Область техники, к которой относится изобретение
Настоящее изобретение относится к способу лечения резистентных к препаратам на основе платины злокачественной опухоли яичника, первичной перитонеальной карциномы или карциномы фаллопиевой трубы комбинацией антитела против HER2, которое эффективно подавляет димеризацию HER, а также гемцитабина.
Уровень техники
Антитела против HER
Семейство HER рецепторных тирозинкиназ является семейством важных медиаторов роста, дифференциации и выживаемости клеток. Семейство рецепторов включает четыре различных члена, включающих рецептор эпидермального фактора роста (EGFR, ErbB1 или HER1), HER2 (ErbB2 или p185neu), HER3 (ErbB3) и HER4 (ErbB4 или tyro2).
EGFR, кодируемый геном erbB1, в основном принимает участие в развитии злокачественных образований у человека. В частности, наблюдали повышенную экспрессию EGFR в злокачественных опухолях молочной железы, мочевого пузыря, легких, головы, шеи и желудка, а также в глиобластоме. Повышенная экспрессия EGFR-рецептора часто связана с увеличенной продукцией лиганда EGFR, трансформирующего фактора роста альфа (ТФР-α, ТGF-α), теми же опухолевыми клетками, приводя к активации рецептора аутокринным стимулирующим путем. Baselga and Mendelsohn, Pharmac. Ther., 64: 127-154 (1994). Оценивали эффективность моноклональных антител против EGFR или его лигандов, ТФР-α и EGF, в качестве терапевтических средств для лечения таких злокачественных образований. Смотри, например, Baselga and Mendelsohn, выше; Masui et al., Cancer Research, 44: 1002-1007 (1984) и Wu et al., J. Clin. Invest., 95: 1897-1905 (1995).
Второй член семейства HER, p185neu, первоначально был установлен в качестве продукта трансформирующего гена из нейробластомы у обработанных химическими соединениями крыс. Активная форма протоонкогена neu возникает в результате точечной мутации (валин в глутаминовую кислоту) в трансмембранной области кодируемого белка. Амплификацию человеческого гомолога neu наблюдают в злокачественных опухолях молочной железы и яичника, и она коррелирует с плохим прогнозом (Slamon et al., Science, 235: 177-182 (1987); Slamon et al., Science, 244: 707-712 (1989) и патент США № 4968603). К настоящему времени не сообщалось о точечных мутациях, подобных в протоонкогене neu, для человеческих опухолей. Также наблюдали сверхэкспрессию HER2 (часто, но не во всех случаях в результате амплификации гена) в других карциномах, включая карциному желудка, эндометрия, слюнной железы, легких, почки, ободочной кишки, щитовидной железы, поджелудочной железы и мочевого пузыря. Смотри, например, среди прочих King et al., Science, 229: 974 (1985); Yokota et al., Lancet, 1: 765-767 (1986); Fukushige et al., Mol. Cell Biol., 6: 955-958 (1986); Guerin et al., Oncogene Res., 3: 21-31 (1988); Cohen et al., Oncogene, 4: 81-88 (1989); Yonemura et al., Cancer Res., 51: 1034 (1991); Borst et al., Gynecol. Oncol., 38: 364 (1990); Weiner et al., Cancer Research, 50: 421-425 (1990); Kern et al., Cancer Research, 50: 5184 (1990); Park et al., Cancer Res., 49: 6605 (1989); Zhau et al., Mol. Carcinog., 3: 254-257 (1990); Aasland et al., Br. J. Cancer, 57: 358-363 (1988); Williams et al., Pathobiology, 59: 46-52 (1991) и МcCann et al., Cancer, 65: 88-92 (1990). HER2 может сверхэкспрессироваться в злокачественной опухоли простаты (Gu et al., Cancer Lett., 99: 185-9 (1996); Ross et al., Hum. Pathol. 28: 827-33 (1997); Ross et al., Cancer 79: 2162-70 (1997) и Sadasivan et al., J. Urol., 150: 126-31 (1993)).
Описаны антитела против крысиного p185neu и белковых продуктов человеческого HER2. Drebin и соавторы получили антитела против продукта крысиного гена neu, p185neu. Смотри, например, Drebin et al., Cell, 41: 695-706 (1985); Myers et al., Meth. Enzym. 198: 277-290 (1991) и WO 94/22478. Drebin et al., Oncogene, 2: 273-277 (1988) сообщают, что смеси антител, реагирующих с двумя различными областями p185neu, приводят к синергическим противоопухолевым эффектам в отношении трансформированных neu клеток NIH-3T3, имплантированных голым мышам. Также смотри патент США № 5824311 от 20 октября 1998 г.
Hudziak et al., Mol. Cell Biol., 9(3): 1165-1172 (1989) описывают получение панели антител против HER2, которые были охарактеризованы с использованием клеток опухоли молочной железы SK-BR-3. Определяли относительную клеточную пролиферацию клеток SK-BR-3 после воздействия антител по окрашиванию монослоев кристаллическим фиолетовым через 72 ч. С использованием данного теста максимальное ингибирование получали с антителом, названным 4D5, которое подавляло клеточную пролиферацию на 56%. Другие антитела в данной панели снижали клеточную пролиферацию в меньшей степени в данном тесте. Дополнительно было установлено, что антитело 4D5 сенсибилизирует клетки опухоли молочной железы со сверхэкспрессией HER2 к цитотоксическому действию ТФР-α. Смотри также патент США № 5677171 от 14 октября 1997 г. Антитела против HER2, обсужденные Hudziak et al., были дополнительно охарактеризованы Fendly et al., Cancer Research., 50: 1550-1558 (1990); Kotts et al., In vitro 26(3): 59A (1990); Sarup et al., Growth Regulation 1: 72-82 (1991); Shepard et al., J. Clin. Immunol., 11(3): 117-127 (1991); Kumar et al., Mol. Cell Biol., 11(2): 979-986 (1991); Lewis et al., Cancer Immunol. Immunother., 37: 255-263 (1993); Pietras et al., Oncogene, 9: 1829-1838 (1994); Vitetta et al., Cancer Research, 54: 5301-5309 (1994); Sliwkowski et al., J. Biol Chem., 269(20): 14661-14665 (1994); Scott et al., J. Biol. Chem., 266: 14300-5 (1991); D'souza et al., Proc. Natl. Acad. Sci., 91: 7202-7206 (1994); Lewis et al., Cancer Research, 56: 1457-1465 (1996); и Schaefer et al., Oncogene, 15: 1385-1394 (1997).
Рекомбинантный гуманизированный вариант мышиного антитела против HER2 4D5 (huMAb4D5-8, rhuMAb, трастузумаб или HERCEPTIN®; патент США № 5821337) является клинически эффективным у пациенток с метастазирующими злокачественными опухолями молочной железы со сверхэкспрессией HER2, которые до этого получали продолжительную противоопухолевую терапию (Baselga et al., J. Clin. Oncol., 14: 737-744 (1996)). Трастузумаб разрешен к применению Администрацией по санитарному надзору за продуктами питания и медикаментами от 25 сентября 1998 г. для лечения пациенток с метастазирующими опухолями молочной железы со сверхэкспрессией белка HER2.
Описаны другие антитела к HER2 с различными свойствами Tagliabue et al., Int. J. Cancer, 47: 933-937 (1991); McKenzie et al., Oncogene, 4: 543-548 (1989); Mair et al., Cancer Res., 51: 5361-5369 (1991); Bakus et al., Molecular Carcinogenesis, 3: 350-362 (1990); Stancovski et al., PNAS (USA) 88: 8691-8695 (1991); Bakus et al., Cancer Research, 52: 2580-2589 (1992); Xu et al., Int. J. Cancer, 53: 401-408 (1993); WO 94/00136; Kasprzyk et al., Cancer Research, 52: 2771-2776 (1992); Hancock et al., Cancer Res., 51: 4575-4580 (1991); Shawver et al., Cancer Res., 54: 1367-1373 (1994); Arteaga et al., Cancer Res., 54: 3758-3765 (1994); Harwerth et al., J. Biol. Chem., 267: 15160-15167 (1992); патент США № 5783186 и Klapper et al., Oncogene, 14: 2099-2109 (1997).
В результате скрининга на гомологию было идентифицировано два других члена семейства рецепторов HER: HER3 (патенты США № 5183884 и 5480968, а также Kraus et al., PNAS (USA) 86: 9193-9197 (1989)) и HER4 (заявка на Европейский патент № 599274; Plowman et al., Proc. Natl. Acad. Sci. USA, 90: 1746-1750 (1993) и Plowman et al., Nature, 366: 473-475 (1993)). Для обоих данных рецепторов характерна повышенная экспрессия, по меньшей мере, в некоторых клеточных линиях опухолей молочной железы.
Как правило, рецепторы HER обнаруживают в различных комбинациях в клетках, и полагают, что гетеродимеризация повышает разнообразие ответных реакций на различные лиганды HER (Earp et al., Breast Cancer Research and Treatment, 35: 115-132 (1995)). EGFR связывается с шестью различными лигандами: эпидермальным фактором роста (EGF), трансформирующим фактором роста альфа (ТФР-α), амфирегулином, связывающимся с гепарином эпидермальным фактором роста (НВ-EGF), бетацеллюлином и эпирегулином (Groenen et al., Growth Factors, 11: 235-257 (1994)). Семейство белков герегулинов, происходящих в результате альтернативного сплайсинга одного гена, представляют лиганды для HER3 и HER4. Семейство герегулинов включает альфа, бета и гамма герегулины (Holmes et al., Science, 256: 1205-1210 (1992); патент США № 5641869 и Schaefer et al., Oncogene, 15: 1385-1394 (1997)); факторы дифференциации neu (NDF); глиальные факторы роста (GGF); рецептор, индуцирующий активность ацетилхолина (ARIA); и факторы сенсорных и двигательных нейронов (SMDF). Обзор смотри у Groenen et al., Growth Factors, 11: 235-257 (1994); Lemke, G. Molec. & Cell Neurosci., 7: 247-262 (1996); и Lee et al., Pharm. Rev., 47: 51-85 (1995). Недавно было идентифицировано три дополнительных лиганда HER: нейрегулин-2 (NRG-2), о котором сообщалось, что он связывается с HER3 или HER4 (Chang et al., Nature, 387: 509-512 (1997) и Carraway et al., Nature, 387: 512-516 (1997)); нейрегулин-3, который связывается с HER4 (Zhang et al., PNAS (USA), 94(18): 9562-7 (1997)); и нейрегулин-4, который связывается с HER4 (Harari et al., Oncogene, 18: 2681-89 (1999)) HB-EGF, бетацеллюлин и эпирегулин, которые также связываются с HER4.
Несмотря на то, что EGF и ТФРα не связываются с HER2, EGF стимулирует EGFR и HER2 к образованию гетеродимера, который активирует EGFR и приводит к трансфосфорилированию HER2 в гетеродимере. Оказалось, что димеризация и/или трансфосфорилирование активируют тирозинкиназу HER2. Смотри Earp et al., выше. Аналогично, когда HER3 коэкспрессируется с HER2, то образуется активный сигнальный комплекс, и антитела против HER2 способны разрушать данный комплекс (Sliwkowski et al., J. Biol. Chem., 269(20): 14661-14665 (1994)). Дополнительно аффинность HER3 для герегулина (HRG) повышается до состояния более высокой аффинности при коэкспрессии с HER2. Также смотри Levi et al., Journal of Neuroscience, 15: 1329-1340 (1995); Morrissey et al., Proc. Natl. Acad. Sci. USA, 92: 1431-1435 (1995) и Lewis et al., Cancer Res., 56: 1457-1465 (1996) в отношении белкового комплекса HER2-HER3. HER4, подобно HER3, образует активный сигнальный комплекс с HER2 (Carraway and Cantley, Cell, 78: 5-8 (1994)).
Злокачественная опухоль яичника
Злокачественная опухоль яичника является наиболее частой причиной смерти от злокачественных образований органов репродуктивной системы у женщин. Ежегодно в США регистрируется 24000 новых диагнозов при смертности от данного заболевания на уровне примерно 13000. Пациентки с далеко зашедшим раком яичника часто подвергаются лечению химиотерапией препаратами на основе платины, часто в комбинации с таксаном. При отсутствии эффективности данных препаратов имеется небольшой выбор терапевтических средств. Часто пациентки с опухолями, чувствительными к препаратам на основе платины, повторно подвергаются лечению платиной, но у значительной части пациенток наблюдается незначительная продолжительность ответной реакции на повторное лечение. Результативность лечения у пациенток с опухолью, резистентной к препаратам на основе платины, еще ниже. Топотекан разрешен к применению Администрацией по санитарному надзору за продуктами питания и медикаментами (FDA) для пациенток с отсутствием эффективности первоначальной или последующей химиотерапии; липосомальный доксорубицин разрешен только для пациенток со злокачественной опухолью яичника, которая резистентна к химиотерапии препаратами платины и паклитакселом. Было показано, что топотекан и липосомальный доксорубицин вызывают частичную ответную реакцию соответственно у 6 и 12% пациенток с опухолью, резистентной к препаратам на основе платины, при среднем значении выживаемости без прогрессирования заболевания, составляющем 14-18 недель. Совсем недавно сообщалось об обещающих результатах с гемцитабином при опухоли яичника с резистентностью к препаратам платины при наличии частичной ответной реакции на уровне 16%, что привело к повышенному применению данного средства в качестве 2 линии терапии. Однако существует четкая потребность в новых и усовершенствованных лекарственных препаратах для пациенток с далеко зашедшим раком яичника, для которых имеющиеся в настоящее время средства терапии не эффективны.
ErbB или семейство рецепторов человеческого эпидермального фактора роста (HER) принимает участие в патогенезе рака яичника. Для целенаправленного воздействия на сигнальный путь с участием HER был разработан пертузумаб (rhuMAb 2C4) в качестве гуманизированного антитела, которое подавляет димеризацию HER2 с другими HER-рецепторами, тем самым ингибируя лиганд-регулируемые фосфорилирование и активацию и даунстрим активацию путей RAS и АКТ.
Гемцитабин применяли для лечения различных опухолей, и он показан для применения при раке поджелудочной железы и легких. Наиболее частым проявлением токсичности при применении гемцитабина в качестве одного средства является цитопения с частотой проявления анемии и нейтропении, равной соответственно 68 и 63%. Другим обычным видом токсичности является тошнота и рвота при общей частоте на уровне 69%, при 13% III степени и 1% IV степени. Диарея имеет место реже в 19% случаев. Кожные высыпания проявляются чаще в 30% случаев при том, что III степени только в 1% случаев. Гемцитабин комбинировали со многими другими химиотерапевтическими средствами, такими как таксаны, антрациклины и препараты на основе платины, без какой-либо существенной повышенной или неожиданной токсичности.
Трастузумаб сочетали с гемцитабином в нескольких комбинациях различных химиотерапевтических средств на фазе II клинических испытаний, и он также хорошо переносился без проявления сердечной или неожиданной токсичности. Safran et al., Proc. Am. Soc. Clin. Oncol., 20: 130a (2001); Miller et al., Oncology, 15(2): 38-40 (2001). Смотри также Zinner et al., Proc. Am. Soc. Clin. Oncol., 20: 328a (2001); Nagourney et al., Breast Cancer Res. Treat., 57: 116, Abstract, 475 (1999); Bun et al., Proc. Am. Assoc. Canc. Res., 41: 719, Abstract #4571 (2000); Konecny et al., Breast Cancer Res. Treat., 57: 114, Abstract 476 (1999); O'Shaugnessy et al., Sem. Oncol., 2(suppl 3): 22-26 (2004); Sledge et al., Sem. Oncol., 2(suppl 3): 19-21 (2003); Zinner et al., Lung Cancer, 44(1): 99-110 (2004); Gatzemeier et al., Ann. Of Oncol., 15: 19-27 (2004), касающиеся комбинации трастузумаба и гемцитабина.
В фазе I клинических испытаний омнитарга в качестве единственного средства для лечения солидных опухолей 3 пациентки с далеко зашедшим раком яичника подверглись лечению пертузумабом. У одной была длительная частичная ответная реакция и у одной пациентки наблюдали стабилизацию заболевания в течение 15 недель. Agus et al., Proc. Am. Soc. Clin. Oncol., 22: 192, Absract 771 (2003).
Сущность изобретения
Настоящее изобретение относится, в первом аспекте, к способу лечения злокачественной опухоли, резистентной к препаратам на основе платины, выбранной из группы, состоящей из злокачественной опухоли яичника, первичной перитонеальной карциномы и карциномы фаллопиевых труб, включающему введение пациентке антитела против HER2, которое подавляет димеризацию HER более эффективно по сравнению с трастузумабом, и химиотерапевтического средства антиметаболита в количествах, эффективных для лечения злокачественной опухоли.
В другом аспекте изобретение относится к способу лечения злокачественной опухоли, резистентной к препаратам на основе платины, выбранной из группы, состоящей из злокачественной опухоли яичника, первичной перитонеальной карциномы и карциномы фаллопиевых труб, включающему введение пациентке антитела против HER2, которое связывается с гетеродимерным связывающим сайтом в HER2, и гемцитабина в количествах, эффективных для лечения злокачественной опухоли.
В еще одном аспекте изобретение относится к способу лечения злокачественной опухоли, резистентной к препаратам на основе платины, выбранной из группы, состоящей из злокачественной опухоли яичника, первичной перитонеальной карциномы и карциномы фаллопиевых труб, включающему введение пациентке антитела против HER2, которое связывается с доменом II HER2, и гемцитабина в количествах, эффективных для лечения злокачественной опухоли.
Краткое описание чертежей
На фиг.1А и 1В представлено картирование эпитопа остатков 22-645 во внеклеточном домене (ECD) HER2 (аминокислотная последовательность, включая сигнальную последовательность, представленная на фиг.1А; SEQ ID NO: 13), по данным анализа усеченния мутанта и сайт-направленного мутагенеза (Nakamura et al., J. of Virology 67(10): 6179-6191 (1993); и Renz et al., J. Cell Biol., 125(6): 1395-1406 (1994)). Различные усечения HER2-ECD или точечные мутации получали из кДНК с использованием технологии полимеразной цепной реакции. Мутанты HER2 экспрессировали в виде гибридных белков gD в экспрессирующей плазмиде млекопитающих. В данной экспрессирующей плазмиде используется промотор/энхансер цитомегаловируса с сигналами терминации и полиаденилирования SV40, расположенными справа (в положении даунстрим) от вставленной кДНК. Плазмидную ДНК трансфектировали в клетки 293. Через 1 сутки после трансфекции в клетки метаболически вводили метку в течение ночи в среде DMEM без метионина и цистеина с низкой концентрацией глюкозы, содержащей 1% диализованной фетальной телячьей сыворотки и 25 мкКи каждого 35S-метионина и 35S-цистеина. Собирали супернатанты и к супернатантам добавляли моноклональные антитела против HER2 или контрольные антитела и инкубировали в течение 2-4 ч при 4°С. Комплексы преципитировали, наносили на 10-20% градиентный гель трицин-SDS и подвергали электрофорезу при 100 В. Гель электрофоретически переносили на мембрану и авторадиографировали. Как показано на фиг.1В, антитела против HER2 7С2, 7F3, 2C4, 7D3, 3Е8, 4D5, 2H11 и 3Р4 связываются с различными эпитопами HER2 ECD.
На фиг.2А и 2В показано влияние моноклональных антител против HER2 2C4 и 7F3 на активацию rHRGβ1 в клетках МCF7. На фиг.2А представлены кривые зависимости для ингибирования под действием 2C4 или 7F3 стимулированного HRG фосфорилирования тирозина. На фиг.2В представлены кривые зависимости для ингибирования связывания 125I-меченого rHRGβ177-244 с клетками МCF7 под действием антител 2C4 или 7F3.
На фиг.3 показано ингибирование специфического связывания 125I-меченого rHRGβ177-244 с панелью человеческих опухолевых клеток под действием моноклональных антител 2C4 или 7F3 против HER2. Моноклональными контрольными антителами являются изотипные мышиные моноклональные антитела, которые не блокируют связывание rHRG. Неспецифическое связывание 125I-меченого rHRGβ177-244 определяли с использованием параллельных инкубационных смесей в присутствии 100 нМ rHRGβ1. Значения неспецифического связывания 125I-меченого rHRGβ177-244 составляло менее 1% от общего значения для всех тестированных клеток.
На фиг.4А и 4В показано влияние моноклональных антител 2C4 или 4D5 на пролиферацию клеток MDA-MB-175 (фиг.4А) и SK-BR-3 (фиг.4В). Клетки MDA-MB-175 и SK-BR-3 высевали в 96-луночные планшеты и давали прикрепиться в течение 2 ч. Опыт проводили в среде, содержащей 1% сыворотки крови. Вносили антитела против HER2 или одну среду и клетки инкубировали в течение 2 ч при 37°С. Затем добавляли rHRGβ1 (1 нМ) или одну среду и клетки инкубировали в течение 4 суток. Монослои промывали и окрашивали/фиксировали 0,5% кристаллическим фиолетовым. Пролиферацию клеток оценивали по поглощению при 540 нм.
На фиг.5А и 5В показано влияние моноклонального антитела 2C4, антитела трастузумаба или анти-EGFR-антитела на зависимую от герегулина (HRG) ассоциацию HER2 с HER3 в клетках MCF7, экспрессирующих низкий/нормальный уровень HER2 (фиг.5А), и клетках SK-BR-3, экспрессирующих высокие уровни HER2 (фиг.5В); смотри пример 2 ниже.
На фиг.6А и 6В приводится сравнение активности интактного мышиного моноклонального антитела 2С4 (mu 2С4) и химерного Fab-фрагмента антитела 2С4. На фиг.6А показано ингибирование связывания 125I-HRG с клетками MCF7 химерным Fab-фрагментом 2С4 или интактным мышиным моноклональным антителом 2С4. Клетки MCF7 высевали в 24-луночные планшеты (1×105 клеток/лунку) и культивировали до 85% слияния в течение 2 суток. Опыты по связыванию проводили, как описано Lewis et al., Cancer Researh, 56: 1457-1465 (1996). На фиг.6В показано ингибирование активации rHRGβ1 фосфорилирования тирозина р180 в клетках MCF7, проводимое, как описано Lewis et al., Cancer Researh, 56: 1457-1465 (1996).
На фиг.7А и 7В представлено сопоставление аминокислотных последовательностей вариабельного домена легкой цепи (VL) (фиг.7А) и вариабельного домена тяжелой цепи (VН) (фиг.7В) мышиного моноклонального антитела 2С4 (соответственно SEQ ID NO 1 и 2); доменов VL и VН гуманизированного варианта 2С4 574 (соответственно SEQ ID NO 3 и 4) и консенсусных каркасных областей человеческих VL и VН (hum κ1, подгруппа I легкой цепи каппа; humIII, подгруппа III тяжелой цепи) (соответственно SEQ ID NO 5 и 6). Звездочки указывают различия между гуманизированным вариантом 2С4 574 и мышиным моноклональным антителом 2С4, или гуманизированным вариантом 2С4 574 и человеческой каркасной областью. В рамках находятся определяющие комплементарность области (CDR).
На фиг.8А-С показано связывание химерного Fab 2С4 (Fab.v1) и нескольких гуманизированных вариантов 2С4 с внеклеточным доменом HER2 (ECD) по данным определения ELISA в примере 3.
Фиг.9 представляет ленточную схему доменов VL и VН моноклонального антитела 2С4 с представленным в белом цвете остовом CDR (L1, L2, L3, H1, H2, H3). Также показаны боковые цепи VН, определенные мутагенезом во время гуманизации (смотри пример 3, таблица 2).
На фиг.10 показано влияние моноклонального антитела 2С4 или трастузумаба на опосредуемую EGR, ТФР-α и HRG активацию митоген-активированной протеинкиназы (МАРК).
На фиг.11А и 11В представлены аминокислотные последовательности соответственно легкой цепи трастузумаба (SEQ ID NO: 14) и тяжелой цепи трастузумаба (SEQ ID NO: 15).
На фиг.12А и 12В представлены аминокислотные последовательности соответственно легкой цепи пертузумаба (SEQ ID NO: 16) и тяжелой цепи пертузумаба (SEQ ID NO: 17).
На фиг.13 схематично показано связывание 2С4 с гетеродимерным связывающим сайтом HER2, посредством чего предупреждается гетеродимеризация с активированными EGFR и HER3.
На фиг.14 показана связь HER2/HER3 с путями МАРК и Akt.
На фиг.15 приводится сравнение активности трастузумаба и пертузумаба.
На фиг.16 схематично представлены различные домены HER2.
Подробное описание предпочтительных вариантов осуществления
I. Определения
"Рецептор HER" представляет рецепторную протеин-тирозинкиназу, которая относится к семейству рецепторов HER и включает рецепторы EGFR, HER2, HER3 и HER4 и другие члены данного семейства, которые будут установлены в будущем. Как правило, рецептор HER содержит внеклеточный домен, который может связываться с лигандом HER; липофильный трансмембранный домен; консервативный внутриклеточный домен тирозинкиназы и С-концевой сигнальный домен, содержащий несколько остатков тирозина, которые могут фосфорилироваться. Рецептор HER может быть рецептором HER с "природной последовательностью" или его "вариантом аминокислотной последовательности". Предпочтительно рецептор HER представляет рецептор HER с природной человеческой последовательностью.
Внеклеточный домен HER2 включает четыре домена, домен I (аминокислотные остатки примерно 1-195), домен II (аминокислотные остатки примерно 196-320), домен III (аминокислотные остатки примерно 321-488) и домен IV (аминокислотные остатки примерно 489-632) (нумерация остатков без сигнального пептида). Смотри Garrett et al., Mol. Cell., 11: 495-505 (2003); Cho et al., Nature, 421: 756-760 (2003); Franklin et al., Cancer Cell., 5: 317-328 (2004) и Рlowman et al., Рroc Natl. Acad. Sci., 90: 1746-1750 (1993), и фиг.16 здесь.
Термины "ErbB1", "HER1", "рецептор эпидермального фактора роста" и "EGFR" используются здесь взаимозаменяемо и относятся к EGFR, раскрытому, например, Carpenter et al. Ann. Rev. Biochem. 56:881-914 (1987), включающему его природные мутантные формы (например, смотри мутантный EGFR как результат делеции у Humphrey et al. PNAS (USA) 87: 4207-4211 (1990)). erbB1 относится к гену, кодирующему белковый продукт, EGFR.
Выражения "ErbB2" и "HER2" используются здесь взаимозаменяемо и относятся к человеческому белку HER2, описанному, например, Semba et al., PNAS (USA), 82: 6497-6501 (1985) и Yamamoto et al., Nature, 319: 230-234 (1986) (номер для доступа в Genеbank X033363). Термин "erbB2" относится к гену, кодирующему человеческий ErbB2, и "neu" относится к гену, кодирующему крысиный р185neu. Предпочтительный HER2 представляет природную человеческую последовательность HER2.
"ErbB3" и "HER3" относятся к рецепторному полипептиду, раскрытому, например, в патентах США № 5183884 и 5480968, а также Kraus et al., PNAS (USA) 86: 9193-9197 (1989).
"ErbB4" и "HER4" относятся здесь к рецепторному полипептиду, раскрытому, например, в заявке на Европейский патент № 599274; Plowman et al., Proc. Natl. Acad. Sci. USA, 90: 1746-1750 (1993); и Plowman et al., Nature, 366: 473-475 (1993), включая его изоформы, например, раскрытые в WO 99/19488, опубликованной 22 апреля 1999 г.
Под "лигандом HER" понимается полипептид, который связывается и/или активирует рецептор HER. Представляющим особый интерес лигандом HER является лиганд HER с человеческой природной последовательностью, такой как эпидермальный фактор роста (EGF) (Savage et al., J. Biol. Chem., 247: 7612-7621 (1972)); трансформирующий фактор роста-альфа (TGF-α) (Marquardt et al., Science, 223: 1079-1082 (1984)); амфирегулин, также известный как шванома или аутокринный фактор роста кератиноцитов (Shoyab et al., Science, 243: 1074-1076 (1989)); Kimura et al., Nature, 348: 257-260 (1990) и Cook et al., Mol. Cell Biol., 11: 2547-2557 (1991)); бетацеллюлин (Shing et al., Science, 259: 1604-1607 (1993)) и Sasada et al., Biochem. Biophys. Res. Commun., 190: 1173 (1993)); гепарин-связывающий эпидермальный фактор роста (HB-EGF) (Higashiyama et al., Science, 251: 936-939 (1991)); эпирегулин (Toyoda et al., J. Biol. Chem., 270: 7495-7500 (1995) и Komurasaki et al., Onkogene, 15: 2841-2848 (1997); герегулин (смотри ниже); нейрегулин-2 (NRG-2) (Carraway et al., Nature, 387: 512-516 (1997); нейрегулин-3 (NRG-3) (Zhang et al., Proc. Natl. Acad. Sci., 94: 9562-9567 (1997); нейрегулин-4 (NRG-4) (Harari et al. Oncogene, 18: 2681-89 (1999) или крипто (CR-1) (Kannan et al. J. Biol. Chem., 272(6): 3330-3335 (1997)). Лиганды HER, которые связываются с EGFR, включают EGF, TGF-α, амфирегулин, бетацеллюлин, НВ-EGF и эпирегулин. Лиганды HER, которые связываются с HER3, включают герегулины. Лиганды, которые способны связываться с HER4, включают бетацеллюлин, эпирегулин, НВ-EGF, NRG-2, NRG-3, NRG-4 и герегулины.
