Исследование относится к микробиологическому производству, в нем описывается способ получения L-треонина с помощью ферментации. L-треонин относится к незаменимым аминокислотам и в этой связи используется как компонент различных питательных смесей, используемых для медицинских целей. Более того, он используется как пищевая добавка в корм животных и как фактор роста для процесса получения с помощью микроорганизмов таких аминокислот, как лизин и гомосерин.
Ранее в качестве микроорганизмов, используемых для получения L-треонина путем ферментации, использовались бактерии, выделенные из окружающей среды, или полученные искусственным путем мутанты таких бактерий. Известно множество искусственных L-треонин-продуцирующих мутантов, большинство из которых обладает устойчивостью к α-амин-β-оксивалериановой кислоте и принадлежат к родам Escherichia, Serratia, Brevibacterium или Corynebacterium. Что касается Escherichia, то существуют патенты JPA 55-131397, JPA 59-31691, JPA 56-15696 и JPW 3-501682, в которых описаны способы получения L-треонина с помощью бактерий, несущих рекомбинантную плазмиду, содержащую треониновые опероны.
К тому же, до этого был известен штамм треонин-продуцирующей бактерии Escherichia coli ВКПМ (V KPM) B-3996, характеризующийся наиболее высоким уровнем продукции треонина и коэффициентом потребления (JPW3-501682). В соответствии с настоящим изобретением, "коэффициент потребления" определяет количество граммов сахара, необходимое для продукции одного грамма треонина. Если культивирование бактерий производится в экспериментальном ферменте, то в ответ на сигналы от pH сенсора, в культуральную среду добавляется сахар-аммонийная смесь, при этом максимальный уровень биосинтеза треонина достигает 85 г/л, а коэффициент потребления составляет 2 г. сахара на один грамм треонина.
Аспартокиназа (ниже именуемая в сокращенном виде как AK) представляет собой фермент, который превращает аспарагиновую кислоту в β-фосфоаспарагиновую кислоту, и этот этап представляет собой единственный регулируемый участок на пути биосинтеза аспарагиновой кислоты и производных от нее аминокислот. Как показано на фиг. 1, существует три типа AK в E. coli (AKI, AK II, AK III), при этом первые два являются бифункциональными ферментами, имеющими гомосериндегидрогеназу (ниже именуемую как НД) активность. Один из них AK I - НД I кодируется thr A геном, а другой AK II-H II кодируется metL(M) геном.
Только AK III является монофункциональным ферментом, он представляет собой продукт гена lys C и известно, что он подвергается репрессии и ингибированию лизином по типу обратной связи. С другой стороны, AK I подвергается координированной репрессии треонином и изолейцином и ингибированию треонином, тогда как AK II подвергается репрессии метионином. В результате складывается следующая внутриклеточная пропорция активностей : AK I : AK II : AK III = приблизительно 5:1:4.
Ранее уже было проведено клонирование lys C гена E. coli и определена последовательность оснований (Cassan, M., Parsot, C., Cohen, G.N. и Patte J. C. , J.Biol., 261, 1052, 1986). Мицуками с сотр. описано также получение L-треонина путем ферментации с использованием штаммов E. coli, в которых активность AK III значительно увеличена (Mizukami, T. et al, Agric. Bjol. Chem., 50, 1015, 1986). Однако выделение достаточных количеств лизин-зависимого AK III в процессе ингибирования по типу обратной связи Мицуками с сотр. достигнуто не было.
Исходя из всего этого, предметом настоящего изобретения является достижение достаточно высокой степени освобождения AK III от ингибирования по типу обратной связи лизином и разработка способа получения L-треонина ферментативным путем с улучшенными показателями в сравнении с известным уровнем техники.
Разработчики настоящего изобретения в результате тщательных исследований пришли к удачному решению поставленной выше проблемы, получив ген (мутант lys C или lys C*), который кодирует AK III в E. coli и с помощью которого осуществляется снятие ингибирования этого фермента лизином по типу обратной связи и обнаружили, что при ведении этого гена в треонин - продуцирование бактерии, L-треонин достаточно эффективно накапливается, и на этом настоящее изобретение завершается.
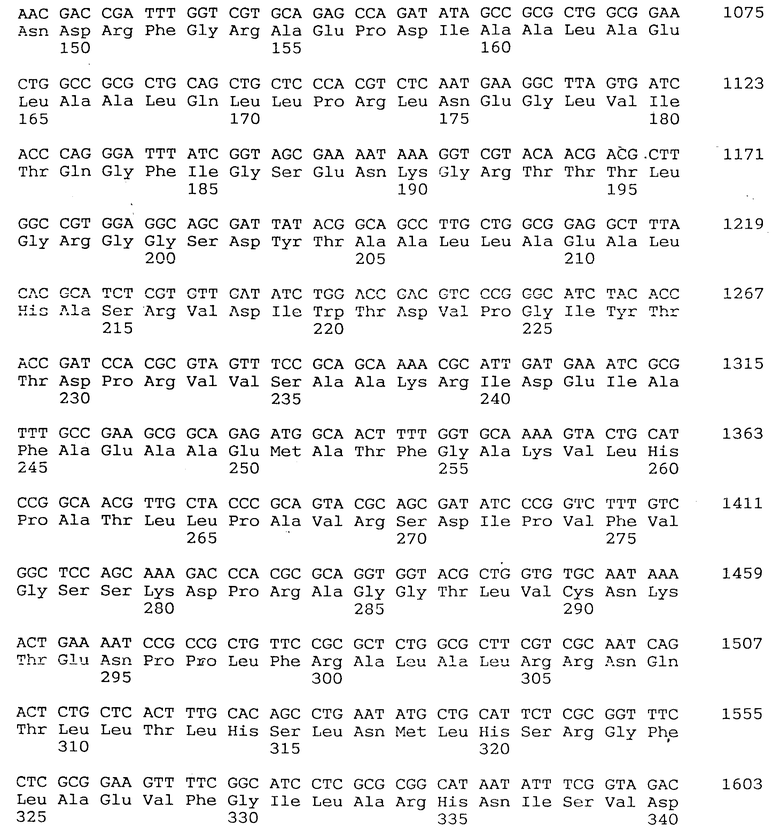
Иными словами, настоящее изобретение относится к ДНК, содержащей ген, который кодирует аспартокиназу III, найденную в бактериях рода Escherichia и которая имеет мутацию в кодирующем регионе, ответственную за снятие ингибирования лизином по типу обратной связи лизином вышеуказанной AK III, а более конкретно, изобретение относится к упомянутой выше ДНК, в которой найдено положение мутации, определяющей освобождение от ингибирования лизином по типу обратной связи AK III, а именно, в A или B домене аспартокиназы III, а точнее, относится к упомянутой ДНК, в которой мутация, определяющая рилизинг аспартокиназы III в процессе ингибирования лизином по типу обратной связи, отнесена к нижеуказанной последовательности: SEQ IDNO: I, отобранной из группы, содержащей различные мутации : мутации, в которой 323-й Гли заменен на Асп и 408-й Гли заменен Асп; мутации, в которой 34-й Арг заменен Цис и 323-й Гли заменен Асп; мутации, в которой 325-й Лей заменен Фен; мутации, в которой 318-й Мет заменен Иле; мутации, в которой 318-й Мет заменен Иле и 349-й Вал заменен Мет; мутации, в которой 345-й Сер заменен Лей; мутации, в которой 347-й Вал заменен Мет; мутации, в которой 352 Тре заменен Иле и 369-й Сер заменен Фен; мутации, в которой 164-й Глу заменен Лиз; мутации, в которой 417-й Мет заменен Иле и 419-й Цис заменен Три.
Кроме того, наносящее изобретение относится к упомянутой выше ДНК, которая является рекомбинантной ДНК, связанной с векторной ДНК, способной к автономной репликации в бактериях Escherichia. Оно относится также к микроорганизмам, принадлежащим к роду Escherichia и которые трансформированы введенной указанной выше рекомбинантной ДНК в клетки. Предметом настоящего изобретения являются также микроорганизмы, трансформированные введением в их хромосомальную ДНК, кодирующую аспартокиназу из упомянутой выше Escherichia и которая содержит ген с мутацией в регионе, кодирующем выделение указанной выше аспартокиназы III в процессе ингибирования лизином по типу обратной связи.
Изобретение также относится к способу получения L-треонина, характерными чертами которого является культивирование упомянутых выше микроорганизмов в сбраживаемой среде, продуцирование и аккумуляция L-треонина в культуре и выделение L-треонина из культуры.
Ниже дано детальное описание изобретения.
В качестве донора для получения ДНК, содержащей ген, кодирующей AK III (lys C) может быть использован любой микроорганизм, принадлежащий к роду Escherichia. А более конкретно, те микроорганизмы, которые отмечены в работах Нейдхардта (Neidhardt, F. C. etal. , Escherichia coli и Salmonella Typhimurium, American Society for Microbiology, Washington, D.C., 1208, табл. 1). Эти примеры включают E. coli, штаммы JM 109 и MC1061.
