Перекрестная ссылка на связанные заявки
Настоящая заявка испрашивает приоритет предварительной заявки на патент США 60/808088, поданной 24 мая 2006 г., все содержание которой включено в настоящее описание посредством ссылки.
Область техники, к которой относится изобретение
Изобретение относится к способам и материалам для биосинтеза таких соединений, как симвастатин, включая методики с использованием микроорганизмов-хозяев.
УРОВЕНЬ ТЕХНИКИ
Симвастатин является полусинтетическим производным природного соединения ловастатина, которое может быть выделено из среды культивирования Aspergillus terreus. Ловастатин и симвастатин представляют собой лекарственные средства, понижающие уровень холестерина, которые существенно понижают риск возникновения сердечных заболеваний у взрослых. Ловастатин и симвастатин поставляются фирмой Merck Со. под марками Mevacor и Зокор, соответственно. Симвастатин является более активным производным ловастатина и вторым, наиболее продаваемым препаратом в Соединенных Штатах в 2005 с ожидаемым уровнем продаж в $4,5 миллиардов только в США.
Кластер генов, регулирующих биосинтез ловастатина в А. terreus (см., например, J.Kennedy, К. et. al., Science, 1999, 284, 1368-1372; и C.R.Hutchinson, J. et. al., Antonie Van Leeuwenhoek, 2000, 78, 287-295), был описан ранее (см., например, патент США 6391583, содержание которого включено в настоящее описание посредством ссылки). Кластером генов кодируется белок LovD (46 кДа), который ранее был идентифицирован как гомолог эстеразы. Монаколин J, непосредственный предшественник в биосинтезе ловастатина, собирается расположенной против хода транскрипции мегасинтазой LovB (см., например, L. Hendrickson, C.R. et. al., Chem. Biol. 1999, 6, 429-439), (также известной, как ловастатин нонакетидсинтаза, LNKS), еноилредуктазой LovC и оксигеназами CYP450. Боковая цепь из пяти атомов углерода синтезируется LovF (ловастатин дикетидсинтазой, LDKS) посредством конденсации между ацетил-CoA и малонил-CoA. Конденсированный дикетид подвергается метилированию и восстановительному сшиванию отдельными доменами LovF с получением α-S-метилбутирилтиоэфира, ковалентно связанного с фосфопантетеиновой субъединицей ацилпереносящего белкового домена (ACP) LovF (см., например, J. Kennedy, K. et. al., Science, 1999, 284, 1368-1372 и C. R. Hutchinson, J. et. al., Antonie Van Leeuwenboek 2000, 78, 287-295), в результате из монаколина J образуется Ловастатин. Инактивация либо LovD, либо LovF у A. terreus приводит к накоплению предшественника монаколина J (см., например, J. Kennedy, K. et. al., Science, 1999, 284, 1368-1372 and C. R. Hutchinson, J. et. al., Antonie Van Leeuwenboek 2000, 78, 287-295).
После образования ловастатина, например, посредством ферментации реципиента A. terreus, из ловастатина может быть получен симвастатин. В настоящее время симвастатин является полусинтетическим производным ловастатина. Ловастатин получают в результате ферментации реципиента A. terreus. После очистки соединения полусинтез может быть проведен следующим образом: 1) 2-метилбутиратная боковая цепь может быть гидролизована в присутствии основания с получением в качестве промежуточного соединения монаколина J; 2) лактонизируют свободную кислоту; 3) спиртовую функциональную группу на C13 блокируют защитной группой (такой как трет-бутилдиметилсилильная); 4) этерифицируют доступный спирт C8 ацильным соединением, таким как 2-диметилбутирилхлорид, с получением C13-защищенного варианта симвастатина, и 5) снимают защиты с OH-группы на С13 с получением симвастатина (Фигура 3).
Различные способы многостадийного синтеза симвастатина были описаны ранее (см., например, РСТ WO 2005/066150 и заявки на патент США №№ 20050080275 и 20040068123, содержание которых включено в настоящее описание посредством ссылки). Например, широко используемый способ начинается с гидролиза C8 сложноэфирной группы ловастатина с образованием триола монаколина J с последующим селективным силилированием С13 спирта, этерификацией C8 спирта диметилбутирилхлоридом и снятием защиты с C13 спирта с получением симвастатина (см., например, W. F. Hoffman, et. al., J. Med. Chem. 1986, 29, 849-852). Ферментативные преобразования, применение липаз и эстераз были исследованы как альтернативы химическому синтезу симвастатина (см., например, РСТ WO 2005/040107, РСТ WO 94/26920 и T. G. Schimmel, et. al., Appl. Environ. Microbiol 1997, 63, 1307-1311, содержание которых включено в настоящее описание посредством ссылки). Однако необходимость региоселективной этерификации неизменно требует защиты других гидроксильных групп и часто приводит к снижению общего выхода. Таким образом, специфичный реагент, способный выборочно ацилировать C8-позицию монаколина J, важен для эффективного синтеза симвастатина и дополнительных аналогов статина.
Изменения вышеописанных схем являются обычными, однако большинство методик сначала неизменно требует выделения ловастатина, гидролиза метилбутиратной боковой цепи, защиты свободного спирта, реакции с ацильным соединением и снятия защиты. Хотя используемые химические превращения являются относительно простыми, они неэффективны, включают много стадий и поэтому вносят вклад в текущую высокую стоимость производства симвастатина ($3 за таблетку).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение обеспечивает способы и материалы, разработанные с целью выгодного применения биологических способов получения ловастатина в получении производного ловастатина, симвастатина. Настоящее изобретение также обеспечивает способы и материалы, разработанные с целью выгодного применения биологических способов, посредством которых получают ловастатин, для получения родственных соединений, таких как производное правастатина, хувастатина. Как было указано выше, биологические способы получения ловастатина при ферментации A. terreus известны в уровне техники. В типовых способах получения ловастатина декалиновую основу и группы HMG-CoA, которые воспроизводят фрагменты молекулы ловастатина, синтезируются ловастатин нонакетидсинтазой (LNKS) и тремя дополнительными ферментами in vivo. 2-метилбутиратная боковая цепь синтезируется ловастатин дикетидсинтазой (LDKS) in vivo. 2-метилбутират ковалентно связан с ацил-переносящим доменом LovF посредством тиоэфирной связи (Фигура 6). Затем ацилтрансфераза LovD способна селективно переносить 2-метилбутират на C8-гидроксильную группу от LDKS в одну стадию с получением ловастатина. Как подробно описано ниже, стало возможным получать симвастатин и родственные соединения как in vitro, так и in vivo, посредством управления указанными процессами.
Варианты осуществления настоящего изобретения включают способы получения симвастатина без использования большого количества химических стадий синтеза, которые в настоящее время применяют при получении указанного соединения. Обычные варианты осуществления изобретения не требуют очистки ловастатина на первой стадии с последующими полусинтетическими методиками, и вместо этого используется единственная стадия ферментации с получением в результате симвастатина. Способы, раскрытые в настоящем описании, разработаны так, чтобы ферментационные предприятия, которые в настоящее время производят ловастатин, могли быть переоборудованы для получения симвастатина и родственных соединений с минимальными модификациями. Материалы, используемые в способах, раскрытых в настоящем описании, относительно недороги, а стадии очистки известны из уровня техники и легко осуществляются.
Специалисты, квалифицированные в данной области техники, поймут, что настоящее описание позволяет выполнить большое разнообразие вариантов осуществления изобретения. Типичным вариантом осуществления изобретения является способ получения симвастатина посредством взаимодействия монаколина J; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и LovD ацилтрансферазы; с последующим использованием ацилтрансферазой LovD ацильной группы, источником которой является ацилтиоэфир для региоселективного ацилирования C8-гидроксильной группы монаколина J; в результате чего образуется симвастатин. В типовых вариантах осуществления изобретения ацилтрансфераза LovD имеет аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2. Другим вариантом осуществления изобретения является способ получения симвастатина, включающий стадии взаимодействия ловастатина; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD. В данном варианте осуществления настоящего способа ацилтрансфераза LovD гидролизует ловастатин до монаколина J; с последующим региоселективным ацилированием C8-гидроксильной группы монаколина J; в результате чего образуется симвастатин. В некоторых вариантах осуществления симвастатин получают in vitro в отсутствии выделенного организма.
Как подробно описано ниже, способы и материалы изобретения, которые применяются для получения симвастатина, могут быть приспособлены для получения соединений, структурно подобных симвастатину, например хувастатина. В данном контексте один из вариантов осуществления изобретения относится к способу получения хувастатина, включающему стадии взаимодействия гидролизованного правастатина в форме тетраола; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе гидролизованного правастатина в форме тетраола в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD; с последующим региоселективным ацилированием C8-гидроксильной группы гидролизованного правастатина в форме тетраола ацилтрансферазой LovD, переносящей ацильную группу от ацилтиоэфира, в результате чего образуется хувастатин. Другой вариант осуществления изобретения относится к композиции веществ, включающей: гидролизованный правастатин в форме тетраола; ацилтиоэфир, который отдает ацильную группу C8-гидроксильной группе гидролизованного правастатина в форме тетраола в присутствии ацилтрансферазы LovD; ацилтрансферазу LovD; и хувастатин. Таким образом, варианты осуществления изобретения включают способы получения композиции веществ, включающей симвастатин или хувастатин, как по существу раскрыто и подтверждено примерами в настоящем описании.
В некоторых вариантах осуществления изобретения монаколин J, ацилтиоэфир и ацилтрансфераза LovD взаимодействуют в среде культивирования в присутствии выделенного организма, который продуцирует ацилтрансферазу LovD, и где дополнительно организм представляет собой: Escherichia coli, Aspergillus terreus, Monascus ruber, Monascus purpureus, Monascus pilosus, Monascus vitreus, Monascus pubigerus, Candida cariosilognicola, Aspergillus oryzea, Doratomyces stemonitis, Paecilomyces virioti, Penicillum citrinum, Penicillin chrysogenum, Scopulariopsis brevicaulis или Trichoderma viride. Необязательно выделенный организм представляет собой Aspergillus terreus, который экспрессирует полипептид LovD SEQ ID NO: 1. В некоторых вариантах осуществления Aspergillus terreus не экспрессирует полипептид LovF SEQ ID NO: 3. В альтернативном варианте организм может представлять собой Escherichia coli, которая экспрессирует полипептид LovD SEQ ID NO: 1 или SEQ ID NO: 2. В некоторых вариантах осуществления Escherichia coli не экспрессирует полипептид bioH SEQ ID NO: 4. Как подробно описано ниже, выделенный организм может быть выращен при одних из многих условий культивирования, известных в уровне техники, при этом подходящие условия могут быть выбраны, например, с учетом требований культивирования конкретного используемого организма. В типовых вариантах осуществления изобретения организм выращивают при температуре 30-40°C в течение периода времени, по меньшей мере, от 4 до, по меньшей мере, 48 часов, и при pH 6,5-8,5. В примерах осуществления организм выращивают в среде культивирования, включающей среды LB, F1 или TB.
Необязательно монаколин J, который взаимодействует с другими компонентами в способах изобретения, продуцируется выделенным организмом в среде культивирования, например одним из организмов, указанных выше, который также продуцирует ацилтрансферазу LovD. В альтернативном варианте монаколин J, который взаимодействует с другими компонентами в способах изобретения, продуцируется другим организмом, который продуцирует указанное соединение и добавляется к среде культивирования, и растет вместе с организмом, который продуцирует ацилтрансферазу LovD. В данном контексте множество организмов, которые, как известно из уровня техники, продуцируют монаколин J, может подходить для применения в указанных вариантах осуществления изобретения (см., например Endo et al., J Antibiot (Tokyo). 1985 Mar; 38(3):420-2; и Kennedy et al., 1999 May 21; 284(5418): 1368-72). В другом варианте осуществления изобретения монаколин J получают из альтернативного внешнего источника (например, химического синтеза) и добавляют к ферментационной смеси.
В типовых вариантах осуществления изобретения ацилтиоэфир, который может выступать донором ацильной группы для C8-гидроксильной группы монаколина J в присутствии LovD ацилтрансферазы, получают из внешнего источника (например, процесса химического синтеза) и добавляют к ферментационной смеси. Ряд таких ацилтиоэфиров раскрыт в настоящем описании. Обычно ацилтиоэфир представляет собой бутирил-тиоэфир, N-ацетилцистамин-тиоэфир или метил-тиогликолевый тиоэфир. Необязательно ацилтиоэфир включает ацильные группы с цепью средней длины (C3-C6). В некоторых вариантах осуществления изобретения ацилтиоэфир способен проходить через клеточные мембраны Escherichia coli или Aspergillus terreus, растущих в среде культивирования. Как правило, ацилтиоэфир выбран из группы, состоящей из α-диметилбутирил-S-метил-меркаптопропионата (DMB-S-MMP), диметилбутирил-S-этилмеркаптопропионата (DMB-S-EMP), а также диметилбутирил-S-метилтиогликолята (DMB-S-MTG) и диметилбутирил-S-метилмеркаптобутирата (DMB-S-MMB). В примере осуществления ацилтиоэфир является α-диметилбутирил-S-метил-меркаптопропионатом, который содержится в среде культивирования в концентрации 1 мМ-100 мМ.
В некоторых вариантах осуществления изобретения способ приводит к превращению, по меньшей мере, 95%, 96%, 97%, 98% или 99% монаколина J, добавленного в комбинацию, в симвастатин. Необязательно способ изобретения обеспечивает получение композиции веществ, включающей 0%-1% монаколина J, который был добавлен в комбинацию изначально. Конкретные варианты осуществления способов получения симвастатина и родственных соединений включают дополнительные стадии для очистки указанных соединений. Например, варианты осуществления изобретения могут включать, по меньшей мере, одну стадию очистки, включающую лизис клеток выделенного организма, присутствующего в комбинации. Варианты осуществления могут также включать, по меньшей мере, одну стадию очистки, включающую центрифугирование клеток или лизатов клеток выделенного организма, присутствующего в комбинации. Кроме того, варианты осуществления могут включать, по меньшей мере, одну стадию очистки, включающую осаждение одного или нескольких соединений, присутствующих в комбинации. Один из вариантов осуществления стадии осаждения включает осаждение свободной кислотной формы симвастатина. Необязательно, в таких вариантах осуществления можно затем превращать указанную свободную кислотную форму симвастатина в соль симвастатина. Варианты осуществления изобретения могут также включать, по меньшей мере, одну стадию очистки, включающую фильтрацию одного или нескольких соединений, присутствующих в комбинации. Кроме того, варианты осуществления могут включать, по меньшей мере, одну стадию очистки, включающую анализ с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) одного или нескольких соединений, присутствующих в комбинации.
Варианты осуществления изобретения включают композиции веществ, используемые при осуществлении и/или получаемые посредством способов, раскрытых в настоящем описании. Например, один из вариантов осуществления изобретения относится к композиции веществ, включающей: монаколин J; ацилтиоэфир, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; ацилтрансферазу LovD; и симвастатин. Необязательно композиция дополнительно включает выделенный организм, такой как Escherichia coli, Aspergillus terreus, Monascus ruber, Monascus purpureus, Monascus pilosus, Monascus vitreus, Monascus pubigerus, Candida cariosilognicola, Aspergillus oryzea, Doratomyces stemonitis, Paecilomyces virioti, Penicillum citrinum, Penicillin chrysogenum, Scopulariopsis brevicaulis или Trichoderma viride. В типовых вариантах осуществления организм в композиции представляет собой Aspergillus terreus или Escherichia coli, которые экспрессируют полипептид LovD SEQ ID NO: 1. В одном из вариантов осуществления изобретения организм представляет собой Aspergillus terreus, который не экспрессирует полипептид LovF SEQ ID NO: 3. В другом варианте осуществления изобретения организм представляет собой Escherichia coli, который не экспрессирует полипептид bioH SEQ ID NO: 4. В некоторых вариантах осуществления изобретения выделенный организм в композиции был трансформирован экспрессионным вектором, кодирующим LovD Aspergillus terreus (SEQ ID NO: 1).
Ряд ацилтиоэфиров, которые могут использоваться в композициях изобретения, раскрыт в настоящем описании. Обычно ацилтиоэфир представляет собой бутирил-тиоэфир, N-ацетилцистаминтиоэфир или метилтиогликолевый тиоэфир. Необязательно ацилтиоэфир включает ацильные группы с цепью средней длины (C3-C6). В некоторых вариантах осуществления изобретения ацилтиоэфир способен проходить через клеточные мембраны Escherichia coli или Aspergillus terreus, растущие в среде культивирования. Как правило, ацилтиоэфир выбран из группы, состоящей из α-диметилбутирил-S-метил-меркаптопропионата (DMB-S-MMP), диметилбутирил-S-этилмеркаптопропионата (DMB-S-EMP) и диметилбутирил-S-метилтиогликолята (DMB-S-MTG) и диметилбутирил-S-метилмеркаптобутирата (DMB-S-MMB). В примере осуществления ацилтиоэфир является α-диметилбутирил-S-метил-меркаптопропионатом, который содержится в среде культивирования в концентрации 1 мМ-100 мМ. В некоторых вариантах осуществления изобретения композиция дополнительно включает ловастатин, при этом количество симвастатина в композиции больше, чем количество ловастатина в композиции.
В некоторых вариантах осуществления изобретения дополнительные компоненты и/или стадии методик могут объединять в себе один или несколько способов и материалов, описанных выше. Например, способы могут дополнительно включать использование ферментации при высокой плотности клеток с целью повышения эффективной концентрации ацилтрансферазы LovD, а также оптимизацию условий ферментации или повышение каталитической активности ацилтрансферазы LovD в целях белковой инженерии одного или нескольких тиоэфиров. Много других компонентов или способов могут использоваться в целях повышения продукции симвастатина, либо промежуточных или родственных соединений, которые облегчают получение симвастатина.
Варианты осуществления изобретения также включают изделия и/или наборы, разработанные в целях облегчения осуществления способов изобретения. Обычно такие наборы включают инструкции по использованию элементов набора согласно способам настоящего изобретения. Такие наборы могут включать поддерживающее приспособление, разделенное на ячейки, плотно удерживающее одну или несколько емкостей, таких как флаконы, пробирки и т.п., при этом каждая емкость включает один из отдельных элементов, используемых в способе. Одна из емкостей может включать флакон, например, содержащий ацилтрансферазу LovD и/или А. terreus, а другой флакон при этом содержит тиоэфирное соединение или подобное, оба из которых могут быть добавлены к ферментационной смеси в целях получения симвастатина и/или хувастатина, или подобного. Дополнительные варианты осуществления изобретения описаны ниже.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фигура 1. A. Ловастатин. B. Симвастатин. Два соединения отличаются заменой одного метила в α положении.
Фигура 2. Ловастатин (1), Симвастатин (2), Правастатин (5), Хувастатин (6) и родственные соединения.
Фигура 3. Текущий способ производства симвастатина.
Фигура 4. Реакция переноса ацила в A. terreus согласно изобретению. Серин 76 может быть нуклеофилом в активном центре.
Фигура 5. A) Кривая ВЭЖХ (238 нм), показывающая образование 4 в результате LovD-опосредованного переноса ацила. Градиент: 60%B-95%B, 5 мин; 95%B, 15 мин; A: H2O+0,1% ТФУ; B: ацетонитрил+0,1% ТФУ; a) зависимость от времени LovD + монаколин J+бутирил-CoA (0, 1, 3, 6 и 10 часов); b) LovD S76A+монаколин J+бутирил-CoA, 10 часов; c) LovD+бутирил-SNAC, 10 часов; d) LovD+бутирил-SMTG, 10 часов. Условия анализа: 1 мМ монаколин J, 4 мМ бутирил-CoA, 10 мкМ LovD, 50 мМ HEPES, pH 7,9, 25°C. (B) Превращение в зависимости от времени для (◇) бутирил-CoA (♦) бутирил-SNAC, (■) бутирил-SMTG. Истинные значения kcat, приведенные в тексте настоящего описания, представляют собой начальное число оборотов в линейном диапазоне. (C) График Михаэлиса-Ментен катализируемого LovD гидролиза ловастатина до монаколина J. Ход реакции контролировали с помощью ВЭЖХ. kcat: 0,21±0,01 мин-1; Kм: 0,56±0,05 мМ.
Фигура 6. Катализируемый LovD перенос ацила с образованием ловастатина. Донор ацила присоединен к LDKS.
Фигура 7. Ацил-SNAC. Заштрихованный круг обозначает любую функциональную группу.
Фигура 8. Кривая ВЭЖХ (238 нм), показывающая образование симвастатина и ловастатина в результате LovD-опосредованного переноса ацила. Градиент такой же, как описанный в Фигуре 1; a) внутренние стандарты ловастатина и симвастатина; b) монаколин J+α-J-метилбутирил-SNAC; c) монаколин J+α-диметилбутирил-SNAC. Условия анализа: 1 мМ монаколин J, 10 мМ ацил-SNAC, 100 мкМ LovD, 50 мМ HEPES, pH 7,9, 25°C, 6 часов.
Фигура 9. Кривая ВЭЖХ (238 нм), показывающая образование правастатина и хувастатина в результате LovD-опосредованного ацилирования тетраола 7. Градиент: 5%B-95%B, 5 мин; 95%B, 15 мин; A: H2O+0,1% ТФУ; B: ацетонитрил+0,1% ТФУ; a) внутренние стандарты правастатина и 7; b) 7+α-S-метилбутирил-SNAC; c) 7+α-диметилбутирил-SNAC. Условия анализа: 1 мМ монаколин J, 10 мМ ацил-SNAC, 100 мкМ LovD, 50 мМ HEPES, pH 7,9, 25°C, 6 часов.
Фигура 10. Микробное превращение монаколина J в симвастатин.
Фигура 11. Катализируемый LovD гидролиз и катализируемое LovD ацилирование. Фигура 11 охватывает варианты осуществления изобретения, включающие композиции веществ, из которых в присутствии LovD и донора ацила получают хувастатин или симвастатин. Фигура 11 также иллюстрирует способ получения симвастатина, включающий гидролиз ловастатина в присутствии LovD с получением монаколина J. Монаколин J превращается, таким образом, в симвастатин в присутствии донора ацила. Аналогично фигура 11 также иллюстрирует способ получения хувастатина, включающий гидролиз правастатина в присутствии LovD с получением гидролизованного правастатина. Гидролизованный правастатин, таким образом, превращается в хувастатин в присутствии донора ацила.
Фигура 12. Продуцирование хувастатина в микроорганизме, экспрессирующем LovD. На Фигуре 12 приведены примеры осуществления изобретения, включающие продуцирование хувастатина в организме, экспрессирующем LovD.
Фигура 13. (A) Цельноклеточное биокаталитическое превращение симвастатина в кислотной форме из монаколина J в кислотной форме и DMB-S-MMP. Штамм E. coli с оверэкспрессией ацилтрансферазы LovD. Гидролиз DMB-S-MMP с получением DMB-S-MPA является конкурентной реакцией. (B) Верхняя кривая: обычный профиль реагентов и продуктов в эксперименте цельноклеточного превращения. Нижние кривые: стандарты для DMB-S-MMP и DMB-S-MPA. Спектры регистрировали при 238 нм.
Фигура 14. (A) Тонкослойная хроматография, показывающая органические экстракты шестидесяти штаммов E. coli при добавлении DMB-S-MMP. Описание штаммов приведено в Таблице 2. Пластинку ТСХ поместили в 20%-ный ЭА в гексане с целью разделения DMB-S-MMP (верх) и DMB-S-MPA (низ). Среднее пятно соответствует неизвестному соединению. Во всех штаммах, кроме #56, DMB-S-MMP гидролизовался до DMB-S-MPA через 10 часов инкубации. Штамм ΔbioH не гидролизировал DMB-S-MMP. (B) Подтверждение результатов ТСХ проводили с помощью ВЭЖХ. В экстракте штамма ΔbioH следы DMB-S-MPA не были обнаружены. Спектры регистрировали при 234 нм.
Фигура 15. (A) Кинетика Михаэлиса-Ментен превращения BioH в DMB-S-MMP. Приведены усредненные результаты из трех опытов. Значения kcat и Км - 260±45 с-1 и 229±26 мкМ, соответственно. Высокое стандартное отклонение, наблюдаемое при высокой концентрации DMB-S-MMP, может быть обусловлено плохой растворимостью субстрата в буфере (50 мМ HEPES, pH 7,9, 10% ДМСО). (B) Сравнение скорости гидролиза трех различных диметилбутирилтиоэфиров. Реакции проводили при 1 мМ тиоэфира и 10 нМ BioH.
Фигура 16. (A) Сравнение скорости роста и конечной плотности культур для BL21(DE3)/pAW31 (серый ◇) и YT2/pAW31 (без биотина: o; с биотином (0,15 мг/л): ●) в минимальной среде F1. Клетки выращивали до OD600 ~0,5, индуцировали 100 мкМ ИПТГ и ставили в шейкер на 20°C в течение 12 часов. Штамм ΔbioH YT2 требовал добавления биотина, необходимого для поддерживания устойчивого роста клеток в синтетической среде. (B) Сравнение превращения симвастатина в зависимости от времени (15 мМ MJ, 25 мМ DMB-S-MMP). YT2/pAW31 (o) значительно быстрее, чем BL21(DE3)/pAW31 (●) (данные взяты из Xie et al., (2007) Appl Environ Microbiol 73: 2054-2060), достигает 99% превращения. У обоих штаммов лаг-фазу регистрировали сразу после добавления субстрата (0 часов) и незадолго до завершения реакции. Скорости реакции между лаг-фазами являлись линейными. Число оборотов для YT2/pAW31 (1,5 мМ/ч) было в два раза выше, чем для BL21(DE3) (0,75 мМ/ч).
Фигура 17. ВЭЖХ анализ (238 нм) стадий очистки симвастатина. Симвастатин в кислотной форме лактонизировали с образованием симвастатина перед введением. Кривая a: неочищенный экстракт (смесь культуральной жидкости и клеточного экстракта) YT2/pAW31 после завершения реакции (99% превращения MJ в кислотной форме в симвастатин в кислотной форме). Пик при 6,80 соответствует DMB-S-MMP; Кривая b: Неочищенный образец после промывки равным объемом гексана с целью удаления непрореагировавшего DMB-S-MMP и осаждения; Кривая c: Очищенный симвастатин после промывки dH2O и солюбилизации в AcN. Маленький пик, зарегистрированный при 6,7 мин, соответствует симвастатину в кислотной форме, который не полностью лактонизировался (~1%).
