Эта заявка является частичным продолжением одновременно рассматриваемой патентной заявки США N 08/567375, зарегистрированной 4 декабря 1995 г., которая является частичным продолжением предварительной заявки N 60/004645. Она также является частичным продолжением одновременно рассматриваемой патентной заявки США N 08/475891, зарегистрированной 7 июня 1995 г., которая является частичным продолжением одновременно рассматриваемой патентной заявки N 08/373374, зарегистрированной 17 января 1995 г. Эти заявки включены сюда в виде ссылок.
Данное изобретение в основном относится к молекулярной биологии растений. В частности, оно относится к нуклеиновым кислотам и способам придания растениям устойчивости к заболеваниям.
Это изобретение было сделано при правительственной поддержке по гранту N GM47907, предоставленному Национальными Институтами Здоровья, и по гранту N 9300834, предоставленному Министерством сельского хозяйства США. Правительство имеет определенные права на это изобретение.
Локусы, придающие устойчивость к заболеваниям, были идентифицированы у многих видов растений. Генетический анализ взаимодействий возбудителя заболевания и растения продемонстрировал, что растения содержат локусы, которые придают устойчивость к конкретным расам возбудителей, содержащим комплементарный ген авирулентности. Молекулярная характеристика этих генов должна обеспечить средства для придания устойчивости к заболеваниям широкому ряду культурных растений.
Те гены устойчивости растений, которые были охарактеризованы на молекулярном уровне, попадают в четыре класса генов. Один ген, Hml у зерновых, кодирует редуктазу и эффективен против грибкового возбудителя Cochliobolus carbonum (Johal et al., Science 258: 985-987 (1992)). У томатов ген Pto придает устойчивость против Pseudomonas syringae, которая экспрессирует ген авирулентности avrPto (Martin et al., Science 262: 1432 (1993)). Предсказанный ген Pto кодирует серин-треонин-протеинкиназу. Ген томатов Cf-9 придает устойчивость к расам гриба Cladosporium fulvum, который несет ген авирулентности Avr9 (Jones et al., Science 266:789-793 (1994)). Ген томатов Cf-9 кодирует предположительно экстраклеточный белок LRR. И, наконец, ген RPS2 Arabidopsis thaliana придает устойчивость к Р. syringae, которая экспрессирует ген авирулентности avRpt2 (Bent et al., Science 265: 1856-1860 (1994)). RPs2 кодирует белок с мотивом LRR и мотивом Р-петли. Бактериальное гнилостное заболевание, вызываемое Xanthomonas spp. , поражает практически все культурные растения и приводит к обширным потерям урожая по всему миру.
Бактериальное гнилостное заболевание риса (Oryza sativa), вызываемое Xanthomonas oryzae pv. Oryzae (Xoo), является важным заболеванием этого культурного растения. Были идентифицированы расы Xoo, которые вызывают реакции устойчивости или чувствительности у культивируемых вариантов риса с различными генами устойчивости (Ха). Один источник устойчивости (Ха21) был идентифицирован у диких видов Oryza longistaminata (Khush et al., Proceedings of the International Workshop on Bacterial Blight of Rice. (International Rice Research Institute, 1989) и Ikeda et al., Jpn J. Breed. 40 (Suppl. 1): 280-281 (1990)). Xa21 является доминантным локусом устойчивости, который придает устойчивость ко всем известным штаммам Хоо и является единственным охарактеризованным геном Ха, который несет устойчивость к Хоо, раса 6. С помощью генетического и физического анализа локуса Ха21 был идентифицирован ряд тесно связанных маркеров на хромосоме 11 (Ronald et al., Mol. Gen. Genet. 236: 113-120 (1992)). Молекулярные механизмы, с помощью которых локус Ха21 придает устойчивость к этому возбудителю, однако, не идентифицированы.
Значительные усилия были направлены на клонирование генов растений, придающих устойчивость к ряду бактериальных, грибковых и вирусных заболеваний. Только один ген устойчивости к вредителям был клонирован у однодольных. Так как урожаем однодольных культурных растений питается большинство людей и животных в мире, идентификация генов устойчивости к заболеваниям у этих растений особенно важна. Данное изобретение направлено на эти и другие нужды.
Данное изобретение представляет конструкции из выделенной нуклеиновой кислоты, состоящие из полинуклеотидной последовательности RRK, которая гибридизуется с ПОСЛ. ИД. N 1 или ПОСЛ. ИД. N 3 в строгих условиях. Примерами полинуклеотидных последовательностей RRK являются последовательности Ха21, которые кодируют полипептид Ха21, который показан в ПОСЛ. ИД. N 4. Полинуклеотиды RRK кодируют белок, имеющий повторяющийся богатый лейцином мотив и/или домен цитоплазматической протеинкиназы. Конструкции нуклеиновой кислоты этого изобретения могут, кроме того, включать промотор, операбельно связанный с полинуклеотидной последовательностью RRK. Промотор может быть тканеспецифичным промотором или структурным промотором.
Изобретение также представляет конструкции нуклеиновой кислоты, состоящие из промоторной последовательности из RRK гена, связанной с гетерологичной полинуклеотидной последовательностью. Примеры гетерологичных полинуклеотидных последовательностей включают структурные гены, которые придают устойчивость к возбудителям болезней растениям.
Это изобретение, кроме того, представляет трансгенные растения, включающие рекомбинантный полигенный экспрессирующий кластер, состоящий из промотора из гена RRK, операбельно связанного с полинуклеотидной последовательностью, а также трансгенные растения, включающие рекомбинантный полигенный экспрессирующий кластер, состоящий из промотора растения, операбельно связанного с полинуклеотидной последовательностью RRK. Хотя в этом изобретении может быть использовано любое растение, может быть наиболее удобно использовать растения риса и томатов.
Это изобретение, кроме того, представляет способы повышения устойчивости к Xanthomonas у растений. Способы включают введение в растение рекомбинантного полигенного экспрессирующего кластера, включающего промотор растения, операбельно связанный с полинуклеотидной последовательностью RRK. Способы могут быть соответственно осуществлены с использованием растений риса и томатов.
Определения
Термин "растение" включает растения целиком, органы растений (например, листья, стебли, корни и тд.), семена и клетки растений и их потомство. Класс растений, который может использоваться при способах этого изобретения, в основном также широк, как и класс высших растений, поддающихся методам трансформации, включая как однодольные, так и двудольные растения.
"Гетерологичная последовательность" является такой последовательностью, которая происходит из другого вида или, если из того же самого вида, то она существенно модифицирована по сравнению с первоначальной формой. Например, промотор, операбельно связанный с гетерологичным структурным геном, происходит из другого вида, а не из того, из которого получен структурный ген, или, если они из одного и того же вида, один или оба существенно модифицированы по сравнению с исходной формой.
"Ген RRK" является представителем нового класса генов устойчивости к заболеваниям, которые кодируют полипептиды RRK, включающие экстраклеточный домен LRR, трансмембранный домен, домен цитоплазматической протеинкиназы (как показано, например, в RLK5, Pto и Fen (Martin et al., Plant Cell 6: 1543-1552 (1994)). Так, как он использован здесь, термин домен LRR является областью повторов единицы из примерно 24 остатков, которая показана на фиг.1 и обнаружена в Cf-9 и RLK5. Используя последовательности, раскрытые здесь, и стандартные методики гибридизации и/или амплификации нуклеиновых кислот, опытный специалист может идентифицировать представителей этого класса генов. Например, проба нуклеиновой кислоты из гена Ха21 выявляет полиморфизм, который различается у гена устойчивости к пирикуляриозу риса (Pyricularia oryzae) (Pi7) в 58 рекомбинантных инбредных линиях риса. Та же самая проба также выявила полиморфизм у близких изогенных линий, несущих гены устойчивости Ха5 и Ха10.
При некоторых предпочтительных осуществлениях представители этого класса генов устойчивости к заболеваниям могут быть идентифицированы по их способности амплифицироваться с помощью вырожденных праймеров ПЦР, которые соответствуют доменам киназы и LRR. Например, праймеры использовались для выделения гомологичных генов у томатов. Примерами праймеров для этой цели являются tcaagcaacaatttgtcaggnca a/g в a/c/t cc (для последовательности GOIP LRR домена) и taacagcacattgcttgatttnan g/a tcncg g/a tg (последовательность HCDIK домена киназы).
"Полинуклеотидная последовательность Ха21" является последовательностью или полинуклеотидной последовательностью полной длины гена Ха21, такого как ген Ха21 риса, которая, когда она присутствует в трансгенном растении, придает растению устойчивость к Xanthomonas spp. (например, к X. Oryzae). Примеры полинуклеотидов этого изобретения включают кодирующую область ПОСЛ. ИД. N 3. Полинуклеотид Ха21 имеет обычно длину по крайней мере от примерно 3100 нуклеотидов до примерно 6500 нуклеотидов, обычно от примерно 4000 до примерно 4500 нуклеотидов.
"Полипептид Ха21" является генным продуктом из Ха21 полинуклеотидной последовательности, который обладает активностью Ха21, т.е. способностью придавать устойчивость к Xanthomonas spp. Полипетиды Ха21, подобно другим полипептидам RRK, характеризуются наличием экстраклеточного домена, включающего область повторов, богатых лейцином (LRR) и/или домен цитоплазматической протеинкиназы. Примеры Ха21 полипептидов этого изобретения включают ПОСЛ. ИД. N 4.
Опытный специалист поймет, что при экспрессии трансгенов нет необходимости, чтобы вставленная полинуклеотидная последовательность была идентична, а она может быть "по существу идентичной" последовательности гена, от которого она была произведена. Как объяснено ниже, эти варианты специфично охватываются этим термином.
В случае, когда вставленная полинуклеотидная последовательность считывается и транслируется с получением функционального пептида RRK, опытный специалист поймет, что из-за вырожденности кодона ряд полинуклеотидных последовательностей будет кодировать тот же самый полипептид. Эти варианты специфично охватываются термином "полинуклеотидная последовательность RRK". Кроме того, термин специфично включает такие последовательности полной длины, по существу идентичные (определяемые, как описано ниже) с последовательностью гена RRK, и те, которые кодируют белки, сохраняющие функцию белка RRK. Таким образом, в случае генов RRK риса, раскрытых здесь, вышеуказанный термин включает варианты полинуклеотидных последовательностей, которые обладают существенной идентичностью с последовательностями, раскрытыми здесь, и с теми, которые кодируют белки, способные придавать устойчивость к Xanthomonas или другим болезням растений и вредителям трансгенным растениям, содержащим эту последовательность.
