Настоящее изобретение относится к области биологии. В частности, настоящее изобретение относится к области направления экспрессии генов, а более конкретно к конструированию и разработке новой системы специфической экспрессии трансгенов. Объектом настоящего изобретения являются, в частности, новые промоторные последовательности, способные регулировать уровень и специфичность экспрессии трансгена in vivo в клетках сердечной мышцы. В настоящем изобретении также описаны новые композиции, конструкции и векторы, которые позволяют регулировать и направлять экспрессию нуклеиновой кислоты в клетках сердечной мышцы. Настоящее изобретение может иметь множество применений, например, в области экспериментальной и клинической медицины, а также в области терапии и диагностики, а более конкретно, оно может быть использовано для лечения и/или предупреждения некоторых патологий сердца.
Регуляция уровня и направленности экспрессии трансгенов необходима для достижения многих целей. Так, например, в генотерапии для успешного лечения может потребоваться осуществление направленности транспорта белка, синтезированного из трансгена, что позволяет тем самым ограничить появление побочных эффектов. Конструирование трансгенных животных и исследование эффектов гена являются примерами, в которых может быть использована соответствующая регуляция специфичности экспрессии белка и может быть достигнуто ее увеличение.
Исходя из этого было протестировано множество промоторов на их способность регулировать кардиоспецифическую экспрессию. Такими промоторами, в частности, являются промоторы генов, кодирующих: легкую цепь сердечного миозина (MLC-2) крыс (Henderson S.A. et al., J.Biol. Chem. 264 (1989) 18142-8; Lee K.J. et al., J.Biol.Chem, 126 (1992) 15875-85), сердечный α-актин мышей (Biben C. et al., Dev Biol, 173 (1996) 200-12), натрийуретический фактор (ANF) (Harris A.N. et al., J. Mol. Cell. Cardiol, 29 (1997) 515-25), тяжелую цепь α- или β-миозина (α- или β-МНС) (Colbert M.C. et al., J. Clin. Invest, 100 (1997) 1958-68), мышечную креатинкиназу кроликов (МСК) (Vincent C.K. et al., Mol. Cell. Biol, 13 (1993) 567-74) или сердечный тропонин Т (патент США № 5266488).
Хотя известно, что указанные промоторы обеспечивают определенную степень специфичности к ткани, однако известно также, что уровни их активности значительно ниже, примерно в 10 и 100 раз, чем уровни активности так называемых сильных промоторов, а поэтому, фактически, они не могут быть использованы в терапевтических целях.
Так, например, Franz W.M. et al. (Cardiovasc. Res, 35 (1997) 560-6) и Griscelli F. et al. (C.R. Acad Sci III, 320 (1997) 103-12) показали, что уровни активности последовательностей, расположенных выше (положение "апстрим") генов, кодирующих крысиные α-МНС и MLC-2, в аденовирусных конструкциях остаются значительно ниже, примерно в 10 раз, чем уровни промотора RSV (вируса саркомы Рауса).
Поэтому, более конкретно, настоящее изобретение относится к новой промоторной последовательности, происходящей от области, расположенной выше (положение "апстрим") гена CARP (сердечного белка анкерина с повторяющимися доменами). Эта последовательность не только способна направлять кардиоспецифическую экспрессию, но также способна обеспечивать высокий уровень экспрессии in vivo, сопоставимый с уровнем экспрессии сильного промотора, такого как промотор CMV (цитомегаловируса).
Были проведены исследования белка CARP, который представляет собой один из первых маркеров для дифференциации кардиомиоцитов, действующих ниже (положение "даунстрим") гена гомеобокса Nbx2.5 в регуляции экспрессии гена MLC-2v, и была секвенирована кодирующая последовательность этого гена у мышей (Zou Y. et al., Development, 24 (1997) 794-804), у кроликов (Aihara Y. et al., Biochim Biophys Acta, 28 (1999) 318-24) и у человека (Chu W. et al., J. Biol. Chem. 270 (1995) 10236-45).
Kuo H. et al. (Development, 126 (1999) 4223-34) клонировали 10 т.п.н.-фрагмент и секвенировали 2,5 т.п.н.-фрагмент, расположенный выше кодирующей последовательности мышиного гена CARP. В этом фрагменте у 5'-конца была сделана делеция, в результате чего было показано, что область промотора, состоящая из 213 п.н. и находящаяся между нуклеотидами -166 и +47 относительно положения транскрипции +1, является достаточной для осуществления кардиоспецифической экспрессии in vitro, что, вероятно, указывает на присутствие у 5'-конца элемента, регулирующего специфичность данного промотора. Kuo H. et al. также получили трансгенные линии мышей, несущих фрагмент в 2,5 т.п.н., расположенный выше гена CARP и обеспечивающий специфическую экспрессию трансгена в клетках сердечной и скелетной мышцы на ранней стадии развития эмбриона, а затем эта экспрессия ингибируется в процессе развития.
В заявке WO 00/15821 описана часть последовательности, расположенная выше (со стороны 5') кодирующей последовательности мышиного гена CARP и находящаяся между нуклеотидами -2285 и +62 относительно положения транскрипции +1. Эта последовательность, в частности, была оценена на ее in vivo активность с помощью аденовирусных векторов. Однако уровни наблюдаемой активности были настолько низки, что был сделан вывод, что для детекции активности in vivo необходимо выделить промоторную последовательность между двумя инвертированными концевыми повторами адено-ассоциированного вируса (AAV-ITR).
Заявители попытались более точно охарактеризовать 5'-область гена, кодирующего белок CARP. Таким образом, они идентифицировали новую последовательность, расположенную выше указанного гена CARP, и продемонстрировали неожиданные и преимущественные свойства этой новой последовательности, а именно ее способность значительно увеличивать уровни активности in vivo.
Действительно, заявителями было неожиданно обнаружено, что хотя эта только что идентифицированная последовательность не обеспечивала значительной экспрессии in vitro, однако, в противоположность этому, она обеспечивала очень высокие уровни активности in vivo, эквивалентные уровням активности так называемых сильных промоторов, сохраняя при этом высокий уровень селективности экспрессии в сердечной ткани.
Поэтому объектом настоящего изобретения является полинуклеотид, содержащий часть, расположенную выше кодирующей последовательности гена белка CARP, или часть, гибридизующуюся в условиях высокой жесткости с указанной вышерасположенной последовательностью, причем указанный полинуклеотид обладает способностью индуцировать специфическую экспрессию трансгена в сердечной ткани, находящегося под его контролем.
Настоящее изобретение также относится к любому природному полинуклеотиду или полинуклеотиду, полученному путем химического синтеза и являющемуся, по крайней мере, на 93%, а предпочтительно, по крайней мере, на 95% идентичным последовательности SEQ ID NO:1. Еще более предпочтительно, чтобы полинуклеотид настоящего изобретения был, по крайней мере, на 98% идентичен последовательности SEQ ID NO:1.
Понятие "природный полинуклеотид" означает фрагмент геномной ДНК, полученный путем расщепления клеточной ДНК рестриктирующим ферментом.
Понятие "полинуклеотид, полученный методом химического синтеза" означает ДНК-фрагмент, созданный с помощью автоматического синтеза, например с использованием подходящего автоматического устройства.