Фигура 18. Химические структуры ловастатина в кислотной форме, монаколина J в кислотной форме и симвастатина в кислотной форме. Исследованная биокаталитическая реакция - ферментативное превращение монаколина J в симвастатин (большая стрелка). LovD способен региоселективно ацилировать C8-гидроксильную группу. Два обычно используемых полусинтетических способа показаны пунктирными стрелками.
Фигура 19. Кинетический анализ катализируемого LovD ацилирования монаколина J с получением симвастатина, где DMB-S-MMP используется в качестве ацилтиоэфира. Ось Y выражает каталитическое превращение (V/Eo) (A) кинетика Михаэлиса-Ментен LovD в зависимости от концентрации монаколина J, при постоянной концентрации DMB-SMMP 2 мМ. Ингибирования субстрата не наблюдали. Км (монаколин J) = 0,78±0,12 мМ. (B) Кинетика Михаэлиса-Ментен LovD в зависимости от концентрации DMB-S-MMP, при постоянной концентрации монаколина J 2 мМ. Км (DMB-S-MMP) = 0,67±0,04 мМ. В обоих анализах значение kcat составило 0,66±0,03 мин-1.
Фигура 20. Ферментация и биокатализ при низкой плотности. Штамм E. coli BL21(DE3)/pAW31, экспрессирующий LovD в течение ночи, сконцентрировали в 10 раз до конечной плотности OD600 22. Субстраты монаколин J и DMB-S-MMP добавляли до конечной концентрации 15 и 40 мМ, соответственно. Превращение в течение времени сопровождалось анализом ВЭЖХ. (A) Кривые ВЭЖХ при исследовании в зависимости от времени. Отмеченные пики - 1: монаколин J (в форме лактона); 2: DMB-S-MPA в качестве продукта гидролиза DMB-S-MMP; 3: DMB-S-MMP; и 4: симвастатин (в форме лактона). (B) Превращение монаколина J в симвастатин в зависимости от времени. Конечное превращение за 24 часа составило 99%. Точки кривой соответствуют усредненным значениям двух опытов.
Фигура 21. Ферментация и биокатализ при высокой плотности. (A) Периодическая ферментация (500 мл) в минимальной среде F1. При OD600 (круги) 5,93 (I) температуру ферментации понижали и поддерживали при КТ. ИПТГ добавляли до конечной концентрации 200 мкМ, после чего начинали подачу исходных веществ, которую поддерживали на уровне 0,08 мл/мин. Измеряли эффективные концентрации LovD при различной продолжительности ферментации (квадраты). (B) Число оборотов монаколина J в симвастатин клетками при четырех различных значениях продолжительности ферментации (обозначены 1, 2, 3 и 4) на Фигуре 21A. Клетки переводили в фазу "покоя", промывая 50 мМ HEPES, pH 7,9, или не переводили, не меняя среду.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Описанные или указанные в настоящем описании методы и методики представляют собой общеизвестные и широко используемые в качестве стандартных методик специалистами, квалифицированными в данной области техники, такие как, например, широко используемые методики молекулярного клонирования, описанные в Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995). При необходимости, методики, включающие использование коммерческих наборов и реактивов, обычно выполняются в соответствии с протоколами и/или параметрами, определенными изготовителем, если не указано иное. Если не определено иное, все термины из уровня техники, примечания и другая научная терминология, используемая в настоящем описании, имеет стандартные значения, известные специалистам в данной области техники, к которым относится настоящее изобретение. В некоторых случаях термины со стандартными значениями определены в настоящем описании в целях ясности и/или в качестве справки, и при включении таких определений в настоящее описание их не следует толковать как существенно отличающимися от общепринятого в данной области техники.
Определения некоторых терминов приведены ниже.
"Ловастатин" (Мевакор) - поликетид, продуцируемый грибом Aspergillus terreus (см., например, A. W. Alberts, J. et. al., Proc. Natl. Acad. Sd. USA., 1980, 77, 3957-3961 и A. Endo, J. Antibiot. 1980, 33, 334-336; а также J. K. Chan, et. al., J. Am. Chem. Soc. 1983, 105, 3334-3336; Y. Yoshizawa, et. al., J. Am. Chem. Soc. 1994, 116, 2693-2694). Ловастатин является фармацевтически важным соединением благодаря его мощной ингибирующей активности в отношении гидроксиметилглутарил-кофермента А редуктазы (HMGR), лимитирующей скорость стадии биосинтеза холестерина, и поэтому широко применяется при лечении гиперлипидемии, гиперхолестеринемии и т.п. Ловастатин также упоминается как Мевакор.
"Симвастатин" - аналог ловастатина. Он находит более широкое применение по сравнению с ловастатином из-за отсутствия нежелательных побочных эффектов и его высокой всасываемости в желудке. Кроме того, сообщалось, что симвастатин предотвращает и снижает риск возникновения болезни Альцгеймера (AD), замедляя синтез Ab42, β-амилоидного пептида, связанного с AD. Из уровня техники известно, что симвастатин может быть получен синтетическим путем посредством прямого метилирования 8'-метилбутирилокси боковой цепи молекулы ловастатина при использовании метилгалогенида в присутствии основания (амида металла). Стадию C-метилирования необходимо проводить при очень низких температурах (от -75 до -30°), используя сильное основание в безводных условиях, что весьма трудно выполнять при массовом производстве (см., например, патент США 5393893, патент США 4582915, патент США 5763646, патент США 5763653, Европейский патент 299656 и Международную патентную публикацию WO 99/45003, содержание которых включено в настоящее описание посредством ссылки). В уровне техники также известны другие способы синтетического получения симвастатина. Например, ловастатин можно гидролизовать избыточным количеством гидроксида лития с целью удаления 2-метилбутириловой боковой цепи и одновременного размыкания 6-членного лактонового кольца с получением триолкислоты. Затем триолкислотное соединение может быть нагрето с получением диоллактона. Гидроксильная группа на лактоновом кольце диоллактона может быть защищена с получением трет-бутилдиметилсилильного эфира, после чего гидроксильная группа в положении C8 кольцевой системы гексагидронафталина может быть ацилирована 2,2-диметилбутановой кислотой в присутствии дициклогексилкарбодиимида, или 2,2-диметилхлорида с получением соединения. Затем на последней стадии защитная т-бутилдиметилсилильная группа соединения может быть удалена с помощью фторида тетрабутиламмония с получением симвастатина (см., например, патент США 4444784, содержание которого включено в настоящее описание посредством ссылки).
Используемое в настоящем описании понятие "производные ловастатина" включает производные или предшественники ловастатина, например правастатин, хувастатин, симвастатин или гидролизованный правастатин в форме тетраола. Понятие "варианты монаколина J" относится к вариантам монаколина J, описанным в уровне техники, например гидролизованному правастатину в форме тетраола или 6-гидрокси-6-деметилмонаколин J и т.п. В некоторых вариантах осуществления изобретения понятие "варианты монаколина J" относится к соединениям монаколина J, имеющим замены в положении C6 на Фигуре 2. При описании таких соединений, как симвастатин, правастатин, монаколин J и варианты, и т.д., специалисты в данной области будут осведомлены, что указанная формулировка охватывает не только указанные соединения, но и соли указанных соединений (например, фармацевтически приемлемые соли, известные в уровне техники). Например, как известно из уровня техники, симвастатин может находиться как в свободной кислотной форме, так и в форме солей натрия, калия или аммония, а также других солей симвастатина, полученных со щелочноземельными элементами, или солей других металлов.
"Aspergillus terreus" или "A. terreus" представляет собой нитчатый аскомицет, обычно присутствующий в почве. Различные штаммы A. terreus известны в уровне техники, например депонированные как, например, ATCC 20542 и ATCC 20541.
Как известно из уровня техники, гены, отвечающие за биосинтез вторичных метаболитов нитчатых грибов, могут образовывать кластер в геноме гриба и упоминаются как "кластеры генов". Например, "ловастатин-продуцирующий кластер генов" может относиться к набору генов, регулирующих синтез ловастатина, который включает P450I (LovA); дегидрогеназу (LovC); эстеразу и ацилтрансферазу (LovD); а также ScPKS или LDKS (LovF). Ранее было определено, что каждый из указанных четырех генов (LovA, LovC, LovD и LovF) требуется для синтеза ловастатина (см., например, патент США 6943017, содержание которого включено в настоящее описание посредством ссылки). LovF (ген LDKS) охарактеризован как ген поликетидсинтазы. LovD является предполагаемым эстераза/карбоксипептидаза-подобным геном. Ранее было выполнено прерывание гена LovF (см., например, Патент США 6943017, содержание которого включено в настоящее описание посредством ссылки). В результате взаимодействия LovD с LovF синтезируется ловастатин, однако для синтеза симвастатина взаимодействия LovD-LovF не требуется. Кроме того, другой ген в ловастатин-продуцирующем кластере генов, LovE, кодирует цинковый палец, который может регулировать транскрипцию других генов. Ловастатин-продуцирующий кластер генов также включает LovB (ген NPKS).
Термины "LDKS" или "ген LDKS" относятся к белку, кодируемому геном LovF, членом ловастатин-продуцирующего кластера генов. LDKS означает ловастатин-дикетидсинтазу. LovF представляет собой ген, который кодирует LDKS. LovF также является геном, который кодирует белок LovF. "Ген LDKS" также относится к гену, который кодирует LDKS. В синтезе ловастатина LDKS синтезирует боковую цепь из пяти атомов углерода монаколина J посредством конденсации между ацетил-CoA и малонил-CoA. Конденсированный дикетид подвергается метилированию и восстановительному сшиванию отдельными доменами LovF с получением α-S-метилбутирилтиоэфира, ковалентно связанного с фосфопантетеиновой субъединицей ацилпереносящего белкового домена (ACP) LovF.
Используемый в настоящем описании термин "ацилтрансфераза LovD" относится к таким полипептидам, как полипептид LovD A. terreus (например SEQ ID NO: 1), который может использовать ацилтиоэфир для региоспецифичного ацилирования гидроксильной группы монаколина J в положении C8 с получением симвастатина. Кроме того, в настоящем описании раскрыто, что указанный фермент LovD может также использовать ацилтиоэфир для региоспецифичного ацилирования C8-гидроксильной группы гидролизованного правастатина в форме тетраола с получением хувастатина.
Ацилтрансферазы LovD включают гомологичные полипептиду LovD A. terreus ферменты (например SEQ ID NO: 1), которые могут быть обнаружены, помимо прочего, например, в кластерах поликетидных генов грибов. Например, в уровне техники можно найти свидетельство, что Mlc в пути биосинтеза компактина катализирует идентичную реакцию переацилирования (см., например, Y. Abe, T. et al., Mol Genet Genomics. 2002, 267, 636-646), тогда как ацилтрансфераза в сквалестатиновом пути может катализировать подобную реакцию между ACP-связывающим тетракетидтиоэфиром и агликоном (см., например, R. J. Cox, F. et. al., Chem Commun (Camb) 2004, 20, 2260-2261). Аминокислотная последовательность полипептида LovD A. terreus (например SEQ ID NO: 1) подобна β-лактамазам C типа, которые катализируют гидролитическую инактивацию β-лактамных антибиотиков (см., например, E. Lobkovsky, E. M. et. al., Biochemistry, 1994, 33, 6162-6112 и A. Dubus, D. et. al., Biochem. J. 1993, 292, 537-543). Выравнивание полипептида LovD A. terreus (например, SEQ ID NO: 1) с лактамазой P99 из Enterobacter cloacae (см., например, S. D. Goldberg, et. al., Protein Sci. 2003, 12, 1633-1645) показало среднюю гомологичность последовательности, включая потенциально консервативные остатки активного центра, например, каталитические Ser76, Lys79, Tyr188 и Lys315 (см., например, S. D. Goldberg, et. al., Protein Sci. 2003, 12, 1633-1645).
Ацилтрансферазы LovD могут также относиться как к генноинженерным, так и природным ферментам, которые подобны полипептиду LovD A. terreus (например, SEQ ID NO: 1) по последовательности, но содержат небольшие различия в аминокислотном составе (например, мутации с заменой 1-10 аминокислот). Симвастатин, например, может быть получен из природных ферментов, которые подобны полипептиду LovD A. terreus (например SEQ ID NO: 1) по последовательности (например MICH из кластера компактина). Термин "ацилтрансферазы LovD" может также относиться к мутантным формам полипептида LovD A. terreus (SEQ ID NO: 1). Из уровня техники известно, что мутантные формы могут быть получены путем использования стандартных методов молекулярной биологии с получением, например, мутантных форм SEQ ID NO: 1, которые улучшают каталитическую активность и т.п. Например, на данный момент авторы настоящего изобретения используют рациональные и направленные методы изменения для улучшения скорости каталитического превращения LovD A. terreus. Обычно подобные мутанты имеют последовательность с подобием 50%-99% к SEQ ID NO: 1. В данном контексте термин "фермент, гомологичный LovD" включает полипептид LovD, последовательность которого, по меньшей мере, на 80%, 85%, 90%, 95%, 97%, 98% или 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO: 1, где указанный полипептид обладает способностью использовать ацилтиоэфир для региоспецифического ацилирования C8-гидроксильной группы монаколина J с получением симвастатина и/или использовать ацилтиоэфир для региоспецифического ацилирования C8-гидроксильной группы гидролизованного правастатина в форме тетраола с получением хувастатина. Такие мутанты легко получают, а затем идентифицируют в анализе, который позволяет обнаруживать требуемое соединение, например симвастатин (обычно в качестве контроля используется полипептид LovD A. terreus (например SEQ ID NO: 1)). Указанные мутанты могут применяться согласно способам настоящего изобретения, например, для получения симвастатина или хувастатина.
Термин "гетерологичный", в случае если относится к последовательностям нуклеиновых кислот, таким как кодирующие последовательности и регуляторные последовательности, обозначает последовательности, которые обычно не связываются с областью рекомбинантной конструкции и/или обычно не связываются с отдельной клеткой. Таким образом, "гетерологичная" область сконструированной нуклеиновой кислоты может представлять собой идентифицируемый фрагмент в нуклеиновой кислоте или присоединенный к другой молекуле нуклеиновой кислоты, которая, как известно, не связана с остальной молекулой в природе. Например, гетерологичная область конструкции может включать кодирующую последовательность между последовательностями, не связанными с кодирующей последовательностью в природе. Точно так же клетка-хозяин, трансформированная конструкцией, которая обычно не присутствует в клетке-хозяине, может считаться гетерологичной (см., например, патенты США 5712146, 6558942, 6627427, 5849541, содержание которых включено в настоящее описание посредством ссылки). Например, конструкцию с Lov-генами можно выделить и экспрессировать в непродуцирующих ловастатин грибковых или дрожжевых клетках-хозяевах, в результате, таким образом, может быть получен ловастатин (см., например, патенты США 6391583 и 6943017, содержание которых включено в настоящее описание посредством ссылки). В качестве другого примера, прокариоты, такие как бактерии, также могут являться клетками-хозяевами, если известны в уровне техники. Гены грибков могут быть также клонированы в экспрессионный вектор для экспрессии в прокариотах (см., например, патент США 5849541, содержание которого включено в настоящее описание посредством ссылки).
Прокариот, такой как E. coli, может использоваться в качестве гетерологичного организма-хозяина. Можно сконструировать несущую целевой ген плазмиду, которой можно трансформировать E. coli. Целевой ген может быть транслирован с получением соответствующего целевому гену белка, который затем может быть очищен. Указанный способ экспрессии и очистки белка известен в уровне техники. Например, экзоны LovD A. terreus можно отдельно амплифицировать из геномной ДНК A. terreus и соединить, получив непрерывную открытую рамку считывания, используя сплайсинг при помощи ПЦР с удлинением перекрывающихся фрагментов. Могут быть введены сайты рестрикции, а кассета гена может быть лигирована с вектором с получением экспрессионной конструкции, которой можно трансформировать E. coli. Таким образом, E. coli может использоваться в качестве гетерологичного организма-хозяина для экспрессии генов A. terreus. E. coli можно культивировать вместе с другим штаммом, продуцирующим другой целевой субстрат. Дополнительно к указанной культуре или сокультуре могут быть добавлены субстраты. Гетерологичная экспрессия генов биосинтеза ловастатина известна в уровне техники (см., например, патенты США 6391583 и 6943017, содержание которых включено в настоящее описание посредством ссылки).
В качестве другого примера некоторые поликетиды, например поликетиды грибов или других организмов, можно гетерологично экспрессировать в E. coli, дрожжах и других организмах-хозяевах. В среды для указанных организмов-хозяев могут быть добавлены другие субстраты, так как для них может быть необходима как гетерологичная экспрессия нужной PKS, так и ферменты, которые синтезируют, по меньшей мере, некоторые из молекул-субстратов, требуемых для PKS (см., например, патент США 7011959, содержание которого включено в настоящее описание посредством ссылки). Точно так же Lov-гены грибков могут быть экспрессированы в E. coli или другой бактерии, при этом в среды для указанных бактерий-хозяев могут быть внесены другие субстраты, такие как ацил-SNAC или другие доноры ацильной группы. Указанные доноры ацильной группы могут проходить через клеточную мембрану и проникать в бактериальную клетку.
Термин "экспрессионный вектор" относится к нуклеиновой кислоте, которая может быть введена в клетку-хозяина. Как известно из уровня техники, экспрессионный вектор может сохраняться в клетке постоянно или транзиентно, либо как часть хромосомной или другой ДНК в клетке, либо в любой клеточной структуре, например, как реплицирующийся вектор в цитоплазме. Экспрессионный вектор также включает промотор, который регулирует экспрессию РНК, которая обычно транслируется в полипептид в клетке или клеточном экстракте. Например, подходящие промоторы для включения в экспрессионные векторы изобретения включают такие промоторы, которые работают в эукариотических или прокариотических клетках-хозяевах. Промоторы могут включать регуляторные последовательности, которые обеспечивают регуляцию экспрессии по отношению к росту клетки-хозяина или которые вызывают включение или выключение экспрессируемого гена в ответ на химическое или физическое воздействие. В E. coli и некоторых других бактериальных клетках-хозяевах могут использоваться промоторы, происходящие из генов биосинтетических ферментов, ферментов, придающих устойчивость к антибиотикам, а также фаговых белков, которые включают, например, галактозный, лактозный (lac), мальтозный, триптофановый (trp), бета-лактамазный (b/a) промоторы, а также PL-промотор бактериофага лямбда и промотор бактериофага T5. Кроме того, могут также использоваться синтетические промоторы, такие как tac-промотор (патент США 4551433). Экспрессионные векторы E. coli предпочтительно включают начало репликации E. coli, например, из pUC, p1P, p1I и pBR. Для эффективной трансляции РНК в белок экспрессионный вектор также обычно содержит последовательность сайта связывания рибосомы, расположенную перед стартовым кодоном кодирующей последовательности экспрессируемого гена. Также в экспрессионном векторе могут присутствовать другие элементы, такие как энхансеры, последовательности сигналов секреции, последовательности завершения транскрипции, а также один или несколько маркерных генов, с помощью которых клетки-хозяева, содержащие вектор, могут быть идентифицированы и/или отобраны. Могут использоваться селективные маркеры, то есть гены, придающие устойчивость или чувствительность к антибиотикам, которые придают селективный фенотип трансформированным клеткам, когда клетки выращивают на соответствующей селективной среде. Например, экспрессионный вектор, содержащий кластер Lov-генов или его части, может быть введен в гетерологичного хозяина, например E. coli. Таким образом, рекомбинантные векторы экспрессироваия могут содержать, по меньшей мере, одну систему экспрессии, которая, в свою очередь, может быть сконструирована, по меньшей мере, из части Lov и/или другого биосинтетического гена, кодирующего последовательности, функционально связанные с промотором и необязательно терминирующими последовательности, которые регулируют экспрессию кодирующей последовательности в совместимых клетках-хозяевах.
"Кодирующая последовательность" может представлять собой последовательность, которая "кодирует" специфический ген, такой как, например, ген из кластера Lov-генов. Кодирующая последовательность является последовательностью нуклеиновых кислот, которая транскрибируется (в случае ДНК) и транслируется (в случае мРНК) в полипептид in vitro или in vivo под контролем соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяются стартовым кодоном на 5' (амино) конце и терминирующим кодоном на 3' (карбокси) конце. Последовательность терминации транскрипции обычно расположена на 3'-конце кодирующей последовательности.
ДНК "регуляторные последовательности" в совокупности относятся к промоторным последовательностям, сайтам связывания рибосомы, сигналам полиаденилирования, последовательностям терминации транскрипции, регуляторным областям перед целевым геном, энхансерам и т.п., которые в совокупности обеспечивают транскрипцию и трансляцию кодирующей последовательности в клетке-хозяине.
Понятие "функционально связанный" относится к расположению элементов, где вышеописанные компоненты расположены так, чтобы выполнять свою обычную функцию. Таким образом, регуляторные последовательности, функционально связанные с кодирующей последовательностью, способны регулировать экспрессию кодирующей последовательности. Регуляторные последовательности не должны обязательно контактировать с кодирующей последовательностью, пока их функцией является управление экспрессией последней.
"Ловастатин-продуцирующий организм" относится к большому ряду различных организмов, как известно из уровня техники, продуцирующих ловастатин. Указанные организмы, продуцирующие ловастатин, могут быть модифицированы с целью продуцирования симвастатина способами настоящего изобретения. A. terreus является одним из примеров ловастатин-продуцирующего организма. Помимо A. terreus сообщалось о других микроорганизмах, продуцирующих ловастатин (мевинолин), включающих виды Monascus, например M. ruber, M. purpureus, M. pilosus, M. vitreus, M. pubigerus, а также виды Penicillium, Hypomyces, Doratomyces, Pboma, Eupenicillium, Gymnoascus и Trichoderma, Pichia labacensis, Candida cariosilognicola, Aspergilus oryzea, Doratomyces stemonitis, Paecilomyces virion, Penicillum citrinum, Penicillin chrysogenum, Scopulariopsis brevicaulis и Tricboderma viride (см., например, патент США 6391583; Juzlova et al., J. Ind. Microbiol. 16:163-170; Gunde-Cimerman et al., FEMS Microbiol. Lett. 132:39-43 (1995); а также Shindia et al., Folio Microbiol. 42:477-480 (1997), содержание которых включено в настоящее описание посредством ссылки).
Используемое в настоящем описании понятие "организмы, не продуцирующие ловастатин" относится к ряду организмов, которые не продуцируют ловастатин без манипуляций человека (например, E. Coli). На указанные организмы можно воздействовать так, чтобы они продуцировали LovD, или же, например, культивировать их в присутствии LovD с получением ловастатина или симвастатина способами настоящего изобретения.
"A. terreus с прерваным геном LDKS" включает A. terreus без гена LDKS, с мутированным геном LDKS, с нокаутированным геном LDKS, с делецией гена LDKS, с геном LDKS, экспрессия которого прервана, или с прерванным геном LDKS. "A. terreus с прерваным геном LDKS" включает A. terreus с геном LDKS, транскрипция которого была подавлена способами, известными из уровня техники. "A. terreus с прерваным геном LDKS" относится к A. terreus, не способному продуцировать функциональный LDKS. "A. terreus с прерваным геном LDKS" может также относиться к A. terreus, который продуцирует функциональный LDKS. LDKS может быть инактивирован или ингибирован способами, известными из уровня техники, например, с использованием методик нокаута гена. Количество присутствующего LDKS может быть уменьшено способами, известными из уровня техники. Другие способы ингибирования, инактивации или прерывания гена LDKS или белка включают, помимо прочих, антисмысловую, миРНК, РНКи или РНК-интерференцию, известные в уровне техники. Используемый в настоящем описании термин "ген LDKS" может также относиться к гену LovF. Прерывание гена LovF известно в уровне техники (см., например, патент США 6391583, содержание которого включено в настоящее описание посредством ссылки). "A. terreus с прерваным геном LDKS" обычно является генетически модифицированным организмом. Генетическая модификация A. terreus известна в уровне техники. Прерывание Lov-генов в A. terreus было выполнено ранее (см., например, патенты США 6391583 и 6943017, содержание которых включено в настоящее описание посредством ссылки). Прерывание только гена LovF (продуцирующего LDKS) в A. terreus было выполнено ранее (см., например, патент США 6943017, содержание которого включено в настоящее описание посредством ссылки). Прерывание гена LovF можно выполнить другими способами, известными из уровня техники. A. terreus с прерваным геном LDKS может присутствовать в ферментационной смеси. К ферментационной смеси A. terreus с прерваным геном LDKS могут быть добавлены субстраты с целью получения аналогов ловастатина.
Используемое в настоящем описании понятие "компонент или способ, направленный на повышение продукции симвастатина" относится к соединению или субстрату, синтетическому или природному, которые повышают продукцию некоторых промежуточных соединений с увеличением количества получаемого симвастатина в масштабном и крупномасштабном синтезе симвастатина. Компоненты и способы, направленные на повышение продукции некоторых промежуточных соединений, известны в уровне техники. Например, соединения, которые добавляют к ферментационной смеси для увеличения количества промежуточных соединений, таких как монаколин J в производстве ловастатина, известны в уровне техники (см., например, патент США 6943017, содержание которого включено в настоящее описание посредством ссылки). Некоторые из указанных промежуточных соединений, такие как монаколин J, могут также использоваться в производстве симвастатина. Соединения для повышения продукции монаколина J, таким образом, могут быть добавлены с целью повышения продукции симвастатина. Например, соединения для повышения продукции монаколина J могут быть непосредственно добавлены к ферментационной смеси с целью увеличения количества получаемого симвастатина. Примером компонента, направленного на повышение продукции симвастатина, является клон, содержащий фрагмент D4B ловастатин-продуцирующего кластера генов, который депонирован в ATCC под номером 98876. Указанным клоном можно трансформировать организм, не продуцирующий ловастатин, с получением монаколина J, как известно из уровня техники. Указанным клоном также можно трансформировать ловастатин-продуцирующий организм с целью повышения продукции монаколина J и, таким образом, повышения продукции симвастатина. Кроме того, другим примером компонента, направленного на повышение продукции симвастатина, является ген LovE/цинковый палец, которым можно трансформировать ловастатин-продуцирующий организм с целью повышения продукции симвастатина. Предпочтительно указанный ловастатин-продуцирующий организм должен иметь прерванный ген LDKS (см., например, патент США 6391583, содержание которого включено в настоящее описание посредством ссылки). Компоненты и способы, направленные на повышение продукции симвастатина, могут относиться ко многим другим компонентам и способам, и не должны ограничиваться приведенными выше примерами.
