Изобретение связано с производством микроорганизмов. Более конкретно, настоящее изобретение относится к способу получения L-аминокислоты путем ферментации, и к ДНК и микроорганизму, которые используются в этом способе.
При получении L-лизина путем ферментации штаммы, выделенные из природных источников, или его синтетические штаммы используют для улучшения продуктивности. Известно большое число L-лизин-производящих синтетических мутантов и многие из них являются аминоэтилцистеин (АЕС)-устойчивыми мутантами, принадлежащими к роду Brevibacterium, Corynebacterium, Bacillus или Escherichia. Далее, также используются трансформанты, полученные при использовании рекомбинантных ДНК (U.S.Patent No 4278765). Таким образом, раскрыто большое число методов для увеличения продуктивности аминокислот.
Например, в отношении рода Escherichia, в Japanese Laid-Open (Kokai) No 18596/1981, US Patent No 4346170 и Applied Microbiology and Biotechnology 15, 227 (1982) описан способ получения L-лизина путем увеличения синтетазы дигидродипиколиновой кислоты (далее обозначаемой аббревиатурой "DDPS").
Синтетаза дигидродипиколиновой кислоты (DDPS) является ферментом, катализирующим дегидроконденсацию аспартосемиальдегида и пировиноградной кислоты с образованием дигидродипиколиновой кислоты. Эта реакция представляет собой начало биосинтеза L-лизина при биосинтезе аминокислот типа аспарагиновой кислоты. Известно, что этот фермент служит в качестве важного регуляторного участка биосинтеза L-лизина в бактериях рода Escherichia наряду с аспартокиназой.
DDPS закодирована в гене, называемом dapA в Escherichia соli (E.coli). Этот dapA уже был клонирован, а его основная последовательность определена [Richaud, F. et al., J.Bacteriol., 297 (1986)].
Между тем, аспартокиназа (далее обозначаемая аббревиатурой "АК") является ферментом, катализирующим реакцию превращения аспарагиновой кислоты в β-фосфоаспарагиновую кислоту, и представляет собой основной регуляторный фермент в системе биосинтеза аминокислот типа аспарагиновой кислоты. АК E.coli включает в себя три типа (AKI, AKII, AKIII) и два из них являются конъюгированными ферментами с гомосерин дегидрогеназой (далее обозначаемой аббревиатурой "HD"). Один из этих конъюгированных ферментов является AKI-HDI, закодированным в гене thrA, а другой является AKII-HDII, закодированным в гене metLM. AKI подвергается согласованной супрессии треонином и изолейцином и ингибированию треонином, а АКII подвергается супрессии метионином.
Между тем, одна AKIII является ферментом единственной функции и она является продуктом гена, называемого 1ysС. Известно, что он подвергается супрессии и ингибированию по типу обратной связи L-лизином. Соотношение их активностей в клетках составляет AKI:AKII:АКIII = приблизительно 5:1:4. Этот 1ysС уже был клонирован и последовательность его оснований определена [Cassan, M., Parsot, С., Cohen, G.N. и Patte, J.C., J.Biol.Chem., 261, 1052 (1986)].
Способ получения L-лизина с использованием E.coli, содержащих dapA с мутацией, по которой снимается ингибирование лизином по типу обратной связи, и lysC с мутацией, по которой снимается ингибирование лизином по типу обратной связи, раскрыт в International Laid-Open Pamphlet WО 95/16042.
При получении L-треонина путем ферментации штаммы, выделенные из природных источников или его синтетических мутантов, используются как микроорганизмы. Известно большое число L-треонин-продуцирующих синтетических мутантов, принадлежащих к роду Escherichia, Serratia, Brevibacterium или Corynebacterium, и многие из них устойчивы к α-амино-β-гидроксивалериановой кислоте. Что касается рода Escherichia, в Japanese Laid-Open (Kokai) 131397/1980, 31691/1984 и 15696/1981, и Japanese Patent Announcement No 501682/1991 описан способ получения L-треонина с использованием штамма, трансформированного рекомбинантной плазмидой, содержащей оперон треонина.
Далее, в International Laid-Open Pamphlet WО 94/11517 раскрыт способ получения L-треонина с использованием Е.соli, содержащих lysC с мутацией, по которой снимается ингибирование лизином по типу обратной связи.
Проблемы, решаемые в изобретении
Настоящее изобретение направлено на получение AKIII, полученной из бактерий рода Escherichia, в которых хорошо снимается ингибирование L-лизином по типу обратной связи, и обеспечение способа получения L-аминокислоты путем ферментации, улучшенного по сравнению с предшествующим.
Способы решения этих проблем
Изобретатели проводили тщательные исследования для того, чтобы решить упомянутые выше проблемы, и последовательно добились успеха в получении ДНК, кодирующей AKII, полученную из бактерий рода Escherichia, в которой хорошо снимается ингибирование L-лизином по типу обратной связи. ДНК, кодирующую AKIII, полученную из бактерий рода Escherichia, в которой хорошо снимается ингибирование L-лизином по типу обратной связи, называют иногда мутантным геном lysC или мутантным геном AKIII в настоящем описании.
ДНК, кодирующую DDPS, полученную из E.coli, в которой хорошо снимается ингибирование L-лизином по типу обратной связи, называют иногда мутантным геном dapA или мутантным геном DDPS в настоящем описании.
Далее, изобретатели получили бактерии рода Escherichia, содержащие в клетках мутантный lysC, и обнаружили, что в этой культуре может быть получено и накоплено значительное количество L-лизина и L-треонина.
Таким образом, настоящее изобретение относится к ДНК, кодирующей аспартокиназу III бактерий, принадлежащих роду Escherichia, и содержащей в кодирующем участке мутацию, через посредство которой снимается ингибирование лизином по типу обратной связи упомянутой аспартокиназы III, причем упомянутая мутация является:
мутацией, при которой 318-й метиониновый остаток аспартокиназы III замещается остатком другой аминокислоты и 323-й глициновый остаток - остатком другой аминокислоты,
мутацией, при которой 325-й лейциновый остаток замещается остатком другой аминокислоты, и 347-й валиновый остаток - остатком другой аминокислоты,
мутацией, при которой 323-й глициновый остаток замещается остатком другой аминокислоты, и 347-й валиновый остаток - остатком другой аминокислоты,
мутацией, при которой 325-й лейциновый остаток замещается остатком другой аминокислоты, и 345-й сериновый остаток - остатком другой аминокислоты,
мутацией, при которой 323-й глициновый остаток замещается остатком другой аминокислоты, и 358-й сериновый остаток - остатком другой аминокислоты,
мутацией, при которой 344-й треониновый остаток замещается остатком другой аминокислоты,
мутацией, при которой 250-й остаток глутаминовой кислоты замещается остатком другой аминокислоты,
мутацией, при которой 346-й остаток глутаминовой кислоты замещается остатком другой аминокислоты, и 347-й лейциновый остаток - остатком другой аминокислоты,
мутацией, при которой 250-й остаток глутаминовой кислоты замещается остатком другой аминокислоты, и 364-й треониновый остаток - остатком другой аминокислоты,
мутацией, при которой 202-й остаток аспарагиновой кислоты замещается остатком другой аминокислоты, и 321-й сериновый остаток - остатком другой аминокислоты,
мутацией, при которой 283-й аргининовый остаток замещается остатком другой аминокислоты, 333-й аланиновый остаток - остатком другой аминокислоты, 338-й сериновый остаток - остатком другой аминокислоты, 346-й остаток глутаминовой кислоты - остатком другой аминокислоты и 414-й аспарагиновый остаток - остатком другой аминокислоты, или
мутацией, при которой 318-й метиониновый остаток замещается остатком другой аминокислоты, 321-й сериновый остаток - остатком другой аминокислоты, 328-й валиновый остаток - остатком другой аминокислоты, 349-й валиновый остаток - остатком другой аминокислоты и 405-й остаток глутаминовой кислоты - остатком другой аминокислоты.
