Изобретение относится к способам определения статуса гормона роста индивидуума. Определение проводят путем измерения циркулирующего в кровообращении двойного или тройного комплекса инсулиноподобного фактора роста (комплекса 150 кДа, содержащего IGF, IGFBP и ALS). Этот комплекс сначала захватывают моноклональным антителом, которое связано с твердым носителем и называется здесь "захватывающим антителом". Один из членов этого комплекса детектируют моноклональным антителом, называемым здесь "детектирующим антителом".
В идеале захватывающее антитело и детектирующее антитело не мешают друг другу или образованию комплекса. По этой причине авторы изобретения получили ряд моноклональных антител и картировали их эпитопы. Данное изобретение относится также к указанным моноклональным антителам и содержащим их наборам.
Суперсемейство инсулиноподобных факторов роста (IGF) включает в себя IGF-I и IGF-II, по меньшей мере шесть высокоаффинных IGF-связывающих белков (IGFBP-1 - IGFBP-6), кислотолабильную (не устойчивую к кислоте) белковую субъединицу (ALS) и различные рецепторы клеточной поверхности, протеазы и антагонисты. Суперсемейство IGF тесно связано с регуляцией клеточного роста и метаболизма [1-4]. IGF продуцируются многочисленными нормальными и злокачественными тканями и присутствуют в крови и других биологических жидкостях в тесной ассоциации с IGFBP, которые специфически связывают и модулируют их биоактивности [4-9] . IGF-I, пептид 7,5 кДа, является главным медиатором in vivo митогенных и метаболических эффектов гормона роста (GH) и, как в случае IGF-II, его действия опосредованы как эндокринными, так и аутокринными-паракринными механизмами [1-4].
Большая часть IGF циркулирует в кровотоке с тройным белковым комплексом приблизительно 150 кДа, состоящим из IGFBP-3 и ALS [4-9]. Связывание ALS с IGFBP-3 зависит от занятия IGFBP-3 факторами IGF-I или IGF-II, и при нормальных условиях почти весь циркулирующий в кровотоке IGF (>95%) связан в этом тройном комплексе [4-12]. IGFBP-3 в норме является главным носителем IGF в сыворотке и его ассоциация, по-видимому, стабилизирует суточные уровни IGF (период полувыведения равен ~ 12-15 часам) и ограничивает доступ IGF к внеклеточным компартментам [4-9]. Предполагается, что большая часть IGF обнаруживается в тройном комплексе, потому что ALS присутствует в значительно более высоком молярном избытке, чем IGFBP-3 или пептиды IGF [13, 14]. Меньшие доли циркулирующих IGF связаны с другими IGF-связывающими белками, и определено, что менее 1% существует в несвязанной или "свободной" форме.
Хотя точная биологическая роль тройного комплекса не была четко определена, циркулирующие уровни его компонентов и, следовательно, образование тройного комплекса, являются высоко-GH-зависимыми [4-9]. Сывороточные уровни IGF-I, IGFBP-3 и ALS являются низкими при недостаточности GH и являются повышенными в субъектах с акромегалией [4-9, 13, 15]. Посттрансляционные модификации IGFBP, в том числе протеолиз [16, 17] и фосфорилирование [18, 19], по-видимому, участвуют в модулировании биодоступности этих IGF-пептидов.
Клиническая оценка статуса GH была дискуссионной прежде всего вследствие эпизодического характера секреции GH, его относительно короткого полупериода существования в кровотоке и значительной вариабельности в измерении GH различными способами [20, 21]. В настоящее время клиническая оценка достаточности GH может включать в себя множественные взятия проб венозной крови для определения секреции GH в ответ на ряд физиологических или фармакологических стимулов. Вследствие сообщенных недостатков провокационного тестирования GH, которые включают в себя произвольное определение диагностических уровней отсечения, и потенциальный риск для здоровья, и высокую стоимость, предпринимались поиски альтернативных процедур скрининга [20, 21].
Поскольку уровни IGF-I, IGFBP-3 и ALS в крови являются высокозависимыми от секреции GH [4-9, 13, 15], подходящее определение их уровней в сыворотке было одобрено как наиболее эффективный способ в оценке статуса GH-IGF-оси, в частности, у детей с низким ростом [21-23]. Определения IGF-I и ALS могут быть также ценными в диагностике недостаточности GH у взрослых и мониторинге терапевтической ответной реакции на заместительную терапию GH [24, 25].
Высокие уровни ассоциации с IGFBP были основной проблемой в рутинном иммуноанализе сывороточного IGF-I, поскольку IGFBP может маскировать реактивные эпитопы или конкурировать с антителами за связывание метки. Достоверное определение IGF-I в сыворотке требует диссоциации и удаления IGFBP перед анализом [26] . Существующие в настоящее время стратегии основываются на подкислении сыворотки для необратимой денатурации ALS и разрушения тройного комплекса с последующими процедурами для удаления большей части или всех IGFBP. Вытеснительная (гель-фильтрационная) хроматография в кислоте рассматривается как "золотой стандартный" способ для последней стадии [27], но является непрактичной для эффективной обработки проб с высоким объемом.
Наиболее часто используемой альтернативой является осаждение смесью кислота-этанол [28] . Однако этот способ может оставлять существенные остаточные количества IGFBP [19, 29-32]. Авторы изобретения недавно подтвердили, что IGFBP, в частности низкомолекулярные IGFBP, такие как IGFBP-1, могут сохраняться в значительных количествах после осаждения смесью кислота-этанол [19]. Остаточные количества IGFBP могли бы конкурировать с детектирующим антителом, в частности в конкурентных РИА [29], где количество антитела и меченого IGF-I является небольшим относительно концентрации аналита и остаточных IGFBP. Этот феномен может быть ответственным за хорошо известную склонность РИА IGF-I давать ошибочные оценки уровней IGF-I в пробах с высокими уровнями эндогенных IGFBP [27, 29, 30, 32].
Различные средства были придуманы для минимизации вмешательства остаточных IGFBP, в том числе добавление IGF-II к тест-смеси, криопреципитация с использованием смеси кислота-этанол, использование меток-аналогов с низкой аффинностью в отношении IGF или кислотная (acid-Bio-Spin) хроматография [29, 30, 32, 33]. Однако эти способы дополнительно осложняют этот анализ и не могут полностью решить проблему вызываемых IGFBP помех.
Клиническая оценка IGF-I, IGFBP-3 и ALS дополнительно осложнена различающимися влияниями возраста, пубертатной стадии развития и факторов питания на их уровни в кровообращении [21, 34, 35]. Поэтому существующие рекомендации предлагают использование IGF-I, IGFBP-3 и ALS в группе более молодого возраста и IGF-I и ALS в случае взрослых субъектов [21-26, 36]. Так как почти все сывороточные IGF-I, IGFBP-3 [4-12] и около 50%, ALS [13-15] присутствуют в высокомолекулярном тройном белковом комплексе IGFBP-3, прямое определение этого комплекса может представлять идеальную и потенциально превосходящую альтернативу скрининга. Однако до сих пор никто не представил надежного, точного и простого способа количественного определения уровня IGFBP-комплекса.
Аббревиатуры и определения
ALS - кислотолабильная (не устойчивая к кислоте) субъединица - белок, обнаруженный в тройном комплексе 150 кДа, где обнаружена большая часть циркулирующего в крови IGF. ALS является чувствительным к инактивации кислотой.
Антитела - описанные здесь антитела включают в себя десять мышиных моноклональных антител против рекомбинантного IGFBP-3 человека, названных ВI-В10; поликлональное антитело против рекомбинантного IGFBP-3 человека, названное В-11, два мышиных моноклональных антитела к С- и N-концам рекомбинантного IGF-I человека, названные I-1 и I-2 соответственно; поликлональное антитело к рекомбинантному IGF-II человека, названное р1-3, и два поликлональных антитела к N- и С-концевым пептидным фрагментам ALS человека, названные рА-1 и рА-2. Дополнительные антитела могут быть получены способами, хорошо известными в данной области, и оценены, как описано здесь.
Двойной комплекс - состоящий из двух частей комплекс IGFBP и ALS или IGFBP и IGF.
Жидкость тела - любая биологическая жидкость, в том числе, но не только: сыворотка, плазма, лимфатическая жидкость, синовиальная жидкость, фолликулярная жидкость, семенная жидкость, амниотическая жидкость, молоко, жидкость молочной железы, цельная кровь, моча, спинальная жидкость, слюна, мокрота, слезы, пот, культуральная среда ткани слизи, экстракты тканей и клеточные экстракты.
БСА - бычий сывороточный альбумин.
Захватывающее антитело - антитело, которое может быть использовано для захвата IGFBP-комплекса.
Захватывающее антитело, связанное с твердым носителем - связывание антитела с твердым носителем может производиться любым из хорошо известных способов связывания, в том числе ковалентным, нековалентным связыванием, магнитными способами и т.п. Обычно используемые сшивающие агенты для прикрепления захватывающего антитела к твердофазному субстрату включают в себя, например, 1,1-бис(диазоацетил)-2-фенилэтан, глутаровый альдегид, эфиры N-гидроксисукцинимида, гомобифункциональные имидоэфиры и бифункциональные малеимиды. Производные агенты, такие как метил-3-[(п-азидофенил)дитио]пропиоимидат, дают фотоактивируемые промежуточные продукты, способные образовывать перекрестные связи в присутствии света. Связывание антител с различными твердыми носителями хорошо известно в данной области и нет необходимости его подробного описания здесь.
Контрольный стандарт - проба IGFBP-комплекса известной концентрации.
Детектирующее антитело - антитело, которое может быть детектировано каким-либо образом.
Детектирующее антитело, связанное с меткой - детектирующее антитело, меченное любым способом. Детектирующее антитело может само быть меченным, например, посредством включенного радиоактивного изотопа или модифицированной аминокислоты или посредством видоспецифических антигенных детерминант. Кроме того, антитело может быть помечено посредством детектируемой части молекулы (метки), связанной с этим антителом.
ELISA 1 - ELISA, основанный на захвате IGFBP-комплекса посредством антитела анти-IGFBP В3 и детектировании при помощи антитела анти-IGF-I I-1.
ELISA 2 - ELISA, основанный на захвате IGFBP-комплекса посредством антитела анти-IGFBP В2 и детектировании при помощи антитела анти-IGF-I рА-1.
GH - гормон роста.
GHRD - дефект рецептора гормона роста.
GHD - недостаточность гормона роста.
Статус GH - статус гормонов роста индивидуума отражается в уровнях GH, IGF, IGFBP, IGFBP-комплекса или ALS, а также другими молекулами, связанными с GH- и IGF-каскадами. Известно, что статус GH индивидуума варьируется с повышением или понижением в определенных связанных с ростом патологических состояниях, таких как гипогликемия, диабет, состояния, связанные с недостаточным питанием, некоторые раковые заболевания, такие как рак молочной железы и рак предстательной железы, иммунодефицитные состояния, задержка роста плода, гигантизм, акромегалия, гиперпитуитаризм, гипофизарная карликовость, недостаточность гормона роста, избыток гормона роста, дефект рецептора гормона роста, тиомегалия и определенные заболевания центральной нервной системы (ЦНС), такие как болезнь Альцгеймера. Таким образом, многие имеющие заболевания индивидуумы (люди или животные) могли бы получить пользу от мониторинга статуса гормонов роста. Кроме того, имеются другие приложения, такие как мониторинг состояния питания, репродуктивного здоровья, показателей состязаний и других показателей активности индивидуумов, улучшенного племенного поголовья животных и т.п.
HRP - пероксидаза хрена.
IGF - инсулиноподобный фактор роста.
IGFBP - IGF-связывающий белок, в том числе IGFBP-1 - IGFBP-6 и до сих пор несеквенированные IGFBP. Предпочтительно, IGFBP представляет собой IGFBP-3 в контексте описанного здесь анализа.
IGFBP-3 - основной циркулирующий в кровотоке IGF-связывающий белок.
IGFBP-комплекс - этот термин включает в себя либо двойной комплекс IGFBP и ALS или IGF, либо тройной комплекс IGFBP и ALS и IGF.
Метка - любой детектируемый маркер или детектируемая функциональная группа, которая не препятствует связыванию антитела с его эпитопом, в том числе, но не только, радиоактивный изотоп, фермент, флуоресцентная метка или другая измеряемая метка (хвост), описанные в патенте США 4275140 и патенте США 4318980. Многочисленные метки для применения в иммуноанализе являются известными, в том числе, но не только, метки, которые могут детектироваться непосредственно, такие как флуорохром, хемилюминесцентные и радиоактивные метки, а также части молекул, такие как ферменты, которые должны прореагировать или должны быть превращены в их производные для детектирования. Примеры меток включают в себя радиоактивные изотопы 32P, 14С, 125I, 3H и 131I, флуорофоры, такие как редкоземельные металлы или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, люциферин, зеленый флуоресцентный белок, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, бета-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазы, например глюкозооксидазу, галактозооксидазу, и глюкозо-6-фосфатдегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, связанные с ферментом, который использует пероксид водорода для окисления предшественника красителя, таким как пероксидаза хрена, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, метки-бактериофаги, стабильные свободные радикалы и т.п.
Кроме того, антитело само может содержать метку, например, когда детектирующее средство включает в себя второе антитело, направленное против антител из вида, из которого получено это антитело. Предпочтительной меткой является пероксидаза хрена или биотин. Количество и типы меток являются многочисленными и нет необходимости полного описания их здесь.
PSA - специфический антиген предстательной железы.
RT - комнатная температура.
SD - стандартное отклонение.