Два полинуклеотида или полипептида называются "идентичными", если последовательность нуклеотидов или аминокислотных остатков соответственно в двух последовательностях является одинаковой при сравнении первичной структуры на максимальное соответствие, как описано ниже. Термин "комплементарный" используется здесь для обозначения того, что комплементарная последовательность идентична всей сравниваемой полинуклеотидной последовательности или ее части.
Сравнение последовательности двух (или более) полинуклеотидов или полипептидов обычно выполняется путем сравнения последовательностей двух последовательностей по сегменту или "окну сравнения" для идентификации и сравнения локальных участков сходства последовательностей. Сегмент, использованный для целей сравнения, может состоять из по крайней мере примерно 20 смежных положений, обычно из от примерно 50 до примерно 200, более обычно от примерно 100 до примерно 150, в которых можно провести сравнение последовательности с эталонной последовательностью из того же числа смежных положений после того, как оптимально проведен анализ первичной структуры двух последовательностей.
Оптимальный анализ первичной структуры последовательностей для сравнения может быть проведен с помощью алгоритма локальной гомологии Smith and Waterman, Adv. Appl. Math. 2: 482 (1981), с помощью алгоритма гомологичного выравнивания Needleman and Wunsch, J. Mol. Biol, 48:443 (1970), путем исследования на подобие методом Pearson and Lipman, Proc. Natl. Acad. Sci. (U. S. A) 85: 2444 (1988), с помощью компьютерного воплощения этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group (GGG), 575 Science Dr., Madison, WI) или с помощью просмотра.
"Процент идентичности последовательности" определяется путем сравнения двух оптимально выравненных последовательностей по окну сравнения, причем часть полинуклеотидной последовательности в окне сравнения может включать добавки или делении (т. е. пропуски) по сравнению с эталонной последовательностью (которая не содержит добавок или делений), для оптимального выравнивания двух последовательностей. Процент рассчитывается путем определения числа положений, в которых идентичные основание нуклеиновой кислоты или аминокислотный остаток присутствует в обеих последовательностях, с получением числа соответствующих положений, делением числа соответствующих положений на общее число положений в окне сравнения и умножением результата на 100 с получением процента идентичности последовательностей.
Термин "существенная идентичность" полинуклеотидных последовательностей означает, что полинуклеотид включает последовательность, которая имеет по крайней мере 60% идентичность последовательности, предпочтительно по крайней мере 80%, более предпочтительно по крайней мере 90% и наиболее предпочтительно по крайней мере 95% по сравнению с эталонной последовательностью при использовании программ, описанных выше (предпочтительно BESTFIT), с использованием стандартных параметров. Опытный специалист поймет, что эти значения могут быть соответственно установлены для определения соответствующей идентичности белков, кодируемых двумя нуклеотидными последовательностями, принимая во внимание вырожденность кодона, сходство аминокислот, положения рамки считывания и тому подобного. Существенная идентичность аминокислотных последовательностей для этих целей обычно означает идентичность последовательностей, равную по крайней мере 40%, предпочтительно по крайней мере 60%, более предпочтительно по крайней мере 90% и наиболее предпочтительно по крайней мере 95%. Полипептиды, которые являются "существенно сходными" долевыми последовательностями, которые отмечены выше, за исключением тех положений остатков, которые не идентичны, могут отличаться заменами консервативных аминокислот. Замещения консервативных аминокислот относятся к взаимозаменяемости остатков, имеющих сходные боковые цепи. Например, группу аминокислот, имеющих алифатические боковые цепи, составляют глицин, аланин, валин, лейцин и изолейцин; группу аминокислот, имеющих алифатическо-гидроксильные боковые цепи, составляют серин и треонин; группу аминокислот, имеющих ароматические боковые цепи, составляют фенилаланин, тирозин и триптофан; группу аминокислот, имеющих серосодержащие боковые цепи, составляют цистеин и метионин. Предпочтительными группами замещения консервативных аминокислот являются валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин и аспарагин-глютамин.
Существует другой показатель того, что нуклеотидные последовательности являются по существу идентичными, если две молекулы гибридизуются друг с другом в соответствующих условиях. Соответствующие условия могут быть высокой или низкой жесткости и будут различными в разных обстоятельствах. В основном строгие условия выбираются такими, чтобы температура была примерно на 5-20oС ниже, чем температура плавления (Тп) для конкретной последовательности при определенных ионной силе и рН. Тп является температурой (при определенной ионной силе и рН), при которой 50% целевой последовательности гибридизуется с совершенно подходящей пробой. Обычно строгие условия отмывания являются такими, при которых концентрация соли равна примерно 0,02 молярной при рН 7 и температура равна по крайней мере 60oС. Однако нуклеиновые кислоты, которые не гибридизуются друг с другом в строгих условиях, являются все еще по существу идентичными, если полипептиды, которые они кодируют, являются по существу идентичными. Это может происходить, например, когда копия нуклеиновой кислоты создается с использованием максимальной вырожденности кодона, допускаемой генетическим кодом. Для гибридизации по Саузерну высоко строгие условия отмывки будут включать по крайней мере одну отмывку 0,1 Х SSC при 65oС.
Нуклеиновые кислоты этого изобретения могут быть идентифицированы из кДНК или геномной библиотеки, полученной по стандартным методикам, а нуклеиновые кислоты, раскрытые здесь (например, ПОСЛ. ИД. N 1 или 3), использовали в качестве пробы. Условия гибридизации с низкими требованиями обычно включают по крайней мере одно отмывание с использованием 2Х SSC при 65oС. Отмывания предпочтительно сопровождаются последующим промыванием с использованием IX SSC при 65oС.
Так, как он использован здесь, гомолог конкретного гена RRK (например, ген риса Ха21, раскрытый здесь) является вторым геном (или в одном и том же виде, или в разных видах), который кодирует белок, имеющий аминокислотную последовательность, обладающую по крайней мере 25% идентичностью или 45% подобием (определенными, как описано выше) с полипептидной последовательностью в первом генном продукте. Считают, что в основном гомологи имеют общее эволюционное прошлое.
На фиг.1 показано сравнение богатых лейцином повторов белков.
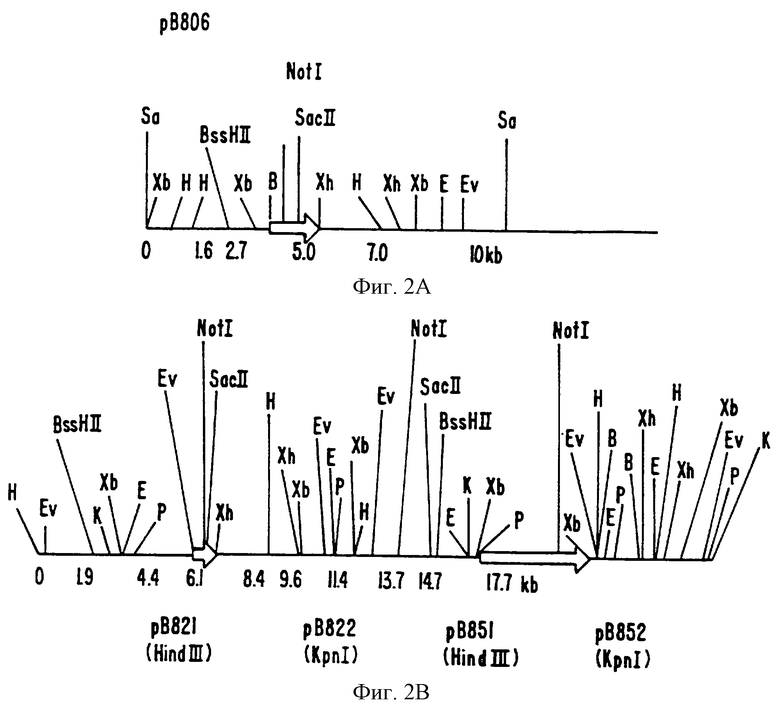
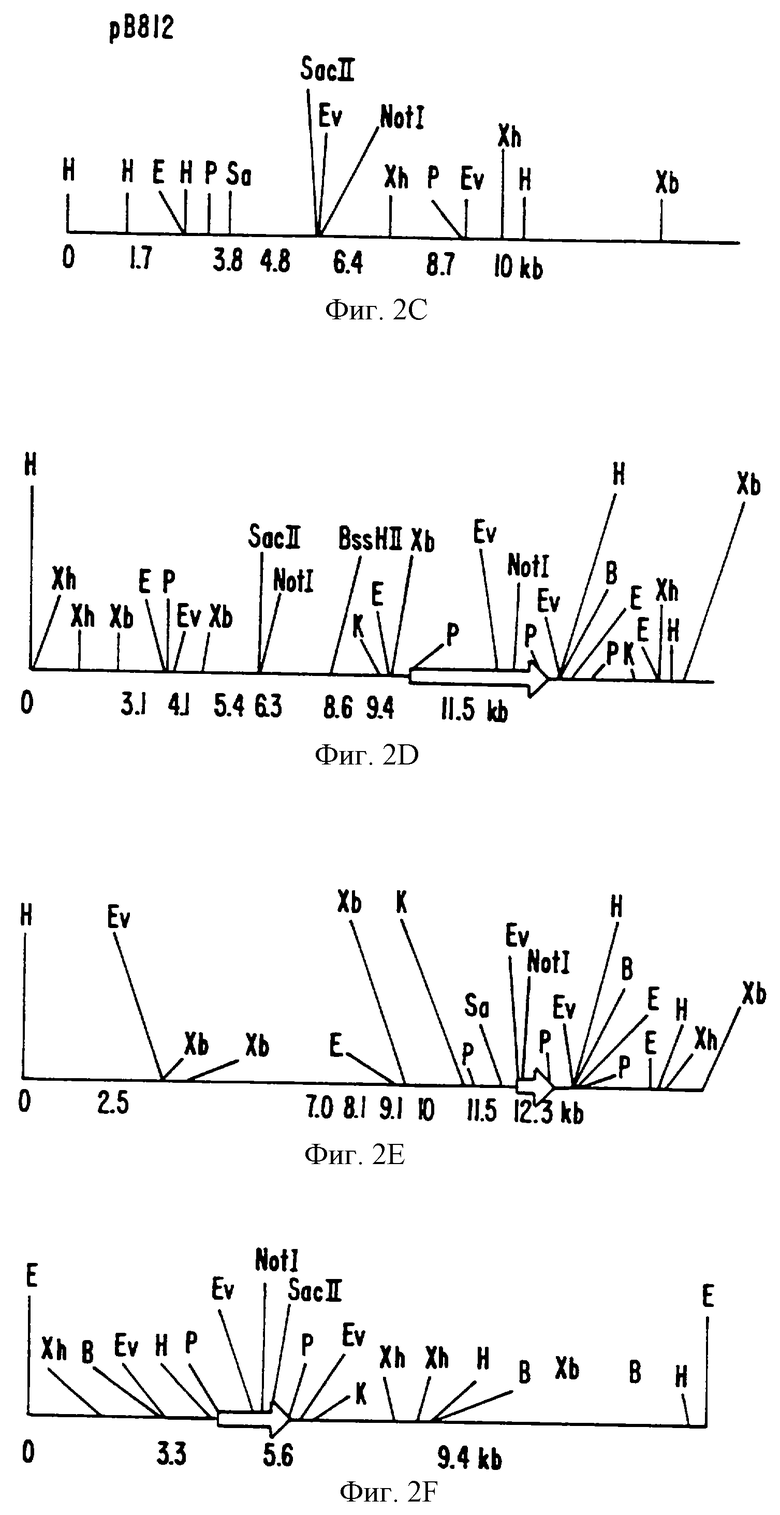
На фиг. 2A-F показаны частичные рестрикционные карты БАК и космидные клоны, содержащие области, которые гибридизуются с Ха21-специфичными пробами.
