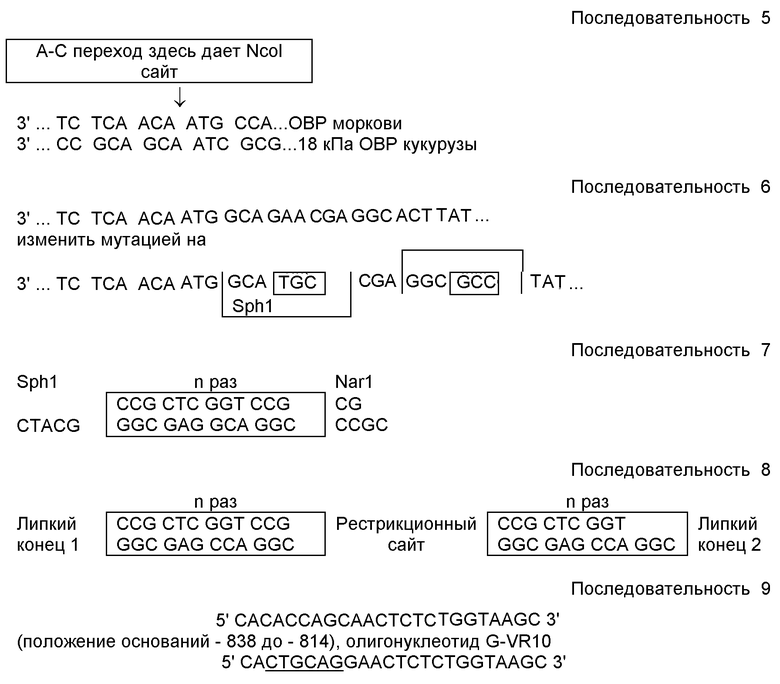
Область изобретения
Настоящее изобретение относится к рекомбинантному способу получения представляющих интерес протеинов, которые легко выделять из компонентов клеток хозяев. Способ иллюстрируется экспрессией представляющего интерес протеина в растениях, особенно в семенах в форме химерного пептида, содержащего протеин масляного тела и представляющий интерес протеин.
Предпосылки изобретения
В растениях были экспрессированы различные протеины. Однако хотя и была продемонстрирована общая доступность осуществления экспрессии чужеродных протеинов в растениях, получение очищенных протеинов из таких источников имеет некоторые ограничения. Эти ограничения включают стадию очистки, необходимую для получения чистого протеина, практически не содержащего растительных материалов и продуктов разложения, которые могут образовываться в экстрактах, полученных в процессе очистки, когда полученные рекомбинантные протеины контактируют с водными буферами.
Растения, имеющие масличные семена, такие как соя, рапс, подсолнечник и ряд других видов растений, таких как кукуруза, морковь и т.д., хранят в своих семенах триглицериды. В растениях эти триглицериды выполняют роль источника энергии для прорастания семян и последующего роста сеянца. Триглицериды широко используют в качестве растительных масел в пище и пищевых продуктах, а также в некоторых областях промышленности.
Триглицериды не смешиваются с водой и выделяются за счет флотации на поверхности водных растворов или образуют небольшие глобулы или липосомы в виде суспензии в водной фазе. Такие глобулы естественно сливаются, если они не стабилизированы модифицированным поверхностным слоем. Такое слияние может привести к образованию суспензии из глобул случайного размера. В семенах же при сохранении триглицеридов масляные глобулы на деле представляют собой капсулированные липидные или масляные тела обычно одинакового размера. С поверхностью этих масляных тел ассоциированы полуфрагменты (half unit) мембран, усыпанные несколькими протеинами, которые обычно называют протеинами масляного тела.
По крайней мере, один класс протеинов масляного тела имеет несколько характеристик, которые существенно консервативны от вида к виду. Этот класс протеинов масляного тела называют "олеозинами". Гидрофильные N- и С- концы этих протеинов, по-видимому, совершенно различны, тогда как липофильные внутренние участки (центральное ядро), по-видимому, строго консервативны для различных видов. Олеозины прочно связаны с масляными телами, эта прочная связь с масляными телами может быть, в основном, связана с липофильным характером этих центральных ядер. Поэтому представляет интерес определение возможности использования таких протеинов масляного тела, как олеозины, для создания средств для выделения рекомбинантных протеинов из растительных материалов.
Известный уровень техники
Продуцирование чужеродных (рекомбинантных) пептидов в растениях было исследовано с использованием различных подходов, включая транскрипционные слияния с применением сильного конститутивного растительного промотора (например, из вируса мозаики цветной капусты - Sijmons et al. (1990) Bio/Technology, 8: 217-221) и кодирование чужеродного протеина; транскрипционные слияния с органо-специфическими последовательностями (Radke et al. (1988) Theoret. Appl. Genet., 75:685-694); и трансляционные слияния, которые требуют последующего отщепления рекомбинантного протеина (Vander Kerkove et al. (1989) Bio/ Technology, 7:929-932). Чужеродные протеины, которые были экспрессированы в растительных клетках, включают активные протеины из бактерий (Fraley et al. (1983) Proc. Nat'l. Acad. Sci. USA, 80:4803-4807), животных (Misra and Gedamu (1989) Theor. Appl. Genet., 78:161-168), грибков и других видов растений (Fraley et al. (1983) Proc. Nat'l. Acad. Sci. USA, 80: 4803-4807).
Некоторые протеины, обычно маркеры интеграции, были экспрессированы ткане-специфическим образом, причем некоторые были экспрессированы в семенах (Sen Gupta et al. (1985) Proc. Nat'l. Acad. Sci. USA, 82:3320-3324); Radke et al. ( 1988) Theor. Appl. Genet., 75:685-694). Эти сообщения концентрируются специфически на использовании промоторов протеинов хранения семян как средства обеспечения специфической для семян экспрессии. Используя эти системы, Vanderkerkove et al. (1989) Bio/Technol., 7: 929-932, экспессировал очень ценный пептид (leu энкефалин) в семенах Arabidopsis thaliana and Brassica napus. Выход этих пептидов чрезвычайно низок, но он демонстрирует возможность экспрессии пептидного гормона животного в ткани растения. Олеозин кукурузы был экспрессирован в масляных телах семян Brassica napus трансформированных геном олеозина кукурузы. Этот ген был экспрессирован под контролем регуляторных элементов из гена Brassica, кодирующего напин (napin), основной протеин хранения семян. Как сообщается, временная регуляция и тканеспецифичность экспрессии правильны для промотора/терминатора гена напина. См. Lee et al., Proc. Nat'l. Acad. Sci. USA, (1991) 88:6181-6185.
Масляные глобулы, которые образуются в семенах, по-видимому, все одинакового размера, что указывает на их стабилизацию (Huang А.Н.С (1985) в Modern Meths. Plant Analysis v. 1:145-151 Springer-Verlag, Berlin). При более тщательном исследовании было обнаружено, что они представляют собой не просто масляные глобулы, а скорее масляные тела, окруженные мембраной. Такие масляные тела называли по разному электронные микроскописты: олеосомы, липидные тела и сферосомы (Gurr М.I., (1980) в The Biochemistry of Plants, 4:205-248, Acad. Press, Orlando, Pla). Были исследованы масляные тела нескольких видов, и пришли к выводу, что они заключены в необычные "полуфрагментарные" мембраны, которые представляют собой не классический липидный бислой, а скорее один амфофильный слой с гидрофобными группами с внутренней стороны и гидрофильными группами с внешней (Huang А. Н. С. (1985) в Modern Meths. Plant Analysis, vol. 1:145-151 Springer-Verlag, Berblin).
Анализ содержания липидных тел продемонстрировал, что помимо триглицеридов и материала мембран они содержат также несколько полипептидов/протеинов, связанных с поверхностью или полостью масляного тела (Bowman-Vance and Huang (1987), J. Biol. Chem. 262:11275-11279, Murphy et al. (1989) Biochem. J., 258: 285-293, Taylor et al. (1990) Planta, 181:18-26). Протеины масляных тел были идентифицированы для широкого круга таксономически различных видов (Moreau et al. (1980) Plant Physiol., 65: 1176-1180; Qu et al. (1986) Biochem. J. , 235:57-65) и было показано, что они расположены только в масляных телах, но не были обнаружены в органеллах растительных тканей. В Brassica napus (семена рапса) имеются, по крайней мере, три полипептида, связанные с масляными телами в развивающихся семенах Taylor et al. (1990), Planta, 181: 18-26). Количество и размеры протеинов, связанных с масляными телами, могут меняться от вида к виду. Так например, в кукурузе существуют два иммунологически различных класса полипептидов, обнаруженных в масляных телах (Bowman-Vance and Huang (1988) J. Biol. Chem. 263:1476-1481).
Было показано, что олеозины содержат участки чередующейся гидрофобности и гидрофильности (Bowman-Vance and Huang (1987) J. Biol. Chem., 262:11275-11279). Аминокислотные последовательные олеозинов из кукурузы, семян рапса и моркови были получены. См. Qu and Huang (1990) J. Biol. Chem. 265:2238-2243, Hatzopoulos et al (1990) Plant Cell., 2:457-467 соответственно. В таких семенах масличных, как рапс, олеозин может составлять от 8% (Taylor et al. (1990) Planta, 181: 19-26) до 20% (Murphy et al. (1989) Biochem. J., 258: 285-293) от полного содержания протеина в семенах. Столь высокое содержание сопоставимо с тем, что было обнаружено для многих протеинов хранения семян.
Сообщалось, что получены гены, кодирующие протеины масляных тел для двух видов: кукуруза (Zea mays, Bowman-Vance and Huang (1987) J. Biol. Chem. 262: 11275-11279; и Qu and Huang (1990), J.Biol.Chem., 265: 2238-2243) и морковь (Hatzopoulos et al. (1990) Plant Cell., 2:457-467).
Краткое содержание изобретения
Предложены способы и композиции для осуществления этих способов получения пептидов, которые можно легко выделить из протеинов хозяев. Этот способ включает стадии получения химерной конструкции ДНК, которая включает последовательность, кодирующую специфическую последовательность масляного тела, содержащую кодирующую последовательность гена протеина масляного тела, специфического для семян, или последовательность, кодирующую, по крайней мере, участок гидрофобного ядра протеина масляного тела, и последовательность, кодирующую нужный пептид, из которых можно получить кассету экспрессии, содержащую химерную конструкцию ДНК; трансформация клеток хозяина кассетой экспрессии в условиях геномной интеграции; и выращивания полученного трансгенного растения до получения семян, в которых нужный полипептид экспрессирован как протеин слияния с олеозином.