Твердый носитель - твердая фаза, используемая для иммобилизации захватывающего антитела, которая может быть любой инертной подложкой или любым инертным носителем, которые являются по существу нерастворимыми в воде и применимы в иммуноанализах, в том числе носители в форме поверхностей, частиц, пористых матриксов и т. д. Примеры общепринятых носителей включают в себя небольшие полоски, Сефадекс, поливинил-хлорид, пластиковые гранулы и тест-планшеты или тест-пробирки, изготовленные из полиэтилена, полипропилена, полистирола и т. п., в том числе 96-луночные микротитрационные планшеты, а также состоящий из частичек материал, такой как фильтровальная бумага, агароза, сшитый поперечными связями декстран и другие полисахариды. Альтернативно реакционноспособные нерастворимые в воде матриксы, такие как активированные цианогенбромидом углеводы, и реакционноспособные субстраты, описанные в патентах США с номерами 3969287; 3691016; 4195128; 4247642; 4229537 и 4330440, являются подходящими для применения в иммобилизации захватывающего антитела. Предпочтительной твердой фазой, используемой здесь, является многолуночный микротитрационный планшет, который может быть использован для анализа одновременно нескольких проб. Количество и типы твердых носителей являются многочисленными, и нет необходимости их подробного описания здесь.
Тройной комплекс - комплекс 150 кДа, состоящий из IGF, IGFBP и ALS.
Лечение (обработка), предназначенное для влияния на статус гормонов роста, включает в себя любое медицинское воздействие, которое предназначено для влияния на GH- или IGF-каскады. Такие воздействия могут включать в себя воздействия такими агентами, как GH, GHBP, IGF, IGFBP, ALS, IGFBP-комплекс, рецепторы GH, рецепторы IGF, антитела или ингибиторы любого из предыдущих компонентов, антагонисты рецепторов для GH или IGF или любое лекарственное средство, которое действует, модулируя статус гормонов роста индивидуума, в том числе тамоксифен, соматостатин, аналоги соматостатина, антагонисты GH, антагонист IGF, стимуляторы IGFBP, ретиноиды, TGF-бета, и аналоги витамина D и т. п. Термин "индивидуумы" включает в себя как человека, так и животных, таких как свиньи, крупный рогатый скот, овцы, козы, лошади, домашняя птица, кошки, собаки, рыбы и т.д.
Недавно авторы изобретения описали неконкурентные способы ELISA для определения IGF-I [37], IGF-II [38], IGFBP-1 [19], IGFBP-3 [39] и ALS [15] в сыворотке и других физиологических жидкостях. Клиническая оценка связанных с GH нарушений и мониторинг лечебных схем (программ лечения) включали в себя увеличенное использование биохимических определений уровней в кровотоке IGF-I, IGFBP-3 и ALS [21-26, 36]. Поскольку почти все IGF-I и IGFBP-3 в крови [4-12] и около 50%, ALS [13, 15] в норме ограничиваются содержанием в GH-зависимом тройном комплексе 150 кДа, прямое определение этого комплекса может иметь значительные преимущества. В данном изобретении авторы сообщают разработку высокоспецифических и быстрых способов ELISA для прямого измерения высокомолекулярных IGF-связывающих белковых комплексов.
Говоря в общем, способ количественного измерения IGFBP-комплекса включает в себя захват этого комплекса первым захватывающим антителом, отмывание всех несвязавшихся компонентов и детектирование оставшегося комплекса вторым детектирующим антителом. Возможные схемы иммуноанализа основываются на многочисленных комбинациях "захватывающего и детектирующего антитела" и могут включать в себя комбинации (а) антител анти-IGFBP-3, (b) антител анти-IGF-I, (с) антител анти-ALS и (d) антител против комплекса при условии, что каждая пара (или триплет) антител взаимодействует с отдельными эпитопами и не препятствует образованию комплекса.
Более конкретно, авторы изобретения представили изобретение примером, в котором захватывающим антителом является анти-IGFBP-3 и детектирование осуществляется антителом, направленным либо против ALS, либо против IGF. Подход авторов исключает необходимость процедур предобработки пробы комплекса, делает возможным объединенное определение компонентов IGFBP-комплекса в одном анализе и потенциально обеспечивает лучшую клиническую информацию.
В зависимости от схемы анализа тройной, а также любой двойной комплекс может быть количественно определен при одновременном исключении вкладов, обусловленных фрагментами и менее прочно связанными или диссоциируемыми разновидностями, в это определение.
Две пары антител специфического захвата-детектирования приводятся в качестве примеров ниже, и эти тесты названы ELISA-1 и ELISA-2. ELISA-1 включает в себя комбинацию антител против IGFBP-3/IGF-I и является специфическим как для тройных комплексов IGFBP-3/IGF-I/ALS, так и для любого двойного комплекса IGFBP-3-/IGF-I, который мог бы потенциально присутствовать. Специфичность этого анализа для таких двойных комплексов устанавливали анализом проб до, и после нейтрализации кислотой, которая, как известно, функционально инактивирует ALS. ELISA-1 измерял одинаковые уровни IGFBP-3-комплекса в необработанных и обработанных кислотой пробах.
ELISA-2 включает в себя комбинацию антител против IGFBP-3/ALS и определяет количественно тройные комплексы на основе как IGF-I, так и IGF-II и бинарный комплекс IGFBP-3/ALS. Как ожидалось, ELISA-2 не детектировал никакой иммунореактивности в обработанных кислотой пробах.
Можно также модифицировать этот анализ для детектирования других молекулярных частиц. Например, можно детектировать общий IGFBP (в комплексе или свободный) вариацией ELISA-1 и ELISA-2. Сначала весь IGFBP захватывается антителом анти-IGFBP. Затем захваченный IGFBP насыщают экзогенным лигандом (IGF и/или ALS в зависимости от используемого детектирующего антитела) перед детектированием детектирующим антителом. В этой схеме анализа как находящийся в комплексе IGFBP, так и свободный IGFBP будут захватываться захватывающим антителом, доводиться до полной ассоциации с экзогенным лигандом и детектироваться детектирующим антителом анти-IGF или анти-ALS.
Далее, хотя авторы привели примеры анализов, в которых захватывающим антителом является антитело анти-IGFBP, можно также использовать антитела анти-IGF, анти-ALS или антитело против комплекса в качестве захватывающего антитела. Очевидно, что различные схемы анализа могут быть оптимизированы специалистом в данной области, как описано здесь.
Данное изобретение включает в себя набор для определения IGFBP-комплекса. Набор содержит захватывающее антитело и детектирующее антитело при условии, что захватывающее и детектирующее антитела не связываются с перекрывающимися эпитопами и не препятствуют образованию IGFBP-комплекса. Захватывающее антитело может быть связано с твердым носителем, а детектирующее антитело обычно является меченным тем или иным образом.
Комбинации захватывающего и детектирующего антител могут быть выбраны из группы, состоящей из В1, В2, В3, В4, В5, В6, В7, В8, В9, В10, рВ11, I-1, I-2, pI-3, pA-1 и рА-2. Однако предпочтительно парой антител является В3 и I-1. В дополнительном предпочтительном варианте парой антител является В2 и рА-1.
Набор может содержать другие полезные реагенты, такие как средство для детектирования метки, буфер для связывания, промывной буфер, буфер для детектирования и контрольный стандарт и т.п.
Данное изобретение описывает также способ детектирования IGFBP-комплекса в жидкости тела. Способ состоит в a) контактировании твердой фазы, связанной с захватывающим антителом, с жидкостью тела для захвата IGFBP-комплекса; b) промывание этой твердой фазы первым промывочным буфером; c) контактирование твердой фазы с детектирующим антителом, связанным с меткой; d) промывание этой твердой фазы вторым промывочным буфером и е) детектирование метки, остающейся с твердой фазой после этой стадии промывания.
Этот способ может быть выгодно использован для определения статуса GH индивидуума, в том числе, но не только, человека, коров, свиней, представителей семейств лошадиных, собачьих, кошачьих или овечьих. Другие специфические применения включают в себя, но не ограничиваются ими: а) мониторинг эффекта способов лечений, предназначенных для воздействия на GH-статус или IGF-статус индивидуума; b) мониторинг клинического статуса и/или лечения индивидуумов с различными заболеваниями, на которые влияет GH-статус, в том числе гипогликемией, диабетом, состояниями, связанными с недостаточным питанием, некоторыми раковыми заболеваниями, такими как рак молочной железы и рак предстательной железы, иммунодефицитными состояниями, задержкой роста плода, гигантизмом, акромегалией, гиперпитуитаризмом, гипофизарной карликовостью, недостаточностью гормона роста, избытком гормона роста, дефектом рецептора гормона роста, тиомегалией и определенными заболеваниями центральной нервной системы (ЦНС), такими как болезнь Альцгеймера; с) определение подверженности индивидуума патологическим состояниям, связанным с ростом, причем подверженность характеризуется отклоняющимся от нормы или повышенным статусом GH; или d) различные ветеринарные применения, такие как мониторинг состояния питания, репродуктивного здоровья, показателей в состязаниях или других показателей активности, детектирование улучшенного племенного поголовья животных и т.п.
Фиг.1. Карта эпитопов IGFBP-3-комплекса.
Были определены четыре антигенных района. Перекрывающиеся кружки указывают на отсутствие образования сэндвичей, касающиеся кружки указывают на препятствие образованию сэндвичей и отдельные кружки указывают на независимые эпитопы.
Фиг. 2. Ингибирование связывания антител с IGFBP-3 посредством IGF-I и IGF-II.
Показано связывание моноклональных и поликлональных антител анти-IGFBP-3 с IGFBP-3 до и после прединкубирования с 125IGF-I или 125IGF-II. Антитело анти-PSA использовали в качестве отрицательного контроля. Величины являются средним из трех измерений.
Фиг.3. Калибровочная кривая для ELISA IGFBP-3-комплекса.
Показаны кусочно-линейные графики типичных калибровочных кривых ELISA-1 и ELISA-2.
Фиг.4. Стабильность IGFBP-3-комплекса, анализируемого ELISA-1.
Повторные аликвоты трех различных проб (S1-S3), хранящихся при различных температурах и анализированных, как указано. Показаны изменения в процентах от величины дня 0.
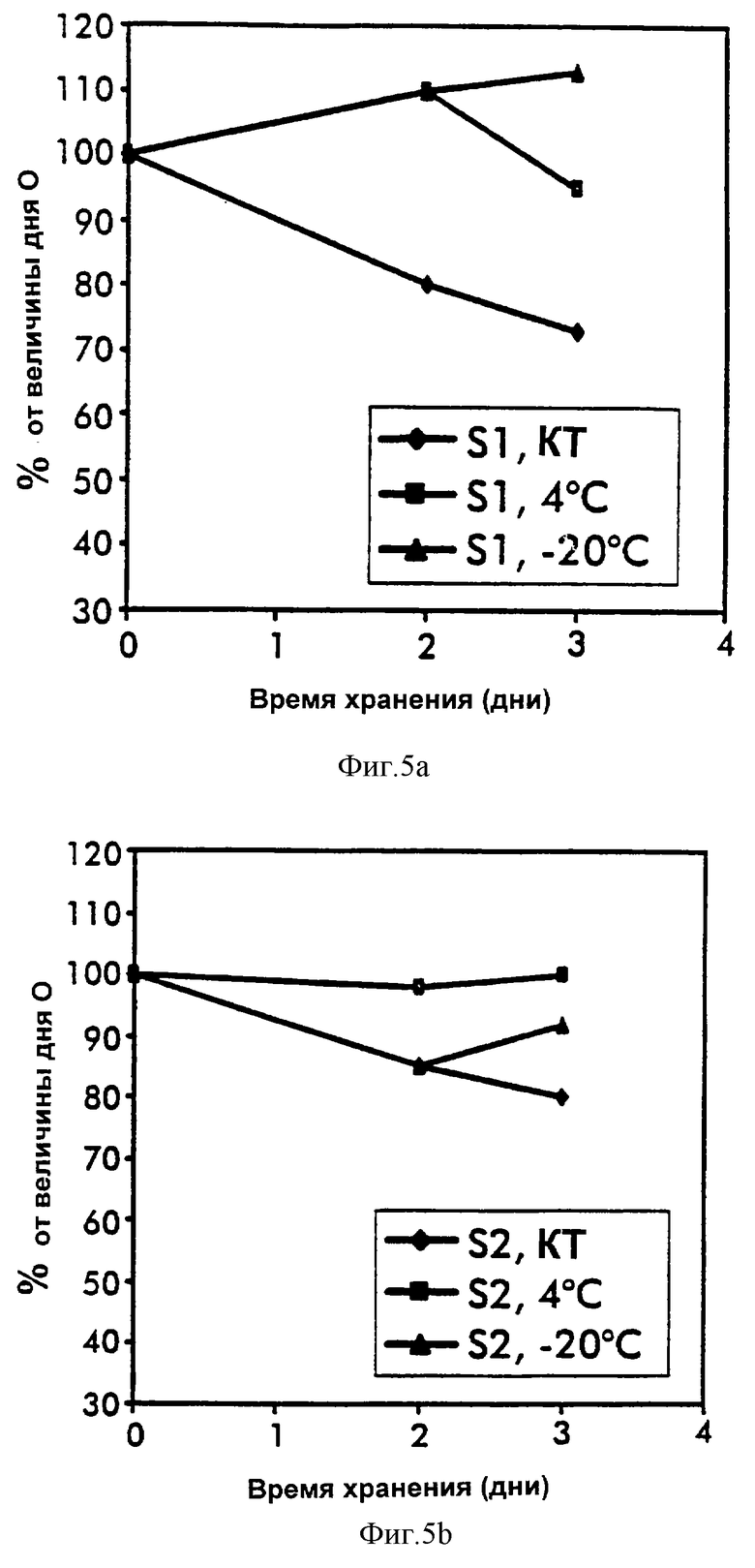
Фиг.5. Стабильность IGFBP-3-комплекса, анализируемого ELISA-2.
Повторные аликвоты трех различных проб (S1-S3), хранящихся при различных температурах и анализированных, как указано. Показаны изменения в процентах от величины дня 0.