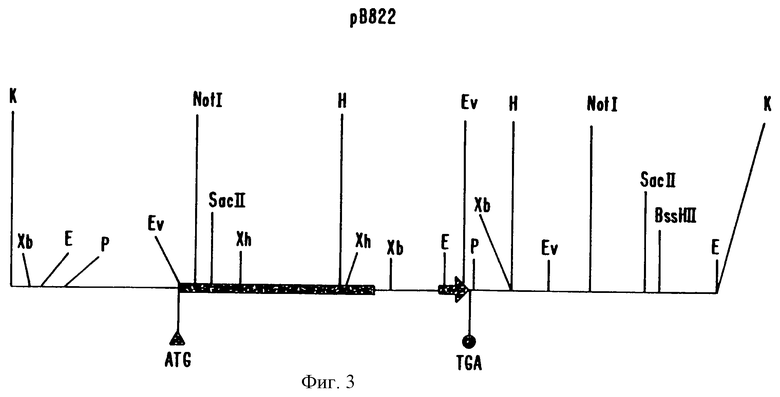
На фиг.3 показана рестрикционная карта рВ822, наиболее активной копии.
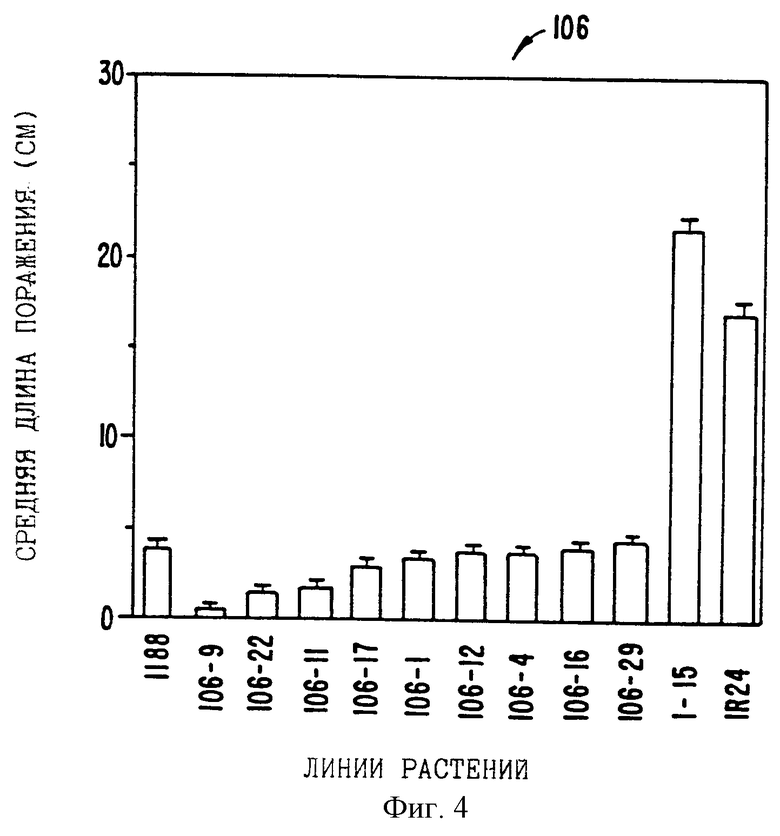
На фиг.4 показаны результаты исследований по определению устойчивости к Xanthomonas у трансгенных растений, содержащих ген Ха21 из клона рВ822.
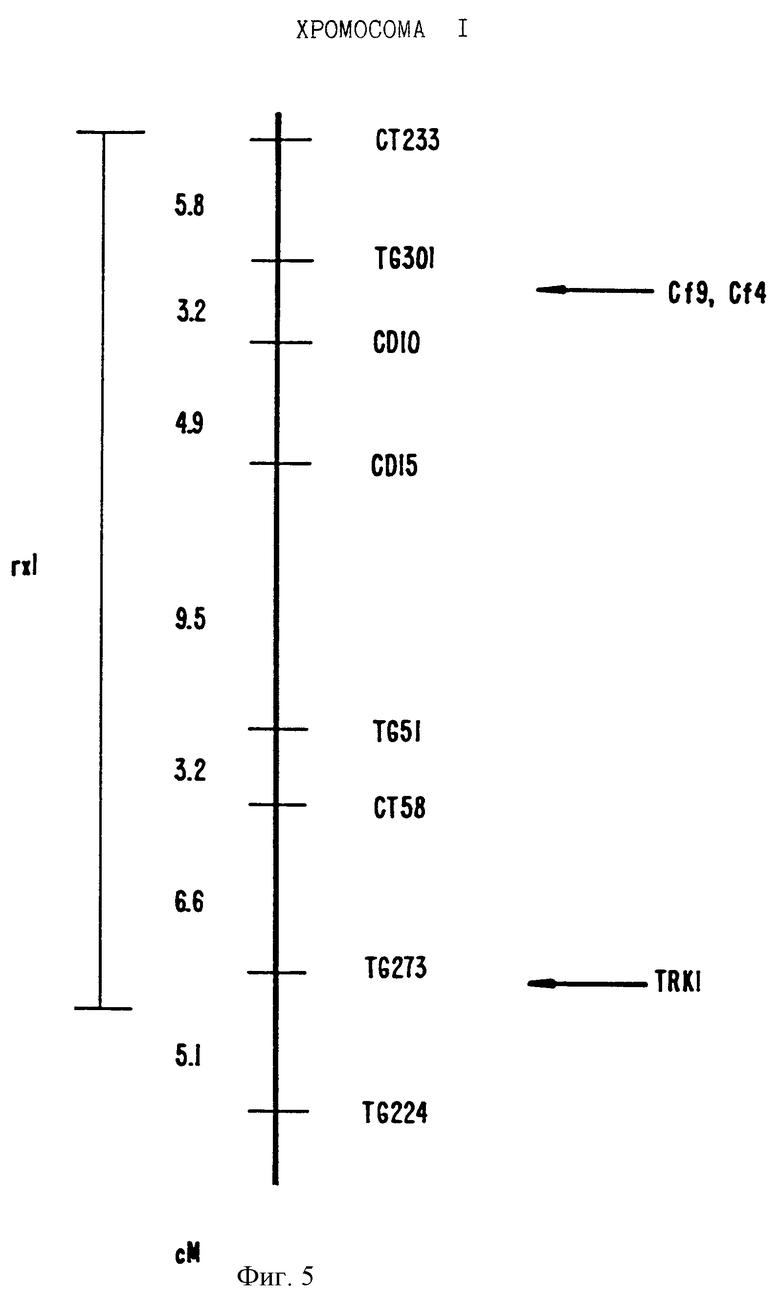
На фиг.5 показано положение на карте TRK1.
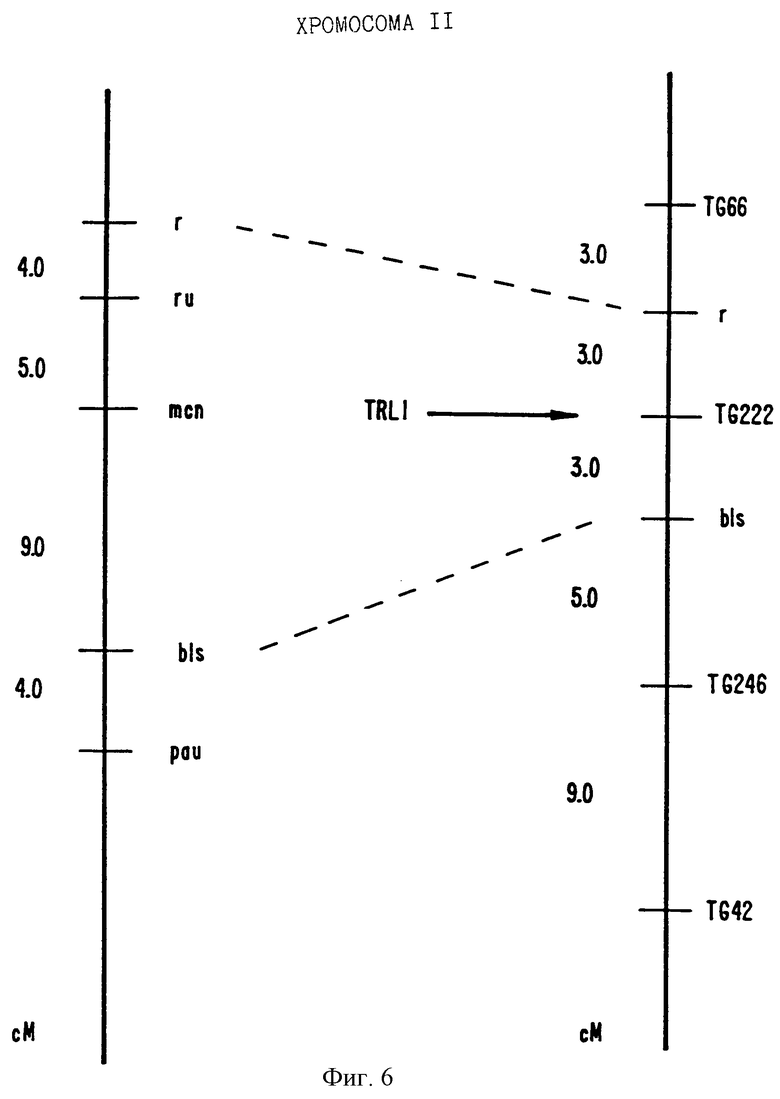
На фиг.6 показано положение на карте TRL1.