При использовании в качестве донора для получения ДНК, содержащей ген, кодирующий AK III (lys C) дикого штамма бактерий, будет получена ДНК, содержащая дикий тип гена AK III. Для введения мутации в этот ген с целью получения гена AK III, который определяет необходимое освобождение фермента от ингибирования по типу обратной связи L-лизином, можно провести прямую мутацию ДНК в процессе обработки in vitro гидроксиламином. Гидроксиламин представляет собой химический мутаген, который индуцирует мутации C ---> T типа за счет превращения цитозина в N4 - оксицитозин.
Кроме того, ДНК, содержащая ген AK III, который определяет освобождение фермента от ингибирования L-лизином по типу обратной связи, может быть получена из мутантного штамма бактерий, который характеризуется способностью выделять активный AK III в результате снятия ингибирования L-лизином по типу обратной связи. Такой мутант может быть получен из клеток, подвергнутых традиционному мутагенезу: облучению ультрафиолетовыми лучами или обработке таким мутагеном, как N-метил-N'-нитро-N-нитрозогуанидин (НТГ).
Ниже дано описание способа получения ДНК, содержащей ген, кодирующий AK III (lys C). Во-первых, E. coli имеющий дикий тип lesC, например, штамм MC1061, выращивается для получения культуральных клеток. Выращивание упомянутых выше микроорганизмов может производится традиционным методом культивирования на твердом субстрате, то с точки зрения выделения и сбора клеток предпочтительно использовать жидкие культуры. Культуральная среда может быть приготовлена путем добавления, например, одной или больше неорганических солей, таких как первичный кислый фосфат калия (KH2 + O3), вторичный кислый фосфат калия (K2HPO3), сульфат натрия, хлорид натрия, хлорид магния, хлорид железа, сульфат железа, или сульфат марганца, к одному или более источников азота, таких как дрожжевой экстракт, пептон, мясной экстракт, кукурузный экстракт или вытяжка из сои или пшеницы и затем добавляют углеводы, витамины и т.д., если это необходимо. Первоначально pH среды доводится до 7-8. Культивирование производится при температуре 30 - 42oC, предпочтительно при 37oC в течение 4-24 ч методом погруженной культуры при аэрации, перемешивании, в спокойной культуре и др. Продукт, получаемый при культивировании таким способом, подвергается с целью отделения от среды центрифугирования, в частности, при 3000 об/мин в течение 5 мин для получения клеток E.coli штамма MC 1061.
Хромоомальная ДНК может быть получена из этих же клеток по методу, например, Саито и Миура (Biochem. Biophys. Acta. 72, 619, 1963), методу К.С. Кирби (Biochem. J., 64, 405, 1955) и др.
С целью получения lys C гена, полученная описанным Выше методами хромосомальная ДНК расщепляется с использованием подходящего рестрикционного фермента. Если степень расщепления зависит от временной длительности процесса рестрикции и т.д., то может быть к тому же использовано множество рестриктаз. Затем ген может быть соединен с векторной ДНК, способной к репликации в бактериях рода Escherichia, а полученная рекомбинантная ДНК может быть трансформирована в мутантный штамм Escherichia, например, штамм GT3, который лишен аспартокиназы I, II, III (для создания библиотеки генов), при этом из полученных трансформантов может быть выделен штамм, способный к росту в минимальной культуральной среде без лизина и таким образом, выделяется рекомбинантная ДНК, содержащая lys C ген.
Более конкретно, хромосомальная ДНК подвергается перевариванию рестрикционным ферментом, например, Sau 3A I при температуре 30oC или выше, предпочтительно при 37oC при концентрации фермента 1-10 ед./мл в течение различного периода времени (1 мин - 2 ч) для полного или частичного переваривания с целью получения множества фрагментов хромосомальной ДНК. Векторная ДНК, способная к репликации в бактериях рода Escherichia, подвергается также расщеплению рестрикционным ферментом, например, Bam H1, который создает те же концевые последовательности нуклеотидов, что и рестриктаза Sau 3A, использованная для расщепления хромосомальной ДНК, при температуре 30oC или выше, при конвентрации фермента 1 - 100 ед./мл в течение 1 ч или больше, предпочтительно 1 - 3 ч для достижения полного прерывания и получения расщепленной ДНК. Затем смесь, содержащая фрагменты ДНК E. col штамм MC 1061 с включенным описанным выше способом lys C геном, смешивается с расщепленной векторной ДНК и с ДНК-лигазой, предпочтительно ДНК-лигазой T4, при этом устанавливается температурный режим 4 - 16oC при концентрации фермента 1 - 100 ед. /мл в течение 1 ч или более, предпочтительно 6 - 24 ч, с целью получения рекомбинантной ДНК.
Векторная ДНК, используемая в настоящем изобретении, преимущественно является плазмидной векторной ДНК, например pUC19, pUC18, pBR 322, pHSG299, pHSG399, RSF1010 и др. K тому же может быть использована фаговая ДНК. Промоторы, функционирующие в микроорганизмах, например, lac, trp, PL и подобные могут быть использованы для эффективной экспрессии гена, необходимого для поставленной цели. Используемый здесь термин "рекомбинантная ДНК" означает ДНК, полученную после включения указанного выше гена в хромосомы по методу, включающему использование транспозона (Berg, D.E. и Berg, C.M., Bio/Technol. , 1, 417, 1983), мю-фага (раскрытие Японской заявки на патент HE 1 2-109985) или гомологической рекомбинации (Experiments in Molecular Genetics, Cold Spring Harbor Lab., 1972).
Эта рекомбинантная ДНК используется для трансформации, например, штамма K-12 E. coli или лучше штамма GT 3 и др., и затем штамм, имеющий рекомбинантную ДНК с lys C геном, выбирается из штаммов, имеющих повышенный уровень активности AK, или из штаммов с комплементарными питательными потребностями. Трансформация может быть проведена по методу Д.М.Моррисона (Methods in Enrymoligy, 68, 326, 1979) или по методу, согласно которому рециппентные клетки обрабатываются хлоридом кальция для повышения уровня прохождения в них ДНК (Mandel, M. и Higa, A., J.Mol. Biol., 53, 159, 1970). Далее рекомбинатная ДНК, полученная вставкой ДНК, содержащей lys C ген, в векторную ДНК, может быть выделена из указанного выше штамма по методу П.Гуери с сотр. (J. Bacteriol. , 116, 1064, 1973 или по методу Д.Б.Клевелл (Clewell, D.B., J.Bacteriol., 110, 667, 1972).
Можно подтвердить, что предлагаемый штамм действительно обладает рекомбинатной ДНК, содержащей lys C, приготовив раствор клеточного экстракта, а затем из него неочищенный энзиматический раствор для измерения и подтверждения активности аспартокиназы. Метод измерения активности аспортокиназы может быть проведен в соответствии с техникой Штадтмана с. сотр. (Stadtman, E. R. , Cohen, G.N., LeBras, G., и Robichon - Szulmajster, H.,J.Biol. Chem., 236, 2033, 1961).
Lys C ген может быть получен альтернативным способом: за счет амплификации lys C гена хромосомальной ДНК в результате ЦПР (цепной полимеразной реакции, см. White, T. J.et al Trends Genet. 5, 185, 1989). Для амплификации используется праймер, комплементарный 3- концу двухцепочечной ДНК, содержащей целиком регион lys C гена или его сегмент. Если амплифицируется только часть гена lys C, то в этом случае необходимо провести скрининг библиотеки генов с целью отыскания фрагментов ДНК, содержащих целиком этот регион, используя данный фрагмент ДНК в качестве праймера. Если амплифицируется целиком нужный регион, то затем фрагмент ДНК подвергается электрофорезу в агарозном геле и далее нужная полоса вырезается для получения ДНК фрагмента, содержащего lys C ген.
ДНК - праймер может быть соответствующим образом приготовлен на основе, например, известной нуклеотидной последовательности E. coli (Cassan, M., Parsot, C. , Cohen, G.N. и Patte, J.C., J.Biol, Chem., 261, 1052, 1986) и предпочтительны два типа праймеров: 5' CTTCCCTTGTGCCAAGGCTG-3' (SEQ IDNO : 2) и 5' - GAATTCCTTTGCGAGCAG-3' (SEQIDNO : 3), которые способны к амплификации региона 1347-ного основания, кодирующего les C ген. Синтез ДНК может быть выполнен традиционным способом с использованием Модели для синтеза ДНК 380В, производимой Эпплайд Биосистемз Ко (Applied Biosistems Co.) или с использованием фосфоамидного метода (см. Tetrahedron Letters 22, 1859, 1981 ЦПР может быть проведена с использованием Модели термоблока для работы с ДНК PJ2000, производимой Такара Щузо Ко (Takara Shuzo Co.) и Tag ДНК - полимеразы, производимой Такара Шузо Ко в соответствии с методом, указанным поставщиком.
Амплифицированный по ЦПР методу lys C ген соединяется с векторной ДНК, способной к репликации в бактериях Escherichia, и после этого вводится в клетку. Для трансформации в хозяина векторной ДНК и подтверждения присутствия lys C гена используются те же самые методы, что и описаны выше.
