Рак простаты является наиболее распространенным злокачественным заболеванием у мужчин и занимает второе место среди причин смертности в западном мире. В настоящее время не существует радикального лечения нерезектабельных опухолей простаты. Из-за высокой смертности и болезненности, связанной с прогрессированием заболевания, крайне необходимо развитие новых способов направленной терапии. В отличие от рака других систем органов, рак простаты, по ряду причин, является отличной мишенью для терапии с применением антител, поскольку i) простата экспрессирует тканеспецифические антигены, ii) простата является второстепенным органом и ее деструкция не причиняет вреда организму хозяина, iii) областями метастазирования являются лимфоузлы и кости, в которых аккумулируются высокие уровни циркулирующих антител, iv) сывороточные уровни ПСА являются средством контроля терапевтического ответа.
Среди некоторых потенциальных маркеров, которые были идентифицированы при раке простаты, наиболее известным, по-видимому, является специфический для простаты мембранный антиген (ПСМА). Этот трансмембранный гликопротеид II типа, молекулярной массой примерно 100 кДа, состоит из короткого внутриклеточного участка (аминокислоты 1-18), трансмембранного домена (аминокислоты 19-43) и протяженного внеклеточного домена (аминокислоты 44-750). ПСМА может служить мишенью для иммунотерапии, поскольку он соответствует следующим критериям: i) экспрессия главным образом ограничена простатой, ii) в качестве белка ПСМА широко экспрессируется на всех стадиях заболевания, iii) он представлен на клеточной поверхности, однако не поступает в систему кровообращения, iv) экспрессия связана с ферментной или сигнальной активностью. ПСМА также экспрессируется во вновь образованной сосудистой сети большинства других солидных опухолей и поэтому может подходить в качестве мишени для специфической доставки антиангиогенных лекарственных средств.
Благодаря специфичности моноклональных антител (monoclonal antibodies - mAb) по отношению к мишеням, особое значение придавалось их разработке для диагностического и терапевтического применения в онкологии. Однако из-за их размера и иммуногенности применение mAb in vivo связано с серьезными проблемами. Поэтому исследование было сосредоточено на разработке более мелких терапевтических антител с меньшим побочным действием, большей доступностью для опухоли и более высокими показателями клиренса. Благодаря генной инженерии появилась возможность создания одноцепочечных фрагментов антител (single chain antibody fragments - scFv), которые потенциально являются мощным средством для терапии рака. Эти мелкие антитела состоят из вариабельных доменов легкой цепи (light chain - VL) и тяжелой цепи (heavy chain - VH), связанных линкерным пептидом. Они проявляют слабую иммуногенность, почти не имеют токсических эффектов, показывают повышенный уровень клиренса, повышенное накопление в опухолях и лучшее проникновение в опухолевые клетки. Рекомбинантные мышиные scFv можно получить в соответствии со стандартными методами, используя либо экспрессионные библиотеки гибридом, либо клетки селезенки специфически иммунизированных мышей (Chowdhury и др., Mol. Immunol., 4, 1997, c.9-20).
Первое опубликованное mAb (7E11-С5) против ПСМА нацелено на внутриклеточный домен белка, и было установлено, что оно является в высокой степени простатспецифическим (Horoszewicz и др., Anticancer Res., 7, 1987, c.927-935). Кроме того, были получены моноклональные антитела против внеклеточного домена ПСМА после иммунизации этим антигеном. Однако все же есть различие между связыванием с антигеном на фиксированных клетках и гистологических срезах, с одной стороны, и связыванием с жизнеспособными опухолевыми клетками, с другой стороны.
Простатспецифический мембранный антиген (ПСМА) является маркером простаты, который высоко экспрессируется в нормальной простате, а также при раке простаты. Его экспрессия при раке простаты повышается и в основном обнаруживается в простате.
Простатспецифический мембранный антиген (ПСМА) является уникальным связанным с мембраной клеточным белком, который сверхэкспрессирован на раковых клетках простаты, а также во вновь образованной сосудистой сети многих других солидных опухолей, в отличие от сосудистой сети нормальных тканей. Благодаря этой уникальной экспрессии ПСМА является важным маркером, а также крупной внеклеточной мишенью предполагаемых агентов. ПСМА может служить в качестве мишени для доставки терапевтических агентов, например цитотоксинов или радионуклеотидов. ПСМА несет две уникальные ферментные функции, фолатгидролазы и NAALADазы, и установлено, что он рециркулируется через окаймленные клатрином ямки, подобно другим мембраносвязанным рецепторам.
Анти-ПСМА моноклональное антитело (mAb) 7E11 в форме радиоиммуноконъюгата коммерчески доступно в качестве продукта «ProstaScint®», который в настоящее время применяется для выявления метастазов и рецидивов рака простаты. Эпитоп ПСМА, распознаваемый моноклональным антителом 7E11-С5.3, локализован в цитоплазматическом домене простатспецифического мембранного антигена.
Однако также есть сообщения, описывающие экспрессию ПСМА в тканях, не относящихся к простате, включая почки, печень и мозг. Возможное объяснение этому факту дано O′Keefe и др. в Prostate, 58, 2004, с.200-210, а именно, что существует ПСМА-подобный ген, который обладает 98% идентичностью с геном ПМСА на уровне нуклеотидов и который экспрессируется в почках и печени под контролем другого промотора, чем у гена ПСМА.
В WO 01/009192 описывается разработка моноклональных антител человека к простатспецифическому мембранному антигену. Человеческие моноклональные антитела были получены путем иммунизации мышей очищенным ПСМА или обогащенными препаратами антигена ПСМА. Такой очищенный антиген является денатурированным ПСМА, поскольку его очистка проводилась путем иммуноадсорбции после лизиса клеток ионными детергентами.
В WO 97/35616 описываются моноклональные антитела, специфические для внеклеточного домена простатспецифического мембранного антигена. Иммунизации проводились с применением С-концевого пептида или мембранного препарата опухоли, экспрессирующей ПСМА. Эти mAb не связываются специфически с ПСМА-экспрессируюшими клетками и поэтому не могут использоваться для диагностических или терапевтических целей.
Bander и др., Seminars in Oncology, 2003, с.667-677 и US 2004/0213791 соответственно, описывают моноклональные антитела, направленные против простатспецифического мембранного антигена. Поскольку иммунизация проводилась с использованием очищенного антигена, моноклональные антитела не обладали в высокой степени способностью связываться с клетками, и из этих mAb нельзя было получить scFv.
В WO 98/03873 описываются антитела, подобные тем, которые описаны в US 2004/0213791, или их связывающие фрагменты, которые распознают внеклеточный домен простатспецифического мембранного антигена. Нельзя утверждать, что связывающие фрагменты этих антител в действительности связываются с антигеном. Fracasso и др. в The Prostate, 2002, c.9-23, описывают моноклональные антитела против ПСМА, которые химически соединены с А-цепью рицина. Конструкты, описанные в этой статье, не обладают достаточной специфичностью связывания с мишенью и имеют обычные описываемые недостатки поколения иммунотоксинов.
Одной из задач настоящего изобретения является предоставление вышеуказанных средств и конструктов, которые помогут с более высокой достоверностью проводить различия между опухолевыми клетками и здоровыми клетками, которые экспрессируют ПСМА или подобный белок, и ПСМА-отрицательными клетками. Такие конструкты могут применяться для более специфического нацеливания на рак простаты.
Другая задача заключается в предоставлении конструктов, которые уничтожают специфические раковые клетки простаты, экспрессирующие ПСМА.
Простатспецифический мембранный антиген (ПСМА) является привлекательной мишенью для иммунотерапии рака простаты. Однако на клетках простаты ПСМА экспрессируется со специфической третичной и четвертичной структурой, и антитела, специфические для изолированного денатурированного ПСМА, недостаточно хорошо распознают опухолевые клетки, экспрессирующие ПСМА. Антитела и scFv, связывающиеся с денатурированным ПСМА, могут быть получены после иммунизации выделенным очищенным антигеном. Настоящее изобретение, однако, позволяет получать высокоафинные антитела и scFv против нативного клеточного ПСМА путем другого метода иммунизации, который дает начало только незначительному количеству положительных клонов. Только антитела, полученные на основе нативного ПСМА, могут взаимодействовать с клеточно-поверхностным ПСМА и могут применяться в качестве диагностических и терапевтических средств.
Моноклональные антитела (mAb), одноцепочечные фрагменты антител (scFv) и двойные антитела настоящего изобретения были получены в соответствии с традиционными методами из клеток селезенки мышей. Однако мышей иммунизировали клетками LNCaP и лизатом клеток LNCaP, содержащими полноразмерный нативный ПСМА. В предпочтительном варианте осуществления настоящего изобретения антиген, а именно полноразмерный нативный ПСМА, был получен после обработки клеток, предпочтительно клеток LNCaP, специальным лизисным буфером M-PER, реагентом для экстракции белков млекопитающих, который серийно выпускается фирмой Pierce, Рокфор, Иллинойс. В буфере M-PER используется патентованный детергент в 25 ммолярном бициновом буфере (рН 7,6). Скрининг и селекция гибридом и scFv проводились путем проточной цитометрии на ПСМА-положительных клетках LNCaP после абсорбции ПСМА-отрицательными клетками простаты DU 145. Дополнительно их тестировали на реактивность с очищенным ПСМА. Полученные в результате моноклональные антитела и scFv были охарактеризованы с помощью проточной цитометриии на LNCaP и ПСМА-трансфецированных DU 145 клетках и вестерн-блоттингом с очищенным гликозилированным и дегликозилированным ПСМА. Кроме того, проводили иммуноцитологические исследования с клетками LNCaP и иммуногистохимические исследования на парафиновых срезах образцов рака простаты.
В ходе настоящего изобретения удалось получить три mAb (3/F11, 3/А12 и 3/Е7), которые взаимодействовали с жизнеспособными клетками LNCaP и ПСМА-трансфецированными клетками DU 145, но не обладали такой способностью в отношении других клеточных линий, которые не экспрессировали ПСМА. Связывание с клетками LNCaP было очень сильным. При концентрациях насыщения (100 нмоль) среднее значение интенсивности флюоресценции (СИФ) РЕ составляло от 1000 до 1600. Реактивность в отношении очищенного ПСМА была более сильной в случае нативной формы (установлено методом иммуноферментного анализа - ИФА), чем в случае денатурированного и дегликозилированного белка (вестерн-блоттинг). Иммуногистохимия на парафиновых срезах была специфически положительной для эпителиальных клеток с mAb E7.
Путем селекции рекомбинантных фагов на клетках LNCaP и очищенном ПСМА из mAb 3/А12 были получены два scFv, обозначенные Е8 и А5. Последовательность scFv Е8 была идентична scFv A4, который был получен из библиотеки В-клеток той же самой мыши. ScFv E8 был высоко реактивен в отношении клеток LNCaP, показывая значение СИФ примерно 100 при концентрациях насыщения, в то время как значение СИФ для scFv A5 составляло только примерно 40 при тех же условиях. Не было обнаружено или было обнаружено минимальное связывание с другими клеточными линиями, не экспрессирующими ПСМА. Связывание обоих scFv с очищенным денатурированным гликозилированным и дегликозилированным ПСМА было слабым. Кроме того, из mAb 3/F11 был получен scFv, обозначенный D7, а из mAb 3/E7 был получен scFv, обозначенный H12.
В настоящей заявке описывают три mAb, которые отличаются от моноклональных антител, описанных другими авторами, высокой аффинностью связывания и высоким окрашиванием экспрессирующих ПСМА раковых клеток простаты. Путем иммунофлуоресцентной цитологии и детекции с применением конфокальной лазерной сканирующей микроскопии показано, что антитела 3/F11, 3/А12 и 3/E7 не только проявляют сильную связывающую активность, но также интернализуются в клетки LNCaP. Эти mAb были получены после иммунизации полноразмерным нативным ПСМА, в отличие от других опубликованных методов иммунизации.
После иммунизации очищенным денатурированным ПСМА были получены mAb, которые были высоко специфическими по отношению к антигену, но обладали только ограниченным связыванием с экспрессирующими ПСМА клетками LNCaP и к тому же не включались в эти клетки. Эти контрольные данные не показаны в настоящей заявке. В литературе описано несколько mAb против ПСМА. Однако средние значения интенсивности флюоресценции были гораздо ниже, чем с антителами настоящего изобретения.
Подобно mAb, после иммунизации денатурированным и нативным ПСМА были получены scFv против ПСМА. С помощью денатурированного ПСМА авторы изобретения получили scFv, высоко специфические к антигену, но не связывающиеся с клетками LNCaP (данные не показаны в настоящей заявке). В противоположность этому, с помощью нативного ПСМА авторы изобретения получили scFv с высокой активностью связывания с клетками, но с низким связыванием с изолированным денатурированным антигеном.
