1 вводили внутрибрюшинно раствор 30 мкг КПГ в объеме 0,5 мл. Контрольным животным вводили 0,5 мл физиологического раствора. Через 24 часа после инъекции получали клетки перитонеального экссудата путем промывания брюшной полости мышей 5 мл среды 199. Полученные суспензии инкубировали при температуре 37oС и 5% СO2 в течение 2 часов в пластиковых чашках Петри диаметром 100 мм, по 10 мл суспензии КПЭ на чашку. После удаления не прилипших клеток оставшиеся прилипшие клетки снимали с поверхности чашки с помощью резинового шпателя, доводили концентрацию КПЭ до 2 млн. клеток в объеме 50 мкл. Этот объем суспензии вносили в лунки 96-луночных планшетов, добавляли 5'-аденозимонофосфат, являющийся субстратом 5'-НТД, и инкубировали 60 минут в указанных выше условиях. Активность фермента определяли фотометрически (длина волны 620 нм) по интенсивности окрашивания молибденового реактива, внесенного в образцы клеток после инкубации. Результаты выражали в единицах оптической плотности в пересчете на 1 млн КПЭ и в виде относительной активности 5'-НТД в процентах от контрольного уровня.
Активность КПГ сравнивали с активностью иммуномодулятора "Полиоксидоний", который хорошо известен как активатор фагоцитов (табл. 3). Полученные результаты свидетельствуют об эффективной активации макрофагов in vivo как КПГ, так и полиоксидонием. При этом КПГ превосходил препарат сравнения по своему активационному действию на макрофаги. Так, под влиянием полиоксидония активность 5'-НТД снижалась в 2 раза, а под влиянием КПГ - в 6 раз, достигая 16% от контрольного уровня. Эти данные свидетельствуют как об активации резидентных макрофагов, так и о привлечении в место введения новых активных макрофагов с высоким бактерицидным потенциалом, то есть доказывают высокую иммуномодулирующую активность КПГ.
Пример 9. Иммуномодулирующая активность: активация клеток иммунной системы человека (NK-клетки, моноциты, гранулоциты).
В условиях in vitro исследовали влияние КПГ на клетки иммунной системы, циркулирующие в периферической крови человека.
Активация NK-клеток человека
Методом трехцветной лазерной проточной цитометрии было обнаружено быстрое появление маркера активации на NK-клетках после воздействия КПГ.
Кровь из локтевой вены получали от здоровых доноров утром натощак в пробирки с антикоагулянтом гепарином натрия с помощью системы "Вакутейнер" (фирма "Бектон Дикинсон"). Препарат КПГ разводили в концентрации 1 мг/мл в среде RPMI-1640 и фильтровали через фильтр 0,22 мкм. Готовили необходимые разведения препарата в полной среде RPMI-1640 в диапазоне от 0,016 до 100 мкг/мл и вносили по 0,2 мл в лунки 48-луночного культурального плейта (фирма "Нунк"). В качестве отрицательного контроля вносили 0,2 мл полной среды RPMI-1640. Во все лунки добавляли по 0,2 мл цельной гепаринизированной крови. Образцы инкубировали при 37oС и 5% СО2 в течение 3-48 часов. После инкубации 0,05 мл образца крови помещали в 1,2 мл полиэтиленовую пробирку. Вносили по 5 мкл следующих антител: анти-СD16-ФИТЦ (фирма "Сорбент"), анти-СD69-фикоэритрин ("Фарминген"), анти-СD3-РеrСR (фирма "Бектон Дикинсон"). Пробирки встряхивали 5 сек, а затем инкубировали при комнатной температуре в темноте в течение 20 минут. В каждую пробирку приливали по 1 мл лизирующего-фиксирующего раствора (фирмы "Бектон Дикинсон") и выдерживали 15 минут при комнатной температуре. Пробирки центрифугировали 10 мин при 300 g, надосадочную жидкость удаляли, а клеточный осадок ресуспензировали в 0,5 мл изотонического раствора. Анализ флуоресценции осуществляли на проточном лазерном цитофлуориметре "ФАКС Калибур" фирмы "Бектон Дикинсон" с использованием программного обеспечения "Cell Quest".
Без добавления КПГ только 7% NK-клеток экспрессировали маркер активации CD69, а при действии КПГ в концентрации 10 мкг/мл практически все (96%) NK-клеток были активированы.