Введение мутации в полученный lys C ген, таких как замещение, вставка или делеция, может быть достигнуто с помощью метода рекомбинатной ЦПР (Higuchi, R. , 61, in PCR Technology (Erlich H.A., Eds., Stockton Press, 1989)), методом сайт- направленного мутагенеза (Kramer, W. and Frits, H.J., Meth. in Enzymol. , 154, 350, 1987; Kunkel, T.A. et al., Meth in Enzymol., 154, 367, 1987) и др. С использованием этих методов можно получить нужную мутацию в нужном месте. Кроме того, с целью получения случайных мутаций можно использовать прямую обработку гена, хромосомной ДНК или плазмиды гидроксиламином (Hashimoto, T, и Sekiguchi, M.,J.Bacteriol, 159, 1039, 1984); традиционным методом, включающим облучение клеток, содержащих данную для работы ДНК, ультрафиолетовыми лучами, или обработку такими химическими агентами, как N-метил-N'-нитрозогуанидин, азотистая кислота и т.д.; а также методом химического синтеза заданного гена.
Метод отбора мутанта lys C* гена по снятию ингибирования включает, во-первых, трансформацию AK - зависимого штамма мутированной рекомбинатной ДНК, например, штамма GT3 E. coli. Затем трансформант культивизируется в минимально среде, например М9, содержащей значительное количество лизина. Штаммы, которые обладают плазмидами, имеющими дикий тип lys C, демонстрируют ингибирование AK III лизином, а поскольку это единственная AK а штамме, то синтез треонина, изолейцина, метионина и диаминопимелиновой кислоты (ДАП) становятся уже невозможными и рост останавливается. В отличие от них штаммы, которые имеют плазмиды с lys C*, который в свою очередь способен снимать ингибирование лизином, должны расти на минимальной среде, содержащей значительное количество лизина. Используя это явление, нужны штаммы, т.е., штаммы, содержащие плазмиды с lys C*, которые снимают ингибирование, т.е. те, которые устойчивы к лизину или к S 2-аминоэтилцистеину (АЭЦ), аналогу лизина, могут быть отобраны.
Упомянутый выше мутантный ген, полученный указанным способом, может быть использован в качестве рекомбинантной ДНК для введения в соответствующий микроорганизм хозяина в экспрессии в нем для получения микроорганизма, обладающего AK, способной к рилизингу в процессе ингибирования по типу обратной связи.
Упомянутый выше дикий тип бактерий Escherichia можно назвать одним из таких хозяев, в которых полученный lys C ген или мутант lys C гена (lys C*) может быть введен и затем амплифицирован для продукции треонина, в дополнение к ним, однако, могут использоваться и другие бактерии в качестве микроорганизма-хозяина при том условии, что в них функционируют и сайт инициации репликации сконструированной рекомбинантной векторной ДНК, и lys C ген или мутированный lys C ген (lys C*), к тому же рекомбинантная векторная ДНК должна быть способна к репликации в них, а действие lys C гена или мутированного lys C гена (lys C*) может быть усилено в них. Наиболее предпочтительным хозяином является E.coli штамм В-3996.
Трансформанты, полученные указанным выше способом и которые обладают рекомбинатной векторной ДНК, содержащей ген, кодирующий аспартокиназу III, способную к рилизингу в процессе ингибирования по типу обратной связи лизином, культивируются, при этом продуцируется необходимый L-треонин, который накапливается в культуральном растворе и затем выделяются из него и собираются.
Культуральная среда, требуемая для продуцирования L-треонина, представляет собой обычную культуральную среду, содержащую источник углерода, источник азота, неорганические ионы и, если нужно, другие компоненты органического характера.
В качестве источника углерода могут быть использованы сахара, такие как глюкоза, лактоза, галактоза, фруктоза, частично гидролизованный крахмал, а также спирты, такие как глицерин, сорбитол, или органические кислоты, такие как фумаровая кислота, лимонная кислота, янтарная кислота и т.п.
В качестве источника азота могут быть использованы неорганические соли аммония, такие как сульфат аммония, хлорид аммония, фосфат аммония и др., а также источники азота органического происхождения, например соевый гидролизат, это могут быть и аммиак, и аммиачная вода и т.п.
Необходимые вещества, такие как витамин B1, L-гомосерин и др., или дрожжевой экстракт лучше добавлять в соответствующих количествах как органические пищевые добавки. В дополнение к ним, в небольших количествах, добавляют фосфат калия, сульфат магния, ионы железа и т.д., если в этом возникает необходимость.
Культивирование предпочтительно проводить в условиях аэрации, при контролируемой температуре 25 - 45oC при pH культуры, равном 5 - 8. Для контроля pH могут использоваться неорганическая или органическая кислоты, щелочное вещество, аммиак и др. Выделение L-треонина из ферментативного сусла производится способом, сочетающим традиционный ионный обмен на смоле, метод осаждения и другие хорошо известные методы.
Фиг. 1 представляет схематически путь биосинтеза треонина.
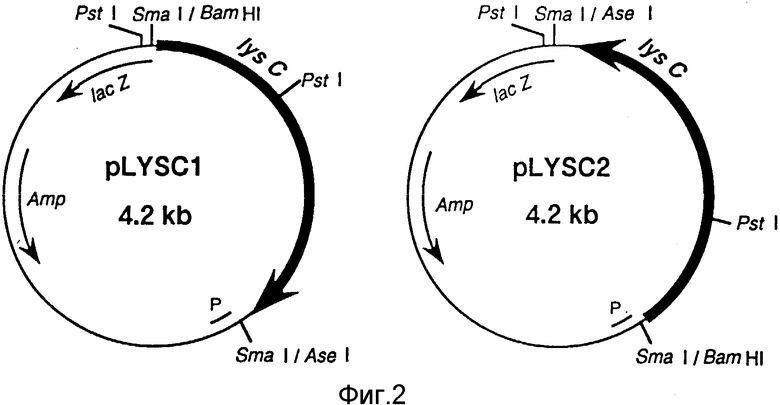
Фиг. 2 представляет собой рестрикционную карту для PLYS C1 и PLYS C2 (см. пример 1).
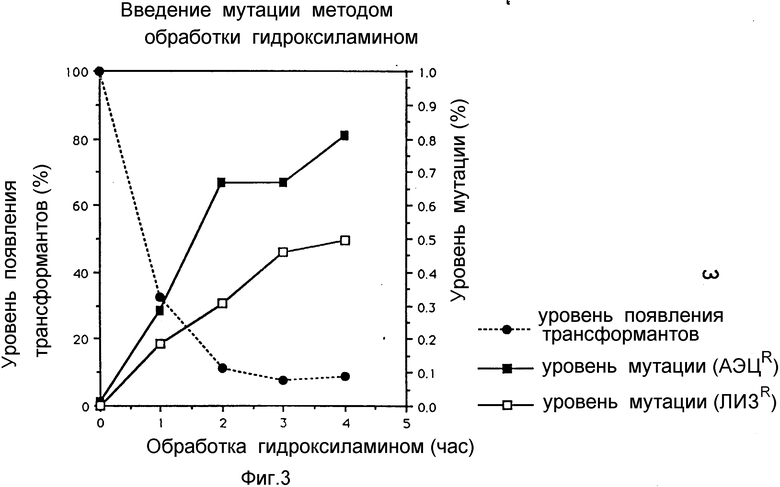
Фиг. 3 представляет собой графическое изображение эффекта введения мутации, индуцированных гидроксиламином, на lys C (пример 2 (2-4)).
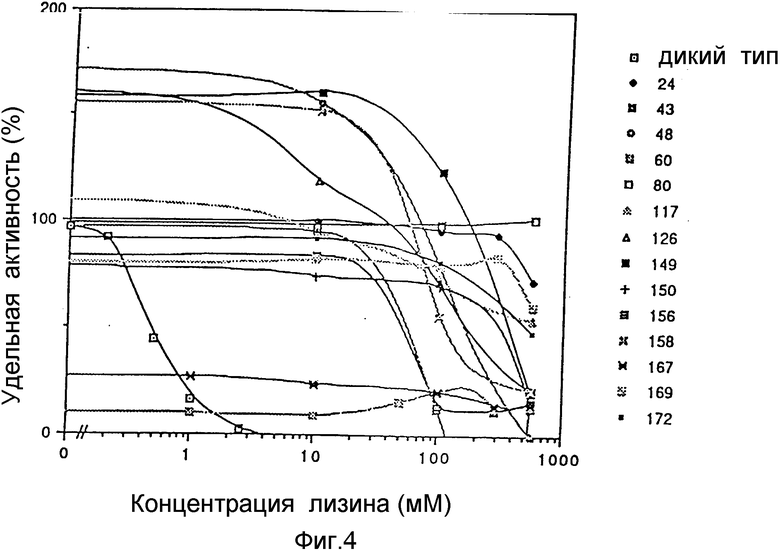
Фиг. 4 демонстрируется степень лизин-зависимого ингибирования аспартокиназ, кодируемых различными lys C* генами (пример 3 (3-4)). Концентрация лизина на горизонтальной оси выражена в виде логарифмов их значений. Удельная активность на вертикальной оси выражается в виде отношения к активности AK III дикого типа с 0 мМ добавкой лизина, принятой за 100%.