Представляющий интерес полипептид можно очистить, выделяя масляные тела из клеток семян, разрушая масляные тела, так что при этом высвобождается протеин слияния. Затем протеины масляного тела легко отделяются от других протеинов и полученных из растений материалов за счет разделения фаз. При желании сайт расщепления может быть расположен, по крайней мере, перед N-концом или после С-конца представляющего интерес полипептида, что обеспечит возможность отщеплять полипептид слияния и выделять его за счет фазового разделения на составляющие его пептиды. Таким образом, создана система, обеспечивающая целенаправленный перенос химерного пептида за счет функциональности протеина его масляного тела к масляным телам, что в свою очередь позволяет быстро очистить представляющий интерес полипептид. Эта система получения находит применение при получении многих пептидов, например таких пептидов, которые обладают нужными фармацевтическими, энзиматическими, реологическими и адгезивными характеристиками.
В соответствии с настоящим изобретением предложен полипептид слияния, отличающийся тем, что может быть обеспечена его доставка к масляному телу, содержащий:
а) первый пептид формулы (последовательность 1, см. в конце описания).
при условии, что указанный первый пептид отличается от полноразмерного природного 16Кb олеозина из моркови, или 18Кb или 16Кb олеозина из кукурузы, где
аа25 представляет любую аминокислоту;
аа26 представляет нейтральную алифатическую аминокислоту;
аа31 представляет нейтральную незамещенную алифатическую аминокислоту, состоящую из 3-6 атомов углерода;
аа33 представляет нейтральную незамещенную алифатическую аминокислоту, содержащую 3-6 атомов углерода;
аа36 представляет нейтральную алифатическую незамещенную аминокислоту, содержащую 3-5 атомов углерода;
аа37 представляет нейтральную незамещенную аминокислоту;
аа39 представляет нейтральную алифатическую незамещенную аминокислоту;
аа41 представляют нейтральную алифатическую незамещенную или оксизамещенную аминокислоту;
аа44 представляет нейтральную алифатическую незамещенную или оксизамещенную аминокислоту;
аа46 представляет нейтральную алифатическую незамещенную аминокислоту или оксизамещенную аминокислоту;
аа47 представляет нейтральную алифатическую незамещенную аминокислоту;
аа59 представляет нейтральную алифатическую или ароматическую незамещенную аминокислоту;
аа76 представляет нейтральную алифатическую незамещенную или тиозамещенную аминокислоту;
аа78 представляет нейтральную алифатическую незамещенную аминокислоту;
аа83 представляет нейтральную алифатическую незамещенную или оксизамещенную аминокислоту;
аа92 представляет нейтральную алифатическую аминокислоту с оксизамещением;
аа96 представляет нейтральную алифатическую тиозамещенную аминокислоту или нейтральную ароматическую гетероциклическую аминокислоту;
аа97 представляет нейтральную алифатическую незамещенную или тиозамещенную аминокислоту;
аа98 представляет нейтральную алифатическую незамещенную аминокислоту или ароматическую оксизамещенную аминокислоту;
аа99 представляет любую аминокислоту;
аа100 представляет оксизамещенную аминокислоту, либо алифатическую, либо ароматическую;
аа101 представляет нейтральную незамещенную алифатическую или ароматическую аминокислоту;
слитый с
b) вторым пептидом, при условии, что этот второй пептид отличается от части природного олеозинового протеина из Arabidopsia или Brassica.
В настоящем изобретении предложен также полипептид слияния, характеризуемый как способный осуществлять целенаправленный перенос к масляному телу, содержащий:
а) первый пептид, выбранный из группы, состоящей из:
(I) пептида, содержащего, по крайней мере, восемь последовательных аминокислот, включенных в аминокислотную последовательность 2, см. в конце описания.
(2) пептид, который кодируется ДНК последовательностью, идентифицированной с помощью олигонуклеотидного зонда, сконструированного на основе указанной аминокислотной последовательности (I), или ее фрагмента, при условии, что указанный первый пептид отличается от полноразмерного природного 16Кb олеозина из моркови или 18Кb или 16Кb олеозина из кукурузы, слитый с
b) вторым пептидом, при условии, что указанный второй пептид отличается от части природного олеозинового протеина из Arabidopsis или Brassica.
Далее в настоящем изобретении предложена химерная ДНК конструкции, содержащая:
а) первую ДНК последовательность, кодирующую олеозин или его часть, достаточную для обеспечения целенаправленного переноса к масляному телу, и
b) вторую ДНК последовательность, кодирующую пептид, при условии, что указанный пептид отличается от части природного олеозинового протеина из Arabidopsis или Brassica.
Далее в настоящем изобретении предложена кассета экспрессии, содержащая:
в качестве компонентов в направлении транскрипции:
- регуляторную ДНК последовательность, содержащую достаточную часть участка 5' до сайта начала трансляции гена, экспрессируемого в семенах, для обеспечения экспрессии ДНК последовательности в семенах;
- химерную ДНК последовательность, содержащую
а) первую ДНК последовательность, кодирующую олеозин, или достаточную ее часть для обеспечения целенаправленного переноса к масляному телу, причем указанная ДНК последовательность включает, по крайней мере, один рестрикционный сайт, и
b) вторую ДНК последовательность, кодирующую пептид, при условии, что указанный пептид отличается от природного олеозинового протеина из Arabidopsis или Brassica; и
- участок окончания трансляции и транскрипции;
где указанные компоненты операбельно связаны, а экспрессия указанной химерной последовательности ДНК регулируется указанной регуляторной ДНК последовательностью.
В настоящем изобретении предложена также кассета экспрессии, содержащая:
- ген протеина масляного тела ( oil body protein = ОВР), который включает достаточную часть участка 5' до сайта начала трансляции для обеспечения экспрессии указанного гена в клетки семян и который включает, по крайней мере, один рестрикционный сайт между 5'-концом кодона инициации метионина и 5'-сигналом окончания трансляции указанного ОВР гена, и
- ДНК последовательность, встроенную в указанный рестрикционный сайт в рамке считывания с указанным ОВР геном, где указанная ДНК последовательность кодирует пептид, который отличается от части нативного олеозинового протеина из Arabidopsis или Brassica.
В настоящем изобретении предложена также кассета экспрессии, содержащая: первую ДНК последовательность, кодирующую пептид, при условии, что указанный пептид отличается от части природного олеозиинового протеина из Arabidopsis или Brassica, встроенную в рамке считывания в ген протеина масляного тела, которая включает достаточную часть регуляторного участка 5' до сайта начала трансляции указанного ОВР гена для обеспечения экспрессии указанного гена в семенах, где указанная последовательность встроена по такому сайту указанного гена, чтобы быть экспрессированной под контролем указанного регуляторного участка.
В настоящем изобретении предложен способ достижения экспрессии целевого пептида в семенах, причем указанный способ включает трансформацию клеток растения-хозяина экспрессионной кассетой в условиях геномной интеграции, где указанная кассета экспрессии содержит в качестве компонентов в направлении транскрипции первую ДНК последовательность, содержащую достаточную часть участка 5' до сайта начала трансляции гена, экспрессируемого в семенах, для обеспечения экспрессии ДНК последовательности в семенах, вторую ДНК последовательность, кодирующую олеозин или достаточную его часть, для обеспечения нацеленного переноса к масляному телу, причем указанная вторая ДНК последовательность включает, по крайней мере, один природный или синтетический рестрикционный сайт, в который встроена в рамке считывания третья ДНК последовательность, кодирующая целевой пептид, при условии, что указанный пептид отличается от части природного олеозинового протеина из Arabidopsis или Brassica; и участок окончания трансляции и транскрипции; где указанные компоненты операбельно связаны, и экспрессия указанной второй ДНК последовательности регулируется указанной первой ДНК последовательностью, для обеспечения экспрессии в семенах.
В настоящем изобретении предложен способ получения очищенного целевого пептида, причем указанный способ включает:
трансформацию клеток растения-хозяина ДНК конструкцией в условиях геномной интеграции, где указанная ДНК конструкция содержит первую ДНК последовательность, кодирующую целевой пептид, при условии, что указанный пептид отличается от части природного олеозинового протеина из Arabidopsis или Brassica, встроенную в считывающую рамку гена протеина масляного тела, которая содержит достаточную часть регуляторного участка 5' до сайта начала трансляции указанного ОВР гена для обеспечения экспрессии указанного гена в семенах, причем указанная последовательность встроена по такому сайту указанного гена, что экспрессия указанной ДНК последовательности контролируется указанным регуляторным участком, за счет чего указанная ДНК конструкция интегрируется в геном указанных клеток растения;
выращивание указанного растения до получения семян, в которых целевой пептид экспрессирован как протеин слияния с продуктом экспрессии указанного ОВР гена;
выделение масляных тел из клеток указанных семян;
разрушение указанных масляных тел, в результате чего выделяется указанный протеин слияния;
и очистку указанного целевого пептида.
Краткое описание чертежей
Фиг. 1А представляет нуклеотидную последовательность и выведенную аминокислотную последовательность (17 кДа протеин) гена протеина масляного тела (олеозина) из Arabidopsis thaliana. Подчеркнуты прямые повторы (R1 и R2) инвертный повтор (Т), САСА, ТАТА, ТААТ и сигналы полиаденилирования.
Интронная последовательность напечатана мелким шрифтом, а предполагаемый АВА-связывающий сайт указан жирным шрифтом.
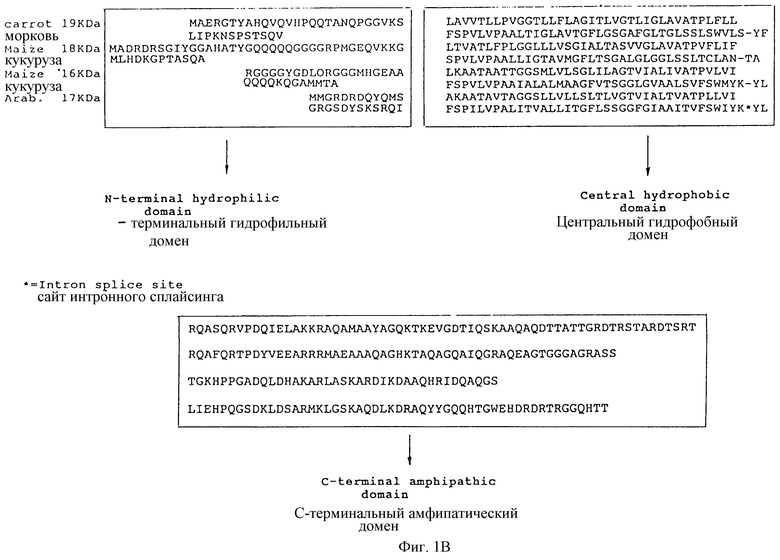
На фиг.1В представлено сравнение последовательностей протеина 16Кb масляного тела из моркови и 18Кb и 16Кb протеинов масляного тела из кукурузы и 17Кb протеина масляного тела из Arabidopsis thaliana, где показаны сохраняющиеся (консервативные) и различающиеся участки этих протеинов; аминокислотные последовательности выравнены, чтобы показать консервативность последовательности в центральном участке протеинов.