Раскрытые в настоящем описании "донор ацила" или "переносчик ацила" представляют собой соединения с ацильной группой, которая может быть перенесена на симвастатин и/или предшественник симвастатина, или родственное соединение. Как правило, "донор ацила" или "переносчик ацила" является ацилтиоэфиром, который отдает ацильную группу C8-гидроксильной группе монаколина J. В уровне техники известно большое разнообразие соответствующих агентов, которые, как показано далее в настоящем описании, обладают указанной активностью (см. примеры ацилтиоэфиров в Таблице 1). В дополнение к известным в уровне техники и, как показано далее в настоящем описании, обладающий указанной активностью, любой потенциальный донор/переносчик ацила, известный в уровне техники (или синтезированный de novo), обладающий способностью ацилировать монаколин J в C8 положении с получением симвастатина, может быть легко идентифицирован с помощью сравнительных экспериментов с донорами ацила, раскрытыми в настоящем описании (например ацил-SNAC). Как известно в уровне техники, ацильная группа может иметь формулу RCON, в которой R может являться алкилом или арилом, а N может представлять собой -Cl, -OOCR, -NH2, -OR и т.п. Соединения, которые имеют ацильную группу, включают, помимо прочих, хлорангидриды, сложные эфиры, амиды или ангидриды и т.п. Указанные соединения могут быть алифатическими или ароматическими, замещенными или незамещенными. Соответствующие примеры включают, помимо прочих, бензоилхлорид, бензойный ангидрид, бензамид или этилбензоат и т.п. Другие примеры доноров ацила включают, помимо прочих, α-диметилбутирил-SNAC, ацил-тиоэфиры, ацил-CoA, бутирил-CoA, бензоил-CoA, ацетоацетил-CoA, β-гидроксибутирил-CoA, малонил-CoA, пальмитоил-CoA, бутирил-тиоэфиры, N-ацетилцистаминтиоэфиры (SNAC), метил-тиогликолят (SMTG), бензоил-SNAC, бензоил-SMTG, или α-S-метилбутирил-SNAC. Указанные соединения могут быть получены в природе или синтетическим путем, а в некоторых случаях могут проникать через клеточную мембрану. Некоторое количество указанных соединений может быть добавлено, например, к LovD в присутствии монаколина J с получением симвастатина.
Используемое в настоящем описании обозначение "ацил-SNAC" относится к α-диметилбутирил-SNAC. Как известно в уровне техники, ацил-SNAC может проникать через клеточную мембрану in vivo. LovD может использовать ацил-SNAC в качестве субстрата, чтобы начать реакцию с превращением монаколина J в симвастатин посредством региоспецифического ацилирования C8-гидроксильной группы монаколина J. Ацил-SNAC может отдавать свою ацильную группу LovD.
ТИПОВЫЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Специалисты, квалифицированные в данной области техники, поймут, что настоящее описание позволяет разработать большое разнообразие вариантов осуществления настоящего изобретения. Типовым вариантом осуществления изобретения является способ получения симвастатина посредством взаимодействия монаколина J; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD; с последующим использованием ацилтрансферазой LovD ацильной группы, источником которой является ацилтиоэфир для региоселективного ацилирования C8-гидроксильной группы монаколина J; в результате чего образуется симвастатин. В примерах осуществления изобретения ацилтрансфераза LovD имеет аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2. Другим вариантом осуществления изобретения является способ получения симвастатина, включающий стадии взаимодействия ловастатина; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD. В данном варианте осуществления настоящего способа ацилтрансфераза LovD гидролизует ловастатин до монаколина J; с последующим региоселективным ацилированием C8-гидроксильной группы монаколина J; в результате чего образуется симвастатин. В некоторых вариантах осуществления симвастатин получают in vitro в отсутствии выделенного организма.
В некоторых вариантах осуществления изобретения монаколин J; ацилтиоэфир и ацилтрансфераза LovD взаимодействуют в среде культивирования в присутствии выделенного организма, который продуцирует ацилтрансферазу LovD, и где дополнительно организм представляет собой: Escherichia coli, Aspergillus terreus, Monascus ruber, Monascus purpureus, Monascus pilosus, Monascus vitreus, Monascus pubigerus, Candida cariosilognicola, Aspergillus oryzea, Doratomyces stemonitis, Paecilomyces virioti, Penicillum citrinum, Penicillin chrysogenum, Scopulariopsis brevicaulis или Trichoderma viride. Необязательно выделенный организм представляет собой Aspergillus terreus, который экспрессирует полипептид LovD SEQ ID NO: 1. В некоторых варианты осуществления Aspergillus terreus не экспрессирует полипептид LovF SEQ ID NO: 3. В некоторых вариантах осуществления изобретения активность экспрессируемых полипептидов, таких как в SEQ ID NO: 3 и 4, снижена, по меньшей мере, до 90, 95 или 99% их эндогенной активности. В альтернативном варианте организм может представлять собой Escherichia coli, экспрессирующую полипептид LovD SEQ ID NO: 1. В некоторых вариантах осуществления Escherichia coli не экспрессирует полипептид bioH SEQ ID NO: 4. Таким образом, организм-хозяин может представлять собой либо организм, который продуцирует ацилтрансферазу LovD эндогенно, либо, в качестве альтернативы, гетерологичный организм-хозяин, в который был введен ген ацилтрансферазы LovD, например, с помощью метода клонирования, известного в уровне техники и дополнительно рассмотренного в настоящем описании. В типовом варианте осуществления гетерологичный организм-хозяин, который экспрессирует ацилтрансферазу LovD, представляет собой бактерию, дрожжи или грибы, которые, как известно из уровня техники, используются в таких целях.
Как подробно рассмотрено ниже, выделенный организм может быть выращен в одних из различных условий ферментации, известных в уровне техники, при этом необходимые условия выбраются, например, в зависимости от параметров ферментации, связанных с оптимальным ростом конкретного организма, используемого в варианте осуществления изобретения (см., например. Miyake et al., Biosci. Biotechnol. Biochem., 70(5): 1154-1159 (2006) и Hajjaj et al., Applied and Environmental Microbiology, 67: 2596-2602 (2001), содержание которых включено посредством ссылки). Обычно организм выращивают при температуре 30-40°C, в течение периода времени, по меньшей мере, между 4 и, по меньшей мере, 48 часами. Обычно организмы выращивают при pH в диапазоне 6,5-8,5. В некоторых вариантах осуществления изобретения pH среды культивирования может составлять 6,9, 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9, 8,0 или 8,1. В примерах осуществления организм выращивают в среде культивирования, включающей среды LB, F1 или TB.
Необязательно монаколин J, который взаимодействует с другими компонентами в способах изобретения, продуцируется выделенным организмом в среде культивирования, например, одним из организмов, указанных выше, который также продуцирует ацилтрансферазу LovD. В альтернативном варианте монаколин J, который взаимодействует с другими компонентами в способах изобретения, продуцируется другим организмом, который продуцирует указанное соединение и добавляется к среде культивирования, и растет вместе с организмом, который продуцирует ацилтрансферазу LovD. В другом варианте осуществления изобретения монаколин J получают из внешнего источника и добавляют к ферментационной смеси. Необязательно способ изобретения обеспечивает получение композиции веществ, включающей 0%-1% монаколина J, который был первоначально добавлен к комбинации. В некоторых вариантах осуществления изобретения способ приводит к превращению, по меньшей мере, 95% монаколина J, добавленного к комбинации, в симвастатин.
В типовых вариантах осуществления изобретения ацилтиоэфир, который может выступать донором ацильной группы для C8-гидроксильной группы монаколина J в присутствии ацилтрансферазы LovD, получают из внешнего источника (например, процесса химического синтеза) и добавляют к ферментационной смеси. Ряд таких ацилтиоэфиров раскрыт в настоящем описании. Обычно ацилтиоэфир представляет собой бутирил-тиоэфир, N-ацетилцистамин-тиоэфир или метил-тиогликолевый тиоэфир. Необязательно ацилтиоэфир включает ацильные группы с цепью средней длины (C3-C6). В некоторых вариантах осуществления изобретения ацилтиоэфир способен проходить через клеточные мембраны Escherichia coli или Aspergillus terreus, растущих в среде культивирования. Как правило, ацилтиоэфир выбран из группы, состоящей из α-диметилбутирил-S-метил-меркаптопропионата (DMB-S-MMP), диметилбутирил-S-этилмеркаптопропионата (DMB-S-EMP), а также диметилбутирил-S-метилтиогликолята (DMB-S-MTG) и диметилбутирил-S-метилмеркаптобутирата (DMB-S-MMB). В примере осуществления ацилтиоэфир является α-диметилбутирил-S-метил-меркаптопропионатом, который содержится в среде культивирования в концентрации 1 мМ-100 мМ.
Некоторые варианты осуществления способов получения симвастатина включают дополнительные стадии для очистки симвастатина в комбинации. Например, некоторые варианты осуществления изобретения включают, по меньшей мере, одну стадию очистки, включающую лизис клеток выделенного организма, присутствующего в комбинации. Варианты осуществления могут включать, по меньшей мере, одну стадию очистки, включающую центрифугирование клеток или лизатов клеток выделенного организма, присутствующего в комбинации. Варианты осуществления могут включать, по меньшей мере, одну стадию очистки, включающую осаждение одного или нескольких соединений, присутствующих в комбинации. Варианты осуществления могут включать, по меньшей мере, одну стадию очистки, включающую фильтрацию одного или нескольких соединений, присутствующих в комбинации. Варианты осуществления могут включать, по меньшей мере, одну стадию очистки, включающую анализ с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) одного или нескольких соединений, присутствующих в комбинации.
В настоящем описании показаны разнообразные вариации указанных способов, которые могут использоваться для получения симвастатина и хувастатина, и т.п. Например, в некоторых вариантах осуществления изобретения организм-хозяин продуцирует ацилтиоэфир и/или монаколин J. В альтернативном варианте ацилтиоэфир и/или монаколин J вводят в организм в качестве части способа получения симвастатина. Способы могут дополнительно включать введение экспрессионного вектора, несущего один или несколько генов A. terreus, которые, как известно, способствуют продукции симвастатина и/или хувастатина, или и т.п., такие как гены, которые кодируют SEQ ID NO: 1 или SEQ ID NO: 2, и трансформацию указанным вектором гетерологичного организма-хозяина, в котором, таким образом, экспрессируется полипептид, обладающий ацилтрансферазной активностью. В другом варианте осуществления монаколин J может быть получен из гетерологичного организма-хозяина.
Другим вариантом осуществления изобретения является способ получения симвастатина из монаколина J в организме, который экспрессирует ген ацилтрансферазы LovD, включающий совместное культивирование указанного первого организма, который экспрессирует ацилтрансферазу LovD, со вторым организмом (например в ферментационной смеси), который продуцирует ацилтиоэфир и/или монаколин J, где ацилтиоэфир взаимодействует с продуктом гена ацилтрансферазы LovD в присутствии монаколина J с получением симвастатина. Необязательно первый организм представляет собой организм, который не продуцирует ловастатин in vivo (например, E. coli, трансформированная геном ацилтрансферазы LovD). В альтернативном варианте первый организм представляет собой ловастатин-продуцирующий организм, такой как A. terreus (например, A. terreus с инактивированным геном LovF/LDKS). Способ может дополнительно включать добавление одного или нескольких экзогенных компонентов к ферментационной смеси в целях повышения продукции предшественников симвастатина, таких как монаколин J, с повышением в результате продукции симвастатина.
Способы изобретения могут дополнительно включать добавление дополнительных компонентов к ферментационной смеси в целях повышения продукции монаколина J с повышением в результате продукции симвастатина и/или хувастатина. В качестве одного из примеров осуществления данного способа компонент может представлять собой клон с геном LovE, где организм трансформируют клоном, в результате чего транслируется LovE и, таким образом, повышается уровень продукции симвастатина. В качестве другого примера осуществления данного способа компонент может представлять собой клон, содержащий фрагмент D4B гена A. terreus (номер в ATCC 98876), где организм трансформируют клоном с повышением в результате продукции монаколин J.
Еще одним вариантом осуществления изобретения является способ превращения монаколина J в симвастатин in vitro или in vivo в присутствии экзогенного ацилтиоэфира. Предпочтительно ацилтиоэфир способен проникать через мембрану клетки. Еще одним вариантом осуществления изобретения является способ превращения монаколина J в симвастатин непосредственно в организме в присутствии ацилтиоэфира, где организм продуцирует LovD. В примере осуществления изобретения организм представляет собой A. terreus с прерванным геном LDKS.
Еще одним вариантом осуществления изобретения является симвастатин в качестве продукта, полученный способом, включающим стадии взаимодействия монаколина J; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD; с последующим использованием ацилтрансферазой LovD ацильной группы, источником которой является ацилтиоэфир для региоселективного ацилирования C8-гидроксильной группы монаколина J; в результате чего в качестве продукта образуется симвастатин. Другим вариантом осуществления изобретения является хувастатин в качестве продукта, полученный способом, включающим стадии взаимодействия гидролизованного правастатина в форме тетраола; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе гидролизованного правастатина в форме тетраола в присутствии ацилтрансферазы LovD; и LovD ацилтрансферазы; с последующим региоселективным ацилированием C8-гидроксильной группы гидролизованного правастатина в форме тетраола ацилтрансферазой LovD, переносящей ацильную группу от ацилтиоэфира; в результате чего в качестве продукта образуется хувастатин.
Другим вариантом осуществления изобретения является способ получения симвастатина, включающий гидролиз ловастатина с образованием монаколина J в присутствии LovD согласно SEQ ID NO: 1 или гомолога LovD, с последующим ацилированием монаколина J, где указанный гидролиз и ацилирование приводят к получению симвастатина. Другим вариантом осуществления изобретения является способ получения хувастатина, включающий гидролиз правастатина в гидролизованный правастатин в присутствии LovD согласно SEQ ID NO: 1 или гомолога LovD, с последующим ацилированием варианта монаколина J, где указанный гидролиз и ацилирование приводят к получению хувастатина.
Варианты осуществления изобретения включают композицию веществ, используемую для получения и/или получаемую способами, раскрытыми в настоящем описании. Например, одним из вариантов осуществления изобретения является композиция веществ, включающая монаколин J; ацилтиоэфир, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD. Конкретные варианты осуществления указанных композиций веществ дополнительно включают ацилтрансферазу LovD. Конкретные варианты осуществления указанных композиций веществ дополнительно включают симвастатин. Необязательно композиция дополнительно включает выделенный организм, такой как Escherichia coli, Aspergillus terreus, Monascus ruber, Monascus purpureus, Monascus pilosus, Monascus vitreus, Monascus pubigerus, Candida cariosilognicola, Aspergillus oryzea, Doratomyces stemonitis, Paecilomyces virioti, Penicillum citrinum, Penicillin chrysogenum, Scopulariopsis brevicaulis или Trichoderma viride. В типовых вариантах осуществления организм в композиции представляет собой Aspergillus terreus или Escherichia coli, которые экспрессируют полипептид LovD SEQ ID NO:1. В одном из вариантов осуществления изобретения организм представляет собой Aspergillus terreus, который не экспрессирует полипептид LovF SEQ ID NO: 3. В другом варианте осуществления изобретения организм представляет собой Escherichia coli, которая не экспрессирует полипептид bioH SEQ ID NO: 4. В некоторых вариантах осуществления изобретения выделенный организм в композиции был трансформирован экспрессионным вектором, кодирующим полипептид LovD Aspergillus terreus SEQ ID NO: 1.
Различные ацилтиоэфиры, которые могут использоваться в композициях изобретения, раскрыты в настоящем описании. Как правило, ацилтиоэфир представляет собой бутирил-тиоэфир, N-ацетилцистаминтиоэфир или метил-тиогликолевый тиоэфир. Необязательно ацилтиоэфир включает ацильные группы с цепью средней длины (C3-C6). В некоторых вариантах осуществления изобретения ацилтиоэфир способен проходить через клеточные мембраны Escherichia coli или Aspergillus terreus, растущих в среде культивирования. Как правило, ацилтиоэфир выбран из группы, состоящей из α-диметилбутирил-S-метил-меркаптопропионата (DMB-S-MMP), диметилбутирил-S-этилмеркаптопропионата (DMB-S-EMP), а также диметилбутирил-S-метилтиогликолята (DMB-S-MTG) и диметилбутирил-S-метилмеркаптобутирата (DMB-S-MMB). В примере осуществления ацилтиоэфир является α-диметилбутирил-S-метил-меркаптопропионатом, который содержится в среде культивирования в концентрации 1 мМ-100 мМ и обычно может присутствовать в концентрации приблизительно 10, 20, 30, 40, 50, 60, 70, 80 или 90 мМ. В некоторых вариантах осуществления изобретения композиция дополнительно включает ловастатин, при этом количество симвастатина в композиции больше, чем количество ловастатина в композиции.
Как подробно рассматривается ниже, способы и материалы изобретения, которые используются для получения симвастатина, могут быть изменены с целью получения соединений, которые структурно подобны симвастатину, например хувастатина. В данном контексте одним из вариантов осуществления изобретения является способ получения хувастатина, включающий стадии взаимодействия гидролизованного правастатина в форме тетраола; ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе гидролизованного правастатина в форме тетраола в присутствии ацилтрансферазы LovD; и LovD ацилтрансферазы; с последующим региоселективным ацилированием C8-гидроксильной группы гидролизованного правастатина в форме тетраола ацилтрансферазой LovD, переносящей ацильную группу от ацилтиоэфира; в результате чего в качестве продукта образуется хувастатин. Другим вариантом осуществления изобретения является композиция веществ, включающая: гидролизованный правастатин в форме тетраола; ацилтиоэфир, который отдает ацильную группу C8-гидроксильной группе гидролизованного правастатина в форме тетраола в присутствии ацилтрансферазы LovD; ацилтрансфераза LovD; и хувастатин.
Еще одим вариантом осуществления изобретения является композиция веществ, включающая один или несколько предшественников хувастатина, например гидролизованный правастатин в форме тетраола или 6-гидрокси-6-диметилмонаколина J в присутствии тиоэфира, выбранного по его способности ацилировать C8-гидроксильную группу монаколина J или варианта монаколина J, такого как гидролизованный правастатин в форме тетраола. Обычно такие композиции могут дополнительно включать организм, как рассмотрено выше. Таким образом, варианты осуществления изобретения включают способы получения композиции веществ, включающей симвастатин или хувастатин, как по существу раскрыто и подтверждено примерами в настоящем описании.
В случаях, когда модифицированный организм, который продуцирует симвастатин (например, A. terreus с прерванным геном LDKS), также продуцирует ловастатин (например, некоторое минимальное остаточное количество), способы и материалы, раскрытые в настоящем описании, обеспечивают модификацию биохимических путей/процессов в организме таким образом, чтобы симвастатин или родственное соединение, такое как хувастатин, являлось основным продуктом указанных путей. Другим вариантом осуществления является композиция веществ, включающая организм и симвастатин и/или хувастатин, продуцированный указанным организмом, где количество симвастатина и/или хувастатина в композиции больше, чем количество ловастатина в композиции.
Другим вариантом осуществления изобретения является композиция веществ, включающая LovD, гидролизированный правастатин и ацилтиоэфир. В примерах осуществления композиция веществ включает E. coli, которая продуцирует хувастатин. В других вариантах осуществления композиция веществ представляет собой A. terreus (например A. terreus с прерванным геном LDKS), который продуцирует хувастатин. В случаях, когда модифицированный организм, который продуцирует хувастатин (например, A. terreus с прерванным геном LDKS), также продуцирует ловастатин (например, некоторое минимальное остаточное количество), способы и материалы, раскрытые в настоящем описании, обеспечивают модификацию биохимических путей/процессов в организме таким образом, чтобы хувастатин являлся основным продуктом указанных путей. Другим вариантом осуществления является композиция веществ, включающая организм и хувастатин, продуцированный указанным организмом, где количество хувастатина в композиции больше, чем количество ловастатина в композиции.
В некоторых вариантах осуществления изобретения дополнительные компоненты и/или методологические стадии могут быть объединены с одним или несколькими способами и материалами, рассмотренными выше. Например, способы могут дополнительно включать использование ферментации при высокой плотности культуры с целью повышения эффективной концентрации ацилтрансферазы LovD и оптимизации условий ферментации и/или повышения каталитической активности ацилтрансферазы LovD по отношению к одному или нескольким ацилтиоэфирам посредством белковой инженерии. Много других компонентов или способов может использоваться для повышения продукции симвастатина или промежуточного соединения, что способствует продукции симвастатина.
Как указано выше, синтез ловастатина (который требует активности ацилтрансферазы LovD) наблюдается в ряде организмов, таких как Aspergillus terreus, Monascus ruber, Monascus purpureus, Monascus pilosus, Monascus vitreus, Monascus pubigerus, Candida cariosilognicola, Aspergillus oryzea, Doratomyces stemonitis, Paecilomyces virioti, Penicillum citrinum, Penicillin chrysogenum, Scopulariopsis brevicaulis или Trichoderma viride. Ввиду сущности изобретения, раскрытого в настоящем описании, применяя известные способы и материалы в комбинации с приводимым настоящим описанием, специалист в данной области техники может объединить культуру одного или нескольких указанных организмов (или любого другого организма, как известно, продуцирующего ловастатин) с монаколином J и ацилтиоэфиром, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD (например, α-диметилбутирил-SNAC) с целью применения ацилтрансферазы LovD при получении симвастатина. Например, как показано в настоящем описании, сущность настоящего изобретения позволяет специалисту получить симвастатин и родственные соединения в большом разнообразии ловастатин-продуцирующих организмов, известных в уровне техники, без какого-либо конкретного знания относительно характеристик конкретной ацилтрансферазы LovD, экспрессируемой в указанных организмах.
В примере методики получения симвастатина в новом организме (например, в грибе родственного A. terreus вида) и/или исследования организма на предмет соответствующей способности, первая стадия заключается в приготовлении смеси монаколина J и ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе монаколина J. Затем во второй стадии полученную смесь объединяют с организмом (например, ловастатин-продуцирующим организмом) в ферментационной смеси и выращивают культуру. Далее, в третьей стадии смесь анализируют на присутствие симвастатина, например, с использованием анализа ВЭЖХ, как описано в примерах ниже. Ввиду высокой производительности методов скрининга, известных в данной области техники (см., например, Kittel et al., Metab. Eng. 2005, 7(1): 53-58, включенное в настоящее описание посредством ссылки), подобные способы могут выполняться при большом количестве анализируемых проб, например, огромном числе грибковых культур, известных в уровне техники и легкодоступных для специалиста, что позволяет легко определять, как и какой вид организмов экспрессирует полипептиды, которые обладают активностью ацилтрансферазы LovD, что позволяет использовать их в вариантах осуществления изобретения, раскрытого в настоящем описании.
Способы, раскрытые в настоящем описании, также позволяют специалистам выделить и клонировать другие варианты ацилтрансферазы LovD, например, такие варианты, которые могут обладать дополнительными желательными качествами, такими как повышенная стабильность при различных условиях реакции и/или благоприятная ферментативная кинетика при различных условиях реакции. В данном контексте другим вариантом осуществления изобретения является способ идентификации других вариантов ацилтрансферазы LovD согласно изобретению, причем указанный способ включает простые стадияи объединения организма, способного экспрессировать ацилтрансферазу LovD (например, ловастатин-продуцирующего организма) или клеточного экстракта из указанного организма с ацилтиоэфиром, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD (например, α-диметилбутирил-SNAC) с последующим анализом указанной комбинации на присутствие симвастатина. Присутствие симвастатина является показателем присутствия нужной активности ацилтрансферазы LovD. Ввиду высокой производительности методов скрининга, известных в данной области техники (см., например, Kittel et al., Metab. Eng. 2005, 7(1): 53-58, включенное в настоящее описание посредством ссылки), подобные способы могут выполняться при большом количестве анализируемых проб, например, огромном числе грибковых культур, известных в уровне техники и легкодоступных для специалиста (например, ловастатин-продуцирующие культуры), при минимальном объеме экспериментальной работы. Таким образом, большое количество ловастатин-продуцирующих организмов, известных в уровне техники, высокая производительность методов скрининга, известных в данной области техники, и настоящее описание позволяют специалисту легко определить, как и какой вид полипептидов обладает активностью ацилтрансферазы LovD, что позволяет использовать их в вариантах осуществления изобретения, раскрытого в настоящем описании.
При обнаружении активности ацилтрансферазы LovD с помощью указанных способов можно применить стандартные в данной области техники способы клонирования гена, кодирующего белок, обладающий нужной активностью. В альтернативном варианте затем может быть проскринирована библиотека генов организма, используя полинуклеотидную последовательность ацилтрансферазы LovD (например, из A. terreus), либо отдельно, либо в комбинации с функциональными исследованиями, как рассмотрено выше, с целью идентифицировать новую полинуклеотидную последовательность ацилтрансферазы LovD, обладающую гомологией с SEQ ID NO: 1 или SEQ ID NO: 2. В данном контексте гомологичные LovD ферменты могут быть обнаружены, например, помимо прочего, в кластерах поликетидных генов грибов. Например, Mlc в пути биосинтеза компактина включен в катализ аналогичной реакции переацилирования (см., например, Y. Abe, T. et. al., Mol Genet Genomics. 2002, 267, 636-646), тогда как ацилтрансфераза в сквалестатиновом пути может катализировать подобную реакцию между ACP-связывающим тетракетидтиоэфиром и агликоном (см., например, R. J. Cox, F. et. al., Chem Commun (Camb) 2004, 20, 2260-2261). Аминокислотная последовательность LovD напоминает β-лактамазы типа C, которые катализируют гидролитическую инактивацию β-лактамных антибиотиков (см., например, E. Lobkovsky, E. M. et. al., Biochemistry, 1994, 33, 6762-6772 и A. Dubus, D. et. al., Biochem. J. 1993, 292, 537-543). Выравнивание полипептида LovD A. terreus (например, SEQ ID NO: 1) с лактамазой P99 из Enterobacter cloacae (см., например, S. D. Goldberg, et. al., Protein Sci. 2003, 12, 1633-1645) показало среднюю гомологичность последовательности, включая потенциально консервативные остатки активного центра, например каталитические Ser76, Lys79, Tyr188 и Lys315 (см., например, S. D. Goldberg, et. al., Protein Sci. 2003, 12, 1633-1645). Ввиду указанных способов еще одним вариантом осуществления изобретения является выделенный полипептид ацилтрансферазы LovD, обладающий, по меньшей мере, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологии последовательности с аминокислотной последовательностью, приведенной в SEQ ID NO:1, где полипептид обладает способностью ацилировать C8-гидроксильную группу монаколина J в присутствии соответствующего ацилтиоэфирного донора (например, α-диметилбутирил-SNAC).