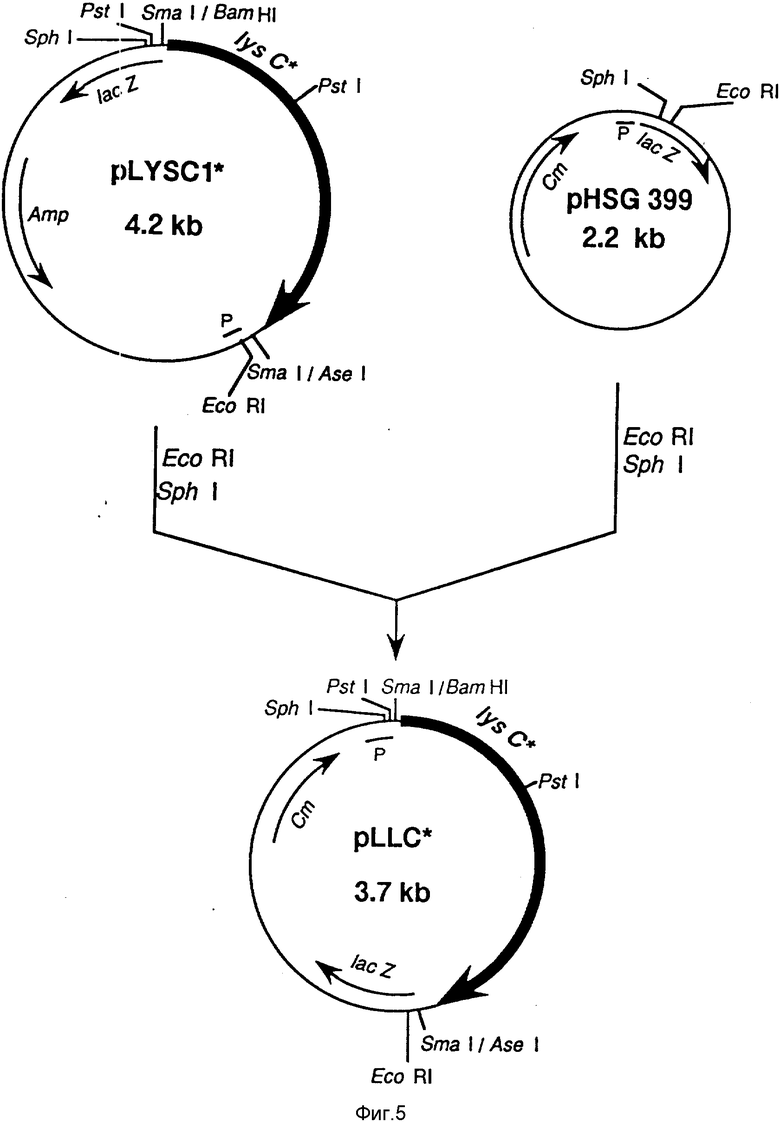
Фиг. 5 демонстрирует схему конструирования pLLC* (пример 4 (4-1)).
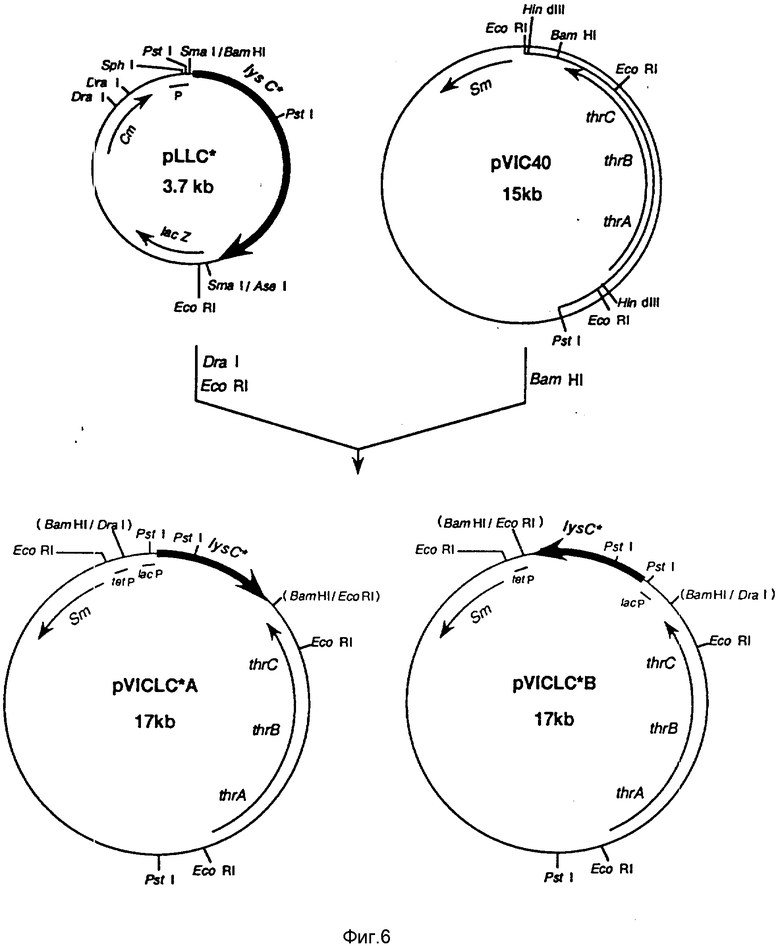
Фиг. 6 демонстрирует схему конструирования p W I CLC*A и p VICLC*B (пример 4 (4-3)).
Фиг. 7 указывает точки мутации каждого LYS C* (пример 5 (5-3)).
Лучший способ выполнения изобретения.
Пример 1. Клонирование lys C гена дикого типа.
Последовательность оснований в lys C гене E/ coli была уже определена (Cassan, M. , Parsot, C. , Cohen, G.N. n Patte, J.C., J.Biol. Chem., 261, 1052, 1986), известно также, что его открытая рамка для считывания (OPC) составляет 1347 оснований и кодирует 449 аминокислот. В случае наличия оператора, ответственного за репрессию лизином, этот оператор должен быть удален, и для клонирования подвергнут амплификации только регион, содержащий SD последовательность и OPC, используя метод ЦПР. Два разных праймера: 5'-CTTCCCTTGTGCCAAGGCTG-3' (последовательность N 2) и 5'-GAATTCCTTTGCGAGCAG-3' (последовательность N 3) приготавливаются, а полная ДНК E.col K-12 штамма ИС 1061 выделяется по методу Саито и Миира (Biochem. Biophys. Acta., 72, 619, 1963). Все эти компоненты используются в ЦПР по методу Эрлиха с сотр. (PCK Technology, Stockton Press, 1989) для амплификации нужной ДНК. Полученная ДНК переваривается рестриктазами Bam HI и AseI, затем их концы делаются десфосфолированными и ДНК встраивается в Sm aI - сайт мультикопийного вектора pUC18. Получается два типа плазмид: одна - нечувствительная (pLYS CI) и вторая - чувствительная (pLYS C2) в отношении IacZ промотора (фиг. 2).
Эти плазмиды были использованы для трансформации E.coli штаммов GT3 (thr A1016b, metL M1005, lys C1004),абсолютно дефицитных в отношении AK I, II, III, и поскольку потребности штамма GT3 в гомосерине + диаминопимелиновая кислота были удовлетворены, этот факт подтверждает присутствие гена lys C, который кодирует активную AK III,
Пример 2. Получение мутантного гена lys C*, способного снимать ингибирование.
(2-1);
Для эффективного получения lys C гена, способного снимать ингибирование (lys C*) ген рекомбинатной плазмидной ДНК, полученной в примере 1, подвергается мутагенной обработке.
Способ получения мутантного гена lys C, способного снимать ингибирование, включает, во-первых, включение с целью трансформации рекомбинантной плазмиды с диким lys C в абсолютно AK-дефицитный штамм E.cpli GT3. К минимальной питательной среде М9, имеющей состав, приведенный ниже, добавляется значительное количество лизина, и на этой среде культивируется трансформанты. Штаммы, обладающие плазмидами с диким типом lys C, испытывают ингибированием единственной AK, т.е. AK III, лизином, и поэтому синтез треонина, изолейцина, метионина и диаминопимелиновой кислоты (ДАП) становится далее невозможным, и рост останавливается.
Минимальная питательная среда М9
A : 20 • M9 - г/л
Na2HPO4 - 303
KH2PO4 - 60
NaCl - 10
NH4Cl - 20
B : I M MgSO4
C : 50% глюкозы
D : I г/л тиамина
A, B, C, D и вода стерилизуются отдельно и смешиваются в следующей пропорции A:B:CoCD : вода = 5 : 0,1 : 0,1 : 95.
В отличие от них, штаммы, обладающие плазмидами с геном lys C*, который снимает ингибирование лизином, должны быть способны к росту на минимальной питательной среде, содержащей значительное количество лизина. Используя это явление, провели селекцию штаммов, рост которых устойчив к лизину или 2-аминоэтилцистеину (АЭЦ), аналогу лизина, т.е. штаммов с плазмидами, имеющими lys C*, который снимает ингибирование.
(2-2);
Изучение условий селекции lys C мутанта (lys C*), устойчивого к ингибированию по типу обратной связи.