Это изобретение относится к генам растений RRK, таким как гены риса Ха21. Последовательности нуклеиновых кислот из генов RRK, в частности генов Ха21, могут использоваться для придания устойчивости к Xanthomonas и другим возбудителям болезней растений. Изобретение применимо для придания устойчивости высшим растениям, чувствительным к инфекции, вызываемой возбудителем. Изобретение, таким образом, применимо в отношении широкого круга видов растений, включая виды из родов Juglans, Fragaria, Lotus, Medicago, Onobrychis, Trifolium, Trigonella, Vigna, Citrus, Linum, Geranium, Manihot, Daucus, Arabidopsis, Brassica, Raphanus, Sinapis, Atropa, Capsicum, Datura, Hyoscyamus, Lycopersicon, Nicotiana, Solanum, Petunia, Digitalis, Majorana, Ciahorium, Helianthus, Lactuca, Bromus, Asparagus, Antirrhinum, Heterocallis, Nemesis, Pelargonium, Panieum, Pennisetum, Ranunculus, Senecio, Salpiglossis, Cucumis, Browaalia, Glycine, Pisum, Phaseolus, Lolium, Zea, Avena, Hordeum, Secale, Triticum и Sorghum.
Нижеследующий раздел примеров, в котором описано выделение и определение характеристик генов риса Ха21, является примером общего подхода к выделению генов Ха21 и других генов RRK. Выделенные гены могут затем использоваться для конструирования рекомбинантных векторов для переноса экспрессии гена RRK в трансгенные растения.
В основном номенклатура и лабораторные методики для генной инженерии, описанные ниже, хорошо известны и обычно используются в этой области технологии. Для клонирования используются стандартные методики выделения ДНК и РНК, амплификации и очистки. Обычно ферментативные реакции с участием ДНК-лигазы, ДНК-полимеразы, рестрикционных эндонуклеаз и тому подобного выполняются по описаниям изготовителей. Эти методики и различные другие в основном выполняются по Sambrook et al., Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York (1989).
Выделение Ха21 и родственных генов RRK может выполняться с помощью ряда методик. Например, для идентификации желаемого гена в кДНК или геномной библиотеке ДНК могут использоваться олигонуклеотидные зонды на основе последовательностей, раскрытых здесь. Для создания геномных библиотек получают большие сегменты геномной ДНК путем случайной фрагментации, например, с использованием рестрикционных эндонуклеаз, и их лигируют с помощью векторной ДНК с образованием конкатемеров, которые могут упаковываться в соответствующий вектор. Для получения библиотеки кДНК выделяют мРНК из желаемого органа, такого как лист, и библиотеку кДНК, которая содержит транскрипт гена RRK, получают с мРНК. Или же кДНК может быть получена с мРНК, извлеченной из других тканей, в которых экспрессируются гены RRK или гомологи.
Затем кДНК и геномную библиотеку можно отсортировать с использованием пробы на основе последовательности клонированного гена RRK, такого как гены Ха21 риса, описываемые здесь. Пробы могут использоваться для гибридизации с последовательностями геномной ДНК или кДНК с целью выделения гомологичных генов в одних и тех же или различных видах растений.
Или же иначе, интересующие нуклеиновые кислоты могут быть амплифицированы из образцов нуклеиновых кислот с применением методик амплификации, например, по методу полимеразной цепной реакции (ПЦР) для амплификации последовательностей RRK и родственных генов непосредственно из геномной ДНК, из кДНК, из геномных библиотек или библиотек кДНК. ПЦР и другие методы амплификации in vitro также могут применяться, например, для клонирования последовательностей нуклеиновых кислот, кодирующих белки, которые нужно экспрессировать, чтобы получить нуклеиновые кислоты для использования в качестве проб для определения присутствия желаемых мРНК в образцах, для секвенирования нуклеиновых кислот или для других целей.
Получены соответствующие праймеры и пробы для идентификации последовательностей RRK из тканей растений в результате сравнения последовательностей, представленных здесь. Для общего представления о ПЦР см. PCR Protocols: A Guide to Methods and Applications (Innis M., Gelfand D., Sninsky J., and White Т., eds.), Academic Press, San Diego (1990), представлено здесь в виде ссылки.
Полинуклеотиды могут также синтезироваться по хорошо известным методикам, как описано в технической литературе. Смотрите, например, Carruthers et al.. Cold Spring Harbor Symp. Quant. Biol. 47:411-418 (1982), и Adams et al. , J. Am. Chem. Soc. 105:661 (1983). Затем могут быть получены фрагменты двунитевой ДНК или путем синтеза комплементарной цепи и совместной ренатурации цепей в соответствующих условиях, или путем добавления комплементарной цепи с использованием ДНК-полимеразы с соответствующей праймерной последовательностью.
Выделенные последовательности, полученные, как описано здесь, можно затем использовать для обеспечения экспрессии гена RRK и, следовательно, устойчивости к Xanthomonas у желаемых растений. Специалист поймет, что нет необходимости, чтобы нуклеиновая кислота, кодирующая функциональный белок RRK (например, ПОСЛ. ИД. N 2 и 4), имела последовательность, идентичную приведенной в качестве примера последовательности гена, описанной здесь. Кроме того, полипептиды, кодируемые генами RRK, подобно другим белкам, имеют различные домены, которые выполняют разные функции. Таким образом, нет необходимости в том, чтобы последовательности гена RRK имели полную длину, при условии, что экспрессируется желаемый функциональный домен белка. Как объяснено в деталях ниже, белки этого изобретения включают экстраклеточный богатый лейцином повторный домен, а также внутриклеточный киназный домен. Модифицированные белковые цепи можно также легко создать, используя методы генной инженерии, хорошо известные специалистам. Например, цепи могут быть изменены по сравнению с встречающимися в природе последовательностями на уровне первичной структуры путем замен аминокислот, добавок, делений и тому подобного. Модификации могут также включать обменные домены из белков этого изобретения с родственными доменами из других генов устойчивости к вредителям. Например, экстраклеточный домен (включающий богатую лейцином область повторов) белков этого изобретения может быть заменен таким доменом из гена томатов Cf-9 и таким образом придавать устойчивость к грибковым возбудителям болезней риса. Эти модификации могут использоваться в ряде комбинаций для получения конечной модифицированной белковой цепи.
Чтобы использовать выделенные последовательности RRK в вышеприведенных методиках, получены векторы рекомбинантной ДНК, пригодные для трансформации растительных клеток. Методики для трансформации широкого ряда видов высших растений хорошо известны и описаны в технической и научной литературе. Смотрите, например, Weising et al., Ann. Rev. Genet. 22:421-477 (1988).
Последовательность ДНК, кодирующая желательный полипептид RRK, например кДНК или геномная последовательность, кодирующая белок полной длины, будет использоваться для построения рекомбинантного полигенного экспрессирующего кластера, который может быть введен в желаемое растение. Полигенный экспрессирующий кластер будет обычно включать полинуклеотид RRK, оперативно связанный с регуляторными последовательностями инициации транскрипции и трансляции, которые будут управлять транскрипцией последовательности из гена RRK в намеченных тканях трансформированного растения.
Например, может использоваться фрагмент промотора растения, который будет управлять экспрессией RRK во всех тканях регенерированного растения. Такие промоторы называются здесь "конститутивными" промоторами и они активны в большинстве случаев в условиях окружающей среды и при состояниях развития и дифференциации клеток. Примеры конститутивных промоторов включают область инициации транскрипции 35S вируса мозаики цветной капусты (CaMV), 1'- или 2'-промотор, происходящий из Т-ДНК Agrobacterium tumifaciens и другие области инициации транскрипции из различных генов растений, известных специалистам.
Или же промотор растения может управлять экспрессией гена RRK в конкретной ткани или, в другом случае, при более строго определенных условиях окружающей среды или регуляции развития. Такие промоторы называются здесь "индуцируемыми" промоторами. Примерами условий окружащей среды, которые могут воздействовать на транскрипцию с помощью индуцируемых промоторов, включают поражение возбудителями заболеваний, влияние анаэробных условий или наличие освещения.
Примеры промоторов, находящихся под управлением развития, включают промоторы, которые инициируют транскрипцию только в определенных тканях, таких как листья, корни, фрукты, семена или цветы. Работа промотора может также изменяться в зависимости от его локализации в геноме. Таким образом, индуцируемый промотор может стать полностью или частично конститутивным в определенных положениях.
Эндогенные промоторы из генов RRK этого изобретения могут использоваться для управления экспрессией генов. Эти промоторы также могут использоваться для управления экспрессией гетерологичных структурных генов. Таким образом, эти промоторы могут использоваться в рекомбинантных полигенных экспрессионных кластерах для направления экспрессии генов, придающих устойчивость к какому-либо числу возбудителей заболеваний, включая грибы, бактерии и тому подобное.
Чтобы идентифицировать промоторы 5'-части клонов, описанных здесь, анализировали на последовательности, характерные для промоторных последовательностей. Например, элементы промоторной последовательности включают консенсусную последовательность ТАТА-бокса (ТАТААТ), которая находится обычно на 20 - 30 пар оснований в обратном направлении от стартового сайта транскрипции. У растений, кроме того, в обратном направлении от ТАТА-бокса в положениях с -80 по -100 существует обычно промоторный элемент с серией аденинов, окружающих тринуклеотид G (или Т) NG. J. Messing et al., in Genetic Engineering in Plants, pp. 221-227 (Kosage, Meredith and Hollaender, eds. 1983).