Однако проблемы, выявленные в этом и других испытаниях с химически связанными иммунотоксинами, заключаются в разработке антител против иммунотоксинов, гепатотоксичностью и синдромом повышенной проницаемости сосудов, а также трудностями в получении больших количеств определенного материала. Эти проблемы, по меньшей мере частично, преодолеваются с помощью генной инженерии, которая позволяет осуществить создание менее иммуногенных более мелких иммунотоксинов, и дает возможность более легкого получения иммунотоксинов в больших количествах. Предполагается также, что более мелкие белки должны лучше проникать в опухоли, чем крупные конъюгаты. Поэтому были созданы рекомбинантные иммунотоксины путем слияния кодирующей последовательности scFv Е8, H12, D7 и А5 и токсина РЕ40. Главным открытием было то, что все рекомбинантные иммунотоксины эффективно убивали культивируемые клетки рака простаты дозозависимым способом. Сильный цитолиз был обнаружен не только с высоко связывающимся Е8-, с IC50 примерно 0,05 нМ, но также с более слабо связывающимся А5-гибридным белком, с IC50 примерно 0,09 нМ. Цитолиз клеток рака простаты, не экспрессирующих ПСМА, был ниже, более чем в 2000 раз. Это может происходить от остаточных бактериальных белков в препаратах иммунотоксинов, поскольку такой же фон можно было наблюдать в равновысоких концентрациях с одним scFv. Понятие IC50 в контексте настоящего изобретения означает концентрацию токсина, выраженную в нмолях, которая редуцирует клеточную пролиферацию на 50% по сравнению с клеточной пролиферацией без добавления токсина.
Антитела и scFv, описанные в настоящей заявке, специфически связываются с нативным ПСМА на поверхности клеток и поэтому являются ценными для диагностического и терапевтического применения, фокусирующегося на ПСМА в качестве антигена-мишени при раке простаты.
Поскольку ПСМА экспрессируется на раковых клетках простаты со специфической третичной и четвертичной структурой, только антитела против этой клеточной конформации могут распознавать и прочно связываться с жизнеспособными раковыми клетками простаты и ПСМА-экспрессирующей тканью. Поэтому, цель настоящего исследования заключалась в получении таких mAb и scFv, которые можно применять для направленной терапии и диагностики рака простаты.
Таким образом, настоящее изобретение предусматривает получение изолированного моноклонального антитела, или его антигенсвязывающего фрагмента, которое связывается со специфическим для простаты мембранным антигеном в его нативной форме, встречающимся на поверхности опухолевых клеток, которое соединено с меткой или цитотоксическим агентом.
Понятие «изолированное моноклональное антитело» относится к гликопротеину, включающему по меньшей мере две тяжелые (heavy - Н) цепи и две легкие (light - L) цепи, взаимосвязанные дисульфидными связями. Каждая тяжелая цепь состоит из вариабельного участка тяжелой цепи (variable region - VH) и константного участка тяжелой цепи. Константный участок тяжелой цепи содержит три домена, а именно СН1, СН2 и СН3. Каждая легкая цепь содержит вариабельный участок легкой цепи (variable region - VL) и константный участок легкой цепи (constant region - CL). Участки VH и VL дополнительно могут подразделяться на гипервариабельные участки, которые также называют участками, определяющими комплементарность (complementary determining region - CDR), рассеянные между участками, которые более консервативны. Такие участки также называются каркасными участками (framework region - FR). Каждый участок VH и VL составлен из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные участки тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном.
На фиг.13, 14 и CDR 20 и 21 заштрихованы серым цветом. Эти области важны для связывания моноклонального антитела или его антигенсвязывающего фрагмента. Другие области являются каркасными участками, которые могут быть замещены другими последовательностями, при условии, что не нарушается трехмерная структура, необходимая для связывания. Если изменить структуру конструкта, не будет достаточного связывания с антигеном. Моноклональные антитела, полученные от мыши, могут вызывать нежелательные иммунологические побочные эффекты, благодаря тому, что они содержат белок от другого вида, который может вызывать выработку антител. Для преодоления этой проблемы моноклональные антитела или их антигенсвязывающие фрагменты можно гуманизировать. Способ получения гуманизированных моноклональных антител известен специалистам в данной области. Каркасные участки мышиного mAb замещают соответствующими каркасными участками человека. Чтобы сохранить предпочтительные связывающие свойства, последовательности участков CDR нужно, по возможности, сохранить. Однако, возможно, что для улучшения связывающих свойств потребуется проведение некоторых аминокислотных замен. Это может выполнить специалист в данной области с помощью обычных методов. Кроме того, при введении структуры человека для улучшения свойств конструкта, возможно, понадобится проведение аминокислотных замен и/или делеций.
Понятие «антигенсвязывающий фрагмент» моноклонального антитела относится к одному или нескольким фрагментам такого антитела, которое сохраняет способность специфически связываться со специфическим мембранным антигеном простаты в его нативной форме. Примеры антигенсвязывающих фрагментов антитела включают фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1 фрагмент F(ab′)2, бивалентный фрагмент, включающий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области, фрагмент Fd, состоящий из доменов VH и CH1, фрагмент FV, состоящий из доменов VL и VH единственной ветви антитела, фрагмент dAb, который состоит из домена VH, и изолированный участок, определяющий комплементарность (CDR).
Изолированное моноклональное антитело или его антигенсвязывающий фрагмент по настоящему изобретению предпочтительно могут поглощаться опухолевой клеткой, если они применяется для терапевтических целей. Для диагностических целей поглощение может не требоваться.
Изолированное моноклональное антитело или его антигенсвязывающий фрагмент по настоящему изобретению прочно связываются с клетками LNCAP, но не связываются с клетками, которые не экспрессируют специфический для простаты мембранный антиген.
Связывание изолированного моноклонального антитела или его антигенсвязывающего фрагмента оценивают по интенсивности флюоресценции фикоэритина (ФЭ) (СИФ), которая предпочтительно равна или выше 40 для scFv и предпочтительно выше 1000 для mAb при концентрациях насыщения.
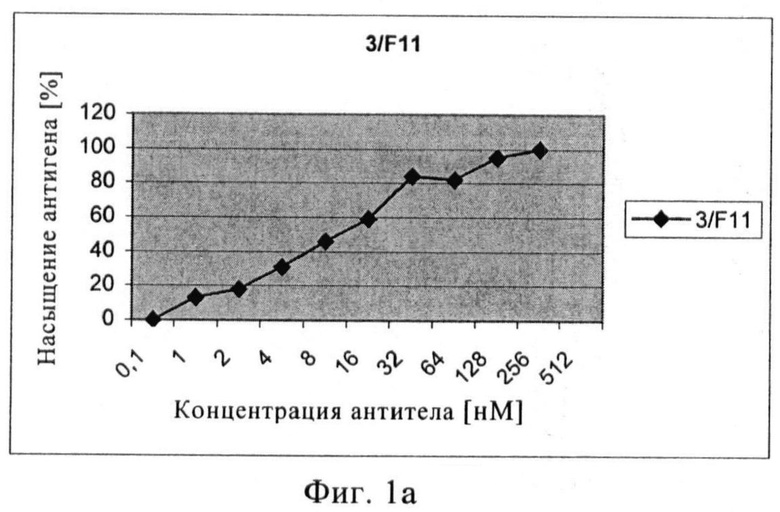
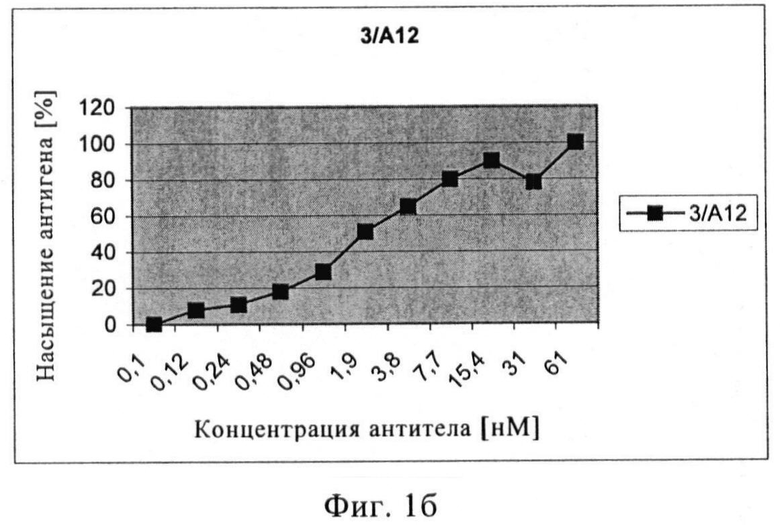
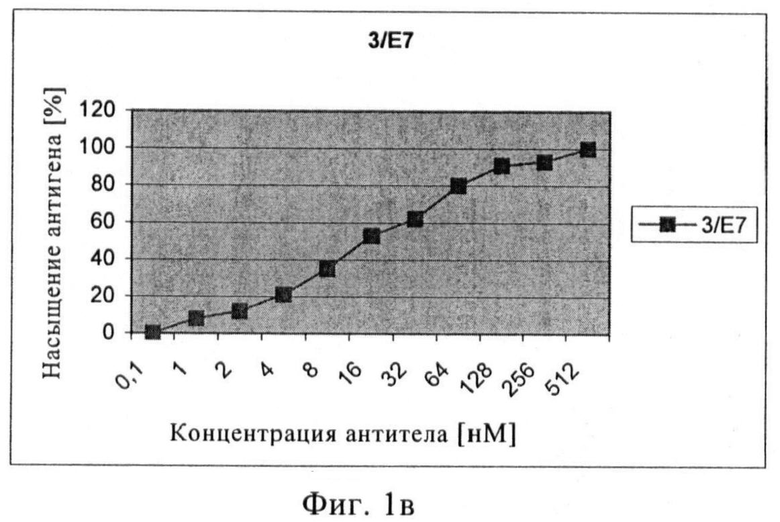
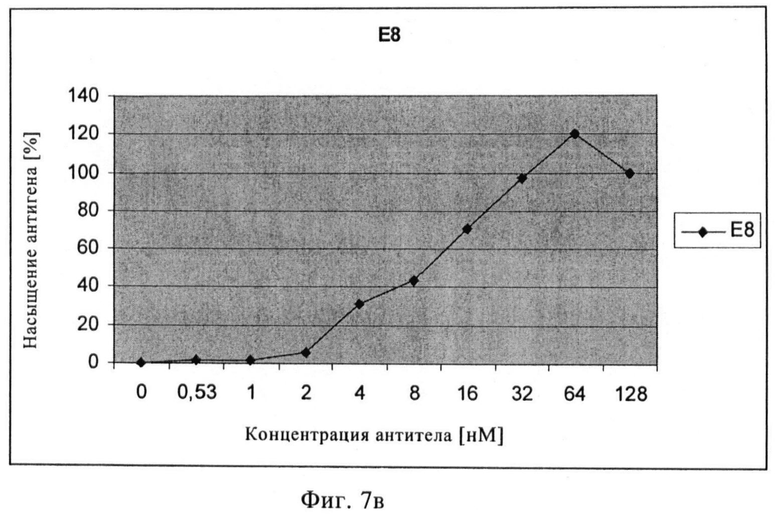
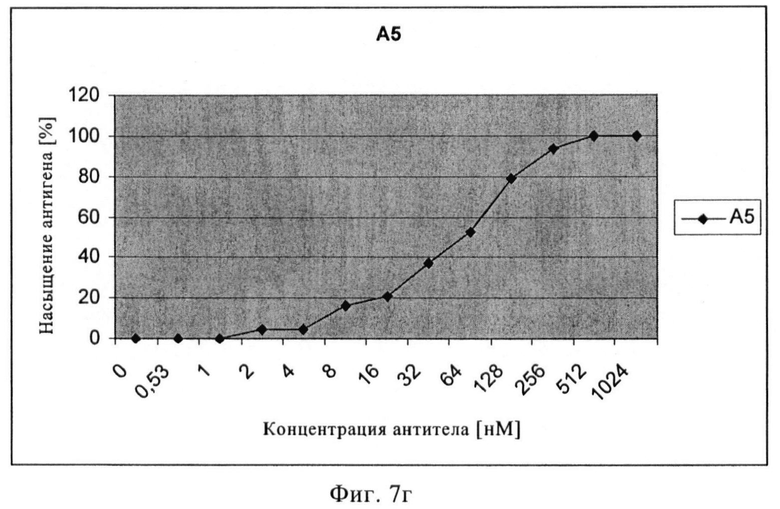
Связывающие свойства моноклональных антител или их антигенсвязывающих фрагментов с нативным ПСМА сравнивали путем обработки клеток LNCAP увеличивающимися концентрациями первичного анти-ПСМА антитела с последующей инкубацией с вторичным ФЭ-меченым антителом. На основе полученных кривых насыщения может быть рассчитана концентрация антитела, при которой достигается 50% насыщение участков ПСМА. Три mAb 3/F11, 3/А12 и 3/Е7 показывали высокую связывающую активность, достигая 50% насыщение участков ПСМА приблизительно при концентрации 16 нМ (3/F11), 2 нМ (3/А12) и 30 нМ (3/Е7). В случае scFv 50% насыщение участков ПСМА было обнаружено при 10 нМ (Е8) и 60 нМ (А5).
