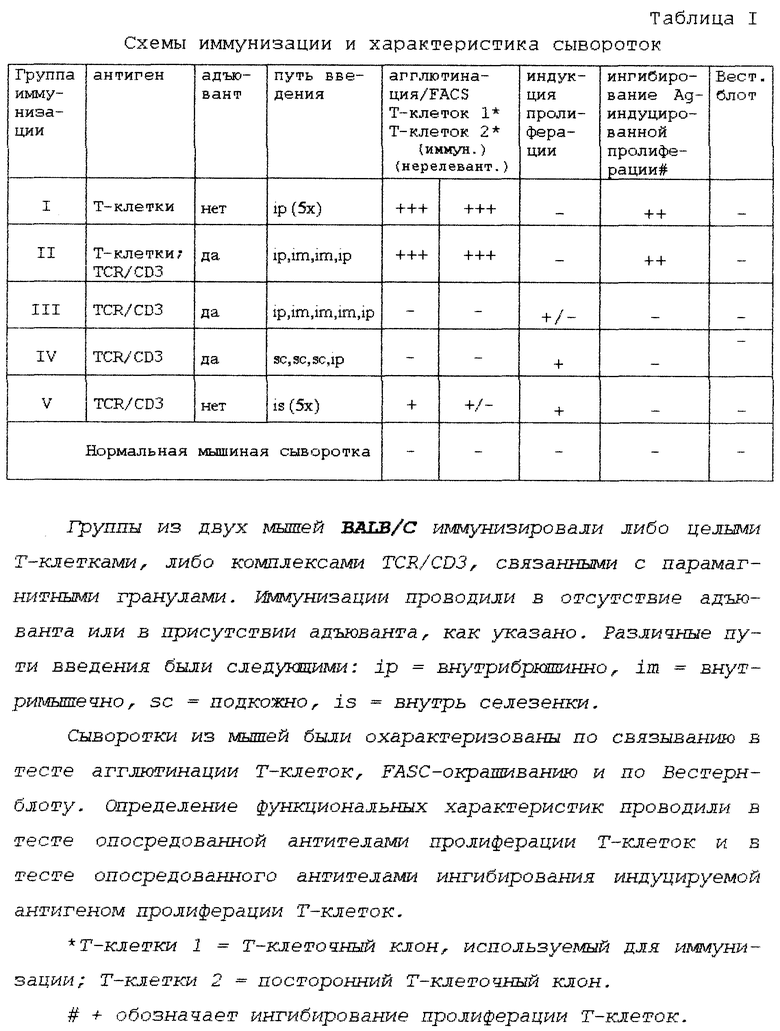
Изобретение относится к способу получения моноклонального антитела против поверхностного антигена, в частности такого поверхностного антигена, который составляет только незначительную часть общего антигена, инъецированного в млекопитающее, и поверхностного антигена, который легко теряет присущую ему in vivo конформацию. Проблема незначительного количества антигена относительно общего количества антигена возникает, например, когда антиген, используемый для инъецирования млекопитающего, происходит из другого вида млекопитающего.
Проблемы, связанные с антигенами, описанными выше, становятся очевидными, когда, например, желательно получить моноклональные антитела против определяющей специфичность части Т-клеточного рецептора (TCR). Такие (моноклональные) антитела известны как антиклонотипические антитела, так как они узнают антигенспецифическую часть (или структуру клонотипа) Т-клеточного рецептора одного конкретного Т-клеточного клона. В то время как о (мышиных) моноклональных антиклонотипических антителах против мышиных Т-клеточных рецепторов сообщалось лишь время от времени, еще даже менее известно о (мышиных) моноклональных антиклонотипических антителах против Т-клеточных рецепторов человека. Т-клетки трудно культивировать, что делает трудным получение и очистку достаточного количества антигена, т.е. белка Т-клеточного рецептора. Это, в свою очередь, делает затруднительным получение иммунного ответа и также делает трудным скрининг. В одном из небольшого числа случаев, где доступным является моноклональное антиклонотипическое антитело против TCR человека, эти проблемы не возникают вследствие того, что этот антиген присутствует на лейкозных клетках Jurkat [1]. Поскольку лейкозные клетки можно культивировать в неограниченных количествах, проблема недостатка антигена не возникала. Таким образом, до сих пор не было способа получения моноклональных антител против редких и/или очень нестабильных антигенов без необходимости скрининга неудобно больших количеств гибридомных клонов.
Целью данного изобретения является обеспечение способа получения моноклонального антитела против поверхностного антигена клеток при неблагоприятных обстоятельствах, описанных выше, причем этот способ очень сильно уменьшает число гибридомных клонов, которые должны быть подвергнуты скринингу, что делает способ согласно данному изобретению более эффективным или даже успешным в тех случаях, когда известные в данной области способы терпят неудачу.
Для этой цели способ данного изобретения включает в себя стадии:
1) инъецирования млекопитающего содержащим клеточный поверхностный антиген материалом, выбранным из группы, состоящей из i) целых клеток и ii) фракции мембран, полученной обработкой целых клеток;
2) выделения клеточной фракции, содержащей В-клетки, из селезенки данного млекопитающего;
3) обогащения клеточной фракции, полученной в стадии 2, В-клетками, специфическими для указанного клеточного поверхностного антигена, путем контактирования этой клеточной фракции со связанным с носителем материалом клеток, родственных указанным целым клеткам, причем связанный с носителем материал клеток не содержит поверхностного антигена клеток, и отделения В-клеток, связанных со связанным с носителем материалом этих клеток, от обогащенной, несвязанной клеточной фракцией, содержащей В-клетки, которая должна использоваться в следующей стадии;
4) лимитирующего разведения обогащенной, содержащей В-клетки клеточной фракции, полученной в предыдущей стадии, с последующей клональной экспансией (размножением);
5) отбора В-клеточного клона и иммортализации отобранного В-клеточного клона при помощи низкомасштабного способа слияния; и
6) отбора и клонирования гибридомы, способной продуцировать антитела, специфически связывающие клеточный поверхностный антиген, с последующим выделением содержащей моноклональные антитела фракции из супернатанта этой гибридомы.
С целью поддержания описания и формулы изобретения легко читаемыми и понятными термин "клетка" используется здесь для указания не только клеток млекопитающих, но также вирусов и, в частности, содержащих мембрану вирусов.
Термин "связанный с носителем материал клеток" включает интактные или целые клетки (и вирусы), мембранные фракции интактных целых клеток (и вирусов) или по существу очищенные поверхностные антигены или их комплексы.
Если нет других указаний, термин "целая клетка" относится к клеткам, имеющим представляющий интерес (целевой) поверхностный антиген. Термин "родственные клетки" относится к клеткам, которые предпочтительно отличаются только тем, что они не содержат представляющего интерес поверхностного антигена или, более конкретно, не содержат иммунологической детерминанты, против которой желательно получить моноклональное антитело.
Неожиданно оказалось возможным обогащение клеточной фракции В-клетками, специфическими для клеточного поверхностного антигена, без использования клеточного поверхностного антигена, путем контактирования выделенной, содержащей В-клетки клеточной фракции с материалом клеток из того же самого вида, из которого был получен клеточный поверхностный антиген, но не имеющим клеточного поверхностного антигена. Другими словами, обогащение достигалось путем контактирования В-клеток с не относящимися к ним клетками или клеточным материалом и удаления связанных В-клеток.
Таким образом, можно получить моноклональное антитело против минорного клеточного поверхностного антигена, даже если этот клеточный поверхностный антиген обладает конформационной нестабильностью.
