Родственная заявка
Настоящая заявка притязает на приоритет согласно статье 119(e) на основании предварительной заявки на выдачу патента США с регистрационным №60/589875, поданной 20 июля 2004, описание которой включено в данную заявку в полном объеме.
Область техники, к которой относится изобретение
Изобретение относится к ангиопоэтинподобному белку 4 (ANGPTL4). Изобретение относится к композициям и способам применения ANGPTL4 и его агонистов и антагонистов для диагностики и лечения заболеваний или расстройств.
Уровень техники
Ангиопоэтинподобный белок 4 (ANGPTL4) является представителем семейства секретируемых белков ангиопоэтинов. Консервативные области семейства ангиопоэтинов включают биспиральный домен и C-концевой фибриногенподобный (FBN-подобный) домен. См., например, Kim et al., Biochem. J. 346: 603-610 (2000). Другие представители семейства включают ангиопоэтин 1, ангиопоэтин 2 и ангиопоэтин 3. Ангиопоэтин 1, ангиопоэтин 2 и ангиопоэтин 3/ангиопоэтин 4 связываются с рецептором Tie2. См., например, Davis et al., Cell 87, 1161-1169 (1996); Maisonpierre et al., Science 277, 55-60 (1997); Valenzuela et al, Proc. Natl. Acad. Sci. USA 96, 1904-1909 (1999) и патенты США №5521073, 5650490 и 5814464. Ангиопоэтины 1 и 4, по-видимому, являются агонистами рецептора Tie2, тогда как ангиопоэтины 2 и 3, по-видимому, являются антагонистами (а может быть, агонистами) рецептора Tie2. См., например, Folkman & D'Amore, Cell, 87: 1153-1155 (1996); Suri et al., Cell, 87: 1171-1180 (1996); Masionpierre et al., Science 277: 55-60 (1997) и Ward & Dumont, Seminars in Cell and Developmental Biology, 13: 19-27 (2002). Рецептор Tie2 относится к семейству специфичных для эндотелиальных клеток тирозинкиназных рецепторов, которые также включают сиротский рецептор Tie1. Обнаружено, что другой представитель семейства, ангиопоэтинподобный белок 3, связывается с интегрином αvβ3. См., например, заявку на выдачу патента США 2003/0215451 и Camenisch et al., J. Biol. Chem., 277 (19): 17281-17290 (2002).
ANGPTL4 известен под другими терминами. Например, ANGPTL4 также известен как фибриноген печени/ангиопоэтин-родственный белок (HFARP) (Kim et al., Biochem. J. 346:603-610 (2000)), PPARγ-ангиопоэтин-родственный белок (PGAR) (Yoon, et al., Mol. Cell Biol., 20: 5343-5349 (2000)) и индуцируемый голоданием адипозный фактор (FIAF) (Kerten et al., J. Biol. Chem., 275: 28488-28493 (2000)).
Исследования in vitro и in vivo и характеристики ANGPTL4 могут обеспечить полезную идентификацию и открытие терапевтических средств и/или лекарственных средств, применимых для профилактики, улучшения состояния или корректировки заболеваний или дисфункций, связанных с активностью и/или экспрессией ANGPTL4. Например, подтверждено, что исследования на культурах ткани и генно-инженерных мышах представляют собой полезные средства для функционального анализа биологических процессов, имеющих отношение к заболеванию человека, включая иммунологию, злокачественные опухоли, нейробиологию, сердечно-сосудистую биологию, ожирение и многие другие. Существует необходимость в раскрытии и понимании многих биологических функций ANGPTL4. Изобретение обращено к указанным и других потребностям, которые будут очевидны при рассмотрении следующего описания.
Сущность изобретения
Изобретение относится к ангиопоэтинподобному белку 4 (ANGPTL4). Изобретение относится к применению ANGPTL4 или его подпоследовательности, или его агониста или антагониста для лечения состояний или заболеваний, характеризуемых аномальной экспрессией или активностью ANGPTL4, и/или состояний или заболеваний, в которые вовлечены экспрессия и/или активность ANGPTL4.
Предлагаются способы модулирования пролиферации гепатоцитов посредством ANGPTL4 или его агонистов или антагонистов. В некоторых вариантах способы включают в себя индукцию пролиферации гепатоцитов. Например, способ включает в себя введение эффективного количества ANGPTL4 или агониста ANGPTL4 в популяцию гепатоцитов или предшественников гепатоцитов, индуцируя таким образом пролиферацию. В одном аспекте стадия введения включает в себя введение нуклеиновой кислоты, которая кодирует ANGPTL4. Альтернативно или дополнительно может быть введено эффективное количество средства, которое индуцирует продукцию ANGPTL4 в гепатоците или предшественнике гепатоцита, чтобы стимулировать пролиферацию. ANGPTL4 или агонисты ANGPTL4 можно применять для лечения дисфункции, заболеваний и повреждения печени с помощью введения эффективного количества ANGPTL4 или агониста. В одном аспекте ANGPTL4 обеспечивается нуклеиновой кислотой, кодирующей ANGPTL4. В одном варианте изобретения агонистом ANGPTL4 является агонист рецептора αvβ5.
Также предлагаются способы ингибирования пролиферации гепатоцитов. В некоторых вариантах способ включает в себя введение эффективного количества композиции, содержащей антагонист ANGPTL4, в популяцию гепатоцитов или предшественников гепатоцитов. В одном аспекте антагонистом ANGPTL4 является средство, которое ингибирует продукцию белка ANGPTL4, например, антисмысловая молекула или молекула рибозима. В одном аспекте антагонистом ANGPTL4 является анти-ANGPTL4-антитело. В другом аспекте антагонистом ANGPTL4 является антагонистическое анти-αvβ5-антитело. В одном варианте антагонистом ANGPTL4 является ANGPTL4-миРНК. Антагонисты ANGPTL4 могут быть использованы для лечения, например, рака печени или нежелательной гипертрофии печени посредством введения эффективного количества антагониста ANGPTL4 в гепатоциты.
Также предлагаются способы модулирования клеточной адгезии гепатоцитов. В некоторых вариантах способы включают в себя индукцию клеточной адгезии гепатоцитов посредством введения эффективного количества композиции, содержащей ANGPTL4 или агонист ANGPTL4, в популяцию гепатоцитов. В других вариантах способы включают в себя ингибирование клеточной адгезии гепатоцитов посредством введения эффективного количества композиции, содержащей антагонист ANGPTL4, в популяцию гепатоцитов с ингибированием при этом клеточной адгезии гепатоцитов.
Кроме модулирования пролиферации и клеточной адгезии гепатоцитов, которые вовлечены в гомеостаз липидов, ANGPTL4 модулирует уровни триглицеридов и холестерина в сыворотке и стимулирует пролиферацию преадипоцитов, которые также вовлечены в гомеостаз липидов. Изобретение относится к способам модулирования ряда различных аспектов гомеостаза липидов. Например, способы согласно изобретению заключаются в стимуляции пролиферации преадипоцитов посредством введения эффективного количества композиции, содержащей ANGPTL4 или агонист ANGPTL4, в популяцию преадипоцитов, индуцируя таким образом пролиферацию преадипоцитов. Также предлагаются способы ингибирования пролиферации преадипоцитов. Например, способы включают в себя введение эффективного количества композиции, содержащей антагонист ANGPTL4, в популяцию преадипоцитов. Также включены способы модулирования клеточной миграции преадипоцитов. Например, способы согласно изобретению включают в себя индукцию клеточной миграции преадипоцитов посредством введения эффективного количества ANGPTL4 или агониста ANGPTL4 в популяцию преадипоцитов. Также предлагаются способы ингибирования клеточной миграции преадипоцитов, которые заключаются, например, во введении эффективного количества антагониста ANGPTL4 в популяцию преадипоцитов с ингибированием при этом клеточной миграции.
Также в изобретении предлагаются способы модулирования уровней триглицеридов или холестерина в сыворотке у субъекта. Например, способы заключаются во введении эффективного количества композиции, содержащей ANGPTL4, или агонист ANGPTL4, или антагонист ANGPTL4 субъекту, модулируя таким образом уровни триглицеридов и/или холестерина в сыворотке у субъекта. В одном варианте вводят ANGPTL4 или антагонист ANGPTL4, который приводит к накоплению триглицеридов и/или холестерина в сыворотке субъекта по сравнению с контролем. В другом варианте субъекту вводят эффективное количество антагониста ANGPTL4, снижая таким образом уровень в сыворотке субъекта по меньшей мере одного из следующих компонентов: триглицеридов, свободных жирных кислот и/или холестерина. В некоторых вариантах изобретения контролем является сыворотка субъекта до лечения или сыворотка субъекта, не подвергаемого лечению или меньше подвергаемого лечению, и т.д.
ANGPTL4 и модулятор ANGPTL4 (его агонист или антагонист) можно использовать для лечения расстройств гомеостаза липидов посредством введения эффективного количества молекулы субъекту. См. «расстройство гомеостаза липидов» в разделе «Определения» в данном описании. Например, способ включает в себя введение субъекту композиции, содержащей антагонист ANGPTL4 в количестве, эффективном для лечения гиперлипидемии.
Также предлагаются способы лечения ожирения и/или уменьшения массы тела субъекта. Например, способ включает в себя введение субъекту эффективного количества модулятора ANGPTL4, осуществляя таким образом лечение ожирения и/или уменьшение массы тела субъекта по сравнению с отсутствием лечения или лечением с помощью контроля. В одном варианте у субъекта снижают ожирение (жир). Таким образом, также можно лечить состояния, связанные с ожирением, например, сердечно-сосудистые заболевания, диабет и т.д.
В некоторых вариантах изобретение осуществляют на клетках, например, гепатоцитах, преадипоцитах, у субъекта. Обычно субъектом является человек.
ANGPTL4 согласно изобретению включает полноразмерный белок, а также биологически активные молекулы, например, остатки, соответствующие N-концу, N-концевому биспиральному домену, C-концу, C-концевому фибриногенподобному домену, или аминокислотную подпоследовательность ANGPTL4 (1-183), ANGPTL4 (23-183), ANGPTL4 (от 1 до примерно 162), ANGPTL4 (примерно 162-406), ANGPTL4 (23-406) или ANGPTL4 (184-406) человеческого ANGPTL4 и/или аминокислотную подпоследовательность mANGPTL4 (1-183), mANGPTL4 (23-183), mANGPTL4 (от 1 до примерно 165), mANGPTL4(от 23 до примерно 165), mANGPTL4 (23-410) или mANGPTL4 (184-410) мышиного ANGPTL4. Другие подпоследовательности также включают без ограничения, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-406, 60-406, 80-406, 100-406, 120-406, 140-406 и 160-406 hANGPTL4 и, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-410, 60-410, 80-410, 100-410, 120-410, 140-410 и 160-410 mANGPTL4. Агонисты ANGPTL4 включают молекулы, которые активируют ANGPTL4 или создают активности ANGPTL4, например, активные полипептиды, малые молекулы (синтетические аналоги) и молекулы, которые увеличивают активность или экспрессию ANGPTL4. Агонисты ANGPTL4 также включают агонисты αvβ5.
Антагонистами ANGPTL4 согласно изобретению являются молекулы, которые ингибируют или уменьшают активность ANGPTL4. Ингибитор ANGPTL4 может включать вещество с низкой молекулярной массой, полинуклеотид, антисмысловые молекулы, аптамеры РНК, рибозимы против ANGPTL4 или полипептидов его рецептора, полипептид, антагонистические варианты ANGPTL4, изолированный белок, рекомбинантный белок, антитело или его конъюгаты или слитые белки, которые ингибируют активность ANGPTL4 прямо или опосредованно. В некоторых вариантах осуществления изобретения антагонистическим ANGPTL4-антителом является антитело, которое ингибирует или уменьшает активность ANGPTL4 в результате связывания со специфичной подпоследовательностью или областью белка ANGPTL4, например, N-концом, N-концевым биспиральным доменом, C-концом, C-концевым фибриногенподобным доменом или аминокислотной подпоследовательностью ANGPTL4 (1-183), ANGPTL4 (23-183), ANGPTL4 (от 1 до примерно 162), ANGPTL4 (примерно 162-406), ANGPTL4 (23-406) или ANGPTL4 (184-406) человеческого ANGPTL4 и/или аминокислотной подпоследовательностью mANGPTL4 (1-183), mANGPTL4 (23-183), mANGPTL4 (от 1 до примерно 165), mANGPTL4 (от 23 до примерно 165), mANGPTL4 (23-410) или mANGPTL4 (184-410) мышиного ANGPTL4. Другие подпоследовательности также включают без ограничения, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-406, 60-406, 80-406, 100-406, 120-406, 140-406 и 160-406 hANGPTL4 и, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-410, 60-410, 80-410, 100-410, 120-410, 140-410 и 160-410 mANGPTL4. В некоторых вариантах осуществления изобретения антагонист ANGPTL4 включает анти-αvβ5-антитело, например, антагонистическое анти-αvβ5-антитело. В некоторых вариантах антитела согласно изобретению являются гуманизированными антителами. В некоторых вариантах осуществления изобретения антагонистом ANGPTL4 является молекула миРНК. В одном варианте молекулой миРНК является молекула ANGPTL4-миРНК, при этом мишенью молекулы является последовательность ДНК (например, GTGGCCAAGCCTGCCCGAAGA) (SEQ ID NO: 3) нуклеиновой кислоты, кодирующей ANGPTL4. Иммуноадгезин ANGPTL4 содержит, по меньшей мере, связывающую рецептор область ANGPTL4, слитую с последовательностью иммуноглобулина. В некоторых вариантах ANGPTL4, агонист или антагонист находится в сочетании с носителем, например, фармацевтически приемлемым носителем.
Описаны трансгенные и нокаутированные по ANGPTL4 животные и также предлагаются применения указанных трансгенных животных. Изобретение также относится к изолированной клетке, полученной из трансгенного животного, отличного от человека, геном которого содержит нарушение гена, который кодирует ANGPTL4. В некоторых вариантах изолированной клеткой является мышиная клетка (например, эмбриональная стволовая клетка).
Нарушения в мутантном гене ANGPTL4 привели к фенотипическим наблюдениям, связанным с различными патологическими состояниями или дисфункциями, включая сердечно-сосудистые, эндотелиальные или ангиогенные расстройства, включая атеросклероз; расстройства, связанные с аномальным метаболизмом, включая расстройства гомеостаза липидов; или иммунологические и воспалительные расстройства. Способы согласно изобретению включают лечение сердечно-сосудистого, эндотелиального или ангиогенного расстройства; расстройства, связанного с аномальным метаболизмом, иммунологического расстройства; расстройства гомеостаза липидов или онкологического заболевания, связанного с нарушением гена, который кодирует ANGPTL4, или связанного с активностью ANGPTL4, посредством введения субъекту эффективного количества ANGPTL4, агониста или антагониста ANGPTL4, таким образом осуществляя эффективное лечение указанного расстройства или заболевания.
Также предлагаются способы идентификации фенотипа, связанного с нарушением гена, который кодирует ANGPTL4. Например, способ включает в себя (a) измерение физиологического признака трансгенного животного, отличного от человека, геном которого содержит нарушение гена, который кодирует ANGPTL4; и (b) сравнение измеренного физиологического признака с признаком животного дикого типа соответствующего пола. Фенотип, возникающий в результате нарушения гена, идентифицируют в виде физиологического признака трансгенного животного, отличного от человека, который отличается от физиологического признака животного дикого типа. Трансгенное животное, отличное от человека, может быть гомозиготным или гетерозиготным в отношении нарушения гена, который кодирует ANGPTL4.
Также предлагаются способы идентификации средства, которое модулирует фенотип, связанный с нарушением гена, который экспрессирует ANGPTL4. Например, способ включает в себя (a) измерение физиологического признака трансгенного животного, отличного от человека, геном которого содержит нарушение гена, который кодирует ANGPTL4; и (b) сравнение измеренного физиологического признака (a) с признаком животного дикого типа соответствующего пола. Фенотип, возникающий в результате нарушения гена у трансгенного животного, отличного от человека, является физиологическим признаком трансгенного животного, отличного от человека, который отличается от физиологического признака животного дикого типа. Тестируемое средство вводят трансгенному животному, отличному от человека (a); и определяют, модулирует ли тестируемое соединение идентифицированный фенотип, связанный с нарушением гена. Тестируемое средство, которое модулирует фенотип, является средством, которое модулирует такой фенотип.
В некоторых вариантах фенотип, связанный с нарушением гена ANGPTL4, или фенотип, проявляемый трансгенным животным, отличным от человека, по сравнению с однопометными животными дикого типа соответствующего пола, представляет собой, по меньшей мере, один из следующих фенотипов, но не ограничен указанным; например, сердечно-сосудистое, эндотелиальное или ангиогенное расстройство; иммунологическое расстройство; расстройство гомеостаза липидов или расстройство, связанное с аномальным метаболизмом.
Также предлагаются способы идентификации средства, которое модулирует физиологический признак, связанный с нарушением гена, который кодирует ANGPTL4.
В некоторых вариантах способ включает в себя (a) измерение физиологического признака, проявляемого трансгенным животным, отличным от человека, геном которого содержит нарушение гена, который кодирует ANGPTL4; и (b) сравнение измеренного физиологического признака согласно (a) с физиологическим признаком животного дикого типа соответствующего пола. Физиологический признак, проявляемый трансгенным животным, отличным от человека, который отличается от физиологического признака, проявляемого животным дикого типа, идентифицируют как физиологический признак, связанный с нарушением гена. Тестируемое средство вводят трансгенному животному, отличному от человека (a), и определяют, модулируется ли физиологический признак, связанный с нарушением гена. Тестируемое средство, которое модулирует физиологический признак, является средством, которое модулирует такой признак.
В некоторых вариантах трансгенное животное, отличное от человека, проявляет, по меньшей мере, один из следующих физиологических признаков по сравнению с однопометными животными дикого типа соответствующего пола, например, модулирование средних уровней холестерина в сыворотке, модулирование средних уровней триглицеридов в сыворотке, модулирование в тесте толерантности к глюкозе, модулирование гомеостаза глюкозы, пониженный средний уровень глюкозы в сыворотке; повышенный средний уровень инсулина в сыворотке; пониженный средний уровень инсулина в сыворотке; повышенный средний уровень IgM в сыворотке и повышенное абсолютное количество нейтрофилов, повышенное среднее процентное содержание жира в организме; пониженная масса тела и длина, пониженная общая масса ткани и безжировая компонента массы тела, пониженная общая жировая масса, задержка роста с пониженной массой тела и длиной и/или пониженное среднее процентное содержание общего жира в организме, общей массы ткани. В одном варианте модулирование средних уровней холестерина в сыворотке представляет собой пониженный средний уровень холестерина в сыворотке. В одном варианте модулирование среднего уровня триглицеридов в сыворотке представляет собой снижение среднего уровня триглицеридов в сыворотке. В другом варианте модулирование в тесте толерантности к глюкозе представляет собой повышенную толерантность к глюкозе.
Предлагаются способы идентификации средства, которое улучшает состояние при сердечно-сосудистом, эндотелиальном или ангиогенном расстройстве; иммунологическом расстройстве; онкологическом заболевании; расстройстве метаболизма липидов; или аномальном метаболическом расстройстве, связанном с нарушением гена, который кодирует ANGPTL4. Например, способ включает в себя (a) введение тестируемого средства трансгенному животному, отличному от человека, имеющему нарушение в гене ANGPTL4; и (b) определение того, улучшает ли тестируемое средство состояние при сердечно-сосудистом, эндотелиальном или ангиогенном расстройстве; иммунологическом расстройстве; онкологическом заболевании; расстройстве метаболизма липидов; или метаболическом расстройстве, связанном с нарушением гена у трансгенного животного, отличного от человека.
Изобретение относится к способам оценки терапевтического средства, способного влиять на состояние, связанное с нарушением гена, который кодирует ANGPTL4. Например, способ включает в себя (a) измерение физиологического признака трансгенного животного, отличного от человека, геном которого содержит нарушение гена, который кодирует ANGPTL4; (b) сравнение измеренного физиологического признака согласно (a) с физиологическим признаком животного дикого типа соответствующего пола; (c) введение тестируемого средства трансгенному животному, отличному от человека, согласно (a); и (d) оценку влияния тестируемого средства на идентифицированное состояние, связанное с нарушением гена, у трансгенного животного, отличного от человека. Физиологический признак трансгенного животного, отличного от человека, который отличается от физиологического признака животного дикого типа, идентифицируют как состояние, возникающее в результате нарушения гена, у трансгенного животного, отличного от человека. Например, состояние представляет собой сердечно-сосудистое, эндотелиальное или ангиогенное расстройство; иммунологическое расстройство; онкологическое заболевание; расстройство гомеостаза липидов или метаболическое расстройство.
Также предлагаются способы идентификации средства, которое модулирует экспрессию ANGPTL4. Например, способ включает в себя (a) осуществление контакта тестируемого средства с клеткой-хозяином, экспрессирующей ANGPTL4; и (b) определение того, модулирует ли тестируемое средство экспрессию ANGPTL4 клеткой-хозяином.
Средство, идентифицированное любым из указанных выше способов, также включено в изобретение. В одном варианте средство является агонистом. В другом варианте средство является антагонистом ANGPTL4. Средства, которые являются терапевтическими средствами, также включены в изобретение наряду с фармацевтической композицией, содержащей терапевтическое средство.
В различных способах согласно изобретению молекула согласно изобретению, например ANGPTL4, агонист или антагонист ANGPTL4, средство и т.д., могут быть введены субъекту с помощью системы для системной доставки. В одном аспекте система для системной доставки содержит препарат клеток, содержащий клетки млекопитающих (например, клетки CHO), экспрессирующие рекомбинантную форму данного средства. В другом аспекте система для системной доставки может содержать препарат замедленного высвобождения, включающий в себя очищенное средство и полимерный матрикс. В некоторых вариантах молекулу вводят субъекту с использованием фармацевтически приемлемого носителя. Альтернативно молекулу согласно изобретению можно вводить с помощью вектора для целенаправленной доставки генов в ткани (например, в адипоциты, печень и т.д.), содержащего нуклеиновую кислоту, кодирующую молекулу. Можно использовать хорошо разработанные вирусные и невирусные векторы для генной терапии в качестве вектора для целенаправленной доставки генов в ткани в изобретении.
Краткое описание фигур
Фиг.1 иллюстрирует последовательность нуклеиновой кислоты ANGPTL4 человека (SEQ ID NO: 1).
Фиг.2 иллюстрирует аминокислотную последовательность ANGPTL4 человека (SEQ ID NO: 2), полученную на основе кодирующей последовательности SEQ ID NO: 1, показанной на фиг.1.
На фиг.3 панель A иллюстрирует очищенный рекомбинантный ANGPTL4 мыши (23-410) при разделении с помощью электрофореза в SDS-полиакриламидном геле (SDS-ПААГ) (4-20%) в присутствии (10 мМ) или в отсутствие дитиотреитола (DTT). На фиг.3 панель B иллюстрирует hANGPTL4 дикого типа (дорожка 1) и вариант hANGPTL4 (дорожка 2), после разделения в SDS-геле и выявления с помощью Вестерн-блоттинга, где вариант hANGPTL4 имеет замену R162G и R164E.
Фиг.4 схематично иллюстрирует, что ANGPTL4 индуцирует клеточную адгезию гепатоцитов человека.
Фиг.5 схематично иллюстрирует, что ANGPTL4 индуцирует пролиферацию гепатоцитов.
На фиг.6 панели A и B схематично иллюстрируют, что внеклеточный ANGPTL4 индуцирует висцеральную пролиферацию первичных преадипоцитов человека (панель A) и подкожную пролиферацию преадипоцитов (панель B).
Фиг.7 схематично показывает, что ANGPTL4 (23-406) и химерные формы IgG-ANGPTL4 человека связываются с подкожными первичными адипоцитами человека по данным анализа FACS.
На фиг.8 панели A, B и C иллюстрируют, что ANGPTL4 индуцирует клеточную миграцию первичных подкожных преадипоцитов человека. Панели A и B иллюстрируют индукцию ANGPTL4 клеточной миграции первичных преадипоцитов в течение ночи (панель A) и в течение 7 часов (панель B). Панель C схематично иллюстрирует миграцию первичных преадипоцитов под влиянием ANGPTL4 в течение 7 часов, где (1) без добавления сыворотки, (2) с 10% фетальной сыворотки теленка (FCS), (3) с PDGF-BB и (4) с mANGPTL4.
На фиг.9 панели A, B, C, D и E иллюстрируют связывание ANGPTL4 с интегрином αvβ5. Панель A иллюстрирует адгезию клеток 293-1953 (αvβ5) с планшетом, покрытым либо mANGPTL4, либо витронектином в концентрации, указанной внизу в (мкг/мл), где БСА используют в качестве контроля. Панель B иллюстрирует, что анти-αvβ5-антитело и анти-hANGPTL4-антитело удаляют активность ANGPTL4 в клеточной адгезии, где (1) означает БСА, (2) означает вибронектин и (3) означает mANGPTL4. Панель C иллюстрирует связывание белка (mANGPTL4, hANGPTL4-Nконец или hANGPTL4-Cконец) при использовании указанного количества с планшетами, покрытыми αvβ5. Панель D иллюстрирует ингибирование связывания белка (mANGPTL4, hANGPTL4-Nконец или hANGPTL4-Cконец) с планшетами, покрытыми αvβ5, с использованием анти-hANGPTL4, где в качестве контролей использовали контрольное антитело против белка критической для синдрома Дауна области 1 (Dscr), 5G7 или среду. Панель E иллюстрирует связывание ANGPTL4 и αvβ5, где (1) означает планшет, покрытый hANGPTL4-Cконец, (2) означает планшет, покрытый hANGPTL4-Cконец и инкубированный с анти-hANGPTL4, (3) означает планшет, покрытый hANGPTL4-Cконец и инкубированный с анти-Dscr, (4) означает планшет, покрытый витронектином, и (5) означает планшет, покрытый БСА перед добавлением αvβ5.
Фиг.10 иллюстрирует уровни триглицеридов у мышей при инъекции в хвостовую вену ANGPTL4 и вариантов ANGPTL4, где (1) означает Ad-GFP, (2) означает Ad-Gd, (3) означает ANGPTL4(1-406), (4) означает ANGPTL4(1-183), (5) означает ANGPTL4 (184-406), (6) означает вариант ANGPTL4 R1162G и R164E, (7) означает ANGPTL4 (1-408) и (8) означает контроль.
Подробное описание
Определения
Перед подробным описанием изобретения следует понять, что данное изобретение не ограничено конкретными композициями или биологическими системами, которые, конечно, могут варьировать. Также следует понимать, что терминология, используемая в данном описании, предназначена только для целей описания конкретных вариантов и не предназначена для ограничения. В используемом в данном описании и прилагаемой формуле изобретения смысле формы единственного числа включают множественные объекты, если содержание ясно не диктует иное. Таким образом, например, ссылка на «молекулу» необязательно включает комбинацию двух или более таких молекул, и т.п. Если не оговорено особо, подразумевается, что все научные и технические термины имеют такое же значение, которое обычно используют в области, к которой они относятся. В целях изобретения ниже определены следующие термины.
Термин «ANGPTL4» или «Angptl4» относится к ангиопоэтинподобному полипептиду или белку 4 вместе с его встречающимися в природе аллельными, секретируемыми и процессированными формами. Например, ANGPTL4 человека является белком из 406 аминокислот, тогда как мышиный ANGPTL4 является белком, состоящим из 410 аминокислот.Термин «ANGPTL4» также используют для указания фрагментов (например, подпоследовательностей, укороченных форм и т.д.) полипептида, содержащих, например, N-концевой фрагмент, биспиральный домен, C-концевой фрагмент, фибриногенподобный домен, аминокислоты 1-183, 23-183, от 1 до примерно 162, от 23 до примерно 162, 23-406, 184-406, примерно 162-406 или 23-184 ангиопоэтинподобного белка 4 человека и аминокислоты 1-183, 23-183, от 1 до примерно 165, от 23 до примерно 165, 23-410 или 184-410 мышиного ангиопоэтинподобного белка 4. Другие фрагменты включают без ограничения, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-406, 60-406, 80-406, 100-406, 120-406, 140-406, и 160-406 hANGPTL4 и, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-410, 60-410, 80-410, 100-410, 120-410, 140-410 и 160-410 mANGPTL4. Ссылка на любые такие формы ANGPTL4 также может быть указана в заявке, например, в виде «ANGPTL4 (23-406)», «ANGPTL4 (184-406)», «ANGPTL4 (23-183)», «mANGPTL4 (23-410)», «mANGPTL4 (184-410)» и т.д., где m указывает мышиную последовательность. Положения аминокислот для фрагмента нативного ANGPTL4 пронумерованы, как указано в нативной последовательности ANGPTL4. Например, положение аминокислоты 22(Ser) во фрагменте ANGPTL4 также является положением 22(Ser) в нативном ANGPTL4 человека, например, см. фиг.2. Как правило, фрагмент нативного ANGPTL4 обладает биологической активностью.
Полипептид с «нативной последовательностью» включает полипептид, имеющий такую же аминокислотную последовательность, как и полипептид, полученный из природного источника. Таким образом, полипептид с нативной последовательностью может иметь аминокислотную последовательность встречающегося в природе полипептида любого млекопитающего. Такой полипептид с нативной последовательностью можно выделить из природного источника и можно получить способами рекомбинации или синтеза. Термин полипептид с «нативной последовательностью» специально охватывает встречающиеся в природе укороченные или секретируемые формы полипептида (например, последовательность внеклеточного домена), встречающиеся в природе вариантные формы (например, альтернативно сплайсируемые формы) и встречающиеся в природе аллельные варианты полипептида.
«Вариант» полипептида означает биологически активный полипептид, имеющий, по меньшей мере, примерно 80% идентичность аминокислотной последовательности с соответствующим полипептидом с нативной последовательностью или его фрагментом. Такие варианты включают, например, полипептиды, в которых один или несколько аминокислотных остатков добавлены или делетированы на N- и/или C-конце полипептида. Обычно вариант будет обладать, по меньшей мере, примерно 80% идентичностью аминокислотной последовательности или, по меньшей мере, примерно 90% идентичностью аминокислотной последовательности, или, по меньшей мере, примерно 95% или большей идентичностью аминокислотной последовательности с полипептидом с нативной последовательностью или его фрагментом.
Термин «вариант ANGPTL4» в используемом в данном описании смысле относится к варианту, который описан выше, и/или ANGPTL4, который содержит одну или несколько аминокислотных мутаций в нативной последовательности ANGPTL4. Необязательно одна или несколько аминокислотных мутаций включают аминокислотную замену(ы). ANGPTL4 и его варианты для применения в изобретении могут быть получены множеством способов, хорошо известных в данной области. Варианты аминокислотной последовательности ANGPTL4 могут быть получены в результате мутаций в ДНК ANGPTL4. Такие варианты включают, например, делеции, инсерции или замены остатков в аминокислотной последовательности ANGPTL4, например в аминокислотной последовательности человека, кодируемой нуклеиновой кислотой, депонированной в ATCC под номером депозита 209284, или показанной на фиг.2. Может быть осуществлена любая комбинация делеции, инсерции и замены, чтобы добиться конечной конструкции, обладающей требуемой активностью. Мутации, которые будут получены в ДНК, кодирующей вариант, не должны выводить последовательность из рамки считывания и предпочтительно не будут создавать комплементарные области, которые могут создавать вторичную структуру мРНК. EP 75444A.
Варианты ANGPTL4 необязательно получают сайт-специфичным мутагенезом нуклеотидов в ДНК, кодирующей нативный ANGPTL4, или способами фагового дисплея, получая таким образом ДНК, кодирующую вариант, и после этого экспрессируя ДНК в культуре рекомбинантных клеток.
В то время как сайт введения изменения аминокислотной последовательности определяют предварительно, мутация как таковая необязательно должна быть определена предварительно. Например, чтобы оптимизировать эффективность мутации в данном сайте, может быть проведен случайный мутагенез в кодоне-мишени или области-мишени и экспрессированные варианты ANGPTL4 подвергнуты скринингу в отношении оптимальной комбинации требуемой активности. Способы получения мутаций в виде замен в предварительно определяемых сайтах ДНК, имеющей известную последовательность, хорошо известны, например, такие как сайт-специфичный мутагенез. Получение вариантов ANGPTL4, описанных в данной публикации, можно осуществить с помощью способов фагового дисплея, таких как способы, описанные в публикации PCT WO 00/63380.
После отбора такого клона область мутантного белка может быть извлечена и помещена в соответствующий вектор для продукции белка, как правило, экспрессирующий вектор такого типа, который может быть использован для трансформации подходящего хозяина.
Делеции аминокислотной последовательности обычно находятся в пределах примерно от 1 до 30 остатков, необязательно от 1 до 10 остатков, необязательно от 1 до 5 или менее и обычно являются непрерывными.
Инсерции аминокислотной последовательности включают слияния на амино- и/или карбоксильном конце от одного остатка до полипептидов по существу неограниченной длины, а также инсерции внутри последовательности одного или множества аминокислотных остатков. Инсерции внутри последовательности (т.е. инсерции в нативной последовательности ANGPTL4) могут быть в пределах, как правило, примерно от 1 до 10 остатков, необязательно от 1 до 5, или необязательно от 1 до 3. Пример концевой инсерции включает слияние сигнальной последовательности, либо гетерологичной, либо гомологичной по отношению к клетке-хозяину, с N-концом, чтобы облегчить секрецию из рекомбинантных хозяев.
Дополнительными вариантами ANGPTL4 являются такие варианты, в которых, по меньшей мере, один аминокислотный остаток в нативном ANGPTL4 был удален, а другой остаток встроен на его место. В одном варианте осуществления изобретения вариант ANGPTL4 содержит замену в положении 162 и/или 164 ANGPTL4 или замену в положении 169 mANGPTL4. Такие замены могут быть осуществлены в соответствии с заменами, показанными в таблице. Варианты ANGPTL4 также могут содержать неприродные аминокислоты, которые описаны в данной публикации.
Аминокислоты могут быть сгруппированы в соответствии со сходствами в свойствах их боковых цепей (в A. L. Lehninger, Biochemistry, second ed., pp.73-75, Worth Publishers, New York (1975)):
(1) неполярные: Ala (A), Val (V), Leu (L), Ile (I), Pro (P), Phe (F), Trp (W), Met (M)
(2) незаряженные полярные: Gly (G), Ser (S), Thr (T), Cys (C), Tyr (Y), Asn (N), Gln (Q)
(3) кислотные: Asp (D), Glu (E)
(4) основные: Lys (K), Arg (R), His (H)
Альтернативно встречающиеся в природе остатки могут быть разделены на группы на основе общих свойств боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.

«Встречающиеся в природе аминокислотные остатки» (т.е. аминокислотные остатки, кодируемые генетическим кодом) могут быть выбраны из группы, состоящей из аланина (Ala); аргинина (Arg); аспарагина (Asn); аспарагиновой кислоты (Asp); цистеина (Cys); глутамина (Gln); глутаминовой кислоты (Glu); глицина (Gly); гистидина (His); изолейцина (Ile); лейцина (Leu); лизина (Lys); метионина (Met); фенилаланина (Phe); пролина (Pro); серина (Ser); треонина (Thr); триптофана (Trp); тирозина (Tyr) и валина (Val). «Не встречающийся в природе аминокислотный остаток» относится к другому остатку, отличному от встречающихся в природе аминокислотных остатков, перечисленных выше, который способен ковалентно связывать соседний аминокислотный остаток(ки) в полипептидной цепи. Примеры не встречающихся в природе аминокислотных остатков включают, например, норлейцин, орнитин, норвалин, гомосерин и другие аналоги аминокислотных остатков, такие как остатки, описанные в Ellman et al. Meth. Enzym. 202:301-336 (1991), и публикации заявок на выдачу патентов США 2003/0108885 и 2003/0082575. Коротко, указанные способы включают активацию супрессорной тРНК не встречающимся в природе аминокислотным остатком с последующей транскрипцией и трансляцией РНК in vitro или in vivo. См., например, публикации заявок на выдачу патентов США 2003/0108885 и 2003/0082575; Noren et al. Science 244: 182 (1989); и Ellman et al., выше.
«Идентичность аминокислотных последовательностей в процентах (%)» в данном описании определяют как процентное содержание аминокислотных остатков в исследуемой последовательности, которые идентичны аминокислотным остаткам в выбранной последовательности после выравнивания последовательностей и введения при необходимости пробелов, чтобы достичь максимального процента идентичности последовательностей, и без учета каких-либо консервативных замен в качестве части идентичности последовательностей. Выравнивание в целях определения идентичности аминокислотных последовательностей в процентах можно осуществить различными способами, которые возможны в данной области, например, используя общедоступную компьютерную программу, такую как компьютерная программа BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для измерения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания на протяжении полной длины сравниваемых последовательностей. Однако в целях данного изобретения значения идентичности аминокислотных последовательностей в % получают, как описано ниже, с использованием компьютерной программы сравнения последовательностей ALIGN-2. Компьютерная программа сравнения последовательностей ALIGN-2 разработана Genentech, Inc. и подана с документацией для пользователей в Бюро регистрации авторских прав США, Washington D.C., 20559, где зарегистрирована с регистрационным № авторского права США TXU510087, и общедоступна от Genentech, Inc., South San Francisco, California. Программа ALIGN-2 должна быть компилирована для применения с операционной системой UNIX, например, цифровой UNIX V4.0D. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и не изменяются.
В целях данного изобретения идентичность аминокислотной последовательности в % для данной аминокислотной последовательности A по сравнению или по отношению к данной аминокислотной последовательности B (что альтернативно может быть выражено как данная аминокислотная последовательность A, которая имеет или содержит определенную идентичность аминокислотных последовательностей в % по сравнению или по отношению к данной аминокислотной последовательности B) рассчитывают следующим образом:
100 × дробь X/Y,
где X означает количество аминокислотных остатков, оцененных как идентичные совпадения программой выравнивания последовательностей ALIGN-2, при выравнивании указанной программой A и B, и где Y означает общее количество аминокислотных остатков в B. Будет понятно, что в том случае, когда длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, идентичность аминокислотной последовательности A по сравнению с B в % не будет равна идентичности аминокислотной последовательности B по сравнению с A в %.
«Изолированным» полипептидом является полипептид, который был идентифицирован и отделен и/или извлечен из компонента его природной среды. Загрязняющие компоненты его природной среды представляют собой вещества, которые могут мешать диагностическим или терапевтическим применениям полипептида и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В некоторых вариантах полипептид будет очищен (1) более чем на 95% мас. полипептида, который определяют способом Лоури, или более чем на 99% мас., (2) в такой степени, которая достаточна для получения, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся цилиндром или (3) до гомогенности с помощью SDS-ПААГ в восстанавливающих или невосстанавливающих условиях с использованием окраски Кумасси синим или серебром. Изолированный полипептид включает полипептид, находящийся in situ в рекомбинантных клетках, так как, по меньшей мере, один компонент из природной среды полипептида не будет присутствовать. Однако обычно изолированный полипептид будет получен, по меньшей мере, с помощью одной стадии очистки.
Термин «модулятор ANGPTL4» относится к молекуле, которая может активировать ANGPTL4 или его экспрессию, например, к агонисту, или которая может ингибировать активность ANGPTL4 или его экспрессию, например, к антагонисту (или ингибитору). Агонисты ANGPTL4 включают антитела и активные фрагменты. Антагонист ANGPTL4 относится к молекуле, способной нейтрализовать, блокировать, ингибировать, отменять, уменьшать или служить препятствием для активностей ANGPTL4, например, пролиферации или роста клеток, миграции, адгезии или модулирования метаболизма, например, липидов, или его экспрессии, включая его связывание с рецептором ANGPTL4, например, αVβ5. Антагонисты ANGPTL4 включают, например, анти-ANGPTL4-антитела и их антигенсвязывающие фрагменты, молекулы рецепторов и производные, которые специфично связываются с ANGPTL4, таким образом предотвращая его связывание с одним или несколькими рецепторами, антитела против рецептора ANGPTL4 и антагонисты рецептора ANGPTL4, такие как низкомолекулярные ингибиторы рецептора. Другие антагонисты ANGPTL4 также включают варианты антагонистов ANGPTL4, антисмысловые молекулы (например, ANGPTL4-миРНК), РНК-аптамеры и рибозимы против ANGPTL4 или его рецептора. В некоторых вариантах антагонистическими ANGPTL4-антителами являются антитела, которые ингибируют или уменьшают активность ANGPTL4 посредством связывания со специфичной подпоследовательностью или областью ANGPTL4, например, N-концевым фрагментом, биспиральным доменом, C-концевым фрагментом, фибриногенподобным доменом, аминокислотами 1-183, 23-183, от 1 до примерно 162, от 23 до примерно 162, 23-406, 184-406, примерно 162-406 или 23-184 ангиопоэтинподобного белка 4 человека и аминокислотами 1-183, 23-183, от 1 до примерно 165, от 23 до примерно 165, 23-410 или 184-410 ангиопоэтинподобного белка 4 мыши. Другие подпоследовательности также включают, но не ограничены указанным, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-406, 60-406, 80-406, 100-406, 120-406, 140-406 и 160-406 hANGPTL4 и, например, 40-183, 60-183, 80-183, 100-183, 120-183, 140-183, 40-410, 60-410, 80-410, 100-410, 120-410, 140-410 и 160-410 mANGPTL4.