Усиление цитолитической активности NK-клеток под влиянием КПГ
Из цельной гепаринизированной крови вьщеляли мононуклеарную фракцию путем центрифугирования (30 мин 300g при комнатной температуре) в ступенчатом градиенте плотности фиколла ("Фармация"). Выделенные клетки отмывали в среде 199 и разводили до концентрации 2 млн/мл среде RPMI-1640, содержащей 10% фетальной телячьей сыворотки, 20 мМ Хепес-буфера (рН 7.4), 10 мкг/мл гентамицина (полная среда, ПС).
В лунки 12-луночного планшета (фирмы "Нунк") вносили по 1 мл клеточной суспензии и добавляли по 1 мл ПС (контроль) или 1 мл раствора КПГ 20 мкг/мл, или 1 мл раствора стандартного активатора NK-клеток интерлейкина-2 (ИЛ-2, 20 МЕ/мл). Образцы инкубировали в течение 3 часов при 37oС в атмосфере 5% СO2. После инкубации клетки собирали в центрифужные пробирки, осаждали центрифугированием (10 мин 300g) и доводили концентрацию до 5 млн. клеток в 1 мл ПС. Полученные клетки служили эффекторами цитолиза. Серию 2-кратных разведений в ПС клеток-эффекторов разливали по 0,1 мл в триплетах в лунки 96-луночного круглодонного планшета. Во все лунки добавляли по 10 тыс. меченных 3H-уридином клеток-мишеней К-562 в объеме 0,1 мл ПС. Смесь клеток-эффекторов и клеток-мишеней инкубировали в течение 4 часов при 37oС в атмосфере 5% СО2. После инкубации клетки переносили на бумажные фильтры с помощью сборщика клеток Titertek Cell Harvester 550 и подсчитывали радиоактивность на счетчике Wallac 1409.
После 3-часовой инкубации клеток-эффекторов в присутствии КПГ 29% NK-клеток экспрессировали CD69, то есть были активированы. Тестирование цитолитической активности клеток-эффекторов, предварительно активированных КПГ, против К562 клеток-мишеней показало резкое нарастание способности NK-клеток лизировать мишень. Это заметно при низких отношениях эффектор : мишень, в частности, при отношении 6,25 и 12,5, когда эффективность лизиса возрастала под влиянием КПГ примерно в три раза. Если сравнивать действие КПГ с ИЛ-2, известным стимулятором NK-клеток, то видно, что КПГ в несколько раз более эффективен, чем ИЛ2, как по экспрессии CD69, так и по лизису клеток-мишеней К562.
Активация гранулоцитов
При изучении влияния КПГ на экспрессию активационных маркеров на различных типах клеток было обнаружено, что, кроме моноцитов и NK-клеток, активационный маркер CD69 экспрессируется и на гранулоцитах. Через 24 часа инкубации в присутствии КПГ активация гранулоцитов становилась значительной, к этому сроку около 60% гранулоцитов экспрессировали CD69.
Пример 10. Иммуностимулирующее действие: активация синтеза и секреции цитокинов.
КПГ активирует выработку цитокинов клетками крови человека. В частности, КПГ запускает продукцию интерлейкина-1β (ИЛ-1β), фактора некроза опухоли (α-ФНО), интерлейкина-8 (ИЛ-8). Кровь из локтевой вены получали от здоровых доноров утром натощак в пробирки с антикоагулянтом гепарином натрия с помощью системы "Вакутейнер" (фирма "Бектон Дикинсон"). Препарат КПГ разводили в концентрации 1 мг/мл в среде RPMI-1640 и фильтровали через фильтр 0,22 мкм. Готовили необходимые разведения препарата в полной среде RPMI-1640 в диапазоне от 0,016 до 100 мкг/мл и вносили по 0,2 мл в лунки 48-луночного культурального плейта (фирма "Нунк"). В качестве отрицательного контроля вносили 0,2 мл полной среды RPMI-1640. Во все лунки добавляли по 0,2 мл цельной гепаринизированной крови. Образцы инкубировали при 37oС и 5% СO2 в течение 3-48 часов. После инкубации отбирали по 0,2 мл культуральной среды в 0,5 мл центрифужные пробирки и центрифугировали при 10000 об/мин 15 минут. Супернатант замораживали при -70oС и использовали для определения секретируемых цитокинов. Клетки, оставшиеся в лунке, тщательно пипетировали и переносили в 1,2 мл полиэтиленовые пробирки для определения поверхностных маркеров активации.