В соответствии с настоящим изобретением термин "условия высокой жесткости" используется в том значении, как он был определен в руководстве Maniatis et al., 1982 (Molecular Cloning, A Laboratory manual, Cold Spring Harbor, CSH, N.Y, USA) или в одном из его последних изданий. Так, например, для элиминации негибридизованных фрагментов необходимы следующие условия гибридизации: три промывки при 65°С в присутствии 0,2% SSC (стандартный солевой раствор) и 0,1% ДСН.
"Специфический" характер экспрессии означает, что активность промотора в клетках сердечной ткани является значительно более высокой. Хотя неспецифическая экспрессия может происходить и в других клетках, однако в этих клетках, по сравнению с уровнем активности, наблюдаемым в клетках сердца, соответствующий уровень активности остается, в основном, очень низким (пренебрежимо малым), обычно ниже, по крайней мере, в 10 раз.
Результаты, представленные в примерах, показали, что в этом случае разница в экспрессии, которая в клетках сердца может достигать 1000-кратного увеличения, указывает на высокую селективность полинуклеотидов настоящего изобретения in vivo.
Кроме того, результаты, представленные в нижеследующих примерах, ясно указывают на то, что полинуклеотиды настоящего изобретения представляют собой систему, обеспечивающую высокую степень экспрессии, превышающей экспрессию, наблюдаемую в присутствии других промоторов, которые, как известно, являются специфическими по отношению к сердечной ткани, и в этом случае разница в экспрессии составляет более чем в 100 раз. Следовательно, указанные элементы иллюстрируют преимущества и неожиданные свойства полинуклеотида настоящего изобретения с точки зрения их активности и специфичности в отношении экспрессии нужных нуклеиновых кислот в ткани сердца. Полинуклеотид настоящего изобретения, предпочтительно, содержит часть последовательности, расположенной между -2266 и +92 относительно положения транскрипции +1, и эта часть последовательности представлена в приложении как SEQ ID NO:1.
Следовательно, объектом настоящего изобретения являются последовательности, гибридизующиеся в условиях высокой жесткости с последовательностью SEQ ID NO:1.
При этом настоящее изобретение не ограничивается полинуклеотидами, содержащими фрагменты, расположенные выше мышиного гена, и оно относится к любому функциональному варианту или к какой-либо другой последовательности любого другого вида, имеющей аналогичные свойства, а именно способность специфически индуцировать экспрессию трансгена in vivo в ткани сердца.
Таким образом, каждый специалист в этой области может преимущественно использовать последовательность, расположенную выше человеческого гена, депонированного в Genbank под номером AF131884, и эта последовательность представлена в приложении как SEQ ID NO:2. Поэтому настоящее изобретение охватывает любую последовательность, содержащую фрагменты последовательностей, расположенных выше гена белка CARP, которые были модифицированы, например, путем делеции определенных структур и которые сохраняют при этом идентичные или аналогичные функции последовательности SEQ ID NO:1.
Предпочтительно, чтобы полинуклеотид настоящего изобретения был, по крайней мере, на 80%, а более предпочтительно, по крайней мере, на 90% идентичен последовательности SEQ ID NO:2.
Термин "функциональный вариант" означает модифицированную последовательность, сохраняющую свойства полинуклеотидов, упомянутых выше. Указанными модификациями в рассматриваемых последовательностях могут быть одно или несколько добавлений, мутаций, делеций и/или замен нуклеотидов. Эти модификации могут быть введены стандартными методами молекулярной биологии, такими как, например, сайт-направленный мутагенез или, в более конкретном применении, искусственный синтез данной последовательности на синтезаторе. Полученные варианты тестируют на их способность регулировать специфичность экспрессии в клетках сердечной мышцы по сравнению со специфичностью полинуклеотида, имеющего последовательность SEQ ID NO:1.
В другом своем аспекте настоящее изобретение относится к экспрессирующей кассете, содержащей определенный выше полинуклеотид, функционально присоединенный к трансгену так, чтобы экспрессия этого трансгена специфически направлялась в сердечную мышцу.
При этом, предпочтительно, чтобы кассета настоящего изобретения, кроме того, содержала сигнал терминации транскрипции, расположенный со стороны 3'-конца от нуклеотидной последовательности этого трансгена.
Предпочтительно также, чтобы указанный трансген содержал нуклеиновую кислоту, представляющую терапевтическую ценность и кодирующую белок, или РНК, которые могут быть ассоциированы с сердечно-сосудистыми патологиями, такими как сердечная недостаточность, гипертрофия сердца, гипоксия, ишемия или отторжения сердечного трансплантата.
В качестве примеров белка, представляющего интерес для терапии, могут быть упомянуты, inter alia:
- белки, индуцирующие ангиогенез, такие как, например, члены семейства VEGF, члены семейства FGF, а более конкретно FGF1, FGF2, FGF4, FGF5, ангиогенин, EGF, TGFα, TGFβ, TNFα, фактор рассеяния/HGF, члены семейства ангиопоэтинов, цитокины, а в частности интерлейкины, включая IL-1, IL-2, IL-8, ангиотензин-2, активатор плазминогена (ТРА), урокиназу (uPA) и молекулы, участвующие в синтезе активных липидов (простагландины, Сох-1);
- белки, участвующие в регуляции сердечных сокращений, такие как фосфоламбан, ингибиторы фосфоламбана, SERCA-2a, β2-адренергический рецептор или дистрофин или минидистрофин (FR 91 11947);
- белки с криопротективной активностью, которые, в частности, блокируют апоптоз, например такие белки, которые являются членами семейства bcl, и протеинкиназы, такие как АКТ/РКВ;
- факторы транскрипции, такие как, например, природные или химерные ядерные рецепторы, содержащие ДНК-связывающий домен, лиганд-связывающий домен и домен активации или ингибирования транскрипции, такие как, например, гибридные белки tetR-NLS-VP16; гибридные белки, происходящие от рецепторов эстрогена; гибридные белки, происходящие от рецепторов стероидных гормонов; гибридные белки, происходящие от рецепторов прогестерона; белки системы CID (химерный индуктор димеризации), описанные Rivera et al. (Rivera et al., Nature Medicine, 2 (1996) 1028-1032). Более конкретно, в качестве примеров химерных ядерных рецепторов могут быть упомянуты ядерные рецепторы PPAR (активируемый пролифератором рецептор пероксисомы), а в частности, PPARγ2, описанный в заявках WO 96/23884 и FR 99 07957 и в работе Frohnert et al., (J. Biol. Chem. 274 (1999) 3970-3977) и Mukherjee et al., (J. Biol. Chem. 272 (1997) 8071-8076), либо в его нативной форме, не содержащей модификации первичной структуры, либо модифицированный PPARγ2, содержащий один или несколько лиганд-связывающих сайтов или Е/F-доменов (Schoonjans et al. Biochem. Biophys. Acta. 1302 (1996), 93-109), такой как PPARγ2γ2, имеющий последовательность SEQ ID NO:3;
- иммуносупрессоры, такие как, например, интерлейкины-2 и -10, которые, возможно, полностью или частично ингибируют путь передачи иммунного сигнала и таким образом увеличивают срок жизни трансплантатов сердца;
- белки, действующие как агенты для снижения гипоксии, такие как NOS (нитрооксидсинтетаза), белок В-клеточной лейкемии/лимфомы 2 (bcl-2), супероксид-дисмутаза (СОД) и каталаза.
