ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
В данной заявке заявляется приоритет, согласно 35 U.S.С. §119, предварительной заявки США, серийный номер U60/750,085, поданной 13 декабря, 2005 г.; предварительной заявки США, серийный номер 60/750,772, поданной 14 декабря 2005 г.; предварительной заявки США, серийный номер 60/774,747, поданной 17 февраля 2005 г.; и предварительной заявки США, серийный номер 60/808,183, поданной 24 мая 2006 г., каждая из которых полностью включена в данную заявку путем ссылок.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Область изобретения Изобретение относится к связывающим протеинам, которые связывают инсулиноподобный фактор роста - 2 (IGF-II) имеющим перекрестную реактивность к инсулиноподобному фактору роста - 1 (IGF-I) и к использованию таких связывающих протеинов. Более конкретно, изобретение относится к моноклональным антителам, направленным на IGF-II имеющим перекрестную реактивность к IGF-I и к использованию таких антител. Аспекты данного изобретения также относятся к гибридомам или другим линиям, которые экспрессируют такие антитела.
Описание уровня техники
Инсулиноподобные факторы роста IGF-I и IGF-II представляют собой небольшие полипептиды, которые задействованы в регулировании клеточной пролиферации, выживании, дифференциации и трансформации. IGFs различными способами влияют, путем, в основном, взаимодействия со специфическим рецептором клеточной поверхности, рецептором IGF-I (IGF-IR) и активирования различных межклеточных сигнальных каскадов. IGFs циркулирует в сыворотке, в основном связывая IGF-связывающие протеины (IGFBP-1-6). Взаимодействие IGFs с IGF-IR регулируется IGFBPs, a IGFs может связывать только IGF-IR после освобождения из IGFBPs (главным образом, путем протеолиза IGFBPs). IGF-I может также связывать гибридный рецептор, состоящий из IGF-IR и инсулинового рецептора (IR). Было показано, что IGF-II связывает "А" изоформу рецептора инсулина.
Злокачественные преобразования включают в себя дисбаланс различных процессов, таких как рост клеток, дифференциация, апоптоз и преобразование. IGF-I и IGF-II были задействованы в патофизиологии широкого диапазона условий, и считается, что они принимают участие в мутагенезе, что вызвано их митогенными и антиапоптозными свойствами, опосредованными рецептором IGF-IR. LeRoith and Roberts, Cancer Lett. 195: 127-137 (2003).
IGF-I был открыт как фактор роста, который вырабатывается печенью под регулированием гормона роста слизистой, и изначально получил название соматомедин - С.Salmon et al., J. Lab. Clin. Med. 49: 825-826 (1957). Как IGF-I, так и IGF-II экспедируются повсеместно и действуют в качестве эндокринных, паракринных и аутокринных факторов роста, посредством их взаимодействия с IGF-IR, транс-мембранной тирозин-киназой, которая структурно и функционально относится к рецепторам инсулина (IR). IGF-I функционирует, в основном, путем активации IGF-IR, где IGF-II может действовать через IGF-IR или через изоформу IR-A. LeRoith and Roberts, Cancer Lett. 195: 127-137 (2003). Дополнительно, взаимодействие как IGF-I, так и IGF-II с IGF-связывающими протеинами может оказывать влияние на полупериод жизни и бионакопление IGFs, также как и их непосредственное взаимодействие с рецепторами в аналогичных случаях. Rajaram et al., Endocr. Rev. 18: 801-831 (1997).
IGF-I оказывает продолжительное влияние на клеточную пролиферацию, дифференциацию, и апоптоз. Эксперименты с культивированной остеосаркомой и раком молочной железы предположили, что IGF-I является протеиновым митогеном и проявляет свое митогенное действие путем возрастания синтеза DNA и путем стимулирования экспрессии циклинового D1, ускоряющего прогрессию клеточного цикла от G1 до S фазы. Furlanetto et al., Mol. Endocrinol. 8: 510-517 (1994); Dufourny et al., J. Biol. Chem. 272: 311663-31171 (1997). Подавление экспрессии D1 в раковых клетках поджелудочной железы упраздняет митогенное влияние IGF-I. Kornmann et al., J. Clin. Invest. 101: 344-352 (1998). Кроме стимулирования прогрессии клеточного цикла, IGF-I также ингибирует апоптоз. Было показано, что IGF-I стимулирует экспрессию Bсl протеинов и подавляет экспрессию Вах, что приводит к возрастанию относительного количества гетеродимера Bcl/Bax, блокируя, таким образом, путь апопотоза. Minshall et al., J. Immunol. 159: 1225-1232 (1997); Parrizas etal, Endocrinology 138: 1355-1358 (1997); Wang et al., Endocrinology 139: 1354-1360 (1998).
Аналогично IGF-I, IGF-II также имеет мутагенное и антиапоптическое действие и регулирует клеточную пролиферацию и дифференциацию. По сравнению с IGF-I, в сыворотке циркулируют высокие концентрации IGF-II. Высокое содержание IGF-II в сыворотке было найдено у пациентов, имеющих рак кишечника, с тенденцией к увеличению концентрации при развитии болезни. Renehan et al., Br. J. Cancer 83:1344-1350. Кроме того, большинство первичных опухолей и перерожденных клеточных линий избыточно экспрессируют IGF-II и РНК и протеин. Werner and LeRoith Adv. Cancer Res. 68:183-223 (1996). Избыточная экспрессия IGF-II рака толстой кишки связана с агрессивным фенотипом, потеря отпечатков (потерей аллель-специфичной экспрессии) IGF-II гена может быть важна в карциногенезе толстой кишки. Michell et al., Br. J. Cancer 76:60-66 (1997); Takano et al., Oncology 59: 210-216 (2000). Раковые клетки, которые имеют сильную тенденцию к образованию метастаз, имеют уровни экспрессии IGF-, в четыре раза превышающие экспрессию в тех клетках, которые имеют слабую тенденцию к образованию метастаз Guerra et al., Int. J. Cancer 65: 812-820 (1996).
Научные и клинические исследования высветили роль членов семейства IGF в развитии, сохранении и прогрессировании рака. Было показано, что многие раковые клетки имеют избыточную экспрессию h IGF-IR и/или IGF лигандов. Например, IGF-I и IGF-II являются сильными митогенами широкого диапазона раковых клеточных линий, включая саркому, лейкемию, и рак простаты, молочной железы, легких, толстой кишки, желудка, пищевода, печени, поджелудочной железы, почек, щитовидной железы, мозга, яичников, и матки. Macaulay et al., Br. J. Cancer 65: 311-320 (1992); Oku et al., Антиcacancer Res. 11: 1591-1595 (1991); LeRoith et al., Ann. Intern. Med. 122:54-59 (1995); Yaginuma et al., Oncology 54: 502-507 (1997); Singh et al., Endocrinology 137: 1764-1774 (1996); Frostad et al., Eur. J. Haematol 62: 191-198 (1999). Когда IGF-I вводили в злокачественные раковые клетки пищевода, они становились резистентными к апоптозу, вызываемому цитокинами. Remacle-Bonnet et al., Cancer Res. 60: 2007-2017 (2000).
Роль IGFs в развитии рака также подтверждена эпидемиологическими исследованиями, в которых было показано, что высокие уровни циркуляции IGF-I и низкие уровни IGFBP-3 связаны с повышенным риском развития нескольких видов обычного рака (простаты, молочной железы, толстой кишки и легких). Mantzoros et al., Br. J. Cancer 76: 1115-1118 (1997); Hankinson et al., Lancet 351: 1393-1396 (1998); Ma et al., J. Natl. Cancer Inst. 91: 620-625 (1999); Karasik et al., J. Clin. Endocrinol Metab. 78: 271-276 (1994). Эти результаты предполагают, что IGF-I и IGF-II действуют как мощные митогенные и анти-апоптозные сигналы, и их избыточная экспрессия коррелирует с неблагоприятными прогнозами для пациентов с несколькими видами рака.
Используя модели нокаутированных мышей, в нескольких исследованиях была дополнительно установлена роль IGFs при росте опухолей. С развитием технологии тканеспецифической, условной генной делеции, была разработана мышиная модель печеночной IGF-I недостаточности (LID). Печеночно-специфическая делеция гена igfl упраздняла экспрессию IGF-I иРНК и вызывала сильное уменьшение уровней циркуляции IGF-I. Yakar et al., Proc. Natl. Acad. Sci. USA 96: 7324-7329 (1999). Когда опухоли млекопитающих индуцировали у мышей LID, уменьшение уровней циркуляции IGF-1 приводило к значительному уменьшению развития рака, роста и образования метастаз, в то время как увеличение уровней циркуляции IGF-1 было связано с усилением роста опухолей. Wu et al., Cancer Res. 63: 4384-4388 (2003).
В нескольких статьях сообщалось о том, что ингибирование IGF-IR экспрессии и/или сигналов приводит к ингибированию роста опухолей, как in vitro, так и in vivo. Было также показано, что ингибирование передачи сигналов IGF увеличивает чувствительность опухолевых клеток к химиотерапевтическим агентам. Было разработано множество стратегий (антисмысловые олигонуклеотиды, растворимый рецептор, пептидные ингибиторы, доминантные негативные рецепторы-мутанты, малые молекулы - ингибиторы киназной активности и анти-hIGF-IR антитела) для ингибирования IGF-IR путей передачи сигнала в опухолевых клетках. Один из подходов состоял в поражении киназной активности IGF-IR небольшими молекулами-ингибиторами. Недавно были идентифицированы два соединения в качестве небольших молекул - ингибиторов киназы, способных селективно ингибировать IGF-IR. Garcia-Echeverria et al., Cancer Cell 5: 231-239 (2004); Mitsiades et al., Cancer Cell 5: 221-230 (2004). Ингибирование киназной активности IGF-IR не разрешало IGF-I-опосредованное выживание и образование колоний в мягком агаре раковых клеток MCF-7 человеческой молочной железы. Garcia-Echeverria et al., Cancer Cell 5: 231-239 (2004). При введении киназного ингибитора IGF-IR мышам с опухолевыми ксенотрансплантатами, передача сигналов IGF-IR в опухолевых ксенотрансплантантах ингибировалось, а рост вызываемых IGF-IR фибросарком значительно уменьшился. Garcia-Echeverria et al., Cancer Cell 5: 231-239 (2004). Аналогичный эффект наблюдался при гематологических злокачественных опухолях, особенно множественной миеломы. В клетках множественной миеломы (болезнь Рустицкого-Калера), небольшая молекула IGF-IR, ингибитор киназы, проявляла в >16-большую мощность по отношению к IGF-IR, по сравнению с рецептором инсулина, и проявляла аналогичную эффективность при ингибировании роста и выживания клеток. Mitsiades et al., Cancer Cell 5: 221-230 (2004). Аналогичные соединения вводили мышам интраперитонеально и ингибировали рост клеток множественной миеломы и усиливали выживаемость мышей Mitsiades et al., Cancer Cell 5: 221-230 (2004). В сочетании с другими химиотерапевтическими агентами в субтерапевтических дозах, ингибирование киназной активности IGF-IR синергетически уменьшало массу опухолей Mitsiades et al., Cancer Cell 5: 221-230 (2004).
