Настоящее изобретение относится к белку плазматических мембран адипоцитов, который обладает специфическим сродством связывания к фосфоинозитолгликанам.
Роль фосфолипидов и фосфолипаз в трансмембранной передаче сигналов твердо установлена. В равной степени хорошо обоснована концепция заякоривания белков в клеточные мембраны с помощью ковалентно присоединяемого гликозилфосфатидилинозитола (GPI), причем точная химическая структура GPI-якоря определена для нескольких GPI-заякориваемых белков, таких как ацетилхолинэстераза (AchE) эритроцитов человека, Thy-1 крыс и нескольких оболочечных белков паразитов, аналогичных варианту поверхностного гликопротеина (VSG) из Trypanosoma brucei. Заякоривание липида происходит через фосфатидилинозитол (PI), который состоит из диацил- или алкилацилглицеринового типа фосфолипида. Поскольку последний встречается, в числе других, среди якорей млекопитающих и отличается от основной части PI, представленного в мембранах, можно было бы создать новые виды молекул, участвующих в образовании вторичных посредников, происходящих из GPI. Передача сигналов с помощью молекул GPI представляет особый интерес, так как данные заякориваемые липидами молекулы не пронизывают насквозь мембрану, а в большинстве случаев заключены во внешнюю половину липидного бислоя. Сигнал-опосредуемое высвобождение молекул GPI из клеточной мембраны продемонстрировано для разнообразных эндокринных и паракринных молекул, начиная от гормонов и включая факторы роста. Участие молекул GPI в трансмембранной передаче сигналов и их внутриклеточные эффекты в настоящее время установлены, однако меньше известно о сигнальном пути, ведущем к наблюдаемым метаболическим эффектам.
Идея о том, что GPI-заякориваемые молекулы обладают сигнальными свойствами, появилась из ранних экспериментов, в которых было показано, что связывание инсулина со своим рецептором активирует гидролиз молекул GPI. Идентифицировали низкомолекулярное вещество, которое имитирует некие механизмы действия инсулина на ферменты обмена веществ. Данное вещество обладает структурой инозитолгликана и образуется в плазматической мембране в результате гидролиза GPI, чувствительного к инсулину. Хотя первоначально думали, что GPI-предшественник инозитолгликанового ферментного модулятора представляет собой структурный аналог GPI-мембранного белкового якоря, существуют ясно выраженные отличия углеводной составляющей между GPI, преобразующим сигнал, и GPI-якорем мембранных белков. GPI-якорный мембранный белок неизменно состоит из триманнозной сердцевины, сопровождаемой этаноламинфосфатом, который обеспечивает соединение с С-концевой аминокислотой присоединяемого белка.
Регулируемый гидролиз GPI не ограничивается только инсулином, но наблюдается также при действии ряда других гормонов.
Практически во всех случаях стимуляция клеток гормонами или факторами роста приводит к временному высвобождению GPI-заякоренных белков из клеточной поверхности. Большинство рецепторов этих агонистов представляют собой либо тирозинкиназные рецепторы, либо рецепторы, связанные с тирозинкиназами.
Многие белки, участвующие в действии инсулина, идентифицированы на молекулярном уровне. Инсулиновый рецептор представляет собой трансмембранную тирозинкиназу, которая при активации в результате связывания с инсулином подвергается быстрому аутофосфорилированию и фосфорилирует ряд внутриклеточных субстратов, среди которых один или несколько 50-60 кДа белков, в том числе Shc, 15 кДа белок, связывающий жирную кислоту, и несколько так называемых инсулин-рецепторных субстратных белков, IRS-1/2/3/4. После фосфорилирования тирозина IRS-полипептиды действуют в качестве "стыковочных" белков для нескольких Src-гомологичных, содержащих 2 домена, адапторных молекул и ферментов, включая фосфатидилинозитол-3-киназу (PI-3-K), Grb2, SHP2, Nck, и Fyn. Взаимодействие между IRS-белками и PI-3-K происходит через регуляторную субъединицу p85 данного фермента и ведет к увеличению каталитической активности субъединицы p110. Фермент PI-3-K существенен для многих чувствительных к инсулину метаболических процессов, включая стимуляцию транспорта глюкозы и синтеза гликогена. Во всех случаях, в которых имеет место стимуляция фосфорилирования тирозина IRS-белков, имеет место и сопутствующая «стыковка» этих белков с субъединицей p85 фермента PI-3-K, и, за исключением пересечения между сигнальными системами инсулина и ангиотензина, данная «стыковка» ассоциируется со стимуляцией активности PI-3-K.
Помимо идентификации преобразующих сигнал путей, идущих непосредственно из инсулинового рецептора на ниже лежащие мишени, были разграничены несколько пересечений между передачей сигнала с помощью инсулина и другими гормонами/факторами роста или различными экзогенными раздражителями, которые либо имитируют (до некоторой степени) или модулируют положительным или отрицательным образом метаболическое и/или митогенное действие инсулина на разные клеточные системы. Поскольку ни один из этих лигандов прямо не активирует киназный рецептор инсулина, их сигнальные пути могут сходиться с сигнальным путем инсулина на более дистальной сигнальной стадии. Данное свойство является общим для молекул фосфоинозитолгликанпептида (PIG-P) разного вида, например, для PIG-P, полученного из гликозилфосфатидилинозитольного якоря из дрожжей Gce1p, который в существенной степени имитирует метаболическое действие инсулина, не сопровождаемое индукцией киназной активности рецептора инсулина.
Положительное взаимное влияние фосфоинозитолгликанов (PIG) и PIG-пептидов (PIG-P) на каскадное преобразование сигнала инсулина в чувствительных к инсулину клетках-мишенях включает перераспределение гликозилфосфатидилинозитол (GPI)-заякоренных белков плазматической мембраны (GPI-белок) и дважды ацилированных нерецепторных тирозинкиназ из содержащихся в плазматической мембране, детергент-устойчивых, гликолипид-обогащенных совокупностей доменов с высоким содержанием холестерина (hcDIG) в совокупность доменов с низким содержанием холестерина (lcDIG).
В выделяемых адипоцитах крыс первичная мишень PIG-P локализована в hcDIG. Меченный радиоактивной меткой PIG-P, Tyr-Cys-Asn-NH-(CH2)2-O-PO(OH)O-6Manα1-2)-2Manα1-6Manα1-4GluN1-6lno-1,2-(циклический)-фосфат (YCN-PIG), а также меченный радиоактивной меткой и липолитически расщепляемый GPI-белок (lcGce1p) из Saccharomyces cerevisiae, из которого был получен YCN-PIG, насыщающим образом связываются с hcDIG, но не с lcDIG, микросомами или с общими плазматическими мембранами. Связывание и YCN-PIG, и lcGce1 является специфичным, поскольку оно полностью аннулируется либо избытком химически синтезированного немеченого YCN-PIG, или предварительной обработкой адипоцитов с помощью трипсина, и затем NaCl или N-этилмалеимида (NEM), что свидетельствует о том, что YCN-PIG распознается поверхностным рецептором клетки. Связывание PIG-P существенно увеличивается в hcDIG из адипоцитов, предварительно подвергнутых обработке с помощью GPI-специфичных фосфолипаз С, совместно с липолитическим удалением эндогенных лигандов, таких как GPI-белки/липиды. Связывание по сродству является наибольшим для YCN-PIG, за которым следует комбинация отдельных составляющих, Tyr-Cys-Asn- NH-(CH2)2-OH(YCN) и HO-PO(H)O-6Manα1(Manα1-2)-2Manα1-6Manα1-4GluN1-6Ino-1,2-(циклический)-фосфат (PIG37), а также пептидный вариант, YMN-PIG. PIG37 и YCN порознь демонстрируют промежуточное и низкое сродство. Инкубация адипоцитов с YCN-PIG уменьшает последующее меченье с помощью [C14]NEM 115 кДа-полипептида, высвобождаемого с клеточной поверхности последовательной обработкой трипсин/NaCl. Эти данные показывают, что в адипоцитах крыс имитирующий инсулин PIG(-P) распознается трипсин/NaCl/NEM-чувствительным 115 кДа-белком из hcDIG, который действует в качестве рецептора GPI-белков.
В одной и той же клетке, по-видимому, существует несколько видов DIG. Ямки на поверхности терминально дифференцированных клеток представляют собой специальные DIG, которые образуют колбообразные впячивания, приводимые в движение чрезмерной экспрессией маркера и структурного белка кавеолина 1-3.
Ямки, которые составляют в адипоцитах 20% площади поверхности плазматической мембраны, принимают участие в рецептор-опосредованном потоцитозе (potocytosis), эндоцитозе, трансцитозе и передаче сигнала. В выделяемых крысиных адипоцитах lcDIG с низким содержанием холестерина/кавеолина, проявляющих высокую плавучую плотность (в соответствии с градиентным центрифугированием в плотности сахарозы), могут отличаться от типичных hcDIG с высоким содержанием холестерина/кавеолина, характеризуемых низкой плавучей плотностью. Основная фракция GPI-белков, таких как Gce1 и Nuc, а также дважды ацилированных белков, таких как NRTK, нерецепторная тирозинкиназа, pp59Lyn, локализована в hcDIG. В ответ на имитирующие инсулин стимулы, такие как синтетический PIG или сульфонилмочевина, глимепирид, и GPI-белки и NRTK перемещаются из hcDIG в lcDIG. Данное перераспределение не связано с потерей их липидной модификации.
Полярная центральная гликановая головная группа без (PIG)- или с (PIG-P)-примыкающими аминокислотами на карбоксиконце GPI-белковой полипептидной составляющей обеспечивают молекулярную основу распределения GPI-белков между hcDIG и lcDIG в основном состоянии и их перераспределение в ответ на имитирующие инсулин стимулы.
GPI-белки представляют собой антигены клеточной поверхности, эктоферменты, рецепторы или клеточные молекулы адгезии, экспрессируемые у эукариот от дрожжей до человека, и заякориваемые в наружный листок плазматической мембраны с помощью ковалентно присоединяемой гликозилфосфатидилинозитольной (GPI) липидной составляющей. Несмотря на отсутствие трансмембранного домена, они вовлечены в передачу сигнала через данную плазматическую мембрану.
Установление того факта, что GPI-белки ассоциированы с множеством специализированных липидных доменов, так называемой детергент-нерастворимой богатой гликолипидами массой, DIG, а не с особыми трансмембранными связывающими/линкерными белками, свидетельствует о возможности липид-липидных взаимодействий в качестве основного связывающего механизма передачи сигнала, опосредуемого GPI-белками.