В качестве примеров РНК, представляющей интерес с точки зрения терапии, могут быть упомянуты антисмысловые РНК, которые могут быть использованы для регуляции экспрессии генов или транскрипции клеточных мРНК и, следовательно, для блокирования трансляции белка методом, описанным в патенте ЕР 140308; а также рибозимы, способные селективно разрушать РНК-мишени, как описано в ЕР 321201.
При этом следует отметить, что настоящее изобретение не ограничивается конкретными примерами белков или РНК и каждый специалист может использовать их для осуществления экспрессии любой нуклеиновой кислоты в клетках сердца с применением простых операций рутинного экспериментирования.
Объектом настоящего изобретения являются также векторы, содержащие полинуклеотид или экспрессирующую кассету настоящего изобретения. Такой вектор может содержать любую другую ДНК-последовательность, необходимую для экспрессии трансгена в нужных тканях, а в частности, он может содержать сайт инициации репликации, который является эффективным в клетках сердца.
Вектор настоящего изобретения может иметь различную природу и/или происхождение, а в частности, он может быть плазмидным, эписомным, хромосомным, вирусным или фаговым вектором. Предпочтительным вектором является плазмида или рекомбинантный вирус.
В качестве иллюстративных примеров плазмид, содержащих полинуклеотид или экспрессирующую кассету, могут быть упомянуты, например, плазмиды, pXL3634, pXL3728 и pXL3759, которые будут описаны ниже.
В соответствии с первым вариантом осуществления изобретения векторами настоящего изобретения являются векторы плазмидного типа. В качестве примеров плазмидных векторов могут быть упомянуты, inter alia, любые клонирующие или экспрессирующие плазмиды, которые известны специалистам и которые обычно включают сайты инициации репликации. Могут быть также упомянуты плазмиды новой генерации, несущие сайты инициации репликации и/или маркеры, которые были точно определены и описаны, например, в заявке WO 96/26270.
В соответствии с предпочтительным вариантом осуществления изобретения плазмидный вектор представляет собой миниплазмиду и содержит сайт инициации репликации, причем этому вектору для его функционирования в клетке-хозяине требуется присутствие, по крайней мере, одного белка, являющегося специфичным и чужеродным для указанной клетки. Такие клетки описаны, в частности, в заявке WO 97/10343.
В соответствии со вторым вариантом осуществления изобретения векторами настоящего изобретения являются вирусные векторы. В качестве примеров этих векторов могут быть упомянуты, inter alia, рекомбинантные аденовирусы, рекомбинантные аденоассоциированные вирусы, рекомбинантные ретровирусы, лентивирусы, герпесвирусы и вирус коровьей оспы, которые могут быть получены методами, известными специалистам. В качестве химерных вирусных векторов, предпочтительно, используются химерные векторы, полученные на основе аденовирусов-ретровирусов и описанные, inter alia, в заявке WO 95/22617, а также эписомные/аденовирусные векторы, которые были описаны Leblois et al. (Mol Ther (2000) 1(4), 314-322) и в заявке WO 97/47757.
Если в данном варианте осуществления изобретения используются аденовирусы, то предпочтительными являются векторы, полученные из дефектных аденовирусов, то есть такие векторы, которые неспособны к автономной репликации в клетке-мишени. Конструирование этих дефектных вирусов, а также их инфекционные свойства были широко описаны в литературе (см., в частности, S. Baeck & K.L. March, Circul. Research, 82, (1998) 295-305); T. Shenk, B.N. Fields, D.M. Knipe, P.M. Howley et al. (1996), Adenoviridae: Viruses and Replication (in Virology) 211-2148, EDS - Ravens publishers Philadelphia; Yeh, P. et al. FASEB 11 (1997) 615-623).
Были охарактеризованы различные серотипы аденовирусов, структуры и свойства которых до определенной степени варьируются. В соответствии с настоящим изобретением среди таких серотипов предпочтительно использовать аденовирус человека типа 2 или 5 (Ad 2 или Ad 5) или аденовирусы животных, такие как вирусы, описанные в заявке Франции 9305954, или аденовирусы смешанного типа. Из аденовирусов животных, подходящих для использования в настоящем изобретении, могут быть упомянуты аденовирусы собак, коров, мышей (Beard et al., Virology 75 (1990) 81), овец, свиней, птиц или обезьян. Предпочтительным аденовирусом животного является аденовирус собак, а более предпочтительным - аденовирус CAV2 (Manhattan или штамм А26/61), описанный в заявке WO 94/26914.
Дефектные аденовирусы настоящего изобретения на каждом из своих концов обычно содержат инвертированный концевой повтор (ITR), последовательность, обеспечивающую инкапсидацию вируса (Psi), ген Е1 и, по крайней мере, один из генов Е2, Е4 и L1-L5, который, кроме того, может быть инактивирован любым методом, известным специалистам (Levero et al., Gene, 101 (1991) 195, EP 185573; Graham, EMBO J. 3 (1984) 2917).
При этом, предпочтительно, чтобы аденовирус, используемый в настоящем изобретении, содержал делецию в области Е1 своего генома. Еще более предпочтительно, чтобы он содержал делецию в областях Е1а и Е1b. В качестве примера могут быть упомянуты делеции 454-3328, 383-3446 или 357-4020 (по сравнению с геномом Ad5).
В соответствии с предпочтительным вариантом рекомбинантный аденовирус, используемый в настоящем изобретении, кроме того, имеет в своем геноме делецию в области Е4. Более конкретно, делеция в области Е4 влияет на все открытые рамки считывания. В качестве конкретного примера могут быть упомянуты делеции 33466-35535 или 33093-35535. Другие типы делеций в области Е4 были описаны в заявках WO 95/02697 и WO 96/22378, которые вводятся в настоящее описание посредством ссылки.
Что касается адено-ассоциированных вирусов (AAV), то они представляют собой ДНК-вирусы относительно небольшого размера, которые интегрируются в геном инфицируемых ими клеток стабильным и сайт-специфическим способом. Они способны инфицировать клетки широкого ряда, не оказывая при этом какого-либо влияния на рост, морфологию или дифференцировку клеток. Кроме того, эти вирусы, очевидно, не причастны к возникновению патологий у человека. Геном AAV был клонирован, секвенирован и охарактеризован. Он включает примерно 4700 оснований и на каждом своем конце содержит инвертированный концевой повтор (ITR), состоящий примерно из 145 оснований и служащий в качестве сайта инициации репликации для данного вируса. Остальной геном разделен на 2 главные области, несущие функции инкапсидации: левая часть генома, которая содержит ген rep, ответственна за репликацию вируса и экспрессию вирусных генов; а правая часть этого генома содержит ген сар, кодирующий вирусные капсидные белки.
Использование AAV-векторов для переноса генов in vitro и in vivo описано в литературе (см., в частности, WO 91/18088; WO 93/09239; патент США № 4797368, патент США № 5139941, ЕР 488528). В этих заявках описаны различные AAV-конструкции, в которых гены rep и/или сар были делетированы и заменены нужным геном и их использование для переноса in vitro (на клетках в культуре) или in vivo (непосредственно в организме) указанного нужного гена. Дефектные рекомбинантные AAV настоящего изобретения могут быть получены путем ко-трансфекции плазмиды, содержащей последовательности нуклеиновой кислоты настоящего изобретения, фланкированные двумя инвертированными концевыми повторами AAV (ITR), и плазмиды, несущей гены инкапсидации (гены rep и сар), в клеточные линии, инфицированные человеческим вирусом-хелпером (например, аденовирусом). Затем полученные рекомбинантные AAV очищают стандартными методами.