Европейский патент ЕР 0488470 и ссылка 3, в основе которой лежит этот Европейский патент, описывают способ, в котором I) млекопитающее инъецируют антигеном, II) выделяют фракцию, содержащую В-клетки, III) отбирают В-клетки, специфические для этого антигена, IV) обогащенную фракцию подвергают клональному размножению и после V) отбора В-клеточного клона и иммортализации при помощи низкомасштабного способа слияния VI) гибридому отбирают и клонируют с последующим выделением фракции, содержащей моноклональные антитела. В стадии III В-клетки отбирают связыванием их на пластиковой поверхности, покрытой антигеном, или методом розеткообразования с покрытыми антигеном парамагнитными гранулами. Неспецифические В-клетки удаляют промыванием. Таким образом, кроме других различий, этот способ основан на вполне достаточной доступности и использовании антигена - во время отбора используют 2 мкг/мл для покрытия планшетов антигеном, тогда как данное изобретение решает проблему для случая, в котором антиген доступен в очень ограниченных количествах и даже является очень нечистым.
Предпочтительный вариант характеризуется тем, что млекопитающее, инъецируемое поверхностным антигеном, относится к другому виду, чем вид млекопитающего, из которого происходит поверхностный антиген.
Способ согласно данному изобретению очень удобен для получения моноклональных антител в случае присутствия многих антигенов, способных индуцировать иммунный ответ.
Эта ситуация существует, в частности, когда поверхностный антиген имеет константный сегмент и вариабельный сегмент, причем по меньшей мере часть вариабельного сегмента определяют как определяющую специфичность часть поверхностного антигена.
Согласно предпочтительному варианту в качестве материала, содержащего клеточный поверхностный антиген, используют материал, содержащий молекулу рецептора.
Способ согласно данному изобретению особенно пригоден для получения моноклональных антител против определяющей специфичность части молекулы рецептора. Определяющая специфичность часть молекулы рецептора является только минорной частью молекулы рецептора, которая сама является минорным компонентом всех молекул - и, следовательно, антигенов - на поверхности клетки.
Предпочтительной мишенью согласно данному изобретению является клон Т-клеток, который используют для получения материала, содержащего молекулы рецептора.
Т-клетка может быть использована в виде целой клетки или может быть использована для получения фракции ее мембран. Так, в первом случае термин "получение" может относиться здесь к простому выделению Т-клетки или ресуспендированию в отличающейся среде.
Согласно предпочтительным вариантам мембранную фракцию в стадии 1 получают механической обработкой целых клеток и материал, содержащий клеточный поверхностный антиген, инъецируют в млекопитающее в отсутствие адъюванта.
Обе меры помогают предотвратить потерю существующей in vivo конформации поверхностного антигена.
Кроме того, обогащенную, содержащую В-клетки клеточную фракцию предпочтительно обогащают дополнительно (стадия 3) путем контактирования этой клеточной фракции с материалом, содержащим клеточный поверхностный антиген, связанный с носителем, выбранным из группы j) целых клеток, jj) мембранной фракции, полученной из целых клеток, и jjj) по существу очищенного клеточного поверхностного антигена, и затем отделения В-клеток, не связанных с материалом, связанным с носителем, от В-клеток, связанных с материалом, связанным с носителем, содержащим дополнительно обогащенную клеточную фракцию.
Это дополнительное обогащение может быть названо способом положительного отбора, со специальным отбором специфических В-клеток. Таким образом, хотя едва ли доступен какой-либо антиген, дополнительное обогащение может быть достигнуто.
Согласно предпочтительному варианту в качестве носителя предпочтительно использовать парамагнитные гранулы.
Использование парамагнитных гранул во время обогащения облегчает разделение нежелательных и желательных В-клеток.
Незначительное количество антигена представляет также проблему во время скрининга. Согласно предпочтительному варианту отбор по меньшей мере в одной из стадий 5 и 6 проводят с применением реакции агглютинации, в которой супернатант клона В-клеток приводят в контакт с носителем, покрытым антителами, способными связывать антитела вида инъецированного млекопитающего, используемого в стадии 1, и целыми клетками, несущими клеточный поверхностный антиген, и агглютинацию детектируют.
Простое смешивание супернатанта культуры В-клеток, целых клеток и носителя, покрытого антителами, способными связывать антитела вида млекопитающего, используемого в стадии 1, делает возможным очень чувствительное и быстрое детектирование пригодных клонов, причем промывание не является необходимым.
В качестве контроля можно выгодно использовать целые клетки, родственные указанным целым клеткам, но не имеющие поверхностного антигена.
Это позволяет отклонить ложноположительные клоны и экономит время, поскольку позволяет избежать сверхтекучих низкомасштабных слияний.
Согласно предпочтительному варианту данного способа изобретения, отобранный клон В-клеток стадии 5 смешивают с миеломными клетками и подвергают мини-электрослиянию.
Мини-электрослияние делает возможным эффективное слияние очень малых количеств (например, сотен) клеток.
Данное изобретение относится также к фармацевтической композиции, содержащей моноклональное антитело, полученное согласно изобретению, смешанное с подходящим наполнителем.
Кроме того, данное изобретение относится к моноклональному антителу, реактивному с клонотипической структурой Т-клеточного рецептора.
Такое моноклональное антитело можно использовать для диагностических целей, а также для приготовления фармацевтической композиции.
Более конкретно, Т-клеточный рецептор представляет собой Т-клеточный рецептор, ассоциированный с аутоиммунным заболеванием и, в частности, ревматоидным артритом.
Предпочтительно моноклональное антитело является реактивным с Т-клеточным рецептором НС gp-39-реактивного клона Т-клеток и, в частности, с клоном Т-клеток Н.243 (ЕСАСС accession 96103122).
Характерными примерами подходящих моноклональных антител являются моноклональные антитела, продуцируемые гибридомой, выбранной из группы, состоящей из TCR 69 (ЕСАСС accession 96103118), TCR 70 (ЕСАСС accession 96103119), TCR 72 (ЕСАСС accession 96103120) и TCR 83 (ЕСАСС accession 96103121).
Как указано выше, данное изобретение относится также к фармацевтической композиции, содержащей моноклональное антитело согласно данному изобретению, смешанное с приемлемым наполнителем, пригодной для лечения ревматоидного артрита.
Наконец, данное изобретение относится к диагностическому применению моноклонального антитела, выбранного из группы, состоящей из моноклонального антитела, полученного при помощи способа согласно данному изобретению, и моноклонального антитела согласно данному изобретению. Вариантом данного изобретения является также диагностический реагент, содержащий это антитело.
Теперь изобретение будет объяснено более детально со ссылкой на следующий далее пример, показывающий наилучший способ проведения данного изобретения в применении к получению мышиного моноклонального антитела, специфического для клонотипа Т-клеточного рецептора человека.
Подписи к чертежам
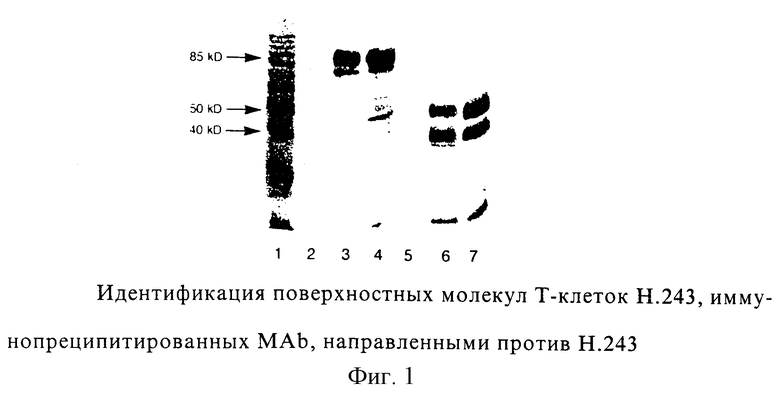
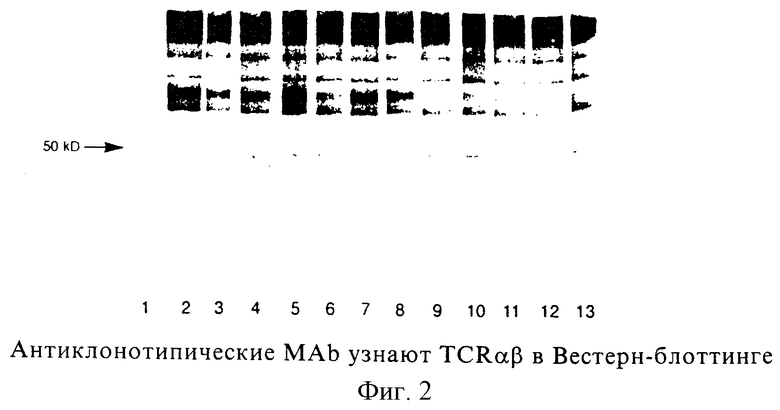
Фиг. 1: Идентификация Н.243 Т-клеточных поверхностных молекул, иммунопреципитированных моноклональными антителами (MAb), направленными против Н. 243. Электрофорез в ДСН-ПААГ проводили при невосстанавливающих условиях (дорожки 2, 3 и 4) или при восстанавливающих условиях (дорожки 5, 6 и 7) в 10% геле. Дорожка 1: 10 кД-ступенчатый маркер; дорожки 2 и 5: контрольные МАb; дорожки 3 и 6: TCR 83; дорожки 4 и 7: анти-CD3, ОКТ3.