Основным структурным элементом DIG является латеральная совокупность (глико)сфинголипидов и холестерина, которая принимает вид упорядоченной жидкости (lo) в отличие от смежных участков неупорядоченной жидкости (ld) в липидном бислое данной мембраны. Плазматические мембраны клеток млекопитающих содержат холестерин (30-50 мол.%) и смесь липидов преимущественно в ld-доменах (например, фосфатидилхолины с ненасыщенными хвостовыми фракциями), а также липидов, несущих насыщенные ацильные цепи преимущественно в lo-доменах (например, [глико]сфинголипиды и GPI-липиды). Считают, что холестерин способствует плотной упаковке липидов в lo-доменах путем заполнения интерстициального пространства между липидными молекулами, и образование lo-доменов отмечается лишь при определенных концентрациях холестерина.
Инсулин является чрезвычайно важным гормоном, который оказывает существенное влияние на обмен веществ в организме. В общих чертах, он усиливает анаболические процессы и подавляет катаболические процессы. В частности, он увеличивает скорость синтеза гликогена, жирных кислот и белка, и тормозит распад белка и гликогена. Жизненно важное действие гормона заключается в стимуляции клеток печени, мышц и жировых клеток для удаления из кровотока глюкозы, некоторых других сахаров и аминокислот.
Бычий инсулин состоит из двух полипептидных цепей, полипептида А, содержащего 21 АК, и полипептида В, содержащего 30 АК, которые соединены двумя -S-S- (дисульфидными мостиками). Такой же характер структуры отмечен для инсулина многих млекопитающих, в том числе и для человека.
Его структура является компактной, подобной цилиндру с только карбоксильным концом В-цепи, выступающим из остального белка. Имеется много гидрофобных остатков, которые взаимодействуют с образованием центрального гидрофобного ядра, а внешнедиспергированными являются полярные остатки на любой стороне, которые дополнительно стабилизируют данный белок. Три дисульфидных мостика, два внутрицепочечных и один межцепочечный, скрепляют данную структуру.
Общей характерной чертой биосинтеза многих белков, и особенно белков, экспортируемых из клеток, является то, что белок, продуцируемый в виде предшественника, затем модифицируется, принимая окончательную форму в период сохранения и перед высвобождением. Инсулин синтезируется группой клеток поджелудочной железы, называемых островками Лангерганса, сохраняется в гранулах и при необходимости высвобождается в кровоток.
Когда инсулин синтезируется впервые, он состоит из 100 АК единственной полипептидной цепи, состоящей из 16 АК сигнальной последовательности, В-цепи, С-цепи из 33 АК, именуемой соединительной цепью, и А-цепи. Данную структуру называют пре-проинсулином (PPI). Полагают, что сигнальная область ответственна за направление PPI из участка синтеза в ER (эндоплазматический ретикулум) данной клетки, который накапливает и пакует инсулин в виде депонируемых гранул. При помещении в ER указанный сигнальный пептид удаляется с помощью протеазного фермента.
Сахарный диабет представляет собой хроническое заболевание, которое требует обращения к врачу на протяжении длительного времени, чтобы ограничить развитие его разрушительных осложнений и уметь обращаться с ними, когда они случаются. Диабет связан с острыми и хроническими осложнениями, такими как гипогликемия, диабетический кетоацидоз и гиперосмолярный некетозный синдром.
Диабет 1 типа случается у молодых, худощавых пациентов и характеризуется отчетливой неспособностью поджелудочной железы секретировать инсулин, вследствие аутоиммунной деструкции бета-клеток. У пациентов с диабетом 1 типа при отмене инсулина наблюдают развитие кетоза и в конечном итоге - развитие кетоацидоза. Следовательно, эти пациенты зависимы от экзогенного инсулина, который поддерживает их жизнь.
Диабет 2 типа, как правило, возникает у индивидов старше 40 лет, которые обладают семейным анамнезом диабета. Диабет 2 типа характеризуется периферической инсулинорезистентностью с нарушением секреции инсулина, которая варьирует по тяжести. Данные дефекты приводят к повышенному глюконеогенезу печени, что вызывает гипергликемию натощак. Большинство пациентов (90%), у которых развился диабет 2 типа, страдают ожирением, а само ожирение связано с инсулинорезистентностью, которая ухудшает состояние диабетика.
Различные другие типы диабета, ранее названные «вторичным диабетом», вызываются другими заболеваниями или лекарственными препаратами. В зависимости от исходного рассматриваемого процесса (а именно, деструкции бета-клеток поджелудочной железы или развития периферической инсулинорезистентности), данные типы диабета ведут себя подобно диабету 1 типа или 2 типа. Наиболее обычными являются заболевания поджелудочной железы, которые разрушают бета-клетки (например, гемохроматоз, панкреатит, кистозный фиброз, злокачественная опухоль поджелудочной железы), гормональные синдромы, которые препятствуют секреции инсулина (например, феохромоцитома), либо вызванны периферической инсулинорезистентностью (например, акромегалия, синдром Кушинга, феохромоцитома), а также диабет, индуцированный лекарственным средством (например, фенитоин, глюкокортикоиды, эстрогены).
Сахарный диабет характеризуется неправильной регуляцией уровня глюкозы в сыворотке. При диабете 1 типа аутоиммунная атака на эндокринную поджелудочную железу приводит к прогрессирующему и необратимому разрушению секретирующих инсулин бета-клеток. Утрата инсулина воздействует на поглощение глюкозы восприимчивой к инсулину клеткой-мишенью и на течение обмена веществ. Диабет 2 типа имеет несколько этиологий, большей частью сказывается на клеточной резистентности к действию инсулина, и сопровождается также изменениями регуляции уровня глюкозы в сыворотке. Инсулин действует через связанный с дисульфидом гетеротетрамерный рецептор клеточной поверхности, который включает внеклеточную альфа-субъединицу, связанную с помощью дисульфидных связей с трансмембраной и внутриклеточной бета-субъединицей. При диабете 1 типа отсутствие лиганда с нормальной клеточной рецепторной структурой и функцией в большинстве случаев вызывает последующие метаболические дефекты. Гормон-заместительная терапия в виде ежедневных инсулиновых инъекций обеспечивает организм лигандом с действием на рецептор, хотя и не обязательно нормальным физиологическим образом. При диабете 2 типа резистентность к действию инсулина часто лежит в основе заболевания с некоторой резистентностью, обусловленной нарушениями рецепторного действия.
Известно, что в случае инсулинорезистентности, требуется больше инсулина для того, чтобы с помощью инсулинового рецептора запустить инсулиновый сигнальный каскад. Настоящее изобретение относится к белку клеточной мембраны адипоцитов, который способен стимулировать поглощение глюкозы в обход сигнального пути, запускаемого рецептором инсулина. Это обеспечивает действенное решение проблемы в отсутствие инструмента скринирования, чтобы идентифицировать соединения, которые могли бы действовать в качестве альтернативы инсулину.
Поэтому настоящее изобретение относится к белку плазматической мембраны адипоцита, который, по-видимому, стабилизируется благодаря одновременному присутствию плазматических мембран и/или липидных пузырьков, и/или множества доменов с высоким содержанием холестерина и/или липидных пузырьков, и который обладает специфическим сродством связывания с фосфоинозитолгликаном или фосфоинозитолгликан-пептидом, отличающимся
а] способностью запускать в адипоците tyr-фосфорилирование субстрата 1 или 2 рецептора инсулина после специфического связывания фосфоинозитолгликана или фосфоинозитолгликан-пептида с данным белком и
b] способностью стимулировать в адипоците поглощение глюкозы после специфического связывания фосфоинозитолгликана или фосфоинозитолгликан-пептида с данным белком.
Количество данного белка по сравнению с другими белками и/или стабилизирующими компонентами и/или иными соединениями (например, солями, ионом, распылителем) находится в диапазоне между 0,01-10% в пересчете на сырую массу.
Количество данного белка предпочтительно находится в диапазоне от 0,1 до 5% в расчете на влажную массу, но наиболее предпочтительно находится в диапазоне от 0,1 до 1% в пересчете на влажную массу.
В естественных условиях количество указанного белка в плазматических мембранах находится в диапазоне менее 10-6% в пересчете на влажную массу.
В предпочтительных вариантах настоящего изобретения фосфоинозитолгликан или фосфоинозитолгликан-пептид состоит, по меньшей мере, из одного из следующих соединений:
YCN-PIG, YMN-PIG, PIG37, YCN или lcGce1.
Связывание фосфоинозитолгликана или фосфоинозитолгликан-пептида с данным белком происходит, предпочтительно, с константой связывания (KD) от 0,001 до 10 мкМ.
Константа связывания представляет собой термодинамически упорядоченное количественное описание равновесия между диссоциированными и недиссоциированными формами комплексов между данным белком и фосфоинозитолгликаном или фосфоинозитолгликан-пептидом.
Константа связывания выражается отношением констант скорости прямой и обратной реакции. Высокие значения константы связывания (например, более 10 мМ) указывают на слабое и неспецифическое связывание, а низкие значения (например, не более чем 100 мкМ) указывают на сильное и специфическое связывание.
Константы связывания можно определить разными способами, например, с помощью равновесного диализа, спектроскопии или графическими методами (Scatchard-Plot).
Что касается плазматической мембраны адипоцита, то она предпочтительно происходит от крысы, мыши или человека.
Молекулярная масса данного белка находится между 100-120 кДа, предпочтительно между 110-120 и наиболее предпочтительно равна 115 кДа. Следует отметить, что определение молекулярной массы белков любым способом, в частности с помощью SDS-ПААГ, имеет место с неопределенностью ±5-10%.
Кроме того, настоящее изобретение относится к комплексу, который образуется из вышеизложенного белка настоящего изобретения и, по меньшей мере, одного соединения из следующей группы: YCN-PIG, YMN-PIG, PIG37, YCN или lcGce1.
Предпосылкой образования комплекса является специфическое связывание данного лиганда с данным белком. Полученный комплекс можно стабилизировать, создавая ионную или ковалентную взаимосвязь между лигандом и белком.
Настоящее изобретение относится также к получению белка настоящего изобретения, в котором:
а] адипоциты получают из ткани крысы, мыши или человека,
b] из п. a] выделяют плазматические мембраны адипоцитов,
c] совокупность доменов с высоким холестерином (hcDIG) получают из плазматических мембран п. b],
d] hcDIG из п. c] обрабатывают раствором трипсин/NaCl,
e] инкубационную смесь из п. d] центрифугируют и белки полученного супернатанта разделяют при помощи электрофореза в SDS-ПААГ (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия),
f] из данного геля элюируют белковую фракцию размером 100-120 кДа и по возможности солюбилизируют раствором или суспензией, содержащими детергент или биологические мембраны.
