Данное изобретение касается мутантной фосфоенолпируваткарбоксилазы, кодирующего ее гена и способа получения аминокислоты, и, в частности, касается гена, имеющего мутацию, вызывающую десенсибилизацию (снижение чувствительности) к ингибированию аспарагиновой кислотой по типу обратной связи, и его применение.
Предпосылки изобретения
Фосфоенолпируваткарбоксилаза является ферментом, который обнаружен почти во всех бактериях и во всех растениях. Этот фермент играет роль в биосинтезе аспарагиновой кислоты и глутаминовой кислоты и подаче C4-дикарбоновой кислоты в цикл лимонной кислоты для поддержания его оборота. Однако, в области ферментативного получения аминокислот при помощи микроорганизмов было несколько сообщений о действии этого фермента, которые не прояснили его роли (Atsushi Yokota and Isamu Shiio, Agric. Biol. Chem., 52, 455-463 (1988), Josef Cremer et al., Appl. Environ. Microbiol., 57, 1746-1752 (1991), Petra, G. Peters-Weintisch, FEMS Microbiol. Letters, 112, 269-274 (1993)).
Между прочим, аминокислота является соединением, которое повсеместно существует в клетках в виде компонентов белков, однако, в целях экономичного метаболизма энергии и метаболизма веществ ее образование строго контролируется. Этот контроль представляет собой в принципе контроль по типу обратной связи, в котором конечный продукт метаболического пути ингибирует активность фермента, катализирующего более раннюю стадию этого пути. Фосфоенолпируваткарбоксилаза также подвергается различным типа регуляции в проявлении ее активности.
Например, в случае фосфоенолпируваткарбоксилазы микроорганизмов, принадлежащих к роду Corynebacterium или к роду Escherichia, активность фермента ингибируется аспарагиновой кислотой. Таким образом, вышеупомянутый биосинтез аминокислоты, в котором участвует фосфоенолпируваткарбоксилаза, также ингибируется аспарагиновой кислотой.
В предшествующих исследованиях были разработаны различные способы эффективного получения аминокислот при помощи ферментации и ферментативным путем получали лейцин, изолейцин, триптофан, фенилаланин и т.п. с применением мутантных штаммов, которые были сделаны нечувствительными к контролю по типу обратной связи. Однако, не было известно ни о мутантной фосфоенолпируваткарбоксилазе, превращенной в фермент, нечувствительный к ингибированию аспарагиновой кислотой, ни о попытках использовать этот фермент для ферментативного получения аминокислот из семейства аспарагиновой кислоты и семейства глутаминовой кислоты.
С другой стороны, уже был клонирован ppc ген, который является геном, кодирующим фосфоенолпируваткарбоксилазу Escherichia coli, и была также определена его нуклеотидная последовательность (Fujita, N., Miwa, T., Ishijima, S. , Izui, K. and Katsuki, H., J. Biochem., 95, 909-916 (1984). Однако нет сообщений о мутанте, в котором снижена чувствительность к ингибированию аспарагиновой кислотой.
Данное изобретение осуществлено на основании указанной точки зрения и целью данного изобретения является создание мутантной фосфоенолпируваткарбоксилазы с существенно сниженной чувствительностью к ингибированию аспарагиновой кислотой по типу обратной связи, обеспечению гена, кодирующего фермент, и способа его применения.
Описание изобретения
В результате усердного исследования для достижения указанной выше цели авторы данного изобретения обнаружили, что ингибирование аспарагиновой кислоты существенно десенсибилизируется посредством замены аминокислоты в специфическом сайте фосфоенолпируваткарбоксилазы Escherichia coli другой аминокислотой, что привело к получению гена, кодирующего мутантный фермент, что и явилось завершением данного изобретения.
Конкретно, данное изобретение заключается в получении мутантного фосфоенолпируваткарбоксилазы, происходящей из микроорганизма рода Escherichia и имеющей мутацию для снижения чувствительности (десенсибилизации) к ингибированию аспарагиновой кислотой по типу обратной связи, и последовательности ДНК, кодирующей мутантную фосфоенолпируваткарбоксилазу.
Далее данное изобретение обеспечивает микроорганизмы, принадлежащие к роду Escherichia или к коринеформным бактериям, несущие в себе этот фрагмент ДНК, и способ получения аминокислоты, предусматривающий культивирование любого из этих микроорганизмов в предпочтительной среде и выделение аминокислот, выбранных из L-лизина, L-треонина, L-метионина, L-изолейцина, L-глутаминовой кислоты, L-аргинина и L-пролина, из этой среды.
В применении к этому описанию ДНК последовательность, кодирующую мутантную фосфоенолпируваткарбоксилазу, или последовательность ДНК, содержащую, кроме того, промотор, иногда называют просто "последовательность ДНК данного изобретения", "мутантным геном" или "геном фосфоенолпируваткарбоксилазы".
Далее данное изобретение будет объяснено подробно.
<1> Мутантная фосфоенолпируваткарбоксилаза.
Мутантная фосфоенолпируваткарбоксилаза данного изобретения (далее называемая "мутантным ферментом") основана на фосфоенолпируваткарбоксилазе микроорганизма, принадлежащего к роду Escherichia, имеющему мутацию, вызывающую десенсибилизацию (снижение чувствительности) к ингибированию по типу обратной связи аспарагиновой кислоты.
Такая мутация может быть любой мутацией при условии, что чувствительность к ингибированию по типу обратной связи существенно снижается без потери ферментативной активности фосфоенолпируваткарбоксилазы. В качестве примера можно привести мутацию, которая в случае существования мутантной фосфоенолпируваткарбоксилазы с такой мутацией в клетках микроорганизма Escherichia придает этим клеткам устойчивость к соединению, имеющему следующие свойства:
соединение проявляет ингибирующее рост действие против микроорганизма рода Escherichia, продуцирующего немутантную фосфоенолпируваткарбоксилазу (дикого типа);
указанное ингибирующее рост действие снимается присутствием L-глутаминовой кислоты или L-аспарагиновой кислоты; и
соединение ингибирует фосфоенолпируваткарбоксилазную активность дикого типа.
Более конкретно, в качестве примеров таких мутацией могут быть приведены следующие мутации (нумерация от N-конца фосфоенолпируваткарбоксилазы):
(1) мутация для замены глутаминовой кислоты 625 лизином;
(2) мутация для замены 222 гистидином и глутаминовой кислоты 223 лизином, соответственно;
(3) мутация для замены серина 288 фенилаланином, глутаминовой кислоты 289 лизином, метионина 551 изолейцином и глутаминовой кислоты 804 лизином соответственно;
(4) мутация для замены аланина 867 треонином;
(5) мутация для замены аргинина 438 цистеином; и
(6) мутация для замены лизина 620 серином.
Для фосфоенолпируваткарбоксилазы микроорганизма, принадлежащего к роду Escherichia, аминокислотная последовательность, выведенная из гена фосфоенолпируваткарбоксилазы Escherichia coli (Fujita, N., Miwa, T., Ishijima, S., Izui, K. и Katsuki, H., J. Biochem., 95, 909-916 (1984)), показана в SEQ ID N 2 в Списке последовательностей. Кроме того, полная нуклеотидная последовательностью плазмиды pT2, содержащей ген фосфоенолпируваткарбоксилазы Escherichia coli, показана в SEQ ID N 1 вместе с аминокислотной последовательностью (см. в конце описания).
Указанные выше мутантные ферменты кодируются последовательностями ДНК данного изобретения, описанными ниже, которые образуются путем экспрессии этих последовательностей ДНК в Escherichia coli и т.п.
<2> Последовательность ДНК данного изобретения и содержащие ее микроорганизмы.
Последовательность ДНК данного изобретения включает последовательности ДНК, кодирующие вышеупомянутые мутантные ферменты, и имеет мутацию, вызывающую десенсибилизацию к ингибированию по типу обратной связи фосфоенолпируваткарбоксилазы аспарагиновой кислотой, в кодирующих районах во фрагментах ДНК, кодирующих фосфоенолпируваткарбоксилазу микроорганизма, принадлежащего к роду Escherichia.
Конкретно можно провести в качестве примера последовательность ДНК, кодирующую фосфоенолпируваткарбоксилазу, имеющую любую из указанных выше мутаций (1) - (6). В отношении нуклеотидной последовательности SEQ ID N 1 можно привести в качестве примера последовательность ДНК, имеющую любую из следующих мутаций:
i) мутацию для превращения GAA с номерами оснований 2109-2111 в AAA или AAG;
ii) мутацию для превращения CGC с номерами оснований 900-902 в CAT или CAC и GAA 903-905 в AAA или AAG соответственно;
iii) мутацию для превращения TCT с номерами оснований 1098-1100 в TTT или TTC, GAA 1101-1103 в AAA или AAC, ATG 1887-1889 в ATT, ATC или ATA и GAA 2646-2648 в AAA или AAG соответственно:
iv) мутацию для превращения GCG 2835-2837 в любой из кодонов ACT, ACC, ACA и ACG; и
v) мутацию для превращения CGT 1548-1550 в TGT или TGC; и
vi) мутацию для превращения AAA 2094-2096 в TCT, TCC, TCA или TCG.
Такой мутантный ген получают таким образом, что рекомбинантную ДНК, полученную лигированием гена фосфоенолпируваткарбоксилазы в виде немутантного гена фермента или имеющего другую мутацию гена, с вектором ДНК, адаптированным к хозяину, подвергают мутирующей обработке для проведения скрининга трансформантов при помощи рекомбинантной ДНК. Альтернативно приемлемо также подвергнуть микроорганизм, продуцирующий немутантный фермент, мутирующей обработке, в результате которой получают мутантный штамм, продуцирующий мутантный фермент, а затем мутантный ген скринируют из мутантного штамма. Для получения мутации рекомбинантной ДНК применяют гидроксиламин и т.п. Далее при обработке самого микроорганизма условиями мутагенеза можно применять лекарственное средство или способ, обычно применяемые для искусственного мутагенеза.
Далее в соответствии с такими способами, как способ перекрывающегося удлинения (Ho, S.N., Hunt, H.D., Horton, R.M., Publen, J.K. and Pease, L.R., Gene, 77, 51-59 (1989)), способ сайт-специфического мутагенеза (Kramer, W. and Frits, H. J., Meth. in Enzymol., 154, 350 (1987); Kunkel, T.A. et al., Meth. in Enzymol., 154, 367 (1987)) и др., указанный мутантный ген может быть также получен путем введения мутации, такой как замена аминокислоты, инсерция, делеция и т.п., в ген фосфоенолпируваткарбоксилазы в виде немутантного гена фермента или в виде гена, имеющего другую мутацию. Эти способы основаны на принципе, что немутированный ген ДНК применяют в качестве матрицы, а синтетическую ДНК, содержащую ошибочное спаривание оснований в точке мутации, применяют в качестве одного из праймеров для синтеза комплементарных цепей указанного гена ДНК, вследствие чего происходит введение мутации. С применением этих способов можно получить предполагаемую мутацию в целевом сайте.
Альтернативно синтезируют последовательность, которая имеет сайты расщепления рестриктазами на обоих концах и включает обе стороны точки мутации, и заменяют ею соответствующую часть немутированного гена, в результате чего может быть введена мутация (кассетный способ мутирования).
Ген фосфоенолпируваткарбоксилазы, представляющий немутантный ген фермента или имеющий другую мутацию, используемый для введения мутации, может быть любым геном этого фермента при условии, что он является геном, кодирующим фосфоенолпируваткарбоксилазу микроорганизма, принадлежащего к роду Escherichia, который предпочтительно имеет определенную последовательность оснований (т. е. секвенирован) и клонирован. Если он не был клонирован, можно амплифицировать и выделить фрагмент ДНК, содержащий этот ген, при помощи PCR-метода и т.п. с последующим применением подходящего вектора для достижения клонирования.