Как раскрыто в настоящем описании, "ацилтиоэфир для региоселективного ацилирования C8-гидроксильной группы монаколина J" является соединением, содержащим ацильную группу, которая может быть перенесена на монаколин J или родственное соединение с получением симвастатина или родственного соединения, как раскрыто в настоящем описании. В уровне техники известно большое разнообразие подобных агентов, которые, как показано далее в настоящем описании, обладают указанной активностью (см. примеры ацилтиоэфиров в Таблице 1). В дополнение к известным в уровне техники и, как показано далее в настоящем описании, обладающим указанной активностью, любой потенциальный донор/переносчик ацила, известный в уровне техники (или синтезированный de novo), обладающий способностью ацилировать монаколин J в C8 положении с получением симвастатина, может быть легко идентифицирован с помощью сравнительных экспериментов с донорами ацила, раскрытыми в настоящем описании (например, ацил-SNAC). Обычно в таких экспериментах ацилтиоэфир представляет собой бутирил-тиоэфир, N-ацетилцистаминтиоэфир или метилтиогликолевый тиоэфир. Необязательно ацилтиоэфир включает ацильные группы с цепью средней длины (C3-C6). В некоторых вариантах осуществления изобретения ацилтиоэфир способен проходить через клеточные мембраны клеток Escherichia coli или Aspergillus terreus, растущие в среде культивирования. Как правило, ацилтиоэфир выбирается из группы, состоящей из α-диметилбутирил-S-метил-меркаптопропионата (DMB-S-MMP), диметилбутирил-S-этил-меркаптопропионата (DMB-S-EMP) и диметилбутирил-S-метилтиогликолята (DMB-S-MTG), и диметилбутирил-S-метилмеркаптобутирата (DMB-S-MMB).
Как показано настоящим описанием, сущность настоящего изобретения позволяет специалисту легко идентифицировать соединение как "ацилтиоэфир для региоселективного ацилирования C8-гидроксильной группы монаколина J" с минимальным объемом экспериментальной работы. В одном из примеров методики для идентификации соединения как "ацилтиоэфир для региоселективного ацилирования C8-гидроксильной группы монаколина J" первая стадия включает приготовление смеси монаколина J и A. terreus. Затем во второй стадии полученную смесь объединяют с анализируемым соединением в ферментационной смеси и выращивают культуру. В третьей стадии смесь анализируют на присутствие симвастатина, например, с использованием анализа ВЭЖХ, как рассмотрено в примерах ниже, где присутствие симвастатина указывает на то, что соединение обладает указанной активностью. Ввиду высокой производительности методов скрининга, известных в данной области техники (см., например, Kittel et al., Metab. Eng. 2005, 7(1): 53-58, включенное в настоящее описание посредством ссылки), подобные способы могут выполняться при большом количестве анализируемых проб, в результате чего можно легко определить, как и какой вид соединений-доноров ацильной группы обладает активностью ацилтрансферазы LovD, что позволяет использовать их в вариантах осуществления изобретения, раскрытого в настоящем описании.
Варианты осуществления настоящего изобретения дополнительно включают изделия и/или наборы, разработанные в целях облегчения осуществления способов изобретения. Обычно такие наборы включают инструкции по использованию элементов набора согласно способам настоящего изобретения. Такие наборы могут включать поддерживающее приспособление, разделенное на ячейки, плотно удерживающее одну или несколько емкостей, таких как флаконы, пробирки и т.п., при этом каждая емкость включает один из отдельных элементов, используемых в способе. Например, одна из емкостей может включать флакон, содержащий, например, ацилтрансферазу LovD и/или А. terreus, при этом другой флакон содержит ацил-SNAC соединение или подобное, оба из которых могут быть добавлены к ферментационной смеси в целях получения симвастатина.
В типовом варианте осуществления изобретения обеспечивается изделие, содержащее материалы, применяемые для производства симвастатина. Изделие включает контейнер и этикетку. Подходящие контейнеры включают, например, бутылочки, флаконы, шприцы и пробирки. Контейнеры могут быть изготовлены из разнообразных материалов, таких как стекло или пластик. Контейнер может содержать композицию веществ (например, переносчик ацильной группы и организм), из которой, например, можно получить симвастатин. Этикетка на контейнере, или имеющая отношение к контейнеру, указывает, что композиция используется для того, чтобы исследовать клеточные полипептиды. Изделие может дополнительно включать второй контейнер, включающий, например, другое соединение или субстрат для добавления к ферментационной смеси. Указанное соединение или субстрат, например, могут использоваться для повышения продукции некоторых промежуточных соединений в синтезе симвастатина, таких как монаколин J. Изделие может дополнительно включать другие материалы, необходимые в технических целях, а также в потребностях потребителя, включающие другие буферы, разбавители, фильтры, иглы, шприцы и вложенные пакеты с инструкциями по применению.
Дополнительные биологические аспекты изобретения рассмотрены в нижеследующих разделах.
БИОХИМИЧЕСКИЕ АСПЕКТЫ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Понимание некоторых аспектов изобретения облегчается описанием биохимических аспектов изобретения. В следующих разделах и примерах описания ацилтрансфераза LovD представляет собой полипептид LovD A. terreus, приведенный в SEQ ID NO: 1, и обычно упоминается как "LovD" или "фермент LovD".
Была исследована неспецифичность фермента LovD в отношении альтернативных доноров ацильной группы. Авторы настоящего изобретения обнаружили, что переносчик/донор ацильной группы не должен быть обязательно присоединен к LDKS. Подходящими являются альтернативные тиол-содержащие переносчики, включая ацил-CoA (кофермент A) и, что более важно, проникающий через мембрану ацил-SNAC (Фигура 7). Авторы настоящего изобретения обнаружили, что LovD не должен обязательно взаимодействовать с LovF и может акцептировать ацильную группу от различных доноров. Авторы настоящего изобретения также обнаружили, что LovD может переносить различные ацильные субстраты к C8-гидроксильной группе монаколина J с получением ряда аналогов ловастатина. Авторы настоящего изобретения обнаружили, что LovD может региоселективно ацилировать гидроксильную группу в C8 положении монаколина J с различными ацильными субстратами. Среди эффективных ацильных групп находится α-диметилбутирил-SNAC, который приводит к получению симвастатина в присутствии LovD и монаколина J. Также авторы настоящего изобретения обнаружили, что активность LovD при переацилировании подтверждается in vitro и может быть воссоздана в гетерологичном организме-хозяине. Авторы настоящего изобретения обнаружили, что LovD непосредственно ацилирует монаколин J α-диметилбутиратом с получением фармацевтически важного симвастатина. α-Диметилбутирил-SNAC проникает в клетку. Способность α-диметилбутирил-SNAC проникать в клетку имеет важное значение: соединение может поступать в качестве предшественника in vivo в организм, например прокариот или эукариот, экспрессирующий LovD. Прокариот или эукариот при культивировании в присутствии монаколина J (синтезируемого эндогенно или поступающего из среды культивирования) могут непосредственно продуцировать симвастатин. E. coli, например, исследовали в качестве микроорганизма-хозяина для биоконверсии монаколина J в симвастатин. И монаколин J, и диметилбутирил-SNAC добавляли к растущей культуре штамма E. coli с оверэкспрессией фермента LovD (Фигура 10). Симвастатин выделяли из среды культивирования с хорошим выходом. Данная методика может использоваться в природном продуценте ловастатина Aspergillus terreus. Штамм, который является дефицитным по LDKS, может быть сконструирован так, что он не способен синтезировать 2-метилбутиратную боковую цепь. α-Диметилбутирил-SNAC может быть добавлен к среде культивирования. Симвастатин может синтезироваться в ходе данной единственной стадии культивирования.
Как отмечено выше, LovD может катализировать последнюю стадию переноса ацила в процессе биосинтеза ловастатина и может региоспецифически ацилировать C8-гидроксильную группу монаколина J. LovD может проявлять широкую субстратную специфичность в отношении декалинагликона, переносчика ацила и ацильной группы. При добавлении синтетического субстрата α-диметилбутирил-SNAC LovD может продуцировать фармацевтически важный симвастатин in vitro и in vivo. Когда α-диметилбутирилтиоэфирный предшественник добавляется к LovF-дефицитному штамму A. terreus, путь биосинтеза ловастатина может быть перенаправлен с непосредственным синтезом в результате симвастатина. Настоящее изобретение обеспечивает: 1) микробное превращение монаколина J в симвастатин (Фигура 10); 2) мовместное культивирование оверэкспрессирующего штамма LovD со штаммом, продуцирующим монаколин J; и 3) единственную стадию культивирования Aspergillus terreus (ΔLDKS) с получением симвастатина. В каждом случае ацильный субстрат α-диметилбутирил-SNAC может быть синтезирован химически и добавлен в среду культивирования.
LOVD НЕ ДОЛЖЕН ВЗАИМОДЕЙСТВОВАТЬ С LOVF, А LOVD МОЖЕТ КАТАЛИЗИРОВАТЬ РЕАКЦИЮ ПЕРЕАЦИЛИРОВАНИЯ АЦИЛ-COA И ДРУГОГО АЛЬТЕРНАТИВНОГО ТИОЛСОДЕРЖАЩЕГО ПЕРЕНОСЧИКА (ФИГУРА 5)
Вначале авторы настоящего изобретения исследовали, может ли ацил-CoA использоваться в качестве субстрата в реакции переацилирования. Непрерванный ген LovD амплифицировали из геномной ДНК A. terreus с использованием сплайсинга посредством ПЦР с удлинением перекрывающихся фрагментов и встраивали в экспрессионный вектор pET28a. Оверэкспрессию растворимого, слитого с N-концевым 6×His-тагом LovD выполняли в штамме E. coli BL21(DE3)/pAW31. LovD очищали, используя аффинную колонку Nickel-NTA, с получением гомогенного вещества (конечный выход ~40 мг/л). Субстрат монаколин J (1 мм) получали путем гидролиза ловастатина LiOH и добавляли к реакционной смеси, содержащей чистый LovD (10 мкМ) и бутирил-CoA (4 мМ), который является коммерческим ацил-CoA, наиболее схожим с природной α-метилбутиратной боковой цепью. Реакционную смесь инкубировали при комнатной температуре, экстрагировали этилацетатом и анализировали с помощью ВЭЖХ и ЖХ/МС (Фигура 5).
В смеси реакции было образовано одно, более гидрофобное соединение с идентичным ловастатину поглощением в УФ-диапазоне, при одновременном исчезновении монаколина J (Фигура 5, график a). Масса нового соединения, как установили, составляла 391 (M+H), что соответствует присоединению бутирильной группы к монаколину J. Селективную этерификацию C8-гидроксильной группы монаколина J с получением 4 подтверждали протонной ЯМР-спектроскопии (CDCl3, 500 МГц). Протонный ЯМР-спектр 4 (лактонная форма) практически идентичен спектру ловастатина за исключением сигналов алифатических атомов линейной ацильной боковой цепи. Характерный мультиплет H8 (определенный с помощью 1H-1H COSY, 1H-13C HMQC и 1H-13C HMBC) в 4 сдвинут в сторону слабого поля до δ 5,38, по сравнению с δ 4,23, наблюдаемой для того же протона в монаколине J. Это соответствует эффекту деэкранирования ацилокси замещения по C8. Протоны H11 и H13 углеродов, несущие другие гидроксильные группы, были сдвинуты с δ 4,71 и δ 4,38 в монаколине J до δ 4,51 и δ 4,36 в 4, соответственно.
Как показано на Фигуре 5A (график a), 87% превращение монаколина J в 4 наблюдали через 10 часов, используя в качестве донора ацильной группы бутирил-CoA. Исследование в зависимости от времени дало кажущееся число оборотов 0,18 мин-1 (Фигура 5B). Наблюдаемая степень превращения 87%, по-видимому, приближается к равновесию, как видно из Фигуры 5B. Чтобы исследовать биохимическую природу равновесного превращения, авторы настоящего изобретения оценивали, катализирует ли LovD также обратную реакцию гидролиза. Действительно, когда ловастатин использовали в качестве единственного субстрата, образование монаколина J четко детектировали со значением kcat и Км 0,21±0,01 мин-1 и 0,56±0,05 мМ, соответственно (Фигура 5C). Когда в предполагаемом активном сайте LovD серин (Ser76) заменяли на аланин, образование 4 или гидролиз ловастатина не наблюдали (Фигура 5A, график b).
Ферментативный синтез 4 подтверждает, что LovD действительно катализирует реакцию переноса ацильной группы, показанный на Фигуре 4. Кроме того, полученный результат показывает, что прямая взаимосвязь между доменами LovF и LovD не требуется для каталитического оборота, в противоположность ранее предполагаемому механизму катализа LovD (см., например, J. Kennedy, K. et. al., Science, 1999, 284, 1368-1372 и C. R. Hutchinson, J. et. al., Antonie Van Leeuwenboek 2000, 78, 287-295). Ацил-S-CoA может заменять ацил-S-LovF, хотя вероятно со значительно более высоким значением Kм, из-за потери потенциальных белок-белковых взаимодействий.
ЭКСПЕРИМЕНТЫ ПО ПЕРЕАЦИЛИРОВАНИЮ С НЕСКОЛЬКИМИ РАЗЛИЧНЫМИ КОММЕРЧЕСКИМИ АЦИЛЬНЫМИ ЗАМЕСТИТЕЛЯМИ ВЫЯВИЛИ ПРЕДПОЧТЕНИЕ LOVD К АЦИЛЬНЫМ ГРУППАМ С ЦЕПЬЮ СРЕДНЕЙ ДЛИНЫ (C3-C6)
Авторы настоящего изобретения оценивали чувствительность LovD к различным ацильным заместителям, проводя эксперименты по переацилированию с различными коммерческими ацил-CoAs (Таблица 1A). Все эксперименты выполняли при 1 мМ монаколина J, 4 мМ ацил-CoA и 10 мкМ LovD в течение 10 часов. Полученные результаты четко указывают, что LovD проявляет предпочтение к ацильным группам с цепью средней длины (C3-C6), причем оптимальным алкилацил-CoA субстратом является бутирил-CoA. Неожиданно более объемный бензоил-CoA являлся одним из лучших исследованных ацильных субстратов со степенью превращения симвастатина в соответствующий 8-бензокси-аналог ловастатина около 70% (кажущаяся kcat=0,16 мин-1). Введение α-β ненасыщенности значительно снижало скорость реакции, что подтверждалось 6% ацилирования монаколина J в присутствии кротонил-CoA. Ацетоацетил-CoA и β-гидроксибутирил-CoA являлись превосходными субстратами для LovD, что хорошо соответствовало выделению монаколина X (см., например, A. Endo, K. et. al., J. Antibiot., 1986, 38, 321-327) и монаколина М. (см., например, A. Endo, D. et. al., J. Antibiot. 1986, 39, 1670-1673) из природного организма-хозяина, соответственно. Из проанализированных субстратов CoA LovD была неактивна в отношении малонил- и пальмитоил-CoA.
ДАЛЬНЕЙШИЕ ЭКСПЕРИМЕНТЫ ПО ПЕРЕАЦИЛИРОВАНИЮ С АЛЬТЕРНАТИВНЫМ ПЕРЕНОСЧИКОМ АЦИЛА, АЦИЛ-SNAC, КОТОРЫЙ ПРОЩЕ ПОЛУЧАТЬ СИНТЕТИЧЕСКИ И КОТОРЫЙ МОЖЕТ ПРОНИКАТЬ ЧЕРЕЗ КЛЕТОЧНУЮ МЕМБРАНУ IN VIVO, ВЫЯВИЛИ, ЧТО АЦИЛ-SNAC ЯВЛЯЕТСЯ КОМПЕТЕНТНЫМ СУБСТРАТОМ ДЛЯ LOVD
В целях дальнейшего исследования субстратной специфичности LovD в отношении альтернативных переносчиков ацильной группы, в особенности таких, которые проще получать синтетически и которые могут проникать через клеточную мембрану in vivo, авторы настоящего изобретения оценивали два варианта бутирил-тиоэфиров в качестве субстратов для LovD (Фигура 5). N-ацетилцистаминтиоэфиры (SNAC) широко использовали в качестве проб и предшественников в исследовании биохимии природных соединений (см., например, Auclair et. al., Science, 1997, 277, 367-369). Метил-тиогликолят (SMTG), как было недавно показано, является экономически выгодным заменителем SNAC в направленном биосинтезе эритромицина из предшественника (см., например, S. Murli, K. S. et. al., Appl. Environ. Microbiol. 2005, 71, 4503- 4509). На Фигуре 5 (графики c и d) показано превращение монаколина J в 4, когда в качестве доноров ацила использовались указанные бутирил-тиоэфиры. Эффективное использование тиоэфиров SNAC и SMTG вместо бутирил-CoA, с кажущимися значениями kcat 0,09 мин-1 и 0,23 мин-1 (Фигура 5B), соответственно, дополнительно подтверждает, что белок-белковые взаимодействия между LovD и LovF, так же как и взаимодействие между LovD и фосфопантетеиновой субъединицей, не требуются для переноса ацила. Впрочем, бутирил-тиоэтан не являлся компетентным субстратом для LovD и способствовал только 4% превращения монаколина J в 4. Аналогичным образом бензоил-SNAC и бензоил-SMTG эффективно использовали вместо бензил-CoA, с кажущимися значениями kcat 0,12 мин-1 и 0,15 мин-1, соответственно (Таблица 1A).
ЭКСПЕРИМЕНТ ПО IN VITRO ХЕМОЭНЗИМАТИЧЕСКОМУ СИНТЕЗУ ЛОВАСТАТИНА И СИМВАСТАТИНА С ИСПОЛЬЗОВАНИЕМ α-S-МЕТИЛБУТИРИЛ-SNAC И α-ДИМЕТИЛБУТИРИЛ-SNAC ПОКАЗАЛ, ЧТО МОНАКОЛИН J ПРЕВРАЩАЕТСЯ В СИМВАСТАТИН В ПРИСУТСТВИИ АЦИЛ-SNAC
Далее авторы настоящего изобретения синтезировали α-S-метилбутирил-SNAC и α-диметилбутирил-SNAC и оценивали их в in vitro хемоэнзиматическом синтезе ловастатина и симвастатина, соответственно. Результаты показаны на фигуре 8 и в Таблице 1A. Стандартные пробы ловастатина и симвастатина использовали в качестве контроля для ВЭЖХ-детектирования (Фигура 8, график a). Природная, α-S-метилбутиратная боковая цепь неожиданно оказалась более плохим субстратом по сравнению с бутирил-, пентаноил- и гексаноил-SNAC. Кажущаяся kcat (~0,04 мин-1) в синтезе ловастатина оказалась более чем на 50% меньше, чем kcat LovD в отношении бутирил-SNAC. Предложили, что дикий тип LovD не был оптимизирован для переноса разветвленного субстрата. Введение второго метильного заместителя в α-положение дополнительно снижало скорость ацилирования, что по-видимому обусловлено возросшим стерическим затруднением диметильной группы. В стандартных условиях эксперимента приблизительно 10% монаколина J было превращено в симвастатин при использовании в качестве субстрата α-диметилбутирил-SNAC (кажущаяся kcat=0,02 мин-1). Можно было достичь степени равновесного превращения >70%, когда 100 мкМ LovD и 10-кратный избыток α-диметил-SNAC или α-диметил-SMTG добавляли к реакционной смеси in vitro (Фигура 8, графики b и c).
LOVD IN VITRO МОЖЕТ ПЕРЕНОСИТЬ РАЗЛИЧНЫЕ АЦИЛСУБСТРАТЫ (НЕ ТОЛЬКО LDKS) К C-8-ГИДРОКСИЛЬНОЙ ГРУППЕ МОНАКОЛИНА J С ПОЛУЧЕНИЕМ РЯДА АНАЛОГОВ ЛОВАСТАТИНА
Для оценки субстратной специфичности LovD к вариантам монаколина J исследовали превращение тетраола 7 в 8, правастатина (5) и хувастатина (6) (Фигура 9). Ранее было показано, что когда 8-диметил-монаколин J добавляется к мутантному A. terreus с заблокированным биосинтезом монаколина J, может быть выделен компактин (см., например, J. L. Sorensen, K. et. al., Org. Biomol. Chem. 2003, 1, 50-59). Это указывает, что LovD может быть нечувствителен к замещению в C6 положении декалинового ядра. Действительно, LovD показывает слабую специфичность к замещению гидроксильной группы в C6 положении и катализирует ацилирование 7 с более высокой эффективностью. Кажущееся число оборотов в синтезе 8, правастатина или хувастатина с использованием соответствующих ацил-тиоэфиров, составляло 0,18, 0,11, 0,03 мин-1, соответственно. Время удержания нативного правастатина было идентично энзиматически синтезированному соединению. Масса вновь образованных соединений проверяли с помощью ЖХ/МС. В реакционной смеси каких-либо ди-ацилированных продуктов не обнаруживали.
ДАННЫЕ IN VIVO ПОКАЗАЛИ, ЧТО LOVD МОЖЕТ ИСПОЛЬЗОВАТЬСЯ ДЛЯ ПРЕПАРАТИВНОГО БИОСИНТЕЗА АНАЛОГОВ ЛОВАСТАТИНА
С целью демонстрации того, что LovD может использоваться для препаративного биосинтеза аналогов ловастатина, сначала предприняли попытку провести реакцию бензилирования in vivo, используя E. coli в качестве гетерологичного организма-хозяина. Оверэкспрессирующий штамм BL21(DE3)/pAW31 выращивали во встряхиваемой колбе (200 мл) до OD600 1,0, после чего в культуру добавляли 1 мМ ИПТГ, 0,8 мМ монаколина J и 4 мМ бензоил-SNAC или бензоил-SMTG. Экспрессию LovD и биоконверсию проводили при 18°C. Культуру экстрагировали и анализировали на предмет образования 8-бензоил-монаколина J. При добавлении бензоил-SNAC наблюдали 84% превращение монаколина J в течение 20 часов после индукции. Продукт подвергали лактонизации и очищали в одну стадию с помощью хроматографии на силикагеле. Спектры ЯМР подтверждали региоселективное бензоилирование C8-гидроксильной группы, так как характерный мультиплет H8 сдвигался в слабое поле до δ 5,61. Напротив, для культуры, в которую добавляли бензоил-SMTG, наблюдали только 40% бензоилирования. Бензоил-SMTG был быстро расщеплен E. coli, и по истечении 24 часов в среде культуры его следов не обнаруживали.
АЦИЛЬНЫЙ СУБСТРАТ α-ДИМЕТИЛБУТИРИЛ-SNAC ПРИВЕЛ К ПОЛУЧЕНИЮ СИМВАСТАТИНА В ПРИСУТСТВИИ ОЧИЩЕННОЙ LOVD И МОНАКОЛИНА J
Ферментацию проводили при низкой плотности культуры с α-диметилбутирил-SNAC в качестве предшественника с получением симвастатина из монаколина J в одной стадии биосинтеза. После двух дней культивирования наблюдали превращение ~90% монаколина J в симвастатин (см., например, приведенные ниже "условия ферментации"). Продукт очищали, причем протонные и углеродные спектры ЯМР были идентичны спектрам коммерческого соединения. Более низкий выход симвастатина соответствовал с более низким числом оборотов, наблюдаемым in vitro, который может быть также уменьшен при более низкой внутриклеточной концентрации предшественника SNAC. Другие факторы, например обратимый гидролиз симвастатина (наблюдали гидролиз симвастатина в присутствии LovD, при этом kcat (0,02 мин-1), гидролиза ~10 раз ниже, чем при гидролизе ловастатина, показано на фигуре 5C), и инактивация LovD после длительной ферментации может привести к наблюдаемому превращению. Ожидалось, что выход может быть значительно повышен за счет нескольких факторов, например: 1) использования ферментации при высокой плотности культуры с целью увеличения эффективной концентрации LovD и оптимизации условий ферментации; 2) увеличения каталитической эффективности LovD в отношении неестественного предшественника путем белковой инженерии.
По всему тексту настоящей заявки были приведены ссылки на различные публикации. Описания указанных публикаций, таким образом, посредством ссылки включены в настоящее описание в полном объеме.
ПРИМЕРЫ
Примеры, приводимые ниже, обеспечивают примеры способов и материалов, которые могут использоваться в практическом осуществлении различных вариантов осуществления изобретения, раскрытого в настоящем описании.
ПРИМЕР 1: КЛОНИРОВАНИЕ, ЭКСПРЕССИЯ И ОЧИСТКА ПРИМЕРА АЦИЛТРАНСФЕРАЗЫ LOVD
Три экзона LovD A. terreus отдельно амплифицировали из геномной ДНК A. terreus и соединяли с получением непрерывной открытой рамки считывания с использованием сплайсинга посредством ПЦР с удлинением перекрывающихся фрагментов. Сайты рестрикции NdeI и HindIII вводили в 5' и 3' фланкирующие праймеры, соответственно. Кассету гена лигировали в pET28 (Novagen) с получением экспрессионной конструкции pAW31. Штамм E. coli BL21(DE3), трансформированный pAW31, выращивали в среде LB при 37°C до OD600 0,5, после чего к культуре добавляли 1 мМ ИПТГ и проводили экспрессию при 18°C в течение 24 часов. Клетки собирали центрифугированием, ресуспендировали в Буфере А (50 мМ Трис, pH 8,0, 2 мМ DTT, 2 мМ EDTA) и лизировали ультразвуком. Клеточный дебрис и нерастворимые белки удаляли с помощью центрифугирования (17000g, 4°C, 1 час). К осветленному лизату добавляли 2 мл смолы Ni-NTA (Qiagen). LovD очищали, используя ступенчатый градиент буфера А с растущей концентрацией имидазола. Чистые (>95%) белки LovD элюировали в буфере А, содержащем 250 мМ имидазола, меняли буфер на буфер А без имидазола, концентрировали, разливали на аликвоты и быстро замораживали. Замороженные аликвоты LovD использовали однократно. Наблюдали существенное снижение активности фермента после повторных циклов размораживания-замораживания.
ПРИМЕР 2: ПРИМЕРЫ УСЛОВИЙ ФЕРМЕНТАЦИИ
Условия ферментации: 500 мл культуры на среде LB с 30 мг/л канамицина. При OD600 1,0 клетки концентрировали до конечной плотности OD600 5,0 и индуцировали 1 мМ ИПТГ. Субстраты монаколин J и α-диметилбутирил-SNAC добавляли до конечной концентрации 1 мМ и 4 мМ. В различные моменты времени отбирали пробы культуры, центрифугировали, фильтровали и вводили в ВЭЖХ (20 мкл). Условия экстракции: после достижения максимальной степени превращения среду подкисляли до pH 2,0, экстрагировали этилацетатом, сушили и повторно растворяли в толуоле. Лактоную форму монаколина J получали путем кипячения с дефлегматором, используя аппарат Сокслета, как было описано ранее (см., например, J. L. Sorensen, K. et. al., Org. Biomol. Chem. 2003, 1, 50-59).