В соответствии с настоящим изобретением могут быть также использованы лентивирусы, поскольку они способны обеспечивать перенос и стабильную интеграцию нужного гена в покоящиеся клетки.
В качестве примеров таких вирусов могут быть упомянуты HTLV-I лентивирусов животных, таких как FIV (вирус кошачьей инфекции), EIAV (вирус инфекционной анемии лошадей; WO 98/51810), BIV (вирус коровьего иммунодефицита), CAEV (вирус козьего артрита-энцефалита)(WO 98/39463; Naldini et al., Science 272 (1996) 263-267; Schnele et al., Hum Gen Ther 11 (2000) 439-447) или лентивирус ВИЧ-2, родственный вирусу, вызывающему СПИД, но не являющийся в высокой степени патогенным для человека (Kundra et al., Hum Gen Ther. 9 (1998) 1371-1380).
Экспрессирующие кассеты могут быть введены в различные сайты рекомбинантного генома. Они могут быть введены на уровне области Е1, Е3 или Е4 в качестве замены для ингибированных или избыточных последовательностей. Они могут быть также введены в любой другой сайт, находящийся за пределами последовательностей, необходимых для цис-активации продуцирования вирусов (последовательности ITR и последовательность инкапсидации).
Однако следует отметить, что введение последовательностей настоящего изобретения в вышеописанные векторы не является обязательным и клетки сердца могут быть непосредственно трансфицированы ДНК, содержащей эти последовательности.
Последовательности нуклеиновой кислоты настоящего изобретения могут быть введены путем ковалентного связывания с соединениями, стимулирующими их проникновение в клетки или их транспорт в ядро, при этом полученные конъюгаты могут быть, но необязательно, инкапсидированы в полимерные микрочастицы, как это описано в Международной заявке WO 94/27238.
В соответствии с другим вариантом осуществления изобретения последовательности нуклеиновой кислоты могут быть включены в систему трансфекции, содержащую полинуклеотиды, стимулирующие их проникновение в клетки, как это описано в Международной заявке WO 95/10534.
Указанные полинуклеотиды, кассеты и векторы могут быть введены in situ любым способом, известным специалистам, например путем коронарной инфузии (Barr et al., Gene Ther, 1, (1994) 51-58), интракардиальной инъекции, эпикардиальной инъекции, которая может быть осуществлена, например, через вентрикулярную стенку (Guzman et al., Cir Res, 73 (1993) 1202-1207), интраперикардиальной инъекции (Fromes et al., Gene Ther, 6 (1999), 683-688) или путем ретроинфузии коронарных вен (Boeckstegers et al., Circulation, 100 (Suppl.I (1999), I-815).
Указанные полинуклеотиды, кассеты и векторы могут быть преимущественно введены в форме содержащих их композиций, например, с помощью агента химического или биохимического переноса, облегчающего их трансфекцию в клетки сердца. Выражение "агент химического или биохимического переноса" означает любое соединение, облегчающее проникновение нуклеиновой кислоты в клетку. Такими агентами могут быть катионные агенты, такие как катионные липиды, пептиды, полимеры (полиэтиламин, полилизин) и наночастицы, либо некатионные агенты, такие как некатионные липосомы, некатионные наночастицы или полимеры. Указанные агенты хорошо известны специалистам и, в частности, описаны в заявках WO 95/18863, WO 97/18185 и WO 98/15639.
Кроме того, настоящее изобретение относится к лекарственным препаратам, содержащим указанные полинуклеотиды, экспрессирующие кассеты или векторы, а также к фармацевтическим композициям, содержащим указанные полинуклеотиды, экспрессирующие кассеты или векторы в фармацевтически эффективном количестве и фармацевтически приемлемые носители.
Указанные полинуклеотиды, кассеты и векторы могут быть преимущественно использованы в целях изготовления лекарственных препаратов для их доставки в сердечную ткань, в которой, в частности, может экспрессироваться ген, кодирующий нужный белок; и для лечения и/или предупреждения сердечной недостаточности, гипоксии, гипертрофии сердца, миокардита, ишемии сердца или для предупреждения отторжения трансплантата при пересадке сердца.
Указанные лекарственные препараты могут, например, включать кассету или вектор настоящего изобретения, который способен экспрессировать функциональную форму дефектного гена, соответствующую сердечной патологии, на которую направлено данное лечение.
Фармацевтическая композиция, предпочтительно, содержит фармацевтически приемлемые носители, подходящие для инъекций, а в частности, для внутрисердечных инъекций. Такими носителями могут быть, в частности, изотонические, стерильные физиологические растворы (мононатрийфосфата или динатрийфосфата, хлорида кальция или магния и т.п. или смесей указанных солей) или осушенные, а в частности лиофилизованные, композиции, из которых при добавлении стерилизованной воды или физиологического раствора, в зависимости от цели применения, можно получить растворы для инъекций. Могут быть использованы и другие носители, например гидрогель. Такой гидрогель может представлять собой любой биологически совместимый и нецитотоксичный (гомо или гетеро) полимер. Такие полимеры описаны, например, в заявке WO 93/08845. Некоторые из них, в частности такие, как полимеры, полученные из окиси этилена и/или пропилена, являются коммерчески доступными. Дозы, используемые для инъекций, могут быть скорректированы в соответствии с различными параметрами, а в частности в соответствии с преследуемыми целями (мечение, лечение патологий, скрининг и т.п.), например, в зависимости от экспрессируемого трансгена или продолжительности экспрессии нужного гена.
В общих чертах, рекомбинантные аденовирусы настоящего изобретения получают и вводят в дозах 104-1014 б.о.е., а предпочтительно 106-1010 б.о.е. Термин "б.о.е." (бляшкообразующая единица) соответствует инфекционной активности вирусного раствора и определяется инфекционностью соответствующей клеточной культуры, которая измеряется числом бляшек инфицированных клеток. Методы определения титра б.о.е. вирусного раствора хорошо известны специалистам.
Настоящее изобретение также относится к способу экспрессии трансгена, представляющего интерес с точки зрения терапии, где указанный способ предусматривает использование полинуклеотидов, кассет или векторов настоящего изобретения, так чтобы мог экспрессироваться указанный трансген.
Кроме того, настоящее изобретение также относится к любой клетке, модифицированной кассетой или вектором (а в частности, аденовирусом), описанным выше. Термин "модифицированная клетка" означает любую клетку, содержащую полинуклеотид или кассету настоящего изобретения. Эти клетки могут быть имплантированы в организм в соответствии с методикой, описанной в заявке WO 95/14785. Такими клетками являются, в основном, клетки человеческого сердца.
Настоящее изобретение также относится к трансгенным животным, а в частности к мышам, несущим полинуклеотид или кассету, определенные выше, в которых ген, кодирующий белок, представляющий терапевтическую ценность, был заменен репортерным геном. Такая трансгенная мышь может быть использована для скрининга молекул на их активность под действием регуляторных последовательностей гена, кодирующего белок CARP.
Эти молекулы могут быть введены мышам, а затем после их умерщвления могут быть получены гистологические срезы для идентификации тканей, окрашиваемых репортерным геном.