В качестве примера гена, описанного выше, можно привести ген Escherichia coli, который был клонирован и секвенирован (Fujita, N., Miwa, T., Ishijima, S. , Izui, K. and Katsuki, H., J. Biochem., 95, 909-916 (1984)). Последовательность в кодирующем районе этого гена показана в SEQ ID N 1 (основания N 237-2888).
Скрининг хозяина, несущего этот мутантный ген, может быть выполнен с применением соединения-аналога аспарагиновой кислоты. Соединение-аналог имеет предпочтительно следующие свойства. Оно проявляет ингибирующее рост действие против микроорганизма, принадлежащего к роду Escherichia, продуцирующего немутантную фосфоенолпируваткарбоксилазу, это ингибирующее действие снимается присутствием L-глутаминовой кислоты или L-аспарагиновой кислоты, и аналог ингибирует фосфоенолпируваткарбоксилазу дикого типа.
Если из микроорганизмов рода Esсherichia выбран мутантный штамм, устойчивый к соединению-аналогу, указанному выше, например, Escherichia coli HB101, продуцирующий фосфоенолпируваткарбоксилазу дикого типа, с применением ингибирования роста этого микроорганизма в качестве критерия, то существует большая вероятность получения микроорганизма-хозяина, который продуцирует фосфоенолпируваткарбоксилазу с пониженной чувствительностью к ингибированию по типу обратной связи аспарагиновой кислотой.
В качестве общей структуры ингибитора фосфоенолпируваткарбоксилазы предлагается в основном структура C4-карбоновых кислот. С этой точки зрения, авторы данного изобретения подвергали скринингу различные соединения. Escherichia coli HB101 культивировали в LB среде и переносили на M9 среды (содержащие 20 мкг/мл тиамина и 3 мкг/мл каждого из Leu и Pro), содержащие также одно из следующих соединений: DL-2-амино-4-фосфономасляную кислоту, бромянтарную кислоту, мезо-2,3-дибромянтарную кислоту, 2,2-дифторянтарную кислоту, 2-бромпировиноградную кислоту, α-кетомасляную кислоту, α-кетоадипиновую кислоту, DL-трео-β-гидроксиаспарагиновую кислоту, β-метиловый эфир L-аспарагиновой кислоты, α-метил-DL-аспарагиновую кислоту, 2,3-диаминоянтарную кислоту или β-гидразид аспарагиновой кислоты, и поглощение этой среды измеряли при 660 нм по мере увеличения времени, наблюдая за ростом.
Далее, когда эти соединения присутствовали при концентрациях, ингибирующих рост, исследовали, снималось ли это ингибирование добавлением нуклеиновых кислот (уридином, аденозином, каждый в концентрации 10 мг/дл), глутаминовой кислоты или аминокислотами семейства аспарагиновой кислоты (Asp: 0,025%, каждый из Met, Thr, Lys: 0,1%).
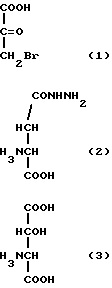
В результате были выбраны три соединения: 3-бромпируват (3BP) (1), аспартат-β-гидразид (AHУ) (2) и DL-трет-β-гидроксиаспартат (βHA) (3).
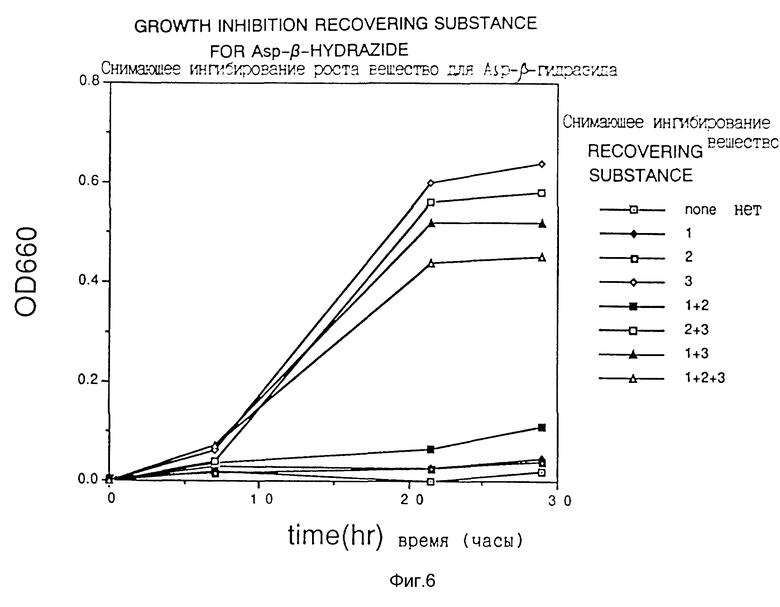
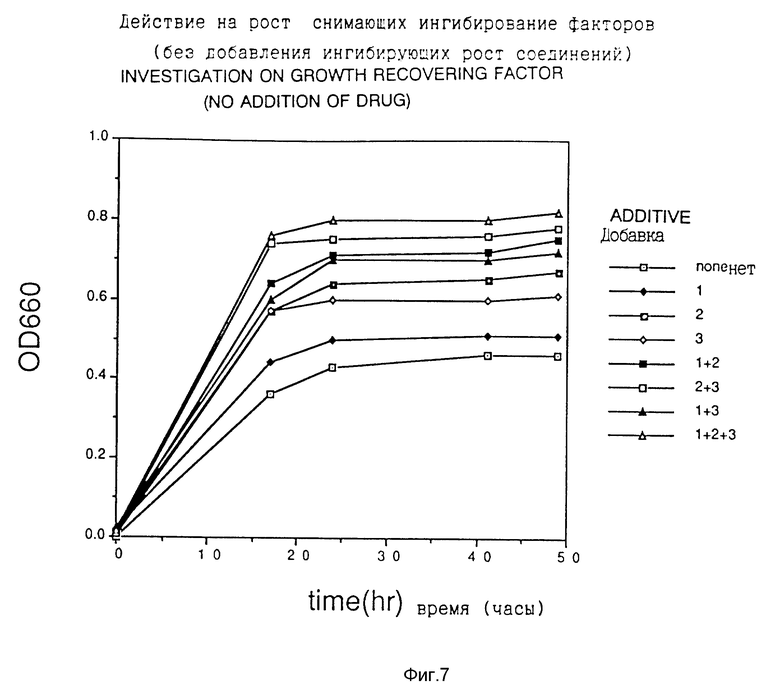
Ингибирование роста Escherichia coli этими соединениями-аналогами показано на фиг. 1-3. Далее восстановление роста Escherichia coli в случае добавления снимающих ингибирование веществ, отдельно или в виде смеси 2 или 3 типов веществ, показано на фиг. 4-6. Кроме того, в качестве контроля рост в случае добавления снимающих ингибирование веществ в отсутствие ингибирующего вещества показан на фиг. 7. На фиг. 4-7 добавки 1, 2 и 3 обозначают нуклеиновые кислоты, глутаминовую кислоту или аминокислоты семейства аспарагиновой кислоты соответственно.
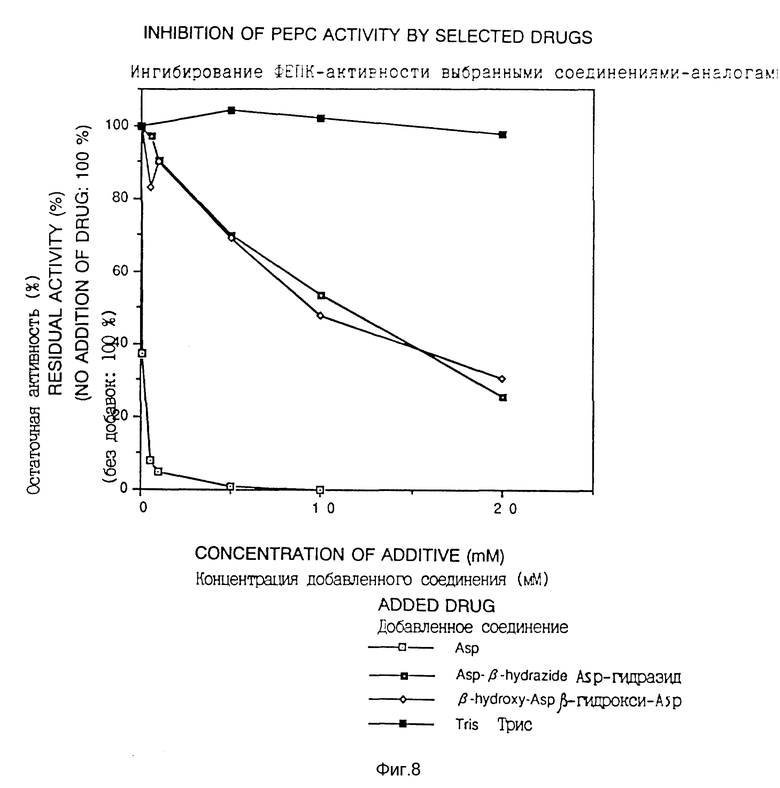
Далее ингибирование активности соединением-аналогом фосфоенолпируваткарбоксилазы также исследовали. Неочищенный фермент получали из штамма Escherichia coli HB101 согласно способу, описанному в Journal of Biochemistry, vol. 67, N 4 (1970) и активность фермента измеряли согласно методу, описанному в Eur. J. Biochem., 202, 797-803 (1991).
Клетки Escherichia coli HB101, культивируемые в LB среде, разрушали, и суспензию центрифугировали для получения супернатанта, который применяли в качестве раствора неочищенного фермента. Измеряли активность фермента измерением уменьшения в поглощении при 340 нм, добавляя ацетил-кофермент A, который, как известно, влияет на активность фермента, в концентрации 0,1 мМ в систему для измерения, содержащую 2 мМ фосфоенолпируват калия, 0,1 мМ НАДН, 0,1 М трис-ацетат (pH 8,5), 1,5 E малатдегидрогеназы и неочищенный фермент. Результаты представлены на фиг. 8.
В соответствии с представленными выше результатами очевидно, что указанные три соединения ингибируют рост Escherichia coli, это ингибирование не может быть снято добавлением только нуклеиновых кислот, но оно может быть снято добавлением глутаминовой кислоты или аминокислот семейства аспарагиновой кислоты. Таким образом, эти соединения-аналоги являются избирательными ингибиторами фосфоенолпируваткарбоксилазы. Как показано в описанных ниже примерах, с применением этих соединений в данном изобретении удалось выбрать Escherichia coli, которая продуцирует мутантную фосфоенолпируваткарбоксилазу.
При скрининге трансформанта, имеющего целевой ген мутантного фермента, при помощи указанных соединений и выделении рекомбинантной ДНК получают мутантный ген фермента. Альтернативно в случае мутирующей обработки самого микроорганизма, когда мутантный штамм, имеющий целевой ген мутантного фермента, скринируют при помощи указанных соединений, из штамма выделяют фрагмент ДНК, содержащий целевой ген мутантного фермента, и его лигируют с подходящим вектором и затем получают ген мутантного фермента.
С другой стороны, в результате обстоятельного исследования авторов данного изобретения, обращающих внимание на важность остатка аргинина в аспартатсвязующем белке Esсherichia coli (Krikos, A., Mouth, N., Boyd, A. and Simon, M. I. Cell, 33, 615-622 (1983), Mowbray, S.L. and Koshland, D.E. J. Biol. Chem., 264, 15638-15643 (1990), Milburn, M.V., Prive. G.G., Milligan, D.L., Scott, W.G., Yeh, J., Jancarik, J., Koshland, D.E. and Kim, S.H., Science, 254, 1342-1347 (1991)), было обнаружено, что ингибирование аспарагиновой кислотой существенно десенсибилизируется превращением аргинина 438 фосфоенолпируваткарбоксилазы в цистеин. Для превращения аргинина 438 в цистеин кодон аргинина 438 гена, кодирующего фосфоенолпируваткарбоксилазу, может быть превращен в кодон цистеина. Например, в SEQ ID N 1 CGT нуклеотиды 1548-1550 могут быть превращены в TGT или TGC.