Модуляторами ANGPTL4 являются молекулы, которые модулируют активность ANGPTL4, например, агонисты и антагонисты. Термин «агонист» используют по отношению к пептидным и непептидным аналогам ANGPTL4 и к антителам, специфично связывающим такие молекулы ANGPTL4, при условии, что они обладают способностью передавать сигнал через нативный рецептор ANGPTL4 (например, интегрин αVβ5). Термин «агонист» определяют в контексте биологической роли рецептора ANGPTL4 (например, αVβ5). В некоторых вариантах агонисты обладают биологическими активностями нативного ANGPTL4, которые определены выше, такими как стимуляция пролиферации, миграции и/или адгезии клеток и/или модулирование гомеостаза липидов.
Термин «антагонист» используют по отношению к молекулам, которые обладают способностью ингибировать биологическую активность ANGPTL4, независимо от того, обладают ли они способностью связывать ANGPTL4 или его рецептор, например, αVβ5. Например, антагонисты, которые обладают способностью связывать ANGPTL4 или его рецептор, включают анти-ANGPTL4-антитела и анти-αVβ5-антитела. В изобретение включены антагонисты, которые ингибируют экспрессию ANGPTL4, например, ANGPTL4-миРНК. Антагонист ANGPTL4 можно оценить, например, по ингибированию активности ANGPTL4, например, активности ANGPTL4 в адгезии, миграции, пролиферации и/или модулировании гомеостаза липидов. Что касается активности рецептора интегрина αVβ5, то модулятор рецептора интегрина αVβ5 можно определить способами, известными в данной области. Например, можно использовать способ, описанный J. W. Smith et al. в J. Biol. Chem. 265: 12267-12271 (1990).
Термин «анти-ANGPTL4-антитело» означает антитело, которое связывается с ANGPTL4 с достаточной аффинностью и специфичностью. В некоторых вариантах осуществления изобретения анти-ANGPTL4-антитело согласно изобретению можно применять в качестве терапевтического средства для целенаправленного воздействия и вмешательства в заболевания или состояния, в которые вовлечена активность ANGPTL4. В общем, анти-ANGPTL4-антитело, как правило, не будет связываться с другими гомологами ANGPTL4, например, ANGPTL3.
Термин «антитело» используют в самом широком смысле, и термин охватывает моноклональные антитела (включая полноразмерные или интактные моноклональные антитела), поликлональные антитела, поливалентные антитела, полиспецифичные антитела (например, биспецифичные антитела) и фрагменты антител (см. ниже) при условии, что они проявляют требуемую биологическую активность.
Если не оговорено особо, то выражение «поливалентное антитело» используют на протяжении данного описания, чтобы указать антитело, содержащее три или более антигенсвязывающих участков. Поливалентное антитело обычно конструируют так, чтобы оно имело три или более антигенсвязывающих участков, и, как правило, оно не является нативной последовательностью IgM- или IgA-антитела.
«Фрагменты антитела» содержат только часть интактного антитела, обычно включая антигенсвязывающий участок интактного антитела и поэтому сохраняя способность связывать антиген. Примеры фрагментов антитела, охватываемых приведенным определением, включают: (i) Fab-фрагмент, имеющий домены VL, CL, VH и CH1; (ii) Fab'-фрагмент, который представляет собой Fab-фрагмент, имеющий один или несколько остатков цистеина на C-конце домена CH1; (iii) Fd-фрагмент, имеющий домены VH и CH1; (iv) Fd'-фрагмент, имеющий домены VH и CH1 и один или несколько остатков цистеина на C-конце домена CH1; (v) Fv-фрагмент, имеющий домены VL и VH одного плеча антитела; (vi) dAb-фрагмент (Ward et al., Nature 341, 544-546 (1989)), который состоит из домена VH; (vii) изолированные CDR-области; (viii) F(ab')2-фрагменты, бивалентные фрагменты, содержащие два фрагмента Fab', связанные дисульфидным мостиком в шарнирной области; (ix) молекулы одноцепочечных антител (например, одноцепочечный Fv; scFv) (Bird et al., Science 242: 423-426 (1988) и Huston et al., PNAS (USA) 85: 5879-5883 (1988)); (x) «диантитела» с двумя антигенсвязывающими участками, содержащие вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (см., например, EP 404097, WO 93/11161 и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)); (xi) «линейные антитела», содержащие пару тандемных участков Fd (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют пару антигенсвязывающих областей (Zapata et al. Белок Eng. 8(10): 1057 1062 (1995) и патент США №5641870).
Термин «моноклональное антитело» в используемом в данном описании смысле относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в минорных количествах. Моноклональные антитела являются высоко специфичными, направленными против одного антигена. Кроме того, в отличие от препаратов поликлональных антител, которые обычно содержат разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Определение «моноклональное» не следует рассматривать как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела, применяемые согласно изобретению, могут быть получены способом на основе гибридом, впервые описанным Kohler et al., Nature 256: 495 (1975), или могут быть получены способами на основе рекомбинантной ДНК (см., например, патент США №4816567). «Моноклональные антитела» также могут быть выделены из фаговых библиотек антител с использованием способов, описанных, например, в Clackson et al., Nature 352: 624-628 (1991) или Marks et al., J. Mol. Biol. 222: 581-597 (1991).
Моноклональные антитела в данном описании специально включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных от конкретного вида, или относящимся к конкретному классу или подклассу антител, тогда как остальная часть цепи (цепей) идентична или гомологична соответствующим последовательностям в антителах, полученных от другого вида или относящихся к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют требуемую биологическую активность (патент США №4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)).
«Гуманизированные» формы антител животных, отличных от человека (например, мышей), представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина животного, отличного от человека. Главным образом гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области вида, отличного от человека (донорное антитело), такого как мышь, крыса, кролик или примат, отличный от человека, обладающей требуемой специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими остатками животного, отличного от человека. Кроме того, гуманизированные антитела могут содержать остатки, которые не встречаются ни в реципиентном антителе, ни в донорном антителе. Такие модификации осуществляют для того, чтобы дополнительно улучшить эффективность антитела. В общем, гуманизированное антитело будет содержать в основном целиком, по меньшей мере один, и обычно два вариабельных домена, в которых все или в основном все гипервариабельные петли соответствуют петлям иммуноглобулина животного, отличного от человека, и все или в основном все FR являются FR из последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также будет содержать, по меньшей мере, часть константной области иммуноглобулина (Fc), обычно константной области иммуноглобулина человека. Более подробное описание см. в Jones et al., Nature 321: 522-525 (1986); Riechmann et al., Nature 332: 323-329 (1988) и Presta, Curr. Op.Struct. Biol. 2: 593-596 (1992).
«Человеческим антителом» является антитело, которое имеет аминокислотную последовательность, соответствующую аминокислотной последовательности антитела, образуемого в организме человека, и/или которое получено с использованием любого способа получения человеческих антител, который описан в данной публикации. Такое определение человеческого антитела специально включает гуманизированное антитело, содержащее антигенсвязывающие остатки животного, отличного от человека. Человеческие антитела могут быть получены с использованием различных способов, известных в данной области. В одном варианте человеческое антитело выбрано из фаговой библиотеки, при этом такая фаговая библиотека экспрессирует человеческие антитела (Vaughan et al. Nature Biotechnology 14: 309-314 (1996): Sheets et al. PNAS (USA) 95: 6157-6162 (1998)); Hoogenboom and Winter, J. Mol. Biol., 227: 381 (1991); Marks et al., J. Mol. Biol., 222: 581 (1991)). Человеческие антитела также могут быть получены введением локусов иммуноглобулинов человека в трансгенных животных, например мышей, у которых эндогенные гены иммуноглобулинов были частично или полностью инактивированы. При стимуляции наблюдают продукцию антител человека, которая очень похожа на продукцию антител, наблюдаемую у людей во всех отношениях, включая реаранжировку генов, сборку и репертуар антител. Такой способ описан, например, в патентах США №5545807, 5545806, 5569825, 5625126, 5633425, 5661016 и в следующих научных публикациях: Marks et al., Bio/Technology 10: 779-783 (1992); Lonberg et al., Nature 368: 856-859 (1994); Morrison, Nature 368: 812-13 (1994); Fishwild et al., Nature Biotechnology 14: 845-51 (1996); Neuberger, Nature Biotechnology 14: 826 (1996); Lonberg and Huszar, Intern. Rev. Immunol. 13: 65-93 (1995). Альтернативно человеческое антитело может быть получено посредством иммортализации B-лимфоцитов человека, продуцирующих антитело, направленное против антигена-мишени (такие B-лимфоциты могут быть извлечены из организма человека или могут быть иммунизированы in vitro). См., например, Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985); Boerner et al., J. Immunol., 147 (1): 86-95 (1991) и патент США №5750373.
Термин «вариабельный» относится к тому факту, что некоторые части вариабельных доменов в значительной степени отличаются по последовательности среди антител и используются в связывании и обеспечении специфичности каждого конкретного антитела по отношению к его конкретному антигену. Однако вариабельность неравномерно распределена на протяжении вариабельных доменов антител. Она сконцентрирована в трех участках, называемых гипервариабельными областями, в вариабельных доменах как легкой цепи, так и тяжелой цепи. Более высококонсервативные части вариабельных доменов называют каркасными областями (FR). Каждый из вариабельных доменов нативных тяжелой и легкой цепей содержит четыре FR, в основном принимающих конфигурацию бета-складок, соединенных тремя гипервариабельными областями, которые образуют петли, связывающие и в некоторых случаях образующие часть бета-складчатой структуры. Гипервариабельные области в каждой цепи удерживаются вместе в непосредственной близости посредством FR и с гипервариабельными областями из другой цепи вносят вклад в образование антигенсвязывающего участка антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Константные домены непосредственно не вовлечены в связывание антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в зависимой от антител опосредованной клетками цитотоксичности (ADCC).
Термин «гипервариабельная область» при использовании в данном описании относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельная область обычно содержит аминокислотные остатки из «области, определяющей комплементарность» или «CDR» (например, остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)) и/или остатки из «гипервариабельной петли» (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (H1), 53-55 (H2) и 96-101 (H3) в вариабельном домене тяжелой цепи; Chothia and Lesk J. Mol. Biol. 196: 901-917 (1987)). Остатки «каркасной области» или «FR» представляют собой другие остатки вариабельного домена, отличные от остатков гипервариабельной области, которые определены в данном описании.
В зависимости от аминокислотной последовательности константного домена своих тяжелых цепей интактные антитела могут быть отнесены к разным «классам». Существует пять основных классов интактных антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно разделены на «подклассы» (изотипы), например, IgG1 (включая не-A- и A-аллотипы), IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам антител, названы α, δ, ε, γ и μ, соответственно. Субъединичные структуры и трехмерные конфигурации разных классов иммуноглобулинов хорошо известны.
Легкие цепи антител из любого вида позвоночных могут быть отнесены к одному из двух явно отличающихся типов, названных каппа (κ) и лямбда (λ), на основании аминокислотных последовательностей их константных доменов.
Термин «Fc-область» используют для определения C-концевой области тяжелой цепи иммуноглобулина, которая может быть создана при расщеплении интактного антитела папаином. Fc-область может представлять собой Fc-область с нативной последовательностью или вариант Fc-области. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи IgG человека обычно определяют в промежутке от аминокислотного остатка примерно в положении Cys226 или примерно от положения Pro230 до карбоксильного конца Fc-области. Fc-область иммуноглобулина обычно содержит два константных домена, домен CH2 и домен CH3 и необязательно содержит домен CH4. Под «цепью Fc-области» в данном описании подразумевают одну из двух полипептидных цепей Fc-области.
«Домен CH2» Fc-области IgG человека (также называемый доменом «Cg2») обычно простирается от аминокислотного остатка примерно в положении 231 до аминокислотного остатка примерно в положении 340. Домен CH2 является уникальным в том, что он не спарен непосредственно с другим доменом. Вернее, две N-связанные разветвленные углеводные цепи размещены между двумя доменами CH2 интактной нативной молекулы IgG. Было выдвинуто предположение, что углевод может обеспечивать замену спариванию домен-домен и помогать стабилизировать домен CH2. Burton, Molec. Immunol. 22: 161-206 (1985). Домен CH2 согласно настоящему изобретению может представлять собой домен CH2 с нативной последовательностью или вариант домена CH2.
«Домен CH3» содержит участок из остатков, расположенных на C-конце по отношению к домену CH2 в Fc-области (т.е. от аминокислотного остатка примерно в положении 341 до аминокислотного остатка примерно в положении 447 IgG). Область CH3 согласно настоящему изобретению может представлять собой домен CH3 с нативной последовательностью или вариант домена CH3 (например, домен CH3 с введенным «выступом» в его одной цепи и соответствующей введенной «впадиной» в его другой цепи; см. патент США №5821333, специально включенный в данное описание в виде ссылки). Такие варианты домена CH3 могут быть использованы для получения полиспецифичных (например, биспецифичных) антител, которые рассмотрены в данном описании.
«Шарнирную область», как правило, определяют как участок примерно от Glu216 или примерно от Cys226 до примерно Pro230 IgG1 человека (Burton, Molec. Immunol. 22: 161-206 (1985)). Шарнирные области других изотипов IgG могут быть выравнены с последовательностью IgG1 при размещении первого и последнего остатков цистеина, образующих S-S-связи между тяжелыми цепями, в одних и тех же положениях. Шарнирная область согласно настоящему изобретению может представлять собой шарнирную область с нативной последовательностью или вариант шарнирной области. Две полипептидные цепи варианта шарнирной области обычно сохраняют, по меньшей мере, один остаток цистеина на полипептидную цепь, так что две полипептидные цепи варианта шарнирной области могут образовывать дисульфидную связь между двумя цепями. Предпочтительной шарнирной областью в настоящем изобретении является шарнирная область человека с нативной последовательностью, например, шарнирная область IgG1 человека с нативной последовательностью.
«Функциональная Fc-область» обладает, по меньшей мере, одной «эффекторной функцией» Fc-области с нативной последовательностью. Примеры «эффекторных функций» включают связывание C1q; комплементзависимую цитотоксичность (CDC); связывание рецептора Fc; зависимую от антител опосредованную клетками цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора, BCR) и т.д. Такие эффекторные функции обычно требуют комбинирования Fc-области со связывающим доменом (например, вариабельным доменом антитела) и могут быть оценены с использованием различных анализов, известных в данной области для оценки таких эффекторных функций антитела.
«Fc-область с нативной последовательностью» содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, встречающейся в природе.
«Вариант Fc-области» содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности Fc-области с нативной последовательностью вследствие, по меньшей мере одной, аминокислотной модификации. Предпочтительно вариант Fc-области имеет, по меньшей мере одну, аминокислотную замену по сравнению с Fc-областью с нативной последовательностью или с Fc-областью исходного полипептида, например, от примерно одной до примерно десяти аминокислотных замен и предпочтительно от примерно одной до примерно пяти аминокислотных замен в Fc-области с нативной последовательностью или в Fc-области исходного полипептида. Вариант Fc-области согласно настоящему изобретению обычно будет, например, по меньшей мере, примерно на 80% идентичен по последовательности Fc-области с нативной последовательностью и/или Fc-области исходного полипептида, или, по меньшей мере, примерно на 90% идентичен им по последовательности, или, по меньшей мере, примерно на 95% или более идентичен им по последовательности.
«Зависимая от антител опосредованная клетками цитотоксичность» и «ADCC» относятся к опосредованному клетками ответу, при котором неспецифичные цитотоксические клетки, которые экспрессируют рецепторы Fc (FcR) (например, природные клетки-киллеры (NK), нейтрофилы и макрофаги), узнают связанное антитело на клетке-мишени и затем вызывают лизис клетки-мишени. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках суммирована в таблице 3 на странице 464 в Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-92 (1991). Чтобы оценить активность в ADCC представляющей интерес молекулы, можно осуществить анализы ADCC in vitro, такие как анализы, описанные в патентах США №5500362 или 5821337. Применимые эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (PBMC) и природные клетки-киллеры (NK). Альтернативно или дополнительно ADCC-активность представляющей интерес молекулы можно оценить in vivo, например, в животной модели, такой как модель, описанная в Clynes et al. PNAS (USA) 95: 652-656 (1998).
«Эффекторными клетками человека» являются лейкоциты, которые экспрессируют один или несколько FcR и осуществляют эффекторные функции. Обычно клетки экспрессируют, по меньшей мере, FcγRIII и осуществляют ADCC-эффекторную функцию. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (PBMC), природные клетки-киллеры (NK), моноциты, цитотоксические T-клетки и нейтрофилы; при этом обычно предпочтительны PBMC и NK-клетки. Эффекторные клетки могут быть выделены из их природного источника, например, из крови или PBMC, как описано в данной публикации.
Термины «Fc-рецептор» и «FcR» используют для описания рецептора, который связывается с Fc-областью антитела. Предпочтительным FcR является FcR человека с нативной последовательностью. Кроме того, предпочтительным FcR является FcR, который связывает IgG-антитело (гамма-рецептор), и к предпочтительным рецепторам относятся рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и альтернативно сплайсируемые формы указанных рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые имеют сходные аминокислотные последовательности, которые отличаются главным образом своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит в своем цитоплазматическом домене основанный на тирозине мотив активации иммунорецептора (ITAM). Ингибирующий рецептор FcγRIIB содержит в своем цитоплазматическом домене основанный на тирозине мотив ингибирования иммунорецептора (ITIM) (обзор в Daлron, Annu. Rev. Immunol. 15: 203-234 (1997)). Обзор, посвященный FcR, представлен в Ravetch and Kinet, Annu. Rev. Immunol 9: 457-92 (1991); Capel et al., Immunomethods 4: 25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126: 330-41 (1995). Другие FcR, включая FcR, идентифицируемые в природе, включены в данном описании в термин «FcR». Термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG в плод (Guyer et al., J. Immunol. 117: 587 (1976) и Kim et al., J. Immunol. 24: 249 (1994)).
«Комплементзависимая цитотоксичность» и «CDC» относятся к лизису мишени в присутствии комплемента. Путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом) в комплексе со своим антигеном. Чтобы оценить активацию комплемента, можно осуществить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996).
Термин «иммуноадгезин» относится к молекулам, подобным антителам, которые объединяют специфичность связывания гетерологичного белка («адгезина») с эффекторными функциями константных доменов иммуноглобулинов.
Структурно иммуноадгезины содержат слияние аминокислотной последовательности с требуемой специфичностью связывания, которая отлична от участка антигенного узнавания и связывания антитела (т.е. является «гетерологичной») и последовательности константного домена иммуноглобулина. Адгезиновая часть молекулы иммуноадгезина обычно представляет собой непрерывную аминокислотную последовательность, содержащую, по меньшей мере, связывающий сайт рецептора или лиганд. Последовательность константного домена иммуноглобулина в иммуноадгезине может быть получена из любого иммуноглобулина, такого как подтипы IgG1, IgG2, IgG3 или IgG4, IgA (включая IgA1 и IgA2), IgE, IgD или IgM.
«Активный» или «активность» в целях настоящего изобретения относится к форме(ам) ANGPTL4, которые сохраняют биологическую и/или иммунологическую активность нативного или встречающегося в природе ANGPTL4, при этом «биологическая» активность относится к биологической функции (либо ингибирующей, либо стимулирующей), вызванной нативным или встречающимся в природе ANGPTL4, отличной от способности индуцировать продукцию антитела против антигенного эпитопа, которой обладает нативный или встречающийся в природе ANGPTL4, и «иммунологическая» активность относится к способности индуцировать продукцию антитела против антигенного эпитопа, которой обладает нативный или встречающийся в природе ANGPTL4.
Антитело со «зрелой аффинностью» представляет собой антитело с одним или несколькими изменениями в одной или нескольких CDR, которые приводят к улучшению аффинности антитела по отношению к антигену по сравнению с исходным антителом, в котором нет указанного изменения(ий). Предпочтительные антитела со зрелой аффинностью будут иметь наномолярные или даже пикомолярные аффинности по отношению к антигену-мишени. Антитела со зрелой аффинностью получают способами, известными в данной области. В Marks et al. Bio/Technology 10: 779-783 (1992) описано созревание аффинности в результате перетасовки доменов VH и VL. Случайный мутагенез остатков CDR и/или каркаса описан в Barbas et al. Proc. Nat. Acad. Sci, USA 91: 3809-3813 (1994); Schier et al. Gene 169: 147-155 (1995); Yelton et al. J. Immunol. 155: 1994-2004 (1995); Jackson et al., J. Immunol. 154(7): 3310-9 (1995) и Hawkins et al, J. Mol. Biol. 226: 889-896 (1992).
«Функциональный антигенсвязывающий участок» антитела представляет собой участок, который способен связывать антиген-мишень. Аффинность связывания антигена антигенсвязывающего участка необязательно такая же высокая, как у исходного антитела, из которого антигенсвязывающий участок получают, но способность связывать антиген должна быть измеримой с использованием любого из множества способов, известных для оценки связывания антитела с антигеном. Кроме того, аффинность связывания антигена каждого из антигенсвязывающих участков поливалентного антитела согласно настоящему изобретению необязательно должна быть одинаковой количественно. В случае мультимерных антител согласно изобретению количество функциональных антигенсвязывающих участков можно оценить с использованием анализа на основе ультрацентрифугирования. Согласно указанному способу анализа в разных соотношениях объединяют антиген-мишень и мультимерное антитело и рассчитывают среднюю молекулярную массу комплексов, допуская разные количества функциональных участков связывания. Полученные теоретические значения сравнивают с полученными реальными экспериментальными значениями, чтобы оценить количество функциональных участков связывания.
Антитело, обладающее «биологическим свойством» указанного антитела, представляет собой антитело, которое имеет одно или несколько биологических свойств указанного антитела, которые отличают его от других антител, которые связываются с тем же самым антигеном. Чтобы провести скрининг антител, которые связываются с эпитопом на антигене, связываемом представляющим интерес антителом, можно осуществить обычный анализ перекрестного блокирования, такой как анализ, описанный в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988).
«Полипептидной цепью» является полипептид, в котором каждый из его доменов связан с другим доменом(ами) пептидной связью(ями), которые противоположны нековалентным взаимодействиям или дисульфидным связям.
«Гибкий линкер» в данном описании относится к пептиду, содержащему два или более аминокислотных остатков, связанных пептидной связью(ями), и он обеспечивает больше свободы вращения для двух соединенных им полипептидов (таких как две Fd-области). Такая свобода вращения позволяет двум или более антигенсвязывающим участкам, соединенным гибким линкером, более эффективно достигать антигена-мишени. Примеры подходящих последовательностей гибких линкерных пептидов включают gly-ser, gly-ser-gly-ser, ala-ser и gly-gly-gly-ser.
«Домен димеризации» образован в результате связывания, по меньшей мере двух, аминокислотных остатков (как правило, остатков цистеина) или, по меньшей мере, двух пептидов или полипептидов (которые могут иметь одну и ту же или разные аминокислотные последовательности). Пептиды или полипептиды могут взаимодействовать друг с другом посредством ковалентной и/или нековалентной связи(связей). Примеры доменов димеризации включают Fc-область; шарнирную область; домен CH3; домен CH4; пару CH1-CL; «область контакта», имеющую сконструированное «выпячивание» и/или «выступ», которая описана в патенте США №5821333, специально включенном в данное описание в виде ссылки; лейциновую молнию (например, лейциновую молнию jun/fos, см. Kostelney et al., J. Immunol., 148: 1547-1553 (1992); или лейциновую молнию GCN4 дрожжей); изолейциновую молнию; димерную пару рецепторов (например, рецептора интерлейкина-8 (IL-8R); и гетеродимеры интегрина, такие как LFA-1 и GPIIIb/IIIa) или область(и) их димеризации; димерные полипептиды лигандов (например, фактора роста нервов (NGF), нейтротрофина-3 (NT-3), интерлейкина-8 (IL-8), фактора роста эндотелия сосудов (VEGF), VEGF-C, VEGF-D, представителей PDGF и полученного из головного мозга нейротрофического фактора (BDNF); см. Arakawa et al. J. Biol. Chem. 269(45): 27833-27839 (1994) и Radziejewski et al. Biochem. 32(48): 1350 (1993)), или область(и) их димеризации; пару остатков цистеина, способных образовывать дисульфидную связь; пару пептидов или полипептидов, каждый из которых содержит, по меньшей мере, один остаток цистеина (например, от примерно одного, двух или трех до примерно десяти остатков цистеина), так чтобы могла образоваться дисульфидная связь(связи) между пептидами или полипептидами (в дальнейшем «синтетический шарнир»); и вариабельные домены антител. Наиболее предпочтительным доменом димеризации согласно изобретению является Fc-область или шарнирная область.
Фраза «стимуляция пролиферации клетки» охватывает стадию увеличения показателя роста и/или размножения клетки по сравнению с необработанной клеткой или клеткой, обработанной в меньшей степени in vitro или in vivo. Увеличение клеточной пролиферации в культуре клеток можно выявить подсчетом количества клеток до и после воздействия представляющей интерес молекулой. Степень пролиферации можно количественно оценить при микроскопическом исследовании степени слияния слоя. Пролиферацию клеток также можно количественно оценить, используя анализы, известные в данной области, например, анализ включения тимидина и коммерчески доступные анализы. Фраза «ингибирование пролиферации клетки» охватывает стадию снижения показателя роста и/или размножения клетки по сравнению с необработанной клеткой или клеткой, обработанной в меньшей степени, либо in vitro, либо in vivo. Ингибирование можно количественно оценить, как описано выше.
Введение «в комбинации с» одним или несколькими дополнительными терапевтическими средствами включает в себя совместное (одновременное) и/или последовательное введение в любом порядке.
«Субъект» в целях лечения относится к любому животному. Как правило, животное является млекопитающим. «Млекопитающее» в целях лечения относится к любому животному, классифицируемому как млекопитающее, включая человека, домашних и сельскохозяйственных животных и животных в зоопарках, спортивных животных или комнатных животных, таких как собаки, лошади, кошки, коровы, овцы, свиньи и т.д. Обычно млекопитающим является человек.
Термин «улучшает» или «улучшение» в используемом в данном описании смысле относится к уменьшению, ослаблению или элиминации состояния, заболевания, расстройства или фенотипа, включая аномалию или симптом.
«Расстройство» означает любое состояние, при котором может быть полезно лечение с использованием молекулы согласно изобретению. Такое состояние включает хронические и острые расстройства или заболевания, включая такие патологические состояния, которые обусловливают предрасположенность субъекта к рассматриваемому расстройству.
Термин «эффективное количество» или «терапевтически эффективное количество» относится к количеству лекарственного средства, эффективному при лечении заболевания или расстройства у субъекта.
«Лечение» относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. К субъектам, нуждающимся в лечении, относятся субъекты, уже имеющие расстройство, а также субъекты, у которых необходимо предотвратить расстройство.
«Гипертрофию» в используемом в данном описании смысле определяют как увеличение массы органа или структуры, независимое от природного роста, в которое не вовлечено образование опухоли. Гипертрофия органа или ткани является следствием либо увеличения массы отдельных клеток (истинная гипертрофия), либо следствием увеличения количества клеток, составляющих ткань (гиперплазия), либо и того и другого. Например, гипертрофический рост адипоцитов представляет собой увеличение размера адипоцита, стимулированное накоплением липидов. Гиперпластический рост адипоцитов представляет собой увеличение количества адипоцитов в ткани.
Фразы «сердечно-сосудистое и эндотелиальное расстройство», «сердечно-сосудистая и эндотелиальная дисфункция» и «сердечно-сосудистое, эндотелиальное или ангиогенное расстройство» используют взаимозаменяемо, и они относятся к расстройствам, обычно системным, которые стимулируют ангиогенез и/или кардиоваскуляризацию. К ним относятся заболевания, которые поражают сосуды, а также заболевания самих сосудов, таких как артерии, капилляры, вены и/или лимфатические сосуды. Такие расстройства включают без ограничения, например, болезни артерий, такие как атеросклероз, сахарный диабет, гипертония, воспалительные васкулитиды, болезнь Рейно и синдром Рейно, аневризмы и рестеноз артерий; венозные и лимфатические расстройства, такие как тромбофлебит, воспаление лимфатических узлов и лимфедема; злокачественную опухоль, такую как опухоли сосудов, например, гемангиома (капиллярная и кавернозная), гломангиомы, телангиэктазия, бациллярный ангиоматоз, гемангиоэндотелиома, ангиосаркома, гемангиоперицитома, саркома Капоши, лимфангиома и лимфангиосаркома; ангиогенез опухоли и другие сосудистые расстройства, такие как болезнь периферических сосудов, травму, такую как раны, ожоги и другие повреждения ткани, фиксация имплантата, рубцы, ишемическое реперфузионное повреждение, ревматоидный артрит, болезнь сосудов головного мозга, почечные заболевания, такие как острая почечная недостаточность; инсульт, болезнь коронарных артерий, гиперхолестеринемию, гипертриглицеридемию и/или остеопороз. Расстройства также могут включать стенокардию, инфаркты миокарда, такие как острые инфаркты миокарда, гипертрофию сердца и сердечную недостаточность, такую как застойная сердечная недостаточность (CHF). Также включены сердечно-сосудистые заболевания, связанные с дислипидемией, например, без ограничения, гипертония, атеросклероз, сердечная недостаточность, инсульт, различные болезни коронарных артерий, ожирение, диабет и т.д.
Термин «расстройство гомеостаза липидов» включает расстройство, заболевание или состояние, ассоциированное, вызванное и/или связанное с аномальной регуляцией (например, повышающей регуляцией или понижающей регуляцией) метаболизма липидов. Расстройства гомеостаза липидов могут быть вызваны или связаны с аномальным липолизом, аномальным поглощением липидов, аномальным синтезом и/или секрецией липидов, аномальным внутриклеточным высвобождением и/или обменом липидов, аномальным внутриклеточным высвобождением и/или обменом триглицеридов, аномальной внутриклеточной массой липидов и/или триглицеридов, и/или аномальной секретируемой массой липидов и/или триглицеридов в клетке или из клетки, например, клетки печени. Расстройства гомеостаза липидов включают без ограничения атеросклероз, ожирение, состояния, связанные с ожирением, диабет, резистентность к инсулину, гиперлипидемию, гиполипидемию, дислипидемию, гиперхолестеринемию, гипохолестеринемию, болезнь накопления триглицеридов, сердечно-сосудистое заболевание, болезнь коронарных артерий, гипертонию, инсульт, избыточный вес, анорексию, кахексию, гиперлипопротеинемию, гиполипопротеинемию, болезнь Ниманна-Пика, гипертриглицеридемию, гипотриглицеридемию, панкреатит, диффузный идиопатический скелетный гиперостоз (DISH), фенотип атерогенного липопротеина (ALP), эпилепсию, болезнь печени, жировую печень, стеатогепатит, синдром поликистозных яичников, злокачественную опухоль и т.д. Термин «расстройство метаболизма липидов» относится к аномальных клиническим химическим уровням холестерина и триглицеридов. Термин «гиперлипидемия» или «гиперлипемия» относится к состоянию, при котором имеют место более высокие, чем в норме, уровни липидов в сыворотке. Липиды сыворотки включают холестерин (сложный эфир и свободный), липопротеины, триглицериды, свободные жирные кислоты и другие стерины. В одном аспекте повышенные уровни указанных липидов являются показателем атеросклероза.
Термин «ожирение» относится к состоянию, при котором млекопитающее имеет индекс массы тела (BMI), который рассчитывают в виде массы (кг) на рост2 (метры2), составляющий по меньшей мере 25,9. Условно люди с нормальной массой имеют BMI от 19,9 до менее чем 25,9. Ожирение согласно изобретению может быть вызвано любой причиной, либо генетической, либо окружающей средой. Примеры расстройств, которые могут приводить к ожирению или могут быть причиной ожирения, включают, например, без ограничения, переедание и булимию, поликистоз яичников, краниофарингиому, синдром Прадера-Вилли, синдром Фрелиха, диабет типа II, субъекты с дефицитом GH, нормальный вариант низкорослости, синдром Тернера и другие патологические состояния, при которых проявляется пониженная метаболическая активность или снижение потребления энергии в покое в виде процента от общей нежировой массы, например, у детей с острым лимфобластным лейкозом. К «определяющим ожирение признакам» относятся жировые клетки и ткань, например, жировые тела, общая масса тела, уровни триглицеридов в мышцах, печени и жировой ткани и уровни лептина, свободных жирных кислот и триглицеридов в крови при голодании и в отсутствие голодания.
Термин «состояния, связанные с ожирением» относится к состояниям, которые являются результатом или которые обостряются при ожирении, такие как, без ограничения, дерматологические расстройства, такие как инфекции, варикозные вены, акантокератодермию и экзему, непереносимость физической нагрузки, сахарный диабет, резистентность к инсулину, гипертонию, гиперхолестеринемию, желчнокаменную болезнь, остеоартрит, повреждение опорно-двигательного аппарата, тромбоэмболическое заболевание, злокачественную опухоль (например, рак молочной железы, рак ободочной кишки, рак простаты и т.д.) и коронарную (или сердечно-сосудистую) болезнь сердца, в частности, сердечно-сосудистые состояния, связанные с высокими уровнями триглицеридов и свободных жирных кислот у субъекта.
Ожирение представляет собой наиболее распространенное из расстройств массы тела, поражающее по оценкам от 30 до 50% популяции среднего возраста в странах Запада. Другие расстройства массы тела, такие как нервная анорексия и нейрогенная булимия, которые вместе поражают примерно 0,2% женской популяции в западных странах, также представляют собой серьезную опасность для здоровья. Кроме того, такие расстройства, как анорексия и кахексия (истощение) также являются заметными признаками других заболеваний, таких как злокачественная опухоль, кистозный фиброз и СПИД.
Термин «истощающие» расстройства (например, синдром атрофии, кахексия, саркопения) относится к расстройству, вызванному нежелательной и/или болезненной потерей массы или потерей клеточной массы тела. У пожилых людей, а также у пациентов со СПИД и злокачественной опухолью истощающее заболевание может приводить к нежелательной потере массы тела, включая как жировую, так и нежировую составляющие. Истощающие заболевания могут быть результатом неадекватного потребления пищи и/или метаболических изменений, связанных с болезнью и/или процессом старения. Пациенты со злокачественной опухолью и пациенты со СПИД, а также пациенты после обширной хирургической операции или имеющие хронические инфекции, иммунологические болезни, гипертиреоидизм, внекишечные проявления болезни Крона, психогенное заболевание, хроническую сердечную недостаточность или другую тяжелую травму, часто страдают от истощающего заболевания, которое иногда также называют кахексией, метаболическим расстройством и иногда расстройством пищевого поведения. Кахексия дополнительно характеризуется гиперметаболизмом и гиперкатаболизмом. Хотя кахексию и истощающее заболевание часто используют взаимозаменяемо по отношению к состояниям, вызывающим истощение, существует, по меньшей мере, одно исследование, которое отличает кахексию от синдрома истощения, как например, потеря нежировой массы и, особенно, клеточной массы тела (Mayer, 1999, J. Nutr. 129 (1S Suppl.): 256S-259S). Саркопения, еще одно такое расстройство, которое может поражать стареющего человека, обычно характеризуется потерей мышечной массы. Конечная стадия истощающего заболевания, которое описано выше, может развиваться у людей, страдающих либо от кахексии, либо от саркопении.
Диабет является хроническим расстройством, поражающим метаболизм углеводов, жиров и белков у животных. Диабет является главной причиной слепоты, почечной недостаточности и ампутаций нижних конечностей у взрослых людей и является основным фактором риска для сердечно-сосудистого заболевания и инсульта.
Сахарный диабет типа I (или инсулинозависимый сахарный диабет («IDDM») или юношеский диабет) составляет примерно 10% всех случаев диабета. Заболевание характеризуется прогрессирующей потерей функции секреции инсулина бета-клетками поджелудочной железы. Указанная характеристика также является общей и для неидиопатического или «вторичного» диабета, берущего свое начало от болезни поджелудочной железы. Сахарный диабет типа I связан с такими клиническими признаками или симптомами, например, со стойко повышенной концентрацией глюкозы в плазме или гипергликемией; полиурией; полидипсией и/или гиперфагией; хроническими капиллярными осложнениями, такими как ретинопатия, нефропатия и нейропатия; и такими осложнениями макрососудов, как гиперлипидемия и гипертония, которые могут приводить к слепоте, заболеванию почек в конечной стадии, ампутации конечностей и инфаркту миокарда.
Сахарный диабет типа II (инсулиннезависимый сахарный диабет или NIDDM) является метаболическим расстройством, в которое вовлечена нарушенная регуляция метаболизма глюкозы и нарушенная чувствительность к инсулину. Сахарный диабет типа II обычно развивается во взрослом состоянии и связан с неспособностью организма использовать или достаточно производить инсулин. Кроме резистентности к инсулину, наблюдаемой в тканях-мишенях, пациенты, страдающие от сахарного диабета типа II, имеют сравнительную недостаточность инсулина, т.е. пациенты имеют более низкие уровни инсулина, чем прогнозируемые для данной концентрации глюкозы в плазме. Сахарный диабет типа II характеризуется следующими клиническими признаками или симптомами, например, со стойко повышенной концентрацией глюкозы в плазме или гипергликемией; полиурией; полидипсией и/или гиперфагией; хроническими микрокапиллярными осложнениями, такими как ретинопатия, нефропатия и нейропатия; и осложнениями макрососудов, такими как гиперлипидемия и гипертония, которые могут приводить к слепоте, болезни почек в конечной стадии, ампутации конечностей и инфаркту миокарда.
Синдром X, также называемый синдромом резистентности к инсулину (IRS), метаболическим синдромом или метаболическим синдромом X, распознают приблизительно в 2% случаев диагностических катетеризаций коронарных артерий. Часто приводящий к нетрудоспособности указанный синдром представляет собой симптомы и факторы риска развития сахарного диабета типа II и сердечно-сосудистого заболевания, включая, например, нарушенную толерантность к глюкозе (IGT), нарушенный уровень глюкозы натощак (IFG), гиперинсулинемию, резистентность к инсулину, дислипидемию (например, высокие уровни триглицеридов, низкие уровни ЛВП), гипертонию и ожирение.
Иммунологические расстройства включают без ограничения, например, системную красную волчанку; ревматоидный артрит; ювенильный хронический артрит; спондилоартропатии; системный склероз (склеродерму); идиопатические воспалительные миопатии (дерматомиозит, полимиозит); синдром Шегрена; системный васкулит; саркоидоз; аутоиммунную гемолитическую анемию (иммунную панцитопению, пароксизмальную ночную гемоглобинурию); аутоиммунную тромбоцитопению (идиопатическую тромбоцитопеническую пурпуру, опосредованную иммунной системой тромбоцитопению); тиреоидит (диффузный токсический зоб, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит, атрофический тиреоидит); сахарный диабет; опосредованное иммунной системой заболевание почек (гломерулонефрит, тубулярно-интерстициальный нефрит); демиелинизирующие заболевания центральной и периферической нервной системы, такие как рассеянный склероз, идиопатическая димиелинизирующая полинейропатия или синдром Гийена-Барре и хроническая воспалительная демиелинизирующая полинейропатия; гепатобилиарные заболевания, такие как инфекционный гепатит (гепатит A, B, C, D, E и другие негепатотропные вирусы), аутоиммунный хронический активный гепатит, первичный билиарный цирроз, гранулематозный гепатит и склерозирующий холангит; воспалительное заболевание кишечника (язвенный колит, болезнь Крона); глутенчувствительную энтеропатию и болезнь Уиппла; аутоиммунные или опосредованные иммунной системой заболевании кожи, включая буллезные заболевании кожи, мультиформную эритему и контактный дерматит, псориаз; аллергические болезни, такие как астма, аллергический ринит, атопический дерматит, гиперчувствительность к пищевым продуктам и крапивницу; иммунологические заболевания легких, такие как эозинофильная пневмония, идиопатический легочный фиброз и гиперчувствительный пневмонит; и/или заболевания, связанные с трансплантацией, включая отторжение трансплантата и болезнь трансплантат-против-хозяина. Могут присутствовать другие расстройства, такие как расстройство развития (например, эмбриональная смертность), неврологическое расстройство (например, пониженная подобная тревожности реакция при тестировании активности в открытом поле, аномальный циркадный ритм при тестировании активности в случае клеточного содержания и т.д.), аномалии глаз (например, аномалии сетчатки); и/или метаболическое нарушение или расстройство костей (например, артрит, остеопороз и/или остеопетроз).