Трансгенные животные настоящего изобретения также являются объектом молекулярно-биологических исследований, проводимых в целях выяснения молекулярных механизмов, лежащих в основе наследственных патологий сердца, таких как сердечная недостаточность, гипертрофия сердца, гиперплазия сердца и инфаркт миокарда.
В качестве примера могут быть упомянуты мышиные модели для исследования миокардита, в которых был инактивирован ген, кодирующий интерферон-1 (IFN-1) (Aitken et al., Circulation, 90 (1994) 1-139).
Другие представляющие интерес животные-модели настоящего изобретения могут содержать полинуклеотид настоящего изобретения, присоединенный к трансгенам, например таким, как протоонкогены или онкогены, например, с-myc, для конструирования модели гиперплазии (Jackson et al., Mol.Cell.Biol, 10 (1990), 3709-3716); р21-ras для конструирования моделей вентрикулярной гипертрофии (Hunter et al., J. Biol. Chem., 270 (1995), 23176-23178), и ядерный антиген вируса Эпштейна-Барра для исследования некоторых кардиомиопатий (Huen et al., J.Gen.Virol, 74 (1993) 1381-1391). В соответствии с другим вариантом осуществления изобретения трансгенные животные настоящего изобретения представляют собой экспериментальные модели гипертрофии сердца и содержат экспрессирующую кассету, в которой данный трансген кодирует, например, кальмодулин (Gruver et al., Endocrinology, 133 (1993) 376-388), интерлейкин-6 или рецептор интерлейкина-6 (Hirota et al., Proc. Natl. Acad. Sci, 92 (1995), 4862-4866), кардиотропин-1 (Pennica et al., Proc. Natl. Acad. Sci, 92 (1995) 1142-1146) и, наконец, α-адренергический рецептор (Milano et al., Proc. Natl. Acad. Sci, 92 (1994) 10109-10113).
Кроме того, частью настоящего изобретения также являются полинуклеотиды настоящего изобретения, которые были модифицированы так, что они повышали уровень экспрессии гена CARP. Полученные таким образом трансгенные животные также являются экспериментальным инструментом для исследования инфаркта миокарда (Stanton et al., Circul Res, 86 (2000) 939-945).
Для осуществления настоящего изобретения каждый специалист может обратиться преимущественно к руководству "Sambrook et al. (Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York 1989) или к одному из его последних изданий.
Настоящее изобретение более подробно описано в нижеследующих примерах, которые приводятся лишь в иллюстративных целях и не ограничивают объема изобретения.
Описание графического материала
На фигуре 1 представлена нуклеотидная последовательность (SEQ ID NO:1) полинуклеотида, расположенного выше гена, кодирующего мышиный белок CARP;
На фигуре 2 представлена нуклеотидная последовательность (SEQ ID NO:2) полинуклеотида, расположенного выше гена, кодирующего человеческий белок CARP;
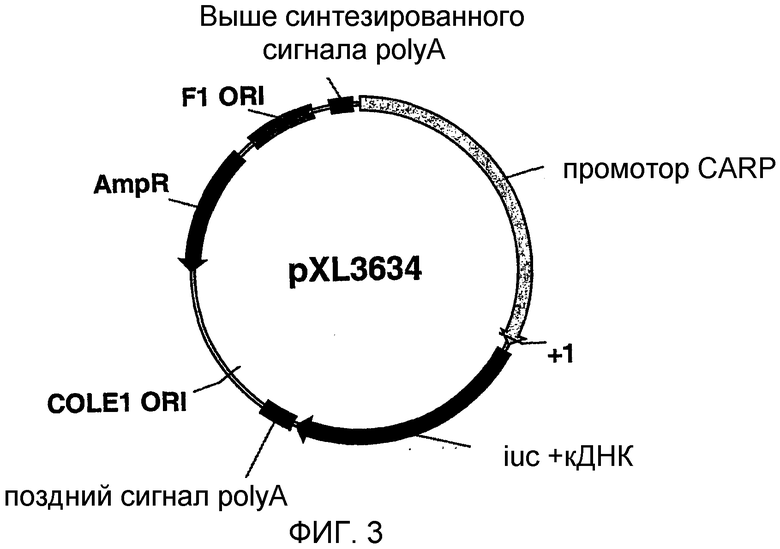
На фигуре 3 схематически показана плазмида pXL3634;
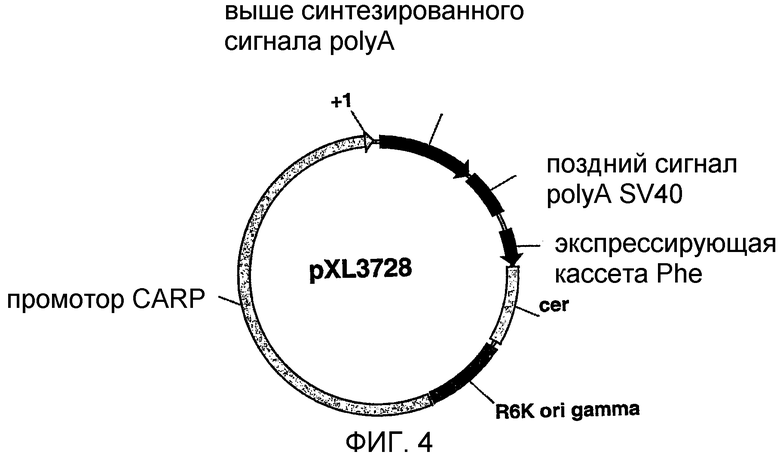
На фигуре 4 схематически показана плазмида pXL3728;
На фигуре 5 проиллюстрирована относительная активность in vitro плазмид pXL3635 и pXL3634 по сравнению с эталонной активностью промотора CMV (pRL-CMV). Активность каждого промотора представляет собой люциферазную активность Photinus pyralis, нормализованную по люциферазной активности Renilla reniformis.