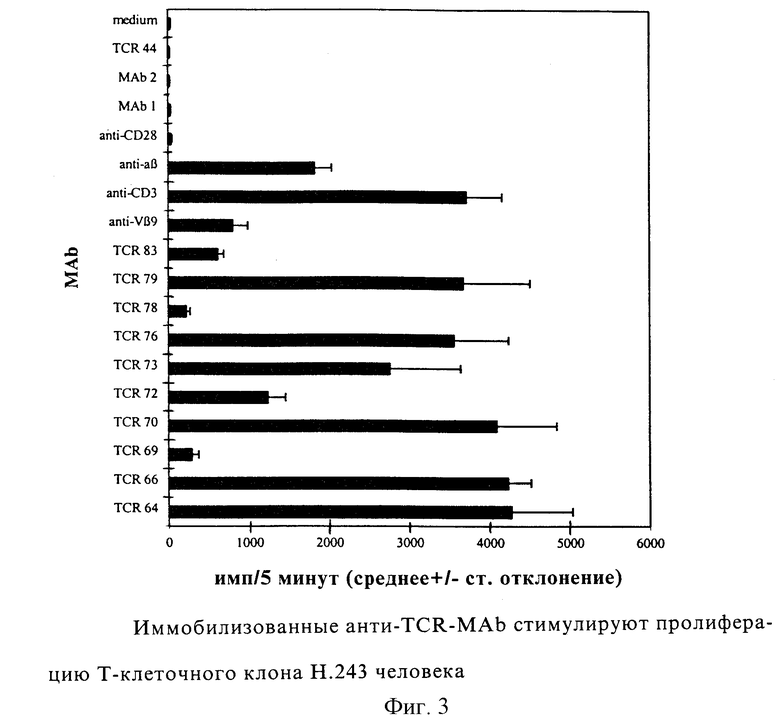
Фиг. 2: Антиклонотипические МАb узнают ТСВαβ в Вестерн-блоттинге. Иммунопреципитированные комплексы TCR/CD3 Н.243 разделяли на 10% геле ДСН-ПААГ при невосстанавливающих условиях и затем переносили на мембраны PVDF. Мембраны инкубировали с МАb и, наконец, связанные MAb детектировали с использованием конъюгированного со щелочной фосфатазой второго козьего антитела против мышиного Ig. Дорожка 1: 10 кД-ступенчатый маркер, дорожка 2: TCR 64, дорожка 3: TCR 66, дорожка 4: TCR 69, дорожка 5: TCR 70, дорожка 6: TCR 72, дорожка 7: TCR 73, дорожка 8: TCR 76, дорожка 9: TCR 78, дорожка 10: TCR 79, дорожка 11: TCR 83, дорожка 12: контрольные MАb, дорожка 13: среда в качестве контроля.
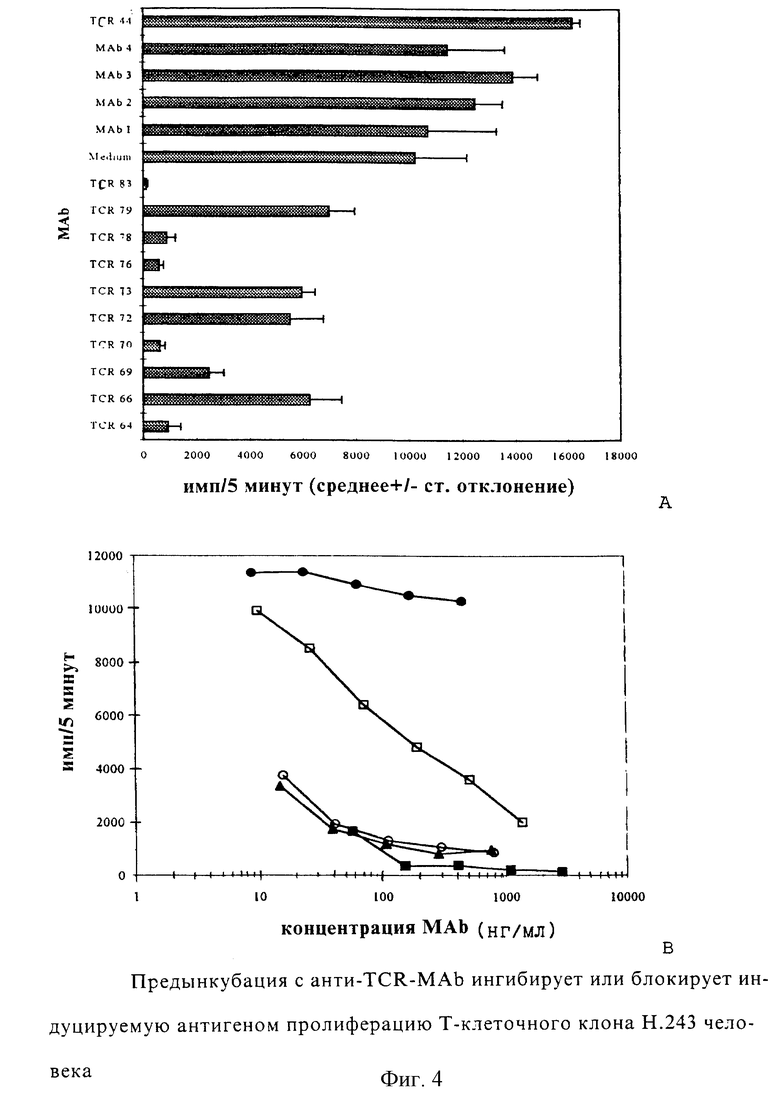
Фиг.3: Иммобилизованные моноклональные антитела против TCR (анти-TCR MАb) стимулируют пролиферацию Т-клеточного клона Н.243 человека. Пролиферацию, индуцированную МАb, направленными против Н.243 (TCR 64 - TCR 83), сравнивали с контрольными MAb, как указано, и с TCR 44, который представляет собой антиклонотипические MАb, направленные против другого TCR. Каждая величина представляет собой среднее число импульсов за 5 минут для четырех культур +/- среднее квадратичное отклонение.