Кроме того, настоящее изобретение относится к способу идентификации соединения, которое специфически связывается с белком настоящего изобретения, в котором:
a] получают фракцию клеток, которая содержит белок настоящего изобретения,
b] получают соединение,
c] фракцию клеток из п. a] приводят в контакт с соединением п. b],
d] определяют связывание данного соединения с фракцией клеток из п. a],
e] о специфичности связывания делают вывод путем сравнения результатов из п. d] с результатами эксперимента, в котором такое же соединение, что и в п. b] приводят в контакт с фракцией клеток того же вида и/или той же тканевой специфичности, что и клетки из п. a], но которая не содержит белка настоящего изобретения, указывающего, таким образом, о более высокой специфичности связывания в случае большего количества данного соединения из п. b], связывающегося с фракцией клеток, которая содержит белок настоящего изобретения, по сравнению с фракцией клеток, которая не содержит белка настоящего изобретения.
Данную фракцию клеток берут предпочтительно из адипоцитов, клеток скелетных мышц, клеток сердечной мышцы или клеток печени. Каждый вид этих клеток можно получить предпочтительно из мыши, крысы или от человека. Данная фракция клеток состоит предпочтительно из клеточных мембран любых клеток или более предпочтительно, из совокупности доменов с высоким содержанием холестерина (hcDIG). Данное соединение, которое используют для осуществления способа идентификации соединения, которое специфически связывается с белком настоящего изобретения, можно пометить радиоактивным изотопом (например, С14, Н3, Р32, J121 и другие) или флуоресцентным маркером.
Далее, настоящее изобретение относится к способу идентификации соединения, которое специфически связывается с белком настоящего изобретения, в котором:
a] получают клетку, транспортирующую глюкозу, которая содержит белок настоящего изобретения,
b] получают соединение,
c] клетку из п. a] приводят в контакт с соединением п. b],
d] определяют связывание данного соединения с клеткой, транспортирующей глюкозу,
e] вывод о специфичности связывания делают на основании сравнения результатов из п. d] с результатами эксперимента, в котором такое же соединение, что и в п. b], приводят в контакт с транспортирующей глюкозу клеткой того же вида и/или той же тканевой специфичности, что и клетки из п. a], но которая не содержит белка настоящего изобретения, указывающего, таким образом, о более высокой специфичности связывания в случае большего количества соединения из п. b], связывающегося с транспортирующей глюкозу клеткой, которая содержит белок настоящего изобретения, по сравнению с транспортирующей глюкозу клеткой, которая не содержит белка настоящего изобретения.
Транспортирующую глюкозу клетку, которая не содержит белка настоящего изобретения, можно получить из транспортирующей глюкозу клетки, которая содержит белок настоящего изобретения путем обработки данной клетки, которая содержит белок настоящего изобретения, раствором трипсин/NaCl и/или гликозидазой.
Транспортирующая глюкозу клетка предпочтительно представляет собой адипоцит, клетку скелетной мышцы, клетку сердечной мышцы или клетку печени. Данные клетки предпочтительно берут из культуры тканей или клеток человека, мышиного или человеческого происхождения.
Данное используемое соединение предпочтительно метят радиоактивным изотопом или флуоресцентным маркером.
Кроме того, настоящее изобретение относится к способу идентификации соединения, которое является агонистом или антагонистом белка настоящего изобретения, в котором
a] получают транспортирующую глюкозу клетку, в которой представлен белок настоящего изобретения,
b] получают природный лиганд белка настоящего изобретения,
c] получают химическое соединение,
d] транспортирующую глюкозу клетку п. a] приводят в контакт с лигандом из п. b] и химическим соединением из п. c],
e] определяют поглощение глюкозы транспортирующей глюкозу клеткой из п. d],
f] определяют поглощение глюкозы транспортирующей глюкозу клеткой из п. d], в которой стимуляция поглощения глюкозы подразумевает агонистическую активность, а ингибирование поглощения глюкозы подразумевает антагонистическую активность соединения п. c].
Лиганд вышеприведенного способа идентификации агониста или антагониста белка настоящего изобретения предпочтительно представляет собой YCN-PIG, YMN-PIG, PIG37, YCN или lcGce1. Транспортирующая глюкозу клетка в способе идентификации агониста или антагониста белка настоящего изобретения предпочтительно представляет собой адипоцит, клетку скелетной мышцы, клетку сердца или клетку печени, и предпочтительно происходят от видов - человек, мышь или крыса.
Настоящее изобретение относится также к лекарственному средству, содержащему соединение, которое было идентифицировано способом идентификации соединения, которое связывается с белком настоящего соединения, или которое является агонистом, либо антагонистом белка настоящего изобретения, а также вспомогательные соединения для создания лекарственного средства. Данное лекарственное средство содержит в предпочтительном варианте осуществления настоящего изобретения, по меньшей мере, одно соединение из следующей группы: YCN-PIG, YMN-PIG, PIG37, YCN или lcGce1.
Данное лекарственное средство может также содержать часть или производное, по меньшей мере, одного соединения из следующей группы: YCN-PIG, YMN-PIG, PIG37, YCN или lcGce1.
Кроме того, настоящее изобретение относится к использованию соединения, которое было идентифицировано в качестве связывающегося с белком настоящего изобретения или в качестве агониста или антагониста белка настоящего изобретения, для получения лекарственного средства для лечения инсулинорезистентности или диабета.
Таким соединением может предпочтительно быть YCN-PIG, YMN-PIG, PIG37, YCN или lcGce1, или же часть либо производное одного из данных соединений.
ПРИМЕРЫ
Химический синтез PIG(-P): синтез YCN-PIG (общий метод, смотрите Фиг. 1, 2, 3)
Синтез продукта 2 (Фиг. 4; i, ii), продукт 1 (8,0 г, 20,6 ммоль) от Bachem (Hedelberg, Германия) растворяли в 200 мл пиридина, и добавляли 5 г (81,8 ммоль) этаноламина и 5 мл N-этилморфолина. После отстаивания (16 ч, комнатная температура) при перемешивании и при 5°С по каплям приливали 50 мл уксусного ангидрида. Данную реакционную смесь перемешивали (2 ч, комнатная температура), и затем концентрировали в высоком вакууме. Сконцентрированный остаток растворяли в 150 мл горячего метанола, и полученный раствор концентрировали. Затем приливали 100 мл метиленхлорида/метанола (15/1) и 200 мл н-гептан/этилацетата (2/1), и данный продукт кристаллизовали. Выход продукта 2: 6,1 г (84%) белых кристаллов с т. пл. 175°С. ТСХ (тонкослойная хроматография): метиленхлорид/метанол (9/1), Rf=0,7. МС: (M + Li)+=358,2, расчетное значение для C16H21N3O6, M=351,36.
Синтез продукта 3 (Фиг. 4; iii), 2,0 г палладия на угле (10% Pd) вносили в раствор продукта 2 (12,0 г, 34,0 ммоль) в 200 мл метанол/уксусной кислоты (1/1), и данную смесь гидрировали (2 ч, комнатная температура). Полученный раствор фильтровали на силикагеле и концентрировали, а остаток выделяли очисткой с помощью флэш-хроматографии (метиленхлорид/метанол/концентрированный аммиак 30/5/1). Выход продукта 3: 7,3 г (98%) желтоватого масла. ТСХ: метиленхлорид/метанол/концентрированный аммиак (30/5/1), Rf=0,5. МС: (M + Li)+=224,2, расчетное значение для C8H15N3O4, M=217,23.
Синтез продукта 4 (Фиг. 4; iv), 1,5 г (4,5 ммоль) 1(o-(циано(этоксикарбонил)-метилиден)амино-1,1,3,3-тетраметилуронийтетрафторборат (TOTU), 0,64 г (4,5 ммоль) этил-(гидроксиимино)-цианоацетата (оксим) и 1,7 мл (13,5 ммоль) N-этилморфолина вносили при 0°С с перемешиванием в раствор из 0,8 г (3,7 ммоль) продукта 3 и 2,8 г (4,5 ммоль) TrtCys(Trt)OH в диметилформамиде, и данную смесь перемешивали (2 ч, 0°С). После добавления 200 мл этилацетата данную смесь трижды промывали насыщенным раствором NaHCO3, обезвоживали над MgSO4 и концентрировали. Полученный остаток растирали в н-гептан/этилацетате (6/1) и этот продукт кристаллизовали. Выход продукта 4: 2,2 г (74%) белых кристаллов с т. пл. 185°С. ТСХ: метиленхлорид/метанол (15/1), Rf=0,4. МС: (M + Li)+=811,7, расчетное значение для C49H48N4O5S, M=805,0.
Синтез продукта 6 (Фиг. 4; v, vi), 4,0 г (5,0 ммоль) продукта 4 растворяли в 200 мл метиленхлорида. Приливали 4 мл воды и 3 мл трифторуксусной кислоты. Через 15 мин данную смесь трижды промывали насыщенным раствором NaHCO3, обезвоживали над MgSO4 и концентрировали с 99%-ным выходом неочищенного продукта 5. Данный неочищенный продукт растворяли в 50 мл метанола и затем каплями добавляли 0,5 мл 1 М раствора метанолята натрия. Через 15 мин приливали 50 мл метиленхлорида, и данную смесь фильтровали на силикагеле. После концентрирования отфильтрованного раствора остаток выделяли очисткой с помощью флэш-хроматографии (метиленхлорид/метанол (9/1)). Выход продукта 6: 2,2 г (85%) аморфного твердого вещества. ТСХ: метиленхлорид/метанол (5/1), Rf=0,7. МС: (M + Li)+=527,3, расчетное значение для C28H32N4O4S, M=520,6.
Синтез продукта 7 (Фиг. 4; vii), 2,7 г (5,2 ммоль) продукта 6, 4,2 г (10,4 ммоль) Ztyr(Bn)OH, 3,4 г (10,4 ммоль) TOTU, 1,5 г (10,4 ммоль) оксима и 2 мл N-этилморфолина в 50 мл диметилформамида аналогичным образом подвергали взаимодействию с препаратом продукта 4. Выход продукта 7: 4,2 г (89%) белых кристаллов. ТСХ: метиленхлорид/метанол (15/1), Rf=0,25. МС: (M + Li)+=914,8, расчетное значение для C25H53N5O8S, M=908,1.