Другой подход в ингибировании передачи сигналов IGF состоял в разработке эффективных нейтрализующих антител, направленных против рецептора IGF-IR. Различными группами были разработаны антитела к IGF-IR для ингибирования стимулированного рецепторами IGF-I аутофосфорилирования, индуцирования рецепторной интернализации и деградации и уменьшения пролиферации и выживаемости различных человеческих раковых клеточных линий. Hailey et al., Mol Cancer Ther. 1: 1349-1353 (2002); Maloney et al., Cancer Res. 63: 5073-5083 (2003); Benini et al., Clin. Cancer Res. 7: 1790-1797 (2001); Burtrum et al., Cancer Res. 63: 8912-8921 (2003). Кроме того, в ксенотрансплантатных опухолевых моделях, блокада IGF-IR приводила к значительному ингибированию роста опухолей молочной железы, почек и поджелудочной железы in vivo. Burtrum et al., Cancer Res. 63: 8912-8921 (2003); Maloney et al., Cancer Res. 63: 5073-5083 (2003). Эксперименты, в которых использовали химерные гуманизированные антитела IGF-IR привели к аналогичным результатам, ингибируя рост раковых клеток молочной железы in vitro и в опухолевых ксенотрансплантатах. Sachdev et al., Cancer Res. 63: 627-635 (2003). Другие гуманизированные IGF-IR антитела блокировали вызванное IGF-I фосфорилирование щитовидной железы и ингибирование роста опухолей в молочной железе и не мелких клетках легких, так же как и in vivo. Cohen et al., Clin. Cancer Res. 11: 2063-2073 (2005); Goetsch et al., Int. J. Cancer 113: 316-328 (2005).
Увеличенные уровни IGF-I также были связаны с несколькими не-раковыми патологическими состояниями, включая акромегалию и гигантизм (Barkan, Cleveland Clin. J. Med. 65: 343, 347-349, 1998), в то время как ненормальное функционирование рецептора IGF-I/IGF-II было задействовано в псориазе (Wraight et al., Nat. Biotech. 18: 521-526, 2000), атеросклерозе и рестенозе гладкой мускулатуры кровяных сосудов после ангиопластики (Bayes-Genis et al., Circ. Res. 86: 125-130, 2000). Увеличенные уровни IGF-I были задействованы в диабете или осложнениях, связанных с диабетом, таких как микрососудистая пролиферация (Smith et al., Nat. Med. 5: 1390-1395, 1999).
Антитела к IGF-I и IGF-II описаны в уровне техники. См., например, Goya et al., Cancer Res. 64: 6252-6258 (2004); Miyamoto et al., Clin. Cancer Res. 11: 3494-3502 (2005). Дополнительно, см. WO 05/18671, WO 05/28515 и WO 03/93317.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Воплощения данного изобретения относятся к связывающим протеинам, которые специфично связывают инсулиноподобные факторы роста и уменьшают рост опухолей. В одном воплощении, связывающие протеины являются полностью человеческими моноклональными антителами, или их связывающими фрагментами, которые специфично связывают инсулиноподобные факторы роста и уменьшают рост опухолей. Механизмы достижения этого могут включать и не ограничиваются либо ингибированием связывания IGF-I/II и его рецептора IGF-IR, ингибированием вызванной IGF-I/II передачи сигналов IGF-IR, либо увеличением клиренса IGF-I/II, таким образом, уменьшая эффективную концентрацию IGF-I/II.
Таким образом, некоторые воплощения обеспечивают полностью человеческий выделенный специфичный связывающие протеин, который, предпочтительно связывает инсулиноподобный фактор роста-II (IGF-II) и имеет перекрестную реактивность к инсулиноподобному фактору роста I (IGF-I), нейтрализует активность IGF-I и IGF-II. В некоторых аспектах, связывающий протеин связывает IGF-II с афинностью, которая, как минимум, в 2,5 раза превышает его афинность к IGF-I. В других аспектах, связывающий протеин, связывающий IGF-II с афинностью, которая, как минимум, в 3, как минимум, в 4, как минимум, в 5, как минимум, в 7, как минимум, в 10, как минимум, в 50, как минимум, в 60, как минимум, в 100 или, как минимум, в 150 раз превышает его афинность к IGF-I.
В некоторых воплощениях, специфический связывающий протеин имеет ЕС50 не более 15 нМ для ингибирования IGF-I-зависимого фосфорилирования IGF-I рецептора в NIH3T3 клетках, которые эктопически экспрессируют IGF-1R. В некоторых аспектах, специфический связывающий протеин имеет ЕС50 не более 15 нМ, не более 10 нМ, или не более 8 нМ для ингибирования IGF-I-зависимого фосфорилирования IGF-I рецептора в NIH3T3 клетках, которые эктопически, экспрессируют IGF-1R.
В некоторых воплощениях, специфический связывающий протеин имеет ЕС50 не более 5 нМ, не более 4 нМ, или не более 3 нМ для ингибирования IGF-I-зависимого фосфорилирования IGF-I рецептора в NIH3T3 клетках, которые эктопически экспрессируют IGF-1R.
В других воплощениях, специфический связывающий протеин ингибирует более чем 70% IGF-II зависимой пролиферации клеток NIH3T3, которые экспрессируют рекомбинантные hIGF-IR при значениях ЕС50, не превышающих 25 нМ, не превышающих 20 нМ, не превышающих 15 нМ, или не превышающих 10 нМ.
В других воплощениях, специфический связывающий протеин ингибирует более 70% IGF-I зависимой пролиферации клеток NIH3T3, которые экспрессируют рекомбинантные hIGF-IR при значениях ЕС50, не превышающих 40 нМ, не превышающих 30 нМ, не превышающих 25 нМ.
В некоторых воплощениях, специфический связывающий протеин конкурирует за связывание с моноклональным антителом, которое содержит последовательность вариабельной тяжелой цепи, которую выбирают из группы, состоящей из SEQ ID №: 2, SEQ ID №: 6, SEQ ID №: 10, SEQ ID №: 14 и SEQ ID №: 18, последовательность вариабельной легкой цепи, которую выбирают из группы, состоящей из SEQ ID №: 4, SEQ ID №; 8, SEQ ID №: 12 и SEQ ID №: 16.
Одним из воплощений данного изобретения является полностью человеческое антитело, которое связывает IGF-I при Kd, которое имеет значение менее чем 500 пикомолей (пМ). Более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 450 пикомолей (пМ). Более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 410 пикомолей (пМ). Более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 350 пМ. Даже более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 300 пМ. Измерения афинности и/или авидности могут быть проведены при помощи BIACORE®, как описано в данной заявке.
Еще одно воплощение данного изобретения представляет собой полностью человеческое антитело, которое связывает IGF-II при Kd, которое имеет значение менее чем 175 пикомолей (пМ). Более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 100 пикомолей (пМ). Более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 50 пикомолей (пМ). Более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 5 пикомолей (пМ). Даже более предпочтительно, антитело связывается при Kd, которое имеет значение менее чем 2 пМ.
В некоторых воплощениях, специфический связывающий протеин представляет собой полностью человеческое моноклональное антитело или связывающий фрагмент представляет собой полностью человеческого моноклонального антитела. Связывающие фрагменты могут включать Fab, Fab(или F(ab′)2 и Fv.
Одно из воплощений данного изобретения включает полностью человеческие моноклональные антитела 7.251.3 (номер доступа АТСС РТА-7422), 7.34.1 (номер доступа АТСС РТА-7423) и 7.159.2 (номер доступа АТСС РТА-7424) которые специфически связывают IGF-I/II, как более подробно обсуждается ниже.
В некоторых воплощениях специфический связывающий протеин, который связывает инсулиноподобный фактор роста-II (IGF-II) и имеет перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), или его связывающий фрагмент может включать полипептид тяжелой цепи, имеющий последовательность SEQ ID №: 6, и полипептид легкой цепи, имеющий последовательность SEQ ID №: 8.
Специфический связывающий протеин может включать полипептид тяжелой цепи, имеющий последовательность SEQ ID №: 10, и полипептид легкой цепи, имеющий последовательность SEQ ID №: 12.
Специфический связывающий протеин в соответствии с данным изобретением может включать полипептид тяжелой цепи, имеющий последовательность SEQ ID №: 14 и полипептид легкой цепи, имеющий последовательность SEQ ID №: 16.
В некоторых воплощениях, специфический связывающий протеин может представлять собой смесь с фармацевтически приемлемым носителем.
Другое воплощение включает выделенные молекулы нуклеиновой кислоты, кодирующие любые специфические связывающие протеины, описанные в данной заявке, вектора, имеющие выделенные молекулы нуклеиновой кислоты, кодирующие специфические связывающие протеины, или клетки-хозяева, трансформированные с любыми такими молекулами нуклеиновой кислоты и векторами.
В некоторых воплощениях специфический связывающий протеин, который связывает инсулиноподобный фактор роста-II (IGF-II) и имеющий перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), или его связывающий фрагмент не связывает специфически IGF-II или IGF-I протеины, если указанные протеины связаны со связывающими протеинами инсулинового фактора роста.