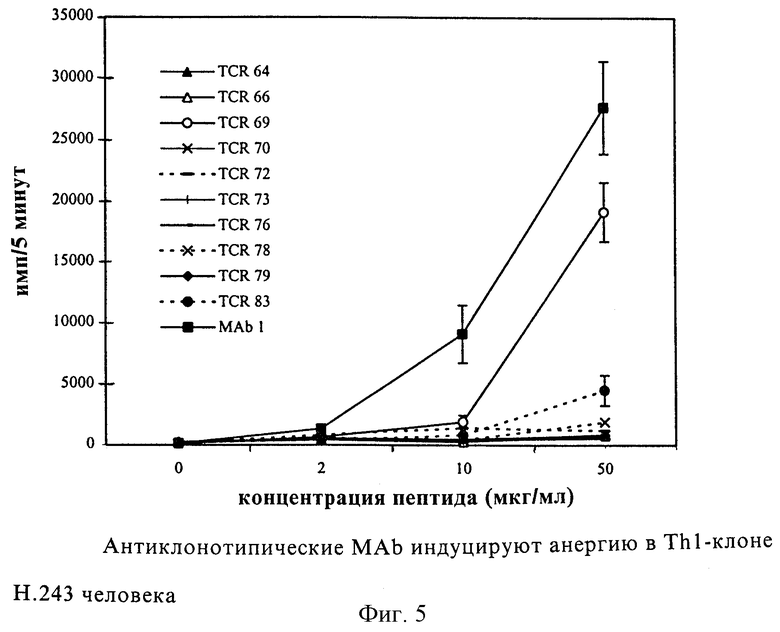
Фиг. 4: Предынкубация с анти-TCR MAb ингибирует или блокирует индуцируемую антигеном пролиферацию Т-клеточного клона Н.243 человека. а) Ингибирование MАb, направленными против Н.243 (TCR 64 - TCR 83), сравнивали с контрольными MAb, как указано, и с TCR 44, который представляет собой антиклонотипические МАb, направленные против другого TCR. Каждая величина представляет собой среднее число импульсов за 5 минут для четырех культур +/- среднее квадратичное отклонение. b) Кривая дозовой зависимости сильного ингибирования MАb; TCR 64 (светлые кружки), TCR 70 (темные треугольники), TCR 78 (светлые квадраты), TCR 83 (темные квадраты) и контрольные MАb 1 (темные кружки). Каждая величина обозначает среднее число импульсов за 5 минут для трех культур.
Фиг. 5: Антиклонотипические MАb индуцируют анергию (толерантность) в TH1-клоне Н.243 человека.
Иммобилизованные антиклонотипические MАb или контрольные MАb 1 инкубировали в течение ночи с Т-клетками Н7243. После удаления клеток из контакта с анергическим стимулом клеткам предоставляли полный стимул увеличивающихся концентраций пептида и DRB1*0401 - совместимых АПК (антигенпредставляющих клеток). Пролиферацию оценивали по включению 3H-тимидина. Каждая величина представляет собой среднее число импульсов за 5 минут для трех культур +/- среднее квадратичное отклонение.
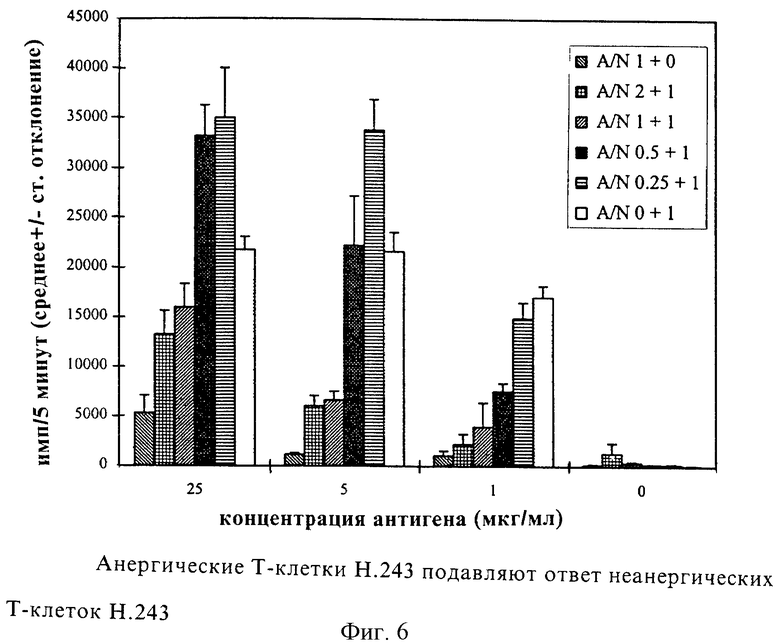
Фиг. 6: Анергические Т-клетки Н.243 подавляют ответ неанергических Т-клеток Н.243.
Иммобилизованные TCR 76 или контрольные MAb 1 инкубировали в течение ночи с Т-клетками Н.243. Затем обе популяции Т-клеток использовали в тесте пролиферации с увеличивающимися концентрациями пептида (или фитогемагглютинина (РHА)) и АПК с использованием 2•104 Т-клеток на лунку. Кроме того, различные количества анергических клеток, полученных инкубацией с TCR 76, смешивали с 2•104 неанергических клеток, полученных инкубацией с MАb 1, и проводили тест пролиферации. А/N: отношение анергических клеток к неанергическим клеткам; 1= 2•104 клеток. Пролиферацию оценивали по включению 3H-тимидина. Каждая величина представляет собой среднее число импульсов за 5 минут для трех культур + среднее квадратичное отклонение.
ПРИМЕРЫ
Материалы и способы
Реагенты
Культуральная среда: DMEM/HAM's F12 (Gibco cat. no. 32500), дополненная 2500 мг/л бикарбоната натрия, 2,3 мг/л 2-меркаптоэтанола, 55 мг/л пирувата натрия, 1,22 мг/л этаноламина, 360 мг/л L-глутамина, 4,5•104 мг/л селенита натрия, 62,5 мг/л натрий-пенициллина и 62,5 мг/л стрептомицин-сульфата. В экспериментах по слиянию к среде добавляли еще 13,61 мг/л гипоксантина и 3,83 мг/л тимидина. Эта среда названа DMEM/HAM's F12/HT. Отбор гибридом проводили в DMEM/HAM's F12/HT, дополненной 1% (об./об.) содержащим ИЛ-6 супернатантом линии Т24 (Т24СМ) клеток карциномы мочевого пузыря человека и 0,4 мкМ аминоптерином.
Среда для слияния: 280 мМ инозит, 0,1 мМ ацетат кальция, 0,5 мМ ацетат магния и 1 мМ гистидин; удельное сопротивление: 1,11•104 Ом.см. Ингредиенты растворяли в MilliQ-воде и затем электропроводность доводили до 90 мкС/см MilliQ-водой или раствором, содержащим 1 мМ ацетат кальция и 5 мМ ацетат магния.
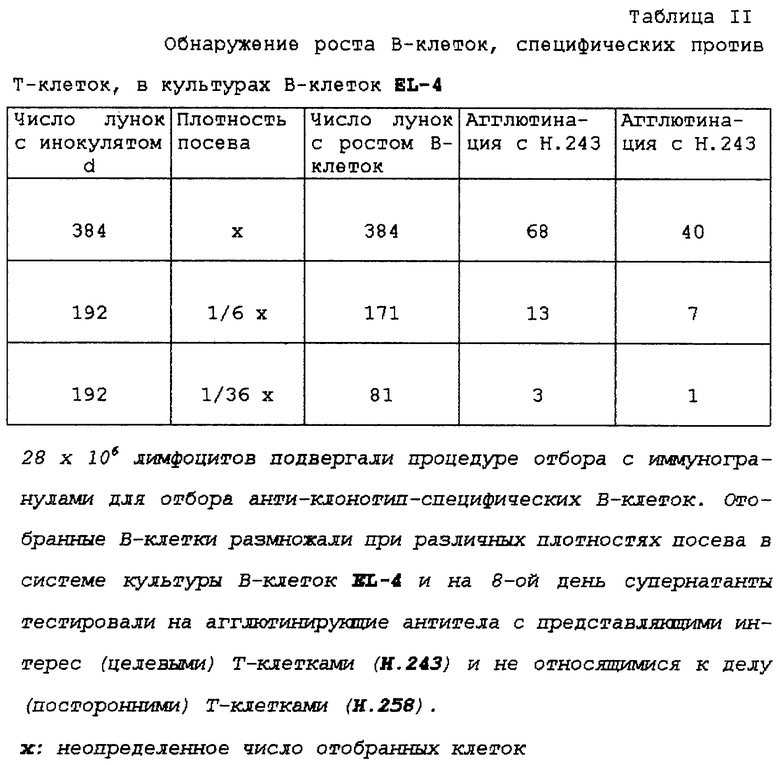
Культуры клеток
Т-клеточный клон H. 243 получали от больного, страдающего ревматоидным артритом. Этот Th1-подобный Т-клеточный клон узнает эпитоп RSFTLASSETGVG (SEQ ID 1) из gр-39 хряща человека (НС gp-39) в контексте DRB1*0401 (принадлежит к МНС класса II). TCR этого клона характеризуется как Vα8- и Vβ9-положительный. Клетки культивировали общепринятым способом в DMEM/HAM's F12 с добавлением 10% фетальной телячьей сыворотки, 20 Е/мл ИЛ-2, 5 Е/мл ИЛ-4 и периодически повторно стимулировали антигеном и гистосовместимыми антигенпредставляющими клетками (АПК) или фитогемагглютинином (РНА) и питающими клетками. Для тестов пролиферации фетальную телячью сыворотку (PCS) заменяли нормальной человеческой сывороткой (NHS).