Синтез продукта 8 (Фиг. 5; viii), 6,0 г (73 ммоль) фосфористой кислоты подвергали четырехкратному концентрированию с пиридином, после чего помещали в 180 мл безводного пиридина. При 10°С по каплям добавляли 13 мл пивалоилхлорида. Данный реакционный раствор оставляли на 45 мин при комнатной температуре. В этот реакционный раствор вводили 16,4 г (18,1 ммоль) продукта 7, как описано выше. Через 5 ч. его разбавляли с помощью 200 мл толуола и 150 мл метиленхлорид/метанол/33% NH3 (30/10/3). После концентрирования остаток пиридина трижды перекристаллизовывали с 200 мл толуола. Данный остаток суспендировали в 200 мл метиленхлорид/метанола (20/1). Нерастворившиеся компоненты фильтровали и дважды промывали с помощью 50 мл метиленхлорид/метанола (20/1). Полученный фильтрат концентрировали и выделяли очисткой с помощью флэш-хроматографии. Выход продукта 8: 11,6 г (66%) белых кристаллов. ТСХ: метиленхлорид/метанол/33% NH3 (30/5/1), Rf=0,25. МС: (M + Li)+=978,4, расчетное значение для C52H54N5O10SP, M=972,08.
Синтез продукта 10 (Фиг. 6; ix, x), 4,5 г продукта 8 (4,6 ммоль) и 6,0 г продукта 9 (2,3 ммоль; синтез осуществляли, как описано раньше в ссылке 47), растворяли в 80 мл безводного пиридина. Через 30 мин при комнатной температуре данную реакцию охлаждали до 0°С и добавляли 5 мл воды и 1,3 г йода. Эту реакционную смесь перемешивали (30 мин, 10°С) и затем разбавляли с помощью 500 мл метиленхлорида, 150 мл насыщенного раствора NaCl и 30 мл насыщенного раствора тиосульфата и перемешивали в течение 5 мин. Органическую фазу обезвоживали над MgSO4 и концентрировали. Полученный остаток выделяли очисткой с помощью флэш-хроматографии с метиленхлорид/метанол/конц.NH3 (30/5/1-30/10/3). Выход продукта 10: 8,0 г в виде аморфного твердого вещества. ТХС: метиленхлорид/метанол (20/1), Rf=0,5. МС: (M + Li)+=3583,6, расчетное значение для C207H214N8O42SP2, M=3580,0.
Синтез продукта 11 (Фиг. 6; xi), 300 мл аммиака концентрировали при -78°С. В нем растворяли 2,1 г (91 ммоль) натрия. Данный раствор разбавляли 150 мл безводного тетрагидрофурана, после чего 8,0 г продукта 10 (2,2 ммоль) из защищенного конечного продукта, растворенного в 50 мл безводного тетрагидрофурана, медленно по каплям добавляли в реакционную смесь с температурой -78°С. Через 15 мин течения реакции (голубая окраска не должна исчезать), данную смесь осторожно обрабатывали с помощью 5 г хлорида аммония. При исчезновении голубой окраски данную смесь осторожно разбавляли с помощью 50 мл воды и 150 мл метанола. Давали ей возможность растаять, а затем концентрировали до около 100 мл. Данный раствор разбавляли с помощью 500 мл метиленхлорид/метанол/33% NH3 (3/3/1) и вносили во флэш-силикагельную колонку (500 мл силикагеля). Ее последовательно элюировали с помощью 1 л, каждого, метиленхлорид/метанол/33% NH3 (3/3/2) и (3/3,5/3). Затем данный элюируемый продукт хроматографировали с использованием н-бутанол/этанол/вода/33% NH3 (2/2/2/1). Выход продукта 11: 2,4 г (67% от продукта 9) в виде белого твердого вещества. ТХС: н-бутанол/этанол/вода/33% NH3 (2/2/2/1), Rf=0,5. МС: (M + NH3)+=1572,6; расчетное значение для C54H88N6O40P2S, M=1555,31. P31-ЯМР (D2O)=15,3 м.д. для циклофосфата и 0,3 для эфира фосфорной кислоты. Данные для H1- и C13-ЯМР представлены в Таблице 1.
Синтез продукта YCN (Фиг. 7; xii), 11,0 г (11,3 ммоль) продукта 7 подвергали снятию защиты аналогичным образом, как и препарат продукта 11. Выход YCN: 4,5 г (90%) белых кристаллов. ТСХ: метиленхлорид/метанол/концентрированный аммиак (30/15/5), Rf=0,25. МС: (M + Li)+=448,3, расчетное значение для C18H27N5O6S, M=441,51.
Синтез продукта YMN-PIG, YMN-PIG синтезировали с помощью той же последовательности реакций, которая представлена на Фиг. 2. Использование BocMetOH вместо TrtCys(Trt)OH давало YMN-PIG со сходным выходом в виде белого твердого вещества. ТХС: н-бутанол/этанол/вода/33% NH3 (2/2/2/1), Rf=0,5. МС: (M + NH3)+=1600,6; расчетное значение для C56H92N6O40P2S, M=1583,38. P31-ЯМР (D2O)=15,3 м.д. для циклофосфата и 0,3 для эфира фосфорной кислоты.
Получение радиоактивно меченого и липолитически расщепляемого Gce1p (lcGce1p)
Gce1p с интактным GPI-якорем выделяли очисткой из растущих на лактате дрожжевых клеток, которые метаболически метили с помощью мио-[C14]инозита, а затем ферментативно преобразовали в сферопласты. Плазматически мембраны получали выделением очисткой с помощью центрифугирования в градиенте фиколла, солюбилизировали с использованием 0,35% β-амидотаурохолата и подвергали TX-114-разделению. Gce1p, содержащийся в богатой детергентом фазе, выделяли очисткой с помощью гель-фильтрующей хроматографии на Сефадексе S-300, хроматографии по сродству на Сефарозе N6-(2-аминоэтил)-cAMP и хроматографии на фенил-Сефарозе. Элюирование с колонок сопровождалось непрерывным мониторингом H3-радиоактивности. Частично выделенный очисткой Gce1p осаждали (12% полиэтиленгликоль 6000), затем ресуспендировали в буфере G (25 мМ Трис/ацетат, рН 7,4, 144 мМ NaCl, 0,1% β-амидотаурохолат, 0,5 мМ DTT, 0,2 мМ ЭДТА, 5% глицерин, 0,1 мМ PMSF, 5 мкМ лейпептин, 1 мМ йодацетамид, 10 мкг/мл ингибитор трипсина из соевых бобов) при концентрации 0,2 мг белка/мл и впоследствии инкубировали (3 ч, 25°С) в присутствии 6 ед./мл PI-специфичного PLC (B. cereus). После добавления ледяного раствора, состоящего из 2% Тритона Х-114, 10 мМ Трис/HCl (рН 7,4), 144 мМ NaCl и отделенной фазы, (инкубирования в течение 2 мин при 37°С и центрифугирования при 12000×g в течение 1 мин при 25°С), lcGce1p извлекали из верхней бедной детергентом фазы. После двух дополнительных извлечений из нижней богатой детергентом фазы путем добавления равного объема 10 мМ Трис/HCl, 144 мМ NaCl, перерастворения на льду и последовательного отделения фазы, объединенные бедные детергентом фазы преципитировали (12% полиэтиленгликолем 6000).
Меченный радиоактивным изотопом lcGce1p суспендировали в буфере в отсутствие β-амидотаурохолата при 200-1000 распадах в минуту/мкл.
Получение меченого радиоактивным изотопом YCN-PIG
Меченый радиоактивным изотопом YCN-PIG готовили из Gce1p путем последовательной обработки с помощью V8-протеазы (S. aureus) и PI-PLC (B. cereus). YCN-PIG извлекали из бедной детергентом фазы после TX-114-распределения, а затем последовательно выделяли очисткой с помощью катионообменной хроматографии (Дауэкс 50W-X8), гель-фильтрации на BioGel-P4, анионообменной хроматографии на SAX-колонке ВЭЖХ, двух тонкослойных хроматографических разгонок на Si-60-пластинках HPTLC с использованием разных систем растворителей и, в заключение, гель-фильтрации на BioGel-P4. Элюирование материала в течение каждого хроматографического разделения сопровождалось измерением H3-радиоактивности, УФ-поглощения (А220) и инсулиноподобной активности по стимуляции транспорта глюкозы в изолированных адипоцитах крысы. Для демонстрации радиохимической чистоты конечный препарат YCN-PIG подвергали анионообменной ВЭЖХ на Dionex CarboPac PA-1, калибруемой на Dionex-комплексах путем включения стандартной смеси глюкозного олигомера. Внутренние эталоны детектировали с использованием импульсного амперометрического детектора. Появление С14-меченных фрагментов отслеживали, непрерывно контролируя радиоактивность при помощи Raytest Ramona. Для определения концентрации YCN-PIG гидролизовали (6 М HCl, 16 ч, 110°С) и определяли количество неорганического фосфата (2 моля/молекулу) и тирозина (1 моль/молекулу). Обезвоженный YCN-PIG хранили при -80°С до использования, а затем суспендировали в H2O, содержащей 2 мМ DTT, в конечной концентрации 100 мкМ.
Получение крысиных адипоцитов и их инкубация с PIG(-P)/YCN
Из жировых тел придатка яичка самцов крыс Sprague Dawley (140-160 г, накормленные досыта) путем обработки коллагеназой выделяли адипоциты и инкубировали их в KRH-буфере (0,14 мМ NaCl, 4,7 мМ KCl, 2,5 мМ CaCl2, 1,2 мМ MgSO4, 1,2 мМ KH2PO4, 20 мМ Hepes/KOH, рН 7,4), содержащем 1% (мас./об.) BSA, 100 мкг/мл гентамицина, 100 мМ 1-метил-2-фенилэтиладенозина, 0,5 ед./мл аденозиндезаминазы, 0,5 мл натрийпирувата и 5 мМ D-глюкозы, в присутствии PIG(-P)/YCN (растворенного в 20 мМ Hepes/KOH, рН 7,4, 2 мМ DTT) при 37°С на качалке с водяным термостатом при постоянном барботировании с помощью 5% СО2/95% О2 в течение указанных периодов.
