Настоящее изобретение относится к применению антагонистов лептина для лечения резистентности к инсулину при диабете II типа и к лекарственному средству для лечения такой резистентности.
Диабет является одним из наиболее распространенных заболеваний обмена веществ в промышленно развитых странах. Во всем мире диабетом страдает около 110 миллионов человек; в то время как приблизительно 10 миллионов из них имеет диабет I типа, подавляющее большинство (приблизительно 100 миллионов) страдает диабетом II типа. Это заболевание вызывается нарушением метаболизма глюкозы. При диабете I типа поражение β-клеток поджелудочной железы приводит к прекращению выработки инсулина. Это отсутствие инсулина приводит к повышению содержания глюкозы в крови и, если не проводить заместительное лечение инсулином, к кетоацидозу, диабетической коме или смерти пациента. При диабете II типа причинные взаимоотношения различны и характеризуются первичным развитием резистентности к инсулину, т.е. к уменьшению способности клеток адекватно реагировать на инсулин. Избыточный вес и недостаточная физическая активность, в частности, считаются причинами появления резистентности к инсулину. Последнее состояние сначала остается незамеченным, так как компенсируется избыточной секрецией инсулина. Однако длительно существующая резистентность к инсулину приводит в результате процесса, длящегося много лет, к декомпенсации этого эндогенного механизма и последующему развитию диабета II типа. В то время как диета и физическая активность могут замедлить развитие этих последовательных явлений, предупредить проявление заболевания они зачастую не могут. В таких случаях для адекватного контроля содержания глюкозы в крови требуется медикаментозное лечение.
Ключевым моментом долгосрочной успешной терапии является поддержание глюкозы в крови в пределах физиологических значений при минимальных колебаниях. В настоящее время считают, что повышенные в течение десятилетий уровни глюкозы, как это происходит при плохом контроле диабета (как I, так и II типа), играют важную роль в развитии поздних осложнений диабета. Эти поздние осложнения заключаются, в частности, в повреждении кровеносных сосудов, что приводит к заболеванию почек, потере зрения и сердечно-сосудистым заболеваниям. Это так называемое позднее поражение является важным фактором, который вносит свой вклад в смертность при диабете.
В 1994 г. был описан новый гормон, лептин, который образуется в жировых клетках и отсутствует у мышей с наследственным ожирением (мыши ob/ob) (Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L., and Friedman, J. M. (1994). Positional cloning of the mouse obese gene and its human homologue. Nature 372, 425-432). Человеческий лептин и мышиный лептин в большой степени идентичны. Инъецирование мышам ob/ob рекомбинантного лептина приводит к сокращению потребления пищи и снижению веса (Pelleymounter, M.A., Cullen, M. J., Baker, M.B., Hecht, R., Winters, D., Boone, Т., and Collins, F. (1995). Effects of the obese gene product on body weight regulation in ob/ob mice. Science 269, 540-543). До настоящего времени не существует указаний на то, что мутации гена ob могут быть ответственны за распространенность ожирения у людей (в США приблизительно 30% популяции имеют выраженный избыточный вес). Систематические исследования показали, что уровни сывороточного лептина у тучных людей повышены; то же наблюдается и на различных экспериментальных животных моделях ожирения (Dagogo-Jack, S., Fanelli, С., Paramore, D., Brothers, J., and Landt, M. (1996). Plasma leptin and insulin relationships in obese and nonobese humans. Diabetes 45, 695-698; Considine, R.V., Sinha, M.K., Heiman, M.L., Kriauciunas, A., Stephens, T.W., Nyce, M.R. , Ohannesian, J. P., Marco, C.C., McKee, L.J., Bauer, T.L., and Caro, J.F. (1996). Serum immunoreactive leptin concentrations in normal-weight and obese humans. N. Engl. J. Med. 334, 292-295). По этой причине были сделаны выводы о том, что лептин является сигналом обратной связи, который информирует головной мозг о количестве энергии, которая хранится в жировой ткани. Согласно этому выводу, именно головной мозг осуществляет функцию снижения потребления пищи путем подавления аппетита, с одной стороны, и стимуляции основного обмена, с другой. При ожирении у человека эта регуляторная цепочка, как полагают, прерывается.
Помимо этого полагают, что лептин действует также непосредственно на ткани вне головного мозга.
К настоящему времени опубликованы результаты трех исследований, касающихся прямого действия лептина на изолированные клетки:
Kroder et al. (Kroder, G. , Kellerer, M., and  H. (1996) Exp. Clin. Endocrin. Diabetes 104 Suppl. 2, 66, (Abstract)) высказали предположение о том, что лептин устанавливает связь между резистентностью к инсулину и ожирением, и, сообщают о том, что лептин снижает индуцированное инсулином фосфорилирование инсулиновых рецепторов и субстрата инсулиновых рецепторов 1 (IRS-1) фибробластов крыс 1, которые избыточно экспрессируют рецепторы инсулина человека. Степень, в которой лептин также оказывает влияние на конечный результат действия инсулина, например, стимуляцию транспорта глюкозы или синтез гликогена, не исследовалась и не обсуждалась в литературе.