Предпочтительно, настоящее изобретение относится к ДНК, кодирующей аспартокиназу III бактерий, принадлежащих к роду Escherichia, и содержащей в кодирующем участке мутацию, через посредство которой снимается ингибирование лизином по типу обратной связи упомянутой аспартокиназы III, причем упомянутая мутация является:
мутацией, при которой 318-й метиониновый остаток аспартокиназы III замещается изолейциновым остатком и 323-й глициновый остаток - остатком аспарагиновой кислоты,
мутацией, при которой 325-й лейциновый остаток замещается остатком фенилаланина и 347-й валиновый остаток - метиониновым остатком,
мутацией, при которой 323-й глициновый остаток замещается остатком аспарагиновой кислоты и 347-й валиновый остаток - метиониновым остатком,
мутацией, при которой 325-й лейциновый остаток замещается остатком фенилаланина и 345-й сериновый остаток - лейциновым остатком,
мутацией, при которой 323-й глициновый остаток замещается остатком аспарагиновой кислоты и 358-й сериновый остаток - лейциновым остатком,
мутацией, при которой 344-й треониновый остаток замещается метиониновым остатком,
мутацией, при которой 250-й остаток глутаминовой кислоты замещается лизиновым остатком,
мутацией, при которой 346-й остаток глутаминовой кислоты замещается лизиновым остатком и 347-й лейциновый остаток - остатком фенилаланина,
мутацией, при которой 250-й остаток глутаминовой кислоты замещается лизиновым остатком и 364-й треониновый остаток - метиониновым остатком,
мутацией, при которой 202-й остаток аспарагиновой кислоты замещается глициновым остатком и 321-й сериновый остаток - пролиновым остатком,
мутацией, при которой 283-й аргининовый остаток замещается сериновым остатком, 333-й аланиновый остаток - треониновым остатком, 338-й сериновый остаток - треониновым остатком, 346-й остаток глутаминовой кислоты - остатком аспарагиновой кислоты и 414-й аспарагиновый остаток - сериновым остатком, или
мутацией, при которой 318-й метиониновый остаток замещается лизиновым остатком, 321-й сериновый остаток - пролиновым остатком, 328-й валиновый остаток - остатком фенилаланина, 349-й валиновый остаток - глициновым остатком и 405-й остаток глутаминовой кислоты - валиновым остатком.
Настоящее изобретение относится к упомянутой выше рекомбинантной ДНК, которую получают при лигировании ДНК пункта 1 с векторной ДНК, способной к автономной репликации в клетках бактерии рода Escherichia, а также к микроорганизмам рода Escherichia, имеющим эту ДНК.
Настоящее изобретение относится к способу получения L-аминокислоты, который включает инкубирование упомянутого выше микроорганизма, обладающего способностью продуцировать L-аминокислоту в ферментативной среде, получение и накопление L-аминокислоты в культуре и извлечение L-аминокислоты из этой культуры.
В настоящем описании ДНК, кодирующую DDPS или AKIII, или ДНК, содержащую указанные ДНК и промотор, называют иногда "ген DDPS" или "ген AKIII". Далее, мутантный фермент, в котором снимается ингибирование L-лизином по типу обратной связи, иногда называют "мутантным ферментом", а ДНК, содержащую то же, или ДНК, содержащую то же и промотор, - "мутантным геном". Еще далее, "снять ингибирование L-лизином по типу обратной связи" означает, что это ингибирование является, по существу, снятым и оно не обязательно снято полностью.
Настоящее изобретение подробно описано ниже.
ДНК, кодирующая мутантную аспартокиназу (AKIII) настоящего изобретения
ДНК, кодирующая мутантную аспартокиназу (AKIII) настоящего изобретения, представляет собой ДНК, кодирующую AKIII дикого типа, имеющую мутацию, через посредство которой снимается ингибирование L-лизином по типу обратной связи AKIII, которая закодирована. Под AKIII имеется в виду AKIII, полученная из бактерий рода Escherichia, особенно AKIII, полученная из Е.соli. Мутация, через посредство которой снимается ингибирование L-лизином по типу обратной связи AKIII, включает в последовательности аминокислот AKIII, представленной последовательностью 1 Таблицы Последовательностей, считая от N-конца AKIII,
(а) мутацию, при которой 318-й метиониновый остаток аспартокиназы III замещается остатком другой аминокислоты, предпочтительно изолейциновым остатком, и 323-й глициновый остаток - остатком другой аминокислоты, предпочтительно аспарагиновой кислоты,
(b) мутацию, при которой 325-й лейциновый остаток замещается другой аминокислотой, предпочтительно остатком фенилаланина, и 347-й валиновый остаток - остатком другой аминокислоты, предпочтительно метиониновым остатком,
(c) мутацию, при которой 323-й глициновый остаток замещается остатком другой аминокислоты, предпочтительно остатком аспарагиновой кислоты, и 347-й валиновый остаток - остатком другой аминокислоты, предпочтительно метиониновым остатком,
(d) мутацию, при которой 325-й лейциновый остаток замещается остатком другой аминокислоты, предпочтительно фенилаланина, и 345-й сериновый остаток - остатком другой аминокислоты, предпочтительно лейциновым остатком,
(e) мутацию, при которой 323-й глициновый остаток замещается остатком другой аминокислоты, предпочтительно остатком аспарагиновой кислоты, и 358-й сериновый остаток - остатком другой аминокислоты, предпочтительно лейциновым остатком,
(f) мутацию, при которой 344-й треониновый остаток замещается остатком другой аминокислоты, предпочтительно метиониновым остатком,
(g) мутацию, при которой 250-й остаток глутаминовой кислоты замещается остатком другой аминокислоты, предпочтительно лизиновым остатком,
(h) мутацию, при которой 346-й остаток глутаминовой кислоты замещается остатком другой аминокислоты, предпочтительно лизиновым остатком, и 347-й лейциновый остаток - остатком другой аминокислоты, предпочтительно остатком фенилаланина,
(i) мутацию, при которой 250-й остаток глутаминовой кислоты замещается остатком другой аминокислоты, предпочтительно лизиновым остатком, и 364-й треониновый остаток - остатком другой аминокислоты, предпочтительно метиониновым остатком,
(j) мутацию, при которой 202-й остаток аспарагиновой кислоты замещается остатком другой аминокислоты, предпочтительно глициновым остатком, и 321-й сериновый остаток - остатком другой аминокислоты, предпочтительно пролиновым остатком,
(k) мутацию, при которой 283-й аргининовый остаток замещается остатком другой аминокислоты, предпочтительно сериновым остатком, 333-й аланиновый остаток - остатком другой аминокислоты, предпочтительно треониновым остатком, 338-й сериновый остаток - остатком другой аминокислоты, предпочтительно треониновым остатком, 346-й остаток глутаминовой кислоты - остатком другой аминокислоты, предпочтительно остатком аспарагиновой кислоты и 414-й аспарагиновый остаток - остатком другой аминокислоты, предпочтительно сериновым остатком, или
(l) мутацию, при которой 318-й метиониновый остаток замещается остатком другой аминокислоты, предпочтительно лизиновым остатком, 321-й сериновый остаток - остатком другой аминокислоты, предпочтительно пролиновым остатком, 328-й валиновый остаток - остатком другой аминокислоты, предпочтительно остатком фенилаланина, 349-й валиновый остаток - остатком другой аминокислоты, предпочтительно глициновым остатком и 405-й остаток глутаминовой кислоты - остатком другой аминокислоты, предпочтительно валиновым остатком.
ДНК, кодирующая AKIII дикого типа, в частности, не ограничена. Упоминается ДНК, кодирующая AKIII, полученная из бактерий рода Escherichia, например Е.соli. Точнее, упоминается ДНК, кодирующая последовательность аминокислот, представленную Последовательностью 1, и, кроме того, последовательность, представленную основаниями от 584 до 1930 в последовательности оснований, представленной Последовательностью 1. AKIII Е.соli закодирована в гене lysC.
Из упомянутых выше последовательностей, последовательность, имеющая мутацию в последовательности оснований, которая вызывает замещение остатка аминокислоты, является ДНК, кодирующей мутантную AKIII настоящего изобретения. Тип кодона, соответствующего замещенному остатку аминокислоты, особенно не ограничен, пока он кодирует этот остаток аминокислоты. Последовательность аминокислот AKIII дикого типа незначительно меняется в зависимости от бактерий или штаммов. Последовательность, имеющая замещение, делецию или инсерцию остатка аминокислоты в положении, которое не участвует в ферментативной активности, также включена в мутантный ген АК III настоящего изобретения.