Различные микробиологические среды, такие как LB, F1 или TB, известны в уровне техники и могут быть использованы или приспособлены к использованию в соответствии с вариантами осуществления изобретения, раскрытыми в настоящем описании, включая LB, F1 и TB. Другие среды, специально приспособленные для выращивания таких организмов, как A. terreus и M. Pilosus, также известны в уровне техники (см., например, Miyake et al., Biosci. Biotechnol. Biochem., 70(5): 1154-1159 (2006) и Hajjaj et al., Applied and Environmental Microbiology, 67: 2596-2602 (2001), содержание которых включено посредством ссылки).
Стандартная пропись для приготовления среды Луриа-Бертани (LB):
• 10 г Триптона
• 5 г Дрожжевого Экстракта
• 10 г NaCl
• 1 л дистиллированной воды
pH ~7,3-7,5.
Стандартная пропись для приготовления TB:
• 3 г PIPES (10 мМ)
• 2,2 г CaCl2·H2O (15 мМ)
• 18,6 г KCl (250 мМ)
• 10,9 г MnCl2 (55 мМ)
• 1 л дистиллированной воды
pH 6,7-6,8 (KOH)
Стандартную среду F1 можно найти в патенте США 5064856, содержание которого включено в настоящее описание посредством ссылки.
Обычно такие среды стерилизуют после приготовления посредством фильтрации или автоклавирования.
ПРИМЕР 3: УЛУЧШЕНИЕ БИОКОНВЕРСИИ СИМВАСТАТИНА В ESCHERICHIA COLI ПОСРЕДСТВОМ ДЕЛЕТИРОВАНИЯ BIOH
Данный пример дополнительно характеризует полипептид LovD, кодируемый кластером генов ловастатина (см. также Xie et. al., (2006) Chem. Biol. 13: 1161-1169). LovD катализирует последнюю стадию биосинтеза ловастатина и отвечает за перенос 2-метилбутиратной боковой цепи от мегасинтазы LovF к непосредственному биосинтетическому предшественнику, монаколину J (MJ) в кислотной форме (см., например, Kennedy et. al., (1999) Science 284: 1368-1372). Здесь было показано, что LovD обладает широкой субстратной специфичностью в отношении декалинового ядра, тиоэфирной ацильной единицы и тиоэфирного переносчика ацила. Используя штамм Escherichia coli с оверэкспрессией LovD и проникающий через клеточную мембрану тиоэфир диметилбутирил-S-метилмеркаптопропионат (DMB-S-MMP) (Фигура 13A), авторы настоящей заявки разработали цельноклеточный биокаталитический способ, с помощью которого можно превратить кислотную форму MJ в кислотную форму симвастатина в одну стадию с высоким выходом (см., например, Xie et. al., (2007) Appl. Environ. Microbiol. 73: 2054-2060). Способ ферментации может быть конкурентоспособной альтернативой в экономическом отношении текущим путям синтеза.
Тиоэфир DMB-S-MMP представляет собой важный компонент биоконверсии симвастатина. Он является одним из наиболее каталитически эффективных доноров ацила, используемых в реакции переноса ацила, и наименее дорогим в синтезе. Одним существенным недостатком, связанным с данным соединением, является гидролиз метилсложноэфирной связи в DMB-S-MMP, который приводит к образованию диметилбутирилмеркаптопропионовой кислоты (DMB-S-MPA) (Фигура 13). Реакция гидролиза является ферментативной, поскольку в отсутствие E. coli какого-либо расщепления не наблюдается, и значительно ускоряется в процессе ферментации при высокой плотности культуры. Побочная реакция нежелательна по трем причинам: 1) гидролиз субстрата приводит к снижению количества DMB-S-MMP, доступного для LovD-катализируемого переацилирования, что требует внесения тиоэфира с высоким молярным избытком и частого добавления в ходе ферментации; 2) так как DMB-S-MPA менее эффективен по сравнению с DMB-S-MMP в качестве донора диметилбутиратной группы (~10 раз медленнее) (см., например, Xie et. al., (2007) Appl. Environ. Microbiol. 73: 2054-2060), накапливающийся более растворимый DMB-S-MPA может служить конкурентным ацильным субстратом для LovD. Это сильно снижает скорость реакции, и было продемонстрировано in vitro с использованием очищенной LovD. Таким образом, общая продолжительность биоконверсии излишне продливается; и 3) карбоксильная кислотная группа DMB-S-MPA мешает очистке симвастатина в кислотной форме из среды культивирования после завершения биоконверсии. Оба соединения выпадают в осадок при подкислении среды, в результате чего перед кристаллизацией симвастатина требуются дополнительные стадии разделения. Хотя DMB-S-MPA может быть удален путем промывания фильтрата избыточным количеством воды, значительное количество симвастатина в кислотной форме теряется в течение стадий промывания, что приводит к уменьшению общего выхода.
Исключение нежелательной побочной реакции гидролиза может, таким образом, улучшить общую экономичность цельноклеточного биокаталитического способа и последующих стадий очистки. Поэтому наиболее безотлагательной целью является точное выявление фермента (ферментов), ответственного за нежелательную побочную реакцию. В настоящей заявке описана идентификация BioH как карбоксиэстеразы E. coli, которая гидролизует DMB-S-MMP до DMB-S-MPA. Конструируя ΔbioH штамм на основе штамма с оверэкспрессией LovD, была полностью устранена конкурентная реакция и дополнительно улучшена надежность цельноклеточного биокаталитического синтеза симвастатина в кислотной форме.
МАТЕРИАЛЫ, ШТАММЫ И ПЛАЗМИДЫ
Монаколин J и DMB-S-MMP были получены, как описано ранее (см., например, Xie et. al., (2007) Appl. Environ. Microbiol 73: 2054-2060). Все реактивы были получены из стандартных источников. Штамм BL21(DE3) [F- omp hsdSB (rB- mB-) gal dcm λ(DE3)] был получен от Novagen. Коллекция Keio была получена из Национального Института Генетики, Япония (см., например, Baba et al., (2006). Mol Syst Biol. 2: 2006 0008). Мутанты с одним нокаутированным геном были получены из штамма BW25113 [rrnB3 DElacZ4787 hsdR514 DE(araBAD)567 DE(rhaBAD)568rph-1]. WA837 (rB-, mB+, gal met), штамм E. coli B без системы рестрикции и с системой модификации был получен из Coli Genetic Stock Center (CGSC) (см., например, Wood, W.B. (1966) J. Mol. Biol. 16: 118-133). Плазмиды pAW31 (kanr) и pXK8 (kanr) были получены из pET28a и содержат lovD ген из A. terreus и bioH ген из E. coli, соответственно.
ЭКСПЕРИМЕНТ ПО ЦЕЛЬНОКЛЕТОЧНОМУ ГИДРОЛИЗУ
Отобранные 57 мутантов из коллекции Keio с BW25113, BL21(DE3) и BL21(DE3)/pAW31 выращивали до насыщения в 96-луночном глубоком планшете в 1 мл среды LB при 37°C. Чистый DMB-S-MMP (5 мкл) добавляли к каждой культуре до конечной концентрации 20 мМ. После инкубирования на качалке (20°C, 300 об/мин) в течение 10 часов каждую культуру экстрагировали равным объемом этилацетата (ЭА)/1%-ной уксусной кислоты (AcOH). Органическую фазу сушили, повторно растворяли в 20 мкл ацетонитрила (CH3CN), после чего 1 мкл наносили на ТСХ-пластину (силикагель 60 F254). ТСХ-пластины элюировали в 20%-ном р-ре ЭА в гексане и проявляли йодом.
Анализ соединений также выполняли с помощью ВЭЖХ, используя аналитическую колонку С18 (Alltech Apollo 5u, 150 мм × 4,6 мм); линейный градиент: 60%-ный CH3CN в воде (0,1% р-р трифторуксусной кислоты [ТФУ]) до 95%-ного CH3CN в воде (0,1% ТФУ) в течение 5 мин, 95%-ный CH3CN в воде (0,1% ТФУ) в течение 10 мин, при расходе 1 мл/мин. Время удержания ВЭЖХ (tR) было следующим: лактонная форма MJ: 3,40 мин; DMB-S-MPA: 3,82 мин; DMB-S-MMP: 6,80 мин; лактонная форма симвастатина: 8,45 мин. И MJ, и симвастатин в кислотной форме были лактонизированы перед ВЭЖХ анализом.
ОЧИСТКА BIOH И ФЕРМЕНТАТИВНЫЙ АНАЛИЗ
Ген bioH амплифицировали из геномной ДНК E. coli с помощью ПЦР, вводя фланкирующие сайты рестрикции NdeI и EcoRI с использованием праймеров 5'-AACATATGAATAACATCTGGTGGCA-3' (SEQ ID NO: 5) и 5'-AAGAATTCTACACCCTCTGCTTCAACG-3' (SEQ ID NO: 6). Кассету гена расщепили и лигировали в pET28a с получением экспрессионной конструкции pXK8. Штамм E. coli BL21(DE3), трансформированный pXK8, выращивали в среде LB при 37°C до OD600 0,5, после чего к культуре добавляли 0,1 мМ ИПТГ и проводили экспрессию при 20°C в течение 16 часов. Клетки ресуспендировали в Буфере А (50 мМ Трис-HCl, pH 8,0, 2 мМ DTT, 2 мМ EDTA) и лизировали ультразвуком. Очистку BioH проводили с использованием N-концевого 6xHis-тага и смолы Ni-NTA (Qiagen). Практически чистый (>95%) BioH элюировали в Буфере А, содержащим 250 мМ имидазола, затем буфер заменяли на Буфер A, концентрировали, разливали на аликвоты и быстро замороживали.
Гидролизный анализ выполняли при комнатной температуре в 50 мМ HEPES, pH 7,9. Тиоэфирный субстрат DMB-S-MMP добавляли до конечной концентрации в диапазоне 0,1-1,0 мМ. С целью облегчения солюбилизации DMB-S-MMP во все пробы добавляли ДМСО до конечной концентрации 10%. Реакцию начинали добавлением BioH (0,01 мкМ) и останавливали в заданные моменты времени (10, 20, 30 мин), добавляя равный объем ЭА/1% AcOH. Органическую фазу отделяли, сушили, повторно растворяли в 20 мкл ACN и анализировали с помощью ВЭЖХ. Степени превращения DMB-S-MMP в DMB-S-MPA определяли количественно, интегрируя пики при 234 нм. Сравнение каталитической эффективности BioH в отношении трех диметилбутирилтиоэфиров выполняли при концентрации субстрата 1 мМ и концентрации BioH 10 нМ.
ТРАНСДУКЦИЯ Р1
Согласно стандартным протоколам (см., например, Miller, J.H. (1992) A short course in bacterial genetics: A laboratory manual and handbook for Escherichia coli and related bacteria. Cold Spring Harbor, NY: Cold Spring Harbor Press), трансдукцию Р1 использовали для создания ΔbioH мутанта BL21(DE3). Из-за различной системы рестрикции у штамма E. coli K (BW25113) и штамма E. coli B (BL21), штамм B WA837 (rB-, mB+) использовали в качестве промежуточного хозяина для трансдукции. Маркер ΔbioH::FRT-kan-FRT сначала трансдуцировали из JW3375 в WA837 с получением YT0 (WA837 ΔbioH::FRT-kan-FRT). Используя BL21(DE3) в качестве реципиента, а YT0 в качестве донора, был создан штамм YT1 (BL21 ΔbioH::FRT-kan-FRT λ(DE3)). Штамм YT1 трансформировали вспомогательной плазмидой pCP20, которая содержит термочувствительное начало репликации и термоиндуцируемый ген FLP (см., например, Datsenko et. al., (2000) Proc. Natl. Acad. Sci. USA 97: 6640-6645; и Wanner, 2000). Удаление маркера kan с получением YT2 (BL21 ΔbioH::FRT λ(DE3)) проводили согласно опубликованным методикам (см., например, Datsenko et. al., (2000) Proc. Natl. Acad. Sci. USA 97: 6640-6645; и Wanner, 2000).
Для проверки генетических изменений YT2 использовали ПЦР. Создавали два праймера. Праймеры В1: 5'-TGACGGCTTCGCTATCCCAT-3' (SEQ ID NO: 7); B2: 5'-TACACCCTCTGCTTCAACG-3' (SEQ ID NO: 8); и B3: 5'-GCTGGATTGTTTCGCCGATC-3'(SEQ ID NO: 9) отжигались на гене yhgA слева, на 3'-конце bioH, который оставляли интактным, и на гене gntX справа, соответственно. Ожидаемые продукты в реакции ПЦР с использованием геномной ДНК YT2 в качестве матрицы: B1/B2: 602 п.н.; B1/B3: 1002 п.н. Ожидаемые продукты в реакции ПЦР с использованием геномной ДНК BL21(DE3) в качестве матрицы: B1/B2: 1271 п.н.; B1/B3: 1771 п.н. Ожидаемые продукты ПЦР регистрировали для каждого штамма.
ЦЕЛЬНОКЛЕТОЧНЫЙ БИОКАТАЛИЗ
Цельноклеточный каталитический синтез кислотной формы симвастатина из кислотной формы MJ и DMB-S-MMP проводили, как описано (см., например, Xie et. al., (2007) Appl. Environ. Microbiol. 73: 2054-2060). Штаммы E. coli BL21(DE3)/pAW31 и YT2/pAW31 культивировали одновременно для сравнения. Одиночную колонию только что трансформированных штаммов использовали для инокуляции 5 мл среды LB с добавкой 35 мг/л канамицина и выращивали за ночь при 37°C. Следующим утром 100 мкл ночной культуры инокулировали 50 мл культур, содержащих среду LB, минимальную среду F1 и среду F1 с 0,15 мг/л биотина. Скорости роста контролировали, периодически измеряя плотность OD600. Когда OD600 достигла ~0,5, к культуре добавляли 100 мкМ ИПТГ и проводили экспрессию LovD при 20°C в течение 16 часов. Для имитации условий ферментации при высокой плотности культуры сконцентрировали в 10 раз перед добавлением субстратов. Обычно аликвоту культуры объемом 14 мл переносили в центрифужную пробирку на 15 мл, после чего клетки собирали центрифугированием (4°C, 4000g, 10 мин). Осадок клеток мягко ресуспендировали в 1316 мкл супернатанта с последующим добавлением 84 мкл стокового раствора кислотной формы MJ (250 мМ в H2O) (конечная концентрация 15 мМ). Затем сконцентрированную культуру разделяли на семь проб по 200 мкл и добавляли к каждой пробе 1,2 мкл чистого DMB-S-MMP (конечная концентрация ~25 мМ). После этого культуру качали при 300 об/мин при комнатной температуре. В каждый момент времени проводили суммарную экстракцию, добавляя 10 мкл 20% ДСН, чтобы лизировать клетки, с последующей жидкостной экстракцией 500 мкл ЭА/1% AcOH. Органическую фазу удаляли, выпаривали и повторно растворяли в 50 мкл ACN с целью анализа с помощью ВЭЖХ. В пробу BL21(DE3) через 12 часов добавляли дополнительную аликвоту (1,2 мкл) DMB-S-MMP для восполнения гидролизуемых субстратов.
ИДЕНТИФИКАЦИЯ BIOH В КАЧЕСТВЕ DMB-S-MMP-ЭСТЕРАЗЫ
Вначале авторы настоящего изобретения старались идентифицировать фермент E. coli, ответственный за наблюдаемый гидролиз DMB-S-MMP с образованием DMB-S-MPA в ходе ферментации. При использовании в качестве тиоэфирных субстратов различных переносчиков ацила, таких как диметилбутирил-S-этилмеркаптопропионат (DMB-S-EMP) и диметилбутирил-S-метилтиогликолят (DMB-S-MTG), отмечали присутствие в ферментационной среде соответствующих гидролизованных карбоновых кислот. Это дает возможность предположить, что ответственный фермент является эстеразой или гидролазой с нестрогой субстратной специфичностью в отношении функциональных сложноэфирных групп.
Для идентификации ответственного фермента применяли высокоэффективный метод, используя библиотеку мутантных штаммов E. coli K-12 с нокаутом одного гена внутри рамки считывания (коллекция Keio) (см., например, Baba et. al., (2006) Mol. Syst. Biol. 2: 2006 0008). Предполагали, что какой-либо мутантный штамм неспособен гидролизовать DMB-S-MMP непосредственно из-за делеции конкретного гена. Проверка аннотаций геномов E. coli (см., например, Blattner et. al., (1997) Science 277: 1453-1474) позволила выявить 23 эстеразы/эстеразоподобных фермента, 94 гидролазы/гидролазоподобных фермента и 16 ацилтрансфераз (всего 133 гена-кандидата). Ферменты с подтвержденной активностью и субстратами, которые, по-видимому, не будут включены в гидролиз DMB-S-MMP, не были исследованы на первом этапе анализа. Список из 57 исследованных мутантных штаммов BW25113 приведен в Таблице 2.
Мутанты, дикий тип BW21113 и BL21(DE3), выращивали до насыщения в среде LB в 96-луночном планшете. К каждой культуре (1 мл) добавили 5 мкл чистого DMB-S-MMP, после чего планшет инкубировали с энергичным встряхиванием в течение 10 дополнительных часов при комнатной температуре. Культуры подкисляли, экстрагировали этилацетатом и анализировали органическую фазу с помощью тонкослойной хроматографии (ТСХ), используя подвижную фазу (20%-ный этилацетат в гексане) с разделением в результате DMB-S-MMP и DMB-S-MPA (Фигура 14A). Дикий тип BW25113 (линия 58) и BL21 (DE3) (линия 59) показали сопоставимый уровень гидролиза субстрата и накопления DMB-S-MPA. Все исследованные мутанты показывали гидролитическую активность в отношении DMB-S-MMP за иключением ΔbioH (линия 56, штамм JW3357). Указанное неожиданное открытие позволяет предположить, что BioH (см., например, O'Regan et. al., (1989) Nucleic Acids Res 17: 8004), который вовлечен в биосинтез биотина (витамин H), может являться единственным ферментом, ответственным за наблюдаемый гидролиз in vivo. Дополнительный анализ с помощью ВЭЖХ также подтверждал, что DMB-S-MPA не обнаруживается в органическом экстракте ΔbioH мутанта, тогда как в bioH+ штаммах регистрировали почти 30% гидролиза (Фигура 14B).
ПРОВЕРКА СВОЙСТВ BIOH IN VITRO
С целью доказательства того, что BioH непосредственно вовлечен в гидролиз DMB-S-MMP в ходе ферментации, клонировали ген bioH и экспрессировали его в N-his-меченной форме в BL21(DE3) (см., например, Tomczyk et al., (2002) FEBS Lett 513: 299-304). Белок очищали до гомогенности с помощью аффинной хроматографии Ni-NTA с конечным выходом 9 мг/л. Оценивали каталитические свойства BioH в отношении DMB-S-MMP, а также измеряли степень гидролиза с помощью ВЭЖХ (234 нм). Исследовали кинетику Михаэлиса-Ментен для BioH в отношении DMB-S-MMP, при этом определенные значения kcat и Км составляли 260±45 с-1 и 229±26 мкМ, соответственно (Фигура 15A).
Также с помощью анализа ВЭЖХ исследовали активность BioH в отношении других диметилбутирилтиоэфиров, полученные значения показаны на фигуре 15B. При идентичных условиях реакции (1 мМ субстрата, 10 нМ BioH) отмечали, что наибольшую эстеразную активность BioH проявлял в отношении DMB-S-MMP. Уменьшение длины основной цепи карбоновой кислоты приводило к 2,5-кратному снижению скорости гидролиза (DMB-S-MTG), тогда как увеличение сложноэфирной цепи, до этилового эфира, значительно снижало скорость гидролиза субстрата (DMB-S-EMP). Полученные результаты коррелируют с результатом in vivo, где оба субстрата гидролизовались в присутствии клеток, хотя и в меньшей степени, чем DMB-S-MMP.
BioH является ключевым ферментом в биосинтезе биотина в E. coli (см., например, Lemoine et al., (1996) Mol Microbiol 19: 645-647). Было предположено, что BioH является ответственным за синтез пимелоил-CoA (см., например, Guillen-Navarro et al., (2005) FEMS Microbiol Lett 246: 159-165), но его точная биохимическая функция еще не была подтверждена. Примечательно, что кристаллическая структура BioH E. coli была определена при разрешении 1,7Å в попытке предсказать функцию белка на основе структурных свойств (см., например, Sanishvili et al., (2003) J Biol Chem 278: 26039-26045). Высокоэффективный структурный анализ привел к обнаружению в BioH каталитической триады, которая также была найдена в известных гидролазах, что указывает на то, что BioH может обладать гидролитической активностью. Анализы с использованием сложных эфиров п-нитрофенола показали, что BioH проявляет карбоксиэстеразную активность с предпочтением в отношении сложных эфиров жирных кислот с короткой цепью (см., например, Sanishvili et al., (2003) J Biol Chem 278: 26039-26045). Настоящая работа, in vivo и in vitro, дополнительно позволила изучить биохимические свойства BioH и показала, что фермент обладает очень широкой субстратной специфичностью в отношении сложноэфирных групп. Напротив, BioH не проявлял какой-либо тиоэстеразной активности в отношении тиоэфирной связи, присутствовавшей в субстратах, проанализированных в настоящей работе. Какого-либо дальнейшего расщепления гидролизованных тиоэфирных кислот не наблюдали.
СОЗДАНИЕ BL21(DE3) ΔBIOH МУТАНТА YT2
После идентификации и проверки BioH как фермента, ответственного за гидролиз DMB-S-MMP в ходе ферментации, стало очевидно, что штамм E. coli с дефицитом BioH должен использоваться в качестве организма-хозяина в цельноклеточном биосинтезе кислотной формы симвастатина. Сконструировали различные векторы экспрессии LovD, не требующие T7-полимеразы, и ввели их в JW3357. Оценка скорости биоконверсии кислотной формы симвастатина показала, что в полученных штаммах активность LovD значительно ниже, чем в BL21(DE3)/pAW31. Более низкая скорость реакции была в значительной степени обусловлена пониженными уровнями экспрессии LovD в полученных комбинациях хозяин/вектор, что было определено с помощью ДСН-ПААГ электрофореза. В результате был сделан вывод, что для того, чтобы достичь максимальной скорости превращения, устранив гидролиз субстрата, необходим ΔbioH мутант BL21(DE3).
Каждый из мутантов с нокаутом одного гена из Коллекции Keio содержал ген устойчивости к канамицину вместо целевого гена (см., например, Baba et al., (2006) Mol Syst Biol 2: 2006 0008). Маркер фланкирован сайтами FRT, которые способствуют свободному удалению маркера ферментом FLP. Для переноса маркера ΔbioH::FRT-kan-FRT из JW3357 в BL21(DE3) использовали P1 трансдукцию. Из-за различий в системе рестрикции между донорным штаммом E. coli К и реципиентным штаммом E. coli В использовали E. coli WA837 (без системы рестрикции, с системой модификации) в качестве промежуточного хозяина в Р1 трансдукции (см., например, Dien et al., (2001) J Ind Microbiol Biotechnol 27: 259-264). После переноса маркера в BL21(DE3) с получением YTl вспомогательную плазмиду pCP20, которая содержит термочувствительный репликон и термоиндуцируемый ген FLP (см., например, Datsenko et al., (2000) Proc Natl Acad Sci U S A 97: 6640-6645; и Wanner, 2000), использовали для удаления гена kan с получением YT2 (BL21(DE3) ΔbioH::FRT). ПЦР-анализ с использованием праймеров, отжигающихся слева и справа, подтверждал удаление bioH, а также удаление маркера kan (данные не показаны).
Затем YT2 культивировали в среде LB и проводили анализ на гидролиз DMB-S-MMP. Как ожидали, новый штамм не катализировал какого-либо заметного гидролиза тиоэфира, при этом следов DMB-S-MPA в культуральной жидкости не обнаруживали. DMB-S-MMP можно почти количественно выделить из насыщенной культуры, выращенной в течение >24 часов, причем субстрат гарантированно может оставаться незатронутым в ходе длительной ферментации с использованием данного штамма.
ЦЕЛЬНОКЛЕТОЧНЫЙ БИОКАТАЛИЗ С ИСПОЛЬЗОВАНИЕМ YT2
Вначале исследовали устойчивость YT2 как хозяина в условиях цельноклеточного биокаталитического синтеза кислотной формы симвастатина. YT2 трансформировали плазмидой экспрессии pAW31 посредством электропорации с получением YT2/pAW31. BL21(DE3)/pAW31 и YT2/pAW31 выращивали до OD600 0,5 с последующей индукцией синтеза белка (100 мкМ ИПТГ) при 20°C в течение 16 часов. Два штамма показали идентичную кинетику роста при использовании среды LB, достигая одинаковой плотности OD600 (3,9~4,0) в конце периода экспрессии белка. При использовании минимальной среды F1 два штамма росли с сопоставимой скоростью до индукции (Фигура 16A). Напротив, в среде без добавки биотина после индукции YT2/pAW31 рос значительно медленнее, чем BL21(DE3)/pAW31. Плотность культуры после индукции для мутантного штамма составляла ~60% от плотности исходного штамма. Низкую скорость роста связывали с неспособностью YT2/pAW31 синтезировать биотин и поддерживать устойчивый рост клеток в минимальной среде. Когда штамм YT2/pAW31 выращивали в среде F1 с добавкой биотина (0,15 мг/л), кинетика роста мутантного штамма была такой же, как и у BL21(DE3)/pAW31.