Чтобы установить силу связывания проводилось измерение интенсивности флюоресценции ФЭ (фикоэритина) (СИФ). Значения СИФ наносили на график против концентрации антитела (или его связывающих фрагментов), при этом значение плато СИФ соответствует 100% насыщению антигена. После определения верхнего значения (плато, соответствующее 100% насыщению антигена) можно легко установить значение, соответствующее 50% насыщению. С помощью графика можно узнать соответствующие концентрации в нмолях антител или их связывающих фрагментов.
Изолированное моноклональное антитело или его антигенсвязывающий фрагмент включает метку, которая может представлять частицу, которая испускает радиоактивное излучение. Эта частица может быть радиоактивным элементом в форме, которая может быть присоединена к конструкту, предпочтительно в форме комплекса. Например, для детекции отдаленных метастатических опухолей у пациентов с раком простаты в качестве иммуносцинтиграфического агента можно использовать mAb, меченное 111индием. Также можно использовать и другие пригодные радиоактивные элементы, например 35S или 131I.
В другом варианте изолированное моноклональное антитело или его антигенсвязывающий фрагмент могут включать цитотоксический агент, который является клеточным токсикантом, выбранным из группы, состоящей из токсинов, например таксола, цитохалазина В, грамицидина D, этидиум бромида, эметина, митомицина, этопсида, тенопсида, винкристина, винбластина, колхицина, доксирубицина, даунорубицина, дигидроксиантрациндиона, митоксантрона, митрамицина, актиномицина D, 1-дегидротестостерона, гликокортикоидов, прокаина, тетракаина, лидокаина, пропранолола и/или пуромицина.
В предпочтительном варианте осуществления настоящего изобретения изолированное моноклональное антитело или его антигенсвязывающий фрагмент включают частичную аминокислотную последовательность по меньшей мере из 10 последовательных аминокислот SEQ ID NO:1 (scFv Е8), SEQ ID NO:10 (scFv А5), SEQ ID NO:20 (scFv H12) и/или SEQ ID NO:22 (scFv D7). В предпочтительном варианте осуществления настоящего изобретения изолированное моноклональное антитело или его антигенсвязывающий фрагмент включают по меньшей мере 25 или, более предпочтительно по меньшей мере 35 и особо предпочтительно по меньшей мере 50 последовательных аминокислот SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:20 и/или SEQ ID NO:22 соответственно.
В предпочтительном варианте осуществления настоящего изобретения изолированное моноклональное антитело или его антигенсвязывающий фрагмент включают по меньшей мере один из CDR, имеющий последовательности SEQ ID NO:2 - SEQ ID NO:7 и/или SEQ ID NO:11-16. Более предпочтительно такой конструкт включает по меньшей мере 3 и более предпочтительно 5, их этих последовательностей CDR. Аналогичным образом CDR можно вывести из фиг.20 и 21, на которых обозначены последовательности CDR.
В настоящем изобретении также предусмотрены последовательности ДНК, которые можно применять для получения моноклональных антител или их связывающих фрагментов. SEQ ID NO:8 и 9 относятся к scFv E8, a SEQ ID NO:17 и 18 относятся к scFv A5. SEQ ID NO:19 и 23 относятся к scFv H12, a SEQ ID NO:21 и 24 относятся к scFv D7. Последовательности также описывают кодирующую цепь и комплементарную цепь. SEQ ID NO:9 и 18 показаны в 5′→3′ ориентации. Полинуклеотиды настоящего изобретения включают непрерывную последовательность по меньшей мере из 20, предпочтительно 50, более предпочтительно 75 и особо предпочтительно по меньшей мере 100 нуклеотидов из группы, состоящей из SEQ ID NO: 8, 9, 17, 18, 19, 21, 23 и 24. Особенно предпочтительны последовательности, кодирующие CDR.
Также особенностью настоящего изобретения является то обстоятельство, что в нем предусматривается фармацевтическая композиция, включающая изолированное моноклональное антитело или его антигенсвязывающий фрагмент, описанные в настоящей заявке. Фармацевтическая композиция настоящего изобретения включает моноклональное антитело или его антигенсвязывающий фрагмент вместе с фармацевтически приемлемыми вспомогательными веществами. Предпочтительно такую композицию получают для внутримышечного или внутривенного введения. В другом варианте антитело может быть предусмотрено в составе для пролонгированного действия, который позволяет проводить замедленное высвобождение биологически активного агента в течение определенного времени, которое может колебаться предпочтительно от одного до шести месяцев. Такой состав замедленного высвобождения может включать полимер, поддающийся биодеструкции, например, полилактид или сополимер полилактида/полигликолида, который в организме человека разлагается в течение длительного времени, при этом происходит контролируемое высвобождение в течение определенного времени антитела, или его антигенсвязывающего фрагмента, предпочтительно несущего токсин.
Изолированное моноклональное антитело или его антигенсвязывающий фрагмент может применяться для получения лекарственного средства для лечения рака, в частности рака простаты.
В другом варианте в настоящем изобретении предусматривается диагностический набор для выявления опухолевых клеток, включающий изолированное моноклональное антитело или его антигенсвязывающий фрагмент. В таких вариантах осуществления настоящего изобретения метка позволяет выявлять конструкцию с помощью соответствующих устройств.
Настоящее изобретение также предусматривает способ для идентификации in vitro опухолевых клеток, при котором опухолевые клетки, подлежащие идентификации, подвергают контакту с изолированным моноклональным антителом или его антигенсвязывающим фрагментом, несущим метку, которую можно обнаружить соответствующими аналитическими приборами. Метка позволяет проводить диагностическую идентификацию опухолевых клеток, например, в срезе опухолевой ткани, полученной после хирургии или биопсии.
Краткое описание фигур
Фиг.1. FACS-анализ связывания mAb 3/F11, 3/А12 и 3/Е7 с поверхностью ПСМА-экспрессирующих клеток LNCaP при концентрациях насыщения.
Фиг.1а-в. Кривые насыщения антигена антителами 3/F11 (а), 3/А12 (б), 3/Е7 (в).
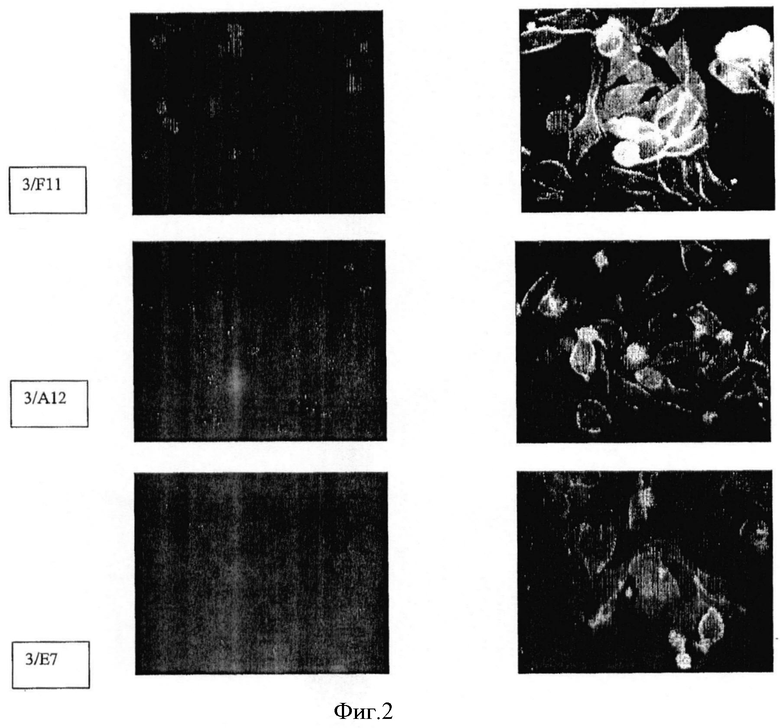
Фиг.2. Иммунофлюоресцентная цитология: Связывание а) mAb 3/F11 б) mAb 3А/12 в) 3Е7 с клетками LNCaP. Рисунки слева показывают контрольное окрашивание 4′,6-диамидино-2-фенилиндолом (ДАПИ).
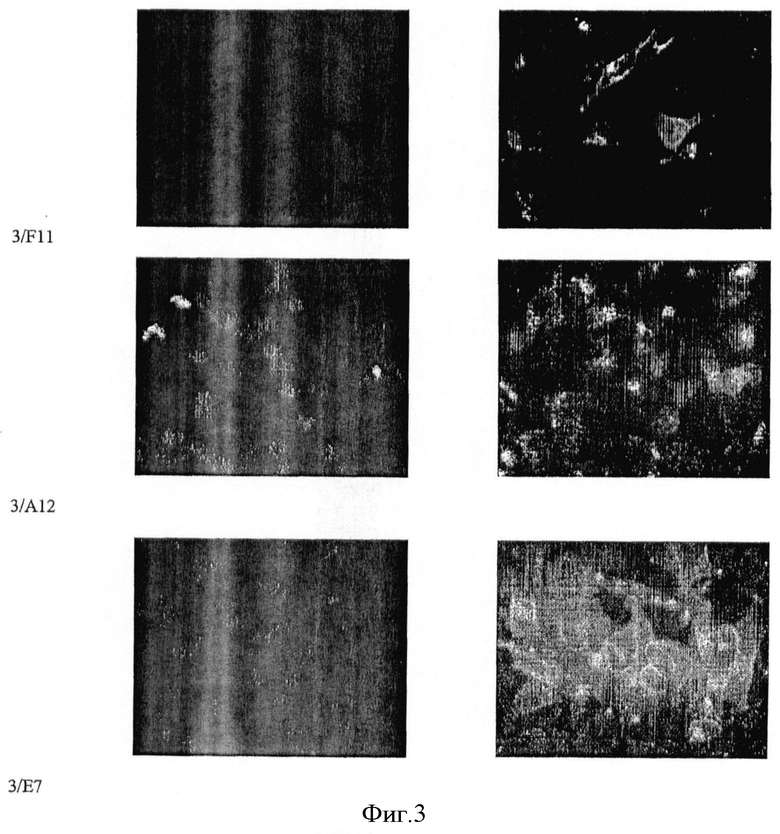
Фиг.3. Иммунофлюоресцентная цитология: Интернализация a) mAb 3/F11 б) mAb 3А/12 в) 3Е7 в клетки LNCaP. Рисунки слева показывают контрольное окрашивание 4′,6-диамидино-2-фенилиндолом (ДАПИ).
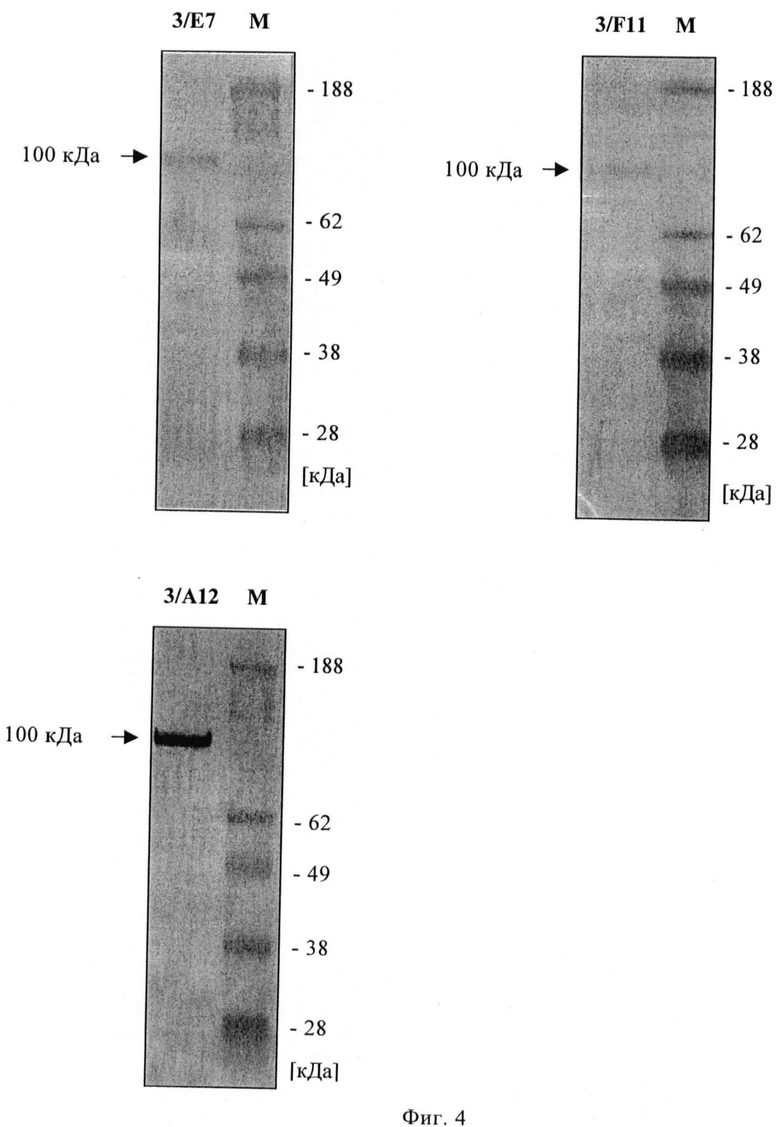
Фиг.4. Вестерн-блоттинг с очищенным ПСМА и mAb 3/E7, 3/А12 и 3/F11.