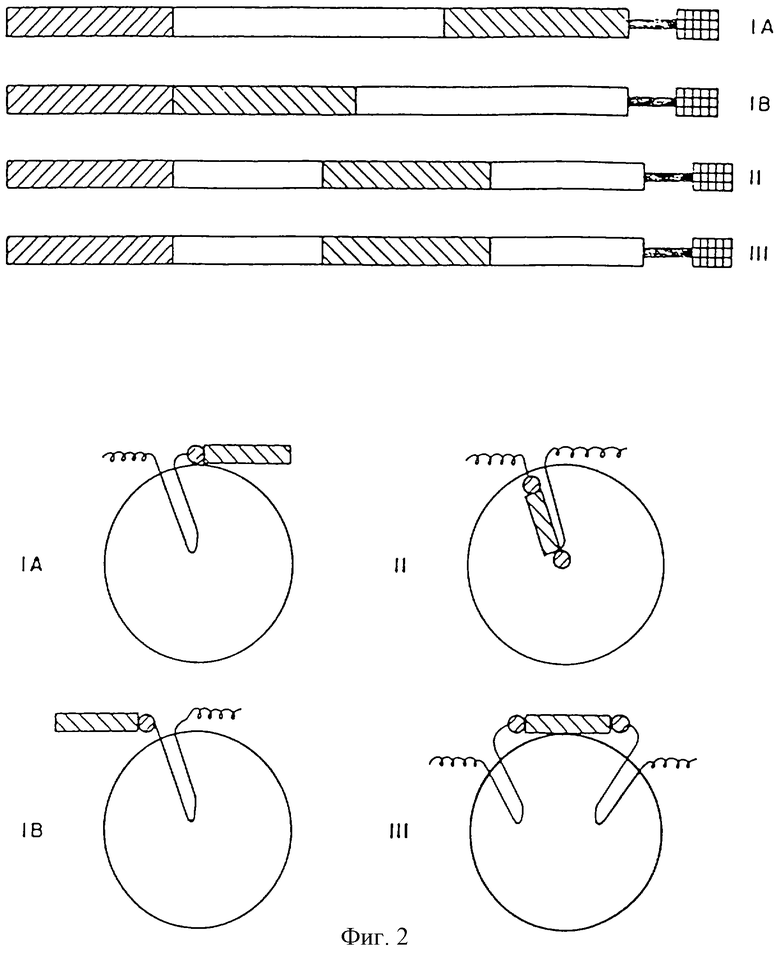
На фиг.2 представлены конструкции, использованные для слияния генов протеинов масляного тела с генами, кодирующими чужеродные пептиды. IА представляет С-терминальное слияние целевого пептида с ОВР; IВ представляет N-терминальное слияние целевого пептида с ОВР; II представляет внутреннее слияние целевого пептида с ОВР; и III представляет междимерное трансляционное слияние целевого пептида, заключенного между двумя практически полными последовательностями протеина масляного тела, обеспечивающими его направленную доставку. В верхней части фиг. (А) представлена ДНК конструкция, использованная для трансляционного слияния целевых пептидов с протеинами масляного тела. В нижней части фиг. (В) представлены конфигурации генных продуктов, показанных на верхнем участке трансляции и доставки к масляным телам.
Ключ к фиг. следующий: заштрихованный снизу слева вверх направо прямоугольник представляет ОВР промотор или другой специфический для семян промотор; заштрихованный снизу справа вверх налево прямоугольник представляет последовательность, кодирующую целевой пептид; незаштрихованный прямоугольник представляет последовательность, кодирующую протеин масляного тела или синтетическую последовательность, обеспечивающую направленную доставку, основанную на ОВР консервативных фрагментах; заштрихованные в клеточку прямоугольники представляют терминатор гена, содержащий сигнал полиаденилирования; заштрихованный кружок представляет фрагмент распознавания протеазы; спиральная линия представляет нативный С- или N-конец ОВР.
Фиг. 3 представляет подробную структуру конструкции С-терминального слияния. Изображена структура последовательности, кодирующей участок распознавания коллагеназы, в качестве линкора при слиянии типичного гена протеина масляного тела и пептида слияния, который нужно связать, используя Ncol для клонирования и экспрессии в растениях.
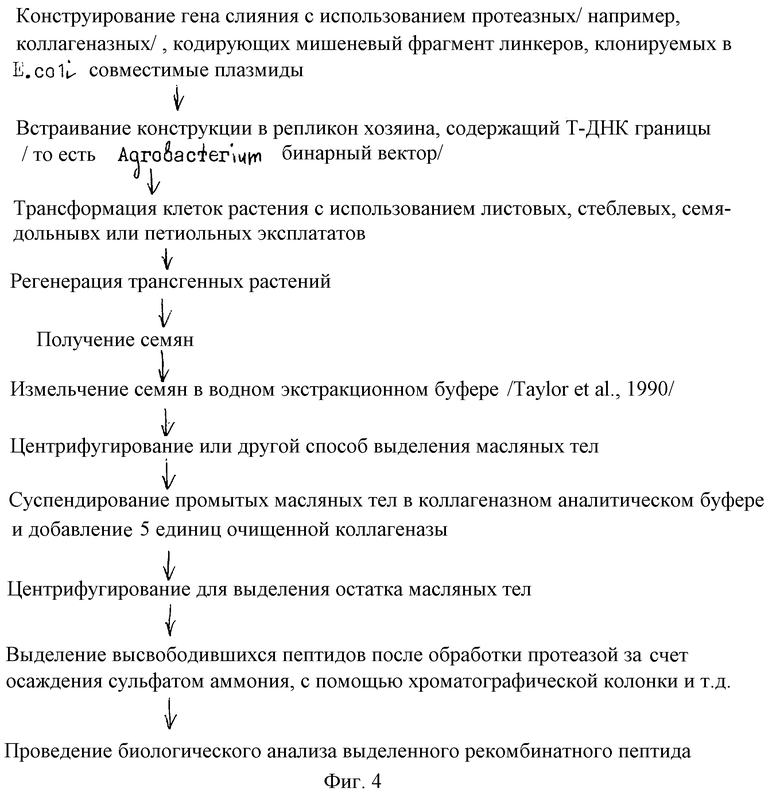
На фиг.4 схематически представлен процесс конструирования векторов пептида слияния, их введение в растения и последующую экстракцию и анализ целевого рекомбинантного пептида.
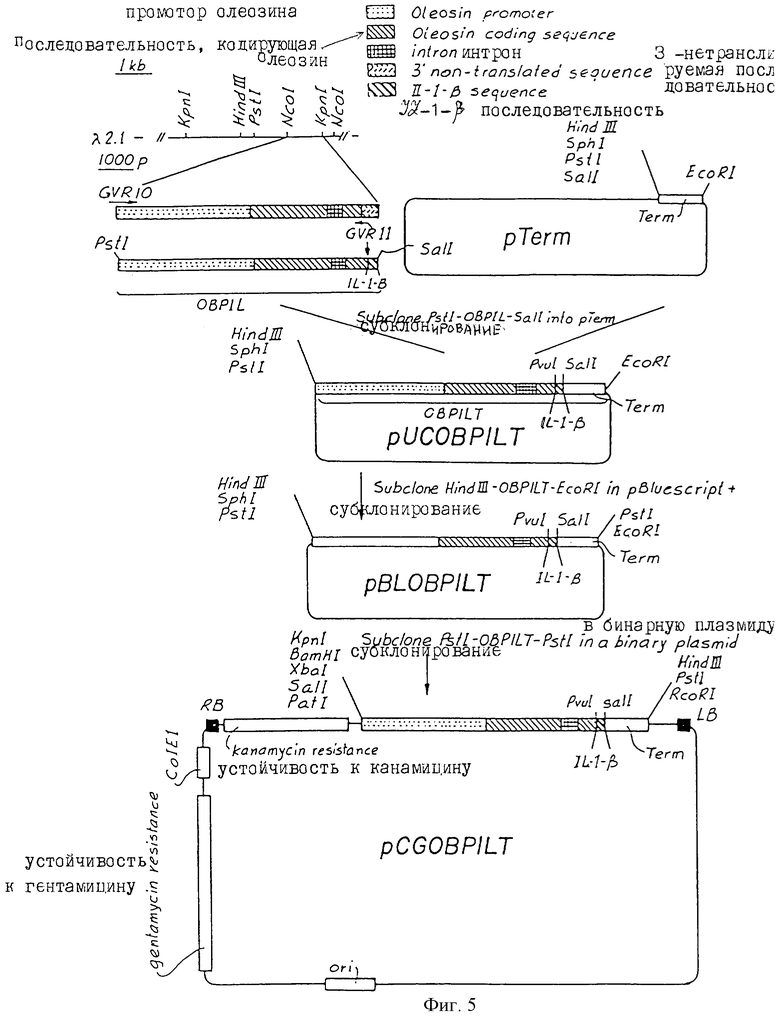
На фиг. 5 схематически представлено конструирование рCGOBPILT. Прямоугольник с пунктиром представляет олеозиновый промотор; прямоугольник, заштрихованный сверху слева-вправо вниз, представляет олеозиновую кодирующую последовательность; заштрихованный в клеточку прямоугольник представляет интрон; прямоугольник со штрихами представляет 3'- нетранслируемую последовательность; прямоугольник с редко расположенными полосами сверху слева - вправо вниз, представляет последовательность интерлейкина - I-β, снабженную последовательностью, кодирующей сайт расщепления протеазы ( Фактор Ха или тромбин, расположенный непосредственно в обратном направлении).
На фиг.6 представлена конструкция олигонуклеотида GVR11. На фиг.3А представлена 3' кодирующая последовательность А.thaliana oleosin, трансляционно слитая с кодирующей последовательностью фактора Ха/IL-1-β, после которой следует ТАА стоп-кодон. Для целей дальнейшего кодирования включают PvuI и SalI рестрикционные сайты. Создание PvuI рестрикционного сайта приводит к дополнительной кодирующей последовательности для аланина (ala). Подчеркнуты последовательности распознавания рестрикционного энзима. Друг над другом расположены последовательности олеозина А. thaliana и последовательность распознавания фактора Ха. Действительный сайт расщепления обозначен звездочкой. На фиг. 3В представлена последовательность GVR11 для осуществления слияния с А. thaliana олезином, праймер CVR11 должен быть комплементарен последовательности верхней нити.
На фиг. 7 представлена нуклеотидная последовательность OBPILT. Подчеркнута последовательность, кодирующая IL-1-β; последовательность, кодирующая сайт распознавания фактора Ха, указана жирным шрифтом. Терминаторная последовательность нопалинсинтетазы указана мелкими буквами.