ИЛ-1β определяли в культуральной среде методом иммуноферментного анализа. Для измерения концентрации ИЛ-1β использовали набор фирмы "Иммунотех" (Франция) в соответствии с инструкцией по его использованию. α-ФНО определяли с помощью наборов фирмы "Иннодженетикс" (Бельгия). Интерлейкин-8 (ИЛ-8) определяли с помощью наборов фирмы "Иннодженетикс" (Бельгия). Интенсивность окраски измеряли при длине волны 450 нм на автоматическом ридере "Дайнатек".
Определение внутриклеточного содержания цитокинов в моноцитах крови человека проводили методом трехцветной лазерной проточной цитофлуорометрии.
Цельную кровь разводили 1:1 в среде RPMI-1640, содержащей различные концентрации препарата и ингибитор транспорта белка брефельдин А ("Сигма") в конечной концентрации 10 мкг/мл. Образцы инкубировали при 37oС и 5% СО2 в течение 5 часов. После окончания инкубации отбирали по 0,1 мл суспензии и добавляли по 1 мл лизирующего-фиксирующего раствора (фирмы "Бектон Дикинсон"). Выдерживали 15 минут при комнатной температуре. Пробирки центрифугировали 10 мин при 300 g, надосадочную жидкость отсасывали, добавляли к клеточному осадку 0,5 мл раствора для повышения проницаемости клеточной мембраны (фирмы "Бектон Дикинсон") и выдерживали 10 минут при комнатной температуре. Эта процедура позволяет окрашивать внутриклеточное содержимое. После добавления 5 мл изотонического раствора, содержащего 0,5% БСА и 0,1% азида натрия, клетки осаждали центрифугированием (10 минут 300 g), надосадочную жидкость отсасывали. К клеточному осадку добавляли по 5 мкл антител. Для определения содержания ИЛ-1β в моноцитах использовали следующие антитела: анти-ИЛ-1β-FITC ("Калтак"), анти-СD14-РЕ ("Калтак") и анти-CD45-PerCP ("Бектон Дикинсон"). Аналогичным образом определяли внутриклеточное содержание α-ФНО в моноцитах.
Из полученных данных следует, что КПГ стимулирует производство α-ФНО в моноцитах и что именно моноциты являются источником α-ФНО в среде. Действительно, если в инкубационную среду добавлен брефельдин, ингибитор секреции белка из клеток во внеклеточную среду, наблюдается накопление α-ФНО внутри моноцитов, при этом содержание α-ФНО в среде - на уровне фона. Если же ингибитор транспорта белка отсутствует, то накопления α-ФНО в моноцитах не происходит, а концентрация α-ФНО в среде нарастает прямо пропорционально концентрации КПК. Из дозовой зависимости накопления α-ФНО в моноцитах видно, что уже при концентрации 40 нг/мл КПГ моноциты заметно усиливают синтез α-ФНО. При дозе 0,4 мкг/мл происходит достаточно интенсивная выработка ИЛ-1β, α-ФНО и ИЛ-8. Если сравнить концентрации цитокинов во внеклеточной среде, то видно, что ИЛ-8 вырабатывается в несколько раз больше, чем ИЛ-1β и α-ФНО.
Пример 11. Индукция синтеза интерферонов.
Изучение интерферон-индуцирующей активности КПГ проводили in vitro в перевиваемых культурах клеток человека, инфицированных вирусом энцефаломиелита (ВЭМ). Интенсивность индукции интерферона оценивали по противовирусному действию КПГ. В экспериментах использовали линии клеток человека L-41 (фибробластоподобные клетки, используются для титрования ИФН-α и ИФН-γ) и J-96 (линия моноцитоподобных клеток). Клетки в концентрации 200000 клеток/мл высевались в 96-луночные плоскодонные культуральные планшеты в среде 199 (J-96) или Игла (L-41) с добавлением 10% сыворотки эмбрионов крупного рогатого скота, глутамина (300 мкг/мл) и антибиотика (100 ЕД/мл пеницилина).
В качестве контрольного индуктора интерферонов использовали Ридостин (НПО "Вектор", Новосибирск, Россия). Противовирусный (интерфероногенный) эффект оценивался по минимально эффективной концентрации (наибольшее разведение препарата, эффективно подавляющее рост вируса) препарата, защищающей 50% клеток от цитопатогенного действия ВЭМ (ЦПД50). Интерфероногенным принято считать вещество, снижающее концентрацию вируса в культуре клеток на 1,7-2,0 lg. Для определения ЦПД вируса использовали инвертированный микроскоп (Leitzб увеличение 200х). Каждый исследуемый препарат в культуральных планшетах с клетками L-41 или J-96 клеток титровали с шагом 1:2 в 24 лунках и 3 повторах каждого разведения. Эксперименты повторяли 3 раза.