Получение лизата Т-клеток и загрузка гранул комплексами TCR/CD3
Спустя 10-14 дней после повторной стимуляции Т-клетки промывали PBS (ЗФР) и солюбилизировали инкубированием их (30 минут, 0oС) при концентрации 108 клеток/мл в буфере для экстракции согласно Oettgen et al. [2] (10 мМ триэтанол-амин, 150 мМ NaCl, 1 мМ Na2-ЭДТА, 1% дигитонин, 10 мкг/мл лейпептин, 10 мкг/мл апротинин, 1 мкг/мл пепстатин, 1 мМ Pefabloc® AEBSF и 1,8 мг/мл йодацетамин рН 7,8). Мягкий детергент дигитонин позволяет экстрагировать интактные комплексы TCR/CD3. Ядерные и клеточные остатки удаляли центрифугированием при 16000 g в течение 15 минут при 4oС и супернатант хранили в виде аликвот при -20oС до использования для отбора В-клеток или для иммунопреципитации.
Для отбора специфических В-клеток парамагнитные гранулы нагружали комплексами TCR/CD3.
Вкратце, 100 мкл парамагнитных гранул, связанных с овечьими антителами против мышиного Ig (SAM-гранулы; Dynabead® 110,02), инкубировали (в течение ночи, 4oС) с 22 мкг анти-CD3, ОКТ3, в PBS (ЗФР) с 1% БСА. После промывания гранул ЗФР/БСА добавляли 1,4•108 клеточных эквивалентов Т-клеточного лизата и равный объем (1,4 мл) ЗФР/БСА с 1% нормальной мышиной сывороткой. Последнюю добавляли для предотвращения связывания В-клеток со свободными SAM-связывающими сайтами на гранулах. Суспензию гранул инкубировали в течение 2-4 часов при 4oС и промывали ЗФР/БСА перед использованием.
Иммунизация
Шестинедельных самок мышей BALB/C иммунизировали с интервалами 3-7 недель либо 5•106 целыми Т-клетками (изобретение), либо парамагнитными гранулами, нагруженными комплексами TCR/CD3 5•107 клеточных эквивалентов (контрольный эксперимент). Использовали различные пути введения, как указано в Таблице I. В случае использования адъюванта (контрольный эксперимент) первую инъекцию выполняли с полным адъювантом Фрейнда, последующие инъекции - с неполным адъювантом Фрейнда и конечную повторную инъекцию - без адъюванта.
Генерирование антиклонотипических MАb
Шестинедельных самок мышей BALB/C инъецировали внутрибрюшинно 4 раза с интервалами 3-7 недель, каждый раз 5•106 покоящимися Т-клетками в ЗФР. Спустя 5 дней после последней инъекции мышей умерщвляли и получали истощенные по эритроцитам и моноцитам популяции клеток селезенки. Это приводило к получению клеточной фракции, обогащенной В-клетками, но не приводило к обогащению В-клеток, специфических в отношении указанного клеточного поверхностного антигена, по сравнению с неспецифическими В-клетками в соответствии со стадией 2 данного изобретения.
Для предварительной очистки этих популяций клеток селезенки с целью получения В-клеток, реактивных с CD3 или константным районом ТСНαβ человека, приблизительно 3•107 клеток инкубировали дважды с 20 мкл SAM-гранул, нагруженных комплексами TCR/CD3 не относящегося к делу (постороннего) Т-клеточного клона (Vα3,Vβ14-положительного Т-клеточного клона, SCRO.08A, но любые другие не относящиеся к делу Т-клетки также можно использовать). Затем В-клетки, специфические для вариабельного района TCR, отбирали путем инкубирования полученной клеточной суспензии с гранулами SAM, нагруженными комплексами TCR/CD3 представляющего интерес Т-клеточного клона (Н.243). Каждую инкубацию проводили в течение 90 минут при комнатной температуре в DMEM/HAM's F12, дополненной 10% фетальной телячьей сывороткой (Hyclone®) и 0,2% нормальной мышиной сывороткой. Во время этих инкубаций суспензию осторожно гомогенизировали каждые 5 минут. После последней инкубации гранулы осторожно промывали пять раз DMEM/HAM's F12, 10% телячьей сывороткой и ресуспендировали в той же самой среде.
Гибридомы, продуцирующие моноклональные антитела, получали из этих отобранных В-клеток при помощи клонального размножения и мини-электрослияния. Вкратце, отобранные В-клетки смешивали с Т-клеточным супернатантом (ТСН) и 50000 облученных (2500 рад) EL-4 В5-клеток при конечном объеме 200 мкл DMEM/HAM's F12, дополненной 10% фетальной телячьей сывороткой (FCS), в 96-луночных плоскодонных планшетах для культуры ткани. На 8-й день супернатанты испытывали в одноступенчатом тесте агглютинации Т-клеток, как описано ниже, с использованием либо целевого Т-клеточного клона, либо постороннего Т-клеточного клона. Культуры В-клеток, продуцирующие дискриминирующие (различающие) MАb, подвергали процедуре мини-электрослияния. Содержимое этих В-клеточных культур смешивали с 106 NS-1 миеломных клеток и освобождали от сыворотки промыванием DMEM/HAM's F12/HT. Затем эти клетки обрабатывали раствором проназы в течение 3 минут и затем промывали средой для слияния. Электрослияние проводили в камере для слияния на 50 мкл при помощи переменного электрического поля 30 с, 2 МГц, 400 В/см с последующим плоским импульсом поля 10 мкс, 3 кВ/см и опять переменным электрическим полем 30 с, 2 МГц, 400 В/см. Наконец, содержимое камеры для слияния переносили в 20 мл селективной среды и высевали в 96-луночный микротитрационный планшет. На 8-й день после слияния культуры испытывали на рост гибридомы и снова подвергали скринингу в одноступенчатом тесте агглютинации Т-клеток.
Тест агглютинации Т-клеток
Скрининг гибридомных культур на антитела против Т-клеток и определение перекрестной реактивности MАb с другими Т-клетками проводили в одноступенчатом тесте агглютинации. С использованием микротитрационных планшетов с половиной площади 50 мкл гибридомного супернатанта смешивали с 20 мкл суспензии гранул/клеток, содержащей 2•105 парамагнитных гранул с ковалентно связанными овечьими антителами против мышиного Ig и 5•104 целевых Т-клеток в DMEM/HAM's F12. После 2-3 часов инкубации при 37oС агглютинацию оценивали под микроскопом, наблюдением агрегатов клеток и гранул.