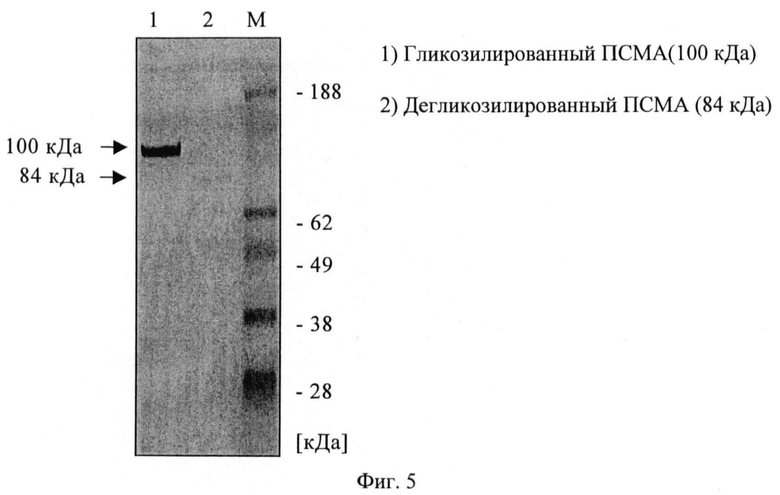
Фиг.5. Вестерн-блоттинг с гликозилированным и дегликозилированным ПСМА и mAb 3/A12.
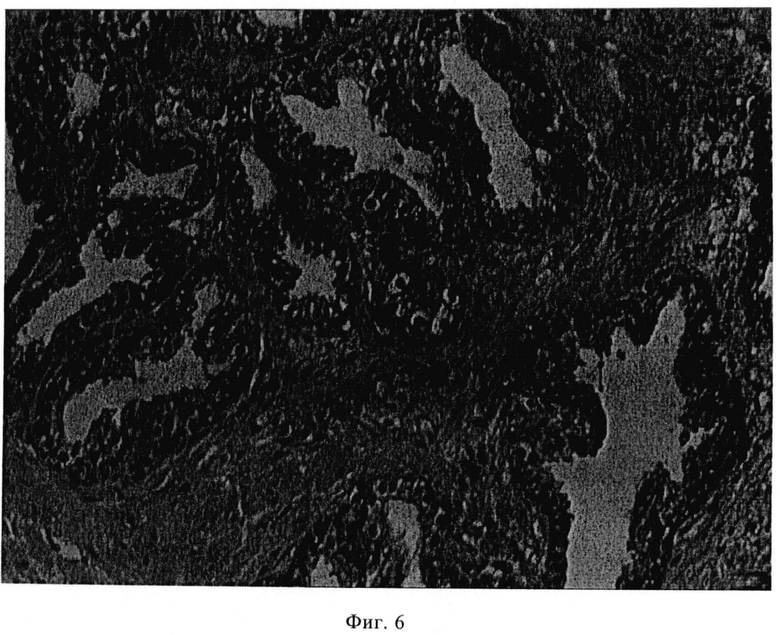
Фиг.6. Иммуногистохимия mAb 3/E7 на парафиновом срезе раковой опухоли простаты.
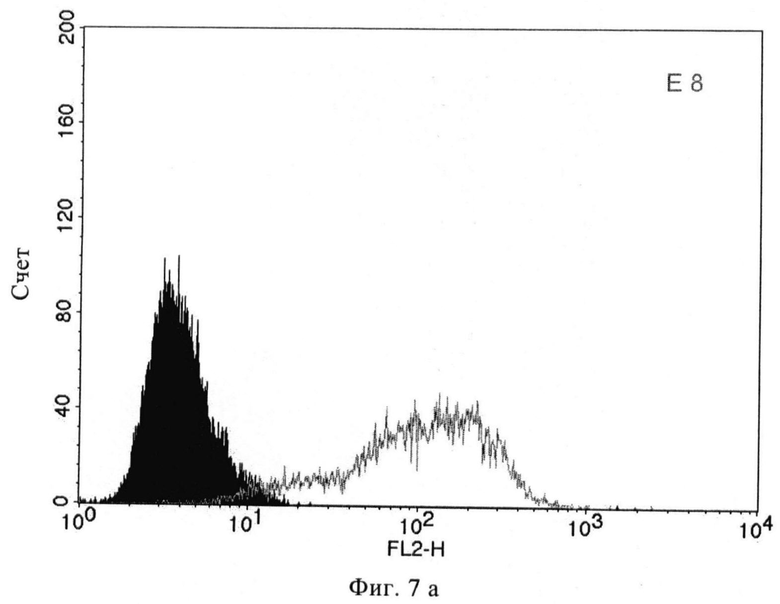
Фиг.7а/б. FACS-анализ scFv Е8 (а) и А5 (б) на ПСМА-экспрессирующих клетках LNCaP при концентрациях насыщения.
Фиг.7в/г. Кривые насыщения антигена scFv Е8 (в) и А5 (г).
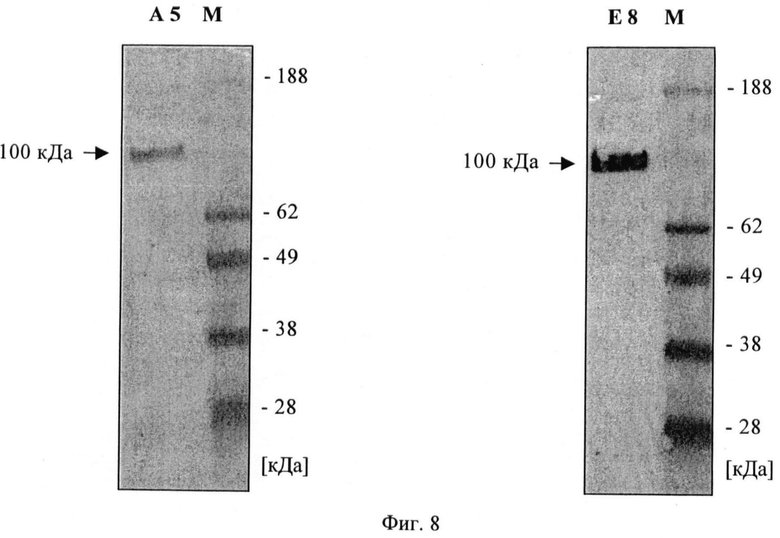
Фиг.8. Вестерн-блоттинг с очищенным ПСМА и scFv А5 и Е8.
Фиг.9. Иммуноцитология scFv Е8 на клетках LNCaP.
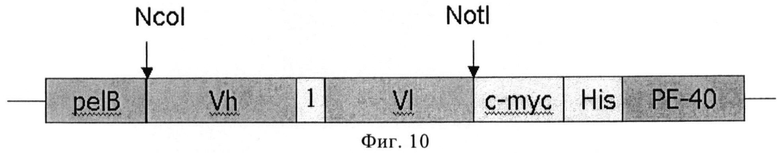
Фиг.10. Конструкция иммунотоксина Е8-РЕ40.
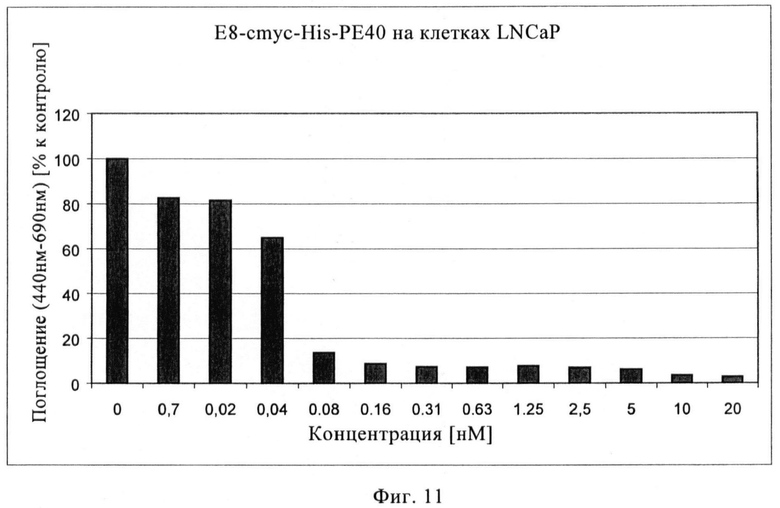
Фиг.11. Цитотоксический эффект рекомбинантного иммунотоксина Е8-РЕ40 на клетках LNCaP.
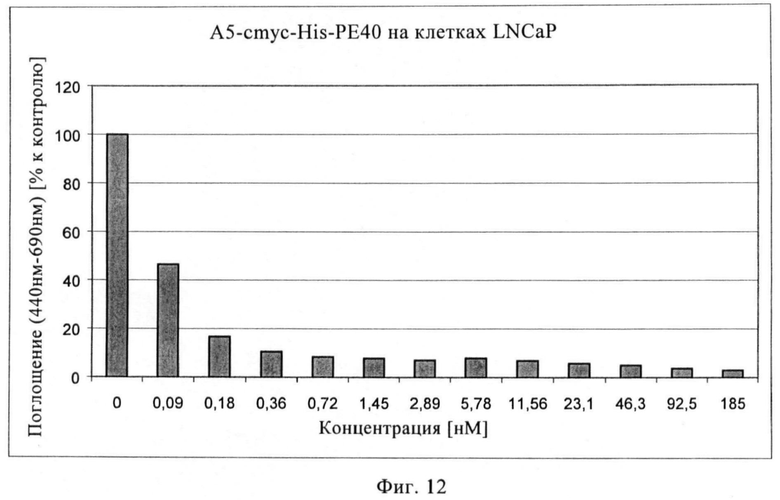
Фиг.12. Цитотоксический эффект рекомбинантного гибридного белка А5-РЕ40 на клетках LNCaP.
Фиг.13. Последовательность scFv Е8. Приведена последовательность ДНК, а также аминокислотная последовательность, при этом помеченные зоны означают участки CDR.
Фиг.14. Последовательность scFv Е5. Приведена последовательность ДНК, а также аминокислотная последовательность, при этом помеченные зоны означают участки CDR.
Фиг.15. Связывание scFv А5, Н12 и D7 с ПСМА-отрицательными клетками DU145 (А) и ПСМА-положительными клетками LNCaP (А5=А, Н12=Б, D7=В). Клетки окрашивали mAb и РЕ-конъюгированным mAb к IgG мыши. Гистограммы представляют логарифмы флуоресценции ФЭ на проточном цитометре. Отрицательный контроль был выполнен только с вторичным антителом.
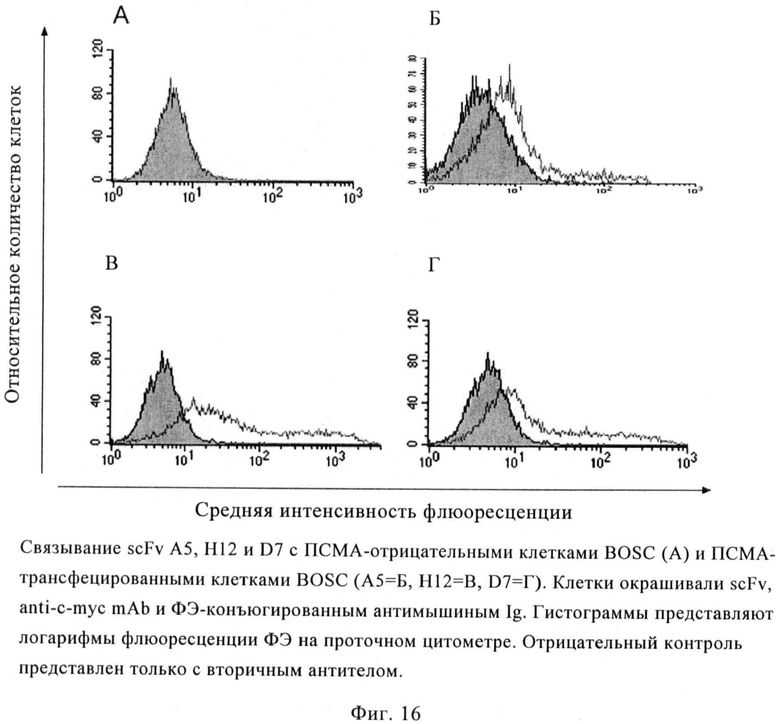
Фиг.16. Связывание scFv А5, Н12 и D7 с ПСМА-отрицательными клетками BOSC (А) и ПСМА-трансфецированными клетками BOSC (А5=Б, Н12=В, D7=Г). Клетки окрашивали scFv анти-с-myc mAb и ФЭ-конъюгированным антимышиным Ig. Гистограммы представляют логарифмы флуоресценции ФЭ на проточном цитометре. Отрицательный контроль был выполнен только с вторичным антителом.