Слово «метка» при использовании в данном описании относится к регистрируемому соединению или композиции, которые конъюгированы прямо или опосредованно с полипептидом. Метку можно регистрировать как таковую (например, радиоизотопные метки или флуоресцентные метки) или, как в случае ферментной метки, она может катализировать химическое изменение субстратного соединения или композиции, которое регистрируется.
«Изолированной» молекулой нуклеиновой кислоты является молекула нуклеиновой кислоты, которую идентифицируют и отделяют, по меньшей мере, от одной примесной молекулы нуклеиновой кислоты, с которой она обычно ассоциирована в природном источнике нуклеиновой кислоты полипептида. Изолированная молекула нуклеиновой кислоты имеет другие форму или окружение, отличные от той формы и окружения, в которых она встречается в природе. Поэтому изолированные молекулы нуклеиновой кислоты отличаются от молекулы нуклеиновой кислоты, в виде которой они существуют в природных клетках. Однако изолированная молекула нуклеиновой кислоты включает в себя молекулу нуклеиновой кислоты, которая содержится в клетках, которые обычно экспрессируют полипептид, в которых молекула нуклеиновой кислоты находится, например, в положении хромосомы, отличном от положения в хромосоме природных клеток.
Выражение «регуляторные последовательности» относится к последовательностям ДНК, необходимым для экспрессии оперативно связанной кодирующей последовательности в организме конкретного хозяина. Регуляторные последовательности, которые подходят для прокариот, например, включают в себя промотор, необязательно последовательность оператора и участок связывания рибосомы. Эукариотические клетки, как известно, используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является «оперативно связанной», когда она размещена в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК для препоследовательности или лидера секреции является оперативно связанной с ДНК для полипептида, если она экспрессируется в виде пребелка, который принимает участие в секреции полипептида; промотор или энхансер является оперативно связанным с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или участок связывания рибосом является оперативно связанным с кодирующей последовательностью, если он расположен так, чтобы способствовать трансляции. Обычно «оперативно связанные» означает, что последовательности ДНК, которые связаны, являются соседними и в случае лидера секреции являются соседними и находятся в рамке считывания. Однако энхансеры не должны быть соседними. Связывание осуществляют лигированием в подходящих сайтах рестрикции. Если таких сайтов не существует, то используют синтетические олигонуклеотидные адаптеры или линкеры, согласно обычной практике.
В используемом в данном описании смысле выражения «клетка», «линия клеток» и «культура клеток» используют взаимозаменяемо, и все такие определения включают потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают первичную клетку субъекта и полученные из нее культуры, независимо от количества переносов. Также понятно, что все потомство может быть не строго идентичным по содержанию ДНК вследствие преднамеренных или случайных мутаций. В изобретение включено мутантное потомство, которое имеет такую же функцию или биологическую активность, которые выявлены в исходно трансформированной клетке. Случаи, когда имеют в виду особые отличительные признаки, будут понятны из контекста.
Термин «ген» относится к (a) гену, содержащему последовательность ДНК, кодирующую ANGPTL4, например, депозит ATCC с номером 209284, или см. фиг.1; (b) любой последовательности ДНК, которая кодирует аминокислотную последовательность ANGPTL4 (см., например, фиг.2) и/или; (c) любой последовательности ДНК, которая гибридизуется с комплементом кодирующих последовательностей, описанных в данной публикации. В некоторых вариантах термин включает кодирующие, а также некодирующие области, и предпочтительно включает все последовательности, необходимые для нормальной экспрессии гена.
Термин «целенаправленное изменение гена» относится к типу гомологичной рекомбинации, которая происходит, когда фрагмент геномной ДНК вводят в клетку млекопитающего, и указанный фрагмент располагается и рекомбинирует с эндогенными гомологичными последовательностями. При целенаправленном изменении гена посредством гомологичной рекомбинации используют способы на основе рекомбинантной ДНК, чтобы заменить специфичные геномные последовательности экзогенной ДНК конкретной конструкции.
Термин «гомологичная рекомбинация» относится к обмену фрагментами ДНК между двумя молекулами ДНК или хроматидами в участке гомологичных нуклеотидных последовательностей.
Термин «ген-мишень» (альтернативно называемый «последовательностью гена-мишени» или «последовательностью ДНК мишени») относится к любой молекуле нуклеиновой кислоты, полинуклеотиду или гену, который модифицируют посредством гомологичной рекомбинации. Последовательность-мишень содержит интактный ген, экзон или интрон, регуляторную последовательность или любую область между генами. Ген-мишень может содержать часть конкретного гена или генетического локуса в геномной ДНК индивидуума.
«Нарушение» гена ANGPTL4 происходит, когда фрагмент геномной ДНК располагается и рекомбинирует с эндогенной гомологичной последовательностью, при этом нарушение представляет собой делецию нативного гена или его части, или мутацию нативного гена, или при этом нарушение представляет собой функциональную инактивацию нативного гена. Альтернативно нарушения последовательности могут быть созданы посредством неспецифичной инсерционной инактивации с использованием вектора генной ловушки (т.е. трансгенные животные, отличные от человека, содержащие и экспрессирующие случайно встроенный трансген; см., например, патент США №6436707, опубликованный 20 августа 2002). Указанные нарушения или модификации последовательности могут включать инсерции, миссенс-мутации, сдвиг рамки, делеции или замены или перестановки последовательности ДНК или любую их комбинацию. К инсерциям относится инсерция полных генов, которые могут иметь происхождение из животного, растения, гриба, насекомого, прокариота или вируса. Нарушение, например, может изменять нормальный генный продукт посредством ингибирования его продукции, частично или полностью, или посредством усиления активности нормального генного продукта. В одном варианте нарушение представляет собой нарушение до нуля, когда нет значимой экспрессии гена ANGPTL4.
Термин «нативная экспрессия» относится к экспрессии полноразмерного полипептида, кодируемого геном ANGPTL4, на уровне экспрессии, имеющейся у мыши дикого типа. Таким образом, нарушение, при котором «нет нативной экспрессии» эндогенного гена ANGPTL4, относится к частичному или полному снижению экспрессии, по меньшей мере, части полипептида, кодируемого эндогенным геном ANGPTL4, в одной клетке, в выбранных клетках или всех клетках млекопитающего.
Термин «нокаут» относится к нарушению гена ANGPTL4, при котором нарушение приводит к функциональной инактивации нативного гена; делеции нативного гена или его части; или мутации в нативном гене.
Термин «вставка» («knock-in») относится к замене мышиного ортолога (или другого мышиного гена) кДНК человека, кодирующей гены, кодирующие ANGPTL4, или их варианты (т.е. нарушение приводит к замене нативного гена мыши нативным геном человека).
Термин «конструкция» относится к искусственно собранному участку ДНК для переноса в ткань-мишень, линию клеток или животное. Как правило, конструкция будет содержать ген или последовательность нуклеиновой кислоты, представляющие конкретный интерес, маркерный ген и соответствующие регуляторные последовательности. Как предлагается в данном изобретении, направляющая к мишени конструкция ANGPTL4 содержит последовательность ДНК, гомологичную, по меньшей мере, одной части гена ANGPTL4 и способную вызывать нарушение в гене ANGPTL4 в клетке-хозяине.
Термин «трансгенная клетка» относится к клетке, содержащей в своем геноме ген ANGPTL4, который был разрушен, модифицирован, изменен или заменен полностью или частично способом целенаправленного изменения гена.
Термин «трансгенное животное» относится к животному, которое содержит в своем геноме конкретный ген, который был разрушен или иным образом модифицирован или мутирован с помощью способов, описанных в данной публикации, или же способов, хорошо известных в данной области. В некоторых вариантах трансгенным животным, отличным от человека, является млекопитающее. В одном варианте млекопитающее является грызуном, таким как крыса или мышь. Кроме того, «трансгенное животное» может быть гетерозиготным животным (т.е. один дефектный аллель и один аллель дикого типа) или гомозиготным животным (т.е. два дефектных аллеля). Считается, что эмбрион входит в объем определения животного. Указание животного включает в себя указание эмбриона или плода in utero, либо в результате спаривания, либо в результате иных обстоятельств, и независимо от того, достигает ли он или не достигает срока родов.
В используемом в данном описании смысле термины «маркер селекции» и «маркер селекции положения» относится к гену, кодирующему продукт, который позволяет только тем клеткам, которые несут ген, выживать и/или расти в определенных условиях. Например, растительные и животные клетки экспрессируют введенный ген резистентности к неомицину (Neor), резистентный к соединению G418. Клетки, которые не несут маркерного гена Neor, погибают при действии G418. Другие маркеры позитивной селекции известны или находятся в компетенции специалистов в данной области.
Термин «модулирует» или «модулирование» в используемом в данном описании смысле относится к уменьшению, ингибированию, снижению, улучшению, увеличению или усилению функции, экспрессии активности гена ANGPTL4 или, альтернативно, фенотипа, связанного с геном ANGPTL4.
Термин «аномалия» относится к любому заболеванию, расстройству, состоянию или фенотипу, в которые вовлечен ANGPTL4, включая патологические состояния и поведенческие проявления.
ANGPTL4
Изобретение является результатом осуществления желания дополнительно выяснить биологическую функцию ANGPTL4 и его роль в патологических состояниях. Экспрессия ANGPTL4 главным образом обнаружена в плаценте, жировой ткани, ткани печени и почек. Настоящее изобретение относится к дополнительным применениям ANGPTL4 и модуляторов ANGPTL4 в области гепатоцитов, адипоцитов и гомеостаза липидов. Изобретение также описывает трансгенных или нокаутированных мышей, содержащих нарушение в гене ANGPTL4, и их применения.
Ангиопоэтинподобный белок 4 (ANGPTL4) является секретируемым белком и является членом семейства ангиопоэтина. Он также известен как фибриноген печени/ангиопоэтинродственный белок (HFARP) (Kim et al., Biochem. J. 346: 603-610 (2000)), PGAR (PPARγ-ангиопоэтинродственный белок) (Yoon, et al., Mol. Cell Biol., 20: 5343-5349 (2000)), индуцируемый голоданием адипозный фактор (FIAF) (Kerten et al., J. Biol. Chem., 275: 28488-28493 (2000)); ангиопоэтинродственный белок (ARP-4); NL2 (см. патенты США №6348350, 6372491 и 6455496); и Ang6.
Белок ANGPTL4 человека является белком из 406 аминокислот (например, патенты США №6348350, 6372491 и 6455496), тогда как мышиный ANGPTL4 является белком из 410 аминокислот (Kim et al., Biochem. J. 346: 603-610 (2000)). Белки мыши и человека обладают примерно 75% идентичностью на уровне аминокислот. Kim et al., Biochem. J. 346: 603-610 (2000). ANGPTL4 имеет сигнальный пептид, три потенциальных сайта N-гликозилирования и четыре остатка цистеина, которые могут быть вовлечены в образование внутримолекулярных дисульфидных связей. ANGPTL4 образует более высокомолекулярные структуры, например, как показано на фиг.3, панель A. См. также, например, Ge et al., J. Biol. Chem., 279(3): 2038-2045 (2004); Ge et al., J. Lipid Res. 45: 2071-2079 (2004) и Mandard et al., J. of Biol. Chem., 279(33): 34411-34420 (2004). ANGPTL4 также может протеолитически процессироваться, например, замена R162G и R164E в ANGPTL4 приводит к варианту ANGPTL4, разгоняемому на уровне более высокой молекулярной массы в SDS-геле, чем белок дикого типа (см., фиг.3, панель B). См., также, например, Ge et al., J. Biol. Chem., 279(3): 2038-2045 (2004) и Mandard et al., J. of Biol. Chem., 279(33): 34411-34420 (2004).
Консервативные области семейства ангиопоэтина включают биспиральный домен и C-концевой фибриноген (FBN)-подобный домен. См., например, Kim et al., Biochem. J. 346: 603-610 (2000). Предполагается, что ANGPTL4 протеолитически процессируется регулируемым образом с высвобождением C-концевого фибриногенподобного домена. См., например, Ge et al., J. Biol. Chem., 279(3): 2038-2045 (2004).
ANGPTL4 связывается с интегрином αVβ5. См., например, фиг.9, панели A-E. Другой представитель семейства ангиопоэтин-3-подобный белок (ANGPTL3) является ангиогенным фактором, который связывается с интегрином αVβ3. См., например, заявку на выдачу патента США 2003/0215451, опубликованную 20 ноября 2003, и Camenisch et al., J. Biol. Chem., 277(19): 17281-17290 (2002). ANGPTL3, по-видимому, не связывается с рецептором Tie2. Camenish et al., Journal of Biol. Chem. 277(19): 17281-17290 (2002). ANGPTL3 также является регулятором уровней липидов в плазме. См., например, Koishi et al., Nat. Genetics 30: 151-157 (2002).
Интегрин αVβ5 является рецептором белков внеклеточного матрикса, включая витронектин и Del-1 (см., например, Stupack and Cheresh, Journal of Cell Science 115: 3729-3738 (2002)). Альфа-v-интегрины вовлечены в прогрессирование опухолей и метастазы. См., например, Marshall, J. F. and Hart, I. R. Semin. Cancer Biol. 7(3): 129-38 (1996). Кроме того, также была показана роль альфа-v-интегринов в ходе ангиогенеза. См., например, Eliceiri, B.P. and Cheresh, D.A. Molecular Medicine 4: 741-750 (1998). Например, было показано, что моноклональное антитело к αVβ5 ингибирует VEGF-индуцированный ангиогенез в роговице кроликов и в модели на основе хориоаллантоисной оболочки цыпленка. См., например, M.C. Friedlander, et al., Science 270: 1500-1502 (1995). Также показано, что антагонисты αVβ3 и αVβ5 ингибируют ангиогенез, индуцированный фактором роста и опухолью. См., например, Eliceiri and Cheresh, Current Opinion in Cell Biology, 13: 563-568 (2001).
Применение ANGPTL4 и модуляторов ANGPTL4
Изобретение относится к применениям ANGPTL4 или его агониста или антагониста для модулирования разнообразных активностей и процессов в клетках, например, пролиферации и/или клеточной адгезии гепатоцитов и пролиферации преадипоцитов и/или клеточной миграции преадипоцитов. ANGPTL4 вовлечен в модулирование уровней триглицерида и холестерина в сыворотке. Кроме того, ANGPTL4 также может быть негативным регулятором воспалительных реакций. Модуляторы ANGPTL4 могут быть использованы для лечения расстройств и заболеваний, связанных с указанными активностями.
Печень
ANGPTL4 стимулирует пролиферацию гепатоцитов и адгезию гепатоцитов. Печень является основным органом для гомеостаза холестерина. См. также раздел «гомеостаз липидов» в данном описании. Печень отвечает за биосинтез холестерина и катаболизм холестерина. Печень синтезирует и секретирует липопротеины очень низкой плотности (VLDL). В кровообращении VLDL метаболизируются, становясь липопротеинами низкой плотности (LDL), которые являются основными несущими холестерин липопротеинами в плазме.
Печень действует в качестве стража, находящегося между пищеварительным трактом и остальной частью организма. Основная функция печени заключается в эффективном поглощении, хранении, метаболизме и поставке в кровь и желчь больших количеств таких веществ, как углеводы, липиды, аминокислоты, витамины и микроэлементы. Другая функция печени заключается в детоксикации ксенобиотических загрязняющих веществ, лекарственных средств и эндогенных метаболитов посредством механизмов фазы I (окисление/восстановление) и фазы II (конъюгирование).
Печень является основным контролирующим метаболизм органом в организме человека, который содержит тысячи мельчайших долек (lobuli hepatis), функциональных единиц органа. Печеночная ткань содержит два основных типа дифференцированных клеток: паренхимные клетки (т.е. гепатоциты) и непаренхимные клетки. Сложные функции печени в большой степени осуществляются гепатоцитами, тогда как непаренхимные клетки, такие как клетки Купфера, клетки Ито и эндотелиальные клетки синусоидов печени (LSEC) играют важные роли в поддержании и поставке материалов к гепатоцитам. Mochida et al. Biochem. Biophys. Res. Comm. 226: 176-179 (1996).
Кроме нормального роста в ходе раннего развития, ткань печени обладает уникальной способностью регенерировать во взрослом состоянии. Регенерация печени после потери печеночной ткани является основной составляющей процесса восстановления в ответ на различные формы повреждения печени, такие как гепатотоксичность, вирусная инфекция, повреждение сосудов и частичная гепатэктомия. Например, после частичной гепатэктомии размер печени обычно восстанавливается до своей исходной массы примерно за шесть дней. В процесс роста и регенерации печени вовлечена пролиферация как гепатоцитов, так и непаренхимных клеток, таких как эндотелиальные клетки синусоидов. Обычно первыми пролиферируют гепатоциты, а другие клетки печени вступают в фазу синтеза ДНК примерно через 24 часа после гепатоцитов. Michalopoulos and DeFrances Science 276: 60-65 (1997).
Изобретение относится к способам стимуляции роста печени и/или клеточной пролиферации гепатоцитов посредством введения эффективного количества ANGPTL4 или его агониста. Стимулирующее действие согласно изобретению можно оценить либо in vitro, либо in vivo, используя способы, известные в данной области. См., например, Drakes et al. J. Immunol. 159: 4268 (1997); Omori et al. Hepatology 26: 720 (1997) и патент США №5227158. Например, пролиферацию клеток оценивают во время культивирования с использованием способов, известных в данной области, включая, но не ограничивая указанным, измерение скорости синтеза ДНК (см., например, Nakamura et al. Biochem. Biophys. Res. Comm. 122: 1450 (1984), подсчет в гематоцитометре с исключением при окрашивании красителем трипановым синим (см., например, Omiri et al. (1997) выше) или проточная цитометрия (см., например, Drakes (1997) выше).
В некоторых вариантах осуществления изобретения ANGPTL4 или его агонист вводят для того, чтобы индуцировать клеточную адгезию гепатоцитов. Адгезию гепатоцитов можно оценить способами, известными в данной области, включая, например, анализ с использованием кристаллического фиолетового. См. также Landegren, U. J. Immunological Methods, 67: 379-388 (1984). В одном варианте осуществления изобретения гепатоциты и другие непаренхимные клетки печени выделяют из проверяемой печени и ресуспендируют в подходящей среде для культуры ткани с ANGPTL4 или его агонистом, чтобы индуцировать адгезию клеток. При необходимости разные клеточные фракции могут быть дополнительно разделены (например, паренхимные клетки отделены от непаренхимных клеток) центрифугированием с разными скоростями в течение разных периодов времени.
В другом варианте пролиферативное действие ANGPTL4 или агониста ANGPTL4 на клетки печени и орган печени в целом измеряют in vivo, используя, например, гистохимические анализы образцов ткани печени. В одном аспекте пролиферацию клеток печени in vivo оценивают по реактивности по отношению к антителу, направленному против белка, который, как известно, присутствует в более высоких концентрациях в пролиферирующих клетках, чем в не пролиферирующих клетках, такого как ядерный антиген пролиферирующих клеток (PCNA или циклин). Rodgers et al. J. Burn Care Rehabil. 18: 381-388 (1997). В другом аспекте можно использовать иммуногистохимический анализ BrdU, который описан Gerber et al. Development 126: 1149-1159 (1999).
Вследствие важной роли для жизни дисфункция и заболевания печени часто являются ослабляющими здоровье и угрожающими жизни. Ряд острых или хронических патологических состояний связаны со структурными и/или функциональными аномалиями печени. Такие состояния включают без ограничения печеночную недостаточность, гепатит (острый, хронический или алкогольный), цирроз печени, токсическое повреждение печени, медикаментозное повреждение печени, печеночную энцефалопатию, печеночную кому или некроз печени. Усиление клеточного роста гепатоцитов может быть полезным при лечении болезни печени. Соединения и способы согласно изобретению могут обеспечивать репарацию повреждений печени. Не будучи связанными с теорией, полагают, что это можно осуществить либо непосредственно, либо косвенно в результате стимуляции клеток печени к росту и делению. Согласно одному варианту изобретение относится к способам лечения патологического состояния печени у субъекта посредством введения эффективного количества ANGPTL4 или агониста ANGPTL4 согласно изобретению.
Фразу «патологическое состояние печени» используют взаимозаменяемо с фразами «расстройство печени» или «заболевание печени», чтобы указать какое-либо структурное и/или функциональное отклонение в печени от нормы. Неограничивающие примеры патологического состояния печени включают состояния, связанные с печеночной недостаточностью, гепатитом (острым, хроническим или алкогольным), циррозом печени, токсическим повреждением печени, медикаментозным повреждением печени, печеночной энцефалопатией, печеночной комой или некрозом печени.
В одном аспекте изобретение относится к способам защиты печени от повреждения у субъекта, чувствительного к условиям или факторам, вызывающим повреждение печени. Фразу «повреждение печени» используют в данном описании в самом широком смысле, и она указывает на любое структурное или функциональное повреждение печени, возникающее непосредственно или опосредованно в результате действия внутренних или внешних факторов или их комбинаций. Повреждение печени может быть индуцировано рядом факторов, включая, но не ограничиваясь указанным, воздействие гепатотоксических соединений, воздействие излучения, механические повреждения печени, генетическую предрасположенность, вирусные инфекции, аутоиммунное заболевание, такое как аутоиммунный хронический гепатит, и в результате повышенных уровней in vivo таких белков, как активин и TGF-β. Повреждения печени, индуцированные гепатотоксическими соединениями, включают прямую цитотоксичность, в том числе реакции гиперчувствительности к лекарственным средствам, холестаз и повреждение эндотелия сосудов.
Многие химические и биологические средства, либо терапевтические, либо совершенно опасные, могут индуцировать повреждения печени и следовательно являются гепатотоксичными. Гепатотоксичные соединения также являются важной причиной хронического заболевания печени, включая жировую печень, гепатит, цирроз и сосудистые и неопластические поражения печени. (Sinclair et al., Textbook of Internal Medicine, 569-575 (1992) (editor, Kelley; Publisher, J. B. Lippincott Co.). В изобретении предлагаются способы защиты печени субъекта от повреждения вследствие воздействия гепатотоксического агента, включающие в себя введение субъекту ANGPTL4 или агониста, при этом указанный ANGPTL4 или агонист ANGPTL4 эффективно защищает печень от повреждения. В одном аспекте ANGPTL4 или агонист ANGPTL4 вводят до или одновременно с воздействием на указанного субъекта гепатотоксического агента, при этом указанным гепатотоксическим агентом является терапевтическое средство, такое как химиотерапевтическое средство или излучение для лечения злокачественных опухолей. В таком случае способы служат для повышения эффективности лечения, обеспечивая толерантность субъекта к высоким дозам терапевтических средств. В другом аспекте ANGPTL4 или агонист ANGPTL4 вводят после воздействия на субъекта гепатотоксических агентов, но до появления какого-либо регистрируемого повреждения печени у субъекта. Такие способы могут быть применимы для лечения повреждений печени вследствие случайного воздействия гепатотоксического агента на субъекта.
Гепатотоксические агенты могут индуцировать повреждение печени непосредственно в результате цитотоксичности по отношению к печени или в результате образования токсических метаболитов (к указанной категории относится реакция гиперчувствительности, которая имитирует аллергию на лекарственные средства); холестаз, задержку протекания желчи вследствие обструкции желчных протоков и сосудистые повреждения, такие как окклюзионное заболевание вен (VOD), при котором повреждение эндотелия сосудов приводит к тромбозу печеночных вен. На индивидуальную чувствительность к повреждению печени, индуцированному гепатотоксическими агентами, влияют генетические факторы, возраст, пол, питание, воздействие других лекарственных средств и системные заболевания (Sinclair et al., Textbook of Internal Medicine, выше).
Многие гепатотоксические соединения вызывают непредсказуемые повреждения печени в небольшой части реципиентов. У некоторых пациентов повреждение печени называют реакцией гиперчувствительности, и она подобна реакции на лекарственные средства, при этом у пациента имеет место жар, сыпь и эозинофилия и происходят рецидивы симптомов при повторной стимуляции лекарственным средством. В других ситуациях механизм повреждения неизвестен и может представлять собой аномальный метаболизм у чувствительных пациентов, который создает возможность для образования или накопления гепатотоксических метаболитов.
Лекарственные средства, индуцирующие цитотоксичность в результате прямого химического воздействия, включают следующие средства: анестезирующие средства, такие как энфлуран, флуроксен, галотан и метоксифлуран; нейропсихотропные средства, такие как кокаин, гидразиды, метилфенидат и трициклические средства; противосудорожные средства, такие как фенитоин и вальпроевая кислота; анальгетики, такие как ацетаминофен, хлорзоксазон, дантролен, диклофенак, ибупрофен, индометацин, салицилаты, толметин и зоксазоламин; гормоны, такие как ацетогексамид, карбутамид, глипизид, метагексамид, пропилтиоурацил, тамоксифен, диэтилстилбестрол; противомикробные средства, такие как амфотерицин B, клиндамицин, кетоконазол, мебендазол, метронидазол, оксациллин, парааминосалициловая кислота, пенициллин, рифампицин, сульфонамиды, тетрациклин и зидовудин; сердечно-сосудистые лекарственные средства, такие как амиодарон, дилитиазем, a-метилдопа, мексилетин, гидразалин, никотиновая кислота, папаверин, пергексилин, прокаинамид, хинидин и токаинамид; и иммунодепрессанты и противоопухолевые средства, такие как аспарагиназа, цисплатин, циклофосфамид, дакарбазин, доксорубицин, фторурацил, метотрексат, митрамицин, 6-MP, нитрозомочевины, тамоксифен, тиогуанин и винкристин; и разнообразные лекарственные средства, такие как дисульфирам, ион йодида, оксифенизатин, витамин A и парааминобензойная кислота.
Гепатотоксические соединения, вызывающие реакцию гиперчувствительности в печени, включают следующие соединения: фенитоин, парааминосалициловую кислоту, хлорпромазин, сульфонамиды, эстолат эритромицина, изониазид, галотан, метилдопа и вальпроевую кислоту.
Повреждения гепатотоксическими соединениями, включая холестаз, задержку желчи, могут иметь несколько форм. Центрилобулярный холестаз сопровождается портальными воспалительными изменениями. Сообщалось об изменениях желчного протока при действии некоторых лекарственных средств, таких как эритромицин, тогда как строго канальцевый холестаз является характерной особенностью действия других лекарственных средств, таких как анаболические стероиды. Хронический холестаз был связан с такими лекарственными средствами, как метилтестостерон и эстрадиол.
Гепатотоксические соединения, индуцирующие холестатическое заболевание, включают следующие соединения: контрацептивные стероиды, андрогенные стероиды, анаболические стероиды, ацетилсалициловую кислоту, азатиоприн, бензодиазепин, хенодезоксихолевую кислоту, хлордиазэпоксид, эстолат эритромицина, флуфеназин, фуросемид, гризеофулвин, галоперидол, имипрамин, 6-меркаптопурин, метимазол, метотрексат, метилдопа, метилендиамин, метилтестостерон, напроксен, нитрофурантоин, пеницилламин, перфеназин, прохлорперазин, промазин, тиобендазол, тиоридазин, толбутамид, триметопримсульфаметоксазол, мышьяк, медь и гербициды.
Некоторые лекарственные средства, хотя и являются главным образом холестатическими, также могут вызывать гепатотоксичность, и поэтому повреждение печени, которое они вызывают, является смешанным. Лекарственные средства, вызывающие смешанное повреждение печени, включают, например, следующие средства: хлорпромазин, фенилбутазон, галотан, хлордиазэпоксид, диазепам, аллопуринол, фенобарбитал, напроксен, пропилтиоурацил, хлорамфеникол, триметопримсульфаметоксазол, амринон, дизопирамид, азатиоприн, циметидин и ранитидин.
Повреждения сосудов печени, включая тромбоз печеночных вен, окклюзию печеночных венул или веноокклюзионное заболевание (VOD) и печеночная пурпура, могут быть вызваны лекарственными средствами. Кроме того, могут возникать повреждения, включающие расширение синусоидов, перисинусоидальный фиброз и гепатопортальный склероз. Вначале сообщалось, что расширение синусоидов промежуточной и перицентральной зон является осложнением пероральной контрацептивной терапии. Печеночная пурпура является состоянием, при котором имеются большие заполненные кровью полости, которые возникают в результате просачивания эритроцитов через эндотелиальный барьер, с последующим перисинусоидальным фиброзом. Печеночная пурпура была описана у пациентов, принимающих пероральные контрацептивы, анаболические стероиды, азатиоприн и даназол. Также известно, что повреждение и окклюзия центральных печеночных венул связана с приемом внутрь пирролизидиновых алкалоидов, таких как травяные чаи. Начальным повреждением является центральный некроз, сопровождаемый прогрессирующим заболеванием, связанным с калибром венул. Все указанные повреждения могут быть обратимы только частично в случае прекращения приема лекарственного средства, и может развиваться цирроз.
Несколько типов доброкачественного и злокачественного новообразования печени могут быть результатом введения гепатотоксических соединений. Аденома, повреждение, ограниченно выявляемое у женщин в детородном возрасте, связано с применением контрацептивных стероидов, и риск увеличивается с увеличением продолжительности применения. Гепатоклеточная карцинома также может наблюдаться у пациентов, принимающих андрогенные гормоны по поводу апластической анемии или гипопитуитаризма.
Известные гепатотоксические соединения, вызывающие повреждения печени, включают следующие соединения: контрацептивные стероиды, пирролизидиновые алкалоиды, уретан, азатиоприн, 6-меркаптопурин, 6-тиогуанин, митомицин, BCNU, винкристин, адриамицин, внутривенный витамин E, анаболические-андрогенные стероиды, азатиоприн, ацетат медроксипрогестерона, сульфат эстрона, тамоксифен, неорганические соединения мышьяка, диоксид тория, витамин A, метотрексат, гидрохлорид метиламфетамина, витамин A, кортикостероиды, диоксид тория и терапию радием.
Повреждение печени, вызванное другими факторами, обычно принимает сходные формы. Повреждение печени, независимо от того, вызвано ли оно гепатотоксичностью соединения, лучевой терапией, генетической предрасположенностью, механическим повреждением или любой комбинацией указанных и других факторов, можно выявить несколькими способами. Биохимические тесты использовали клинически в течение многих лет в качестве стандартного показателя гепатотоксичности. Большинство биохимических тестов обычно относятся к двум категориям: тесты, в которых измеряют специфичные маркеры печени, например, протромбиновое время свертывания и/или печеночный кровоток, или тесты, в которых анализируют маркеры сыворотки, для выявления некроза, холестаза, прогрессирующего фиброгенеза или гепатомы (Cornelius, C. in Hepatotoxicology, Meeks et al. eds., pgs. 181-185 (1991)). Значимость таких тестов основана на их простоте и том факте, что они являются неинвазивными. Обоснование применения ферментов сыворотки для оценки повреждения печени заключается в том, что указанные ферменты, в норме находящиеся в клетках печени, получают доступ в общее кровообращение в случае повреждения клеток печени.
Повышенная активность ферментов сыворотки свидетельствует о некрозе и/или холестазе. Повышенные уровни конъюгатов билирубина в сыворотке свидетельствуют о внутрипеченочном или внепеченочном холестазе. Однако существуют некоторые ограничения для применения уровней ферментов в сыворотке в качестве единственных средств диагностики повреждения печени. Уровни ферментов в сыворотке могут увеличиваться в результате просачивания из клеток с измененной проницаемостью вследствие системного действия средства, а не специфичного повреждения печени, вызванного химическим веществом. Гистопатологическое исследование печени является следующей логической стадией идентификации и количественной оценки природы и степени повреждения печени.
Ферменты сыворотки в качестве маркеров повреждения печени можно разделить на четыре группы на основании специфичности и чувствительности к повреждению печени (Kodavanti et al., Toxicologic Pathology 20(4): 556-69 (1992); Kodavanti et al., Archives of Toxicology 63(5): 367-75 (1989).
Группа I: такие ферменты показывают более избирательно печеночный холестаз в случае их повышения, например, щелочная фосфатаза (AP), 5'-нуклеотидаза (5'-ND) и a-глутамилтранспептидаза (G-GT) и лейцинаминопептидаза (LAP). Группа II: такие ферменты в случае повышения показывают повреждение паренхимы, например, аспартаттрансаминаза (AST), аланинтрансаминаза (ALT), фруктозо-1,6-дифосфатальдолаза (ALD), лактатдегидрогеназа (LDH), изоцитратдегидрогеназа (ICDH), орнитинкарбамоилтрансфераза (OCT) и сорбитдегидрогеназа (SDH), аргиназа и гуаназа. Группа III: такие ферменты при повышении свидетельствуют о повреждении другой ткани, например, креатинфосфокиназа (CPK). Группа IV: такие ферменты подавлены при повреждении печени, например, холинэстераза (ChE).
Другие сывороточные маркеры включают уровни пептида проколлагена типа III (PIIIP) для оценки того, является ли фиброгенез печени активным; уровни аммиака в крови при гепатоэнцефалопатии; уровни лиганда при некрозе и гепатоме; уровни гиалуроната вследствие повреждения эндотелиальных клеток печени; уровни a-1-фетопротеина (AFP) для выявления гепатомы; уровни карциноэмбрионального антигена (CEA) для выявления метастазов злокачественной опухоли в печени; повышение титра антител против ряда клеточных компонентов, таких как митохондриальные, ядерные и специфичные для мембран печени белки; и выявление белков, таких как альбумин, глобин, аминокислот, холестерина и других липидов. Также биохимический анализ различных минеральных веществ, метаболитов и ферментов, полученных из образцов биопсии печени, может быть применим для исследования специфичных биохимических дефектов в случае наследственных, приобретенных и экспериментально индуцированных расстройств печени.
Могут быть осуществлены тесты в отношении функций печени, чтобы оценить повреждение печени. Тесты функций печени включают следующие анализы: к группе I относится оценка печеночного клиренса органических анионов, таких как билирубин, индоцианин зеленый (ICG), сульфобромфталеин (BSP) и желчные кислоты; к группе II относится оценка печеночного кровотока посредством измерений клиренса галактозы и ICG; и к группе III относится оценка функции микросом печени посредством использования дыхательного теста на аминопурин и тестирования клиренса кофеина. Например, можно измерить билирубин в сыворотке, чтобы подтвердить наличие и тяжесть желтухи и определить степень гипербилирубинемии, которая наблюдается при паренхимном заболевании печени. Оценки аминотрансферазы (трансаминазы) отражают тяжесть активного гепатоклеточного повреждения, тогда как по оценкам щелочной фосфатазы выносят решение о холестазе и инфильтратах печени (Isselbacher, K. and Podolsky, D. in Harrisson's Principles of Internal Medicine, 12th edition, Wilson et al. eds., 2: 1301-1308 (1991)). Способы осуществления анализа ферментов в сыворотке известны в данной области и описаны, например, в Kodavanti et al., выше.
Так как обширное повреждение печени может приводить к сниженным уровням в крови альбумина, протромбина, фибриногена и других белков, синтезируемых исключительно гепатоцитами, то уровни указанных белков могут быть измерены в качестве показателей повреждения печени. В отличие от измерений ферментов в сыворотке уровни белков в сыворотке отражают синтетическую функцию печени, а не точно повреждение клеток (Podolsky, D. Harrison's Principles of Internal Medicine, 12th edition, Wilson et al. eds., 2: 1308-1311 (1991)).
У многих пациентов компьютерная томография (CT), ультразвуковое исследование, сцинтиграммы или биопсия печени могут быть необходимы для определения природы заболевания печени (Isselbacher, K. and Friedman, L. and Needleman, L. in Harrison's Principles of Internal Medicine, 12th edition, Wilson et al. eds., 2: 1303-1307 (1991)).
Изобретение относится к способам усиления эффекта терапии у субъекта, при этом указанные способы включают в себя введение субъекту ANGPTL4 или агониста ANGPTL4 способом, эффективным для защиты печени субъекта от повреждения, вызываемого гепатотоксическим соединением, перед или одновременно с терапией, тем самым повышая толерантность субъекта к терапии. Например, химиотерапевтические средства, используемые в курсе химиотерапии, могут оказывать цитотоксическое воздействие на клетки печени, таким образом ограничивая дозу и/или продолжительность введения химиотерапевтического средства пациенту. При воздействии на печень композиции, содержащей ANGPTL4 или агонист ANGPTL4, такие токсические воздействия могут быть предотвращены или уменьшены. При этом дозу химиотерапевтических средств можно увеличить, тем самым повышая эффективность терапии злокачественной опухоли.
ANGPTL4 или агонист ANGPTL4 можно комбинировать с другими средствами в способах, описанных в данной публикации. Например, были привлечены несколько факторов роста и цитокинов, в качестве средств, способных индуцировать регенерацию печени, особенно фактор роста гепатоцитов (HGF), эпидермальный фактор роста (EGF), трансформирующий фактор роста (TGF), интерлейкин-6 (IL-6), фактор невроза опухолей-α (TNF-α), основные и кислотные факторы роста фибробластов, CTGF, HB-EGF и норэпинефрин. Fujiwara et al. Hepatol. 18: 1443-9 (1993); Baruch et al. J. Hepatol. 23: 328-32 (1995); Ito et al. Biochem. Biophys. Res. Commun. 198: 25-31 (1994); Suzuma et al. J. Biol. Chem. 275: 40725-31 (2000); и Michalopoulos and DeFrances (1997), выше. Указанные средства можно комбинировать с ANGPTL4 или агонистом ANGPTL4.
HGF, один из наиболее сильных митогенов печени, сначала был идентифицирован как фактор, способный стимулировать синтез ДНК в культивируемых гепатоцитах, но в настоящее время известно, что он имеет множество разных функций, влияя на многие эпителиальные клетки. Nakamura et al. Biochem. Biophys. Res. Comm. 122: 1450 (1984); и Russell et al. J. Cell. Physiol. 119: 183-192 (1984). HGF также известен как рассеивающий фактор (SF), что привело к обозначению HGF/SF. Stoker and Perryman J. Cell Sci. 77: 209-223 (1985) и Gherardi and Stoker Nature 346: 228 (1990). Биологические эффекты HGF преобразуются через единственный тирозинкиназный рецептор Met, продукт протоонкогена Met. В печени HGF экспрессируется в других клетках, отличных от гепатоцитов, таких как клетки Ито и LSEC, тогда как транскрипты met экспрессируются строго в гепатоцитах. Hu et al. Am. J. Pathol. 142: 1823-1830 (1993). После химического или механического повреждения печени уровни HGF резко возрастают, приводя к сильной пролиферации гепатоцитов. Horimoto et al. J. Hepatol. 23: 174-183 (1995). Печень трансгенных мышей со специфичной для печени сверхэкспрессией HGF в два раза больше по размеру, чем печень контрольных животных, и печень регенерирует намного быстрее после частичной гепатэктомии. Sakata et al. (1996) Cell Growth Differ. 7: 1513-1523; Shiota et al. (1994) Hepatol. 19: 962-972.