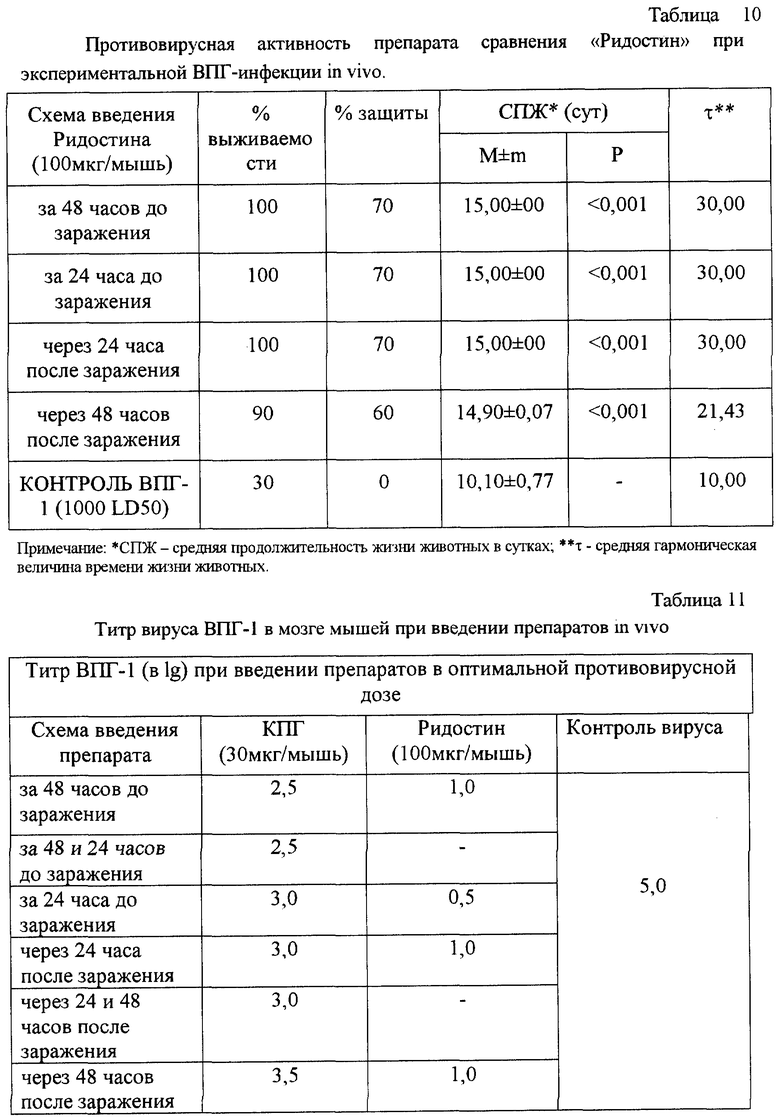
В табл. 4 и 5 представлены данные, показывающие минимально эффективные концентрации и максимально эффективные разведения, при которых исследуемые препараты проявляют интерфероногенную активность в культурах клеток L-41 и J-96. Представленные данные доказывают, что КПГ обладает выраженной противовирусной активностью in vitro в концентрациях, выше 6-50 мкг/мл. Сравнение КПГ с Ридостином, известным противовирусным препаратом - индуктором интерферона, свидетельствует об интерфероногенном действии КПГ.
Пример 12. Активация антибактериальной защиты.