Далее авторы данного изобретения осуществили химическую модификацию остатков лизина фосфоенолпируваткарбоксилазы при помощи 2,4,6-тринитробензолсульфоновой кислоты (TNBS), которая является соединением, химически модифицирующим остатки лизина в белке. Во время реакции модификации присутствовала также яблочная кислота, способная действовать как ингибитор фосфоенолпируваткарбоксилазы. Предполагалось, что остаток лизина вблизи связывающего положения фосфоенолпируваткарбоксилазы мог бы быть защищенным путем связывания яблочной кислоты и не подвергаться химической модификации. В результате было сделано предположение, что остаток лизина 620 был важен для связывания яблочной кислоты с фосфоенолпируваткарбоксилазой, и было обнаружено, что ингибирование по типу обратной связи аспарагиновой кислотой было десенсибилизировано при сохранении ферментативной активности фосфоенолпируваткарбоксилазы посредством превращения остатка лизина 620 в остаток серина. Для превращения остатка лизина в остаток серина кодон лизина 620 гена, кодирующего фосфоенолпируваткарбоксилазу, может быть превращен в кодон серина. Например, в SEQ ID N 1 AAA с номерами нуклеотидов 2094-2096 может быть заменен TCT, TCC, TCA, TCG, AGT или AGC.
В соответствии с такими способами, как способ перекрывающегося удлинения (Ho, S. N. , Hunt, H.D., Horton, R.M., Pullen, J.K., and Pease, L.R., Gene, 77, 51-59 (1989)), способ сайт-специфической мутации (Kramer, W. and Frits, H. J. , Meth. in Enzymol., 154, 350 (1987); Kunkel, T.A. et al., Meth. in Enzymol., 154, 367 (1987)) и т.п., превращение этого кодона может быть достигнуто введением такой мутации, как замена аминокислоты, инсерция, делеция и т. п. , в ген фосфоенолпируваткарбоксилазы в виде гена фермента дикого типа или в виде гена, имеющего другую мутацию. Эти способы основаны на принципе, что немутированный ген ДНК используют в качестве матрицы, а синтетическую ДНК, содержащую ошибочное спаривание при точке мутации, используют в качестве одного из праймеров для синтеза комплементарных цепей вышеупомянутого гена ДНК, тем самым вводя мутацию. При помощи этих способов можно получить желаемую мутацию в целевом сайте.
Альтернативно синтезируют последовательность, имеющую сайты расщепления рестриктазами на обоих концах и содержащую обе стороны точки мутации, и заменяют ею соответствующую часть немутированного гена, посредством чего может быть введена мутация (кассетный способ мутирования).
Фрагмент ДНК, кодирующий фосфоенолпируваткарбоксилазу с введенной, как описано выше, мутацией экспрессируют с применением подходящей системы хозяин-вектор, при помощи которой можно получить мутантный фермент. Альтернативно даже выполнением трансформации посредством встраивания фрагмента ДНК данного изобретения в хромосомную ДНК хозяина можно получить целевой мутантный фермент.
В качестве хозяина могут применяться микроорганизмы, принадлежащие к роду Escherichia, например Escherichia coli, коринеформные бактерии и т.п. Коринеформные бактерии включают в себя бактерии, принадлежащие к роду Corynebacterium, бактерии, принадлежащие к роду Brevibacterium, классифицированные до настоящего времени как принадлежащие к роду Brevibacterium, но в настоящее время объединенные как бактерии, принадлежащие к роду Corynebacterium, и бактерии, действительно принадлежащие к роду Brevibacterium, близко родственные бактериям, принадлежащим к роду Corynebacterium. Хозяева, предпочтительные для получения аминокислоты, будут описаны ниже.
С другой стороны, в качестве вектора ДНК предпочтителен плазмидный вектор и предпочтительны плазмидные векторы, способные самореплицироваться в клетке-хозяине. В случае хозяина Escherichia coli, например, применимы pUC19, pUC18, pBR322, pHSG299, pHSG399, RSF1010 и т.п. Альтернативно можно также использовать в качестве вектора ДНК фага.
Далее в случае, если хозяином является коринеформная бактерия, могут использоваться векторы и несущие их хозяева, указанные ниже. В этих примерах номера депозитов в международных хранилищах (депозитариях) показаны в скобках.
pAj655 Escherichia coli Aj11882 (FERM BP-136) Corynebacterium glutamicum SR8201 (ATCC 39135)
pAj1844 Escherichia coli Aj11883 (FERM BP-137) Corynebacterium glutamicum 8202 (ATCC 39136)
pAj611 Escherichia coli Aj11884 (FERM BP-138)
pAj3148 Corynebacterium glutamicum 8203 (ATCC 39137)
pAj440 Bacillus subtilis Aj11901 (FERM BP-140)
Эти векторы могут быть получены из депонированных микроорганизмов следующим образом. Клетки, собранные в логарифмической фазе роста, подвергают бактериолизу с применением лизозима и додецилсульфата натрия (SDS) и центрифугируют при 30000g для получения раствора супернатанта из лизата, к которому добавляют полиэтиленгликоль для выполнения отделения и очистки векторов при помощи равновесного центрифугирования в градиенте плотности хлорид цезия-бромида этидия.
Для трансформации Escherichia coli рекомбинантным вектором, полученным встраиванием последовательности ДНК данного изобретения в указанный выше вектор, можно использовать способ, обычно применяемый для трансформации Escherichia coli, например способ, в котором клетки обрабатывают хлоридом кальция для усиления проникаемости ДНК (Mandel, M. and Higa, A., J. Mol. Biol., 53, 159 (1977)) и т.п.
Далее в качестве способа трансформации коринеформных бактерий также применяют указанный выше способ, в котором клетки обрабатывают хлоридом кальция, или способ, в котором включение выполняют при специфическом периоде роста, в котором клетки могут включать в себя ДНК (сообщение в отношении Bacillus subtilis Duncan, C. H. et al.). Далее включение в бактериальные клетки может быть достигнуто путем образования протопластов или сферопластов реципиентов ДНК, которые легко включают плазмидную ДНК. Они известны для Bacillus subtilis, Actinomyces и дрожжей (Chang, S. et al., Molec. Gen. Genet. , 168, 111 (1979), Bibb et al., Nature 274, 398 (1978), Hinnen, A. et al. , Proc. Natl. Acad. Sci. USA, 75 1929 (1978)). Дополнительно способ трансформации коринеформных бактерий описан в JP Laid-open N 2-207791.
Для экспрессии последовательности ДНК данного изобретения в указанных выше хозяевах можно использовать такие промоторы, как lac, trp, PL и т.п., которые эффективно работают в микроорганизмах, или когда последовательность данного изобретения содержит промотор гена фосфоенолпируваткарбоксилазы, этот промотор можно использовать как таковой. Альтернативно в случае применения коринеформной бактерии в качестве хозяина можно также использовать известный trp промотор из бактерии, принадлежащей к роду Brevibacterium (JP Laid-open N 62-244382) и др.
Далее, как описано выше, допустимо, чтобы последовательность ДНК данного изобретения была встроена в вектор ДНК, способный самореплицироваться, и введена в хозяина, чтобы хозяин удерживал ее в виде плазмиды. Также допустимо, чтобы последовательность ДНК данного изобретения была интегрирована в хромосому микроорганизма при помощи способа с применением транспозона (Berg, D. E. and Berg, C.M., Bio/Technol, 1, 417 (1983)) Mu-- фага (Japanese Patent Laid-open N 2-109985) или гомологичной рекомбинации (Experiments in Molecular Genetics, Cold Spring Harbor Lab. (1972)). Кроме того, для интеграции ДНК данного изобретения в коринеформные бактерии можно использовать чувствительную к температуре плазмиду, описанную в Japanese Patent Laid-open N 5-7491.
При культивировании микроорганизма, трансформированного последовательностью ДНК данного изобретения, описанной выше, и экспрессии этой последовательности ДНК получают мутантный фермент. В результате измерения активности путем добавления аспарагиновой кислоты к реакционной системе фермента становится ясно, имеет ли полученный таким образом мутантный фермент сниженную чувствительность к ингибированию по типу обратной связи аспарагиновой кислотой. Для измерения активности фермента можно использовать спектрометрический способ (Yoshinage, T. , Izui, K. and Katsuki, H., J. Biochem., 68, 747-750 (1970)) и т.п.
Далее последовательность ДНК данного изобретения кодирует мутантный фермент, в котором десенсибилизировано ингибирование по типу обратной связи аспарагиновой кислотой, так что микроорганизм, несущий эту последовательность ДНК, можно использовать для эффективного ферментативного продуцирования аминокислот семейства аспарагиновой кислоты и семейства глутаминовой кислоты, как описано ниже.
Escherichia coli AJ12907, AJ12908, AJ12909 и AJ12910, несущие гены мутантного фермента, полученные в примерах, описанных ниже, депонированы в National Institute of Bioscience and Human Technology of Agancy of Industrial Science and Technology (1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan; zip code 305) 3 августа 1993 г. под депозитными номерами FERM P-13774, FERM P-13775, FERM P-13776 и FERM P-13777, перенесены из первоначальных депозитариев в международный депозитарий на основе Budapest Treaty 11 июля 1994 г. и депонированы под депозитными номерами FERM BP-4734, FERM BP-4735, FERM BP-4736, FERM BP-4737 соответственно в этой последовательности.
<3> Способ получения аминокислот.
Аминокислоты могут быть получены культивированием микроорганизма, несущего в себе последовательность ДНК данного изобретения, в предпочтительной среде и выделением образовавшихся аминокислот. Такими аминокислотами могут быть, например, L-лизин, L-треонин, L-метионин, L-изолейцин, L-глутаминовая кислота, L-аргинин и L-пролин.
Примеры предпочтительных хозяев, в которые вводят последовательность ДНК данного изобретения для использования для получения каждой из этих аминокислот, и способа культивирования приводятся ниже.
(1) Хозяева, предпочтительные для получения L-лизина.
Примером хозяина, который может быть использован для получения L-лизина согласно данному изобретению, могут быть бактерии, принадлежащие к роду Escherichia, предпочтительно продуцирующая L-лизин Escherichia coli. Конкретно может быть использован мутантный штамм, устойчивый к аналогу лизина. Такими аналогами лизина являются аналоги, которые ингибируют рост микроорганизмов, принадлежащих к роду Escherichia, однако эта супрессия полностью или частично десенсибилизируется при условии, что L-лизин также присутствует в среде. Например, имеются такие аналоги, как оксализин, лизингидроксамат-S-(2-аминоэтил)-цистеин (далее сокращаемый как "AEC"), γ-метиллизин, α-xлоркапролактам и др. Мутантные штаммы, обладающие устойчивостью к этим аналогам лизина, могут быть получены путем применения обычной искусственной мутационной обработки к микроорганизмам рода Escherichia. Конкретно в качестве бактериального штамма, который можно применять для получения L-лизина, можно привести Escherichia coli AJ11442 (депонированный как FERM P-5084, см. нижний левый столбец на стр. Japanese Patent Laid-open N 56-18596).