Дополнительные воплощения включают способы определения уровней инсулиноподобного фактора роста-II (IGF-II) и инсулиноподобного фактора роста I (IGF-I) в пробе пациента. Эти способы могут включать обеспечение пробы пациента; контактирование пробы со специфическим связывающим протеином, который связывает инсулиноподобный фактор роста-II (IGF-II) и имеющий перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), или его связывающий фрагмент; и определение уровня IGF-I и IGF-II в указанной пробе. В некоторых аспектах, пробой пациента была кровь.
Дополнительные воплощения включают способы лечения злокачественных опухолей у млекопитающих. Эти способы могут включать отбор млекопитающего, который нуждается в лечении злокачественной опухоли; и введение млекопитающему терапевтически эффективной дозы специфически связывающего протеина, который связывает инсулиноподобный фактор роста-II (IGF-II) и имеет перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), или их связывающего фрагмента. В некоторых аспектах животным является человек. В некоторых аспектах, связывающий протеин представляет собой полностью человеческое моноклональное антитело, и его выбирают из группы, состоящей из mAb 7.251.3 (номер доступа АТСС РТА-7422), mAb 7.34.1 (номер доступа АТСС РТА-7423) и mAb 7.159.2 (номер доступа АТСС РТА-7424).
Заболевания, поддающиеся лечению, могут включать меланому, немелкоклеточный рак легких, глиому, печеночно-клеточный рак, опухоль щитовидной железы, рак желудка, рак простаты, рак молочной железы, рак яичников, рак мочевого пузыря, рак легких, глиобластому, рак эндометрия, рак почек, рак толстой кишки, рак поджелудочной железы и плоскоклеточный рак.
Дополнительные воплощения включают способы лечения заболевания, зависящие от фактора роста у млекопитающих. Эти способы включают отбор млекопитающего, который нуждается в лечении заболевания; и введение указанному млекопитающему терапевтически эффективной дозы специфически связывающего протеина, который связывает инсулиноподобный фактор роста-II (IGF-II) и имеет перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), или их связывающего фрагмента. В некоторых аспектах, млекопитающим может быть человек. В некоторых аспектах, связывающий протеин представляет собой полностью человеческое моноклональное антитело, и его выбирают из группы, состоящей из mAb 7.251.3 (номер доступа АТСС РТА-7422), mAb 7.34.1 (номер доступа АТСС РТА-7423) и mAb 7.159.2 (номер доступа АТСС РТА-7424).
Поддающиеся лечению заболевания, зависимые от фактора роста, могут включать остеопороз, диабет и сердечно-сосудистые заболевания. Другие поддающиеся лечению болезненные состояния включают акромегалию и гигантизм, псориаз, атеросклероз и рестеноз гладкой мускулатуры кровяных сосудов, а также диабет.
Дополнительные воплощения включают конъюгат, состоящий из полностью человеческого моноклонального антитела, которое связывает инсулиноподобный фактор роста-II (IGF-II) и имеет перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), или его связывающего фрагмента, и терапевтическое средство. В некоторых аспектах, терапевтическим средством может быт токсин, радиоактивный изотоп, или фармацевтическая композиция.
В других воплощениях, изобретение обеспечивает полностью человеческие моноклональные антитела, или их связывающий фрагмент, которые связывают инсулиноподобный фактор роста-II (IGF-II) и имеют перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), и содержат гипервариабельный участок 1 (CDR1) тяжелой цепи, имеющий аминокислотную последовательность "Ser Tyr Tyr Trp Ser" (SEQ ID №: 21); гипервариабельный участок 2 (CDR2) тяжелой цепи, имеющий аминокислотную последовательность "Туr Phe Phe Tyr Ser Gly Tyr Thr Asn Tyr Asn Pro Ser Leu Lys Ser" (SEQ ID №: 22); и гипервариабельный участок 3 (CDR3) тяжелой цепи, имеющий аминокислотную последовательность "Ile Thr Gly Thr Thr Lys Gly Gly Met Asp Val" (SEQ ID №: 23).
Дополнительные воплощения включают полностью человеческие моноклональные антитела, или их связывающий фрагмент, содержащие гипервариабельный участок 1 (CDR1) легкой цепи, имеющий аминокислотную последовательность "Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His" (SEQ ID №: 24). Антитела в данной заявке могут также включать гипервариабельный участок 2 (CDR2) легкой цепи, имеющий аминокислотную последовательность "Gly Asn Asn Asn Arg Pro Ser" (SEQ ID №: 25); и гипервариабельный участок 3 (CDR3) легкой цепи, имеющий аминокислотную последовательность "Gin Ser Phe Asp Ser Ser Leu Ser Gly Ser Val" (SEQ ID№: 26).
В других воплощениях, изобретение обеспечивает полностью человеческие моноклональные антитела, или их связывающий фрагмент, которые связывают инсулиноподобный фактор роста-II (IGF-II) и имеют перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), и содержат гипервариабельный участок 1 (CDR1) тяжелой цепи, имеющий аминокислотную последовательность "Ser Tyr Tyr Trp Ser" (SEQ ID №: 27); гипервариабельный участок 2 (CDR2) тяжелой цепи, имеющий аминокислотную последовательность "Tyr Phe Phe Tyr Ser Gly Tyr Thr Asn Tyr Asn Pro Ser Leu Lys Ser" (SEQ ID №: 28); и гипервариабельный участок 3 (CDR3) тяжелой цепи, имеющий аминокислотную последовательность "Ile Thr Gly Thr Thr Lys Gly Gly Met Asp Val" (SEQ ID №: 29).
Дополнительные воплощения включают полностью человеческие моноклональные антитела, или их связывающий фрагмент, содержащий гипервариабельный участок 1 (CDR1) легкой цепи, имеющий аминокислотную последовательность "Thr Gly Arg Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His" (SEQ ID №: 30); гипервариабельный участок 2 (CDR2) легкой цепи, имеющий аминокислотную последовательность "Gly Asn Ser Asn Arg Pro Ser" (SEQ ID №: 31); и гипервариабельный участок 3 (CDR3) легкой цепи, имеющий аминокислотную последовательность "Gln Ser Tyr Asp Ser Ser Leu Ser Gly Ser Val" (SEQ ID №: 32).
В других воплощениях, изобретение обеспечивает полностью человеческие моноклональные антитела, или их связывающий фрагмент, которые связывают инсулиноподобный фактор роста-II (IGF-II) и имеют перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), и содержат гипервариабельный участок 1 (CDR1) тяжелой цепи, имеющий аминокислотную последовательность "Ser Tyr Asp Ile Asn" (SEQ ID №: 33); гипервариабельный участок 2 (CDR2) тяжелой цепи, имеющий аминокислотную последовательность "Trp Met Asn Pro Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe Gln Gly" (SEQ ID №: 34); и гипервариабельный участок 3 (CDR3) тяжелой цепи, имеющий аминокислотную последовательность "Asp Pro Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val" (SEQ ID №: 35).
Дополнительные воплощения включают полностью человеческие моноклональные антитела, или их связывающий фрагмент, содержащий гипервариабельный участок 1 (CDR1) легкой цепи, имеющий аминокислотную последовательность "Ser Gly Ser Ser Ser Asn Ile Glu Asn Asn His Val Ser" (SEQ ID №: 36); гипервариабельный участок 2 (CDR2) легкой цепи, имеющий аминокислотную последовательность "Asp Asn Asn Lys Arg Pro Ser" (SEQ ID №: 37); и гипервариабельный участок 3 (CDR3) легкой цепи, имеющий аминокислотную последовательность "Glu Thr Trp Asp Thr Ser Leu Ser Ala Gly Arg Val" (SEQ ID №: 38).
В других воплощениях, изобретение обеспечивает полностью человеческие моноклональные антитела, bkb их связывающий фрагмент, которые связывают инсулиноподобный фактор роста-II (IGF-II) и имеют перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), и содержат гипервариабельный участок 1 (CDR1) тяжелой цепи, имеющий аминокислотную последовательность "Ser Ser Ser Tyr Tyr Trp Gly" (SEQ ID №: 81); гипервариабельный участок 2 (CDR2) тяжелой цепи, имеющий аминокислотную последовательность "Gly Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu Lys Ser" (SEQ ID №: 82); и гипервариабельный участок 3 (CDR3) тяжелой цепи, имеющий аминокислотную последовательность "Gln Arg Gly His Ser Ser Gly Trp Trp Tyr Phe Asp Leu" (SEQ ID №: 83).
Дополнительные воплощения включают полностью человеческие моноклональные антитела, или их связывающий фрагмент, содержащий гипервариабельный участок 1 (CDR1) легкой цепи, имеющий аминокислотную последовательность "Arg Ala Ser Gln Gly Ile Ser Ser Tyr Leu Ala" (SEQ ID №: 84); гипервариабельный участок 2 (CDR2) легкой цепи, имеющий аминокислотную последовательность "Ala Ala Ser Ser Leu Gln Ser" (SEQ ID №: 85); и гипервариабельный участок 3 (CDR3) легкой цепи, имеющий аминокислотную последовательность "Gln Gln Ala Asn Asn Phe Pro Phe Thr" (SEQ ID №: 86).
В других воплощениях, изобретение обеспечивает полностью человеческие моноклональные антитела, или их связывающий фрагмент, которые связывают инсулиноподобный фактор роста-II (IGF-II) и имеют перекрестную реактивность к инсулиноподобному фактору роста-I (IGF-I), и содержат гипервариабельный участок 1 (CDR1) тяжелой цепи, имеющий аминокислотную последовательность "Ser Ser Ser Asn Tyr Trp Gly" (SEQ ID №: 87); гипервариабельный участок 2 (CDR2) тяжелой цепи, имеющий аминокислотную последовательность "Gly Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu Arg Ser" (SEQ ID №: 88); и гипервариабельный участок 3 (CDR3) тяжелой цепи, имеющий аминокислотную последовательность "Gln Arg Gly His Ser Ser Gly Trp Trp Tyr Phe Asp Leu" (SEQ ID №: 89).
Дополнительные воплощения включают полностью человеческие моноклональные антитела, или их связывающий фрагмент, содержащие гипервариабельный участок 1 (CDR1) легкой цепи, имеющий аминокислотную последовательность "Arg Ala Ser Arg Gly Ile Ser Ser Trp Leu Ala" (SEQ ID №: 90); гипервариабельный участок 2 (CDR2) легкой цепи, имеющий аминокислотную последовательность "Thr Ala Ser Ser Leu Gln Ser" (SEQ ID №: 91); и гипервариабельный участок 3 (CDR3) легкой цепи, имеющий аминокислотную последовательность "Gln Gln Ala Asn Ser Phe Pro Phe Thr" (SEQ ID №: 92).