"Герегулин" (HRG) в том смысле, в котором здесь используется данный термин, относится к полипептиду, кодируемому продуктом гена герегулина, раскрытому в патенте США № 5641869 или Marchionni et al., Nature, 362: 312-318 (1993). Примеры герегулинов включают герегулин-α, герегулин-β1, герегулин-β2 и герегулин-β3 (Holmes et al., Science, 256: 1205-1210 (1992) и патент США № 5641869); фактор дифференциации neu (NDF) (Peles et al., Cell, 69: 205-216 (1992)); индуцирующий активность ацетилхолинового рецептора (ARIA) (Falls et al., Cell, 72: 801-815 (1993)); глиальные факторы роста (GGF) (Marchionni et al., Nature, 362: 312-318 (1993)); факторы, полученные из сенсорных и двигательных нейронов (SMDF) (Ho et al., J. Biol. Chem., 270: 14523-14532 (1995)); γ-герегулин (Schaefer et al., Oncogene, 15: 1385-1394 (1997)). Термин включает биологически активные фрагменты и/или варианты аминокислотных последовательностей полипептида HRG с природной последовательностью, такие как EGF-подобные фрагменты его домена (например, HRGβ1177-244).
"Димер HER" представляет нековалентно связанный димер, содержащий, по меньшей мере, два различных рецептора HER. Такие комплексы могут образоваться, когда клетка, экспрессирующая два или более рецепторов HER, подвергается воздействию лиганда HER, и их можно выделить, например, иммунопреципитацией и подвергнуть анализу электрофорезом в SDS-PAGE, как описано Sliwkowski et al., J. Biol. Chem., 269(20): 14661-14665 (1994). Примеры таких димеров HER включают гетеродимеры EGFR-HER2, HER2-HER3 и HER3-HER4. Кроме того, димер HER может включать два или более рецепторов HER2 в комбинации с другим рецептором HER, таким как HER3, HER4 или EGFR. С димером могут быть связаны другие белки, такие как субъединица рецептора цитокина (например, gp 130).
"Гетеродимерный связывающий сайт" в HER относится к области во внеклеточном домене HER2, который контактирует или граничит с областью во внеклеточном домене EGFR, HER3 или HER4 при образовании с ним димера. Данная область обнаружена в домене II HER2. Franklin et al., Cancer Cell, 5: 317-328 (2004).
"Активация HЕR" или "активация HER2" относится к активации или фосфорилированию одного или более рецепторов HЕR или рецепторов HЕR2. Как правило, активация HЕR приводит к передаче сигнала (например, который индуцируется внутриклеточным доменом киназы рецептора HЕR, фосфорилируя остатки тирозина в рецепторе HЕR или полипептиде-субстрате). Активация HЕR может опосредоваться лигандом HЕR, связывающимся с димером HЕR, содержащим интересующий HЕR-рецептор. Связывание лиганда HЕR с димером HЕR может активировать домен киназы одного или более рецепторов HЕR в димере и, тем самым, приводить к фосфорилированию остатков тирозина в одном или более рецепторов HЕR и/или фосфорилированию остатков тирозина в дополнительном субстрате-полипептиде(ах), таком как внутриклеточные киназы Akt или МАРК.
Полипептид с "природной последовательностью" представляет таковой, который имеет такую же аминокислотную последовательность в виде полипептида (например, рецептор HЕR или лиганд HЕR), полученного из природы. Такие полипептиды с природной последовательностью можно выделить из природы или можно получить рекомбинантными или синтетическими путями. Таким образом, полипептид с природной последовательностью может иметь аминокислотную последовательность природного человеческого полипептида, мышиного полипептида или полипептида от любого другого вида млекопитающих.
Термин "вариант аминокислотной последовательности" относится к полипептидам, имеющим аминокислотные последовательности, которые до некоторой степени отличаются от полипептида с природной последовательностью. Как правило, варианты аминокислотных последовательностей будут обладать, по меньшей мере, примерно 70% гомологией, по меньшей мере, с одним доменом связывания рецептора природного лиганда HЕR или, по меньшей мере, с одним доменом связывания лиганда природного рецептора HЕR, и, предпочтительно, они будут, по меньшей мере, на 80%, более предпочтительно, по меньшей мере, примерно на 90% гомологичны с доменами связывания такого рецептора или лиганда. Варианты аминокислотных последовательностей включают замены, делеции и/или вставки в определенных положениях аминокислотной последовательности природной аминокислотной последовательности.
"Гомология" определяется как процент остатков в варианте аминокислотной последовательности, которые являются идентичными после сопоставления последовательностей и введения пробелов, если это необходимо, для достижения максимальной процентной гомологии. Способы и компьютерные программы для сопоставления хорошо известны в данной области. Одной такой компьютерной программой является "Align 2" производства Genentech, Inc., которая была подана с документацией для пользователя в United States Copyright Office, Washington, DC 20559, 10 декабря 1991.
Термин "антитело" в том смысле, в котором он здесь используется, применятся в самом широком смысле, и, в частности, он включает интактные моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифически антитела), образованные, по меньшей мере, из двух интактных антител, и фрагменты антитела, при условии, что они обладают желаемой биологической активностью.
Термин "моноклональное антитело" в том смысле, в котором он здесь используется, относится к антителу, полученному из популяции в основном однородных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными и/или связываются с одним эпитопом, за исключением возможных вариантов, которые могут появиться во время получения моноклонального антитела, где такие варианты, как правило, присутствуют в минорных количествах. В противоположность препаратам поликлональных антител, которые, как правило, включают различные антитела против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты в антигене. Помимо их специфичности моноклональные антитела обладают тем преимуществом, что они не содержат примесей других иммуноглобулинов. Определение "моноклональное" указывает на характер антитела как полученного в основном из однородной популяции антител и не сводится к необходимости получения антитела каким-либо конкретным методом. Например, моноклональные антитела для применения по настоящему изобретению можно получить гибридомным методом, впервые описанным Kohler et al., Nature, 256: 495 (1975), или можно получить методами рекомбинантной ДНК (смотри, например, патент США № 4816567). "Моноклональные антитела" также можно выделить из фаговых библиотек антител с использованием, например, методов, описанных Clackson et al., Nature, 352: 624-628 (1991) и Marks et al., J. Mol. Biol., 222: 581-597 (1991).
Моноклональные антитела здесь, в частности, включают "химерные" антитела, в которых область тяжелой и/или легкой цепи является идентичной или гомологичной с соответствующими последовательностями в антителах, полученных от определенного вида или относящимся к определенному классу или подклассу антител, в то время как остальная часть цепи(ей) идентична или гомологична соответствующим последовательностям в антителах, полученных от другого вида или относящимся к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют желаемую биологическую активность (патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984)). Интересующие химерные антитела здесь включают "приматизированные" антитела, содержащие антигенсвязывающие последовательности вариабельного домена, полученные от примата, не относящегося к человеку (например, низшие узконосые обезьяны, высшие обезьяны и т.д.), и последовательности человеческой константной области.
"Фрагменты антитела" включают участок интактного антитела, предпочтительно содержащий его антигенсвязывающую или вариабельную область. Примеры фрагментов антитела включают фрагменты Fab, Fab', F(ab')2 и Fv; диатела; линейные антитела; молекулы одноцепочечных антител и мультиспецифические антитела, полученные из фрагмента(ов) антитела.
"Интактное антитело" представляет таковое, которое содержит антигенсвязывающую вариабельную область, а также константный домен легкой цепи (CL) и константные домены тяжелой цепи СН1, СН2 и СН3. Константные домены могут представлять собой константные домены с природной последовательностью (например, константные домены с человеческой природной последовательностью) или их вариант аминокислотной последовательности. Предпочтительно интактное антитело обладает одной или более эффекторных функций.
"Эффекторные функции" антитела относятся к таким биологическим активностям, присущим области Fс (Fс-область природной последовательности или Fс-область варианта аминокислотной последовательности) антитела. Примеры эффекторных функций антитела включают связывание C1q; комплемент-зависимую цитотоксичность; связывание Fс-рецептора; антителозависимую клеточноопосредованную цитотоксичность (ADCC); фагоцитоз; даун-регуляцию рецепторов клеточной поверхности (например, рецептора В-клеток; BCR) и т.д.
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей интактные антитела можно подразделить на различные "классы". Существует пять основных классов интактных антител: IgA, IgD, IgE, IgG и IgМ, и некоторые из них можно дополнительно подразделить на "подклассы" (изотипы), например IgG1, IgG2, IgG3, IgG4, IgА1 и IgА2. Константные домены тяжелой цепи, которые соответствуют различным классам антител, обозначают соответственно α, δ, ε, γ и µ. Структуры субъединиц трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
"Антителозависимая клеточноопосредованная цитотоксичность" и "АDCC" относятся к клеточноопосредованной реакции, в которой неспецифические цитотоксические клетки, которые экспрессируют Fс-рецепторы (FсR) (например, природные клетки-киллеры (NK), нейтрофилы и макрофаги), распознают связанное антитело на клетке-мишени и затем вызывают лизис клетки-мишени. Основные клетки для опосредования АDCC, клетки NK, экспрессируют только FсγRIII, в то время как моноциты экспрессируют FсγRI, FсγRII и FсγRIII. Экспрессия FсR на гематопоэтических клетках в обобщенном виде представлена в таблице 3 на странице 464 у Ravetch and Kinet, Annu. Rev. Immunol., 9: 457-92 (1991). Для оценки активности АDCC интересующей молекулы можно провести тест АDCC в условиях in vitro, такой как описан в патентах США № 5500362 или 5821337. Подходящие эффекторные клетки для постановки таких тестов включают мононуклеарные клетки периферической крови (РВМС) и природные клетки-киллеры (NK). Альтернативно или дополнительно активность АDCC интересующей молекулы можно оценить в условиях in vivo на животной модели, такой как раскрыта Clynes et al. PNAS (USA), 95: 652-656 (1998).
"Человеческие эффекторные клетки" представляют лейкоциты, которые экспрессируют один или более FсR и осуществляют эффекторные функции. Предпочтительно клетки экспрессируют, по меньшей мере, FсγRIII и осуществляют эффекторные функции АDCC. Примеры человеческих лейкоцитов, которые опосредуют АDCC, включают мононуклеарные клетки периферической крови (РВМС), природные клетки-киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы; при том, что РВМС и NK являются предпочтительными. Эффекторные клетки можно выделить из их природного источника, например из крови или РВМС, как здесь описано.
Термины "Fс-рецептор" или "FсR" используются для обозначения рецептора, который связывается с Fс-областью антитела. Предпочтительный FсR представляет человеческий FсR с природной последовательностью. Кроме того, предпочтительным FсR является таковой, который связывается с IgG антителом (гамма-рецептор) и включает рецепторы подклассов FсγRI, FсγRII и FсγRIII, в том числе аллельные варианты и альтернативно сплайсированные формы таких рецепторов. Рецепторы FсγRII включают FсγRIIА ("активирующий рецептор") и FсγRIIВ ("ингибирующий рецептор"), которые имеют аналогичные аминокислотные последовательности, в основном различающиеся по их цитоплазматическим доменам. Активирующий рецептор FсγRIIА содержит иммунорецепторный мотив активации на основе тирозина (ITAM) в его цитоплазматическом домене. Ингибирующий рецептор FсγRIIВ содержит иммунорецепторный мотив ингибирования на основе тирозина (ITIM) в его цитоплазматическом домене (смотри обзор Daëron, Annu. Rev. Immunol., 15: 203-234 (1997)). Имеются обзоры по FсR у Ravetch and Kinet, Annu. Rev. Immunol., 9: 457-92 (1991); Capel et al., Immunomethods, 4: 25-34 (1994) и de Haas et al., J. Lab. Clin. Med., 126: 330-41 (1995). Другие FсR, включая таковые, которые будут открыты в будущем, объединяются здесь термином "Fс". Термин также включает неонатальный рецептор, FсRn, который ответственен за передачу материнских IgG плоду (Guyer et al., J. Immunol., 117: 587 (1976) и Kim et al., J. Immunol., 24: 249 (1994)).
"Комплементзависимая цитотоксичность" или "CDD" относится к способности молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента инициируется связыванием первого компонента комплементной системы (C1q) с молекулой (например, антителом), образующей комплекс со "своим" антигеном. Для оценки активации комплемента можно поставить тест CDD, например, описанный Gazzano-Santoro et al., J. Immunol. Methods, 202: 163 (1996).
"Природные антитела" обычно представляют гетеротетрамерные гликопротеины с молекулярной массой примерно 150000 дальтон, состоящие из двух идентичных легких цепей (L) и двух тяжелых цепей (Н). Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как количество дисульфидных связей варьируется среди тяжелых цепей различных изотипов иммуноглобулинов. Между каждой тяжелой и легкой цепью регулярно располагаются внутрицепочечные дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым следует несколько константных доменов. Каждая легкая цепь имеет на одном конце вариабельный домен (VL) и константный домен - на другом. Константный домен легкой цепи сопоставим с первым константным доменом тяжелой цепи, и вариабельный домен легкой цепи сопоставим с вариабельным доменом тяжелой цепи. Полагают, что определенные аминокислотные остатки образуют поверхность раздела между вариабельными доменами легкой цепи и тяжелой цепи.
Термин "вариабельный" относится к факту, что определенные участки вариабельных доменов в значительной степени различаются по последовательности среди антител и используются при связывании и определении специфичности каждого конкретного антитела для его конкретного антигена. Однако вариабельность неравномерно распределена в вариабельных доменах антител. Она концентрируется в трех сегментах, называемых гипервариабельными областями в вариабельных доменах легкой цепи и тяжелой цепи. Более высококонсервативные участки вариабельных доменов называются каркасными областями (FR). Каждый вариабельный домен природной тяжелой и легкой цепей содержит четыре FR, в основном принимающих конфигурацию β-складки, соединенную тремя гипервариабельными областями, которые при объединении образуют петли и в некоторых случаях образуют часть структуры β-складки. Гипервариабельные области в каждой цепи поддерживаются в тесной близости друг к другу через FR и с гипервариабельными областями из другой цепи, внося свой вклад в образование антигенсвязывающего сайта антител (смотри Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institute of Health, Bethеsda, MD (1991)). Константные домены непосредственно не участвуют в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (ADCC).
Термин "гипервариабельная область" в том смысле, в котором он здесь используется, относится к аминокислотным остаткам антитела, которые ответственны за связывание антигена. Как правило, гипервариабельная область содержит аминокислотные остатки из "определяющего комплементарность участка" или "CDR" (например, остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (Н1), 50-65 (Н2) и 95-102 (Н3) в вариабельном домене тяжелой цепи; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethеsda, MD (1991)) и/или таковые остатки из "гипервариабельной петли" (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3) в вариабельном домене тяжелой цепи; Chothia and Lesk, J. Mol. Biol., 196: 901-917 (1987)). Остатки "каркасной области" или "FR" представляют остатки вариабельного домена, иные, чем остатки гипервариабельной области, имеющие определенные здесь значения.
Переваривание антител папаином приводит к образованию двух идентичных антигенсвязывающих фрагментов, называемых фрагментами "Fab", каждый с одним антигенсвязывающим сайтом, и остаточного фрагмента "Fс", название которого отражает его способность к легкой кристаллизации. Обработка пепсином дает фрагмент F(ab')2, который содержит два антигенсвязывающих сайта и по-прежнему способен к перекрестному связыванию с антигеном.
"Fv" представляет минимальный фрагмент антитела, который содержит полный антигенраспознающий и антигенсвязывающий сайт. Данная область состоит из димера вариабельного домена одной тяжелой цепи и одной легкой цепи в тесной, нековалентной связи. Она находится в такой конфигурации, что три гипервариабельных области каждого вариабельного домена взаимодействуют с образованием антигенсвязывающего сайта на поверхности димера VH-VL. Вместе шесть гипервариабельных областей придают антителу антигенсвязывающую специфичность. Однако даже один вариабельный домен (или половина Fv, содержащая только три гипервариабельных области, специфичные для антигена) обладает способностью распознавать и связывать антиген, хотя с более низкой аффинностью по сравнению с целым связывающим сайтом.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков в С-конце домена СН1 тяжелой цепи, включая один или более цистеинов из шарнирной области антитела. Fab'-SH обозначает здесь Fab', в котором остаток(и) цистеина константных доменов несут, по меньшей мере, одну свободную тиольную группу. Фрагменты антитела F(ab')2 первоначально были получены в виде пар Fab'-фрагментов, которые содержали шарнирные цистеины между ними. Также известны другие химические сочетания фрагментов антител.
"Легкие цепи" антител от любого вида позвоночных можно подразделить на два четко различающихся типа, называемых каппа (κ) и лямбда (λ), основываясь на аминокислотных последовательностях их константных доменов.
"Одноцепочечные Fv" или "scFv" фрагменты антитела содержат домены VH и VL антитела, где данные домены находятся в одной полипептидной цепи. Предпочтительно полипептид Fv дополнительно содержит полипептидный линкер между доменами VH и VL, который способствует scFv образовывать желаемую структуру для связывания антигена. Обзор по scFv смотри у Plückthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New-York, pp. 269-315 (1994). Фрагменты scFv антитела против HER2 описаны в WO 93/16185; патенте США № 5571894 и патенте США № 5587458.
Термин "диатела" относится к небольшим фрагментам антитела с двумя антигенсвязывающими сайтами, где данные фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH-VL). При использовании линкера, который является слишком коротким, чтобы имело место объединение двух доменов в одной цепи, домены вынуждены объединяться с комплементарными доменами другой цепи и образуют два антигенсвязывающих сайта. Более подробно диатела описаны, например, в ЕР 404097; WO 93/11161 и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993).
"Гуманизированные" формы не относящихся к человеческим антител (например, грызунов) представляют химерные антитела, которые содержат минимальную последовательность, полученную из нечеловеческого иммуноглобулина. В большинстве случаев гуманизированные антитела представляют человеческие иммуноглобулины (реципиентное антитело), в которых остатки гипервариабельной области реципиента заменены на остатки из гипервариабельной области вида, не относящегося к человеку (донорное антитело), такого как мышь, крыса, кролик или примат, не относящийся к человеку, обладающие желаемой специфичностью, аффинностью и активностью. В некоторых случаях остатки каркасной области (FR) человеческого иммуноглобулина заменены на соответствующие нечеловеческие остатки. Кроме того, гуманизированные антитела могут содержать остатки, которые отсутствуют в реципиентном антителе или в донорном антителе. Подобные модификации проводят для дополнительного повышения функциональной способности антитела. Как правило, гуманизированное антитело будет содержать, по меньшей мере, один и обычно два вариабельных домена, в которых все или в основном все гипервариабельные петли соответствуют таковым в нечеловеческом иммуноглобулине или все или в основном все FR представляют последовательность человеческого иммуноглобулина. Гуманизированное антитело необязательно также будет включать, по меньшей мере, участок константной области иммуноглобулина (Fc), как правило, человеческого иммуноглобулина. Дополнительные детали смотри у Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-329 (1988); и Presta, Curr. Op. Struct. Biol., 2: 593-596 (1992).
Гуманизированные антитела против HER2 включают huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 или трастузумаб (HERCEPTIN®), представленные в таблице 3 патента США № 5821337, специально включенного здесь для сведения; гуманизированные 520С9 (WO 93/21319) и гуманизированные 2С4 антитела, описанные здесь.
Для настоящих целей "трастузумаб" (HERCEPTIN®) и "huMAb4D5-8" относятся к антителу, содержащему аминокислотные последовательности легкой и тяжелой цепи, соответственно SEQ ID NO 14 и 15.
Здесь "пертузумаб" и "OMNITARG™" относятся к антителу, содержащему аминокислотные последовательности легкой и тяжелой цепи, соответственно SEQ ID NO 16 и 17.
"Голое антитело" представляет антитело (имеющее определенные здесь значения), которое не конъюгировано с гетерологичной молекулой, такой как цитотоксическая группа или радиоактивная метка.
"Выделенное" антитело представляет таковое, которое было идентифицировано и выделено и/или отделено от компонента его природной среды. Загрязняющие компоненты его природной среды представляют вещества, которые будут мешать диагностическому или терапевтическому применениям антитела, и они могут включать ферменты, гормоны и другие белковые или небелковые растворенные соединения. В предпочтительных вариантах осуществления антитело будет очищено 1) до более чем 95% по массе антитела по данным метода Лоури и, наиболее предпочтительно, до более чем 99% по массе, 2) до степени, достаточной для получения, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности при использовании спинового чашечного секвенатора или 3) до гомогенности электрофорезом SDS-PAGE в восстанавливающих и невосстанавливающих условиях с использованием Кумасси голубого или предпочтительно окрашивания серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, поскольку, по меньшей мере, один компонент из природной среды антитела будет отсутствовать. Однако, как правило, выделенное антитело будет получено, по меньшей мере, одной стадией очистки.
Антитело против HER2, которое "подавляет димеризацию HER2 более эффективно, чем трастузумаб", представляет таковое, которое уменьшает или исключает образование димеров HER2 более эффективно (например, по меньшей мере, в 2 раза эффективнее) по сравнению с трастузумабом. Предпочтительно такое антитело подавляет димеризацию HER2, по меньшей мере, примерно так же эффективно, как антитело, выбранное из группы, состоящей из мышиного моноклонального антитела 2С4, Fab-фрагмента мышиного моноклонального антитела 2С4, пертузумаба и Fab-фрагмента пертузумаба. Можно оценить подавление димеризации HER при непосредственном исследовании димеров HER2 или при оценке активации HER, или даунстрим передачу сигнала, которая возникает в результате димеризации HER, и/или при оценке связывания антитело-HER2 и т.д. Тесты для скрининга антител, способных подавлять димеризацию HER более эффективно по сравнению с трастузумабом, описаны Agus et al., Cancer Cell 2: 127-137 (2002) и в примерах 1-2 и 4 здесь. Только в качестве примера, можно определить подавление димеризации HER по оценке, например, ингибирования образования димера HER (смотри, например, фиг.1А-В у Agus et al., Cancer Cell, 2: 127-137 (2002) и в примере 2 здесь); уменьшения активации лиганда HER в клетках, которые экспрессируют димеры HER (пример 1 здесь и фиг.2А-В у Agus et al., Cancer Cell 2: 127-137 (2002), например); блокирования связывания лиганда HER с клетками, которые экспрессируют димеры HER (пример 1 здесь и фиг.2Е у Agus et al., Cancer Cell 2: 127-137 (2002), например); подавления роста опухолевых клеток (например, клеток MCF7, MDA-MD-134, ZR-75-1, MD-MB-175, T-47D), которые экспрессируют димеры HER в присутствии (или отсутствии) лиганда HER (пример 1 здесь и фиг.3A-D у Agus et al., Cancer Cell 2: 127-137 (2002), например); ингибирования даунстрим передачи сигнала (например, ингибирования HRG-зависимого фосфорилирования АКТ и ингибирования HRG- или TФРα-зависимого фосфорилирования МАРК) (пример 1 здесь и фиг.2С-D у Agus et al., Cancer Cell, 2: 127-137 (2002), например). Также можно оценить, насколько антитело подавляет димеризацию HER при исследовании связывающего сайта антитела против HER2, например, оценкой структуры или модели, такой как кристаллическая структура антитела, связанного с HER2 (смотри, например, Franklin et al., Cancer Cell 5: 317-328 (2004)).
Антитело против HER2 может "ингибировать HRG-зависимое фосфорилирование АКТ" и/или и ингибировать "HRG- или TФРα-зависимое фосфорилирование МАРК" более эффективно (например, по меньшей мере, в 2 раза эффективнее) по сравнению с трастузумабом (смотри Agus et al., Cancer Cell, 2: 127-137 (2002) и пример 4 здесь, в качестве примера).
Антитело против HER2 может представлять таковое, которое "не ингибирует отщепление эктодомена HER2" (Molina et al., Cancer Res. 61: 4744-4749 (2001)).
Антитело против HER2, которое "связывается с гетеродимерным связывающим сайтом" HER2, связывается с остатками в домене II (и необязательно также связывается с остатками в других доменах внеклеточного домена HER2, таких как домены I и III), и может стерически затруднять, по меньшей мере, в некоторой степени образование гетеродимера HER2-EGFR, HER2-HER3 или HER2-HER4. Franklin et al., Cancer Cell 5: 317-328 (2004) охарактеризовали кристаллическую структуру димера HER2-пертузумаба, депозированного в банке данных RCSB Protein (ID Code IS78), показав примерное антитело, которое связывается с гетеродимерным связывающим сайтом HER2.
Антитело, которое "связывается с доменом II" HER2, связывается с остатками в домене II и необязательно с остатками в другом домене(ах) HER2, таких как домены I и III. Предпочтительно антитело, которое связывается с доменом II, связывается с сочленением между доменами I, II и III HER2.
"Яичник" представляет собой один из двух небольших, миндалевидной формы органов, расположенных с каждой стороны матки у самок.
"Фаллопиевая труба" или "яйцевод" представляет одну из двух тонких труб, ведущих из яичников в матку у самок млекопитающих.
"Брюшина" представляет эпителиальную оболочку, выстилающую полость в организме, такую как брюшная полость.
"Злокачественная опухоль яичника" представляет потенциально угрожающее жизни злокачественное образование, которое развивается в одном или обоих яичниках. К моменту появления симптомов злокачественной опухоли яичника опухоль яичника может стать достаточно большой для того, чтобы опухолевые клетки распространились по брюшной полости. Клетки опухоли яичника, которые вышли за пределы яичников, относятся к метастазирующим опухолям яичника. Опухоли яичника имеют тенденцию распространяться на диафрагму, тонкий кишечник и/или сальник (жировой слой, который покрывает и служит защитной подкладкой для органов в брюшной полости). Также опухолевые клетки могут распространиться на другие органы по лимфатическим сосудам и кровяному руслу. Опухоль яичника для лечения по изобретению включает три основных группы злокачественных новообразований яичника, а именно эпителиальную опухоль, герминальную опухоль и стромальную опухоль.
"Первичная перитонеальная карцинома" относится к злокачественной опухоли, которая развивается в брюшине. Первичная перитонеальная карцинома может быть очень похожа на эпителиальную опухоль яичника в отношении микроскопической картины, симптомов, характера распространения и прогноза. У женщины с удаленными яичниками может развиться первичная перитонеальная карцинома.
"Злокачественная опухоль фаллопиевой трубы" относится к злокачественной опухоли фаллопиевой трубы и/или широкой связки матки.
"Эпителиальная опухоль" развивается в слое клеток кубовидной формы, известных как герминативный эпителий, которые окружают яичники с внешней стороны. Эпителиальные опухоли составляют до 90% от всех злокачественных опухолей яичника.
"Герминальную опухоль" обнаруживают в созревающих яйцеклетках яичника. Герминальные опухоли, которые составляют 3% от всех злокачественных опухолей яичника, наиболее часто развиваются у подростков и молодых женщин.
"Стромальная опухоль" развивается из клеток соединительной ткани, которые поддерживают структуру яичника и которые продуцируют женские половые гормоны, эстроген и прогестерон. Стромальные опухоли составляют 6% от всех злокачественных опухолей яичника.
"Образец опухоли" здесь представляет образец, полученный из или содержащий опухолевые клетки из опухоли пациентки. Примеры образцов опухоли здесь включают, но не ограничиваются этим, биопсийный материал опухоли, циркулирующие опухолевые клетки, циркулирующие белки плазмы крови, асцитную жидкость, первичные клеточные культуры или клеточные линии, полученные из опухолей или проявляющими опухолеподобные свойства, а также законсервированные образцы опухолей, такие как зафиксированные в формалине, залитые в парафин образцы опухолей.
"Ингибирующее рост средство" в том смысле, в котором здесь используется данный термин, относится к соединению или композиции, которые ингибируют рост клетки, в частности экспрессирующей HER опухолевой клетки в условиях in vitro или in vivo. Таким образом, ингибирующее рост средство может представлять таковое, которое существенно снижает процент экспрессирующих HER клеток в S-фазе. Примеры ингибирующих рост средств включают средства, которые блокируют протекание клеточного цикла (в другой точке, чем S-фаза), такие как средства, которые вызывают задержку в фазе G1 и М. Классические блокаторы М-фазы включают алкалоиды растения барвинок (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Такие средства, которые останавливают G1, также вызывают задержку в S-фазе, например ДНК-алкилирующие агенты, такие как тамоксифен, преднизон, дакарбазин, меклоретамин, цисплатин, метотрексат, 5-фторурацил и ара-С. Дополнительную информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1, под заголовком "Cell cycle regulation, oncogenes and antineoplastic drugs" by Murakami et al. (WB Saunders: Philadelphia, 1995), особенно стр. 13.
Примерами "ингибирующих рост" антител являются таковые, которые связываются с HER2 и подавляют рост опухолевых клеток со сверхэкспрессией HER2. Предпочтительные ингибирующие рост антитела против HER2 подавляют рост клеток злокачественной опухоли молочной железы SK-BR-3 в культуре клеток более чем на 20% и, предпочтительно, более чем на 50% (например, в пределах примерно от 50% до 100%) при титре антител примерно от 0,5 до 30 мкг/мл, когда ингибирование роста определяют через 6 суток после воздействия на клетки SK-BR-3 антителом (смотри патент США № 5677171 от 14 октября 1997 г.). Постановка теста ингибирования роста клеток SK-BR-3 более подробно описана в указанном патенте и здесь ниже. Предпочтительное ингибирующее рост антитело представляет гуманизированный вариант мышиного моноклонального антитела 4D5, например трастузумаб.