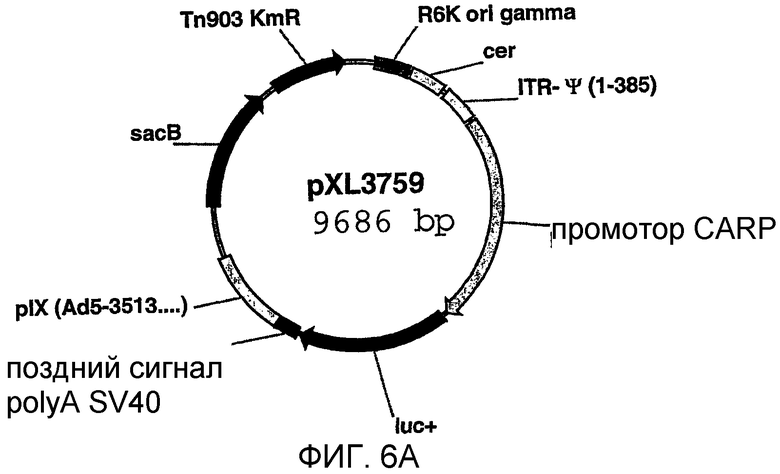
На фигуре 6А схематически показана плазмида pXL3759;
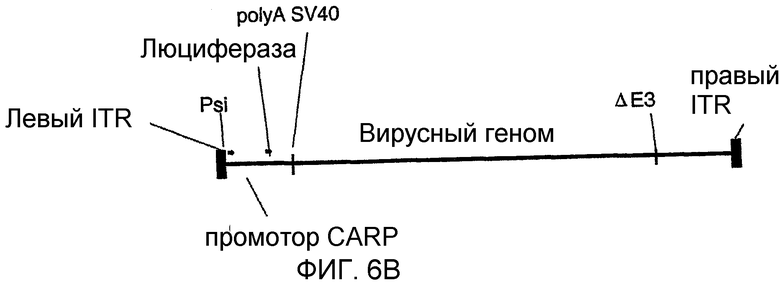
На фигуре 6В схематически показан аденовирус AV1.0 CARP-Luc+;
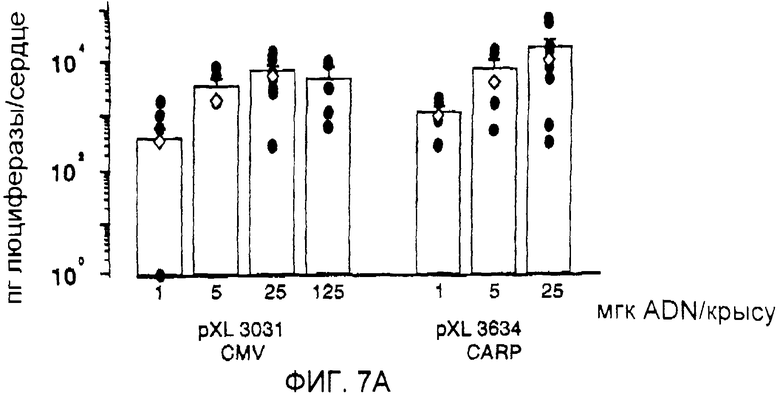
На фигуре 7А проиллюстрирована люциферазная активность (пг люциферазы/сердце) через 7 дней после введения крысам различных количеств плазмид pXL3031 и pXL3634 путем внутрисердечной инъекции через диафрагму;
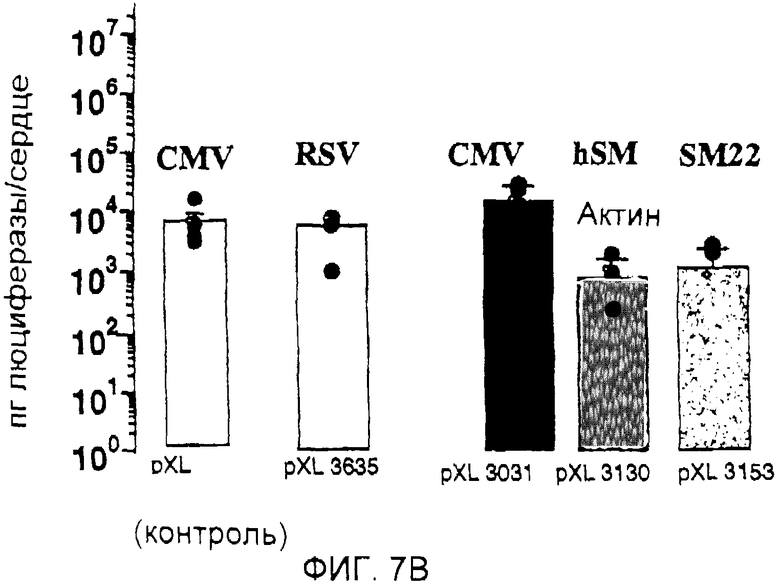
На фигуре 7В проиллюстрирована экспрессия люциферазы (пг люциферазы/сердце) через 7 дней после введения крысам 25 мкг плазмид pXL3031 и pXL3635, pXL3130 и pXL3153 путем внутрисердечной инъекции через диафрагму;
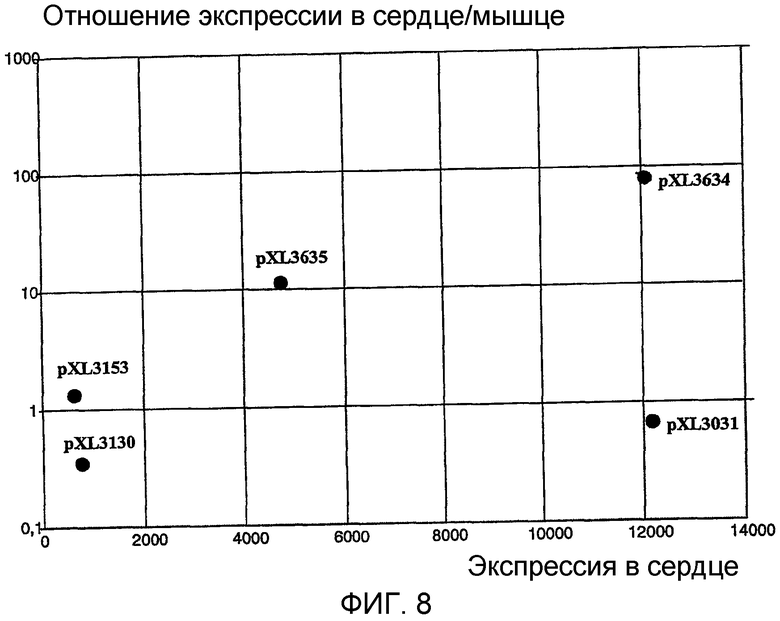
На фигуре 8 показано отношение экспрессии люциферазы в сердце к экспрессии люциферазы в мышце, которое определяет экспрессию в сердце, наблюдаемую после внутрисердечного введения плазмид pXL3031, pXL3634, pXL3635, pXL3153 и pXL3130.
Примеры
Пример 1: Характеризация полинуклеотида, расположенного выше гена CARP
BamHI-XhoI-фрагмент размером в 2,3 т.п.н. последовательности 5' мышиного гена, кодирующего белок CARP, клонировали и секвенировали по обеим цепям методом терминации цепи (Sanger et al., 1977, PNAS, 74, 5463) с использованием набора Sequenase Kit (United States Biochemical, Cleveland, Ohio). Эта последовательность представлена на фиг. 1 и содержит часть, расположенную выше гена, кодирующего мышиный белок CARP, между нуклеотидами -2266 и +92 относительно положения транскрипции +1 (SEQ ID NO:1).
Пример 2: Конструирование плазмидных векторов CARP
2.1 Плазмида pXL3634
BamHI-XhoI-фрагмент размером в 2,3 т.п.н., охарактеризованный в примере 1, клонировали после достраивания BamHI-сайта в плазмиду рGL3-Basic (Promega), предварительно гидролизованную ферментами XhoI и SmaI, и получали плазмиду pXL3634. Эта плазмида схематически представлена на фигуре 3.
2.2 Плазмида pXL3728
Плазмида pXL3728 была получена из плазмиды pXL3179, которая представляет собой вектор, происходящий от плазмиды pXL2774 (WO 97/10343), где ген, кодирующий гибрид между сигнальным пептидом интерферона человеческого фибробласта и кДНК FGF1 (фактор роста фибробластов-1)(sp-FGF1, Jouanneau et al., PNAS 88 (1991), 2893-2897), был введен под контроль промотора, полученного из ранней области цитомегаловируса человека (hCMV IE) и сигнала полиаденилирования поздней области вируса SV40 (Genbank SV4CG).
2,3 т.п.н.-BamHI-XhoI-фрагмент, который был охарактеризован в примере 1 и концы которого были достроены, клонировали в плазмиду рХL3179 (pCOR CMV-FGF), предварительно гидролизованную ферментами XbaI и EcoRI, и получали плазмиду pXL3728. Эта плазмида схематически представлена на фигуре 4.