Ангиогенез является важным клеточным событием, при котором эндотелиальные клетки сосудов пролиферируют, мигрируют и подвергаются реорганизации, образуя новые сосуды, отходящие от уже существующей сосудистой сети. Существуют убедительные доказательства того, что развитие кровоснабжения является важным для нормальных и патологических пролиферативных процессов (Folkman and Klagsbrun (1987) Science 235: 442-447). Регенерирующая печень по аналогии с быстро растущими опухолями должна синтезировать новую строму и кровеносные сосуды. См., например, WO 03/103581; Yamane et al. Oncogene 9: 2683-2690 (1994); Mochida et al. Biochem. Biophy. Res. Comm. 226: 176-179 (1996); Ajioka et al. Hepatology 29: 396-402 (1999) и Assy et al. J. Hepatol. 30: 911-915 (1999), Michalopoulos and DeFrances (1997) выше; Mochida et al. (1996). В одном варианте осуществления изобретения ANGPTL4 или агонист ANGPTL4 вводят в комбинации с ангиогенным средством, например, VEGF или активаторами VEGFR. «Ангиогенным фактором или средством» является фактор роста, который стимулирует развитие кровеносных сосудов, например, стимулирует ангиогенез, рост эндотелиальных клеток, стабильность кровеносных сосудов и/или васкулогенез и т.д. Например, ангиогенные факторы включают без ограничения, например, VEGF и представителей семейства VEGF (A, B, C, D и E), PlGF, семейства PDGF, семейства факторов роста фибробластов (FGF), лиганды TIE (ангиопоэтины), ANGPTL3, эфрины и т.д. К ним также относятся факторы, которые ускоряют заживление ран, такие как гормон роста, инсулиноподобный фактор роста-I (IGF-I), VIGF, эпидермальный фактор роста (EGF), CTGF и представители его семейства и TGF-α и TGF-β. См., например, Klagsbrun and D'Amore, Annu. Rev. Physiol., 53: 217-39 (1991); Streit and Detmar, Oncogene, 22: 3172-3179 (2003); Ferrara and Alitalo, Nature Medicine 5(12): 1359-1364 (1999); Tonini et al., Oncogene, 22: 6549-6556 (2003) (например, таблицу 1, в которой перечислены известные ангиогенные факторы); и Sato Int. J. Clin. Oncol., 8: 200-206 (2003).
Гомеостаз липидов
ANGPTL4, кроме печени, вовлечен в модулирование других аспектов гомеостаза энергии. ANGPTL4 связан с дифференцировкой адипоцитов, системным метаболизмом липидов и ангиогенезом. См., например, Yoon et al., Molecular and Cellular Biology, 20(14): 5343-5349 (2000); Le Jan et al., American Journal of Pathology, 162(5): 1521-1528 (2003); и EP 1403367. Экспрессию ANGPTL4 также индуцируют PPAR-гамма и -альфа в жировой ткани, и она индуцируется голоданием. См., например, Yoon et al., Mol. Cell. Biol., 20: 5343-5349 (2000); и Kersten et al., J. Biol. Chem., 275: 28488-28493 (2000). Экспрессия ANGPTL4 подвергается повышающей регуляции во время голодания, и избыток белка в плазме снижается при потреблении пищи с высоким содержанием жиров.
Кроме того, ANGPTL4 ингибирует активность липопротеинлипазы (LPL). См., например, EР 1403367. Липопротеинлипаза (LPL) является секретируемым гликопротеином, который опосредует метаболизм липопротеинов посредством гидролиза триглицеридов, присутствующих в хиломикронах и липопротеинах очень низкой плотности (VLDL), с образованием свободных жирных кислот и фосфолипидов.
Как показано в данном описании, нокаутированные по ANGPTL4 мыши имеют пониженные уровни холестерина и триглицеридов в сыворотке по сравнению с однопометными животными дикого типа, совпадающие с ними по полу. См. разделы, озаглавленные «Трансгенные нокаутированные животные» и «пример 4» в данном описании. Кроме того, внутривенная инъекция ANGPTL4 увеличивает уровни липидов в циркулирующей плазме у мышей и увеличивает уровни липопротеинов очень низкой плотности. См., например, Yoshida et al., Journal of Lipid Research, 43: 1770-1772 (2002) и см. фиг.10.
В изобретении предлагаются способы модулирования уровней триглицеридов или холестерина в сыворотке субъекта. Например, способы включают в себя введение субъекту эффективного количества композиции, содержащей ANGPTL4, или агонист ANGPTL4, или антагонист ANGPTL4, таким образом модулируя уровни триглицеридов или холестерина в сыворотке субъекта. В одном варианте вводят ANGPTL4 или агонист ANGPTL4, который приводит к накоплению триглицеридов или холестерина в сыворотке. В другом варианте субъекту вводят эффективное количество антагониста ANGPTL4, таким образом снижая уровень по меньшей мере одного из компонентов: триглицеридов, свободных жирных кислот и/или холестерина в сыворотке субъекта по сравнению с субъектом до лечения или субъектом в отсутствие лечения или более низкого уровня лечения. Средние уровни холестерина и триглицеридов в сыворотке можно оценить способом, известным в данной области.
ANGPTL4 также может модулировать адипоциты. Например, ANGPTL4 может стимулировать пролиферацию преадипоцитов или индуцировать клеточную миграцию преадипоцитов. Жировая ткань главным образом состоит из адипоцитов, которые также играют ключевую роль в гомеостазе энергии. Адипоциты синтезируют и хранят липиды, когда питательных веществ имеется в изобилии, и высвобождают жирные кислоты в кровообращение, когда требуются питательные вещества. У позвоночных обнаружена белая жировая ткань (WAT) и бурая жировая ткань (BAT). WAT хранит и высвобождает жир в зависимости от потребностей животного в питательных веществах. Жир, который хранится в WAT, используется для (1) теплоизоляции (например, подкожный жир), (2) механической прокладки (например, окружающей внутренние органы) и (3) в качестве источника энергии. BAT сжигает жир, высвобождая энергию в виде тепла в ходе теплообразования для поддержания гомеотермии за счет увеличения теплообразования в ответ на пониженные температуры, и для поддержания энергетического баланса за счет увеличения потребления энергии в ответ на увеличение потребления калорий. См., например, Sears, I. B. et al. (1996) Mol. Cell. Biol. 16(7): 3410-3419 (1996). В общем, BAT уменьшается с возрастом, но может быть повторно активирован при некоторых состояниях, например, при длительном воздействии холода, поддержании диеты с высоким содержанием жира и при наличии опухолей, продуцирующих норадреналин.
В адипогенез вовлечены морфологические изменения, задержка роста, экспрессия липогенных ферментов, накопление липидов и приобретенная чувствительность к различным гормонам, например, инсулину. Предлагаются способы, которые включают в себя стимуляцию пролиферации адипоцитов посредством введения эффективного количества ANGPTL4 или агониста ANGPTL4. Пролиферацию клеток можно оценить в ходе культивирования, используя способы, известные в данной области, включая, но не ограничивая указанным, измерение скорости синтеза ДНК (см., например, Nakamura et al. Biochem. Biophy. Res. Comm. 122: 1450 (1984), подсчет в гемоцитометре с исключением при окрашивании трипановым синим (см., например, Omiri et al. (1997), выше) или проточную цитометрию (см., например, Drakes (1997), выше). ANGPTL4 или агонисты ANGPTL4 могут быть применимы для индукции пролиферации адипоцитов при расстройствах, когда может быть полезным дополнительное количество адипоцитов, например, включая, но не ограничивая указанным, например, истощающие заболевания (например, такие как различные типы злокачественной опухоли, пациенты с иммунной недостаточностью (например, пациенты, страдающие СПИД, и т.д.), пожилые люди) и т.д. Также предлагаются способы, которые включают в себя индукцию миграции преадипоцитов посредством введения эффективного количества ANGPTL4 или агониста ANGPTL4.
ANGPTL4 или агонисты ANGPTL4 также можно комбинировать с другими факторами, которые стимулируют дифференцировку адипоцитов. Такие факторы включают без ограничения, например, IGF-1, инсулин, глюкокортикоиды, 3,3',5-трийодтиронин, ретиноевую кислоту, PGF2α, PGI2 и т.д. Дополнительные факторы также можно комбинировать с ANGPTL4 или агонистом ANGPTL4. Например, адипогенез подвергается гормональной и транскрипционной регуляции. Например, адипогенез может быть опосредован каскадом транскрипционных факторов, включая, например, представителей семейства активируемого пролифераторами пероксисом рецептора (PPAR), например, (PPARα,γ), семейства CCAAT/энхансерсвязывающего белка (C/EBP) и семейства основной лейциновой молнии, имеющей структуру спираль-петля-спираль (bHLH), например, ADD1/SREBP1. См., например, Wu et al. Current Opin. Cell Biol. 11: 689-694 (1999); Rosen and Spiegelman Annu. Rev. Cell Dev. Biol. 16: 145-171 (2000); Gregoire et al., Physiological Reviews 78(3) (1998); и Kim and Spiegelman Genes Devel. 10: 1096-1107 (1996)). PPAR (действует в жировой ткани и стимулирует адипогенез и хранение липидов. См., например, Rosen et al., Annu. Rev. Cell Dev. Biol., 16: 145-171 (2000); Rosen et al., Mol. Cell. 4: 611-617 (1999); Ren et al., Genes Dev. 16: 27-32 (2002); Rosen et al., Genes Dev., 16: 22-26 (2002); и Fukumura et al., Circ. Res. 93: e88-e97 (2003). PPAR также опосредует уровни мРНК и белка липопротеинлипазы в адипоцитах и других клетках (см., например, Gbaguidi et al., FEBS Letters 512: 85-90 (2002)), и PPAR также вовлечены в злокачественную опухоль (см., например, Yoshimura et al., Int. J. Cancer 104: 597-602 (2003); и Kubota et al., Cancer Research 58: 3344-52 (1998)).
Однако часто рост и/или образование жировой ткани нежелателен. Например, ожирение обычно возникает в том случае, когда поступление энергии превышает потребление энергии, приводя к росту и/или образованию жировой ткани в результате гипертрофического и гиперпластического роста. Гипертрофическим ростом является увеличение размера адипоцитов, стимулированное накоплением липидов. Гиперпластический рост определяют как увеличение количества адипоцитов в жировой ткани.
Ожирение является хроническим заболеванием, которое весьма распространено в современном обществе и связано не только с клеймом позора в обществе, но также со сниженной продолжительностью жизни и многочисленными медицинскими проблемами, включая неблагоприятное психологическое развитие, репродуктивные расстройства, такие как поликистоз яичника, дерматологические расстройства, такие как инфекции, варикозные вены, акантокератодермия и экзема, непереносимость физической нагрузки, резистентность к инсулину, гипертонию, гиперхолестеринемию, желчнокаменную болезнь, остеоартрит, повреждение опорно-двигательного аппарата, тромбоэмболическое заболевание, злокачественную опухоль и коронарную болезнь сердца. Rissanen et al., British Medical Journal, 301: 835-837 (1990). Лечение ожирения заключается в использовании средств, подавляющих аппетит, и других индукторов потери массы, модификациях диеты и т.д., но подобно пациентам с резистентностью к инсулину большинство пациентов с ожирением со временем отказываются от исходной диеты, тем самым терпят неудачу в достижении идеальной массы тела. Антагонисты ANGPTL4 могут применяться для лечения ожирения и/или снижения общей массы тела у субъекта с использованием эффективного количества антагониста ANGPTL4. Ожирение может быть определено по BMI и/или определяющим ожирение характеристикам, которые известны в данной области и описаны в данной публикации. Например, лечение ожирения в общем относится к снижению BMI млекопитающего до уровня меньше примерно 25,9 и поддержания такой массы, по меньшей мере, в течение 6 месяцев. Лечение соответственно приводит к снижению потребления пищи и калорий у млекопитающих. Кроме того, лечение в данном контексте относится к профилактике появления ожирения, если лечение проводят до наступления состояния ожирения. Лечение включает в себя ингибирование и/или полное подавление липогенеза у млекопитающих с ожирением, т.е. избыточного накопления липидов в жировых клетках или накопления жировых клеток, который является одним из основных признаков ожирения у человека и животного, а также потерю общей массы тела. Снижение общей массы тела можно измерить, используя стандартные способы (например, весы). В одном варианте снижают ожирение (жир) у субъекта. Таким образом также можно лечить состояния, связанные с ожирением, например, сердечно-сосудистое заболевание, диабет и т.д.
ANGPTL4 также вовлечен в модулирование лептина, который является гормоном, получаемым из адипоцитов. Лептин, который структурно родственен цитокинам, действует на рецепторы, которые относятся к суперсемейству рецепторов цитокинов. См., например, Zhang F., et al., Nature 387: 206-209 (1997); Tartaglia L. A., et al., Cell 83: 1263-1271 (1995); и Lee G.-H., et al., Nature 379: 632-635 (1996). Лептин кодируется геном, затрагиваемым мутацией, приводящей к ожирению (ob) (Zhang F, et al., Nature 387: 206-209 (1997)). Длинная форма рецептора лептина кодируется геном, затрагиваемым диабетической мутацией (db) (Tartaglia L. A., et al., Cell 83: 1263-1271 (1995)). Рецептор лептина, который имеет несколько изоформ, наиболее близок трансдуцирующим сигнал субъединицам gp130 и LIFR, которые активируются такими цитокинами, как IL-6, LIF и CNTF, и рецепторам гормонов для такого гормона роста, как эритропоэтин. См., например, Tartaglia L. A., et al., выше. Отсутствие функционального лептина или его рецептора вызывает тяжелое ожирение. См., например, Zhang et al., выше; Lee et al., выше; и Chen H et al, Cell 84: 491-495 (1996). Известно, что лептин действует в некоторых областях головного мозга (например, в гипоталамусе), регулируя потребление пищи, расход энергии и нейроэндокринную функцию, например, показано, что он является ключевым регулятором запасания жиров, при этом уровни лептина увеличиваются с увеличением запасов жира. См., например, Zhang Y., et al., Nature 372: 425-432 (1994); Halaas J. L. et al., Science 269: 543-546 (1995); Campfield L. A., et al., Science 269: 546-549 (1995); и Pellymounter M. A., et al., Science 269: 540-543 (1995).
Также обнаружено, что лептин является ангиогенным фактором. См., например, Sierra-Honigmann et al., «Biological Action of Leptin as an Angiogenic Factor» Science 281: 1683-1686 (1998); и Bouloumie et al., Circ. Res. 83: 1059-1066 (1998). Рост жировой ткани зависит от неоваскуляризации. См., например, Rupnick et al., PNAS USA 99(16): 10730-10735 (2002). Лептин также играет роль в иммунитете. См., например, La Cava and Matarese, «The Weight of Leptin in Immunity» Nature Reviews 4: 371-379 (2004). Другие активности лептина включают модулирование размножения, модулирование гематопоэза, модулирование метаболизма глюкозы и модулирование провоспалительных иммунных ответов. См., например,

ANGPTL4 подвергается повышающей регуляции у мышей ob/ob (нокаутированные по лептину) и db/db (нокаутированные по рецептору лептина). Изобретение относится к способам модулирования лептина и/или активностей лептина посредством введения эффективного количества ANGPTL4, агониста ANGPTL4 или антагониста ANGPTL4. Уровни лептина можно анализировать с использованием стандартных способов, например, SDS-ПААГ, иммуноблотов и т.д.
ANGPTL4, агонисты ANGPTL4 и/или антагонисты ANGPTL4 можно применять для лечения заболеваний и расстройств, связанных с нарушениями гомеостаза липидов и метаболизмом жира, к которым относятся без ограничения, например, метаболические заболевания, такие как сердечные расстройства, сердечно-сосудистые, эндотелиальные или ангиогенные расстройства, дислипидемия, гипертония, атеросклероз, артериосклероз, болезнь коронарных артерий (CAD), коронарная болезнь сердца, гиперхолестеринемия, сердечная недостаточность, инсульт, диабет, дисфункции поджелудочной железы, остеоартрит, желчные камни, злокачественная опухоль, глаукома, ожирение, а также родственные расстройства, такие как тучность, расстройства пищевого поведения, синдромы истощения (кахексия), апноэ во сне и другие. Например, некоторые состояния человека характеризуются особенным составом липидов в тканях, клетках, мембранах и внеклеточных областях или структурах. Например, при атеросклерозе холестерин (неэтерифицированные, этерифицированные и окисленные формы) и другие липиды накапливаются в клетках и во внеклеточных областях стенки артерий и в других местах. Такие липиды оказывают потенциально опасное биологическое действие, например, изменяя клеточные функции, в том числе экспрессию генов, и сужая просвет сосудов, препятствуя потоку крови. Регулирование уровней липидов может обеспечивать многочисленные существенные полезные эффекты. Влияние введения ANGPTL4, агониста или антагониста можно также измерить с помощью множества анализов, известных в данной области, включая анализ жировых клеток и ткани, например, жировых тел, общей массы тела, уровней триглицеридов в мышце, печени и жировой ткани, уровней лептина натощак и не натощак и уровней свободных жирных кислот и триглицеридов в крови. Агонист ANGPTL4 также можно применять для ингибирования миграции преадипоцитов посредством введения эффективного количества антагониста ANGPTL4.
В некоторых аспектах изобретения требуется комбинирование терапевтических средств на основе ANGPTL4, агониста ANGPTL4 или антагониста ANGPTL4 с другими терапевтическими схемами. ANGPTL4 или агонисты ANGPTL4 можно комбинировать с введением других факторов, например, таких как факторы, описанные в данной публикации. Что касается антагонистов, то антагонисты ANGPTL4 можно комбинировать с введением, например, терапевтических средств для лечения гиперлипидемии (и заболеваний, связанных с гиперлипидемией, например, ожирения, гиперхолестеринемии, атеросклероза, сердечно-сосудистого заболевания, сахарного диабета, гипотиреоидизма, синдрома Кушинга), например, включая, но не ограничивая указанным, ниацин, холестирамин, колестипол, гемфиброзил, клофибрат, статины, флувастатин (лескол), правастатин, симвастатин, розувастатин кальция (ZD-4522), питавастатин (NK104), премарин/правахол (эстроген/правастатин), эзетимб/симвастатин, суперстатин, липитор, вакцина CETi-1, антитела против CETP (белок-переносчик сложного эфира холестерина), BMS-201038 (микросомальный белок-переносчик триглицеридов), FM-VP4 (ингибитор транспорта холестерина), фиостанол, гипогликемические средства, инсулин, прамлинтид, амилин, AC2993 синтетический эксендин-4, ксеникал (орлистат), цилиарный нейротрофический фактор, аксокин, метформин XT, метформин, глюкованс (метформин/глибурид), декслипотам (R+/- альфа-липоевая кислота), агонисты PPAR, агонисты бета-3-адренергических рецепторов, ингибиторы липазы, ATL-962, лептин, анорексигенные препараты или средства, подавляющие аппетит, фентермин, меридиа (силбутрамин), веллбутрин (бупроион), проциглем (диазоксид), тенуат (диэтилпропион), ревиа (налтрексон), бонтрил (фендиметразин), золофт (сертралин), цилиарный нейротрофический фактор (CNTF), аксокин, антагонисты CB1-каннабиноидного рецептора, SR 141716, фитофарм, AOD9604, hGH 177-191, средство для потери массы и его производные (например, соли, пэгилированные варианты и т.д.). См. также WO96/04260 (соединения для лечения диабета типа II), WO94/01420, WO95/17394, WO97/36579, WO97/25042, WO99/08501, WO99/19313 и WO99/16758. Изменения образа жизни также можно комбинировать с терапевтическими средствами согласно изобретению. Изменения образа жизни включают без ограничения, например, диету, физические нагрузки, ограниченное потребление холестерина, прекращение курения и т.д. См. также WO91/19702 (гипогликемические и гипохолестеринемические средства). В некоторых аспектах антагонисты ANGPTL4 можно комбинировать, например, с цитокинами и другими провоспалительными молекулами и некоторыми факторами роста, которые ингибируют адипогенез. Такие факторы включают без ограничения, например, фактор некроза опухолей (TNF)-α, IL-1, PDGF, FGF, EGF, трансформирующий фактор роста (TGF)-α, -β, фактор преадипоцитов-1 (pref-1) и т.д. См., например, Gregoire et al., Physiological Reviews 78(3): 783-809 (1998).
Термин «агент для потери массы» относится к молекуле, применимой для лечения или профилактики ожирения. Такие молекулы включают, например, гормоны (катехоламины, глюкагон, ACTH и гормон роста в комбинации с IGF-1; белок Ob; клофибрат; галогенат; цинхокаин; хлорпромазин; лекарственные средства, подавляющие аппетит, действующие на норадренергические нейромедиаторы, такие как мазиндол и производные фенэтиламина, например, фенилпропаноламин, диэтилпропион, фентермин, фендиметразин, бензфетамин, амфетамин, метамфетамин и фенметразин; лекарственные средства, действующие на нейромедиаторы серотонина, такие как фенфлурамин, триптофан, 5-гидрокситриптофан, флуоксетин и сертралин; центрально действующие лекарственные средства, такие как налоксон, нейропептид-Y, галанин, кортикотропин-рилизинг гормон и холецистокинин; холинергический агонист, такой как пиридостигмин; сфинголипид, такой как лизосфинголипид или его производное; термогенные лекарственные средства, такие как тиреоидный гормон; эфедрин; бета-адренергические агонисты; лекарственные средства, влияющие на желудочно-кишечный тракт, такие как ингибиторы ферментов, например, тетрагидролипостатин, неперевариваемые продукты питания, такие как сложный полиэфир сахарозы, и ингибиторы опорожнения желудка, такие как треохлорлимонная кислота или его производные; β-адренергические агонисты, такие как изопротеренол и йохимбин; аминофиллин для повышения эффектов йохимбина, подобных бета-адренергическим эффектам, лекарственное средство, блокирующее α2-адренергические рецепторы, такие как клонидин отдельно или в комбинации с пептидом, высвобождающим гормон роста; лекарственные средства, которые препятствуют всасыванию в кишечнике, такие как бигуаниды, например, метформин и фенформин; наполнители, такие как метилцеллюлоза; лекарственные средства, блокирующие метаболизм, такие как гидроксицитрат; прогестерон; агонисты холецистокинина; малые молекулы, которые имитируют кетокислоты; агонисты кортикотропин-рилизинг гормона; родственное алкалоидам спорыньи ингибирующее пролактин соединение для снижения жировых запасов в организме (патент США №4783469, опубликованный 8 ноября 1988); бета-3-агонисты; бромкриптин; антагонисты опиоидных пептидов; антагонисты нейропептида Y; антагонисты рецепторов глюкокортикоидов; агонисты гормона роста; их комбинации и т.д.
Другие применения
По-видимому, ANGPTL4 также является негативным регулятором воспалительных ответов. В некоторых вариантах согласно изобретению ANGPTL4 или агонисты ANGPTL4 могут быть использованы для ингибирования иммунного ответа, например, в случае нежелательного или опасного иммунного ответа, например, при отторжении трансплантата или заболевании «трансплантат против хозяина». Антагонисты ANGPTL4 могут быть применимы для стимуляции иммунной системы. Например, стимуляция иммунной системы может быть желательна при лейкозе, других типах злокачественной опухоли, у пациентов с иммунной недостаточностью (например, пациентов, страдающих СПИД и т.д.) и т.д.
ANGPTL4 также вовлечен в развитие злокачественной опухоли. ANGPTL4 при экспрессии в опухолевых клетках вызывает пролиферацию опухолевых клеток in vitro и in vivo (см. предварительную заявку 60/589782 и с номером в реестре поверенного P2144R1, поданную одновременно с настоящей заявкой, которая включена в виде ссылки для всех целей). Когда ANGPTL4 экспрессируется в опухолях, подвергаемых лечению фактором против ангиогенеза, например анти-VEGF-антителом, то опухоль по-прежнему сохраняет способность к росту. Также было показано, что ANGPTL4 подвергается повышающей регуляции в злокачественных опухолях почек. См., например, документ с номером в реестре поверенного P5032R1; WO 02/07941 и Le Jan et al., American Journal of Pathology, 162(5): 1521-1528 (2003). Кроме того, ANGPTL4 является проангиогенным фактором (см., например, S. Le Jan et al., Am. J. Pathol., 162(5): 1521-1528 (2003)), который представляет собой мишень для терапии злокачественной опухоли, и является фактором выживания при апоптозе для эндотелиальных клеток (см., например, Kim et al., Biochem K. 346: 603-610 (2000). Подобно VEGF (Shweiki et al., Proc. Natl. Acad. Sci, USA 92: 768-772 (1995) экспрессия ANGPTL4 возрастает в ответ на гипоксию. См., например, Le Jan et al., American Journal of Pathology, 162(5): 1521-1528 (2003). Исследователи сообщали о связях между ангиогенезом и адипогенезом или ростом жировой ткани. См., например,
Sierra-Honigmann et al., "Biological Action of Leptin as an Angiogenic Factor" Science 281:1683-1686 (1998); Rupnick et al., "Adipose tissue mass can be regulated through the vasculature" Proc. Nat. Acad. Sci. USA, 99(16):10730-10735 (2002); Kolonin et al., "Reversal of obesity by targeted ablation of adipose tissue" Nature Medicine Advance Online publication May 9, 2004: 1-8; and, Fukumura et al., "Paracrine Regulation of Angiogenesis and Adipocyte Differentiation During In Vivo Adipogenesis." Circ. Res. 93:e88-e97 (2003).
ANGPTL4 также можно использовать в диагностических анализах. Множество различных анализов и форм анализов можно использовать для выявления количества ANGPTL4 в образце по сравнению с контрольным образцом. Такие формы, в свою очередь, применимы в диагностических анализах согласно изобретению, которые используются для выявления наличия или появления у субъекта расстройств, описанных в данной публикации.
Любой способ, известный в данной области для измерения растворимых анализируемых веществ, можно применить при практическом осуществлении настоящего изобретения. Такие способы включают без ограничения системы конкурентного и неконкурентного анализа с использованием таких методик, как радиоиммуноанализ, ферментный иммуноанализ (EIA), например, ELISA, иммуноанализы типа «сэндвич», реакции с использованием преципитинов, реакции диффузии в геле, иммунодиффузионные анализы, анализы фиксации комплемента, иммунорадиометрические анализы, флуоресцентные иммуноанализы, иммуноанализы белка A и иммуноэлектрофоретические анализы. См., например, патенты США №4845026 и 5006459.
Трансгенные по ANGPTL4 нокаутированные животные
Нуклеиновые кислоты, которые кодируют ANGPTL4 или его модифицированные формы, также могут быть использованы для создания либо трансгенных животных, либо «нокаутированных» животных, которые, в свою очередь, применимы для разработки и скрининга терапевтически полезных реагентов. Трансгенное животное (например, мышь или крыса) представляет собой животное, имеющее клетки, которые содержат трансген, и при этом указанный трансген был введен в животное или предка животного в пренатальной, например, эмбриональной стадии. Трансген представляет собой ДНК, которую интегрируют в геном клетки, из которой развивается трансгенное животное. Изобретение относится к кДНК, кодирующей ANGPTL4, которая может быть использована для клонирования геномной ДНК, кодирующей ANGPTL4, согласно разработанным методикам, и геномным последовательностям, применяемым для создания трансгенных животных, которые содержат клетки, которые экспрессируют ДНК, кодирующую ANGPTL4.
Любой способ, известный в данной области, можно использовать для введения трансгена гена-мишени животным, чтобы получить родоначальные линии трансгенных животных. Такие способы включают, без ограничения, микроинъекцию в пронуклеус (патенты США №4873191, 4736866 и 4870009); опосредованный ретровирусами перенос генов в зародышевые линии (Van der Putten, et al., Proc. Natl. Acad. Sci., USA, 82: 6148-6152 (1985)); целенаправленное изменение генов в эмбриональных стволовых клетках (Thompson, et al., Cell, 56: 313-321 (1989)); неспецифичную инсерционную инактивацию с использованием вектора генной ловушки (патент США №6436707); электропорацию эмбрионов (Lo, Mol. Cell. Biol., 3: 1803-1814 (1983)) и опосредованный сперматозоидами перенос генов (Lavitrano, et al., Cell, 57: 717-723 (1989)) и т.д.
Обычно конкретные клетки могут быть целенаправленно изменены в направлении включения трансгена ANGPTL4 с использованием тканеспецифичных энхансеров. Трансгенных животных, которые содержат копию трансгена, кодирующего ANGPTL4, введенную в зародышевую линию животного на эмбриональной стадии, можно использовать для исследования влияния повышенной экспрессии ДНК, кодирующей полипептиды ANGPTL4. Таких животных можно использовать в качестве животных для испытаний реагентов, которые предположительно обеспечивают защиту, например, от патологических состояний, связанных с их сверхэкспрессией. Согласно указанному аспекту изобретения животного подвергают обработке реагентом, и сниженная частота возникновения патологического состояния по сравнению с необработанными животными, несущими трансген, будет свидетельствовать о потенциальном терапевтическом воздействии на патологическое состояние.
Альтернативно можно использовать гомологи ANGPTL4 животного, отличного от человека, для конструирования «нокаутированного» по ANGPTL4 животного, которое имеет дефектный или измененный ген, кодирующий белок ANGPTL4, в результате гомологичной рекомбинации между эндогенным геном, кодирующим ANGPTL4, и измененной геномной ДНК, кодирующей ANGPTL4, введенной в эмбриональную стволовую клетку животного. В некоторых вариантах нокаутированным животным является млекопитающее, например, грызун, такой как крыса или мышь. Например, кДНК, кодирующую ANGPTL4, можно использовать для клонирования геномной ДНК, кодирующей ANGPTL4, согласно разработанным способам. Часть геномной ДНК, кодирующей ANGPTL4, можно делетировать или заменить другим геном, таким как ген, кодирующий маркер селекции, который можно использовать для контроля интегрирования. Обычно в вектор включают несколько тысяч пар нуклеотидов неизмененной фланкирующей ДНК (как на 5'-, так и на 3'-конце) (см., например, Thomas and Capecchi, Cell, 51: 503 (1987) в отношении описания векторов для гомологичной рекомбинации).
Вектор вводят в линию эмбриональных стволовых клеток (например, электропорацией) и отбирают клетки, в которых введенная ДНК гомологично рекомбинировала с эндогенной ДНК (см., например, Li et al., Cell, 69: 915 (1992)). Затем отобранные клетки инъецируют в бластоцисту животного (например, мыши или крысы) для образования агрегационных химер (см., например, Bradley, in Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), pp.113-152). Затем химерный эмбрион может быть имплантирован подходящей псевдобеременной самке приемного животного и эмбрион доведен до родов для создания «нокаутированного» животного. Потомство, несущее гомологично рекомбинированную ДНК в своих зародышевых клетках, можно идентифицировать стандартными способами и использовать для разведения животных, у которых все клетки животного содержат гомологично рекомбинированную ДНК. Нокаутированных животных можно охарактеризовать, например, по их способности защищаться от некоторых патологических состояний и по развитию у них патологических состояний вследствие отсутствия гена, кодирующего ANGPTL4.
Кроме того, нокаутированные мыши могут быть высоко информативными в отношении раскрытия функции генов и фармацевтической применимости исследуемого лекарственного средства, а также при определении возможных точных побочных эффектов, связанных с данным исследуемым лекарственным средством. Функция генов и физиология являются высоко консервативными при сравнении мыши и человека, так как они оба являются млекопитающими и содержат сходные количества генов, которые являются высоко консервативными у указанных видов. Недавно было хорошо подтверждено документально, что 98% генов хромосомы 16 мыши имеют ортолог человека (Mural et al., Science 296: 1661-71 (2002)).
Хотя целенаправленное изменение генов в эмбриональных стволовых клетках (ES) сделало возможным конструирование мышей с нуль-мутациями во многих генах, связанных с заболеванием человека, не все генетические заболевания можно отнести к нуль-мутациям. Можно сконструировать полезные мышиные модели заболеваний человека с помощью разработки способа замены генов (knock-in), которая будет нарушать мышиный локус и вводить человеческую копию с мутацией. Затем можно проводить исследования лекарственных средств in vivo, целью которых является белок человека (Kitamoto et al., Biochemical and Biophysical Res. Commun., 222: 742-47 (1996)).
Применения трансгенных животных
В некоторых вариантах изобретение относится к способам скрининга соединений для идентификации соединений, которые имитируют ANGPTL4 (агонисты) или предотвращают действие ANGPTL4 (антагонисты). Агонисты, которые имитируют ANGPTL4, могут быть особенно полезными с терапевтической точки зрения для индукции активностей ANGPTL4, например, таких, которые описаны в данной публикации, и в тех случаях, когда наблюдают негативный фенотип на основе данных, полученных с использованием трансгенного животного, отличного от человека, геном которого содержит нарушение гена, который кодирует ANGPTL4. Антагонисты, которые предотвращают действие ANGPTL4, могут быть особенно полезны с терапевтической точки зрения для предотвращения активностей ANGPTL4, например, таких, которые описаны в данной публикации, и в тех случаях, когда наблюдают позитивный фенотип на основе наблюдений с использованием трансгенного нокаутированного животного, отличного от человека. Разработаны скрининговые анализы предполагаемых антагонистических лекарственных средств для идентификации соединений, которые связываются или образуют комплекс с ANGPTL4, кодируемым генами, идентифицированными в данном описании, или иным образом мешают взаимодействию кодируемого полипептида с другими клеточными белками, например, с рецептором ANGPTL4 (например, αVβ5), белком липолидазой и т.д.
Например, влияние антагониста ANGPTL4 можно оценить при введении антагониста ANGPTL4 мыши дикого типа для того, чтобы имитировать известный нокаутированный фенотип.Таким образом, сначала можно нокаутировать представляющий интерес ген ANGPTL4 и наблюдать полученный в результате фенотип как следствие нокаута или нарушения гена ANGPTL4. Затем можно оценить эффективность антагониста ANGPTL4 посредством введения антагониста ANGPTL4 мыши дикого типа. Можно ожидать, что эффективный антагонист будет имитировать фенотипический эффект, который сначала наблюдали у нокаутированного животного.
Подобным образом можно оценить влияние агониста ANGPTL4 посредством введения агониста ANGPTL4 трансгенному животному, отличному от человека, например мыши, чтобы улучшить известный негативный нокаутированный фенотип. Соответственно можно сначала нокаутировать представляющий интерес ген ANGPTL4 и наблюдать полученный в результате фенотип как следствие нокаута или нарушения гена ANGPTL4. Затем можно оценить эффективность агониста ANGPTL4 посредством введения агониста ANGPTL4 трансгенному животному, отличному от человека, например мыши. Можно ожидать, что эффективный агонист будет улучшать негативный фенотипический эффект, который сначала наблюдали у нокаутированного животного.
В другом анализе антагонистов клетки млекопитающих или препараты мембран, экспрессирующие рецептор, можно инкубировать с меченым ANGPTL4 в присутствии выбранного для исследования соединения. Затем можно измерить способность соединения усиливать или блокировать такое взаимодействие.
Антитела
Антитела согласно изобретению включают анти-ANGPTL4-антитела или антигенсвязывающие фрагменты для ANGPTL4, анти-αVβ5-антитела или другие антитела, описанные в данной публикации. Примеры антител включают, например, поликлональные, моноклональное, гуманизированные антитела, фрагменты антител, полиспецифичные, гетероконъюгированные, поливалентные, обладающие эффекторной функцией и т.д. антитела. Антитела могут быть агонистами или антагонистами.
Поликлональные антитела
К антителам согласно изобретению могут относиться поликлональные антитела. Способы получения поликлональных антител известны специалисту в данной области. Например, поликлональные антитела против ANGPTL4 вырабатываются у животных при одной или нескольких подкожных (п/к) или внутрибрюшинных (в/б) инъекциях соответствующего антигена и адъюванта. Может быть полезным конъюгирование соответствующего антигена с белком, который является иммуногенным для иммунизируемого вида, например, с гемоцианином лимфы улитки, сывороточным альбумином, бычьим тиреоглобулином или ингибитором трипсина сои, с использованием бифункционального или дериватизирующего агента, например, сложного эфира малеимидобензоилсульфосукцинимида (конъюгирование через остатки цистеина), N-гидроксисукцинимида (через остатки лизина), глутаральдегида, янтарного ангидрида, SOCl2 или R1N=C=NR, где R и R1 означают разные алкильные группы.
Животных иммунизируют ANGPTL4, иммуногенными конъюгатами или производными, комбинируя, например, 100 мкг или 5 мкг белка или конъюгата (для кроликов или мышей, соответственно) с 3 объемами полного адъюванта Фрейнда и инъецируя раствор внутрикожно в нескольких местах. Спустя месяц животных подвергают бустер-иммунизации, используя от 1/5 до 1/10 исходного количества пептида или конъюгата в полном адъюванте Фрейнда посредством подкожной инъекции в нескольких местах. Через 7-14 дней у животных берут кровь и анализируют сыворотку в отношении титра антител. Животных подвергают бустер-иммунизации вплоть до выхода титра на плато. Обычно животного подвергают бустер-иммунизации конъюгатом того же самого антигена, но конъюгированного с другим белком и/или с помощью другого поперечно сшивающего реагента. Конъюгаты также могут быть получены в культуре рекомбинантных клеток в виде слияний белков. Также соответствующим образом используют агрегирующие агенты, такие как квасцы, для усиления иммунного ответа.
Моноклональные антитела
Моноклональные антитела могут быть получены с использованием способа на основе гибридом, впервые описанного Kohler et al., Nature, 256: 495 (1975), или могут быть получены способами на основе рекомбинантной ДНК (патент США №4816567).
В способе на основе гибридом мышь или другое подходящее животное-хозяин, такое как хомячок или макака, иммунизируют, как описано выше, чтобы вызвать появление лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфично связываться с белком, используемым для иммунизации. Альтернативно лимфоциты можно иммунизировать in vitro. Затем лимфоциты сливают с клетками миеломы, используя подходящий агент для слияния, такой как полиэтиленгликоль, чтобы образовать гибридомную клетку (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)).
Полученные таким образом клетки гибридомы высевают и выращивают в подходящей культуральной среде, которая обычно содержит одно или несколько веществ, которые ингибируют рост или жизнеспособность неслитых исходных клеток миеломы. Например, если в исходных клетках миеломы отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), то в культуральную среду для гибридом обычно будут включать гипоксантин, аминоптерин и тимидин (среда HAT), и такие вещества предотвращают рост HGPRT-дефицитных клеток.
Обычные клетки миеломы представляют собой клетки, которые эффективно сливаются, поддерживают на стабильно высоком уровне продукцию антител отобранными продуцирующими антитела клетками и чувствительны к такой среде, как среда HAT. Среди них предпочтительными линиями клеток миеломы являются линии клеток миеломы мышей, такие как линии, полученные из опухолей мышей MOPC-21 и MPC-11, доступные из Salk Institute Cell Distribution Center, San Diego, California USA, и клетки SP-2 или X63-Ag8-653, доступные из Американской коллекции типов культур, Rockville, Maryland USA. Линии клеток миеломы человека и гетеромиеломы мыши-человека также были описаны для получения моноклональных антител человека (Kozbor, J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, в которой выращивают клетки гибридомы, анализируют в отношении продукции моноклональных антител, направленных против ANGPTL4. Специфичность связывания моноклональных антител, продуцируемых клетками гибридомы, можно определить с помощью иммунопреципитации или анализом связывания in vitro, таким как радиоиммуноанализ (РИА) или твердофазный иммуноферментный анализ (ELISA). Такие способы и анализы известны в данной области. Аффинность связывания моноклонального антитела, например, можно определить анализом по Скэтчарду, как описано в Munson and Pollard, Anal. Biochem., 107: 220 (1980).
После того как идентифицировано, что клетки гибридомы продуцируют антитела требуемой специфичности, аффинности и/или активности, клоны могут быть субклонированы способами лимитирующего разведения и выращены стандартными способами (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)). Подходящие культуральные среды для указанной цели включают, например, среду D-MEM или RPMI-1640. Кроме того, клетки гибридомы могут быть выращены in vivo в виде асцитных опухолей у животного.
Моноклональные антитела, секретируемые подклонами, соответствующим образом отделяют от культуральной среды, асцитной жидкости или сыворотки обычными способами очистки иммуноглобулинов, такими как, например, хроматография с использованием белка A-сефарозы, гидроксилапатита, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела также могут быть получены способами на основе рекомбинантной ДНК, такими как способы, описанные в патенте США №4816567. ДНК, кодирующую моноклональные антитела, легко выделяют и секвенируют, используя обычные способы (например, используя олигонуклеотидные зонды, которые способны специфично связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител). Клетки гибридомы служат в качестве источника такой ДНК. После выделения ДНК может быть помещена в экспрессирующие векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки E.coli, клетки COS обезьян, клетки яичника китайского хомячка (CHO) или клетки миеломы, которые в ином случае не продуцируют белок иммуноглобулина, чтобы получить синтез моноклональных антител в рекомбинантных клетках-хозяевах. Рекомбинантная продукция антител будет более подробно описана ниже.