Описание предпочтительного варианта
В соответствии с целью изобретения предложены способы и композиции для получения пептидов, которые легко поддаются очистке. Целевой способ включает стадии получения экспрессионной кассеты, содержащей ДНК последовательность, кодирующую достаточную часть специфической последовательности масляного тела, например олеозина, для обеспечения направленного транспорта к масляному телу, и представляющий интерес пептид; трансформации экспрессионной кассетой клеток растения-хозяина; создания трансгенного растения и выращивание его для получения семян, в которых химерный протеин экспрессирован и перемещен к масляным телам. Химерный пептид включает целевой пептид и такой протеин масляного тела, как олеозин. Целевой пептид обычно является чужеродным пептидом, который обычно не экспрессируется в семенах или не находится на масляных телах. Использование протеина масляного тела в качестве носителя или в качестве средства для направленной доставки обеспечивает простой механизм очистки чужеродного протеина. Химерный протеин выделяют из массы клеточных протеинов в одну стадию (например, за счет центрифугирования или флотации); этот протеин также защищен от разложения в процессах экстракции, так как при разделении удаляются неспецифические протеазы из контакта с масляными телами. Ген, кодирующий чужеродный пептид, может быть получен из любого источника, включая растительные, бактериальные, грибковые или животные источники.
Желательно, чтобы химерный пептид содержал последовательности, которые позволяли бы отщеплять целевой пептид от олеозина. Такой метод можно использовать для экспрессии различных пептидов, которые затем легко поддаются очистке.
Обеспечение направленной доставки чужеродного рекомбинантного протеина к масляному телу обеспечивает несколько преимуществ, включая следующие. Протеины можно выделить из клеточной массы после лизиса клеток за счет центрифугирования. Частицы масляных тел будут плавать на поверхности экстракта. Протеин можно необязательно снабдить пептидным линкером, содержащим сайт распознавания протеазы. Это позволяет отделить пептид от масляного тела. Протеин можно ввести в рекомбинантный полипептид таким образом, чтобы он был внутри липофильного консервативного участка. Это приводит к интернализации рекомбинантного пептида в масляное тело, таким образом защищая его от атаки протеазы.
Экспрессионная кассета обычно будет включать в 5'-3' направлении транскрипции транскрипционный и трансляционный регуляторный участок, обеспечивающие возможность экспрессии в развивающихся семенах, типичным примером которых служит промотор, и расположенные в обратном направлении участки, связанные с протеином масляного тела, что обеспечит экспрессию химерного протеина в семенах; ДНК последовательность, кодирующую химерный пептид, содержащий аминокислотную последовательность для обеспечения направленной доставки к масляному телу, и целевой протеин, транскрипционный и трансляционный терминальные участки, функциональные в растениях. Может также присутствовать один или более из интронов.
Специфическая последовательность масляного тела встречает аналогию во фрагментах протеинов масляного тела, в частности олеозинов. Специфическая последовательность масляного тела может быть той же, что и последовательность, получаемая из протеина масляного тела, и они имеют достаточную гомологию, чтобы обеспечить направленную доставку целевого протеина к масляному телу. Под "получаемой" подразумевают аминокислотную последовательность, которая может быть природной, синтетической или комбинированной, достаточно близкой к аминокислотной последовательности протеина нативного масляного тела, для достижения нужного обеспечения направленной доставки. Особый интерес представляют центральный гидрофобный домен протеинов масляного тела, который, по-видимому, в высшей степени консервативен среди различных видов растений, и его фрагментов и гомологичных последовательностей на аминокислотном уровне.
Выведенную последовательность 3 аминокислот для Arabidopsis thaliana протеина масляного тела см. в конце описания.
Аминокислоты от около 25 до 101 содержат центральный гидрофобный домен.
Особый интерес представляют в качестве средств, обеспечивающих направленную доставку для некоторых применений, специфические последовательности 4 масляного тела или их фрагменты последовательности, которые обеспечивают направленную доставку к масляному телу (см. в конце описания).
где:
рр1 и рр2 одинаковы или различны и могут быть такими же или могут отличаться от природного протеина масляного тела и обычно отличаются; они могут представлять водороды, что указывает на терминальную часть указанного полипептида, или они могут быть полипептидами, содержащими вплоть до 1000 аминокислот, обычно вплоть до около 500 аминокислот, и могут содержать лишь одну аминокислоту или могут индивидуально или раздельно быть полипептидами, содержащими от 1 до 100 аминокислот, обычно от около 1 до 75 аминокислот и особенно от около 5 до 50 аминокислот; эти полипептиды могут иметь специфические применения при модификации специфически описанных последовательностей для заранее определенных целей;
аа25 может быть любой аминокислотой, в частности нейтральной алифатической аминокислотой, обычно содержащей от 3 до 6 атомов углерода, более конкретно лейцином или аланином;
аа26 представляет нейтральную алифатическую аминокислоту, особенно аланин или гидроксизамещенную аминокислоту, содержащую от 3 до 4 атомов углерода, в частности треонин, или основную аминокислоту, содержащую от 5 до 6 атомов углерода, в частности лизин;
аа31 представляет нейтральную незамещенную алифатическую аминокислоту, содержащую от 3 до 6 атомов углерода, в частности аланин, валин или лейцин, или ароматическую незамещенную аминокислоту, особенно фенилананин;
аа33 представляет нейтральную незамещенную алифатическую аминокислоту, содержащую от 3 до 6 атомов углерода, особенно аланин, валин или лейцин, или оксизамещенную алифатическую аминокислоту, особенно треонин;
аа36 представляет нейтральную алифатическую незамещеную аминокислоту, содержащую от 3 до 5 атомов углерода, особенно лейцин, или нейтральную алифатическую оксизамещенную аминокислоту, содержащую от 3 до 4 атомов углерода, особенно треонин или серин;
аа37 представляет нейтральную незамещенную аминокислоту, особенно лейцин или тиозамещенную аминокислоту, особенно метиомин;
аа39 представляет нейтральную алифатическую незамещенную аминокислоту, особенно валин, или ароматическую незамещенную аминокислоту, особенно фенилаланин;
аа41 представляет нейтральную алифатическую незамещенную или оксизамещенную аминокислоту, особенно аланин, лейцин или серин;
аа44 представляет нейтральную алифатическую незамещенную или оксизамещенную аминокислоту, в частности аланин, изолейцин или треонин;
аа46 представляет нейтральную алифатическую незамещенную аминокислоту или оксизамещенную аминокислоту, особенно аланин, валин или треонин;
аа47 представляет нейтральную алифатическую незамещенную аминокислоту, особенно глицин или аланин;
аа59 представляет нейтральную алифатическую или ароматическую незамещенную аминокислоту, особенно лейцин или фенилаланин;
аа76 представляет нейтральную алифатическую незамещенную или тиозамещенную аминокислоту, особенно аланин, лейцин или метионин;
аа78 представляет нейтральную алифатическую незамещенную аминокислоту, особенно аланин, или нейтральную алифатическую аминокислоту, содержащую тио- или окси-замещения, особенно метионин или треонин;
аа83 представляет нейтральную алифатическую незамещенную или оксизамещенную аминокислоту, особенно глицин, серин или треонин;
аа92 представляет нейтральную алифатическую аминокислоту с окси-замещением, особенно серин или треонин;
аа96 представляет нейтральную алифатическую тиозамещенную аминокислоту или нейтральную ароматическую гетероциклическую аминокислоту, особенно триптофан;
аа97 представляет нейтральную алифатическую незамещенную или тиозамещенную аминокислоту, особенно валин, лейцин, изолейцин или метионин;
аа98 представляет нейтральную алифатическую незамещенную аминокислоту или ароматическую окси-замещенную аминокислоту, особенно аланин, лейцин или тирозин;
аа99 может представлять любую аминокислоту;
аа100 представляет окси-замещенную аминокислоту, либо алифатическую, либо ароматическую, особенно тирозин или треонин;
аа101 представляет нейтральную незамещенную алифатическую или ароматическую аминокислоту, особенно аланин, лейцин или фенилаланин.
Особый интерес в качестве источника ДНК кодирующей последовательности, способной обеспечить направленную доставку к протеину масляного тела, представляют гены протеинов масляных тел, получаемые из Arabidopsis или Brassica napus, которые обеспечивают экспрессию целевого протеина в семенах (см. Taylor et al. (1990), Planta 181: 18-26). Нужные участки и аминокислотные последовательности, необходимые для обеспечения способности направленной доставки к масляным телам, должны быть, по-видимому, существенно гидрофобным центральным участком протеинов масляного тела.
Для идентификации других генов протеинов масляного тела, обладающих нужными характеристиками, где протеин масляного тела уже был выделен, этот протеин можно частично секвенировать, с тем чтобы можно было сконструировать зонд для идентификации мРНК. Такой зонд особенно ценен, если он сконструирован таким образом, чтобы нацелить его к кодирующему району центрального гидрофобного домена, который в высшей степени консервативен среди различающихся видов растений. Следовательно, ДНК или РНК зонд для этого участка может быть особенно полезен для идентификации кодирующих последовательностей протеинов масляного тела из других видов растений. Для дальнейшего повышения концентрации мРНК можно получить кДНК и эту кДНК вычесть с мРНК или кДНК из клеток, продуцирующих не масляные тела. Оставшуюся кДНК можно затем использовать для зондирования генома для комплементарных
последовательностей, используя соответствующие библиотеки, полученные из растительных клеток. Затем можно выделить последовательности, которые гибридизуются с кДНК в жестких условиях.
В некоторых случаях, как указано ранее, использующих зонд гена протеина масляного тела (консервативный участок), зонд можно использовать непосредственно для скринирования геномной библиотеки кДНК и идентификации последовательностей, которые гибридизуются с зондом. Выделение можно также осуществить стандартным иммунологическим скринированием библиотеки экспрессии кДНК специфической для семян. Антитела можно легко получить к протеинам масляных тел, используя способы очистки и схемы получения антител, описанные Taylor et al. (Planta, (1990), 181, 18-26). Библиотеку экспрессии кДНК скринируют, используя антитела по способу Huynh et al. (1985 в DNA Cloning, Vol.1, а Practical Approach, ed. D.М. Glover, IRL Press, рр. 49-78). Подтверждение структуры последовательности облегчается за счет высокого консерватизма, существующего в центральном гидрофобном участке (фиг.1). ДНК секвенирование по способу Sander et al. (Proc. Natl. Acad. Sci. USA, (1997) 74:5463-5467 или Maxam and Gilbert (1980), Meth. Ensymol., (1980) 65:497-560) можно осуществить на всех предполагаемых клонах и осуществить поиски гомологов. Гомологи последовательностей, кодирующих центральный гидрофобный домен, обычно составляет ≥ 70% как на аминокислотном, так и на нуклеотидном уровнях между различающимися видами. Если доступно антитело, подтверждение идентичности последовательности можно осуществить в экспериментах отбора гибридов и трансляции из препаратов мРНК семян по способу Sambrook et al. (Molecucar Cloning, (1990), 2nd Ed., Cold Spring Harbor Press, рр.8-49 до 8-51).