С другой стороны, для данного изобретения можно использовать различные искусственные мутантные штаммы коринеформных бактерий в качестве продуцирующих лизин бактерий. Такими искусственными мутантными штаммами являются следующие: AEC-резистентный мутантный штамм; мутантный штамм, требующий для его роста аминокислоты, такой как L-гомосерин (Japanese Patent Publication N 48-28078 и N 56-6499); мутантный штамм, проявляющий резистентность к AEC и требующий аминокислоты, такой как L-лейцин, L-гомосерин, L-пролин, L-серин, L-аргинин, L-аланин, L-валин и т.п. для его роста (US Patent N 3708395 и N 3825472); продуцирующий L-лизин мутантный штамм, проявляющий резистентность к DL α-амино ε-капролактаму, α-амино-лауриллактаму, хиноиду и N-лауроиллейцину; продуцирующий L-лизин мутантный штамм, проявляющий резистентность к ингибитору оксалоацетатдекарбоксилазы или фермента дыхательной системы (Japanese Patent Laid-open N 50-53588, 52-102498, 53-86089, 55-9783, 55-9759, 56-32995, 56-39778 и Japanese Patent Publication N 53-43591 и 53-1833); продуцирующий L-лизин мутантный штамм, требующий инозита или уксусной кислоты (Japanese Patent Laid-open N 55-9784 и 56-8692); продуцирующий L-лизин мутантный штамм, проявляющий чувствительность к фторпирувату или температуре не менее 34oC (Japanese Patent Laid-open N 55-9783 и 53-86090); и мутантный штамм Brevibacterium или Corynebacterium, проявляющий резистентность к этиленгликолю и продуцирующий L-лизин (см. US Patent Application Serial N 333455).
Далее приведены конкретные примеры коринеформных бактерий, применимых для получения лизина:
Brevibacterium lactofermentum AJ12031 (FERM-BP277), см. стр. 525 в Japanese Patent Laid-open N 60-62994;
Brevibacterium lactofermentum ATCC 39134, см. нижнюю правую колонку на стр. 473 в Japanese Patent Laid-open N 60-62994;
Brevibacterium lactofermentum AJ3463 (FERM-P1987), см. Japanese Patent Publication N 51-34477.
Кроме того, дикие штаммы коринеформных бактерий, описанные ниже, также можно применять таким же образом для данного изобретения.
Corynebacterium acetoacidophilum ATCC 13870
Corynebacterium acetoglutamicum ATCC 15806
Corynebacterium callunae ATCC 14991
Corynebacterium glutamicum ATCC 13032
ATCC 13060
(Brevibacterium divaricatum) ATCC 14020
(Brevibacterium Lactofermentum) ATCC 13869
(Corynebacterium lilium) ATCC 15990
Corynebacterium melassecola ATCC 17965
Brevibacterium saccharolyticum ATCC 14066
Brevibacterium immariophilum ATCC 14068
Brevibacterium roseum ATCC 13825
Brevibacterium flavum ATCC 13826
Brevibacterium thiogenitalis ATCC 19240
Microbacterium ammoniaphilum ATCC 15354
(ii) Хозяева, предпочтительные для получения L-треонина.
Escherichia coli B-3996 (RIA 1867), см. Japanese Patent Laid-open N 3-501682 (PCT);
Escherichia coli AJ12349 (FERM-P9574), см. верхнюю левую колонку на стр. 887 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ12351 (FERM-P9576), см. нижнюю правую колонку на стр. 887 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ12352 (FERM P-9577), см. верхнюю левую колонку на стр. 888 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ11332 (FERM P-4898), см. верхнюю левую колонку на стр. 889 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ12350 (FERM P-9575), см. верхнюю левую колонку на стр. 889 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ12353 (FERM P-9578), см. верхнюю правую колонку на стр. 889 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ12358 (FERM P- 9764), см. верхнюю левую колонку на стр. 890 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ12359 (FERM P-9765), см. верхнюю левую колонку на стр. 890 в Japanese Patent Laid-open N 2-458;
Escherichia coli AJ11334 (FERM P-4900), см. колонку 6 на стр. 201 в Japanese Patent Publication N 1-29559;
Escherichia coli AJ11333 (FERM P-4899), см. колонку 6 на стр. 201 в Japanese Patent Publication N 1-29559;
Escherichia coli AJ11335 (FERM P-4901), см. колонку 7 на стр. 202 в Japanese Patent Publication N 1-29559.
Следующие бактериальные штаммы приведены в качестве примеров коринеформных бактерий:
Brevibacterium lactofermentum AJ11188 (FERM P-4190), см. верхнюю правую колонку на стр. 473 в Japanese Patent Laid-open N 60-87788;
Corynebacterium glutamicum AJ11682 (FERM BP418), см. колонку 8 на стр. 230 в Japanese Patent Publication N 2-31956;
Brevibacterium flavum AJ11683 (FERM BP-119), см. колонку 10 на стр. 231 в Japanese Patent Publication N 2-31956.
(iii) Хозяева, предпочтительные для получения L-метионина.
Следующие бактериальные штаммы приведены в качестве примеров бактерий для получения L-метионина:
Escherichia coli AJ11457 (FERM P-5175), см. верхнюю правую колонку на стр. 552 в Japanese Patent Laid-open N 56-35992;
Escherichia coli AJ11458 (FERM P-5176), см. верхнюю правую колонку на стр. 552 в Japanese Patent Laid-open N 56-35992;
Escherichia coli AJ11459 (FERM P-5177), см. верхнюю правую колонку на стр. 552 в Japanese Patent Laid-open N 56-35992;
Escherichia coli AJ11539 (FERM P-5479), см. нижнюю левую колонку на стр. 435 в Japanese Patent Laid-open N 56-144092;
Escherichia coli AJ11540 (FERM P-5480), см. нижнюю левую колонку на стр. 435 в Japanese Patent Laid-open N 56-144092;
Escherichia coli AJ11541 (FERM P25481), см. нижнюю левую колонку на стр. 435 в Japanese Patent Laid-open N 56-144092;
Escherichia coli AJ11542 (FERM P-5482), см. нижнюю левую колонку на стр. 435 в Japanese Patent Laid-open N 56-144092.
(iv) Хозяева, предпочтительные для получения L-аспарагиновой кислоты.
Следующие бактериальные штаммы приведены в качестве примеров бактерий для получения L-аспарагиновой кислоты:
Brevibacterium flavum AJ3859 (FERM P-2799), см. верхнюю левую колонку на стр. 524 Japanese Patent Laid-open N 51-61689;
Brevibacterium lactofermentum AJ3860 (FERM P-2800), см. верхнюю левую колонку на стр. 524 в Japanese Patent Laid-open N 51-61689;
Corynebacterium acetoacidophilum AJ3877 (FERM P-2803), см. верхнюю левую колонку на стр. 524 в Japanese Patent Laid-open N 51-61689;
Corynebacterium glutamicum AJ3876 (FERM P-2802), см. верхнюю левую колонку на стр. 524 в Japanese Patent Laid-open N 51-61689;
(v) Хозяева, предпочтительные для получения L-изолейцина.
Escherichia coli КХ141 (VKPM-B4781) (см. параграф 45 в Japanese Patent Laid-open N 4-33027) приведена в качестве бактерии, принадлежащей к роду Escherichia, и Brevibacterium lactofermentum AJ12404 (FERM P-10141) (см. нижнюю левую колонку на стр. 603 в Japanese Patent Laid-open N 2-42988) и Brevibacterium flavum AJ12405 (FERM P-10142) (см. нижнюю левую колонку на стр. 524 в Japanese Patent Laid-open N 2-42988) приведены в качестве примеров коринеформных бактерий.
(vi) Хозяева, предпочтительные для получения L-глутaминoвoй кислоты.
Следующие бактериальные штаммы приведены в качестве бактерий, принадлежащих к роду Escherichia:
Escherichia coli AJ12628 (FERM P-12389), см. Franch Patent Publication N 2 680 (1993);
Escherichia coli AJ12624 (FERM P-12379), см. French Patent Publication N 2 680 178 (1993).
Следующие бактериальные штаммы приведены в качестве примеров коринеформных бактерий:
Brevibacterium lactofermentum AJ12745 (FERM BP-2922), см. нижнюю правую колонку на стр. 561 в Japanese Patent Laid-open N 3-49690;
Brevibacterium lactofermentum AJ12746 (FERM BP-2923), см. верхнюю левую колонку на стр. 562 в Japanese Patent Laid-open N 3-49690;
Brevibacterium lactofermentum AJ12747 (FERM BP-2924), см. верхнюю левую колонку на стр. 562 в Japanese Patent Laid-open N 3-49690;
Brevibacterium lactofermentum AJ12748 (FERM BP-2925), см. верхнюю левую колонку на стр. 562 в Japanese Patent Laid-open N 3-49690;
Brevibacterium flavum ATCC 14067, см. таблицу 1 на стр. 3 в Japanese Patent Laid-open N 5-3793;
Corynebacterium glutamicum ATCC 21492, см. таблицу 1 на стр. 3 в Japanese Patent Laid-open N 5-3793.
(vii) Хозяева, предпочтительные для получения L-аргинина.
Следующие бактериальные штаммы приведены в качестве бактерий, принадлежащих к роду Escherichia:
Escherichia coli AJ11593 (FERM P-5616), см. верхнюю левую колонку на стр. 468 в Japanese Patent Laid-open N 57-5693;
Escherichia coli AJ11594 (FERM P-5617), см. верхнюю правую колонку на стр. 468 в Japanese Patent Laid-open N 57-5693.
Следующие бактериальные штаммы приведены в качестве примеров коринеформных бактерий:
Brevibacterium flavum AJ12144 (FERM P-7642), см. колонку 4 на стр. 174 в Japanese Patent Publication N 5-27388;
Corynebacterium glutamicum AJ12145 (FERM P-7643), см. колонку 4 на ст. 174 в Japanese Patent Publication N 5-27388;
Brevibacterium flavum ATCC 21493, см. таблицу 1 на стр. 3 в Japanese Patent Laid-open M 5-3793;
Corynebacterium glutamicum ATCC 21659, см. таблицу 1 на стр. 3 в Japanese Patent Laid-open N 5-3793.
(viii) Хозяева, предпочтительные для получения L-пролина.
Следующие бактериальные штаммы приведены в качестве бактерий, принадлежащих к роду Escherichia:
Escherichia coli AJ11543 (FERM P-5483), см. нижнюю левую колонку на стр. 435 в Japanese Patent Laid-open N 56-144093;
Escherichia coli AJ11544 (FERM P-5484), см. нижнюю левую колонку на стр. 435 в Japanese Patent Laid-open N 56-144093.
Следующие бактериальные штаммы приведены в качестве примеров коринеформных бактерий:
Brevibacterium lactofermentum AJ11225 (FERM P-4370), см. верхнюю левую колонку на стр. 473 в Japanese Patent Laid-open N 60-87788;
Brevibacterium flavum AJ11512 (FERM P-5332), см. колонку 2 на стр. 185 в Japanese Patent Publication N 62-36679;
Brevibacterium flavum AJ11513 (FERM P-5333), см. колонку 2 на стр. 185 в Japanese Patent Publication N 62-36679;
Brevibacterium flavum AJ11514 (FERM P-5334), см. колонку 2 на стр. 185 в Japanese Patent Publication N 62-36679;
Corynebacterium glutamicum AJ11522 (FERM P-5342), см. колонку 2 на стр. 185 в Japanese Patent N 62-36679;
Corynebacterium glutamicum AJ11523 (FERM P-5343), см. колонку 2 на стр. 185 в Japanese Patent Publication N 62-36679.
(2) Способ культивирования.
Способ культивирования вышеуказанных хозяев не особенно отличается от способа культивирования продуцирующих аминокислоты микроорганизмов в предшествующих описаниях. Конкретно используют стандартную среду, содержащую источник углерода, источник азота и неорганические ионы и иногда органические питательные элементы в микроколичествах, такие как аминокислоты, витамины и т.п.