Например, последовательность оснований (Последовательность 1) дикого типа гена lysC, полученного в Примере 1, отличается от последовательности оснований 1ysС штамма Е.coli К-12 JC411, который уже опубликован [Cassan, M. , Parsot, С. , Cohen, G.N. и Patte J.C., J.Biol.Chem., 261, 1052 (1986)] 6 сайтами. Кодируемые аминокислотные остатки различаются в 2 сайтах из 6 (в lysC штамма JC411 58-й глициновый остаток, считая от N-конца, замещен цистеиновым остатком, и 401-й глициновый остаток аланиновым остатком в последовательности аминокислот lysC, представленной Последовательностью 1). Если любая из упомянутых выше мутаций от а) до l) вводится в lysC, обладающий той же последовательностью, что и lysC штамма E.coli К-12 JC411, ожидается, что будет получен lysC, имеющий мутацию, через посредство которой снимается ингибирование L-лизином по типу обратной связи.
ДНК, кодирующую мутантную AKIII, в которой снимается ингибирование L-лизином по типу обратной связи, получают следующим образом. Сначала ДНК, содержащую ген AKIII дикого типа или ген AKIII, имеющий другую мутацию, подвергают мутации in vitro и мутантную ДНК лигируют с векторной ДНК, адаптируемой к хозяину, получая при этом рекомбинантную ДНК. Рекомбинантную ДНК вводят в микроорганизм хозяина, получая трансформант. Когда трансформант, экспрессирующий мутантную AKIII, отбирают, то этот трансформант сохраняет мутантный ген. Далее, ДНК, содержащую ген AKIII дикого типа или ген АК III, имеющий другую мутацию, лигируют с векторной ДНК, адаптируемой к хозяину, получая при этом рекомбинантную ДНК. Эту рекомбинантную ДНК затем подвергают мутации in vitro, и мутантную ДНК вводят в микроорганизмы хозяев, получая трансформант. Когда трансформант, экспрессирующий AKIII, отбирают, этот трансформант также сохраняет мутантный ген. По-другому, также возможно подвергать мутации микроорганизмы, вырабатывающие фермент дикого типа с образованием мутантного штамма, производящего мутантный фермент, и затем получать мутантный ген из этого мутантного штамма.
В качестве агента для прямого мутагенеза ДНК упоминается гидроксиламин и т. п. Гидроксиламин является химическим мутагеном, который вызывает мутацию цитозина в тимин, превращая цитозин в N4-гидроксицитозин. Далее, если микроорганизм уже является мутантом, производят облучение ультрафиолетовым светом или обработку мутагеном, используемым обычно при осуществлении искусственной мутации, таким как N-метил-N'-нитро-N-нитрозогуанидин (NTG).
В качестве донорных бактерий ДНК, содержащей ген AKIII дикого типа или ген AKIII, имеющий другую мутацию, можно использовать любые микроорганизмы, принадлежащие к роду Escherichia. Точнее, проигодны те, которые описаны в Documents of F.C.Neidhardt et al. (Neidhardt, F.C., et al., Escherichia coli and Salmonella Typhimurium, American Society for Microbiology, Washington D. C. , p.1208, Таблица 1). Их примерами являются штамм E.coli JM109 и штамм E. coli MC1061.
(1) Получение гена AKIII дикого типа
Пример получения ДНК, содержащей ген AKIII, описан ниже. Сначала, например, дикого типа штамм E.coli MC1061, содержащий lysC, инкубируют для получения культуры. Упомянутые выше микроорганизмы можно инкубировать в твердой культуре. В свете эффективности сбора клеток предпочтительно использование жидкой культуры. В этом случае культуральной средой является, например, среда, полученная при добавлении одного или более типов неорганических солей, выбираемых из моногидрофосфата калия, дигидрофосфата калия, сульфата магния, хлорида натрия, хлорида магния, хлорида железа, сульфата железа и сульфата марганца, к одному или более из источников углерода, выбираемому из экстракта дрожжей, пептона, экстракта мяса, экстракта замоченных зерен и стока соевых бобов или пшеницы, с дальнейшим добавлением подходящих количеств источника сахаров, витамина и т.п., как полагается. Начальный рН культуральной среды полезно установить между 6 и 8. Инкубирование ведут при температуре от 30 до 42oС, предпочтительно, при приблизительно 37oС, в течение от 4 до 24 ч в погруженно-воздушной культуре при перемешивании, во встряхиваемой культуре или в стационарной культуре. Полученную таким образом культуру разделяют, например, при 3000 об/мин в течение 5 мин для получения штамма Е.соli МС1061. Хромосомную ДНК можно получить из этого штамма, например по методу Saito и Miura [Biochem.Biophys.Acta., 72, 619 (1963)] или по методу K. S.Kirby [Biochem.J., 64, 405 (1956)]. Для того чтобы выделить ген AKIII из полученной хромосомной ДНК, готовят хромосомную библиотеку ДНК. Сначала хромосомную ДНК частично расщепляют подходящей рестрикционной эндонуклеазой, получая при этом различные смеси фрагментов. Если степень расщепления контролировать путем контроля времени разрезания или т.п., получается большое число смесей фрагментов. Например, хромосомная ДНК взаимодействует с Sau 3AI при концентрации фермента от 1 до 10 ед./мл при температуре от 30oС или выше, предпочтительно при 37oС, за период времени от 1 мин до 2 ч для ее переваривания.
Затем расщепленный фрагмент хромосомной ДНК лигируют с вектором ДНК, способным к автономной репликации в клетках бактерий рода Escherichia, получая при этом рекомбинантную ДНК. Точнее, вектор ДНК взаимодействует с рестрикционной эндонуклеазой, которая допускает образование такой же концевой последовательности оснований, как рестрикционная эндонуклеаза Sau 3AI, используемая для расщепления хромосомной ДНК, например Bam HI, при концентрации фермента от 1 до 100 ед./мл при температуре от 30oС или выше, в течение 1 ч или больше, предпочтительно от 1 до 3 ч, для полного переваривания, и она расщепляется. Потом полученную выше смесь фрагментов хромосомной ДНК смешивают с расщепленной векторной ДНК. Эта смесь взаимодействует с лигазой ДНК, предпочтительно с Т4 лигазой ДНК, при концентрации фермента от 1 до 100 ед. /мл, при температуре от 4 до 116oС, в течение 1 ч или больше, предпочтительно от 6 до 24 ч, для образования рекомбинантной ДНК.
Микроорганизм рода Escherichia, например штамм E.coli К-12, предпочтительно штамм, полностью лишенный AKI, AKII и AKIII, например штамм E.coli GT3 [который можно получить из E.coli Genetic Stock Center (Connecticut, U. S. А)] трансформируют для получения библиотеки хромосомной ДНК. Эта трансформация может быть проведена по методу D.M. Morrison [Methods in Enzymology 68, 326 (1979)] или по методу, в котором проницаемость ДНК повышается при обработке клеток реципиентных бактерий хлоридом кальция [Mandel, M. и Higa, A., J.Mol. Biol., 53, 159 (1970)].
Штамм, обладающий рекомбинантной ДНК гена AKIII, получают из штамма, имеющего повышенную AKIII-активность, или штамма, в котором ауксотрофия, вызванная делецией гена АК III, скомпенсирована в полученной библиотеке хромосомной ДНК. Например, когда мутант, полностью лишенный АК, используется в качестве хозяина, рекомбинантный штамм, который может быть выращен в среде, свободной от L-лизина, L-треонина, L-метионина и диаминопимелиновой кислоты, или в культуральной среде, свободной от гомосерина и диаминопимелиновой кислоты, и рекомбинантную ДНК выделяют из этого штамма, делая возможным получение фрагмента ДНК, содержащего ген AKIII. Клеточный экстракт готовят из штамма-кандидата, а из клеточного экстракта образуется сырой ферментный раствор. Затем определяется активность AKIII. Ферментативную активность АК III можно измерить по методу E.R. Stadtman [Stadtman, E.R., Cohen, G.N., LeBras, G., и Robichon-Szulmajster, H., J.Biol. Chem., 236, 2033 (1961)].
Рекомбинантную ДНК, полученную при встраивании ДНК, содержащей ген AKIII, в векторную ДНК, можно выделить, например, по методу Р. Guerry et al. [J. Bacteriol. , 116, 1064 (1973)] или по методу D.B.Clewell [J.Bacteriol., 110, 667 (1972)].