H. (1996) Exp. Clin. Endocrin. Diabetes 104 Suppl. 2, 66, (Abstract)) высказали предположение о том, что лептин устанавливает связь между резистентностью к инсулину и ожирением, и, сообщают о том, что лептин снижает индуцированное инсулином фосфорилирование инсулиновых рецепторов и субстрата инсулиновых рецепторов 1 (IRS-1) фибробластов крыс 1, которые избыточно экспрессируют рецепторы инсулина человека. Степень, в которой лептин также оказывает влияние на конечный результат действия инсулина, например, стимуляцию транспорта глюкозы или синтез гликогена, не исследовалась и не обсуждалась в литературе.
Было показано, что чувствительность к липогенным гормонам (дексаметазону и инсулину) понижена у трансформированных предшественников адипоцитов 30А5, которые избыточно экспрессируют лептин (Bai, Y.L., Zhang, S.Y., Kim, K.S., Lee, J.K., and Kim, K.H. (1996) J. Biol. Chem. 271, 13939-13942). Синтез жирных кислот и синтез нейтральных жиров понижались при избыточной экспрессии лептина даже без стимуляции. В то время как контрольные клетки демонстрировали выраженное повышение скорости синтеза липидов после обработки дексаметазоном или инсулином, или комбинацией этих двух гормонов, клетки, которые экспрессировали избыточное количество лептина, с трудом стимулировались при этих обстоятельствах. Помимо этого, было также проведено исследование, которое касалось ингибирования активности глицерофосфат-дегидрогеназы и экспрессии ацетил-КоА-карбоксилазы, что наблюдается после обработки клеток комбинацией дексаметазона и инсулина. Было установлено, что невозможно стимулировать клетки, экспрессирующие лептин. Наблюдавшиеся эффекты указывают, что лептин подавляет жировой обмен в целом. Упоминания о какой-либо возможной связи между ожирением и резистентностью к инсулину не было.
В другой экспериментальной системе, т. е. на мышечных трубочках мыши C2C12, было показано, что лептин вызывает инсулиноподобные эффекты (Berti, L. , Kellerer, M. , and  H. (1996) Diabetologia 39 Suppl. 1, A59 (Abstract)). Это исследование показало, что как транспорт глюкозы, так и синтез гликогена стимулируются лептином. Эти наблюдения вступают в противоречие с данными других авторов и с результатами, представленными в настоящем документе. Возможно, они имели дело со специфичным для этого типа клеток эффектом.
H. (1996) Diabetologia 39 Suppl. 1, A59 (Abstract)). Это исследование показало, что как транспорт глюкозы, так и синтез гликогена стимулируются лептином. Эти наблюдения вступают в противоречие с данными других авторов и с результатами, представленными в настоящем документе. Возможно, они имели дело со специфичным для этого типа клеток эффектом.
В наших исследованиях, посвященных действию лептина на изолированные адипоциты крысы, экспериментальную модель жировой ткани, в настоящее время обнаружено, что чувствительность к инсулину важных метаболических путей жировой клетки, таких как стимуляция липогенеза, транспорт глюкозы и гликогенез, резко снижается (пример 4), в то время как базальные значения остаются без изменений. То же относится и к ингибированию липолиза, стимулированного изопротеренолом. Транспорт глюкозы в изолированных крысиных адипоцитах добавлением инсулина (10 нМ) стимулируется приблизительно в 14 раз. Эта способность к стимуляции снижается дозозависимым образом путем предварительной инкубации с лептином в различных концентрациях в течение 15 часов. Лептин десенсибилизирует клетки, т. е. вырабатывается резистентность к инсулину. Кривые доза-эффект для инсулина при различных концентрациях лептина (пример 5) показывают, что концентрации, при которых in vitro уже можно выявить эффекты, что касается как инсулина (0,1-0,2 нМ), так и лептина (0,5-1 нМ), находятся в пределах физиологических значений (Dagogo-Jack et al., 1996; Considine et al., 1996). Более высокие уровни лептина (2-4 нМ) (Dagogo-Jack et al. , 1996; Considine et al., 1996) обнаружены у тучных людей, поэтому возможно, что у этих людей эффект инсулина нарушен сильнее. Эти выводы предполагают, что хронически повышенный уровень лептина, как это можно видеть у тучных субъектов, приводит к развитию инсулиновой резистентности. Как уже объяснялось выше, резистентность к инсулину является важным фактором патогенеза диабета II типа.
Таким образом, целью настоящего изобретения является обеспечение новых антагонистов лептина, из которых можно составлять фармацевтические композиции. Другой целью настоящего изобретения является обеспечение способов лечения диабета II типа и других нарушений, связанных с инсулином.
Настоящее изобретение, следовательно, связано с применением антагонистов лептина, в частности, тех из них, которые получены из самого лептина, для изготовления лекарственного средства для применения при диабете II типа. Антагонисты лептина для этого применения более подробно описаны ниже.
Согласно первой цели этого изобретения обеспечены фармацевтические композиции, которые включают антагонист лептина. Согласно той же цели раскрываются фармацевтические композиции, которые включают антагонисты лептина, полученные из самого лептина. Далее, согласно той же цели, раскрываются фармацевтические композиции, которые включают антагонист лептина, который является растворимым рецептором лептина или его производным.