В качестве источника углерода можно применять глюкозу, сахарозу, лактозу и т. п. , а также гидролизат крахмала, молочную сыворотку, мелассы и другие содержащие эти компоненты продукты. В качестве источника азота можно использовать газообразный аммиак, водный аммоний, соль аммония и т.п. В некоторых случаях при применении в качестве хозяина мутантного штамма, имеющего потребность в аминокислотах или др. веществах, следует добавлять такой питательный элемент, как аминокислоты или другие необходимые вещества, к среде. Пример среды для получения лизина приведен в табл. 1 в качестве среды, которая может применяться для получения аминокислот. В некоторых случаях к другим компонентам добавляют карбонат кальция после отдельной стерилизации.
Культивирование проводят до тех пор, пока генерирование и накопление аминокислот по существу не останавливается при контролировании pH и температуры среды при аэробных условиях. Для сбора накопленных таким образом аминокислот в среде можно использовать обычный способ.
Краткое описание чертежей.
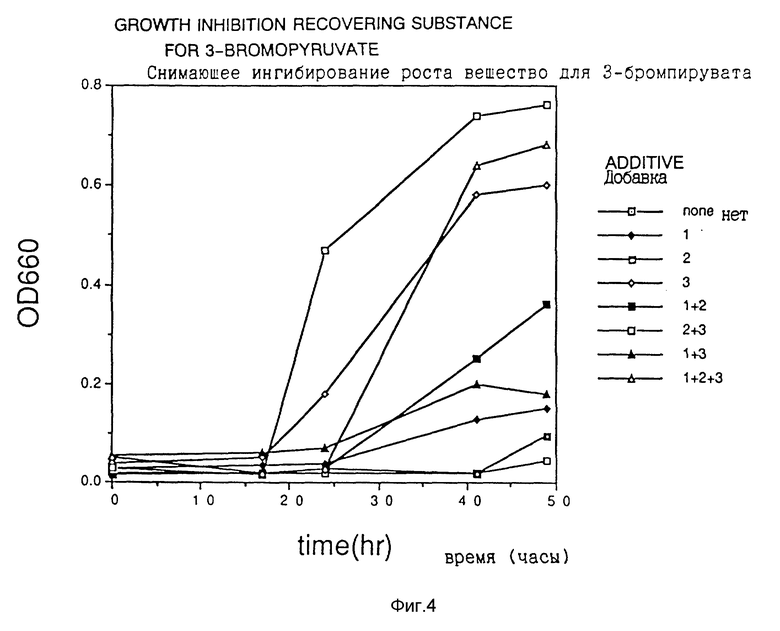
Фиг. 1 показывает ингибирование роста 3-бромпируватом.
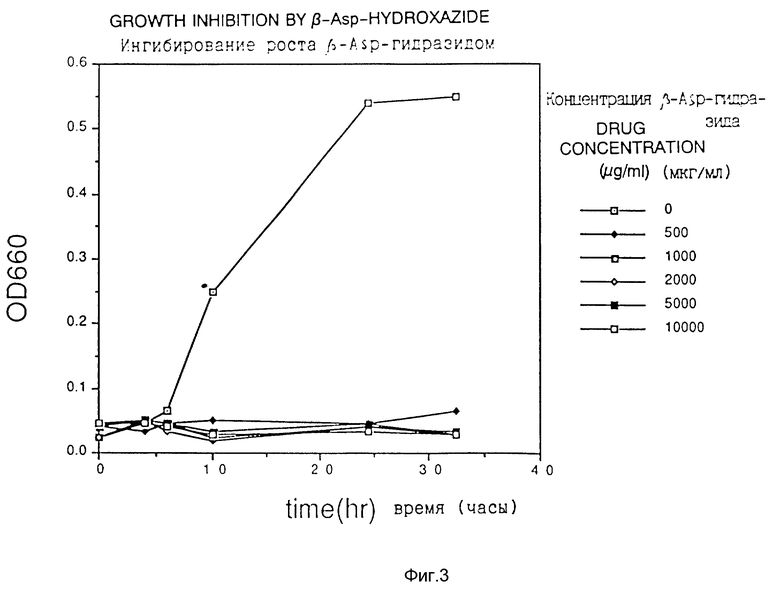
Фиг. 2 показывает ингибирование роста аспартат-β-гидразидом.
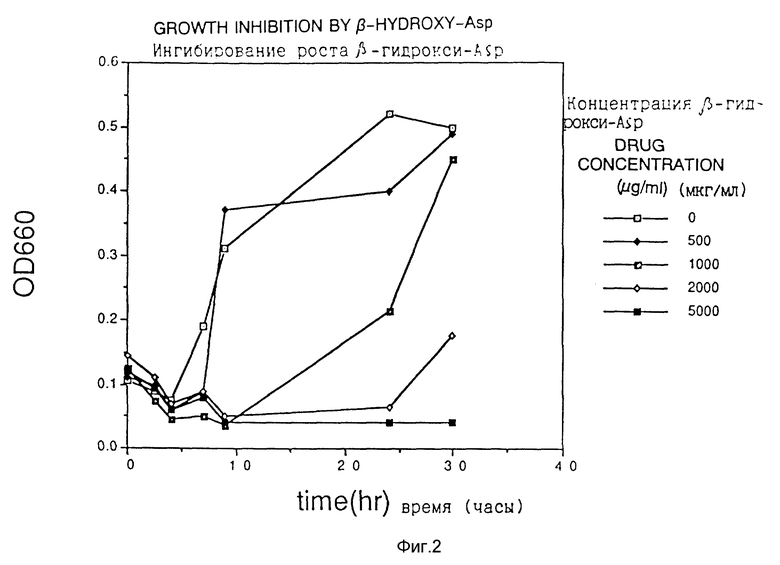
Фиг. 3 показывает ингибирование роста DL-трео-β -гидроксиаспартатом.
Фиг. 4 показывает действие снимающих ингибирование веществ на 3-бромпируват.
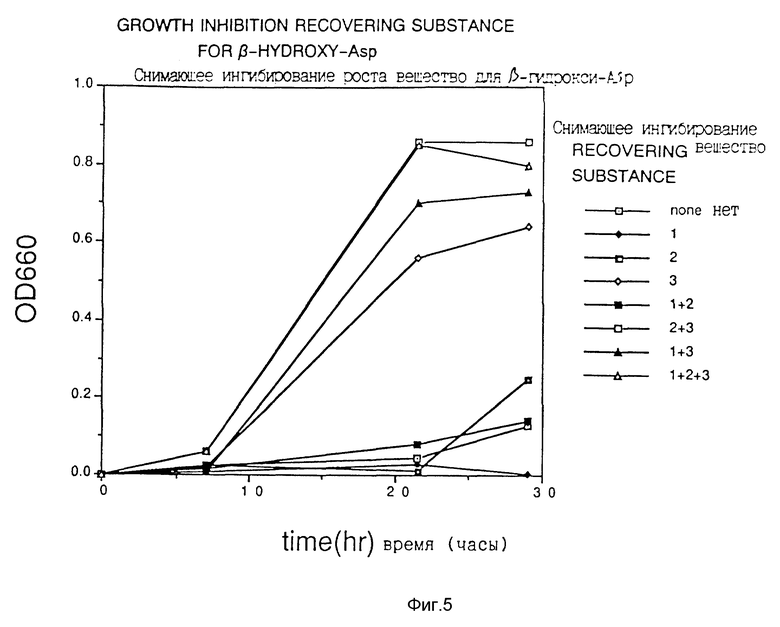
Фиг. 5 показывает действие снимающих ингибирование веществ на аспартат-β-гидразид.
Фиг. 6 показывает действие снимающих ингибирование веществ на DL-трео-β-гидроксиаспартат.
Фиг. 7 показывает действие на рост снимающих ингибирование факторов.
Фиг. 8 показывает ингибирование фосфоенолпируваткарбоксилазы ингибирующими рост веществами.
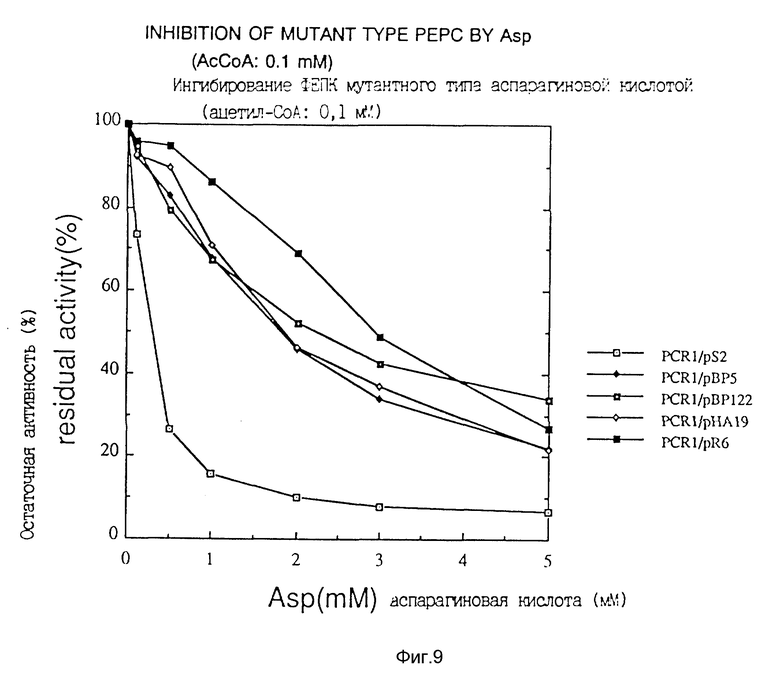
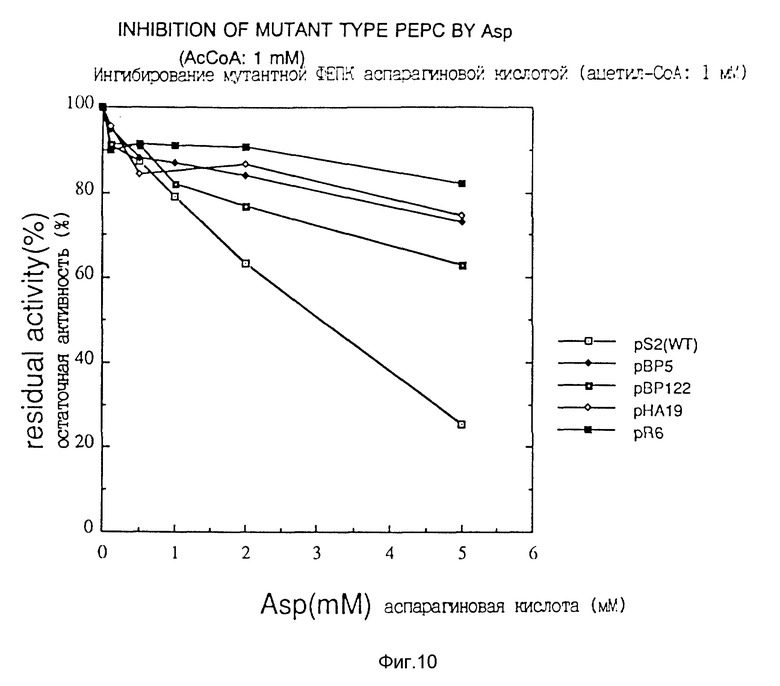
Фиг. 9 показывает ингибирование фосфоенолкарбоксилазы данного изобретения аспарaгиновой кислотой.
Фиг. 10 показывает ингибирование фосфоенолпируваткарбоксилазы данного изобретения аспарагиновой кислотой.
Фиг. 11 показывает способ введения мутации в ген фосфоенолпируваткарбоксилазы.
Фиг. 12 показывает влияние, оказываемое аспарагиновой кислотой, на активность фосфоенолкарбоксилазы дикого типа и мутантной фосфоенолпируваткарбоксилазы, в которой аргинин 438 заменен цистеином (нумерация от N-конца).
Фиг. 13 показывает влияние, оказываемое аспарагиновой кислотой на активности фосфоенолпируваткарбоксилазы дикого типа и мутантной фосфоенолпируваткарбоксилазы, в которой лизин 620 заменен серином (нумерация с N-конца).