Если желательна экспрессия соответствующего полипептида, должна быть включена область полиаденилирования на 3'-конце области, кодирующей RKK. Область полиаденилирования может быть получена из природного гена, из ряда других генов растения или из Т-ДНК.
Вектор, включающий эти последовательности из гена RRK, будет обычно включать маркерный ген, который придает селективный фенотип растительным клеткам. Например, маркер может кодировать биоцидную устойчивость, в частности устойчивость к антибиотикам, такую как устойчивость к канамицину, G418, блеомицину, гигромицину, или гербицидную устойчивость, такую как устойчивость к хлорослюфорону или Баста.
Такие конструкции ДНК можно вводить в геном желательного растения-хозяина с помощью ряда общепринятых методик. Например, конструкцию ДНК можно непосредственно ввести в геномную ДНК клетки растения, используя такие методики, как электропорация, ПЭГ-порация, бомбардировка частицами и микроинъекция протопластов растительной клетки или эмбриогенного каллюса, или конструкции ДНК можно вводить непосредственно в ткань растения, используя баллистические методы, такие как бомбардировка частицами ДНК. Или же конструкции ДНК могут быть соединены с подходящими ограничивающими областями Т-ДНК и введены в обычный вектор-хозяин Agrobacterium tumefaciens. Функции вирулентности хозяина Agrobacterium tumefaciens будут направлять вставку (генно-инженерной) конструкции и примыкающего маркера в ДНК растительной клетки, когда клетка инфицирована бактерией.
Методики трансформации известны в этой области технологии и хорошо описаны в научной и патентной литературе. Введение генно-инженерных конструкций ДНК с использованием преципитации полиэтиленгликолем описано у Paszkowski et al., Embo J. 3:2717-2722 (1984). Методики электропорации описаны у Fromm et al., Natl. Acad. Sci. USA 82:5824 (1985). Методики баллистической трансформации описаны у Klein et al., Nature 327:70-73 (1987). При использовании ряда подходов можно трансформировать виды зерновых, такие как рожь (de la Репа et al., Nature 325:274-276 (1987)), пшеница (Rhode et al., Science 240:204-207 (1988)) и рис (Shimamoto et al., Nature 338:274-276 (1989), путем электропорации; Li et al., Plant Cell Rep. 12:250-255 (1993) путем баллистических методов).
Методики трансформации, опосредуемые Agrobacterium tumefaciens, хорошо описаны в научной литературе. Смотрите, например, Horsch et al., Science 233: 496-498 (1984), Fraley et al. , Proc. Natl. Acad. Sci. USA 80:4803 (1983). Хотя Agrobacterium сначала применены у двудольных, некоторые однодольные могут трансформироваться с помощью Agrobacterium. Например, трансформация риса с помощью Agrobacterium описана Hiei et al., Plant J. 6:271-282 (1994).
Трансформированные растительные клетки, которые получены с помощью вышеназванных методик трансформации, можно культивировать с регенерацией целого растения, которое обладает трансформированным генотипом, и таким образом, желаемым фенотипом под контролем RRK. Такие методики регенерации основаны на процедурах с применением некоторых фитогормонов в среде для роста тканевой культуры, обычно с учетом маркера биоцидных и/или гербицидных свойств, который был введен вместе с нуклеотидными последовательностями RRK. Регенерация растения из культивируемых протопластов описана у Evans et al., Protoplasts Isolation and Culture, Handbook of Plant Cell Culture, pp. 124-176, MacMillian Publishing Company, New York, 1983; и Binding, Regeneration of Plants, Plant Protoplasts, pp. 21-73, CRC Press, Boca Raton, 1985. Регенерация может быть получена из растительного каллюса, эксплантатов, органов или их частей. Методики такой регенерации описаны в основном у Klee et al., Ann. Rev. of Plant Phys. 38:467-486 (1987).
Методы данного изобретения особенно пригодны для включения полинуклеотидов RRK в трансформированные растения таким образом и при таких условиях, которые не встречаются в природе. В частности полипептиды RRK могут экспрессироваться в такое время и в таких количествах, которые не характерны для растений в естественном виде.
Специалист поймет, что после того как полигенный экспрессирующий кластер устойчиво включен в трансгенные растения и подтверждено, что он является операбельным, он может быть введен в другие растения путем полового скрещивания. Может применяться любая из ряда стандартных методик селекции в зависимости от видов, которые должны быть скрещены.
Эффект модификации экспрессии гена RRK может быть оценен количественно путем определения повышения или снижения уровней мРНК при использовании, например, нозерн-блоттинга. Кроме того, фенотипические эффекты экспрессии гена можно обнаружить путем количественного определения протяженности поражения, как в случае растений. Соответствующие исследования для определения устойчивости описаны ниже.
Следующие примеры предлагаются в виде иллюстрации, но не ограничения.
Пример 1
Гены растений могут быть также выделены при использовании методов клонирования на основе картирования. Этот метод состоит в идентификации маркеров ДНК, которые тесно связаны с интересующим(и) геном или генами. Одно из требований для успеха клонирования на основе картирования и физического анализа больших хромосомных участков состоит в доступности библиотек, содержащих большие вставки геномной ДНК. Недавно Shizuya H., et al., Рrос. Natl. Acad. Sci. 89, 8794-8797 (1992), описали бактериальную искусственную хромосомную (ВАС) систему для клонирования больших фрагментов ДНК из человеческого генома. В этой системе использован вектор на основе F-фактора, и она способна к удерживанию фрагментов человеческой геномной ДНК размером >300 т.п.н. ДНК может быть клонирована с высокой эффективностью, ей легко манипулировать и она устойчиво сохраняется в Е. coli. Последующее является описанием применения этой методики для выделения генов этого изобретения.
Выделение ВАС и космидных клонов, несущих последовательности, родственные Ха21
ВАС клоны
А. МАТЕРИАЛЫ И МЕТОДЫ
Получение ДНК риса с высоким молекулярным весом
Линия риса Международного научно-исследовательского института риса (IRRI) IR-BB21 была использована в качестве растительного материала. Растения выращивали в теплице в течение 3-5 недель. Листовую ткань собирали и перед растиранием промывали дистиллированной водой. ДНК с высоким молекулярным весом экстрагировали из ткани риса по существу так, как описано Hatano S. , et al., Plant Sciences, 83, 55-64 (1992) и Zhang H.B., et.al., Plant J. 7: 175-184 (1994), со следующими модификациями: примерно 20 грамм листовой ткани растирали в порошок, используя холодную ступку и пестик в жидком азоте. Порошок суспендировали путем взбалтывания в 200 холодного буфера для экстрагирования ядер (ЭЯ) (1 мМ спермидина, 1 мМ спермина, 10 мМ Na2 ЭДТУ, 10 мМ Trizma, основания, 80 мМ КС1, 0,5% тритон-Х 100 и 0,4 М сахарозы, рН 9,4). Смесь фильтровали через два слоя серпянки в бутыль GSA и центрифугировали при 1200 g при 4oС в течение 20 мин. Супернатант сливали, а осадок из ядер (бледно-зеленый) снова суспендировали в 50 мл холодного буфера ЭЯ. Повторно суспендированный осадок затем отфильтровывали через 80-микронное сито в 50-мл пробирку для удаления остатков зеленой ткани и затем центрифугировали при 1000 g в течение 10 минут. Осадок повторно суспендировали и центрифугировали, как описано выше, но без пропускания через 80-микронное сито. Ядерный остаток (примерно 5•108 ядер/мл) снова суспендировали в 2,5 мл СЦЭ буфера (1 М сорбит, 0,1 М цитрат натрия, 60 мМ ЭДТУ, рН 7,0) и вводили в 2,5 мл 1% агарозы с низкой температурой плавления (НТП) (Ultrapure). 80-мл образцы инкубировали в 25 мл раствора ЭСП (0,5 М ЭДТУ, рН 9,3, 1% лаурилсаркозина натрия, 5 мг/мл протеиназы К, Boehringer Mannheim) при 50oС в течение двух дней с одной заменой буфера. Каждый образец содержал примерно 5 мкг ДНК.
Частичное расщепление ДНК с высоким молекулярным весом и фракционирование по размеру с помощью ГЭПЭП (гель-электрофореза в пульсирующем электрическом поле)
Агарозные образцы диализировали дважды в ТЭ (10 мМ Трис-НСl и 1 мМ ЭДТУ, рН 8,0) плюс 1 мм ФМСФ (фенилметилсульфонилфторид) при 50oС в течение 1 часа и затем уравновешивали буфером HindIII (50 мМ NaCl, 10 мМ Трис-HCl, 10 мМ MgCl2 и 1 мМ дитиотреитол, рН 7,9) дважды при комнатной температуре в течение одного часа. Образцы расплавляли при 65oС в течение 15 минут и выдерживали при 37oС в течение 5 минут перед частичным расщеплением. К раствору ДНК добавляли 5-7 единиц HindIII (NEB, USA) на образец и инкубировали при 37oС в течение 30 мин. Реакцию останавливали путем добавления 1/10 объема 0,5 М ЭДТУ, рН 8,0. Частично расщепленную ДНК сразу же помещали на 0,8% агарозный гель НТП с помощью пипетки с обрезанным концом до внутреннего диаметра 2 мм и разделяли с помощью ГЭПЭП (система CHEF DR II, BioRad, USA). Для составления библиотеки использовали два различных метода ГЭПЭП. Сначала гель подвергали электрофорезу при 150 V, применяя 8-секундное первоначальное и 8-секундное конечное время включения в течение 16 часов при 14oС. Нераспавшуюся ДНК (≥200 т. п.н.) фокусировали в тонкую полосу. Во-вторых гель подвергали электрофорезу при 150 V, линейном увеличении времени включения от 60 до 90 секунд. В течение 16 часов при 14oС. При обоих методах гель, содержащий частично расщепленную ДНК, вырезали и выдерживали в ТЭ, пока маркерные полосы геля окрашивались этидийбромидом. Из геля вырезали агарозный слой, содержащий фрагменты более 200 т.п.н. (первый метод ГЭПЭП), или агарозный слой, содержащий фрагменты в 250-350 т.п.н. (второй метод). Агарозный слой уравновешивали в ТЭ в течение 2 часов при 4oС, помещали в 1,5 мл пробирку, расплавляли при 65oС в течение 10 минут, расщепляли желатиназой (Epicentre, USA) (одна единица фермента на 100 мг агарозы) и инкубировали при 45oС в течение одного часа. Раствор ДНК непосредственно использовали для реакции лигирования.