Согласно второй цели этого изобретения, обеспечены способы, которые используют изобретенные фармацевтические композиции для лечения диабета II типа. Также согласно той же цели, обеспечены способы восстановления или усиления физиологического эффекта инсулина.
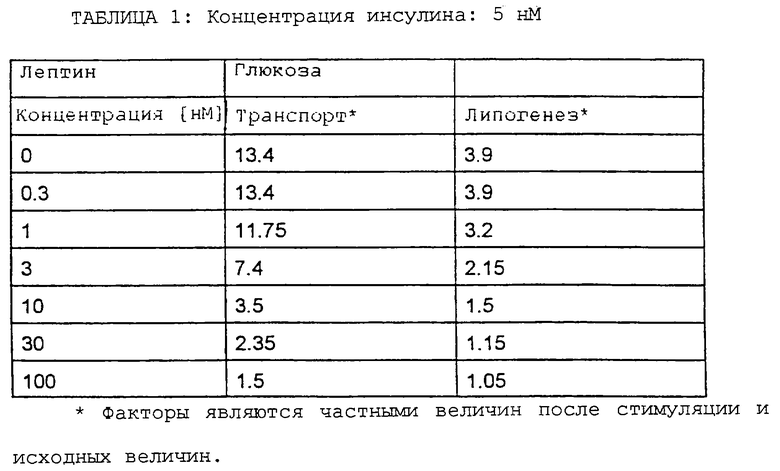
Таблица 1: Ингибирование индуцированного инсулином транспорта глюкозы лептином в адипоцитах крысы. Этот эксперимент подробно описан в примере 8.
Таблица 2: Захват 2-дезоксиглюкозы адипоцитами крысы в зависимости от концентрации лептина. Этот эксперимент подробно описан в примере 9.
Таблица 3: Антагонизм в отношении эффекта лептина со стороны фрагмента 116-167 лептина на примере индуцированного инсулином транспорта глюкозы в адипоцитах крысы. Этот эксперимент подробно описан в примере 10.
Таблица 4: Аминокислотная последовательность лептина человека. Два остатка цистеина связаны дисульфидным мостиком.
Таблица 5: Аминокислотная последовательность лептина мыши. Два остатка цистеина связаны дисульфидным мостиком.
Таблица 6: Аминокислотная последовательность фрагмента 116-167 лептина мыши. Два остатка цистеина связаны дисульфидным мостиком.
Изобретение, представленное в настоящей заявке, направлено на композиции и способы снижения или полного устранения инсулиновой резистентности путем ингибирования эффекта лептина. Для этой цели можно применять пептиды, которые действуют как антагонисты лептина, что приводит in vitro в изолированных жировых клетках к устранению индуцированной лептином инсулиновой резистентности. Эти пептиды, таким образом, удобны для лечения инсулиновой резистентности, предпочтительно у тучных пациентов.
АНТАГОНИСТЫ ЛЕПТИНА
Антагонисты лептина согласно изобретению включают, конкретно, пептидные антагонисты. Указанные пептиды являются производными фрагментами лептина и могут быть получены, например, путем химического или ферментативного расщепления интактного лептина (например, с помощью лизилэндопептидазы, трипсина, эндо-Аrg С или цианогенбромида) или путем экспрессирования, прямо или в форме слитого белка, микроорганизмами. В связи с получением этих пептидов с использованием микроорганизмов нет необходимости полагаться на наличие натуральных сайтов расщепления при выборе фрагментов, которые предназначены для экспрессирования. Для получения этих пептидов удобно использовать лептин человека или животных, например, лептин крыс, свиней или человекообразных обезьян.
Один подходящий в качестве примера пептид охватывает с 116 по 167 аминокислоту или с 116 по 166 аминокислоту (таблица 6, SEQ ID NO: 4), согласно последовательности, которая была опубликована Zhang et al., (1994) (примеры 3 и 6). В примере 6 адипоциты инкубировали в течение приблизительно 15 часов в присутствии 10 нМ лептина и различных концентраций антагонистического фрагмента лептина 116-167. Эти клетки затем стимулировали 5 нМ инсулином. В этом эксперименте было показано, что повышение количества антагониста приводит к восстановлению способности отвечать на стимуляцию инсулином, и в присутствии высоких концентраций антагониста лептина резистентность не развивается. Таким образом, используя эти или подобные исследования, специалист может легко убедиться в наличии антагонистических свойств у любого антагониста лептина, который может быть пригоден для использования согласно настоящему изобретению.
Помимо этого можно использовать также аналогию антагонистических фрагментов лептина, в которых имеет место одна или более аминокислотных замен или пропусков. Предпочтительные замены являются обычными аминокислотными заместителями. Такие обычные заместители включают заряженные-заряженные, полярные-полярные и гидрофобные-гидрофобные аминокислотные заместители. Например, один или более остатков аспарагиновой кислоты могут быть заменены остатками глутаминовой кислоты и/или наоборот, и/или один или более остатков лейцина могут быть заменены остатками изолейцина и/или наоборот. Другие замены и пропуски могут быть рационально произведены на основе стереомерных или структурных соображений, таких как размер аминокислот и склонность образовывать или нарушать спиральную структуру.