В другом варианте антитела или фрагменты антител могут быть выделены из фаговых библиотек антител, созданных с использованием способа, описанного в McCafferty et al., Nature, 348: 552-554 (1990). В Clackson et al., Nature, 352: 624-628 (1991) и Marks et al., J. Mol. Biol., 222: 581-597 (1991) описано выделение мышиных и человеческих антител, соответственно, с использованием фаговых библиотек. В последующих публикациях описано получение высокоаффинных (нМ-диапазон) антител человека с помощью перетасовки цепей (Marks et al., Bio/Technology, 10: 779-783 (1992)), а также комбинаторной инфекции и рекомбинации in vivo в качестве методики конструирования очень больших фаговых библиотек (Waterhouse et al., Nuc. Acids. Res., 21: 2265-2266 (1993)). Таким образом, указанные способы являются приемлемой альтернативой традиционным способам на основе продуцирующих моноклональные антитела гибридом для выделения моноклональных антител.
ДНК также может быть модифицирована, например, подстановкой кодирующей последовательности константных доменов тяжелой и легкой цепей человека вместо гомологичных мышиных последовательностей (патент США №4816567; Morrison, et al., Proc. Natl. Acad. Sci. USA, 81: 6851 (1984)) или ковалентным связыванием с кодирующей иммуноглобулин последовательностью всей или части последовательности, кодирующей неиммуноглобулиновый полипептид.
Обычно такими неиммуноглобулиновыми полипептидами заменяют константные домены антитела или ими заменяют вариабельные домены одного антигенсвязывающего участка антитела, чтобы создать химерное бивалентное антитело, содержащее один антигенсвязывающий участок, имеющий специфичность по отношению к антигену, и другой антигенсвязывающий участок, имеющий специфичность по отношению к другому антигену.
Гуманизированные и человеческие антитела
Антитела согласно изобретению могут включать гуманизированные антитела или человеческие антитела. Гуманизированное антитело имеет один или несколько аминокислотных остатков, введенных в него из источника, который отличен от человека. Указанные аминокислотные остатки животного, отличного от человека, часто называют «импортируемыми» остатками, которые обычно берут из «импортируемого» вариабельного домена. Гуманизацию можно по существу выполнить, следуя способу Винтера и соавторов (Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-327 (1988); Verhoeyen et al., Science, 239: 1534-1536 (1988)), заменой CDR или последовательностями CDR грызунов соответствующих последовательностей антитела человека. Соответственно такие «гуманизированные» антитела являются химерными антителами (патент США №4816567), в которых значительно меньшая часть, чем интактный вариабельный домен человека, была заменена соответствующей последовательностью вида, отличного от человека. На практике гуманизированные антитела обычно представляют собой антитела человека, в которых некоторые остатки CDR и возможно некоторые остатки FR заменены остатками из аналогичных участков антител грызунов.
Выбор вариабельных доменов человека как легкой, так и тяжелой цепи, используемых для получения гуманизированных антител, очень важен для снижения антигенности. Согласно так называемому способу «оптимальной подгонки» последовательность вариабельного домена антитела грызуна подвергают скринингу по сравнению с полной библиотекой известных последовательностей вариабельных доменов человека. Последовательность человека, которая наиболее близка последовательности грызуна, затем берут в качестве каркаса (FR) человека для гуманизированного антитела (Sims et al., J. Immunol., 151: 2296 (1993); Chothia et al., J. Mol. Biol., 196: 901 (1987)). В другом способе используют конкретный каркас, полученный из консенсусной последовательности всех антител человека конкретной подгруппы легких или тяжелых цепей. Один и тот же каркас можно использовать для нескольких разных гуманизированных антител (Carter et al., Proc. Natl. Acad. Sci. USA, 89: 4285 (1992); Presta et al., J. Immunol., 151: 2623 (1993)).
Кроме того, важно, чтобы антитела были гуманизированы с сохранением высокой аффинности по отношению к антигену и других благоприятных биологических свойств. Для достижения указанной цели согласно обычному способу гуманизированные антитела получают, проводя анализ исходных последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов общедоступны и известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и показывают возможные трехмерные конформационные структуры выбранных для исследования последовательностей иммуноглобулина. Исследование таких изображений позволяет проанализировать вероятную роль остатков в функционировании выбранной для исследования последовательности иммуноглобулина, т.е. анализировать остатки, которые влияют на способность выбранного для исследования иммуноглобулина связывать соответствующий ему антиген. Таким образом могут быть выбраны и объединены остатки FR из реципиентных и импортируемых последовательностей так, чтобы достичь требуемых характеристик антитела, таких как повышенная аффинность по отношению к антигену-мишени. В общем, остатки CDR непосредственно и в наибольшей степени влияют на связывание антигена.
Альтернативно в настоящее время можно получать трансгенных животных (например, мышей), которые способны после иммунизации продуцировать полный репертуар антител человека в отсутствие продукции эндогенных иммуноглобулинов. Например, описано, что гомозиготная делеция гена соединяющей области тяжелой цепи (JH) антитела у химерных и мутантных по зародышевой линии мышей приводит к полному ингибированию продукции эндогенных антител. Перенос ряда генов иммуноглобулина зародышевой линии человека в таких мутантных по зародышевой линии мышей будет приводить к продукции антител человека при антигенной стимуляции. См., например, Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551 (1993); Jakobovits et al., Nature, 362: 255-258 (1993); Bruggermann et al., Year in Immuno., 7: 33 (1993); и Duchosal et al. Nature 355: 258 (1992). Антитела человека также могут быть получены из библиотек в фаговом дисплее (Hoogenboom et al., J. Mol. Biol., 227: 381 (1991); Marks et al., J. Mol. Biol., 222: 581-597 (1991); Vaughan et al. Nature Biotech 14: 309 (1996)).
Антитела человека также могут быть получены с использованием различных способов, известных в данной области, включая библиотеки в фаговом дисплее (Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222: 581 (1991)). Согласно указанному способу гены V-доменов антител клонируют в рамке с геном либо основного, либо минорного белка оболочки нитчатого бактериофага, такого как M13 или fd, и выводят в виде функциональных фрагментов антител на поверхность фаговой частицы. Поскольку нитчатая частица содержит однонитевую копию ДНК фагового генома, то селекция на основе функциональных свойств антитела также приводит к селекции гена, кодирующего антитело, проявляющее такие свойства. Таким образом, фаг имитирует некоторые свойства B-клетки. Фаговый дисплей может быть выполнен в разных формах, обзор которых приведен, например, в Johnson, K. S. and Chiswell, D J., Cur. Opin. in Struct. Biol. 3: 564-571 (1993). Для фагового дисплея можно использовать несколько источников участков V-генов. Например, Clackson et al., Nature, 352: 624-628 (1991) выделили ряд разнообразных антител против оксазолона из небольшой случайной комбинаторной библиотеки V-генов, полученной из селезенок иммунизированных мышей. Можно сконструировать репертуар V-генов неиммунизированных людей-доноров и можно выделить антитела к ряду разнообразных антигенов (включая аутоантигены) по существу, следуя, например, способу, описанному Marks et al., J. Mol. Biol. 222: 581-597 (1991) или Griffith et al., EMBO J. 12: 725-734 (1993). См. также патенты США №5565332 и 5573905. Способы Cole et al. и Boerner et al. также доступны для получения человеческих моноклональных антител (Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985) и Boerner et al., J. Immunol., 147(1): 86-95 (1991)). Антитела человека также могут быть образованы активированными in vitro B-клетками (см. патенты США 5567610 и 5229275).
Фрагменты антител
Фрагменты антител также включены в объем изобретения. Разработаны различные способы получения фрагментов антител. Традиционно такие фрагменты получали в результате протеолитического расщепления интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24: 107-117 (1992) и Brennan et al., Science, 229: 81 (1985)). Однако в настоящее время такие фрагменты могут непосредственно продуцироваться рекомбинантными клетками-хозяевами. Например, фрагменты антител могут быть выделены из фаговых библиотек антител, обсуждаемых выше. Альтернативно Fab'-SH-фрагменты могут быть непосредственно извлечены из клеток E.coli и химически связаны с образованием F(ab')2-фрагментов (Carter et al., Bio/Technology 10:163-167 (1992)). Согласно другому способу F(ab')2-фрагменты могут быть выделены непосредственно из культуры рекомбинантных клеток-хозяев. Другие способы получения фрагментов антител будут известны специалистам в данной области. В других вариантах предпочтительное антитело представляет собой одноцепочечный Fv-фрагмент (scFv). См. WO 93/16185, патент США №5571894 и патент США №5587458. Fv и sFv являются единственными видами с интактными связывающими участками, которые лишены константных областей; таким образом, они являются подходящими с точки зрения пониженного неспецифичного связывания в ходе применения in vivo. Могут быть сконструированы слитые белки SFv, чтобы получить слияние эффекторного белка либо с амино-, либо с карбоксильным концом sFv. См. Antibody Engineering, ed. Borrebaeck, выше. Фрагмент антитела также может представлять собой «линейное антитело», например, такое, как описано в патенте США 5641870. Такие линейные фрагменты антител могут быть моноспецифичными или биспецифичными.
Полиспецифичные антитела (например, биспецифичные)
Антитела согласно изобретению также включают, например, полиспецифичные антитела, которые обладают специфичностями связывания, по меньшей мере, по отношению к двум разным антигенам. Несмотря на то, что такие молекулы, как правило, будут связывать только два антигена (т.е. биспецифичные антитела, BsAb), антитела с дополнительными специфичностями, такие как триспецифичные антитела, входят в указанное определение при его использовании в данном описании. Примеры BsAb включают антитела с одним плечом, направленным против клеточного антигена, и другим плечом, направленным против запускающей цитотоксической молекулы, такие как анти-FcγRI/анти-CD15, анти-p185HER2/FcγRIII (CD16), анти-CD3/антитело против злокачественной B-клетки (1D10), анти-CD3/анти-p185HER2, анти-CD3/анти-p97, анти-CD3/антитело против почечно-клеточной карциномы, анти-CD3/анти-OVCAR-3, анти-CD3/L-D1 (антитело против карциномы ободочной кишки), анти-CD3/антитело против аналога меланоцитостимулирующего гормона, антитело против рецептора EGF/анти-CD3, анти-CD3/анти-CAMA1, анти-CD3/анти-CD19, анти-CD3/MoV18, антитело против молекулы адгезии нервных клеток (NCAM)/анти-CD3, антитело против связывающего фолат белка (FBP)/анти-CD3, антитело против пан-антигена, связанного с карциномой (AMOC-31)/анти-CD3; BsAb с одним плечом, которое специфично связывается с антигеном на клетке, и одним плечом, которое связывается с токсином, такие как анти-сапорин/анти-Id-1, анти-CD22/анти-сапорин, анти-CD7/анти-сапорин, анти-CD38/анти-сапорин, анти-CEA/антитело против цепи A рицина, анти-интерферон-α(IFN-α)/анти-гибридомный идиотип, анти-CEA/антитело против алкалоида барвинка; BsAb, относящиеся к превращению активируемых ферментами пролекарств, такие как анти-CD30/анти-щелочная фосфатаза (которая катализирует превращение пролекарства фосфата митомицина в спирт митомицина); BsAb, которые могут быть использованы в качестве фибринолитических средств, такие как анти-фибрин/антитело против тканевого активатора плазминогена (tPA), анти-фибрин/антитело против активатора плазминогена урокиназного типа (uPA); BsAb для направления иммунных комплексов к рецепторам клеточной поверхности, такие как антитело против липопротеинов низкой плотности (LDL)/анти-Fc-рецептор (например, FcγRI, FcγRII или FcγRIII); BsAb для применения в терапии инфекционных заболеваний, такие как анти-CD3/антитело против простого вируса герпеса (HSV), антитело против комплекса T-клеточный рецептор:CD3/антитело против вируса гриппа, анти-FcγR/анти-ВИЧ; BsAb для выявления опухолей in vitro или in vivo, такие как анти-CEA/анти-EOTUBE, анти-CEA/анти-DPTA, анти-p185HER2/анти-гаптен; BsAb в качестве адъювантов вакцин; и BsAb в качестве диагностических средств, такие как антитело против IgG кролика/анти-ферритин, антитело против пероксидазы хрена (HRP)/антигормон, анти-соматостатин/антитело против вещества P, анти-HRP/анти-FITC, анти-CEA/анти-β-галактозидаза. Примеры триспецифичных антител включают анти-CD3/анти-CD4/анти-CD37, анти-CD3/анти-CD5/анти-CD37 и анти-CD3/анти-CD8/анти-CD37. Биспецифичные антитела могут быть получены в виде полноразмерных антител или фрагментов антител (например, биспецифичные F(ab')2-антитела).
Способы получения биспецифичных антител известны в данной области. Традиционное получение полноразмерных биспецифичных антител основано на коэкспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулина, при этом две цепи имеют разные специфичности (Millstein et al., Nature, 305: 537-539 (1983)). Вследствие случайного выбора тяжелых и легких цепей иммуноглобулина такие гибридомы (квадромы) продуцируют возможную смесь 10 разных молекул антител, из которых только одна имеет правильную биспецифичную структуру. Очистка правильной молекулы, которую обычно осуществляют, используя стадии аффинной хроматографии, является довольно громоздкой, а выходы продукта низкими. Сходные способы описаны в WO 93/08829 и в Traunecker et al., EMBO J., 10: 3655-3659 (1991).
Согласно другом способу вариабельные домены антител с требуемыми специфичностями связывания (участки связывания антитело-антиген) сливают с последовательностями константных доменов иммуноглобулина. Слияние предпочтительно осуществляют с константным доменом тяжелой цепи иммуноглобулина, содержащим, по меньшей мере, часть шарнирной области, областей CH2 и CH3. Предпочтительно иметь первую константную область тяжелой цепи (CH1), содержащую сайт, необходимый для связывания легкой цепи, присутствующий, по меньшей мере, в одном из слияний. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина и, в случае необходимости, легкой цепи иммуноглобулина, встраивают в отдельные экспрессирующие векторы и котрансфицируют в подходящий организм-хозяин. Это дает большую гибкость при корректировке взаимных соотношений трех полипептидных фрагментов в таких вариантах, когда неравные соотношения трех полипептидных цепей, используемых в конструкции, дают оптимальные выходы. Однако можно встраивать кодирующие последовательности для двух или всех трех полипептидных цепей в один экспрессирующий вектор в том случае, когда экспрессия, по меньшей мере двух, полипептидных цепей в равных соотношениях дает высокие выходы или когда соотношения не имеют особого значения.
В одном варианте указанного способа биспецифичные антитела состоят из гибридной тяжелой цепи иммуноглобулина с первой специфичностью связывания в одном плече и гибридной пары тяжелой цепи-легкой цепи иммуноглобулина (обеспечивающей вторую специфичность связывания) в другом плече. Обнаружено, что такая асимметричная структура облегчает отделение требуемого биспецифичного соединения от нежелательных комбинаций цепей иммуноглобулина, так как наличие легкой цепи иммуноглобулина только в одной половине биспецифичных молекул обеспечивает простой способ разделения. Такой способ описан в WO 94/04690. Более подробное описание создания биспецифичных антител см., например, в Suresh et al., Methods in Enzymology, 121: 210 (1986).
Согласно другому способу, описанному в WO 96/27011, область контакта между парой молекул антител может быть сконструирована так, чтобы максимизировать процент гетеродимеров, которые извлекают из культуры рекомбинантных клеток. Предпочтительная область контакта содержит, по меньшей мере, часть домена CH3 константного домена антитела. В указанном способе одну или несколько небольших боковых цепей аминокислот из области контакта первой молекулы антитела заменяют более крупными боковыми цепями (например, тирозин или триптофан). Создают компенсирующие «впадины», имеющие размер, идентичный или сходный с размером большой боковой цепи(цепей), на области контакта второй молекулы антитела, заменяя большие боковые цепи аминокислот меньшими боковыми цепями (например, аланин или треонин). Это обеспечивает механизм увеличения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Способы создания биспецифичных антител из фрагментов антител также описаны в литературе. Например, биспецифичные антитела могут быть получены с использованием химического связывания. В Brennan et al., Science, 229: 81 (1985) описан способ, при котором интактные антитела протеолитически расщепляют, чтобы создать F(ab')2-фрагменты. Полученные фрагменты восстанавливают в присутствии агента, образующего комплексы дитиола, арсенита натрия, чтобы стабилизировать соседние дитиолы и предотвратить образование межмолекулярного дисульфида. Затем созданные Fab'-фрагменты превращают в производные тионитробензоата (TNB). Один из Fab'-TNB-производных затем снова превращают в Fab'-тиол восстановлением с использованием меркаптоэтиламина и смешивают с эквимолярным количеством другого Fab'-TNB-производного с образованием биспецифичного антитела. Полученные биспецифичные антитела можно использовать в качестве агентов для избирательной иммобилизации ферментов.
Недавние достижения облегчили прямое извлечение Fab'-SH-фрагментов из E.coli, которые могут быть химически связаны с образованием биспецифичных антител. В Shalaby et al., J. Exp.Med., 175: 217-225 (1992) описано получение полностью гуманизированной молекулы биспецифичного антитела на основе F(ab')2. Каждый Fab'-фрагмент отдельно секретировался из клеток E.coli, и его подвергали химическому связыванию in vitro с образованием биспецифичного антитела. Образованное таким образом биспецифичное антитело способно связываться с клетками, сверхэкспрессирующими рецептор VEGF, и нормальными T-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека против опухолевых мишеней молочной железы человека.
Также описаны различные способы получения и выделения фрагментов биспецифичных антител непосредственно из культуры рекомбинантных клеток. Например, были получены биспецифичные антитела с использованием лейциновых молний. Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992). Пептиды лейциновой молнии из белков Fos и Jun связывали с Fab'-частями двух разных антител посредством слияния генов. Гомодимеры антител восстанавливали в шарнирной области, чтобы образовать мономеры, и затем снова окисляли с образованием гетеродимеров антител. Такой способ также можно использовать для получения гомодимеров антител. Способ «диантител», описанный Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993), дал альтернативный механизм получения фрагментов биспецифичных антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) линкером, который слишком короткий, чтобы обеспечить возможность спаривания между двумя доменами на одной и той же цепи. Соответственно, домены VH и VL одного фрагмента вынуждены спариваться с комплементарными доменами VL и VH другого фрагмента, образуя при этом два антигенсвязывающих участка. Также сообщалось о другой методике получения фрагментов биспецифичных антител с использованием димеров одноцепочечного Fv (sFv). См. Gruber et al., J. Immunol., 152: 5368 (1994).
Предполагаются антитела с более чем двумя валентностями. Например, могут быть получены триспецифичные антитела. Tutt et al. J. Immunol. 147: 60 (1991).
Гетероконъюгаты антител
Биспецифичные антитела включают поперечно сшитые или «гетероконъюгированные» антитела, которые являются антителами согласно изобретению. Например, одно из антител в гетероконъюгате может быть связано с авидином, а другое с биотином. Такие антитела, например, были предложены для того, чтобы направить клетки иммунной системы к нежелательным клеткам-мишеням (патент США №4676980), и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373 и EP 03089). Гетероконъюгаты антител могут быть получены с использованием любых способов образования поперечных сшивок. Подходящие поперечно сшивающие агенты хорошо известны в данной области и описаны в патенте США №4676980 вместе с рядом способов образования поперечных сшивок.
Поливалентные антитела
Антитела согласно изобретению включают поливалентное антитело. Поливалентное антитело может быть быстрее интернализовано (и/или подвергнуто катаболизму) клеткой, экспрессирующей антиген, с которым связываются антитела, чем бивалентное антитело. Антитела согласно изобретению могут представлять собой поливалентные антитела (которые не относятся к классу IgM) с тремя или более антигенсвязывающими участками (т.е. тетравалентные антитела), которые могут быть легко получены посредством рекомбинантной экспрессии нуклеиновой кислоты, кодирующей полипептидные цепи антитела. Поливалентное антитело может содержать домен димеризации и три или более антигенсвязывающих участка. Предпочтительный домен димеризации содержит (или состоит из) Fc-области или шарнирной области. По такому плану антитело будет содержать Fc-область и три или более антигенсвязывающих участка, аминоконцевых по отношению к Fc-области. Предпочтительное поливалентное антитело согласно изобретению содержит (или состоит из) от трех до примерно восьми, но предпочтительно четыре антигенсвязывающих участка. Поливалентное антитело содержит, по меньшей мере, одну полипептидную цепь (и предпочтительно две полипептидных цепи), при этом полипептидная цепь(и) содержит два или более вариабельных домена. Например, полипептидная цепь(и) может содержать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 означает первый вариабельный домен, VD2 означает второй вариабельный домен, Fc означает одну полипептидную цепь Fc-области, X1 и X2 означают аминокислоту или полипептид, и n равно 0 или 1. Например, полипептидная цепь(и) может содержать цепь VH-CH1-гибкий линкер-VH-CH1-Fc-область или цепь VH-CH1-VH-CH1-Fc-область. Поливалентное антитело согласно изобретению предпочтительно дополнительно содержит, по меньшей мере, два (и предпочтительно четыре) полипептида вариабельного домена легкой цепи. Поливалентное антитело согласно изобретению может, например, содержать от примерно двух до примерно восьми полипептидов вариабельного домена легкой цепи. Полипептиды вариабельного домена легкой цепи, предусмотренные в настоящем изобретении, содержат вариабельный домен легкой цепи и необязательно дополнительно содержат домен CL.
Конструирование эффекторных функций
Может быть желательной модификация антитела согласно изобретению в отношении эффекторной функции так, чтобы, например, повысить эффективность антитела для лечения злокачественной опухоли. Например, остаток(ки) цистеина могут быть введены в Fc-область, таким образом обеспечивая возможность образования межцепочечного дисульфида в данной области. Созданное таким образом гомодимерное антитело может иметь улучшенную способность к интернализации и/или повышенному опосредованному комплементом цитолизу клеток и антителозависимой клеточной цитотоксичности (ADCC). См. Caron et al., J. Exp Med. 176: 1191-1195 (1992) и Shopes, B. J. Immunol. 148: 2918-2922 (1992). Гомодимерные антитела с повышенной направленной к мишени активностью также могут быть получены с использованием гетеробифункциональных поперечно сшивающих агентов, которые описаны в Wolff et al. Cancer Research 53: 2560-2565 (1993).
Альтернативно может быть сконструировано антитело, которое имеет двойные Fc-области и в связи с этим обладает повышенной способностью к зависимому от комплемента лизису и ADCC. См. Stevenson et al. Anti-Cancer Drug Design 3: 219-230 (1989). Чтобы увеличить время полужизни антитела в сыворотке, можно включить в антитело (в частности, во фрагмент антитела) эпитоп связывания рецептора спасения, как описано, например, в патенте США 5739277. В используемом в данном описании смысле термин «эпитоп связывания рецептора спасения» относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который ответственен за увеличение времени полужизни молекулы IgG в сыворотке in vivo.
Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам, содержащим антитело, описанное в данной публикации, конъюгированное с цитотоксическим средством, таким как химиотерапевтическое средство, токсин (например, ферментативно активный токсин, происходящий из клеток бактерий, грибов, растений или животных, или его фрагменты) или радиоактивный изотоп (т.е. радиоконъюгат). Имеется множество радионуклидов для получения радиоконъюгатов антител. Примеры включают без ограничения, например, 212Bi, 131I, 131In, 90Y и 186Re.
Химиотерапевтические средства, применимые для создания таких иммуноконъюгатов, были описаны выше. Например, BCNU, стрептозоицин, винкристин, 5-фторурацил, семейство агентов, вместе известных как комплекс LL-E33288, описанных в патентах США 5053394, 5770710, эсперамицины (патент США 5877296) и т.д. (см. также определение химиотерапевтических средств в данном описании) могут быть конъюгированы с анти-ANGPTL4-антителами или антителами против ангиогенеза или их фрагментами.
Для избирательного разрушения клетки антитело может содержать высокорадиоактивный атом. Множество радиоактивных изотопов доступно для получения радиоконъюгированных анти-ANGPTL4 или их фрагментов. Примеры включают без ограничения например, 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 32P, 212Pb, 111In, радиоактивные изотопы Lu и т.д. В том случае, когда конъюгат используют для диагностики, он может содержать радиоактивный атом для сцинтиографических исследований, например, 99mTc или 123I, или спиновую метку для ядерной магнитно-резонансной (ЯМР) визуализации (также известной как магнитно-резонансная визуализация, MRI), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки могут быть включены в конъюгат известными способами. Например, пептид может быть биологически синтезирован или может быть синтезирован посредством химического синтеза из аминокислот с использованием подходящих предшественников аминокислот, содержащий, например, фтор-19 вместо водорода. Такие метки как 99mTc или 123I, 186Re, 188Re и 111In, могут быть связаны с пептидом через остаток цистеина. Иттрий-90 может быть связан через остаток лизина. Способ IODOGEN (Fraker et al. Biochem. Biophys. Res. Commun. 80: 49-57 (1978) можно использовать для включения йода-123. См., например, Monoclonal Antibodies in Immunoscintigraphy (Chatal, CRC Press, 1989), где подробно описаны другие способы.
Ферментативно активные токсины и их фрагменты, которые могут быть использованы, включают цепь A дифтерийного токсина, не связывающиеся активные фрагменты дифтерийного токсина, цепь A экзотоксина (из Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модексина, альфа-сарцин, белки Aleurites fordii, белки диантины, белки Phytolacca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, неомицин и трикотецены. См., например, WO 93/21232, опубликованную 28 октября 1993.
Конъюгаты антитела и цитотоксического средства получают, используя множество бифункциональных связывающих белки агентов, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат, иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат·HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(пара-азидобензоил)гександиамин), производные бис-диазония (такие как бис(пара-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин может быть получен, как описано в Vitetta et al. Science 238: 1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего агента для конъюгирования радионуклида с антителом. См. WO94/11026. Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать кислотолабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, линкер на основе диметила или линкер, содержащий дисульфид (Chari et al., Cancer Research 52: 127-131 (1992); патент США №5208020).
Альтернативно можно получить слитый белок, содержащий анти-ANGPTL4 и цитотоксическое средство, например, способами на основе рекомбинации или пептидным синтезом. В длину ДНК могут быть включены соответствующие области, кодирующие две части конъюгата, либо расположенные рядом друг с другом, либо разделенные областью, кодирующей линкерный пептид, который не нарушает требуемые свойства конъюгата.
В некоторых вариантах антитело конъюгируют с «рецептором» (например, стрептавидином) в случае применения для предварительного направления к клетке-мишени, когда конъюгат антитело-рецептор вводят пациенту с последующим удалением несвязанного конъюгата из кровообращения с использованием хелатирующего агента и затем введением «лиганда» (например, авидина), который конъюгируют с цитотоксическим средством (например, радионуклидом). В некоторых вариантах иммуноконъюгат образуют между антителом и соединением с нуклеолитической активностью (например, рибонуклеазой или ДНК-эндонуклеазой, такой как дезоксирибонуклеаза; ДНКаза).
Мейтанзин и мейтанзиноиды
Изобретение относится к антителу согласно изобретению, которое конъюгировано с одной или несколькими молекулами мейтанзиноидов. Мейтанзиноиды являются ингибиторами митоза, которые действуют, ингибируя полимеризацию тубулина. Мейтанзин впервые был выделен из восточно-африканского кустарника Maytenus serrata (патент США №3896111). Затем было обнаружено, что некоторые микроорганизмы также продуцируют мейтанзиноиды, такие как мейтанзинол и сложные C-3-мейтанзиноловые эфиры (патент США №4151042). Синтетический мейтанзинол и его производные и аналоги описаны, например, в патентах США №4137230, 4248870, 4256746, 4260608, 4265814, 4294757, 4307016, 4308268, 4308269, 4309428, 4313946, 4315929, 4317821, 4322348, 4331598, 4361650, 4364866, 4424219, 4450254, 4362663 и 4371533.
Например, анти-ANGPTL4-антитело или анти-αVβ5-антитело конъюгируют с молекулой мейтанзиноида без существенного снижения биологической активности антитела или молекулы мейтанзиноида. Показано, что в среднем 3-4 молекулы мейтанзиноида, конъюгированных на молекулу антитела, эффективны для усиления цитотоксичности для клеток-мишеней без негативного влияния на функцию или растворимость антитела, хотя можно ожидать, что даже одна молекула токсин/антитело усиливает цитотоксичность по сравнению с применением «голого» антитела. Мейтанзиноиды хорошо известны в данной области и могут быть синтезированы известными способами или выделены из природных источников. Подходящие мейтанзиноиды описаны, например, в патенте США №5208020 и в других патентах и непатентных публикациях, ссылки на которые указаны выше. В одном варианте мейтанзиноиды представляют собой мейтанзинол и аналоги мейтанзинола, модифицированные в ароматическом цикле или в других положениях молекулы мейтанзинола, такие как различные сложные мейтанзиноловые эфиры.
Существует много связывающих групп, известных в данной области для получения конъюгатов антитело-мейтанзиноид, включая, например, группы, описанные в патенте США №5208020 или патенте EP 0425235 B1 и в Chari et al., Cancer Research 52: 127-131 (1992). Связывающие группы включают дисульфидные группы, тиоэфирные группы, кислотолабильные группы, фотолабильные группы, неустойчивые к действию пептидазы группы или неустойчивые к действию эстеразы группы, которые описаны в указанных выше патентах, при этом предпочтительны дисульфидные и тиоэфирные группы.
Конъюгаты антитела и мейтанзиноида могут быть получены с использованием множества бифункциональных, связывающих белки агентов, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат, иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат·HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(пара-азидобензоил)гександиамин), производные бис-диазония (такие как бис(пара-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Обычные связывающие агенты включают N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173: 723-737 [1978]) и N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP) для образования дисульфидной связи.
Линкер может быть связан с молекулой мейтанзиноида в разных положениях, в зависимости от типа связи. Например, сложноэфирная связь может быть образована при взаимодействии с гидроксильной группой с использованием обычных способов связывания. Реакция может осуществляться в положении C-3, имеющим гидроксильную группу, в положении C-14, модифицированном гидроксиметилом, в положении C-15, модифицированном гидроксильной группой, и в положении C-20, имеющем гидроксильную группу. Связь образуется в положении C-3 мейтанзинола или аналога мейтанзинола.
Калихеамицин
Другой представляющий интерес иммуноконъюгат содержит анти-ANGPTL4-антитело или анти-αVβ5-антитело, конъюгированное с одной или несколькими молекулами калихеамицина. Калихеамициновое семейство антибиотиков способно образовывать двунитевые разрывы ДНК в субпикомолярных концентрациях. Получение конъюгатов семейства калихеамицина см. в патентах США 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001, 5877296 (все American Cyanamid Company). Структурные аналоги калихеамицина, которые могут быть использованы, включают без ограничения γ1 I, α2 I, α3 I, N-ацетил-γ1 I, PSAG и θI 1 (Hinman et al., Cancer Research 53: 3336-3342 (1993), Lode et al., Cancer Research 58: 2925-2928 (1998) и указанные выше патенты США, принадлежащие American Cyanamid).
Другим противоопухолевым лекарственным средством, с которым может быть конъюгировано антитело, является QFA, который является антифолатом. Как калихеамицин, так и QFA имеют внутриклеточные места действия и с трудом проходят через плазматическую мембрану. Поэтому поглощение клетками указанных средств посредством опосредованной антителами интернализации значительно усиливает их цитотоксическое действие.
Другие модификации антител
В данном изобретении предполагаются другие модификации антитела. Например, антитело может быть связано с одним из множества небелковых полимеров, например, полиэтиленгликолем, полипропиленгликолем, полиоксиалкиленами или сополимерами полиэтиленгликоля и полипропиленгликоля. Антитело также можно заключать в микрокапсулы, полученные, например, способами коацервации или полимеризацией на границе фаз (например, микрокапсулы из гидроксиметилцеллюлозы или желатиновые микрокапсулы и микрокапсулы из поли(метилметакрилата), соответственно), в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences, 16th edition, Oslo, A., Ed., (1980).
Липосомы и наночастицы
Полипептиды согласно изобретению могут быть приготовлены в липосомах. Например, антитела согласно изобретению также могут быть приготовлены в виде иммунолипосом. Липосомы, содержащие антитело, получают способами, известными в данной области, такими как способы, описанные в Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688 (1985); Hwang et al., Proc. Natl Acad. Sci. USA, 77: 4030 (1980) и патентах США №4485045 и 4544545. Липосомы с повышенным периодом циркуляции описаны в патенте США №5013556. В общем, создание и применение липосом известно специалистам в данной области.
Особенно применимые липосомы могут быть созданы способом обращенно-фазового выпаривания с использовании композиции липидов, содержащей фосфатидилхолин, холестерин и ПЭГ-дериватизованный фосфатидилэтаноламин (PEG-PE). Липосомы подвергают экструзии через фильтры с определенным размером пор, чтобы получить липосомы требуемого диаметра. Fab'-фрагменты антитела согласно изобретению можно конъюгировать с липосомами, как описано в Martin et al. J. Biol. Chem. 257: 286-288 (1982), посредством реакции дисульфидного обмена. Наночастицы или нанокапсулы также можно использовать для улавливания полипептидов согласно изобретению. В одном варианте биоразрушаемые наночастицы полиалкилцианоакрилата можно использовать с полипептидами согласно изобретению.
Другие применения
Анти-ANGPTL4-антитела имеют разнообразные применения. Например, анти-ANGPTL4-антитела можно использовать в диагностических анализах в отношении ANGPTL4 или фрагментов ANGPTL4, например, для выявления их экспрессии в конкретных клетках, тканях или сыворотке, для выявления заболеваний, например, расстройств, описанных в данной публикации, и т.д. В одном варианте антитела к ANGPTL4 используют для отбора популяции пациентов для лечения способами, предлагаемыми в настоящем изобретении. Могут быть использованы различные способы диагностического анализа, известные в данной области, такие как анализы конкурентного связывания, прямые или непрямые анализы типа «сэндвич» и анализы иммунопреципитации, проводимые либо в гетерогенных, либо в гомогенных фазах (Zola, Monoclonal Antibodies: A Manual of Techniques, CRC Press, Inc. (1987) pp.147-158). Антитела, используемые в диагностических анализах, можно метить регистрируемым компонентом. Регистрируемый компонент должен обладать способностью прямо или косвенно производить регистрируемый сигнал. Например, регистрируемым компонентом может быть радиоизотоп, такой как 3H, 14C, 32P, 35S или 125I, флуоресцирующее или хемилюминесцентное соединение, такое как изотиоцианат флуоресцеина, родамин или люциферин, или фермент, такой как щелочная фосфатаза, бета-галактозидаза или пероксидаза хрена. Можно использовать любой способ, известный в данной области для конъюгирования антитела с регистрируемым компонентом, включая способы, описанные в Hunter et al., Nature, 144: 945 (1962); David et al., Biochemistry, 13: 1014 (1974); Pain et al., J. Immunol. Meth., 40: 219 (1981) и Nygren, J. Histochem. and Cytochem., 30: 407 (1982).
Анти-ANGPTL4-антитела также применимы для аффинной очистки ANGPTL4 из культуры рекомбинантных клеток или природных источников. В таком способе антитела против ANGPTL4 иммобилизуют на подходящей подложке, такой как смола сефадекс или фильтровальная бумага, используя способы, хорошо известные в данной области. Иммобилизованное антитело затем подвергают контакту с образцом, содержащим ANGPTL4, который необходимо очистить, и затем положку промывают подходящим растворителем, который будет удалять в основном все вещества в образце, кроме ANGPTL4, который связан с иммобилизованным антителом. Наконец, подложку промывают другим подходящим растворителем, который будет высвобождать ANGPTL4 из антитела.
Векторы, клетки-хозяева и способы рекомбинации
Полипептиды согласно изобретению могут быть получены рекомбинантно с использованием способов и веществ, которые могут быть легко получены.
Для рекомбинантного получения полипептида согласно изобретению, например ANGPTL4, анти-ANGPTL4-антитела или анти-αVβ5-антитела, кодирующую его нуклеиновую кислоту, выделяют и встраивают в реплицируемый вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую полипептид согласно изобретению, легко выделяют и секвенируют, используя обычные способы. Например, ДНК, кодирующую моноклональное антитело, выделяют и секвенируют, например, используя олигонуклеотидные зонды, которые способны специфично связываться с генами, кодирующими тяжелые и легкие цепи антитела. Имеется множество векторов. Компоненты вектора обычно включают, без ограничения, один или несколько из указанных ниже компонентов: сигнальную последовательность, начало репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
Компонент - сигнальная последовательность
Полипептиды согласно изобретению могут быть получены рекомбинантно не только непосредственно, но также в виде слитого полипептида с гетерологичным полипептидом, который обычно представляет собой сигнальную последовательность или другой полипептид, имеющий специфичный сайт расщепления на N-конце зрелого белка или полипептида. Выбранная гетерологичная сигнальная последовательность обычно представляет собой последовательность, которая узнается и процессируется (т.е. отщепляется сигнальной пептидазой) клеткой-хозяином. В случае прокариотических клеток-хозяев, которые не узнают и не процессируют сигнальную последовательность нативного полипептида, сигнальную последовательность заменяют прокариотической сигнальной последовательностью, выбранной, например, из группы лидеров щелочной фосфатазы, пенициллиназы, lpp или термостабильного энтеротоксина II. Для секреции дрожжами нативная сигнальная последовательность может быть заменена, например, лидером инвертазы дрожжей, лидером α-фактора (включая лидеры α-факторов Saccharomyces и Kluyveromyces) или лидером кислой фосфатазы, лидером глюкоамилазы C. albicans или сигналом, описанным в WO 90/13646. Для экспрессии в клетках млекопитающих имеются сигнальные последовательности млекопитающих, а также вирусные лидеры секреции, например, сигнал gD простого герпеса.
ДНК для такой области предшественника лигируют в рамке считывания с ДНК, кодирующей полипептид согласно изобретению.
Компонент - начало репликации
И экспрессирующие и клонирующие векторы содержат последовательность нуклеиновой кислоты, которая делает возможной репликацию вектора в одной или нескольких выбранных клетках-хозяевах. В общем, в клонирующих векторах такая последовательность представляет собой последовательность, которая обеспечивает возможность репликации вектора независимо от хромосомной ДНК хозяина, и включает в себя начала репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для многих бактерий, дрожжей и вирусов. Начало репликации из плазмиды pBR322 подходит для большинства грамотрицательных бактерий, начало из 2-мкм-плазмиды подходит для дрожжей и различные вирусные начала репликации (SV40, полиомы, аденовируса, VSV или BPV) подходят для клонирующих векторов в клетках млекопитающих. В общем, компонент, представленный началом репликации, не является необходимым для экспрессирующих вектором млекопитающих (начало SV40 обычно можно использовать только потому, что оно содержит ранний промотор).
Компонент - ген для селекции
Экспрессирующие и клонирующие векторы могут содержать ген для селекции, также называемый маркером селекции. Обычно гены селекции кодируют белки, которые (a) придают резистентность к антибиотикам или другим токсинам, например, к ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют ауксотрофные дефициты, или (c) снабжают необходимыми питательными веществами, недоступными из сложных сред, например, ген, кодирующий D-аланинрацемазу для Bacilli.
В одном примере схемы селекции используют лекарственное средство для задержки роста клетки-хозяина. Те клетки, которые успешно трансформированы гетерологичным геном, продуцируют белок, придающий резистентность к лекарственному средству, и таким образом выживают в режиме селекции. В примерах такой доминантной селекции используют лекарственные средства неомицин, микофеноловую кислоту и гигромицин.
Другим примером подходящих маркеров селекции для клеток млекопитающих являются маркеры, которые обеспечивают возможность идентификации клеток, компетентных в отношении поглощения нуклеиновой кислоты антитела, такие как гены DHFR, тимидинкиназы, металлотионеина-I и -II, обычно гены металлопротионеинов приматов, аденозиндезаминазы, орнитиндекарбоксилазы и т.д.
Например, клетки, трансформированные геном селекции DHFR, сначала идентифицируют культивированием всех трансформантов в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. Подходящей клеткой-хозяином при использовании DHFR дикого типа является линия клеток яичника китайского хомячка (CHO), дефицитных по активности DHFR.