Во-первых, pLYS C1 и pLYS C2 оба включаются в E.coli с целью получения двух различных трансформантов, далее культивирование проводится на минимальной М9 агаризованной твердой питательной среде в бактериологических чашках, содержащей лизин или АЭЦ. При этом определяется ингибирующая рост концентрация лизина или АЭЦ, и далее изучаются условия селекции lys C*.
Как показано в табл. 1, видно в отношении pLYS C2, что уровень экспрессии амплифицирован LacZ промотором, и в этом случае наблюдается устойчивость к достаточно высоким концентрациям лизина или АЭЦ даже в случае дикого типа lys C, но если lys C ген в pLYS C2 нечувствителен к LacZ промотору или отсутствует сам промотор lys C гена, то уровень экспрессии оказывается низким и рост ингибируется даже при более низких концентрациях лизина или АЭЦ (рост полностью ингибируется при добавлении лизина или АЭЦ в концентрации около 0,2 мМ. "+" в таблице означает рост трансформантов, а "-" означает отсутствие их роста). Было показано, что такое ингибирование роста снимается и рост восстанавливается одновременным добавлением гомосерина и ДАП.
Таким образом, для эксперимента введения мутации использована плазмида pLYS C1, а в качестве среды, используемой с целью селекции lys C*, применяется М9 минимальная питательная среда, в которую добавляется 10 мМ лизина или 0,2 мМ АЭЦ.
(2-3); Мутагенная обработка
Гидроксиламин представляет собой химический мутаген, который индуцирует мутацию Ц Т типа за счет превращения цитозина в N4 - оксицитозин. Для введения мутации в плазмиду используются два метода: один представляет собой прямую обработку плазмиды in vitro гидроксиламином, а другой метод, включающий мутагенную обработку in vivo, обеспечивает разнообразие мутаций, т.е. отличных от мутаций ЦТ типа, в этом случае клетки, обладающие плазмидами, обрабатываются нитрозогуанидином (НТГ), и затем плазмиды выделяются.
(2-4); Мутационная обработка in vitro гидроксиламином
Порция ДНК в 2 г ДНК обрабатывается реакционным раствором, состав которого приведен ниже, содержащем 0,4 М гидроксиламина, при 75oC в течение 1-4 ч. Обработанная ДНК очищается с помощью стеклянного порошка и затем используется для трансформации в полностью AK - дефицитный штамм GT3, который помещается на полную среду (среда Луриа; 1% бактотриптон, 0,5% дрожжевой экстракт, 0,5% NaCl, 1,5% агар) для образования колоний на чашке Петри. Полученные колонии перенесли для выращивания на среду для селекции, описанную в (2-1). Как показано на фиг. 3, уровень выявленных трансформантов и мутаций варьирует. В случае четырехчасовой обработки выход мутантного штамма достаточно высокий и составляет 0,5 - 0,8%.
Состав реакционного раствора
0,1 M KH2PO4 - I мМ ЭДТА (pH 6,0) - 100 μл
1M гидроксиламин - 1 мМ ЭДТА (pH 6,0) - 80 μл
ДНК - 2 μг
Вода - Баланс
Всего - 200 μл
(2-5);
Мутационная обработка нитрозогуанидином (НТГ) in vivo
Для трансформации pLYS C1 используется E. coli штамм МС 1061, при этом клетки подвергаются прямой обработке НТГ. Для закрепления мутации обработанные клетки культивируются в течение ночи, после чего из них извлекаются плазмиды и используются далее для трансформации в GT3, после чего способом, описанным в (2-4), проводится скрининг для получения устойчивых к лизину (LysR) или устойчивых к АЭЦ (AECR) мутантов. Ниже представлена схема такой обработки.
Схема обработки
(1) Культивирование клеток, содержащих нужную плазмиду в 2 х TY среде (1,6% бактотриптон, 1% дрожжевой экстракт, 0,5% NaCl), затем проводится сбор клеток при оптической плотности, равной примерно 0,3 при 660 нм.
(2) Промывание ТМ буфером.
(3) Суспендирование в растворе НТГ (0,2 мг/мл в ТМ буфере), после чего обработка при 37oC продолжительностью 0-90 мин.
(4) Промывание ТМ буфером и 2 х Т средой, затем проводится культивирование в течение ночи 2 х Т среде и закрепление мутации.
(5) Выделение плазмидой ДНК из клеток, трансформации GT3 штамма и скрининг рекомбинантных штаммов.
TM буфер имеет следующий состав:
Трис - 50 мМ
Малеиновая кислота - 50 мМ
(NH4)2SO4 - 1 г/л
MgSO4•7H2O - 0,1г/л
Ca(No3)2 - 5 мг/л
FeSO4•7H2O - 0,25 мг/л
pH доводится до 6,0 с помощью NaOH.
Пример 3. Выделение lys C* гена
(3-1);
Все 180 штаммов, полученных в примере 2 (обработанные гидроксиламином = 48 штаммов и обработанные ТНГ = 132 штамма) снова помещали в среду для селекции, и с использованием АЭЦ и лизина было получено 153 устойчивых к ним штамма. Невзирая на имеющиеся различия в плане накопления аминокислоты в среде, 153 штамма разделили на 14 групп, представители каждой из которых были отобраны для измерения AK - активности. Поскольку не было существенных различий между мутантами, обработанными соответственно гидроксиламином и НТГ, различия не делались между ними и в дальнейшем в ходе эксперимента.
(3-2); Измерение аспартокиназой активности
Полностью AK - дефицитный штамм GT3 был использован в качестве хозяина, в отношении которого проводилась трансформация соответственно указанными выше 14-ю мутантными плазмидами PLYS C1 (названными PLYS C1* серией) и плазмидами pLYS C1 дикого типа, после чего из трансформантов приготовили бесклеточный экстракт, и было проведено определение активности AK III. Метод измерения энзиматической активности представлен ниже.
(3-3); Метод измерения активности AK III
(3-3-1);
Метод приготовления неочищенного энзиматического раствора
(1) Культивирование клеток в 2 х TY среде, затем сбор клеток в фазе оптической плотности примерно 0,3 при 660 нм.
(2) Промывание при 0oC 0,02 M KH2PO4 (pH 6,75) - 0,03 M β -меркаптоэтанолом.
(3) Звуковое разрушение клеток (0oC, 100 Вт, 30 с х 4 раза).
(4) Центрифугирование в течение 1 ч при 0oC при 33 тыс. об/мин, затем к супернатанту добавляется сульфат аммония до 80% насыщения.
(5) Центрифугирование, растворение пеллет в буфере (2) и сохранение ( - 20oC).
(3-3-2); Метод измерения энзиматической активности
Метод измерения энзиматической активности соответствует приведенному Штадтманом с соавт. (Stadtman, E.R., Cohen, G.N., Lebras, G. и Robichon-Czulmajster, H., J. Biol. Chem., 236, 2033, 1961).
Реакционный раствор имеет состав, приведенный ниже
Состав реакционного раствора, мл:
Реакционная смесь(*1) - 0,3
Раствор гидроксиламина(*2 - 0,2
0,1 М аспартат калия pH 7,0 - 0,1
Энзиматический раствор
Вода - Баланс
Всего - 1,0
*1 : 9 мл 1 М Трис-HCl (pH 8,1) oC 0,5 мл 0,3 М MgSO4 + 5 мл 0,2 M АТФ (pH 7,0)
*2 : 8 М гидроксиламина, нейтрализованный перед самим применением с помощью KOH.
Реакционный раствор без аспартата калия определяется как чистый.
Ниже дано схематическое описание метода измерения активности.
Схема метода измерения.
(1) Инкубирование реакционного раствора при 27oC в течение 45 мин.
(2) Добавление раствора FeCl3 (0,4 мл 2,8N HCl + 0,4 мл 12% TXY + 0,7 мл 5% FeCl3•6H2O/0,1N HCl) для окрашивания.
(3) Центрифугирование, затем измерение величины поглощения супернатанта при 540 нм (A540).
Активность выражается как количество гидроксамовой кислоты, продуцируемой в одну минуту.
I U + I μ мол/мин. Коэффициент молярного поглощения = 600.
Результаты представлены на фиг. 4
(3-4) Степень рилизинга лизин-зависимого ингибирования
Для измерения ферментативной активности AK в энзиматические реакционные растворы добавляется в различных концентрациях лизин, и далее определяются уровни лизин-зависимого ингибирования. AK III дикого типа очень сильно ингибируется лизином, т.е. на 50% при 0,45 мМ концентрации лизина и практически на 100% при 5 мМ концентрации лизина (фиг. 4).
В противоположность им, полученные здесь мутанты AK III демонстрируют различную степень релизинга, но все 14 имели некоторый уровень освобождения от лизин-зависимого ингибирования (фиг. 4 и табл. 2). Особенно хорошо это заметно на NN 24, 80, 117, 169 и 172, в которых практически не наблюдалось ингибирования при концентрации лизина 100 мМ и всего на 50% при концентрации лизина в 200 раз выше.