Выделение и получение вектора и реакция лигирования
Вектор pBeloBAC был предоставлен докторами Н. Shizuya и М. Simon (California Institute of Technology, USA). Этот вектор содержит ген lacz, встроенный в вектор pBAC108L Shizuya et al. (1992). Одиночную колонию засевали в 5 мл среды LB, содержащей 12,5 мкг/мл хлорамфеникола и подращивали при 37oС в течение 4-5 часов перед добавлением к 6 литрам среды LB. Посев подращивали в течение примерно 16 часов при 37oС до ОП600нм 1,3-1,5. Плазмиду выделяли, используя набор для макси-выделения плазмид Qiagen (Qiagen, USA). Векторную ДНК дополнительно очищали путем равновесного центрифугирования в хлориде цезия/этидийбромиде при 45000 об/мин в течение 60 часов. Скорость вращения ротора снижали до 35000 об/мин в течение 1 часа, чтобы дать возможность градиенту релаксироваться, используя фиксированный ангерный ротор 70,1 (Beckman, USA). Плазмиду расщепляли HindIII до завершения и исследовали с помощью гель-электрофореза. Концы вектора дефосфорилировали с помощью НК фосфатазы (Epicenter, USA) при 30oС в течение 1 часа, используя 1 единицу фермента на 1 мкг векторной ДНК. Фосфатазу НК инактивировали путем нагревания при 65oС в течение 30 мин. Лигирование проводили в 100 мкл объеме, в котором примерно 40 нг селектированной по размеру ДНК риса (примерно 85 мкл) лигировали с 10 нг, расщепленного HindIII вектора (1 мкл, молярное отношение равно примерно 10 к 1 при избытке вектора) с помощью 400 единиц ДНК-лигазы Т4 (NEB, USA) при 16oС в течение ночи. Перед трансформацией смесь лигирования подвергали диализу в ТЭ в фильтровальной пробирке ULTRAFREE-MC (Millipore, USA) при 4oС в течение ночи.
ВАС трансформация
Трансформацию клеток компетентной Е. coli DH10B (GIBCO-BRL, USA) проводили путем электропорации, применяя клеточный поратор (GIBCO-BRL, USA) при следующих параметрах: напряжение 400 Вольт; скорость заряда: быстрая; устойчивость усилителя к напряжению: 4000; емкость 330 μ; импеданс: низкий. Тринадцать мкл компетентных клеток смешивали с 0,5-1,0 мкл раствора лигирования для каждой электропорации. После электропорации клетки переносили в 1 мл раствора SOC (2% бактотриптона, 0,5% бакто-дрожжевого экстракта, 10 мМ NaCl, 2,5 мМ КСl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкозы, рН 7,0) и инкубировали при 37oС при легком перемешивании (90-95 об/мин) в течение 45 мин. Клетки высевали на чашки со средой LB, содержащей хлорамфеникол (12,5 мкг/мл), X-gal (40 мкг/мл) и ИПТГ (изопропилтио-β-D-галактозид) (0,072 мкг/мл). Чашки инкубировали при 37oС в течение 24 часов. Белые колонии, содержащие вставки ДНК риса, снимали на новую чашку с LB для повторного отбора по цвету. Клоны ВАС переносили на 384-ячеечные микротитровальные платы (Genetix, UK), содержащие 60 мкл буфера LB для замораживания (36 мМ К2НРO4, 13,2 мМ КН2РO4, 1,7 мМ цитрата, 0,4 мМ MgSO4, 6,8 мМ (NH4)2SO4, 4,4% об. глицерина, 12,5 мкг/мл хлорамфеникола, LB) и инкубировали при 37oС в течение 24 часов. Так как более 95% колоний были все еще белыми при втором отборе, при последующих экспериментах применяли только один отбор и белые колонии снимали непосредственно на 384-ячеечные платы. Библиотеку реплицировали в двух повторностях и хранили в двух разных морозильниках при -80oС.
Препарат на фильтре
Клоны ВАС с каждой 384-ячеечной титровальной платы реплицировали на фильтр Hybond N+ (Amersham, USA). Фильтр помещали в пластиковый контейнер, содержащий агар LB с 12,5 мкг/мл хлорамфеникола, и контейнер выдерживали при 37oС в течение ночи до тех пор, пока колонии не достигали размера 2-3 мм в диаметре. Обработка фильтров была описана Nizetic D., et al., Nucl. Acids Res. 19, 182 (1990); Hoheeisel J.D., et al., Cell, 73, 109-120 (1993). Условия гибридизации и отмывания были такими же, как и описанные у Hoheisel, et al. (1993). Пробы метили, используя случайное праймерное удлинение. Feinberg A.P. and Vogelstein В., Anal. Biochem 132, 6-13 (1983); Addendum 137, 266-267 (1984).
Б.РЕЗУЛЬТАТЫ
Библиотека ВАС, описанная выше, состоит из 11000 клонов. Библиотека была составлена с использованием двух различных подходов. Первая половина библиотеки, имеющая 7269 клонов, была создана с помощью одной селекции по размеру с использованием метода зоны сгущения, который описан у Ramsay M. and Wicking С. , Protocols in Human Molecular Genetics, 197-221 (1991). Вторая половина библиотеки, имеющая 3731 клон, была создана с использованием двойного отбора по размеру частично расщепленной ДНК. Двойной отбор по размеру, однако, не давал возможности увеличить средний размер вставочной ДНК. Очевидно, небольшие молекулы ДНК все еще присутствовали в растворе селекционированной по размеру ДНК (выделенной ДНК только 250-350 т.п.н.). Последующие эксперименты продемонстрировали, что двойной отбор ДНК размером 350-500 т. п. н. для лигирования давал большее увеличение размера вставки в клонах ВАС. Из 54 случайных клонов ВАС, выбранных из библиотеки, 50 клонов содержали ДНК риса (93,0%). Некоторые из этих клонов (7%) не содержали вставок. Размеры вставок ДНК колебались в пределах 30-250 т.п.н. при среднем размере в 125 т.п.н.
ДНК с высоким молекулярным весом, использованную для создания библиотеки ВАС, выделяли из очищенных ядер клеток риса. Большую часть хлоропластов и митохондрий удаляли путем центрифугирования при низкой скорости (<1000 g). Низкая частота встречаемости хлоропластных или митохондриальных клонов, обнаруженная в библиотеке ВАС в этом изобретении (<0,3%), снижает возможность колигирования ДНК органелл и ядер.
Библиотеку ВАС использовали для составления примыкающих наборов клонов (примык. ), перекрывающих локус Ха21. Для сортировки библиотеки ВАС использовали два маркера ДНК, связанные с Ха21, RG103 (1 т.п.н. см. Ronald et al., Mol. Gen. Genet. 236, 113-120 (1992)) и рТА818 (1,2 т.п.н., эквивалент RAPD818 у Ronald et al.). RG103 обнаружен в 8 копиях в содержащих Ха21 копиях и гибридизуется с 8 геномными фрагментами ДНК HindIII в этой линии. Все эти фрагменты генетически и физически связаны с локусом устойчивости к заболеваниям Ха21. рТА818 гибридизуется с 2 фрагментами ДНК, и по крайней мере один из этих фрагментов связан с локусом Ха21. Ronald et al. (1992).
Клоны ВАС 7296 испытывали с пробами рТА818 (2 копии) и RG103 (8 копий). Было идентифицировано семь и пять клонов ВАС, гибридизующихся с RG103 и рТА818 соответственно. ДНК ВАС выделяли из этих клонов и расщепляли HindIII. Фрагменты ДНК разделяли с помощью ГЭПЭП. Анализ по Саузерну показал, что 7 ВАС клонов, гибридизующихся с RG103, несли 4 разных копии RG103 геномных HindIII фрагментов. Проба гибридизовалась с фрагментом ДНК в 4,3 т.п.н. и фрагментом в 9,5 т.п.н., фрагментом в 9,6 т.п.н. и фрагментом в 6,2 т.п.н. Размер фрагментов ДНК установлен по лямбда ДНК, расщепленной HindIII.
Выделено четыре клона ВАС, которые несли одну копию HindIII фрагмента рТА818, и идентифицирован один клон ВАС, который содержал другую копию. Один из рТА818, содержащих ВАС, гибридизовался с маркером рТА248 (эквивалентом RAPD248 у Ronald et al. (1992), подтверждающий, что эти два клонированных маркера RAPD составляют 60 т.п.н. друг друга. Ronald et al.(1992).
Идентификация 12 ВАС клонов, гибридизующихся с 2 клонированными последовательностями ДНК (соответствующими 10 фрагментам ДНК в геноме риса), немного ниже, чем 20 клонов, ожидаемых на основе скрининга 2х геномных эквивалентов (7296 клонов, геном 450000 т.п.н., средний размер вставки 125 т.п.н. ). Конкретно, последовательности рТА818 и четыре (из восьми) гибридизующихся с RG103 последовательности излишне представлены в этой части библиотеки. В противоположность этому другие четыре гибридизующихся с RG103 последовательности недостаточно представлены. Размеры вставок ДНК из этих клонов находились в пределах от 40 до 140 т.п.н.