Некоторые воплощения обеспечивают использование специфических связывающих протеинов, описанных в этой заявке, для получения лекарственных средств для лечения злокачественных опухолей. В некоторых аспектах, специфический связывающий протеин может представлять собой полностью человеческое моноклональное антитело. В определенных аспектах, связывающий протеин представляет собой mAb 7.251.3 (номер доступа АТСС РТА-7422) или mAb 7.34.1 (номер доступа АТСС РТА-7423) или mAb 7.159.2 (номер доступа АТСС РТА-7424). В некоторых аспектах, лекарственное средство предназначено для использования в комбинации со вторым антинеопластическим агентом, который выбирают из группы, состоящей из антитела, химиотерапевтического агента, или радиоактивного средства. В некоторых аспектах, лекарственное средство предназначено для использования в сочетании или после проведения традиционной хирургии, трансплантации стволовых клеток костного мозга или трансплантации периферических стволовых клеток.
Злокачественной опухолью может быть, например, меланома, немелкоклеточный рак легких, глиома, печеночно-клеточный рак, опухоль щитовидной железы, рак желудка, рак простаты, рак молочной железы, рак яичников, рак мочевого пузыря, рак легких, глиобластома, рак эндометрия, рак почек, рак толстой кишки, рак поджелудочной железы и плоскоклеточный рак.
Другие воплощения обеспечивают использование специфических связывающих протеинов, описанных в данной заявке, для получения лекарственного средства для лечения заболеваний, зависящих от фактора роста. В некоторых аспектах, специфический связывающий протеин представляет собой полностью человеческое моноклональное антитело и может быть выбран из группы, состоящей из mAb 7.251.3 (номер доступа АТСС РТА-7422), mAb 7.34.1 (номер доступа АТСС РТА-7423) и mAb 7.159.2 (номер доступа АТСС РТА-7424).
Заболеваниями, зависящими от фактора роста, могут быть, например, остеопороз, диабет и сердечно-сосудистые заболевания.
Преимущественно, антитело содержит аминокислотную последовательность тяжелой цепи, имеющую гипервариабельный участок (CDR) с одной или более последовательностями, приведенными в Таблице 11. Например, антитело может содержать аминокислотную последовательность тяжелой цепи, имеющую CDR1, CDR2, или CDR3 одной или более последовательностей, приведенных в Таблице 11, или их комбинацию. Отметим, что специалист в данной области сможет легко выполнить определения CDR. См., например, Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3.
Воплощения в соответствии с данным изобретением, описанные в данной заявке, относятся к моноклональным антителам, связывающим IGF-I/II и влияющим на функцию IGF-I/II. Другие воплощения относятся к полностью человеческим анти-IGF-I/II антителам и получению анти-IGF-I/II антител с желаемыми свойствами, с точки зрения терапевтической перспективы, включая высокую афинность связывания с IGF-I/II, способность к нейтрализации IGF-I/II in vitro и in vivo, и способность к ингибированию клеточной пролиферации, вызванной IGF-I/II.
ПОДРОБНОЕ ОПИСАНИЕ
Воплощения в соответствии с данным изобретением, описанные в данной заявке, относятся к связывающим протеинам, которые специфически связывают IGF-II и имеют перекрестную реактивность к IGF-I (в данной заявке называются "IGFI/II"). В некоторых воплощениях, связывающие протеины являются антителами, или их связывающими фрагментами, и связывают IGF-II и имеют перекрестную реактивность к IGF-I и ингибирую связывание этих протеинов с их рецептором, IGF-IR. Остальные воплощения в соответствии с данным изобретением включают полностью человеческие нейтрализующие анти-IGF-I/II антитела, и препараты антител, терапевтически полезные и связывают оба инсулиноподобные факторы роста. Такие препараты анти-IGF-I/II антител, преимущественно, имеют желательные терапевтические свойства, включая сильную афинность связывания IGF-I/II, способность к нейтрализации IGF-I/II in vitro, и способность к ингибированию вызванной IGF-I/II клеточной пролиферации in vivo.
Воплощения в соответствии с данным изобретением также включают выделенные связывающие фрагменты анти-IGF-I/II антител. Преимущественно, связывающие фрагменты получают из полностью человеческих анти-IGF-I/II антител. Иллюстративные фрагменты включают Fv, Fab(или другие хорошо известные фрагменты антител, как более подробно описано ниже. Воплощения в соответствии с данным изобретением также включают клетки, которые экспрессируют полностью человеческие антитела к IGF-I/II. Примеры клеток включают гибридомы, или рекомбинантно созданные клетки, такие, как клетки яичников хомячков (СНО), которые производят антитела к IGF-I/II.
Кроме того, воплощения в соответствии с данным изобретением включают способы использования данных антител для лечения заболеваний. Анти-IGF-I/II антитела полезны для профилактики IGF-I/II опосредованной IGF-I/II передачи сигналов, ингибируя, таким образом, клеточную пролиферацию. Механизм действия такого ингибирования IGF-I/II вследствие связывания его рецептора, IGF-IR, ингибирования IGF-I/II-вызванной IGF-IR передачи сигнала, или усиленного клиренса IGF-I/II, что приводит к понижению эффективной концентрации IGF-I/II для связывания IGF-IR. Болезни, поддающиеся лечения благодаря этому механизму ингибирования включают, не ограничиваясь приведенным, неопластические заболевания, такие как меланома, немелкоклеточный рак легких, глиома, печеночно-клеточный рак, гинекологические опухоли, рак головы и шеи, рак пищевода, глиобластома, и различные виды рака и опухолей щитовидной железы, желудка, простаты, молочной железы, яичников, мочевого пузыря, легких, матки, почек, толстой кишки и поджелудочной железы, слюнной железы и ободочной и прямой кишки.
Другие воплощения в соответствии с данным изобретением включают диагностические анализы для специфического определения количества IGF-I/II в биологической пробе. Аналитический набор может включать анти-IGF-I/II антитела наряду с необходимыми маркировками для метки таких антител. Эти диагностические анализы полезны для скрининга заболеваний, связанных с фактором роста, включая, но не ограничиваясь приведенным, такие неопластические заболевания, как меланома, немелкоклеточный рак легких, глиома, печеночно-клеточный рак, гинекологические опухоли, рак головы и шеи, рак пищевода, глиобластома и карцинома щитовидной железы, желудка, простаты, молочной железы, яичников, мочевого пузыря, легких, матки, почек, толстой кишки, и поджелудочной железы, слюнной железы, и ободочной и прямой кишки. Другими не-неопластическими состояниями могут быть акромегалия и гигантизм, псориаз, остеопороз, атеросклероз и рестеноз гладкой мускулатуры кровяных сосудов, а также диабет.
Дополнительные воплощения, характеристики и т.п., относительно анти-IGF-I/II антител дополнительно подробно описаны ниже.
Перечень последовательностей
Воплощения в соответствии с данным изобретением включают специфические анти-IGF-I/II антитела, приведенные ниже в Таблице 1. В данной таблице сообщается идентификационный номер каждого анти-IGF-I/II антитела, наряду с SEQ ID номер соответствующих генов легких и тяжелых цепей. Кроме того, последовательности зародышевых линий каждой производной легкой и тяжелой цепи также приведены ниже в Таблице 1.
Каждому антителу был присвоен идентификационный номер, который включает два или три номера, разделенные одной или двумя десятичными точками. В таких случаях, получают несколько клонов одного антитела. Хотя клоны содержат те же нуклеиновокислотные и аминокислотные последовательности, что и родительские последовательности, они также могут быть перечислены отдельно, а номер клона показан номером справа от второй десятичной точки. Таким образом, например, нуклеиновокислотные и аминокислотные последовательности антитела 7.159.2 идентичны последовательностям антитела 7.159.1.
Как можно увидеть путем сравнения последовательностей в перечне последовательностей, SEQ ID №№: 1-20 отличаются от SEQ ID №№: 39-58, поскольку SEQ ID №№: 39-58 включает нетранслированный сигнальный пептид и константный регион для каждой секвенированной тяжелой и легкой цепи.
Определения
Если не определено иное, то научные и технические термины, которые используются в данной заявке, будут иметь значения, как правило, понятные специалистам в данной области. Кроме того, если из контекста не следует иное, то единственное число будет включать множественное число, а множественное число - единственное. В общем, номенклатуры и методы, которые использовали в связи с клеточными и тканевыми культурами, молекулярной биологией и химией протеинов и олиго- и полинуклеотидов и гибридизации, описанные в данной заявке, являются хорошо известными и широко используемыми в данной области.
Для рекомбинантных ДНК, олигонуклотидного синтеза и тканевой культуры и трансформации используют стандартные методы (например, электропорацию и липофекцию). Ферментные реакции и методы очистки выполняли в соответствии с инструкциями производителя или общепринятыми методами, известными из уровня техники, или так, как описано в данной заявке. Вышеуказанные методы и способы, в общем, выполняли в соответствии с традиционными способами, хорошо известными в уровне техники и так, как описано в различных общих и более конкретных ссылках, процитированных в данной заявке, которые обсуждаются в описании к ней. См., например, Sambrook et al. Molecular Cloning: A Laboratory Manual (3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001)), которая включена в данную заявку путем ссылки. Номенклатуры и лабораторные способы и методы, которые использовали в связи с аналитической химией, синтетической органической химией, медицинской и фармацевтической химией, описанные в данной заявке, хорошо известны и широко используются в данной области. Для химического синтеза, химических анализов, фармацевтических препаратов, композиций и доставки и лечения пациентов использовали стандартные методы.
Как используется в соответствии с данным описанием, следующие термины, если не указано иное, должны быть истолкованы как имеющие следующие значения.