Антитело, которое "индуцирует апоптоз", представляет таковое, которое индуцирует запрограммированную гибель клеток, что определяют по связыванию аннексина V, фрагментации ДНК, сморщиванию клеток, расширению эндоплазматического ретикулюма, фрагментации клетки и/или образованию мембранных пузырьков (называемых апоптозными телами). Как правило, клетка представляет таковую со сверхэкспрессией рецептора HER2. Предпочтительно клетка является опухолевой клеткой, например клеткой опухоли молочной железы, яичника, желудка, эндометрия, слюнной железы, легких, почки, ободочной кишки, щитовидной железы, поджелудочной железы или мочевого пузыря. В условиях in vitro клетка может представлять собой клетку SK-BR-3, BT474, Calu 3, MDA-MB-453, MDA-MB-361 или SKOV3. Имеются различные методы оценки клеточных событий, связанных с апоптозом. Например, передвижение фосфатидилсерина (PS) можно определить по связыванию аннексина; фрагментацию ДНК можно оценить по наличию ступенчатости ДНК и конденсацию ядра/хроматина наряду с фрагментацией ДНК можно оценить по увеличению количества гиподиплоидных клеток. Предпочтительно антитело, которое вызывает апоптоз, представляет таковое, которое приводит к повышению связывания аннексина примерно в 2-50 раз, предпочтительно, примерно в 5-50 раз и, наиболее предпочтительно, примерно в 10-50 раз по сравнению с необработанными клетками в тесте определения связывания аннексина при использовании клеток ВТ474 (смотри ниже). Примерами антител против HER2, которые индуцируют апоптоз, являются 7С2 и 7F3.
"Эпитоп 2С4" представляет область во внеклеточном домене HER2, с которой связывается антитело 2С4. Для скрининга антител, которые связываются с эпитопом 2С4, можно провести обычный тест перекрестного блокирования, такой как описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). Альтернативно можно провести картирование эпитопа для оценки того, насколько антитело связывается с эпитопом 2С4 HER2 (например, любой один или более остатков в области примерно от 22 до 584 остатка HER2 включительно; смотри фиг.1А-В). Эпитоп 2С4 содержит остатки из домена II во внеклеточном домене HER2. 2С4 и пертузумаб связываются со внеклеточным доменом HER2 по сочленению доменов I, II и III. Franklin et al., Cancer Cell 5: 317-328 (2004).
"Эпитоп 4D5" представляет область во внеклеточном домене HER2, с которой связываются антитело 4D5 (АТСС CRL 10463) и трастузумаб. Данный эпитоп расположен близко к трансмембранному домену HER2 и внутри домена IV HER2. Для скрининга антител, которые связываются с эпитопом 4D5, можно провести обычный тест перекрестного блокирования, такой как описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). Альтернативно можно провести картирование эпитопа для оценки того, насколько антитело связывается с эпитопом 4D5 HER2 (например, любой один или более остатков в области примерно от остатка 529 до примерно остатка 625 включительно; смотри фиг.1А-В).
"Эпитоп 7С2/7F3" представляет область в N-конце, в домене I, внеклеточного домена HER2, с которой связываются антитела 7С2 и/или 7F3 (каждое находится на хранении в АТСС, смотри ниже). Для скрининга антител, которые связываются с эпитопом 7С2/7F3, можно провести обычный тест перекрестного блокирования, такой как описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). Альтернативно можно провести картирование эпитопа для оценки того, насколько антитело связывается с эпитопом 7С2/7F3 HER2 (например, любой один или более остатков в области примерно от остатка 22 до примерно остатка 53 HER2 включительно; смотри фиг.1А-В).
"Лечение" относится к терапевтическому лечению и профилактическим или превентивным мероприятиям. Нуждающиеся в лечении включают тех, у которых уже имеется злокачественная опухоль, а также тех, у кого образование опухоли необходимо предотвратить. Следовательно, пациентке, которой предназначено настоящее лечение, может быть уже поставлен диагноз злокачественной опухоли, или она предрасположена или восприимчива к развитию злокачественной опухоли.
Термин "эффективное количество" относится к количеству лекарственного препарата, эффективному для лечения злокачественной опухоли у пациентки. Эффективное количество лекарственного препарата может снизить количество опухолевых клеток; уменьшить размеры опухоли; подавить (т.е. до некоторой степени замедлить и предпочтительно остановить) инфильтрацию опухолевых клеток в периферические органы; подавить (т.е. до некоторой степени замедлить и, предпочтительно, остановить) метастазирование опухоли; до некоторой степени подавить рост опухоли и/или до некоторой степени ослабить один или более симптомов, связанных со злокачественной опухолью. Лекарственный препарат может до некоторой степени предупреждать рост и/или приводить к гибели имеющихся опухолевых клеток, он может оказывать цитостатическое и/или цитотоксическое действие. Эффективное количество может привести к увеличению выживаемости без прогрессирования заболевания (например, при определении Оценочными критериями ответной реакции для солидных опухолей, RECIST, или изменениями по СА-125), привести к развитию объективной ответной реакции (включая частичную ответную реакцию, PR, или полную ответную реакцию, СR), повысить продолжительность выживаемости в целом и/или ослабить один или более симптомов злокачественного заболевания (например, при оценке по FOSI).
"Выживаемость в целом" относится к пациентке, остающейся живой в течение определенного периода времени, такого как 1 год, 5 лет и т.д., например, со времени постановки диагноза или начала лечения.
"Выживаемость без прогрессирования заболевания" относится к пациентке, остающейся живой, без ухудшения состояния в результате развития злокачественного заболевания.
"Объективная ответная реакция" относится к определяемой ответной реакции, включая полную ответную реакцию (СR) или частичную ответную реакцию (PR).
Под "полной ответной реакцией" или "полной ремиссией" понимается исчезновение всех признаков злокачественной опухоли в ответ на лечение. Это не всегда означает излечение от злокачественного заболевания.
"Частичная ответная реакция" относится к уменьшению размера одной или более опухолей или участков поражения или степени развития опухоли в организме в ответ на лечение.
"Экспрессирующая HER злокачественная опухоль" представляет таковую, содержащую клетки, которые имеют белок HER на их клеточной поверхности.
"Экспрессирующая HER2 злокачественная опухоль" представляет таковую, которая продуцирует достаточные концентрации HER2 на поверхности ее клеток, так, что антитело против HER2 может связываться с ними и оказывать терапевтическое действие в отношении злокачественной опухоли.
Злокачественная опухоль, которая "сверхэкспрессирует" рецептор HER, представляет таковую, которая имеет достоверно более высокие концентрации рецептора HER, такого как рецептор HER2, на их поверхности клеток по сравнению с неопухолевыми клетками такого типа ткани. Такая сверхэкспрессия может быть вызвана амплификацией гена или повышенной транскрипцией или трансляцией. Сверхэкспрессию рецептора HER можно определить в диагностическом или прогностическом тесте при оценке повышенных уровней белка HER, находящегося на поверхности клетки (например, при использовании иммуногистохимического метода; IHC). Альтернативно или дополнительно можно определить уровень нуклеиновой кислоты, кодирующей HER, в клетке, например, флуоресцентной гибридизацией in situ (FISH; смотри WO 98/45479, опубликованную в октябре 1988), саузерн-блоттингом или полимеразной цепной реакцией (ПЦР), такой как количественная ПЦР в режиме реального времени (RT-ПЦР). Также можно исследовать сверхэкспрессию рецептора HER определением "слущивающегося" антигена (например, внеклеточного домена HER) в биологической жидкости, такой как сыворотка крови (смотри, например, патент США № 4933294 от 12 июня 1990 г.; WO 91/05264, опубликованную 18 апреля 1991 г.; патент США № 5401638 от 28 марта 1995 г. и Sias et al., J. Immunol. Methods, 132: 73-80 (1990)). Помимо вышеуказанных тестов специалистам в данной области известны различные методы в условиях in vivo. Например, можно подвергнуть клетки в организме пациентки воздействию антитела, которое необязательно содержит детектируемую метку, например радиоактивный изотоп, и оценить связывание антитела с клетками у пациентки, например, внешним сканированием радиоактивности или анализом биопсийного материала, взятого у пациентки после воздействия антитела.
В противоположность, опухоль, которая "не сверхэкспрессирует рецептор HER", представляет таковую, которая не экспрессирует более высокие, чем нормальные, уровни рецептора HER по сравнению с неопухолевой клеткой такого же типа ткани.
Злокачественная опухоль, которая "сверхэкспрессирует" лиганд HER, представляет таковую, которая продуцирует достоверно более высокие концентрации данного лиганда по сравнению с неопухолевой клеткой такого же типа ткани. Такая сверхэкспрессия может быть вызвана амплификацией гена или повышенной транскрипцией или трансляцией. Сверхэкспрессию лиганда HER можно определить в диагностическом тесте при оценке уровней лиганда (или нуклеиновой кислоты, кодирующей его) у пациентки, например, в биопсийном материале опухоли или различными диагностическими тестами, такими как IHC, FISH, саузерн-блоттинг, ПЦР, или с использованием тестов in vivo.
Термин "цитотоксическое средство" в том смысле, в котором он здесь используется, относится к соединению, которое подавляет или предупреждает функцию клеток и/или вызывает разрушение клеток. Термин предназначен для включения радиоактивных изотопов (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu), химиотерапевтических средств и токсинов, таких как токсины с небольшой молекулой или токсины с ферментативной активностью бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты.
"Химиотерапевтическое средство" представляет химическое соединение, пригодное для лечения злокачественной опухоли. Примеры химиотерапевтических средств включают алкилирующие агенты, такие как тиотепа и циклофосфамид CYTOXAN®; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквуон, метуредопа и уредопа; этиленимины и метиламеламины, включая альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфамид и триметилоломеламин; ацетогенины (особенно буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®; бета-лапахон; лапахол; кольхицины; бетулиновая кислота; камфотецин (включая синтетический аналог топотекан (HYCAMTIN®), СРТ-11 (иринотекан, CAMPTOSAR®), ацетилкамфотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; СС-1065 (включая адозелезин, карзелезин и синтетические аналоги бизелезина); подофиллотоксин; подофиллиновая кислота; тенипозид; криптофицины (особенно криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, меклоретамин, меклоретамин оксид гидрохлорид, мельфалан, новембицин, фенестерин, пренестерин, преднимустин, трофосфамид, урациловый иприт; нитрозмочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, особенно калихеамицин гамма1I и калихеамицин омегаI1 (смотри, например, Agnew, Chem. Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, включая динемицин А; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарзиностатин хромофор и близкие хромопротеиновые антибиотики-хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин ADRIAMYCIN® (включая морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофенольная кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квуламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пуринов, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидинов, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан, тестолактон; антагонисты гормонов надпочечников, такие как аминоглутетимид, митотан, трилостан; восполнитель фолиевой кислоты, такой как фролиниковая кислота; ацеглатон; альдофосфамид гликозид; аминолевулиновая кислота; энилурацил; амсакрин; бестрабуцил; бизантрен; эдатраксат; дефофамин; демеколцин; диазиквуон; элфорнитин; эллиптиний ацетат; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и анзамитоцины; митогуазон; митоксантрон; мопиданмол; нитраерин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазиквуон; 2,2',2''-трихлортриэтиламин; трихотецены (особенно токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин (ELDISINE®, FILDESIN®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-C"); циклофосфамид; тиотепа; таксаны, например паклитаксел TAXCOL® (Bristol-Myers Squibb Oncology, Princeton, N.J.), композиция паклитаксела на основе наночастиц из генно-инженерного альбумина, не содержащая кремофор ABRAXANETM (American Pharmaceutical Patners, Schamberg, Illinois) и доксетаксел ТAXOTERE® (Rhône-Poulenc Rorer, Antony, Франция); хлоранбуцил; гемцитабин (GEMZAR®); 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин (VELBAN®); платина; этопозид (VP-16); ифосфамид; митоксанрон; винкристин (ONCOVIN®); оксалиплатин; лейкововин; винорелбин (NAVELBINE®); новантрон; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных препаратов; а также комбинации двух или более вышеуказанных препаратов, такие как СНОР, сокращенное обозначение комбинированной терапии циклофосфамидом, доксорубицином, винкристином и преднизолоном, и FOLFOX, сокращенное обозначение схемы лечения оксалиплатином (ELOXATIN™) в комбинации с 5-FU и лейкововином.
Также в данное определение включены антигормональные средства, которые действуют для регуляции или ингибирования действия гормонов на опухоли, такие как антиэстрогены и селективные модуляторы рецепторов эстрогенов (SERM), включая, например, тамоксифен (в том числе, тамоксифен NOLVADEX®), ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен FARESTON®; ингибиторы ароматазы, которые ингибируют фермент ароматазу, регулирующую продукцию эстрогенов в надпочечниках, например, такие как 4(5)-имидазолы, аминоглутетимид, мегестрол ацетат MEGASE®, эксеместан AROMASIN®, форместан, фадрозол, ворозол RIVISOR®, летрозол FEMARA® и анастрозол ARIMIDEX®; и антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; а также троксацитабин (аналог нуклеозида цитозина 1,3-диоксолан); антисмысловые олигонуклеотиды, в частности, таковые, которые ингибируют экспрессию генов в сигнальных путях, связанных с аномальной пролиферацией клеток, например, такие как РКС-альфа, Raf, H-Ras и рецептор эпидермального фактора роста (EGF-R); вакцины, например вакцины для генной терапии, такие как вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; PROLEUKIN® rIL-2; ингибитор топоизомеразы 1 LURTOTECAN®; ABARELIX® rmRH; и фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных препаратов.
"Химиотерапевтическое средство антиметаболит" представляет средство, которое в структурном отношении близко к метаболиту, но оно не может использоваться в организме продуктивным образом. Многие химиотерапевтические средства антиметаболиты оказывают отрицательное действие на продукцию нуклеиновых кислот, РНК и ДНК. Примеры химиотерапевтических средств антиметаболитов включают гемцитабин (GEMZAR®), 5-фторурацил (5-FU), капецитабин (XELODATM), 6-меркаптопурин, метотрексат, 6-тиогуанин, пеметрексед, ралтитрексед, арабинозилцитозин ARA-C цитарабин (CYTOSAR-U®), дакарбазин (DTIC-DOME®), азоцитозин, дезоксицитозин, пиридмиден, флударабин (FLUDARA®), кладрабин, 2-дезокси-D-глюкоза и т.д. Предпочтительным химиотерапевтическим средством антиметаболитом является гемцитабин.
"Гемцитабин" или "2'-дезокси-2',2'-дифторцитидин моногидрохлорид (b-изомер)" представляет аналог нуклеозида, проявляет противоопухолевую активность. Эмпирической формулой гемцитабина HCl является С9Н11F2N3O4·HCl. Гемцитабин HCl производится Eli Lilly под торговым названием GEMZAR®.
"Химиотерапевтическое средство на основе платины" включает органическое соединение, которое содержит платину в качестве составной части молекулы. Примеры химиотерапевтических средств на основе платины включают карбоплатин, цисплатин и оксалиплатину.
Термин "химиотерапия препаратами на основе платины" представляет терапию одним или более химиотерапевтическим средством на основе платины, необязательно в комбинации с одним или более другим химиотерапевтическим средством.
Под термином "резистентная к препаратам на основе платины" злокачественная опухоль понимается, что у пациентки с опухолью имело место прогрессирование заболевания при получении химиотерапии препаратами на основе платины (т.е. пациентка является "резистентной к платине"), или у пациентки произошло прогрессирование заболевания в течение 12 месяцев (например, в течение 6 месяцев) после завершения схемы химиотерапии препаратами на основе платины.
В том смысле, в котором здесь используется данный термин, "лекарственный препарат с целенаправленным действием против EGFR" относится к терапевтическому средству, которое связывается с EGFR и необязательно ингибирует активацию EGFR. Примеры таких средств включают антитела и небольшие молекулы, которые связываются с EGFR. Примеры антител, которые связываются с EGFR, включают MAb 579 (ATCC CRL HB 8506), MAb 455 (ATCC CRL HB 8507), MAb 225 (ATCC CRL 8508), MAb 528 (ATCC CRL 8509) (смотри патент США № 4943533, Mendelsohn et al.) и их варианты, такие как химерное антитело 225 (С225 или Cetuximab; ERBUTIX® и реконструированное человеческое антитело 225 (Н225) (смотри WO 96/40210, Imclone Systems Inc.); антитела, которые связываются с мутантным EGFR типа II (патент США № 5212290); гуманизированные и химерные антитела, которые связываются с EGFR, как описано в патенте США № 5891996; и человеческие антитела, которые связываются с EGFR, такие как ABX-EGF (смотри WO 98/50433, Abgenix). Анти-EGFR-антитело можно конъюгировать с цитотоксическим средством, получив, таким образом, иммуноконъюгат (например, смотри ЕР659439А, Merck Patent GmbH). Примеры небольших молекул, которые связываются с EGFR, включают ZD1839 или гефитиниб (IRESSATM; Astra Zeneca), CP-358774 или эрлотиниб HCl (TARCEVATM; Genentech/OSI) и AG1478, AG1571 (SU 5271; Sugen).
"Ингибитор тирозинкиназы" представляет молекулу, которая до некоторой степени ингибирует активность тирозинкиназы, такой как рецептор HER. Примеры таких ингибиторов включают лекарственные препараты с целенаправленным действием против EGFR, упомянутых в предыдущем абзаце, а также ингибитор HER2 с небольшой молекулой, такой как ТАК 165, производства Takeda, двойные ингибиторы HER, такие как ЕКВ-569 (производства Wyeth), которые предпочтительно связываются с EGFR, но ингибируют клетки со сверхэкспрессией HER2 & EGFR, ингибитор тирозинкиназы HER2 и EGFR для приема внутрь GW572016 (производства Glaxo) и PKI-166 (производства Novartis); общие ингибиторы HER, такие как канертиниб (CI-1033; Pharmacia); ингибиторы Raf-1, такие как антисмысловой агент ISIS-5132 производства ISIS Pharmaceuticals, который ингибирует сигнальный путь с участием Raf-1; ингибиторы ТК, без целенаправленного действия на HER, такие как иматиниб мезилат (GleevacTM) производства Glaxo; ингибитор внеклеточной регулируемой МАРК киназы I CI-1040 (производства Pharmacia); хиназолины, такие как PD 153035, 4-(3-хлоранилино)хиназолин; пиридопиримидины; пиримидопиримидины; пирролопиримидины, такие как CGP 59326, CGP 60261 и CGP 62706; пиразолопиримидины, 4-(фениламино)-7Н-пирроло[2,3-d]пиримидины; куркумин (диферулоилметан, 4,5-бис(4-фторанилино)фталимид); тирфостины, содержащие группы нитротиофена; PD-0183805 (Warner-Lamber); антисмысловые молекулы (например, таковые, которые связываются с нуклеиновой кислотой, кодирующей HER); хиноксалины (патент США № 5804396); трифостины (патент США № 5804396); ZD6474 (Astra Zeneca); РТК-787 (Novartis/Schering AG); общие ингибиторы HER, такие как CI-1033 (Pfizer); аффинитак (ISIS 3521; Isis/Lilly); иматиниб мезилат (Gleevac; Novartis); PKI 166 (Novartis); GW2016 (Glaxo SmithKline); CI-1033 (Pfizer); EKB-569 (Wyeth); семаксиниб (Sugen); ZD6474 (AstraZeneca); PTK-787 (Novartis/Schering AG); INC-1C11 (Imclone) или описанные в любой из последующих патентных публикаций: патент США № 5804396; WO 99/09016 (American Cyanimid); WO 98/43960 (American Cyanimid); WO 97/38983 (Warner Lambert); WO 99/06378 (Warner Lambert); WO 99/06396 (Warner Lambert); WO 96/30347 (Pfizer, Inc.); WO 96/33978 (Zeneca); WO 96/3397 (Zeneca) и WO 96/33980 (Zeneca).
"Антиангиогенное средство" относится к соединению, которое блокирует или до некоторой степени оказывает отрицательное влияние на развитие кровеносных сосудов. Антиангиогенный фактор может, например, представлять собой небольшую молекулу или антитело, которые связываются с фактором роста или рецептором фактора роста, принимающим участие в ангиогенезе. Предпочтительным антиангиогенным фактором здесь является антитело, которое связывается с васкулярным эндотелиальным фактором роста (VEGF), такой как бевацизумаб (AVASTIN®).
Термин "цитокин" представляет общий термин для белков, высвобождаемых одной клеточной популяцией, которые оказывают воздействие на другую клетку в качестве межклеточных медиаторов. Примерами таких цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. В число цитокинов входит гормон роста, такой как человеческий гормон роста, N-метиониловый человеческий гормон роста и бычий гормон роста; гормон паращитовидной железы; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (ФСГ), тироидстимулирующий гормон (ТСГ) и лютеинизирующий гормон (ЛГ); фактор роста печени; фактор роста фибробластов; пролактин; плацентарный лактоген; фактор-α и -β некроза опухолей; муллериан-ингибирующее вещество; мышиный связанный с гонадотропином пептид; инхибин; активин; васкулярный эндотелиальный фактор роста; интегрин; тромбопоэтин (ТРО); факторы роста нервов, такие как NGF-β; тромбоцитарный фактор роста; трансформирующие факторы роста (TФР), такие как TФР-α и TФР-β; инсулиноподобный фактор роста I и II; эритропоэтин (ЕРО); остеоиндуктивные факторы; интерфероны, такие как интерферон-α, -β и -γ; колониестимулирующие факторы роста (CSF), такие как макрофагальный CSF (М-CSF); гранулоцитарный-макрофагальный CSF (GM-CSF) и гранулоцитарный CSF (G-CSF); интерлейкины (IL), такие как IL-1, IL-1α, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12; фактор некроза опухолей, такой как TNF-α или TNF-β; и другие полипептидные факторы, включая LIF и набор-лиганд (KL). В том смысле, в котором он здесь используется, термин цитокин включает белки из природных источников или из культуры рекомбинантных клеток и биологически активные эквивалентные варианты цитокинов с природной последовательностью.
II. Получение антител против HER2
Далее следует описание примерных способов получения антител, используемых по настоящему изобретению. Антиген HER2 для использования в целях получения антител может представлять собой, например, растворимую форму внеклеточного домена HER2 или его область, содержащую желаемый эпитоп. Альтернативно клетки, экспрессирующие HER2 на их клеточной поверхности (например, клетки NIH-3T3, трансформированные для сверхэкспрессии HER2; или линия клеток карциномы, таких как клетки SK-BR-3, смотри Stancovski et al., PNAS (USA), 88: 8691-8695 (1991)), можно использовать для получения антител. Специалистам в данной области будут очевидны другие формы HER2, пригодные для получения антител.
i) Поликлональные антитела
Образование поликлональных антител у животных предпочтительно индуцируют многократными подкожными (п/к) или внутрибрюшинными (в/б) инъекциями "своего" антигена и адъюванта. Может быть пригодным конъюгировать "свой" антиген с белком, который является иммуногенным для вида, предназначенного для иммунизации, например гемоцианином из морского моллюска, сывороточным альбумином, бычьим тироглобулином или соевым ингибитором трипсина с использованием бифункционального или дериватизирующего реагента, например малеимидобензоилсульфосукцинимидового эфира (конъюгация идет по остаткам цистеина), N-гидроксисукцинимида (через остатки лизина), глутаральдегида, янтарного ангидрида, SOCl2 или R1N=C=NR, где R и R1 представляют различные алкильные группы.
Животных иммунизируют против антигена, иммуногенных конъюгатов или производных смешением, например, 100 мкг или 5 мкг белка или конъюгата (соответственно для кроликов или мышей) с 3 объемами полного адъюванта Фрейнда и внутрикожным введением раствора во много мест. Через 1 месяц животных иммунизируют повторно 1/5 до 1/10 от первоначального количества пептида или конъюгата в полном адъюванте Фрейнда подкожным введением раствора во много мест. Через 7-14 суток у животных отбирают кровь и анализируют сыворотку для определения титра антител. Животным проводят бустер-иммунизацию до получения плато титра антител. Предпочтительно бустер-иммунизацию животным проводят конъюгатом того же антигена, но конъюгированным с другим белком, и/или с помощью другого реагента для перекрестного сшивания. Также конъюгаты можно получить в культуре рекомбинантных клеток в виде гибридных белков. Также для усиления иммунного ответа подходят агрегирующие агенты, такие как квасцы.
ii) Моноклональные антитела
Моноклональные антитела получают из популяции в основном однородных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных природных мутаций, которые могут иметь место в минорных количествах. Таким образом, определение "моноклональное" указывает на характер антитела, а не на то, что это смесь отдельных антител.
Например, моноклональные антитела можно получить с использованием гибридомного метода, впервые описанного Kohler et al., Nature, 256: 495 (1975), или методами рекомбинантной ДНК (патент США № 4816567).
В гибридомном методе мышь или другое подходящее животное-хозяин, такое как хомяк, иммунизируют, как описано выше, для получения лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфически связываться с белком, используемым для иммунизации. Альтернативно лимфоциты можно иммунизировать в условиях in vitro. Затем лимфоциты гибридизуют с миеломными клетками с использованием реагента для гибридизации, такого как полиэтиленгликоль, с получением гибридомных клеток (Goding, Monoclonal Antibodies: Principles and Practice, рр. 59-103 (Academic Press, 1986)).
Полученные таким образом гибридомные клетки высевают и культивируют в подходящей культуральной среде, которая предпочтительно содержит одно или более веществ, ингибирующих рост или выживаемость негибридных, исходных миеломных клеток. Например, если в исходных миеломных клетках отсутствует фермент гипоксантин-гуанинфосфорибозилтрансфераза (HGPRT или HPRT), то культуральная среда для гибридом, как правило, содержит гипоксантин, аминоптерин и тимидин (среда НАТ), где данные вещества предупреждают рост HGPRT-дефицитных клеток.
Предпочтительными миеломными клетками являются таковые, которые эффективно гибридизуются, поддерживают стабильно высокий уровень продукции антитела отобранными продуцирующими антитело клетками и которые чувствительны к среде, такой как среда НАТ. Среди них предпочтительными миеломными клеточными линиями являются мышиные миеломные линии, такие как полученные из мышиных опухолей МОРС-21 и МРС-11, из Salk Institute Cell Distribution Center, San Diego, California USA, и клетки SP-2 или X63-Ag8-653 из Американской коллекции типовых культур, Rockville, Maryland USA. Также описаны человеческие миеломные и мышиные-человеческие гетеромиеломные клеточные линии для получения человеческих моноклональных антител (Kozbor, J. Immunol., 133: 3001 (1984); и Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, в которой культивируют гибридомные клетки, анализируют на продукцию моноклональных антител, направленных против антигена. Предпочтительно специфичность связывания моноклональных антител, продуцированных гибридомными клетками, определяют иммунопреципитацией или в тесте связывания в условиях in vitro, таком как радиоиммуноанализ (RIA), или твердофазным иммуноферментным анализом (ELISA).
Аффинность связывания моноклонального антитела можно определить, например, анализом Scatchard по Munson et al., Anal. Biochem., 107: 220 (1980).
После идентификации гибридомных клеток, которые продуцируют антитела с желаемой специфичностью, аффинностью и/или активностью, клоны можно субклонировать методами серийных разведений и культивировать обычными методами (Goding, Monoclonal Antibodies: Principles and Practice, pp. 59-103 (Academic Press, 1986)). Подходящие культуральные для этой цели среды включают, например, среду DMEM или RPMI-1640. Кроме того, гибридомные клетки можно культивировать в условиях in vivo в виде асцитных опухолей у животных.
Моноклональные антитела, секретированные субклонами, выделяют из культуральной среды, асцитной жидкости и сыворотки крови обычными методами очистки антител, например, такими как хроматография на протеин А-сефарозе, гидроксиапатите, гель-электрофорез, диализ или аффинная хроматография.
ДНК, кодирующую моноклональные антитела, легко выделить и секвенировать с использованием обычных методик (например, при использовании олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи мышиных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно вставить в экспрессирующие векторы, которыми затем трансфектируют клетки-хозяева, такие как E. coli, клетки обезьяны COS, клетки яичника китайского хомяка (СНО) или миеломные клетки, которые иначе не продуцируют белок антитела, для синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Статьи с обзором по рекомбинантной экспрессии ДНК, кодирующей антитело, в бактериях включают Skerra et al., Curr. Opinion in Immunol., 5: 256-262 (1993) и Plückthun, Immunol. Revs., 139: 151-188 (1992).
В дополнительном варианте осуществления моноклональные антитела или фрагменты антител можно выделить из фаговых библиотек антител, полученных с использованием методов, описанных McCafferty et al., Nature, 348: 552-554 (1990). Clackson et al., Nature, 352: 624-628 (1991) и Marks et al., J. Мol. Biol., 222:581-597 (1991) описывают выделение соответственно мышиных и человеческих антител с использованием фаговых библиотек. В последующих публикациях описано получение человеческих антител с высокой аффинностью (на уровне нМ) перестановкой цепей (Marks et al., Bio/Technology, 10: 779-783 (1992)), а также комбинаторным инфицированием и рекомбинацией в условиях in vivo для конструирования очень крупных фаговых библиотек (Waterhouse et al., Nuc. Acids Res., 21: 2265-2266 (1993)). Таким образом, данные методы представляют жизнеспособные альтернативы для традиционных гибридомных методик для моноклональных антител в целях выделения моноклональных антител.
ДНК также можно модифицировать, например, заменой кодирующей последовательности константных доменов человеческой тяжелой цепи и легкой цепи вместо гомологичных мышиных последовательностей (патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851 (1984)) или ковалентным связыванием с кодирующей иммуноглобулин последовательностью всей или части кодирующей последовательности для неиммуноглобулинового полипептида.