Для изменения и оптимизации антагонистических свойств указанных пептидов определенным образом можно применять методы молекулярной биологии и биотехнологии. Помимо этого, эти пептиды можно модифицировать химически, например, посредством ацетилирования, карбамоилирования, формилирования, биотинилирования, ацилирования или дериватизации с использованием полиэтиленгликоля или гидрофильных полимеров, с целью повышения их стабильности или модулирования их периода полужизни в плазме и фармакокинетики.
Антитела против лептина, в частности, их лептинсвязывающие домены также пригодны в качестве антагонистов лептина для указанных целей. Помимо этого, растворимые рецепторы лептина и/или фрагменты рецепторов лептина и их слитые с другими белками формы (например, с участком Fc IgG) также пригодны. Подобно антагонистам пептидной природы, любой антагонист лептина, который является белком, может быть изменен молекулярно-биологическими средствами. Подобно этому, эти антагонисты могут изготавливаться путем экспрессирования, прямо или в форме слитого белка, микроорганизмами или целым рядом стандартных экспрессирующих систем.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ
Настоящее изобретение также относится к лекарственному средству, которое включает антагонисты лептина, описанные в настоящей патентной заявке.
Эти лекарственные средства могут использоваться, например, в форме фармацевтических препаратов, которые можно вводить перорально, например, в форме таблеток, таблеток в оболочке, твердых или мягких желатиновых капсул, растворов, эмульсий или суспензий. Их можно также вводить ректально, например, в форме суппозиториев или парентерально, например, в форме растворов для инъекций. Эти лекарственные средства могут также вводиться через слизистые оболочки носа, рта или через легкие. Чтобы изготовить эти фармацевтические препараты, эти соединения можно помещать в терапевтические инертные, органические и неорганические, наполнители. Лактоза, кукурузный крахмал или его производные, тальк и стеариновая кислота или ее соли являются примерами таких наполнителей для таблеток, таблеток в оболочке и твердых желатиновых капсул. Вода, многоатомные спирты, сахароза, инвертированный сахар и глюкоза являются удобными наполнителями для приготовления растворов. Вода, одноатомные и многоатомные спирты, глицерин и растительные масла являются удобными наполнителями для приготовления растворов для инъекций. Растительные и отвержденные масла, воски, жиры и полутвердые многоатомные спирты являются удобными наполнителями для приготовления суппозиториев. Фармацевтические препараты могут также включать консерванты, растворители, стабилизаторы, увлажняющие агенты, эмульгаторы, подсластители, красители, корригенты, соли для изменения осмотического давления, буферы, покрывающие агенты, антиоксиданты и, если это необходимо, другие терапевтические активные соединения.
Предпочтительно пероральное введение и инъекции. Для инъекций новые антагонисты лептина приготавливают в виде жидкого раствора, предпочтительно в физиологически приемлемом буфере, таком как раствор Хенка или раствор Рингера. Однако новые антагонисты лептина могут изготовляться также в твердой форме для растворения или суспендирования непосредственно перед использованием.
Типичные композиции содержат терапевтически благоприятное количество антагониста лептина. Терапевтически благоприятное количество может быть тем же, что и терапевтически эффективное количество, как обсуждается ниже. Помимо этого, терапевтически благоприятное количество может представлять собой стандартную дозу, которая одна или в количестве несколько штук может использоваться для обеспечения терапевтически эффективного количества антагониста лептина. Таким образом, терапевтически благоприятное количество будет зависеть, помимо прочего, от природы заболевания, подвергаемого лечению.
СПОСОБЫ ЛЕЧЕНИЯ
Способы настоящего изобретения пригодны для лечения любого состояния, при котором вовлечено действие лептина. С учетом настоящего описания и наблюдения того факта, что лептин ингибирует некоторые физиологические эффекты инсулина, изобретенные способы являются особенно пригодными для лечения состояний, при которых имеются нарушения активности инсулина, и особенно для лечения диабета II типа. Такие нарушения активности инсулина включают изменения в липогенезе, транспорте глюкозы, гликогенезе и липолизе. Соответственно, способы настоящего изобретения включают способы лечения диабета II типа и способы восстановления или усиления физиологических эффектов инсулина.
Типичный способ включает введение пациенту, нуждающемуся в подобном лечении, терапевтически эффективного количества антагониста лептина. Пациент будет нуждаться в таком лечении, если он страдает от состояния, при котором вовлечено действие лептина. Это лечение особенно показано, если пациент страдает диабетом II типа. Это лечение также показано, если пациент страдает от состояния, при котором нарушена активность инсулина, такого как изменения в липогенезе, транспорте глюкозы, гликогенезе и липолизе. Для лечения состояний, при которых имеет место такое нарушение, способы восстановления или усиления физиологических эффектов инсулина являются полезными.