кДНК клоны, полученные из семян, можно скринировать, используя кДНК зонды, полученные из консервативных кодирующих участков любого доступного гена протеина масляного тела (например, Bowman-Vance and Huang (J. Biol. Chem., (1987) 262: 11275-11279). Отбирают клоны, которые обладают степенью гибридизации с ДНК семян, нежели с кДНК сеянцев. Скринирование повторяют для идентификации конкретной кДНК, связанной с масляными телами развивающихся семян, используя прямое скринирование антителами или отбор гибридов и трансляцию. мРНК, комплементарные специфическим кДНК, отсутствуют в других тканях, подвергавшихся тестированию. Затем кДНК используют для скринирования геномной библиотеки и отбирают фрагмент, который гибридизуется с нужной кДНК.
Для достижения экспрессии химерного гена в семена участок, регулирующий начало транскрипции, и участок, регулирующий начало трансляции нетранслируемых 5' последовательностей, "рибосомосвязывающие сайты", ответственные за связывание мРНК с рибосомами, и начало трансляции, получаемые из любого гена, предпочтительно экспрессируемого в семенах, могут быть использованы. Примеры таких генов включают гены протеинов хранения семян, например из напина (Josefsson et al., J. Biol. Chem., (1987) 262:12196-12201; Scoield S. R. and Crouch N. L. J. Biol. Chem. (1987) 262: 12203-12208). Предпочтительно, чтобы участок был получен из протеина масляного тела (протеины масляных тел из Arabidopsis, моркови (Hatzopoulos et al., см. ранее) или кукурузы (Huang et al. 1987 и 1990, см. ранее). Такой участок обычно содержит, по крайней мере, 100 bp 5' до начала трансляции последовательности, кодирующей структурный ген, вплоть до 2,5 кb 5' до того же начала трансляции. Предпочтительно, чтобы в се транскрипционные и трансляционные функциональные элементы участка, контролирующего инициацию, были получены или их можно было бы получить из одного и того же гена. Под выражением "можно получить" подразумевают ДНК последовательность, достаточно сходную с нативной последовательностью для того, чтобы обеспечить целевую специфичность транскрипции ДНК последовательности, кодирующей химерный протеин. Она включает природные и синтетические последовательности и может быть комбинацией синтетических и природных последовательностей.
Уровень транскрипции должен быть достаточным для обеспечения такого количества РНК, которое способно обеспечить получение модифицированных семян. Под термином "модифицированные семена" подразумевают семена, обладающие заметно отличающимся фенотипом от фенотипа семян нетрансформированного растения того же вида, например не содержащих рассматриваемой кассеты экспрессии и в его геноме. Представляют интерес различные изменения в фенотипе. Эти изменения включают избыточную экспрессию протеина масляного тела или ОВР-накопление на масляном теле или полученного химерного протеина в цитоплазме.
Представляющим интерес полипептидом может быть любой протеин, и они включают, например, энзим, антикоагулянт, нейропептид, гормон или адгезивный предшественник. Примеры протеинов включают интерлейкин - I-β, антикоагулянт Hirudin, энзим β-глюкуронидазу или одноцепочечное антитело, содержащее трансляционное слияние VН или VL цепей иммуноглобулина. ДНК последовательность, кодирующая целевой полипептид, может быть синтетической или природной или их комбинацией. В зависимости от характера источника ДНК кодирующей целевой полипептид может оказаться желательным синтезировать ДНК последовательность с предпочтительными растительными кодонами. Предпочтительно растительные кодоны модно определить из кодонов, наиболее часто встречающихся в протеинах, экспрессированных в больших количествах в конкретные виды растений, представляющих интерес в качестве растений-хозяев.
Используемый терминаторный участок должен быть преимущественно одним из обычных, так как во многих случаях терминационные участки, по-видимому, относительно взаимозаменяемы. Терминационный участок может быть нативным с участком инициирования транскрипции, может быть нативным с ДНК последовательностью, кодирующей целевой полипептид или может быть получен из другого источника. Удобные терминационные участки доступны из Тi - плазмид А. tumefaciens, такие как октопин-синтазные и нопалин-синтазные терминационные участки.
Легирование ДНК последовательности, кодирующей обеспечивающую направленную доставку последовательность с геном, кодирующим целевой пептид, может осуществляться различными способами, включая терминальное слияние, внутреннее слияние и полимерное слияние. Во всех случаях такие слияния осуществляют таким образом, чтобы не прерывать считывающую рамку протеина масляного тела, и с тем, чтобы избежать каких-либо сигналов прекращения трансляции внутри или вблизи этих соединений. Различные типы терминальных и внутренних слияний представлены на фиг.2 наряду с представлением их конфигураций in vivo.
Во всех описанных случаях лигирование гена, кодирующего пептид, предпочтительно должно включать линкер, кодирующий протеазный мишеневый фрагмент. Это позволит осуществить выделение пептила после его экстрагирования в виде протеина слияния. Потенциальные сайты расщепления. которые можно использовать, являются фрагментами для тромбина (leu-val-pro-arg-gly) (Fujlkawa et al. , Biochemistry (1972) 11:4892-4899), фактора Ха (phe-glu-gly-arg-аа) (Nagat et al. , Proc. Nat'l Acad. Set. USA, (1985 82:7252-7255) или коллагеназы (pro-leu-gly-pro) (Scholtissek and Grosse Gene (1988) 62:55-64).
За счет соответствующих манипуляций, таких как рестрикция, переваривание (chewing back) или заполнение липких концов для создания тупых концов, легирование линкеров или т.п., комплементарные концы фрагментов можно получить соединением и легированием. При проведении различных стадий используют клонирование с тем, чтобы умножить количество ДНК и обеспечить возможность анализа ДНК, для того чтобы убедиться, что операция прошла должным образом. Доступно множество векторов клонирования, когда вектор клонирования включает функциональную в Е. coli систему репликации и маркер, который обеспечивает селекцию трансформированных клеток. Иллюстративные векторы включают рВR332, рUC серии, М13mp серии, PACYC 184 и т.д. Так, последовательность можно встроить в вектор по соответствующему рестрикционному сайту (сайтам), полученную плазмиду использовать для трансформации Е.coli хозяина, вырастить Е. coli в соответствующей питательной среде, собрать клетки, провести лизис и выделить плазмиду. Анализ может включать анализ последовательности, рестрикционный анализ, электрофорез или т.п. После каждой манипуляции ДНК последовательность, которая должна быть использована в окончательной конструкции, может быть рестриктирована и соединена со следующей последовательностью, где каждая из частичных конструкций может быть клонирована в ту же самую или в отличающуюся плазмиду.
Для введения ДНК в растительные клетки хозяина существует множество возможных методик. Так например, химерные ДНК конструкции можно вводить в клетки хозяев, полученные из двудольных растений, например табака, и таких масличных видов, как Brassica napus, используя стандартные Agrobacterium векторы, по такой схеме трансформации, как например та, которая описана Moloney et al. Plant Cell Rep., (1989) 8:238-242 или Hinchee et al/ Bio/Technol., (1988) 6:915-922; или другие методики, известные специалистам. Так например, использование Т-ДНК для трансформации растительных клеток интенсивно исследуется и подробно описано в ЕРА 120516; Hcekema, В "The Binary Plant Vector System Offset-drukkerij Kanters В.V., Alblasserdam, 1985, Chapter V., Knauf, et al., Genetic Analysis of Host Range Expression by Agrobacterium, В "Molecular Genetics of the Bacteria. Plant Interaction, Puhler, А.ed., Springer-Verlag, NY, 1983, р.245, and An et al., EMBO J. (1985), 4:277-284.
Удобно эксплантаты культивировать с А. tumefacietns или А. rhizogenes для того, чтобы обеспечить перенос транскрипционной конструкции в растительные клетки. После трансформации с использованием Agrobacteria растительные клетки диспергируют в подходящей селективной среде для отбора, выращивают до образования каллуса, проростки и молодые растения регенерируют из каллуса, выращивая в среде для образования корней. Agrobаcterium хозяин будет содержать плазмиду, содержащую Vir гены, необходимые для переноса Т-ДНК в растительные клетки, и может содержать или не содержать Т-ДНК. Для инъекции и электропорации (см. далее) обнаруженные Т1 - плазмиды (не содержащие опухолевых генов, особенно Т-ДНК участка) можно вводить в растительные клетки.
Использование не-Agrobаcterium методик позволяет использовать описанные здесь конструкции для осуществления трансформации и экспрессии в широкий круг однододольных и двудольных растений. Эти методики особенно подходят для таких видов, которые не дают результатов в Agrobаcterium трансформационной системе. Другие методики для переноса генов включают биолизис (Sanford, Trends in Biotech. (1988) 6:299-302), электропорацию (Fromm et al. (1985) Proc. Nat'l. Асаd. Sci. USA 82:5824-5828; Riggs and Batec (1986), Proc. Nat'l. Acad. Sci. (USA) 83, 5603-5606 или захват ДНК с помощью PEG (Potrykus et al. (1985), Mol. Gen. Genet. 199:169-177).
В качестве клеток хозяев можно использовать клетки любых растений, имеющих семена, причем эти клетки получают на таких частей растений, как стебли, листья, корни или семена, или соответствующих виду репродуктивных структур. Клетки могут быть как выделенными клетками, так и частями растений, например листовыми дисками. В специфических применениях, таких как с Brassica napus, клетки хозяева обычно получают из семядольных петиолей, как указано Moloney et al. Plant Cell Rep. (1989) 8:238-242). В других примерах с использованием коммерческих маслосодержащих семян используют трансформацию семядолей в соевые эксплантаты (Hihchee et al. Вiotechnology, (1988) 6: 915-922) и трансформацию стебля хлопка (Umbeck et al. Biotechnology, (1981) 5:263-266).
После трансформации клетки, например, в виде дисков листьев, выращивают на селективной среде. После того как начинают образовываться ростки, их иссекают и помещают на корнеобразующую среду. После достаточного развития корней растения переносят в почву. Затем предположительно трансформированные растения тестируют на присутствие маркера. Осуществляют Саузерн-блоттинг на геномной ДНК, используя соответствующий зонд, например, А.thaliena олеозиновый ген для доказательства интеграции целевой последовательности в геном клетки хозяина.
Экспрессионную кассету обычно соединяют с маркером для селекции в растительных клетках. Удобно, чтобы маркер был устойчивым к гербицидам, например, антибиотику, такому как канамицин, G41В, блеомицин, гигромицин, хлорамфеникол и т.п. Конкретным используемым маркером должен быть такой, который обеспечит селекцию трансформированных клеток при сравнении с клетками, которые не содержат ДНК, которую вводили.