Как правило, такие неиммуноглобулиновые полипептиды заменяют на константные домены антитела или их заменяют на вариабельные домены одного антигенсвязывающего сайта антитела с получением химерного бивалентного антитела, содержащего один антигенсвязывающий сайт, обладающий специфичностью для антигена, и другой антигенобъединяющий сайт, обладающий специфичностью для другого антигена.
iii) Гуманизированные антитела
В данной области описаны способы получения гуманизированных, нечеловеческих антител. Предпочтительно гуманизированное антитело содержит один или более аминокислотных остатков, введенных в него из источника, не относящегося к человеческому. Данные нечеловеческие аминокислотные остатки часто относят к "импортным" остаткам, которые, как правило, взяты из "импортного" вариабельного домена. Гуманизацию в основном можно проводить по методу Winter и соавторов (Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-327 (1988); Verhoeyen et al., Science, 239: 1534-1536 (1988)) заменой последовательностей гипервариабельной области на соответствующие последовательности человеческого антитела. Следовательно, "гуманизированные" антитела являются химерными антителами (патент США № 4816567), в которых в основном минимум чем интактный человеческий вариабельный домен замещен на соответствующую последовательность от видов, не относящихся к человеку. На практике гуманизированные антитела, как правило, являются человеческими антителами, в которых некоторые остатки гипервариабельной области и возможно остатки FR замещены на остатки из аналогичных сайтов в антителах грызунов.
Выбор человеческих вариабельных доменов, обеих легкой и тяжелой цепей, для применения в получении гуманизированных антител является очень важным для снижения антигенности. Согласно методу "наилучшей подгонки" последовательность вариабельного домена антитела грызунов подвергается скринингу против целой библиотеки известных последовательностей человеческих вариабельных доменов. Затем человеческую последовательность, которая наиболее близка к таковой у грызунов, принимают в качестве каркасной области (FR) для гуманизированного антитела (Sims et al., J. Immunol., 151: 2296 (1993); Chothia et al., J. Mol. Biol., 196: 901 (1987)). В другом способе используется определенная каркасная область, полученная из консенсусной последовательности всех человеческих антител определенной подгруппы легкой и тяжелой цепей. Одну и ту же каркасную область можно использовать для нескольких различных гуманизированных антител (Carter et al., Proc. Natl. Acad. Sci. USA, 89: 4285 (1992); Presta et al., J. Immunol., 151: 2623 (1993)).
Также важно, чтобы антитела были гуманизированы с сохранением высокой аффинности для антигена и других полезных биологических свойств. Для достижения данной цели, по предпочтительному способу, гуманизированные антитела получают анализом исходных последовательностей и различных виртуальных гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Обычно имеются трехмерные модели иммуноглобулинов, и они известны специалистам в данной области. Имеются компьютерные программы, которые показывают и воспроизводят возможные трехмерные конформационные структуры выбранных последовательностей-кандидатов иммуноглобулинов. Изучение данных дисплеев позволяет проанализировать вероятную роль остатков в функционировании последовательности-кандидата иммуноглобулина, т.е. проанализировать остатки, которые оказывают влияние на способность иммуноглобулина-кандидата связываться с его антигеном. Таким образом, можно отобрать остатки FR и объединить с реципиентными и "импортными" последовательностями так, чтобы обеспечивалось антитело с желаемыми свойствами, такими как высокая аффинность для целевого антигена(ов). Как правило, остатки гипервариабельной области непосредственно и наиболее существенно влияют на связывание антигена.
В примере 3 ниже описано получение примерных гуманизированных антител против HER2, которые связываются с HER2 и блокируют активацию лиганда HER-рецептора. Гуманизированное антитело, представляющее в данном случае особый интерес, блокирует опосредуемую EGF, TФР-α и/или HRG активацию МАРК в основном так же эффективно, как мышиное моноклональное антитело 2С4 (или его Fab-фрагмент), и/или связывается с НЕR2 в основном так же эффективно, как мышиное моноклональное антитело 2С4 (или его Fab-фрагмент). Гуманизированное антитело может, например, содержать остатки нечеловеческой гипервариабельной области, включенные в человеческий вариабельный домен тяжелой цепи и может дополнительно включать замену каркасной области (FR) в положении, выбранном из группы, состоящей из 69Н, 71Н и 73Н при использовании системы нумерации вариабельных доменов, представленной Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991). В одном варианте осуществления гуманизированное антитело включает замены FR в двух или всех положениях 69Н, 71Н и 73Н.
Примерное интересующее гуманизированное антитело содержит определяющие комплементарность остатки вариабельного домена тяжелой цепи GFTFTDYTMX, где Х предпочтительно представляет D или S (SEQ ID NO: 7); DVNPNSGGSIYNQRFKG (SEQ ID NO: 8) и/или NLGPSFYFDY (SEQ ID NO: 9), необязательно содержащие аминокислотные модификации данных остатков CDR, например, где в результате модификаций в основном сохраняется или повышается аффинность антитела. Например, вариант интересующего антитела может включать примерно от 1 до примерно 7 или примерно 5 аминокислотных замен в вышеуказанных CDR последовательностей вариабельного домена тяжелой цепи. Такие варианты антитела можно получить "созреванием аффинности", например, описанной ниже. Наиболее предпочтительное гуманизированное антитело содержит аминокислотную последовательность вариабельного домена тяжелой цепи SEQ ID NO: 4.
Гуманизированное антитело может включать определяющие комплементарность остатки вариабельного домена легкой цепи KASQDVSIGVA (SEQ ID NO: 10); SASYX1X2X3, где X1 предпочтительно представляет R или L, X2 предпочтительно является Y или Е и X3 предпочтительно представляет T или S (SEQ ID NO: 11) и/или QQYYIYPYT (SEQ ID NO: 12); например, в дополнении к остаткам CDR вариабельного домена тяжелой цепи, описанным в предыдущем абзаце. Такие гуманизированные антитела необязательно включают аминокислотные модификации вышеуказанных остатков CDR, например, где в результате модификаций в основном сохраняется или повышается аффинность антитела. Например, вариант интересующего антитела может включать примерно от 1 до примерно 7 или примерно 5 аминокислотных замен в вышеуказанных CDR последовательностей вариабельного домена легкой цепи. Такие варианты антитела можно получить "созреванием аффинности", например, описанной ниже. Наиболее предпочтительное гуманизированное антитело содержит аминокислотную последовательность вариабельного домена легкой цепи SEQ ID NO: 3.
Настоящая заявка также включает антитела со "зрелой аффинностью", которые связываются с HER2 и блокируют активацию лиганда рецептора HER. Исходное антитело может представлять собой человеческое антитело или гуманизированное антитело, например, таковое, содержащее последовательности вариабельного домена легкой и/или тяжелой цепи соответственно SEQ ID NO: 3 и 4 (т.е. вариант 574). Антитело со "зрелой аффинностью" предпочтительно связывается с рецептором HER2 с аффинностью, превышающей таковую для мышиного антитела 2С4 или варианта 574 (например, примерно в 2-4 раза, примерно в 100 раз или примерно в 1000 раз увеличенной аффинностью, например, при оценке HER2-внеклеточного домена (ECD) с использованием ELISA). Примеры вариабельных остатков CDR тяжелой цепи для замены включают Н28, Н30, Н34, Н35, Н64, Н96, Н99 или комбинации двух или более (например, два, три, четыре, пять, шесть или семь из данных остатков). Примеры остатков CDR вариабельного домена легкой цепи для замены включают L28, L50, L53, L56, L91, L92, L93, L94, L96, L97 или комбинации двух или более (например, два, три, четыре, пять, шесть или семь из данных остатков).
Предусматриваются различные формы гуманизированного антитела или антитела со "зрелой аффинностью". Например, гуманизированное антитело или антитело со "зрелой аффинностью" может представлять фрагмент антитела, такой как Fab, который необязательно конъюгирован с одним или более цитотоксическим(и) средством для получения иммуноконъюгата. Альтернативно гуманизированное антитело или антитело со "зрелой аффинностью" может представлять интактное IgG1 антитело.
iv) Человеческие антитела
В качестве альтернативы гуманизации можно получить человеческие антитела. Например, в настоящее время возможно получить трансгенных животных (например, мышей), которые способны, при иммунизации, продуцировать весь спектр человеческих антител при отсутствии продукции эндогенных иммуноглобулинов. Например, было описано, что гомозиготная делеция гена сочленяющего сегмента тяжелой цепи (JH) у химерных и мутантных мышей зародышевой линии приводит к полному ингибированию продукции эндогенных антител. Перенос спектра генов человеческих иммуноглобулинов зародышевой линии в мутантных мышей зародышевой линии будет приводить к продукции человеческих антител при стимуляции антигеном. Смотри, например, Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggermann et al., Year in Immuno., 7: 33 (1993) и патенты США № 5591669, 5589369 и 5545807.
Альтернативно можно использовать технологию фагового дисплея (McCafferty et al., Nature, 348:552-553 (1990)) для получения человеческих антител и фрагментов антител в условиях in vitro из спектра генов вариабельного домена (V) иммуноглобулинов. По данному методу гены доменов V антитела клонируют в рамке считывания в ген большой или малой оболочки нитчатого бактериофага, такого как М13 или fd, и воспроизводят в виде функциональных фрагментов антитела на поверхности фаговой частицы. Поскольку нитчатая частица содержит копию одноцепочечной ДНК фагового генома, то селекции, основывающиеся на функциональных свойствах антитела, также приводят к отбору гена, кодирующего антитело, проявляющее такие свойства. Таким образом, фаг имитирует некоторые свойства В-клетки. Фаговый дисплей можно осуществить в различных форматах; обзор по ним смотри, например, у Johson, Kevin S. and Chiswell, David J., Current Opinion in Structural Biology, 3: 564-571 (1993). Можно использовать несколько источников сегментов V-гена для фагового дисплея. Clackson et al., Nature, 352: 624-628 (1991) выделили различный спектр анти-оксазолон-антител из небольшой произвольной комбинаторной библиотеки V-генов, полученных из селезенки иммунизированных мышей. Можно сконструировать спектр V-генов от иммунизированных людей-доноров и можно выделить антитела к различному спектру антигенов (включая аутоантигены), в основном следуя методикам, описанным Marks et al., J. Mol. Biol., 222: 581-597 (1997) или Griffith et al., EMBO J., 12:725-734 (1993). Также смотри патенты США № 5565332 и 5573905.
Как описано выше, человеческие антитела также можно получить в условиях in vitro с использованием В-клеток (смотри патенты США № 5567610 и 5229275).
Человеческие антитела против HER2 описаны в патенте США № 5772997 от 30 июня 1998 и WO 97/00271, опубликованной 3 января 1997.
v) Фрагменты антител
Были разработаны различные способы получения фрагментов антител. Традиционно данные фрагменты были получены протеолитическим перевариванием интактных антител (смотри, например, Morimoto et al., Journal of Biochemical and Biophysical Methods, 24: 107-117 (1992) и Brennan et al., Science, 229: 81 (1985)). Однако в настоящее время данные фрагменты можно получить непосредственно с использованием рекомбинантных клеток-хозяев. Например, фрагменты антител можно выделить из фаговых библиотек антител, которые уже обсуждались выше. Альтернативно Fab'-SH-фрагменты можно непосредственно выделить из E. coli и химически связать с F(ab')2-фрагментами (Carter et al., Bio/Teсhnology, 10: 163-167 (1992)). При использовании данного подхода F(ab')2-фрагменты можно выделить непосредственно из культуры рекомбинантных клеток-хозяев. Специалистам в данной области будут очевидны другие методы получения фрагментов антител. В других вариантах осуществления антителом выбора является одноцепочечный Fv-фрагмент (scFv). Смотри WO 93/16185; патент США № 5571894 и патент США № 5587458. Фрагмент антитела также может представлять собой "линейное антитело", например, описанное в патенте США № 5641870. Такие линейные фрагменты антитела могут быть моноспецифическими и биспецифическими.
vi) Биспецифические антитела
Биспецифические антитела представляют антитела, которые обладают специфичностью связывания, по меньшей мере, для двух различных эпитопов. Типичные биспецифические антитела могут связываться с двумя различными эпитопами белка HER2. В других таких антителах сайт связывания с HER2 может быть объединен с сайтом(и) связывания для EGFR, HER3 и/или HER4. Альтернативно плечо HER2 можно объединить с плечом, которое связывается с "запускающей" молекулой на лейкоците, такой как молекула рецептора Т-клетки (например, CD2 или CD3), или Fc-рецепторами для IgG (FcγR), такими как FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16), так, чтобы сфокусировать механизмы клеточной защиты на экспрессирующей HER2 клетке. Также биспецифические антитела можно использовать для расположения цитотоксических средств в клетках, экспрессирующих HER2. Данные антитела содержат HER2-связывающее плечо и плечо, которое связывается с цитотоксическим средством (например, сапонином, анти-интерфероном-α, алкалоидом из растения барвинок, цепью рицина А, метотрексатом или гаптеном радиоактивного изотопа). Биспецифические антитела можно получить в виде полноразмерных антител или фрагментов антител (например, F(ab')2 биспецифических антител).
В WO 96/16673 описывается биспецифическое антитело против HER2/FcγRIII, и в патенте США № 5837234 раскрывается биспецифическое антитело против HER2/FcγRI IDM1 (Osidem). Биспецифическое антитело против HER2/Fcα представлено в WO 98/02463. В патенте США № 5821337 раскрывается биспецифическое антитело против HER2/CD3. MDX-210 является биспецифическим антителом против HER2-FcγRIII.
В данной области известны способы получения биспецифических антител. Традиционное получение полноразмерных биспецифических антител основано на коэкспрессии двух пар иммуноглобулиновых тяжелой цепи-легкой цепи, где две цепи обладают различной специфичностью (Millstein et al., Nature, 305: 537-539 (1983)). В результате произвольного набора тяжелой и легкой цепей иммуноглобулина данные гибридомы (квадромы) продуцируют потенциальную смесь из 10 различных молекул антител, из которых только одно обладает правильной биспецифической структурой. Очистка правильной молекулы, которую обычно проводят стадиями аффинной хроматографии, является довольно трудоемкой, и выход продукта довольно низким. Аналогичные методы раскрыты в WO 93/08829 и у Traunecker et al., EMBO J., 10: 3655-3659 (1991).
Согласно другому подходу вариабельные домены антитела с желаемой специфичностью связывания (сайты объединения антитела-антигена) гибридизуют с последовательностями константного домена иммуноглобулина. Гибридизованный продукт предпочтительно представляет константный домен тяжелой цепи иммуноглобулина, включающий, по меньшей мере, шарнирные области, СН2 и СН3. Предпочтительно иметь первую константную область тяжелой цепи (СН1) с сайтом, необходимым для связывания с легкой цепью, присутствующей, по меньшей мере, в одном гибридизованном продукте. ДНК, кодирующие слитые тяжелые цепи иммуноглобулина и, если желательно, легкую цепь иммуноглобулина, вставляют в отдельные экспрессирующие векторы и котрансфектируют в подходящий организм-хозяин. Это обеспечивает высокую пластичность при подгонке общих участков трех полипептидных фрагментов в вариантах осуществления, когда используются неравные соотношения трех полипептидных цепей в конструкции с обеспечением оптимального выхода. Однако возможно вставить кодирующие последовательности для двух или всех трех полипептидных цепей в один экспрессирующий вектор, когда экспрессия, по меньшей мере, двух полипептидных цепей в равных соотношениях приводит к высокому выходу или когда соотношения не имеют особого значения.
В предпочтительном варианте осуществления данного подхода биспецифические антитела состоят из гибридной тяжелой цепи иммуноглобулина с первой специфичностью связывания в одном плече и гибридной парой тяжелая цепь-легкая цепь иммуноглобулина (с обеспечением второй специфичности связывания) в другом плече. Было установлено, что данная асимметричная структура облегчает отделение желаемого биспецифического соединения от нежелательных комбинаций иммуноглобулиновых цепей, поскольку присутствие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий путь отделения. Данный подход раскрыт в WO 94/04690. Например, дополнительные детали получения биспецифических антител смотри у Suresh et al., Methods in Enzymology, 121: 210 (1986).
Согласно другому подходу, описанному в патенте США № 5731168, можно сконструировать поверхность раздела между парой молекул антител для получения максимального процента гетеродимеров, которые выделяют из культуры рекомбинантных клеток. Предпочтительная поверхность раздела содержит, по меньшей мере, часть домена СН3 константного домена антитела. В данном методе одну или более небольших боковых аминокислотных цепей из поверхности раздела первой молекулы антитела заменяют на более крупные боковые цепи (например, тирозин или триптофан). Создают компенсаторные "полости" идентичного или одинакового размера с крупной боковой цепью(и) на поверхности раздела второй молекулы антитела при замещении крупных аминокислотных боковых цепей на более мелкие (например, аланин или треонин). Это обеспечивает механизм повышения выхода гетеродимера по сравнению с нежелательными конечными продуктами, такими как гомодимеры.
Биспецифические антитела включают перекрестносшитые или "гетероконъюгированные" антитела. Например, одно из антител в гетероконъюгате можно сочетать с авидином, другое - с биотином. Такие антитела, например, были предложены для целенаправленного воздействия на иммунные клетки, а не на нежелательные клетки (патент США № 4676980), и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373 и ЕР 03089). Гетероконъюгированные антитела можно получить любыми методами перекрестного сшивания. Подходящие агенты для перекрестного сшивания хорошо известны в данной области, и они описаны, например, в патенте США № 4676980, наряду с рядом методик поперечного сшивания.
В литературе также описаны методы получения биспецифических антител из фрагментов антител. Например, биспецифические антитела можно получить с использованием химической связи. Brennan et al., Science, 229: 81 (1985) описывают методику, в которой интактные антитела протеолитически расщепляют с получением F(ab')2-фрагментов. Данные фрагменты восстанавливают в присутствии дитиол-комплексообразующего реагента арсенита натрия для стабилизации соседних дитиолов и предупреждения образования межмолекулярных дисульфидных связей. Затем полученные Fab'-фрагменты превращают в производные тионитробензоата (TNB). Затем одно из производных Fab'-TNB вновь превращают в Fab'-тиол восстановлением меркаптоэтиламином и смешивают с эквимолярным количеством другого производного Fab'-TNB с образованием биспецифического антитела. Полученные биспецифические антитела можно использовать в качестве средств для селективной иммобилизации ферментов.
Недавние достижения способствуют непосредственному выделению фрагментов Fab'-SH из E. coli, которые можно объединить химическим путем с образованием биспецифических антител. Shalaby et al., J. Exp. Med., 175: 217-225 (1992) описывают получение молекулы полностью гуманизированного биспецифического антитела F(ab')2. Каждый Fab'-фрагмент отдельно секретируется из E. coli и подвергается прямому химическому сочетанию в условиях in vitro с образованием биспецифического антитела. Полученное таким образом биспецифическое антитело способно связываться с клетками, сверхэкспрессирующими рецептор HER2, и нормальными человеческими Т-клетками, а также "запускать" литическую активность человеческих цитотоксических лимфоцитов в отношении человеческих клеток-мишеней опухоли молочной железы.
Также описаны различные методики получения и выделения фрагментов биспецифического антитела непосредственно из культуры рекомбинантных клеток. Например, были получены биспецифические антитела с использованием лейциновых "застежек". Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992). Пептиды лейциновой "застежки" из белков Fos и Jun связывали с участками Fab' двух различных антител гибридизацией генов. Гомодимеры антител восстанавливали в шарнирной области с образованием мономеров и затем вновь окисляли с образованием гетеродимеров антител. Данный метод также можно использовать для получения гомодимеров антител. Технология получения "диател", описанная Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993), обеспечивает альтернативный механизм получения фрагментов биспецифических антител. Фрагменты включают вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) с помощью линкера, который является слишком коротким, чтобы происходило объединение двух доменов в одной цепи. Следовательно, домены VH и VL одного фрагмента вынуждены объединяться с комплементарными доменами VH и VL другого фрагмента, образуя, таким образом, два антигенсвязывающих сайта. Имеются сообщения о другой стратегии получения фрагментов биспецифических антител при использовании димеров одноцепочечных Fv (sFv). Смотри Gruber et al., J. Immunol., 152: 5368 (1994).
Предусматриваются антитела с более чем двумя валентностями. Например, можно получить триспецифические антитела. Tutt et al., J. Immunol., 147: 60 (1991).
vii) Другие модификации аминокислотной последовательности
Предусматривается модификация(и) аминокислотной последовательности антител против HER2, описанных здесь. Например, может быть желательным улучшить аффинность связывания и/или другие биологические свойства антитела. Варианты аминокислотной последовательности антитела против HER2 получают введением соответствующих нуклеотидных замен в нуклеиновую кислоту антитела против HER2 или пептидным синтезом. Такие модификации включают, например, делеции из и/или вставки в и/или замены остатков в аминокислотных последовательностях антитела против HER2. Проводят любую комбинацию делеции, вставки и замены для их воспроизведения в конечной конструкции, при условии, что конечная конструкция обладает желаемыми характеристиками. Замены аминокислот также могут изменить посттрансляционный процессинг антитела против HER2, такой как изменение количества или положения сайтов гликозилирования.
Пригодный метод для идентификации некоторых остатков или областей антитела против HER2, которые являются предпочтительными положениями для мутагенеза, называется "аланин-санирующим мутагенезом", описанным Cunningham and Wells, Science, 244: 1081-1085 (1989). В данном случае идентифицируют остаток или группу целевых остатков (например, заряженные остатки, такие как arg, asp, his, lys и glu) и замещают на нейтральную или отрицательно заряженную аминокислоту (наиболее предпочтительно аланин или полиаланин) для воздействия на взаимодействие аминокислот с антигеном HER2. Затем данные положения аминокислот, показывающие функциональную чувствительность к заменам, усовершенствуют введением дополнительных или других вариантов в, или вместо, сайты замены. Таким образом, несмотря на то, что сайт для введения вариации аминокислотной последовательности определен заранее, природу самой мутации не требуется определять заранее. Например, для анализа эффективности проведения мутации в данном сайте проводят ala-сканирование или произвольный мутагенез в целевом кодоне или целевой области и экспрессированные варианты антитела против HER2 подвергают скринингу на желаемую активность.
Вставки аминокислотных последовательностей включают амино- и/или карбоксиконцевые гибридные продукты, колеблющиеся по длине от одного остатка до полипептидов, содержащих сотни или более остатков, а также вставки внутри последовательности одного или многих аминокислотных остатков. Примеры концевых вставок включают антитело против HER2 с N-концевым остатком метионина или антитело, гибридизованное с цитотоксическим полипептидом. Другие инсерционные варианты молекулы антитела против HER2 включают гибридизацию N- или С-конца антитела против HER2 с ферментом (например, ADEPT) или полипептидом, который повышает период полураспада антитела в сыворотке крови.
Другим типом варианта является вариант замены аминокислоты. Данные варианты содержат, по меньшей мере, один остаток аминокислоты в молекуле антитела против HER2, замещенный другим остатком. Сайты, представляющие наибольший интерес для мутагенеза замещением, включают гипервариабельные области, но также предусматриваются изменения в FR. Консервативные замены представлены в таблице 1 под заголовком "предпочтительные замены". Если такие замены приводят к изменению биологической активности, то тогда можно ввести более значительные замены, обозначенные в таблице 1 как "типовые замены", или дополнительно описанные ниже в отношении групп аминокислот, и продукты подвергнуть скринингу.

Существенные модификации в биологических свойствах антитела проводят при отборе замен, которые значительно отличаются по их влиянию на сохранение а) структуры полипептидного остова в области замены, например, в виде складчатой или спиральной конформации, b) заряда или гидрофобности молекулы в сайте-мишене или с) объема боковой цепи. Аминокислоты можно объединить в группы по аналогии свойств их боковых цепей (A.L. Lehninger, Biochemistry, second ed., pp. 73-75, Worth Publishers, New York (1975)):
1) неполярные: Ala (A), Val (V), Leu (L), Ile (I), Pro (P), Phe (F), Trp (W), Met (M);
2) незаряженные полярные: Gly (G), Ser (S), Thr (T), Cys (С), Tyr (Y), Asn (N), Gln (Q);
3) кислые: Asp (D), Glu (E);
4) основные: Lys (K), Arg (R), His (H).
Альтернативно, природные остатки можно подразделить на группы на основе общих свойств боковой цепи:
1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
3) кислые: Asp, Glu;
4) основные: His, Lys, Arg;
5) остатки, влияющие на ориентацию цепи: Gly, Pro;
6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены будут включать обмен члена одной из данных групп на другую группу.
Можно также заместить любой остаток цистеина, не участвующий в поддержании правильной конформации антитела против HER2, как правило, на серин для повышения устойчивости к окислению молекулы и предупреждения аберрантного перекрестного сшивания. В противоположность цистеиновую связь(и) можно добавить к антителу для повышения его стабильности (особенно, когда антитело является фрагментом антитела, таким как Fv-фрагмент).
Особенно предпочтительный тип заместительного варианта включает замену одного или более остатков гипервариабельной области исходного антитела (например, гуманизированного или человеческого антитела). Как правило, полученный вариант(ы), отобранный для дальнейшей разработки, будет обладать улучшенными биологическими свойствами по сравнению с исходным антителом, из которого он получен. Подходящий путь для получения таких заместительных вариантов включает "созревание аффинности" с использованием фагового дисплея. Кратко, несколько сайтов гипервариабельной области (например, 6-7 сайтов) подвергают мутагенезу с получением всех возможных аминокислотных замен в каждом сайте. Полученные таким образом варианты антитела воспроизводят моновалентным образом из нитчатых фаговых частиц в виде гибридов с продуктом гена III М13, упакованных в каждой частице. Затем воспроизведенные на фаге варианты подвергают скринингу на их биологическую активность (например, аффинность связывания), как здесь уже было раскрыто. Для идентификации сайтов-кандидатов гипервариабельной области для модификации можно провести аланин-сканирующий мутагенез для идентификации остатков гипервариабельной области, вносящих значительный вклад в связывание антигена. Альтернативно или дополнительно, может быть полезным провести анализ кристаллической структуры комплекса антиген-антитело для идентификации контактных точек между антителом и человеческим HER2. Такие контактные остатки и смежные остатки являются кандидатами для замены с использованием разработанных здесь методик. После получения таких вариантов панель вариантов подвергают скринингу, как здесь описано, и для дальнейшей разработки можно отобрать антитела с наилучшими свойствами, проявленными в одном или более соответствующих тестов.
В другом типе аминокислотного варианта антитела изменяется первоначальный характер гликозилирования антитела. Под изменением понимается делеция одной или более углеводных групп, обнаруженных в антителе, и/или добавление одного или более сайтов гликозилирования, которые отсутствуют в антителе.
Гликозилирование антител, как правило, является N-связанным или О-связанным. N-связанное относится к присоединению углеводной группы к боковой цепи остатка аспарагина. Последовательности трипептида аспарагин-Х-серин и аспарагин-Х-треонин, где Х представляет любую аминокислоту, за исключением пролина, представляют последовательности распознавания для ферментативного присоединения углеводной группы к боковой цепи аспарагина. Таким образом, наличие любой из данных трипептидных последовательностей в полипептиде приводит к образованию потенциального сайта гликозилирования. О-связанное гликозилирование относится к присоединению одного из сахаров, N-ацетилгалактозамина, галактозы или ксилозы, к гидроксиаминокислоте, как правило, серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление сайтов гликозилирования в антитело преимущественно проводят изменением аминокислотной последовательности таким образом, чтобы она содержала одну или более вышеописанных трипептидных последовательностей (для сайтов N-связанного гликозилирования). Изменение можно также осуществить добавлением или заменой одного или более остатков серина или треонина в последовательности исходного антитела (для сайтов О-связанного гликозилирования).
Нуклеиновокислотные молекулы, кодирующие варианты аминокислотных последовательностей антитела против HER2, получают различными методами, известными в данной области. Данные методы включают, но не ограничиваются, выделение из природного источника (в случае вариантов природных аминокислотных последовательностей) или получение олигонуклеотид-опосредованным (или сайт-направленным) мутагенезом, ПЦР-мутагенезом и кассетным мутагенезом ранее полученного варианта или невариантной версии антитела против HER2.
Может быть желательным модифицировать антитело по изобретению в отношении эффекторной функции, например, так, чтобы повысить антигензависимую клеточноопосредованную цитотоксичность (ADCC) и/или комплемензависимую цитотоксичность (CDC) антитела. Этого можно достичь введением одной или более аминокислотных замен в Fc-область антитела. Альтернативно или дополнительно можно ввести остаток(и) цистеина в Fc-область, тем самым обеспечив образование межцепочечных дисульфидных связей. Полученное таким образом гомодимерное антитело может обладать повышенной способностью к интернализации и/или более высокой комплементопосредуемой активностью индукции гибели клеток и антителозависимой клеточной цитотоксичностью (ADCC). Смотри Caron et al., J. Exp. Med., 176: 1191-1195 (1992) и Shopes, B. J. Immunol., 148: 2918-2922 (1992). Также можно получить гомодимерные антитела с повышенной противоопухолевой активностью с использованием гетеробифункциональных перекрестносшивающих агентов, как описано Wolff et al., Cancer Reseach, 53: 2560-2565 (1993). Альтернативно можно сконструировать антитело, которое содержит двойные Fc-области, в результате обладает повышенной способностью к комплементопосредованному лизису и ADCC. Смотри Stevenson et al., Anti-Cancer Drug Design, 3: 219-230 (1989).
Для увеличения периода полураспада антитела в сыворотке крови можно включить в антитело (особенного фрагмент антитела) эпитоп связывания "спасательного" рецептора, как описано, например, в патенте США № 5739277. В том смысле, в котором он здесь используется, термин "эпитоп связывания спасательного рецептора" относится к эпитопу Fc-области в молекуле IgG (например, IgG1, IgG2, IgG3 или IgG4), который ответственен за увеличение периода полураспада молекулы IgG в сыворотке крови.
viii) Скрининг антител с желаемыми свойствами
Выше описаны способы получения антител. Затем можно отобрать антитела с определенными желаемыми биологическими характеристиками.
Для идентификации антитела, которое блокирует активацию лиганда HER-рецептора, можно оценить способность антитела блокировать связывание лиганда HER с клетками, экспрессирующими HER-рецептор (например, при конъюгации с другим HER-рецептором, с которым интересующий HER-рецептор образует гетероолигомер HER). Например, клетки, естественно экспрессирующие или трансфицированные для экспрессии HER-рецепторов гетероолигомера HER, можно инкубировать с антителом и затем подвергнуть воздействию меченого лиганда HER. Затем можно оценить способность антитела против HER2 блокировать связывание лиганда с HER-рецептором в гетероолигомере HER.