То же самое наблюдается и в отношении удельной активности; активность фермента на единицу белка у мутантов была либо равной, либо выше, чем у дикого типа, при этом условии роста клеток и приготовления образцов не сказывались на ее снижении, равно как не возникало проблем со снижением активности фермента в связи с введением мутации (табл. 2). Из этих данных можно сделать заключение, что центр активности AK III и центр, ответственный за регуляцию лизином, независимым друг от друга.
В табл. 2 уровень освобождения от ингибирования изображается как % активности фермента в присутствии 100 мМ лизина и от его активности в реакционном растворе без лизина, а термическая стабильность изображается как % сохранения активности после обработки при 55oC в течение 1,5 ч от его активности без нагревания.
(3-5); Термическая стабильность
Когда есть необходимость повышать уровень активности какого-либо фермента, важно, чтобы тот или иной фермент стабильно поддерживался в клетке. Так, например, протеазы имеют различия в их активности внутри и за пределами клетки, и в этой связи, а также в связи с влиянием сохраняющего буфера, измерения из активности предпочтительно проводить in vivo, но здесь ради удобства термическая стабильность как параметр для оценки AK III фермента в каждом мутанте изучалась in vitro.
В результате различных подходов к выбору температуры, при которой происходит инактивация AK III была выбрана температура 56oC, и через 90 мин после начала такой обработки определяется уровень сохранения активности фермента. Как показано в табл. 2, половина мутантов имеет уровень выше, чем дикий тип. Как правило, мутированные белки имеют меньшую стабильность, чем белки дикого типа, но некоторые из видов мутантов, полученных здесь, имеют более высокую стабильность, акт, чем дикий тип, и многие из них, мы полагаем, представляются весьма полезными в плане практического получения L-треонина.
Пример 4. Ферментативное получение L-треонина с использованием штаммов с введенным lys C*.
(4-1);
Из известных к настоящему времени треонин-продуцирующих бактерий E. coli, наибольшей способностью к продукции треонина обладает штамм В-3996. Было решено использовать штамм В-3996 в качестве хозяина для оценки возможностей lys C*. Штамм В-3996 находился на хранении в научно-исследовательском Институте Генетики и Селекции Промышленных Микроорганизмов и был зарегистрирован под N ВКПМ (V KPM) В-3996. Далее, для оценки lys C* было выделено 6 типов с различной степенью снятия ингибирования и с различной удельной активностью (NN 24, 43, 60, 80, 149 и 167 в табл. 2) для использования их в следующем эксперименте.
Во-первых, с целью повышения уровня экспрессии lys C*, каждый из упомянутых 6-и типов lys C* на pLYS C1* был подвергнут векторному переносу генов на вектор pH SG 399 (продукт Такара Шузо Ко) в прямом направлении LacZ промотора (в направлении 5' - 3') для усиления способности каждого lys C* к инвертируемой инсерции. Полученные таким способом новые плазмиды получили коллективное название pLLC* серии (фиг. 5). E. coli штамм HB101, в которые были вставлены указанные выше lys C* NN 80 и 167, обозначали соответственно как AJ 12750 и AJ 12751, и которые были первоначально депонированы в Национальном Институте Бионаук и Технологии в отношении Человека (Япония) с 1 сентября 1992 года. AJ 12750 был присвоен N FERM P-13136 (зарегистрированный в Международном депозите с 4 сентября 1993 г. под N FERM BP-4462), а AJ 12751 был присвоен номер FERM 13137 (измененный с того же самого времени, что и указанный выше на N FERM BP-4463). Остальные не были депонированы, поскольку все точки мутации для каждого lys C* были определены, как описано ниже, каждый, обладающий знаниями среднего специалиста в данной области, может легко восстановить плазмиды из названных депонированных бактерий по методу Маниатиса с сотр. (Sambrook, J., Fritsch, E.F. Maniatis, T., Molecular Cloning, Cold Spring Harbor laboratory Press, 1, 21, 1989), получить остальные lys C* гены с использованием метода сайт-направленного мутагенеза (Sambrook, J. , Fritsch, E.F., Maniatis, T., Molecular Cloning, Cold Spring Harbor Laboratory Press, 15, 83, 1989). Для вставки этих плазмид в штамм В-3996 с целью оценки был использован традиционный метод.
Культивирование микроорганизмов проводилось на среде, состав которой приведен ниже (при этом компоненты A, B и C стерилизуются отдельно) в течение 38 ч при 37oC при перемешивании со скоростью 114-116 об/мин.
Среда для продукции треонина
A : - г/л
(NH4)2SO4 - 16
KH2PO4 - 1
MgO4•7H2O - 1
FeSO4•7H2O - 0,01
MnO4•5H2O - 0,01
Дрожжевой экстракт (Difco) - 2
L - Мет - 0,5
pH доводится до 7,0 с помощью KOH, и компонент A стерилизуется при 115oC в течение 10 мин (16/20 объем).
B : 20% глюкоза, автоклавируется при 115oC в течение 10 мин (4/20 объем. )
C : Фармакопейный CaCO3, автоклавируется при 180oC в течение двух дней (30 г/л).
D : Антибиотик (100 г/мл стрептомицина и 5 г/мл канамицина)
(4-2); Оценка lys C*
Каждая из шести плазмид pLL C* серии была использована для трансформации В-3996 штамма, и каждый трансформант культивировался как в условиях с добавкой 1 г/л лизина, так и без него. В качестве контроля использовали штамм-хозяин В-3996, обладающий и не обладающий плазмидой с диким типом lys C (pLLC1).
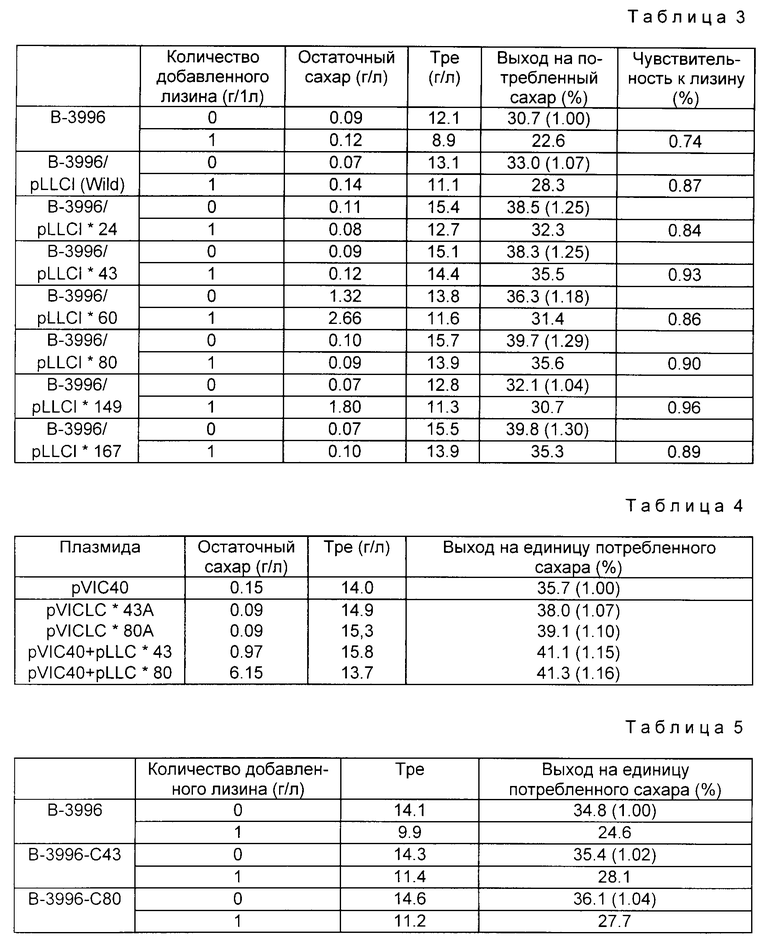
Результаты представлены ниже в табл. 3. В этой таблице значения в круглых скобках означают отношение выхода лизина (г) на 1 г потребленного сахара к выходу лизина (г) на 1 г потребленного сахара, полученного в том случае, когда В-3996 культивируется без добавления лизина (контроль). Это означает, что последний компонент рассматривается равным 1,00. Чувствительность к лизину определяется как (выход лизина на единицу потребленного сахара с добавлением лизина) / (выход лизина на единицу потребленного сахара без добавления лизина). В отношении штамма В-3996 снижение выхода на единицу потребленного сахара в культуре с лизином составляло примерно 0,74 по сравнению с величиной, полученной в культуре без добавления лизина.
Однако в случае штаммов, в которые были введен lys C*, к примеру в штамме В-3996 / pLLC* 149, снижение выхода на единицу потребленных сахара в культуре с лизином уже составило 0,96 по сравнению с аналогичным показателем в культуре без добавления лизина. Из этого можно предположить, что AK III, кодируемая lys C, вносит определенный вклад в биосинтез треонина и что ингибирование активности AK III при добавлении лизина, в свою очередь, связано с ингибированием биосинтеза треонина. Этот феномен уменьшает свое влияние, когда используется полученный в настоящем изобретении lys C*
При этом, рассматривая продукцию треонина культурой без добавления лизина, видно, что штаммы, имеющие плазмиды с диким типом lys C, демонстрируют более высокую продуктивность по сравнению со штаммами без плазмид в отношении треонина, более того, уровень этой продукции еще более возрастает у штаммов, имеющих мутантные lys C* (примерно в 1,3 раза, чем у хозяина в случае N 80). Эти результаты показывают, что AK III является фактором, лимитирующим уровень продукции треонина штаммов В-3996. Применяя клетки, в которые был введен мутантный lys C*, для культивирования без лизина, получается более высокий выход, чем в случае клеток, в которые был введен дикий тип lys C, что дает основание считать, что последние подвергают ингибированию содержащуюся в них AK III лизином, который сами клетки синтезируют.