2.3 Плазмида pXL3729
EcoRI-SalI-фрагмент плазмиды pXL3634 клонировали в плазмиду pXL3728, предварительно гидролизованную ферментами EcoRI-SalI, и получали плазмиду pXL3729.
Пример 3: Сравнительные плазмиды
3.1. Плазмиды pXL3130 и pXL3153
Плазмиды pXL3130 и pXL3153 содержат соответственно промотор α-актина гладкой мышцы человека (-680, +30) и промотор мышиного SM22 (-436, +43), присоединенный к энхансеру CMV (-522, -63), как описано в заявке WO 00/18908.
3.2. Плазмида pXL3635
Промотор RSV -229, +34 клонировали из конструкции, содержащей более длинный вариант промотора RSV (содержащегося в Ad1.0RSVLAcZ, Statford-Perricaudet et al., J.Clin. Invest. 90 (1992) 626-30), посредством ПЦР с использованием праймеров 5'-GGC GAT TTA AAT AAT GTA GTC TTA TGC AAT 3' и 5'-GGG GTC TAG AAG GTG CAC ACC AAT GTG GTG A-3', которые вводят соответственно SwaI и XbaI-сайт в 5' и 3' ПЦР-фрагмент. Затем эти два рестрикционных сайта использовали для введения промоторного фрагмента в основной рGL3 с получением pXL3635.
3.2. Плазмида pXL3031
Плазмида pXL3031 описана Soubrier et al., Gene Ther. 6 (1999), 1482-8. Эта плазмида представляет собой вектор, происходящий от плазмиды pXL2774 (WO 97/10343), в которой ген luc, кодирующий модифицированную люциферазу Photinus pyralis (цитоплазматическую), полученную из базового pGL3 (GenBank: CVU47295), вводили под контроль промотора, полученного из ранней области цитомегаловируса человека (hCMV IE, GenBank HS5IEE) и сигнала полиаденилирования поздней области вируса SV40 (Genbank SV4CG).
Пример 4: Клеточные культуры
Получали первичные культуры крысиных кардиомиоцитов. Для этого беременных крыс умерщвляли в камере, насыщенной CO2. После вскрытия брюшины рога матки удаляли и промывали в PBS при комнатной температуре. Эмбрионы удаляли из их оболочки и плаценту разрезали (10-12 эмбрионов на крысу). Сердце этих эмбрионов удаляли и промывали в смеси ADS/глюкоза. Предсердия и крупные сосуды удаляли под бинокулярной линзой, а затем сердца снова очищали в смеси ADS/глюкоза так, чтобы оставались только желудочки, после чего три раза промывали в стерильной смеси ADS/глюкоза.
Затем сердца трипсинизировали в 0,3 мл смеси ADS/глюкоза/трипсин на одно сердце с использованием трипсина Т 4674 (Sigma, St Louis, Missouri) в конечной концентрации 0,1 мг/мл в течение 20 минут при 37°С при мягком перемешивании (60-100 оборотов в минуту).Затем супернатант удаляли и трипсин инактивировали добавлением 1 мл декомплементированной FCS. После центрифугирования при 1500 об/мин в течение 10 минут супернатант удаляли и клетки сердца растворяли в 1 мл декомплементированной FCS. Параллельно стадии обработки трипсином повторяли 5-6 раз до тех пор, пока не происходила полная диссоциация клеток. Пул клеток центрифугировали при 1500 об/мин в течение 10 минут, а затем дважды промывали FCS, после чего клетки фильтровали на сетчатом фильтре.
Затем выделенные таким образом клетки помещали в культуру при концентрации 106 клеток/лунку в 24-луночный планшет или при концентрации 2х106 клеток/лунку в 12-луночный планшет. При этом каждая лунка содержала 1 мл культуральной среды.
Культуральная среда при полном объеме 100 мл содержала 68 мл DMEM (без пирувата) (Gibco BRL), 17 мл М199 (Sigma М 4530), 10 мл декомплементированной лошадиной сыворотки (Sigma Н6762), 5 мл декомплементированной смеси FCS (Gibco BRL) и 1 мл 100х смеси пенициллин/стрептомицин/глутамин (Peni/Strepto/glutamine Gibco BRL).
Кардиомиоциты культивировали в течение приблизительно 1 или 2 дней.
Пример 5: Трансфекция первичных культур кардиомиоцитов
Первичные культуры кардиомиоцитов котрансфицировали общим количеством ДНК, равным 500 мг на лунку и содержащим 1 нг плазмиды pRL-CMV (Promega Corp., Madison, WI), различные количества, варьирующиеся от 1 до 100 нг каждой из плазмид pXL3635 и pXL3634, описанных выше, и достаточное количество (до 500 нг) pUC19.
Для этого указанную смесь плазмид инкубировали вместе с 6 нмоль RPR 120535B (Byk et al., J. Med. Chem. 41 (1998) 229-35) на мкг ДНК (0,3 мкг раствора липида при 10 мМ) в конечном объеме 20 мкл в 150 мМ NaCl, 50 мМ бикарбоната, после чего интенсивно перемешивали в течение 5 минут и снова инкубировали в течение примерно 20-30 минут при комнатной температуре.
Затем полученную смесь добавляли к 250 мкл бессывороточной среды и инкубировали с клетками, по крайней мере, в течение 2 часов. И наконец, среду удаляли и клетки инкубировали в течение периода времени от 24 часов до 7 дней при температуре 37°С в присутствии 5% CO2.
Клетки собирали через 24 часа или 48 часов после трансфекции и активности люциферазы Renillia и люциферазы Firefly анализировали с помощью набора Promega Dual Luc в соответствии с инструкциями производителей. Эти активности считывали на аппарате Victor.
Пример 6: Сравнительная оценка in vitro активности полинуклеотида
Относительные активности полинуклеотида CARP (pXL3634) и промоторов RSV (pXL3635) оценивали in vitro при транзитной трансфекции в первичные культуры кардиомиоцитов крыс и результаты выражали в виде отношения к активности плазмиды pRL-CMV (фигура 5).
Результаты показали, что полинуклеотид, расположенный выше используемого гена CARP (pXL3634), имел очень низкую in vitro активность, составляющую примерно 0,04% по отношению к активности промотора CMV.
Относительная активность неспецифического сильного промотора RSV (pXL3635) также была низкой и составляла соответственно примерно 0,05% и 0,68% по отношению к активности эталонного промотора CMV.
Пример 7: Конструирование аденовируса
Аденовирус, обеспечивающий экспрессию люциферазы под контролем промотора CARP, конструировали методом, описанным Crouzet et al. (PNAS, 94 (1997) 1414-1419), при этом экспрессирующая кассета была идентична кассете плазмиды pXL3634 (фигура 3).
Челночный вектор, обеспечивающий рекомбинацию в E.coli, конструировали в две стадии. Сначала промотор CARP (фрагмент XhoI, достроенный ферментами Кленова/BamHI), вводили в плазмиду pXL3474 (гидролизованную ферментами Scal и BglII) между областями ITR-ψ и pIX и получали плазмиду pXL3758. Затем генерировали плазмиду pXL3759 путем введения в плазмиду pXL3758, гидролизованную ферментами BstBII (достроенную фрагментом Кленова) и BstEII, фрагмента, содержащего кДНК люциферазы и сайт полиаденилирования SV40 (BamHI-фрагмент pXL3634, достроенный ферментами Кленова/BstEII). pXL3759 схематично показана на фигуре 6А.