Наилучший способ проведения изобретения
Данное изобретение будет объяснено более подробно ниже на примерах.
Пример 1: получение мутантного гена фосфоенолпируваткарбоксилазы.
Мутантный ген получали с применением плазмиды pS2, полученной встряхиванием клонированного и секвенированного гена фосфоенолпируваткарбоксилазы в Sall сайт векторной плазмиды pBR322. pS2 имеет ген устойчивости к ампициллину в качестве маркерного гена устойчивости к лекарственному средству (Sabe, H. et al., Gene, 31, 279-283 (1984)). Нуклеотидная последовательность гена фосфоенолпируваткарбоксилазы, содержащаяся в pS2, такая же, которая содержится в упомянутой выше плазмидe pT2.
pS2 ДНК обрабатывали при 75oC в течение 2 ч раствором гидроксиламина, содержащим 20 мкг/мл pS2 ДНК, 0,05 М фосфат натрия (pH 6,0), 1 мМ ЭДТА, 0,4 М гидроксиламин. Вследствие влияния pH на обработку гидроксиламином смешивали 80 мкл раствора 1М гидроксиламина HCl и 1 мМ ЭДТА, имеющего pH, доведенный до 6,0 гидроксидом натрия, 100 мкл раствора 0,1 М фосфата натрия (pH 6,0) и 1 мМ ЭДТА, и ТЕ-буфер (10 мМ Трис-HCl, 1 мМ ЭДТА), содержащий 2 мкг pS2 ДНК, доводя конечный объем до 200 мкл водой.
В указанных условиях трансформанты имеют пропорцию выживших организмов 0,2% в расчете на состояние перед обработкой в содержащей ампициллин среде при трансформации плазмидой pS2 Escherichia coli HB101 после этой обработки.
Клетки Escherichia coli HB101, трансформированные плазмидой pS2, обработанной гидроксиламином, наносили на твердую среду для чашек Петри, получая приблизительно 10000 колоний трансформантов. Их суспендировали в жидкой среде и высеивали на твердую среду для чашек Петри, содержащую один из аналогов аспарагиновой кислоты (3-бромпируват (3BP), аспартат- β-гидроксамат (АНХ), аспартат-β-гидразид (АНУ) и DL-трео-β-гидроксиаспартат (βHA) при концентрации, близкой к минимальной ингибирующей концентрации, получая 103 - 105 клеток на одну чашку со средой, и отбирали растущие колонии.
Из 100 полученных таким образом устойчивых к соединению-аналогу штаммов продуцируемую каждым из них фосфоенолпируваткарбоксилазу частично очищали в соответствии со способом, описанным в Journal of Biochem., vol. 67, N 4 (1970), и исследовали ингибирование активности фермента соединениями-аналогами. Измерение активности фермента проводили как описано выше.
Далее плазмиды выделяли из бактериальных штаммов, продуцирующих мутантные ферменты с активностями, не ингибируемыми соединениями-аналогами, и эти плазмиды вводили в Escherichia coli PCR1, являющийся дефектным в отношении фосфоенолпируваткарбоксилазы штаммом (Sabe, H. et al., Gene 31, 279 283 (1984)), для подтверждения образования мутантных ферментов.
Таким образом, получили 5 трансформантов, несущих мутантные гены фермента. Определение последовательностей оснований этих генов показало, что 2 штамма имели одну и ту же мутацию, и были получены 4 типа мутантных генов. Трансформанты, несущие эти гены, были названы AJ12907, AJ12908, AJ12909 и AJ12910 и были депонированы в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology (1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan; zip code 305) 3 августа 1993 г. под депозитными номерами FERM P-137, FERM P-13775, FERM P-13776 и FERM P-13777, перенесены из первоначального депозитария в международный депозитарий (хранилище) на основе Budapest Treaty 11 июля 1994 г. и были депонированы в виде депозитных номеров FERM BP-4734, FERM BP-4735, FERM BP-4736 и FERM BP-4737 соответственно в этой последовательности. Далее плазмиды, имеющие эти гены, были обозначены как pBP5, pHA19, pBP122 и pR6 соответственно. Мутации генов фосфоенолпируваткарбоксилазы, содержащиеся в каждой из этих плазмид, показаны в табл. 2. Цифровые величины в таблице указывают номера нуклеотидов или номера аминокислот в SEQ ID N 1.
Обычно отбор для AJ12907 и AJ12909 выполняли в среде, содержащей 500 мкг/мл 3BP, для AJ12908 в среде, содержащей 1000 мкг/мл βHA, и для AJ12910 в среде, содержащей 500 мкг/мл AHУ.
Пример 2: мутантная фосфоенолпируваткарбоксилаза.
Для фосфоенолпируваткарбоксилаз, продуцируемых вышеупомягутыми 4 трансформантами, исследовали чувствительность к аспарагиновой кислоте. Эти бактериальные штаммы дефектны по гену фосфоенолпируваткарбоксилазы, происходящему из хозяина, так что фосфоенолпируваткиназа этих штаммов является фосфоенолпируваткиназой плазмиды.
Чувствительность к аспарагиновой кислоте исследовали в соответствии с известным способом (Yoshinaga, T., Izui, K. and Katsuki, H., J. Biochem., 68, 747-750 (1970)), a именно посредством измерения активности фермента, продуцируемого каждым из трансформаторов или Escherichia coli, несущей pS2, в присутствии ацетил-кофермента A, влияющего на активность в системе измерения активности фермента, при концентрации 0.1 или 1 мМ, исследовали чувствительность к аспарагиновой кислоте, и эти данные представлены в фиг. 9 и фиг. 10.
Согласно этим данным видно, что фермент дикого типа теряет свою активность в присутствии высокой концентрации аспарагиновой кислоты, тогда как мутантная фосфоенолпируваткарбоксилаза данного изобретения продолжает по существу сохранять свою активность.
Пример 3: ферментативное продуцирование L-треонина Escherichia coli с введенной мутантной фосфоенолпируваткарбоксилазой.
Как продуцирующий треонин штамм Escherichia coli B-3996 (Japanese Patent Laid-open N 3-501682 (PCT)) имеет наивысшую производительность среди известных в настоящее время продуцирующих треонин бактерий. Таким образом, при оценке мутантной фосфоенолпируваткарбоксилазы B-3996 использовали в качестве хозяина. Этот штамм B-3996 был депонирован в Research Institute for Genetics and Industrial Microorganism Breeding под регистрационным номером R1A 1867. Кроме того, для оценки мутантной фосфоенолпируваткарбоксилазы был выбран плазмидный вектор pBH5, который был подвергнут эксперименту.
Плазмиду pBP5, имеющую мутантную фосфоенолпируваткарбоксилазу (ген), вводили в Escherichia coli B-3996 согласно способу Hanahan (J.Mol. Biol., vol. 106, 577 (1983)) и выделяли трансформант. В качестве контроля Escherichia coli B-3996 трансформировали таким же способом при помощи pS2 в качестве плазмиды, экспрессирующей ген фосфоенолпируваткарбоксилазы дикого типа.
При инокулировании Escherichia coli B-3996 и полученных из нее трансформантов соответственно в колбы Sakaguchi на 500 мл, залитые 20 мл среды, имеющей состав, приведенный в табл. 3, и культивировании при 37oC в течение 40 ч для исследования продуцирования L-треонина, были получены результаты, представленные в табл. 4. Обычно вышеупомянутую среду готовили в виде двух частей: сначала глюкозу и MgSO4•7H2O и другие компоненты смешивали и доводили pH до 7,0 при помощи KOH с последующим автоклавированием при 115oC в течение 10 мин, а затем после их смешивания отдельно стерилизовали CaCO3 и добавляли до 30 г/л.
Как выяснилось из этих данных, Escherichia coli B-3996/pBP5, несущая экспрессирующую мутантный фермент плазмиду, имеющую последовательность ДНК данного изобретения, имела улучшенную способность продуцирования треонина по сравнению с Escherichia coli B-3996/pS2, несущей плазмиду, экспрессирующую фермент дикого типа.
Пример 4: ферментативное продуцирование L-глутаминовой кислоты Escherichia coli с введенной мутантной фосфоенолпируваткарбоксилазой
Как продуцирующая глутаминовую кислоту бактерия штамм Escherichia coli AJ-12628, описанный в Japanese Patent Laid-open N 4-11461, имеет наивысшую продукционную способность среди известных в настоящее время бактерий. Поэтому при оценке мутантной фосфоенолпируваткарбоксилазы AJ-12628 использовали в качестве хозяина.
Штамм AJ-12628 был депонирован в National Institute of Bioscience and Human Tachnology of Agency of Industrial Science and Technology под регистрационным номером FERM BP-385. Далее была выбрана плазмида pBP5 в качестве оцениваемой мутантной фосфоенолпируваткарбоксилазы, которую подвергали эксперименту.
Плазмиду pВP5, имеющую мутантную фосфоенолпируваткарбоксилазу, вводили в Escherichia coli AJ-12628 согласно способу Hanahan (J. Mol. Biol., vol. 106, 577 (1983)) и выделяли трансформант. Таким же способом был выделен трансформант Escherichia coli AJ-12628 с pS2.
При инокулировании Escherichia coli AJ-12628 и полученных из нее трансформантов соответственно в колбы Sakaguchi на 500 мл, залитые 20 мл среды, имеющей состав, приведенный в табл. 5, и культивировании при 37oC в течение 36 ч для исследования продуцирования L-глутаминовой кислоты были получены результаты, представленные в табл. 6. Вышеупомянутую среду готовили в виде двух частей: глюкозу и MgSO4•7H2O и другие компоненты смешивали, доводили pH до 7,0 при помощи KOH с последующим автоклавированием при 115oC в течение 10 мин, а затем после перемешивания добавляли отдельно простерилизованный CaCO3 до 30 г/л.
Как выяснилось из полученных данных, Escherichia coli AJ-126/pBP5, несущая экспрессирующую мутантный фермент плазмиду, имеющую последовательность ДНК данного изобретения, имела улучшенную способность продуцирования глутаминовой кислоты по сравнению с Escherichia coli AJ-12628/pS2, несущей плазмиду, экспрессирующую фермент дикого типа.
Пример 5: продуцирование L-лизина коринеформной бактерией с введенной мутантной. фосфоенолпируваткарбоксилазой
Для введения и экспрессии мутантного гена в коринеформную бактерию получали промотор, происходящий из бактерии, принадлежащей к роду Brevibacterium, и лигировали его с мутантным геном для получения плазмиды экспрессирующего типа. Далее эту конструкцию вводили в бактерию рода Brevibacterium для продуцирования L-лизина.
<1> Получение гена аспартокиназы (АК), происходящего из бактерии, принадлежащей к роду Brevibacterium.
Хромосомную ДНК получали согласно стандартному способу из Brevibacterium lactofermentum (Corynebacterium glutamicum) дикого типа (ATCC 13869). АК ген амплифицировали из хромосомной ДНК при помощи PCR (полимеразной цепной реакции: см. White, T.J. et al., Trends Genet., 5, 185 (1989)). Для ДНК праймеров, применяемых в амплификации, синтезировали 23-членный олигонуклеотид (SEQ ID N 3) и 21-членный олигонуклеотид (SEQ ID N 4) для амплификации района приблизительно из 1643 п.н., кодирующего АК ген, на основе последовательности, известной в Corynebacterium glutamicum (см. Molecular Microbiology (1991) 5 (5), 1197-1204, Mol. Gen. Genet. (1990) 224, 317-324).
Синтез ДНК проводили согласно стандартному фосфоамидитному способу (см. Tetrahedron Letters (1981), 22, 1859) с использованием синтезатора ДНК модели 380B, производимой Applied Biosystems Co. В реакции PCR применяли DNA Thermal Cycler PJ2000, производимый Takara Shuzo Co., Ltd., и амплификацию гена выполняли с применением Taq ДНК-полимеразы согласно инструкции изготовителя.