Фиг.17. Цитотоксическое действие рекомбинантного иммунотоксина НЕ12-РЕ40 на клетки LNCaP (черные) и клетки DU (белые).
Фиг.18. Схема строения двойного антитела A5-CD3.
Фиг.19. Цитотоксический эффект двойного антитела, построенного из scFv А5 (A5/CD3), при различных концентрациях, и лимфоцитов периферической крови (целевое соотношение 10:1) на клетки LNCaP после инкубации в течение 48 ч.
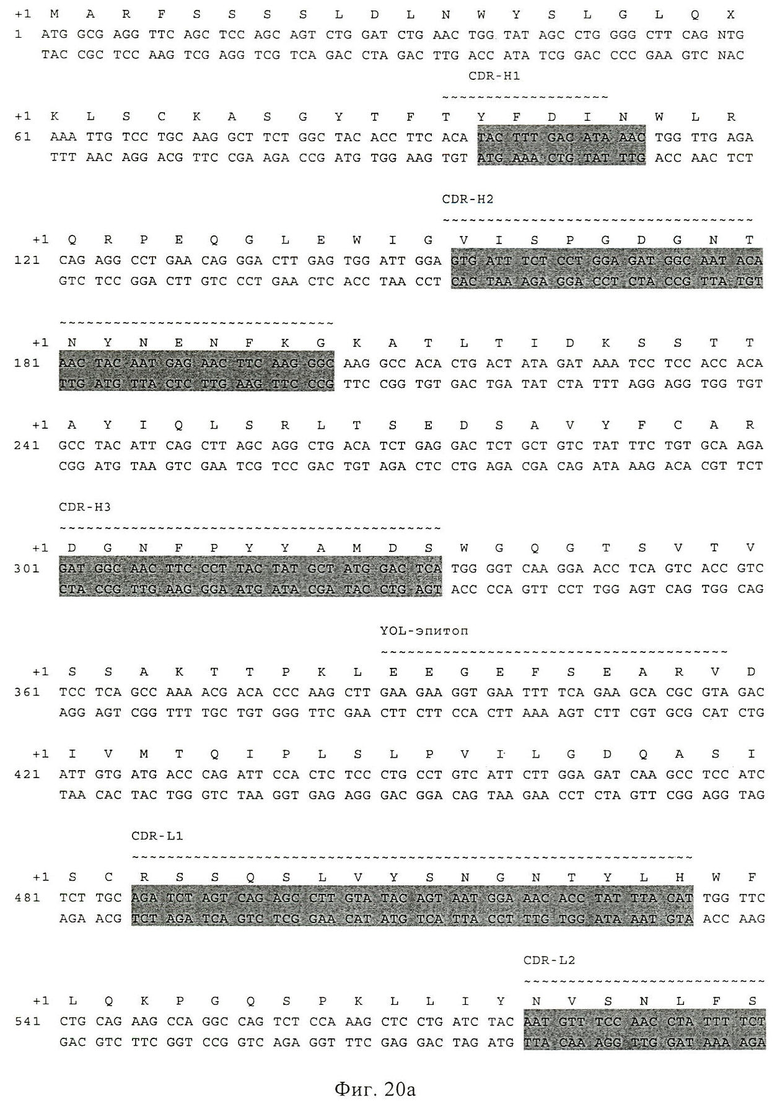
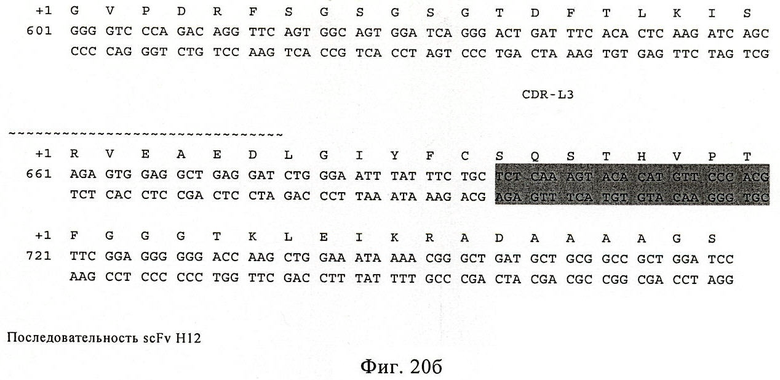
Фиг.20. Последовательность scFv HI 2. Аминокислотная последовательность показана в виде однобуквенного кода на первой линии (соответствует последовательности SEQ ID NO:20). Кодирующая цепь показана на второй линии (SEQ ID NO:19) и комплементарная цепь показана на третьей линии. Эта последовательность соответствует SEQ ID NO:23. CDR специфически обозначены в качестве CDR H1, H2, Н3, L1, L2 и L3. Последовательности нуклеиновых кислот, кодирующие области CDR, показаны на сером фоне.
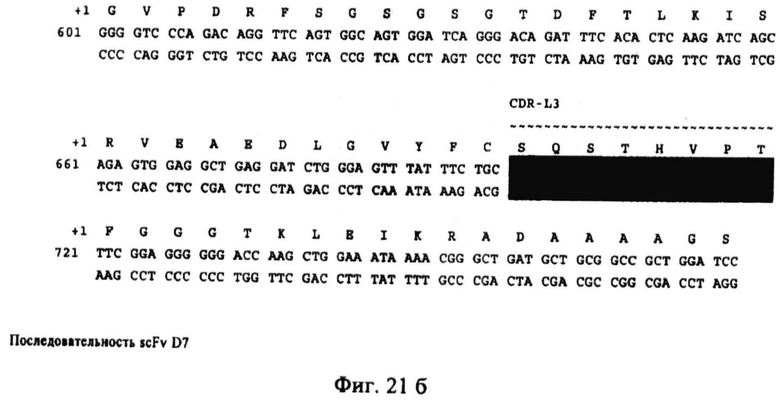
Фиг.21. Последовательность scFv D7. Аминокислотная последовательность показана на первой линии в виде однобуквенного кода. Эта последовательность соответствует SEQ ID NO:22. Кодирующая цепь нуклеиновой кислоты показана на первой линии. Эта последовательность соответствует SEQ ID NO:21 и комплементарная цепь показана на третьей линии. Эта последовательность соответствует SEQ ID NO:24. В последовательности показаны области CDR H1, H2, Н3, L1 и L2. Последовательности нуклеиновых кислот, кодирующие эти области, показаны на сером фоне.
Далее настоящее изобретение иллюстрируется следующими примерами.
Пример 1
а) Получение ПСМА
Клеточные линии карциномы простаты человека LNCaP, DU 145, PC-3 и HeLa, а также гибридома 7Е11-С5.3 (IgG1-k, ПСМА) получают в Американской коллекции типовых культур (АТСС), Роквилл, Мэриленд, США. Клетки LNCaP, DU 145 и HeLa культивируют в среде RPMI 1640, PC-3 культивируют в среде Nutrimix F12, обе среды с добавлением пенициллина (100000 Ед/л), стрептомицина (100 мг/л) и 10% ФСТ при 37°С во влажной атмосфере с 5% СO2. Для генерирования клеток LNCaP, экспрессирующих негликозилированный ПСМА на своей поверхности, в среду добавляют 2 мкг/мл туникамицина (фирма ICN Biomedicals) в течение 48 ч.
Фиксированные клетки LNCaP получают путем обработки 4% параформальдегида в течение 10 мин при комнатной температуре и затем тщательно промывают ФСБ.
Для получения очищенного ПСМА 108 клеток LNCaP промывают ФСБ и затем лизируют в ФСБ, содержащем 1% IGEPAL в течение 20 мин при комнатной температуре. После центрифугирования при 10000 g супернатант наносят на колонку для аффинной хроматографии 7Е11-С5 (фирма Amersham Biosciences, Упсала, Швеция) и элюируют ПСМА 100 ммолярным глициновым буфером, рН 2,5, содержащим 1% Тритона Х-100. После нейтрализации белок экстенсивно диализируют с применением ФСБ.
Для приготовления дегликозилированного ПСМА, 1/10 объема гликопротеин-денатурирующего буфера (фирма BioLabs) добавляют к раствору, содержащему очищенный ПСМА, и нагревают в течение 10 мин при 100°С. Затем добавляют 1/10 объема 10% NP-40 (10%) и 50 Ед фермента PNGase на мкг ПМСА и инкубируют при 37°С в течение 1 ч.
Для получения лизата клеток LNCaP, содержащего полноразмерный нативный ПСМА, клетки лизируют реагентом M-PER (фирма Pierce) в течение 10 мин и затем центрифугируют при 15000 об/мин в течение 30 мин при 4°С. Супернатант, содержащий нативный полный ПСМА, собирают (M-PER-лизат). Высокомолекулярную фракцию (100-600 кДа) этого лизата разделяют с помощью ВЭЖХ на ионообменной колонке Biosil 250.
б) Трансфекция полноразмерного ПСМА в клетки DU 145 и РС3
Полноразмерный ПСМА клонируют в двух фрагментах (фрагмент 1 от 262 п.о. до специфического сайта рестрикции EcoRI при 1573 п.о. и фрагмент 2 от положения 1574 до 2512) в вектор pCR3.1 (фирма Invitrogen). Временную трансфекцию получают путем добавления смеси 4 мкг ДНК и 10 мкл липофектамина (фирма Invitrogen) в 500 мкл среды RPMI к 106 клеткам по протоколу изготовителя. После инкубации в течение 48 ч временно трансфецированные клетки используют для исследования.
Пример 2
Иммунизация мышей
Четырехмесячных самок мышей линии Balb/c иммунизируют внутрибрюшинно 300 мкг M-PER-лизата клеток LNCaP, или высокомолекулярной фракцией ВЭЖХ лизата, или 106 клетками LNCaP, фиксированными 2% параформальдегидом. Эти препараты смешивают в отношении 1:1 с полным адъювантом Фрейнда. Каждую мышь иммунизируют 4 или 5 раз с 2-х недельными интервалами. Спустя четверо суток после последней иммунизации клетки селезенки собирают и используют для получения либо гибридом, либо библиотеки В-клеток.
Пример 3
Получение библиотеки В-клеток
Селезенку мыши промывают в фосфатно-солевом буферном растворе (ФСБ), измельчают до мелких кусочков, снова промывают в ФСБ и затем мягко гомогенизируют в «свободном» ручном гомогенизаторе. Полученную в результате однородную клеточную суспензию наслаивают на Ficoll (фирма Pharmacia, Фрейбург, Германия) и центрифугируют при 400 g в течение 20 мин при комнатной температуре. Межфазные В-клетки изолируют с помощью микрошариков CD 19 по инструкциям производителя (фирма Miltenyi, Гербиш Гладбах, Германия). В-клетки в количестве 106 лизируют в 350 мкл раствора, состоящего из 4 молей гуанидинтиоционата, 25 ммолей цитрата натрия, 0,5% N-лауросилсаркозината натрия и 100 ммолей 2-меркаптоэтанола.
Пример 4
а) Получение гибридом
Селезенку асептические удаляют и готовят суспензию отдельных клеток в среде RPMI-1640, не содержащей сыворотки. Спленоциты добавляют к клеткам миеломы SP2/0 в отношении 10:1 и по общепринятым методам проводят слияние и селекцию (Galfre и др., Nature, 1979, с.131-133).
Супернатанты гибридом исследуют FACS-анализом на клетках LNCaP и DU145 и ИФА с очищенным ПСМА в качестве твердой фазы. Очистку моноклональных антител проводят, используя колонку с белком G (фирма Pharmacia).
б) Определение изотипа mAb
Ig-изотипы mAb к ПСМА определяют ИФА, используя либо немеченые (твердая фаза), либо меченные пероксидазой (следы) специфические антитела против изотипов (фирма Southern Biotechnology Associates, Бирмингем, Алабама).
в) Изоляция и характеристика конформационных моноклональных антител против ПСМА
Проводят слияние клеток селезенки мышей Balb/c, которых 5 раз иммунизировали M-PER-лизатом клеток LNCaP, с клетками SP2/0 по общепринятым методам. Положительные гибридомы отбирают путем проточной цитометрии с клетками LNCaP и ИФА на очищенном ПСМА. Таким способом получают три положительных клона. Соответствующие mAb с их изотипами представляют 3/F11 (IgG2a), 3/A12 (IgG1) и 3/Е7 (IgG2b).
г) Характеристика mAb
Путем проточной цитометрии можно заметить, что три mAb и окрашенные клетки LNCaP очень хорошо связываются с положительными клетками, количество которых находится в диапазоне от 95% до 98%. Форма кривых флюоресценции относительно числа событий показывает, что ПСМА однородно распределяется в популяции клеток LNCaP (фиг.1). Чтобы оценить специфичность связывания анти-ПСМА mAb, ПСМА-отрицательные клетки DU145, РС3, клетки HeLa и Jurkat также окрашивают и анализируют путем проточной цитометрии. Ни один из трех mAb не окрашивает ПСМА-отрицательные клетки (процентное содержание положительных клеток колебалось в пределах от 0,04% до 2%).