Термин "IGF-I" относится к молекуле инсулиноподобного фактора роста-I, а термин "IGF-II" относится к молекуле инсулиноподобного фактора-II. Термин "IGF-I/II" относится к обеим молекулам инсулиноподобного фактора роста s-I и -II, относится к предпочтительному связыванию с IGF-II и имеют перекрестную реактивность к IGF-I. Таким образом, антитело, которое связывает IGF-I/II, будет, предпочтительно, связывать IGF-II, но сможет перекрестно реагировать с IGF-I, связывая IGF-II с афинностью, превышающей афинность к IGF-I. Например, антитело может связывать IGF-II с афинностью, в 2,5 раз превышающей афинность к IGF-I. В некоторых воплощениях, антитело может связывать IGF-II с афинностью, которая, как минимум, в 5, как минимум, в 10, как минимум, в 25, как минимум, в 50 или, как минимум, в 150 раз превышает афинность к IGF-I.
Термин "нейтрализующий" при использовании по отношению к антителу относится к способности антитела уничтожать, или значительно уменьшать, активность антигена-мишени. Соответственно, "нейтрализующее" анти-IGF-I/II антитело способно к уничтожению или значительному уменьшению активности IGF-I/II. Нейтрализующее IGF-I/II антитело может, например, действовать путем блокирования связывания IGF-I/II с его рецептором IGF-IR. Путем блокирования связывания, IGF-IR-опосредованная передача сигнала в значительной степени, или полностью, уничтожается. В идеальном случае, нейтрализующее антитело к IGF-I/II ингибирует клеточную пролиферацию.
Термин "выделенный полинуклеотид", как используется в данной заявке, должен обозначать полинуклеотид, который был выделен из его природного источника. Такие полинуклеотиды могут быть геномными, кДНК или синтетическими. Выделенные полинуклеотиды, преимущественно, не связаны со всеми или с частью полинуклетидов, с которыми они связаны в природе. Выделенный полинуклеотид может быть функционально связан с другим полинуклеотидом, с которым он не связан в природе. Кроме того, выделенные полинуклеотиды, преимущественно, не встречаются в природе как часть большей последовательности.
Термин "выделенный полинуклеотид", указанный в данной заявке, означает протеин, выделенный из природного источника. Такие протеины могут быть получены из геномных ДНК, кДНК, рекомбинантной ДНК, рекомбинантной РНК или иметь синтетическое происхождение или их представлять собой их некоторую комбинацию, где в силу своего происхождения или источника получения "выделенный протеин" (1) не связан с протеинами в природе, (2) не содержит другие протеины, полученные из того же источника, т.е. не содержащие мышиные протеины, (3) экспрессируется клетками из различных видов или (4) не встречается в природе.
Термин "полипептид" используется в данной заявке в качестве родового понятия для обозначения природного протеина, фрагментов или аналогов полипептидной последовательности. Следовательно, природный протеин, фрагменты и аналоги являются видами полипептидного рода. Предпочтительные полипептиды в соответствии с данным изобретением содержат человеческие молекулы иммуноглобулина тяжелой цепи и человеческие каппа молекулы иммуноглобулина легкой цепи, а также молекулы антитела, образованные путем комбинаций, содержащих молекулы иммуноглобулина тяжелой цепи и молекулы иммуноглобулина легкой цепи, такие, как каппа или лямбда молекулы иммуноглобулина легкой цепи, и наоборот, а также их фрагменты и аналоги. Предпочтительные полипептиды, в соответствии с данным изобретением, могут также содержать только молекулы иммуноглобулина тяжелой цепи и их фрагменты и аналоги.
Термин "который встречается в природе", как используется в данной заявке, в применении к объектам, относится к тому факту, что объект может встречаться в природе. Например, полипептидная или полинуклеотидная последовательность, которая присутствует в организме (включая вирусы), которая может быть выделена из природного источника и которая не была намеренно модифицирована человеком в лаборатории или иным образом, является такой, которая встречается в природе.
Термин "функционально связанный", как используется в данной заявке, относится к положениям компонентов, описанных таким образом, что их взаимное расположение позволяет им функционировать по предназначению. Например, контрольная последовательность, которая "функционально связана" с кодирующей последовательностью, связана таким образом, что экспрессия кодирующей последовательности достигается при условиях, совместимых с контрольными последовательностями.
Термин "полинуклеотид", как используется в данной заявке, означает полимерную форму нуклеотидов, имеющих, как минимум 10 оснований в длину, рибонуклеотидов или дезоксирибонуклеотидов, или модифицированную форму любого типа нуклеотида, или гетеро-дуплексы РНК-ДНК. Термин включает одно- и двухцепочечные формы ДНК.
Термин "олигонуклеотид", как используется в данной заявке, включает встречающиеся в природе и модифицированные нуклеотиды, связанные вместе связями, которые встречаются или не встречаются в природе. Олигонуклеотиды представляют собой полинуклеотидные подмножества, в общем состоящие из 200 или менее оснований. Преимущественно, олигонуклеотиды имеют длину в 10-60 оснований и наиболее преимущественно 12, 13, 14, 15, 16, 17, 18, 19, или 20-40 оснований. Олигонуклеотиды обычно являются одноцепочечными, для проб; хотя олигонуклеотиды могут быть двухцепочечными, например, для использования при конструировании генного мутанта. Олигонуклеотиды могут быть смысловыми или антисмысловыми олигонуклеотидами.
Термин "нуклеотиды, которые встречаются в природе", как используется в данной заявке, включает дезоксирибонуклеотиды и рибонуклеотиды. Термин "модифицированные нуклеотиды", как используется в данной заявке, включает нуклеотиды с модифицированными или замещенными сахарными группами и т.п. Термин "олигонуклеотидные связи", как используется в данной заявке, включает олигонуклеотидные связи, такие как фосфоротиоат, фосфородитиоат, фосфороселенат, фосфородисленет, фосфороанилотиоат, фосфоранилидат, фосфороамидат и т.п. См., например, LaPlanche et al. Nucl. Acids Res. 14: 9081 (1986); Stec et al. J. Am. Chem. Soc. 106: 6077 (1984); Stein et al. Nucl. Acids Res. 16: 3209 (1988); Zon et al. Анти-Сапсег Drug Design 6:539 (1991); Zon et al. Onuzouywieomuds and Analogues: A Practical Approach, pp.87-108 (F. Eckstein, Ed., Oxford University Press, Oxford England (1991)); Stec et al. Патент США №5,151,510; Uhlmann and Peyman Chemical Reviews 90: 543 (1990), описания которых включены в данную заявку путем ссылок. Олигонуклеотид может включать метку для детекции, необязательно.
Термин "селективно гибридизовать", как используется в данной заявке, означает детектируемо и селективно связывать. Полинуклеотиды, олигонуклеотиды и их фрагменты селективно гибридизуют в нити нуклеиновых кислот в условиях гибридизации и промывания, которые минимизируют существенные количества детектируемого связывания с неспецифическими нуклеиновыми кислотами. Очень жесткие условия можно использовать для достижения селективных условия гибридизации, как известно из уровня техники и обсуждается в данной заявке. В общем, нуклеиновокислотная гомология между полинуклеотидами, олигонуклеотидами, или фрагментами антител и нуклеиновой кислотой, которая рассматривается, составляет, как минимум, 80%, и более типично, с преимущественно большей гомологией, которая составляет, как минимум, 85%, 90%, 95%, 99% и 100%.
Две аминокислотные последовательности являются "гомологичными", если существует частичная или полная идентичность между их последовательностями. Например, 85% гомология означает, что 85% аминокислот идентичны, если две последовательности линеаризованы для максимального совпадения. Для максимизации совпадения разрешены гэпы (в одной из последовательностей, которые совпадают); предпочтительная длина гэпов составляет 5 или менее, более предпочтительно - 2 или менее. Альтернативно и преимущественно, две протеиновые последовательности (или полипептидные последовательности, полученные из них, длиной, как минимум, приблизительно 30 аминокислот) являются гомологичными, в значении термина, используемого в данной заявке, если счет выравнивания составляет более 5 (в стандартных единицах отклонения) с использованием программы ALIGN с матрицей мутации данных и штрафом за пропуски, составляющим 6 или более. См. Dayhoff, М.О., в Atlas of Protein Sequence and Structure, pp.101-110 (Volume 5, National Biomedical Research Foundation (1972)) и Приложение 2 к этому тому, pp.1-10. Две последовательности или их части, являются, более предпочтительно, гомологичными, если их аминокислоты имеют идентичность в 50% или более при оптимальном выравнивании при помощи программы ALIGN. Должно быть оценено, что могут существовать различные области гомологии в двух ортологических последовательностях. Например, функциональные сайты ортологий мышей и человека могут иметь более высокую степень гомологии, чем нефункциональные регионы.
Термин "соответствует" используют в данной заявке для обозначения гомологичности (т.е. идентичности, не строгого эволюционного отношения) полинуклеотидной последовательности всей или части реперной полинуклеотидной последовательности или того, что полинуклеотидная последовательность идентична реперной полипептидной последовательности.
Наоборот, термин "комплементарный к" используют в данной заявке для обозначения того, что комплементарная последовательность является гомологичной всей или части реперной полинуклеотидной последовательности. В качестве иллюстрации, нуклеотидная последовательность "ТАТАС" соответствует реперной последовательности "ТАТАС" и является комплементарной реперной последовательности "GTATA".