Альтернативно, клетки-хозяева (особенно хозяева дикого типа, которые содержат эндогенную DHFR), трансформированные или котрансформированные последовательностями ДНК, кодирующими полипептид согласно изобретению, белок DHFR дикого типа и другой маркер селекции, такой как аминогликозид-3'-фосфотрансфераза (APH) можно отбирать по росту клеток в среде, содержащей агент для селекции, соответствующий маркеру селекции, такой как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. патент США №4965199.
Подходящим геном селекции для применения у дрожжей является ген trp1, присутствующий в дрожжевой плазмиде Yrp7 (Stinchcomb et al., Nature, 282: 39 (1979)). Ген trp1 обеспечивает маркер селекции для мутантного штамма дрожжей, у которого отсутствует способность расти на триптофане, например, ATCC №44076 или PEP4-1. Jones, Genetics, 85: 12 (1977). В таком случае наличие повреждения trp1 в геноме дрожжевой клетки-хозяина создает эффективные условия для регистрации трансформации по росту в отсутствие триптофана. Подобным образом, штаммы дрожжей, дефицитные по Leu2 (ATCC 20622 или 38626), комплементируются известными плазмидами, несущими ген Leu2.
Кроме того, векторы, полученные на основе кольцевой 1,6-мкм-плазмиды pKD1, могут быть использованы для трансформации дрожжей Kluyveromyces. Альтернативно, сообщалось о системе экспрессии для крупномасштабного производства рекомбинантного химозина теленка для K. lactis. Van den Berg, Bio/Technology, 8: 135 (1990). Также описаны стабильные мультикопийные экспрессирующие векторы для секреции зрелого рекомбинантного сывороточного альбумина человека промышленными штаммами Kluyveromyces. Fleer et al., Bio/Technology, 9: 968-975 (1991).
Промоторный компонент
Экспрессирующие и клонирующие векторы обычно содержат промотор, который распознается организмом-хозяином и оперативно связан с нуклеиновой кислотой, кодирующей полипептид согласно изобретению. Промоторы, подходящие для применения при использовании прокариотических хозяев, включают промотор phoA, промоторные системы β-лактамазы и лактозы, промотор щелочной фосфатазы, промоторную систему триптофана (trp) и гибридные промоторы, такие как промотор tac. Однако подходящими являются и другие известные бактериальные промоторы. Промоторы для применения в бактериальных системах также будут содержать последовательность Шайна-Далгарно (S.D.), оперативно связанную с ДНК, кодирующей полипептид согласно изобретению.
Известны промоторные последовательности для эукариот. Практически все эукариотические гены имеют AT-богатую область, расположенную примерно на 25-30 оснований выше сайта инициации транскрипции. Другой последовательностью, обнаруживаемой на 70-80 оснований выше старта транскрипции многих генов, является область CNCAAT, где N может быть любым нуклеотидом. На 3'-конце большинства эукариотических генов находится последовательность AATAAA, которая может быть сигналом для добавления поли-A-хвоста к 3'-концу кодирующей последовательности. Все такие последовательности соответствующим образом встраивают в эукариотические экспрессирующие векторы.
Примеры подходящих промоторных последовательностей для применения в дрожжевых хозяевах включают промоторы 3-фосфоглицераткиназы или других гликолитических ферментов, таких как енолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другими дрожжевыми промоторами, которые являются индуцируемыми промоторами, имеющими дополнительное преимущество, связанное с транскрипцией, регулируемой условиями роста, являются области промотора алкогольдегидрогеназы 2, изоцитохрома C, кислой фосфатазы, разрушающих ферментов, связанных с метаболизмом азота, металлотионеина, глицеральдегид-3-фосфатдегидрогеназы и ферментов, ответственных за утилизацию мальтозы и галактозы. Подходящие векторы и промоторы для применения для экспрессии у дрожжей дополнительно описаны в EP 73657. С дрожжевыми промоторами также предпочтительно используют дрожжевые энхансеры.
Транскрипция полипептидов согласно изобретению с векторов в клетках-хозяевах млекопитающих регулируется, например, промоторами, полученными из геномов вирусов, таких как вирус полиомы, поксвирус птиц, аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита-B и обычно вирус 40 обезьян (SV40), гетерологичными промоторами млекопитающих, например, промотором актина или промотором иммуноглобулина, промоторами теплового шока, при условии, что такие промоторы совместимы с системами клеток-хозяев.
Ранний и поздний промоторы вируса SV40 легко получают в виде рестрикционного фрагмента SV40, который также содержит начало репликации вируса SV40. Немедленный ранний промотор цитомегаловируса человека легко получают в виде фрагмента рестрикции HindIIIE. Система для экспрессии ДНК в хозяевах-млекопитающих с использованием вируса бычьей папилломы в качестве вектора описана в патенте США №4419446. Модификация такой системы описана в патенте США №4601978. См. также Reyes et al., Nature 297: 598-601 (1982) в отношении экспрессии кДНК β-интерферона человека в мышиных клетках под контролем промотора тимидинкиназы из вируса простого герпеса. Альтернативно, в качестве промотора можно использовать длинный концевой повтор вируса саркомы Рауса.
Компонент - энхансерный элемент
Транскрипцию ДНК, кодирующей полипептид согласно настоящему изобретению, высшими эукариотами часто увеличивают встраиванием последовательности энхансера в вектор. В настоящее время известно много энхансерных последовательностей генов млекопитающих (глобина, эластазы, альбумина, α-фетопротеина и инсулина). Как правило, используют энхансер из вируса эукариотических клеток. Примеры включают энхансер SV40, расположенный после начала репликации (100-270 п.н.), энхансер раннего промотора цитомегаловируса, энхансер вируса полиомы, расположенный после начала репликации, и аденовирусные энхансеры. См. также Yaniv, Nature 297: 17-18 (1982) в отношении усиливающих элементов для активации эукариотических промоторов. Энхансер может быть сплайсирован в вектор в положении 5'- или 3'- по отношению к последовательности, кодирующей полипептид, но обычно расположен в сайте с 5'-стороны от промотора.
Компонент для терминации транскрипции
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах (дрожжей, грибов, насекомых, растений, животных, человека или нуклеированных клетках из других многоклеточных организмов), также будут содержать последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно доступны из 5'- и иногда 3'-нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Указанные области содержат нуклеотидные участки, транскрибируемые в виде полиаденилированных фрагментов в нетранслируемой части мРНК, кодирующей полипептид согласно изобретению. Одним из применимых компонентов для терминации транскрипции является область полиаденилирования бычьего гормона роста. См. WO94/11026 и описанные в заявке экспрессирующие векторы.
Селекция и трансформация клеток-хозяев
Подходящими клетками-хозяевами для клонирования или экспрессии ДНК, кодирующей полипептиды согласно изобретению, в векторах согласно изобретению являются прокариотические, дрожжевые клетки или клетки высших эукариот, описанные выше. Подходящие прокариоты для указанной цели включают эубактерии, такие как грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как Escherichia, например E.coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans, и Shigella, а также Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанная в DD 266710, опубликованном 12 апреля 1989), Pseudomonas, такие как P. aeruginosa, и Streptomyces. Обычно клонирующим хозяином в случае E.coli является E.coli 294 (ATCC 31446), хотя подходящими являются и другие штаммы, такие как E.coli B, E.coli X1776 (ATCC 31537), и E.coli W3110 (ATCC 27325). Указанные примеры являются иллюстративными, а не ограничивающими.
Кроме прокариот, подходящими клонирующими или экспрессирующими хозяевами для векторов, кодирующих полипептид согласно изобретению, являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи. Saccharomyces cerevisiae или обычные пекарские дрожжи являются наиболее широко используемыми из низших эукариотических микроорганизмов-хозяев. Однако ряд других родов, видов и штаммов являются общедоступными и применимыми в настоящем изобретении, такие как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие, например, как K. lactis, K. fragilis (ATCC 12424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906), K. thermotolerans и K. marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитчатые грибы, такие как, например, Neurospora, Penicillium, Tolypocladium, и хозяева Aspergillus, такие как A. nidulans и A. niger.
Подходящие клетки-хозяева для экспрессии гликозилированных полипептидов согласно изобретению получают из многоклеточных организмов. Примерами клеток беспозвоночных являются клетки растений и насекомых. Идентифицированы многочисленные бакуловирусные штаммы и варианты и соответствующие пермиссивные клетки-хозяева насекомых из таких хозяев как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Общедоступно множество вирусных штаммов для трансфекции, например, вариант L-1 NPV Autographa californica и штамм Bm-5 NPV Bombyx mori, и такие вирусы могут быть использованы в качестве вируса согласно изобретению, в частности, для трансфекции клеток Spodoptera frugiperda. Культуры растительных клеток хлопка, кукурузы, картофеля, сои, петунии, томатов и табака также можно использовать в качестве хозяев.
Однако наибольший интерес представляли клетки позвоночных, и размножение клеток позвоночных в культуре (культуре ткани) стало рутинной процедурой. Примерами применимых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированных SV40 (COS-7, ATCC CRL 1651); линия клеток эмбриональной почки человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36: 59 (1977)); клетки почки сирийского хомячка (BHK, ATCC CCL 10); клетки яичника китайского хомячка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 4216 (1980)); клетки Сертоли мышей (TM4, Mather, Biol. Reprod. 23: 243-251 (1980); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой обезьяны (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крыс Buffalo (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383: 44-68 (1982)); клетки MRC 5; клетки FS4 и линия клеток гепатомы человека (Hep G2).
Клетки-хозяева трансформируют описанными выше экспрессирующими или клонирующими векторами для продукции полипептида согласно изобретению и культивируют в обычных питательных средах, соответствующим образом модифицированных для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих требуемые последовательности.
Культивирование клеток-хозяев
Клетки-хозяева, используемые для получения полипептидов согласно изобретению, можно культивировать во многих средах. Коммерчески доступные среды, такие как среда Хама F10 (Sigma), минимальная питательная среда ((MEM), (Sigma), RPMI-1640 (Sigma), модифицированная по Дульбекко среда Игла ((DMEM), Sigma), нормальные ростовые среды для гепатоцитов (Cambrex), среда для роста преадипоцитов (Cambrex) и т.д., подходят для культивирования клеток-хозяев. Кроме того, любую из сред, описанных в Ham et al., Meth. Enz. 58: 44 (1979), Barnes et al., Anal. Biochem. 102: 255 (1980), патентах США №4767704, 4657866, 4927762, 4560655 или 5122469, WO 90/03430, WO 87/00195 или в патенте США Re. 30985, можно использовать в качестве культуральной среды для клеток-хозяев. Любая из указанных сред может быть дополнена при необходимости гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальций, магний и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство GENTAMYCINTM), микроэлементами (определяемыми как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Любые другие необходимые добавки также могут быть включены в соответствующих концентрациях, которые могут быть известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия, используемые ранее в случае клеток-хозяев, отобранных для экспрессии, и условия будут известны специалисту в данной области.
Очистка полипептидов
При использовании рекомбинантных способов полипептид согласно изобретению, например ANGPTL4, анти-ANGPTL4-антитело или анти-αVβ5-антитело, может быть получен внутриклеточно, в периплазматическом пространстве или непосредственно секретирован в среду. Полипептиды согласно изобретению могут быть извлечены из культуральной среды или из лизатов клеток хозяев. Если полипептид связан с мембраной, то его можно высвобождать из мембраны с использованием раствора подходящего детергента (например, тритона-X 100) или ферментативным расщеплением. Клетки, используемые для экспрессии полипептида согласно изобретению, могут быть разрушены различными физическими или химическими способами, такими как циклы замораживания-оттаивания, обработка ультразвуком, механическое разрушение или с использованием агентов, лизирующих клетки.
Может требоваться очистка полипептида согласно изобретению от белков или полипептидов рекомбинантной клетки. Следующие способы являются примером подходящих способов очистки: фракционирование на ионообменной колонке; преципитация этанолом; обращенно-фазовая ВЭЖХ; хроматография на диоксиде кремния, хроматография на гепарин-SepharoseTM, хроматография на анионообменной или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой, DEAE и т.д.); хроматофокусировка; SDS-ПААГ; преципитация сульфатом аммония; гель-фильтрация с использованием, например, сефадекса G-75; колонки с белком A-сефарозой, чтобы удалить такие примеси, как IgG; и металл-хелатных колонок, чтобы связать меченные эпитопом формы полипептидов согласно изобретению. Могут быть использованы различные способы очистки белка и такие способы известны в данной области и описаны, например, в Deutscher, Methods in Enzymology, 182 (1990); Scopes, Protein Purification: Principles and Practice, Springer-Verlag, New York (1982). Выбранная стадия(и) очистки будет зависеть, например, от природы применяемого способа получения и конкретного полученного полипептида согласно изобретению.
Например, композиция антител, полученных из клеток, может быть очищена с использованием, например, хроматографии на гидроксилапатите, гель-электрофореза, диализа и аффинной хроматографии, при этом аффинная хроматография является типичным способом очистки. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, который присутствует в антителе. Белок A можно использовать для очистки антител, которые основаны на тяжелых цепях γ1, γ2 или γ4 человека (Lindmark et al., J. Immunol. Meth. 62: 1-13 (1983)). Белок G рекомендуется для всех мышиных изотипов и для γ3 человека (Guss et al., EMBO J. 5: 1567-1575 (1986)). Матрицей, с которой связывают аффинный лиганд, наиболее часто является агароза, но имеются и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают возможность более высоких скоростей потока и более коротких периодов времени обработки, чем можно достичь в случае агарозы. В том случае, когда антитело содержит домен CH3, для очистки применима смола Bakerbond ABXTM (J. T. Baker, Phillipsburg, NJ). Также доступны другие способы очистки белка, например, указанные выше способы, в зависимости от антитела, которое необходимо извлечь. См. также Carter et al., Bio/Technology 10: 163-167 (1992), где описан способ выделения антител, которые секретируются в периплазматическое пространство E.coli.
Ковалентные модификации полипептидов согласно изобретению
Ковалентные модификации полипептида согласно изобретению, например, ANGPTL4 или полипептидного агониста, или полипептидного антагониста входят в объем настоящего изобретения. Модификации могут быть осуществлены с помощью химического синтеза или при необходимости ферментативным или химическим расщеплением полипептида. Другие типы ковалентных модификаций полипептида вводят в молекулу посредством взаимодействия целевых аминокислотных остатков полипептида с органическим дериватизирующим агентом, который способен взаимодействовать с отдельными боковыми цепями или N- или C-концевыми остатками, или посредством включения модифицированной аминокислоты или неприродной аминокислоты в растущую полипептидную цепь, например, Ellman et al. Meth. Enzym. 202: 301-336 (1991); Noren et al. Science 244: 182 (1989); и заявки на выдачу патента США 2003/0108885 и 2003/0082575.
Остатки цистеинила в большинстве случаев взаимодействуют с α-галогенацетатами (и соответствующими аминами), такими как хлоруксусная кислота или хлорацетамид, давая карбоксиметильные или карбоксиамилометильные производные. Остатки цистеинила также подвергают дериватизации в результате взаимодействия с бромтрифторацетоном, α-бром-β-(5-имидозоил)пропионовой кислотой, хлорацетилфосфатом, N-алкилмалеимидами, 3-нитро-2-пиридилдисульфидом, метил-2-пиридилдисульфидом, пара-хлормеркурибензоатом, 2-хлормеркури-4-нитрофенолом или хлор-7-нитробензо-2-окса-1,3-диазолом.
Остатки гистидила подвергают дериватизации посредством взаимодействия с диэтилпирокарбонатом при pH 5,5-7,0, так как указанный агент является сравнительно специфичным по отношению к боковой цепи гистидила. Также применим пара-бромфенацилбромид; реакцию обычно осуществляют в 0,1 М какодилате натрия при pH 6,0.
Лизинил и аминоконцевые остатки взаимодействуют с янтарным ангидридом или ангидридами других карбоновых кислот. Дериватизация указанными агентами оказывает влияние в виде изменения заряда остатков лизинила. Другие подходящие реагенты для дериватизации α-аминосодержащих остатков включают сложные имидоэфиры, такие как метилпиколинимидат, пиридоксальфосфат, пиридоксаль, хлорборогидрид, тринитробензолсульфоновую кислоту, O-метилизомочевину, 2,4-пентандион и катализируемую трансаминазой реакцию с глиоксилатом.
Остатки аргинила модифицируют посредством взаимодействия с одним или несколькими обычными реагентами, среди которых фенилглиоксаль, 2,3-бутандион, 1,2-циклогександион и нингидрин. Дериватизация остатков аргинина требует, чтобы реакция проходила в щелочных условиях из-за высокого значения pKa функциональной группы гуанидина. Кроме того, указанные реагенты могут взаимодействовать с группами лизина, а также эпсилон-аминогруппой аргинина.
Специфичная модификация остатков тирозила может быть осуществлена в случае особого интереса во введении спектральных меток в остатки тирозила посредством взаимодействия с ароматическими соединениями диазония или тетранитрометаном. В большинстве случаев используют N-ацетилимидазол и тетранитрометан для образования O-ацетилтирозила и 3-нитропроизводных, соответственно. Остатки тирозила подвергают йодированию с использованием 125I или 131I, чтобы получить меченые белки для применения в радиоиммуноанализе.
Карбоксильные боковые группы (аспартил или глутамил) избирательно модифицируют посредством взаимодействия с карбодиимидами (R-N=C=N-R'), где R и R' означают разные алкильные группы, такими как 1-циклогексил-3-(2-морфолинил-4-этил)карбодиимид или 1-этил-3-(4-азониа-4,4-диметилпентил)карбодиимид. Кроме того, остатки аспартила и глутамила превращают в остатки аспарагинила и глутаминила в результате взаимодействия с ионами аммония.
Остатки глутаминила и аспарагинила часто деамидируют до соответствующих остатков глутамила и аспартила, соответственно. Указанные остатки деамидируют в нейтральных или основных условиях. Деамидированная форма указанных остатков входит в объем настоящего изобретения.
Другие модификации включают гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп остатков серила или треонила, метилирование α-аминогрупп боковых цепей лизина, аргинина и гистидина (T.E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman and Co., San Francisco, pp.79-86 (1983)), ацетилирование N-концевого амина и амидирование любой C-концевой карбоксильной группы.
Другой тип ковалентной модификации заключается в химическом или ферментативном связывании гликозидов с полипептидом согласно изобретению. Такие способы имеют преимущества, так как они не требуют получения полипептида в клетке-хозяине, которая обладает способностью к гликозилированию, в частности, к N- или O-связанному гликозилированию. В зависимости от используемого способа связывания сахар(а) может быть связан с (a) аргинином и гистидином, (b) свободными карбоксильными группами, (c) свободными сульфгидрильными группами, такими как сульфгидрильные группы цистеина, (d) свободными гидроксильными группами, такими как гидроксильные группы серина, треонина или гидроксипролина, (e) ароматическими остатками, такими как ароматические остатки фенилаланина, тирозина или триптофана, или (f) амидной группой глутамина. Такие способы описаны в WO 87/05330, опубликованной 11 сентября 1987, и в Aplin and Wriston, CRC Crit. Rev. Biochem., pp.259-306 (1981).
Удаление любых углеводных остатков, присутствующих на полипептиде согласно изобретению, можно осуществить химически или ферментативно. Химическое дегликозилирование требует воздействия на полипептид трифторметансульфоновой кислотой или эквивалентным соединением. Такая обработка приводит к отщеплению большинства или всех сахаров, за исключением связывающего сахара (N-ацетилглюкозамин или N-ацетилгалактозамин), оставляя при этом полипептид интактным. Химическое дегликозилирование описано в Hakimuddin, et al. Arch. Biochem. Biophys. 259: 52 (1987) и в Edge et al. Anal. Biochem., 118: 131 (1981). Ферментативное отщепление углеводных остатков, например, от антитела, можно осуществить с использованием множества эндо- и экзогликозидаз, которые описаны в Thotakura et al. Meth. Enzymol. 138: 350 (1987).
Другой тип ковалентной модификации полипептида согласно изобретению включает в себя связывание полипептида с одним или несколькими небелковыми полимерами, например, полиэтиленгликолем, полипропиленгликолем или полиоксиалкиленами таким образом, как указано в патентах США №4640835, 4496689, 4301144, 4670417, 4791192 или 4179337.
Фармацевтические композиции
Терапевтические препараты молекул согласно изобретению, ANGPTL4, агониста ANGPTL4 или антагониста ANGPTL4, применяемые согласно изобретению, готовят для хранения смешиванием вещества, например, полипептида, имеющего требуемую степень чистоты, с необязательными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в форме лиофилизованных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в применяемых дозах и концентрациях и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол и м-крезол); полипептиды с низкой молекулярной массой (примерно меньше 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как TweenTM, PluronicsTM или полиэтиленгликоль (ПЭГ).
Активные ингредиенты также могут быть заключены в микрокапсулы, приготовленные, например, способами коацервации или полимеризации на границе фаз, например, микрокапсулы из гидроксиметилцеллюлозы или желатиновые микрокапсулы и микрокапсулы из поли(метилметакрилата), соответственно, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences, 16th edition, Osol, A., Ed., (1980). См., также Johnson et al., Nat. Med., 2: 795-799 (1996); Yasuda, Biomed. Ther., 27: 1221-1223 (1993); Hora et al., Bio/Technology, 8: 755-758 (1990); Cleland, «Design и Production of Single Immunization Vaccines Using Polylactide Polyglycolide Microsphere Systems» in Vaccine Design: The Subunit and Adjuvant Approach, Powell and Newman, eds. (Plenum Press: New York, 1995), pp.439-462; WO 97/03692, WO 96/40072, WO 96/07399 и патент США №5654010.
В некоторых вариантах препараты, используемые для введения in vivo, являются стерильными. Это легко осуществить фильтрованием через стерильные фильтрующие мембраны.
Могут быть приготовлены препараты длительного высвобождения. Подходящие примеры препаратов длительного высвобождения включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие полипептид согласно изобретению, и такие матрицы имеют определенную форму, например, пленки или микрокапсулы. Примеры матриц для длительного высвобождения включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, не разрушаемый этилен-винилацетат, разрушаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и ацетата лейпролида), сополимер молочной и гликолевой кислот (PLGA) и поли-D-(-)-3-гидроксимасляная кислота. В то время как такие полимеры, как этилен-винилацетат и молочная кислота-гликолевая кислота способны высвобождать молекулы в течение около 100 дней, некоторые гидрогели высвобождают белки в течение более коротких периодов времени. В том случае, когда инкапсулированные антитела остаются в организме в течение длительного времени, они могут денатурировать или агрегировать в результате воздействия влаги при 37°C, что приводит к потере биологической активности и возможным изменениям в иммуногенности. Могут быть разработаны стандартные методики для стабилизации, в зависимости от вовлекаемого механизма. Например, если обнаружено, что механизмом агрегации является образование межмолекулярной S-S-связи в результате тиодисульфидного обмена, то стабилизация может быть достигнута посредством модификации сульфгидрильных остатков, лиофилизацией из кислых растворов, регуляцией влагосодержания, использованием соответствующих добавок и разработкой специальных композиций на основе полимерной матрицы. См. также, например, патент США №6699501, в котором описаны капсулы с полиэлектролитным покрытием.
Кроме того, предполагается, что терапевтическое белковое средство согласно изобретению (ANGPTL4, агонист ANGPTL4 или антагонист ANGPTL4) может быть введено субъекту способом генной терапии. Генная терапия относится к терапии, осуществляемой введением пациенту нуклеиновой кислоты. В случае применений генной терапии гены вводят в клетки, чтобы добиться синтеза in vivo терапевтически эффективного генетического продукта, например, для замены дефектного гена. «Генная терапия» включает в себя как традиционную генную терапию, когда длительный эффект достигается однократной обработкой, и введение генных терапевтических средств, которое заключается в однократном или многократном введении терапевтически эффективной ДНК или мРНК. Антисмысловые РНК и ДНК можно использовать в качестве терапевтических средств для блокирования экспрессии некоторых генов in vivo. См., например, Ad-ANGPTL4-миРНК, описанные в данной публикации. Уже было показано, что короткие антисмысловые олигонуклеотиды могут быть импортированы в клетки, где они действуют в качестве ингибиторов, несмотря на их низкие внутриклеточные концентрации, вызванные их ограниченным поглощением через клеточную мембрану (Zamecnik et al., Proc. Natl. Acad. Sci. USA 83: 4143-4146 (1986)). Олигонуклеотиды могут быть модифицированы для усиления их поглощения, например, заменой их отрицательно заряженных фосфодиэфирных групп незаряженными группами. Общий обзор способов генной терапии см., например, в Goldspiel et al. Clinical Pharmacy 12: 488-505 (1993); Wu and Wu Biotherapy 3: 87-95 (1991); Tolstoshev Ann. Rev. Pharmacol. Toxicol. 32: 573-596 (1993); Mulligan Science 260: 926-932 (1993); Morgan and Anderson Ann. Rev. Biochem. 62: 191-217 (1993); и в May TIBTECH 11: 155-215 (1993). Способы, общеизвестные в области технологии рекомбинантной ДНК, которые можно использовать, описаны в Ausubel et al. eds. (1993) Current Protocols in Molecular Biology, John Wiley & Sons, NY; и Kriegler (1990) Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY.
Существует множество способов, доступных для введения нуклеиновых кислот в жизнеспособные клетки. Способы варьируют в зависимости от того, переносят ли нуклеиновую кислоту в культивируемые клетки in vitro или в клетки предполагаемого хозяина in vivo. Способы, подходящие для переноса нуклеиновой кислоты в клетки млекопитающих in vitro, включают применение липосом, электропорации, микроинъекции, слияния клеток, DEAE-декстрана, способа преципитации фосфатом кальция и т.д. Например, способы переноса генов in vivo включают, без ограничения, например, трансфекцию вирусных (обычно ретровирусных) векторов и трансфекцию, опосредованную липосомами, содержащими белок вирусного капсида (Dzau et al., Trends in Biotechnology 11, 205-210 (1993)). Например, способы переноса нуклеиновой кислоты in vivo включают трансфекцию с помощью вирусных векторов (таких как аденовирус, вирус простого герпеса I, лентивирус, ретровирус или аденоассоциированный вирус) и основанные на липидах системы (применимыми липидами для опосредованного липидами переноса гена являются, например, DOTMA, DOPE и DC-Chol). Примеры применимых вирусных векторов в генной терапии можно найти в Clowes et al. J. Clin. Invest. 93: 644-651 (1994); Kiem et al. Blood 83: 1467-1473 (1994); Salmons and Gunzberg Human Gene Therapy 4: 129-141 (1993); Grossman and Wilson Curr. Opin. in Genetics and Devel. 3: 110-114 (1993); Bout et al. Human Gene Therapy 5: 3-10 (1994); Rosenfeld et al. Science 252: 431-434 (1991); Rosenfeld et al. Cell 68: 143-155 (1992); Mastrangeli et al. J. Clin. Invest. 91: 225-234 (1993); и Walsh et al. Proc. Soc. Exp.Biol. Med. 204: 289-300 (1993).
В некоторых случаях желательно доставлять источник нуклеиновой кислоты с использованием агента, который направлен к клеткам-мишеням, такого как антитело, специфичное для мембранного белка клеточной поверхности или для клетки-мишени, лиганд рецептора на клетке-мишени и т.д. В случае использования липосом могут быть использованы белки, которые связываются с мембранным белком клеточной поверхности, связанным с эндоцитозом, для направления к мишени и/или облегчения поглощения, например, белки капсида или их фрагменты, тропные к конкретному типу клеток, антитела к белкам, которые подвергаются интернализации при прохождении цикла, белки, которые направляют внутрь клеток и повышают время полужизни в клетках. Способ опосредованного рецепторами эндоцитоза описан, например, Wu et al., J. Biol. Chem. 262, 4429-4432 (1987); и Wagner et al., Proc. Natl. Acad. Sci. USA 87, 3410-3414 (1990). Обзор протоколов мечения генов и генной терапии см. в Anderson et al., Science 256, 808-813 (1992).
Дозировка и введение
Дозировки и требуемые концентрации лекарственного средства фармацевтических композиций согласно изобретению могут варьировать в зависимости от конкретного предполагаемого применения. Определение соответствующей дозы или пути введения входит в компетенцию лечащего врача. Эксперименты на животных дают надежные рекомендации для определения эффективных доз для терапии человека. Межвидовое масштабирование эффективных доз можно осуществить, следуя принципам, установленным Mordenti, J. and Chappell, W. «The use of interspecies scaling in toxicokinetics» In Toxicokinetics and New Drug Development, Yacobi et al., Eds., Pergamon Press, New York 1989, pp.42-96.
В зависимости от типа и тяжести заболевания от примерно 1 мкг/кг до 50 мг/кг (например, 0,1-20 мг/кг) ANGPTL4, агониста ANGPTL4 или антагониста ANGPTL4 является исходной пригодной дозой для введения пациенту, независимо от того, является ли введение однократным или представляет собой несколько отдельных введений или непрерывную инфузию. При использовании введения ANGPTL4 или его агониста или антагониста in vivo нормальная доза может варьировать от примерно 10 нг/кг вплоть до 100 мг/кг массы тела млекопитающего или более в сутки, предпочтительно от примерно 1 мкг/кг/сутки до 10 мг/кг/сутки, в зависимости от пути введения. Рекомендации по конкретным дозам и способам доставки предлагаются в литературе; см., например, патенты США №4657760, 5206344 или 5225212. Предполагается, что разные препараты будут эффективными в случае разных терапевтических соединений и разных расстройств, что для введения, мишенью которого является один орган или ткань, например, может быть необходимой доставка способом, отличным от способа доставки в другой орган или ткань. В случае многократных введений в течение нескольких дней или более в зависимости от состояния лечение продолжают вплоть до того, как происходит требуемое подавление симптомов заболевания. Однако могут быть применимы другие схемы дозирования. Обычно врач будет вводить молекулу(ы) согласно изобретению до достижения дозы(доз), которая обеспечивает требуемый биологический эффект. Ход лечения согласно изобретению легко контролируют, используя обычные способы и анализы.
Терапевтическая композиция согласно изобретению может быть введена любым подходящим способом, включая, но не ограничиваясь указанным, парентеральное, подкожное, внутрибрюшинное, внутрилегочное, интрацереброспинальное, подкожное, внутрисуставное, интрасиновиальное, интратекальное, пероральное, местное и интраназальное введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Кроме того, терапевтическую композицию соответствующим образом вводят импульсной инфузией, в частности, со снижающимися дозами антитела. В некоторых вариантах терапевтическую композицию вводят посредством инъекций, например внутривенных или подкожных инъекций, отчасти в зависимости от того, является ли введение кратким или хроническим.
Как описано в данной публикации, ANGPTL4, агонист ANGPTL4 или антагонист ANGPTL4 можно комбинировать с одним или несколькими терапевтическими средствами. Комбинированное введение включает совместное введение с использованием отдельных препаратов или одного фармацевтического препарата и последовательное введение в любом порядке. Применение нескольких средств также включено в объем изобретения. Например, ANGPTL4 или агонист ANGPTL4 может предшествовать, следовать за, чередоваться с введением дополнительного терапевтического средства или может быть введен одновременно с ним. В одном варианте существует период времени, в течение которого оба (или все) активные средства одновременно проявляют свои биологические активности.
В некоторых вариантах лечение согласно изобретению заключается в комбинированном введении антагониста ANGPTL4 и одного или нескольких терапевтических средств. Изобретение также охватывает введение множества ингибиторов. Комбинированное введение включает совместное введение с использованием отдельных препаратов или одного фармацевтического препарата и последовательное введение в любом порядке. Например, антагонист ANGPTL4 может предшествовать, следовать за, чередоваться с введением дополнительного терапевтического средства или может быть введен одновременно с ним. В одном варианте существует период времени, в течение которого оба (или все) активные средства одновременно проявляют свои биологические активности.
Для профилактики или лечения заболевания подходящая доза ANGPTL4, агониста ANGPTL4 или антагониста ANGPTL4 будет зависеть от типа заболеваний, подвергаемых лечению, которые описаны выше, тяжести и течения заболевания, от того, вводят ли средство для профилактических или терапевтических целей, предыдущей терапии, истории болезни пациента и ответа на средство и решения лечащего врача. Средство соответствующим образом вводят пациенту одновременно или в виде серии обработок. В схеме комбинированной терапии композиции согласно изобретению вводят в терапевтически эффективном количестве или терапевтически синергическом количестве. В используемом в данном описании смысле терапевтически эффективное количество является таким, что совместное введение ANGPTL4, агониста ANGPTL4 или антагониста ANGPTL4 и одного или нескольких других терапевтических средств или введение композиции согласно изобретению приводит к снижению или ингибированию целевого заболевания или состояния. Терапевтически синергетическим количеством является такое количество ANGPTL4, агониста ANGPTL4 или антагониста ANGPTL4 и одного или нескольких других терапевтических средств, например, описанных в данной публикации, которое необходимо для того, чтобы синергетически или существенно уменьшить или исключить состояния или симптомы, связанные с конкретным заболеванием.
Изделия
В другом варианте осуществления изобретения предлагаются промышленные изделия, содержащие вещества, применимые в способах и для лечения расстройств, описанных выше. Промышленное изделие содержит емкость, этикетку и вкладыш в упаковку. Подходящие емкости включают, например, бутылки, флаконы, шприцы и т.д. Емкости могут быть сделаны из множества материалов, таких как стекло или пластмасса. В емкости находится композиция, которая является эффективной для лечения состояния, и может иметь стерильное входное отверстие (например, емкость может представлять собой мешок с внутривенным раствором или флакон, имеющий пробку, прокалываемую иглой для подкожной инъекции). По меньшей мере, одним активным средством в композиции является ANGPTL4, агонист ANGPTL4 или антагонист ANGPTL4. Ярлык на емкости или прикрепленный к емкости указывает, что композиция применима для лечения выбранного состояния. Изделие может дополнительно содержать вторую емкость, содержащую фармацевтически приемлемый буфер, такой как фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Изделие может дополнительно содержать другие материалы, желательные с коммерческой точки зрения и точки зрения пользователя, включая другие буферы, разбавители, наполнители, иглы и шприцы. Необязательно, вкладывают набор инструкций, обычно письменных инструкций, которые относятся к применению и дозам ANGPTL4, агониста или антагониста в случае расстройства, описанного в данной публикации. Инструкции, вложенные в набор, обычно содержат информацию о дозе, схеме дозирования и пути введения для лечения расстройства. Емкости с ANGPTL4, агонистом ANGPTL4 или антагонистом ANGPTL4 могут представлять собой стандартные дозы, упаковки большой массы (например, упаковки с множеством доз) или части стандартной дозы.
Депозит материалов
Следующий материал был депонирован в Американской коллекции типов культур, 10801 University Boulevard, Manassas, VA. 20110-2209, USA (ATCC):
Депонирование осуществляли по условиям Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры и инструкций к нему (Будапештский договор). Он гарантирует сохранение жизнеспособной культуры депозита в течение 30 лет от даты депозита. Депозиты могут стать доступными из ATCC по условиям Будапештского договора и по условиям соглашения между Genentech, Inc. и ATCC, которое гарантирует постоянную и неограниченную доступность потомства культуры депозита для общественности после опубликования соответствующего патента США или после выкладки для общего доступа любой заявки на выдачу патента США или иностранного патента, какой бы ни поступил первым, и гарантирует доступность потомства лицу, определяемому комиссаром по патентам и торговым маркам США как имеющему на это право согласно 35 USC § 122 и соответствующим правилам для комиссара (включая 37 CFR § 1.14 с конкретной ссылкой на 886 OG 638).
Правопреемник настоящей заявки согласен, что если культура материалов на депонировании погибнет, или потеряется, или нарушится при культивировании в подходящих условиях, то материалы при уведомлении будут быстро заменены другими такими же материалами. Доступность депонированного материала не следует толковать как разрешение на практическое осуществление изобретения в нарушение прав, гарантированных полномочиями любого правительства в соответствии с его патентным законодательством.
ПРИМЕРЫ
Пример 1: ANGPTL4 индуцирует клеточную адгезию и пролиферацию гепатоцитов человека
Получение аденовирусных векторов и трансдукция: Аденовирусные конструкции были сконструированы посредством клонирования Not1-Not1-вставки кДНК в полилинкерном сайте с использованием наборов для конструирования вектора Ad-easy из Stratagene (LaJolla, CA), по существу, как описано производителем. См., например, Hesser et al., Blood, 104(1): 149-158 (2004).
Получение меченного одной flag-меткой белка hAngptl4(23-406) (PUR9384), mAngptl4(184-410)-IgG (PUR9388) и mAngptl4(23-410) (PUR9452): Собранную жидкость из культур клеток пропускали в течение ночи через смолу анти-flag M2 (Sigma №A-2220). Колонку промывали до базовой линии PBS, затем элюировали 50 мМ цитратом Na pH 3,0. Полученный объем концентрировали на Amicon-15 с пределом отсечки по молекулярной массе 10000 (Millipore №UFC901024). Конечная стадия представляла собой диализ в 1 мМ HCl/ H2O Super Q и фильтрование через 0,2 мкм-фильтры. SDS-ПААГ-гель в 4-20% трис/глицине (Invitrogen # EC6028box)+/- 10 мМ DTT использовали для определения чистоты. Правильные белки идентифицировали, либо получая масс-спектры, либо N-концевым секвенированием по Эдману.
Получение hAngptl4(184-406)-IgG (PUR 9441) с N-концевой flag-меткой, за которой последовательно располагается N-концевая hu-Fc-метка: Собранную жидкость из культуры клеток пропускали в течение ночи через ProSep A (Amersham №113111835). Колонку промывали до базовой линии PBS. Затем осуществляли стадию промывки четырьмя объемам колонки 0,5 М TMAC/PBS pH 7,5 с последующей промывкой PBS до базовой линии. Стадию элюирования осуществляли в 50 мМ цитрате Na pH 3,0. Полученный объем концентрировали на Amicon-15 с пределом отсечения по молекулярной массе 10000 (Millipore №UFC901024). Конечная стадия представляла собой диализ в 1 мМ HCl/H2O Super Q и фильтрование через 0,2 мкм-фильтры. SDS-ПААГ-гель в 4-20% трис/глицине (Invitrogen №EC6028box)+/- 10 мМ DTT использовали для определения чистоты. Правильные белки идентифицировали, либо получая масс-спектры, либо N-концевым секвенированием по Эдману. Рекомбинантные белки также можно получить, используя стандартные способы, известные в данной области.
Создание Ad-ANGPTL4-миРНК: 4 возможные молекулы ANGPTL4-миРНК (Qiagen) создавали на основе полноразмерной последовательности hANGPTL4. Одну ANGPTL4-миРНК отбирали на основе способности миРНК ингибировать экспрессию hANGPTL4. Она была направлена к следующей последовательности ДНК-мишени GTGGCCAAGCCTGCCCGAAGA (SEQ ID NO: 3) ANGPTL4, например, r(GGCCAAGCCUGCCCGAAGAUU) (SEQ ID NO: 4) и/или r(UCUUCGGGCAGGCUUGGCCAC) (SEQ ID NO: 5). МиРНК клонировали в векторе для переноса CMVpShuttle-H1.1 с РНК-промотором, например, промотором H1 (GenScript). Затем клонировали кассету экспрессии миРНК, чтобы создать аденовирусную конструкцию AdhANGPTL4-миРНК. Аденовирусные конструкции получали клонированием Not1-Not1-вставки кДНК в полилинкерном сайте, используя наборы для конструирования вектора Ad-easy из Stratagene (LaJolla, CA), по существу, как описано производителем. См., например, Hesser et al., Blood, 104(1): 149-158 (2004).
Экспрессию ANGPTL4 подтверждали Вестерн-блот-анализом, используя анти-FLAG-антитело. Отбирали один клон с высокой экспрессией и титры увеличивали согласно инструкции производителя. Вирусные препараты очищали центрифугированием в CsCl и тестировали в отношении ревертантов с помощью ПЦР. Титры вирусов определяли в экспериментах по лизированию клеток в 96-луночных планшетах согласно инструкциям производителя. Указанные векторы наряду с поставляемым pShuttleCMV-lacZ рекомбинировали в электрокомпетентных бактериях BJ5183 с вектором AdEasy, содержащим геном Ad5, делетированный в областях E1 и E3. Первичные вирусные штаммы получали, используя временную трансфекцию рекомбинантных плазмид AdEasy в клетки-хозяева HEK293. Аденовирусные штаммы дополнительно амплифицировали в клетках HEK293 и очищали, используя способ очистки в градиенте CsCl, как описано производителем. Рабочие титры аденовируса получали в анализе ELISA.