Космидные клоны
А. МАТЕРИАЛЫ И МЕТОДЫ
Получение ДНК с высоким молекулярным весом (ВМВ) из листьев риса
В качестве растительного материала для выделения ДНК с ВМВ использовали линию риса 1188, несущую локус Ха21. Собирали 120 г ткани 4-6 недельных листьев и измельчали в тонкий порошок, используя холодную ступку и пестик в жидком азоте. Порошок затем суспендировали путем перемешивания в 800 мл холодного Н буфера [4 мМ спермидина, 1 мМ спермина, 10 мМ ЭДТУ, 10 мМ Трис-HCl, 80 мМ КСl, 0,5 М сахарозы, 1 мМ ФМСФ (фенилметилсульфонилфторида, добавляется непосредственно перед использованием), 0,5% об. тритона-Х 100, 1/1000 (об. /об. ) β-меркаптоэтанола (добавляется непосредственно перед использованием), рН 9,5]. Смесь фильтровали через 80-микронное сито в бутыли GSA и осадок снова суспендировали в 400 мл буфера и снова фильтровали. Два объема фильтрата объединяли и центрифугировали при 3500 об/мин в течение 10 минут при 4oС. Осадок снова суспендировали в 300 мл буфера для отмывания (такого же, как и буфер Н, за исключением ФМСФ и β-меркаптоэтанола) и центрифугировали при 3500 об/мин в течение 10 минут при 4oС. Осадок промывали дополнительно еще два раза, пока цвет осадка не становился бледно-зеленым. Осадок снова суспендировали в 40 мл раствора для отмывания и ядра лизировали путем добавления равного объема буфера для лизиса (2% лаурилсаркозина натрия, 100 мМ Трис-HCl, 0,5 М ЭДТУ, рН 9,5), содержащего 2 мг/мл протеиназы K (Boeringer Mannheim). Белки удаляли путем инкубации при 50oС в течение 5 часов и затем экстрагирования раствора (путем мягкого перевертывания) равным объемом фенола-хлороформаизоамилового спирта (24:24:1) в течение 30 минут при комнатной температуре. ДНК с ВМВ осаждали путем мягкого наслаивания 1/10 объема 3М ацетата натрия (рН 5,5), 2 объемов этанола и перевертывания несколько раз. И, наконец, ДНК удаляли из этанола, используя пипетку с широким просветом на конце, промывали 70% этанолом, сушили и растворяли в 1 мл ТЭ (10 мМ Трис-HCl, 1 мМ ЭДТУ, рН 8,0) при 4oС в течение ночи без перемешивания. В норме из 120 г листьев можно выделить 250 мкг ДНК с ВМВ.
Получение вставочной ДНК
(А) Частичное расщепление ДНК с ВМВ
Вспомогательный эксперимент. 30 мкг (70 мкл) ДНК с ВМВ смешивали с 10 мкл буфера 10•Sau3AI (NEB) и предварительно нагревали при 37oС в течение 5 мин. Затем к раствору ДНК добавляли 20 мкл (2 единицы) Sau3AI, осторожно перемешивали с помощью пипетки с широким просветом на конце и инкубировали при 37oС. Образцы в 15 мкл отбирали через 0, 5, 10, 20, 30 и 70 минут и сразу же смешивали с 5 мкл 0,5 М ЭДТУ (рН 8,0) на льду, чтобы остановить реакцию. Образцы анализировали с помощью электрофореза в 0,3% агарозно/ТБЭ геле при 2 Вольт/см длины геля в течение 36 часов в холодной комнате.
Крупномасштабное частичное расщепление ДНК достигалось путем повторения вспомогательного эксперимента с использованием оптимизированных интервалов времени инкубации, равных 20 мин при 37oС.
(Б) Селекция по размеру
Частично расщепленную ДНК фракционировали на градиенте плотности сахарозы от 5 до 40% путем центрифугирования в роторе SW27 при 26000 об/мин при 20oС в течение 13 часов. Фракции по 0,8 мл (всего 20) собирали, аккуратно поместив капиллярную трубку на дно центрифужной пробирки и выкачивая градиент при очень низкой скорости. 20 мкл каждого образца исследовали на 0,3% агарозном геле при 2 В/см длины геля в течение 36 часов. Фракции ДНК с размерами примерно 35-50 т.п.н. объединяли вместе. После разведения сахарозы равным объемом воды ДНК осаждали 2 объемами этанола. Частичное наполнение в реакции достигалось с использованием стандартных методик.
Лигирование, заполнение и трансфицирование
Космидный вектор рНС80 был любезно предоставлен доктором Scot Hulbert. Вектор и вставочную ДНК лигировали в молярном отношении 2 к 1 при конечной концентрации 0,8 мкг/мкл. Реакцию дотирования проводили с помощью 600 единиц ДНК-лигазы Т4 (NEB, USA) при 16oС в течение ночи. Лигированную ДНК заполняли in vitro с помощью экстракта для заполнения GigapackII (Stratagene, USA) и трансфицировали в компетентные клетки E.coli NM554 в соответствии с руководством Stratagene.
Скрининг библиотеки
61440 космидных колоний (более пяти геномных эквивалентов) из 160 384-ячеечных плат переносили на фильтры Hybond N+ (Amersham, USA) при двух видах плотности. При первом методе космидные клоны реплицировали при низкой плотности (1536 колоний/11,5•15 см фильтр), используя ручные репликаторы (Genetix, U. K. ), и выращивали на LB/агаре со 100 мкг/мл ампициллина в течение ночи. Чтобы охватить всю космидную библиотеку, было сделано сорок фильтров. При втором методе космидные клоны реплицировали в массивы с высокой плотностью, используя автоматическую рабочую установку Beckman BioteckTM, и подращивали, применяя тот же метод, что и описанный выше. При использовании матрицы 3•3 3456 колоний переносили на фильтр 8,5•12 см. Чтобы точно локализовать положительные колонии на негативном фоне в первом положении каждой сетки 3•3 помещали эталонную космидную колонию (содержащую маркер RG103). В остальные восемь положений штампа помещали колонии из восьми микротитровальных плат космидной библиотеки. В этом случае всю библиотеку могли охватить 20 фильтров размером 8,5•12 см каждый. Для гибридизаций с единственной пробой пробу RG103 смешивали со специфичной пробой в соотношении 1:4 для получения эталонного образца.
Бактерии на фильтрах лизировали и фиксировали, используя методику с пароводяной баней в следующей модификации: колонии помещали лицевой поверхностью вверх на верх двух кусков 3 мм ватмана, пропитанного в растворе для лизиса (0,5 М NaOH, 1,5 М NaCl), на 4 минуты при комнатной температуре, пластиковые коробки, содержащие фильтры, инкубировали на водно-паровой бане при 85oС в течение 6 минут, и затем фильтры переносили на 3 мм ватман, пропитанный нейтрализующим буфером (1 М Трис-НСl (рН 7,4), 1,5 М NaCl), на 4 минуты. Белки и остатки клеток удаляли путем погружения в 50 мл раствора протеиназы К (50 мМ Трис-НСl (рН 8,5), 50 мМ ЭДТУ (рН 8,0), 100 мМ NaCl, 1% (весооб. ), Na-лаурилсаркозина, 250 мкг/мл протеиназы К) и инкубировали при 37oС в течение 20 мин. Фильтры осторожно промывали 2 раза раствором SSC в течение 5 мин при комнатной температуре, сушили и фильтры обрабатывали УФ при 10 см в течение 2,5 мин.
Гибридизацию выполняли по стандартным методикам следующим образом: фильтры перед гибридизацией подвергали воздействию раствора (7% ДСН, 0,5 М Na2PO4 (pH 7,2), 1 мМ ЭДТУ, 100 мкг/мл ссДНК) при 65oС от 2 часов до в течение ночи. Пробы метили, используя методику случайного праймерного удлинения, и гибридизацию выполняли при 65oС при перемешивании в течение ночи. Фильтры недолго промывали в (40 мМ Na2PO4 (pH 7,2), 0,1% ДСН) при комнатной температуре, и фильтры инкубировали в том же самом растворе при 65oС в течение 20 минут при осторожном перемешивании.
Б.РЕЗУЛЬТАТЫ
Три связанных с Ха-21 маркера (RG103, RAPD 248 и RAPD 818) использовали для проверки космидной библиотеки. Геномный анализ по Саузерну показал, что число копий этих трех маркеров в резистентных линиях равно 8, 1 и 2 соответственно (неопубликованные результаты). Было идентифицировано и подтверждено последующим анализом по Саузерну шесть положительных космидных клонов, гибридизующихся с маркером RG103. Однако не было идентифицировано положительных клонов, которые содержат RAPD 248 и RAPD 818.
Пример 2
Определение характеристик генов Ха21
Пять космидных клонов и 1, ВАС клон, выделенные в примере 1, были в дальнейшем охарактеризованы путем рестрикционного ферментативного картирования. Фиг.2А-2Е являются частичными рестрикционными картами космидных клонов. Фиг.2F является частичной рестрикционной картой ВАС клона.
Была идентифицирована открытая рамка считывания у одного из клонов, рВ806 (ПОСЛ. ИД. N 1). Она включает промоторную область, предсказанный интрон и частичную 3'-последовательность. ПОСЛ. ИД. N 2 представляет предсказанную аминокислотную последовательность. Предсказанный интрон был удален (сплайсинг).
Предсказанная аминокислотная последовательность проявляла два характерных признака белков, которые показывают, что она кодируется представителем нового класса генов устойчивости растений к болезням, называемым здесь генами RRK. Во-первых, экстраклеточный домен белков, кодируемых этими генами, включает блок из примерно 23 тандемных богатых лейцином повторов (LRR) со средней длиной в 24 аминокислоты. Мотив LRR участвовал во взаимодействиях белков и лигандном связывании у ряда белков. Экстраклеточный домен также включает область между LRR и сигнальным пептидом, который содержит мотив, SWNTS, который сохраняется у ряда белков, включая Cf-9, PGIP и RLK5. Кроме того, белок включает область с высокой идентичностью последовательности рецептороподобным протеинкиназам (РППК), таким как RLK5 и ТМК1 (Walker et al. Plant J. 3:451 (1993)); Chang et al. Plant Cell 4:1263 (1992); Valon et al. Plant Molec. Biol. 23:415 (1993)), а также продукту гена устойчивости томатов, Pto (Martin et al. Science 262: 1432 (1993). Сигнальный домен, экстраклеточный домен (включая область LRR), трансмембранный домен и домен цитоплазматической киназы идентифицированы в ПОСЛ. ИД. N 2.
Фиг. 3 является рестрикционной картой второго клона, рВ822, который был использован для создания плазмиды, использованной в экспериментах по трансформации, описанных в примере 3, ниже. В этом клоне был также секвенирован ген Ха21 (ПОСЛ. ИД. N 3). В предсказанной аминокислотной последовательности (ПОСЛ. ИД. N 4) обнаружены те же самые мотивы, которые идентифицированы в ПОСЛ. ИД. N 2.