Терапевтически эффективное количество будет зависеть, например, от природы состояния, подвергаемого лечению, способа введения, конкретных свойств выбранного антагониста и особенно от оценки лечащего врача. Терапевтически эффективное количество обычно является количеством, достаточным для эффективного лечения нужного состояния или для выполнения заявленной цели способа, например, для восстановления или усиления физиологических эффектов инсулина. В конечном счете терапевтически эффективное количество будет зависеть от клинически оцененной эффективности и токсичности каждого из антагонистов лептина. Такие оценки производятся обычным порядком и хорошо известны клиницистам.
Термин "лечение", в его различных грамматических формах, в отношении настоящего изобретения касается предупреждения, лечения, достижения обратного развития, ослабления, облегчения, минимизации, подавления или прекращения вредных эффектов болезненного состояния, прогрессирования заболевания, причинного агента заболевания или другого патологического состояния.
Дозы, предпочтительные для системного введения, составляют приблизительно от 0,01 мг/кг до 50 мг/кг веса тела в день.
Настоящее изобретение ниже иллюстрируется, но не ограничивается таблицами и примерами.
ПРИМЕРЫ
ПРИМЕР 1: Клонирование лептина мыши
Выделение РНК - У взрослых мышей удаляли эпидидимальные жировые подушечки и подвергали быстрому замораживанию в жидком азоте. 1 г жировой ткани помещали в ступку в жидком азоте, после чего добавляли 15 мл 5 М раствора гуанидина тиоцианата в 50 мМ Трис-буфера (рН 7,5), 10 мМ ЭДТА и 0,1 ДТТ, и все вместе интенсивно гомогенизировали до получения тонкой дисперсии. После полного измельчения тканевых частиц добавляли 10 г твердого CsCl и смесь перемешивали при комнатной температуре. После добавления 10 мл H2O на 9 мл 5,7 М раствора CsCl наслаивали 25 мл этого раствора в центрифужной пробирке. После центрифугирования в течение 15 часов в роторе SW28 при 25 000 об/мин (18oС) пробирки подвергали глубокому замораживанию в жидком азоте, и нижнюю четверть пробирки отсекали с помощью горячего лезвия скальпеля; замороженное содержимое извлекали и соскабливали осадок РНК с донышка. РНК растворяли, а затем осаждали этанолом.
Синтез кДНК - В смеси 1 мкг общей РНК из жировой ткани и 1 мкг специфичного олигонуклеотида-праймера 5'-GAATGCAGAATAAATAAATA (SEQ ID NO.: 1; Zhang et al. , 1994) растворяли 10 мкл H2O, а затем денатурировали нагреванием и инкубировали при 65oС в течение 5 минут. После добавления 0,5 мкл ингибитора РНК-азы, в каждом случае 5 нмоль дНТФ и 0,5 мкл обратной транскриптазы вируса миелобластоза птиц (Boehringer Mannheim), смесь инкубировали при 42oС в течение 1 часа. После этого объем кДНК доводили до 200 мкл водой и хранили при минус 20oС.
ПЦР (полимеразная цепная реакция) - 3 мкл специфично праймированной кДНК амплифицировали 0,5 мкг каждого из двух праймеров 5'-CAAAGAAGGATCCAGTGCCTATCCAGAAAGTCCA (SEQ ID NO: 2) и 5'-GGAGAGAAGCTTGAGGGAGAGAAATGAATGATGG (SEQ ID NO.: 3; Zhang et al., 1994) и 2,5 ED Taq-полимеразы (Perkin Elmer) в течение 30 циклов в реакционном буфере, рекомендованном производителем (1,5 мМ MgCl2, 200 мкМ дНТФ в 100 мкл). Каждый цикл включал 1 мин при 55oС и 2 мин при 72oС.
Лигирование - Специфично амплифицированный продукт ПЦР (583 пары нуклеотидов) из препарата ПЦР расщепляли, в каждом случае при 37oС в течение 2 часов в буферных условиях, оговоренных производителем, с помощью рестрикционных ферментов BamHI и HindIII (Boehringer Mannheim), после чего фрагмент 564 п.н. очищали электрофорезом и выделяли. Затем его инкубировали, при 30oС в течение 2 часов и в 20 мкл, вместе с 0,1 мкг вектора pQE31 (Qiagen), расщепленного BamHI и HindIII, и 20 ЕД Т4 ДНК-лигазы (New England Biolabs).
Клонирование - 5 мкл лигированной смеси держали на льду в течение 30 мин вместе со 100 мкл трансформационно-компетентных клеток E.coli штамма НВ101, и смесь затем слегка перемешивали вращением пробирки в течение 5 мин в водяной бане при 37oС. После добавления 0,9 мл питательной среды, содержавшей 10 мМ MgCl2, смесь встряхивали при 37oС в течение 1 часа. 100 мкл объемы этой смеси в каждом случае помещали на агаровые пластинки, содержащие ампициллин (100 мкг/мл).