Например, ингибирование антителами к HER2 связывания HRG с клетками опухоли молочной железы MCF7 можно провести с использованием монослойных культур MCF7 на льду в 24-луночных планшетах, в основном, как описано в примере 1 ниже. В каждую лунку можно внести моноклональные анти-HER2-антитела и инкубировать в течение 30 мин. Затем можно добавить rHRGβ1177-224, меченный 125I (25 пкМ), и продолжить инкубацию в течение 4-16 ч. Можно получить кривые зависимости концентрация-ответная реакция и рассчитать значение IC50 для интересующего антитела. В одном варианте осуществления антитело, которое блокирует активацию лиганда HER-рецептора, будет иметь значение IC50 для ингибирования связывания HRG с клетками MCF7 в данном тесте, равное примерно 50 нМ или ниже, более предпочтительно, 10 нМ или ниже. В тех случаях, когда антитело является фрагментом антитела, таким как Fab-фрагмент, то значение IC50 для ингибирования связывания HRG с клетками MCF7 в данном тесте может равняться, например, примерно 100 нМ или ниже, более предпочтительно, 50 нМ или ниже.
Альтернативно или дополнительно, можно оценить способность антитела к HER2 блокировать стимулированное лигандом HER фосфорилирование тирозина HER-рецептора в гетероолигомере HER. Например, клетки, эндогенно экспрессирующие HER-рецепторы или трансфектированные для их экспрессии, можно инкубировать с антителом и затем определить активность зависимого от лиганда HER фосфорилирования тирозина с использованием моноклонального анти-фосфотирозин-антитела (которое необязательно конъюгировано с детектируемой меткой). Тест активации рецептора киназы, описанный в патенте США № 5766863, также подходит для оценки активации HER-рецептора и блокирования данной активности антителом.
В одном варианте осуществления можно провести скрининг на антитело, которое ингибирует стимуляцию под действием HRG фосфорилирования тирозина р180 в клетках MCF7, как в основном описано в примере 1 ниже. Например, клетки MCF7 можно высеять в 24-луночные планшеты и в каждую лунку внести моноклональные антитела к HER2 и инкубировать в течение 30 мин при комнатной температуре; затем можно добавить в каждую лунку rHRGβ1177-224 до конечной концентрации 0,2 нМ и продолжить инкубирование в течение 8 мин. Среду можно аспирировать из каждой лунки и реакцию остановить добавлением 100 мкл буфера для проб SDS (5% SDS, 25 мМ DTT и 25 мМ Трис-HCl буфера, рН 6,8). Каждую пробу (25 мкл) подвергают электрофорезу в 4-12% градиентном геле (Novex) и затем электрофоретически переносят на мембрану из поливинилидена дифторида. Можно провести иммуноблоттинг с анти-фосфотирозин-антителами (при концентрации 1 мкг/мл) и количественно определить интенсивность основной реактивной полосы с Mr ≈ 180000 с помощью отражательной денситомерии. Отобранное антитело будет предпочтительно ингибировать стимуляцию HRG фосфорилирования тирозина р180 до уровня, составляющего примерно 0-35% от контроля в данном тесте. Можно построить кривую зависимости концентрация-ответная реакция для стимуляции HRG фосфорилирования тирозина р180 по данным отражательной денситомерии и рассчитать значение IC50 для интересующего антитела. В одном варианте осуществления антитело, которое блокирует активацию лиганда HER-рецептора, будет иметь значение IC50 для стимуляции HRG фосфорилирования тирозина р180 в данном тесте, равное примерно 50 нМ или ниже, более предпочтительно, 10 нМ или ниже. В тех случаях, когда антитело является фрагментом антитела, таким как Fab-фрагмент, то значение IC50 для стимуляции под действием HRG фосфорилирования тирозина р180 в данном тесте может равняться, например, примерно 100 нМ или ниже, более предпочтительно, 50 нМ или ниже.
Также можно оценить ингибирующее рост действие антитела в отношении клеток MDA-MB-175, например, в основном, как описано Schaefer et al., Oncogene, 15: 1385-1394 (1997). По данному тесту клетки MDA-MB-175 можно обработать моноклональным анти-HER2-антителом (10 мкг/мл) в течение 4 суток и окрасить кристаллическим фиолетовым. Инкубирование с антителом против HER2 может выявить ингибирующее рост действие в отношении данной линии клеток, аналогичное проявляемому моноклональным антителом 2С4. В дополнительном варианте осуществления экзогенный HRG не будет существенно "снимать" данное ингибирование. Предпочтительно, чтобы антитело было способно ингибировать пролиферацию клеток MDA-MB-175 в более высокой степени по сравнению с антителом 4D5 (и не обязательно в более высокой степени по сравнению с моноклональным антителом 7F3) как в присутствии, так и в отсутствие HRG.
В одном варианте осуществления интересующее антитело к HER2 может блокировать герегулин-зависимое связывание HER2 с HER3 в клетках МCF7 и SK-BR-3, что определяют коиммунопреципитацией, такой как описана в примере 2, в основном более эффективно по сравнению с моноклональным антителом 4D5 и, предпочтительно, значительно более эффективно по сравнению с моноклональным антителом 7F3.
Для идентификации ингибирующих рост антител против HER2 можно провести скрининг на антитела, которые подавляют рост опухолевых клеток со сверхэкспрессией HER2. В одном варианте осуществления ингибирующее рост антитело выбора способно ингибировать рост клеток SK-BR-3 в культуре клеток примерно на 20-100% и, предпочтительно, примерно на 50-100% при титре антител, равном примерно 0,5-30 мкг/мл. Для идентификации таких антител можно провести тест с клетками SK-BR-3, описанный в патенте США № 5677171. По данному методу клетки SK-BR-3 культивируют в смеси среды F12 и DMEM в соотношении 1:1 с добавлением 10% фетальной телячьей сыворотки, глютамина и смеси пенициллин-стрептомицин. Клетки SK-BR-3 высевают из расчета 20000 клеток на чашку для культивирования диаметром 35 мм (2 мл/чашку диаметром 35 мм). Антитело к HER2 с титром 0,5-30 мкг/мл добавляют на чашку. Через 6 суток подсчитывают количество клеток по сравнению с необработанными клетками с использованием электронного счетчика клеток COULTERTM. Можно отобрать те антитела, которые подавляют рост клеток SK-BR-3 примерно на 20-100% или примерно на 50-100%, в качестве ингибирующих рост антител. Смотри в патенте США № 5677171 методы скрининга на ингибирующие рост антитела, такие как 4D5 и 7F3.
Для отбора антител, индуцирующих апоптоз, имеется тест определения связывания аннексина с использованием клеток BT474. Клетки BT474 культивируют и высевают на чашки, как уже описано в предыдущем абзаце. Затем среду удаляют и заменяют на новую порцию одной среды и среду, содержащую 10 мкг/мл моноклонального антитела. Через 3 суток инкубации монослои промывают PBS и разделяют обработкой трипсином. Затем клетки центрифугируют, ресуспендируют в Ca2+ связывающем буфере и аликвотные порции вносят в пробирки, как уже описывалось выше, для постановки теста определения гибели клеток. Затем в пробирки вносят меченый аннексин (например, аннексин V-FITC) (1 мкг/мл). Пробы можно анализировать с использованием проточного цитометра FACSCANTM и программного обеспечения FACSONVERTTM CellQuest (Becton Dickson). Данные антитела, которые вызывают статистически достоверно более высокие уровни связывания аннексина по сравнению с контролем, отбирают в качестве индуцирующих апоптоз антител. В дополнение к тесту связывания аннексина имеется тест окрашивания ДНК с использованием клеток BT474. Для постановки теста клетки BT474, обработанные интересующим антителом, как описано в двух предыдущих параграфах, инкубируют с 9 мкг/мл HOECHST 33342™ в течение 2 ч при 37°С, затем пробы анализируют на проточном цитометре EPICSELITETM (Coulter Corporation) с использованием программного обеспечения MODFITLTTM (Verity Software House). Антитела, которые вызывают изменение процента подвергшихся апоптозу клеток в 2 раза или выше (и предпочтительно в 3 раза и выше) по сравнению с необработанными клетками (до 100% апоптозных клеток), можно отобрать в качестве вызывающих апоптоз антител при использовании данного теста. Смотри в WO 98/17797 методы скрининга антител, которые индуцируют апоптоз, такие как 7C2 и 7F3.
Для скрининга антител, которые связываются с эпитопом в HER2, связанном интересующим антителом, можно провести обычный тест перекрестного блокирования, такой как описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988), для оценки того, насколько антитела перекрестно блокируют связывание антитела, такого как 2С4 или пертузумаб, с HER2. Альтернативно или дополнительно, можно осуществить картирование эпитопа методами, известными в данной области (смотри, например, фиг.1А и 1В здесь), и/или можно исследовать структуру комплекса антитело-HER2 (Franklin et al., Cancer Cell, 5: 317-328 (2004)) для установления того, какой домен(ы) в HER2 связывается/связываются с антителом.
ix) Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам, содержащим антитело, конъюгированное с цитотоксическим средством, таким как химиотерапевтическое средство, токсин (например, токсин с небольшой молекулой или токсин с ферментативной активностью бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты) или радиоактивный изотоп (т.е. радиоконъюгат).
Химиотерапевтические средства, пригодные для получения таких иммуноконъюгатов, описаны выше. Конъюгаты антитела и одного или более токсинов с небольшой молекулой, таких как калихеамицин, майтанзин (патент США № 5208020), трихотен и СС1065, также включаются здесь.
В одном предпочтительном варианте осуществления изобретения антитело конъюгировано с одной или более молекулами майтанзина (например, примерно от 1 до примерно 10 молекул майтанзина на молекулу антитела). Например, майтанзин можно превратить в May-SS-Me, который можно восстановить в May-SН3 и подвергнуть взаимодействию с модифицированным антителом (Chari et al., Cancer Research, 52: 127-131 (1992)) с получением иммуноконъюгата майтанзиноид-антитело.
Другой интересующий иммуноконъюгат включает анти-HER-антитело, конъюгированное с одной или более молекулами калихеамицина. Семейство антибиотиков калихеамицинов способно приводить к разрывам двухцепочечной ДНК в субпикомолярных концентрациях. Структурные аналоги калихеамицина, которые можно использовать, включают, но не ограничиваются этим, γ1 I, α2 I, α3 I, N-ацетил-γ1 I, PSAG и θI 1 (Hinman et al., Cancer Research, 53: 3336-3342 (1993) и Lоde et al., Cancer Research, 58: 2925-2928 (1998)). Смотри также патенты США № 5714586; 5712374; 5264586 и 5773001, специально включенные здесь для сведения.
Токсины с ферментативной активностью и их фрагменты, которые можно использовать, включают цепь А токсина возбудителя дифтерии, несвязывающиеся активные фрагменты токсина возбудителя дифтерии, цепь эндотоксина А (из Pseudomonas aeruginosa), цепь рицина А, цепь абрина А, цепь модеццина А, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из момордика карантия, курцин, кротин, ингибитор из сапаонарии официналис, гелонин, метогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Например, смотри WO 93/21232, опубликованную 28 октября 1993 г.
Настоящее изобретение дополнительно включает иммуноконъюгат, образованный антителом и соединением с нуклеолитической активностью (например, рибонуклеазой или ДНК-эндонуклеазой, такой как дезоксирибонуклеаза; ДНКаза).
Для получения радиоконъюгатов анти-HER-антител имеются различные радиоактивные изотопы. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu.
Конъюгаты антитела и цитотоксического средства можно получить с использованием различных бифункциональных реагентов для сочетания с белками, такими как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат, иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глютаральдегид), бис-азидопроизводные (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и производные бис-активного фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, можно получить иммунотоксин рицин, как описано Vitetta et al., Science, 238: 1098 (1987). Меченная углеродом14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет пример комплексообразующего реагента для конъюгации радионуклеотида с антителом. Смотри WO 94/11026. Линкер может представлять "отщепляемый линкер", облегчающий высвобождение цитотоксического препарата в клетке. Например, можно использовать лабильный под действием кислоты линкер, чувствительный к пептидазе линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Research, 52: 127-131 (1992)).
Альтернативно можно получить гибридный белок, включающий анти-HER2-антитело и цитотоксическое средство, например, рекомбинантными методами или пептидным синтезом.
В еще одном варианте осуществления антитело можно конъюгировать с "рецептором" (таким как стрептавидин) для направления к опухоли, где конъюгат антитело-рецептор вводят пациентке с последующим удалением несвязанного конъюгата из крови с использованием очищающего агента и затем введением "лиганда" (например, авидина), который конъюгирован с цитотоксическим средством (например, радионуклеотидом).
х) Антителозависимая опосредуемая ферментом терапия пролекарствами (ADEPT)
Антитела по настоящему изобретению также можно использовать в ADEPT при конъюгации антитела с активирующим пролекарство ферментом, который превращает пролекарство (например, пептидное химиотерапевтическое средство, смотри WO 81/01145) в активный противоопухолевый препарат. Смотри, например, WO 88/07378 и патент США № 4975278.
Ферментный компонент иммуноконъюгата, пригодный для ADEPT, включает любой фермент, способный воздействовать на пролекарство таким образом, что оно превращается в его более активную цитотоксическую форму.
Ферменты, которые подходят для применения в способе по изобретению, включают, но не ограничиваются этим, щелочную фосфатазу, пригодную для превращения фосфат-содержащих пролекарств в свободные препараты; арилсульфатазу, пригодную для превращения сульфат-содержащих пролекарств в свободные препараты; цитозиндеаминазу, пригодную для превращения не токсичного 5-фторцитозина в противоопухолевый препарат, 5-фторурацил; протеазы, такие как серратиа-протеаза, термолизин, субтилизин, карбоксипептидазы и катепсины (такие как катепсины В и L), которые пригодны для превращения пептид-содержащих пролекарств в свободные препараты; D-аланилкарбоксипептидазы, пригодные для превращения пролекарств, содержащих заместители на основе D-аминокислоты; углевод-расщепляющие ферменты, такие как β-галактозидаза и нейраминидаза, пригодные для превращения гликозилированных пролекарств в свободные препараты; β-лактамазу, пригодную для превращения препаратов, дериватизированных β-лактамами, в свободные препараты; и пенициллинамидазы, такие как пенициллин-V-амидаза или пенициллин-G-амидаза, пригодные для превращения препаратов, дериватизированных соответственно по атомам азота их аминогруппы феноксиацетильной или фенилацетильной группами, в свободные препараты. Альтернативно антитела с ферментативной активностью, также известные в данной области как "абзимы", можно использовать для превращения пролекарств по изобретению в свободные активные препараты (смотри, например, Massey, Nature, 328: 457-458 (1987)). Конъюгаты антитело-абзим можно получить, как здесь описано, для доставки абзима в популяцию опухолевых клеток.
Ферменты по данному изобретению можно ковалентно связать с анти-HER2-антителами с использованием методик, хорошо известных в данной области, таких как применение гетеробифункциональных перекрестносшивающих реагентов, указанных выше. Альтернативно гибридные белки, содержащие, по меньшей мере, антигенсвязывающую область антитела по изобретению, связанную, по меньшей мере, с функционально активным участком фермента по изобретению, можно сконструировать с использованием методов рекомбинантной ДНК, хорошо известных в данной области (смотри, например, Neuberger et al., Nature, 312: 604-608 (1984)).
xi) Другие модификации антитела
Предусматриваются другие модификации антитела. Например, антитело можно связать с одним из различных небелковых полимеров, например полиэтиленгликолем, полипропиленгликолем, полиоксиалкиленами или сополимерами полиэтиленгликоля и полипропиленгликоля. Антитело также может быть включено в микрокапсулы, полученные, например, методами коацервации или межфазной полимеризацией (например, соответственно микрокапсулы из гидроксиметилцеллюлозы или желатина и микрокапсулы из поли(метилметакрилата)), и в коллоидные системы для доставки лекарственных препаратов (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или макроэмульсии. Такие методы раскрыты в Remington's Pharmaceutical Sciences, 16th edition, Oslo, A., Ed., (1980).
Антитела против HER2, раскрытые здесь, можно также формулировать в виде иммунолипосом. Липосомы, содержащие антитело, готовят способами, известными в данной области, такими как описаны Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA, 77: 4030 (1980); в патентах США № 4485045 и 4544545; и WO 97/38731, опубликованной 23 октября 1997 г. В патенте США № 5013556 раскрыты липосомы с повышенным временем циркуляции в крови.
Особенно пригодные липосомы можно получить обратнофазовым выпариванием с липидной композицией, содержащей фосфатидилхолин, холестерин и ПЭГ-дериватизированный фосфатидилэтаноламин (ПЭГ-РЕ). Липосомы пропускают через фильтры с определенным размером пор с получением липосом с желаемым диаметром. Фрагменты Fab' антитела по настоящему изобретению можно конъюгировать с липосомами, как описано Martin et al., J. Biol. Chem., 257: 286-288 (1982), посредством реакции взаимообмена дисульфида. Химиотерапевтическое средство необязательно находится внутри липосомы. Смотри Gabizon et al., J. National. Cancer Inst., 81(19): 1484 (1989).
III. Фармацевтические композиции
Терапевтические композиции антител, используемых по настоящему изобретению, готовят для хранения смешением антитела, обладающего желаемой степенью чистоты, с необязательными, фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Oslo, A. Ed. (1980)) в виде лиофилизованных композиций или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмоний хлорид; гексаметоний хлорид; бензалконий хлорид; бензетоний хлорид; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; протеины, такие как сывороточный альбумин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; комплексообразующие реагенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-протеин) и/или неионогенные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (ПЭГ). Предпочтительные лиофилизованные композиции анти-HER2-антитела описаны в WO 97/04801, специально включенной здесь для сведения.
Настоящая композиция также может содержать более чем одно активное соединение, как это необходимо для конкретного показания для лечения, предпочтительно таковые с дополнительной активностью, которые не оказывают отрицательного влияния друг на друга. Например, может быть желательным дополнительно обеспечить антитела, которые связываются с EGFR, HER2 (например, антитело, которое связывается с другим эпитопом в HER2), HER3, HER4 или васкулярным эндотелиальным фактором роста (VEGF), в одной композиции. Альтернативно или дополнительно, композиция может дополнительно содержать химиотерапевтическое средство, цитотоксический агент, цитокин, ингибирующее рост средство, антигормональный препарат, препарат с целенаправленным действием против EGFR, антиангиогенный препарат и/или кардиопротектор. Такие молекулы соответственно находятся в комбинации в количествах, эффективных для предназначенной цели.
Активные ингредиенты можно включить в микрокапсулы, приготовленные, например, методами коацервации или межфазной полимеризацией, например соответственно микрокапсулы из гидроксиметилцеллюлозы или желатина и микрокапсулы из поли(метилметакрилата), в коллоидные системы для доставки препаратов (например, липосомы, альбуминовые микрогранулы, микроэмульсии, наночастицы и нанокапсулы) или макроэмульсии. Такие методики раскрыты в Remington's Pharmaceutical Sciences 16th edition, Oslo, A. Ed. (1980).
Можно приготовить препараты с длительным высвобождением. Подходящие примеры препаратов с длительным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где такие матрицы находятся в виде имеющих форму предметов, например это пленки или микрокапсулы. Примеры матриц с длительным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, не подвергающийся деградации этиленвинилацетат, деградируемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPORTTM (инъекционные микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты), и поли-D-(-)-3-гидроксимасляную кислоту.
Композиции для использования в целях введения in vivo должны быть стерильными. Это легко осуществить фильтрованием через мембраны для стерильного фильтрования.
IV. Скрининг пациентов для терапии
По предпочтительному варианту осуществления изобретения у пациента, выбранного для терапии, имеется опухоль с активацией HER (и предпочтительно HER2). В одном варианте осуществления степень активации HER (или HER2) в опухолевых клетках существенно превышает уровень активации данного рецептора в неопухолевых клетках того же типа ткани. Такая интенсивная активация является результатом сверхэкспрессии рецептора HER и/или более высоких по сравнению с нормой уровней лиганда HER, доступного для активации HER-рецептора в опухолевых клетках. Такая интенсивная активация может привести к и/или быть вызвана злокачественным состоянием опухолевой клетки. В некоторых вариантах осуществления злокачественная опухоль подвергается диагностическому или прогностическому тесту для определения того, насколько имеет место амплификация и/или сверхэкспрессия HER-рецептора, что приводит к такой интенсивной активации HER-рецептора. Альтернативно или дополнительно, опухоль можно подвергнуть диагностическому или прогностическому тесту для определения того, насколько имеет место амплификация и/или сверхэкспрессия лиганда HER в опухоли, что связано с интенсивной активацией рецептора. В подгруппе таких опухолей интенсивная активация рецептора может возникнуть в результате функционирования аутокринного стимулирующего пути. Ниже более подробно описаны различные тесты для определения активации HER.
i) Димеры HER
Образцы опухолей можно оценить на присутствие димеров HER, указывающего на активацию HER или HER2. Можно использовать любой метод, известный в данной области, для детектирования димеров HER2, таких как EGFR-HER2, HER2-HER3 в опухолях. Ниже описано несколько предпочтительных методов. С помощью данных методов можно детектировать нековалентные белок-белковые взаимодействия, или они иначе указывают на близость между интересующими белками.
Методы, основанные на иммуноаффинности, такие как иммунопреципитация или ELISA, можно использовать для детектирования димеров HER. В одном варианте осуществления антитела против HER2 используют для иммунопреципитации комплексов, содержащих HER2 из опухолевых клеток, и затем полученный иммунопреципитант зондируют на присутствие EGFR или HER3 иммуноблоттингом. В другом варианте осуществления антитела к EGFR или HER3 можно использовать для стадии иммунопреципитации и затем иммунопреципитант зондировать антителами против HER2. В дополнительном варианте осуществления лиганды HER, специфичные для EGFR, HER3, комплексов EGFR/HER2 или комплексов HER2/HER3, можно использовать для преципитации комплексов, которые затем зондируют на присутствие HER2. Например, лиганды можно конъюгировать с авидином, и комплексы очистить на колонке с биотином.
В других вариантах осуществления, таких как тесты ELISA или методы на основе антитела "сэндвич"-типа, антитела к HER2 иммобилизуют на твердой подложке, подвергают контакту с опухолевыми клетками или лизатом опухолевых клеток, промывают и затем воздействуют антителом против EGFR или HER3. Связывание последнего антитела, которое можно детектировать непосредственно или с помощью вторичного антитела, конъюгированного с детектируемой меткой, указывает на присутствие гетеродимеров. В некоторых вариантах осуществления антитело против EGFR или HER3 иммобилизуют и антитело к HER2 используют на стадии детектирования. В других вариантах осуществления можно использовать лиганды HER вместо или в комбинации с антителами против HER.
Можно также использовать химическое или индуцированное УФ-светом перекрестное сшивание для ковалентного связывания димеров на поверхности живых клеток. Hunter et al., Biochem. J., 320: 847-53. Примеры химических перекрестносшивающих агентов включают дитиобис(сукцинимидил)пропионат (DSP) и 3,3'дитиобис(сульфосукцинимидил)пропионат (DTSSP). В одном варианте осуществления клеточные экстракты из химически сшитых опухолевых клеток анализируют электрофорезом в SDS-PAGE и проводят иммуноблоттинг с антителами против EGFR и/или HER3. Сверхсмещенная полоса соответствующей молекулярной массы наиболее вероятно представляет димеры EGFR-HER2 или HER2-HER3, поскольку HER2 является наиболее предпочтительным партнером для EGFR и HER3. Данный результат можно подтвердить последующей постановкой иммуноблоттинга с антителами к HER2.
Резонансную передачу энергии флуоресценции (FRET) также можно использовать для детектирования димеров EGFR-HER2 или HER2-HER3. С помощью FRET можно детектировать конформационные изменения и белок-белковые взаимодействия в условиях in vivo и in vitro на основе передачи энергии от флуорофора-донора к флуорофору-акцептору. Selvin, Nat. Struct. Biol., 7: 730-34 (2000). Перенос энергии имеет место только, если флуорофор-донор находится в достаточной близости к флуорофору-акцептору. В обычном опыте с FRET два белка или два сайта в одном белке метят различными флуоресцирующими зондами. Один из зондов, донорный зонд, возбуждается до более высокого энергетического состояния под действием внезапного света определенной длины волны. Затем донорный зонд передает его энергию второму зонду, акцепторному зонду, приводя к снижению интенсивности флуоресценции донора и повышению эмиссии флуоресценции акцептора. Для определения степени передачи энергии проводят сравнение интенсивности флуоресценции донора в пробе, меченной донорным и акцепторным зондами, с интенсивностью флуоресценции пробы, меченной только донорным зондом. Необязательно интенсивность акцептора сравнивают с пробами, содержащими донор/акцептор и только акцептор. В данной области известны подходящие зонды, и они включают, например, проходящие через мембрану красители, такие как флуоресцеин и родамин, органические краски, такие как цианиновые красители, и атомы лантанидов. Selvin, выше. Методы и аппаратура для детектирования и определения передачи энергии также хорошо известны в данной области. Selvin, выше.
Методы, основанные на FRET, пригодные для детектирования и определения белок-белковых взаимодействий в отдельных клетках, также известны в данной области. Например, микроскопию на основе резонансной передачи энергии флуоресценции с фотоотбеливанием донора (pbFRET) и микроскопию с визуализацией существования флуоресценции (FLIM) можно использовать для детектирования димеризации рецепторов клеточной поверхности. Selvin, выше; Gadella & Jovin, J. Cell Biol., 129: 1543-58 (1995). В одном варианте осуществления pbFRET используется на клетках либо "в суспензии", либо "in situ" для детектирования и определения образования димеров EGFR-HER2 или HER2-HER3, как описано Nagy et al., Cytometry, 32: 120-131 (1998). С помощью данных методов определяют снижение существования флуоресценции донора за счет передачи энергии. В конкретном варианте осуществления можно использовать метод проточной цитометрии типа Foerster на основе FRET (FCET) для изучения димеризации EGFR-HER2 или HER2-HER3, как описано Nagy et al., выше, и Brockhoff et al., Cytometry, 44: 338-48 (2001).
Предпочтительно FRET используют в комбинации со стандартными иммуногистохимическими методами с использованием метки. Kenworthy, Methods, 24: 289-96 (2001). Например, антитела, конъюгированные с подходящими флуоресцентными красителями, можно использовать в качестве зондов для введения метки в два различных белка. Если белки находятся близко друг от друга, то флуоресцентные краски функционируют в качестве доноров и акцепторов в FRET. Передача энергии детектируется стандартными средствами. Передачу энергии можно детектировать с помощью проточной цитометрии или цифровых микроскопических систем, таких как конфокальная микроскопия или широкопольная флуоресцентная микроскопия, снабженных камерой с зарядной матрицей (CCD).
В одном варианте осуществления настоящего изобретения антитела против HER2 и антитела против EGFR или HER3 непосредственно метят двумя флуорофорами, например, как описано Nagy et al., выше. Опухолевые клетки или лизаты опухолевых клеток контактируют с антителами, меченными различным образом, которые действуют в качестве доноров и акцепторов для FRET, в присутствии димеров EGFR-HER2 или HER2-HER3. Альтернативно, немеченые антитела против HER2 и антитела против EGFR или HER3 используют вместе со вторичными антителами, меченными различным образом, которые служат в качестве доноров и акцепторов. Смотри, например, Brockhoff et al., выше. Передачу энергии детектируют и присутствие димеров определяют, если красители находятся в тесной близости друг от друга.
В других вариантах осуществления лиганды HER-рецептора, которые являются специфическими для HER2 и HER1 или HER3, метят флуоресцентным красителем и используют для постановки FRET.
В еще одних вариантах осуществления настоящего изобретения присутствие димеров на поверхности опухолевых клеток демонстрируется совместной локализацией HER2 с EGFR или HER3 с использованием обычных методов прямой или непрямой иммунофлуоресценции и конфокальной лазерной сканирующей микроскопии. Альтернативно, применяют лазерную сканирующую визуализацию (LSI) для детектирования связывания и совместной локализации HER2 с EGFR или HER3 в формате с высокой пропускной производительностью, таком как микролуночный планшет, как описано Zuck et al., Proc. Natl. Acad. Sci. USA, 96: 11122-27 (1999).
В дополнительных вариантах осуществления присутствие димеров EGFR-HER2 или HER2-HER3 определяют по идентификации ферментативной активности, которая зависит от близости компонентов димера. Антитело к HER2 конъюгируют с одним ферментом и антитело к EGFR или HER3 конъюгируют со вторым ферментом. Вносят первый субстрат для первого фермента, и в результате реакции образуется второй субстрат для второго фермента. Это приводит к взаимодействию с другой молекулой с получением детектируемого соединения, такого как краситель. Присутствие другого химического соединения приводит к разрушению второго субстрата таким образом, что взаимодействие со вторым ферментом не происходит, если только первый и второй ферменты, и таким образом два антитела, находятся в тесной близости друг к другу. В конкретном варианте осуществления опухолевые клетки или лизаты клеток контактируют с антителом против HER2, которое конъюгировано с глюкозооксидазой, и антителом против HER3 или HER1, которое конъюгировано пероксидазой из хрена. В реакционную смесь вносят глюкозу вместе с предшественником красителя, такого как DAB, и каталазой. Присутствие димера определяют по появлению окраски при окрашивании в случае с DAB.