Ген AKIII дикого типа получают в результате получения хромосомной ДНК из штамма, имеющего на хромосоме ген AKIII по методу Saito и Miura, и амплификации гена AKIII путем полимеразной цепной реакции [PCR, White, T.J., et al. , Trends Genet., 5, 185 (1989)]. ДНК праймер, используемый при амплификации, является праймером, комплементарным к обоим 3'-концам двойной спирали ДНК, содержащей общий участок гена AKIII или его часть. Когда амплифицируется только часть области гена AKIII, необходимо скринировать фрагмент ДНК, содержащий общий участок из библиотеки хромосомной ДНК, используя в качестве праймера фрагмент ДНК, имеющий часть участка AKIII. Если амплифицируют весь участок гена AKIII, раствор PCR, содержащий фрагмент ДНК с амплифицированным геном AKIII, подвергают электрофорезу в агарозном геле, и затем экстрагируют желаемый фрагмент ДНК, в результате чего можно выделить фрагмент ДНК, содержащий ген AKIII.
ДНК праймер может быть образован, например, на основе известной последовательности E.coli [Cassan, М., Parsot, С., Cohen, G.N. и Patte J.C., J.Biol. Chem. , 261, 1052 (1986)]. Подходит праймер, способный амплифицировать участок, состоящий из 1347 оснований, кодирующих ген 1ysС. Например, подходят два типа праймеров, представленных Последовательностями 2 и 3. Праймерную ДНК можно синтезировать обычным методом, например фосфоамидидным методом [Tetrahedron Letters 22, 1859 (1981)] с использованием коммерчески доступного синтезатора ДНК (например, синтезатора ДНК модели 380В, производимого Applied Biosystems). Далее, PCR может быть проведена по методу, указанному изготовителем с использованием коммерчески доступного PCR оборудования (ДНК Thermal Cycler Model PJ2000, производимого Takara Shuzo Co., Ltd.) и Taq-ДНК полимеразы (предоставленной Takara Shuzo Co., Ltd.).
Ген AKIII, амплифицированный путем PCR, лигируют с векторной ДНК, способной к автономной репликации в клетках бактерий, принадлежащих к роду Escherichia, и вводят в клетки бактерий рода Escherichia, посредством чего может быть легко проведено введение мутации в ген AKIII. Векторная ДНК, трансформация и идентификация наличия гена АКТ II являются теми же, что и описанные выше.
(2) Введение мутации в ген AKIII
Полученный выше ген AKIII подвергают такой мутации, как замещение, делеция или инсерция остатка аминокислоты рекомбинантным PCR методом [Higuchi, R. 61 по PCR технологии (Еrlich, H.A. Eds., Stockton Press (1989)], методом сайт-специфического мутагенеза [Kramer, W. and Frits, H.J., Methods in Enzymology, 154, 350 (1987); Kunkel, Т.A. et al. Methods ih Enzymology 154, 367 (1987)], или т.п. Эти методы могут вызвать желаемую мутацию в нужном сайте.
Мутацию или случайную мутацию можно ввести в нужный сайт методом химического синтеза нужного гена.
Далее, существует метод, в котором ген AKIII в хромосоме или плазмиде непосредственно обрабатывается гидроксиламином [Hashimoto, Т. and Sekiguchi, M., J.Bacteriol., 159, 1039 (1984)]. Еще далее, можно применить метод, в котором бактерии рода Escherichia, содержащие ген AKIII, облучают ультрафиолетовым светом, или метод с использованием химического агента, такого как N-метил-N'-нитрозогуанидин или азотная кислота. Мутацию можно вводить произвольно этими методами.
Метод отбора мутантного AKIII гена описан ниже. Так, во-первых, рекомбинантную ДНК, содержащую мутантный ген АК III, трансформируют в штамм, полностью лишенный АК, например в штамм E.coli GT3. Затем, этот трансформант инкубируют в минимальной среде, например М9, содержащей значительное количество L-лизина. Так как исключительно АК ингибируется L-лизином в штамме, содержащем рекомбинантную ДНК, содержащую дикого типа ген AKIII, L-треонин, L-изолейцин, L-метионин и диаминопимелиновая кислота (DAP) не могут быть синтезированы, поскольку они контролируют ее рост. Между тем, штамм, имеющий рекомбинантную ДНК, содержащую мутантный ген AKIII, в котором снимается ингибирование L-лизином, должен расти в минимальной среде, содержащей значительное количество L-лизина. При использовании этого явления возможно выбрать штамм, устойчивый к L-лизину или S-2-аминоэтилцистеину (АЕС), аналогично L-лизину при росте, т.е. штамм с рекомби-нантной ДНК, содержащей мутантный ген AKIII, в котором снимается ингибирование.
Полученный таким образом рекомбинантный ген в виде рекомбинантной ДНК вводят в подходящий микроорганизм (хозяин) и экспрессируют, чтобы получить микроорганизм, содеращий АК III, в котором снимается ингибирование по типу обратной связи.
В качестве хозяина предпочтительны микроорганизмы, принадлежащие к роду Escherichia. Например, упоминается Escherichia coli (E.coli).
Далее, можно использовать субстанцию, полученную при извлечении фрагмента мутантного гена AKIII из рекомбинантной ДНК и введенную в другой вектор. В качестве векторной ДНК, которая может быть использована в настоящем изобретении, предпочтительна плазмидная векторная ДНК. Ее примеры включают pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219 и pMW218. Также пригодны фаговый ДНК-вектор и транспозонный вектор.
Для эффективной экспрессии мутантного гена AKIII другой промотор, действующий внутри микроорганизма, такой как lac, trp и PL, может быть лигирован с расположенным в обратном направлении участком, кодирующим мутантную AKIII, или в качестве такового используется амплифицированный промотор, содержащийся в гене AKIII.
Как отмечалось выше, вещество, полученное при встраивании мутантного гена в векторную ДНК, способную к автономной репликации, может быть введено в хозяина и удерживаться в нем в виде внехромосомной ДНК, как плазмида. Или мутантный ген можно встроить в хромосому микроорганизма хозяина путем трансдукции с помощью транспозона (Berg D.E. and Berg С.М., Bio/Technol., 1, 417 (1983)] с Mu-фагом [Japanese Laid-Open (Kokai) No 109985/1990] или путем комплементарной рекомбинации [Experiments in Molecular Genetics, Cold Spring Harbour Lab. (1972)].
Получение L-аминокислоты по способу настоящего изобретения
L-аминокислоту можно получить с высокой эффективностью в результате инкубирования в подходящей культуральной среде бактерий рода Escherichia, которые были трансформированы при введении полученного выше мутантного гена AKIII, получения и накопления L-аминокислоты в культуре и выделения L-аминокислоты из этой культуры.
Бактерии рода Escherichia, содержащие AKIII, в которой снято ингибирование L-лизином по типу обратной связи, включают в себя бактерии рода Escherichia, которые трансформированы путем встраивания ДНК, кодирующей AKIII с мутацией, при которой снимается ингибирование L-лизином по типу обратной связи в хромосомную ДНК, и бактерии рода Escherichia, которые трансформированы путем введения в клетки рекомбинантной ДНК, образованной при лигировании упомянутой выше ДНК с векторной ДНК, способной к автономной репликации в клетках бактерий рода Escherichia. Далее, также пригодны мутанты бактерий рода Escherichia, обеспечивающие получение мутантной AKIII в результате осуществления мутации клеток бактерий рода Escherichia.
Что касается бактерий рода Escherichia, которые используют в качестве хозяев для трансформации, могут быть использованы любые бактерии, если в их клетках действует промотор мутантного гена AKIII или другой промотор для экспрессии этого гена. Или, если мутантный ген AKIII вводят в плазмиду в виде внехромосомной ДНК, можно использовать любые бактерии, если в клетках может действовать источник репликации векторной ДНК, используемой для введения, и если этот источник может быть реплицирован.
Культуральной средой, используемой для инкубирования микроорганизма с мутантным геном, согласно настоящему изобретению, является обычная культуральная среда, содержащая источник углерода, источник азота, неорганические ионы и другие необходимые органические вещества.
Примеры источника углерода включают сахариды, такие как глюкоза, лактоза, галактоза, фруктоза и гидролизат крахмала; спирты, такие как глицерин и сорбит; и органические кислоты, такие как фумаровая кислота, лимонная кислота и янтарная кислота.
Примеры источника углерода включают неорганические аммонийные соли, такие как сульфат аммония, хлорид аммония и фосфат аммония; гидролизат соевых бобов, газообразный аммиак и водный аммиак.
В качестве источника органических питательных микроэлементов рекомендуется вносить подходящее количество необходимого вещества, такого как витамин B1 или L-изолейцин, или экстракт дрожжей. Далее, в требуемых количествах добавляют фосфат калия, сульфат магния, ионы железа и ионы марганца.