Амплифицированный фрагмент гена 1643 т.п.н. подтверждали электрофорезом в агарозном геле и затем этот фрагмент вырезали из геля и очищали стандартным способом, а затем расщепляли рестриктазами Nrul (производимой Takara Shuzo Co., Ltd.), и Eco RI (производимой Takara Shuzo Co., Ltd.). В качестве клонирующего вектора для этого фрагмента гена примеряли pHSG399 (см. Takeshita S. et al.,; Gene (1987), 61, 63-74). pHSG399 расщепляли рестриктазами Smal (производимой Takara Shuzo Co., Ltd.), и EcoR1 и лигировали с амплифицированным фрагментом АК гена.
Лигирование ДНК проводили известным способом с применением набора для лигирования ДНК (Takara Shuzo Co., Ltd.). Таким образом была изготовлена плазмида, в которой pHSG399 была лигирована с фрагментом АК гена, амплифицированным из хромосомы Bravibacterium. Плазмида, имеющая АК ген, происходящий из ATCC 13869 (штамма дикого типа), была названа p399AKУ.
<2> Определение последовательности оснований АК гена Bravibacterium lactofermentum.
После получения АК плазмиды, p399AKУ, определяли последовательность оснований АК гена. Определение последовательности оснований проводили по способу Sanger et al. (F. Sanger et al.: Proc. Natl. Acad. Sci. US, 74, 5463 (1977) и т.д.). Результаты показаны в SEQ ID N 5 и SEQ ID N 7. Эти фрагменты ДНК имеют две открытые рамки считывания, которые соответствуют α-cубъединице и β-субъединице АК соответственно. В SEQ ID N 5 и SEQ ID N 7 аминокислотные последовательности, соответствующие каждой из открытых рамок считывания, показаны вместе с нуклеотидными последовательностями. Далее только аминокислотные последовательности, соответствующие каждой из открытых рамок считывания, показаны в SEQ ID N 6 и SEQ ID N 8.
<3> Получение экспрессирующей плазмиды фосфоенолпируваткарбоксилазы.
Sall фрагменты приблизительно 4,4 т.п.н., содержащие гены фосфоенолпируваткарбоксилазы, экстрагировали из pS2 в качестве плазмиды, имеющей ген фосфоенолпируваткарбоксилазы дикого типа, и из pBP5 в качестве плазмиды, имеющей полученный мутантный ген фосфоенолпируваткарбоксилазы, и эти фрагменты встраивали в Sall сайт плазмидного вектора pHSG399, универсально применяемого для Escherichia coli. Изготовленные плазмиды были названы pHS2 для дикого типа и pHBP5 для мутанта.
Для получения pHS2 и pHPB5 в плазмиды, способные экспрессироваться в Brevibacterium, вводили промотор и затравку репликации (начало репликации), функционирующие в Brevibacterium. В качестве промотора из p399AKУ экстрагировали фрагмент гена, содержащий последовательность оснований от 1-го сайта Nru 1 до 297-го сайта ApaL1, которую считают промоторным районом клонированного АК гена, и этот фрагмент встраивали в Ava1 сайт, расположенный приблизительно на 60 п.н. впереди структурных генов pHS2 и pHВP5, чтобы направление транскрипции было правильным.
Затем в сайт, расположенный на векторе, вводили фрагмент гена, делающий возможной автономную репликацию этой плазмиды в Brevibacterium, а именно начало репликации этой плазмиды. Фрагмент гена, содержащий начало репликации этой плазмиды, экстрагировали из вектора pHC4 для Brevibacterium (см. 10 в Japanese Patent Laid-open N 5-7491; Escherichia coli AJ12039, несущая такую плазмиду, депонирована в National Institute of Bioscience and Human Technology of Agency Industrial Science and Technology под депозитным номером FERM P12215) и сайты рестриктаз на обоих концах были модифицированы в PSt1 сайты введением линкеров.
Этот фрагмент вводили в PSt1 сайт в векторной части плазмиды с добавленным промотором из Brevibacterium. Сконструированные экспрессирующие фосфоенолпируваткарбоксилазу плазмиды были названы pHS2B для происходящей из pS2 плазмиды с геном фоенолпируваткарбоксилазы дикого типа и pHBP5B для происходящей из pBP5 плазмиды с геном мутантной фосфоенолпируваткарбоксилазы соответственно.
<4> Получение L-лизина при помощи экспрессирующей фосфоенолпируваткарбоксилазу плазмиды.
Полученные плазмиды pHS2B и pHBP5B соответственно вводили в штамм AJ3463, являющийся продуцирующим L-лизин штаммом, Brevibacterium lactofermentum (см. Japanese Patent Publication N 51-24477). Для введения этого гена использовали метод трансформации с применением электрического импульса (см. Japanese Patent Laid-open N 2-207791). Штамм-хозяин и трансформанты культивировали с качанием в течение 72 ч при 31,5oC в среде для продуцирования лизина, имеющей состав, показанный в табл. 7. Вышеупомянутую среду готовили следующим образом: все компоненты, перечисленные в таблице, кроме CaCO3, добавляли в 1 л воды и доводили pH до 8,0 при помощи KOH с последующим автоклавированием при 115oC в течение 15 мин, а затем добавляли CaCO3, подвергнутый стерилизации нагреванием. Накопленные после культивирования количества L-лизина в среде показаны в табл. 8.
Как показали эти данные, Brevibacterium lactofermentum AJ3463, несущий экспрессирующую мутантный фермент плазмиду, имеющую последовательность ДНК данного изобретения, имел улучшенную способность продуцирования лизина по сравнению с Brevibacterium lactofermentum AJ3463/pHS2B, несущим плазмиду для экспрессии фермента дикого типа.
Пример 6: другой пример мутантной фосфоенолпируваткарбоксилазы данного изобретения и ее гена.
<1> Получение гена мутантной фосфоенолпируваткарбоксилазы.
При получении ДНК, кодирующей мутантную фосфоенолпируваткарбоксилазу, в качестве материала использованы ген фосфоенолпируваткарбоксилазы, клонированный в плазмиде pT2.
Хозяин, который должен нести плазмиду pT2, предпочтительно должен быть дефектным по гену фосфоенолпируваткарбоксилазы, чтобы можно было детектировать только активность фосфоенолпируваткарбоксилазы, происходящей из плазмиды. В качестве такого не содержащего ген данного фермента штамма применяли Escherichia coli F15 (Hfr, recAl, met, Δ(ppc-argECBH), Tn10). Escherichia coli AJ-12873, способная удерживать pT2 в F15 штамме, депонирована как FERM P-13752 в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology (1-3, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan; zip code 305) 15 июля 1993, перенесена из первоначального депозитария в международный депозитарий на основе Budapest Treaty 11 июля и депонирована как FERM PB-4732. Кроме того, полная последовательность оснований pT2 показана в SEQ ID N 1.
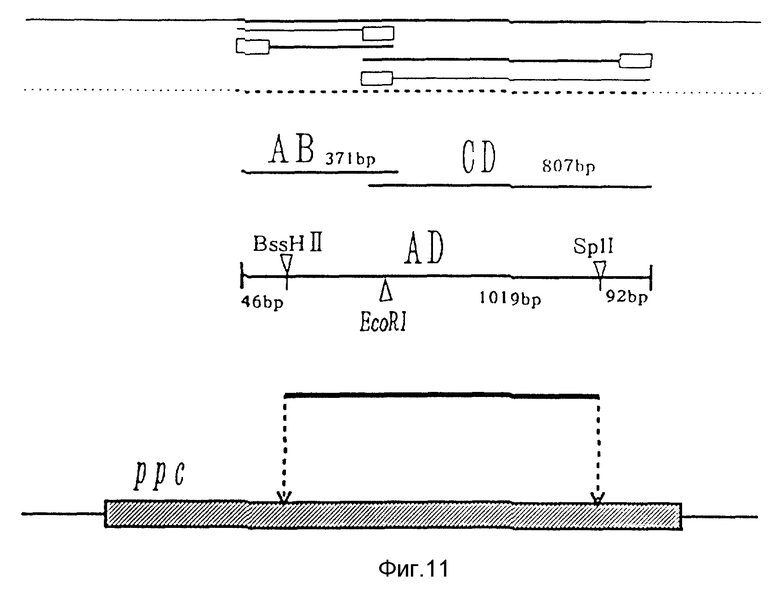
Для замены кодона аргинина 438 фосфоенолпируваткарбоксилазы на кодон цистеина с применением pT2 применяли способ перекрывающего удлинения (Ho, S.N. , Hunt, H. D. , Horton, R.M., Pullen, J.K. и Pease, L.R., Gene, 77, 51-59 (1989)) c применением способа PCR (полимеразной цепной реакции).
PCR способ представляет собой такой способ, в котором повторяется цикл амплификации, включающий в себя термическую денатурацию двухцепочечной ДНК, гибридизацию олигонуклеотидных праймеров, соответствующих последовательностям на обоих концах целевого амплифицируемого сайта и термически денатурированной ДНК, и полимеразную реакцию с использованием этих олигонуклеотидов в качестве праймеров. В результате последовательность ДНК амплифицируется экспоненциально.
Район, подвергнутый сайт-специфической мутации по PCR и способу, показан на фиг. 11. Праймерами, применяемыми в данном изобретении, были 4 вида: праймер с (SEQ ID N 11, соответствующий основаниям 1535-1554 в SEQ ID N 1), имеющий последовательность вблизи кодона аргинина 438, праймер в (SEQ ID N 10), имеющий последовательность, комплементарную праймеру с, праймер а (SEQ ID N 9, соответствующий основаниям 1185-1200 в SEQ ID N 1), имеющий последовательность слева от него, и праймер d (SEQ ID N 12, соответствующий основаниям 2317-2342 в SEQ ID N 1), имеющий последовательность, комплементарную последовательности, расположенной в направлении 5′_→ 3′.
В праймере b и праймере с кодон (CGT) аргинина 438 заменяли кодоном (TGT) цистеина. Такая замена может применять TGC, который является другим кодоном цистеина. Затем С кодона (AAC) аспарагина 435 заменяли Т, и, следовательно, EcoR1 сайт был введен внутрь без замены аминокислоты, так что мутантная плазмида могла бы отбираться с использованием этого сайта в качестве индикатора. Однако эта мутация не является существенной для данного изобретения.
Путем проведения PCR реакции с применением pT2 ДНК в качестве матрицы и праймера а и праймера b в качестве праймеров был амплифицирован фрагмент от положения слева от сайта мутации до сайта мутации (AB фрагмент на фиг. 11). Затем путем проведения PCR реакции с применением праймера с и праймера d был амплифицирован фрагмент справа от сайта мутации (CD фрагмент на фиг. 11). После повторной гибридизации каждого из амплифицированных продуктов (AB, CD) после термической денатурации для проведения полимеразной реакции их лигировали, получая фрагмент AD (AD в фиг. 11). Обычно PCR реакцию проводили повторением 30 циклов, каждый из которых предусматривал нагревание при 94oC 1 мин с последующей денатурацией (94oC, 1,5 мин), гибридизацией (50oC, 2 мин) и реакцией удлинения полимеразой (72oC, 3,5 мин). Кроме того, составы реакции показаны в табл. 9.
<2> Отбор не чувствительнoй к ингибированию фосфоенолпируваткарбоксилазы.