Следующие термины используют для описания взаимосвязи последовательностей двух или более полинуклеотидных или аминокислотных последовательностей: "реперная последовательность", "окно сравнения", "идентичность последовательностей", "процент идентичности последовательностей" и "существенная идентичность". "Реперная последовательность" является определенной последовательностью, которую используют на основе сравнения последовательностей. Реперная последовательность может быть подмножеством большей последовательности, например, сегментом полноразмерной кДНК или генной последовательности, приведенной в перечне последовательностей, или может содержать полноразмерную ДНК или генную последовательность. В общем, реперная последовательность содержит, как минимум, 18 нуклеотидов или 6 аминокислот, часто, как минимум, 24 нуклеотида или 8 аминокислот, и часто, как минимум, 48 нуклеотидов или 16 аминокислот. Поскольку каждый из двух полинуклеотидов или аминокислотных последовательностей может (1) содержать последовательность (т.е. часть полной полинуклеотидной или аминокислотной последовательности), аналогичную у двух молекул и (2) может дополнительно содержать последовательность, которая отличается у двух полинуклеотидов или аминокислотных последовательностей, то сравнения последовательностей между двумя (или более) молекулами типично проводят путем сравнения последовательностей двух молекул в "окне сравнения" для идентификации и сравнения местных областей идентичности последовательностей. "Окно сравнения", как используется в данной заявке, относится к концептуальному сегменту из, как минимум, приблизительно, 18 соседних нуклеотидных положений или, приблизительно, 6 аминокислот, где полинуклеотидную последовательность или аминокислотную последовательность сравнивают с реперной последовательностью из, как минимум 18 соседних нуклеотидных или 6 аминокислотных последовательностей и где часть полинуклеотидной последовательности в окне сравнения может включать добавления, делеции, замещения и т.п. (т.е. гэпы) 20 или менее процентов по сравнению с реперной последовательностью (которая не содержит добавления или делеций) для оптимального выравнивания двух последовательностей. Оптимальное выравнивание последовательностей для выравнивания в окне сравнения может быть проведено при помощи алгоритма локальной гомологии Smith и Waterman Adv. Appl. Math. 2:482 (1981), при помощи алгоритма выравнивания гомологии Needleman и Wunsch J. Mol. Biol. 48: 443 (1970), путем поиска способа подобия Pearson и Lipman Proc. Natl. Acad. Sci. (U.S.A.) 85: 2444 (1988), путем компьютеризированных воплощения этих алгоритмов (GAP, BESTFIT, FASTA, и TFASTA в Wisconsin Genetics Software Package Release 7.0 (Genetics Computer Group, 575 Science Dr., Madison, Wis.), GENEWORKS™, или программное обеспечение MACVECTOR®), или выбирают путем проверки, или наилучшего выравнивания (т.е. приводящему к наиболее высокому проценту гомологии в окне сравнения), которое можно получить путем применения различных способов.
Термин "идентичность последовательностей" означает, что две полинуклеотидные или аминокислотные последовательности являются идентичными (т.е. та основе нуклеотид-нуклеотид или остаток-остаток) в окне сравнения. Термин "процент идентичности последовательностей" рассчитывают путем сравнения двух оптимально выровненных последовательностей в окне сравнения, с определением количества положений, в которых находятся идентичные нуклеиновокислотные основания (например, А, Т, С, G, U, или I) или аминокислотные остатки в двух последовательностях для получения количества совпавших положений, разделив количество совпавших положений на общее количество положений в окне сравнения (т.е. на размер окна) и умножив на 100 для получения процента идентичности последовательностей. Термины "существенная идентичность", как используется в данной заявке, означает характеристику полинуклеотидной или аминокислотной последовательности, где полинуклеотид или аминокислота содержит последовательность, которая имеет, как минимум, 85 процентов идентичности последовательностей, преимущественно, как минимум, 90-95 процентов идентичности последовательностей, более предпочтительно, как минимум, 99 процентов идентичности последовательностей, по сравнению с реперной последовательностью в окне сравнения из, как минимум, 18 нуклеотидных (6 аминокислотных) положений, часто в окне из, как минимум, 24-48 нуклеотидных (8-16 аминокислотных) положений, где процент идентичности последовательностей рассчитывают путем сравнения реперной последовательности с последовательностью, которая может включать делеции или добавления, составляющие в сумме 20 или менее процентов от реперной последовательности в окне сравнения. Реперная последовательность может представлять собой подмножество большей последовательности.
Как используется в данной заявке, двадцать традиционных аминокислот и их аббревиатуры имеют традиционное использование. См. Immunology - A Synthesis (2nd Edition, E.S. Golub and D.R. Gren, Eds., Sinauer Associates, Sunderland, Mass. (1991)), включенную в данную заявку путем ссылок. Стереоизомеры (например, D-аминокислоты) двадцати традиционных аминокислот, неприродных аминокислот, таких, как α-, α-дизамещенные аминокислоты, N-алкильные аминокислоты, лактоновые кислоты, и другие нетрадиционные аминокислоты могут также быть пригодными компонентами для полипептидов в соответствии с данным изобретением. Примеры нетрадиционных аминокислот включают: 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, О-фосфосерин, N-ацетилсерин, N-формулметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие подобные амино- и иминокислоты (например, 4-гидроксипролин). При обозначении полипептидов, используемых в данной заявке, левостороннее направление является N-концевым направлением, а правостороннее направление - карбокси-концевым направлением, в соответствии со стандартным использованием.
Аналогично, если не указано иное, левый конец одноцепочечной полинуклеотидной последовательности является 5(концом; левостороннее направление двухцепочечной полинуклеотидной последовательности называется 5(направление. Направление 5′-3(добавления образующихся РНК транскриптов называется направлением транскрипции; области последовательности нити ДНК, имеющие такие же последовательности, что и РНК, которые являются 5′-5(концами транскрипта РНК, называются "верхними последовательностями"; области последовательности нити ДНК, имеющие такие же последовательности, что и РНК, которые являются 3′-3(концами транскрипта РНК, называются "нижними последовательностями".
Как применяется по отношению к полипептидам, термин "существенно идентичный" означает, что пептидные последовательности, при оптимальном выравнивании, например, при помощи программ GAP или BESTFIT с использованием веса гэпов по умолчанию, имеют, как минимум, 80 процентов идентичности последовательностей, преимущественно, как минимум, 90 процентов идентичности последовательностей, более предпочтительно, как минимум 95 процентов идентичности последовательностей, и наиболее преимущественно, как минимум, 99 процентов идентичности последовательностей. Преимущественно, положения остатков, которые не являются идентичными, отличаются консервативными аминокислотными замещениями. Консервативные аминокислотные замещения относятся к взаимозаменяемости остатков с аналогичными боковыми цепями. Например, группа аминокислот, имеющая алифатические боковые цепи, представляет собой глицин, аланин, валин, лейцин и изолейцин; группа аминокислот, имеющая алифатически-гидроксильные боковые цепи, представляет собой серин и треонин; группа аминокислот, имеющая амид-содержащие боковые цепи, представляет собой аспарагин и глутамин; группа аминокислот, имеющая ароматические боковые цепи, представляет собой фенилаланин, тирозин и триптофан; группа аминокислот, имеющая основные боковые цепи, представляет собой лизин, аргинин и гистидин; группа аминокислот, имеющая серосодержащие боковые цепи, представляет собой цистеин и метионин. Преимущественными группами замещения консервативных аминокислот являются: валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, глутамо-аспаркамовая, и аспарагин-глутамин.
Как обсуждается в данной заявке, минимальные вариации аминокислотных последовательностей антител или молекул иммуноглобулина рассматривают как такие, которые описаны в данном изобретении, при условии, что вариации аминокислотных последовательностей сохраняют, как минимум, 75%, более предпочтительно, как минимум, 80%, 90%, 95%, и наиболее преимущественно, 99% идентичности последовательностей антител или молекул иммуноглобулина, описанных в данной заявке. В частности, рассматриваются консервативные аминокислотные замещения. Консервативными замещениями являются такие, которые происходят в семействе аминокислот с родственными боковыми цепями. Генетически кодированные аминокислоты, в общем, разделяют на семейства: (1) кислотное=аспартат, глутамат; (2) основное=лизин, аргинин, гистидин; (3) неполярное=аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженное полярное=глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин. Более предпочтительными семействами являются: серин и треонин представляют собой алифатически-гидроксильное семейство; аспарагин и глутамин представляют собой амид-содержащее семейство; аланин, валин, лейцин и изолейцин представляют собой алифатическое семейство; и фенилаланин, триптофан и тирозин представляют собой ароматическое семейство. Например, целесообразно ожидать, что выделенное замещение лейцина на изолейцин или валин, аспартата на глутамат, треонина на серин или аналогичное замещение аминокислоты на структурно родственную аминокислоту не будет иметь большого влияния на функцию связывания или свойства результирующей молекулы, особенно если в замещении не участвует аминокислота в каркасном сайте. Может ли замена аминокислоты привести к образованию функционального пептида, можно легко определить путем анализа специфической активности производного полипептида. Анализы подробно описаны в данной заявке. Фрагменты или аналоги антител или молекул иммуноглобулина могут быть легко получены специалистом в данной области. Преимущественные амино- и карбокси-концы фрагментов или аналогов расположены около границ функциональных доменов. Структурные и функциональные домены можно идентифицировать путем сравнения данных нуклеотидных и/или аминокислотных последовательностей, который имеются в государственных или частных базах данных последовательностей. Преимущественно, используют компьютеризованные способы сравнения для идентификации фрагментов последовательностей или предсказанных конформаций доменов протеинов, которые встречаются в других протеинах известной структуры и/или функции. Известны способы идентификации протеиновых последовательностей, подпадающих по известные трехмерные структуры. Bowie et al. Science 253: 164 (1991). Поэтому приведенные примеры демонстрируют, что специалист в данной области может распознать фрагменты последовательностей и структурные конформаций, которые могут быть использованы для определения структурных и функциональных доменов в соответствии с антителами, описанными в данной заявке.
Преимущественными аминокислотными замещениями являются те, при которых: (1) уменьшается чувствительность к протеолизу, (2) уменьшается чувствительность к окислению, (3) изменяется афинность связывания для образования протеиновых комплексов, (4) изменяется афинность связывания, и (5) образуются или изменяются другие физико-химические или функциональные свойства таких аналогов. Аналоги могут включать различные мутантные последовательности, отличные от пептидных последовательностей, встречающихся в природе. Например, одно- или многократные аминокислотные замещения (преимущественно, консервативные аминокислотные замещения) могут быть осуществлены в последовательностях, которые встречаются в природе (преимущественно, в части полипептида вне домена (доменов)) образуют межмолекулярные контакты. Консервативное аминокислотное замещение не должно существенно изменять структурные характеристики родительской последовательности (например, аминокислотное замещение не должно быть направлено на разрушение спирали, имеющейся в родительской последовательности, или разбивание других типов вторичных структур, которые характеризуют родительскую последовательность). Примеры признанных в уровне техники вторичных и третичных структур описаны в Proteins, Structures and Molecular Principles (Creighton, Ed., W.H. Freeman and Company, New York (1984)); Introduction to Protein Structure (C. Branden and J. Tooze, eds., Garland Publishing, New York, N.Y. (1991)); и Thornton et at. Nature 354: 105 (1991), каждая из которых включена в данную заявку путем ссылок.