Получение mANGPTL4: Клетки 293 временно трансфицировали конструкцией, которая содержала нуклеиновую кислоту, кодирующую полноразмерный mANGPTL4 (1-410). mANGPTL4 очищали из надосадка и использовали для экспериментов.
Клеточная адгезия гепатоцитов: Способность ANGPTL4 индуцировать клеточную адгезию первичных гепатоцитов оценивали в 96-луночных планшетах. Планшеты покрывали подпоследовательностью 23-410 мышиного Angptl4, фибронектином или контрольным белком NL4 в разных концентрациях, например, без покрытия, 0,3 мкг/мл, 3,0 мкг/мл или 30 мкг/мл, в 60 мкл при 4°C в течение ночи. Избыток белка удаляли и покрытые лунки блокировали 200 мкл 3% БСА в PBS при 37°C в течение 1 часа. После инкубации надосадок отсасывали и один раз промывали PBS.
Получали первичные гепатоциты человека и выращивали в нормальной ростовой среде (Cambrex). Клетки 3 раза промывали PBS. Клетки обрабатывали трипсином, а затем раствором для нейтрализации трипсина (Clonetics). Затем клетки ресуспендировали в нормальной ростовой среде (Cambrex). Клетки высевали по 1,5×104 клеток/лунку в общем объеме 200 мкл. Клетки делили в 5% сыворотке за 24 часа до внесения дозы. Клетки инкубировали в лунках при 37°C в течение 2 часов. Надосадок удаляли. Прикрепление клеток измеряли, используя анализ с кристаллическим фиолетовым. В лунку добавляли 50 мкл 10% раствора формалина и фиксировали в течение 10 минут.Клетки осторожно один раз промывали PBS. Добавляли 50 мкл 0,5% раствора кристаллического фиолетового, который перед использованием фильтровали. Раствор инкубировали в лунках в течение 30 минут или больше при комнатной температуре. Лунки промывали 3-5 раз в PBS. Из лунок удаляли PBS и сушили. 96-луночный планшет регистрировали при OD550. См. фиг.4. Также можно использовать способ PNAG согласно Landegren. См. Landegren, U. (1984) J. Immunol. Methods 67: 379-388. Как видно на фиг.4, рекомбинантный mAngptl4 (23-410) индуцирует клеточную адгезию первичных гепатоцитов in vitro.
Пролиферация гепатоцитов: Исследовали влияние Angptl4 на пролиферацию первичных гепатоцитов человека. Получали аденовирусные конструкции Ad-человеческий(h)Angptl4, Ad-LacZ и Ad-Angptl3. См., например, Hesser et al, Blood 104(1): 149-158 (2004). Первичные гепатоциты человека трансдуцировали любой из конструкций, включая конструкцию аденовирус-Angptl4 (Ad-Angptl4), конструкцию аденовирус-LacZ (Ad-LacZ) в качестве контроля или конструкцию аденовирус-Angptl3 (Ad-Angptl3) при множественности инфекции (MOI) 10, 100 и 1000. После 5 дней роста гепатоцитов в нормальной среде для роста гепатоцитов (Cambrex) проводили подсчет клеток. Как указано на фиг.5, Ad-Angptl4 индуцирует пролиферацию гепатоцитов in vitro при MOI, равной 10.
Пример 2: ANGPTL4 индуцирует пролиферацию преадипоцитов
Пролиферация преадипоцитов: Оценивали способность ANGPTL4 индуцировать пролиферацию преадипоцитов. Преадипоциты человека (висцеральные или подкожные) выращивали на 6-луночных чашках (Falcon, Primaria), деля клетки до плотности 30000 клеток/лунку в объеме 3 мл ростовой среды, содержащей сыворотку (среда роста преадипоцитов (Cambrex)). Затем непосредственно после посева клеток добавляли 500 мкл кондиционированной среды из культуры клеток COS, которые были трансдуцированы аденовирусными конструкциями, например, Ad-LacZ (4), Ad-человеческий(h)Angptl4 (23-406) (5) или Ad-человеческий(h)Angptl3 (полноразмерный белок) (6), или рекомбинантные белки (рекомбинантный мышиный Angptl4 (23-410) (2); IgG-mAngptl4 (184-410) (3) или без добавок (1) в следующих концентрациях (rmAngptl4 (23-410) (5 мкг/мл); IgG-mAngptl4 (5 мкг/мл)). Клетки выращивали в течение 5 дней при 37°C в инкубаторе с 5% CO2. Клетки трипсинизировали, используя 500 мкл 1× трипсина в течение 3-5 минут. Смесь клеток (0,5 мл) пипетировали в 9,5 мл изотоничного буферного раствора и считали в устройстве для подсчета клеток (учитывая фактор разведения 20). Как показано на фиг.6, панель A, и рекомбинантный мышиный Angptl4 (23-410) (2) и кондиционированная среда клеток COS, содержащая hAngptl4 (23-406) (5), индуцируют пролиферацию первичных висцеральных преадипоцитов человека. Панель B на фиг.6 иллюстрирует, что и рекомбинантный мышиный Angptl4 (23-410) (2) и кондиционированная среда клеток COS, содержащая hAngptl4(23-406) (5), индуцируют пролиферацию первичных подкожных преадипоцитов человека.
FACS-анализ связывания Angptl4 с первичными адипоцитами человека: Связывание ANGPTL4 с первичными адипоцитами человека исследовали, используя FACS-анализ. Первичные подкожные адипоциты человека высевали в чашки для культивирования диаметром 10 см при плотности 500000 - 1×106 клеток/лунку с образцом. Клетки разделяли за день до FACS. Клетки один раз промывали PBS и затем добавляли 10 мл 20 мМ EDTA в PBS и инкубировали в течение 10-20 минут. Через 20 минут клетки соскабливали из планшета. Добавляли 10 мл 5% FCS в PBS и клетки переносили в пробирку Falcon объемом 50 мл. Клетки осаждали центрифугированием при 1,8 тыс.об/мин в течение 5 минут при 4°C. Надосадок удаляли и клетки ресуспендировали в 1 мл 5% FCS в PBS. По 100 мкл суспензии клеток распределяли в пробирки для FACS объемом 5 мл, содержащие 1 мкг белка, и инкубировали в течение 30 минут или больше на льду. Использовали следующие белки: mAngptl4 (23-410), PUR 9452, 0,428 мг/мл (2 мкл/образец); hAngptl4 (23-406), PUR 9384,+/-90 мкг/мл (10 мкл/образец); mAngptl4 (184-410)-IgG, PUR 9388, 8,5 мг/мл (0,2 мкл/образец); hAngptl4 (184-406)-IgG, PUR 9441, 1,5 мг/мл (1 мкл/образец); и контрольный FLAG-BAP (Sigma) 0,1 мг/мл (2 мкл/образец). После инкубации пробирки заполняли 5 мл 5% FCS в PBS на льду. Клетки осаждали центрифугированием в течение 5 минут при 2 тыс.об/мин. Надосадок удаляли. Добавляли анти-FLAG-FITC-антитело (Sigma) (2 мкл антитела (100 мкг/мл исходного раствора) и инкубировали на льду в течение 5 минут или больше. Конечная концентрация антитела составляла 1 мкг/мл. Добавляли 5 мл 5% FCS в PBS и клетки осаждали центрифугированием в течение 5 минут при 1,8 тыс.об/мин при 4°C. Надосадок удаляли и клетки ресуспендировали в 0,25 мл PBS с 5% FCS на льду. Также может присутствовать 0,05% азид натрия, чтобы предотвратить интернализацию рецептора. Добавляли 1 мкл разбавленного 1:50 исходного раствора йодида пропидия (PI) на образец. Затем клетки подвергали FACS-анализу. Как указано на фиг.7, при таких условиях обе формы Angptl4 человека, rhAngptl4 (23-406) и rhIgG-hAngptl4 (184-406), связывались более эффективно с подкожными адипоцитами по сравнению с мышиным ортологом.
ПРИМЕР 3: Angptl4 индуцирует миграцию первичных подкожных преадипоцитов человека
Angptl4 индуцирует клеточную миграцию: Авторы исследовали способность Angptl4 индуцировать клеточную миграцию первичных подкожных преадипоцитов человека. Подвижность клеток измеряли, как описано ранее (см., например, Camenish et al., J. Biol. Chem., 277(19): 17281-17290 (2002)), используя многолуночные вкладыши для культуры ткани HTS с размером пор 3 мкм (Becton Dickinson, NJ). hANGPTL4 (1-406) разбавляли в 50/50/0,1% БСА до 5, 1 и 0,2 мкг/мл. В качестве позитивного контроля мембраны инкубировали либо со средой, содержащей 10% фетальной сыворотки теленка (FCS), либо 0,1 мкг/мл рекомбинантного PDGF-BB человека (R & D Systems). PBM/0,1% БСА использовали в качестве негативного контроля. Первичные адипоциты человека три раза промывали PBS, собирали и суспендировали примерно в концентрации 2-5×105 клеток/мл в PBM/0,1% БСА. Тестировали следующие препараты клеток, где ANGPTL4 указан как NL2.
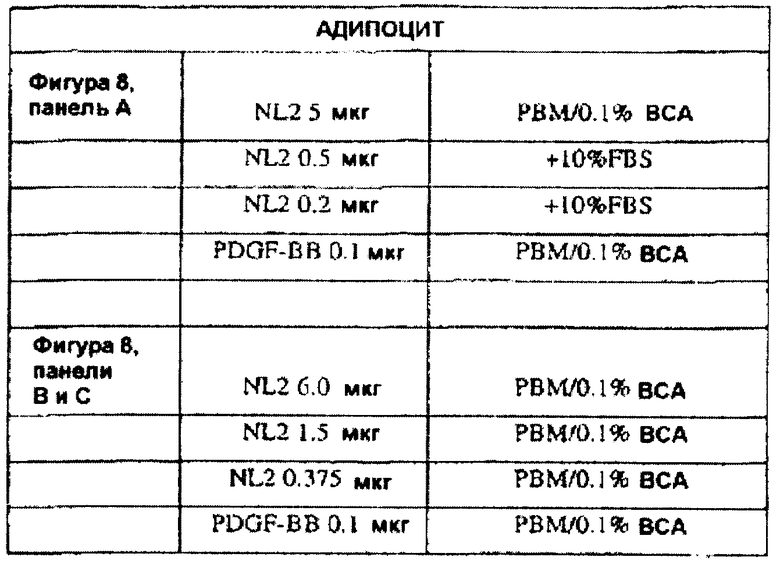
Препараты добавляли в нижнюю камеру и препараты инкубировали при 37°C в течение 19 часов.
Суспензию клеток (250 мкл) добавляли в верхнюю камеру и клеткам давали возможность мигрировать в течение ночи при 37°C в инкубаторе во влажной атмосфере с 5% CO2. После инкубации среду отсасывали как из верхней, так и из нижней камер и клетки, которые мигрировали к нижней поверхности мембраны, фиксировали метанолом (400 мкл MeOH в течение 30 минут при 4°C, удаляли MeOH и сушили на воздухе в течение 40 минут) и красили YO-PRO-1-йодидом (Molecular Probes, OR) (400 мкл YO-PRO-1-йодида в концентрации 10 мкМ (1:100 из 1 мМ исходного раствора)). Результаты миграции количественно оценивали в виде среднего количества клеток/поле микроскопа при 20-кратном увеличении, используя компьютерную программу Openlab (Improvision, MA). Как видно на фиг.8, панель A, hAngptl4, добавленный к первичным подкожным преадипоцитам человека, индуцирует их миграцию. Панель B на фиг.8 иллюстрирует миграцию через 7 часов. Панель C на фиг.8 графически иллюстрирует миграцию адипоцитов после 7 часов обработки без сыворотки (1), 10% фетальной сывороткой теленка (FCS) (2), PDGF-BB (3), mANGPTL4 (4).
Пример 4: Вариант Angptl4
Вариант ANGPTL4 получали, используя стандартный набор для мутагенеза (например, набор для сайт-специфичного мутагенеза QuikChange XL (Invitrogen, Carlsbad, California)), следуя протоколу производителя. Осуществляли две аминокислотных замены в последовательности ANGPTL4 человека (см., например, фиг.2). Получали замены в положении 162 и 164 (R162G и R164E), приводящие к замене RKR на GKE. Белок ANGPTL4 (L280-плазмида, а/к 1-406) или вариант ANGPTL4 выделяли из надосадка временно трансфицированных клеток COS-7. Для очистки надосадок наносили на колонку с никелем. Белок выявляли Вестерн-блот-анализом с помощью анти-FLAG-HRP-антитела. См. фиг.3, панель B. В том случае, когда получали замены и вариант ANGPTL4 сравнивали с нативным белком или белком ANGPTL4 дикого типа, обнаружили, что вариант ANGPTL4 имеет более высокую молекулярную массу, чем нативный ANGPTL4 при Вестерн-блоттинге. Замена RKR на GKE в положении 162 и 164 нативного белка предотвращала протеолитическое разрушение ANGPTL4.
Пример 5: Angptl4 связывается с интегрином α V β 5
Ангиопоэтины являются секретируемыми факторами, которые регулируют ангиогенез в результате связывания со специфичным для эндотелиальных клеток тирозинкиназным рецептором Tie2 посредством фибриноген (FBN)-подобного домена. Обнаружено, что биспиральный домен, присутствующий в семействе секретируемых лигандов, необходим для олигомеризации лиганда (см., например, Procopio et al., J. Biol. Chem., 274: 30196-201(1999)).
Подобно ангиопоэтинам ANGPTL3 и ANGPTL4 являются секретируемыми гликопротеинами, каждый из которых состоит из N-концевого сигнального пептида, за которым следует биспиральный домен и C-концевой FBN-подобный домен. Было определено, что ANGPTL3 связывается с αVβ3 посредством FBN-подобного домена. Авторы определили, что ANGPTL4 связывается с αVβ5. Тестировали линию клеток 293-1953, которые стабильно трансфицированы интегрином αVβ5, в отношении способности связываться или прилипать к планшетам, покрытым ANGPTL4. Клетки собирали и разбавляли до 105 клеток/мл в бессывороточной среде, содержащей PBS, 1% БСА, 1 мМ CaCl2 и 1 мМ MgCl2. Клетки предварительно инкубировали в присутствии или в отсутствие антител против интегрина αVβ5 (MAB1961 (Chemicon, Temecula, CA)) или пептидов в течение 15 минут при 37°C. Рекомбинантным mANGPTL4, БСА или витронектином (1 мкг, 3 мкг, 10 мкг или 30 мкг/мл) покрывали 96-луночные планшеты для микротитрования с плоским дном Nunc Maxisorp в течение ночи при 4°C и блокировали 200 мкл 3% БСА в фосфатно-солевом буфере (PBS), pH 7,4, в течение 1,5 часов при 37°C. Суспензии клеток (5×104 клеток/100 мкл/лунку (5×105/мл)) добавляли в покрытые лунки и планшеты инкубировали при 37°C в течение 5,5 часов. Неприкрепленные клетки удаляли промывками PBS и прикрепление клеток измеряли, добавляя 200 мкл красителя CyQuant GD (молекулярные зонды (Invitrogen detection Technologies (Carlsbad, California)) (1:400)/в буфере для лизиса клеток и инкубировали в течение 2-5 минут. Флуоресценцию образцов измеряли, используя возбуждение при 480 нм и максимум эмиссии при 520 нм. Можно использовать способ PNAG согласно Lanndegren (см., например, Landegren, J. Immunol. Methods, 67: 379-388 (1984)). Наблюдали прилипание клеток, экспрессирующих αVβ5, к ANGPTL4 и витронектину (USBiological, Swampscott, Massachusetts), позитивному контролю, по сравнению с БСА, негативным контролем. См. фиг.9, панель A.
Чтобы определить, является ли интегрин αVβ5 достаточным для того, чтобы опосредовать адгезию клеток к ANGPTL4, тестировали блокирующие антитела по их способности ингибировать адгезию в анализе адгезии клеток. Функциональные блокирующие антитела (анти-αVβ5-антитело (MAB1961 (Chemicon, Temecula, CA)) или анти-hANGPTL4-антитела) добавляли к клеткам 293-1953 перед инкубацией с покрытыми белком лунками (БСА (1), витронектин (2) или ANGPTL4(3)). См. фиг.9, панель B. Анти-αVβ5-антитела и анти-ANGPTL4-антитела отменяли адгезионную активность клеток по отношению к ANGPTL4.
Осуществляли дополнительные эксперименты, чтобы подтвердить, что ANGPTL4 связывает αVβ5. Осуществляли эксперименты ELISA, чтобы выявить, связывается ли mANGPTL4, IgG-hANGPTL4-N-конец (1-183) и/или IgG-hANGPTL4-C-конец (184-406) с планшетами, покрытыми αVβ5 (USBiological, 37K, Swampscott, Massachusetts). 100 мкл/лунку интегрина αVβ5, разбавленного буфером для покрывания (1 мкг/мл буфера для покрывания (50 мМ карбонат/бикарбонат, pH 9,6)), инкубировали в течение ночи при 4°C. Планшеты три раза промывали буфером для промывки (PBS, pH 7,4, 0,05% твин-20) и добавляли 100 мкл/лунку блокирующего буфера (PBS, pH 7,4, 0,5% БСА) на 1 час при комнатной температуре и осторожном встряхивании. Различные количества (0, 0,070 мкг, 0,22 мкг, 0,66 мкг, 2 мкг или 6 мкг) образцов mANGPTL4, IgG-hANGPTL4-N-конец (1-183) и/или IgG-hANGPTL4-C-конец (184-406) готовили в буфере для образцов (0,5% БСА, 50 мМ трис, pH 7,4, 0,05% твин-20, 1 мМ MnCl2, 50 мкМ CaCl2, 50 мкМ MgCl2, 100 мМ NaCl) и инкубировали в течение 30 минут. В планшеты добавляли образцы (100 мкл/лунку в количествах, указанных выше) и инкубировали в течение 2 часов при комнатной температуре и осторожном встряхивании. Планшеты промывали буфером и добавляли 100 мкл/лунку конъюгата антитело против Flag-пероксидаза хрена (HRP) (100 нг/мл) (Jackson №109-036-098) в буфере для анализа (PBS, pH 7,4, 0,5% БСА, 0,05% твин-20) и инкубировали в течение 1 часа при комнатной температуре и осторожном встряхивании. Планшеты промывали. Добавляли 100 мкл/лунку тетраметилбензидина (TMB) (Moss, Inc.) и инкубировали в планшетах до развития хорошей окраски при комнатной температуре. Добавляли 100 мкл/лунку стоп-раствора (1 М H3PO4), чтобы остановить реакцию. Планшеты регистрировали при 630 нм. mANGPTL4, IgG-hANGPTL4-N-конец и IgG-hANGPTL4-C-конец связывались с покрытыми αVβ5 планшетами, при этом немного больше белка IgG-hANGPTL4-C-конец было связано с планшетами. См. фиг.9, панель C.
Анти-ANGPTL4-антитела ингибируют связывание ANGPTL4 с покрытыми αVβ5 планшетами. Осуществляли эксперименты ELISA. 100 мкл/лунку интегрина αVβ5, разбавленного буфером для покрывания (1 мкг/мл буфера для покрывания (50 мМ карбонат/бикарбонат, pH 9,6)), инкубировали в течение ночи при 4°C. Планшеты три раза промывали буфером для промывки (PBS, pH 7,4, 0,05% твин-20) и добавляли 100 мкл/лунку блокирующего буфера (PBS, pH 7,4, 0,5% БСА) на 1 час при комнатной температуре и осторожном встряхивании. От 0,6 мкг до 6,0 мкг образцов, mANGPTL4, IgG-hANGPTL4-N-конец (1-183) и/или IgG-hANGPTL4-C-конец (183-406) в буфере для образцов (0,5% БСА, 50 мМ трис, pH 7,4, 0,05% твин-20, 1 мМ MnCl2, 50 мкМ CaCl2, 50 мкМ MgCl2, 100 мМ NaCl) инкубировали с анти-ANGPTL4-антителами (1,5 мкг) или анти-Dscr (1,5 мкг) в течение 30 минут. После инкубации 100 мкл/лунку образца+/- антитело инкубировали в планшетах в течение 2 часов при комнатной температуре и осторожном встряхивании. Планшеты промывали буфером и добавляли 100 мкл/лунку анти-Flag-HRP (100 нг/мл) в буфере для анализа (PBS, pH 7,4, 0,5% БСА, 0,05% Твин-20) и инкубировали в течение 1 часа при комнатной температуре и осторожном встряхивании. Планшеты промывали и добавляли 100 мкл/лунку TMB и инкубировали в планшетах вплоть до развития хорошей окраски при комнатной температуре. Добавляли 100 мкл/лунку стоп-раствора (1 М H3PO4), чтобы остановить реакцию. Планшеты регистрировали при 630 нм. Анти-ANGPTL4-антитела уменьшали количество mANGPTL4, IgG-hANGPTL4-N-конец и IgG-hANGPTL4-C-конец, связывающегося с покрытыми αVβ5 планшетами, по сравнению с анти-Dscr-антителом, моноклональным антителом 5G7 или средой. См. фиг.9, панель D.
В другом эксперименте связывание ANGPTL4 и интегрина αVβ5 было показано с помощью ELISA. В таком эксперименте в планшеты добавляли 80 мкл/лунку C-концевого hANGPTL4, витронектина или БСА (5 мкг/мл) в буфере для покрывания (50 мМ карбонат/бикарбонат, pH 9,6) и инкубировали в течение ночи при 4°C. Планшеты промывали (буфер для промывки: PBS, pH 7,4, 0,05% твин-20) и добавляли 100 мкл/лунку блокирующего буфера (PBS, pH 7,4, 0,5% БСА) либо со средой, либо с анти-hANGPTL4-антителами (15 мкг/100 мкл), либо с анти-Dscr-антителами (15 мкг/100 мкл) и инкубировали в течение 1 часа при комнатной температуре и осторожном встряхивании. Планшеты промывали и добавляли 100 мкл (3-9 мкг/мл) αVβ5 и инкубировали в течение 2 часов при комнатной температуре и осторожном встряхивании. Планшеты промывали и добавляли 1 мкг/мл (1:1000) анти-αVβ5-антитела (Chemicon) (5 мкг/100 мкл) в буфере для анализа (PBS, pH 7,4, 0,5% БСА, 0,05% твин-20) и инкубировали в течение 1 часа при комнатной температуре и осторожном встряхивании. После инкубации планшеты промывали и добавляли 100 мкл/лунку конъюгированного с пероксидазой хрена (HRP) антимышиного антитела (1:5000) в буфере для анализа. Планшеты промывали и добавляли 100 мкл/лунку тетраметилбензидина (TMB) и инкубировали при комнатной температуре вплоть до развития хорошей окраски. Реакцию останавливали добавлением 100 мкл/лунку 1 М H3PO4 и планшеты регистрировали при 630 нм. αVβ5 связывается с планшетами, покрытыми ANGPTL4 (дорожка 1), и витронектином (дорожка 4). Связывание блокируется анти-ANGPTL4-антителами (дорожка 2), но не блокируется при использовании контрольного анти-Dscr-антитела (дорожка 3) или в том случае, когда планшеты покрывают контрольным белком (дорожка 5). См. фиг.9, панель E.
Следовательно, полученные данные свидетельствуют, что рекомбинантный ANGPTL4 специфично связывается с интегрином αVβ5.
Пример 6: Angptl4 увеличивает уровни триглицеридов у мышей при внутривенной инъекции.
Уровни триглицеридов определяли у мышей C57B1-6, которым инъецировали различные аденовирусные конструкции, которые содержат ANGPTL4. Мышам C57B1-6 внутривенно в хвост инъецировали либо (1) аденовирусную GFP-конструкцию, (2) аденовирусную Gd-конструкцию, (3) аденовирусную ANGPTL4 (1-406)-конструкцию, (4) аденовирусную ANGPTL4 (1-183)-конструкцию, (5) аденовирусную ANGPTL4 (184-406)-конструкцию, (6) аденовирусную конструкцию, содержащую вариант ANGPTL4; (7) аденовирусную ANGPTL4 (1-408)-конструкцию и (8) аденовирусную контрольную конструкцию. Уровни триглицеридов в (мг/дл) измеряли в образцах крови от мышей через семь дней после инъекции. Как видно на фиг.10, конструкция N-конца ANGPTL4 (1-183) оказывала наиболее выраженное воздействие на уровни триглицеридов наряду с конструкцией полноразмерного ANGPTL4 и конструкций варианта ANGPTL4.
Пример 7: Создание и анализ мышей, содержащих нарушение гена ANGPTL4
Чтобы исследовать роль ANGPTL4, получали нарушения гена ANGPTL4 посредством гомологичной рекомбинации. В частности, создавали трансгенных мышей, содержащих нарушения в гене ANGPTL4 (т.е. нокаутированных мышей), либо целенаправленным изменением гена, либо используя улавливание генов (gene trapping). Мутации подтверждали Саузерн-блот-анализом, чтобы подтвердить правильное целенаправленное изменение на обоих 5'- и 3'-концах. Также осуществляли специфичное для гена генотипирование, используя геномную ПЦР, чтобы подтвердить утрату эндогенного нативного транскрипта, которую показывали с помощью ОТ-ПЦР, используя праймеры, которые отжигаются с экзонами, фланкирующими сайт инсерции. Векторы для целенаправленного изменения электропорировали в ES-клетки линии 129 и идентифицировали целенаправленно измененные клоны. Целенаправленно измененные клоны микроинъецировали в бластоцисты хозяина, чтобы получить химеры. Химер скрещивали с животными C57, получая гетерозиготы F1. Гетерозигот взаимно скрещивали, получая потомство F2 животных дикого типа, гетерозиготных и гомозиготных, которые использовали для фенотипического анализа. Иногда, если не было получено достаточно гетерозигот F1, гетерозигот F1 скрещивали с мышами C57 дикого типа, получая достаточно гетерозигот, чтобы получить группы для анализа фенотипа. Все фенотипические анализы проводили через 12-16 недель после рождения.
Результаты
Создание и анализ мышей, содержащих нарушения в гене ANGPTL4: В экспериментах, связанных с нокаутом, нарушали ген, кодирующий ANGPTL4 (PRO197-полипептид, обозначенный как ДНК 22780-1078; UNQ171). Конкретная информация о гене для указанных исследований представляет собой следующее: мутантный ген мыши соответствует ссылке на нуклеотидную последовательность: NM_020581. Доступ: NM_020581 NID:10181163; или ангиопоэтинподобный белок 4 Mus musculus (Angptl4); ссылка на белок: Q9Z1P8. Доступ: Q9SZ1P9 NID; или Mus musculus (мышь). NG27 (ангиопоэтин-родственный белок печени) (гипотетический белок 425O18-1) (фибриноген/ангиопоэтинродственный белок) (ангиопоэтинподобный белок) (ангиопоэтинподобный 4). MOUSESTRNRDB; ссылка на последовательность гена человека: NM_139314. Доступ: NM_139314 NID:21536397 ангиопоэтинподобный 4 Homo sapiens (ANGPTL4); последовательность белка человека соответствует ссылке: Q9BY76. Доступ: Q9BY78 NID: или Homo sapiens (человека). Предшественник ангиопоэтинродственного белка 3 (ангиопоэтинподобный 4) (фибриноген/ангиопоэтинродственный белок печени) (HFARP) (ангиопоэтинподобный белок PP1158). HUMANSTRNRDB.
Нарушенным геном мыши является Angptl4 (ангиопоэтинподобный 4), который является ортологом ANGPTL4 человека. Альтернативные названия включают названия, описанные в данной публикации, и BK89, Bk89, FIAF, NG27, Ng27, HFARP, Farp-pending, фибриноген/ангиопоэтинродственный белок, область главного комплекса гистосовместимости NG27, ARP4, PGAR, PPARG, PP1158, ANGPTL2, индуцированный голоданием адипозный фактор, PPARG-ангиопоэтинродственный белок, ангиопоэтинродственный белок печени и фибриноген/ангиопоэтинродственный белок печени.
Целенаправленные или полученные улавливанием случайные генные мутации создавали в линии полученных из 129SvEVBrd эмбриональных стволовых клеток (ES). Химерных мышей скрещивали с мышами-альбиносами C57BL/6J, получая гетерозиготных животных F1. Полученное потомство скрещивали между собой, получая потомство F2 дикого типа, гетерозиготное и гомозиготное мутантное потомство. В редких случаях, например, когда от химер получали очень мало мышей F1, гетерозиготных мышей F1 скрещивали с гибридными мышами 129SvEVBrd/C57, получая дополнительное количество гетерозиготных животных для взаимного скрещивания, чтобы получить мышей F2. Фенотипический анализ осуществляли на мышах из указанного поколения, как описано ниже.
Хи-квадрат=2,76; значимость=0,26294 (гомоз./n)=0,16; средний размер приплода=7
Ретровирусная инсерция (OST) происходила с нарушением гена между кодирующими экзонами 2 и 3 (NCBI доступ NM_020581.1).
Экспрессию дикого типа гена-мишени регистрировали в эмбриональных стволовых (ES) клетках и во всех образцах тканей взрослого организма, тестируемых с использованием ОТ-ПЦР, за исключением хвоста. Анализ в ОТ-ПЦР выявил, что транскрипт отсутствует у анализируемой (-/-)-мыши.
1. Фенотипический анализ
Общие выводы о фенотипах: Мутация гена, кодирующего ортолог ангиопоэтинподобного белка 4 человека (ANGPTL4), приводила к пониженным уровням холестерина и триглицеридов у (-/-)-мышей. Кроме того, самцы (-/-)-мышей проявляли повышенную толерантность к глюкозе в тесте толерантности к глюкозе. У мутантных (-/-)-мышей также наблюдали иммунологические аномалии, включая повышенные уровни IgM в сыворотке и среднее абсолютное количество нейтрофилов, по сравнению с их однопометными (+/+)-животными. При ОТ-ПЦР транскрипт отсутствовал.
Анализ фенотипа сердечно-сосудистой системы/метаболизма - химии крови: В области сердечно-сосудистой биологии осуществляли фенотипическое тестирование, чтобы идентифицировать потенциальные мишени для лечения сердечно-сосудистых, эндотелиальных или ангиогенных расстройств, таких как гипертония, атеросклероз, сердечная недостаточность, инсульт, различные болезни коронарных артерий, дислипидемии, такие как высокий уровень холестерина (гиперхолестеринемия) и повышенные уровни триглицеридов в сыворотке (гипертриглицеридемия), злокачественная опухоль и/или ожирение. Фенотипические тесты включают измерение холестерина и триглицеридов в сыворотке. Кроме того, фенотипические анализы химии крови также включали тесты толерантности к глюкозе, чтобы измерить чувствительность к инсулину и изменения в метаболизме глюкозы. Отклоняющиеся от нормы результаты теста толерантности к глюкозе являются без ограничения показателем следующих расстройств или состояний: диабета типа 1 и типа 2, синдрома X.
Фенотипические тесты в данном случае включали измерение холестерина и триглицеридов в сыворотке.
Липиды крови
Способ: В данном анализе использовали группу из 4 самцов дикого типа и 8 гомозиготных самцов. Измеряли средние уровни холестерина и триглицеридов в сыворотке и сравнивали с совпадающими по полу однопометными (+/+)-животными. Осуществляли параллельное тестирование толерантности к глюкозе, так как указанный тест является стандартом для определения нарушенного гомеостаза глюкозы у млекопитающих. Тестирование толерантности к глюкозе осуществляли, используя глюкометр Lifescan. Животным инъецировали в/б 2 г/кг D-глюкозы, вводимой в виде 20% раствора, и уровни глюкозы в крови измеряли через 0, 30, 60 и 90 минут после инъекции. Применяли COBAS Integra 400 (Roche) при проведении тестирования химии крови у мышей.
Результаты: У самцов и самок гомозиготных мутантных мышей наблюдали заметно пониженный средний уровень триглицеридов по сравнению с совпадающими по полу однопометными животными дикого типа и по сравнению с историческими средними значениями. Такие мутанты также показали пониженные средние уровни холестерина в сыворотке по сравнению с однопометными животными дикого типа. Одновременно самцы (-/-)-мышей имели повышенную толерантность к глюкозе в присутствии нормального уровня глюкозы натощак во всех 3 тестированных интервалах по сравнению с совпадающими по полу однопометными (+/+)-животными и историческими средними значениями, тогда как самки (-/-)-мышей имели пониженный средний уровень глюкозы в сыворотке. В итоге указанные нокаутированные мыши проявляли позитивный фенотип в отношении метаболизма липидов и/или глюкозы. Таким образом, мутантные мыши, дефицитные по гену ANGPTL4, могут служить в качестве модели для лечения сердечно-сосудистого заболевания. Антагонисты ANGPTL4 или кодирующего его гена могут играть важную роль в регуляции уровня липидов в крови и, в частности, в поддержании нормального метаболизма холестерина и триглицеридов. Такие ингибиторы или антагонисты ANGPTL4 могут быть применимы для лечения таких сердечно-сосудистых заболеваний, связанных с дислипидемией, как гипертония, атеросклероз, сердечная недостаточность, инсульт, различные болезни коронарных артерий, ожирение и/или диабет.
Фенотипический иммунологический анализ: Связанные с иммунной системой и воспалительные заболевания являются проявлениями или последствиями довольно сложных, часто многократно взаимосвязанных биологических путей, которые в случае нормальной физиологии являются важными для ответа на инсульт или повреждение, инициируют восстановление после инсульта или повреждения и обеспечивают установление врожденной и приобретенной защиты против чужеродных организмов. Заболевание или патология возникает в том случае, когда указанные нормальные физиологические пути вызывают дополнительный инсульт или повреждение, либо непосредственно связанное с интенсивностью ответа, либо как следствие аномальной регуляции или избыточной стимуляции, либо в виде реакции на «свое», или комбинации указанного.
Хотя в генез указанных заболеваний часто вовлечены многоступенчатые пути и часто множество разных биологических систем/путей, вмешательство в ключевые точки в одном или нескольких из указанных путей может оказывать улучшающий или терапевтический эффект. Терапевтическое вмешательство может происходить либо в виде антагонизма по отношению к вредному процессу/пути, либо в виде стимуляции полезного процесса/пути.
T-лимфоциты (T-клетки) являются важным компонентом иммунного ответа млекопитающих. T-клетки узнают антигены, которые связаны с собственной молекулой, кодируемой генами главного комплекса гистосовместимости (MHC). Антиген может быть экспонирован вместе с молекулами MHC на поверхности антигенпрезентирующих клеток, инфицированных вирусом клеток, клеток злокачественной опухоли, трансплантатов и т.д. T-клеточная система удаляет указанные измененные клетки, которые представляют опасность для здоровья животного-хозяина. T-клетки включают хелперные T-клетки и цитотоксические T-клетки. Хелперные T-клетки в значительной степени пролиферируют после узнавания комплекса антиген-MHC на антигенпрезентирующей клетке. Хелперные T-клетки также секретируют множество цитокинов, например, лимфокинов, которые играют центральную роль в активации B-клеток, цитотоксических T-клеток и множества других клеток, которые принимают участие в иммунном ответе.
В случае многих иммунных ответов воспалительные клетки инфильтруют место повреждения или инфекции. Мигрирующие клетки могут быть нейтрофильными, эозинофильными, моноцитами или лимфоцитами, которые можно определить с помощью гистологического исследования пораженных тканей. См., например, Current Protocols in Immunology, ed. John E. Coligan, 1994, John Wiley and Sons, Inc.
Известны и широко исследованы многие связанные с иммунной системой заболевания. Такие заболевания включают опосредованные иммунитетом воспалительные заболевания (например, ревматоидный артрит, опосредованное иммунитетом почечное заболевание, гепатобилиарные болезни, воспалительное заболевание кишечника (IBD), псориаз и астму), не опосредованные иммунитетом воспалительные заболевания, инфекционные болезни, иммунодефицитные заболевания, неоплазию и отторжение трансплантата и т.д. В области иммунологии идентифицированы мишени для лечения воспаления и воспалительных расстройств.
В области иммунологии идентифицированы мишени согласно настоящему изобретению для лечения воспаления и воспалительных расстройств. Связанные с иммунной системой заболевания в одном случае можно лечить подавлением иммунного ответа. Использование нейтрализующих антител, которые ингибируют молекулы, обладающие иммуностимулирующей активностью, может быть полезным для лечения опосредованных иммунитетом и воспалительных заболеваний. Молекулы, которые ингибируют иммунный ответ, могут быть использованы (белки непосредственно или с помощью применения агонистов антител), чтобы ингибировать иммунный ответ и таким образом улучшить состояние при связанном с иммунной системой заболевании.
Осуществляли следующий тест:
Анализ изотипов иммуноглобулинов в сыворотке: Анализ изотипов иммуноглобулинов в сыворотке осуществляли, используя набор множества шариков для цитометрического анализа (Cytometric Bead Array (CBA)). Указанный анализ использовали для быстрой идентификации изотипов тяжелой и легкой цепей мышиного моноклонального антитела в одном образце. Значения выражены в виде «относительных единиц флуоресценции» и основаны на выявлении легких каппа-цепей. Любое значение <6 является незначимым.
Результаты: Анализ изотипов иммуноглобулина в сыворотке выявил, что мутантные (-/-)-мыши имели повышение уровня иммуноглобулинов IgM в сыворотке по сравнению с совпадающими по полу однопометными (+/+)-животными. Иммуноглобулины IgM являются первыми иммуноглобулинами, которые продуцируются при гуморальном иммунном ответе для нейтрализации бактериальных токсинов и являются особенно важными для активации системы комплемента. Подобным образом IgG-иммуноглобулины оказывают нейтрализующее действие и в меньшей степени важны для активации системы комплемента. Кроме того, (-/-)-мыши имели повышенное среднее абсолютное количество нейтрофилов по сравнению с их однопометными (+/+)-животными и историческим средним значением. Наблюдаемый фенотип свидетельствует, что ANGPTL4 является негативным регулятором воспалительных ответов. Указанные иммунологические отклонения от нормы свидетельствуют, что ингибиторы (антагонисты) ANGPTL4 могут быть важными средствами, которые могут стимулировать иммунную систему (например, пролиферацию T-клеток) и могут найти применение в случаях, когда такой эффект может быть полезным для человека, например, в случае лейкоза и других типов злокачественной опухоли, и для пациентов с иммунной недостаточностью, таких как пациенты, страдающие от СПИД. Соответственно ANGPTL4 или его агонисты могут играть роль в ингибировании иммунного ответа и могут быть полезными для подавления опасных и иммунных ответов, например, в случае отторжения трансплантата или болезней трансплантат-против-хозяина.
Пример 8: Получение антител, которые связываются с Angptl4
Способы получения поликлональных антител и моноклональных антител известны в данной области и описаны в данной публикации. Антигены (или иммуногены), которые могут быть использованы, включают очищенный белок согласно изобретению, фрагменты белка, слитые белки, содержащие такой белок, и клетки, экспрессирующие рекомбинантный белок и/или фрагменты белка на клеточной поверхности. Выбор антигена может сделать специалист в данной области без чрезмерного экспериментирования.
Мышей, таких как Balb/c, иммунизируют антигеном, эмульгированным в полном адъюванте Фрейнда и инъецированным подкожно или внутрибрюшинно в количестве 1-100 мкг. Альтернативно, антиген эмульгируют в адъюванте MPL-TDM (Ribi Immunochemical Research, Hamilton, Mont.) и инъецируют в подушечки задней лапы животного. Затем иммунизированных мышей подвергают бустер-иммунизации через 10-12 дней дополнительным количеством антигена, эмульгированного в выбранном адъюванте. Затем в течение нескольких недель мыши также могут подвергаться бустер-иммунизациям с помощью дополнительных инъекций. Периодически от мышей можно получать образцы сыворотки, отбирая кровь ретроорбитально, для тестирования в анализах ELISA, чтобы выявить антитела.