Инкубирование ведут в аэробных условиях в течение от 16 до 72 ч. Температура инкубирования находится между 25 и 45oС. рН устанавливается между 5 и 8 во время инкубирования. Для установления рН можно использовать неорганическое или органическое кислотное или щелочное соединение и еще газообразный аммиак.
L-аминокислоту обычно можно выделить из ферментационной жидкости при лигировании методом ионного обмена на смоле, осаждении и другими известными методами.
Примеры
Более конкретно, настоящее изобретение проиллюстрировано со ссылкой на следующие примеры.
Пример 1
Получение мутантного гена AKIII (1)
Клонирование дикого типа гена AKIII
О последовательности оснований гена AKIII E.coli (lysC) уже сообщалось [Cassan, М. , Parsot, С., Cohen, G.N. b Patte J.C., J.Biol.Chem., 261, 1052 (1986)] . Известно, что открытая считывающая рамка (ORF) состоит из 1347 пар оснований, кодирующих протеин, состоящий из 449 аминокислотных остатков. Этот ген содержит оператор и подвергается супрессии L-лизином. Поэтому, для того, чтобы удалить его операторный участок, последовательность SD и участок, содержащий только ORF, были амплифицированы с помощью PCR и клонированы.
Полную геномную ДНК штамма E.coli K-12 MC1061 получают по методу Saito и Miura [Biochem.Biophys.Acta., 72, 619 (1963)], и создают два типа праймеров с последовательностями, представленными Последовательностями 2 и 3. PCR осуществляют по методу H. A. Erlich et al. (PCR Technology, Stockton Press (1989)], используя то же для амплификации гена AKIII.
Полученную ДНК переваривают Bam HI и Ase I, затем ее концы затупляют и встраивают ее в сайт Sma I вектора с малым числом копий pMW119 для конструирования pMLYSC2. Этот сайт Sma I расположен в прямом направлении от промотора lacZ, находящегося в векторе. Когда рекомбинантную ДНК, полученную при встраивании фрагмента ДНК в Sma I сайт pMW119, вводят в E.coli, встроенный фрагмент ДНК транскрибируется путем транскрипции под контролем промотора lacZ (фиг.1).
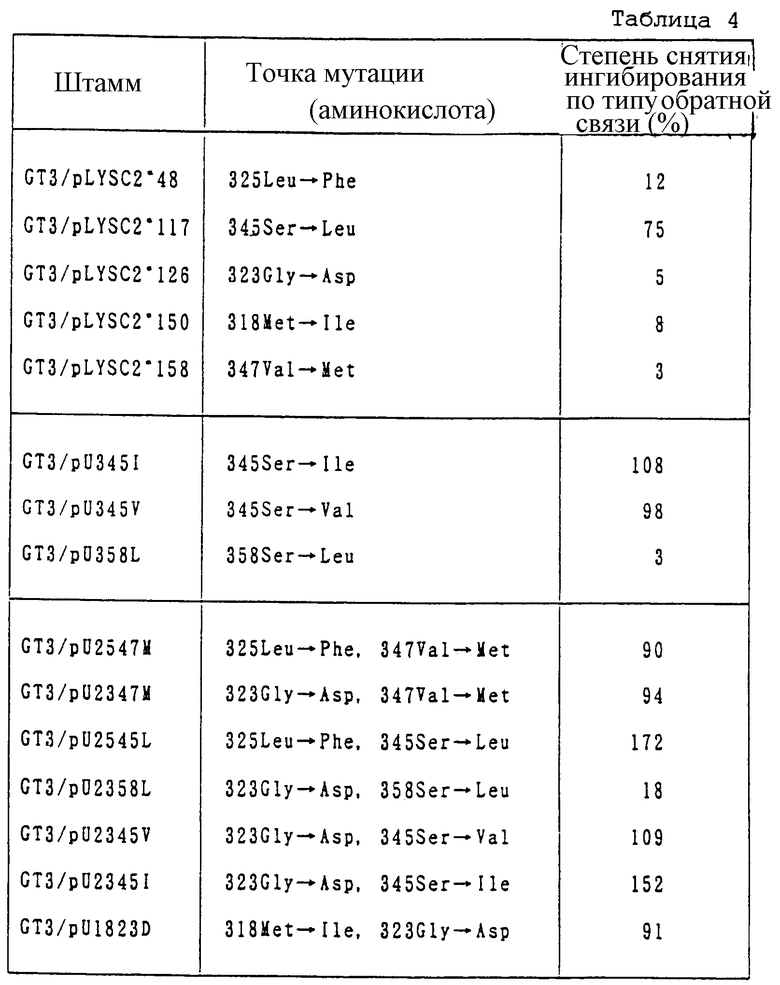
Клонирование мутантного гена AKIII
Плазмиды с мутантным геном AKIII, а именно pLYSCl*80, pLYSCl*117 и pLYSCl*126, как описано в International Laid-Open Pamphlets WО 95/11517 и WО 95/16042, переваривают Есо RI и Hind III для вырезания фрагментов ДНК, содержащих мутантный ген AKIII. Эти фрагменты встраивают в Есо RI-Hind III сайт pMW119 для создания плазмид, обозначенных pMLYSC2*80, pMLYSC2*117 и pMLYSC2*126, соответственно.
Плазмиду pLYSC*80 можно получить из pLLC*8 в соответствии с описанием в International Laid-Open Pamphlet WО 94/11517. Штамм, полученный при введении pLLC*80 в штамм E.coli НВ101, был обозначен AJ12750. Он был депонирован в Национальном Институте Биологических Наук и Технологии Человека Агенства Промышленной Науки и Технологии (No 1-3, Higashi 1-chrome, Tsukuba-shi, Ibaragi-ken, 305) под регистрационным FERM P-13136 1 сентября 1992. Этот штамм был передан в Международный депозитарий по Будапештскому договору 4 ноября 1993 и недавно ему был присвоен регистрационный No FERM ВР-4462. pLYSC2*117 и pLYSC2*126 еще не депонированы. Однако, т.к. точки их мутаций раскрыты в International Laid-Open Pamphlet WО 94/11517, они легко могут быть получены при использовании pLYSCl*80 в качестве исходного материала.
Изучение условий отбора нового мутантного гена AKIII
Трансформант, полученный при введении pMLYSC1 в штамм, полностью лишенный АК, E. coli GT3 (thrA1016B, metLM1005, 1уsС1004) был обозначен GT3/pMLYSC2. Также были получены GT3/pMLYSC2*80, GT3/pMLYSC2*117 и GT3/pMLYSC2*126. Штамм GT3/pMLYSC2 содержит плазмиду, включающую дикого типа 1уsС, и AKIII, закодированная в lysC этой плазмиды, является единственной АК. Так как дикого типа AKIII, являющаяся единственной АК, в присутствии значительных количеств L-лизина в минимальной среде ингибируется L-лизином, L-треонин, L-изолейцин, L-метионин и диаминопимелиновая кислота не могут быть синтезированы, поскольку они подавляют ее рост. Ожидая, что плазмидусодержащий штамм, содержащий мутантный lysC, в котором снято ингибирование L-лизином, можно выращивать в минимальной среде, содержащей значительное количество L-лизина, был выбран штамм, устойчивый при росте к L-лизину, и, таким образом, был отобран штамм, несущий плазмиду, содержащую мутантный lysC, в котором снято ингибирование L-лизином.
Штамм GT3/pMLYSC2, штамм GT3/pMLYSC2*80, штамм GT3/pMLYSC2*-117 и штамм GT3/pMLYSC2*126 инкубируют на агаровой пластине минимальной среды, содержащей L-лизин в различных концентрациях для определения ингибирующей рост концентрации и условий отбора плазмидусодержащих штаммов. Рост трансформантов на агаровой пластине минимальной среды, содержащей L-лизин в различных концентрациях, представлен в табл. 1. В табл. 1 + означает, что трансформант растет, ± означает, что трансформант растет слабо, и - означает, что трансформант не растет.
Рост штамма GT3/pMLYSC2, имеющего дикого типа 1уsС, был полностью подавлен в области добавления, содержащей 0.2-М лизина. Далее, рост штамма GT3/pMLYSC2*80, штамма GT3/pMLY-SC2*117 и штамма GT3/pMLYSC2*126, имеющих мутантный lysC, как хорошо известно, также почти полностью подавлены в области добавления, содержащей 0.4-М L-лизина. В результате было предположено, что мутант, обладающий более высокой степенью снятия ингибирования, чем известный до сих пор 1уsС, получается, по-видимому, при отборе в области добавления, содержащей 0.4-М L-лизина. Было установлено, что это ингибирование роста прекратилось при одновременном добавлении гомосерина и диаминопимелиновой кислоты.