Также димеры можно детектировать с использованием методов, основанных на аналитической системе eTagTM (Aclara Bio Sciences, Mountain View, CA), как описано, например, в заявке на патент США 2001/0049105, опубликованной 6 декабря 2001, оба источника специально включены здесь для сведения. eTagTM или "электрофоретическая метка" содержит детектируемую репортерную группу, такую как флуоресцентная группа. Она также может включать "модификатор подвижности", который в основном состоит из группы, обладающей уникальной электрофоретической подвижностью. Данные группы позволяют отделять и детектировать eTagTM из сложной смеси при определенных электрофоретических условиях, таких как капиллярный электрофорез (СЕ). Участок eTagTM, содержащий репортерную группу, и необязательно модификатор подвижности связан с первой целевой связывающей группой посредством отщепляемой соединяющей группы с получением первого связывающего соединения. Первая целевая связывающая группа специфически распознает определенную первую мишень, такую как нуклеиновая кислота или белок. Первая целевая связывающая группа никоим образом не ограничивается и может представлять, например, полинуклеотид или полипептид. Предпочтительно первая целевая связывающая группа является антителом или фрагментом антитела. Альтернативно первая целевая связывающая группа может быть лигандом HER2-рецептора или его связывающим фрагментом.
Соединяющая группа предпочтительно содержит отщепляемую группу, такую как субстрат фермента, или любую химическую связь, которая в определенных условиях может расщепляться. Когда первая целевая связывающая группа связывается с ее мишенью, то вводится и/или активируется отщепляющий реагент, и соединяющая группа отщепляется, высвобождая, таким образом, участок eTagTM, содержащий репортерную группу и модификатор подвижности. Таким образом, присутствие "свободной" eTagTM указывает на связывание целевой связывающей группы с ее мишенью.
Предпочтительно второе связывающее соединение содержит отщепляющий реагент и вторую целевую связывающую группу, которая специфически распознает вторую мишень. Вторая целевая связывающая группа также никоим образом не ограничивается и может представлять, например, антитело или фрагмент антитела, или лиганд HER-рецептора или связывающий фрагмент лиганда. Отщепляющий реагент является таким, что он будет отщеплять только соединяющую группу в первом связывающем соединении, если первое связывающее соединение и второе связывающее соединение находятся в максимальной близости друг от друга.
В варианте осуществления настоящего изобретения первое связывающее соединение содержит eTagTM, в которой антитело против HER служит в качестве первой целевой связывающей группы. Второе связывающее соединение включает антитело против EGFR или HER3, соединенных с отщепляющим реагентом, способным отщеплять соединительную группу eTagTM. Предпочтительно отщепляющий реагент должен активироваться для того, чтобы быть способным отщепить соединяющую группу. Опухолевые клетки или лизаты опухолевых клеток контактируют с eTagTM, которая связывается с HER2 и с модифицированным антителом против EGFR или HER3, которое связывается с EGFR или HER3 на клеточной поверхности. Не связанное связывающее соединение предпочтительно удаляется и отщепляющий реагент активируется, если это необходимо. Если присутствуют димеры EGFR-HER2 или HER2-HER3, то отщепляющий реагент будет отщеплять соединяющую группу и высвобождать eTagTM за счет близости отщепляющего реагента к соединяющей группе. Затем свободную eTagTM можно детектировать любым известным в данной области методом, таким как капиллярный электрофорез.
В одном варианте осуществления отщепляющий реагент представляет активируемую химическую группу, которая функционирует на соединяющей группе. Например, отщепляющий реагент может быть активирован при воздействии света на пробу.
В другом варианте осуществления eTagTM конструируется с использованием антитела против EGFR или HER3 в качестве первой связывающей группы, и второе связывающее соединение конструируют из антитела против HER2.
В еще одном варианте осуществления димер HER детектируется с использованием антитела или другого реагента, который специфически или предпочтительно связывается с димером по сравнению с его связыванием с рецептором HER в димере.
ii) Фосфорилирование HER2
После иммунопреципитации с антителом EGFR, HER2 или HER3, как уже обсуждалось в предыдущем разделе, может следовать функциональный тест для димеров в качестве альтернативы или дополнения к иммуноблоттингу. В одном варианте осуществления после иммунопреципитации с антителом против HER3 следует тест для определения активности рецепторной тирозинкиназы в иммунопреципитанте. Поскольку HER3 не обладает присущей ему активностью тирозинкиназы, то присутствие активности тирозинкиназы в иммунопреципитанте указывает на то, что HER3 наиболее вероятно связан с HER2. Graus-Porta et al., EMBO J., 16: 1647-55 (1997); Klapper et al., Proc. Natl. Acad. Sci. USA, 96: 4995-5000 (1999). Данный результат может быть подтвержден постановкой иммуноблоттинга с антителами против HER2. В другом варианте осуществления после иммунопреципитации с антителом HER2 следует тест определения активности рецепторной тирозинкиназы EGFR. В данном тесте иммунопреципитант контактирует с радиоактивной АТФ и пептидным субстратом, который имитирует в условиях in vivo сайт транфосфорилирования HER2 с помощью EGFR. Фосфорилирование пептида указывает на коиммунопреципитацию и таким образом димеризацию EGFR с HER2. Тесты определения активности рецепторной тирозинкиназы являются хорошо известными в данной области и включают тесты, с помощью которых детектируют фосфорилирование субстратов-мишеней, например, антителом к фосфотирозину и активацию "своих" путей передачи сигнала, таких как путь с участием МАРК.
Фосфорилирование HER-рецептора можно оценить иммунопреципитацией одного или более HER-рецепторов, таких как HER (HER2) рецептор, и вестерн-блоттингом. Например, о положительных результатах судят по присутствию полосы, соответствующей фосфо-HER2 в геле с использованием анти-фосфотирозин-антитела для детектирования фосфорилированного остатка(ов) тирозина в иммунопреципитированном HER-рецепторе(ах). Анти-фосфотирозин-антитела являются промышленно доступными от Pan Vera (Madison, WI), дочерней компании Invitrogen, Chemicon International Inc. (Temecula, CA) или Upstate Biotechnology (Lake Placid, NY). Об отрицательных результатах судят по отсутствию полосы.
В другом варианте осуществления фосфорилирование HER (HER2) рецептора оценивают иммуногистохимическим методом с использованием специфического анти-фосфо-HER2-антитела (клон PN2A; Thor et al., J. Clin. Oncol., 18(18): 3230-3239 (2000)).
Другие методы детектирования фосфорилирования HER-рецептора(ов) включают, но не ограничиваются, KIRA ELISA (патенты США № 5766863; 5891650; 5914237; 6025145 и 6287784), масс-спектрометрией (сравнение размера фосфорилированного и нефосфорилированного HER2) и тест определения с е-меткой близости с антителом против HER (например, HER2) и фосфо-специфическим или фосфотирозин-специфическим антителом (например, с использованием набора для постановки теста eTagTM производства Aclara BioSciences (Mountain View, CA). Подробности по постановке теста с eTag описаны выше.
Также можно использовать фосфо-специфические антитела в клеточном спектре для оценки состояния фосфорилирования в клеточной пробе белка передачи сигнала (заявка на патент США 2003/0190686).
iii) Лиганды HER2
Можно определить уровни лиганда HER2, такого как TФР-α, в или связанного с опухолью известными методами. С помощью таких методов можно детектировать белок и/или нуклеиновую кислоту, кодирующую его, в пробе, предназначенной для тестирования. В одном варианте осуществления уровни лиганда HER2 в опухоли можно определить с использованием иммуногистохимического метода (IHC); смотри, например, Scher et al., Clin. Cancer Research, 1: 545-500 (1995). Альтернативно или дополнительно можно определить уровни кодирущей лиганд HER2 нуклеиновой кислоты в пробе, предназначенной для тестирования; например, с помощью FISH, саузерн-блоттинга или ПЦР.
iv) Злокачественная опухоль без сверхэкспрессии HER2
Несмотря на то, что для злокачественной опухоли характерна сверхэкспрессия HER2-рецептора, настоящая заявка дополнительно относится к способу лечения злокачественной опухоли, которая не рассматривается как таковая со сверхэкспрессией HER2.
Для определения экспрессии HER2 в злокачественной опухоли имеются различные диагностические/прогностические тесты. В одном варианте осуществления сверхэкспрессию HER2 можно определить IHC, например, с использованием HERCEPTEST® (Dako). Срезы ткани, залитые в парафин, из биопсийной пробы опухоли можно подвергнуть IHC и результаты интенсивности окрашивания на белок HER2 оценить следующим образом:
0 баллов не наблюдается окрашивания или наблюдаются окрашивание мембраны менее чем в 10% опухолевых клеток;
1+ баллов детектируется слабое/едва различимое окрашивание мембраны в более чем 10% опухолевых клеток. Окрашена только часть мембраны клеток;
2+ баллов наблюдается полное окрашивание мембраны по интенсивности от слабого до среднего более чем в 10% опухолевых клеток;
3+ баллов наблюдается полное окрашивание мембраны по интенсивности от среднего до сильного более чем в 10% опухолевых клеток.
Опухоли с баллами 0 или 1+ при оценке сверхэкспрессии HER2 можно охарактеризовать как опухоли без сверхэкспрессии HER2, в то время как опухоли с баллами 2+ или 3+ можно охарактеризовать как опухоли, сверхэкспрессирующие HER2.
Опухоли со сверхэкспрессией HER2 можно оценить в баллах при проведении гистохимических исследований соответственно количеству копий молекул HER2, экспрессированных в расчете на клетку, и можно определить биохимически:
0 = 0-10000 копий/клетку,
1+ = по меньшей мере, примерно 200000 копий/клетку,
2+ = по меньшей мере, примерно 500000 копий/клетку,
3+ = по меньшей мере, примерно 2000000 копий/клетку.
Сверхэкспрессия HER2 на уровне 3+, которая приводит к независимой от лиганда активации тирозинкиназы (Hudziak et al., Proc. Natl. Acad. Sci. USA, 84: 7159-7163 (1987)), имеет место примерно у 30% злокачественных опухолей молочной железы, и у данных пациенток снижены выживаемость без рецедива и выживаемость в целом (Slamon et al., Science, 244: 707-712 (1989); Slamon et al., Science, 235: 177-182 (1987)).
Альтернативно или дополнительно можно проводить тесты FISH, такие как INFORMTM (производства Ventana, Arizona) или PATHVISIONTM (Vysis, Illinois), с зафиксированной в формалине, залитой в парафин опухолевой тканью для определения степени (если таковая имеется) сверхэкспрессии HER2 в опухоли.
В одном варианте осуществления злокачественная опухоль будет таковой, которая экспрессирует (и может сверхэкспрессировать) EGFR, такую экспрессию можно оценить методами, использованными для оценки экспрессии HER2, как указывалось выше.
Сверхэкспрессию или амплификацию рецептора HER2 или лиганда HER2 можно также оценить с использованием диагностического теста в условиях in vivo, например, при введении молекулы (такой как антитело), которая связывается с молекулой, предназначенной для детектирования, и которую метят детектируемой меткой (например, радиоактивным изотопом) и последующем внешнем сканировании пациента с целью установления локализации метки.
V. Лечение антителами против HER2
Предусматривается, что по настоящему изобретению антитела против HER2 можно использовать для лечения резистентной к химиотерапии злокачественной опухоли или резистентной к препаратам на основе платины злокачественной опухоли, такой как опухоль, резистентная к препаратам платины. Злокачественная опухоль, как правило, будет включать экспрессирующие HER2 клетки так, что антитело против HER2 будет способно связываться с опухолевыми клетками.
Пациент подвергается лечению комбинацией антитела против HER2, предпочтительно пертузумаба, и химиотерапевтического средства антиметаболита, предпочтительно гемцитабина. Комбинированное введение включает совместное введение или одновременное введение с использованием отдельных композиций или одной фармацевтической композиции и последовательное введение в любом порядке, где предпочтительно имеется период времени, когда оба (или все) активные средства одновременно проявляют их биологическую активность. Таким образом, химиотерапевтическое средство антиметаболит можно вводить до или после введения антитела к HER2. В данном варианте осуществления период времени, по меньшей мере, между одним введением химиотерапевтического средства антиметаболита и, по меньшей мере, введением антитела против HER2 составляет, предпочтительно, примерно 1 месяц или менее и, наиболее предпочтительно, примерно 2 недели или менее. Альтернативно, химиотерапевтическое средство антиметаболит и антитело против HER2 вводят пациенту одновременно в одной композиции или раздельных композициях.
Лечение комбинацией будет приводить к ослаблению признаков или симптомов злокачественного заболевания. Например, такая терапия может привести к повышению выживаемости (выживаемости в целом и/или выживаемости без прогрессирования заболевания) по сравнению с пациентом, подвергшимся лечению только химиотерапевтическим средством антиметаболитом, и/или может привести к получению объективной клинической ответной реакции (частичной или полной). Кроме того, лечение комбинацией химиотерапевтического средства антиметаболита (например, гемцитабина) и антитела против HER2 (например, пертузумаба) может привести к проявлению синергического или более чем аддитивного терапевтического действия у пациента.
Предпочтительно вводимое антитело к HER2 представляет "голое" антитело. Однако антитело к HER2 можно конъюгировать с цитотоксическим средством. Предпочтительно иммуноконъюгат и/или белок HER2, с которым он связан, интернализуется/интернализуются клеткой, приводя к повышенной терапевтической эффективности иммуноконъюгата в отношении индукции гибели опухолевой клетки, с которой он связывается. В предпочтительном варианте осуществления цитотоксическое средство направлено или оказывает отрицательное действие на нуклеиновую кислоту в опухолевой клетке. Примеры таких цитотоксических средств включают майтанзиноиды, калихеамицины, рибонуклеазы и эндонуклеазы ДНК.
Антитело против HER2 (или иммуноконъюгат антитела против HER2) и химиотерапевтическое средство антиметаболит вводят человеку-пациенту известными способами, такими как внутривенное введение, например, в виде болюса или продолжительной инфузии в течение периода времени, внутримышечным, внутрибрюшинным, интрацереброспинальным, подкожным, внутрисуставным, внутрисиновиальным, интратекальным, пероральным, местным или ингаляционным путями. Антитело против HER2 и химиотерапевтическое средство антиметаболит можно, но без отсутствия необходимости в этом, вводить одним и тем же путем введения. Предпочтительным является внутривенное введение антитела и химиотерапевтического средства антиметаболита.
Для профилактики или лечения заболевания подходящие дозы антитела против HER2 будут зависеть от типа заболевания, которое подвергается лечению, определенного выше, тяжести и течения заболевания, от того, вводят антитело против HER2 с профилактической или лечебной целью, предшествующей терапии, истории болезни пациента и ответной реакции на антитело против HER2 и по усмотрению лечащего врача. Антитело против HER2 соответственно вводят пациенту один раз или несколькими курсами. В зависимости от типа и тяжести заболевания доза примерно от 1 мкг/кг до 50 мг/кг (например, 0,1-20 мг/кг) антитела против HER2 является первоначальной дозировкой-кандидатом для введения пациенту, например, при одном или нескольких введениях или продолжительной инфузией. В одном варианте осуществления продолжительность первоначальной инфузии антитела против HER2 может быть более длительной, чем продолжительность последующих инфузий, например, примерно 90 мин для первоначальной инфузии и примерно 30 мин для последующих инфузий (если хорошо переносится первая инфузия). Предпочтительно дозировка антитела против HER2 будет находиться в пределах примерно от 0,05 мг/кг до примерно 10 мг/кг. Таким образом, пациенту можно вводить одну или более доз, составляющих примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любое их сочетание). Такие дозы можно вводить с перерывом, например каждую неделю или каждые три недели (например, так, чтобы пациент получал примерно от двух до примерно двадцати доз, например, примерно шесть доз антитела против HER2). Можно вводить первоначальную более высокую нагрузочную дозу с последующим введением одной или более низких доз. В одном варианте осуществления антитело против HER2 вводят при нагрузочной дозе, равной примерно 840 мг, с последующим введением примерно 420 мг примерно каждые 3 недели. В другом варианте осуществления антитело против HER2 вводят в дозе, равной примерно 1050 мг, примерно каждые 3 недели.
Химиотерапевтическое средство антиметаболит, как правило, вводят в дозах, известных для него, или их необязательно снижают за счет комбинированного действия лекарственных препаратов или проявления отрицательных побочных эффектов в результате введения химиотерапевтического средства антиметаболита. Можно использовать приготовление и схемы введения для таких химиотерапевтических средств согласно инструкциям производителя, или их могут определить эмпирически специалисты в данной области. Приготовление и схемы введения для такой химиотерапии также описаны в Chemotherapy Service Ed., M. C. Perry, Williams & Wilkins, Baltimore, MD (1992). В тех случаях, когда химиотерапевтическое средство антиметаболит представляет гемцитабин, то предпочтительно вводить его в дозе в пределах примерно от 600 мг/м2 до 1250 мг/м2 (например, примерно 1000 мг/м2), например, на 1 и 8 сутки 3-недельного курса.
Помимо антитела против HER2 и химиотерапевтического средства антиметаболита можно вводить в комбинации другие терапевтические средства. Например, можно вводить второе (третье, четвертое и т.д.) химиотерапевтическое средство(а), где второе химиотерапевтическое средство представляет любое другое, иное химиотерапевтическое средство антиметаболит или химиотерапевтическое средство, которое не является антиметаболитом. Например, второе химиотерапевтическое средство может представлять собой таксан (такой как паклитаксел или доцетаксел), капецитабин или химиотерапевтическое средство на основе платины (такое как карбоплатин, цисплатин или оксалиплатин), антрациклин (такой как доксорубицин, включая липосомальный доксорубицин), топотекан, пеметрексед, алкалоид растения барвинок (такой как винорелбин) и TLK 286. Можно вводить "коктейли" различных химиотерапевтических средств.
Другие терапевтические средства, которые можно комбинировать с антителом против HER2 и химиотерапевтическим средством антиметаболитом, включают одно или более из следующего: второе другое антитело против HER2 (например, ингибирующее рост антитело против HER2, такое как трастузумаб, или антитело против HER2, которое индуцирует апоптоз клеток со сверхэкспрессией HER2, такое как 7С2, 7F3 или их гуманизированные варианты); второе антитело, направленное против другого связанного с опухолью антигена, такого как EGFR, HER3, HER4; антигормональное соединение, например антиэстрогенное соединение, такое как тамоксифен, или ингибитор ароматазы; кардиопротектор (для профилактики или ослабления любой дисфункции миокарда, связанной с терапией); цитокин; направленный против EGFR лекарственный препарат (такой как TARCEVA®, IRESSA® или цетуксимаб); антиангиогенное средство (особенно бевацизумаб производства Genentech под торговой маркой AVASTIN®); ингибитор тирозинкиназы; ингибитор СОХ (например, ингибитор СОХ-1 или СОХ-2); нестероидный противовоспалительный препарат, целекоксиб (CELEBREX®); ингибитор фарнезилтрансферазы (например, типифарниб/ZARNESTRA® R115777 производства Johnson and Johnson или лонафарниб SCH66336 производства Schering-Plough); антитело, которое связывается с онкофетальным белком СА 125, такое как ореговомаб (MoAb B43.13); вакцина против HER2 (такая как вакцина HER2 Auto Vac производства Pharmexia, или белковая вакцина APC8024 производства Dendreon, или пептидная вакцина производства GSK/Corixa); другая целенаправленная на HER терапия (например, трастузамаб, цетуксимаб, гефитиниб, эрлотиниб, CI1033, GW2016 и т.д.); ингибитор Raf и/или ras (смотри, например, WO 2003/86467); топетекан; таксан; GW572016; TLK286; EMD-7200; лекарственный препарат для лечения тошноты, такой как антагонист серотонина, стероид или бензодиазепин; лекарственный препарат для профилактики или лечения кожных высыпаний или обычная терапия воспалительных поражений сальных желез, включая антибиотики для местного применения или приема внутрь; лекарственный препарат для снижения температуры, такой как ацетаминофен, дифенгидрамин или меперидин; гематопоэтический фактор роста и т.д.
Подходящие дозы для каждого из совместно вводимых средств представляют таковые, в которых они применяются в настоящее время, или дозы могут быть снижены за счет комбинированного действия (синергии) средства и антитела против HER2.
В дополнении к вышеуказанным терапевтическим схемам пациента можно подвергнуть хирургическому удалению опухолевых клеток и/или лучевой терапии.
Помимо введения белка антитела пациенту настоящая заявка предусматривает введение антитела генной терапией. Такое введение нуклеиновой кислоты, кодирующей антитело, входит в значение выражения "введение антитела". Смотри, например, WO 96/07321, опубликованную 14 марта 1996 г., касающуюся применения генной терапии для получения внутриклеточных антител.
Существует два основных подхода для доставки нуклеиновой кислоты (необязательно входящей в вектор) в клетки пациента в условиях in vivo и ex vivo. Для доставки в условиях in vivo нуклеиновую кислоту непосредственно вводят пациенту, обычно в место, где необходимо антитело. Для лечения ex vivo клетки пациента выделяют, нуклеиновую кислоту вводят в данные выделенные клетки и модифицированные клетки вводят пациенту непосредственно или, например, инкапсулируют в пористые мембраны, которые имплантируют пациенту (смотри, например, патенты США № 4892538 и 5283187). Имеются различные методики, подходящие для введения нуклеиновых кислот в живые клетки. Методики различаются в зависимости от того, вводят нуклеиновую кислоту в культивированные клетки в условиях in vitro или in vivo в клетки предназначаемого хозяина. Методики, подходящие для переноса нуклеиновой кислоты в клетки млекопитающих в условиях in vitro, включают применение липосом, электропорацию, микроинъекцию, слияние клеток, DEAE-декстрановый метод, метод осаждения фосфатом кальция и т.д. Широко используемым вектором для доставки гена ex vivo является ретровирус.
Предпочтительные в настоящее время методы доставки нуклеиновой кислоты в условиях in vivo включают трансфекцию вирусными векторами (такими как аденовирус, вирус герпеса простого I или адено-связанный вирус) и системы на основе липидов (пригодные липиды для опосредованного липидами переноса гена являются, например, DOTMA, DOPE и DC-Chol). В некоторых ситуациях желательно обеспечить источник нуклеиновой кислоты агентом, который направляет ее в клетки-мишени, таким как антитело, специфическое для мембранного белка клеточной поверхности или клетки-мишени, лиганд для рецептора на клетке-мишени и т.д. В тех случаях, когда используются липосомы, то белки, которые связываются с мембранным белком клеточной поверхности, связанным с эндоцитозом, можно использовать для направленного транспорта и/или облегчения захвата, например капсидные белки или их фрагменты, проявляющие тропность для клеток определенного типа, антитела к белкам, которые подвергаются интернализации в крови, и белки, которые направлены на внутриклеточную локализацию и повышают период полураспада в клетке. Описана методика опосредованного рецептором эндоцитоза, например, Wu et al., J. Biol. Chem. 262: 4429-4432 (1987); Wagner et al., Proc. Natl. Acad. Sci. USA, 87: 3410-3414 (1990). Обзор по известному в настоящее время маркированию генов и протоколам генной терапии, смотри у Anderson et al., Science, 256: 808-813 (1992). Также смотри WO 93/25673 и цитированные там источники.
VI. Депозирование материалов
Следующие гибридомные клеточные линии были депозированы в Американской коллекции типовых культур, 10801 University Boulevard, Manassas, VA 20110-2209, USA (ATCC):

Дополнительные детали изобретения иллюстрируются последующими не ограничивающими примерами. Раскрытие всех цитат в описании специально включено здесь для сведения.
Пример 1
Получение и характеристика моноклонального антитела 2С4
Мышиные моноклональные антитела 2С4, 7F3 и 4D5, которые специфически связываются с внеклеточным доменом HER2, получали, как описано Fendy et al., Cancer Research, 50: 1550-1558 (1998). Кратко, клетки NIH 3T3/HER2-3400 (экспрессирующие примерно 1×105 молекул HER2/клетку), полученные, как описано Hudziak et al., Proc Natl. Acad. Sci., (USA) 84: 7159-7163 (1987), собирали в забуференный фосфатом физиологический раствор (PBS), содержащий 25 мМ ЭДТА и использовали для иммунизации мышей BALB/с. Мышам вводили внутрибрюшинно 107 клеток в 0,5 мл PBS на 0, 2, 5 и 7 недели. Мышам с антисывороткой, которая иммунопреципитировала 32Р-меченый HER2, вводили внутрибрюшинно мембранный экстракт HER2, очищенный на агглютинине из проростков пшеницы/сефарозе (WGA), на 9 и 13 недели. Затем вводили внутривенно 0,1 мл препарата HER2 и спленоциты гибридизовали с клетками мышиной миеломы X63-Ag8.653.
Супернатанты гибридом подвергали скринингу на связывание HER2 с помощью ELISA и радиоиммунопреципитацией.
Эпитопы HER2, связанные с моноклональными антителами 2С4, 7F3 и 4D5, определяли анализом конкурентного связывания (Fendly et al., Cancer Research, 50: 1550-1558 (1990)). Опыты по перекрестному блокированию проводили с антителами прямой флуоресценцией на интактных клетках с использованием устройства PANDEXTM Screen для количественного определения флуоресценции. Каждое моноклональное антитело конъюгировали с флуоресцеином изотиоцианатом (FITC) с использованием принятых методов (Wofsy et al., Selected Methods in Cellular Immunology, p. 287, Mishel and Schiigi (eds.) San Francisco: W. J. Freeman Co. (1980)). Слитые монослои клеток NIH 3T3/HER2-3400 обрабатывали трипсином, промывали один раз и ресуспендировали для получения титра 1,75×106 клеток/мл в холодном PBS, содержащем 0,5% бычьего сывороточного альбумина (BSA) и 0,1% NaN3. Вносили латексные частицы в конечной концентрации 1% (IDC, Portland, OR) для уменьшения забивания мембран планшета PANDEXTM. Клетки в виде суспензии, 20 мкл, и 20 мкл очищенных моноклональных антител (от 100 мкг/мл до 0,1 мкг/мл) вносили в лунки планшета PANDEXTM и инкубировали на льду в течение 30 мин. В каждую лунку вносили заранее определенное разведение меченных FITC моноклональных антител в 20 мкл, инкубировали в течение 30 мин, промывали и количественно определяли флуоресценцию на PANDEXTM. Моноклональные антитела рассматривали как разделяющие эпитоп, если каждое блокировало связывание другого на 50% или выше по сравнению с иррелевантным контрольным моноклональным антителом. В данном опыте моноклональные антитела 4D5, 7F3 и 2С4 представляли соответственно эпитопы I, G/F и F.
Способность моноклональных антител 2С4, 7F3 и 4D5 ингибировать рост оценивали с использованием линии клеток опухоли молочной железы SK-BR-3 (смотри Hudziak et al., Molec. Cell Biol., 9(3): 1165-1172 (1989)). Кратко, клетки SK-BR-3 разделяли при использовании 0,25% раствора (об./об.) трипсина и суспендировали в полной среде при титре 4×105 клеток на мл. Аликвотные порции 100 мкл (4×104 клеток) высевали в 96-луночные титрационные микропланшеты, клеткам давали возможность прикрепиться и затем вносили 100 мкл одной среды и среды, содержащей моноклональное антитело (конечная концентрация 5 мкг/мл). Через 72 ч планшеты дважды промывали PBS (рН 7,5), окрашивали кристаллическим фиолетовым (0,5% раствор в метаноле) и анализировали относительную пролиферацию, как описано Sugarman et al., Science, 230: 943-945 (1985). Моноклональные антитела 2С4 и 7F3 подавляли пролиферацию клеток SK-BR-3 примерно соответственно на 20 и 38% по сравнению с примерным 56% ингибированием, полученным с моноклональным антителом 4D5.
Моноклональные антитела 2С4, 4D5 и 7F3 оценивали на их способность ингибировать стимулированное HRG фосфорилирование тирозина в белках в пределе Mr 180000 из лизатов цельных клеток MCF7 (Lewis еt al., Cancer Research, 56: 1457-1465 (1996)). Сообщалось, что клетки MCF7 экспрессируют все известные HER-рецепторы, но в относительно низких концентрациях. Поскольку HER2, HER3 и HER4 имеют примерно одинаковые размеры молекулы, то невозможно установить, какой белок становится фосфорилированным по тирозину, когда вестерн-блоттингом оцениваются лизаты цельных клеток.
Однако данные клетки являются идеальными для постановки тестов по оценке фосфорилирования тирозина под влиянием HRG, поскольку в используемых условиях анализа, при отсутствии экзогенного добавленного HRG, они показывают низкие или не детектируемые уровни белков с фосфорилированным тирозином в пределе Mr 180000.
Клетки MCF7 высевали в 24-луночные планшеты и моноклональные антитела против HER2 вносили в каждую лунку и инкубировали в течение 30 мин при комнатной температуре; затем в каждую лунку добавляли rHRGβ1177-244 до конечной концентрации 0,2 нМ и инкубирование продолжали в течение 8 мин. Из каждой лунки осторожно аспирировали среду и реакции останавливали добавлением 100 мкл буфера для проб SDS (5% SDS, 25 мМ DTT и 25 мМ Трис-HCl буфера, рН 6,8). Каждую пробу (25 мкл) подвергали электрофорезу в 4-12% градиентном геле (Novex) и затем электрофоретически переносили на мембрану из поливинилидена дифторида. Проводили иммуноблоттинг с антителами против фосфотирозина (4G10 производств UBI с титром 1 мкг/мл) и количественно оценивали интенсивность основной реактивной полосы, соответствующей Mr ≈ 180000, отражательной денситометрией, как описано ранее (Holmes et al., Science, 256: 1205-1210 (1992); Sliwkowski et al., J. Biol. Chem., 269: 14661-14665 (1994)).