Пептид слияния в кассете экспрессии, сконструированной, как указано ранее, экспрессируется, по крайней мере, предпочтительно, в развивающихся семенах. Соответственно, трансформированным растениям, выращенным в соответствии с принятыми способами, дают возможность завязать семена. См. например, McCormick et al. Plant Cell Reporta (1986) 5:81-84. Норзерн-блоттинг можно вести, используя соответствующий генный зонд с РНК, выделенной из ткани, в которой ожидается транскрипция, например в эмбрионе семени. Затем размеры транскриптов можно сравнить с размерами, предсказанными для транскриптов протеина слияния.
Можно вырастить два или более поколений трансгенных растений и либо опылить их пыльцой от того же трансформированного штамма, либо от других штаммов, контролируя в полученном гибриде нужные фенотипические характеристики для подтверждения того, что нужные фенотипические характеристики стабильно сохраняются и наследуются, а затем семена собирают для того, чтобы выделить представляющий интерес пептид или для того, чтобы получить семена, обладающие новыми фенотипическими характеристиками.
Целевой протеин можно экстрагировать из семян, которые могут быть гомо- или гетерозиготными, для последующих испытаний различными способами, которые включают использование водных, забуференных экстракционных страд и различные способы измельчения (размалывания, раскалывания, пульверизации) для разрушения клеток семян. Затем продукты экстракции можно разделить (например, центрифугированием или за счет седиментации) на три фракции: седимент, или нерастворимый осадок, водный супернатант и плавающую пленку, содержащую липид хранения семян и масляные тела. Эти масляные тела содержат как нативные протеины масляных тел, так и химерные протеины масляных тел, причем последние содержат чужеродные пептиды. Масляные тела выделяют из водорастворимых протеинов и повторно суспендируют в водном буфере.
Если в кассету экспрессии был включен линкер, содержащий фрагмент распознавания протеазы, тогда к повторно суспендированному буферу добавляют протеазу, специфичную для фрагмента распознавания, полученного в результате трансляции линкерной последовательности. Это приводит к выделению целевого пептида в водную фазу. Теперь в результате второй стадии центрифугирования снова получают плавающую пленку обработанных масляных тел с присоединенными к ним протеинами и остается водный раствор целевого пептида. Этот целевой пептид можно осадить, химически модифицировать или лиофилизировать в зависимости от предполагаемого применения.
В некоторых случаях применения нет необходимости отделять химерный протеин от протеинов масляного тела. Такие применения включают случаи, в которых пептид слияния включает энзим, который толерантен к N- или С-терминальному слиянию и сохраняет свою активность; такие энзимы могут быть использованы без дальнейшего расщепления и очистки. Химерный энзим-ОВР будет контактировать с субстратом как протеин слияния. Возможно также при желании провести очистку энзим-ОВР протеина слияния, используя иммуноафинную колонку, содержащую иммобилизованные антитела с высоким титром против ОВР (см. например, Taylor et al. (1990 ранее).
Возможно другое применение предмета изображения. Так как ОВР содержат высокий процент протеинов семян, оказывается возможным снабдить семена для некоторых целей такими ценными свойствами, как высокое содержание лизина метионина и т.д., просто обогащая протеин слияния нужными аминокислотами. Этот способ может найти применение особенно при модификации зерновых, которые прямо или косвенно используются для корма домашних животных, включая скот, птицу, а также для продуктов питания людей. Может оказаться возможным включать как пептид слияния энзим, который облегчит последующую обработку масла или муки в обычных процессах измельчения и экстракции семян масличных растений, например, за счет включения термостабильного липид-модифицирующего энзима, который останется активным и при повышенных температурах измельчения, которые обычно наблюдаются в этом процессе обработки семян, и тем самым удастся повысить ценность экстрагированных триглицеридов или протеинов.
Другие применения протеинов слияния включают использование их для повышения устойчивости сельскохозяйственных растений и неблагоприятным условиям. Так например, с протеином масляного тела можно соединить инсектицидный протеин или часть иммуноглобулина, специфичного для сельскохозяйственных вредителей, такую как стенки клеток или мембран грибков, тем самым уменьшая опасность нападения на семена конкретных вредителей растений.
Приводимые далее примеры представлены лишь в целях иллюстрации и не являются ограничивающими.
Экспериментальная часть
Пример 1
Экспрессия терминальных слияний чужеродных пептидов с протеинами масляных тел
А. С-терминальные слияния
Геномный клон гена протеина масляного тела, содержащий, по крайней мере 100bp 5' до начала трансляции, клонируют в плазмидном векторе, способном к репликации в подходящем бактериальном хозяине (например. рUC или рBR322 в Е. coli. ). Рестрикционный сайт расположен в участке, кодирующем гидрофильную С-терминальную часть гена. В 19 кДa ОВР этот участок простирается обычно от кодонов 125 до конца клона. Идеальный рестрикционный сайт является уникальным, но это не является абсолютно необходимым. Если в этом участке нет удобного рестрикционного сайта, его можно ввести за счет сайт-направленного мутагенеза по Kunkel Proc. Nat'l. Acad Sci. USA, (1985) 82:488-492. Единственным основным ограничением по встраиванию этого сайта является то, что он должен быть расположен в направлении 5' от трансляционного стоп сигнала ОВР клона.
Получив такой мутантный клон, можно получить синтетический олигонуклеотидный адаптор, который содержит кодирующую последовательность для сайта распознавания протеазы, например, Pro-Leu-Gly-Pro или его мультимер. Это сайт распознавания для протеазы коллагеназы. Адаптор следует синтезировать таким образом, чтобы обеспечить: 4-основной выступ у 5' конца, совместимый с рестрикционным сайтом 3' конца ОВР клона, 4-основной выступ у 3' конца адаптора для облегчения легирования с последовательностью, кодирующей чужеродный пептид, и дополнительными основаниями при необходимости, чтобы обеспечить отсутствие сдвигов рамки в переходе между ОВР кодирующей последовательностью, сайтом распознавания протеазы и последовательностью, кодирующей чужеродный пептид. Типичная конструкция такого слияния представлена на фиг.3. Представленный пример использует существующий Xhol сайт вблизи стоп кодона ОВР моркови (Hatzopoulos et al. Plant Cell, (1990) 2:457-467. Его переваривают, и он может быть легирован с адаптором, сконструированным из двух описанных олигонуклеотидов. Этот адаптор образует прекрасный Nсol выступ на конце и не будет нарушать трансляционной рамки. Другой конец образует Ncol выступ, который выбирают произвольно (подходят любые отрезки из 6 оснований), но который содержит ATG из целевого чужеродного пептида.
Окончательный продукт легирования содержит самое большее полный ОВР ген, последовательность, кодирующую фрагмент распознавания коллагеназы, и участок, кодирующий целевой пептид, причем все в единой считывающей рамке. Этот состоящий из трех частей фрагмент клонируют в Agrobacterium бинарную плазмиду (Bevan Nucl. Acid Res. (1984) 12: 8711-8721), такую, как обычно используют для переноса чужеродной ДНК в растения Fraley et al. Proc. Nat'l, Acad. Sci. USA (1983) 80:4803-4807), и это используют для трансформации семян масличных растений, таких как рапс, используя способ Moloney et al. Plant Cell Rep. , (1989) 8:238-242) или аналогичную процедуру. Трансгенные растения могут быть получены из этого эксперимента по трансформации и выращены до цветения. Затем получают семена растения в результате самоопыления.
Семенам дают вызреть (60-80 дней), а затем собирают и измельчают в водном экстракционном буфере (Taylor et al. Planta, (1990) 181:18-26). Полученную суспензию центрифугируют при 5000хg в течение 20 минут, в результате получают на поверхности пленку. Ее снова выделяют и суспендируют при интенсивном встряхивании в коллагеназном аналитическом буфере (Scholtissek and Grosse, Gene (1998) 62:55-64). Добавляют 5 ед. коллагеназы и суспензию инкубируют со встряхиванием в течение 4 часов. После этого суспензию снова центрифугируют при 5000 хg в течение 20 минут. Удаляют поверхностную пленку и определяют содержание протеина в водной фазе с помощью электрофореза на SDS-полиакриламидном геле. Если обнаруживают полосу приблизительно того же размера, что и нужный пептид, протеин можно осадить, используя сульфат аммония, концентрирование с использованием ультрафильтрации, или лиофилизировать.
В. N-терминальное слияние
Гидрофильный N-терминальный конец протеинов масляных тел позволяет осуществить слияние пептидов с N-концом, обеспечивая при этом сохранение чужеродного пептида на внешней поверхности масляного тела. Конфигурации таких слияний представлены на фиг. 2IВ.
Такую конфигурацию можно сконструировать из таких же исходных материалов, что были использованы для С-терминальных слияний, но необходима идентификация удобных рестрикционных сайтов вблизи начала трансляции гена протеина масляного тела. Удобный сайт можно создать для многих генов протеинов масляных тел без изменения в кодирующей последовательности за счет введения изменения одного основания сразу в направлении 5' к первому "АТG". В исследованных до настоящего времени протеинах масляных тел второй аминокислотой является аланин, кодон которого начинается с "G". Контекст последовательности 5 представлен в конце описания.
Изменение одного основания у аденина перед "ATG" дает в обоих случаях... ССATGG..., что представляет собой Ncol сайт. Таким образом, модификация этого основания с помощью сайт-направленного мутагенеза по Kunkel (Proc. Nat'l. Acad. Sci. USA, (1985) 82:488-492) дает возможность подготовить этот клон для использования, предполагая, что в последовательности нет других Ncol сайтов.
Кодирующая последовательность для чужеродного пептида может потребовать изменений, которые обеспечат ее лигирование непосредственно по Ncol сайту. Обычно это может потребовать изменения одного или двух оснований за счет сайт-направленного мутагенеза (Kunkel, 1985, ранее) для создания Ncol сайта около начала трансляции чужеродного пептида. Затем этот пептид иссекают из его вектора клонирования, используя Ncol и второй энзим, который разрезает вблизи остановки трансляции мишени. И снова, используя описанный ранее способ, второй удобный сайт можно ввести за счет сайт-направленного мутагенеза. Было предложено, Qu and Huang (1990, ранее), что N-терминальный метионин может быть удален в процессинге протеина in vivo и что аланин, который следует непосредственно в прямом направлении от него, может быть ацилирован. Чтобы учесть эту возможность, может оказаться необходимым сохранить последовательность Met-Ala по N-терминальному концу протеина. Это легко осуществить, используя различные стратегии введения удобного рестрикционного сайта в кодирующую последовательность в Ala кодон или после него. Так например, за счет сайт-направленного мутагенеза последовательность 6 можно модифицировать (см. в конце описания).