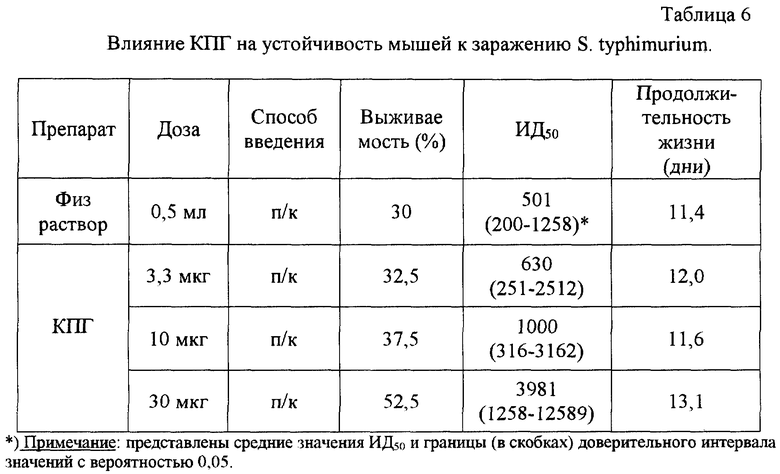
Препарат КПГ исследовался в модели экспериментальной инфекции Salmonella typhimurium у беспородных лабораторных мышей. Эксперименты проводили на мышах обоего пола массой 12-14 г. Препарат вводили подкожно в разных дозах (3,3 мкг, 10 мкг или 30 мкг) за 24 часа до заражения сальмонеллами. Заражение мышей производили внутрибрюшинно дозами 102, 103, 104 или 105 микробных клеток на мышь. Наблюдение за гибелью животных вели в течение 20 дней. Эффективность препарата оценивали по его влиянию на выживаемость инфицированных животных, ЛД50 инфекции (ИД50) и продолжительность жизни.
Полученные результаты представлены в табл. 6. Видно, что однократное подкожное введение КПГ в дозах 10-30 мкг приводило к значительному повышению резистентности мышей к инфекции. Возрастали процент выживших животных, средняя продолжительность жизни инфицированных и, особенно сильно, доза инфекции, приводящая к 50% гибели (ИД50).
Антимикробная защита, усиленная КПГ, не ограничена биологическим видом микроба или хозяина. Например, КПГ эффективно усиливает защиту цыплят от стафилококковой инфекции.
В работе были использованы цыплята яичного направления. КПГ вводили per os в дозе 10 мкг (на цыпленка) дважды с интервалом 96 ч между введениями. Для сравнения в контрольных группах вводили физиологический раствор NaCl или препарат сравнения - иммуномодулятор полиоксидоний. Через 5 дней после второго введения препарата цыплят заражали внутримышечным введением Staphylococcus aureus 209R в дозе 0,75•109, 109 или 2•109 микробных тел. После заражения у цыплят развивалась острая септицемия, они погибали в течение 10 дней. Так, через 10 дней после заражения дозой 2•109 микробных тел погибало 95% инфицированных цыплят (табл. 7). Гибель цыплят в группах, получивших КПГ или полиоксидоний, была существенно меньше, она составила 45 и 55%, соответственно. При использовании сублетальных инфицирующих доз КПГ и полиоксидоний защищали 100% инфицированных цыплят, в то время как в контроле заражения погибало 40% при заражении 0,75•109 S. aureus и 60% - при заражении 109 S. aureus.
В химиотерапевтических опытах применяли антибактериальный препарат нифулин, который вводили в дозе 10 мг/кг per os через 20 минут после заражения цыплят 1,5•109 S. aureus. Полученные результаты представлены в табл. 8. Лечение нифулином позволило увеличить выживаемость цыплят до 70% по сравнению с 40% в контроле заражения. Сочетание КПГ с нифулином защищало всех инфицированных цыплят.
Полученные результаты свидетельствуют о высокой активности препарата КПГ. КПГ значительно повышал устойчивость цыплят к заражению различными дозами Staphylococcus aureus 209R. Для активации противоинфекционного иммунитета КПГ был более эффективен, чем иммуномодулятор полиоксидоний.
Пример 13. Активация противовирусной защиты.
Противовирусный эффект КПГ исследовали in vivo на белых беспородных (б/п) мышах на модели вируса простого герпеса 1 типа (ВПГ-1) штамм Л2 (Шубладзе А. К. , Гайдамович С.Я., "Краткий курс практической вирусологии", 1954 г.; Пшеничнов В.А., Семенов Б.Ф., Зезеров Е.Г. "Стандартизация методов вирусологических исследований", 1974г., стр. 123-128). Предварительно вирус прошел 3 пассажа на белых б/п мышах весом 7-8 г при внутримозговом заражении. Титр вируса при этом составил 7 lg ЛД50/0,03 мл. В основном опыте мышей весом 10-12 г заражали внутрибрюшинно вирусом в виде 10% суспензии мозга мышей в дозе 1000 ЛД50. Титр вируса при внутрибрюшинном заражении был равен 3,5 lg/0,2 мл. Препарат КПГ вводили животным внутрибрюшинно в различных дозах: 1,0 мкг/мышь, 3,0 мкг/мышь, 10 мкг/мышь, 30 мкг/мышь по одной из следующих схем:
1) за 48 часов до заражения,
2) за 48 и 24 часа до заражения,
3) за 24 часа до заражения,
4) через 24 часа после заражения,
5) через 24 и 48 часов после заражения,
6) через 48 часов после заражения.