Связывающие свойства трех антител сравнивают путем обработки клеток LNCaP возрастающими концентрациями первичного анти-ПСМА mAb с последующей инкубацией с насыщающим количеством вторичного ФЭ-(фикоэритин)-меченого козьего антитела и последующим цитофлюорометрическим анализом. При концентрациях антигенного насыщения [100 нМ] скорректированная средняя интенсивность флюоресценции фикоэритина составляет примерно 1000 для mAb 3А 12, примерно 1400 для mAb 3F11 и примерно 1600 для mAb 3E7. Показано, что для mAb 3A12 СИФ в 5-раз ниже на клетках LNCaP, экспрессирующих негликозилированный ПСМА (выращенных с туникамицином).
Путем иммунофлюоресцентной цитологии и детекции с использованием лазерного сканирующего конфокального микроскопа можно показать сильное связывание трех mAb с клетками LNCaP (фиг.2), а также их интернализацию в эти клетки (фиг.3). Все mAb положительны в ИФА с очищенным ПСМА в качестве твердой фазы. С денатурированным ПСМА mAb показывают сигнал примерно при 100 кДа в вестерн-блоттинге (фиг.4), а блот с дегликозилированным ПСМА слабый, дающий сигнал примерно при 84 кДа, что соответствует литературным данным (фиг.5).
Иммуногистохимия на парафиновых срезах раковой ткани простаты положительна в случае mAb 3/E7, но отрицательна в случае mAb 3/F11 и 3/А12 (фиг.6). Данные объединены в таблице 1.

На основании этих данных можно сделать заключение, что 3 mAbs показывают очень сильное и высокоспецифическое связывание с нативным и денатурированным ПСМА. Хотя связывание с дегликозилированным ПСМА является слабым, специфичность сахара может быть исключена на основании того факта, что не наблюдается связывания с клетками, которые не экспрессируют ПСМА.
Пример 5
Получение библиотеки экспрессии scFv в фагмиде pSEX
Из библиотеки В-клеток или из гибридомных клеток выделяют общую РНК и иРНК с использованием мембран на основе силикагеля (реагент Rneasy, фирма Qiagen, Хилден, Германия) по протоколу производителя. Синтез кДНК осуществляют при 42°С в течение 60 мин в конечном объеме 50 мкл, который содержит 25 мкл денатурированной РНК, 10 мкл 5 × буфера (фирма Promega, Гейдельберг, Германия), 5 мкл 10 мМ дНТФ (дАТФ, дЦТФ, дГТФ, дТТФ, фирма Promega), 1,5 мкл РНК-зина (40 Ед/мкл, фирма Promega), 2,5 мкл 150 пМ произвольных гексамерных праймеров и 2,5 мкл обратной транскриптазы AMV (10 Ед/мкл, фирма Promega). Затем кодирующие участки тяжелых цепей и гамма- и каппа-цепей амплифицируют путем ПЦР по приведенному ранее описанию Orum и др. в Nucleic Acies Res., 1993, с.4491-4498. Для каждой цепи проводят 25 отдельных реакций, сочетанием 25 различных прямых праймеров для константных участков с одним соответствующим обратным праймером. Очистку амплифицированных продуктов для легких цепей и тяжелых цепей проводят электрофорезом в агарозном геле.
Продукты ПЦР для легких цепей расщепляют ферментами MluI и NotI, и лигируют к фагмиде pSEX81 (Dübel и др. Gene, 1993, с.97-101) при молярном соотношении 1:3 (2 мкг вектора, 400 нг вставки). Продукты одного лигирования используют для электропорации 50 мкл электрокомпетентных клеток Е. coli XL1-blue (фирма Stratagene) по протоколу поставщика. Бактерии высевают на девять чашек диаметром 80 мм с агарозной средой, содержащей 100 мкг/мл ампициллина и 0,1 М глюкозы (SOB-AG), и инкубируют в течение ночи при 30°С. Бактерии изолируют, добавляя 3 мл 2×YT среды на каждую чашку и соскабливая их стерильным стеклянным шпателем, и осаждают при 3000 g в течение 15 мин. Из этих бактерий выделяют плазмидную ДНК, которая представляет суббиблиотеку Vl. Затем продукты ПЦР для тяжелой цепи и суббиблиотеку Vl расщепляют ферментами NcoI и HindIII. Лигирование осуществляют при соотношении 3:1 (2 мкг суббиблиотеки и 400 нг вставки). Трансформацию путем электропорации, посев и отбор трансформированных бактерий проводят по описанию для суббиблиотеки Vl. С девяти пластин SOB-AG диаметром 80 мм получают в целом 18 мл библиотеки VHVL.
Пример 6
Получение и селекция фага, экспонирующего антитело
а) Получение
В библиотеке VHVL в фагмиде pSEX гены антител гбридизируют в рамке с геном III, который кодирует минорный поверхностный белок gIIIp нитчатого фага. Следовательно, получение рекомбинантных фагмидных частиц, экспонирующих на поверхности антитело, требует заражения несущей фагмиду бактериальной клетки дефектным по репликации фагом M13KO7 [14]. M13KO7 добавляют к 10 мл библиотечной культуры при кратности 10. После инкубации при 37°С в течение 90 мин клетки осаждают центрифугированием и ресуспендируют в 15 мл 2×YT-среды, содержащей 100 мкг/мл ампициллина, 10 мкг/мл тетрациклина и 50 мкг/мл канамицина. Культуру инкубируют в течение ночи при 37°С при 250 об/мин, затем охлаждают на льду и центрифугируют для удаления клеток. Супернатант, содержащий фаги, смешивают с 1/5 объема водного раствора, содержащего 20% ПЭГ 8000 и 14% NaCl, и инкубируют 1 ч при 4°С . Затем центрифугируют 30 мин при 4°С и 6200 g. Осадок, содержащий фаг, ресуспендируют в 2 мл 10 мМ Трис/HCl, рН 7,5, 20 мМ NaCl, 2 мМ ЭДТА, рН 7,5 и используют для пэннинга.
б) Пэннинг для выбора клонов, связывающихся с антигеном и клеткой
Пэннинг на очищенном ПСМА проводят в 96-луночных планшетах для микротитрования Maxi-Sorb (фирма Nunc), покрытых раствором очищенного ПСМА (100 мкл/лунку, 12 мкг/мл ПСМА в ФСБ), и блокируют 4% обезжиренным молоком/ФСБ. Один мл очищенных рекомбинантных фагов (примерно 1011) инкубируют в 1 мл 4% обезжиренного молока/ФСБ с добавлением 15 мкл 10% Тритона X100 в течение 15 мин, затем допускают связывание с 8 лунками, покрытыми ПСМА, в течение 2 ч при 37°С. После 20 циклов промываний с использованием раствора ФСБ/Твин (0,1%) связанные фаги элюируют 0,1 М глициновым буфером, рН 2,2. Для пэннинга на жизнеспособных клетках LNCaP фаги предварительно абсорбируют на клетках DU 145. Для этой процедуры 1 мл (примерно 1011) рекомбинантных фагов инкубируют в 2% обезжиренном молоке/ФСБ в течение 15 мин и затем с клетками DU 145 (107) в течение 1 ч при комнатной температуре на шейкере. После этого клетки центрифугируют и супернатант с неабсорбированными фагами инкубируют с клетками LNCaP (106) в течение 1 ч при комнатной температуре на шейкере. После 10 промываний 2% обезжиренным молоком/ФСБ и пяти промываний ФСБ связанные фаги элюируют 50 М HCl с последующей нейтрализацией 1 М Трис-HCl (рН 7,5).
Клетки Е. coli TG1 инфицируют элюированными фагами, высевают в чашки с SOB-AG и инкубируют в течение ночи при 30°С. Для титрования используют аликвоты элюата. Процедуру селекции повторяют от трех да шести раз.
в) «Спасение» мелких фагов
Изолируют отдельные колонии из 96-луночного планшета для микротитрования и каждую из них переносят в отдельную лунку 96-луночного планшета с глубокими лунками, содержащими по 500 мкл 2×YT-среды, содержащей 100 мкг/мл ампициллина и 0,1 моль глюкозы (YT-AG), и инкубируют в течение ночи при 37°С (главный планшет). Затем по 40 мкл насыщенной культуры из каждой лунки главного планшета переносят в соответствующие лунки нового планшета, содержащие по 400 мкл 2×YT-AG-среды.
К каждой лунке добавляют примерно 1×1010 херперных фагов М13КО7 и инкубируют на шейкере в течение 2 ч при 37°С. Затем планшет центрифугируют, осадок суспендируют в среде 2×YT с добавлением 100 мкг/мл ампициллина, 10 мкг/мл тетрациклина и 50 мкг/мл канамицина и инкубируют при 29°С и 240 об/мин в течение ночи. После центрифугирования супернатант, содержащий спасаемые фагмиды, отбирают и используют для исследования фагов методами ИФА и проточной цитометрии.
г) Изучение фагов методом ИФА
Планшеты для микротитрования покрывают очищенным ПСМА (1,5 мкг ПСМА/мл ФСБ), выдерживают в течение ночи и затем блокируют 2% обезжиренным молоком/ФСБ. К каждой лунке добавляют по 200 мкл спасенных фагмид, предварительно инкубированных 1:1 с 2% обезжиренным молоком/ФСБ, и инкубируют в течение 2 ч при комнатной температуре. После пяти этапов промывания с использованием ФСБ-Твина, связанные фаги выявляют с использованием 200 мкл/лунку анти-М13 антитела, конъюгированного с пероксидазой хрена (фирма Pharmacia), в течение 2 ч при комнатной температуре. В качестве субстрата используют с 3,3′,5′,5′-тетраметилбензидин.
д) Выделение и характеристика конформационного scFv против ПСМА
Для генерации конформационного scFv против ПСМА создают библиотеку VHVL в фагмиде pSEX из библиотеки В-клеток мыши, иммунизированной М-PER-лизатом клеток LNCaP. Численность этой библиотеки составляет 107. Подобным образом готовят библиотеку VHVL из моноклонального антитела 3/А12, которое получают от той же мыши, иммунизированной лизатом клеток LNCaP. Численность этой библиотеки составляет 105. Чтобы выделить фаги, экспонирующие на своей поверхности scFv, связывающий клеточный ПСМА, в другом варианте проводят шесть циклов пэннинга на клетках LNCaP после абсорбции клетками DU-145 в полистероловых пробирках и в микротитровальных планшетах, покрытых 20 мкг/мл очищенного ПСМА. После трех, четырех и шести циклов пэннинга вырастают отдельные фагмидные колонии и фаговые частицы спасают заражением М13КО7. Анализ 800 фаговых клонов из библиотеки В-клеток путем проточной цитометриии с клетками LNCaP и ИФА на очищенном ПСМА выявил один положительный клон, названный Е8. После четвертого цикла пэннинга из библиотеки VHVL от mAb 3/А12 получено два положительных клона, названных А4 и А5. Путем секвенирования обнаружено, что А4 идентичен Е8.
Кодирующие области scFv Е8 и А5 переносят из фагмиды pSEX в вектор экспрессии pHOG, содержащий с-myc и His-tag на С-конце. Последовательности с соответствующими CDR представлены на фиг 13 и фиг.14. Области, кодирующие CDR, антигенсвязывающих частей отмечены на фиг.13 и 14. Эти последовательности не должны подвергаться изменениям, при этом другие части последовательности, которые не отмечены, можно изменять. Однако должна сохраняться соответствующая трехмерная структура.
С помощью проточной цитометрии установлено, что scFv Е8 сильно взаимодействует с жизнеспособными клетками LNCaP, причем значение СИФ примерно равно 100 при концентрациях насыщения, при этом связывание А5 было гораздо слабее и значение СИФ равно примерно 40 при концентрациях насыщения (фиг 7). В противоположность этому, связывание с очищенным ПСМА в качестве твердой фазы в ИФА было слабым для Е8 и несколько более сильным для А5. Подобный характер просматривался в вестерн-блотах с гликозилированным и дегликозилированным ПСМА (фиг.8). С помощью иммунофлюоресцентной цитологии с клетками LNCaP и детекции путем конфокальной лазерной микроскопии можно показать очень хорошее связывание и интернализацию scFv E8 (фиг.9). Данные по scFv суммированы в таблице 2.
Пример 7
Экспрессия и очистка scFv
Фрагменты scFv экспрессируют в Е.coli XL1-Blue (фирма Stratagene), используя вектор секреции pHOG 21, который содержит последовательности для His-6 и c-myc-tag в С-концевом положении scFv (Kipriganov и др., J.Immunol. Methods, 1997, с.69-77). Бактерии Е.coli, трансформированные плазмидами pHOG, выращивают в течение ночи в среде 2×YT-AG, затем разводят в отношении 1:20 и выращивают в виде культур, объемом 600 мл, при 37°С. При достижении культурами оптической плотности 0,8, бактерии осаждают центрифугированием при 1500 g в течение 10 мин и ресуспендируют в том же объеме свежей среды YT, содержащей 50 мкг/мл ампициллина, 0,4 моля сахарозы и 1 ммоль изопропилтиогалактозид (ИПТГ). После этого рост продолжают при комнатной температуре в течение 18-20 ч. Клетки собирают центрифугированием при 5000 g в течение 10 мин и 4°С. Для выделения растворимых периплазматических белков осажденные бактерии ресуспендируют в 5% начального объема ледяного 50 мМ Трис-HCl, 20% сахарозы, 1 мМ ЭДТА, рН 8,0. После инкубации в течение 1 ч на льду сферопласты центрифугируют при 20000 g при 4°С в течение 30 мин, собирая периплазматический экстракт в супернатанте. Периплазматический экстракт концентрируют, используя мембраны Amicon YM10 с верхней границей отсекаемой молекулярной массы 10 кДа (фирма Amicon, Виттен, Германия), с последующим диализом против 50 мМ Трис-HCl, 1 М NaCl, рН 7,0.