Иммунопреципитация и Вестерн-блоттинг
Спустя 10-14 дней после рестимуляции Т-клетки промывали и клеточные поверхностные белки метили биотином. Вкратце, Т-клетки инкубировали при концентрации 5•106 клеток/мл ImmunoPure® sulfo-NHS-биотином в ЗФР в течение 30 минут при комнатной температуре. Затем клетки опять промывали и солюбилизировали, как описано выше. Перед использованием в эксперименте иммунопреципитации лизат клеток предочищали один раз инкубированием с парамагнитными SAM-гранулами. Иммунопреципитацию проводили инкубированием 107 клеточных эквивалентов предочищенного лизата в течение 3-4 часов с 15 мкл парамагнитных SAM-гранул, предварительно нагруженных MАb (1 мл супернатанта гибридомы). Затем иммунопреципитаты промывали трижды ЗФР, кипятили в буфере для проб и подвергали электрофорезу в ДСН-ПААГ на 10% геле либо при восстанавливающих условиях, либо при невосстанавливающих условиях по способу Laemmli [4] с применением системы Mini-protean II (Biorad). Иммуноблоттинг выполняли согласно Towbin et al. [5] с применением мембран PVDF. Сайты неспецифического связывания блокировали инкубированием мембран с 5% сепарированным молоком в ЗФР с 0,5% Твином 20. После промывания блоты зондировали конъюгатом стрептавидин - щелочная фосфатаза в ЗФР, 0,5% Твине 20, 1% БСА, 1% нормальной козьей сыворотке в течение 1 часа при комнатной температуре. Наконец, блоты проявляли при помощи BCIP и NВТ в качестве хромогенного субстрата. Для исследований Вестерн-блотов парамагнитные SAM-гранулы нагружали комплексами TCR/CD3, как описано выше, с применением ОКТ3 в качестве иммунопреципитирующего антитела и лизата нормальных Т-клеток. После электрофореза в ДСН-ПААГ на 10% ПАА-геле при восстанавливающих или невосстанавливающих условиях блоты инкубировали с 2,5 мл супернатанта гибридомы. Связанные антитела детектировали при помощи конъюгированных со щелочной фосфатазой козьих антител против мышиного Ig и BCIP/NBT, как описано выше.
Опосредованная антителами пролиферация Т-клеток
Плоскодонные микротитрационные лунки покрывали (в течение ночи, при комнатной температуре) 100 мкл овечьих антител против мышиного Ig при 40 мкг/мл в ЗФР. Лунки промывали один раз и затем инкубировали со 100 мкл MAb при концентрации 2 мкг/мл в ЗФР в течение 2 часов. Избыток свободных MAb удаляли промыванием и добавляли 2,5•104 покоящихся Т-клеток в 200 мкл DMEM/HAM's F12, дополненной 10% NHS. После двух дней инкубации при 37oС клетки импульсно метили 0,5 мкКи [3Н]-тимидином и инкубировали еще 16-18 часов. Наконец, клетки собирали на фильтры из стекловолокна и включение [3H] -тимидина измеряли счетом бета-излучения на Matrix 96TM (Packard). Каждую переменную определяли в четырех повторностях.
Опосредованное антителами ингибирование индуцированной антигеном пролиферации
Плоскодонные микротитрационные лунки инокулировали 20 мкл супернатанта гибридомы, 2•104 покоящимися Т-клетками и 1•105 гистосовместимыми MNC в 150 мкл DMEM/HAM's F12, дополненной 10% NHS. После 3 часов инкубации при 37oС к культурам добавляли 50 мкл пептида RSFTLASSETGVG (16 мкг/мл). Культуры инкубировали еще в течение двух дней при 37oС и включение [3Н]-тимидина определяли, как описано выше. Каждую переменную определяли в четырех повторностях.
Индукция анергии при помощи клонотипспецифических MАb
24-луночные культуральные планшеты покрывали (в течение ночи, при комнатной температуре) 1 мл овечьих антител против мышиного Ig при 40 мкг/мл в ЗФР. Лунки промывали один раз и затем инкубировали с 1 мл MAb при концентрации 2 мкг/мл в ЗФР в течение 2 часов. Избыток свободных MAb удаляли промыванием и 2•106 промытых, покоящихся Т-клеток добавляли в 2 мл DMEM/HAM's F12, 10% NHS. Где указано, при времени инициирования культуры добавляли циклоспорин A (Sandoz, Bazel Switzerland), rIL-2, циклогексимид (Sigma, St. Louis, МО, USA) или HLA-DRB1*0401-совместимые АПК (облученные 3000 рад). После инкубации в течение ночи планшеты охлаждали на льду и Т-клетки ресуспендировали пипетированием. Клетки промывали один раз полной культуральной средой и использовали для теста пролиферации Т-клеток, анализа цитокинов и FACS-анализа, как описано ниже.
Антиген-специфический пролиферативный ответ Т-клеток оценивали в культурах в плоскодонных лунках, содержащих 2•104 Т-клеток, 1•105 НLА-DRВ1*0401-совместимых облученных (3000 рад) РВМС и вариабельные концентрации антигена, в 200 мкл DMEM/HAM's F12, 10% NHS. После 2 дней инкубации при 37oС клетки импульсно метили 0,5 мкКи [3H] -тимидина и инкубировали еще 16-18 часов. Наконец, клетки собирали на фильтры из стекловолокна и включение [3H]-тимидина измеряли по газовой сцинтилляции на Matrix 96 (Packard, Meriden, CT, USA). Каждую переменную определяли в трех повторностях.
Результаты
Иммунизации
Иммунизации с использованием адъюванта (контрольный эксперимент) не давали сывороток, содержащих антитела, способные дискриминировать между Н.243 и не относящимися к делу Т-клетками (Таблица I; группа иммунизации II, III, IV). Титры сывороток группы V были низкими (1:100 в тесте агглютинации), тогда как титры сывороток группы I и II были гораздо более высокими (1: 1200).
Генерирование антиклонотипических MАb
При использовании клеток селезенки из группы I инкубация парамагнитных гранул, нагруженных комплексами TCR/CD3 из нецелевого Т-клеточного клона, здесь H. 243, приводила ко многим образующим розетки клеткам, что указывало на успешное удаление многих CD3- и TCRαβ-реактивных В-клеток. Когда не образующие розеток клетки подвергали второй предочистке с нецелевыми комплексами TCR/CD3 из того же самого Т-клеточного клона, едва ли можно было видеть какие-либо розеткообразующие клетки.
Микроскопическое исследование после инкубации с парамагнитными гранулами с представляющим интерес Т-клеточным клоном не обнаружило видимого розеткообразования. При ожидаемой частоте встречаемости специфических В-клеток это не является удивительным. После лимитирующего разведения и клональной экспансии (размножения) было обнаружено, что 36 В-клеточных супернатантов агглютинировали Н. 243, но не посторонний Т-клеточный клон Н.258 (Таблица II). Некоторые В-клеточные супернатанты были способны агглютинировать оба клона, что указывало на то, что они избежали процедур предочистки.