Термин "полипептидный фрагмент", как используется в данной заявке, относится к полипептиду, имеющему N-концевую и/или карбокси-концевую делецию, но где оставшаяся аминокислотная последовательность идентична соответствующим положениям последовательности, встречающейся в природе, полученной, например, из полноразмерной кДНК последовательности. Фрагменты типично имеют длину, как минимум, 5, 6, 8 или 10 аминокислот, преимущественно, как минимум, 14 аминокислот, более предпочтительно, как минимум, 20 аминокислот, обычно, как минимум, 50 аминокислот, и даже более предпочтительно, как минимум, 70 аминокислот. Термин "аналог", как используется в данной заявке, относится к полипептидам, которые состоят из сегмента из, как минимум, 25 аминокислот, имеющих существенную идентичность с частью полученной аминокислотной последовательности и которые имеют, как минимум, одно из следующих свойств: (1) специфическое связывание IGF-I/II, в пригодных условиях связывания, (2) способность блокировать соответствующее IGF-I/II связывание, или (3) способность ингибировать активность IGF-I/II. Типично, полипептидные аналоги содержат консервативное аминокислотное замещение (или добавление или делецию) по отношению к последовательности, которая встречается в природе. Аналоги, типично, имею минимальную длину в, как минимум, 20 аминокислот, преимущественно, как минимум, 50 аминокислот или более, и могут часто иметь такую же длину, что и полноразмерный полипептид, встречающийся в природе.
Пептидные аналоги традиционно используют в фармацевтической промышленности в качестве непептидных лекарств, свойства которых аналогичны свойствам темплатных пептидов. Эти типы непептидных соединений называются "пептидомиметики" или "пептидные миметики". Fauchere, J. Adv. Drug Res. 15: 29 (1986); Veber and Freidinger TINS p.392 (1985); и Evans et al. J. Med. Chem. 30: 1229 (1987), которые включены в данную заявку путем ссылок. Такие соединения часто разрабатывают при помощи компьютерного молекулярного моделирования. Пептидомиметики, структурно подобные терапевтически полезным пептидам, могут быть полезными для получения эквивалентного терапевтического или профилактического эффекта. В общем, пептидомиметики, структурно подобны реперным полипептидам (т.е. полипептидам, имеющим биохимические свойства или фармакологическую активность), таким как человеческое антитело, но имеют одну или более пептидных связей, необязательно замещенных связью, которую выбирают из группы, состоящей из: --CH2NH--, --CH2S--, --СН2-СН2--, -CH=CH-- (цис и транс), --COCH2--, --CH(OH)CH2- и -CH2SO--, способами, хорошо известными из уровня техники. Систематическое замещение одной или более аминокислот непрерывной последовательности на D-аминокислоту того же типа (например, D-лизин вместо L-лизина) можно использовать для получения более стабильных пептидов. Кроме того, ограниченные пептиды, содержащие непрерывную последовательность или существенно идентичную вариацию непрерывной последовательности, могут быть получены способами, известными из уровня техники (Rizo and Gierasch Ann. Rev. Biochem. 61: 387 (1992), включена в данную заявку путем ссылок); например, путем добавления внутренних цистеиновых остатков, способных к образованию внутримолекулярных сульфидных мостиков, циклизующих пептид.
Как используется в данной заявке, термин "антитело" относится к полипептиду или группе полипептидов, состоящих из, как минимум, одного связывающего домена, образованного путем сгибания полипептидных цепей, имеющих трехмерные связывающие расстояния, форма внутренних поверхностей и распределение заряда которых комплементарны характеристикам антигенной детерминанты антигена. Антитело, типично, имеет тетрамерную форму, содержит две идентичные пары полипептидных цепей, каждая пара имеет одну "легкую" и одну "тяжелую" цепь. Вариабельные регионы каждой легкой/тяжелой цепи образуют сайт связывания антитела.
"Связывающие фрагменты" антитела получают при помощи рекомбинантных ДНК методик, или ферментным или химическим расщеплением интактных антител. Связывающие фрагменты включают Fab, Fab′, F(ab′)2, Fv, и одноцепочечные антитела. Антитело, иное, чем "биспецифичное" или "бифункциональное" антитело истолковывают как такое, у которого все сайты связывания идентичные. Антитело существенно ингибирует адгезию рецептора к противорецептору, если избыток антитела снижает количество рецептора, связанного с противорецепотором, на, как минимум, приблизительно, 20%, 40%, 60% или 80%, и более традиционно, более чем, приблизительно, 85% (в соответствии с результатами измерений конкурентного анализа связывания in vitro).
Как используется в данной заявке, "связывающий протеин" или "специфически связывающий протеин" представляют собой протеины, которые специфически связывают молекулу-мишень. Антитела и связывающие фрагменты антител являются связывающими протеинами.
Термин "эпитоп" включает любой протеиновый детерминант, способный специфически связывать иммуноглобулин или рецептор Т-клеток. Эпитопные детерминанты обычно состоят из химически активных поверхностных групп молекул, таких, как аминокислоты или сахарные боковые цепи и могут, но не всегда, иметь специфические трехмерные структурные характеристики, а также специфичные зарядные характеристики. Говорят, что антитело специфически связывает антиген, если константа диссоциации ≤1 мкМ, преимущественно ≤100 нМ и наиболее преимущественно ≤10 нМ.
Термин "агент" используют в данной заявке для обозначения химического соединения, смеси химических соединений, биологической макромолекулы, или экстракта, полученного из биоматериала.
"Активный" или "активность" по отношению к IGF-I/II полипептиду относится к части IGF-I/II полипептида, имеющей биологическую или иммунологическую активность природного IGF-I/II полипептида. "Биологический", как используется в данной заявке, относится к биологической функции, которая является результатом активности природного IGF-I/II полипептида. Преимущественная биологическая активность IGF-I/II включает, например, IGF-I/II-вызванную клеточную пролиферацию.
"Млекопитающий", как используется в данной заявке, относится к любому животному, которое считается млекопитающим. Преимущественно, млекопитающим является человек.
Дигестия антител ферментом, папаином, приводит к образованию двух идентичных антиген-связывающих фрагментов, также известных как "Fab" фрагменты, и "Fc" фрагмента, не имеющих антиген-связывающей активности, но имеющих способность к кристаллизации. Дигестия антител ферментом, пепсином, приводит к образованию F(ab′)2 фрагмента, в котором два ответвления антитела остаются связанными и содержат два антиген-связывающих сайта. Фрагмент F(ab′)2 имеет способность перекрестно связывать антиген.
"Fv", как используется в данной заявке, относится к минимальному фрагменту антитела, который сохраняет как антиген-распознающие, так и антиген-связывающие сайты.
"Fab", как используется в данной заявке, относится к фрагменту антитела, который содержит константный домен легкой цепи и CH1 домен тяжелой цепи.
Термин "mAb" относится к моноклональному антителу.
"Липосома", как используется в данной заявке, относится к небольшому пузырьку, который может использоваться для доставки лекарств, и может включать IGF-I/II полипептидов соответствии с данным изобретением ил антитела к такому IGF-I/II полипептиду у млекопитающих.
"Метка" или "меченый", как используется в данной заявке, относится к добавлению детектируемой группы к полипептиду, например, радиоактивной метки, флуоресцентной метки, ферментной метки, хемилюминесцентной метки или биотионильной группы. Радиоактивные изотопы или радионуклиды могут включать 3Н, 14С, 15N, 35S, 90Y, 99Тс, 111In, 125I, 131L флуоресцентные метки могут включать родамин, лантанид фосфора или FITC, а ферментные метки могут включать пероксидазу хрена, β-галактозидазу, люциферазу и щелочную фосфатазу.
Термин "фармацевтическое средство или лекарство", как используется в данной заявке, относится к химическому соединению или композиции, способным вызывать желательный терапевтический эффект при должном введении пациенту. Другие химические термины в данной заявке употребляют так, как это принято в данной области, как показано в The McGraw-Hill Dictionary of Chemical Terms (Parker, S., Ed., McGraw-Hill, San Francisco (1985)), (включена в данную заявку путем ссылок).
Как используется в данной заявке, "существенно чистый" означает доминирующее присутствие видов объектов (т.е. с молярной точки зрения, большее присутствие, чем любой другой отдельный вид в композиции), и, преимущественно, существенно очищенной фракцией является композиция, где виды объектов составляют, как минимум, около 50 процентов (с молярной точки зрения) всех макромолекулярных присутствующих видов. В общем, существенно чистая композиция будет содержать более чем, приблизительно, 80% от всех макромолекулярных видов, присутствующих в композиции, более предпочтительно более чем, приблизительно, 85%, 90%, 95% и 99%. Наиболее преимущественно, виды объектов очищены до существенной гомогенности (загрязнения не могут быть обнаружены в композиции при помощи традиционных способов детекции), и композиция состоит, существенно, из одного макромолекулярного вида.
Термин "пациент" включает человеческие и ветеринарные объекты.
Человеческие антитела и гуманизация антител
У человеческих антител отсутствуют некоторые проблемы, связанные с антителами, которые имеются у мышиных или крысиных вариабельных и/или константных регионов. Наличие таких полученных из мышей или крыс протеинов может привести к быстрому клиренсу антител или получению иммунного ответа пациента на антитело. Во избежание использования антител, полученных из мышей или крыс, могут быть получены полностью человеческие антитела путем введения локусов функционального человеческого антитела в грызуна, другого млекопитающего или животное таким образом, чтобы млекопитающее или животное продуцировало полностью человеческие антитела.
Одним из способов получения полностью человеческих антител является использование штаммов мыши XenoMouse®, сконструированных таким образом, чтобы содержать менее 1000 kb-размерных зародышевых линий сконфигурированных фрагментов локуса человеческой тяжелой цепи и локуса каппа легкой цепи. См. Mendez et al. Nature Genetics 15: 146-156 (1997) и Green and Jakobovits J. Exp. Med. 188: 483-495 (1998). Штаммы XenoMouse® доступны у Abgenix, Inc. (Fremont, CA).