Очистку проводят путем аффинной хроматографии с иммобилизованными катионами металлов. Хроматографик осуществляют, используя колонку объемом 1 мл с хелатообразующей сефарозой (фирма Pharmacia), заряженной Cu2+ и уравновешенной буфером, содержащим 50 ммолей Трис-HCl и 1 моль NaCl, рН 7,0. Загружают периплазматический экстракт, промывают уравновешивающим буфером, в объеме, равном двадцати объемам колонки, содержащим 30 ммолей имидазола и затем элюируют тем же буфером, содержащим 250 ммолей имидазола. Элюированный материал диализируют против ФСБ.
Определение белкового содержимого проводят с использованием набора Micro BCA Protein Reagent Kit (фирма Pierce) в соответствии с инструкциями производителя.
Индукцию синтеза белка получают с помощью ИПТГ и выход scFv составляет примерно 20 мкг от 600 мл культуры Е. coli XL1.
Пример 8
Проточная цитометрия
Из флаконов для тканевых культур собирают свежие клетки LNCaP, DU 145 и РС3 и готовят суспензию однородных клеток в ФСБ с 3% ФТС и 0,1% NaNa. Приблизительно 105 клеток инкубируют с 50 мкл спасенных фагмид, проинкубированных в отношении 1:1 с 2% обезжиренным молоком/ФСБ, в течение 1 ч на льду. После 3-х циклов промывания с использованием ФСБ добавляют 25 мкл/лунку анти-с-myc моноклонального антитела 9Е10 (10 мкг/мл, фирма Becton Dickinson) или, в случае тестирования фагов, 25 мкг/лунку анти-М13 антитела (10 мкг/мл, фирма Pharmacia) и инкубируют 40 мин на льду. После трехкратного промывания ФСБ клетки инкубируют с 100 мкл ФЭ-меченого козьего антимышиного IgG (фирма Becton Dickinson) в течение 40 мин на льду. Затем клетки снова промывают и ресуспендируют в 100 мкл раствора, содержащего 1 мкг/мл иодида пропидиума (фирма Sigma, Deisenhofen) в ФСБ с 3% ФСТ и 0,1% NaN3 для исключения мертвых клеток. Относительную флюоресценцию окрашенных клеток измеряют с использованием проточного цитометра FACScan и прикладной программы CellQuest (фирма Becton Dickinson, Монтейн Вью, Калифорния).
Пример 9
Иммунофлюоресцентная цитология
Клетки LNCaP выращивают на покровных стеклах в течение 24 ч. Для фиксации клетки обрабатывают 2% параформальдегидом в ФСБ в течение 30 мин при комнатной температуре, который не изменяет проницаемость клеточной мембраны, промывают 1% БСА-ФСБ, гасят в течение 10 мин в 50 мМ NH4Cl в ФСБ, и ополаскивают 1% БСА-ФСБ. Добавляют первичное моноклональное антитело в концентрации 4 мкг/мл в 1% БСА-ФСБ и инкубируют в течение 60 мин при 4°С. ФИТЦ-меченое козье антимышиное вторичное антитело (2 мкг/мл, фирма Southern Biotechnology Associates Inc., Бирмингем, США) инкубируют в течение 30 мин и тщательно промывают 1% БСА-ФСБ. Микроскопические препараты хранят в Vectashield (фирма Vector Laboratories, Inc. Берлингейм, Калифорния).
Для исследований интернализации первичное антитело инкубируют в течение 30 мин при 37°С до фиксации клеток 2% параформальдегидом и нарушения проницаемости 0,5% Тритоном X100 в ФСБ.
Пример 10
а) Иммуногистохимия
Заключенные в парафин тканевые срезы сначала депарафинизируют и затем обрабатывают 0,3% Тритоном X100 в ФСБ для восстановления антигена. Замороженные срезы фиксируют в холодном ацетоне. Срезы обрабатывают 30 мин 3% Н2О2 и 10% метанолом для подавления эндогенной пероксидазы. После блокирования с использованием 1% БСА-ФСБ добавляют первичное антитело в концентрации 2 мкг/мл и инкубируют в течение 1 ч при комнатной температуре. Для scFv добавляют вторичное мышиное анти-с-myc антитело и инкубируют в течение 1 ч при комнатной температуре. Затем биотинилированное козье антимышиное антитело инкубируют в течение 30 мин при комнатной температуре и в заключении проявляют АВС-реагентом (продуктом Vectastain).
б) Вестерн-блот анализ
Вестерн-блот анализ проводят с использованием электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН-ПААГ), очищенного ПСМА и лизата клеток LNCaP и переноса на поливинилидендифторидные мембраны. Блоты блокируют в течение ночи в ФСБ, содержащим 5% обезжиренного молока, и инкубируют с очищенными mAb или scFv при концентрации 10 мкг/мл в течение 1 ч. Затем блоты 5 раз промывают в ФСБ-Твине (0,5%) и инкубируют с козьим антимышиным IgG, конъюгированным с пероксидазой хрена, в течение 1 ч при комнатной температуре. После 5-ти промываний в ФСБ-Твине (0,5%) блоты проявляют, используя 3,3′,5′,5′-тетраметилбензидин в качестве субстрата.
Пример 11
Конструирование, экспрессия и очистка scFv-РЕ40-белков
Используемый по настоящему изобретению токсин является усеченным вариантом экзотоксина бактерии Pseudomonas (PE40), у которого отсутствует домен Ia и который содержит только домены Ib, II, и III (Pastan и др., J. Biol. Chem., 1989, ее. 15157-15160). ДНК с кодирующей областью в векторе pSW200 получена от проф. W.Wels, Франкфурт (Wels и др., Biotechnology, 1992, с.1128-1132). Фрагмент ДНК с п.о. в положении от 253 до 613, кодирующий PE40, амплифицируют путем ПЦР с плазмиды pSW200. Затем амплифицированную ДНК лидируют в вектор pHOG-His-scFv в С-концевом положении к scFv, используя сайт рестрикции XbaI. Все этапы клонирования выполняют в соответствии со стандартными методами в Е.coli XL1-blue и продукты контролируют путем секвенирования.
Белковая индукция иммунотоксина и очистка с использованием металл-хелатной хроматографии (IMAC) подобны таковым для scFv. Продукты исследуют и характеризуют путем ДСН-ПААГ, вестерн-блота и проточной цитометрии.
Пример 12
Цитотоксичность иммунотоксинов scFv-PE40
Метаболизм соли тетразолиевого красного WST до водорастворимого формазанового красителя определяют в соответствии с инструкциями производителями (фирма Boehringer). Клетки-мишени (LNCaP и DU 145 в качестве контроля) засевают в количестве 2,5×104/лунку 96-ти луночного планшета и выращивают в течение 24 ч до образования сливающегося слоя клеток. Добавляют различные концентрации рекомбинантных иммунотоксинов в аликвотах 50 мкл/лунку и инкубируют планшеты в течение 48 ч при 37°С в атмосфере 5% CO2. По истечении этого времени культуры перемешивают с 15 мкл/лунку реагента WST и инкубируют в течение 90 мин при 37°С в атмосфере 5% CO2. Затем на спектрофотометре проводят оценку оптической плотности образцов при длине волны 450 нм (контроль 690 нм). Проводят расчет концентрации иммунотоксина, необходимой для 50% снижения жизнеспособности клеток относительно таковой необработанных контрольных культур (50% ингибиторной концентрации = IC50).
Цитотоксические анализы (WST) с иммунотоксинами Е8-Р40 и А5-Р40 проводят с клетками LNCaP, экспрессирующими ПСМА, и контрольными клетками DU 145. На фиг.11 показано, что высокий цитотоксический эффект можно наблюдать с иммунотоксином Е8-РЕ40 на клетках LNCaP, при этом значение IC50 составляет 0,05 нМ. На фиг.12 показан цитотоксический эффект иммунотоксина А5-РЕ40, при этом IC50 составляет примерно 0,09 нМ. Цитотоксический фон на клетках DU 145, не экспрессирующих ПСМА, составляет 5% для Е8-конструкции и только 0,01% для А5-конструкции, свидетельствуя об очень хорошем терапевтическом окне.
Пример 13
Получение scFv H12 и D7 из mAb 3/F11 и 3/Е7
Из каждого mAb по описанию в примере 5 получают библиотеку экспрессии scFv в фагмиде pSEX.
Получение и селекцию фага, экспонирующего антитело, выполняют согласно примеру 6.
После шести циклов пэннинга в разных вариантах на ПСМА и клетках LNCaP из mAB 3/Е7 получен один специфический позитивный клон, названный H12, а из mAB 3/F11 получен один позитивный клон, названный D7. Кодирующую область каждого scFv переносят в вектор экспрессии pHOG-21.
Экспрессию и очистку scFv выполняют по описанию в примере 7.
Пример 14
Характеристика scFv H12 и D7
а) Проточная цитометрия на ПСМА-положительных и ПСМА-отрицательных клеточных линиях
Согласно оценке путем проточной питометрии scFvs H12 и D7 взаимодействуют с жизнеспособными клетками LNCaP.
На основании кривых насыщения установлено, что концентрация антител, достигающая 50% насыщения участков ПСМА, составляет приблизительно 120 нМ (H12) и 20 нМ (D7) соответственно. При концентрациях насыщения достигаются значения СИФ, составляющие 70 (Н12) и 40 (D7) (фиг.15).
Чтобы оценить ПСМА-связывающую специфичность scFv H12 и D7 дополнительно окрашивают и анализируют путем проточной цитометрии ПСМА-отрицательные раковые клетки простаты DU145 и РС3, а также другие ПСМА-отрицательные клеточные линии (HeLa, MCF7, НСТ15 и Jurkat). Ни один из двух scFv не окрашивает ПСМА-отрицательные клетки.
б) Проточная цитометрия на трансфектантах ПСМА
Для подтверждения ПСМА-специфического связывания scFv H12 и D7 также исследуют на клетках BOSC-23, трансфецированных ПСМА. Оба scFv показывают зависимое от концентрации связывание с клетками BOSC, трансфецированными ПСМА полной длины, но не связываются с нетрансфецированными клетками (фиг.16). Условия насыщения достигаются при 100 нМ (D7) и 200 нМ (H12). Подобно mAb, значения СИФ на трансфектанах ниже, чем на клетках LNCaP, и показывают широкий разброс, который может соответствовать изменению молекул ПСМА на поверхности трансфектантов.
в) Иммунофлюоресцентная цитология
Иммунофлюоресцентную цитологию проводят согласно описанию в примере 4. При детекции с помощью лазерного сканирующего конфокального микроскопа наблюдают сильное связывание scFv с клетками LNCaP, а также интернализацию в эти клетки.
г) ИФА и Вестерн-блоттинг
Связывание scFv H12 и D7 с очищенным ПСМА выражено слабо в ИФА и Вестерн-блоттинге.
Последовательности (аминокислот и нуклеиновых кислот) Н12 и D7 приведены на фиг.20 и фиг.21.

Пример 15
Конструирование и цитотоксичность иммунотоксина Н12-РЕ40 и иммунотоксина D7-PE40
Конструирование иммунотоксинов Н12-РЕ40 и D7-PE40 проводят по аналогии с иммунотоксинами А5 и Е8 согласно описанию в примере 11. РЕ-40 представляет фрагмент экзотоксина бактерии Pseudomonas.
Цитотоксичность исследуют согласно описанию в примере 12.
Иммунотоксин стимулировал гибель клеток LNCaP зависящим от времени образом, причем наивысшую цитотоксичность можно наблюдать после 48 ч инкубации.
В это время значения IC50 примерно составляющие 200 пМ обнаружены для Н12-РЕ40 и D7-PE40 (фиг.17).
Дополнительно исследуют цитотоксичность Н12-РЕ40 и D7-PE40 на ПСМА-отрицательных клеточных линиях DU 145, PC-3, MCF7 и НСТ 15. На этих клеточных линиях цитотоксичности не обнаружено при концентрациях до 25000 пМ.