Идентификация клонов - клоны, выросшие за ночь при 37oС, инокулировали в 2 мл объемы жидких культур, содержавших ампициллин, культивировали до стационарной фазы, а затем осаждали центрифугированием. Клетки суспендировали в 0,1 мл 25 мМ Трис-буфера (рН 8), 50 мМ глюкозы, 10 мМ ЭДТА и лизоциме (2 мг/мл) и, после инкубирования при комнатной температуре в течение 5 мин, лизировали добавлением 0,2 мл 0,2 NaOH, 1% ДСН. Хромосомную ДНК осаждали добавлением 150 мкл 3 М ацетата натрия/уксусной кислоты (рН 5,2) и осаждали центрифугированием в течение 5 минут при 4oС (10 000 об/мин на Sigma 2MK). Плазмидную ДНК осаждали добавлением 2,5 объемов этанола, осаждали центрифугированием (см. выше) и, после промывания этанолом, помещали в 100 мкл воды; затем добавляли 10 мкл раствора РНК-азы (10 мг/мл).
Плазмидную ДНК расщепляли рестрикционными ферментами (BgII, Xhol + Pvull; Boehringer Mannheim), в соответствии с инструкциями производителя, и полученные фрагменты ДНК измеряли при сравнении с маркерной ДНК в геле агарозы после электрофореза и окрашивали этидия бромидом. Клоны, имевшие правильные фрагменты, исследовали тем же способом с помощью рестрикционного фермента AfIIII (New England Biolabs) на наличие глутаминового остатка 49.
В каждом случае идентичность одного клона E.coli, содержащего Gln49 (pQEob3-9), и одного - не содержащего Gln49 (pQEob3-4), подтверждалась посредством секвенирования ДНК. Выработку рекомбинантного лептина, содержащего последовательность-предшественник MetArgGlySer(His)6ThrAspPro (от вектора pQE31), вслед за которой идут аминокислоты 22-167 мышиного лептина, проверяли на малых культурах.
В то время как оба рекомбинантных лептина (с Gln49 и без нее) можно использовать для экспериментов, описанных ниже, примеры относятся конкретно к лептину, содержащему Gln49 и полученному экспрессированием pQEob3-9.
ПРИМЕР 2: Получение лептина
Разрушение - Бактерии из 10 л ферментации центрифугировали на 4800 об/мин в течение 20 мин. Осадок замораживали при -20oС, а затем суспендировали в лизисном буфере (6 М гуанидина хлорид, 0,1 М NaH2PO4, 10 мМ Трис/HCl, рН 8) (5 мл лизисного буфера на г осадка), после чего смесь перемешивали при комнатной температуре в течение одного часа, а затем центрифугировали на 4800 об/мин в течение 30 мин.
Ni-NTA-хроматография - 100 мл Ni-NTA-агарозы FF (Qiagen, Hilden) добавляли к неочищенному экстракту, который содержал приблизительно 800 мг лептина, и все вместе перемешивали при 4oС в течение ночи. Суспензию прокачивали через стеклянную колонку диаметром 5 см, содержащую фритту. Колонку вместе с Ni-NTA-агарозой, содержащейся в ней, промывали 300 мл лизисного буфера, а затем 100 мл лизисного буфера, содержащего 10 мМ имидазола (5 мл/мин). Затем происходила фракционная элюция лептина с применением линейного градиента имидазола от 10 до 200 мМ в лизисном буфере (объем градиента: 300 мл со скоростью 5 мл/мин). Фракции анализировали посредством высокоэффективной жидкостной хроматографии (ВЭЖХ) с обращенной фазой, и объединяли фракции, адекватные по чистоте и концентрации (=Ni-NTA-элюат).
Укладка - Ni-NTA-элюат разбавляли лизисным буфером до концентрации 1-3 мг/мл и доводили рН до 9,0 раствором гидроксида натрия. После добавления β-меркаптоэтанола (от 4 до 6 моль-β-меркаптоэтанола на моль лептина) смесь инкубировали при комнатной температуре в течение 2 часов в герметично закрытом контейнере. Для реоксидации и повторной укладки раствор выливали в 9-кратный объем укладочного буфера (0,1 М Трис/HCl, рН 9), и все вместе перемешивали при 16oС от 16 до 24 часов при доступе воздуха. Любое возникавшее замутнение удаляли центрифугированием (4 000 об/мин, 45 мин).
ВЭЖХ с обращенной фазой - рН укладочной смеси доводили до 3,0 с помощью НСl и прокачивали через 2,5•30 см обращеннофазовую колонку (PLRPS 300 ангстрем, 10 μ, Polymer Laboratories, Amherst, USA) со скоростью 20 мл/мин. Колонку затем промывали 300 мл элюента А (0,1% водная ТФУ). Лептин элюировали за 100 минут (скорость тока жидкости 7 мл/мин) с применением градиента от 25 до 50% элюента В (0,09% ТФУ в ацетонитриле). Фракции анализировали посредством аналитической ВЭЖХ. Фракции, адекватные по чистоте и концентрации, объединяли (обращеннофазовый пул). Этот пул обрабатывали 7 ммоль Na2HPO4/l и доводили рН до 3,0 с помощью NaOH, а растворитель удаляли в ротационном испарителе. Водный раствор лептина затем нейтрализовали с помощью NaOH (рН 7,4) и хранили при 4oС в течение ночи. Любое возникавшее замутнение удаляли центрифугированием (4 000 об/мин, 10 мин).