(4-3); Стабилизация плазмид с мутантным lys C*
Как показано на фиг. 6, плазмиды с lys C* приготавливают за счет включения его в обоих направлениях от Bam HI н. сайта в плазмиду pVIC40, выделенную из В-3996 штамма. Метод выделения pV IC40 из штамма В-3996 был уже описан выше. Выбор был сделан в пользу N 80, отличающегося высоким уровнем продукции треонина, так же, как и наличием lys C*, и номера 43 для сравнения, при этом было получено 4 различных плазмиды, т.е. p. V ICL C*80A, pv ICL C* 80B, pV ICL C* 43A и pV ICL C* 43B.
Для использования штамма В-399 в качестве хозяина (В-3996 с удаленной плазмидой pVIC40) он был изготовлен за счет обработки штамма В-3996. Необходимая обработка была достигнута за счет трехкратной обработки средой Луриа без стрептомицина (1/100-кратное разведение) культуральной среды штамма В-3996.
Культивирование проводится при температуре 40oC. В полученный таким образом штамм В-3996 были введены pVICL C* 80A и pV ICL C* 43A, соответственно, из четырех плазмид, имеющих включенный lys C*, и культивирование продолжалось. Результаты представлены в табл. 4.
Штаммы, в которые были введены pV ICLC* 80A или pVICL C* 43A демонстрируют повышенный выход над pVIC40 штаммом-трансформантом (В-3996), использованным в качестве контроля (в 1,10 раз в отношении pVICL C* 80A и в 1,07 раз в отношении pVICL C*43A).
Можно предположить, что снижение степени возрастания выхода в сравнении с результатами в (4-2) связано со снижением числа копий в результате изменения вектора pVIC40, однако рост был такой же, и плазмиды достаточно стабильно поддерживались в штамме. (4-4) Включение мутантного lys C* гена в хромосомы.
Включение мутантного типа lys C* проводилось с использованием явления гомологической рекомбинации в треонин-продуцирующий штамм В-3996 с прицелом на lys C ген дикого типа на хромосоме. Использованный здесь метод представляет модификацию метода Руссела с соавт. (Russel, M. и Model, P.,J. Bacteriol., 159, 1034, 1984).
Если хозяином является мутант E. coli т.е. ММ382 штамм (poIAts, доступный для получения в Центре генетической информации, США), то плазмида pHSG 399 может реплицироваться при 37oC (пермиссивная температура), но не при высоких температурах, таких, как 42oC (непермиссивная температура). Этот фактор был использован в гомологической рекомбинации.
Во-первых, pLL C43* и pLLC80* были введены по отдельности в мутантный штамм ММ383 как в хозяина при 37oC, когда репликация возможна, для того, чтобы получить соответственно трансформанты ММ383/ pLL C* 43 и ММ 383/ pLLC* 80/ Эти трансформанты культивировались при 42oC, после чего были отобраны те из них, которые лишились плазмид, но сохранили устойчивость к хлорамфениколу. Плазмиды включились в хромосомы этих штаммов. Полученные штаммы получили наименование ММ383-С43 и ММ383-С80 соответственно. Эти штаммы были инфицированы PI фагом, был затем приготовлен фаговый раствор, который использовали для инфекции В-3996 штамма. Были отобраны штаммы, содержащие lys C*, трансдуцированные в хромосомы, с использованием признака устойчивости к хлорамфениколу как маркера, полученные штаммы были названы соответственно В-3996-С43 и В-3996-С80.
Культивирование В-3996-С43 и В-3996-С80 проводилось в тех же условиях, что и в случае (4-1), а результаты этого исследования представлены ниже в табл. 5. При этом было получено примерно 4% улучшение выхода треонина.
Пример 5. Определение последовательности оснований дикого типа lys C и мутантного lys C*
(5-1);
Используя секвенатор "Модель ABI 373A" (ex ABI Co.), для определения последовательности оснований в lys C дикого типа, применили традиционный метод такого анализа. Результаты приведены в описании последовательности SEQ IDN : I. Было обнаружено, что эта последовательность отличается от опубликованной ранее последовательности оснований в lys C E. coli K-12 JC411 (Cassan, M. , Parsot, C., Cohen, G.N. и Patta, J.C., J.Biol., Chem., 261, 1052, 1986) по 6 основаниям (в результате изменены 2 аминокислотных остатка) 6 найденных различий, надо полагать, связаны с различиями в используемых штаммах.
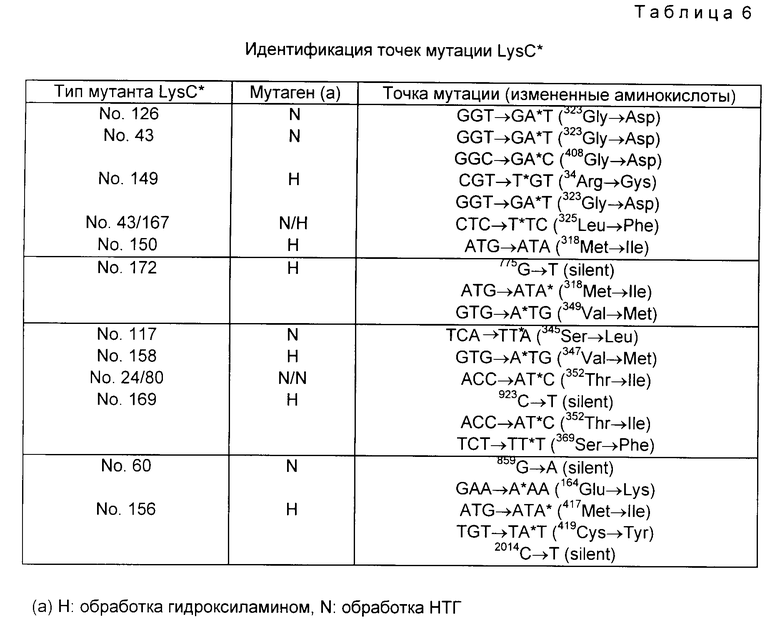
(5-2); Последовательность оснований мутантного lys C*, определяющего устойчивость к ингибированию по типу обратной связи. Последовательности оснований 14 видов lys C*, исследованных в примере 3, были определены тем же самым способом, как и в разделе (5-1), и точно найдены точки мутаций. Результаты приведены в табл. 6. Из 14 определенных видов все оказались истинными мутантами, причем две пары были абсолютно идентичны. Мутанты NN 149, 150, 156, 158, 167, 169 и 172 получены обработкой гидроксиламином, а мутанты NN 24, 43, 48, 60, 80, 117 и 126 - обработкой НТГ. Однако виды мутаций сводились к двум типам: C ---> T или G ---> A; при этом мутация G ---> A образовывались от мутации C ---> T на комплементарной цепи.
(5-3); Определение домена, ответственного за снятие ингибирования по типу обратной связи
Большое число мутаций присутствует на C-конце, и особенно велика их концентрация в A домене (от 318-го Мет остатка до 325-го Лей остатка) и в B домене (от 345-го Сер остатка до 352-го Тре остатка), как приведено на фиг. 7 (см. колонну "Точки мутации" в табл. 6). В добавление к lys C (AK III), E coli имеет еще два типа AK:trhA (AK I - HDI) и metl(M)(AK II - HD II). Мутантный lys C* имеет два участка с высокой степенью гомологии с двумя указанными типами, и эти домены, надо думать, представляют активный центр AK (Cassan, M., Parsot, C., Cohen, G.N.и Patta, J,C., J.Biol, Chem., 261, 1052, 1986).
A и B домены находятся за пределами этого общего домена, и вполне возможно, что именно они являются лизин-контролирующими доменами, специфичными к AK III. Представляется особенно важным тот факт, что мутанты с NN 24/80, 172, 169 и 172, демонстрирующие высокий уровень освобождения от ингибирования, имеют мутации в B домене. Из 12 типов 10 имеют мутацию либо в A, либо в B домене. Номера 60 и 158 представляют исключение, но как раз именно они дали очень низкий уровень снятия ингибирования и низкую термостабильность (табл. 2), что может быть результатом частичного освобождения от ингибирования в связи с изменением структуры фермента и в целом за счет мутации.
В случае мутантных типов, имеющих множественные мутации, остается под вопросом, какие из них являются эффективными, а приведенное ниже объяснение основано на сравнении с мутантными типами, имеющими единичную мутацию в том же сайте.