Такое изменение одного кодона позволяет ввести SphI сайт в кодирующую последовательность. Второе изменение, которое можно осуществить в том же цикле мутагенеза, превращает два основания в кодоне 6 до получения GGC, GCC, что представляет Nar1 сайт. Этот полученный в результате мутаций ген можно затем раскрыть Sph1 и Nar1 до получения отрезка прямого клонирования, который исключает три кодона. В этот сайт можно ввести адаптор, содержащий 3' выступ с последовательностью CATG. .. (совместимую с Sph1) GC 5' выступ с противоположного конца.
Точная последовательность 7 этого адаптора представлена в конце описания.
Этот адаптор будет воссоздавать как Sph1 так и Nar1 рестрикционные сайты, которые можно использовать для диагностических целей. Sph1 сайт теперь можно использовать для раскрытия плазмиды и клона в рамке ДНК фрагмента, содержащего последовательность для нужного пептида. Затем следует проанализировать ориентацию клонирования за счет нарезания по любым несимметрично расположенным сайтам и Nar1 плазмиды.
Полученные из этих N-терминальных слияний конструкции могут быть типичными примерами IВ фиг. 2. Они должны содержать ОВР промоторную последовательность в первых нескольких кодонах ОВР гена, представляющего высокую ценность пептида с его собственной АТG в качестве стартового сигнала при необходимости и остальную часть ОВР гена и терминатор.
Этот модифицированный ген вводят в бинарную Agrobacterium плазмиду (Bevan. (1984, ранее) и мобилизуют в Agrobacterium. Трансформации осуществляют в соответствии с вышеприведенным описанием. Выделение ценного пептида из семян осуществляют по способу, описанному в разделе "С-терминальные слияния".
С. Внутренние транисляционные слияния
Третий тип слияний включает помещение кодирующей ценный пептид последовательности внутрь кодирующей последовательности ОВР. Этот тип слияния требует той же стратегии, что и N-терминальное слияние, но может быть функциональным лишь при модификациях в участках с низкой консервативностью, так как считают, что участки с высокой степенью консервативности в этих ОВР существенны для обеспечения направленной доставки зрелого протеина.
Ключевое отличие в этом типе слияния состоит в необходимости фланкирующих сайтов распознавания коллагеназы для высвобождения протеина. Это означает, что вместо стандартной коллагеназной системы линкер/адаптор, описанной ранее, необходимо иметь линкер вида, представленного в последовательности 8 (см. в конце описания).
Липкие концы 1 и 2 будут использованы для клонирования адаптора в ОВР клон непосредственно. Затем встроенный рестрикционный сайт используют для введения последовательности, кодирующей ценный пептид, фланкированный рестрикционными сайтами или линкерами. Ориентацию контролируют, используя асимметрично расположенный рестрикционный сайт в последовательности, кодирующей ценный пептид, и один из двух рестрикционных сайтов, фланкирующих последовательность, кодирующую фрагмент распознавания коллагеназы.
Мобилизация этих конструкций в Agrobacterium плазмиды, а затем в растения идентична ранее описанной процедуре. Выделение ценного протеина из семян трансгенных растений несколько отличается тем, что после того как масляные тела выделяют и промывают, может понадобиться удалить липиды с масляных тел для того, чтобы сделать доступными сайты распознавания коллагеназы, которые могут быть спрятаны внутри масляных тел в липидной фазе. Эта стадия может уменьшить некоторые преимущества использования протеинов масляных тел в качестве носителей, но может, с другой стороны, оказаться очень удобной для последовательностей протеинов, которые лабильны в водной среде или в растительных цитоплазмах.
Д. Интердимерные трансляционные слияния
Возможно создать конструкцию, в которой целиком повторяется кодирующая последовательность ОВР. Димерный протеин, получаемый из такой конструкции, может все еще содержать все необходимые факторы для обеспечения направленной доставки ОВР к масляным телам. Такая конструкция должна содержать промоторный участок, полную или почти полную открытую рамку считывания для ОВР, но без остановки трансляции, и затем полную открытую считывающую рамку второго ОВР, на этот раз укомплектованного трансляционным "стоп" и терминаторным участком.
При конструировании этого химерного гена пару различающихся рестрикционных сайтов либо находят, либо создают в участке соединения этих двух копий. Эти сайты используют для обеспечения возможности введения такого линкера, как описанный ранее для внутренних трансляционных слияний. Линкеры содержат не только наборы фрагментов распознавания коллагеназы, но также внутренний рестрикционный сайт, в который вставляют последовательность, кодирующую представляющий высокую ценность протеин. Форма такой конструкции представлена на фиг. 2 III. Введение этой конструкции В Agrobacterium, а затем в растении осуществляется описанным ранее способом. Выделение представляющего высокую ценность протеина из семян трансформированных растений осуществляют по способу, описанному ранее для процедуры С-терминального слияния.
Пример 2.
Стратегия клонирования и экспрессии Интерлейкина-1-β (II-1-β) в результате слияния с олеозинами в растениях
А. Клонирование и секвенирование гена олеозина
Arabidopsis thaliana.
Ген олеозина Brassica napus (Murphy et al., (1991) Biochim Biophys Acta 1086: 86-94) используют для скринирования геномной библиотеки А.thliana (cv. Columbia) в ЕМВL3А (Stratagene). В результате скринирования получают выделенный КМВL3А клон (λ 2,1), содержащий 15 kb геномный фрагмент из А. thaliana. Олеозин картируют внутри 6,6 kb Kpn1 вставки, внутри этого 15 kb фрагмента (фиг.5). 1,8 kb Ncol/Kpn1 фрагмент, содержащий ген олеозина, имеет тупой конец, и его субклонируют в SmaI сайт RFM13mp19. 1,8 kb вставку переваривают подходящими рестрикционными энзимами и субклонируют в М13mp19 для секвенирования. Последовательность 1800 kb гена олеозина А.thaliana представлен на фиг.1а. Все процедуры клонирования осуществляют по Sambrook et al. , (1989) (Molecular Cloning: А laboratory manual 2nd ed. Сold Spring Harbor Laboratory Press.).
В. Конструирование олигонуклеотида, кодирующего II-1-β
IL-1-β состоит из 9 аминокислот (аа); val-gln-gly-glu-glu-ser-asn-asp-lys (Antoni et al., (1986) J. Immunol. 137:3201-3204). Протеазный фактор Ха может расщепить последовательность протеина, которая содержит последовательность аминокислот ile-glu-gly-arg. Расщепление происходит после аминокислоты arg. На основании этих последовательностей конструируют олигонуклеотид (GVR11, фиг.5), который содержит дополнительно к IL-1-β кодирующей последовательности кодирующую последовательность для сайта расщепления фактора Ха и 18 нуклеотидов 3' кодирующего участка А. thaliana олеозина (положения оснований 742-759). Сконструирована IL-1-β кодирующая последовательность, используя оптимальное использование кодона для В.napus и А. thaliana олеозина (таблица 1)
С. Создание А. thaliana олеозин IL-1-β слияния. На основании последовательности 9 была проведена следующая операция (см. в конце описания). GVR10 содержит Pst1 рестрикционный сайт (подчеркнут) для облегчения клонирования. Полимеразная цепная реакция (PCR) была использована для амплификации участка между CVR10 и CVR11. Реакционная смесь содержит: 16 мкл dNTRs (1,25 мМ), 10 мкл 10Х PCR буфер (100 мМ Tris-НС1 рН 8,3, 500 мМ, КС1, 15 мМ MgC12, 0,1 % (вес/объем) желатин, 5 мкл Н2О. Реакцию ведут в течение 30 циклов. Каждый цикл включает 1 минуту денатурирования при 92oС, 1 минуту отжига при 45oС и 3 минуты удлинения при 72oС. В результате реакции PCR получают один фрагмент длиной 1652 нуклеотида.
D. Клонирование А. thaliana олеозин - IL-1-β (ОВPIL) слияния
5' SalI-нопалин-синтетазную (nos) терминаторную ЕсоRI 3' последовательность выделяют из рВI121 (Clontech laboratories) и клонируют в Sal/EcoRI сайты рUС19. Плазмиду называют pTerm. Фрагмент 1652 bp (описан в разделе С) выделяют и переваривают рестрикционными энзимами PatI и SalI. Этот фрагмент клонируют в pTerm. Полученную плазмиду называют рUCOBPILT (фиг.5). Эту плазмиду переваривают EсоRI и PstI и получают в переваре рUC19 вектор и ЕсоRI- А. thaliana олеозин-IL-1-β - nos - PstI слитый PstI (OBPILT). Полная последовательность OBPILT представлена на фиг.7. OBPILT субклонируют в ЕсоRI/PstI сайты pBluescript+. Эту плазмиду (рBIOBPILT) переваривают PstI или HindIII и фрагмент PstI-OBPILT-HindIII субклонируют в бинарную Argobacterium плазмиду (Bin 19) (Bevan. М., (1984) Nucl.Acid.Res. 12:8711-8721), содержащую селекционный маркер (неомицинфосфотрансферазу) и PstI-HindIII уникальные сайты. Полученную плазмиду называют рСGOBPILT. Схема процедуры клонирования представлена на фиг. 5. Для описания различных бинарных плазмид смотри рGA642 или 645; An et al. (1985) ЕМВО. J. 4 277-288 или рСGN1558 или 1559; MacBride and Summerfeldt (1990) Plant. Molec. Biol. 14.269-276.
F. Трансформация рСGOBPILT в Agrobacterium штамм ЕНА101.
Отдельную ЕНА101 колонию (Hood et al., (1986) J. Bact. 168:1291-1301) используют для того, чтобы инокулировать 5 мл LB+100 мкг/мл канамицина. Эту культуру выращивают в течение 48 часов при 28oС. 5 мл этой культуры используют для инокулирования 500 мл LB+100 мкг/мл канамицина. Эту культуру выращивают при 28oС до тех пор, пока плотность культуры не достигает ОД6СС= 0,5 (приблизительно 4 часа). В результате центрифугирования клетки осаждают (10 мин, 5000хg) и снова суспендируют в 500 мл стерильной Н2О (повторяют 2 раза). Клетки снова центрифугируют и снова суспендируют в 3 мл стерильной Н2О, содержащей 10% глицерина. Аликвоты по 40 мкл помещают в ампулы Эппендорфа и либо непосредственно используют для электропорации, либо хранят при -80oС до дальнейшего использования. Электропорацию осуществляют по способу Bower et al., Nucl. Acid. Res. (1888) 16 6127-6145. Мощность импульсного генератора устанавливают 25 мкФ, 2,5 кВ и 200 Ом параллельно с камерой образца.