Гель-хроматография - Нейтрализованный и отцентрифугированный раствор лептина концентрировали до 10-15 мг/мл ультрафильтрованием, а затем стерилизовали фильтрованием. От 50 до 75 мг загружали на колонку Superdex 75 (2,6•60 см, Pharmacia, Sweden). В качестве буфера для элюции использовали физиологический раствор с фосфатным буфером (154 мМ NaCl, 10 мМ фосфат натрия, рН 7,4), при скорости тока жидкости 3 мл/мин. Очищенный таким способом лептин затем вновь фильтровали через мембрану 0,22 мкм и хранили при -70oС.
ПРИМЕР 3: Получение антагонистического фрагмента 116-167
1 мл 1 М Трис/HCl, рН 8, добавляли к 40 мл раствора лептина (1 мг/мл в физиологическом растворе с фосфатным буфером) и, после добавления 160 мкг лизилэндопептидазы, лептин расщепляли при комнатной температуре в течение 3 часов. рН смеси доводили до 3,0 и фракционировали посредством ВЭЖХ с обращенной фазой, как описано в примере 2. Фрагмент 116-167 идентифицировали посредством электроспрей-масс-спектрометрии (5532 D), растворитель удаляли, как описано в примере 2, концентрировали и очищали посредством гель-хроматографии.
ПРИМЕР 4: Выделение адипоцитов
Адипоциты получали из эпидидимальной жировой ткани самцов крыс Wistar (140-160 г, виварий Hoechst AG, Kastengrund) посредством расщепления коллагеназой (Rodbell, 1964, J. Biol. Chem. 239, 375-380), дважды промывали раствором KRH (25 мМ свободная кислота Hepes, 25 мМ натриевая соль Hepes, 80 мМ NaCl, 1 мМ MgSO4, 2 мМ СаСl2, 6 мМ КСl, 1 мМ пируват натрия, 0,5% БСА (бычий сывороточный альбумин)) и один раз средой DMEM (минимальная эссенциальная среда Дульбекко), дополнительно содержащей 5,5 мМ глюкозы, 20 мМ Hepes (рН 7,4), 2% эмбриональной бычьей сыворотки, 1% БСА 50 ЕД пенициллина/мл, 10 мг стрептомицина/мл, посредством флотации (800 g, 1 мин, в маленьких пластиковых пробирках) и, наконец, разбавляли до объема 20 мл DMEM/г сырого веса жировой ткани (клеточный титр приблизительно 2,5•105 клеток в мл).
ПРИМЕР 5: Первичная культура адипоцитов и инкубация с лептином
Адипоциты инкубировали при 37oС в течение 15-18 ч в атмосфере с содержанием 5% CO2, при легком встряхивании, в модифицированной (см. выше) среде DMEM в присутствии 100 нМ фенилизопропиладенозина (4 мл DMEM на 1 мл клеточной суспензии, клеточный титр приблизительно 5•104 клеток в мл, в 50 мл стерильных полипропиленовых пробирках), а также в присутствии или отсутствии лептина. Адипоциты затем промывали трижды холодным KRH и доводили клеточный титр приблизительно до 3•105 клеток в мл добавлением 0,7 мл KRH без глюкозы к клеточному слою, который остался после полного удаления последнего промывочного раствора. Для того, чтобы определить способность адипоцитов стимулироваться инсулином, а также их чувствительность к инсулину после первичной культуры, промытые адипоциты инкубировали при 37oС в течение 20 мин в присутствии или отсутствие инсулина человека (конечная концентрация 0,02-50 нМ), а затем определяли транспорт глюкозы или липогенез.
ПРИМЕР 6: Транспорт глюкозы
Транспорт глюкозы определяли как специфичный захват неметаболизирующегося аналога глюкозы - 2-дезоксиглюкозы ( and Wied, 1993, Diabetes 42, 1852-1867). 50 мкл суспензии адипоцитов в KRH, которую предварительно инкубировали или не инкубировали с инсулином (см. выше), инкубировали при 25oС в течение 5 мин с 50 мкл KRH, в который добавляли 2-дезокси-D-[2,6-3H] глюкозу (0,5 мкКи, 0,2 мМ). Инкубационные смеси переносили в тонкие центрифужные пробирки из мягкого пластика, каждая из которых уже содержала 200 мкл масла динонилфталата, и немедленно центрифугировали (2000 g, 30 сек). С помощью специальных ножниц пробирки перерезали внутри слоя масла (вблизи от верхней кромки), и верхние половинки пробирок вместе с клеточными слоями, которые в каждом случае плавали в масляном слое, переносили в сцинтилляционные флаконы. После добавления 10 мл сцинтилляционной жидкости (на водной основе) измеряли радиоактивность, связанную с клетками. С целью поправки на 2-дезоксиглюкозу, заключенную в интерстиции клеток или диффундировавшую в клетки неспецифичным путем, радиоактивность клеток, которые были предварительно инкубированы с цитохалазином В (20 мкМ), вычитали из общей связанной с клетками радиоактивности для каждой отдельной инкубации смеси (Gliemann et al., 1972, Biochim. Biophys. Acta 286, 1-9).