Затем сравнивали эффективность YT2/pAW31 по сравнению с BL21(DE3)/pAW31 в цельноклеточном анализе. Оба штамма выращивали в среде LB, после чего концентрировали в десять раз с целью имитации условий высокой плотности культуры через 12 часов экспрессии LovD. Для инициирования биоконверсии кислотную форму MJ и DMB-S-MMP добавляли до конечных концентраций 15 мМ и 25 мМ, соответственно. На Фигуре 16B показано, что YT2/pAW31 был значительно более эффективен в анализе в результате делеции bioH. Полное превращение (>99%) для мутантного штамма наблюдали меньше чем через 12 часов инкубации без следов гидролиза DMB-S-MMP. Напротив, BL21(DE3)/pAW31 достиг такого же уровня превращения через 24 часа, дополнительно требуя добавления 25 мМ DMB-S-MMP в ходе ферментации в результате гидролиза субстрата. В линейном диапазоне реакции YT2/pAW31 синтезировал кислотную форму симвастатина со скоростью 1,5 мМ/ч, то есть значительно быстрее, чем BL21(DE3)/pAW31 - 0,75 мМ/ч (см., например, Xie et al., (2007) Appl Environ Microbiol 73: 2054-2060). Используя YT2/pAW31, можно было уменьшить минимальную концентрацию DMB-S-MMP, требуемую для завершения реакции. Полного превращения в кислотную форму симвастатина можно достичь с той же скоростью, когда исходное соотношение DMB-S-MMP (18 мМ) к кислотной форме MJ (15 мМ) составляет не более 1,2.
Повышение скорости в значительной степени обусловлено увеличением внутриклеточной концентрации DMB-S-MMP в отсутствии BioH. В исследованиях in vitro было показано, что BioH чрезвычайно быстро гидролизует DMB-S-MMP. Поэтому, даже при низких внутриклеточных концентрациях BioH, реакция гидролиза субстрата может истощать пул субстратов, которые присутствуют в цитоплазме. Сохранение повышенной внутриклеточной концентрации DMB-S-MMP является критическим, особенно предполагая, что LovD может катализировать обратную реакцию, в которой кислотная форма симвастатина может гидролизоваться до кислотной формы MJ в отсутствии ацилтиоэфирных доноров (см., например, Xie et al., (2006) Chem Biol 13: 1161-1169).
Очистка кислотной формы симвастатина после биоконверсии также была улучшена, что демонстрировали при выполнении ферментации в повышенном объеме YT2/pAW31 (200 мл, 15 мМ MJ, 18 мМ DMB-S-MMP). После проверки с помощью ВЭЖХ полного превращения кислотной формы MJ в кислотную форму симвастатина (Фигура 17, график a), объединенную с клеточным экстрактом культуральную жидкость промывали гексаном, подкисляли 6 М HCl и фильтровали с получением осадка кислотной формы симвастатина (график b). После промывания фильтрационного осадка 1 объемом dH2O кислотную форму симвастатина растворяли в ACN, фильтровали и анализировали с помощью ВЭЖХ (график c). Дополнительных стадий промывки для удаления DMB-S-MPA, который присутствовал в BL21(DE3)/pAW31, не требовалось. Конечный выход кислотной формы симвастатина с использованием данной методики составил 94%.
ВЫВОД
Как показано выше, авторы настоящего изобретения использовали библиотеку Keio мутантов с нокаутом одного гена для идентификации BioH как карбоксиэстеразы, которая гидролизует DMB-S-MMP в процессе цельноклеточного биокаталитического превращения кислотной формы MJ в кислотную форму симвастатина. BioH демонстрирует очень высокую скорость гидролиза, в результате которого истощается внутриклеточная концентрация DMB-S-MMP, доступного в качестве донора ацила в LovD-катализируемой переэтерификации. Применение bioH экспрессионного штамма YT2 позволило полностью устранить расщепление DMB-S-MMP и значительно повысить устойчивость цельноклеточного биокатализатора. Указанный штамм может применяться в качестве хозяина в других направленных биосинтезах из предшественника и процессах биокатализа, в которых один или несколько используемых субстратов содержат неустойчивую сложноэфирную связь (см., например, Murli et al., (2005) Appl Environ Microbiol 71: 4503-4509).
ПРИМЕР 4: ЭФФЕКТИВНЫЙ СИНТЕЗ СИМВАСТАТИНА С ИСПОЛЬЗОВАНИЕМ ЦЕЛЬНОКЛЕТОЧНОГО БИОКАТАЛИЗА
В данном примере описан способ одностадийного цельноклеточного биокаталитического синтеза симвастатина из монаколина J. Как подробно описано ниже, используя штамм Escherichia coli с оверэкспрессией LovD A. Terreus, можно было достичь >99% превращения монаколина J в симвастатин без использования каких-либо стадий химической защиты. Ключевым открытием являлся проникающий через мембрану субстрат α-диметилбутирил-S-метил-меркаптопропионат (DMB-S-MMP), который эффективно использовался LovD в качестве донора ацильной группы. Процесс масштабировали для синтеза граммовых количеств симвастатина. Также было показано, что симвастатин, синтезируемый с помощью указанного способа, может быть легко очищен из среды ферментации с >90% выходом и >98% чистотой, согласно анализу ВЭЖХ. Также тестировали биоконверсию при высокой плотности культуры и периодическую ферментацию. Цельноклеточный биокатализ, таким образом, может быть привлекательной альтернативой текущим, многостадийным полусинтетическим способам превращения.
В настоящее время для синтеза симвастатина из ловастатина широко используются два полусинтетических способа (см., например, Askin et al., 1991, J. Org. Chem. 56; и Hoffman et al., 1986 J Med Chem 29: 849-52) (Фигура 18). Один из обычно используемых способов (см., например, Hoffman и др., 1986 J Med Chem 29: 849-52) начинается с гидролиза ловастатина с образованием ключевого промежуточного соединения монаколина J с последующей лактонизацией кислоты для защиты С11 гидроксильной группы и триметилсилильной защиты C13 гидроксила. Затем защищенный монаколин J подвергают ацилированию α-диметилбутирилхлоридом с получением защищенной формы симвастатина, которую затем подвергают снятию защиты с получением симвастатина. Оба многостадийных способа, показанных на Фигуре 18, являются сложными, в результате чего симвастатин почти в пять раз дороже, чем ловастатин. Таким образом, новая схема полусинтетического способа, которая может уменьшить число химических преобразований и увеличить общую эффективность превращения, может являться весьма перспективной.
Ранее авторы настоящего изобретения описывали клонирование и исследование характеристик специальной ацилтрансферазы из кластера генов биосинтеза ловастатина, которая региоселективно переносит α-метилбутирильную группу от ловастатин-дикетидсинтазы (LDKS) к C8-гидроксильной группе монаколина J с получением ловастатина (см., например, Xie et al., 2006, Chem Biol 13: 1161-1169). Было продемонстрировано, что LovD обладает широкой субстратной специфичностью в отношении переносчика ацила, ацильной группы и декалинового ядра. В особенности, LovD была способна катализировать прямое ацилирование монаколина J α-диметилбутирил-S-N-ацетилцистамином (DMB-S-NAC) с получением симвастатина. Реакция является высокорегиоспецифичной только в отношении C8 гидроксила, и поэтому может рассматриваться в качестве потенциального одностадийного способа получения симвастатина из монаколина J без использования защитной химии.
В данном Примере описывается разработка высокоэффективного цельноклеточного биокаталитического способа превращения монаколина J в симвастатин. Используя новый тиоэфир в качестве донора ацила, можно было достичь >99% превращения монаколина J в симвастатин в одну стадию. Ферментационный способ может быть легко масштабирован в целях получения симвастатина в промышленном масштабе.
СИНТЕЗ α-ДИМЕТИЛБУТИРИЛ-S-МЕТИЛ-3-МЕРКАПТОПРОПИОНАТА (DMB-S-MMP):
Диметилбутирилхлорид (1,76 мл, 12 ммоль) медленно добавляли к 50 мл раствора метил-3-меркаптопропионата (1,30 мл, 12 ммоль) и триэтиламина (3,340 мл, 24 ммоль) в диэтиловом эфире при 0°C, после чего раствор перемешивали в течение 2 часов. Реакцию останавливали водным р-ром NH4Cl и дважды экстрагировали 100 мл этилацетата. Органический слой объединяли, сушили и выпаривали с получением бесцветной жидкости (2,6 г). Остаток очищали с помощью хроматографии на силикагеле (EA/гексан, 20/80) с получением чистого DMB-S-MMP (2,35 г, 90% выход). 1H ЯМР: δ 3,87 (с, 3H), 3,09 (т, 2H, 7,1 Гц), 2.58 (т, 2H, 7,0 Гц), 1,59 (кв, 2H, 7,5 Гц), 1,18 (с, 6H), 0,83 (т, 3H, 7,4 Гц); 13C ЯМР: δ 206,16, 172,26, 51,81, 50,04, 34,38, 33,73, 24,71, 23,60, 8,92.
КИНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ LOVD:
Экспрессия и очистка LovD были подробно описаны ранее (см., например, Xie et al., 2006, Chem Biol 13: 1161-1169). Анализы проводили при комнатной температуре в 50 мМ HEPES, pH 7,9. Начальные скорости (ki) ацильных субстратов определяли при 10 мкМ LovD, 1 мМ монаколина J и 4 мМ тиоэфира. Реакционную смесь останавливали в различные моменты времени трифторуксусной кислотой (ТФУ), экстрагировали этилацетатом, выпаривали досуха и повторно растворяли в ацетонитриле. Процент превращения монаколина J в симвастатин определяли с помощью ВЭЖХ-анализа (C18). Линейный диапазон скорости ферментативной реакции обозначали ki в Таблице 1B. Для определения Км для DMB-S-MMP концентрацию монаколина J установливали на уровне 2 мМ, при этом концентрация DMB-S-MMP варьировала от 0,2 мМ до 10 мМ. Диметилсульфоксид добавляли до конечной концентрации 5% с целью облегчения солюбилизации DMB-S-MMP. Для получения Км для монаколина J концентрацию DMB-S-MMP устанавливали на уровне 2 мМ, при этом концентрация монаколина J варьировала от 0,2 мМ до 5 мМ.
АНАЛИЗ АКТИВНОСТИ СУММАРНОГО КЛЕТОЧНОГО ЛИЗАТА
Анализ активности суммарного клеточного лизата использовали для определения уровня активности LovD при различных условиях ферментации. Например, штамм E. coli BL21(DE3), трансформированный pAW31, выращивали в среде LB при 37°C до OD600 0,5, после чего к культуре добавляли 100 мкМ ИПТГ и проводили экспрессию при комнатной температуре (КТ) в течение 16 часов. Затем аликвоту культуры (70 мл) осаждали центрифугированием (4000g, 20 мин). Супернатант отбрасывали, а осадок клеток ресуспендировали в 7 мл буфера для лизиса (20 мМ Трис-HCl, pH 7,9, 500 мМ NaCl). Клетки разрушали ультразвуком на льду, после чего клеточный дебрис удаляли центрифугированием (13000 об/мин, 10 мин, 4°C). Затем лизат подвергали анализу in vitro. Конечные условия анализа: 50 мМ HEPES, pH 7,9, 1 мМ монаколина J, 4 мМ DMB-S-MMP, 5 мкл клеточного лизата, конечный объем 25 мкл. Затем эффективную концентрацию LovD в среде ферментации вычисляли из начальной скорости, используя значение скорости ферментативной реакции 0,6 мин-1.
ФЕРМЕНТАЦИЯ ПРИ НИЗКОЙ ПЛОТНОСТИ КУЛЬТУРЫ
Штамм E. coli BL21(DE3)/pAW31 выращивали в среде LB при 37°C до OD600 0,5, после чего к культуре добавляли 100 мкМ ИПТГ и проводили экспрессию при КТ в течение 16 часов. 10 мл культуры переносили в центрифужную пробирку на 15 мл. Клетки осаждали центрифугированием (4000g, 10 мин). Осадок клеток ресуспендировали в 957 мкл супернатанта. Затем pH доводили до 7,9 1,0Н р-ром NaOH с последующим добавлением 33 мкл 450 мМ р-ра MJ (конечная концентрация 15 мМ) и 6 мкл чистого DMB-S-MMP (конечная концентрация ~25 мМ). Затем культуру встряхивали при 300 об/мин при комнатной температуре. В каждый момент времени отбирали аликвоту (4 мкл) реакционной смеси и останавливали реакцию посредством добавления 300 мкл этилацетата, содержащего 1% ТФУ. Органическую фазу отбирали, выпаривали и повторно растворяли в ацетонитриле с целью проведения ВЭЖХ-анализа.
В целях синтеза симвастатина в большем объеме вышеприведенную методику масштабировали, начиная с культур штамма E. coli BL21(DE3)/pAW31 в 2-х встряхиваемых колбах по 1 л. Через 16 часов экспрессии при КТ культуру сконцентрировали до 200 мл с конечной плотностью OD600 22. Монаколин J (в форме натриевой соли) и DMB-S-MMP добавляли до конечной концентрации 15 мМ и 25 мМ, соответственно. Клетки встряхивали при комнатной температуре, при этом для контроля протекания реакции использовали ВЭЖХ. Для выделения симвастатина из культуры после завершения реакции (>99% превращения согласно ВЭЖХ-анализу) использовали следующую методику. Клетки осаждали центрифугированием. Внутриклеточный симвастатин экстрагировали, суспендируя осадок клеток в ацетоне. Затем ацетон отгоняли при пониженном давлении, а осадок растворяли в dH2O и фильтровали. Фильтрат и осветленную культуральную жидкость объединяли, дважды промывали равным объемом н-гексана и подкисляли до pH 2,0 6Н р-ром HCl. Белый осадок отфильтровывали и промывали избытком ледяной dH2O для удаления одновременно выпавшего DMB-S-MPA. После высушивания осадка на фильтре в вакууме твердые частицы перемешивали в 200 мл ацетонитрила в течение 1 часа с последующей фильтрацией для удаления нерастворимых примесей. Фильтрат выпаривали досуха при пониженном давлении с получением кислотной формы симвастатина.
ПЕРИОДИЧЕСКАЯ ФЕРМЕНТАЦИЯ С ПОДПИТКОЙ ПРИ ВЫСОКОЙ ПЛОТНОСТИ КУЛЬТУРЫ В F1
Периодическую ферментацию с подпиткой в F1 и состав среды заимствовали у Pfeifer et al. (см., например, Pfeifer et al., 2002, Appl Environ Microbiol 68: 3287-92). Раствор витаминов исключили из ферментационной среды и среды для подпитки. Стартовую культуру выращивали за ночь в 5 мл среды LB (с 35 мг/л канамицина) при 37°C и 250 об/мин, а затем 1 мл использовали для инокуляции встряхиваемой колбы на 100 мл с инокуляционной средой F1 (с 35 мг/л канамицина). 10 мл инокуляционной культуры F1 использовали для инокуляции 2-литровой емкости Applikon Biobundle, содержащей 1 л среды F1. Ферментацию проводили при 37°C, а pH поддерживали на уровне 7,1 в ходе эксперимента, добавляя 1 М H2SO4 и 0,5 М NH4OH. Аэрацию регулировали в диапазоне 0,2-0,4 л/мин, а перемешивание поддерживали на уровне 900 об/мин. Когда OD600 достигала 5-10, температуру ферментации уменьшали до КТ с последующим добавлением 200 мкМ ИПТГ для инициирования экспрессии белка. В то же время перистальтический насос подавал в ферментер раствор для подпитки при расходе 0,08 мл/мин.
Каталитическую эффективность LovD и концентрацию на различных стадиях ферментации измеряли, как описано выше. Для подготовки клеток в стадии покоя к экспериментам по биоконверсии клетки центрифугировали при 5000g в течение 10 минут с последующим нежным ресуспендированием в том же объеме фосфатно-солевого буфера с pH 7,4. Затем монаколин J и DMB-S-MMP добавляли к аликвотам клеток для инициирования синтеза симвастатина.
РЕЗУЛЬТАТЫ
ИДЕНТИФИКАЦИЯ ДОНОРА АЦИЛА С ЛУЧШИМИ КИНЕТИЧЕСКИМИ ПАРАМЕТРАМИ
Авторы настоящей заявки ранее показали, что LovD может использовать проникающие через мембрану тиоэфиры в качестве доноров ацильной группы в реакции переэтерификации (см., например, Xie et al., 2006, Chem Biol 13: 1161-1169). И DMB-S-NAC, и DMB-S-MTG (Таблица 1B), как было показано, являются субстратами LovD. Указанные два субстрата, однако, поддерживают низкую скорость ферментативной реакции в синтезе симвастатина, с кажущимся значением ki ~0,02 в минуту. Кроме того, когда используется какой-либо субстрат, реакция замедляется из-за сильного ингибирования субстрата при растущих концентрациях монаколина J. Таким образом, первоочередной задачей в превращении LovD в промышленно применимый катализатор для хемобиосинтеза симвастатина является идентификация лучших субстратов, которые могут преодолеть указанные два ограничения.
Синтезировали несколько дополнительных вариантов DMB-S-MTG и оценивали их каталитические свойства in vitro. DMB-S-метилмеркаптопропионат (DMB-S-MMP), DMB-S-этилмеркаптопропионат (DMB-S-EMP) и DMB-S-метилмеркаптобутират (DMB-S-MMB) синтезировали путем взаимодействия α-диметилбутирилхлорида с соответствующими свободными тиолами. Начальные скорости ферментативной реакции сравниваются в Таблице 1B. Неожиданно увеличение длины тиоэфира-переносчика на один атом углерода (от C2 в S-MTG до C3 в S-MMP и S-EMP) привело к значительному повышению скорости ферментативной реакции. Введение дополнительного атома углерода в S-MMB (C4) привело к дополнительному повышению скорости переэтерификации. При стандартных условиях реакции (1 мМ MJ, 4 мМ ацильного субстрата, 10 мкМ LovD, 50 мМ HEPES, pH 7,9) начальные скорости ферментативной этерификации составили 0,6, 0,7, 0,78 мин-1 для DMB-S-MMP, DMB-S-EMP и DMB-S-MMB, соответственно. Более чем 30-кратное повышение скорости показывает, что относительное положение карбонильных групп к тиоэфирной связи является критическим при связывании с LovD. Напротив, когда DMB-S-MPA использовали в качестве донора ацильной группы, начальная скорость ферментативной реакции значительно уменьшилась до 0,08 в минуту, указывая, что свободная кислота (вероятно, и натриевая соль) плохо связывается с активным сайтом LovD.
DMB-S-MMP выбрали в качестве наиболее подходящего для биокатализа ацилтиоэфира, потому что его тиол-предшественник (метилмеркаптопропионат) значительно более дешевый, чем другие исходные материалы. Были определены кинетические параметры реакции ацилирования при использовании DMB-S-MMP в качестве донора ацила, которые показаны на Фигуре 19. Чтобы получить значения Км для любого субстрата в реакции переацилирования, установили на постоянном уровне концентрацию одного субстрата, изменяя концентрацию другого субстрата с получением кинетической кривой Михаэлиса-Ментен. На Фигуре 19A показана скорость ферментативной реакции (V/Eo) в зависимости от концентрации монаколина J. В отличие от ранее исследованных субстратов какого-либо ингибирования субстрата монаколином J не наблюдали, а полученное значение Км составило 0,78±0,12 мМ. Число оборотов LovD при изменении количеств DMB-S-MMP определяли аналогично, устанавливая концентрацию монаколина J на уровне 2 мМ (Фигура 19B). Значение Км для DMB-S-MMP, как показали, составило 0,67±0,04 мМ. В обоих титрованиях полученное значение kcat составило 0,66±0,03 мин-1. Приведенные результаты кинетического анализа четко показывают, что DMB-S-MMP является субстратом с лучшими кинетическими параметрами по сравнению с тиоэфирами, описанными ранее. Уровень ингибирования субстрата в ферментативной реакции по механизму пинг-понг зависит от относительного значения Км для двух указанных субстратов. Когда DMB-S-MMP используется в качестве ацильного субстрата, его относительно более низкое значение Км позволяет ему легко связываться с LovD и поэтому не подвергаться ингибированию при увеличении концентрации монаколина J. Указанное важное свойство, таким образом, позволяет применять периодический биокаталитический способ в отличие от периодического способа с подпиткой, в котором монаколин J должен непрерывно подаваться для сохранения его концентрации на низком уровне и сведения к минимуму ингибирования субстрата.
Ранее авторы настоящего изобретения отмечали, что ацил-S-MTG быстро гидролизовался при добавлении в среду культивирования E. coli (см., например, Xie et al., 2006, Chem Biol 13: 1161-1169). С целью проверки устойчивости недавно идентифицированного донора ацила in vivo чистый DMB-S-MMP добавляли в среду LB, инокулировали клетками E. coli BL21(DE3) и оставляли культуру на ночь при 37°C (16 часов). Наблюдали расщепление субстрата на ~20% с преобладающим образованием более полярного соединения. При сравнении с известными соединениями, используя ВЭЖХ, основной продукт расщепления идентифицировали как DMB-S-MPA, который может образовываться под действием эндогенных липаз E. coli. Сделали вывод, что при внесении избытка DMB-S-MMP в периодическую реакцию низкая степень расщепления не будет лимитировать биоконверсию монаколина J в симвастатин.
ЦЕЛЬНОКЛЕТОЧНЫЙ БИОКАТАЛИЗ
Получив в свое распоряжение значительно более эффективный DMB-S-MMP тиоэфир, авторы настоящей заявки изучили превращение монаколина J в симвастатин, используя E. coli в качестве цельноклеточного биокатализатора. Штамм BL21(DE3), трансформированный сконструированной на основе pET28a экспрессионной плазмидой pAW31, использовали в качестве микроорганизма-хозяина. Экспрессию LovD выполняли в течение 16 часов при комнатной температуре, причем уровень экспрессии анализировали с помощью ДСН-ПААГ электрофореза. Для определения количества активной LovD, экспрессированной в различных условиях культивирования и средах, использовали анализ активности, в котором суммарный лизат клеток непосредственно добавляли в аналитическую пробу, содержащую 1 мМ монаколина J, 4 мМ DMB-S-MMP в 50 мМ HEPES, pH 7,9. Превращение монаколина J в симвастатин количественно определяли с помощью ВЭЖХ, при этом кажущуюся концентрацию LovD оценивали, используя значение ki 0,6 мин-1 (Таблица 1B). В условиях низкой плотности культуры экспрессия LovD в среде LB почти в три раза выше, чем в минимальной среде F1 (Таблица 3).
Затем исследовали хемобиосинтез симвастатина в среде LB. После ночной экспрессии LovD 10 мл аликвоту клеток сконцентрировали в десять раз до 1 мл в LB. Стадию концентрации выполняли с целью создания условий ферментации, имитирующих высокую плотность культуры, и получения более высокой эффективной концентрации LovD (конечная концентрация приблизительно 20 мкМ). Монаколин J в форме натриевой соли добавляли до конечной концентрации 15 мМ (450 мМ стоковый раствор в воде), после чего чистый DMB-S-MMP добавляли до конечной концентрации 25 мМ. Затем реакционную смесь энергично встряхивали при комнатной температуре. В моменты времени периодически отбирали пробы для анализа превращения монаколина J в симвастатин (Фигура 20A).
Как показано на фигуре 20B, простая культура BL21(DE3)/pAW31 низкой плотности являлась устойчивым цельноклеточным биокатализатором для синтеза симвастатина. Начальную 2-часовую лаг-фазу наблюдали при добавлении указанных двух субстратов с последующим быстрым превращением с линейной скоростью реакции ~0,73 мМ в час. Скорость реакции снизилась при более высоком уровне превращения (>95%) в результате истощения монаколина J. В течение 24 часов 99% монаколина J было превращено в симвастатин. По данным анализа ВЭЖХ какого-либо расщепления монаколина J или симвастатина в результате клеточного метаболизма не наблюдалось. Регистрировали уменьшение на ~20% плотности культуры OD600 через 24 часа, что позволяет предположить, что цельноклеточный процесс можно продолжать в течение длительного катализа.
Авторы настоящей заявки сообщали, что LovD способен гидролизовать ловастатин до монаколина J in vitro при kcat 0,21 мин-1 (см., например, Xie et al., 2006, Chem Biol 13: 1161-1169). С целью исследования, можно ли объединить стадию гидролиза вместе со стадией ацилирования в единый ферментационный процесс, оценивали скорость гидролиза ловастатина под действием BL21(DE3)/pAW31. В тех же in vivo условиях, как и выше, наблюдали образование монаколина J с низкой скоростью. Начиная с 0,5 мМ ловастатина (в форме натриевой соли), достигали 91% гидролиза через 48 часов, при скорости превращения ~0,01 мМ в час, что почти в 75 раз медленнее, чем скорость реакции ацилирования. Это резко контрастирует с кинетическими показателями in vitro, когда скорость гидролиза лишь в три раза медленнее, чем реакция ацилирования. Существенное ослабление скорости гидролиза ловастатина in vivo происходит, по-видимому, из-за мембранного барьера клетки, препятствующего проникновению более гидрофобного ловастатина. Провели попытку облегчить прохождение мембранного барьера путем использования альтернативного штамма BL21(DE3)/pXXK2, в котором LovD клонировали с N-концевой сигнальной последовательностью pelB для локализации в периплазму. Какого-либо улучшения гидролитической активности для данного штамма не наблюдали, что указывает на то, что непроницаемость внешней цитоплазматической мембраны является главным препятствием для транспорта.
ВЫДЕЛЕНИЕ И ОЧИСТКА СИМВАСТАТИНА
С целью получения симвастатина посредством цельноклеточного биокаталитического способа увеличили масштаб биоконверсии до конечного объема 200 мл. Это было достигнуто путем концентрирования 2×1 л культуры экспрессирующего штамма в среде LB после ночной экспрессии LovD. Монаколин J в форме натриевой соли и DMB-S-MMP добавляли до конечной концентрации 15 мМ и 25 мМ соответственно, при этом протекание реакции контролировали с помощью ВЭЖХ. Наблюдали идентичную кинетику превращения в более масштабном процессе, и через 24 часа после добавления субстратов превращение составило >99%. Симвастатин, полученный хемобиосинтетическим путем, может быть легко очищен из среды ферментации без использования стадий хроматографической очистки. Для разделения клеток и культуральной жидкости использовали стадию центрифугирования. Внутриклеточный симвастатин выделяли путем суспендирования осадка клеток в ацетоне с последующим выпариванием растворителя и повторным растворением в dH2O. Водный раствор, содержащий внутриклеточный симвастатин, объединяли с культуральной жидкостью и промывали н-гексаном для удаления DMB-S-MMP. Затем водный раствор подкисляли 6Н HCl до pH 2,0, что привело к осаждению свободных кислотных форм симвастатина и DMB-S-MPA. Как известно из уровня техники, свободные кислотные формы симвастатина могут быть превращены в натриевую, калиевую, аммониевую или любую другую соль, полученную со щелочноземельными элементами, или соли других металлов. Затем указанные соли могут быть превращены в чистый симвастатин, например, с помощью общих методик, известных в уровне техники. Примеси (DMB-S-MPA) можно удалять с помощью фильтрации и промывки осадка на фильтре избытком dH2O. Получали подкисленный фильтрат, содержащий <1% общего количества симвастатина. Почти чистый симвастатин может быть получен путем промывки осадка на фильтре ацетонитрилом с последующым выпариванием фильтрата (Конечный выход: 1,13 г, 90%; конечная чистота по данным анализа ВЭЖХ: 98%).