Обработка крысиных адипоцитов с помощью трипсин/NaCl или NEM
Для обработки трипсин/NaCl-ом, 2 мл суспензии адипоцитов (3,5 х 106 клеток/мл) в KRH-буфере, содержащем 5 мМ глюкозы, инкубировали (20 мин, 30°С) в присутствии 100 мкг/мл трипсина. Приливали ингибитор трипсина из соевых бобов (конечная концентрация 100 мкг/мл) и 2 мл KRH, содержащего 1 М NaCl и 0,5% BSA, и продолжали инкубацию (10 мин, 22°С). При NEM-обработке 1 мл суспензии адипоцитов (3,5 х 106 клеток/мл) в KRH, содержащем 5 мМ глюкозы, инкубировали (30 мин, 25°С) вместе с NEM (1,5 мМ конечная концентрация), а затем с DTT (15 мМ конечная концентрация, 5 мин). После данных обработок клетки центрифугировали (1500 xg, 5 мин, бакет-ротор), а нижний слой удаляли отсасыванием. В оставшуюся суспензию клеток добавляли 10 мл KRH, содержащего 0,5% BSA, после чего вновь центрифугировали (500 xg, 1 мин, бакет-ротор). После двух дополнительных стадий промывки конечную суспензию данных клеток доводили до 25 мл с помощью KRH, содержащего 0,5% BSA, 50 мкМ глюкозы и 1 мМ натрийпирувата. Порции по 0,2 мл анализировали на липогенез для контроля утраты ответной реакции к PIG41. Контрольные клетки подвергали такому же центрифугированию и промывочным операциям, как и обработанные клетки, но с H2O вместо трипсин/NaCl. Для меченья адипоцитов с помощью [C14]NEM данную суспензию клеток центрифугировали (500 xg, 1 мин), а нижний слой удаляли. Порции по 50 мкл (7 х 106 клеток/мл) инкубировали (10 мин, 30°С) с 2,5 мкКи [C14]NEM в общем объеме 60 мкл. После добавления 5 мкл 10 мМ DTT и 55 мкл KRH, содержащего 10 мМ глюкозы, осуществляли трипсин/NaCl-обработку, как описано выше, в общем объеме 200 мкл. Порции по 50 мкл тщательно наслаивали поверх 200 мкл-овых слоев масла, состоящих из динонилфталата, в 0,4 мл-овых центрифужных пробирках. После центрифугирования (5000 xg, 15 сек) указанные пробирки разрезали по масляному слою. Белки данной среды, содержащиеся в нижней части центрифужных пробирок, осаждали (10% ТХУ, две промывки ацетоном), суспендировали в буфере Лэммли для образцов и анализировали с помощью SDS-ПААГ.
Получение плазматических мембран, тотальных клеточных лизатов и микросом
Из выделенных крысиных адипоцитов, которые описаны раньше, получали постядерный нижний слой. Для получения плазматических мебран 1 мл-овые порции наслаивали сверху на 5 мл-овые подушки, состоящие из 38% (мас./об.) сахарозы, 25 мМ Трис/HCl (рН 7,4), 1 мМ ЭДТА, и центрифугировали (110000 xg, 1 ч). Мембраны в интерфазе между двумя слоями (0,5 мл) удаляли отсасыванием, разбавляли четырьмя объемами буфера для гомогенизации, и наслаивали сверху на 8 мл-овую подушку, состоящую из 28% Перколла, 0,25 М сахарозы, 1 мМ ЭДТА, 25 мМ Трис/HCl (рН 7,0). После центрифугирования (45000 xg, 30 мин) плазматические мембраны из нижней трети данного градиента удаляли с помощью пастеровской пипетки, разбавляли 10 объемами буфера для гомогенизации и центрифугировали (200000 xg, 90 мин). Для изучения связывания полученный промытый осадок суспендировали в буфере для связывания до концентрации 1-2 мг белка/мл. Для получения тотальных клеточных лизатов постядерный нижний слой пополняли дезоксихолатом и Nonidet P-40 (конечная концентрация, соответственно, 0,3 и 0,2%), инкубировали (1 ч, 4°С) и окончательно центрифугировали (100000 xg, 1 ч, 4°С). Полученный супернатант использовали для иммунопреципитации. Для получения микросом центрифугировали постядерный супернатант (100000 xg, 1 ч, 4°С). Полученный центрифужный осадок суспендировали в буфере для связывания до концентрации 1-2 мг белка/мл.
Получение hcDIG/lcDIG
Выделенные очисткой осажденные центрифугированием плазматические мембраны (0,5-1 мг) суспендировали в 1,5 мл ледяного 0,5 М Na2CO3 (рН 11,0), содержащем 50 мМ NaF, 5 мМ натрийпирофосфата, 10 мкМ okadaic acid, 1 мМ натрийортованадата, 20 мкМ лейпептина, 5 мкМ пепстатина, 1 мкМ апротинина, 5 мМ йодацетата, 20 мкМ PMSF, 1 мМ ЭДТА, и инкубировали (1 ч, 4°С при циклическом перемешивании встряхиванием и отсасывании с помощью пипетки). Затем данную суспензию смешивали с равным объемом 85%-й сахарозы в 15 мМ MES/KOH (рН 6,5), 75 мМ NaCl и наслаивали ее на 1,5 мл-овые подушки, каждая из которых состоит из 42,5, 35, 28, 22, 15 и 5% сахарозы в одной и той же среде, и центрифугировали (230000 xg, ротор SW41 Beckman, 18 ч). Светорассеивающие опалесцирующие полосы хлопьевидного вещества в 15-22% (фракции 4 и 5) и 28-35% (фракции 8 и 9) сахарозных интерфазах, а также хлопьевидного вещества в 42,5% подушках (фракции 12-15) были собраны в качестве hcDIG, lcDIG и солюбилизированных плазматических мембранных белков, соответственно, с использованием иглы №19 и шприца (0,75 мл на фракцию). Плотность определяли путем измерения коэффициента преломления указанных фракций. hc/lcDIG характеризовали по обогащению/утрате обнаруживаемых маркеров, как описано раньше. Для изучения связывания, hc/lcDIG суспендировали в буфере для связывания (15 мМ Mes/KOH, рН 6,5, 0,25 М сахарозы, 75 мМ NaCl, 2 мМ MgCl2, 0,5 мМ ЭДТА, 0,5 мМ DDT, протеазные ингибиторы).
Связывание меченного радиоактивным изотопом YCN-PIG или lcGce1p с субклеточными фракциями
10 мкл меченного радиоактивным изотопом YCN-PIG или lcGce1p (60000-80000 распадов в минуту/нмоль, конечная концентрация 5 мкМ) приливали к 40 мкл суспендированных плазматических мембран, микросом или hc/lcDIG (40-80 мкг белка) в буфере для связывания в отсутствие или в присутствии немеченого конкурента (как указано в подписи к данной фигуре) в общем объеме 100 мкл, и инкубировали (30 мин, 4°С). Для отделения мембран от инкубационной среды 45 мкл-овые аликвоты осторожно наслаивали поверх 200 мкл-овых слоев масла, состоящих из дибутилфталата и диоктилфталата (1/1 по об., конечная плотность 1,012), в случае плазматических мембран/микросом, или же состоящих из дибутилфталата и динонилфталата (1/9 по об., конечная плотность 9,863) в случае hc/lcDIG в 0,4 мл-овых предварительно охлажденных (4°С) центрифужных пробирках (микропробирки №72.700, Sarstedt, Германия). После центрифугирования (48000 xg, 2 мин), данные пробирки с закрытыми колпачками резали по масляному слою на нижнюю и верхнюю части (с удаленными колпачками), содержащие осажденные центрифугированием плазматические мембраны/микросомы и плавучие hc/lcDIG, которые, соответственно, проникали или не проникали внутрь слоя масла, переносили в 10 мл-овые сцинтиляционные флаконы, содержащие 1 мл 10%-го SDS. После тщательного встряхивания (16 ч, 25°С), считывали радиоактивность в 9 мл ACSII-сцинтилляционном коктейле (Beckman). В этих условиях прилипающие к стенкам пробирки и распределенные в масляном слое радиоактивно меченные YCN-PIG и lcGce1p давали 50-120 распадов в минуту (т.е. меньше чем 0,5% от общей радиоактивности, используемой на инкубацию) и поэтому не учитывались при вычислении данных по связыванию. Обычно, в соответствии с данными определения белка, плазматических мембран и микросом извлекали, соответственно, 78-85; и 65-80%, а hcDIG и lcDIG - соответственно, 83-92% и 70-78%.
Химический синтез PIG(-P)
Гидрофильные GPI-структуры можно получить из природных источников с помощью двух экспериментальных методов: (i) PIG-высвобождением с помощью GPI-специфичного PLC/D из свободных GPI-липидов в виде их полярных центральных гликановых головных групп, и поэтому не имеющих каких-либо аминокислот, и (ii) PIG-P, полученных с помощью сочетанного липолитического и протеолитического расщепления GPI-белка, дающего полярную центральную гликановую головную группу вместе с одной или несколькими аминокислотами, извлеченными из карбоксиконца оставшегося GPI-белка. И GPI-липидный и GPI-белковый остаток в наружном листке плазматической мембраны эукариотических клеток вместе с центральными гликановыми головными группами сохраняются у организмов от дрожжей до человека. Для определения связывания центральной гликановой головной группы GPI использовали синтез с радиоактивной меткой аутентичной PIG(-P)-структуры, который описан у "Müller и соавт., Endocrinology 138, 3459-3475, 1997"; YCN-PIG получали из радиохимически чистого GPI-белка, Gce1p, плазматической мембраны S. cerevisiae, которые метаболически метили с помощью мио-[14С]инозита, путем последовательного протеолитического и липолитического расщепления in vitro. Для определения соотношения структуры и активности в связывании использовали химически синтезируемые YCN-PIG и его производные. (Фиг. 1: YCN-PIG; Фиг. 2: YMN-PIG; Фиг. 3: PIG37; Фиг.4: YCN).
Синтез трипептида YCN-PIG осуществляли посредством известной в данной области техники пептидного синтеза. Гексахарид синтезировали с использованием трихлорацетимидного метода, который описан у "Frick и соавт., Biochemistry 37, 13421-13436, 1998". Ключевая стадия синтеза PIG-P заканчивалась образованием фосфодиэфирной связи. Из разных испытанных методов наиболее продуктивен H-фосфонатный метод.
Снятие защиты с конечных соединений осуществляли под натрием в жидком NH3, усиленное присутствием цистеина (гидратация с палладием невозможна) и кислотонеустойчивым циклофосфатом. Все соединения были охарактеризованы с помощью масс-, H1-ЯМР-, C13-ЯМР- и P31-ЯМР-спектроскопии.