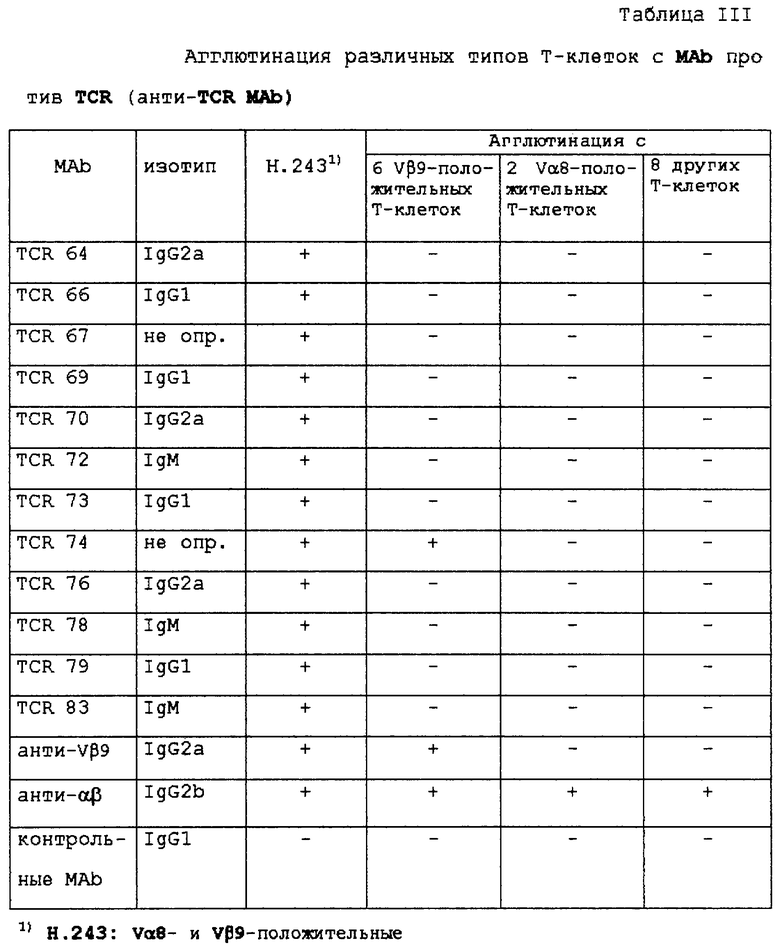
Далее, из Таблицы II можно сделать вывод, что основная часть специфических В-клеточных культур не была клональной, так как выросли также В-клетки, которые не реагируют с какими-либо Т-клетками. Однако это не было проблемой, поскольку после мини-электрослияния можно было отобрать эти неспецифические В-клетки. 18 специфических В-клеточных культур подвергали мини-электрослиянию. Некоторые слияния не приводили к образованию Т-клеточных специфических гибридом, а некоторые другие нельзя было клонировать в стабильные клеточные линии. В конце концов, стабильные гибридомы получили в 11 из 18 мини-электрослияний. Эти 11 MАb характеризовали далее. Изотипы этих МАЬ показаны в Таблице III.
Специфичность MAb
Для дальнейшего исследования специфичности MАb выполняли тесты иммунопреципитации и агглютинации Т-клеток, а также Вестерн-блоттинг.
Электрофорез в ДСН-ПААГ при невосстанавливающих и восстанавливающих условиях показал, что все MAb были способны иммунопреципитировать основную полосу приблизительно 85 кД и минорную полосу 21 кД из дигитонинового лизата Т-клеточного клона Н. 243. Это согласуется с молекулярной массой TCR αβ-комплекса и цепей CD3 ε/δ соответственно. При восстанавливающих условиях полоса 85 кД диссоциирует на две полосы 40 кД и 50 кД, что согласуется с молекулярной массой α- и β-цепей TCR соответственно. Иммуноблот одного из анти-Н. 243-специфического MAb показан в фиг.1, вместе с ОКТ3 в качестве положительного контроля и посторонним MAb в качестве отрицательного контроля.
Таблица III показывает перекрестную реактивность МАb с Vβ9-положительными Т-клеточными клонами, двумя Vα8-положительными Т-клеточными клонами и 8 другими Т-клеточными клонами с отличающимися Vα и Vβ. Антитело TCR 74 должно рассматриваться как Vβ9-специфическое, так как оно агглютинирует все Vβ9-положительные Т-клетки, но не другие Т-клетки. Другие TCR-антитела реагируют только с Н.243, так что очень вероятно, что эти антитела направлены против антигенсвязывающей части Н.243, так как исключается реакция с Vα8, Vβ9, константным районом TCRαβ, CD3 и другими обычными молекулами поверхности Т-клеток. Фиг. 2 показывает, что антитела (MАb) из гибридомных клонов TCR 64, TCR 66, TCR 69, TCR 70, TCR 73 и TCR 76 способны окрашивать TCR на невосстанавливающем Вестерн-блоте (полоса 85 кД). Неспецифические тяжелые полосы на верхней части чертежа происходят из иммунопреципитирующих антител SAM и ОКТ3. При восстанавливающих условиях в Вестерн-блотах не получали окрашивания с антиклонотипическими МАb (результаты не показаны), что предполагает, что эти МАb узнают конформационный эпитоп, образованный интактным комплексом TCRαβ.
Индуцируемая MАb пролиферация Т-клеток
Для исследования, способны ли эти MАb индуцировать пролиферацию Т-клеток Н. 243, MАb иммобилизовали на плоскодонных микротитрационных планшетах и инкубировали с Т-клетками. Хотя между этими МАb существуют большие различия, фиг. 2 показывает, что все они могут индуцировать пролиферацию клеток Н.243. Пролиферацию, сходную с анти-CD3 (ОКТ3), получали с TCR 64, TCR 66, TCR 70, TCR 76 и TCR 79. Пролиферации не получали с MАb TCR 44, которые представляют собой антиклонотипические MАb для другого TCR.
Опосредованное MАb ингибирование индуцируемой антигеном пролиферации Т-клеток
Далее исследовали, может ли данная панель (набор) Mаb ингибировать продуцируемую антигеном пролиферацию т-клеток Н.243. В этом эксперименте MАb предынкубировали с Т-клетками и АПК и три часа спустя импульсно метили различными концентрациями пептида. Фиг. 4 показывает, что TCR 64, TCR 70, TCR 76, TCR 78 и TCR 83 сильно ингибируют индуцируемую антигеном пролиферацию. До 98% ингибирования наблюдали с TCR 83. Кривые дозовой зависимости показывают, что максимальное ингибирование можно получить при такой низкой концентрации, как 100 нг/мл TCR 64, TCR 70 и TCR 83. TCR 78 был примерно в 10 раз менее сильным. Распределение ингибирования не зависело от того, добавляли ли субоптимальные или насыщающие концентрации пептида (результаты не приведены). Антиклонотипические MАb, направленные против другого TCR (TCR 44) не обнаружили какого-либо ингибирования.
Антиклонотипические MАb индуцируют отсутствие отвечаемости Т-клеточного клона Н.245 человека на последующую стимуляцию антигеном и АПК
Как известно, занятие Т-клеточного рецептора в отсутствие костимуляции индуцирует анергию. Ранее мы нашли, что иммобилизованные антиклонотипические MАb к Н.243 могут функционально запускать TCR этого клона. Поэтому представляет интерес исследовать, могут ли те же самые антитела индуцировать анергию. Для этой цели 10 различных антиклонотипических MАb иммобилизовали на 24-луночных планшетах и инкубировали в течение ночи с Т-клетками Н.243 для получения анергического стимула. Затем Т-клетки удаляли из планшетов и определяли, могут ли они отвечать на увеличивающиеся концентрации НС gp-39262-277 (SEQ ID 3), предоставляемого облученными, DRВ1*0401-совместимыми АПК. Фиг. 5 показывает, что этот ответ был полностью уничтожен 8 из 10 MАb, тогда как клетки H.243, инкубированные с контрольными MАb 1, все еще хорошо отвечали на пептид, предоставляемый АПК. Ответ Т-клеток, инкубированных с TCR 69 и TCR 63, был значительно снижен, но не полностью блокирован при более высоких концентрациях пептида.
Анергические клетки подавляют ответ неанергических клеток
Исследовали также, способны ли анергические клетки Н.243 подавлять ответ неанергических клеток Н.243. Анергию индуцировали TCR 76, а неанергические клетки получали инкубацией с контрольными MАb 1. Затем проводили тесты пролиферации с пептидом НС gp-39261-275 (SEQ ID 2) с использованием различных популяций Т-клеток, а именно 2•104 анергических Т-клеток на лунку, 2•104 неанергических Т-клеток на лунку или смесей уменьшающихся концентраций анергических Т-клеток (4•104, 2•104, 1•104 и 0,5•104) и 2•104 неанергических Т-клеток на лунку. Ответ неанергических клеток был значительно снижен зависимым от дозы образом в результате добавления анергических клеток (фиг.6). Приблизительно 90% уменьшения пролиферации получали при концентрации антигена 1 мкг/мл и отношении анергических клеток к неанергическим 2:1. При более высоких концентрациях антигена наблюдали меньшее снижение пролиферации (72% при 5 мкг/мл и 39% при 25 мкг/мл).