В качестве положительного контроля использовали Ридостин (известный противогерпетический препарат, индуктор интерферона), в дозе 100 мкг/мышь. Препарат сравнения вводили мышам по разработанной ранее схеме:
1) за 48 часов до заражения,
2) за 24 часа до заражения,
3) через 24 часа после заражения,
4) через 48 часов после заражения.
Контрольную группу составляли мыши, зараженные ВПГ-1 и получавшие плацебо. Каждая группа состояла из 20 животных. Срок наблюдения - 15 дней.
Противовирусный эффект препаратов оценивали по выживаемости мышей (% выживаемости, % защиты, средняя продолжительность жизни СПЖ), а также в опытах биологического титрования мозга зараженных животных (по 5 мышей на точку) на клетках VERO. Присутствие ВПГ-1 (титр) в ткани мозга опытных и контрольных мышей, взятой на 7 день после заражения, определяли в 3 повторах на клетках VERO. Титр определяли в результате обработки культуры клеток 10-кратными разведениями суспензии мозга. Наблюдение и учет ЦПД осуществляли на 3-4 день после контакта с инфекционным материалом.
Оценку активности препаратов проводили, учитывая:
1) среднюю гармоническую величину времени жизни (τ) для каждой группы мышей, включающую в себя информацию о всех особях, вычисляемую по формуле: 1/τ= (X1/t1+X2/t2+...+Xn/tn):n, где t - день гибели животных, X - количество погибших животных в день t, n - количество животных в группе (20 мышей);
2) кратность защиты (КЗ) - кратность уменьшения количества заболевших и павших животных в опытной группе по сравнению с контролем, вычисляемую по формуле:
КЗ= % павших животных в контрольной группе /% павших животных в опытной группе;
3) индекс эффективности препарата (ИЭ), вычисляемый по формуле:
ИЭ=[(КЗ-1)/КЗ]•100%.
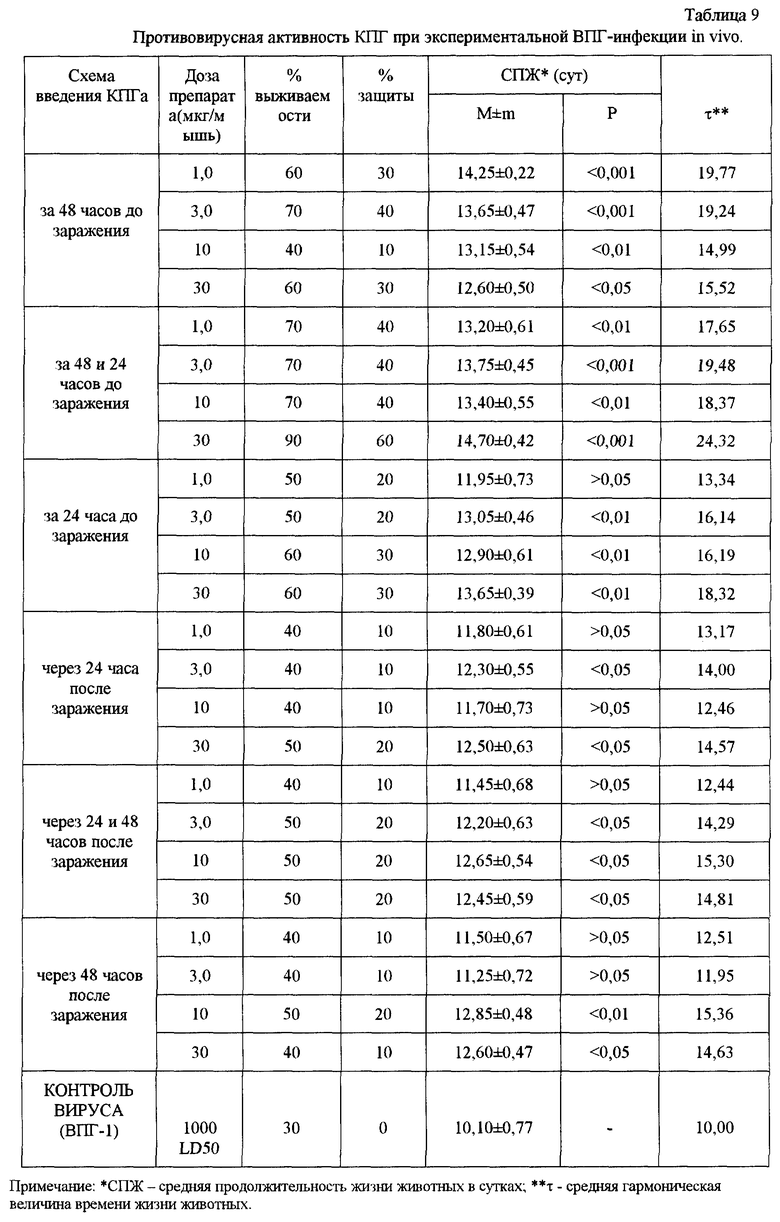
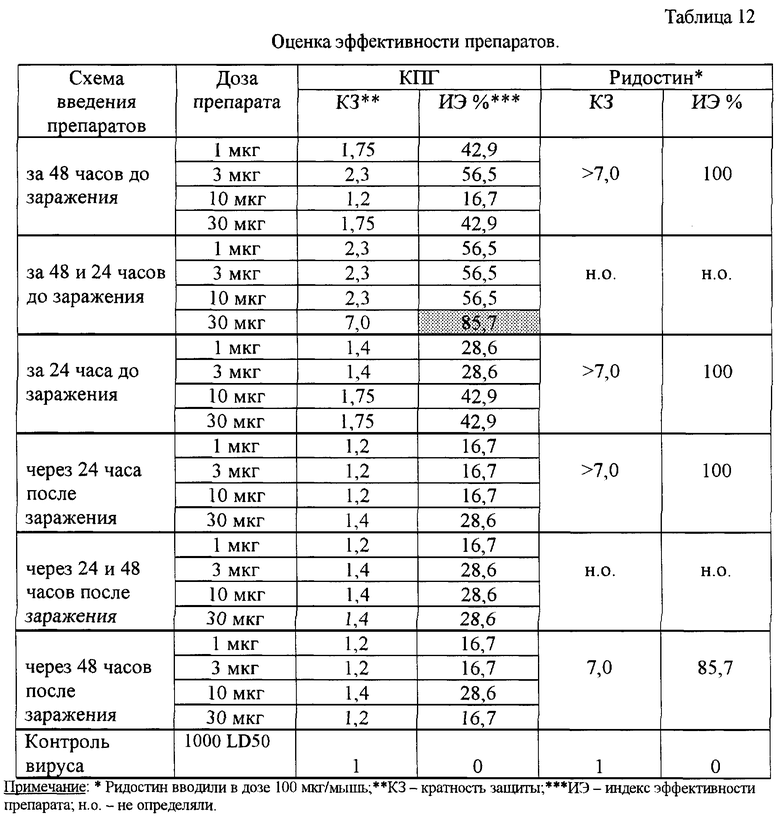
Результаты проведенных экспериментов показали, что препарат КПГ оказывает профилактическое и лечебное действие в модели экспериментальной ВПГ-1 инфекции у мышей. Препарат оказался высоко эффективным (процент защиты 40-60%) при профилактическом введении однократно за 48 часов в дозе 3 мкг/мышь или двукратно за 48 и 24 часа в дозе 3-30 мкг/мышь (табл.9). При этом наблюдали высокие значения СПЖ (р<0,001) и τ по сравнению с контролем. Данные также хорошо согласуются с выявленным подавлением размножения вируса в мозге мышей на 2,5 lg (табл. 10). КЗ и ИЭ также имеют высокие значения - 7,0 и 85,7%, соответственно.