В тесте на введение мутации, минимальная агаровая среда М9, содержащая 0.4-М L-лизина, была изпользована при отборе плазмидусодержащего штамма с мутантным lysC. В примере 1 эта среда называется селективной средой.
Осуществление мутации гена AKIII и получение мутантного гена AKIII
Для введения мутации в плазмиду pMLYSC1 используют два метода, а именно, метод мутагенеза in vitro, в котором плазмиду непосредственно обрабатывают гидроксиламином, и PCR мутагенез для обеспечения различных мутаций, в ожидании других мутаций, кроме мутации из цитозина в тимин с помощью гидроксиламина.
2 мкг pMLYSC2 обрабатывают 0.4-М гидроксиламином [раствор, содержащий 100 мкл смеси 0.1-М КН2РO4 и 1 mM ЕДТА (рН 6.0), и смесь 1-М гидроксиламина и 1 mM ЕДТА (рН 6.0) и 2 мкг ДНК, доводят до объема 200 мкл при добавлении воды] при 75oС в течение от 1 до 4 ч. Обработанную таким образом ДНК очищают стеклянным порошком и затем вводят в штамм E.coli GT3, полностью лишенный АК. Полученный трансформант высевают на полную среду (содержащую 1% L-бульона, 0,5% экстракта дрожжей, 0.5% NaCl, 50 мг/л ампициллина и 1.5% агар-агара) для образования колоний. Эти колонии реплицируют на селективную среду, и штаммы, способные расти на селективной среде, отбирают в качестве кандидатов.
Случайный мутагенез осуществляют с помощью PCR по методу С. Cadwell et al. (Cadwell, C. and G. F.Joyce, PCR methods Applic., 2_, 28 (1982)]. To есть, фрагмент lysC амплифицируют упомянутым выше методом с использованием pMLYSC2 и универсального праймера М13. Этот фрагмент переваривают Есо RI и Hind III и затем встраивают в Есо RI-Hind III сайт pMW119 для того, чтобы ввести его в Е. соli GT3, полностью лишенный АК. Трансформант высевают на полную среду (содержащую 1% L-бульона, 0.5% бактотриптона, 0.5% экстракта дрожжей, 0.5% NaCl, 50 мг/л ампициллина и 1.5% агар-агара) для образования колоний. Эти колонии реплицируют на селективную среду и штаммы, способные расти на селективной среде, выбирают в качестве кандидатов.
Плазмиды выделяют из совокупности 47 штаммов-кандидатов, полученных выше, т. е. 44 штамма, полученных в результате мутагенеза с использованием гидроксиламина, 3 штамма, полученных в результате случайного мутагенеза в результате PCR. Определены последовательности оснований мутантного lysC и были определены точки мутаций. Определение последовательностей оснований проводят обычным методом с использованием секвенсера ДНК ABI модели 373А (представленной ABI). В результате, из штаммов, полученных в результате мутагенеза с использованием гидроксиламина, получают мутантные AKIII штаммы 4-х типов ( 1, 6, 14 и 21) точек мутаций, а из штаммов, полученных в результате случайного мутагенеза в результате PCR, получают мутантные AKIII штаммы 3-х типов ( 28, 29 и 30) точек мутаций (табл. 2).
Эти семь плазмид (pMLYSC2*Yl, pMLYSC2*Y6, pMLYSC2*Y14, pMLYSC2*Y21, pMLYSC2*Y28, pMLYSC2*Y29 и pMLYSC2*Y30) вводят в GT3 штамм, полностью лишенный АК, и из трансформантов приготавливают бесклеточные экстракты. Используя их, определяют ферментативную активность AKIII. Получение бесклеточных экстрактов и определение ферментативной активности AKIII проводят по методу E.R.Stadtman et al. (Stadtman E.R., Cohen, G.N., LeBras G., and Robichon-Szulmajster, H. , J.Biol.Chem., 236, 2033 (1961)]. Далее, при определении ферментативной активности AKIII, к ферментному реакционному раствору добавляют L-лизин в различных концентрациях для определения степени снятия ингибирования L-лизином. Результаты представлены в табл. 3. Степень снятия ингибирования определяют как отношение АК остаточной активности в присутствии 0.4-М L-лизина к АК активности в отсутствие L-лизина.
Как ясно из приведенных выше результатов, мутантную АК III, у которой степень снятия ингибирования выше, чем у обычной типичной мутантной AKIII (мутантная AKIII, закодированная в pMLYSC2*80 и pMLYSC2*117), можно получить по использованному на этот раз методу селекции. На специфическую активность, основанную на полном протеине, обычно влияют условия роста клеток или приготовление образцов. Его специфическая активность равнялась активности исходного протеина и обычных мутантов, и не наблюдалось практически никакого снижения в активности при введении мутации. Соответственно ожидается, что цент активности AKIII и его контрольный сайт с L-лизином независимы друг от друга.
Пример 2
Получение мутантного гена AKIII (2)
Ген lysC, в котором 358-й Ser заменили на Leu, получен при введении сайт-специфичной мутации с использованием LA PCR in vitro Mutagenesis Kit (предоставленного Takara Shuzo Co., Ltd.). В этот раз pLYSC1, описанный в International Laid-Open Pamphlet WО 94/11517 и WО 95/16042, используют в качестве матрицы, а праймер, описанный в таблице Последовательностей 4, - в качестве праймера для введения мутации, соответственно. Оба конца PCR амплифицированного фрагмента расщепляют Eco RI и Hind III, а вырезанный фрагмент лигируют с фрагментом, полученным при расщеплении pUC19 Eco RI и Hind III pUC19 с образованием pLYSC358L. Мутантный ген AKIII, в котором 354-й Ser заменяют в результате мутации на Ilе или Val описанным выше методом, за исключением того, что в качестве праймера используют праймер, описанный в таблице Последовательностей 5 или 6. Этот ген лигируют с фрагментом, полученным при расщеплении pUC19 Eco RI и Hind III. Таким образом создают pLYSC345I и pLYSC354V.
Содержащие lysC фрагменты, полученные при расщеплении pLYSC1*48, pLYSC1*117, pLYSC1*126, pLYSC1*150 и pLYSC1*158 с помощью Eco RI и Hind III, встраивают в Есо-Hind III сайт расщепления pUC19, а полученные фрагменты обозначают pLYSC1*48, pLYSC1*117, pLYSC1*126, pLYSC1*150 и pLYSC1*158, соответственно. International Laid-Open Pamphlets WО 94/11517 и WО 95/16042 раскрывают структуру pLYSC1 и сайты точек мутаций pLYSC1*48, pLYSC1*117, pLYSC1*126, pLYSC1*150 и pLYSC1*158. Поэтому эти плазмиды можно получить из упомянутой выше pLYSC1*80.
Затем, мутантный ген lysC, имеющий два типа мутаций, получают с использованием расщепления по Ssp 1 сайту, находящемуся вблизи от центра точек мутаций полученного таким образом одноосновного мутантного lysC и Ssp 1 сайта расщепления, расположенного в прямом направлении от lysC в pUC19. Таким образом, pU2547M получают при лигировании фрагмента Ssp 1, содержащего lysC ген расположенного в обратном направлении участка pLYSC2*48 с фрагментом Ssp 1, содержащим lysC ген расположенного в прямом направлении участка pLYSC1*158; pU2547M получают при лигировании фрагмента Ssp 1, содержащего lysC ген расположенного в обратном направлении участка pLYSC2*126 с фрагментом Ssp 1, содержащим lysC ген расположенного в прямом направлении участка pLYSC2*158; pU2545L получают при лигировании фрагмента Ssp 1, содержащего lуsС ген расположенного в обратном направлении участка pLYSC2*48 с фрагментом Ssp 1, содержащим lysC ген расположенного в прямом направлении участка pLYSC2*117; pU2358L получают при лигировании фрагмента Ssp 1, содержащего lysC ген расположенного в обратном направлении участка pLYSC2*126 с фрагментом Ssp 1, содержащим lysC ген расположенного в прямом направлении участка pLYSC2358L; pU2345V получают при лигировании фрагмента Ssp 1, содержащего ген lysC расположенного в обратном направлении участка pLYSC2*126 с фрагментом Ssp 1, содержащим ген lysC расположенного в прямом направлении участка pLYSC2345V; и pU2345I получают при лигировании фрагмента Ssp 1, содержащего ген lysC расположенного в обратном направлении участка pLYSC2*l26, с фрагментом Ssp 1, содержащим ген lysC расположенного в прямом направлении участка pLYSC*2345I. Так как точки мутаций pLYSC2*150 и pLYSC2*126 расположены далее в обратном направлении, чем сайты расщепления Sps 1, двухосновный мутантный lysC, имеющий две точки мутаций, получают следующим образом. Сначала фрагмент ДНК с двумя точками мутаций амплифицируют, используя pLYSC2*126 в качестве матрицы, олигонуклеотид, описанный в таблице Последовательностей 7, как праймер, и упомянутый выше набор. Потом оба конца полученного фрагмента ДНК разрезают с помощью Eco RI и Hind III и лигируют с фрагментом, полученным при расщеплении pUC19 с помощью Eco RI и Hind III. Полученный таким образом продукт обозначают как pU1823D.