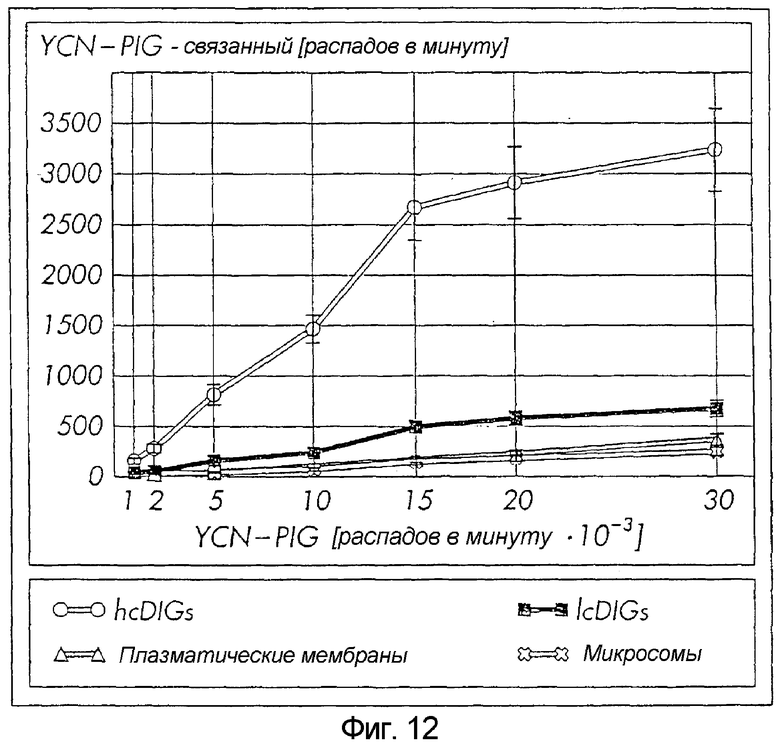
Специфическое связывание PIG(-P) с hcDIG
Тотальные плазматические мембраны, полученные их нестимулированных адипоцитов с помощью дифференциального центрифугирования, были обогащены (в противоположность тотальным клеточным лизатам) специфическими маркерными ферментами данной плазматической мембраны. Уабаин-чувствительная пара-нитрофенилфосфатаза (соответствующая каталитической субъединицы Na+/K+-АТФазы) была обогащена в 9,5 раза, а Nuc в 10,9 раза (в соответствии с ферментными активностями), β1-интегрин - в 13,9 раза и синтаксин-1 - в 16,4 раза (в соответствии с иммуноблотингом), а Gce1 - в 7,8 раза (в соответствии с фотоаффинным меченьем). Одновременно, данный препарат плазматической мембраны был обеднен (в противоположность тотальным клеточным лизатам) маркером саркоплазматического ретикулума, ЭГТА-чувствительной Ca2+-аденозинтрифосфатазой, в 5,7 раза, и эндосомным маркером, SCAMP, («секретарь-переносчик»/ мембранный белок) 37/39, в 8,5 раза, и GLUT4 (транспортер глюкозы 4), в 16,9 раза (в соответствии с иммуноблотингом). Микросомы из нестимулированных адипоцитов были обогащены, в противоположность тотальным клеточным лизатам, по GLUTA4 в 14,4 раза, SCAMP 37/39 в 8,5 раза, рецептору трансферрина в 6,9 раза и IGFII-рецептору в 9,7 раза, но были обеднены, в противоположность тотальным клеточным лизатам, по пара-нитрофенилфосфатазе в 24,6 раза, Gce1 - в 12,5 раза, Nuc - в 15,8 раза, β1-интегрин - в 39,5 раза и синтаксин-1 - в 48,5 раза, в соответствии с иммуноблотингом, а Ca2+-аденозинтрифосфатазной активности - в 19,9 раза. Это указывает на то, что данная фракция представляла первичный эндоплазматический ретикулум и эндосомные структуры и что она практически лишена фрагментов плазматических мембран и саркоплазматического ретикулума. hsDIG и lcDIG получали из нестимулированных адипоцитов на основе их нерастворимости в 0,5 М Na2CO3 (рН 11,0) и низкой плавучей плотности при центрифугировании в градиенте плотности сахарозы. Их характеризовали по утрате ими (в противоположность тотальным плазматическим мембранам) GLUT4 и β-субъединицы рецептора инсулина. hcDIG и lcDIG отличались друг от друга по значительно более высокому обогащению кавеолином, pp59Lyn и Gce1 в hcDIG по сравнению с lcDIG.
Выделенные субклеточные мембранные фракции инкубировали с увеличивающимися количествами меченного радиоактивным изотопом YCN-PIG, и данную инкубацию завершали быстрым отделением от инкубационной среды центрифугированием через слой масла надлежащей плотности.
Мембрано-ассоциированный YCN-PIG преимущественно извлекали с hcDIG, и в меньшей степени с lcDIG, концентрационно-зависимым и насыщающим образом, тогда как плазматические мембраны и микросомы были практически лишены радиоактивной метки (Фиг. 5). В линейном диапазоне неспецифическое связывание YCN-PIG с hcDIG составляло менее чем 20%, что оценивается по 500-кратному избытку немеченного синтетического YCN-PIG или других конкурентов (Фиг. 5). Последующие эксперименты осуществляли с использованием концентрации YCN-PIG, соответствующей окончанию связывания в линейном диапазоне.
Другие способы определения рецептор-лигандного взаимодействия, такие как быстрая фильтрация и центрифугирование на основе седиментации, а не плотности, не позволяли обнаруживать специфическое связывание YCN-PIG с какой-либо мембранной субфракций адипоцита (данные не представлены), вероятно, из-за данной среды связывания по сродству и/или высокой скорости диссоциации. Scatchard-графический анализ показал Kd в диапазоне 50 нМ-500 нМ и Bmax 50-200 пмоль на мг белка hcDIG. Специфичность связывания YCN-PIG с hcDIG продемонстрирована по существенно сниженной эффективности пептидных вариантов, YMN-PIG и PIG37, не содержащих пептидилэтаноламидил-составляющей, а также очень низкой активности одной лишь пептидилэтаноламидил-составляющей, YCN, в конкурентном анализе (Фиг.6).
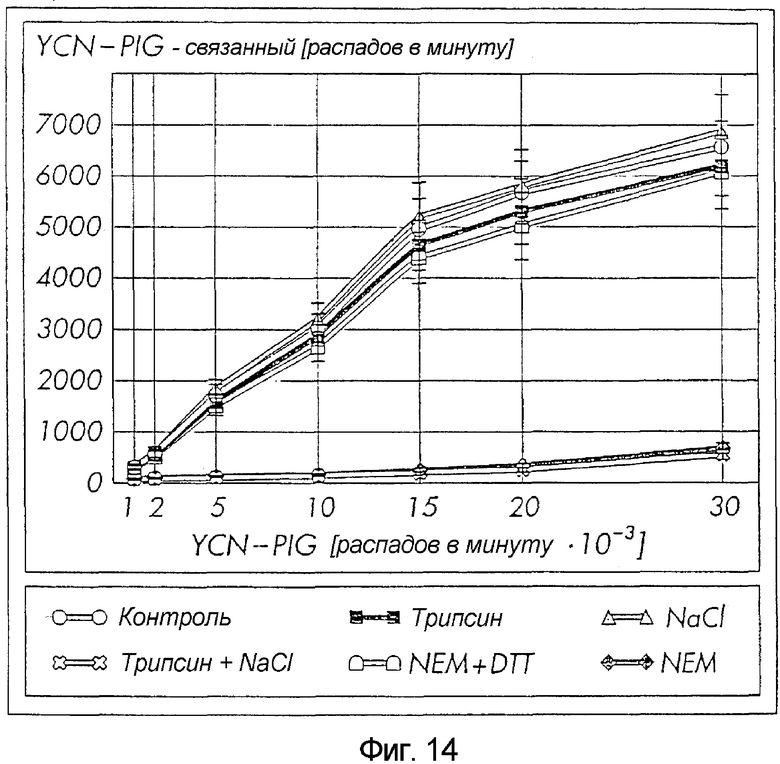
Объединение немеченой YCN и PIG37 (эквимолярное соотношение) смещало связывание радиоактивно меченного YCN-PIG с hcDIG лишь чуть менее эффективно, чем немеченный YCN-PIG, и более сильно, чем только PIG или пептидилэтаноламидил-составляющая, а также YMN-PIG. Этот факт демонстрирует одновременное и синергическое распознавание PIG и пептидилэтаноламидил-составляющих. IC50 конкуренции был лишь в 3-4 раза выше для YCN и PIG37 по сравнению с ковалентно присоединенным YCN-PIG (Фиг. 6). Дополнительно выясняли вопрос, действительно ли идентифицируемый сайт связывания для PIG(-P) имеет белковоподобную природу. hcDIG предварительно обрабатывали трипсин/NaCl или NEM, а затем инкубировали с увеличивающимися концентрациями радиоактивно меченного YCN-PIG в отсутствие или в присутствии избытка немеченого синтетического YCN-PIG (для оценки неспецифического связывания).
Последовательная обработка трипсином и 0,5 М NaCl или обработка с помощью NEM полностью аннулировала специфическое связывание радиоактивно меченного YCN-PIG с hcDIG, а трипсин или NaCl по отдельности или NEM в присутствии DTT не имели существенного эффекта (Фиг. 7). Идентичный пример инактивации наблюдали при более низком взаимодействии по сродству YCN-PIG с lcDIG. Эти данные свидетельствуют о существовании чувствительного к действию трипсина/NaCl и NEM белка, связывающего PIG(-P) с DIG клеточной поверхности адипоцита. Предпочтение YCN-PIG в связывании с hcDIG по сравнению с lcDIG подтверждалось в результате их превращения в процессе истощения холестерина в плазматической мембране адипоцитов с использованием m-βCD и последующего анализа hc/lcDIG на специфическое связывание радиоактивно меченного YCN-PIG. В контрольных адипоцитах основную часть YCN-PIG извлекали вместе с hcDIG, по сравнению с 20%-м остатком, ассоциированным с lgDIG (Фиг. 8). Однако обработка интактных крысиных адипоцитов с помощью m-βCD (1-10 мМ) обнаруживала концентрационно-зависимое снижение количества YCN-PIG, связанного с hcDIG, сопровождающееся соответствующим увеличением lcDIG. Обработка адипоцитов трипсин/NaCl или NEM после истощением холестерина, но перед получением DIG, существенно ухудшала специфическое связывание YCN-PIG и с hcDIG, и с lcDIG (данные не представлены). Эти данные свидетельствуют о преобладающем размещении PIG(-P)-рецептора в hcDIG крысиных адипоцитов, образование которых чрезвычайно зависит от холестерина.