Получено доказательство противовирусной активности КПГ в опытах биологического титрования на клетках VERO. Установлено, что максимальное подавление размножения ВПГ-1 наблюдалось при введении препарата по профилактической схеме, при этом препарат снижал инфекционный титр вируса в 100-1000 раз (то есть на 2-3 lg). При введении препарата по лечебной схеме титр ВПГ-1 снижался на 1,5-2,0 lg.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ВЕЩЕСТВА, ОБЛАДАЮЩЕГО АНТИМИКРОБНОЙ, ПРОТИВОВИРУСНОЙ И ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ, В ЧАСТНОСТИ, В ОТНОШЕНИИ ДЕНДРИТНЫХ КЛЕТОК, ВЕЩЕСТВО, ПОЛУЧЕННОЕ ЭТИМ СПОСОБОМ, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 2013 |
|
RU2563818C2 |
| КОНЪЮГАТ ДЛЯ ИММУНИЗАЦИИ И ВАКЦИНАЦИИ И СПОСОБ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ | 2008 |
|
RU2378015C2 |
| КОМПОЗИЦИЯ ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ ТРАНСГЕНА В ЭУКАРИОТИЧЕСКИХ КЛЕТКАХ И СПОСОБ УВЕЛИЧЕНИЯ ПРОДУКЦИИ ЦЕЛЕВОГО БЕЛКА, КОДИРУЕМОГО ТРАНСГЕНОМ | 2013 |
|
RU2546249C2 |
| ПРОТИВОГРИППОЗНАЯ ВАКЦИНА ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ПРОТИВ ПТИЧЬЕГО ГРИППА А НА ОСНОВЕ ЭКТОДОМЕНА БЕЛКА М2 | 2014 |
|
RU2571944C1 |
| РЕКОМБИНАНТНАЯ ТРЕХВАЛЕНТНАЯ ВАКЦИНА ОТ ГРИППА ЧЕЛОВЕКА | 2012 |
|
RU2485973C1 |
| ПОЛИМЕРНЫЙ ФРАГМЕНТ ПЕПТИДОГЛИКАНА КЛЕТОЧНОЙ СТЕНКИ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ В КАЧЕСТВЕ ИММУНОСТИМУЛЯТОРА | 2006 |
|
RU2412197C2 |
| СПОСОБ ПРОИЗВОДСТВА ФАРМАКОЛОГИЧЕСКИ ПРИЕМЛЕМОЙ СМЕСИ ВЕЩЕСТВ, СОДЕРЖАЩЕЙ НИЗКОМОЛЕКУЛЯРНЫЕ КОМПОНЕНТЫ ПЕПТИДОГЛИКАНА КЛЕТОЧНОЙ СТЕНКИ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ И ОБЛАДАЮЩЕЙ ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2008 |
|
RU2478644C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ЦИТОКИНОВОГО И ИММУНОМОДУЛИРУЮЩЕГО ДЕЙСТВИЯ | 2002 |
|
RU2222345C2 |
| ПРИМЕНЕНИЕ КОМПОЗИЦИИ, СОСТОЯЩЕЙ ИЗ НИЗКОМОЛЕКУЛЯРНЫХ ФРАГМЕНТОВ ПЕПТИДОГЛИКАНА ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ, ДЛЯ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ЗАБОЛЕВАНИЙ ЧЕЛОВЕКА | 2009 |
|
RU2441906C2 |
| ПЕПТИДЫ ПРОИЗВОДНЫЕ ЭЗРИНА И ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ НА ИХ ОСНОВЕ | 2016 |
|
RU2694906C2 |



Изобретение относится к медицине. Проводят экстракцию измельченного растительного сырья, в качестве которого используют растения семейства Dioscoreaceae, Plantaginaceae, Solanaceae, водой. Осуществляют центрифугирование водного экстракта, его концентрирование, осаждение 96%-ным этиловым спиртом в присутствии хлористого натрия. Проводят повторное переосаждение полученного осадка солевым или кислым агентом с последующей обработкой выделенного кислого пептидогликана - сырца раствором щелочи или насыщенным раствором соли щелочного металла. Очистку продукта осуществляют с помощью гельпроникающей хроматографии и сушкой. Вещество, полученное этим способом, представляет собой водорастворимый кислый пептидогликан с молекулярной массой 1200-40000 КД и имеет массовое соотношение между глюкозой и уроновыми кислотами, равное 1: 2-4. Фармацевтическая композиция содержит активное вещество и носитель. Изобретение позволяет получить новое вещество растительного происхождения из доступного сырья, обладающего сильно выраженной иммуностимулирующей, противовирусной и антибактериальной активностью. 3 с. и 5 з.п. ф-лы, 12 табл.
| Хим.-фарм | |||
| журнал, 2000, т.34, № 3, с.51-56 | |||
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ДЕЙСТВИЕМ ПРОТИВ ИНФЕКЦИОННЫХ АГЕНТОВ | 1994 |
|
RU2092177C1 |
| ОУДИ П | |||
| Полный мед | |||
| справочник | |||
| - М.: Дорлинг Киндерсли Лимитед, 2000, с.134-150. | |||