Моноклональные антитела 2С4, 7F3 и 4D5 достоверно ингибировали генерацию индуцированного HRG сигнала фосфорилирования тирозина при Mr 180000. При отсутствии HRG ни одно из данных антител не было способно стимулировать фосфорилирование тирозина в белках в пределе Mr 180000. Также данные антитела не реагировали перекрестно с EGRF (Fendly et al., Cancer Research, 50: 1550-1558 (1990)), HER3 или HER4. Антитела 2С4 и 7F3 достоверно ингибировали стимуляцию под действием HRG фосфорилирования тирозина р180 до <25% от контроля. Моноклональное антитело 4D5 было способно блокировать стимуляцию HRG фосфорилирования тирозина примерно на 50%. На фиг.2А представлены кривые зависимости для ингибирования под действием 2С4 и 7F3 стимуляции HRG фосфорилирования тирозина р180 по данным отражательной денситометрии. Обработка данных кривых ингибирования с использованием 4 параметров давала значение IC50, равное 2,8 ± 0,7 нМ и 29,0 ± 4,1 нМ соответственно, для антител 2С4 и 7F3.
Ингибирование связывания HRG с клетками опухоли молочной железы антителами против HER2 проводили с монослойными культурами на льду в формате 24-луночных планшетов (Lewis et al., Cancer Research, 56: 1457-1465 (1996)). Моноклональные антитела против HER2 вносили в каждую лунку и инкубировали в течение 30 мин. Добавляли rHRGβ1177-244, меченный 125I (25 пкм), и инкубацию продолжали в течение 4-16 ч. На фиг.2В представлены кривые зависимости для ингибирования под действием 2С4 и 7F3 связывания HRG с клетками MCF7. Антитела 2С4 и 7F3 в различных концентрациях инкубировали с клетками MCF7 в присутствии rHRGβ1177-244, меченного 125I, и кривые ингибирования представлены на фиг.2В. Обработка этих данных давала значение IC50, равное 2,4 ± 0,3 нМ и 19,0 ± 7,3 нМ соответственно, для антител 2С4 и 7F3. Максимальное ингибирование, равное примерно 74% для антител 2С4 и 7F3, согласовывалось с данными по фосфорилированию тирозина.
Для определения того, насколько действие антител против HER2, наблюдаемое на клетках MCF7, является общим явлением, человеческие опухолевые клетки инкубировали с 2С4 и 7F3 и определяли степень специфического связывания rHRGβ1, меченного 125I (Lewis et al., Cancer Research, 56: 1457-1465 (1996)). Результаты данного опыта представлены на фиг.3. Связывание rHRGβ1, меченного 125I, достоверно подавлялось под действием антител 2С4 и 7F3 во всех клеточных линиях за исключением клеток опухоли молочной железы MDA-MB-468, о которых сообщалось, что они незначительно экспрессируют или совсем не экспрессируют HER2. Сообщалось, что остальные клеточные линии экспрессируют HER2 при широко варьирующемся уровне экспрессии среди данных клеточных линий. Фактически уровень экспрессии HER2 в данных тестированных клеточных линиях варьировался более чем на 2 порядка. Например, клетки BT-20, MCF7 и Caov3 экспрессируют ≈104 рецепторов HER2/клетку, в то время как клетки BT-4 и SK-BR-3 экспрессируют ≈106 рецепторов HER2/клетку. С учетом колебаний экспрессии HER2 в широких пределах в этих клетках и данных, представленных выше, было сделано заключение, что взаимодействие между HER2 и HER3 или HER4 было само по себе взаимодействием с высокой аффинностью, которое имеет место на поверхности плазматической мембраны.
Оценивали ингибирующее рост действие моноклональных антител 2С4 и 4D5 в отношении клеток MDA-MB-175 и SK-BR-3 в присутствии или отсутствии экзогенного rHRGβ1 (Schaefer et al., Oncogene, 15: 1385-1394 (1997)). Уровень HER2 в клетках MDA-MB-175 был в 4-6 раз выше по сравнению с таковым, обнаруженным в эпителиальных клетках молочной железы, и рецептор HER2-HER4 является конститутивно фосфорилированным по тирозину в клетках MDA-MB-175. Клетки MDA-MB-175 обрабатывали моноклональными антителами 2С4 и 4D5 против HER2 (10 мкг/мл) в течение 4 суток. В тесте окрашивания кристаллическим фиолетовым инкубация с 2С4 показывала сильное ингибирующее рост действие на данную клеточную линию (фиг.4А). Экзогенный HRG не "снимал" достоверно данное ингибирование. С другой стороны, антитело 2С4 не оказывало ингибирующего действия на клетки SK-BR-3 со сверхэкспрессией HER2 (фиг.4В). Моноклональное антитело 2С4 было способно ингибировать пролиферацию клеток MDA-MB-175 в большей степени по сравнению с моноклональным антителом 4D5 в присутствии и при отсутствии экзогенного HRG. Подавление пролиферации клеток под действием антитела 4D5 зависело от уровня экспрессии HER2 (Lewis et al., Cancer Immunol. Immunother., 37: 255-263 (1993)). Можно было детектировать максимальное ингибирование, равное 66%, в клетках SK-BR-3 (фиг.4В). Однако данный эффект невозможно было "снять" экзогенным HRG.
Пример 2
HRG-зависимое связывание HER2 с HER3 блокируется моноклональным антителом 2С4
Тестировали способность HER3 соединяться с HER2 в опыте по коиммунопреципитации. 1,0×106 клеток MCF7 или SK-BR-3 высевали в 6-луночные планшеты для культивирования тканей в среде DMEM/Ham's F12 в соотношении 50:50, содержащей 10% фетальной телячьей сыворотки (FBS) и 10 мМ HEРЕS, рН 7,2 (культуральная среда), и давали прикрепиться в течение ночи. Клетки выращивали в течение 2 ч на бессывороточной культуральной среде перед началом опыта.
Клетки быстро промывали забуференным фосфатом физиологическим раствором (PBS) и затем инкубировали со 100 нМ указанного антитела, разведенного в 0,2% бычьем сывороточном альбумине мас./об. (BSA), среде RPMI со 10 мМ HEРЕS, рН 7,2 (буфер для связывания) или с одним буфером для связывания (контроль). Через 1 ч инкубации при комнатной температуре вносили HRG в конечной концентрации 5 нМ в половину лунок (+). Аналогичный объем буфера для связывания добавляли в другие лунки (-). Инкубацию продолжали примерно в течение 10 мин.
Супернатанты удаляли аспирированием и клетки лизировали в среде RPMI, 10 мМ HEРЕS, рН 7,2, 1,0% TRITON X-100TM об./об., 1,0% мас./об. CHAPS (буфер для лизиса), содержащий 0,2 мМ PMSF, 10 мкг/мл лейпептина и 10 ТЕ/мл апротинина. Лизаты очищали от нерастворимого материала центрифугированием.
HER2 иммунопреципитировали с использованием моноклонального антитела, ковалентно конъюгированного с гелем для аффинной хроматографии (Affi-Prep 10, Bio-Rad). Данное антитело (Ab-3, Oncogene Sciences) распознает эпитоп цитоплазматического домена. Иммунопреципитацию проводили при добавлении 10 мкл суспензии геля, содержащего примерно 8,5 мкг иммобилизованного с каждым лизатом антитела, и пробам давали перемешаться при комнатной температуре в течение 2 ч. Затем гели собирали центрифугированием. Гели промывали порциями три раза буфером для лизиса для удаления несвязанного материала. Затем добавляли SDS-буфер для проб и пробы быстро нагревали на кипящей водяной бане.
Затем супернатанты подвергали электрофорезу в 4-12% полиакриламидном геле и электрофоретически переносили на нитроцеллюлозные мембраны. Присутствие HER3 оценивали гибридизацией блотов с поликлональным антителом против эпитопа цитоплазматического домена (с-17, Santa Cruz Biotech). Блоты визуализировали с использованием хемилюминесцентного субстрата (ECL, Amersham).
Как представлено на контрольных дорожках на фиг.5А и 5В соответственно для клеток MCF7 и SK-BR-3, HER3 присутствовал в иммунопреципитате HER2 только, когда клетки стимулировали HRG. Если клетки вначале инкубировали с моноклональным антителом 2С4, то сигнал HER3 исчезал из клеток MCF7 (фиг.5А, дорожка 2С4+) или существенно уменьшался в клетках SK-BR-3 (фиг.5В, дорожка 2С4+). Как представлено на фиг.5А-В, моноклональное антитело 2С4 блокирует зависимое от герегулина связывание HER3 с HER2 в клетках MCF7 и SK-BR-3 значительно эффективнее по сравнению с трастузумабом. Предварительная инкубация с трастузумабом снижала сигнал HER3 в лизатах клеток MCF7, но оказывала незначительное влияние, или оно отсутствовало, на количество копреципитированного HER3 из лизатов клеток SK-BR-3. Предварительная инкубация с антителом против рецептора EGF (Ab-1, Oncogene Sciences) не оказывала влияния на способность HER3 к коиммунопреципитации с HER2 ни в одной клеточной линии.
Пример 3
Гуманизированные антитела 2С4
Вначале вариабельные домены мышиного моноклонального антитела 2С4 клонировали в вектор, позволяющий получить мышиный/человеческий химерный Fab-фрагмент. Общую фракцию РНК выделяли из гибридомных клеток с использованием набора для экстракции РНК Stratagene, следуя протоколам производителя. Вариабельные домены амплифицировали RT-ПЦР, очищали в геле и вставляли в производное на основе плазмиды pUC119, содержащей человеческий константный домен каппа и человеческий домен СН1, как описано ранее (Carter et al., PNAS (USA), 89: 4285 (1992) и в патенте США № 5821337). Полученную плазмиду трансформировали в штамм 16С9 E. coli для экспрессии Fab-фрагмента. Культивирование культуры, индукцию экспрессии белка и очистку Fab-фрагмента проводили, как описано ранее (Werther et al., J. Immunol., 157: 4986-4995 (1996); Presta et al., Cancer Research, 57: 4593-4599 (1997)).
Очищенный химерный Fab-фрагмент 2С4 сравнивали с мышиным исходным антителом 2С4 в отношении его способности ингибировать связывание 125I-HRG с клетками МCF7 и ингибировать активацию под действием rHRG фосфорилирования тирозина р180 в клетках МCF7. Как представлено на фиг.6А, химерный Fab-фрагмент 2С4 является очень эффективным в разрыве образования сайта связывания HER2-HER3 с высокой аффинностью в клетках опухоли молочной железы МCF7. Относительное значение IC50, рассчитанное для интактного мышиного 2С4, равнялось 4,0 ± 0,4 нМ, в то время как значение для Fab-фрагмента составляло 7,7 ± 1,1 нМ. Как показано на фиг.6В, моновалентный химерный Fab-фрагмент 2С4 является очень эффективным в нарушении HRG-зависимой активации HER2-HER3. Значение IC50, рассчитанное для интактного мышиного антитела 2С4, равнялось 6,0 ± 2 нМ, в то время как значение для Fab-фрагмента составляло 15,0 ± 2 нМ.
Секвенирование ДНК химерного клона позволило идентифицировать остатки CDR (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)) (фиг.7А и В). С использованием олигонуклеотидного сайт-направленного мутагенеза все шесть данных областей CDR вводили в полную человеческую каркасную область (подгруппа VL каппа I и подгруппа VH III), входящую в плазмиду VX4, как описано ранее (Presta et al., Cancer Research, 57: 4593-4599 (1997)). Белок из полученного "CDR-обмена" экспрессировали и очищали, как описано выше. Опыты по связыванию проводили для сравнения двух вариантов. Кратко, планшет NUNC MAXISORP™ покрывали 1 мкг на мл внеклеточного домена HER2 (ECD; полученного, как описано в WO 90/14375) в 50 мМ карбонатном буфере, рН 9,6 в течение ночи при 4°С и затем блокировали разбавителем для ELISA (0,5% BSA, 0,05% полисорбат 20, PBS) при комнатной температуре в течение 1 ч. Серийные разведения проб в разбавителе для ELISA инкубировали в планшетах в течение 2 ч. После промывания детектировали Fab-фрагмент, связанный с биотинилированным мышиным анти-человеческим каппа антителом (ICN 634771), затем с пероксидазой из хрена, конъюгированной со стрептавидином (Sigma), с использованием 3,3',5,5'-тетраметилбензидина (Kirkеgaard & Perry Laboratories, Gaithersburg, MD) в качестве субстрата. Поглощение определяли при 450 нМ. Как представлено на фиг.8А, связывание отсутствовало на конструкции человеческого Fab-фрагмента CDR-обмена.
Для восстановления связывания гуманизированного Fab конструировали мутанты с использованием ДНК из CDR-обмена в качестве матрицы. При использовании созданной компьютерной модели (фиг.9) данные мутации проводили для изменения человеческой каркасной области на ее мышиные копии в положениях, которые могут изменить конформации CDR или поверхность раздела антиген-антитело. Мутанты представлены в таблице 2.

Кривые связывания для различных мутантов приведены на фиг.8А-С. Оказалось, что гуманизированный вариант Fab 574, с изменениями ArgH71Val, AspH73Arg и IleH69Leu, обладал связыванием, восстановленным до исходного химерного Fab-фрагмента 2С4. Дополнительные остатки FR и/или CDR, такие как L2, L54, L55, L56, H35 и/или Н48, можно модифицировать (например, заместить следующим образом - IleL2Thr; ArgL54Leu; TyrL55Glu; ThrL56Ser; AspH35Ser и ValH48Ile) для дополнительного улучшения или повышения связывания гуманизированного антитела. Альтернативно или дополнительно гуманизированное антитело может быть со "зрелой аффинностью" (смотри выше) для дополнительного повышения или улучшения его аффинности и/или других биологических активностей.
Гуманизированному варианту 2С4 574 обеспечивали "зрелую аффинность" при использовании метода фагового дисплея. Кратко, гуманизированный Fab 2С4.574 клонировали в вектор фагового дисплея в виде гибридного продукта гена III. Когда фаговые частицы индуцировали инфицированием фагом-помощником М13КО7, то данный гибридный продукт позволял расположить Fab в N-конце белка фагового отростка, ген III (Baca et al., J. Biol. Chem., 272: 10678 (1997)).
Конструировали отдельные библиотеки для каждого из 6 CDR, указанных выше. В данных библиотеках аминокислоты в CDR, которые идентифицировали с использованием созданной компьютерной модели (фиг.9) как имеющие потенциальное значение для связывания HER2, рандомизировали с использованием олигонуклеотидов, содержащих "NNC" в качестве их кодонов. Затем библиотеки разделяли пэннингом против на покрытых HER2 ECD планшетах NUNC MAXISORP™ с использованием 3% сухого молока в PBS с 0,2% твином 20® (MPBST) вместо всех блокирующих растворов. Для отбора фага с аффинностью, превышающей таковую у 2С4.574, в циклах пэннинга 3, 4 и 5, вносили растворимый HER2 ECD или растворимый Fab 2С4.574 на стадиях промывания в качестве конкурента. Время промывания составляло до 1 ч при комнатной температуре.
После 5 циклов пэннинга отдельные клоны вновь анализировали фаг-ELISA. Отдельные клоны культивировали в планшетах для культивирования тканей с U-образным дном Costar и фаг индуцировали добавлением фага-помощника. После культивирования в течение ночи клетки E. coli центрифугировали и супернатанты, содержащие фаг, переносили в 96-луночные планшеты, где фаг блокировали MPBST в течение 1 ч при комнатной температуре. Планшеты NUNC MAXISORP™, покрытые HER2 ECD, также блокировали MPBST, как описано выше. Блокированный фаг инкубировали на планшетах в течение 2 ч. После промывания связанный фаг детектировали с использованием моноклонального анти-М13-антитела, коньюгированного с пероксидазой из хрена (Amersham Pharmacia Biotech., Inc. 27-9421-01) в разведении 1:5000 в MPBST, затем 3,3',5,5'-тетраметилбензидина в качестве субстрата. Поглощение определяли при 450 нм.
ДНК 48 клонов из каждой библиотеки, которые давали наиболее высокие сигналы, секвенировали. Те клоны, чьи последовательности повторялись наиболее часто, субклонировали в вектор, описанный выше, который обеспечивал экспрессию растворимого Fab. Данные Fab индуцировали, белки очищали и очищенные Fab анализировали на связывание с помощью ELISA, как описано выше, и связывание сравнивали с таковым у исходного гуманизированного варианта 2С4.574.
После идентификации интересующих мутаций в отдельных CDR конструировали дополнительные мутанты, которые представляли различные их комбинации, и тестировали, как описано выше. Мутанты, которые давали повышенное связывание по сравнению с 574, представлены в таблице 3.
Обозначение мутантов, полученных "созреванием аффинности" 2С4.574

*Соотношение количества мутанта, необходимое для получения среднего значения OD на стандартной кривой к количеству 574, необходимого для получения среднего значения OD на стандартной кривой в Erb2-ECD ELISA. Количество менее чем 1,0 указывает, что мутант связывается с Erb2 лучше, чем связывается 574.
Также конструировали и оценивали следующие мутанты:

Конструировали следующие мутанты, предложенные на основе данных по сканированию гомологии:

Предпочтительной аминокислотой в Н34 должен быть метионин. Можно провести замену лейцина, если установлено окисление в данном положении.
Было установлено, что аsnH52 и asnH53 являются очень предпочтительными для связывания. Замена данных остатков на аланин и аспарагиновую кислоту значительно снижает связывание. Было получено интактное антитело, содержащее вариабельные домены легкой и тяжелой цепи гуманизированного варианта 574 с константной областью тяжелой цепи человеческого IgG1 (смотри патент США № 5821337). Интактное антитело получено с использованием клеток яичника китайского хомячка (СНО). Данная молекула обозначена здесь пертузумабом.
Пример 4
Моноклональное антитело 2С4 блокирует опосредованную EGF, TФР-α или HRG активацию киназы МАРК и Akt
Многие рецепторы факторов роста являются сигналом для митоген-активированного пути протеинкиназы (МАРК). Данные киназы с двойной специфичностью представляют одну из ключевых конечных точек в путях передачи сигнала, которые в конечном итоге "запускают" деление опухолевых клеток. Оценивали способность моноклонального антитела 2С4 или трастузумаба ингибировать активацию МАРК, опосредуемую EGF, TФР-α или HRG, следующим образом.
Клетки MCF7 (105 клеток/лунку) высевали в содержащей сыворотку среде в 12-луночных планшетах для культивирования клеток. На следующие сутки среду для клеток удаляли и в каждую лунку вносили новую порцию среды, содержащей 0,1% сыворотки. Затем данную процедуру повторяли на следующие сутки и перед постановкой теста среду заменяли на бессывороточный буфер для связывания (Jones et al., J. Biol. Chem., 273: 11667-74 (1998) и Schaefer et al., J. Biol. Chem., 274: 859-66 (1999)). Клетки уравновешивали при комнатной температуре и затем инкубировали в течение 30 мин с 0,5 мл 200 нМ трастузумаба или моноклонального антитела 2С4. Затем клетки обрабатывали 1 нМ EGF, 1 нМ TФР-α или 0,2 нМ HRG в течение 15 мин. Реакцию останавливали аспирацией среды для клеток и затем добавлением 0,2 мл буфера для проб SDS-PAGE, содержащим 1% DTT. Активацию МАРК оценивали вестерн-блоттингом с использованием антитела против активной МАРК (Promega), как описано ранее (Jones et al., J. Bio. Chem., 273: 11667-74 (1998)).
Как представлено на фиг.10, моноклональное антитело 2С4 достоверно блокировало опосредуемую EGF, TФР-α или HRG активацию МАРК в большей степени по сравнению с трастузумабом. На основании этих данных можно предположить, что моноклональное антитело 2С4 связывается с поверхностью HER2, который используется для его ассоциации с EGFR или HER3, и, таким образом, предупреждает образование сигнального рецепторного комплекса.
Также было показано, что моноклональное антитело 2С4 ингибирует герегулин (HRG)-зависимую активацию Akt. Активация пути передачи сигнала с участием киназы PI3 является важной для выживания клеток (Carraway et al., J. Bio. Chem., 270: 7111-6 (1995)). В опухолевых клетках активация киназы PI3 может играть роль в развитии инвазивного фенотипа (Tan et al., Cancer Research, 59: 1620-1625, (1999)). Путь выживания в основном опосредуется серин/треонинкиназой АКТ (Bos et al., Trends Biochem Sci., 20: 441-442 (1995). Комплексы, образованные между HER2 и HER3 или EGFR, могут инициировать данные пути в ответ соответственно на герегулин или EGF (Olayioye et al., Mol. & Cell Biol., 18: 5042-51 (1998); Karunagaran et al., EMBO Journal, 15: 254-264 (1996) и Krymskaya et al., Am. J. Physiol., 276: L246-55 (1999)). Инкубация клеток опухоли молочной железы MCF7 с антителом 2С4 ингибирует опосредуемую герегулином активацию Akt. Agus et al., Cancer Cell, 2: 127-137 (2002). Кроме того, фоновый уровень активации Akt, который имеет место при отсутствии добавления герегулина, дополнительно уменьшается при добавлении 2С4.
Пример 5
Терапия резистентной к препаратам на основе платины злокачественной опухоли
В данном примере демонстрируется безопасность, переносимость и эффективность пертузумаба в комбинации с гемцитабином у пациентов с резистентными к препаратам на основе платины злокачественной опухолью яичника, первичной перитонеальной карциномой или карциномой фаллопиевой трубы. Действие пертузумаба и гемцитабина на выживаемость без прогрессирования заболевания оценивали у всех пациентов и в подгруппе пациентов, опухоли которых содержали маркеры, которые указывали на активацию HER2.
Для данного исследования отбирали пациентов с прогрессированием заболевания во время получения или в течение 6 месяцев после получения химиотерапии препаратами на основе платины. Пациентов произвольно разделяли для получения гемцитабина в комбинации с пертузумабом или гемцитабина в комбинации с растворителем. Проводили лечение пациентов, которые не получали предшествующего спасающего жизнь лечения резистентной к препаратам платины опухоли перед допуском в исследование и тех, которые получали перед этим один курс лечения резистентной к препаратам платины опухоли.
Гемцитабин вводят в дозе 1000 мг/м2 на 1 и 8 сутки 21 суточного курса. Вначале гемцитабин инфузируют в течение 30 мин. Возможно снижение дозы ввиду проявления токсичности. Растворитель или пертузумаб вводят на 1 сутки 21 суточного курса. Пациентам, произвольно отобранным для получения пертузумаба, вводят первоначальную нагрузочную дозу, равную 840 мг (курс 1), с последующим введением 420 мг в курсах 2 и далее. Пациентам, произвольно отобранным для получения растворителя, растворитель вводят в таком же объеме, какой предназначен для введения пертузумаба для курса 1, курсов 2 и далее. Пациенты без прогрессирования заболевания могут получать до 17 курсов или в течение 1 года.
Пациентам снижают обычную дозу гемцитабина и поддерживают эти дозы в результате развития цитопении. Пертузумаб также поддерживают на уровне поддерживаемых доз гемцитабина для 1 суток. Последующие дозы находятся на уровне пониженных доз, и их не повышают. Если необходимо снижение дозы или поддержание дозы в более чем 4 случаях или если дозы поддерживают более чем в течение 3 недель, то введение гемцитабина прекращают, и по усмотрению лечащего врача или медицинского контроля можно продолжать введение "слепого" препарата до прогрессирования заболевания. Если поддерживают дозы гемцитабина на уровне 8 суток, то тогда дозу на 8 сутки пропускают и последующее лечение начинают со следующего курса (22 сутки предыдущего курса).
Введение гемцитабина продолжают, и рекомендуемые пониженные дозы представлены в последующей таблице:
Последующие дозы для любого пациента, для которого требуется снижение дозы, находятся на уровне пониженной дозы. Если дозы поддерживаются в течение более чем 3 недель в результате развития цитопении, то пациентов оценивают как имеющих неприемлемую токсичность и введение гемцитабина прекращают. В случае, если отсутствует другая дополнительная токсичность III или IV степени, то продолжают введение "слепого" препарата по усмотрению лечащего врача или медицинского контроля. Гематологическая токсичность гемцитабина связана со скоростью введения дозы. Гемцитабин вводят в течение 30 мин независимо от общей дозы. Можно использовать колониестимулирующие средства для цитопении 2 степени по NCI-CTC по усмотрению лечащего врача.
Имеется выбор для перехода на одно средство пертузумаб. Нагрузочную дозу, равную 840 мг, вводят в следующем курсе с продолжением введения дозы 420 мг в последующих курсах каждые 21 сутки. Получают залитые в парафин образцы опухоли или неокрашенные парафиновые срезы репрезентативного образца опухоли и оценивают состояние фосфорилирования HER2. Примерно у 20-40% пациентов со злокачественной опухолью яичника можно обнаружить детектируемое фосфорилирование HER2.
Ответную реакцию оценивают в конце курсов 2, 4, 6, 8, 12 и 17. Определяемое заболевание оценивают с использованием Критериев оценки ответной реакции для солидных опухолей (RECIST) при клиническом осмотре и сканировании СТ или при использовании равноценного метода. Ответную реакцию у пациентов с оцениваемым заболеванием определяют по изменениям СА-125 и признакам заболевания, обнаруживаемым при клиническом и радиологическом обследовании. Ответные реакции подтверждают через 4-8 недель после первоначальной регистрации ответной реакции. Оцениваемые пациенты включают пациентов, которые имели, по меньшей мере, одну оценку ответной реакции и определение состояния фосфорилирования HER2 в образцах их опухолей.
Оценивают следующие результаты лечения.
Первичная эффективность на конечной точке
Выживаемость без прогрессирования заболевания, которая определяется исследователем, проводящим оценку, с использованием изменений по RECIST или СА-125 после начала лечения у всех пациентов.
Выживаемость без прогрессирования заболевания, которая определяется исследователем, проводящим оценку, с использованием изменений по RECIST или СА-125 после начала лечения в каждом варианте в следующих подгруппах:
пациенты с детектируемыми маркерами активации HER2,
пациенты без детектируемых маркеров активации HER2.
Вторичная эффективность на конечной точке
Объективная ответная реакция (PR или CR)
Продолжительность ответной реакции
Время выживания
Отсутствие прогрессирования заболевания в течение 4 месяцев
Данные конечные точки оцениваются у всех пациентов и в следующих подгруппах:
пациенты с детектируемыми маркерами активации HER2,
пациенты без детектируемых маркеров активации HER2.
Для профилактики или лечения возможной тошноты и рвоты пациенту можно проводить премедикацию антагонистами серотонина, стероидами и/или бензодиазепинами. Для профилактики или лечения возможных кожных высыпаний, обычной терапии воспалительного поражения сальных желез можно использовать антибиотики для местного применения или приема внутрь. Другими возможными сопутствующими препаратами являются любые назначенные препараты или противоположные препараты, используемые пациентом в период, начиная за 7 суток до 1 суток и продолжая до последних суток конечного периода лечения. Пациентов с повышением температуры в ответ на инфузию до >38°С или проявлением других симптомов на инфузию можно лечить симптоматически ацетаминофеном, дифенгидрамином или меперидином. Можно вводить не экспериментальные гематопоэтические факторы роста при цитопении 2 степени по NCI-CTC.
У пациентов, подвергшихся лечению, описанному выше, наблюдают улучшение в отношении признаков или симптомов резистентных к препаратам на основе платины злокачественной опухоли яичника, первичной перитонеальной карциномы или карциномы фаллопиевой трубы по одной или более конечным точкам оценки первичной или вторичной эффективности. Кроме того, у пациентов с резистентной к препаратам на основе платины опухолью, подвергшихся лечению, описанному выше, комбинацией пертузумаба и гемцитабина, наблюдают более выраженное улучшение в отношении любого из признаков или симптомов злокачественной опухоли по сравнению с пациентами с резистентной к препаратам на основе платины опухолью при лечении только одним гемцитабином.












| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ АНТИТЕЛ К HER2 | 2005 |
|
RU2361880C2 |
| КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО, КОТОРОЕ СВЯЗЫВАЕТСЯ С ДОМЕНОМ II HER2, И ЕГО КИСЛЫЕ ВАРИАНТЫ | 2009 |
|
RU2543664C2 |
| СПОСОБЫ ИДЕНТИФИКАЦИИ ОПУХОЛЕЙ, ВОСПРИИМЧИВЫХ К ЛЕЧЕНИЮ АНТИТЕЛАМИ ПРОТИВ ErbB2 | 2003 |
|
RU2338751C2 |
| ВВЕДЕНИЕ ФИКСИРОВАННЫХ ДОЗ HER-АНТИТЕЛ | 2005 |
|
RU2438705C2 |
| КОМПОЗИЦИИ АНТИТЕЛ | 2005 |
|
RU2426554C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ ПО ОТНОШЕНИЮ К HER3, И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620068C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО, КОТОРОЕ ОБЛАДАЕТ СПОСОБНОСТЬЮ СВЯЗЫВАТЬ ErbB2 И БЛОКИРОВАТЬ АКТИВАЦИЮ ЛИГАНДОМ РЕЦЕПТОРА ErbB (ВАРИАНТЫ) И КОМПОЗИЦИЯ ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ РАКА, СОДЕРЖАЩАЯ ЭТО АНТИТЕЛО | 2000 |
|
RU2270029C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИ-ТФР-БЕТА-АНТИТЕЛА | 2005 |
|
RU2386638C2 |
| НЕКОНКУРЕНТНЫЕ В ОТНОШЕНИИ НЕЙРЕГУЛИНА АЛЛОСТЕРИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО HER3 И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2704228C2 |
| БИСПЕЦИФИЧНЫЕ АНТИ-HER2/АНТИ-HER3 АНТИТЕЛА | 2016 |
|
RU2653443C2 |

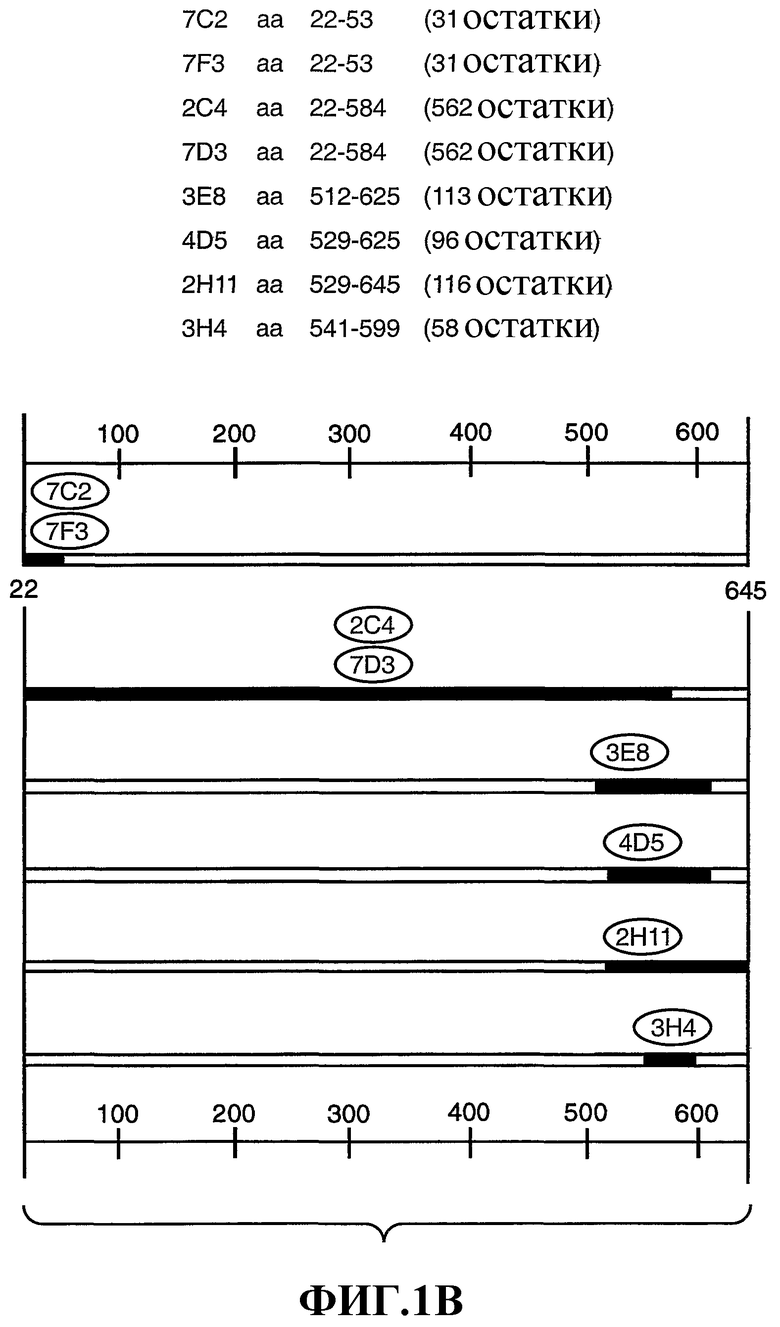

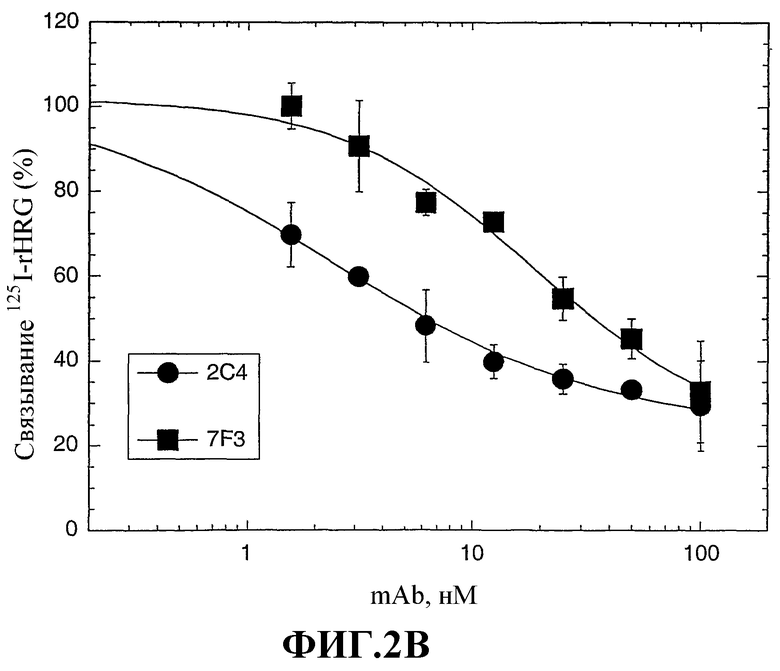

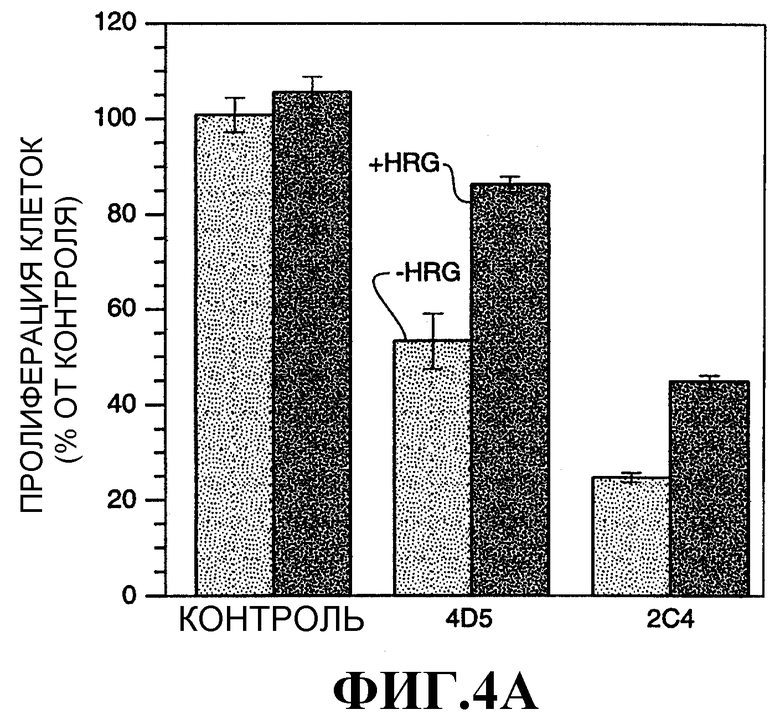

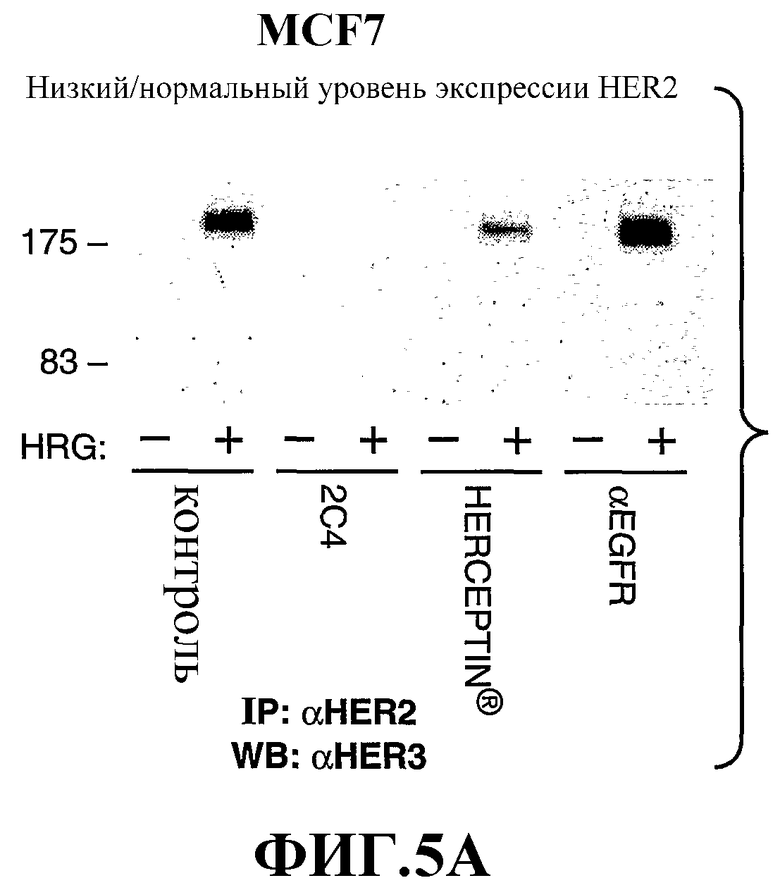
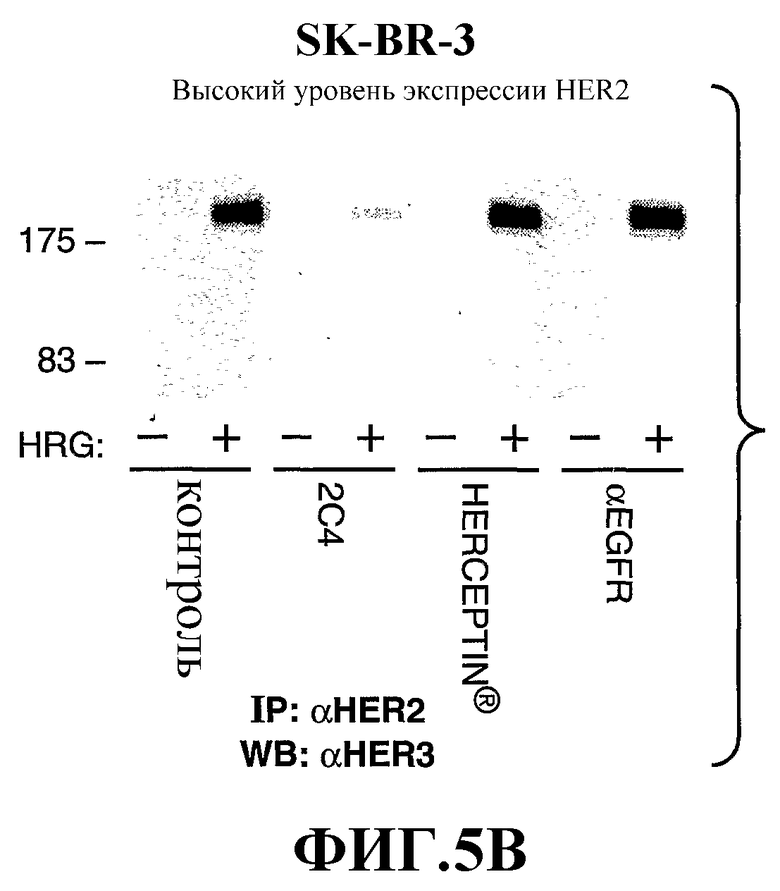
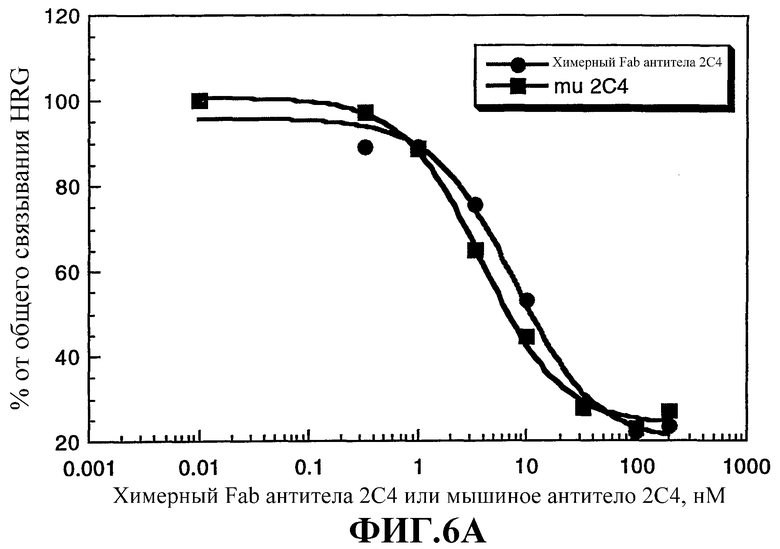
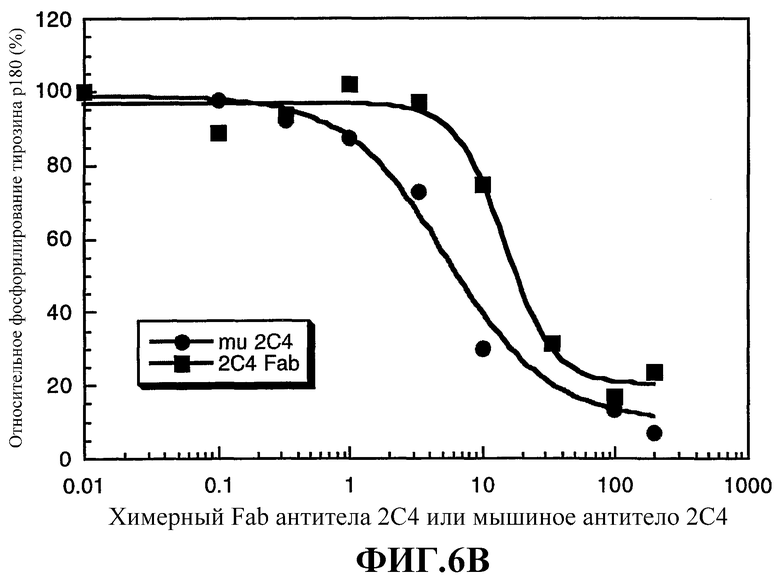



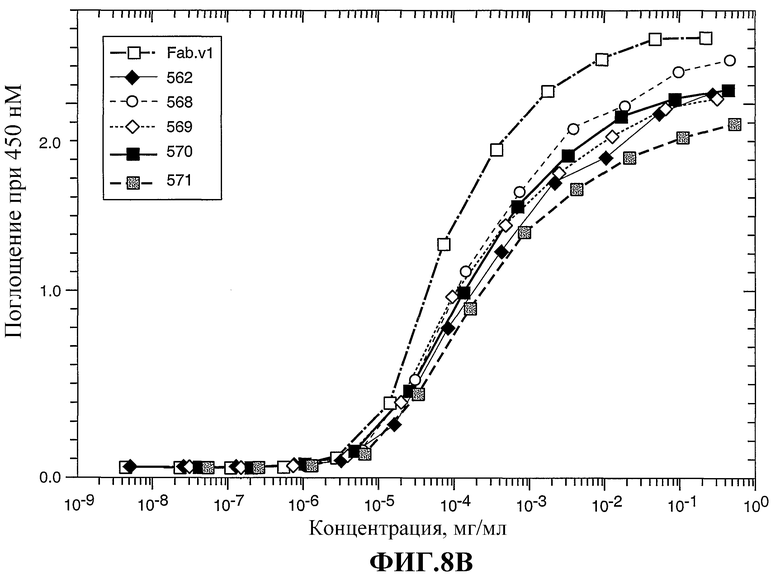
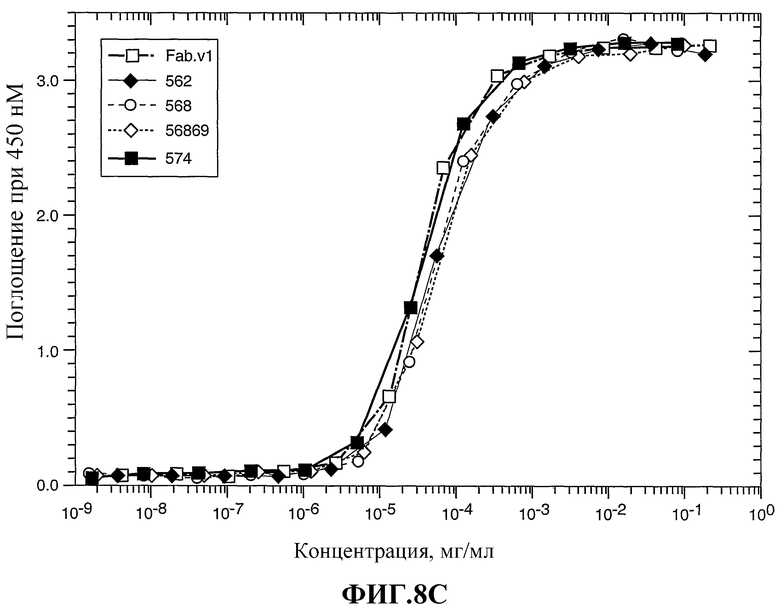
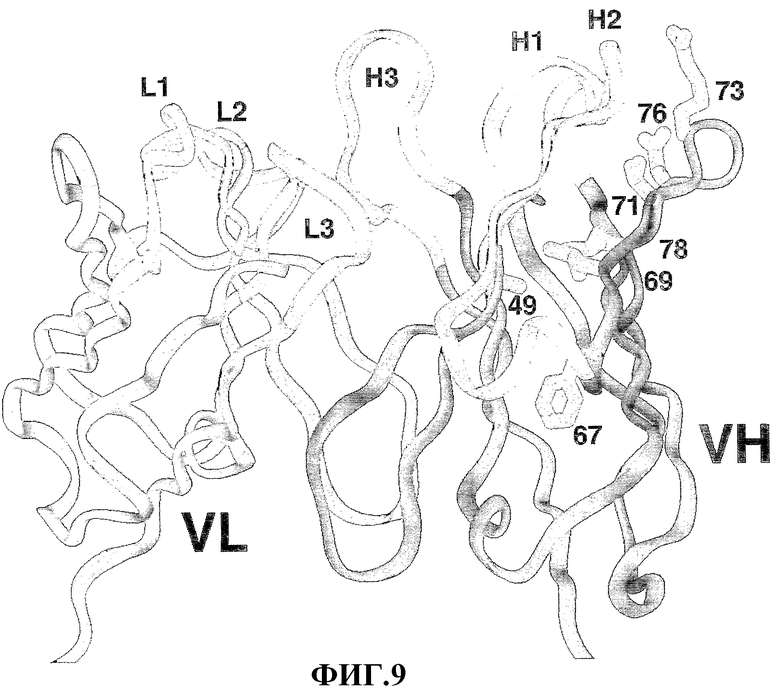
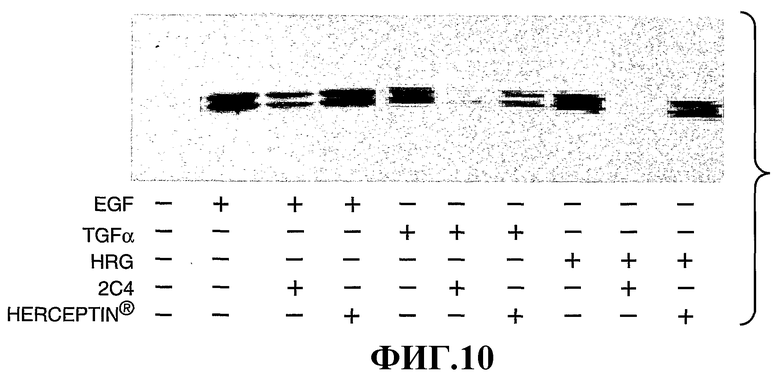





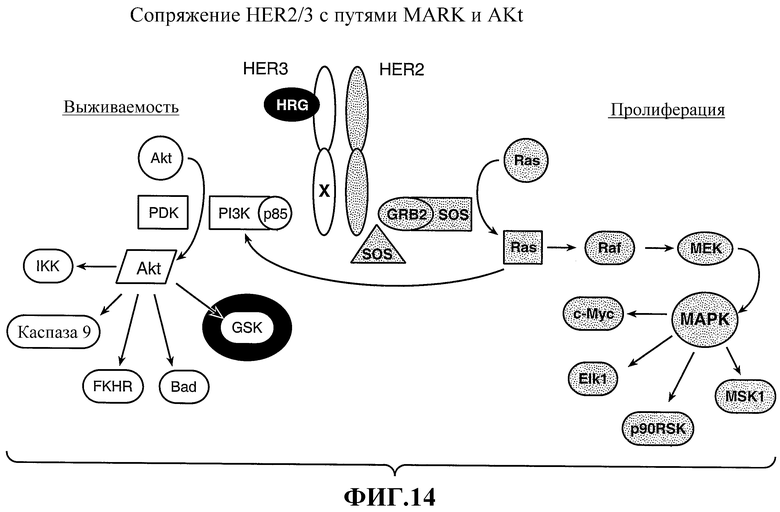

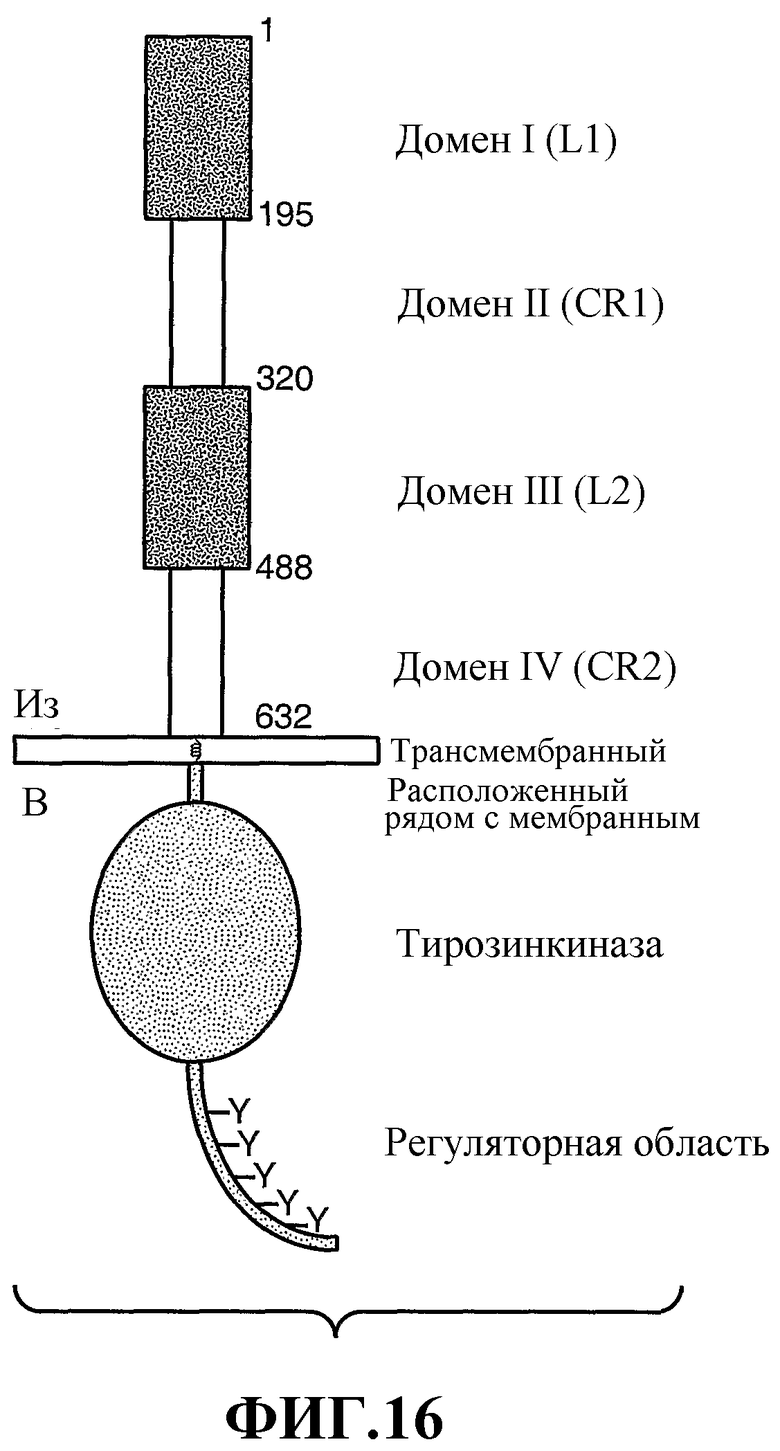
Изобретение относится к медицине, а именно к онкологии, и может быть использовано для лечения злокачественных опухолей, резистентных к препаратам на основе платины. Способ по изобретению включает введение пациенту гуманизированного антитела против 2С4 HER2 или его фрагмента и химиотерапетического средства антиметаболита, каждое в количествах, эффективных для лечения злокачественной опухоли. Использование изобретения позволяет лечить такие опухоли, как злокачественная опухоль яичника, перитонеальная карцинома и карцинома фаллопиевой трубы, резистентные к препаратам на основе платины, за счет более активного подавления антителом 2С4 HER2 димеризации HER. 29 з.п. ф-лы, 4 табл., 16 ил.
1. Способ лечения злокачественной опухоли, выбранной из группы, состоящей из злокачественной опухоли яичника, резистентной к препаратам на основе платины, перитонеальной карциномы, резистентной к препаратам на основе платины, и карциномы фаллопиевых труб, резистентной к препаратам на основе платины, включающий введение пациенту гуманизированного антитела против 2С4 HER2 или его фрагмента, и химиотерапетического средства антиметаболита, каждое в количествах, эффективных для лечения злокачественной опухоли.
2. Способ по п.1, в котором гуманизированное антитело против 2С4 HER2 связывается с доменом II HER2.
3. Способ по п.1, в котором гуманизированное антитело против 2С4 HER2 является пертузумабом.
4. Способ по п.1, в котором химиотерапевтическим средством антиметаболитом является гемцитабин.
5. Способ по п.1, в котором образец опухоли от пациента показывает активацию HER.
6. Способ по п.5, в котором образец опухоли от пациента показывает активацию HER2.
7. Способ по п.1, который приводит к увеличению выживаемости по сравнению с пациентом, подвергшимся лечению только одним химиотерапевтическим средством антиметаболитом.
8. Способ по п.7, который приводит к увеличению выживаемости без прогрессирования заболевания по сравнению с пациентом, подвергшимся лечению только химиотерапевтическим средством антиметаболитом.
9. Способ по п.7, который приводит к увеличению времени выживания в целом по сравнению с пациентом, подвергшимся лечению только одним химиотерапевтическим средством антиметаболитом.
10. Способ по любому из пп.1, 7, 8 или 9, который приводит к объективной ответной реакции.
11. Способ по п.10, который приводит к полной ответной реакции.
12. Способ по п.10, который приводит к частичной ответной реакции.
13. Способ по п.1, в котором гуманизированное антитело против 2С4 HER2 вводят в нагрузочной дозе, равной примерно 840 мг, с последующим введением приблизительно 420 мг примерно каждые 3 недели.
14. Способ по п.1, в котором гуманизированное антитело против 2С4 HER2 вводят в дозе приблизительно 1050 мг примерно каждые 3 недели.
15. Способ по п.4, в котором гемцитабин вводят в дозе в пределах примерно от 600 мг/м2 до 1250 мг/м2 на 1 и 8 сутки 3-недельного цикла.
16. Способ по п.1, в котором химиотерапевтическое средство антиметаболит вводят до или после введения гуманизировнного антитела против 2С4 HER2.
17. Способ по п.16, в котором период времени между, по меньшей мере, одним введением химиотерапевтического средства антиметаболита и, по меньшей мере, одним введением гуманизированного антитела против 2С4 HER2 составляет примерно 1 месяц или менее.
18. Способ по п.17, в котором период времени между, по меньшей мере, одним введением химиотерапевтического средства антиметаболита и, по меньшей мере, одним введением гуманизированного антитела против 2С4 HER2 составляет примерно 2 недели или менее.
19. Способ по п.1, в котором химиотерапевтическое средство антиметаболит и гуманизировнное антитело против 2С4 HER2 вводят пациенту одновременно в одной композиции или разных композициях.
20. Способ по п.1, в котором злокачественная опухоль не сверхэкспрессирует HER2.
21. Способ по п.1, дополнительно включающий введение второго химиотерапевтического средства пациенту.
22. Способ по п.21, в котором второе химиотерапевтическое средство выбрано из группы, состоящей из таксана, капецитабина, химиотерапевтических препаратов на основе платины, антрациклина, липосомального доксорубицина, топотекана, пеметрекседа, алкалоида растения барвинок и TLK 286.
23. Способ по п.1, в котором лечение комбинацией химиотерапевтического средства антиметаболита и гуманизированного антитела против 2С4 HER2 приводит к синергическому или более чем аддитивному, терапевтическому эффекту у пациента.
24. Способ по п.4, в котором лечение комбинацией гемцитабина и гуманизированного антитела против 2С4 HER2 приводит к синергическому или более чем аддитивному эффекту у пациента.
25. Способ по п.1, в котором гуманизированное антитело против 2С4 HER2 представляет "голое" антитело.
26. Способ по п.1, в котором гуманизированное антитело против 2С4 HER2 представляет интактное антитело.
27. Способ по п.1, в котором гуманизированное антитело против 2С4 HER2 представляет фрагмент антитела, содержащий антигенсвязывающую область.
28. Способ по п.1, в котором злокачественная опухоль является злокачественной опухолью яичника, резистентной к препаратам платины.
29. Способ по п.1, в котором злокачественная опухоль является первичной перитонеальной карциномой, резистентной к препаратам платины.
30. Способ по п.1, в котором злокачественная опухоль является карциномой фаллопиевой трубы, резистентной к препаратам платины.
| MARKMAN M | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Gvnecol | |||
| Oncol | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| реферат, он-лайн [Найдено в Интернет на www.pubmed.com 20.02.2009], PMID: 13678730 [PubMed - indexed for MEDLINE] | |||
| ПРОТИВООПУХОЛЕВАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПРОИЗВОДНЫЕ ТАКСАНА | 1993 |
|
RU2131250C1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| CIRISANO | |||