Липолитическое расщепление GPI-белка, специфически связанного с hcDIG
PIG-составляющая, -NH-(CH2)2-O-PO(OH)O-6Manα1(Manα1-2)-2Manα1-6Manα1-4GluN1-6Ino-1,2-(цикло)-фосфат, из YCN-PIG, YMN-PIG и PIG37 (Фиг. 1, 2 и 3), идентична полярной центральной гликановой головной группе у всех эукариотических GPI-белков. Поэтому выясняли, действительно ли белковоподобный сайт связывания для PIG-P взаимодействует с lcGPI-белками, а именно, действительно ли он распознает PIG(-P)-составляющую при присоединении к полной полипептидной части GPI-белка. В целях получения меченного радиоактивным изотопом lcGPI-белка, Gce1p из метаболически меченых клеток S. cerevisiae обрабатывали с помощью PI-специфичного PLC (B. cereus), и гидрофильно расщепленный продукт выделяли очисткой до радиохимической гомогенности. Используя тот же метод центрифугирования в масле, что и для PIG(-P), обнаружили, что lcGce1p концентрационно-зависимым образом ассоциировался с DIG из выделенных крысиных адипоцитов и насыщающим образом - с hcDIG, которые в 11-15 раз более эффективны, чем lcDIG. Неспецифическое связывание в присутствии 200-кратного молярного избытка немеченого lcGce1p составляло менее 15% от тотального lcGce1p, извлеченного с DIG в ненасыщающих концентрациях lcGce1p. В соответствии со Scatchard-графическим анализом, Kd для lcGce1p-связывания с hcDIG находится в диапазоне 0,1-1,0 мкМ и Bmax с 70-200 пмоль на мг белка hcDIG. Тотальные плазматические мембраны и микросомы не обнаруживали специфического связывания lcGce1p. Таким образом, hcDIG плазматических мембран адипоцитов, очевидно, включали сайты специфического связывания для lcGce1p из дрожжей. В дальнейшем анализе идентичности сайтов связывания для PIG(-P) и lcGPI-белков, как показано с помощью аналогичных значений Kd и Bmax, относительное сродство синтетических PIG(-P)-соединений для lcGce1p-связывающего сайта в hcDIG, сравнивали в конкурентных исследованиях (Фиг. 9).
Связывание радиоактивно меченного lcGce1p с hcDIG было смещено избытком (более чем 500-кратным) меченных синтетических YCN-PIG, YMN-PIG и YCN плюс PIG37, более чем на 75% от общего lcGce1p-связывания, подтверждая специфичность взаимодействия lcGce1p и hcDIG. Конкуренция связывания lcGce1p с PIG37 и YCN оказалась существенно менее эффективной. Относительное ранжирование различных PIG(-P), которое отражено средним значением IC50 по вытеснению lcGce1p из hcDIG, составило YCN-PIG>YCN+PIG37>YMN-PIG>PIG37>YCN и, таким образом, идентично тому, которое препятствует связыванию YCN-PIG (Фиг.6). Кроме того, средние значения IC50 весьма сходны для конкуренции lcGce1p- и YCN-PIG-связывания, указывая, что в обоих случаях распознаются одни и те же детерминанты, а остаточная белковая составляющая GPI-белка (за исключением карбоксиконцевого трипептилэтаноламидил-остатка) не способствуют связыванию. Далее, чувствительность взаимодействия lcGce1p c hcDIG к трипсин/NaCl- и NEM-обработке интактных крысиных адипоцитов изучали в условиях, которые почти полностью нарушают связывание меченного изотопом YCN-PIG (Фиг. 7). hcDIG из трипсин/NaCl-, а также NEM-обработанных адипоцитов проявляли связь с радиоактивно меченным lcGce1p, не превышая неспецифическое связывание в присутствии 500-кратного избытка немеченого YCN-PIG (доля которого составляет около 30% от общего Gce1p, извлекаемого с hcDIG из необработанных контрольных клеток) (Фиг. 10). В противоположность этому, инкубация адипоцитов с NEM в присутствии избытка DTT (Фиг. 10), либо отдельно с трипсином или NaCl (данные не представлены), не ухудшала связывание радиоактивно меченного lcGce1p и его конкуренцию с 3 мкМ YCN-PIG37, 5 мкМ PIG37 и 10 мкМ YCN по сравнению с необработанными клетками. Вместе взятые, сайты, специфически связывающие YCN-PIG и lcGec1p, проявляли весьма сходные характеристики относительно локализации в hcDIG плазматической мембране адипоцитов, абсолютного и относительного сродства (со структурными производными), уровня экспрессии и чувствительность в отношении и к трипсину/NaCl и к NEM.
Эндогенные лиганды для рецептора PIG(-P) и lcGPI-белков
Кандидатами в физиологические лиганды несомненно идентичных сайтов связывания для PIG(-P)- и lcGPI-белков являются нерасщепляемые GPI-структуры, а именно GPI-липиды и/или GPI-якорные белки. Для проверки данного предположения выделенные крысиные адипоциты подвергали обработке с помощью различных GPI-специфичных PL и последующей солевой промывке (0,5 М NaCl) до получения hcDIG с целью специфически удалить предполагаемые эндогенные GPI-молекулы, которые взаимодействуют с рецептором и тем самым маскировали сайты связывания для YCN-PIG/lcGce1p.
Инкубация крысиных адипоцитов с увеличивающимися концентрациями PI-специфичного PLC из B. cereus или GPI-специфичного PLD из сыворотки человека давала концентрационно-зависимое увеличение количества радиоактивно меченных YCN-PIG и Gce1p, которые специфически связываются с hcDIG (Фиг. 11). Эффективность липолитических обработок продемонстрирована параллельно, путем потери Gce1p и Nuc у hcDIG.
Их потери, соответственно, на 75% и 65%, коррелируют с увеличением связывания YCN-PIG или lcGce1p с hcDIG до 200% и 260%. Специфичность GPI-расщеплений демонстрировалась полной неспособностью PC-специфичного PLC (B. cereus) и PLD из капусты (которые не атакуют GPI-структуры) к существенному вытеснению Gce1p или Nuc из hcDIG, а также к стимуляции связывания YCN-PIG (lcGce1p) с hcDIG (Фиг. 11, 12). Scatchard-графический анализ специфического связывания с hcDIG из PI-специфичных предварительно обработанных PLC адипоцитов (неспецифическое связывание существенно не изменялось) показал, что увеличенная ассоциация радиоактивно меченного YCN-PIG/lcGce1p обусловлена, главным образом, 2-3-кратным возрастанием Bmax и почти неизменяемой Kd. Эти данные свидетельствуют о том, что около 50% сайтов связывания PIG(-P)- или lcGPI-белков в hcDIG в выделенных крысиных адипоцитах в основном состоянии, заняты эндогенными GPI-структурами, расщепляемыми с помощью (G)PI-специфичного PLC/D. Примечательно, что инсулин в физиологической концентрации имитировал в некоторой степени действие GPI-специфичной PLC/D-обработки в крысиных адипоцитах, вызывая умеренное, но все же существенное снижение количеств Gce1p и Nuc в hcDIG. Инсулин-индуцируемая потеря GPI-белков из hcDIG ведет к отчетливому увеличению связывающих способностей YCN-PIG или lcGce1p (Фиг. 11, 12).
Кроме того, можно было бы продемонстрировать, что рецептор для PIG(-P)- и lcGPI-белков идентичен 115 кДа - белку, чувствительному к действию трипсин/NaCl и NEM, который назвали CIR.
Связывание PIG-P с данным рецептором влияет на его доступность для последующей ковалентной модификации с помощью NEM и/или расщеплению и высвобождению из клеточной поверхности адипоцита с помощью трипсин/NaCl.
Крысиные адипоциты инкубировали с PIG(-P) и затем последовательно подвергали мечению с помощью [C14]NEM и обработке с помощью трипсин/NaCl. С помощью фосфоресцирующего изображения и SDS-ПААГ анализа высвобождаемых радиоактивно меченых полипептидов и (Фиг. 13), обнаруживали, что PIG(-P) уменьшал сшивку 115 кДа-полипептида с помощью [C14]NEM, и/или его извлечение из нижнего слоя адипоцитов после трипсин/NaCl-обработки. Уменьшение с помощью YCN-PIG или PIG37 в 3 мкМ и YCN в 30 мкМ составило, соответственно, 83, 65 и 28%, по сравнению с контрольными клетками. Данный белок представлял лишь основной NEM-меченый компонент, который высвобождался из плазматических мембран с помощью трипсин/NaCl, но не с помощью какой-либо одной обработки (Фиг. 13), и идентичен CIR. В соответствии с экспериментальным доказательством существования эндогенных лигандов (например, GPI-белков) и их удаления из соответствующего связывающего сайта липолитическим отщеплением (смотрите Фиг. 11, 12), обработка адипоцитов с помощью экзогенного PI-специфичного PLC (B. cereus) или инсулина слегка, но воспроизводимо, стимулировала трипсин/NaCl-зависимое высвобождение [C14]NEM-меченного CIR, соответственно, на 30% и 20% (Фиг. 13). Так как относительное соотношение высвобождаемого CIR из клеточной поверхности адипоцита с помощью трипсин/NaCl-обработки, по сравнению с трипсиновой обработкой и с NaCl-обработкой (100/20/10), было приблизительно сопоставимым в контроле, PIG(-P)-стимулированных и PLC/инсулин-обработанных клетках, связывание PIG(-P) и эндогенного GPI-лигандов с hcDIG, по-видимому, ослабляет мечение CIR с помощью NEM, а не его триптическое расщепление. Это вызвано конформационным изменением CIR, выявляемого в результате взаимодействия лигандов с PIG(-P)-рецептором в hcDIG плазматической мембраны адипоцита.
H1- и C13-химические сдвиги сигнала [м.д.] для YCN-PIG в D2O, pD=8,1 (некорр.)
Перечень Фигур
Фиг. 1: Общая схема синтеза PIG, часть 1
Фиг. 2: Общая схема синтеза PIG, часть 2
Фиг. 3: Общая схема синтеза PIG, часть 3
Фиг. 4: Синтез YCN-PIG, часть 1
Фиг. 5: Синтез YCN-PIG, часть 2
Фиг. 6: Синтез YCN-PIG, часть 3
Фиг. 7: Синтез YCN
Фиг. 8: Химическая формула YCN-PIG
Фиг. 9: Химическая формула YMN-PIG
Фиг. 10: Химическая формула PIG37
Фиг. 11: Химическая формула YCN
Фиг. 12: Специфическое связывание PIG(-P) с hcDIG. Увеличенные количества радиоактивно меченного YCN-PIG, выделяемого из S. cerevisae, инкубировали (1 ч, 4°С) с hcDIG (6,5 мкг белка), lcDIG (6,5 мкг), плазматическими мембранами (47,5 мкг) и микросомами (68 мкг) из выделяемых крысиных адипоцитов.