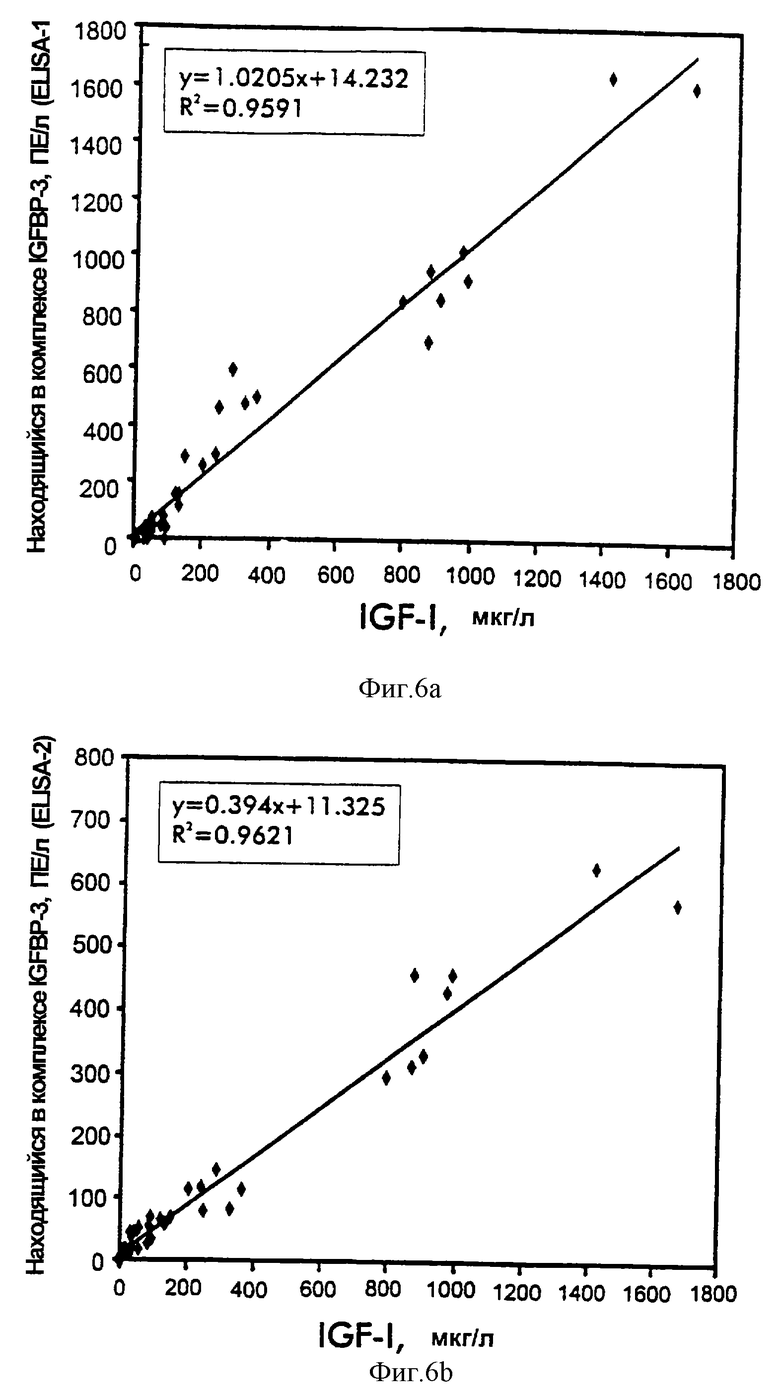
Фиг.6. Сравнение IGFBP-3-комплекса с IGF-I.
Пробы обработанной ЭДТА плазмы из субъектов с не подвергавшимся лечению GHRD (n=11) и нормальных субъектов соответствующего возраста (n=16) и пробы сыворотки из взрослых с акромегалией (n=8) или с GHD (n=5) анализировали на IGF-I и IGFBP-3-комплекс при помощи ELISA-1 и ELISA-2. Величины являются средними из двух повторностей измерений.
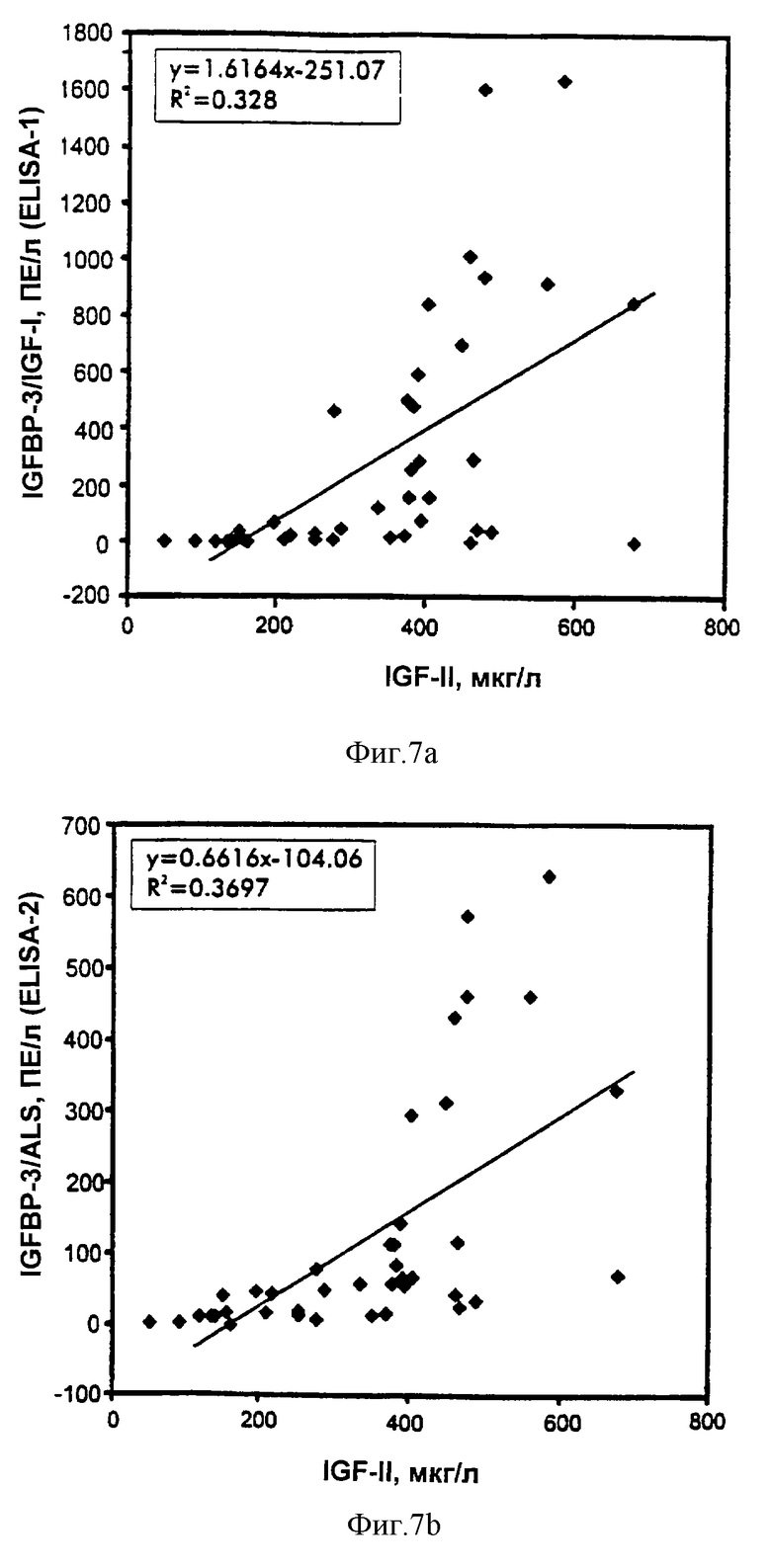
Фиг.7. Сравнение IGFBP-3-комплекса с IGF-II.
Пробы обработанной ЭДТА плазмы из субъектов с не подвергавшимся лечению GHRD (n=11) и нормальных субъектов соответствующего возраста (n=16) и пробы сыворотки из взрослых с акромегалией (n=8) или с GHD (n=5) анализировали на IGF-II и IGFBP-3-комплекс при помощи ELISA-1 и ELISA-2. Величины являются средними из двух повторностей измерений.
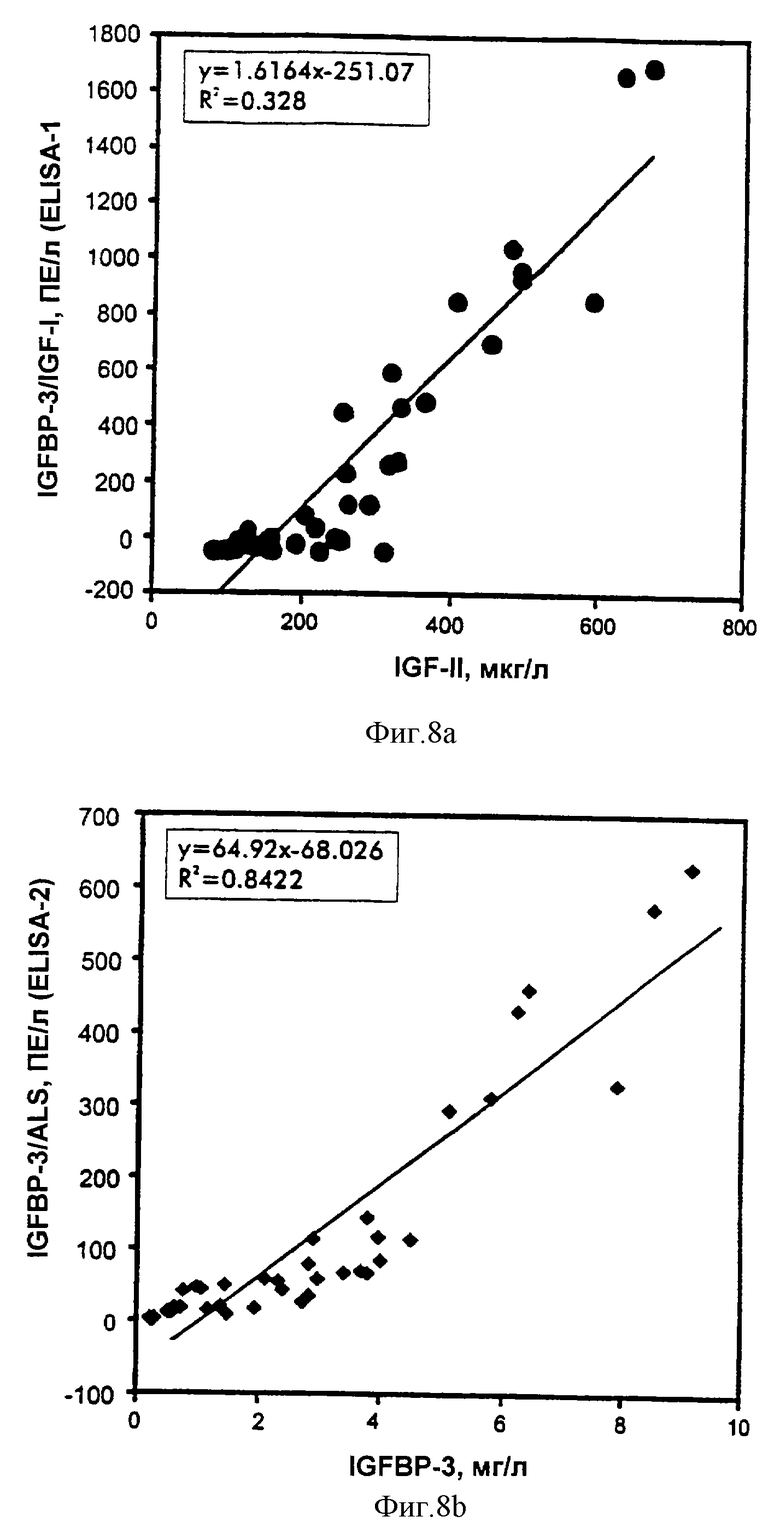
Фиг.8. Сравнение IGFBP-3-комплекса с IGFBP-3.
Пробы обработанной ЭДТА плазмы из субъектов с не подвергавшимся лечению GHRD (n=11) и нормальных субъектов соответствующего возраста (n=16) и пробы сыворотки из взрослых с акромегалией (n=8) или с GHD (n=5) анализировали на IGFBP-3 и IGFBP-3-комплекс при помощи ELISA-1 и ELISA-2. Величины являются средними из двух повторностей измерений.
Фиг.9. Сравнение IGFBP-3-комплекса с общим ALS.
Пробы обработанной ЭДТА плазмы из субъектов с не подвергавшимся лечению GHRD (n=11) и нормальных субъектов соответствующего возраста (n=16) и пробы сыворотки из взрослых с акромегалией (n=8) или с GHD (n=5) анализировали на общий ALS и IGFBP-3-комплекс при помощи ELISA-1 и ELISA-2. Величины являются средними из двух повторностей измерений.
Фиг.10. Уровни компонентов IGF-оси от возраста.
Уровни IGF-I, IGF-II, IGFBP-3, ALS- и IGFBP-3-комплексов, определенные при помощи ELISA-1 и ELISA-2 измеряли в пробах, обработанных ЭДТА плазмы, полученных у 16 нормальных субъектов. Для каждого аналита значения каждой пробы как процент соответствующих средних значений отражают на графике от возраста.