G. Трансформация Nicotina tabacum (табак) с помощью рСGOBPILT
Для трансформации дисков листьев табака используют ЕНА101, содержащий рСGOBPILT. Листья табака длиной 8-10 см берут с растений, растущих в оранжерее, стерилизуют в 70% этаноле в течение 20 секунд, а затем в 10% хлорной извести (например, Javex) в течение 8 минут. Затем листья 6 раз промывают стерильной водой. Края листьев, а также среднюю жилку иссекают, а остальную часть листовой пластины нарезают на квадраты 5х7 мм или диски диаметром 5 мм. Собирают около 30 листовых дисков и помещают в небольшую чашку Петри. Затем на диски выливают раствор Agrobacterium и инкубируют в течение 9 минут. Затем осуществляют блоттинг на стерильной Ватманской фильтровальной бумаге и помещают плоской стороной вниз на среду I (MS, 3% сахарозы и 2 мг/л 2,4-Д). Такое совместное культивирование ведут в течение последующих 48 часов. В этот момент диски листьев переносят на селекционную среду (MD, 3% сахарозы, 2,5 мг/л Ва, 0,1 мг/1 NAA, 500 м г/л карбенициллина и 100 мг/л канамицина), где и оставляют затем на 3-4 недели. После появления проростков их иссекают и помещают на корнеобразующую среду (MS, 3% сахарозы, 0,1 мг/1 NAA, 500 мг/л карбенициллина и 50 мг/л канамицина). После образования достаточной корневой системы растения табака переносят в почву.
Н. Трансформация В. napus с помощью рСGOBPILT
Трансформация на В. napus ведут по способу Noloney et al., (1989) Plant Cell Rep., 8: 238-242 (включено сюда по ссылке).
Процедура трансформации
Отдельные колонии Agrobаcterium tumefaciens штамма ЕНА 101, содержащие бинарную плазмиду, выращивают в течение ночи при 28oС на АВ среде. Образец 50 мкл надосадочной жидкости этой среды выращивают в течение ночи при 28oС в 5 мл бульона MG/L, дополненного соответствующими антибиотиками. Эту бактериальную суспензию осаждают центрифугированием в течение 15 минут при 10000хg, затем повторно суспендируют в 10 мл MS среды, содержащей 3% сахарозы при рН 5,8. Тонкую пленку этой суспензии используют для выстилания дна чашки Петри диаметром 5 см. Отдельные вырезанные семядоли берут с описанных ранее пластин и срезанную поверхность их петиолей погружают в эту бактериальную суспензию на несколько секунд. Их немедленно возвращают на те же самые MS пластины, с которых их взяли. Семядоли культивируют совместно с Agrobacterium в течение 72 часов; при этом не используют питательных слоев.
После совместного культивирования семядоли переносят на регенерационную среду, содержащую MS среду, дополненную 20 мкМ бензиладенина, 3% сахарозы, 0,7% фитагара, рН 5,8 и 500 мг/л карбенициллина (Pyopen, Ayerаt) и 15 мг/л канамицин-сульфата (Boehringer-Mannheim. Снова петиоли осторожно вставляют в агар на глубину 2 мм. Плотность растений поддерживают около 10 эксплантатов на пластину. Более высокая плотность снижает регенерируемость.
Отбор и регенерация растений
Эксплантаты выдерживают на регенерационной среде в специфических условиях освещенности и температуры в течение 2-3 недель. За это время более чем на половине эксплантатов появляются ростки при относительно небольшом образовании каллуса. Некоторые из этих ростков обесцвечиваются к четвертой неделе культивирования. Оставшиеся зеленые ростки субкультивируют на среде для удаления корней, которая представляет собой регенерационную среду, но без бензиладенина. Одна-две недели на этой среде обеспечивают установление апикального доминирования из образовавшихся ростковых кластеров. Полученные таким образом ростки переносят на "корнеобразующую" среду, содержащую MS среду, 3% сахарозы, 2 мг/л индолмасляной кислоты, 0,7% фитагара и 500 мг/ л карбенициллина. На этой стадии не используют канамицин, так как было обнаружено, что без селектирующего агента происходит более быстрое корнеобразование, тогда как очень мало "промахов" реально наблюдается на корнеобразовании после двух раундов отбора на регенерационной и удлиняющей корни среде.
I. Стабильная интеграция ОВPILT в геномы табака и В. napus
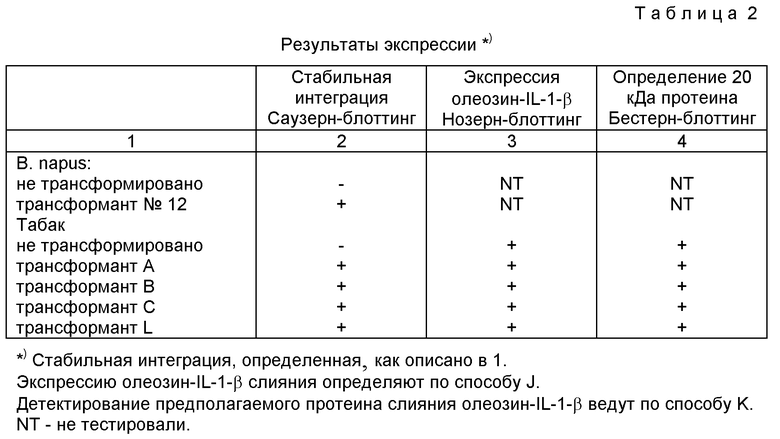
Предположительно трансформированные растения тестируют на активность неомицин-фосфотрансферазы. Выделяют геномную ДНК из растений, демонстрирующих такую активность. Саузерн-блоттинг осуществляют для демонстрации того, что последовательности между Т-ДНК границами (OBPILT и геном неомицин-фосфотрансферазы) стабильно интегрированы в геномы В. napus и табака. Саузерн табака зондируют А. thaliana олеозиновым геном, а геном неомицин-фосфотрансферазы. В. napus Саузерн зондируют геном неомицин-фосфотрансферазы.
J. Экспрессия олеозин-IL-I-β слияния в растениях табака РНК выделяют из развивающихся эмбрионов, полученных из трансформированных и нетрансформированных растений. Нозерн-блотинг осуществляют, используя А.thaliana олеозин в качестве генного зонда. Во всех тестированных трансформированных растениях можно обнаружить 850-нуклеотидный транскрипт. Размеры этих транскриптов соответствуют ожидаемым размерам мРНК олеозин-IL-1-β. Эти транскрипты обнаруживаются в нетрансформированных растениях.
К. Накопление олеозин -IL-1-β протеина
Протеины масляных тел выделяют из трансформированных семян табака (Holbrook et al. , (1991) Plant Phyaical 97:1051-1058. Осуществляют РАGE и протеин переносят из геля на РVDF мембраны. Для определения слияния олеозин-IL-1-β в семенах табака используют антитела, которые вырабатываются против 22 кДа олеозина В. napus. Эти антитела распознают все основные олеозины в В. napus и А. thaliana. Кроме того, эти антитела распознают олеозины табака. Олеозины табака имеют размеры, отличающиеся от олеозинов А.thaliana и В. napus. В трансформированных семенах табака анти-22 кДа антитела распознают 20 кДа - протеин, который отсутствует в семенах нетрансформированных растений. Предсказанный размер олеозин-IL-1-β слияния составляет 20,1 кДа.
Полученные результаты представлены в таблице 2.
За счет экспрессии представляющего интерес пептида, конъюгированного с протеином масляного тела или его частью, достаточной для обеспечения его направленного транспорта к масляным телам, представляющий интерес пептид можно легко очистить, то есть практически отделить его от других клеточных компонентов. Протеин слияния можно расщепить после очистки или его можно использовать далее без расщепления. Рассматриваемые способы и композиции обеспечивают быстрый простой способ очистки представляющего интерес полипептида.
Все публикации и патентные заявки, указанные в этом описании, включены сюда по ссылке.
Далее настоящее изобретение раскрыто полностью, но специалистам должно быть ясно, что не выходя за рамки объема и сути прилагаемой формулы изобретения, можно осуществить множество изменений и модификаций.









Изобретение относится к генетической инженерии и может быть использовано для получения представляющего интерес белка в семенах растения. Предложена химерная ДНК-конструкция, включающая последовательность, кодирующую часть белка масляного тела - олеозина, которая определена как достаточная для обеспечения переноса к масляному телу, и последовательность, кодирующую предназначенный для синтеза в растении чужеродный белок, между которыми дополнительно может быть встроена последовательность, кодирующая сайт протеазного расщепления. Путем помещения указанной ДНК-конструкции под контроль промотора гена, в норме экспрессирующегося в семени растения, получена кассета экспрессии, которая использована для трансформации растительных клеток. Семена выраженного трансгенного растения использованы для получения представляющего интерес белка, синтезированного в виде белка слияния с олеозином. Накопление экспрессированного белка во фракции масляных тел существенно облегчает его выделение и очистку. 3 с. и 12 з.п.ф-лы, 7 ил., 2 табл.
M-M-G-R-D-R-D-Q-Y-Q-M-S-G-R-G-S-D-Y-S-K-S-R-Q-I-A-K-A-A-T-A-V-T-A-G-G-S-L-L-V-L-S-S-L-T-L-V-G-T-V-I-A-L-T-V-A-T-P-L-L-V-I-F-S-P-I-L-V-P-A-L-I-T-A-V-L-L-I-T-G-F-L-S-S-G-G-F-G-I-A-A-I-T-V-F-S-W-I-Y-K-Y-A-T-G-E-H-P-Q-G-S-D-K-L-D-S-A-R-M-K-L-G-S-K-A-Q-D-L-K-D-R-A-Q-Y-Y-G-Q-Q-H-T-G-G-E-H-D-R-D-R-T-R-G-G-Q-H-T-T,
и б) второй пептид, выбираемый из группы, включающей цитокин, фермент, антикоагулянт, нейропептид, гормон, антитело или предшественник молекулы адгезии.
V-Q-G-E-E-S-N-D-K.
M-M-G-R-D-R-D-Q-Y-Q-M-S-G-R-S-G-D-Y-S-K-S-R-Q-I-A-K-A-A-T-A-V-T-A-G-G-S-L-L-V-L-S-S-L-T-L-V-G-T-V-I-A-L-T-V-A-T-P-L-L-V-I-F-S-P-I-L-V-P-A-L-I-T-V-A-L-L-I-T-G-F-L-S-S-G-G-F-G-I-A-A-I-T-V-F-S-W-I-Y-K-Y-A-T-G-E-H-P-Q-G-S-D-K-L-D-S-A-R-M-K-L-G-S-K-A-Q-D-L-K-D-R-A-Q-Y-Y-G-Q-Q-H-T-G-G-E-H-D-R-D-R-T-R-G-G-Q-H-T-T,
и б) вторую последовательность ДНК, кодирующую пептид, выбираемый из группы, включающей цитокин, фермент, антикоагулянт, нейропептид, гормон, антитело или предшественник молекулы адгезии.
V-Q-G-E-E-S-N-D-K.
| МАШИНА ДЛЯ ЗАВЕРТЫВАНИЯ КОНФЕТ | 1940 |
|
SU62353A1 |
| VAN ROOIJEN G.J.H., Plant molecular Biology, v.18, №6, 1992. | |||