После того как выявлен подходящий титр антител, животным, «позитивным» в отношении антител, можно инъецировать конечную внутривенную инъекцию данного лиганда. Через три-четыре дня мышей умерщвляют и собирают клетки селезенки. Затем клетки селезенки сливают (используя 35% полиэтиленгликоль) с выбранной линией клеток миеломы мышей, такой как P3X63AgU.1, доступной из ATCC, №CRL 1597. Слияния дают клетки гибридомы, которые затем можно высевать в 96-луночные планшеты для культуры ткани, содержащие среду HAT (гипоксантин, аминоптерин и тимидин), чтобы ингибировать пролиферацию неслитых клеток, гибридов клеток миеломы и гибридов клеток селезенки.
Клетки гибридомы могут быть подвергнуты скринингу в ELISA в отношении реактивности против антигена. Определение «позитивных» клеток гибридомы, секретирующих требуемые моноклональные антитела против ANGPTL4 согласно изобретению, хорошо известно в данной области.
Позитивные клетки гибридомы могут быть инъецированы внутрибрюшинно сингенным мышам Balb/c, чтобы получить асциты, содержащие моноклональные анти-ANGPTL4-антитела. Альтернативно клетки гибридомы можно выращивать во флаконах для культуры ткани или вращающихся флаконах. Очистку моноклональных антител, полученных в асцитах, можно осуществить, используя преципитацию сульфатом аммония с последующей гель-эксклюзионной хроматографией. Альтернативно можно использовать аффинную хроматографию, основанную на связывании антитела с белком A или белком G.
Например, поликлональные кроличьи антитела получали при иммунизации кролика 500 мкг рекомбинантного белка ANGPTL4 человека (23-406), полученного в E.coli, в 1, 40 и 70 дни. Сыворотку собирали на 80 и 120 день после иммунизации и антитела очищали на колонках, содержащих белок-A-сефадекс.
Пример 9: Блокирующие или нейтрализующие антитела
Антитела против антигенов, описанных в данной публикации, например, ANGPTL4, могут быть идентифицированы различными способами, известными в данной области, например, ELISA. Например, планшеты могут быть покрыты представляющим интерес полипептидом, например ANGPTL4 или его фрагментом, и инкубированы с антителами, созданными против такого полипептида, например ANGPTL4 (см., например, описание в патентах США 6348350, 6372491 и 6455496). Связанное антитело может быть выявлено разными способами.
Антагонистические (например, блокирующие или нейтрализующие) антитела могут быть идентифицированы в конкурентных анализах и/или анализах активности. Например, экспрессия ANGPTL4 стимулирует клеточную пролиферацию гепатоцитов или преадипоцитов, миграцию адипоцитов, регулирует количества триглицеридов или связывание с интегрином αVβ5. Регистрацию блокирующего или нейтрализующего антитела к ANGPTL4 можно показать по способности антитела блокировать указанные активности (см., например, фиг.9, панель B, D и E). Например, гепатоциты или преадипоциты можно высевать и инкубировать с надосадком клеток COS7, трансдуцированных Ad-hAngptl4, вместе с анти-ANGPTL4-антителом или контрольным антителом или PBS. Через несколько дней клетки можно обработать трипсином и подсчитать. Антитела, которые уменьшают количества клеток, идентифицируют как блокирующие или нейтрализующие антитела. Также показано, что ANGPTL4 индуцирует адгезию гепатоцитов и миграцию преадипоцитов, таким образом регистрацию блокирующего или нейтрализующего антитела к ANGPTL4 можно показать по способности антитела блокировать адгезию гепатоцитов и/или клеточную миграцию преадипоцитов. Также показано, что ANGPTL4 является проангиогенным фактором. См., например, Le Jan et al., American Journal of Pathology, 164(5): 1521-1528 (2003). Таким образом, блокирующие или нейтрализующие антитела к ANGPTL4 могут быть идентифицированы с использованием антител в комбинации с ANGPTL4 в анализах ангиогенеза, например, в анализе CAM.
Предполагается, что описание является достаточным, чтобы дать возможность специалисту в данной области практически осуществить изобретение. Понятно, что примеры и варианты осуществления изобретения, описанные в данной публикации, предназначены только для иллюстративных целей. Объем изобретения не ограничен депонированной конструкцией, так как депонированный вариант рассматривается в качестве одной иллюстрации некоторых аспектов изобретения, и любые конструкции, которые являются функционально эквивалентными, входят в объем изобретения. Депозит материала согласно изобретению не является признанием того, что описание является неадекватным, чтобы обеспечить возможность практического осуществления какого-либо аспекта изобретения, включая его наилучший способ, и не рассматривается как ограничивающий объем формулы изобретения конкретными иллюстрациями, которые он представляет. Действительно, различные модификации изобретения в дополнение к модификациям, показанным и описанным в данной публикации, будут очевидны для специалистов в данной области на основании приведенного выше описания, и они входят в объем прилагаемой формулы изобретения. Все публикации, патенты и заявки на выдачу патента, цитированные в данном описании, включены в данное описание в виде ссылки в полном объеме для всех целей.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНГИБИТОРЫ АНГИОПОЭТИНПОДОБНОГО БЕЛКА 4, ИХ КОМБИНАЦИИ И ПРИМЕНЕНИЕ | 2005 |
|
RU2392966C2 |
| МОДУЛЯТОРЫ АКТИВАТОРА ФАКТОРА РОСТА ГЕПАТОЦИТОВ | 2005 |
|
RU2418004C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДУЛЯЦИИ АКТИВАЦИИ ФАКТОРА РОСТА ГЕПАТОЦИТОВ | 2005 |
|
RU2405041C2 |
| НОВАЯ КОМПОЗИЦИЯ И СПОСОБЫ ЛЕЧЕНИЯ СВЯЗАННЫХ С ИММУНИТЕТОМ ЗАБОЛЕВАНИЙ | 2004 |
|
RU2375078C2 |
| СПОСОБЫ ИНГИБИРОВАНИЯ ГЛАЗНОГО АНГИОГЕНЕЗА | 2009 |
|
RU2530583C2 |
| ИНГИБИРОВАНИЕ МЕТАСТАЗОВ ОПУХОЛИ АНТИТЕЛАМИ ПРОТИВ НЕЙРОПИЛИНА-2 | 2007 |
|
RU2437677C2 |
| СПОСОБЫ ИНГИБИРОВАНИЯ АНГИОГЕНЕЗА С ПОМОЩЬЮ АНТАГОНИСТОВ EGFL8 | 2009 |
|
RU2486200C2 |
| ИНГИБИРОВАНИЕ АНГИОГЕНЕЗА | 2008 |
|
RU2471498C2 |
| АНТИТЕЛА ПРОТИВ CCR2 | 2009 |
|
RU2547595C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ИНГИБИРОВАНИЯ αβ-ОПОСРЕДОВАННОГО АНГИОГЕНЕЗА | 1997 |
|
RU2195312C2 |


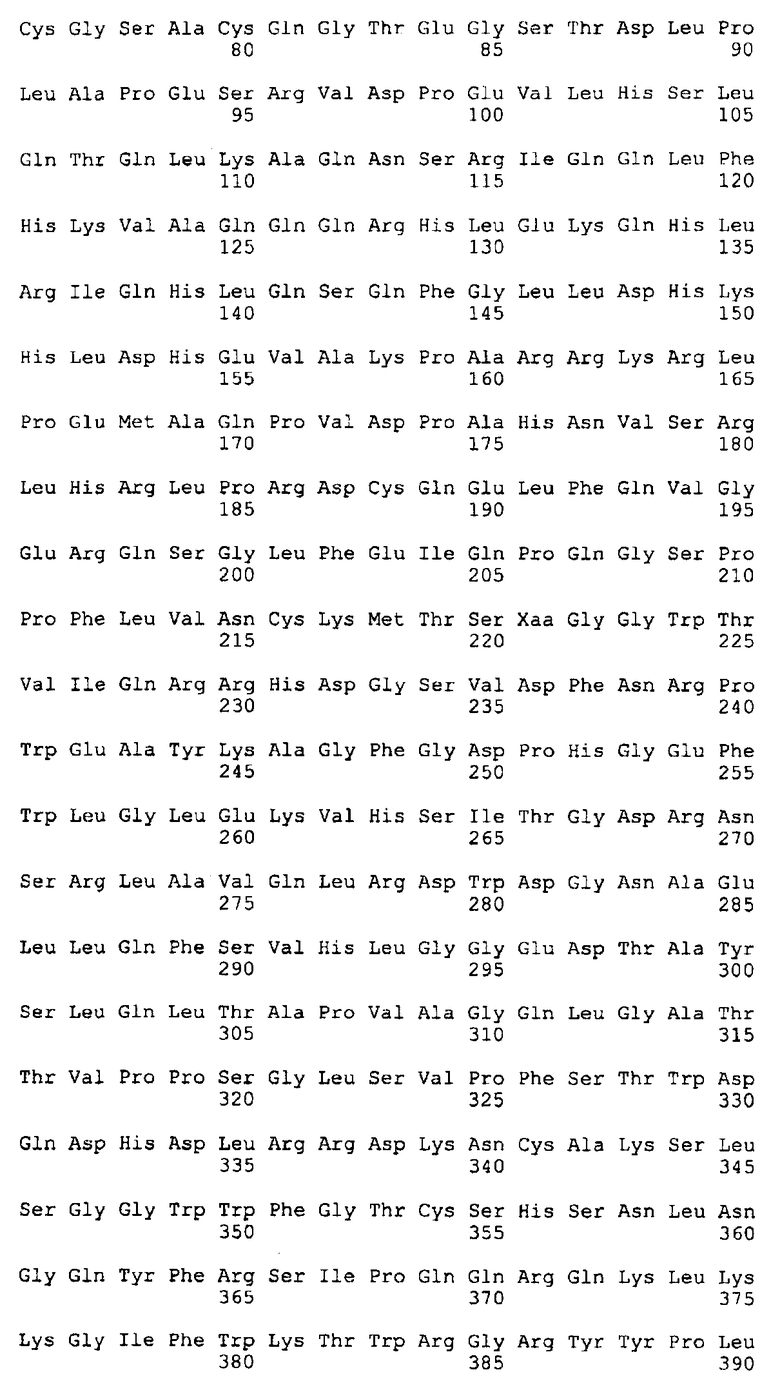




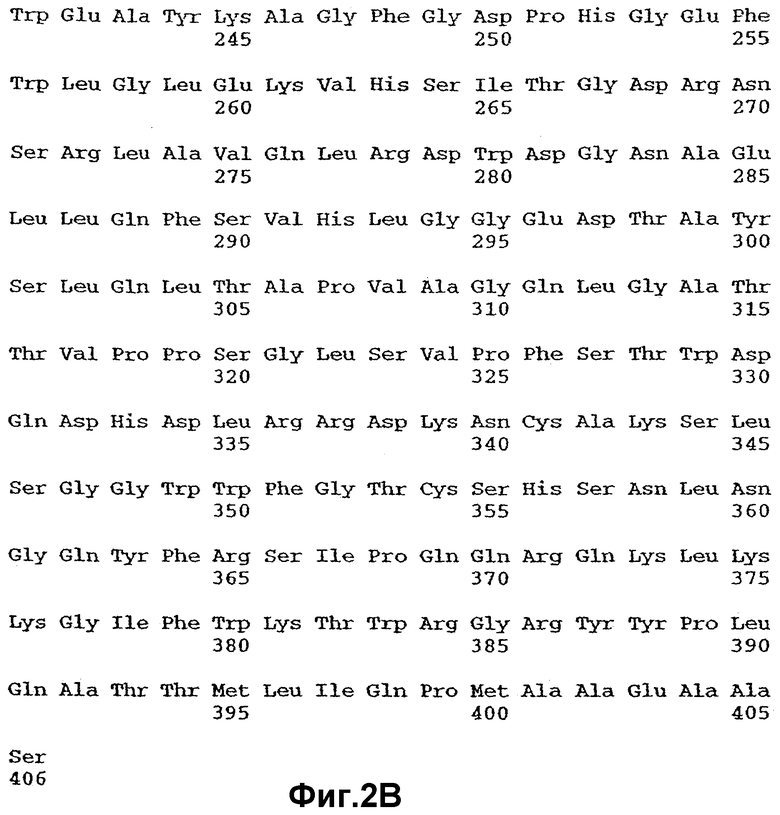
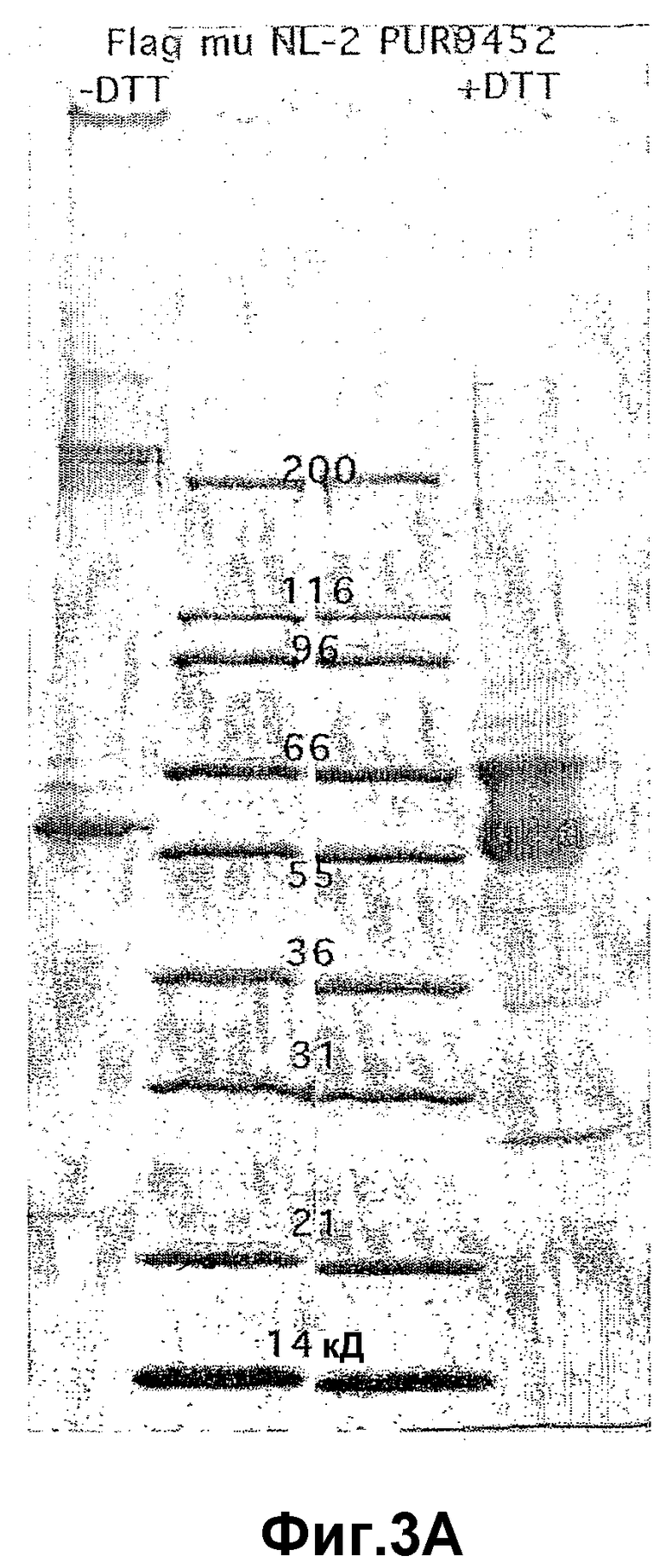
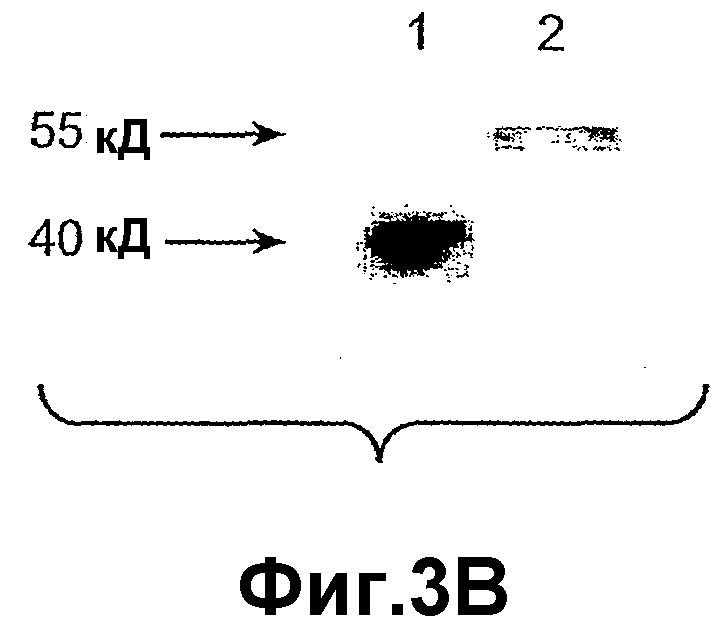
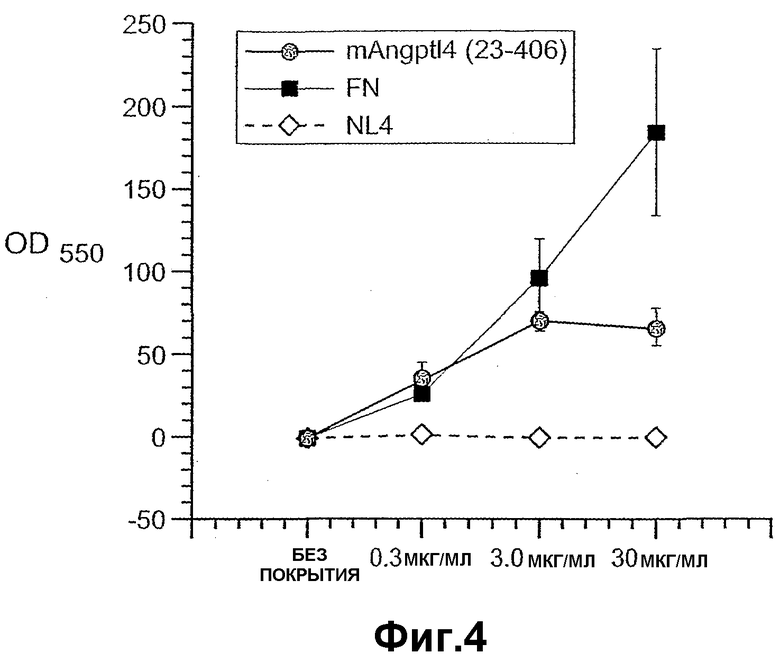
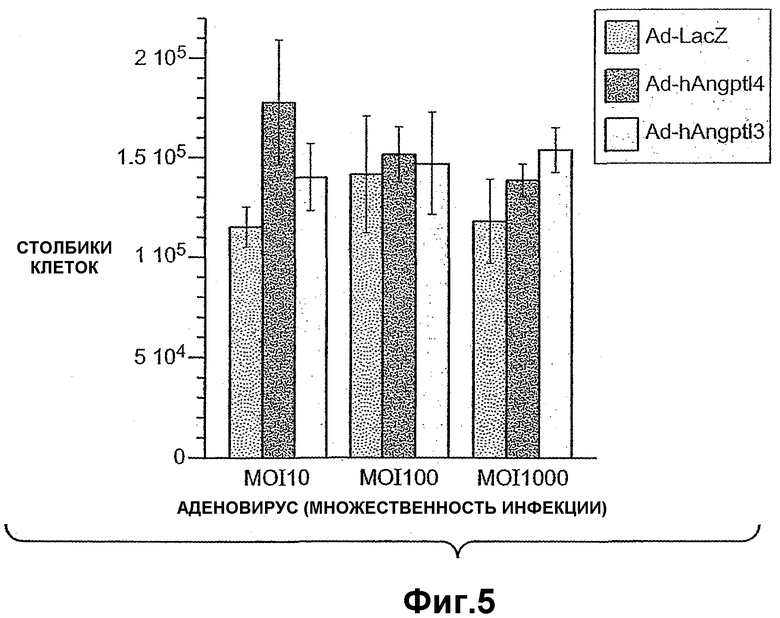
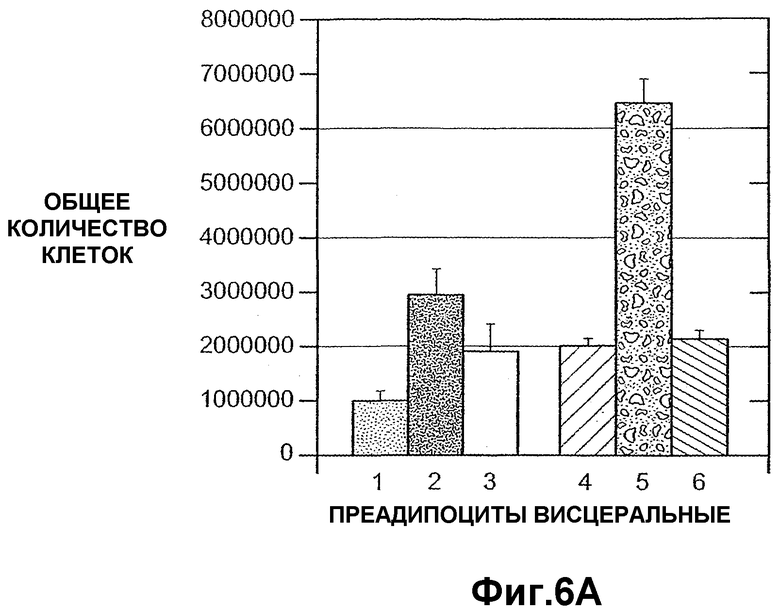
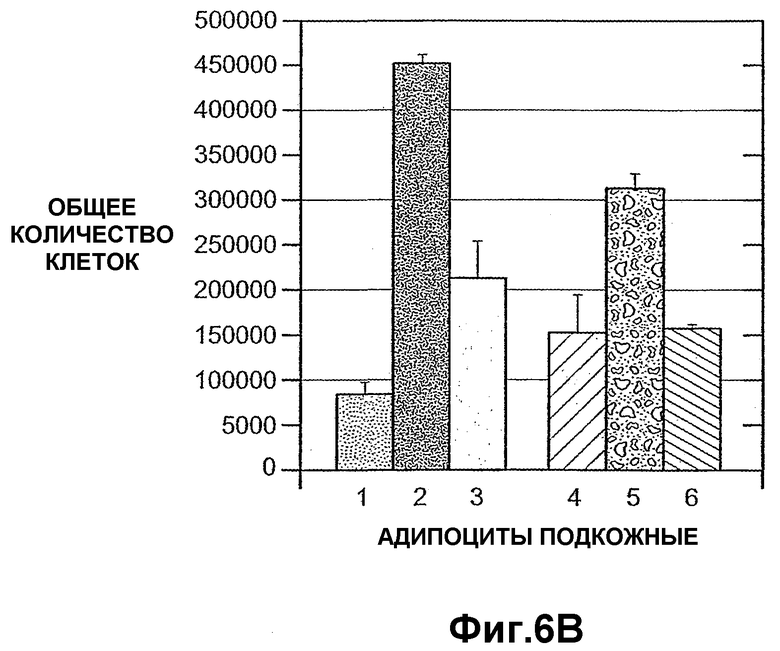
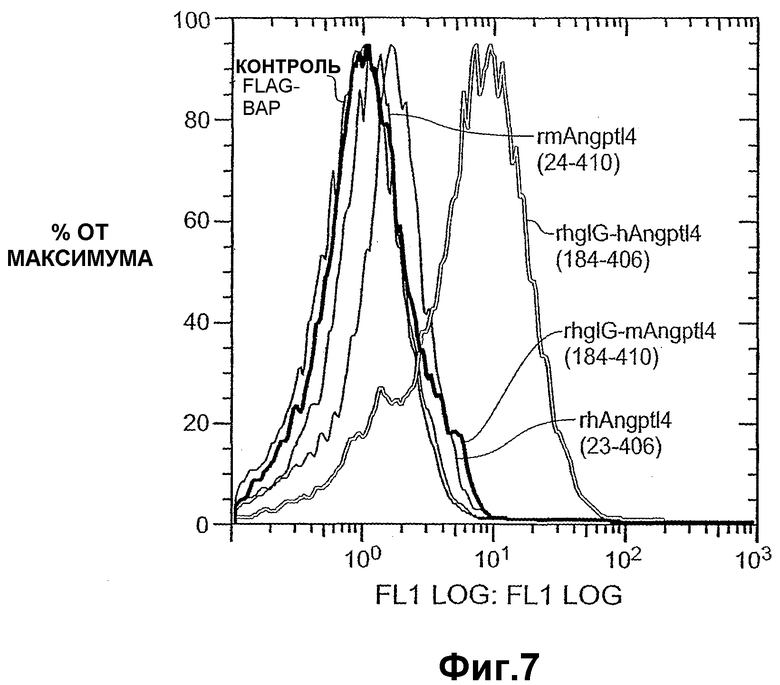
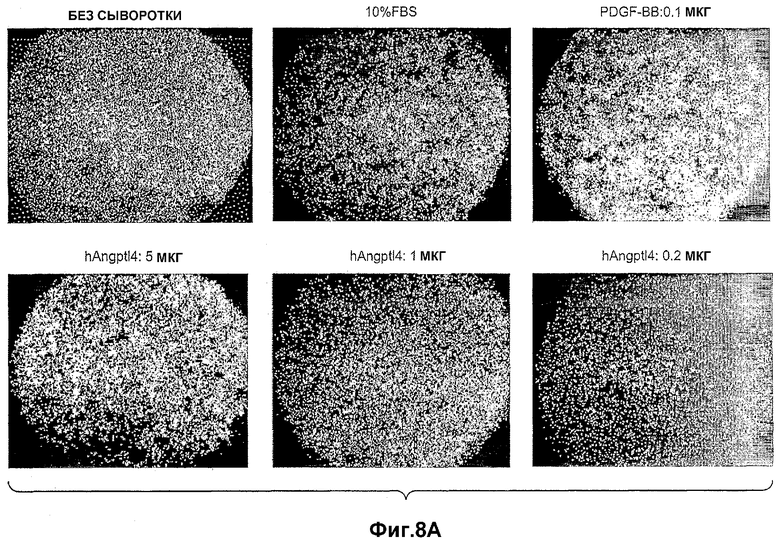

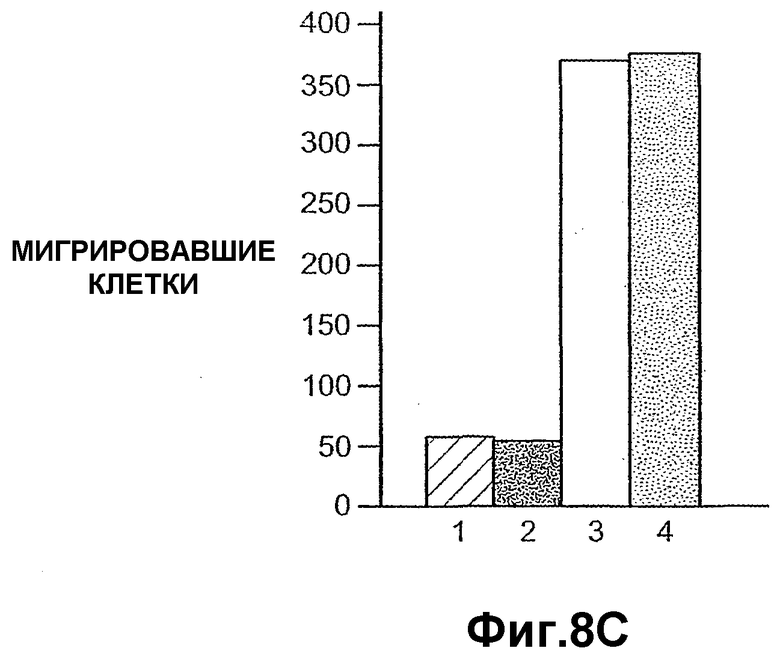
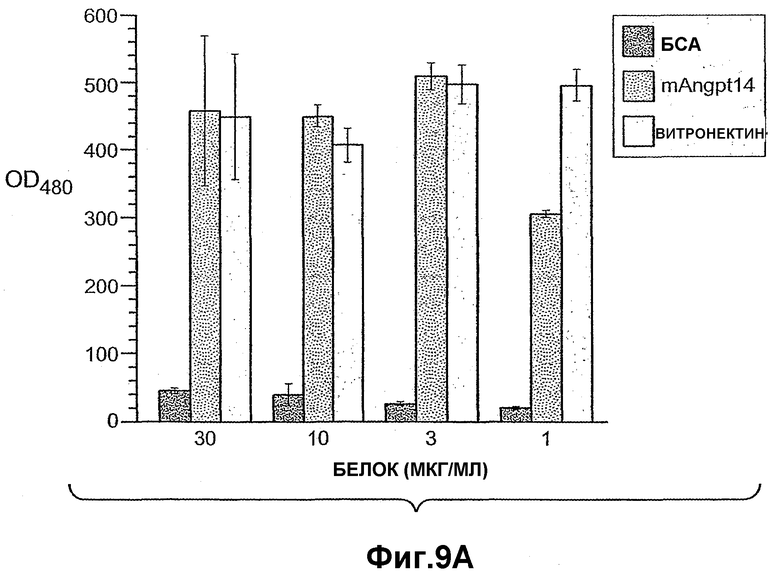
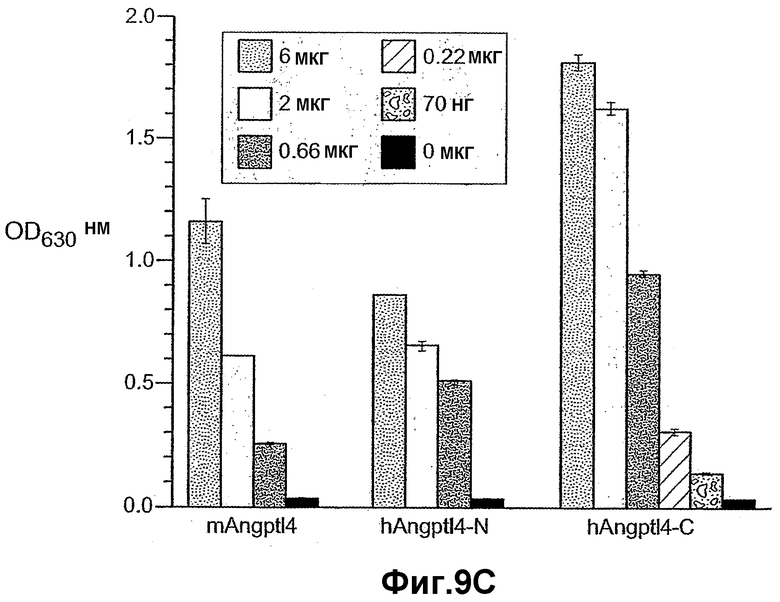
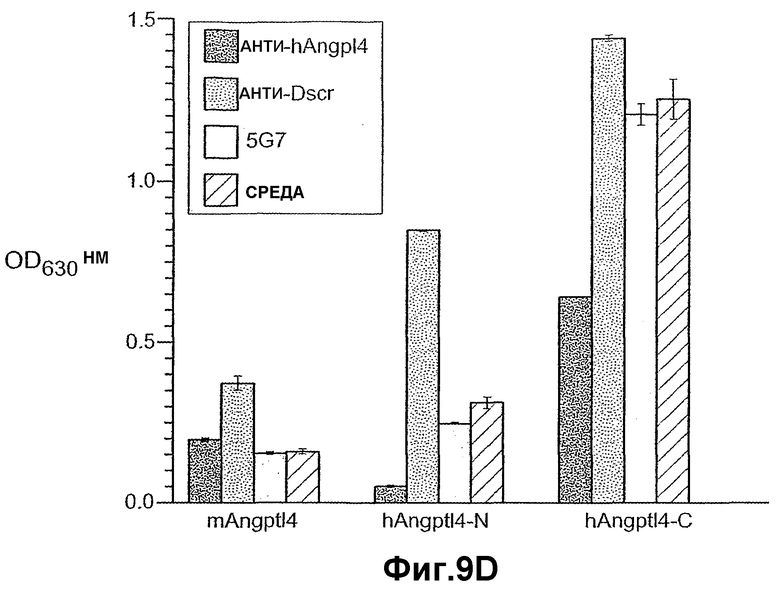
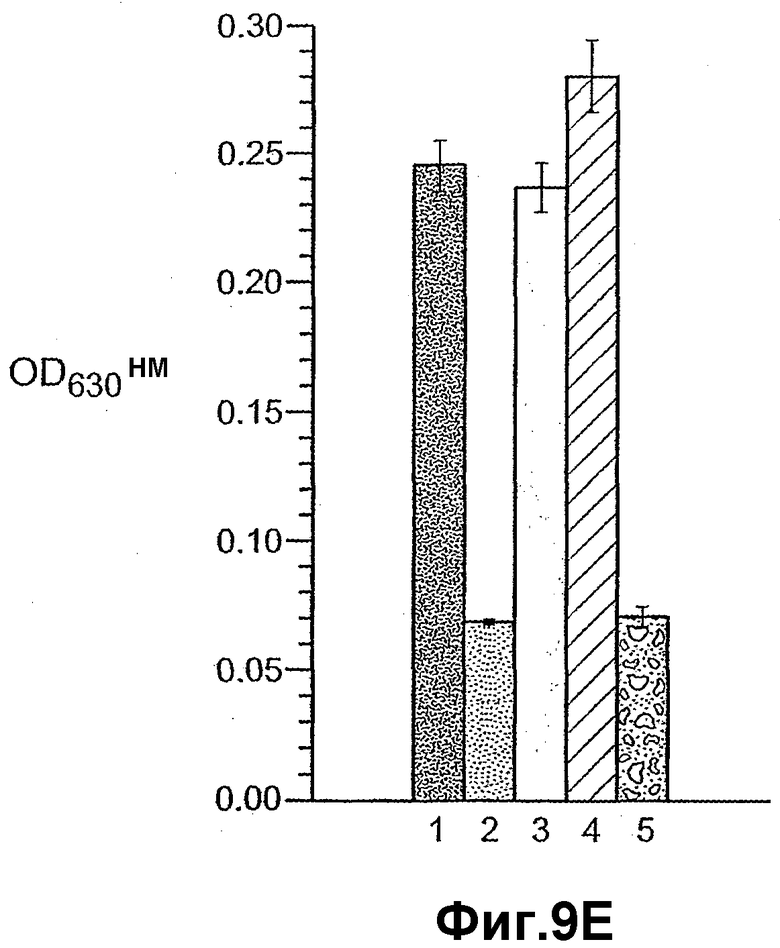
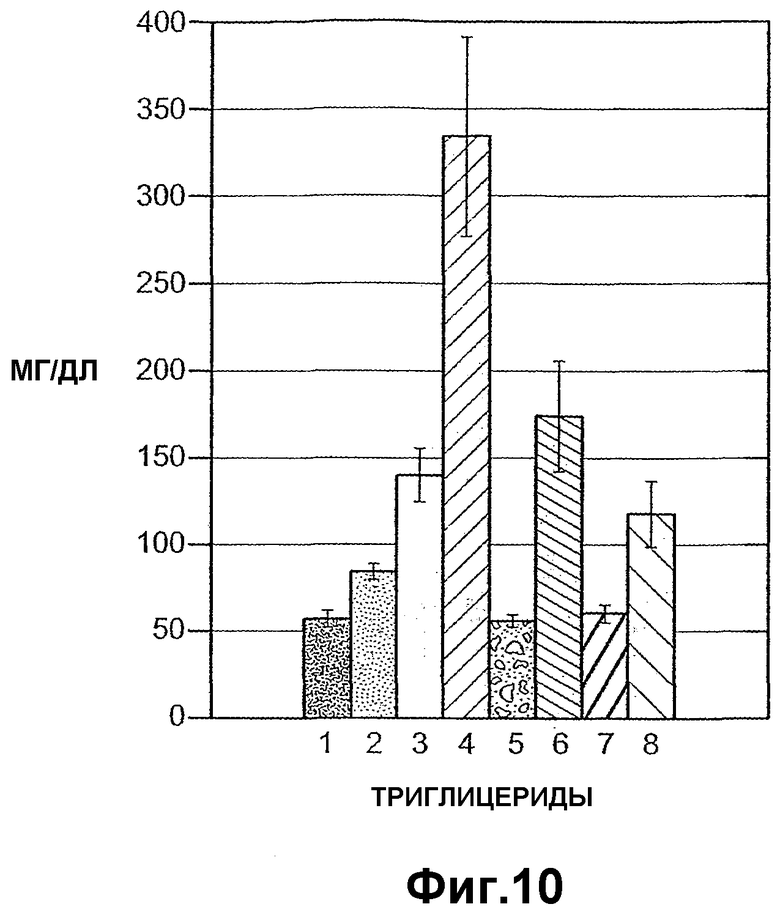
Изобретение относится к области биотехнологии, конкретно к получению антагонистов ангиопоэтинподобного белка-4 (ANGPTL4), и может быть использовано в медицине. В качестве антагониста ANGPTL4 используют анти-ANGPTL4-антитело. Изобретение позволяет получить антагонист ANGPTL4, способный ингибировать пролиферацию и клеточную адгезию гепатоцитов, а также ингибирующий биологическую активность ANGPTL4 в гепатоцитах или предшественниках гепатоцитов. 3 н. и 1 з.п. ф-лы, 20 ил., 1 табл.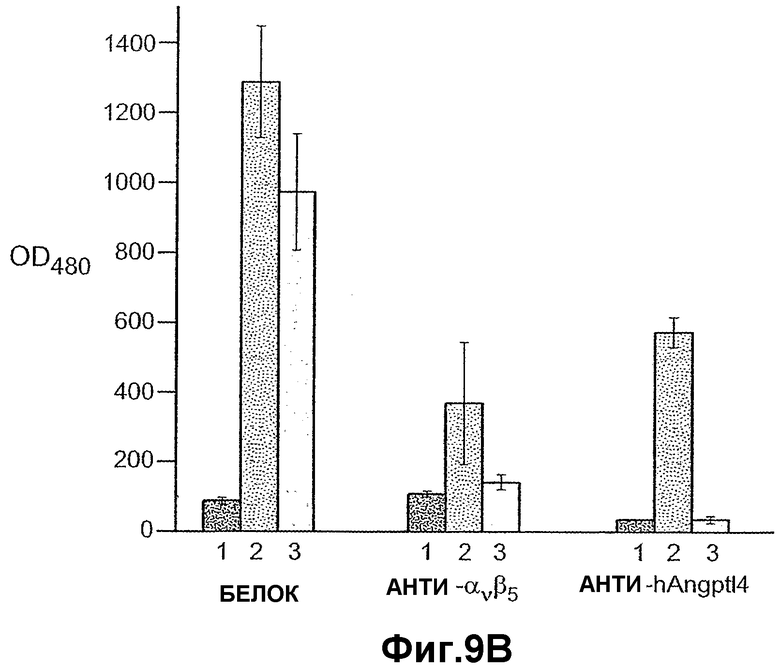
1. Способ ингибирования пролиферации гепатоцитов, где способ включает введение эффективного количества антагониста ANGPTL4 в популяцию гепатоцитов или предшественников гепатоцитов, при этом антагонистом ANGPTL4 является анти-ANGPTL4-антитело.
2. Способ ингибирования клеточной адгезии гепатоцитов, где способ включает введение эффективного количества композиции, содержащей антагонист ANGPTL4, в популяцию гепатоцитов, ингибируя тем самым клеточную адгезию гепатоцитов, при этом антагонистом ANGPTL4 является анти-ANGPTL4-антитело.
3. Способ ингибирования биологической активности ANGPTL4 в гепатоцитах или предшественниках гепатоцитов, где способ включает введение антагониста ANGPTL4, который связывается с С-концом ANGPTL4, при этом антагонистом ANGPTL4 является анти-ANGPTL4-антитело, а указанная биологическая активность ANGPTL4 представляет собой связывание с αvβ5 или индукцию пролиферации клеток.
4. Способ по п.3, где антагонист ANGPTL4 блокирует связывание ANGPTL4 с αvβ5.
| GE H | |||
| et al., Oligomerization and regulated proteolytic processing of angiopoietin-like protein 4, J | |||
| Biol | |||
| Chem., 2004, v.279, n.3, p.2038-2045 | |||
| GE H | |||
| et al., Oligomerization and regulated proteolytic processing of angiopoietin like protein 4, Diabetes, 2004, v.53, Suppl.2, p.A576 | |||
| YOON J.K | |||
| et al., Peroxisome proliferator-activated receptor gamma |