(1) NN 126 (1 точка), 43 (2 точки) и 149 (2 точки)
Точка мутации N 126 в A домене является общей для всех трех типов мутантов, тогда как степень освобождения от ингибирования одинакова для NN 149 и 126, а большее количество точек мутации и в N 43 выражается скорее в противоположном эффекте. Это ведет к тому предположению, что точка мутации и в A домене является единственной, отвечающей за снятие ингибирования.
(2) NN 24/80 (1 точка) и 169 (2 точки)
Из двух точек мутации N 169 одна - в B домене, совпадает с той, что имеет N 24/80. Оба демонстрируют высокий уровень освобождения от ингибирования, из чего можно сделать вывод, что точечной мутации в 24/80 достаточно для этого.
(3) NN 150 (1 точка) и 172 (2 точки)
N 150 имеет одну точку мутации в A домене, тогда как N 172 - две мутации: в A домене (такую же, как и N 150) и в B домене соответственно. Уровень освобождения от ингибирования был выше у N 172, что показывает участие и A домена, и B домена в процессе рилизинга.
Описанным выше способом идентифицировали домены, имеющие отношение к освобождению от лизин-зависимого ингибирования аспартокиназной активности в A домене (выражаясь в терминах аминокислотных остатков, размером от 318 Мет остатка до 325 Лей остатка) и в B домене (от 345 Сер остатка до 352 остатка). Однако создатели настоящего изобретения не пришли к заключению, что освобождение от лизин-зависимого ингибирования аспартокиназной активности происходит только в случае, когда происходит указанное изменение специфических аминокислотных остатков на другие определенные аминокислотные остатки. Так, например, в табл. 6 показано, что освобождение от ингибирования по типу обратной связи происходит также при изменении 323-го остатка глицина на остаток аспарагиновой кислоты, при этом каждому, имеющему средний уровень знаний в данной области известно, что такое снятие ингибирования имеет место при замещении остатка аспарагиновой кислоты на остаток глутаминовой кислоты. Это происходит в связи с тем, что множество аминокислот имеют сходную структуру, такие например, как аспарагиновая и глутаминовая кислоты, которые называют гомологичными аминокислотами, а обмен гомологичных аминокислот не рассматривается как фактор, вызывающий серьезные изменения в функциях белка (список гомологичных аминокислот найден в Protein Engineering, p.31, CMC Co., 1985).
Часто, как было отмечено, обмен негомологичными аминокислотами имеет тот же эффект, что и в случае гомологичного обмена. И с другой стороны, обмен аминокислот на гомологичные может иногда привести к серьезным изменениям в функции белка (Estell, D. A., Graycar, T.P., и Wells, J.A.,J.Biol. Chem., 260, 6518, 1988: Shcultr, S.C., и Richards,J.H., Procedure. Natl. Acad. Sci. USA, 83, 1588, 1986; Yutani, K., etal., J. Biol. Chem., 262, 13429, 1987: Yutani, K. , et al., Procedure. Natl. Acad. Sei. USA, 84, 4441, 1987: Nishiyama, M., et al., J. Biol. Chem., 266, 14294, 1991).
Кроме того, в настоящем изобретении было отмечено, что снятие ингибирования наблюдалось, когда определенные аминокислотные остатки в A и B доменах не были изменены, однако поскольку функции белков определялись на основе излучения доменов, можно предположить, что освобождение от ингибирования по типу обратной связи связано с введением одной или больше аминокислотных мутаций в домены, когда в данный момент не рассматривались. В действительности имеется множество сообщений о примерах такого рода (Furuya, H., et al., Biochemistry, 28, 6848, 1989: Furuya, H., et al. Biochem Biophys. Research. Commun., 160, 699, 1989: Cunningham, B.C. и Wella, J.A., Seience, 244, 1081, 1989).
Короче говоря, настоящее изобретение характеризуется обнаружением и раскрытием его создателями доменов, связанных с ингибированием по типу обратной связи. В приведенных примерах раскрыта лишь часть мутированных участков, однако, как было сказано выше, любому специалисту, имеющему средние знания в данной области, ясно, что мутации в других участках могут вызывать такой же эффект. В связи с этим, такие мутации также входят в гематический охват настоящего изобретения.
Как было отмечено, были получены гены AK III из бактерий рода Escherichia, способные эффективно снимать ингибирование лизином по типу обратной связи. При введении этих генов в треонин-продуцирующие бактерии возможно получение других продуцирующих треонин бактерий, продуктивность которых превышает существующий уровень техники. Успешное применение таких треонин-продуцирующих бактерий дает намного более совершенный и мощный метод ферментативного получения L-треонина, чем традиционные методы.
Перечень последовательностей
SEQ ID N 1
Длина последовательности : 2147
Тип последовательности : нуклеиновая кислота
Характеристика цепи : двухцепочечная
Топология : линейная
Молекулярный тип : гегомная ДНК
Источник происхождения:
Организм : Escherichia coli
Штамм : МС1061
Характеристика особого участка:
Ключевая особенность : - 35 сигналов
Положение : 242 .... 249
Метод определения особенности : S
Характеристика особого участка:
Ключевая особенность : - 10 сигналов
Положение : 265 ... 273
Метод определения особенности : S
Характеристика особого участка:
Ключевая особенность : связывание с праймером
Положение : 536 ... 555
Метод определения особенностей : E
Характеристика особого участка:
Ключевая особенность : связывание с праймером
Положение : 2128 : 2147
Метод определения особенности : E
Характеристика особого участка:
Ключевая особенность : RBS
Положение : 575 ... 578
Метод определения особенности : S
Характеристика особого участка:
Ключевая особенность mat nenmug
Положение : 584 ... 1930
Метод определения особенности : S
Характеристика особого участка:
Ключевая особенность : терминатор
Положение : 1941 ... 1968
Метод определения особенности : S
Описание последовательности см. в конце текста.
SEQ ID N : 2
Длина последовательности : 20
Тип последовательности : нуклеиновая кислота
Характеристика цепи : одноцепочечная
Топология : линейная
Молекулярный тип : другая нуклеиновая кислота, синтетическая ДНК
Описание последовательности:
CTT CCCTT G T G CCA AGGCT G
SEQ ID N : 3
Длина последовательности : 18
Тип последовательности : нуклеиновая кислота
Характеристика цепи : одноцепочечная
Топология : линейная
Молекулярный тип : другая нуклеиновая кислота, синтетическая ДНК
Описание последовательности:
GAA TT CCTTT GCGA GCAGE







Изобретение относится к биотехнологии. Фрагмент ДНК, который кодирует аспартокиназу III, имеет амино-кислотную последовательность дикого типа. В данной последовательности содержится мутация, снимающая ингибирование аспартокиназы III лизином по типу обратной связи. Мутация выбрана из группы: Гли 323 заменен на Асп, Гли323 на Асп и Гли408 на Асп, Арг34 на Цис и Гли323 на Асп, Лей325 на Фен, Мет318 на Иле, Мет318 на Иле и Вал349 на Мет, Сер345 на Лей, Вал347 на Мет, Три 352 на Иле, Три 352 на Иле и Сер369 на Фен, Глу164 на Лиз, Мет 417 на Иле И Цис419 на Тир. В способе получения L-треонина проводят культивирование штамма-продуцента рода Escherichia, трансформированного рекомбинантной плазмидой ДНК, содержащей вышеназванный фрагмент ДНК. Штамм культивируют в ферментационной среде с последующим выделением L-треонина. 2 с.п.ф-лы, 7 ил, 6 табл.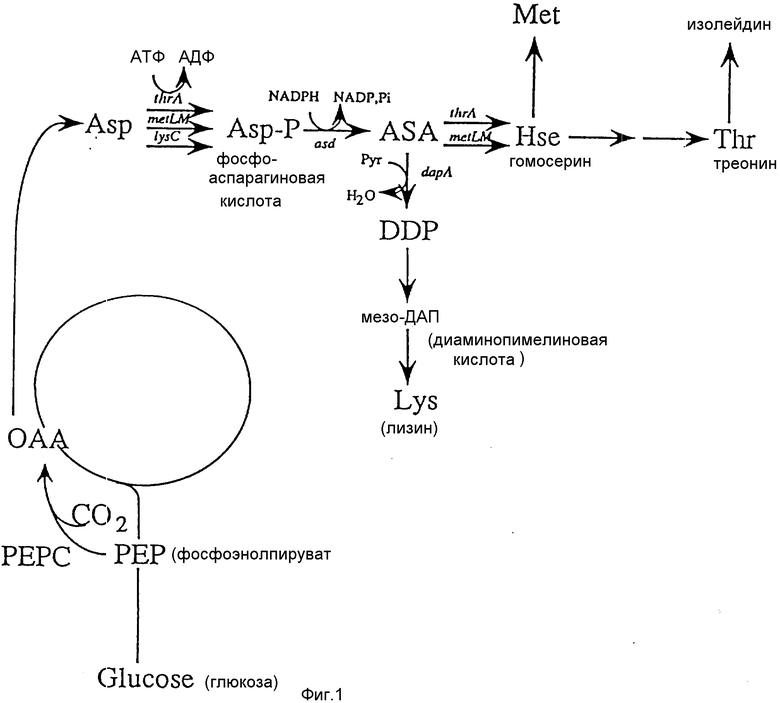
| US, 4321325, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |