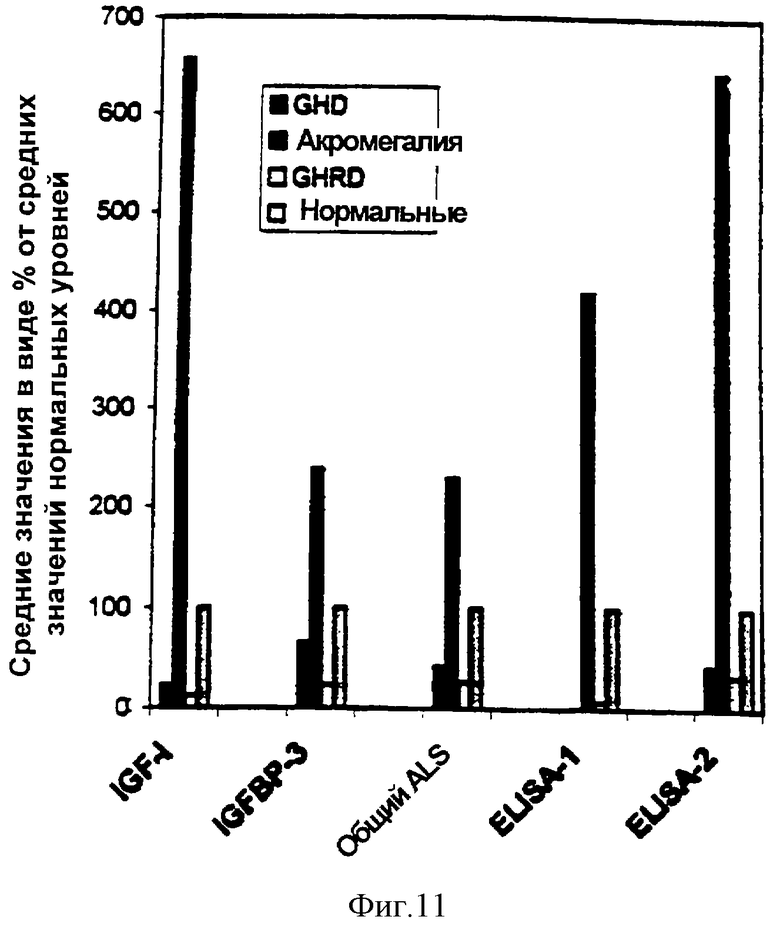
Фиг. 11. Уровни компонентов IGF-оси (компонентов суперсемейства IGF) в виде процента от среднего значения нормальных величин.
Пробы обработанной ЭДТА плазмы из субъектов с не подвергавшимся лечению GHRD (n=11) и нормальных субъектов соответствующего возраста (n=16) и пробы сыворотки из взрослых с акромегалией (n=8) или с GHD (n=5) анализировали на IGF-I, IGF-II, IGFBP-3, общий ALS и IGFBP-3-комплекс при помощи ELISA-1 и ELISA-2. Приведены средние концентрации каждого аналита, измеренные в каждой группе проб, в виде процента от соответствующего среднего значения нормальных величин (рассматриваемых как 100%).
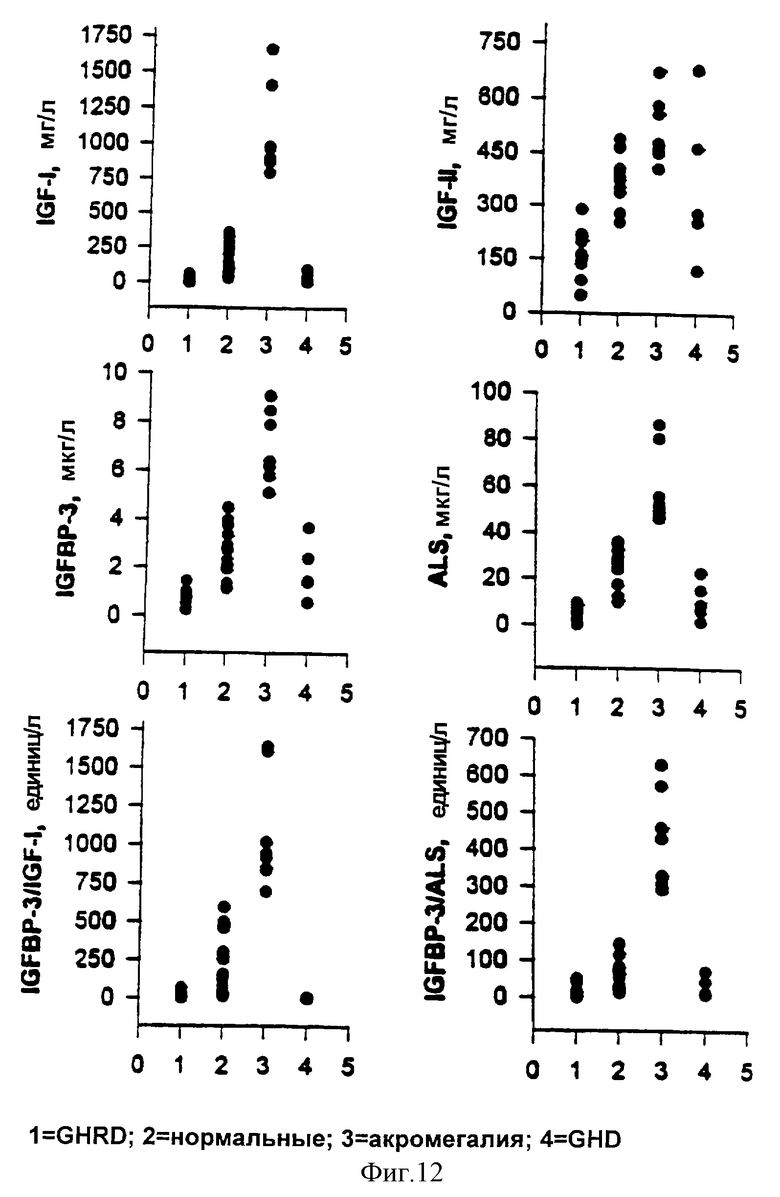
Фиг.12. График распределения компонентов IGF-оси и IGFBP-3-комплекса.
Пробы обработанной ЭДТА плазмы из субъектов с не подвергавшимся лечению GHRD (n=11) и нормальных субъектов соответствующего возраста (n=16) и пробы сыворотки из взрослых с акромегалией (n=8) или с GHD (n=5) анализировали на IGF-I, IGF-II, IGFBP-3, общий ALS и IGFBP-3-комплекс при помощи ELISA-1 и ELISA-2. Величины являются средними из двух повторностей измерений.
Анализ захвата-детектирования IGFBP-комплекса иллюстрируется здесь в примерах двух различных форматов. Однако описаны и картированы множество антител и различные комбинации пар или триплетов антител могут быть использованы в основном формате анализа захвата-детектирования. В следующих примерах описаны эти анализы, картирование эпитопов и антитела данного изобретения.
ПРИМЕР 1. Материалы
А. Образцы
Пробы обработанной ЭДТА плазмы из субъектов с не подвергавшимся лечению GHRD (5 мужчин в возрасте 1-32 лет, средний возраст 17±14,5, и 6 женщин в возрасте 2,3-65 лет, средний возраст 29,9±25) и нормальных контрольных субъектов соответствующего возраста (8 мужчин в возрасте 1-27 лет, средний возраст 13,3±10,4, и 8 женщин в возрасте 2-67 лет, средний возраст 36,6±23,7 лет) были получены от доктора Jaime Guevara-Aguirre, Институт эндокринологии, Метаболизм и репродукция, Кито, Эквадор. Эквадорских пациентов с GHRD выбрали для этого исследования, так как они имели чрезвычайно низкие уровни IGF-I и IGFBP-3 [20]. Протокол взятия проб был одобрен Комитетом по этике Института эндокринологии, Метаболизм и репродукция (Кито, Эквадор) в соответствии с законами и инструкциями Соединенных Штатов и Эквадора. Все субъекты и/или их опекуны подписали испанские варианты форм об одобренном информированном согласии. Эти пробы хранили при -70oС до использования.
Пробы сыворотки из давших согласие взрослых с акромегалией (n=8) и недостаточностью GH (n=5) были любезно предоставлены доктором John Miell, Department of Medicine, Kings College School of Medicine, London, England. Пробы свежей сыворотки (n=42), взятые методом случайного отбора, получали из Клинических лабораторий в Канаде. Эти пробы были остатками от рутинных клинических тест-проб, взятых из популяции взрослых больных. При взятии пробам крови давали свертываться, разделиться и после клинического испытания остатки, хранящиеся при 4oС, использовали для этих исследований в течение 48 часов.
В. Реагенты
Рекомбинантные IGF-I и IGF-II человека получали из GROPEP, PTY, LTD (Adelaide, Australia) и рекомбинантный негликозилированный IGFBP-3- получали из CELTRIX PHARMACEUTICAL, INC. (Santa Clara, CA). Рекомбинантные IGFBP-2, и IGFBP-4 - IGFBP-6 приобретали из AUSTRAL BIOLOGICALS (San Roman, CA). IGFBP-1 очищали из амниотической жидкости человека и калибровали относительно чистого рекомбинантного IGFBP-1 человека, полученного из DSL (Webster, ТХ). Другие материалы и химикалии получали, как описано [40, 41]. Очищенный ALS получали из DSL (Webster, ТХ), как описано ранее [15]. 125I-IGFBP-3 (5x106 имп/мин/мл 0,1 М NaPO4, pH 7,4) получали из DSL (Webster, ТХ). HRP-меченый стрептавидин был приобретен у AMERSHAM INTERNATIONAL (Buckinghamshire, England).
Пероксидазу хрена (HRP) получали из SCRIPPS LABORATORIES (San Diego, CA). Субстратную систему тетраметилбензидинпероксидазы (ТМВ/Н202) получали из KIRKEGAARD AND PERRY LABORATORIES, INC. (Gaithersburg, MD). Сульфосукцинимидил-6-(биотинамидо)гексаноат (NHS-LC-Biotin) получали из PIERCE CHEMICAL CO. (Rockford, IL). Все другие химические реагенты были самого высшего качества и были получены из SIGMA CHEMICAL (St. Louis, МО) или AMRESCO, INC. (Solon, ОН). Полоски и рамки для микротитрования были продуктами COSTAR (Cambridge, МА).
С. Буферы для анализов (тест-буферы)
Анализы IGFBP-3/IGF-I (ELISA-1) и IGFBP-3/ALS (ELISA-2), описанные ниже, основаны на одинаковых принципах и идентичных компонентах, за исключением комбинации антител и выбора тест-буфера. Буферы были следующими:
Тест-буфер для ELISA-1 - 0,05 М Трис-малеат, рН 7,0, 9 г/л NaCl, 5 г/л БСА, 0,001 М Na-ЭДТА, 0,5 мл/л Твина 20, 0,1 г/л тимеросала.
Тест-буфер для ELISA-2 - 0,05 М Трис-малеат, рН 7,0, 0,9 г/л NaCl, 20 г/л БСА, 0,5 г/л бычьего гамма-глобулина, 25 мл/л сыворотки нормальной козы, 100 мг/л ФМСФ, 0,5 мл/л Твина 20, 0,1 г/л тимеросала.
Буфер для нулевого стандартного матрикса - 0,05 М фосфат натрия, рН 7,4, 9 г/л NaCl, 1 г/л БСА, 0,005 М Na-ЭДТА, 2,5 мл/л трасилола, 0,1 г/л тимеросала.
Буфер для концентрата конъюгата антитело-HRP - 0,02 М фосфат натия, рН 7,0, 9 г/л NaCl, 1 г/л CaCl2, 5 г/л БСА, 0,1 г/л тимеросала.
Стоп-раствор - 0,2 М серная кислота в деионизованной воде.
Буфер для покрытия - 50 мМ Трис, 0,1% азид натрия, рН 7,8.
Промывной буфер - 0,05% Твин 20, 5 мМ Трис, рН 7,4.
Буфер для блокирования - 50 мМ Трис, 1% БСА, 0,025% азид натрия, рН 7,4.
D. Антитела
Способы получения как моноклональных, так и поликлональных антител хорошо разработаны в настоящее время [49]. Для этого изобретения моноклональные антитела получали в мышах, а поликлональные антитела индуцировали в козах. Однако эти антитела могли быть индуцированы в различных видах, в том числе, но не только, в мышах, крысе, кролике, козе, овце, осле, лошади и т.д. Анти-IGF и анти-IGFВР-3 индуцировали против негликозилированных рекомбинантных IGF-I, IGF-II или IGFBP-3 человека (см. таблицу 1 в отношении названий и характеристик антител). Антитела анти-ALS индуцировали против синтетических уникальных N- и С-концевых районов ALS человека [15]. ALS-пептиды конъюгировали с овальбумином с использованием глутарового альдегида, смешивали с полным адъювантом Фрейнда и инъецировали в коз (0,1 мг/инъекция) с использованием схемы бустинга и кровоизвлечения один раз в месяц. ALS-пептиды получали с использованием пептидного синтезатора Модели 430А APPLIED BIOSYSTEMS (Foster City, CA) (SYNPEP CORP, Dublin, CA) и очищали жидкостной хроматографией высокого разрешения с обращенной фазой (ВЖХ). Антигены IGF-I, IGF-II и IGFBP-3 инъецировали, как описано выше, без модификации.
Для получения моноклональных антител спленоциты из подходящим образом иммунизированных мышей BALB/c сливали с клетками миеломы по способу с применением полиэтиленгликоля. Жизнеспособные гибридомы отбирали, подвергали скринингу и размножали. Супернатанты из выбранных клонов подвергали скринингу против соответствующего аналита при помощи способа ELISA. Позитивные гибридомы клонировали конечным разведением и клоны, секретирующие специфические антитела, использовали для получения асцитов. Все моноклональные антитела аффинно очищали на протеин А-колонках и соответствующим образом подвергали скринингу на специфичность с использованием хорошо установленных способов вестерн-блоттинга [50].
Поликлональные антитела сначала очищали аффинной хроматографией через колонки с гелями, содержащими соответствующий иммобилизованный иммуноген, и фракции антител дополнительно очищали хроматографией через протеин А-колонки. Как описано ниже, специфичность этих антител доказывали дополнительно посредством исследований с иммуноанализом перекрестной реактивности. Все антитела были получены, охарактеризованы и очищены DSL, INC (Webster, TX). Поликлональные антитела анти-IGF-I, анти-IGF-II, анти-ALS и анти-IGFBP-3 являются высокоспецифическими в отношении соответствующего аналита [15, 37, 38, 39] и они были использованы в соответствующих ELISA, производимых DSL INC. (Webster, TX). Моноклональные антитела анти-IGFВР-3 также являются высокоспецифическими в отношении IGFBP-3 и в настоящее время проходят оценку для разработки дополнительных новых ELISA для IGFBP-3.
ПРИМЕР 2. Способы: процедуры препаративного анализа
Покрытие антителами лунок микротитрационных планшетов выполняли при концентрации 2,5-30 мг/л, если нет иных указаний, с использованием ранее опубликованных способов [40]. Вкратце, 0,1-0,2 мл раствора антитела (5-10 мг/л) добавляли в каждую микротитрационную лунку и давали инкубироваться в течение ночи при комнатной температуре. Затем лунки промывали один раз промывным раствором и добавляли 0,2 мл/лунка блокирующего раствора и давали инкубироваться в течение 1 часа, как описано выше. Лунки промывали один раз перед использованием или хранили до 1 недели в блокирующем буфере при 4oС.
Покрытие IGFBP-3 лунок микротитрационных планшетов выполняли, как описано выше, за исключением того, что IGFBP-3 наносили при концентрации 0,25-2 мг/л.
Связывание антител с пероксидазой хрена (HRP) выполняли, как описано ранее [4]. Реакция связывания включала в себя первоначальную активацию фермента сульфосукцинимидил-4-(N-малеидометил)циклогексан-1-карбоксилатом (SMCC) и его последующую конъюгацию с активированным 2-иминотиоланом антителом. Исходный раствор HRP-конъюгированного антитела хранили при 4oС в темноте и подходящим образом разводили (по меньшей мере в 1000 раз) в подходящем тест-буфере перед использованием.
Связывание антител с биотином выполняли, как описано ранее [42]. Биотинилирование проводили при приблизительно 150-кратном молярном избытке NHS-LC-биотина, добавленного к 0,5 мг/мл раствора антитела. Неконъюгированный биотин удаляли диализом при 4oС в течение 24 часов против нескольких смен 0,1 молярного бикарбоната натрия, рН 8,3, содержащего 9 г/л NaCl и 0,25 г/л азида натрия. Исходный раствор конъюгата антитело-биотин хранили при 4oС и подходящим образом разводили в 0,05 молярном NaPO4, рН 7,2, содержащем 9 г/л NaCl, 2 г/л БСА и 1,0 мл/л антибактериального/противогрибкового консерванта Proclin 300 (Sigma) перед использованием.
Пул свежих проб сывороток брали за 100 произвольных единиц на литр (ПЕ/л) комплекса IGF-связывающего белка (IGFBP-3-комплекса) и использовали для стандартизации ELISA-1 и ELISA-2. Стандарты готовили подходящим разведением этого пула сывороток в буфере для нулевого стандартного матрикса для получения величин ссылочных стандартов 0,78, 1,56, 6,25, 25 и 50 ПЕ/л комплекса IGFBP-3-/IGF-I и 3,13, 6,25, 12,5, 25 и 50 ПЕ/л комплекса IGFBP-3/ALS для применения в ELISA-1 и ELISA-2 соответственно. Эти стандарты были стабильными в течение периода до 24 часов при 4oС и более 2 месяцев при -20oС или при более низкой температуре. Используемые пробы качественного контроля были также подходящим образом разведенными пулами сывороток. Номинальные концентрации контрольных проб устанавливали анализом их в ELISA-1 и ELISA-2 для IGFBP-3-комплекса.
ПРИМЕР 3. Способы: картирование эпитопов IGFBP-3
А. Одновременное связывание антител анти-IGFBP
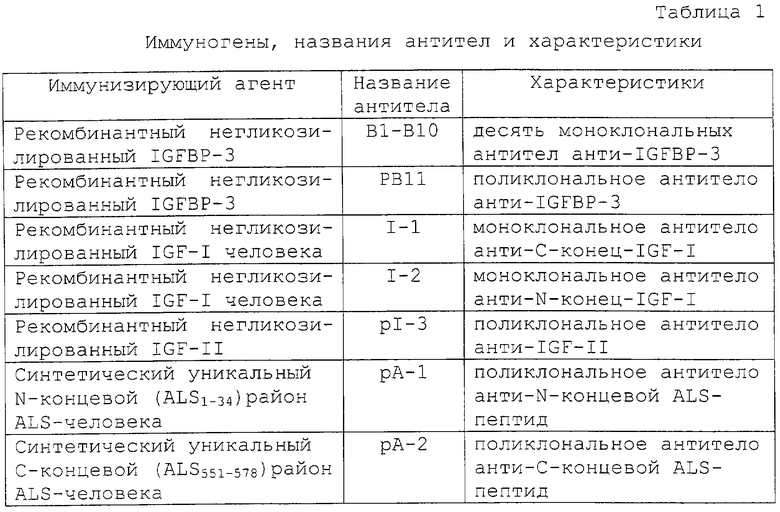
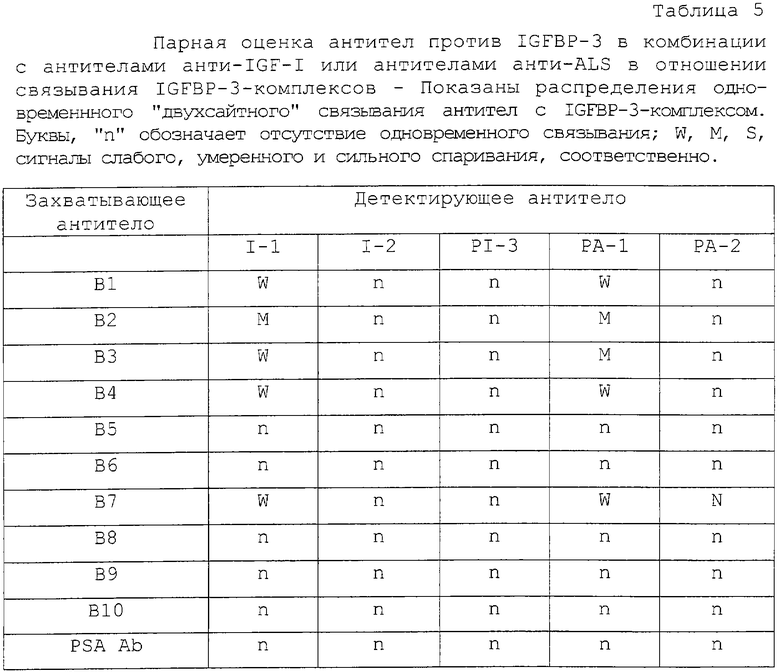
Антитела анти-IGFВР-3 (В1-В10) оценивали на одновременное связывание (спаривание) с IGFBP-3. Вкратце, каждое антитело иммобилизовали на лунки микротитрационных планшетов при 500 нг/100 мкл на лунку и подвергали взаимодействию с IGFBP-3 при 0,0-100 мкг/л (25 мкл/лунка стандартов плюс 100 мкл/лунка тест-буфера). После 1 часа инкубирования со встряхиванием (500-700 об/мин) при комнатной температуре (RT) лунки промывали четыре раза и подвергали взаимодействию с каждым из остальных HRP-меченых антител анти-IGFВР-3, описанных выше, в течение 30 минут. Исходные HRP-антитела разводили приблизительно в 10000 раз в тест-буфере и использовали при 100 мкл/лунка (около 5 нг антитела). После промывания реакцию проявляли 10-минутным инкубированием этих лунок с раствором субстрата ТМВ/Н2О2 (100 мкл на лунку) и добавлением стоп-раствора, как описано ниже. Максимальное увеличение в оптической плотности (OD) ≤ 3Х фон (сигнал нулевого стандарта), между 3Х фона и 1 OD, между 1 OD и ≤2 OD и ≥2 OD было классифицировано как указание на отсутствие одновременного связывания (спаривания) с IGFBP-3, слабое спаривание, умеренное спаривание и сильное спаривание соответственно.
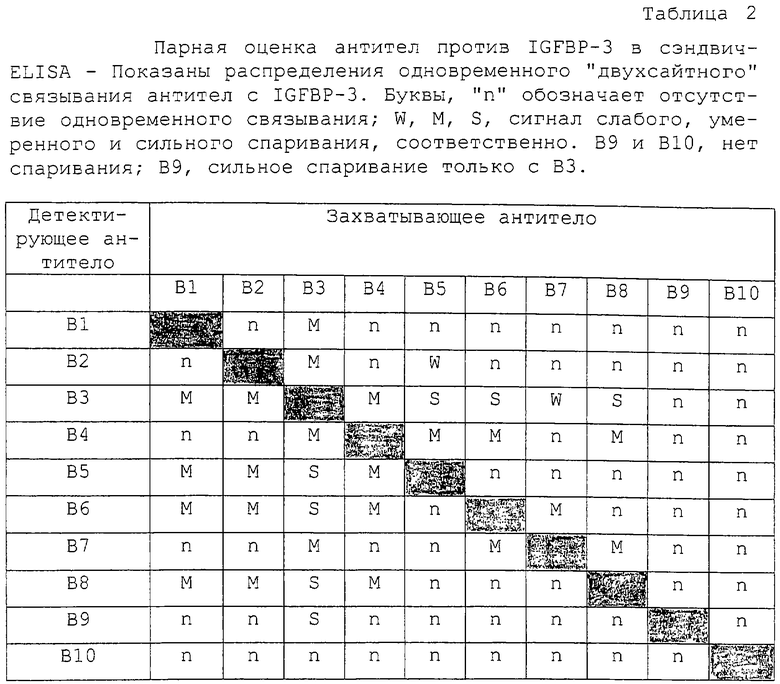
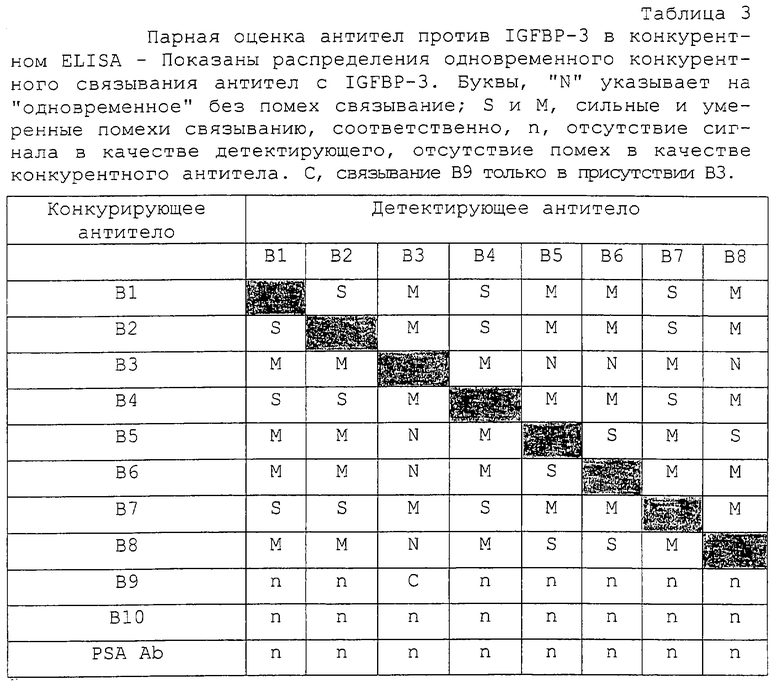
В. Конкурентное связывание антител анти-IGFBP
Антитела анти-IGFВР-3 (В1-В10) оценивали также на конкурентное связывание с IGFBP-3. Вкратце, каждое биотинилированное антитело в заранее заданном разведении смешивали с увеличивающимися концентрациями каждого из остальных немеченых антител (0-50 мкг/мл) и добавляли (50 мкл антитела плюс 100 мкл тест-буфера) в трех повторностях в лунки, предварительно покрытые IGFBP-3 (около 75 нг на лунку). После 2 часов инкубирования со встряхиванием лунки промывали и проявляли 30-минутной реакцией с HRP-меченым стрептавидином (100 мкл на лунку при 2000-кратном разведении в тест-буфере) с последующим добавлением ТМВ/H2O2 и добавлением стоп-раствора. Уменьшение OD ≤20%, 20-60% и 60% были классифицированы как указывающие на препятствующее, умеренно препятствующее и сильно препятствующее связывание пары антител с IGFBP-3.
С. Действие IGF-занятия на связывание анти-IGFBP
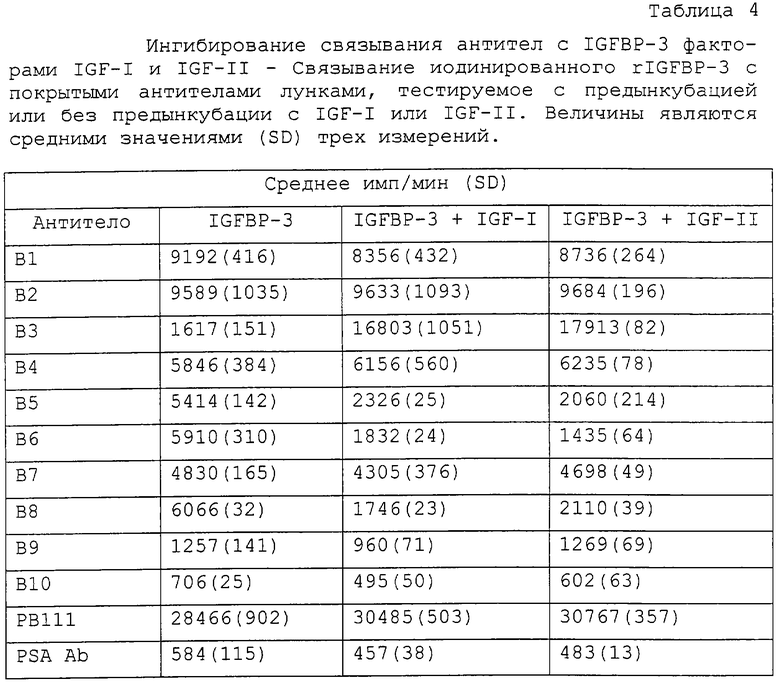
Оценивали также действие IGF-занятия IGFBP-3 на связывание антител анти-IGFВР-3 (В1-В10). Вкратце, 125I-IGFBP-3 (1•106 имп/мин/мл) смешивали с 0,0-0,75 мкг/мл IGF-I или IGF-II (в тест-буфере для ELISA-1), инкубировали в течение 2 часов при комнатной температуре и добавляли 100 мкл на лунку в трех повторностях к покрытым антителом анти-IGFВР-3 лункам. После 2 часов инкубирования при комнатной температуре лунки промывали три раза дН2О и считали на связанную радиоактивность в гамма-счетчике Packard RIASTAR из PACKARD CANADA (Mississauga, Ontario). Антитело связывается при сайте связывания или вблизи сайта связывания IGF, если его IGFBP-3-связывающий сигнал (имп/мин) уменьшается по меньшей мере на 30% в ответ на предынкубирование IGFBP-3 с IGF.
D. Одновременное связывание антител анти-IGFBP и анти-IGF или анти-ALS
Нативный сывороточный IGFBP-3-комплекс оценивали также на одновременное связывание с антителами анти-IGFВР-3 (В1-В10) в комбинации с антителами анти-IGF-I (I-1, I-2), антителами анти-IGF-II (рI-3) или антителами анти-ALS (рА-1, рА-2) в парном сэндвич-ELISA со "смешанными антителами". Вкратце, пул сыворотки человека получали смешиванием аликвот из 15 различных проб сыворотки. Микротитрационные полоски, покрытые каждым из антител, инкубировали, в четырех повторностях, с 50 мкл на лунку пула сыворотки или буфера для стандартного матрикса и 100 мкл на лунку соответствующего тест-буфера в течение 2 часов при комнатной температуре, как описано выше. После промывания каждую серию из четырех повторностей лунок, обработанных пулом сыворотки/буфером для нулевого стандартного матрикса, инкубировали затем со 100 мкл на лунку каждого из HRP-меченых антител анти-IGF-I, анти-IGF-II или анти-ALS в течение 30 минут при комнатной температуре. После промывания реакцию проявляли 10-минутным инкубированием с субстратом ТМВ/Н2О2 и добавлением стоп-раствора. Увеличение OD ≤3Х фон (сигнал нулевого стандарта), между 3Х фона и 1 OD и между 1 OD и ≥ 2 OD указывает на отсутствие связывания, умеренное связывание или сильное одновременное связывание двух антител с находящимся в комплексе IGFBP-3 соответственно. Комбинации смеси антител, обнаруживающие самый сильный сигнал, были отобраны для разработки ELISA для IGFBP-3-комплекса, описанного ниже.
ПРИМЕР 4. Карта эпитопов находящегося в комплексе IGFBP-3
Подробная информация об эпитопах, узнаваемых моноклональными антителами против IGFBP-3, была получена оценкой всех возможных двухсайтных (захват-детектирование) комбинаций в парном сэндвич-ELISA. Затем пространственное распределение эпитопов, узнаваемых каждым антителом, относительно друг друга оценивали в конкурентном парном ELISA, который оценивает связывание рассматриваемого антитела с IGFBP-3 в присутствии избыточных количеств каждого из остальных антител. Последний способ ELISA обеспечивает информацию о том, является ли эпитоп, узнаваемый одним антителом, достаточно отличающимся, чтобы позволить немешающее (независимое) связывание второго антитела, или был ли этот узнаваемый эпитоп полностью или частично перекрывающимся, приводя к помехам при связывании.
В третьей серии экспериментов, оценивали помехи связывания антител с IGFBP-3 факторами IGF-I и IGF-II. Это индентифицировало антигенные домены в сайте связывания IGF или вблизи него и оценивалось мониторингом связывания твердофазных (иммобилизованных) антител с IGFBP-3 до и после инкубирования с радиоактивно мечеными IGF. Наконец, способность IGFBP-3-антител связываться с сывороточным IGFBP-3-комплексом в парной комбинации с антителами анти-IGF-I, анти-IGF-II и анти-ALS оценивали в сэндвич-ELISA со "смешанными антителами". В сходных экспериментах исследовали также одновременное связывание находящегося в комплексе IGFBP-3 с антителами анти-IGF, спаренными с антителами анти-ALS.
Как показано в таблице 2, оценка 10 антител против IGFBP-3 (B1-B10) в сэндвич-ELISA идентифицировала 31 из 100 возможных комбинаций. Распределения связывания, по-видимому, "скапливаются" (образует кластер) в четырех антигенных районах на основе комбинаций реагирующих антител и силы генерируемого сигнала спаривания. Эти антитела группируются следующим образом: Группа I включала в себя В5, В6 и В8; Группа II включала в себя В1, В2, В4 и В7; Группа III включала в себя В3 и Группа IV включала в себя В9. Антитела в одной и той же группе не связывались одновременно с IGFBP-3 в сэндвич-ELISA, но демонстрировали спаривание, от слабого до сильного, с антителами в других группах.
Оказалось, что спаривание антител в группе I и группе II зависит в некоторой степени, от того, использовали ли конкретное антитело для покрытия или для детектирования. Это обусловлено предположительно субоптимальными концентрациями антител и/или конформационными изменениями эпитопов, индуцируемыми связыванием первого антитела с IGFBP-3. Самые сильные сигналы двухсайтного связывания генерировались между антителами Группы I и Группы III (В3). В9 и В10 были неспособны образовать сэндвич друг с другом или с остальными антителами. В9 демонстрировал сильное связывание с IGFBP-3 только в том случае, когда его использовали в качестве детектирующего антитела в комбинации с В3.
Более ясная картина возникала при оценке антител на связывание с твердофазным IGFBP-3 в парном конкурентном ELISA (таблица 3). В этих экспериментах неспаривающиеся, умеренно спаривающиеся и сильно спаривающиеся антитела, идентифицированные в таблице 2, по-видимому, конкурируют друг с другом сильно, умеренно или не конкурируют вообще соответственно. Только комбинации антител Группы I с антителами Группы III могли связываться одновременно с IGFBP-3 без каких-либо помех. Опять В9 мог также сильно связываться с IGFBP-3 в присутствии В3.
Связывание антител Группы I с IGFBP-3 значимо ингибировалось предынкубацией IGFBP-3 с IGF-I или IGF-II. Это давало основание поместить эпитопы узнавания Группы I в сайт связывания IGF или вблизи сайта связывания IGF. В повторяемых экспериментах связывание антител Группы I с комплексами IGF/IGFBP-3 уменьшалось более чем на 50%, тогда как активность антител Группы II и Группы III, а также поликлональных антител против IGFBP-3 (рВ11) оставалась относительно неизмененной (таблица 4, фиг.2). Связывание В9 было приблизительно в 2 раза более высоким, чем связывание постороннего антитела (анти-PSA), и на него не влияло связывание IGF с IGFBP-3. Антитело В10 было нереактивным, так как оно генерировало такой же сигнал, что и антитело анти-PSA.
В двухсайтном ELISA "смешанных антител" (антитело анти-IGFBP, спаренное с антителами анти-ALS или анти-IGF), антитела в Группе II и Группе III продемонстрировали одновременное связывание с природно встречающимся (нативным) сывороточным IGFBP-3-комплексом в комбинации с антителами анти-IGF-I (I-1) или анти-ALS (рА-1), используемыми для детектирования. Самый сильный сигнал спаривания наблюдали в случае пары антител В2/рА-1 и пары антител B3/I-1. Одновременное связывание с сывороточным IGFBP-3-комплексом не наблюдали при оценке антител анти-IGF-I против антител анти-ALS во всех возможных комбинациях смешанных антител. Связывания с другими возможными комбинациями смешанных антител были одинаковыми со связыванием, в котором участвовало антитело негативного контроля анти-PSA (таблица 5).
Характеристики связывания этих четырех групп суммированы следующим образом:
Группа I: Антитела против IGFBP (В5, В6 и В8). Эта группа антител узнавала эпитопы, которые картированы в сайте или вблизи сайта связывания лиганда IGFBP-3, как определено по ингибированию их связывания в ответ на предынкубацию IGFBP-3 с IGF. Эти антитела не могли быть отличены друг от друга, так как они не связывались одновременно с IGFBP-3 в сэндвич-ELISA и сильно конкурировали друг с другом в тестах конкурентного связывания.
Недавно при помощи вестерн-иммуноблоттинга различных фрагментов IGFBP-3 было показано, что два антитела из Группы I (В5 и В8) узнают эпитопы как при N-концевом районе (IGFBP-31-97), так и при С-концевом районе (IGFBP-3~200-264), тогда как В6, как было обнаружено, взаимодействует только с N-концевым (IGFBP-31-97) фрагментом [44] . Результат данного исследования, по-видимому, показывает связывание В6 с С-концевым, а также с N-концевым районами IGFBP-3. В6 сильно ингибировал связывание В5 и В8 с IGFBP-3 в парном конкурентном ELISA, но в случае В5 и В8, продемонстрировал сильное "перекрывание" образования сэндвича с В3.
Возможно также, что В6 связывается с N-концевыми последовательностями, которые образуют часть лиганд (IGF)-связывающего сайта IGFBP-3. Поскольку считают, что IGF-связывающий сайт IGFBP-3 включает в себя как N-, так и С-концевые последовательности, антитела, которые связываются с такими конформационными эпитопами, могли бы сильно конкурировать за связывание с нативной молекулой, но по-другому с денатурированными фрагментами в вестерн-иммуноблот-анализе. Однако обнаружение специфичности в отношении лигандсвязывающего сайта для антител, которые, как сообщалось, связываются как с N-, так и с С-концом IGFBP-3 [44], согласуется с представлением, что как N-, так и С-концевые последовательности IGFBP-3 участвуют в образовании IGF-связывающего сайта [44, 45].
Группа II: антитела IGFBP (В1, В2, В4, В7). Хотя эпитопы, узнаваемые антителами Группы II, перекрываются с эпитопами, узнаваемыми антителами Группы I, связывание IGF с IGFBP-3 не влияет на иммунореактивность антител Группы II. Антигенный кластер, узнаваемый антителами Группы II, был, следовательно, картирован в зоне на молекуле, удаленной от сайта связывания лиганда IGFBP-3. Опять-таки антитела Группы II не могли быть отличены друг от друга, так как они, по-видимому, связываются с перекрывающимися эпитопами. С открытиями авторов согласуется сообщенная специфичность антител Группы II в отношении промежуточных последовательностей IGFBP-3 (IGFBP-398-159), как оценено вестерн-иммуноблот-анализом [44].
Группа III: антитело IGFBP (B3) - только одно антитело определяло третий эпитоп. Детерминанта для этого антитела также перекрывалась с детерминантами Группы II, но отличалась от антител Группы I. B3 обнаруживал сильные неконкурирующие сэндвич-тесты с антителами в Группе I. Неожиданно было обнаружено, что это антитело узнает эпитоп в N-концевом районе IGFBP-3 (IGFBP-31-97) [44] , который согласно открытиям авторов изобретения не должен перекрываться с эпитопами N-концевого IGF-связывающего района.
Группа IV: антитело IGFBP (B9) - четвертый антигенный эпитоп, узнаваемый B9, является, по-видимому, отличающимся конформационным эпитопом, который был доступным только после связывания В3 с IGFBP-3. В связывании ELISA с использованием захватывающих-детектирующих антител, антитела Группы II и Группы III могли связываться одновременно с нативными сывороточными IGFBP-3-комплексами в комбинации с антителами I-1 или рА-1, используемыми для детектирования. Самый сильный сигнал спаривания получали для комбинаций B3/I-1 (ELISA-1) и В2/рА-1 (ELISA-2). Это показывает, что даже в нативных тройных IGFBP-3-комплексах антигенные домены на IGF-I, а также на ALS являются доступными для связывания антител и могут участвовать в двухсайтном "захват-детектирование"-образовании антител. В согласии с этими открытиями являются недавние наблюдения, что связывание rIGF-I с твердофазным IGFBP-3 могло быть детектировано С-конец-специфическими (D-домен, остатки 63-70) мышиными моноклональными антителами против IGF-I в экспериментах по последовательному связыванию [46].
Отсутствие одновременного "двухсайтного" связывания вышеописанных I-1 и рА-1 с сывороточным IGFBP-3-комплексом предполагает тесную близость связывания N-концевой части ALS- и IGF-связывающего сайта IGFBP-3 (фиг.1). Это наблюдение может дать объяснение сообщенного открытия, что связывание ALS с IGFBP-3 является важным модулятором IGFBP-3-аффинности в отношении IGF-пептидов [47]. Подобно действию антитела В3 на иммунореактивность антитела В9, описанному выше, проксимальное связывание ALS с сайтом связывания лиганда IGFBP-3 могло бы вызывать конформационные изменения, приводящие к повышенной IGF-связывающей аффинности IGFBP-3.
ПРИМЕР 5. Способы: ELISA-1 И ELISA-2
А. ELISA-1
Среди возможных комбинаций анти-IGFВР-3/анти-IGF-1 анти-IGFВР-3 В3 и анти-IGF-I I-1 генерируют самый сильный сигнал связывания с сывороточным IGF-связывающим белковым комплексом при использовании в качестве захватывающих и детектирующих антител, соответственно. Протокол ELISA был оптимизирован, как описано ранее [37, 41].
В этом анализе стандарты или пробы сыворотки (0,025 мл 10-40-кратно разведенные в тест-буфере для нулевого стандартного матрикса) добавляли в двух повторностях в покрытые антителом лунки с последующим добавлением тест-буфера для ELISA-1 (0,10 мл) и 1-часового инкубированием при комнатной температуре с непрерывным встряхиванием. Лунки промывали пять раз и инкубировали с 0,1 мл на лунку конъюгата анти-IGF-I-HRP (разведенного в тест-буфере до приблизительно 0,1-0,25 мг/л) в течение 1 часа при комнатной температуре. Эти лунки промывали пять раз промывочным раствором, добавляли 0,1 мл раствора субстрата ТМВ/Н2О2 и выдерживали дополнительно в течение 10 минут инкубации при комнатной температуре. Затем добавляли стоп-раствор (0,1 мл) и измеряли поглощение посредством измерения на двух длинах волн, при 450 нм с коррекцией фоновой длины волны, установленной при 620 нм.
Данные ELISA анализировали с использованием пакетов сжатия данных, включенных в микропланшет-ридер LABSYSTEMS с аппроксимацией кривых кубическим сплайном (сглаженным). Другие статистические анализы выполняли с использованием статистического пакета MICROSOFT EXCEL 97 STATISTICAL PACKAGE (MICROSOFT CORPORATION (USA)) на IBM-совместимом компьютере Pentium. Описательные данные представлены в виде среднего значения и стандартного отклонения (SD), если нет иных указаний. Линейный регрессионный анализ выполняли по методу наименьших квадратов, и коэффициенты корреляции определяли по методу Пирсона.
В. ELISA-2
Среди возможных комбинаций aнти-IGFBP-3/aнти-ALS анти-IGFBP-3 B2 и анти-ALS рА-1 генерируют самый сильный сигнал связывания с сывороточным IGF-связывающим белковым комплексом при использовании в качестве захватывающих и детектирующих антител соответственно. Протокол ELISA был точно такой, как описанный выше, за исключением того, что оптимальные объемы пробы и тест-буфера были 25 мкл и 50 мкл на лунку соответственно.
С. Проверка достоверности ELISA-1 и ELISA-2
Для проверки достоверности ELISA-1 и ELISA-2 IGFBP-3-комплекса использовали пробы подходящим образом разбавленной свежей сыворотки. В обоих анализах нижний предел детектирования (чувствительность) определяли интерполяцией среднего плюс 2 SD 12 повторностей измерений отрицательного контроля (буфера для нулевого стандартного матрикса). Коэффициенты корреляции (CV) внутри теста определяли анализом повторностей (n=12) трех проб при уровнях IGFBP-3/IGF-I 8,6-24,3 ПЕ/л и уровней IGFBP-3/ALS 6,7-18,7 ПЕ/л; коэффициенты корреляции между тестами определяли измерением двух повторностей соответствующих проб в 7-9 отдельных анализах. Выход оценивали добавлением 50 мкл проб, содержащих различные уровни IGFBP-3-комплекса к 450 мкл 10-кратно разведенных проб сыворотки с последующим анализом. Процент извлечения определяли сравнением количества добавленного IGFBP-3-комплекса с количеством, измеренным после вычитания эндогенного уровня IGFBP-3-комплекса. Линейность тестировали анализом проб сыворотки, разведенных сначала в 10 раз, затем последовательно в 2-16 раз в буфере для нулевого стандартного матрикса. Специфичность ELISA для IGFBP-3-комплекса анализировали определением IGF-I (до 300 мкг/л), IGF-II (до 3000 мкг/л), IGFBP-1, 2, 4-6 до 500 мкг/л и IGFBP-3 до 4,3 мг/л.
Для оценки стабильности IGFBP-3-комплекса и, следовательно, практической применимости данного теста, аликвоты проб свежей сыворотки (n=3) хранили при комнатной температуре, 4oС и -20oС и затем анализировали в дни 0, 2 и 3 хранения при помощи ELISA-1 и ELISA-2 после 10-кратного разведения в буфере для нулевого стандартного матрикса. В день анализа пробы анализировали относительно новой серии стандартов, свежеприготовленных из замороженной (при -20oС) аликвоты исходного стандартного пула сывороток. Аликвоты серии стандартов также хранили при вышеуказанных температурах и анализировали таким же образом.
Результаты ELISA-1 и ELISA-2 сравнивали с результатами, полученными из тестов предшествующего уровня техники. IGF-I, IGF-II, IGFBP-3 и ALS анализировали при помощи наборов для иммуноанализа, производимых DSL (Webster, ТХ). Эти анализы основаны на принципах неконкурентного ELISA, проводимого в покрытых антителом микролунках [15, 37, 38, 39], и использовании меченных пероксидазой хрена (HRP) детектирующих антител, как описано в [41].
IGF-I и IGF-II анализировали при помощи ACTIVEТМ Non-Extraction ELISA для IGF-I и IGF-II (DSL, Webster), который включает в себя стадию предобработки пробы для диссоциации IGF-I от IGFBP перед анализом [43]. Стадия предобработки пробы включает в себя смешивание 20 мкл пробы с 1,0 мл подкисляющего IGF-I-буфера с последующим 30-минутным инкубированием при комнатной температуре и добавлением 1,0 мл нейтрализующего буфера. Конечный фактор разведения препарата был 101-кратным и для анализа IGF использовали 20 мкл этой обработанной пробы. Каждый из наборов ELISA для IGF имеет общее время инкубирования менее 3 часов и общую погрешность менее 10%.
ACTIVEТМ ELISA для IGFBP-3 (DSL, Webster) включает в себя 101-кратное разведение пробы и использует 25 мкл предварительно разведенной пробы для анализа IGFBP-3. Анализ имеет общее время инкубирования около 3 часов, диапазон стандартов 2-100 мкг/л (0,20-10 мг/л после поправки на фактор разведения пробы) и общую погрешность менее 10% [39].
ACTIVEТМ ELISA для общего ALS (DSL, Webster) включает в себя стадию предобработки 101-кратно разведенной пробы, которая приводит к развертыванию как находящегося в комплексе, так и не находящегося в комплексе ALS, что позволяет измерить общие уровни ALS [15]. Стадия предобработки пробы включает в себя смешивание 10 мкл пробы с 1,0 мл буфера для предобработки пробы с последующим 30-минутным инкубированием при комнатной температуре. Конечный фактор разведения препарата был 101-кратным и для анализа общего ALS использовали 20 мкл этой обработанной пробы. Анализ имеет общее время инкубирования около 2 часов, диапазон стандартов 6-600 мкг/л (0,6-60 мг/л после поправки на фактор предварительного разведения) и общую погрешность менее 10%.
Поглощение тестов ELISA измеряли микропланшет-ридером LABSYSTEMS MULTISCAN MULTIOFT (LABSYSTEMS, Helsinki, Finland).
ПРИМЕР 6. Результаты: ELISA-1 и ELISA-2
A. ELISA-1 и ELISA-2
На основе вышеописанного картирования эпитопов были разработаны новые иммуноанализы для количественного определения циркулирующих в кровотоке IGFBP-3-комплексов. Как IGFBP-3/IGF-I-ELISA-l, так и IGFBP-3/ALS-ELISA-2 включают в себя двухсайтную неконкурентную (последовательную) иммунореакцию и основаны на твердофазном антителе анти-IGFВР-3 В3 или В2, спаренным с детектирующими антителами анти-IGF-I I-1 или анти-ALS рА-1 соответственно.
Оптимизированные протоколы были установлены посредством оценки действий различных технических манипуляций на аналитическую производительность этих анализов, как описано ранее [37, 41]. Были выбраны концентрация покрывающего микролунки антитела 10 мг/л, концентрация детектирующего антитела около 0,1-0,25 мг/л, 60-минутные инкубации при комнатной температуре первой и второй стадий и 10-минутная стадия проявления субстрата. Среди испытанных переменных, состав тест-буфера и концентрации покрывающего лунки антитела оказывали наиболее очевидное действие на чувствительность и динамический диапазон этих анализов.
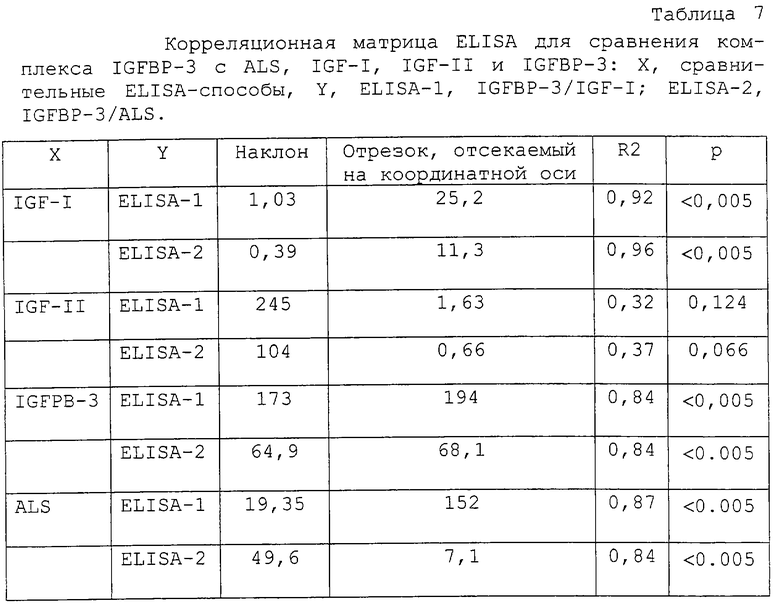
Типичная стандартная кривая и характеристики производительности ELISA-1 и ELISA-2 суммированы на фиг. 3 и в таблице 6. Добавление IGF-I (до 300 мкг/л), IGF-II (до 3000 мкг/л), IGFBP-2 и IGFBP-4-6 (до 500 мкг/л) и IGFBP-3 (до 4,2 мг/л) к буферу для нулевого стандартного матрикса не обнаружили никакой перекрестной реактивности.
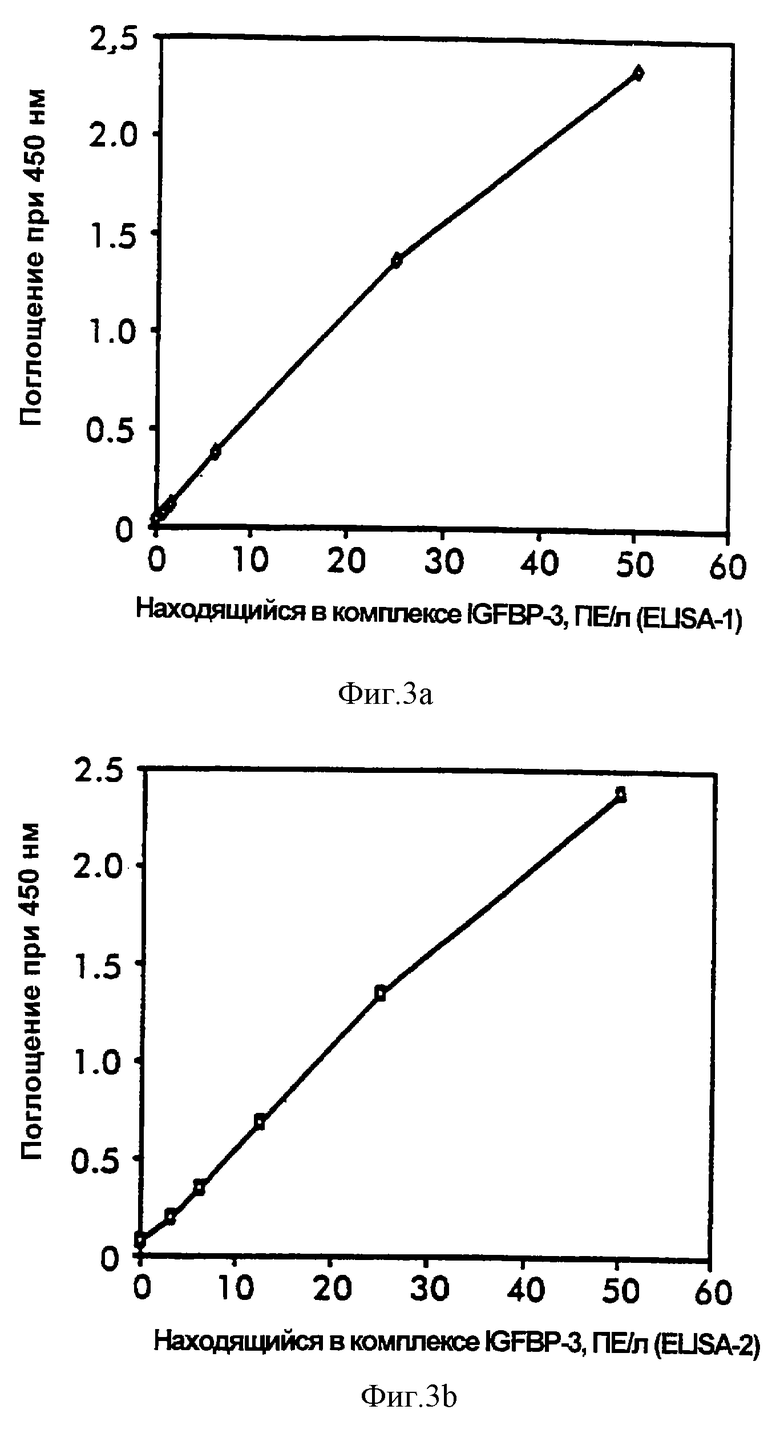
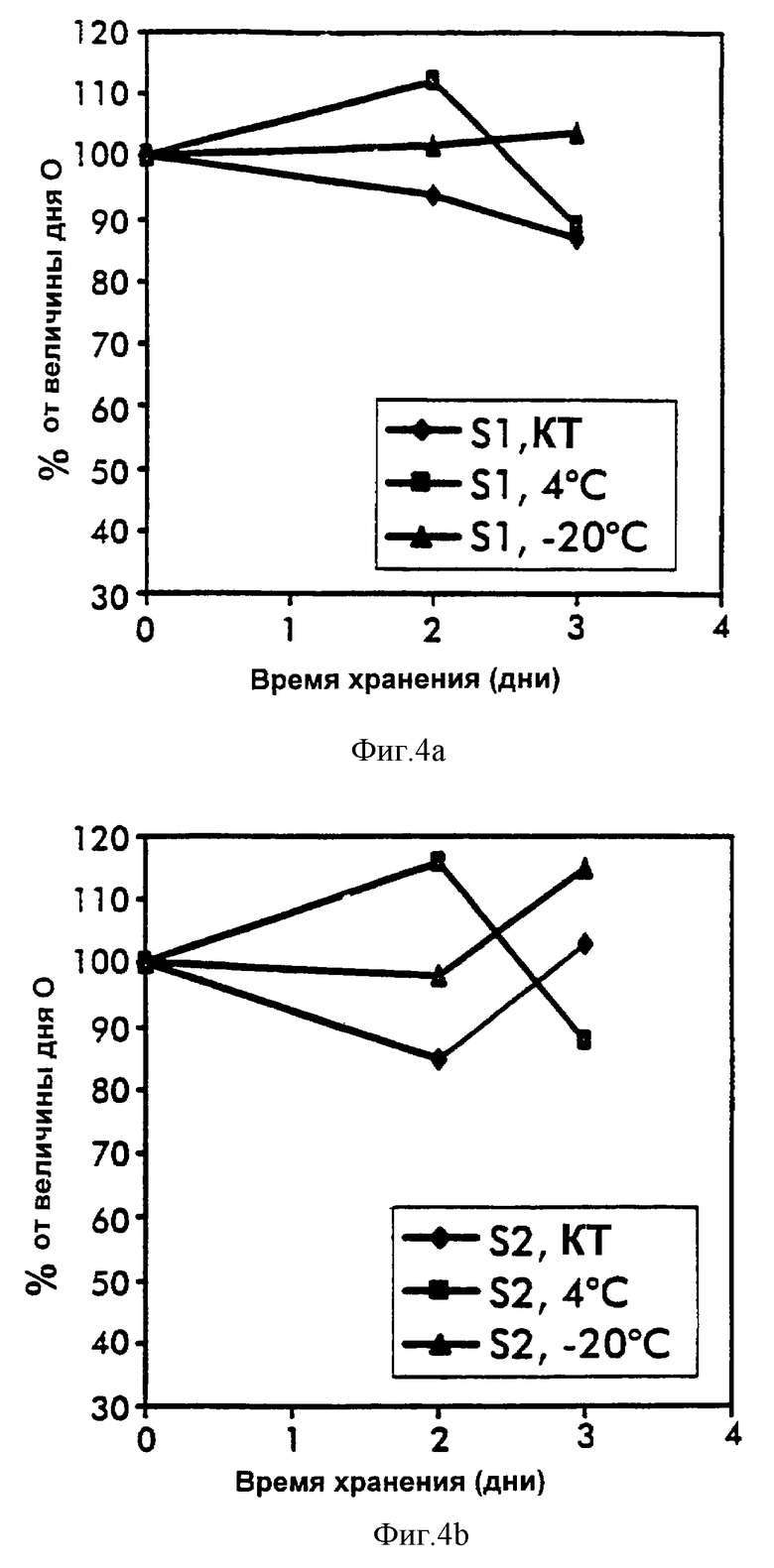
Сывороточные IGFBP-3-комплексы анализировали в повторяемой аликвоте проб, хранимых при комнатной температуре, 4oС и -20oС. Как измерено при помощи ELISA-1 и ELISA-2, IGFBP-3-комплексы продемонстрировали высокую стабильность при всех температурах в течение периода времени хранения до 3 дней и выход при 4oС и -20oС были по меньшей мере 85% от величины дня 0 (фиг.4 и 5). Стандарты на основе сыворотки, хранящиеся и анализированные, как описано выше, обнаружили такую же стабильность и IGFBP-3-комплексы в пуле свежих сывороток, хранящемся при -20oС, обнаруживали стабильность в течение по меньшей мере 2 месяцев.