Escherichia coli трансформировали полученной, как описано выше, плазмидой, и трансформированный штамм культивировали для извлечения плазмиды и отбора плазмиды EcoR1. В отобранной ДНК последовательность оснований района, амплифицированного по описанному PCR способу, определяли дидезокси-секвенированием для подтверждения, что была введена замена оснований, которая была намечена. Эту плазмиду назвали pT2R438C. Штамм (AJ12874), полученный введением этой плазмиды в Escherichia coli F15, был депонирован как FERM P-1375 в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology (1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan; zip code 305) 15 июля 1993 г., перенесен из первоначального депозитария в международный депозитарий на основе Budapest Treaty 11 июля 1994 г. и был депонирован как FERM BP-4733.
Последовательность оснований pT2R438C является последовательностью, в которой нуклеотиды 1541 и 1550 заменены введением Т вместо С соответственно в SEQ ID N 1.
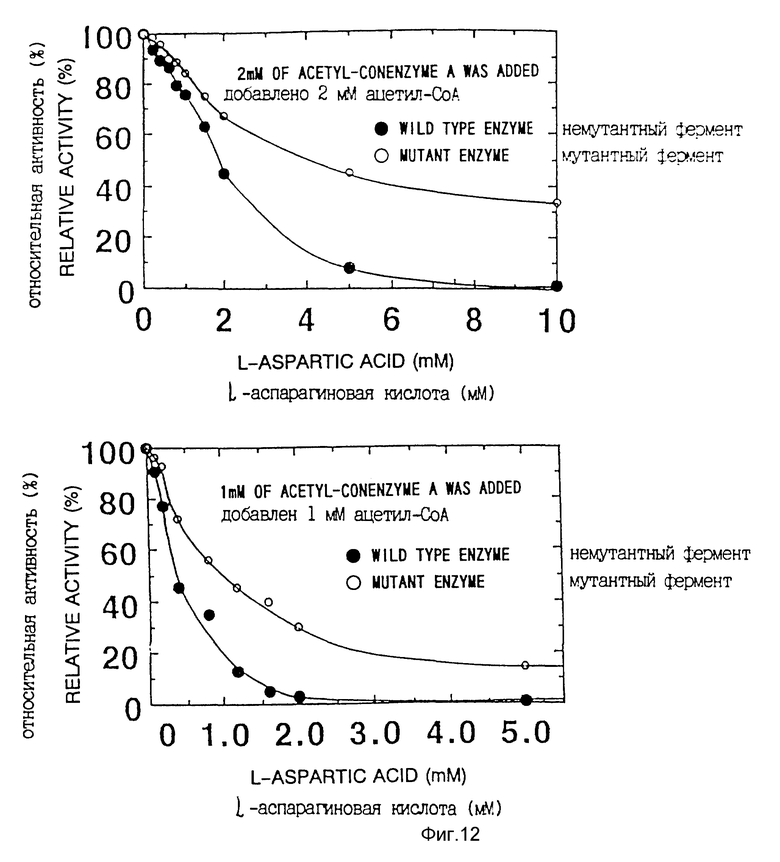
<3> Подтверждение снижения чувствительности (десенсибилизации) к ингибированию мутантной фосфоенолпируваткарбоксилазы аспарагиновой кислотой.
Чувствительность к аспарагиновой кислоте исследовали для фосфоенолпируваткарбоксилазы, продуцируемой Escherichia coli AJ 12874, несущей pT2 438C. Поскольку, как описано выше, Escherichia coli F15 недостаточна в отношении фосфоенолпируваткарбоксилазы, фосфоенолпируваткарбоксилаза, продуцируемая штаммом AJ12874, происходит из плазмиды.
Чувствительность к аспарагиновой кислоте исследовали в соответствии с известным способом (Yoshinaga, T., Izui, K., and Katsuki, H., J. Biochem., 68, 747-750 (1970)), a именно путем измерения активности фермента в присутствии ацетил-кофермента А, влияющего, как известно, на активность в системе для измерения активности, при концентрации 1 мМ или 2 мМ была измерена чувствительность к аспарагиновой кислоте, и полученные результаты представлены на фиг. 12.
Очевидно, что фермент дикого типа значительно снижает свою активность при высокой концентрации аспарагиновой кислоты, тогда как мутантная фосфоенолпируваткарбоксилаза данного изобретения продолжает сохранять свою активность.
<4> Получение гена (11) мутантной фосфоенолпируваткарбоксилазы.
Для замены кодона лизина 620 кодоном серина в гене фосфоенолпируваткарбоксилазы, содержащемся на плазмиде pT2, применяли способ перекрывающегося удлинения (Ho, S.N., Hunt, H.D., Horton, R.M., Pullen, J.K. and Pease, L. R. , Gene, 77, 51-99 (1989)) с использованием PCR способа (полимеразной цепной реакции). Конкретные процедуры аналогичны описанным в <1>. Плазмиду, несущую мутантный ген с целевой заменой, назвали pT2K620S. Полученный мутантный фермент был назван мутантным ферментом K620S.
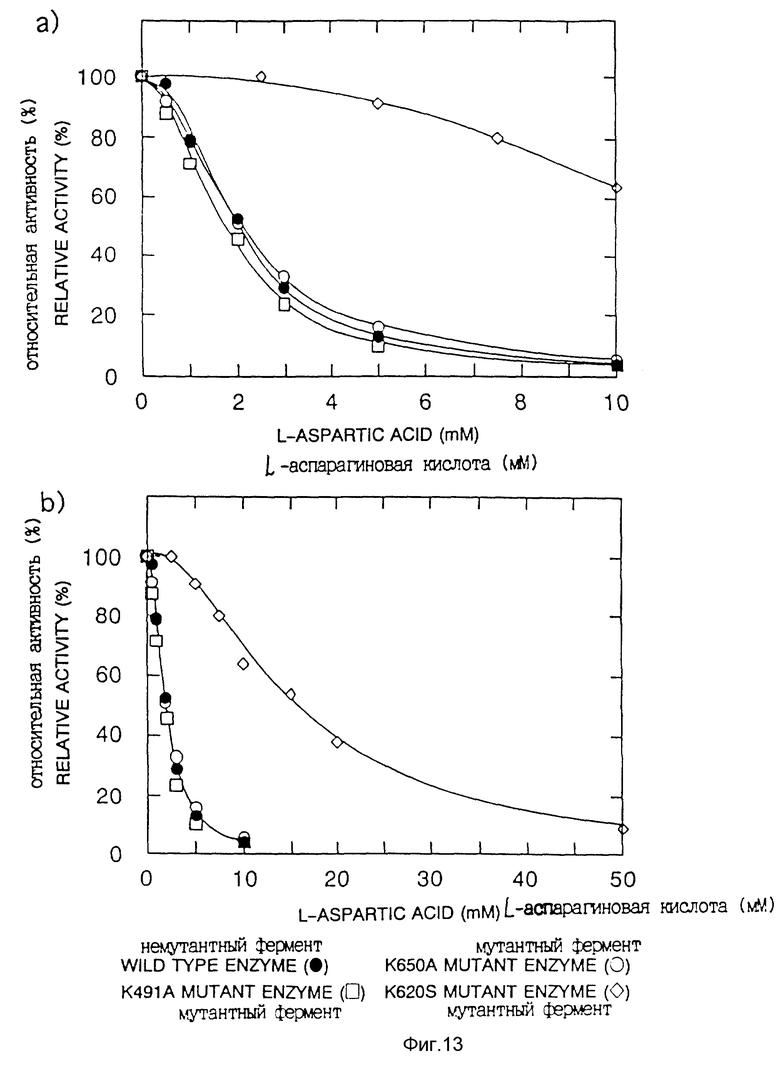
<5> Подтверждение снижения чувствительности к ингибированию аспарагиновой кислотой для мутантной фосфоенолпируваткарбоксилазы.
Для фосфоенолпируваткарбоксилазы, продуцируемой трансформантом, полученным введением плазмиды pT2K620S в указанный штамм Escherichia coli F15, исследовали чувствительность к аспарагиновой кислоте. Поскольку, как описано выше, Escherichia coli F15 не имеет фосфоенолпируваткарбоксилазы, любая фосфоенолпируваткарбоксилаза, продуцируемая трнасформантом, происходит из плазмиды.
Чувствительность к аспарагиновой кислоте исследовали в соответствии с известным способом (Yoshinaga, T., Izui, K. and Katsuki, H., J.Biochem. 68, 747-750 (1970)), a именно путем измерения активности фермента в присутствии ацетил-кофермента А, влияющего, как известно, на активность фермента в системе для измерения активности, при концентрации 1 мМ или 2 мМ измеряли чувствительность к аспарагиновой кислоте, и полученные результаты представлены на фиг. 13.
Очевидно, что фермент дикого типа значительно снижает свою активность при высокой концентрации аспарагиновой кислоты, тогда как фосфоенолпируваткарбоксилаза данного изобретения продолжает сохранять свою активность.
На фиг. 13 показана также чувствительность к аспарагиновой кислоте для мутантной фосфоенолпируваткарбоксилазы, в которой лизин 650 заменен серином (мутантный фермент K650A), и для мутантной фосфоенолпируваткарбоксилазы, в которой лизин 491 заменен серином (мутантный фермент K491A). В случае этих мутантных ферментов ингибирование аспарагиновой кислотой не снималось (не было десенсибилизации).
Промышленная применимость.
Последовательность ДНК данного изобретения кодирует мутантную фосфоенолпируваткарбоксилазу и микроорганизм, несущий эту последовательность ДНК, продуцирует вышеупомянутый фермент.
Мутантная фосфоенолпируваткарбоксилаза данного изобретения по существу не подвергается ингибированию ее активности аспарагиновой кислотой, так что ее можно использовать для ферментативного получения аминокислот, биосинтез которых регулируется аспарагиновой кислотой и т.п.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА (ВАРИАНТЫ) И РЕКОМБИНАНТНАЯ ДНК, ИСПОЛЬЗУЕМАЯ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ (ВАРИАНТЫ) | 1996 |
|
RU2197528C2 |
| ДНК, КОДИРУЮЩАЯ АСПАРТОКИНАЗУ III БАКТЕРИЙ, И СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1997 |
|
RU2204604C2 |
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ АСПАРТОКИНАЗУ III, СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 1993 |
|
RU2113484C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2194076C2 |
| ФЕРМЕНТАЦИОННЫЙ СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1994 |
|
RU2204605C2 |
| СПОСОБ ПОЛУЧЕНИЯ ОСНОВНОГО ВЕЩЕСТВА | 2005 |
|
RU2350655C2 |
| КОРИНЕФОРМНАЯ БАКТЕРИЯ, ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2004 |
|
RU2333247C2 |
| СПОСОБ ПОЛУЧЕНИЯ ОСНОВНОЙ АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2014 |
|
RU2628696C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2288271C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |




















Предложена фосфоенполпируваткарбоксилаза, у которой остаток глутаминовой кислоты в положении 625 замещен на остаток лизина и/или остаток аргинина в положении 222, и/или остаток глутаминовой кислоты в положении 223 на остаток гистидина и остаток лизина соответственно. Кроме того, сериновый остаток в положении 288, остаток глутаминовой кислоты в положении 289, остаток метионина в положении 551 и остаток глутаминовой кислоты в положении 804 могут быть заменены на остаток фенилаланина, остаток лизина, остаток изолейцина и остаток лизина соответственно. Также могут быть заменены остаток аланина в положении 867 на остаток треонина и/или остаток аргинина в положении 438 на остаток цистеина, и/или остаток лизина в положении 620 на остаток серина. Фосфоенолпируваткарбоксилаза продуцируется штаммом Escherichia coli. Модифицированная фосфоенолпируваткарбоксилаза малочувствительна к ингибированию аспарагиновой кислотой. 11 с. и 7 з.п. ф-лы, 9 табл., 13 ил.
| Agric | |||
| Biol | |||
| Chem., 1983, 47, N 7, p.1569-1576 | |||
| J | |||
| Biochem., 1984, v.95, N 4, p.909-916 | |||
| SU 1349250 А1, 30.10.87. |