Степень снятия ингибирования полученного таким образом двухосновного мутантного lуsС генного продукта (AKIII) по сравнению с Lys измеряют по способу примера 1. Соответственно, степень снятия ингибирования любой двухосновной мутант-ной AKIII оказывается выше, чем таковая у одноосновной мутантной AKIII (табл. 4). В данном случае мутантный lysC в pLYSC582L, pLYSC345I и pLYSC345V оказывается новым мутантным геном. В каждом из генных продуктов снимается ингибирование лизином по типу обратной связи, а лигирование с другой мутацией еще больше повышает степень снятия ингибирования. Этот продукт также полезен в качестве интермедиата для конструирования двухосновной мутантной AKIII.
Пример 3
Получение L-лизина путем ферментации с использованием штамма с введенным в него мутантным геном DDPS и мутантным геном AKIII.
Эффект получения L-лизина с учетом мутантного гена DDPS и мутантного гена AKIII описан в International Laid-Open Pamphlet WО 95/16042. Для того чтобы усилить этот эффект, мутантный AKIII ген, полученный в примере 1, заставили сосуществовать с мутантным геном DDPS.
Штамм, полученный при введении плазмиды RSF24P с мутантным геном DDPS, описанным в International Laid-Open Pamphlet WО 95/16042, в штамм E.coli JM109, обозначают как AJ12395. Он был депонирован в Национальном Институте Биологических Наук и Технологии Человека Агенства Промышленной Науки и Технологии (No 1-3, Higashi 1-chrome, Tsukuba-shi, Ibaragi-ken, 305) под регистрационным No FERM P-13935 28 октября 1993. Этот штамм был передан в Международный депозитарий по Будапештскому договору 1 ноября 1994 и ему был присвоен регистрационный No FERM BP-4858.
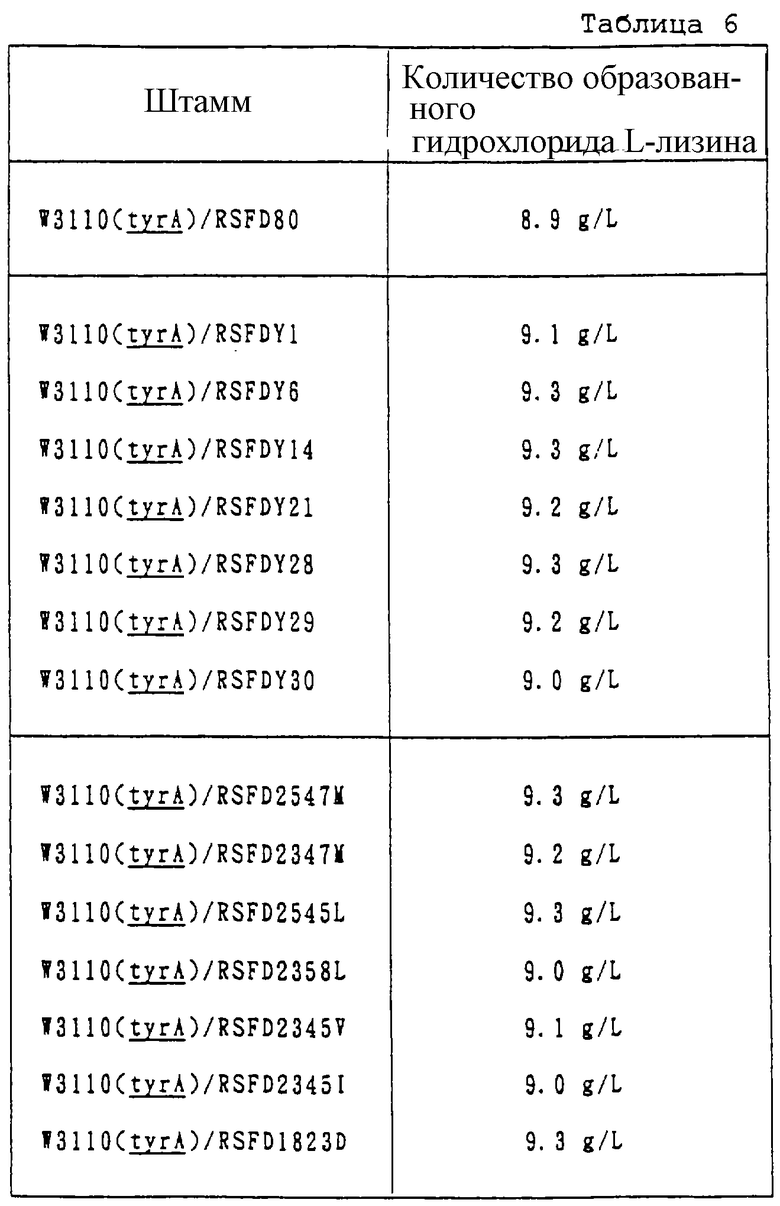
Плазмиды RSFDY1, RSFDY6, RSFDY14, RSFDY21, RSFDY28, RSFDY29, RSFDY30, RSFD2547M, RSFD2347M, RSFD2545L, RSFDY2358L, RSFD2345V, RSFD2345I и RSFD1823D получают, как показано на фиг. 2, из одного типа, выбранного из плазмид, содержащих мутантный ген AKIII, pMLYSC2*Yl, pMLYSC2*Y6, pMLYSC2*Y14, pMLYSC2*Y21, pMLYSC2*Y28, pMLYSC2*Y29, pMLYSC2*Y30, pU2547M, pU2347M, pU2545L, pU2358L, pU2545V, pU2345I и pU1823D.
Плазмиду RSFD80, описанную в International Laid-Open Pamphlet WО 95/16042, используют в качестве контроля. Штамм, полученный при введении плазмиды RSFD80 в штамм E.coli JM-109, обозначают как AJ12396. Он был депонирован в Национальном Институте Биологических Наук и Технологии Человека Агентства Промышленной Науки и Технологии ( 1-3, Higashi 1-chrome, Tsukuba-shi, Ibaragi-ken, 305) под регистрационным No FERM P-13936 28 октября 1993. Этот штамм был передан в Международный депозитарий по Будапештскому договору 1 ноября 1994 и ему был присвоен новый регистрационный No FERM ВР-4859.
Полученные выше плазмиды RSFDY1, RSFDY6, RSFDY14, RSFDY21, RSFDY28, RSFDY29, RSFDY30, RSFD2547M, RSFD2347M, RSFD2545L, RSFDY2358L, RSFD2345V, RSFD2345I и RSFD1823D вводят в штамм В-399 обычным способом для получения L-лизин-производящих штаммов. Проводят оценку производительности по лизину упомянутых выше штаммов. Оценку производительности по лизину также проводят по отношению B-399/RSFD80 в качестве контроля. Метод получения штамма В-399 описан в International Laid-Open Pamphlet WО 95/16042.
Инкубирование ведут в L-лизин-производящей культуральной среде при 37oС в течение 48 ч при перемешивании со скоростью от 114 до 116 об/мин, в соответствии с методом, описанным в International Laid-Open Pamphlet WО 95/16042. Результаты представлены в табл. 5.
Пример 4
Получение L-лизина путем ферментации с использованием штамма с введенным в него мутагенным геном DDPS и мутантным геном AKIII (2)
В примере 3 было установлено, что производство L-лизина можно повысить, вводя в бактерии рода Escherichia мутантный ген DDPS и мутантный ген AKIII. Этот тест проводят для того, чтобы установить, можно ли сохранить этот эффект даже при смене хозяина.