Гомологичную двойную рекомбинацию в E.coli осуществляли, как описано выше для плазмиды pXL3215, содержащей аденовирусный геном ΔЕ1/ΔЕ3, в котором RSV-LacZ-экспрессирующую кассету встраивали в область Е1. Плазмида pXL3215 представляет собой производное плазмиды pXL2689, которое содержит сайт инициации репликации плазмиды RК2 и ген резистентности к тетрациклину (Crouzet et al., PNAS, 1997). Продукт этой двойной рекомбинации, плазмиду pXL3778, оценивали путем секвенирования экспрессирующей кассеты. После расщепления ферментом PacI для выделения линейного вирусного генома эту плазмиду трансфицировали в клеточную линию Per.C6 (WO 97/00326) для получения вируса AV.0CARP-Luc+.
Этот вирус также оценивали путем секвенирования экспрессирующей кассеты посредством рестрикционного анализа и присутствие частиц RCA Е1+ (компетентного по репликации аденовируса) тестировали на гибридизацию с помощью зонда ψ.
Культуры с высоким титром вируса получали путем амплификации вируса в линии Per.C6 и вирусные частицы очищали на градиенте CsCL. Титр этого вируса, выраженный в вирусных частицах/мл (в.ч./мл), получали с помощью хроматографии, а его активность оценивали in vitro путем титрования люциферазной активности после инфицирования скелетных клеток или клеток сердечной мышцы и сравнения с вирусами, используемыми в качестве контроля, содержащего промотор CMV.
Пример 8: Инъекция ДНК in vivo
Крыс CD Sprague весом 200 г анестезировали путем внутрибрюшинной инъекции смеси кетамина (70 мг/мл)/ксилазина (6 мг/мл) при концентрации 1 мл/кг.
После лапаротомии осуществляли интрамиокардиальные инъекции путем введения через диафрагму стеклянного 100 мкл-шприца Гамильтона, подсоединенного с катетеру Steriflex (ном. 167.10 G19V), снабженному выступом-ограничителем и наконечником с иглой BD 26G*3.8 (с короткой скошенной гранью).
50 мкл этого раствора ДНК, доведенного до 0,9% NaCl, инъецировали в течение 5 секунд. После умерщвления животных их сердца удаляли, промывали в 0,9% растворе NaCl и подвергали макроскопической оценке. Затем эти сердца анализировали на люциферазную активность с использованием набора (Promega Е151А) после измельчения на гомогенизаторе (Ultra-thurax, Diax 600 Heidolph) в буфере для лизиса, имеющегося в наборе, в который были добавлены ингибиторы протеазы (CompleteTM, Roche Diagnostics), и последующего центрифугирования в течение 20 минут при 4000 об/мин при 4°С. Считывание проводили на устройстве: LUMAT LB 9501 (10 мкл супернатанта +50 мкл субстрата люциферазы (Promega). Люциферазные активности выражали в единицах массы люциферазы на сердце (пг люциферазы/сердце) с использованием калибровки, описанной Mir et al. (PNAS 96 (1999), 4262-4267).
Альтернативно, сердца фиксировали в 3,7% параформальдегиде и анализировали посредством иммуногистохимического анализа на экспрессию FGF-1.
Пример 9: Сравнительная оценка in vivo активности полинуклеотида CARP
Результаты, систематизированные на фигуре 7А, показали, что уровни экспрессии люциферазы, полученные после инъекции возрастающих доз 1, 5, 25 и 125 мкг плазмид pXL3031 и pXL3634, не имели значительного отличия, что явно свидетельствовало о том, что полинуклеотид, расположенный выше гена CARP, способен индуцировать высокие уровни экспрессии, эквивалентные уровням экспрессии, регулируемой сильным промотором, таким как промотор CMV.
С другой стороны, уровень экспрессии, наблюдаемой с использованием другого сильного промотора, промотора RSV (pXL3635), был ниже, чем уровень экспрессии, наблюдаемый с использованием промотора CMV или полинуклеотида, расположенного выше гена CARP (фигура 7В).
Более того, хотя было продемонстрировано, что присоединение энхансера CMV выше промоторов клеток гладкой мышцы (промотора α-актина SM, pXL3130 или SM22, pXL3153) было в высокой степени эффективным in vitro (WO 00/18908), однако оно оказалось неэффективным в клетках сердца in vivo.
Пример 10: Оценка специфичности экспрессии полинуклеотида CARP
25 мкг каждой из плазмид pXL3634, pXL3635 и pXL3031 вводили крысам путем внутрисердечной инъекции через диафрагму.
Параллельно группам мышей в краниальную большеберцовую мышцу вводили внутримышечные инъекции 10 мкг каждой из указанных плазмид с использованием или без использования электропереноса.
Экспрессию люциферазы анализировали через 7 дней после инъекции, как описано в PNAS 96 (1999), 4262-4267.
Уровни экспрессии люциферазы в сердце выражали в виде отношения к уровням, наблюдаемым в краниальной большеберцовой мышце, и систематизировали на фигуре 8.
Результаты ясно показали, что полинуклеотид, расположенный выше гена CARP, и промотор CMV представляют собой единственные два промотора, которые способны индуцировать самый высокий уровень экспрессии в сердечной ткани. Однако в случае промотора CMV отношение уровней экспрессии в сердце/мышце равно 1, а в случае использования полинуклеотида, расположенного выше гена CARP, это отношение близко к 100, что явно свидетельствует о высокой степени селективности указанного полинуклеотида по отношению к сердечной ткани.
Исключительно высокая специфичность экспрессии полинуклеотида также явно наблюдалась и в других конструкциях, содержащих энхансер и промотор, специфичные для клеток гладкой мышцы, такие как промотор гена, кодирующего белок SM-22, и промотор актина, для иллюстрации которых отношения экспрессии в сердце/мышце также представлены на фигуре 8.



Изобретение относится к области молекулярной биологии и генетической инженерии и может быть использовано в медицине. Определена часть нуклеотидной последовательности, расположенной выше кодирующей последовательности гена CARP, с активностью промотора, обеспечивающего специфическую экспрессию функционально связанного с ним гена в клетках сердца in vivo. Выявленная промоторная область соответствует последовательности гена CARP мыши от нуклеотида в положении (-2266) до нуклеотида в положении (+92). Предложены кассета и векторы для экспрессии представляющего терапевтическую ценность целевого белка в сердечной ткани, отличительной особенностью которых является содержание в них названной регуляторной последовательности, и определены возможности использования этих генетических конструкций в составе лекарственных средств. 9 н. и 10 з.п., 10 ил.
a) получение вектора по любому из пп.11-15 и
b) введение эффективного количества указанного вектора в сердечную ткань при таких условиях, которые обеспечивают экспрессию указанного нужного гена.
| КТО Н | |||
| ет al | |||
| Development | |||
| Ударно-вращательная врубовая машина | 1922 |
|
SU126A1 |
| WO 00/15821 A, 23.03.2000 | |||
| DATABASE EMBL | |||
| Ac | |||
| Автоматическая цепная пила для разделки древесины | 1960 |
|
SU131884A1 |