Несмотря на сложность их построения, анализы ELISA-1 и ELISA-2 для IGFBP-комплекса продемонстрировали приемлемые характеристики аналитической производительности. Обнаружение относительно высокой стабильности IGFBP-3-комплексов при различной температуре и даже в разведенной форме было неожиданным и послужило инструментом в успехе разработки этих анализов. Продемонстрированная линейность этих анализов в ответ на разведение проб может предполагать измерение только прочно связанных тройных IGFBP-3-комплексов. Связывание антител и разведение проб могут индуцировать диссоциацию IGF и, следовательно, удаление рыхло связанных комплексов.
В. Сравнение с анализами предшествующего уровня техники
Пробы плазмы из субъектов с не подвергавшимся лечению GHRD (n=11) и нормальных субъектов соответствующего возраста (n= 16) и пробы сыворотки из взрослых с акромегалией (n=8) или с GHD (n=5) одновременно анализировали на IGF-I, IGF-II, IGFBP-3 и общий ALS при помощи ELISA-1 и ELISA-2.
Регрессионный анализ данных показал высокую степень корреляций между ELISA-1 IGFBP-3-комплекса и ELISA-2 IGFBP3-комплекса в зависимости от уровней IGF, IGFBP-3 и общего ALS (таблица 7 и фиг.6-9). Наилучшие общие корреляции наблюдали в сравнениях с уровнями IGF-I, тогда как корреляции в зависимости от уровней IGF-II были относительно слабые. Уплощенный вид "нижнего конца" графиков корреляции обусловлен значимыми различиями в относительных уровнях различных аналитов как функции возраста (фиг.10), а также клинических условий и не обязательно обусловлен слабыми корреляциями нижнего конца. В самом деле, корреляции в этом диапазоне GHRD были такими же, что и корреляции, включающие величины всех проб.
Хотя количества нормальных проб в группах разного возраста были небольшими, общее распределение уровней IGF, IGFBP-3 и ALS в зависимости от возраста было сходно с распределениями, сообщенными ранее, т.е. уровни обнаруживали значимое повышение во время пубертатного возраста [21, 34, 35]. Как ожидалось, на уровни IGFBP-3-комплекса одинаково влиял возраст. Однако комплексы на основе IGF-I, измеренные при помощи ELISA-1, обнаруживали самые широкие вариации, являясь самыми низкими в самой молодой и самой старой возрастных групп и наивысшими во время пубертатного возраста (фиг.10).
В графиках средних уровней аналитов в виде процента от соответствующего среднего значения нормальных величин, средние значения уровней IGFBP-3-комплекса, измеренные в субъектах с GHRD и GHD при помощи ELISA-1, были менее 8% и 2% от среднего значения нормальных величин соответственно. Следующие наибольшие различия наблюдали для уровней IGF-I; средние уровни IGF-I в субъектах с GHRD и GHD были менее 13% и менее 24% от среднего значения нормальных величин (фиг. 11). В сравнительных графиках распределения индивидуальных величин уровни IGFBP-3-комплекса, измеренные при помощи ELISA-2, показали сходное, если не лучшее, различение между различными группами проб, в частности, между субъектами с GHRD, нормальными субъектами и субъектами с GHD (фиг.12).
В нескольких экспериментах, включающих в себя пробы, выбранные на основе случайного отбора, ELISA для IGFBP-комплексов обнаружили значимую корреляцию с уровнями IGF-I, IGFBP-3 и ALS. Это подтверждали затем посредством предварительных клинических оценок с пробами из субъектов с GHRD, нормальных контролей соответствующего возраста и с образцами из взрослых пациентов с акромегалией и GHD. В целом, как ELISA-1, так и ELISA-2 продемонстрировали высокую корреляцию с уровнями IGF-I, IGFBP-3 и ALS. Это подтверждает сообщенные наблюдения, что большая часть циркулирующих в кровообращении IGF и IGFBP-3 присутствует, прежде всего, в тройном белковом комплексе и что доступность IGF может быть ключевой детерминантой образования тройного комплекса [4-12].
Представляло интерес наблюдение, что IGFBP-комплексы, измеренные при помощи ELISA-1, показали самые широкие относительные вариации как функция от возраста. Уровни IGFBP-комплекса индивидуальных проб были самыми низкими в более молодой и наивысшими в наиболее старшей возрастной группе, пубертатного диапазона (фиг. 10). Подобным образом, в сравнительных графиках распределения величин индивидуальных проб, ELISA-1 продемонстрировала сходное, если не лучшее, различение между различными группами проб (фиг.12). Относительно более низкая величина IGFBP-комплекса, измеренная в группе более молодого возраста при помощи ELISA-1, может предполагать доступность пропорционально более высокого количества IGF-I в свободной (или диссоциируемой) форме в этой возрастной группе, как было сообщено недавно для раннего детского возраста [48].
С. Выводы
Данные, представленные здесь, устанавливают, что описанные здесь антитела могут быть выбраны на основе их эпитопных карт и использованы в успешном формате способа ELISA с захватом-детектированием антигенов. Картирование дополнительных антител может быть выполнено по описанному здесь способу. В частности, была бы выгодной идентификация антител против IGFBP, на связывание которых не влияет связывание IGF-I или ALS. Такие антитела могли бы сделать возможным тест определения общего IGFBP-3 без учета степени комплексообразования.
Кроме двух пар антител, приведенных в качестве примеров выше (ELISA-1 и ELISA-2), другие пары или даже триплеты антител могут быть оптимизированы способом, сходным с описанным здесь способом. Таким образом, приведенные выше примеры построены не как ограничительные, а скорее как иллюстрирующие многие комбинации антител, которые могут быть использованы в формате захвата-детектирования.
В конце описания приведен список источников информации.
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗЫВАЮЩИЕ ПРОТЕИНЫ, СПЕЦИФИЧНЫЕ ПО ОТНОШЕНИЮ К ИНСУЛИН-ПОДОБНЫМ ФАКТОРАМ РОСТА, И ИХ ИСПОЛЬЗОВАНИЕ | 2013 |
|
RU2549703C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ЛИЗИЛОКСИДАЗОПОДОБНЫМ ФЕРМЕНТОМ-2 (LOXL2), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2549684C2 |
| ДЕТЕКТИРОВАНИЕ РАКА ЯИЧНИКА ПО ПОВЫШЕННЫМ УРОВНЯМ BCL-2 В МОЧЕ | 2007 |
|
RU2436098C2 |
| УСТРОЙСТВА, СПОСОБЫ И НАБОРЫ ДЛЯ ИММУНОХРОМАТОГРАФИИ | 2011 |
|
RU2568875C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-21 ЧЕЛОВЕКА | 2008 |
|
RU2504552C2 |
| КОМПОЗИЦИИ И СПОСОБЫ АНАЛИЗА РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2012 |
|
RU2641968C2 |
| ТЕРАПИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ, РЕЗИСТЕНТНОЙ К ПРЕПАРАТАМ НА ОСНОВЕ ПЛАТИНЫ | 2005 |
|
RU2403065C2 |
| СПОСОБ СНИЖЕНИЯ ИММУНОГЕННОСТИ | 2010 |
|
RU2585534C2 |
| СПОСОБ ОБНАРУЖЕНИЯ N-КОНЦЕВОГО ПРО-МНП | 2000 |
|
RU2218568C2 |
| УЛУЧШЕННЫЙ БЕЛОК REP ДЛЯ ПРИМЕНЕНИЯ В ДИАГНОСТИЧЕСКИХ АНАЛИЗАХ | 2018 |
|
RU2736275C1 |













Изобретение относится к области медицины, а именно к способу анализа статуса гормона роста индивидуума. Сущность способа: проводят иммуноанализ тройного комплекса инсулиноподобного фактора роста IGF/IGFBP-3/ALS (150 кДа). Альтернативно может быть определен двойной комплекс. Способ предусматривает захват IGFBP-комплекса одним антителом, связанным с твердой фазой, и детектирование этого комплекса вторым антителом, связанным с меткой. В изобретении описан также набор моноклональных и поликлональных антител, используемый для такого анализа. Технический результат изобретения состоит в расширении арсенала диагностических средств и способов, применяемых при заболеваниях, связанных с нарушением метаболизма гормона роста, и некоторых опухолевых патологиях. 6 с. и 12 з.п.ф-лы, 7 табл., 21 ил.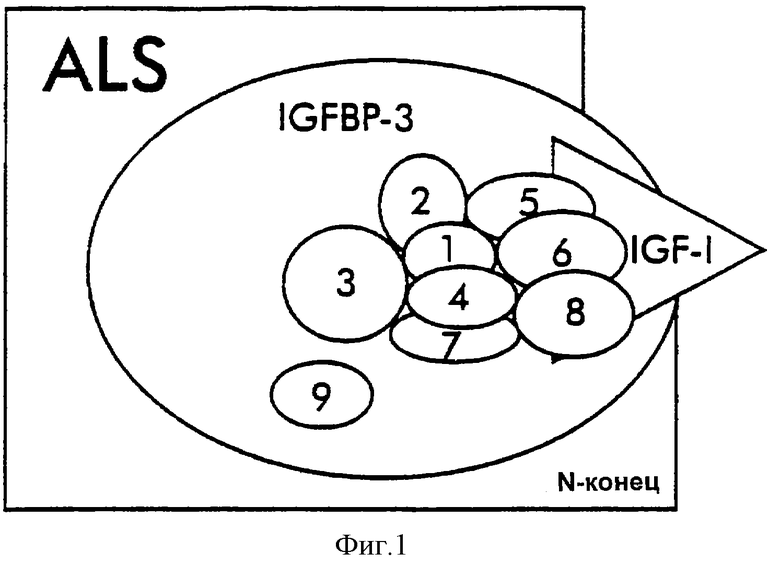
| US 4271140 А, 02.06.1981 | |||
| Baxter R.S | |||
| Circulating levels and molecular distribution.. | |||
| J.Endocrinol | |||
| Metab | |||
| - v | |||
| Деревянный торцевой шкив | 1922 |
|
SU70A1 |