Мембранные фракции/DIG подвергали центрифугированию в слое масла, извлекали с/из осадком / верхнего слоя масла, солюбилизировали и считывали радиоактивность. Специфическое связывание вычисляли в виде разницы между радиоактивностью, измеряемой в отсутствие и в присутствии 10 мкМ немеченого YCN-PIG. Каждая точка соответствует средней ± стандартное отклонение (SD) трех инкубаций с использованием, по меньшей мере, 4-х разных мембранных препаратов.
Фиг. 13: Специфическое связывание PIG-P с hcDIG:
Радиоактивно меченный YCN-PIG (18000-22000 распадов в минуту) инкубировали (1 ч, 4°С) с hcDIG (6,5 мкг белка) в отсутствие или в присутствии увеличивающихся количеств немеченых YCN-PIG, YCN+PIG37, YMN-PIG, PIG37 и YCN (конкуренция). Мембранные фракции/DIG подвергали центрифугированию в слое масла, извлекали с/из осадком / верхнего слоя масла, солюбилизировали и считывали радиоактивность.
Фиг. 14: Характеристика связывающего сайта для PIG-P в hcDIG
Увеличивающиеся количества радиоактивно меченного YCN-PIG, выделяемого из S. cerevisiae, инкубировали (1 ч, 4°С) с hcDIG (6,5 мкг белка) из выделенных крысиных адипоцитов, которые предварительно обрабатывали с помощью трипсин/NaCl, трипсина, NEM+DDT, NaCl или NEM, либо оставляли необработанными (Контроль). DIG подвергали центрифугированию в слое масла, извлекали из верхнего слоя масла, солюбилизировали и считывали радиоактивность. Специфическое связывание вычисляли в виде разницы между радиоактивностью, измеряемой в отсутствие и в присутствии 10 мкМ немеченого YCN-PIG. Каждая точка соответствует средней ± стандартное отклонение (SD) трех инкубаций с использованием, по меньшей мере, 3 разных адипоцитных предварительных обработок.
Фиг. 15: Характеристика связывающего сайта для PIG-P в hcDIG
Радиоактивно меченный YCN-PIG (12000-18000 распадов в минуту) инкубировали (1 ч, 4°С) с определенными (пропорциональными) количествами hcDIG и lcDIG, полученными из выделенных крысиных адипоцитов, которые предварительно обрабатывали (50 мин, 30°С) увеличивающимися концентрациями m-βCD, либо оставляли необработанными. DIG подвергали центрифугированию в слое масла, извлекали из верхнего слоя масла, солюбилизировали и считывали радиоактивность, измеряемую в отсутствие и в присутствии 10 мкМ немеченого YCN-PIG. Каждая точка соответствует средней ± стандартное отклонение (SD) трех инкубаций с использованием, по меньшей мере, 3-х разных предварительных обработок адипоцитов.
Фиг. 16: Специфическое связывание lcGce1p c hcDIG
Радиоактивно меченный Gce1p, полученный из S. cerevisiae и обработанный с помощью PI-специфичного PLC (B. cereus), инкубировали (1 ч, 4°С) с hcDIG (6,5 мкг белка), выделенных из необработанных крысиных адипоцитов, в отсутствие или в присутствии немеченого PIG-P. hcDIG подвергали центрифугированию через слой масла, солюбилизировали и считывали радиоактивность.
Каждая точка соответствует средней ± стандартное отклонение (SD) четырех инкубаций с использованием, по меньшей мере, 3-х разных hcDIG-препаратов и предварительных обработок адипоцитов соответственно.
Фиг. 17: Специфическое связывание lcGce1p c hcDIG.
Радиоактивно меченный Gce1p, полученный из S. cerevisiae и обработанный с помощью PI-специфичного PLC (B. cereus), инкубировали (1 ч, 4°С) с hcDIG (6,5 мкг белка), выделенных из адипоцитов, которые предварительно обрабатывали с помощью трипсин/NaCl, NEM, NEM+DTT или оставляли необработанными (Контроль), в отсутствие или в присутствии немеченного YCN-PIG (конечная концентрация 3 мкМ), YCN+PIG37 (3 мкМ), PIG37 (5 мкМ) и YCN (10 мкМ). hcDIG подвергали центрифугированию через слой масла, солюбилизировали и считывали радиоактивность. Каждая точка соответствует средней ± стандартное отклонение (SD) четырех инкубаций с использованием, по меньшей мере, 3-х разных hcDIG-препаратов и предварительных обработок адипоцитов соответственно.
Фиг. 18: Влияние PL и инсулиновой обработки адипоцитов на связывание YCN-PIG и lcGce1p c hcDIG. Выделенные крысиные адипоциты (7х107 клеток/мл) инкубировали (30 мин, 30°С) с указанными количествами PI-специфического PLC (B. cereus), PC-специфического PLC (B. cereus), GPI-специфического PLD (сыворотка человека) или PLD (капуста) или инсулина человека в общем объеме 2 мл при слабом встряхивании под 5% СО2/95% О2. После добавления 2 мл 1 М NaCl, указанные адипоциты промывали флотацией. Выделяли hcDIG, и 6,5 мл-овые аликвоты инкубировали (1 ч, 4°С) с радиоактивно меченным lcGce1p, получаемым из S. cerevisiae, и YCN-PIG (15000-25000 распадов в минуту) в отсутствие или в присутствии немеченого YCN-PIG (конечная концентрация 10 мкМ), подвергали центрифугированию через слой масла, извлекали из верхнего слоя масла, солюбилизировали и считывали радиоактивность. Специфическое связывание вычисляли в виде разницы между отсутствием и наличием YCN-PIG.
Каждая точка соответствует средней ± стандартное отклонение (SD) трех инкубаций с использованием, по меньшей мере, двух разных hcDIG-препаратов.
Фиг. 19: Влияние PL и инсулиновой обработки адипоцитов на связывание YCN-PIG и lcGce1p c hcDIG. Выделенные крысиные адипоциты (7х107 клеток/мл) инкубировали (30 мин, 30°С) с указанными количествами PI-специфического PLC (B. cereus), PC-специфического PLC (B. cereus), GPI-специфического PLD (сыворотка человека) или PLD (капуста) или инсулина человека в общем объеме 2 мл при слабом встряхивании под 5% СО2/95% О2. После добавления 2 мл 1 М NaCl указанные адипоциты промывали флотацией. Выделяли hcDIG, и 6,5 мл-овые аликвоты инкубировали (1 ч, 4°С) с радиоактивно меченным lcGce1p, полученных из S. cerevisiae, и YCN-PIG (15000-25000 распадов в минуту) в отсутствие или в присутствии немеченого YCN-PIG (конечная концентрация 10 мкМ), подвергали центрифугированию через слой масла, извлекали из верхнего слоя масла, солюбилизировали и считывали радиоактивность.
Фиг. 20: Действие PIG(-P), PI-специфического PLC и инсулина на NEM-мечение CIR. Выделенные крысиные адипоциты инкубировали (30 мин, 37°С) в отсутствие (Контроль) или в присутствии PIG37, YCN-PIG, YCN, PO-PLC (B. cereus) либо инсулина в данной концентрации и затем метили с помощью [C14]NEM. После обработки трипсин/NaCl, как указано, адипоциты отделяли от инкубационной среды центрифугированием через слой масла.
Белки извлекали из данной среды (ниже слоя масла) и разделяли в SDS-ПААГ.
Фосфоресцирующие изображения представлены из обычного эксперимента с тремя повторами со сходными результатами. Количественная оценка четырех разных инкубаций адипоцитов с тремя измерениями дана в качестве независимых единиц измерения (средняя±SD) количества CIR, высвобождаемого из трипсин/NaCl-обработанных контрольных клеток, принятого за 100.


















Изобретение относится к биотехнологии. Предложены белок плазматической мембраны адипоцита, способ его получения и комплекс на основе этого белка. Белок имеет молекулярную массу 115 кДа и обладает способностью запускать tyr-фосфорилирование субстрата инсулин-рецепторных белков в адипоците. Способ получения белка предусматривает получение адипоцитов из ткани крысы, мыши или человека и выделение из них плазматических мембран. Далее выделяют множество доменов с высоким содержанием холестерина hcDIG, которые обрабатывают раствором трипсин/NaCl. Проводят центрифугирование и разделяют белковую фракцию SDS-ПААГ электрофорезом. Полученную белковую фракцию размером 115 кДа элюируют из данного геля. Комплекс представляет собой активированный белок и образуется при его соединении с одним из соединений из группы: YCN-PIG, YMN-PIG, YCN или IcGcel. Белок в его активированной форме позволяет регулировать утилизацию глюкозы в обход инсулинового сигнального каскада. 3 н. и 4 з.п. ф-лы, 20 ил., 1 табл.
сродством специфического связывания с фосфоинозитолгликаном или с фосфоинозитолгликанпептидом, характеризующийся
а) молекулярной массой 115 кДа,
б) способностью запускать tyr-фосфорилирование субстрата инсулинрецепторных субстратных белков IRS-1 и IRS-2 в адипоците после специфического связывания фосфоинозитолгликана или фосфоинозитолгликанпептида с данным белком и
в) способностью стимулировать потребление глюкозы адипоцитом после специфического связывания фосфоинозитолгликана или фосфоинозитолгликанпептида с данным белком.
a) адипоциты получают из ткани крысы, мыши или человека,
b) из адипоцитов выделяют плазматические мембраны,
c) из плазматических мембран получают множество доменов с высоким содержанием холестерина (hcDIG),
d) hcDIG обрабатывают раствором трипсин/NaCl,
e) центрифугируют инкубационную смесь n.d], и белки из полученного супернатанта разделяют при помощи SDS-ПААГ,
f) белковую фракцию размером 115 кДа элюируют из данного геля и, по возможности, солюбилизируют с помощью раствора или суспензии, содержащих детергент или биологические мембраны.
| ЕР 0532915 А, 24.03.1993 | |||
| MULLER G | |||
| et al | |||
| Convergence and divergence of the signaling pathways for insulin and phosphoinositol-glycans | |||
| Т MOLECULAR MEDICINE | |||
| BLACKWELL SCIENCE | |||
| CAMBRIDGE, MA, US, vol | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