В качестве хозяина используют штамм E.coli W3110 (tyrA). Штамм W3110 (tyrA) подробно описан в European Patent Laid-Open 488424/1992. В European Patent Laid-Open No 488424/1992 описано большое число штаммов, полученных при введении плазмиды в штамм W3110 (tyrA). Например, штамм, полученный при введении плазмиды pHATerm, обозначают как штамм E.coli W3110 (tyrA)/pHATerm. Его международно депонировали в Национальном Институте Биологических Наук и Технологии Человека Агенства Промышленной Науки и Технологии ( 1-3, Higa-shi 1-chrome, Tsukuba-shi, Ibaragi-ken, 305) 18 ноября 1991 по Будапештскому договору под регистрационным No FERM BP-3653. Штамм W3110 (tyrA) также можно получить при удалении плазмиды pHATerm из этого штамма W3110 (tyrA)/pHATerm. Удаление плазмиды может быть проведено обычным способом.
Плазмиды, содержащие мутантный ген DDPS и мутантный ген AKIII, полученные в примере 3, а именно RSFDY1, RSFDY6, RSFDY14, RSFDY21, RSFDY28, RSFDY29, RSFDY30, RSFD2547M, RSFD2347M, RSFD2545L, RSFDY2358L, RSFD2345V, RSFD2345I и RSFD1823D, вводят в полученный выше штамм W3110 (tyrA) и проводят оценку производства L-лизина, как в примере 3. В качестве контроля получили W3110 (tyrA)/RSFD80 при введении RSFD80 в штамм W3110 (tyrA), а также провели оценку производства L-лизина этим штаммом. Результаты представлены в табл. 6.
Пример 5
Получение L-треонина путем ферментации с использованием штамма с введенным в него мутантным геном AKIII
Подобно E.coll треонин-производящему штамму, штамм В-3996 обладает самой высокой производительностью среди всех штаммов, известных к настоящему времени. Поэтому, при оценке мутантной AKIII, штамм В-3996 использовали в качестве хозяина. Этот штамм В-3996 описан в патенте США 5175107 и указан как депонированный в Исследовательском Институте Генетики и Промышленного Разведения Микроорганизмов под регистрационным No BKIIM В-3996. Далее, для оценки в качестве мутантного гена AKIII, из мутантных генов, обладающих высокой L-лизин-производительностью, в примере 4 выбирают и тестируют три типа мутантных генов (мутантные гены AKIII RSFDY6, RSFE254M и RSFD1823D). Сначала для того, чтобы повысить количество экспрессируемого мутантного гена AKIII, мутантный ген AKIII, присутствующий в pMLYSC2*Y6, лигируют с расположенным в прямом направлении участком lacZ промотора pUC19 (предоставленного Takara Shuzo Co. , Ltd. ). Образованную таким образом новую плазмиду обозначают рLLС*Y6 (фиг. 3). Так как в pU2547M и pU1823D мутантный ген AKIII первоначально расположен в прямом направлении от lacZ промотора pUC19 (предоставленного Takara Shuzo Co., Ltd.), он был использован как таковой.
Эти плазмиды вводят в штамм В-3996 обычным образом и проводят оценку. Инкубирование проводят по методу, описанному в International Laid-Open Pamphlet WО 94/11517.
PLLC*Y6, pU2547M и pU1823D трансформируют в штамм В-3996 и инкубируют трансформанты в присутствии или в отсутствие 1 г/л лизина. В качестве контролей используют один штамм-хозяин В-3996 и B-3996/pLLC*80, описанный в International Laid-Open Pamphlet WО 94/11517.
Результаты представлены в табл. 7. Восприимчивость к лизину в табл. 7 относится к отношению (выхода сахара, потребленного в присутствии лизина) к (выходу сахара, потребленного в отсутствие лизина). Для штамма В-3996 снижение выхода сахара, потребленного при инкубировании в присутствии лизина, составляет приблизительно 0.74 по отношению к выходу в области инкубирования в отсутствие лизина; а в В-3996/-pLLC*80 снижение выхода сахара, потребленного при инкубировании в присутствии лизина, составляет приблизительно 0.90 по отношению к выходу в области инкубирования в отсутствие лизина. Вновь полученные мутантные гены AKIII на этот раз являются несколько превосходящими по производству треонина и чувствительности к лизину.
Эффекты изобретения
Настоящее изобретение дает возможность получать мутантный ген AKIII, полученный из бактерий рода Escherichia, в котором эффективно снимается ингибирование L-лизином по типу обратной связи. Улучшенный по сравнению с предыдущими штамм, производящий L-аминокислоту, можно получить путем введения упомянутого выше гена в бактерии рода Escherichia. Возможно обеспечить способ получения L-аминокислоты путем ферментации, который превосходит обычный способ в результате использования этого штамма, производящего L-аминокислоту.
Краткое описание чертежей
На фиг.1 показан способ получения pMLYSC2.
На фиг.2 показан способ получения плазмид серий RSFD.
На фиг.3 показан способ получения pLLC*Y6.















Изобретение относится к способу получения L-лизина путем ферментации, а также к ДНК, которая используется в этом способе. Представленная ДНК кодирует аспартокиназу III бактерий, принадлежащих к роду Escherichia и используемых для получения L-аминокислот, и содержит в кодирующем участке мутацию, за счет которой снимается ингибирование лизином по типу обратной связи указанной аспартокиназы III. Способ получения L-лизина включает получение и введение указанной ДНК в клетки для образования бактерий-трансформантов рода Escherichia. Далее проводят инкубирование этих бактерий в соответствующей культуральной среде, в результате чего L-лизин производится и накапливается в этой культуре, и, в дальнейшем, выделение L-лизина из нее. Представленные изобретения позволяют получать аспартокиназу III из бактерий рода Escherichia, в которых хорошо снимается ингибирование L-лизином по типу обратной связи. 2 с.п. ф-лы, 7 табл., 3 ил.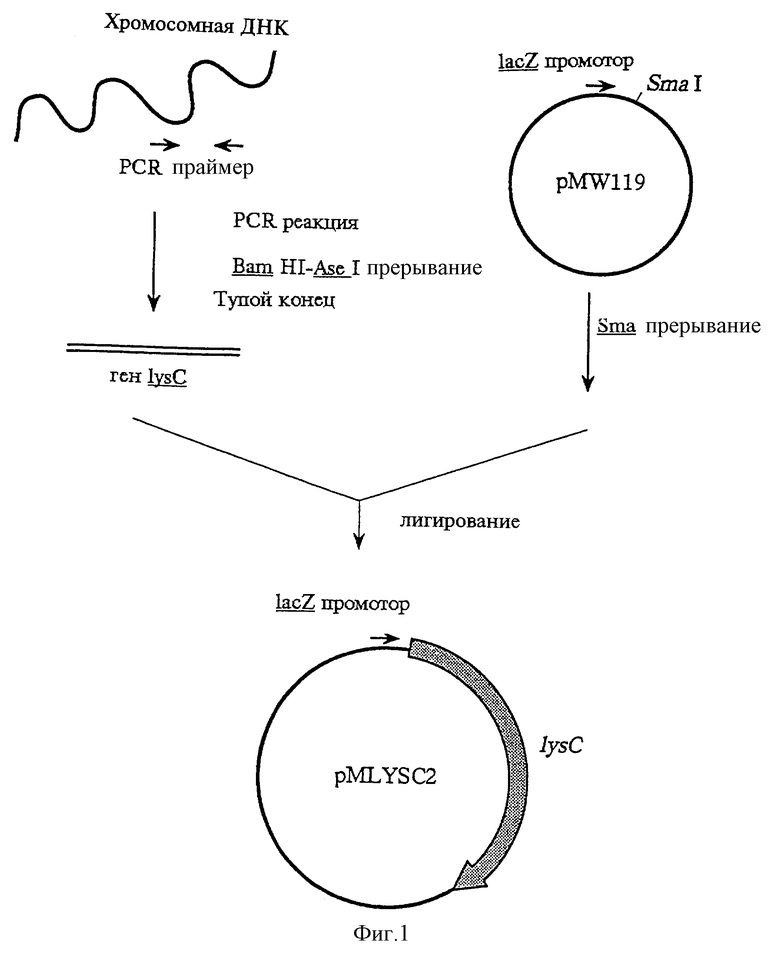
| Экономайзер | 0 |
|
SU94A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ АСПАРТОКИНАЗУ III, СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 1993 |
|
RU2113484C1 |