ФЕРМЕНТАЦИЯ ПРИ ВЫСОКОЙ ПЛОТНОСТИ КУЛЬТУРЫ
Активность цельноклеточного биокатализатора исследовали с использованием ферментера, рассчитанного на высокую плотность культуры. Эффективная концентрация LovD, измеренная в ферментере периодического действия со средой TB, и эффективная концентрация LovD, измеренная в ферментере периодического действия с подпиткой с минимальной средой F1, приведены в Таблице 3. Используя периодический процесс с подпиткой, заимствованный из работы Pfeifer et al (см., например, Pfeifer et al., 2002, Appl Environ Microbiol 68: 3287-92), можно получать культуры с очень высокой плотностью клеток и активностью LovD (Фигура 21A). Можно было достигать уровня экспрессии активной LovD ~1,5 г/л при OD600 75 через 40 часов после инокуляции (Фигура 21A).
Из-за высокой стоимости исходных материалов не было возможности выполнить биоконверсию, используя всю ферментационную среду. Вместо этого в ходе процесса ферментации отбирали аликвоты культуры в различные моменты времени (Фигура 21A). Активность LovD в каждом моменте времени определяли, используя анализ суммарного клеточного лизата, описанный в Материалах и Методах. Затем выполняли биоконверсию, непосредственно используя 1 мл культуры и измеряя скорость образования симвастатина (делящиеся клетки, Фигура 21B). Альтернативно также исследовали свойства цельноклеточного биокатализатора после ресуспендирования осадков клеток в PBS, pH 7,4 (клетки в стадии покоя). Как можно видеть на Фигуре 21 B, в каждый момент времени, высокие скорости ферментативной реакции могут быть достигнуты, когда клетки находятся в стадии покоя. Кроме того, полная активность LovD в культуре линейно не коррелирует со скоростью превращения. Например, клетки в стадии покоя в момент времени 3 показывали наиболее высокую скорость синтеза симвастатина, почти 1 г/л/час, несмотря на меньший уровень полной активности LovD по сравнению с клетками, полученными в момент времени 4. Исключительная стабильность указанных клеток позволила завершить превращение 15 мМ монаколина J в симвастатин менее чем за 8 часов.
Ключевым открытием в данном примере является идентификация еще одного ацилтиоэфира, который отдает ацильную группу C8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD, который значительно более эффективен в данной реакции ацилирования. Авторы настоящей заявки ранее идентифицировали DMB-S-NAC как субстрат LovD (см., например, Xie et al., 2006, Chem Biol 13: 1161-1169). Однако ферментативная реакция ацилирования протекала с меньшей, чем идеальная, скоростью, и в результате слабого связывания субстрата с LovD второй субстрат монаколин J стал конкурентным ингибитором DMB-S-NAC. Последнее свойство особенно нежелательно, так как вынуждает поддерживать концентрацию монаколина J в ходе биоконверсии на минимальном уровне. И DMB-S-MMP, и DMB-S-MMB превосходят DMB-S-NAC как доноры ацильной группы. Значение kcat DMB-S-MMP приблизительно в 30 раз выше в результате небольшого изменения структуры переносчика ацила, тогда как значение Км теперь сопоставимо с Км для монаколина J, в результате чего устраняется ингибирование субстрата монаколином J. Кроме того, тиол-предшественник S-MMP значительно менее дорог по сравнению с S-NAC. Принимая во внимание, что ацил-S-NAC-тиоэфиры широко используются в направленном биосинтезе природных продуктов из предшественника (см., например, Jacobsen et al., 1997, Science 277: 367-9), ацилтиоэфирные переносчики, описанные в настоящей заявке, могут также найти применение в других областях генноинженерного биосинтеза.
Авторы настоящего изобретения пытались разработать биокаталитический способ in vitro превращения монаколина J в симвастатин. Предполагали, что грубо очищенный LovD сможет позволить преодолеть любые трудности, связанные с транспортом и очисткой, с которыми сталкиваются в процессе цельноклеточной ферментации. LovD может быть легко отделен в отдельной фракции от большинства других клеточных белков в одной стадии осаждения 30%-ным сульфатом аммония. Более 98% активности LovD, измеренной в суммарном клеточном лизате, может быть выделено при повторном растворении осадка в 50 мМ HEPES, pH 7,9. Однако LovD выпадает быстро (часы) при высоких концентрациях белка (~100 мкМ) и медленно (дни) при более низких концентрациях (~10 мкМ) в уловиях анализа при комнатной температуре. Примечательно, что выпавший белок LovD может быть заново растворен в том же буфере с сохранением почти полной активности.
Когда в периодическом процессе in vitro использовали умеренные количества монаколина J (>5 мМ), не удавалось достигать >60% равновесного превращения монаколина J в симвастатин, даже после длительной инкубации. Вероятно, это обусловлено протеканием конкурентной реакции, в которой симвастатин обратно гидролизуется до монаколина J под действием LovD. Когда концентрация симвастатина достигает миллимолярного диапазона, скорость ферментативной реакции гидролиза достигает максимального значения (kcat=0,3 мин-1) и, таким образом, значительно препятствует достижению полной чистой скорости ацилирования.
Примечательно, что обратная реакция не являлась лимитирующим фактором in vivo, когда концентрация статина находилась в диапазоне от 10 до 15 мМ (4-6 г/л), что свидетельствовало о высокой степени превращения (>99%) монаколина J в симвастатин, достигнутой в данном исследовании. Таким образом, можно предположить, что после превращения монаколина J в симвастатин униферсальные факторы транспорта, такие как тройной комплекс AcrAB-TolC, способны вытеснять симвастатин из внутренней мембраны или периплазмы в среду (см., например, Murakami et al., 2006, Nature 443: 173-9). Непроницаемость внешней мембраны E. coli препятствует возвращению гидрофобного симвастатина. Более полярный монаколин J способен непрерывно диффундировать через мембрану и служить субстратом для LovD. Одновременно внешняя мембрана E. coli и ее выкачивающие насосы эффективно уменьшают внутриклеточную концентрацию симвастатина, доступного для гидролиза, уменьшая, таким образом, скорость нежелательной обратной реакции и максимизируя превращение в требуемой реакции. Однако выкачивающие насосы, обычно экспрессируемые E. coli, могут быть перегружены, когда концентрация субстрата увеличивается до >20 мМ. При повышенных концентрациях монаколина J (и, следовательно, симвастатина) и DMB-S-MMP можно было достичь максимальной степени превращения 85~90%. Анализ распределения симвастатина в культуре показал, что существенное количество (>20%) локализовано в клетках, вероятно приводя, таким образом, к повышению уровня гидролиза продукта. Неизвестно, был ли симвастатин распределен во внутренней мембране, цитоплазме или периплазме. В настоящее время авторы настоящего изобретения исследуют способы повышения интенсивности экспорта симвастатина посредством оверэкспрессии выбранных универсальных выкачивающих насосов (см., например, Masi et al., 2003, J Chromatogr B Analyt Technol Biomed Life Sci 786: 197-205).
В данном примере был предложен очень стабильный цельноклеточный биокаталитический способ синтеза симвастатина из монаколина J. При использовании E. coli в качестве организма-хозяина был осуществлен лабораторный процесс синтеза граммовых количеств. Дополнительно, простая схема последующей очистки была предложена для легкого выделения и очистки продукта. В настоящее время авторы настоящей заявки используют рациональные и направленные методы изменения, направленные на улучшение числа оборотов LovD. Благодаря оптимизации свойств LovD, а также метаболической инженерии организма-хозяина в целях улучшения эффективности превращения, хемобиосинтетический путь получения симвастатина может стать конкурентоспособной и привлекательной альтернативой синтетическим путям, показанным на Фигуре 18.