and Wied, 1993, Diabetes 42, 1852-1867). 50 мкл суспензии адипоцитов в KRH, которую предварительно инкубировали или не инкубировали с инсулином (см. выше), инкубировали при 25oС в течение 5 мин с 50 мкл KRH, в который добавляли 2-дезокси-D-[2,6-3H] глюкозу (0,5 мкКи, 0,2 мМ). Инкубационные смеси переносили в тонкие центрифужные пробирки из мягкого пластика, каждая из которых уже содержала 200 мкл масла динонилфталата, и немедленно центрифугировали (2000 g, 30 сек). С помощью специальных ножниц пробирки перерезали внутри слоя масла (вблизи от верхней кромки), и верхние половинки пробирок вместе с клеточными слоями, которые в каждом случае плавали в масляном слое, переносили в сцинтилляционные флаконы. После добавления 10 мл сцинтилляционной жидкости (на водной основе) измеряли радиоактивность, связанную с клетками. С целью поправки на 2-дезоксиглюкозу, заключенную в интерстиции клеток или диффундировавшую в клетки неспецифичным путем, радиоактивность клеток, которые были предварительно инкубированы с цитохалазином В (20 мкМ), вычитали из общей связанной с клетками радиоактивности для каждой отдельной инкубации смеси (Gliemann et al., 1972, Biochim. Biophys. Acta 286, 1-9).
ПРИМЕР 7: Липогенез
Липогенез определяли по включению D-глюкозы в липиды, которые поддаются экстракции толуолом (Moody et al., 1974, Horm. Metab. Res. 6, 12-16). 200 мкл суспензии адипоцитов в KRH инкубировали в сцинтилляционных флаконах при 37oС в течение 20 мин, в 680 мкл KRH, в который была добавлена глюкоза в концентрации 3,5 мМ и 20 мкл раствора инсулина. Липогенез начинали путем добавления 100 мкл D-[3-3Н] глюкозы (25 мкКи/мл KRH). После инкубирования при 37oС и легкого встряхивания в атмосфере с содержанием 5% СO2 в течение 90 минут добавляли 10 м сцинтилляционной жидкости (на основе толуола) и определяли радиоактивность толуольной фазы после интенсивного встряхивания и последующего разделения фаз (по меньшей мере, 4 ч инкубации). Радиоактивность липидов в толуольной фазе корректировали радиоактивностью инкубационной смеси, которая содержала такое же количество [3Н]глюкозы, но без клеток.
ПРИМЕР 8: Ингибирование лептином инсулин-индуцированного транспорта глюкозы и инсулин-индуцированного липогенеза
Изолированные адипоциты крысы инкубировали в течение 15 ч в первичной культуре в присутствии или отсутствие возрастающих концентраций лептина. Клетки затем промывали и определяли транспорт глюкозы и липогенез в исходном и простимулированном инсулином (10 нМ) состоянии. Фактор стимуляции инсулина рассчитывали как отношение простимулированной инсулином и исходной активности. Каждая величина представляла среднее от двух независимых культур адипоцитов, а активность определяли в каждом случае два или три раза. Результаты представлены в таблице 1.
ПРИМЕР 9: Влияние лептина на кривую доза-эффект инсулина
Изолированные адипоциты крысы инкубировали в течение 16,5 ч в первичной культуре в присутствии или отсутствие возрастающих концентраций лептина. Клетки затем промывали и тестировали на стимуляцию транспорта глюкозы различными концентрациями инсулина. Активность транспорта глюкозы дана в виде "величины dрм" 2-дезокси-[3H] глюкозы, специфично связанной с клетками. Каждая величина представляет собой среднее от двух независимых культур адипоцитов, а активность определяли в каждом случае четыре раза. Результаты представлены в таблице 2.
ПРИМЕР 10: Антагонизм фрагмента 116-167 по отношению к эффекту лептина
Изолированные адипоциты крысы инкубировали в течение 17,5 ч в первичной культуре в присутствии или отсутствие лептина (10 нМ) и возрастающих концентраций фрагмента лептина 116-167. Клетки затем промывали и тестировали на стимуляцию транспорта глюкозы или липогенеза 5 нМ инсулином. Фактор стимуляции инсулина рассчитывали как отношение простимулированной инсулином и исходной активности. Каждая величина представляла собой среднее от двух независимых культур адипоцитов, а активность определяли в каждом случае три раза. Результаты представлены в таблице 3.
Два остатка цистеина, представленные в последовательностях, показанных в таблицах 4-6, связаны между собой дисульфидным мостиком.










Изобретение относится к медицине. Предложена фармацевтическая композиция, содержащая антагонист лептина, способ лечения диабета II типа с ее использованием, фрагмент лептина мыши и способ его получения. Изобретение позволяет снизить инсулинорезистентность путем ингибирования эффекта лептина. 4 с. и 1 з.п. ф-лы, 6 табл.
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| KIEFFER et al | |||
| Leptin receptors expressed on pancreatic beta-cells Biochem Biophys Res Commun, v | |||
| Фотореле для аппарата, служащего для передачи на расстояние изображений | 1920 |
|
SU224A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Передвижной дровокольный станок | 1913 |
|
SU522A1 |
| СПОСОБ ЛЕЧЕНИЯ САХАРНОГО ДИАБЕТА | 1991 |
|
RU2040926C1 |