Таким образом, этот пример показывает, что можно генерировать антиклонотипические моноклональные антитела против Т-клеточных рецепторов человека. Этот эксперимент привел к получению 10 гибридом, четыре из которых были депонированы с ЕСАСС (Salisbury, UK), как указано в Таблице III. Эта стратегия не требовала больших количеств Т-клеток или больших количеств очищенного TCR. Способ согласно данному изобретению является привлекательной альтернативой для утомительных рекомбинантных процедур, в которых получают растворимый TCR из TCR, экспрессируемых на сингенных мышиных клетках, для использования в процедурах иммунизации. Таким образом, можно избежать также возможных проблем, связанных с конформационной стабильностью. Поскольку моноклональные антитела обнаружили различные ответы в тесте индуцируемой антителами пролиферации Т-клеток и в тесте опосредованного антителами ингибирования индуцируемой антигеном пролиферации Т-клеток (наблюдали до 98% ингибирования), это предполагает, что на одной и той же клонотипической структуре узнаются различные эпитопы.
При перекрестном сшивании все антиклонотипические моноклональные антитела были способны индуцировать пролиферацию Т-клеток Н.243.
Было продемонстрировано, что небольшие количества иммобилизованных антиклонотипических MАb могут индуцировать анергию в аутореактивном клоне. После анергического стимула Т-клетки не могли размножаться при рестимуляции вследствие отсутствия транскрипции гена ИЛ-2. Кроме того, было обнаружено сниженное продуцирование IFNg.
FACS-анализ (анализ с применением клеточного сортера с возбуждением флуоресценции) анергических клеток показал, что анергия не была результатом даун-модулирования (отрицательного модулирования) TCR или отсутствия свободных TCR. В экспериментах по сокультивированию было обнаружено, что анергические Т-клетки подавляют ответ реактивных клеток из того же самого клона. Эта "свидетельская" супрессия приводила к 90% ингибированию пролиферации. Эти данные демонстрируют анергизирующий потенциал клонотипспецифических антител in vitro. Такие MАb можно использовать для индукции и поддержания антигенспецифической толерантности Т-клеток в ревматоидном артрите.
В случае сывороток мышей, иммунизированных инъекцией довольно чистых иммунных комплексов TCR/CD3, связанных с парамагнитными гранулами (Таблица I; группа иммунизации IV), эти сыворотки были более положительными в отношении специфических Т-клеток, которые были использованы для иммунизации, чем в отношении посторонних Т-клеток. Однако невозможно было получение гибридом, продуцирующих антиклонотипические монокло-нальные антитела. Даже хотя в сыворотках мышей, иммунизированных малыми количествами целых Т-клеток, не смогли показать специфичность в отношении целевых Т-клеток в сравнении с посторонними Т-клетками, способ согласно данному изобретению позволил получить гибридомы, способные продуцировать антиклонотипические моноклональные антитела.
Вполне понятно, что из моноклональных антител данного изобретения могут быть получены антигенсвязывающие фрагменты, например, с применением папаина. Моноклональные антитела или их антигенсвязывающие фрагменты могут быть также помечены для облегчения их детектирования, и все эти формы входят в объем данного изобретения.
Также вполне понятно, что не относящиеся к делу клетки, используемые для удаления неспецифических В-клеток, так же подобны, как и клетки, несущие клеточный поверхностный антиген.
ЛИТЕРАТУРА
1. Moretta, A., et al. (1985) Selection and characterization of monoclonal antibodies to the idiotype-like structure of an interleukin-2-producing human leukaemia T-cell line. Int. J. Cancer 36: p. 253-259.
2. Oetgen, H.C. et al. (1986) A T3-like protein complex associated with the antigen receptor on murine T cells. Nature 320: p. 272-275.
3. Steenbakkers, P.G.A., et al. (1994) Efficient generation of monoclonal antibodies from preselected antigen-specific B cells. Molecular Biology Reports 19, p. 125-134.
4. Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of a bacteriophage T4. Nature 227: p. 680-685.
5. Towbin, H. et al. (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 76: p. 4350-4354.
Аббревиатуры
PBS (ЗФР) - забуференный фосфатом физиологический раствор
BCIP - соль 5-бром-4-хлор-3-индоилфосфат п-толуидина
NBT - хлорид п-нитро(синий)-тетразолия
FCS - фетальная телячья сыворотка
SAM - козьи антимышиные
TSM - Т-клеточный супернатант
Mаb - моноклональное антитело (антитела)
Изготовители:
Biorad, Richmond, СА, U.S.A.
Gibco, Paisley, Scotland
Hyclone, Logan, U.S.A.
Packard, Meriden, CT, U.S.A.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СПЕЦИФИЧНОЕ К ЛЕГУМАИНУ | 2013 |
|
RU2574203C2 |
| ИММУНОСТИМУЛЯТОРНЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА | 1995 |
|
RU2155190C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-21 ЧЕЛОВЕКА | 2008 |
|
RU2504552C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-31 И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2444528C2 |
| БИСПЕЦИФИЧЕСКАЯ МОЛЕКУЛА АНТИТЕЛА ДЛЯ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК, СПОСОБ ПОЛУЧЕНИЯ F(AB') 2 ФРАГМЕНТА БИСПЕЦИФИЧЕСКОЙ МОЛЕКУЛЫ АНТИТЕЛА, МОНОКЛОНАЛЬНОЕ АНТИТЕЛО (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ, ФАРМАЦЕВТИЧЕСКИЙ НАБОР ДЛЯ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК (ВАРИАНТЫ), СПОСОБ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК EX VIVO ПРИ АУТОГЕННОЙ ТРАНСПЛАНТАЦИИ КОСТНОГО МОЗГА | 1994 |
|
RU2203319C2 |
| РЕКОМБИНАНТНЫЕ АНТИТЕЛА ПРОТИВ CD4 ДЛЯ ТЕРАПИИ ЧЕЛОВЕКА | 1996 |
|
RU2232773C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ ИНТЕРЛЕЙКИНА-6 ЧЕЛОВЕКА И ГИБРИДОМА, ПРОДУЦИРУЮЩАЯ ДАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО | 2014 |
|
RU2550262C1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К PD-L1 | 2017 |
|
RU2665790C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И ОДНОЦЕПОЧЕЧНЫЕ ФРАГМЕНТЫ АНТИТЕЛ ПРОТИВ КЛЕТОЧНО-ПОВЕРХНОСТНОГО СПЕЦИФИЧЕСКОГО ДЛЯ ПРОСТАТЫ МЕМБРАННОГО АНТИГЕНА | 2006 |
|
RU2458073C2 |
| СПОСОБЫ ИНДУЦИРОВАНИЯ Т-КЛЕТОЧНОЙ ТОЛЕРАНТНОСТИ К ТКАНЕВОМУ ИЛИ ОРГАННОМУ ТРАНСПЛАНТАТУ | 1995 |
|
RU2169009C2 |


Изобретение относится к иммунологии и может быть использовано для получения антител, диагностики аутоиммунного заболевания и лечения ревматоидного артрита. Моноклональное антитело (монАт) продуцируется гибридомами, полученными с использованием материала, содержащего антиген клеточной поверхности, выбранного из группы целых Т-клеток или их мембранной фракции. МонАт специфично в отношении клонотипической структуры Т-клеточного рецептора (TCR). Т-клеточный рецептор ассоциирован с аутоиммунным заболеванием, в частности ревматоидным артритом. На основе монАт получены фармацевтическая композиция для лечения ревматоидного артрита и реагент для диагностики аутоиммунного заболевания. Изобретение позволяет получать антитела против редких и/или нестабильных антигенов, исключая необходимость скрининга больших количеств гибридомных клонов. 4 с. и 16 з.п. ф-лы, 6 ил., 3 табл.
| Моноклональные антитела | |||
| /Под ред | |||
| Р.Г | |||
| Кеннета и др | |||
| - М.: Медицина, 1983, с | |||
| Способ получения нерастворимых лаков основных красителей в субстанции и на волокнах | 1923 |
|
SU132A1 |
| US 5189147 А, 23.02.1993 | |||
| US 4520226 А, 28.05.1985 | |||
| US 4381292 А, 26.04.1983. | |||