Пример 16
Конструирование анти-ПСМА/CD3 двойного антитела
Производят биспецифическое двойное антитело, обладающее специфичностью к ПСМА и цепи CD3 Т-клеточного рецепторного комплекса. Биспецифическое двойное антитело экспрессируют в Е. coli, используя вектор, содержащий дицистронный оперон для косекреции scFv VhCD3-VlA5 и VhA5-V1CD3 (фиг.18). Для конструирования анти-А5/CD3 двойного антитела используют плазмиды pKID19×3 и pKID 3×19 (Kipriyanov, Int. J.Cancer, 1998, с.763). Бактериальная периплазматическая экспрессия и очистка аналогичны этим показателям scFv.
Пример 17
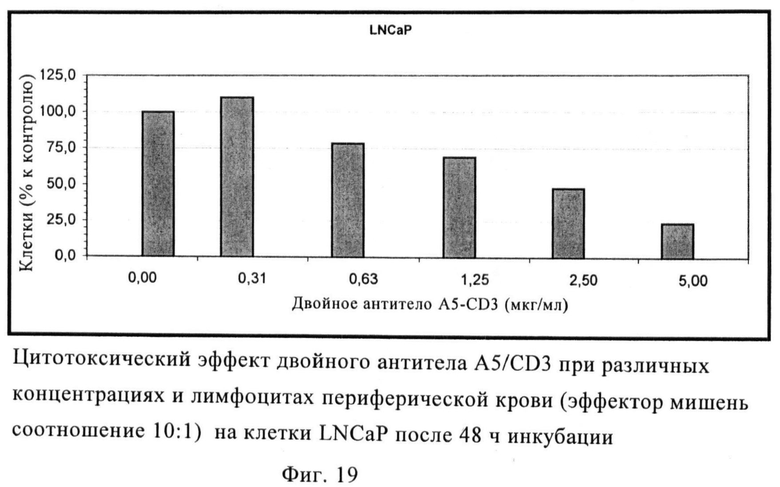
Индуцирование специфической токсичности двойным антителом A5-CD3
Способность биспецифического двойного антитела индуцировать лизис опухолевых клеток путем переадресации опосредованной Т-клетками цитотоксичности исследуют с использованием мононуклеарных клеток периферической крови (МКПК) от здоровых доноров в качестве клеток-эффекторов. После инкубации с ИЛ-2 или без него в течение 4 суток клетки добавляют к клеткам-мишеням LNCaP, которые засеяны при плотности 1,5×104 клеток/лунку в 96-ти луночный планшет. Отношение эффектор-мишень составляет 10:1. Двойное антитело добавляют в различных концентрациях. После инкубации в течение 48 ч культуры перемешивают с 15 мкг/лунку реагента WST и инкубируют в течение 90 мин при 37°С в атмосфере 5% CO2. Затем путем спектрофотометрии оценивают оптическую плотность образцов при длине волны 450 нм (контроль 690 нм).
В этом тесте in vitro двойное антитело, по-видимому, обладает явной способностью в переадресации активированных и неактивированных МКПК вызывать лизис клеток-мишеней LNCaP зависящим от концентрации образом (фиг.19).
Пример 18
Двойное антитело А5-А5
Это бивалентное моноспецифическое двойное антитело получают аналогично получению двойного антитела A5-CD3 (пример 16). Бактериальная периплазматическая экспрессия и очистка аналогичны экспрессии и очистке scFv.
Путем проточной цитометрии можно показать сильное и специфическое связывание двойного антитела А5-А5 с клетками LNCaP.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К НЕКТИНУ-4 ЧЕЛОВЕКА | 2019 |
|
RU2825839C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ СПЕЦИФИЧЕСКИЙ МЕМБРАННЫЙ АНТИГЕН ПРОСТАТЫ, И СООТВЕТСТВУЮЩИЕ КОМПОЗИЦИИ И СПОСОБЫ | 2012 |
|
RU2632647C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, А ИМЕННО АНТИТЕЛА, СПОСОБНЫЕ СВЯЗЫВАТЬСЯ С L1CAM (CD171) | 2015 |
|
RU2754684C2 |
| АНТИТЕЛО ПРОТИВ КЛАУДИНА 18А2 И ЕГО ПРИМЕНЕНИЕ | 2017 |
|
RU2793445C2 |
| ПОЛИПЕПТИДЫ АНТИТЕЛ И ИХ ПРИМЕНЕНИЕ | 2016 |
|
RU2753677C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ПРОТИВ СПЕЦИФИЧЕСКОГО МЕМБРАННОГО АНТИГЕНА ПРОСТАТЫ (PSMA) И СПОСОБ ИНГИБИРОВАНИЯ РОСТА КЛЕТОК, ЭКСПРЕССИРУЮЩИХ PSMA | 2006 |
|
RU2421466C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К СD52 | 2012 |
|
RU2605307C2 |
| АНТИТЕЛА К РОДСТВЕННОЙ РАКОВО-ЭМБРИОНАЛЬНОМУ АНТИГЕНУ МОЛЕКУЛЕ КЛЕТОЧНОЙ АДГЕЗИИ (СЕАСАМ) | 2012 |
|
RU2650869C2 |
| АНТИТЕЛО ПРОТИВ CSF-1R | 2009 |
|
RU2547586C2 |
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
























Настоящее изобретение относится к иммунологии и биотехнологии. Предложены варианты: моноклонального антитела против ПСМА и его антигенсвязывающего фрагмента, которые связаны с меткой или цитотоксическим агентом. Описаны варианты фармацевтической композиции и диагностического набора на основе таких антител. Раскрыты способы идентификации in vitro опухолевых клеток, а также диагностической идентификации опухолевых клеток на основе таких антител. Описан изолированный полинуклеотид для получения моноклональных антител. Использование изобретения обеспечивает антитела, которые могут связывать ПСМА в его нативной форме на поверхности опухолевых клеток, связываются с LNCAP, но не связываются с клетками, у которых утрачена экспрессия ПСМА, что может найти дальнейшее применение в терапии рака простаты. 11 н. и ф-лы, 4 з.п. ф-лы, 21 ил., 3 табл., 18 пр.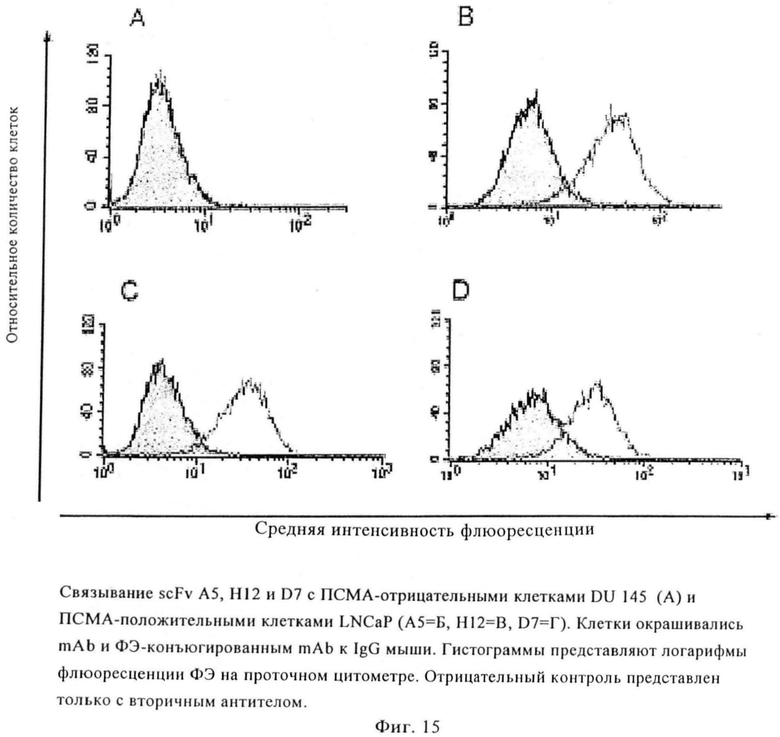
1. Изолированное моноклональное антитело (mAb) или его антигенсвязывающий фрагмент, которые
а) связываются со специфическим для простаты мембранным антигеном (ПСМА) в его нативной форме, присутствующим на поверхности опухолевых клеток,
б) обладают способностью подвергаться интернализации опухолевой клеткой,
в) сильно связываются с клетками LNCAP, но не связываются с клетками, у которых утрачена экспрессия специфического для простаты мембранного антигена,
г) связаны с меткой или цитотоксическим агентом, отличающиеся тем, что
д) они включают аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO:1, SEQ ID NO:10, SEQ ID NO:20 и/или SEQ ID NO:22.
2. Изолированное моноклональное антитело или его антигенсвязывающий фрагмент по п.1, отличающиеся тем, что моноклональное антитело или его антигенсвязывающий фрагмент обладают ФЭ степенью интенсивности флюоресценции (СИФ), которая превышает 1000, и ФЭ степенью интенсивности флюоресценции (СИФ) scFv, которая превышает 40 при насыщении антигена.
3. Изолированное моноклональное антитело или его антигенсвязывающий фрагмент по п.1 или 2, которые показывают высокую активность связывания с клетками LNCAP, при этом 50% насыщение ПСМА-участков достигается при концентрациях 1 нМ-120 нМ.
4. Изолированное моноклональное антитело или его антигенсвязывающий фрагмент по п.1, отличающиеся тем, что меткой является частица, которая испускает радиоактивное или флюоресцентное излучение.
5. Изолированное моноклональное антитело или его антигенсвязывающий фрагмент по п.1, отличающиеся тем, что цитотоксический агент является токсическим для клетки веществом, выбранным из группы, состоящей из таксола, фрагмента экзотоксина Pseudomonas, цитохалазина В, грамицидина D, бромида этидия, эметина, митомицина, этопсида, тенопсида, винкристина, винбластина, колхицина, доксорубицина, даунорубицина, дигидроксиантрацендиона, митоксантрона, митрамицина, актиномицина D, 1-дегидротестостерона, гликокортикоидов, прокаина, тетракаина, лидокаина, пропранолола и/или пуромицина.
6. Изолированное моноклональное антитело или его антигенсвязывающий фрагмент, которые
а) связываются со специфическим для простаты мембранным антигеном (ПСМА) в его нативной форме, присутствующим на поверхности опухолевых клеток,
б) обладают способностью подвергаться интернализации опухолевой клеткой,
в) связываются с клетками LNCAP, но не связываются с клетками, у которых утрачена экспрессия специфического для простаты мембранного антигена, и
г) связаны с меткой или цитотоксическим агентом и отличаются тем, что
д) включают аминокислотную последовательность SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6 и SEQ ID NO:7 или включают аминокислотную последовательность SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:15 и SEQ ID NO:16, или CDRs, обозначенные CDR H1, H2, H3, L1, L2 и L3, так как показано на фиг.20, или соответствующие CDR области, как показано на фиг.21.
7. Фармацевтическая композиция для лечения рака простаты, включающая эффективное количество изолированного моноклонального антитела или его антигенсвязывающего фрагмента по п.1.
8. Фармацевтическая композиция для лечения рака простаты, включающая эффективное количество изолированного моноклонального антитела или его антигенсвязывающего фрагмента по п.6.
9. Диагностический набор для выявления рака простаты, включающий изолированное моноклональное антитело или его антигенсвязывающий фрагмент по п.1.
10. Диагностический набор для выявления рака простаты, включающий изолированное моноклональное антитело или его антигенсвязывающий фрагмент по п.6.
11. Способ идентификации in vitro опухолевых клеток, отличающийся тем, что клетки подвергаются контакту с изолированным моноклональным антителом или его антигенсвязывающим фрагментом по п.1 и идентифицируют клетки, которые связывают указанное изолированное моноклональное антитело или его антигенсвязывающий фрагмент как опухолевые клетки.
12. Способ идентификации in vitro опухолевых клеток, отличающийся тем, что клетки подвергаются контакту с изолированным моноклональным антителом или его антигенсвязывающим фрагментом по п.6 и идентифицируют клетки, которые связывают указанное изолированное моноклональное антитело или его антигенсвязывающий фрагмент как опухолевые клетки.
13. Способ для диагностической идентификации опухолевых клеток, отличающийся тем, что клетки, подлежащие идентификации, подвергаются контакту с изолированным моноклональным антителом или его антигенсвязывающим фрагментом по п.1 и идентифицируют клетки, которые связывают указанное изолированное моноклональное антитело или его антигенсвязывающий фрагмент как опухолевые клетки.
14. Способ диагностической идентификации опухолевых клеток, отличающийся тем, что клетки, подлежащие идентификации, подвергаются контакту с изолированным моноклональным антителом или его антигенсвязывающим фрагментом по п.6 и идентифицируют клетки, которые связывают указанное изолированное моноклональное антитело или его антигенсвязывающий фрагмент как опухолевые клетки.
15. Изолированный полинуклеотид, выбранный из группы, состоящей из SEQ ID NO:8, 9, 17, 18, 19, 21, 23 и 24 и их соответствующей комбинации, для получения моноклональных антител или их связывающих фрагментов по п.1 или 6.
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| FRACASSO et al., "Anti-tumor effects of toxins targeted to the prostate specific membrane antigen", The prostate, v.53, №1, 2002, с.9-23 | |||
| RU 99118510 A, 10.06.2001. | |||