Протеинкиназный домен несет 11 субдоменов, содержащих 15 консервативных остатков, отличительных для протеинкиназ и ограничивается 31 аа перекрывающим мембранным доменом (аа 677-707) и С-концевым доменом. Предполагаемый интрон располагается между двумя высококонсервативными остатками Р и Е (аа 892 и аа 893) в предлагаемом каталитическом домене. Консенсусные последовательности, присутствующие в субдоменах VI (DIKSSN) и VIII(GTIGYAAPE), убедительно свидетельствуют, что Ха-21 обладает серин/треонинкиназной (в противоположность тирозину) активностью.
Предшествующая работа показала, что фосфорилированный белок RLK5 взаимодействует с доменом, взаимодействующим с киназой (KID) типа "С серин-треонин белковой фосфатазой (Stone et al., Science 266: 793-795 (1994)). KID связывает фосфорилированные содержащие LRR белки, RLK и ТМК1, но не связывается с S-связанными рецепторными киназами ZmpK1 и RLK4. Эти результаты свидетельствуют, что KID Arabidopsis функционально аналогичен домену SH2 белков животных. Сравнительный анализ первичной структуры рецептороподобных киназ Arabidopsis RLK5, ТМК1 с Ха-21 выявляет набор консервативных аминокислот (N/Q)X(L/V)S(G/S)(L/A)(F/V)(P/E), соседних по отношению к сериновому остатку, который является карбоксиконцевым для последнего остатка (аргинин), высококонсервативному во всех протеинкиназах (положение 999 в генном продукте Ха21). Карбоксиконцевое положение этого консенсуса в этих белках сходно с фосфотирозином с карбоксилом онкогенного продукта вируса саркомы Рус рр60 c-Src, который является главным для связывания с содержащими домен SH2 белками. Эти консервативные аминокислоты отсутствуют в S-связанных рецепторных киназах ZmpKl, RLK4 и SRK6 и во внутриклеточных киназах, которые не связывают KID. Таким образом, эта область действует как специфичный и с высоким сродством сайт связывания для белков, содержащих KID. Модификация аминокислотной последовательности из этой области Ха21 может, таким образом, использоваться для изменения сродства для белка KID и, таким образом, управляет внутриклеточным сигналом в ответ на лигандное связывание домена LRR.
Пример 3
Трансформация растения с использованием гена Ха-21
Ген Ха21, описанный выше, был использован для трансформации растений риса, чтобы продемонстрировать, что гены могли придавать устойчивость к Xanthomonas восприимчивым растениям. Ген вводили в восприимчивые штаммы риса, используя один из вариантов метода Li et al., Plant Cell Rep. 12: 250-255 (1993). Вкратце котрансформацию проводили, используя гигромициновую конструкцию pMON410 (от Monsato) и вектор-копию, содержащий интересующие последовательности. Кроме того, фрагмент Крn из рВ822 клонировали в вектор рТА818, который получен из вектора Invitrogen, pcr1000 и содержит фрагмент в 1 т.п.н. RAPD 818 (Ronald et al., выше). Полученную плазмиду назвали рС822. Проводили селекцию растений на гигромицин (30 мг/л) и затем отбирали на устойчивость к Хоо, раса 6.
Для определения устойчивости к Xanthomonas у трансформантов использовали стандартные методы. Исследования проводили по методам Kaufman et al. Plant Disease Rep. 57: 537-541 (1973). В кратком изложении, Хоо, раса 6, выращивали на чашках PSA в течение 3 дней. Бактерии снимали, суспендировали в воде и ОП доводили до 109 колониеобразующих единиц на мл. Ножницы погружали в суспензию и листья из трансформированных растений (4 месяца после бомбардировки) обрезали на 5 см от верхушки. Растения оценивали на наличие поражений через 11 дней после заражения.
На фиг.4 показаны данные по длине поражения из экспериментов с использованием вектора экспрессии, содержащего ген из клона рС822. Отдельные растения, полученные от независимых трансформантов 106, -9, -22, -11, -17, -1, -12, -4, -16 и -29, несут конструкцию рС822 и они проявляли повышенную устойчивость по сравнению с чувствительными нетрансформированными контрольными экземплярами (IR24), а также растениями риса, трансформированными вектором (1-15).
Пример 4
Выделение генов RRK из томатов
Как отмечено выше, последовательность Ха21 может использоваться для выделения генов RRK из растений других видов при применении метода вырожденного праймера или метод гибридизации в условиях пониженной жесткости. В этом примере описывается выделение и определение характеристик двух генов RRK из томатов с использованием метода вырожденного праймера.
Вырожденные праймеры были получены так, что продукты ПЦР (полимеразной цепной реакции) должны амплифицироваться между LRR и киназными доменами и поэтому перекрывают трансмембранный домен. Прямые праймеры были взяты из мотивов, сохраненных в области LRR из Ха21 и из нескольких других растительных белков (например, Cf-9, RLK5 и PGIP). Обратные праймеры были взяты из мотивов, сохранных в Ха21 киназном домене, и других серин-треонин-киназных доменов растений (например, RLK5, Pto и Fen (Martin et al. Plant Cell 6:1543-1552 (1994)).
Вырожденные праймеры, использованные для амплификации продуктов ПЦР, были следующими:
1. Область LRR
ТСА AGC ААС ААТ TTG ТСА GGN CA(A/G) АТ(А/С/Т) СС
(ПОСЛ. ИД. N 5)
2. Киназная область
ТАА CAG САС АТТ GCT TGA TTT NAN (G/A)TC NCG(G/A)TG
(ПОСЛ. ИД. N 6)
ТАА CAG САС АТТ GCT TGA TTT NAN (G/A)TC(G/A)CA(G/A)TG
(ПОСЛ. ИД. N 7)
ТАА CAG CAC ATT GCT TGA TTT NAN (G/A)TC (T/C)CT (G/A)TG
(ПОСЛ. ИД. N 8)
ПЦР
Условия ПЦР были следующими (реакция в 20 мкл):
Первый цикл:
94oС в течение 30 с (денатурация)
55oС в течение 30 с (отжиг)
72oС в течение 1 мин (удлинение)
В течение следующих 19 циклов температура отжига снижалась на 1oС каждый цикл. После того как завершалось 20 циклов, реакционную смесь инкубировали в течение 10 мин при 72oС.
После исходной амплификации с использованием этих вырожденных праймеров выполняется второй цикл амплификации со следующими специфическими праймерами:
TAAGCAACAATTTG (ПОСЛ. ИД. N 9)
и
ТАА CAGCACATTGCTTGA (ПОСЛ. ИД. N 10)
Условия для этой амплификации были следующими:
35 циклов
94oС в течение 15 с
55oС в течение 15 с
72oС в течение 15 с
По окончании 35 циклов реакционную смесь инкубировали при 72oС в течение 10 мин.
Продукты ПЦР клонировали и использовали в качестве проб для библиотеки кДНК томатов. Библиотеку создавали из кДНК томатов, для которой затравкой служили олиго-DT праймеры, лигированные с EcoRI адапторами, и которые клонировали в вектор лямбда GT11.
Эти праймеры использовали для выделения двух продуктов ПЦР для томатов и кДНК, принадлежащей к семейству RRK из генов устойчивости к болезням. Первым клоном TRK1 (Tomato Receptor Kinase 1-рецепторная киназа 1 томатов) является продукт ПЦР в 250 т.п.н., и он был использован для выделения частичной кДНК. Эта последовательность ДНК представлена в ПОСЛ. ИД. N11. Выведенная из TRK1 аминокислотная последовательность показана в ПОСЛ. ИД. N12.
Этот клон представлен в виде одной или двух копий в геноме томатов и однокопийных картах к короткому плечу хромосомы 1 вблизи гена устойчивости к Xanthomonas campestris pv. Vesicatoria (Rxl) (Zu et al. Genetics 141:675-682 (1995)) (см. фиг.5).
Вторым клоном TRL1 (Tomato Receptor Like 1 рецептороподобный томатов 1) является продукт ПЦР из 496 т.п.н. Последовательность этой ДНК показана в ПОСЛ. ИД. N 13. Выведенная аминокислотная последовательность представлена ПОСЛ. ИД. N 14 (не показана). TRL1 картируется в нескольких сМ от mcn (см. фиг. 6) мутации на хромосоме 3, которая вызывает пятнистый некроз растений, типичного защитного фенотипа.
Эти результаты показывают, что ген Ха21 может использоваться для выделения генов RRK из других видов растений. Например, гены TRK1 и TRL1, которые были выделены, являются важными компонентами метаболических путей сигнальной трансдукции растения, приводящими к защитной реакции. Эти гены применимы для создания устойчивости к заболеваниям у томатов и других видов растений.
Вышеприведенные примеры представлены для иллюстрации изобретения, но не для ограничения его объема. Другие варианты этого изобретения будут легко видны специалисту в этой области и охватываются прилагаемой формулой изобретений. Все публикации, патенты и патентные заявки, процитированные здесь, включены таким образом путем ссылки.









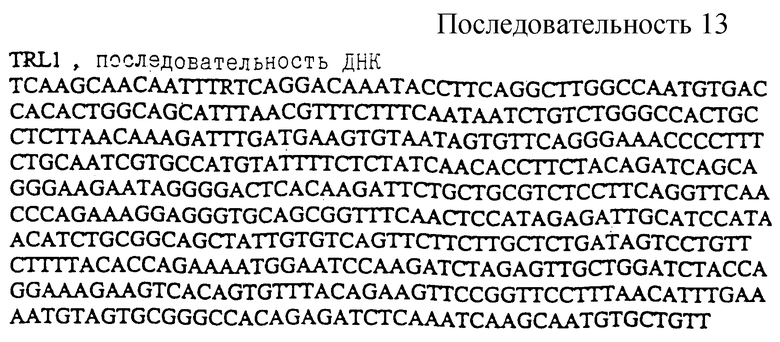

Конструкция нуклеиновой кислоты может быть использована в селекции растений. Трансформация растения посредством конструкции нуклеиновой кислоты позволяет придать ему устойчивость к Xanthomonas spp. за счет экспрессии соответствующих полипептидов. 3 с. и 9 з.п. ф-лы, 6 ил.
Приоритет по пунктам:
17.01.1996 по пп.1 - 10.
| MOLECULAR GENERAL GENETIC | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| Стеклографический печатный станок с ножной педалью | 1922 |
|
SU236A1 |
| EUPHITICA | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| Аппарат, предназначенный для летания | 0 |
|
SU76A1 |