Объем настоящего изобретения не должен ограничиваться вариантами осуществления, раскрытыми в настоящем описании, которые предназначены в качестве простых иллюстраций отдельных аспектов изобретения, при этом любые варианты, которые являются функционально эквивалентными, включены в объем изобретения. Различные модификации образцов и способов изобретения в дополнение к описанным в настоящем описании будут очевидны специалистам, квалифицированным в данной области техники, из предыдущего описания и примеров, и также должны быть включены в объем настоящего изобретения. Подобные модификации или другие варианты осуществления могут быть осуществлены на практике без отступления от объема и сущности изобретения.
Ацилтиоэфиры в качестве субстрата LovD[a]
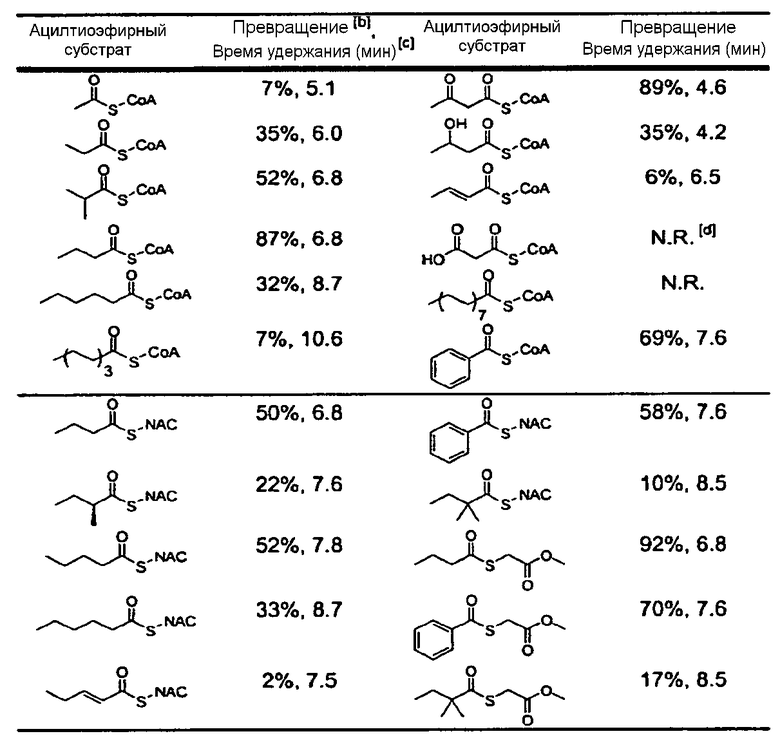
Сравнение начальных скоростей тиоэфирных субстратов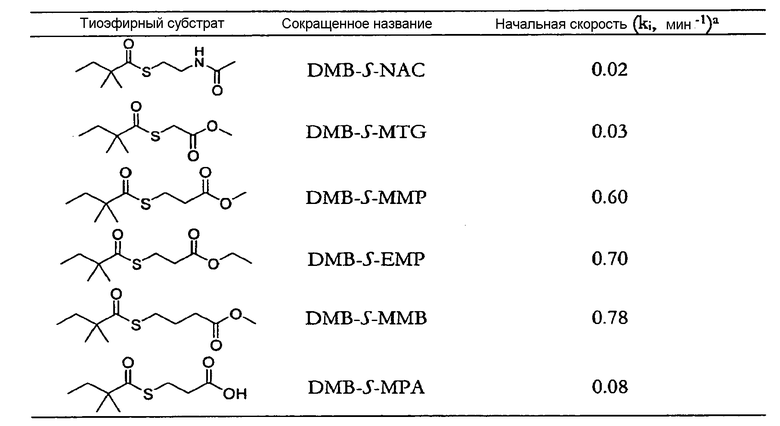
Список мутантов E. coli BW25113, проскринированных в данной работе. Номера соответствуют дорожкам ТСХ, показанным на фигуре 14A. BioH, как было показано, являлся единственным ферментом, ответственным за гидролиз DMB-S-MMP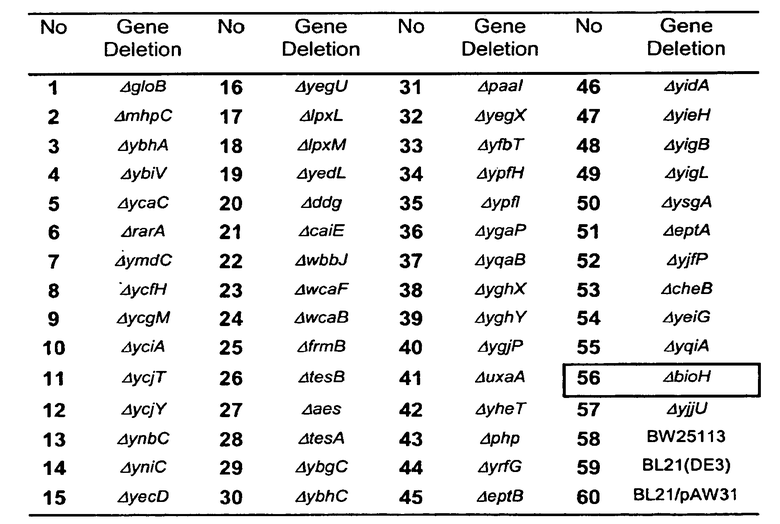
Сравнение выхода белка при различных условиях экспрессииа
b Выход белка оценивали в анализе in vitro, используя суммарный клеточный лизат, как описано в Материалах и Методах. Зарегистрированное превращение использовали для оценки концентрации LovD с учетом числа оборотов (0,6 мин-1).
Список последовательностей полипептидов


| название | год | авторы | номер документа |
|---|---|---|---|
| ФЕРМЕНТАТИВНАЯ ОБРАБОТКА МАСЕЛ | 2004 |
|
RU2377307C2 |
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ, ИХ ПРИМЕНЕНИЕ ПРИ ИСПОЛЬЗОВАНИИ ПИЩЕВЫХ ПРОДУКТОВ | 2010 |
|
RU2538144C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| СПОСОБ | 2004 |
|
RU2376868C2 |
| МОДИФИКАЦИЯ ЛИПИДОВ РАСТЕНИЙ И МАСЕЛ СЕМЯН ПУТЕМ ИСПОЛЬЗОВАНИЯ ДРОЖЖЕВЫХ ГЕНОВ SLC | 1996 |
|
RU2190665C2 |
| МАТЕРИАЛЫ И СПОСОБЫ ЛЕЧЕНИЯ ХРОНИЧЕСКИХ ФИБРОЗНЫХ ЗАБОЛЕВАНИЙ | 2006 |
|
RU2446825C2 |
| ПРОЛЕКАРСТВО ИНТЕРФЕРОНА ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2788736C2 |
| ВАКЦИНА И СПОСОБЫ ОБНАРУЖЕНИЯ И ПРОФИЛАКТИКИ ФИЛЯРИОЗА | 2021 |
|
RU2832185C1 |
| ЭФФЕКТИВНЫЕ АНАЛОГИ КОМПСТАТИНА | 2006 |
|
RU2474586C2 |










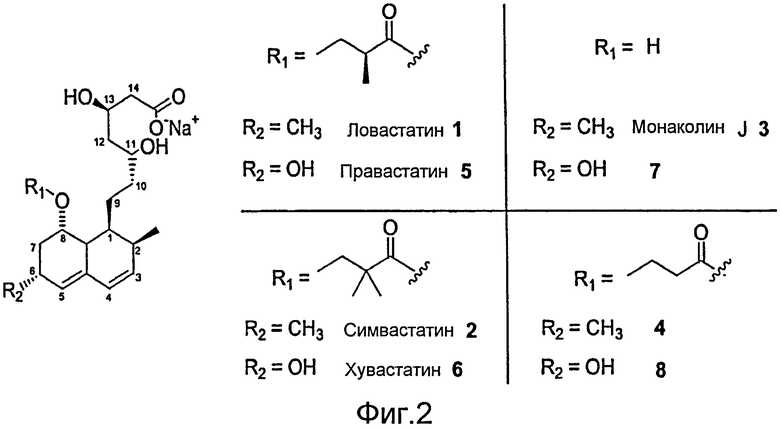
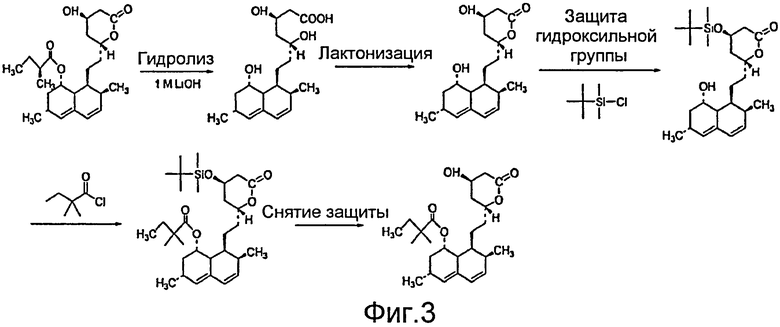
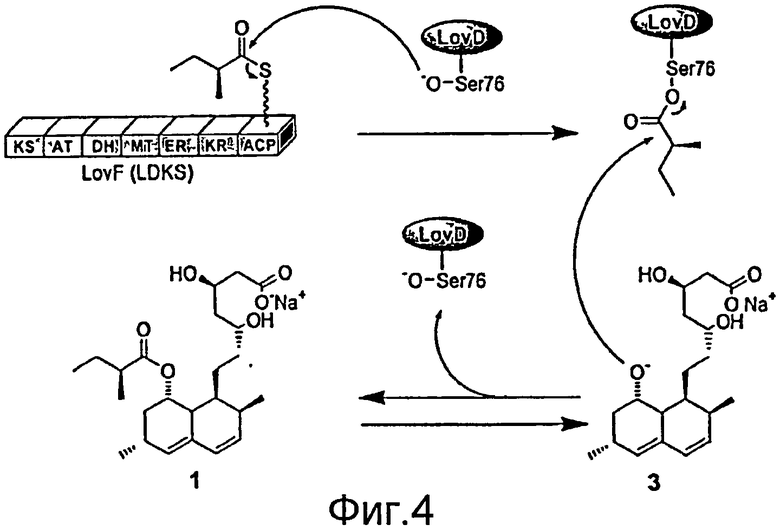
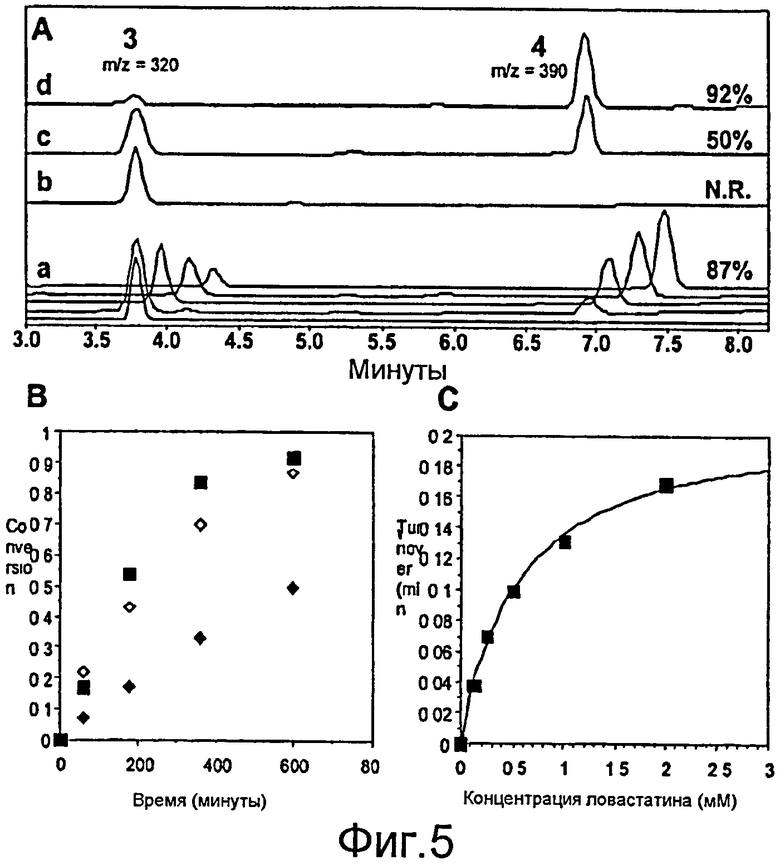
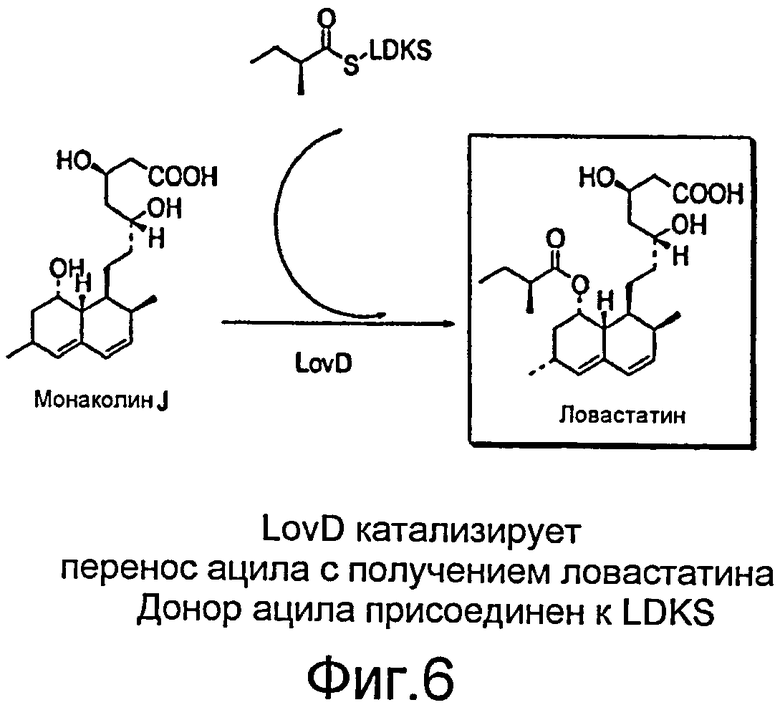
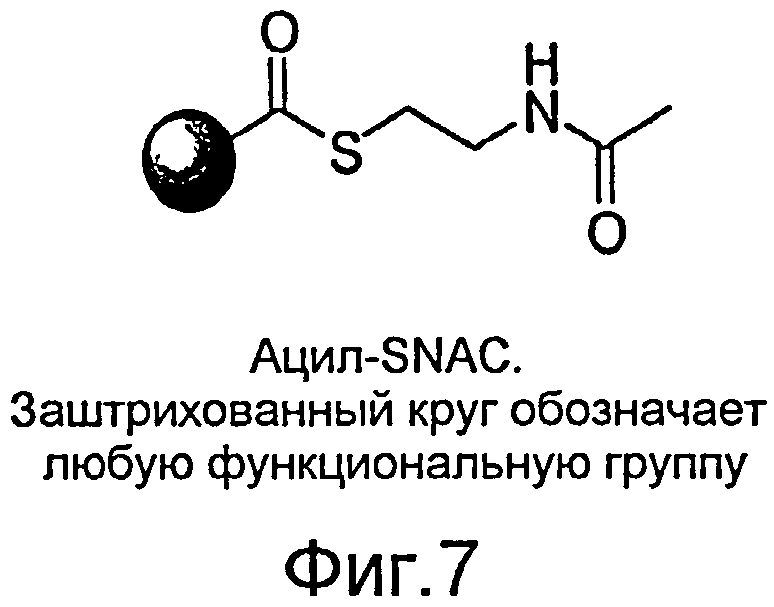

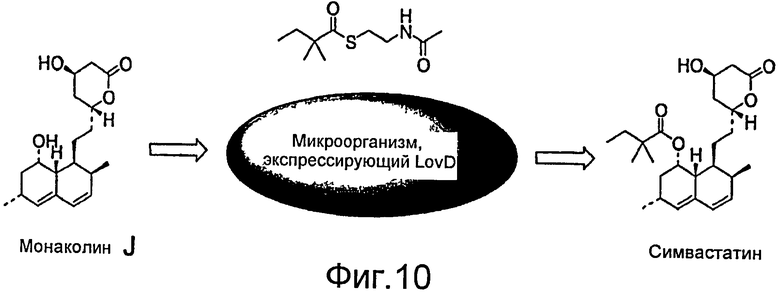
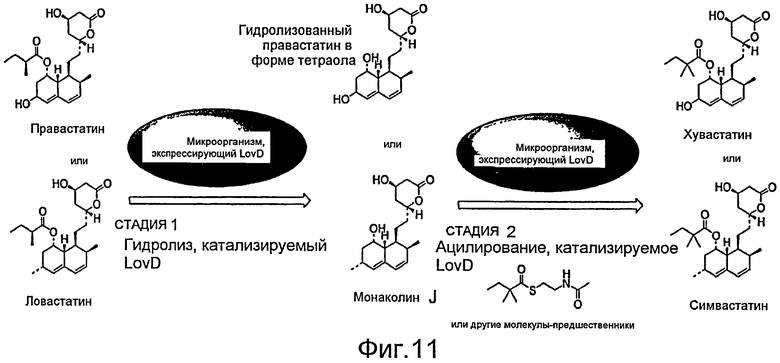
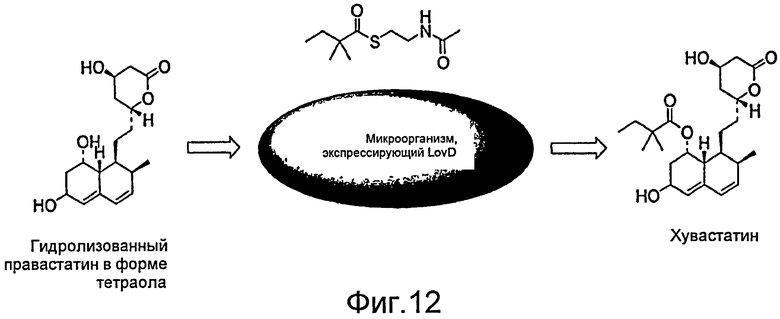
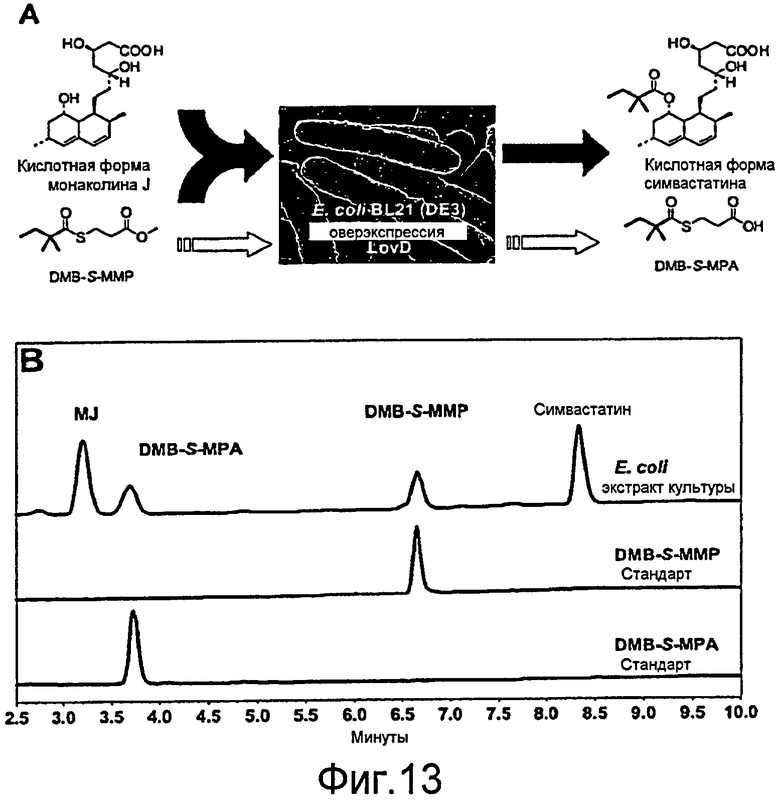
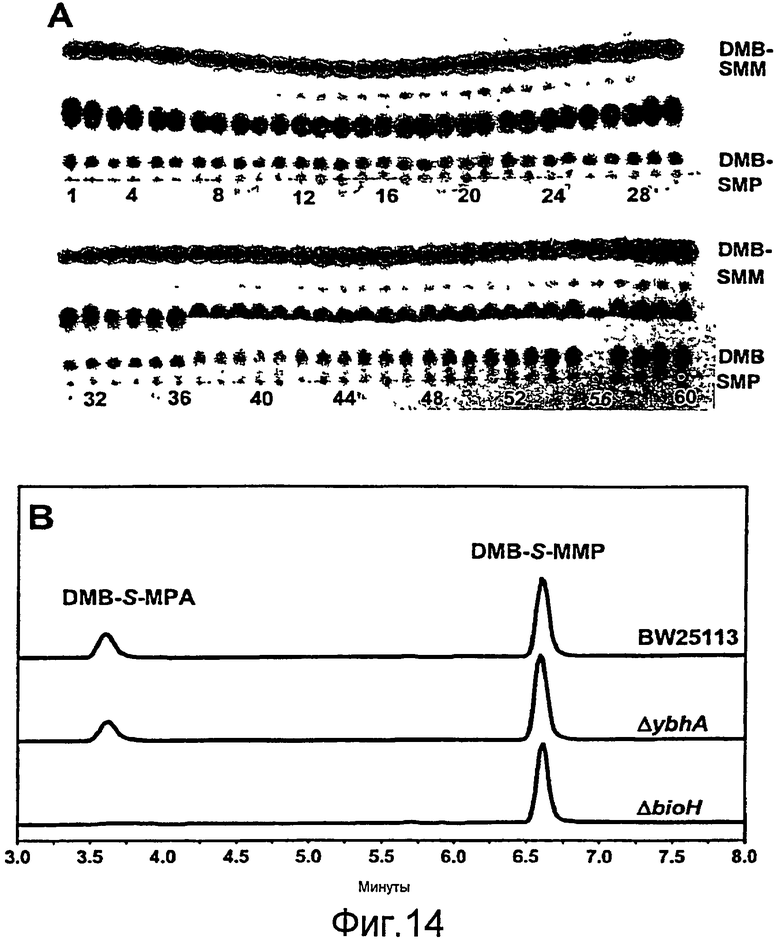
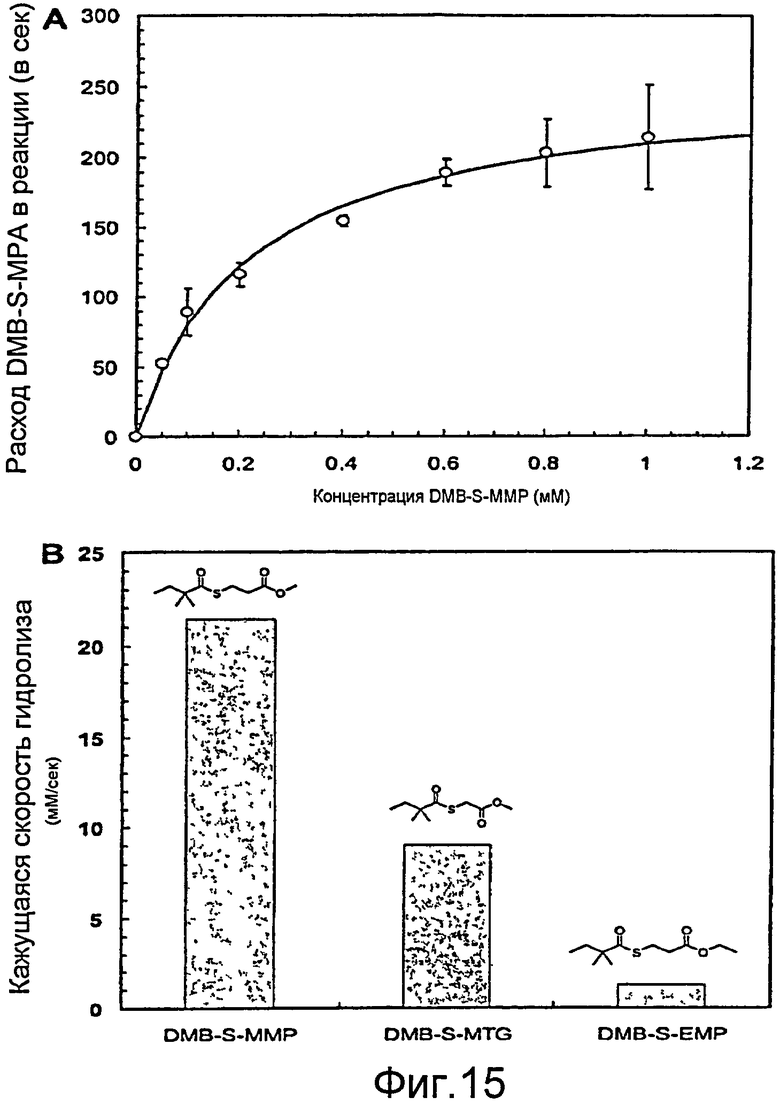
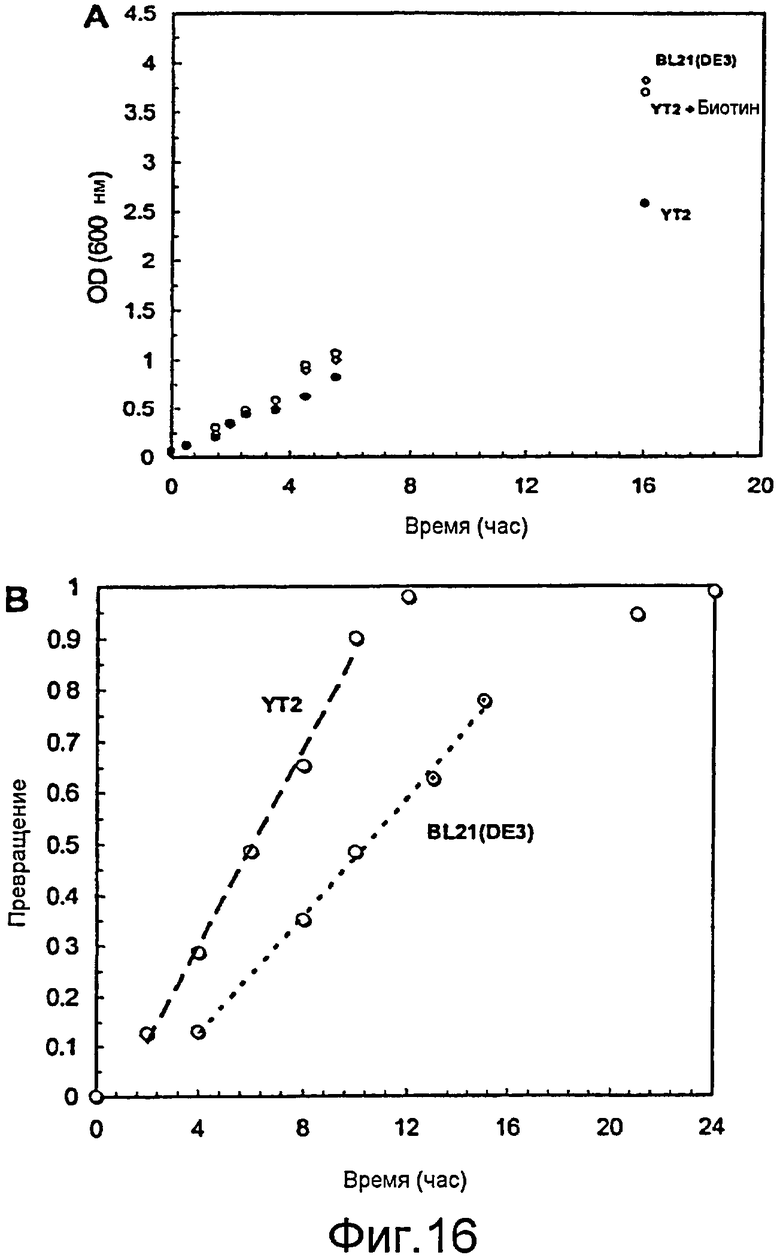
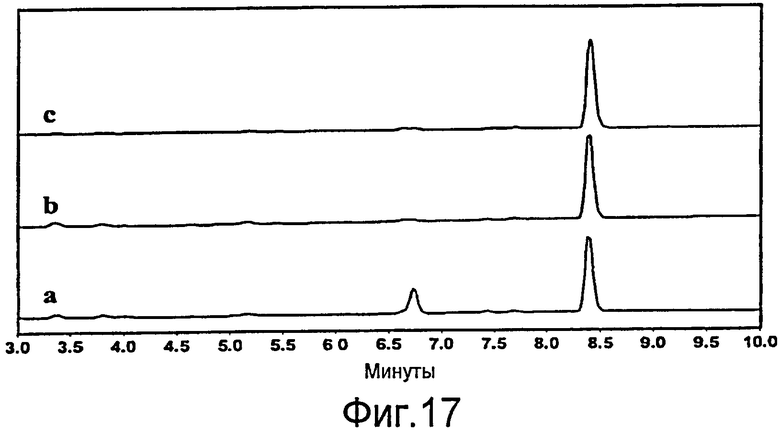
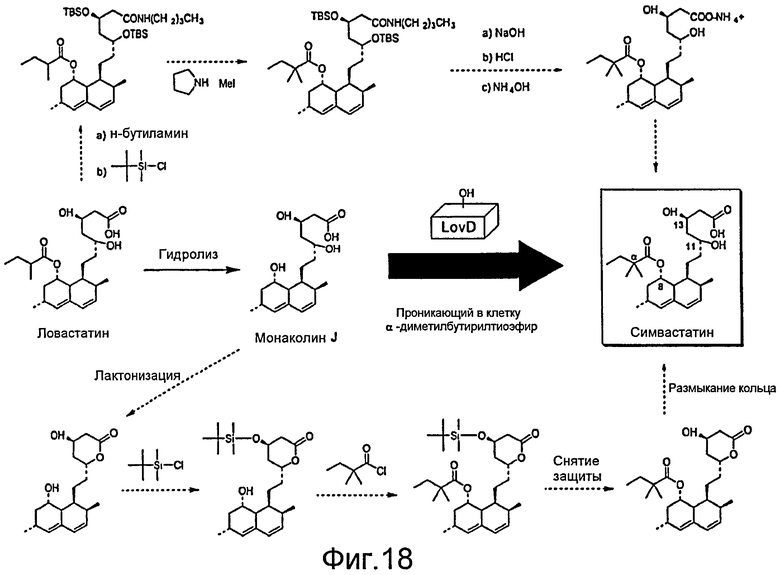
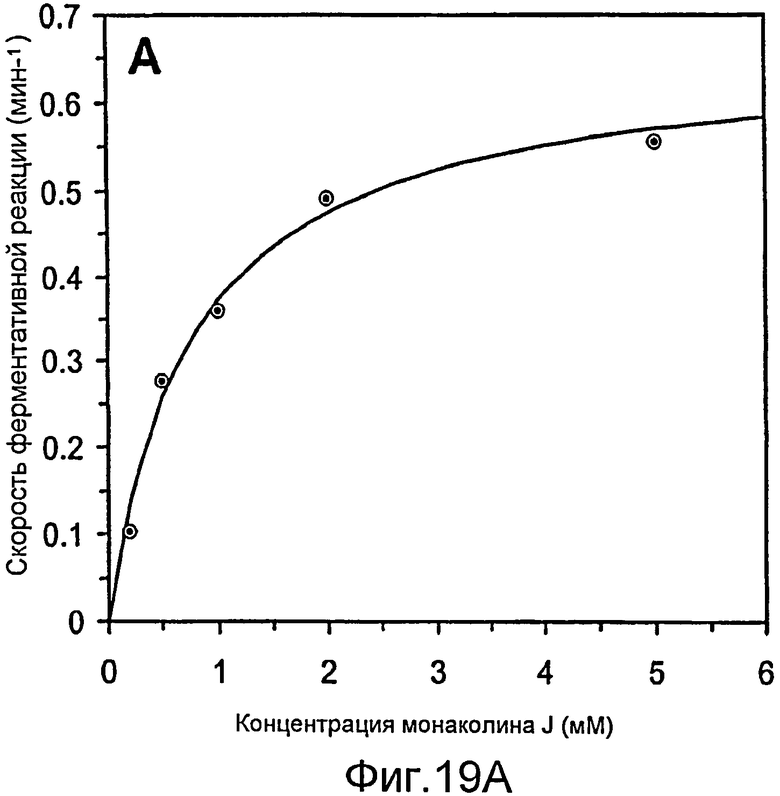

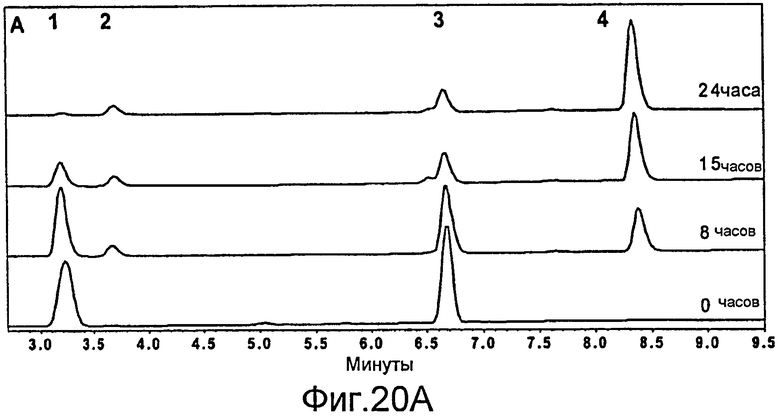
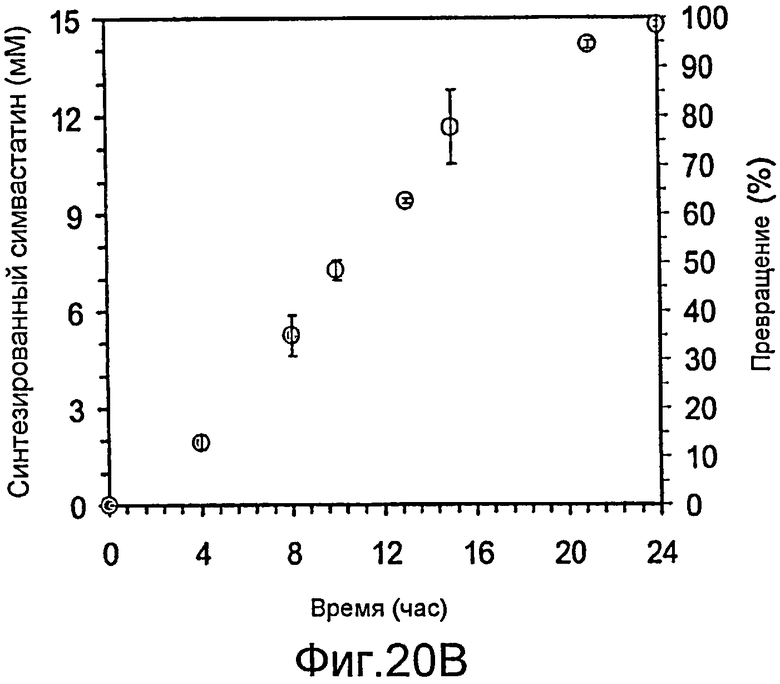
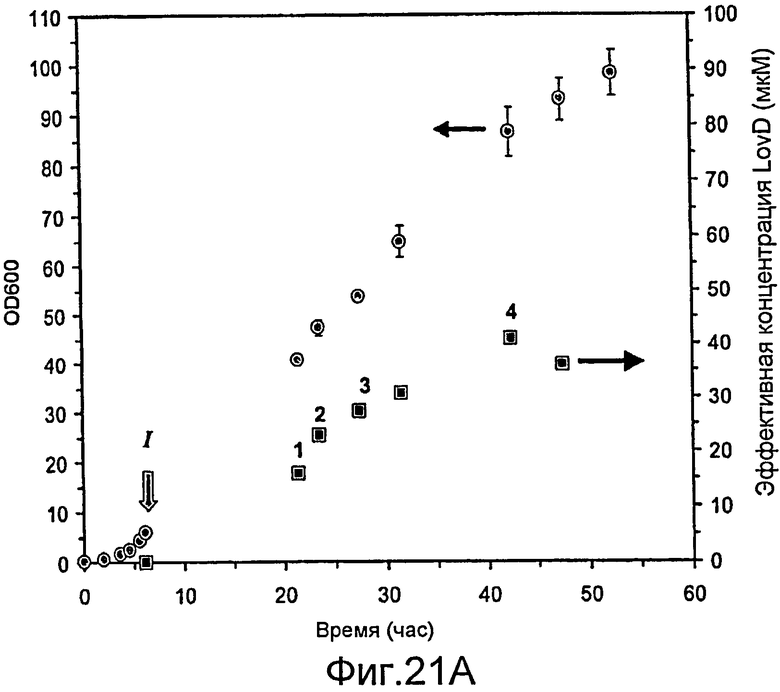
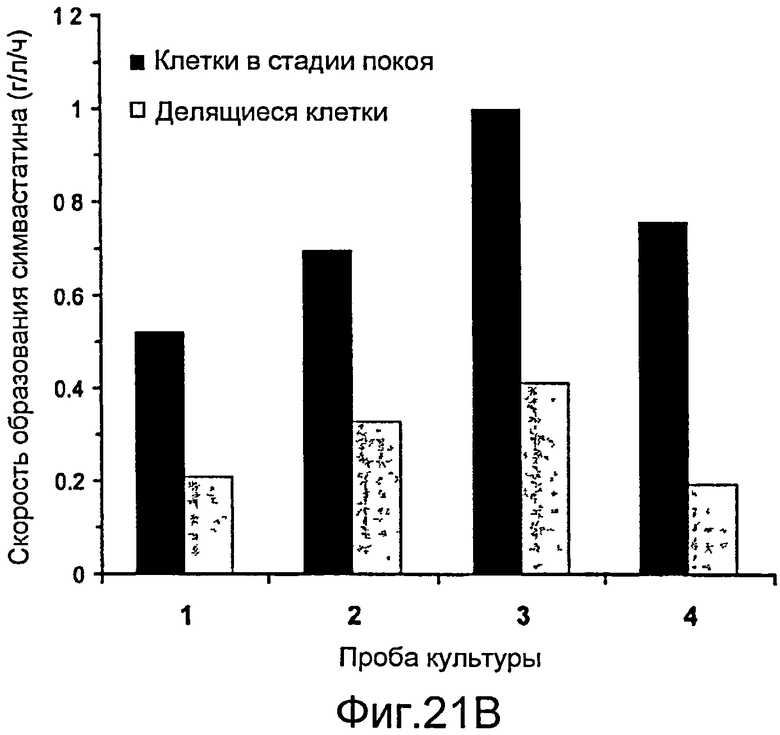
Изобретение относится к биотехнологии. Описаны способы получения симвастатина. В одном варианте способ осуществляют путем взаимодействия монаколина J и ацилтиоэфира, который отдает ацильную группу С8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD и региоселективного ацилирования С8-гидроксильной группы монаколина J ацилтрансферазой LovD, использующей ацильную группу ацилтиоэфира. Во втором варианте - путем взаимодействия ловастатина и ацилтиоэфира, который отдает ацильную группу С8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD и ацилтрансферазы LovD и воздействия ацилтрансферазы LovD, приводящего к гидролизу ловастатина до монаколина J и использованию ацильной группы ацилтиоэфира для региоселективного ацилирования С8-гидроксильной группе монаколина J. Изобретение расширяет арсенал средств получения симвастатина. 2 н. и 10 з.п. ф-лы, 24 ил., 5 табл., 4 пр.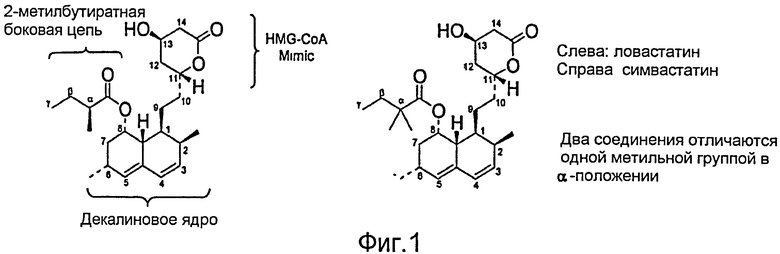
1. Способ получения симвастатина, включающий стадии:
(1) взаимодействия монаколина J; ацилтиоэфира, который отдает ацильную группу С8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD; и
(2) региоселективного ацилирования С8-гидроксильной группы монаколина J ацилтрансферазой LovD, использующей ацильную группу ацилтиоэфира, с получением симвастатина.
2. Способ по п.1, где симвастатин получают in vitro в отсутствии выделенного организма.
3. Способ по п.1, где монаколин J; ацилтиоэфир и ацилтрансфераза LovD взаимодействуют в среде культивирования в присутствии выделенного организма, который продуцирует ацилтрансферазу LovD, и где дополнительно организм представляет собой Escherichia coli, Aspergillus terreus, Monascus ruber, Monascus purpureus, Monascus pilosus, Monascus vitreus, Monascus pubigerus, Candida cariosilognicola, Aspergillus oryzea, Doratomyces stemonitis, Paecilomyces virioti, Penicillum citrinum, Penicillin chrysogenum, Scopulariopsis brevicaulis или Trichoderma viride.
4. Способ по п.3, где выделенный организм представляет собой, по меньшей мере, один из:
(a) Aspergillus terreus, который экспрессирует полипептид LovD SEQ ID NO:1;
(b) Aspergillus terreus, который не экспрессирует полипептид LovF SEQ ID NO:3;
(c) Escherichia coli, которая экспрессирует полипептид LovD SEQ ID NO:1; или
(d) Escherichia coli, которая не экспрессирует полипептид bioH SEQ ID NO:4.
5. Способ по п.1, где ацилтрансфераза LovD имеет аминокислотную последовательность SEQ ID NO:1 или SEQ ID NO:2.
6. Способ по п.1, где выбранный ацилтиоэфир обладает, по меньшей мере, одним из следующих свойств:
(a) является бутирил-тиоэфиром, N-ацетилцистаминтиоэфиром или метилтиогликолевым тиоэфиром;
(b) включает ацильную группу с цепью средней длины (С3-С6);
(c) способен проходить через мембраны клеток Escherichia coli или Aspergillus terreus, растущих в среде культивирования; или
(d) выбран из группы, состоящей из α-диметилбутирил-S-метил-меркаптопропионата (DMB-S-MMP), диметилбутирил-S-этилмеркаптопропионата (DMB-S-EMP) и диметилбутирил-S-метилтиогликолята (DMB-S-MTG) и диметилбутирил-S-метилмеркаптобутирата (DMB-S-MMB).
7. Способ по п.3, где ацилтиоэфир представляет собой α-диметилбутирил-S-метил-меркаптопропионат, и где α-диметилбутирил-S-метил-меркаптопропионат присутствует в среде культивирования в диапазоне концентраций 1-100 мМ.
8. Способ по п.3, где монаколин J продуцируется выделенным организмом в среде культивирования.
9. Способ по п.3, где выделенный организм выращивают, по меньшей мере, при одном из следующих условий:
(a) при температуре в диапазоне 30-40°С,
(b) в течение периода времени, по меньшей мере, от 4 до, по меньшей мере, 48 ч,
(c) при рН 7-8; или
(d) в среде культивирования, включающей LB, F1 или ТВ среды.
10. Способ по п.1, дополнительно включающий очистку симвастатина, полученного указанным способом, посредством, по меньшей мере, одной стадии очистки, включающей:
(a) лизис клеток выделенного организма, присутствующего в смеси,
(b) центрифугирование,
(c) осаждение свободной кислотной формы симвастатина;
(d) превращение свободной кислотной формы симвастатина в соль симвастатина;
(e) фильтрацию; или
(f) высокоэффективную жидкостную хроматографию (ВЭЖХ).
11. Способ по п.1, где способ:
(a) обеспечивает получение композиции веществ, включающей 0-1% монаколина J, который первоначально взаимодействовал с ацилтиоэфиром, который отдает ацильную группу С8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и/или
(b) приводит к превращению, по меньшей мере, 95% монаколина J, добавленного к смеси, в симвастатин.
12. Способ получения симвастатина, включающий стадии:
(1) взаимодействия ловастатина; ацилтиоэфира, который отдает ацильную группу С8-гидроксильной группе монаколина J в присутствии ацилтрансферазы LovD; и ацилтрансферазы LovD; и
(2) воздействия ацилтрансферазы LovD, приводящего к:
(a) гидролизу ловастатина до монаколина J; и
(b) использованию ацильной группы ацилтиоэфира для региоселективного ацилирования С8-гидроксильной группе монаколина J;
с получением симвастатина.
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| СПОСОБЫ ПОЛУЧЕНИЯ СИМВАСТАТИНА ВЫСОКОЙ СТЕПЕНИ ОЧИСТКИ И ПОЛУСИНТЕТИЧЕСКИХ СТАТИНОВ | 2001 |
|
RU2275909C2 |