Получение штаммов мышей XenoMouse® дополнительно обсуждается в патентных заявках США №№07/466,008, поданной 12 января 1990 г., 07/610,515, поданной 8 ноября 1990 г., 07/919,297, поданной 24 июля 1992 г., 07/922,649, поданной 30 июля 1992 г., 08/031,801, поданной 15 марта 1993 г., 08/112,848, поданной 27 августа 1993 г., 08/234,145, поданной 28 апреля 1994 г., 08/376,279, поданной 20 января 1995 г., 08/430, 938, поданной 27 апреля 1995 г., 08/464,584, поданной 5 июня 1995 г., 08/464,582, поданной 5 июня 1995 г., 08/463,191, поданной 5 июня 1995 г., 08/462,837, поданной 5 июня 1995 г., 08/486,853, поданной 5 июня 1995 г., 1995, 08/486,857, поданной 5 июня 1995 г., 08/486,859, поданной 5 июня 1995 г., 08/462,513, поданной 5 июня 1995 г., 08/724,752, поданной 2 октября 1996 г., 08/759,620, поданной 3 декабря 1996 г., публикации США 2003/0093820, поданной 30 ноября 2001 г. и патентах США №№6,162,963, 6,150,584, 6,114,598, 6,075,181, и 5,939,598 и в японских патентах №№3068180 B2, 3068506 B2, и 3068507 B2. См. также Европейский патент №EP 0463151 B1, опубликованный 12 июня 1996 г., международную патентную заявку №WO 94/02602, опубликованную 3 февраля 1994 г., международную патентную заявку №WO 96/34096, опубликованную 31 октября 1996 г., WO 98/24893, опубликованную 11 июня 1998 г., WO 00/76310, опубликованную 21 декабря, 2000. Описания каждого из вышеуказанных патентов, заявок и ссылок включены полностью в данную заявку путем ссылок.
В альтернативном подходе, остальные, включая GenPharm International, Inc., использовали "минилокусный" подход. В этом подходе, экзогенный Ig локус имитировали посредством включения частей (отдельных генов) из локуса Ig. Поэтому один или более VH генов, один или более DH генов, один или более JH генов, mu константный регион, и, обычно, второй константный регион (преимущественно, гамма-константный регион) образуют конструкт для инсерции в животное. Этот патент описан в патенте США №5,545,807, выданном Surani et al., и патентах США №№5,545,806, 5,625,825, 5,625,126, 5,633,425, 5,661,016, 5,770,429, 5,789,650, 5,814,318, 5,877,397, 5,874,299, и 6,255,458, каждый из которых выдан Lonberg и Kау, патенте США №5,591,669 и 6,023.010, выданных Krimpenfort и Berns, патентах США №№5,612,205, 5,721,367 и 5,789,215 выданных Berns et al., и патенте США №5,643,763, выданном Choi и Dunn, и патентных заявках США GenPharm International, серийные №№07/574,748, поданной 29 августа 1990 г., 07/575,962, поданной 31 августа 1990 г., 07/810,279, поданной 17 декабря 1991 г., 07/853,408, поданной 18 марта 1992 г., 07/904,068, поданной 23 июня 1992 г., 07/990,860, поданной 16 декабря 1992 г., 08/053,131, поданной 26 апреля 1993 г., 08/096,762, поданной 22 июля 1993 г., 08/155,301, поданной 18 ноября 1993 г., 08/161,739, поданной 3 декабря 1993 г., 08/165,699, поданной 10 декабря 1993 г., 08/209,741, поданной 9 марта 1994 г., описания которых включены в данную заявку путем ссылок. См. также Европейский патент №0546073 B1, международную патентную заявку №№WO 92/03918, WO 92/22645, WO 92/22647, WO 92/22670, WO 93/12227, WO 94/00569, WO 94/25585, WO 96/14436, WO 97/13852, WO 98/24884 и патент США №5,981,175, описания которых полностью включены в данную заявку путем ссылок. См. также Taylor et al., 1992, Chen et al., 1993, Tuaillon et al., 1993, Choi et al., 1993, Lonberg et al., (1994), Taylor et al., (1994), and Tuaillon et al., (1995), Fishwild et al., (1996), описания которых полностью включены в данную заявку путем ссылок.
Kirin также продемонстрировал получение человеческих антител из мышей, в которых, посредством микроклеточного слияния, были введены большие части хромосом или целые хромосомы. См. европейские патентные заявки №№773288 и 843961, описания которых включены в данную заявку путем ссылок. Дополнительно, были получены KМ™-мыши, полученные в результате скрещивания мышей Kirin Тс с минилокусными Medarex мышами (Humab). Эти мыши имеют человеческие трансхромосомы IgH мышей Kirin и каппа цепной трансген мышей Genpharm (Ishida et al., Cloning Stem Cells (2002), 4: 91-102).
Человеческие антитела также могут быть получены способами in vitro. Приемлемые примеры включают, не ограничиваясь приведенным, фаговый дисплей (CAT, Morphosys, Dyax, Biosite/Medarex, Xoma, Symphogen, Alexion (ранее Proliferon), Affimed) рибосомный дисплей (CAT), дрожжевой дисплей и т.п.
Получение антител
Антитела, как описано в данной заявке, были получены с использованием технологии XenoMouse®, описанной ниже. Такие мыши затем способны к продуцированию человеческих молекул иммуноглобулина и антител, дефицитных при продуцировании мышиных молекул иммуноглобулина и антител. Технологии, использованные для этого, описаны в патентах, заявках и ссылках, описанных в разделе «Предпосылки создания изобретения» данной заявки. В частности, однако, преимущественное воплощение трансгенного продуцирования мышей и их антител, описано в патентной заявке США, серийный номер №08/759,620, поданной 3 декабря 1996 р. и в международной патентной заявке №№WO 98/24893, опубликованной 11 июня 1998 г. и WO 00/76310, опубликованной 21 декабря 2000 г., описания которых включены в данную заявку путем ссылок. См. также Mendez et al., Nature Genetics 15: 146-156 (1997), описаний которых включены в данную заявку путем ссылок.
Посредством использования такой технологии были получены полностью человеческие моноклональные антитела к множеству антигенов. Главным образом, линии мышей XenoMouse® иммунизировали антигеном, который рассматривается (например, IGF-I/II), лимфатические клетки (такие как В-клетки) восстанавливали из гипер-иммунизированных мышей, а восстановленные лимфоциты сливали с линями клеток миелоидного типа для получения иммортализованных гибридомных клеточных линий. Эти гибридомные клеточные линии подвергали скринингу и отбирали для идентификации клеточных линий, которые продуцировали антитела, специфические для антигена, который рассматривается. В данной заявке обеспечиваются способы продуцирования множественных гибридомных клеточных линий, которые продуцируют антитела, специфические к IGF-I/II. Дополнительно, в данной заявке обеспечиваются характеристики антител, продуцируемых такими клеточными линиями, включая анализ нуклеотидных и аминокислотных последовательностей тяжелых и легких цепей таких антител.
Альтернативно, вместо слияния с миеломными клетками для получения гибридом, В клетки могут быть проанализированы непосредственно. Например, CD19+B клетки могут быть выделены из гипериммунных мышей XenoMouse® и оставлены для пролиферации и дифференциации в антитело-секреторные плазмовые клетки. Антитела из клеточных супернатантов затем подвергали скринингу при помощи ELISA на предмет реактивности по отношению к IGF-I/II иммуногену. Супернатанты могут также быть подвергнуты скринингу на иммунореактивность в отношении фрагментов IGF-I/II для дополнительного нанесения на карту различных антител для связывания с доменами функционального интереса IGF-I/II. Антитела могут также быть подвергнуты скринингу на остальные родственный человеческие хемокины и по отношению к крысам, мышам и нечеловеческим приматам, таким, как яванские макаки, ортологичные к IGF-I/II, последний для определения перекрестной реактивности видов. В клетки из лунок, содержащих антитела, которые рассматриваются, могут быть иммортализованы различными способами, включая слияние для получения гибридом от отдельных или объединенных лунок, или путем инфицирования EBV или трансфекции известными генами иммортализации, и последующего помещения в приемлемую среду. Альтернативно, отдельные клетки плазмы, секретирующие антитела с желаемыми свойствами, затем выделяли с использованием IGF-I/II-специфического гемолитического анализа бляшкообразования (Babcook et al., Proc. Natl. Acad. Sci. USA 93: 7843-48 (1996)). Цели-мишени лизиса представляют собой, преимущественно, красные кровяные клетки овец (SRBCs), покрытые антигеном IGF-I/II.
В присутствии культуры B-клеток, содержащей клетки плазмы, секретирующие иммуноглобулин, который рассматривается, и комплемент, образование бляшек указывает на опосредованный специфичным IGF-I/II лизис красных кровяных клеток овец, в окружении клеток плазмы, которые рассматриваются. Отдельная антиген-специфичная клетка плазмы в центре бляшки может быть выделена, а генетическая информация, кодирующая специфичность антитела, выделена из отдельной клетки плазмы. С использованием обратной транскрипции, с последующей PCR (RT-PCR), может быть клонирована ДНК, кодирующая вариабельные области тяжелой и легкой цепей антитела. Такая клонированная ДНК может быть затем вставлена в приемлемый вектор экспрессии, преимущественно, векторную кассету, такую как рсДНК, более предпочтительно, такую, как рсДНК вектор, содержащий константные домены тяжелой и легкой цепей иммуноглобулина. Сгенерированный вектор может быть затем трансфектирован в клетки-хозяева, например, НЕK293 клетки, СНО клетки, и культивирован в традиционной питательной среде, модифицированной надлежащим образом для индуцирования транскрипции, отбора трансформантов или амплификации генов, кодирующих желательные последовательности.
В общем, антителами, которые были продуцированы слитыми гибридомами, были тяжелые цепи человеческого IgG2 с полностью человеческими каппа или лямбда легкими цепями. Антитела, описанные в данной заявке, имеют человеческие IgG4 тяжелые цепи, а также IgG2 тяжелые цепи. Антитела также могут быть антителами других человеческих изотипов, включая IgG1. Антитела имеют высокую афинность, типично, имеют Kd от, приблизительно, 10-6 до, приблизительно, 10-10 М или ниже, при измерениях по твердофазным или жидкофазным методикам. Антитела, имеющие KD, как минимум, 10-11 М являются предпочтительными для ингибирования активности IGF-MI.
Как будет оценено, анти-IGF-I/II антитела могут экспрессироваться в клеточных линиях, отличных от гибридомных клеточных линий. Последовательности, кодирующие конкретные антитела, могут быть использованы для трансформации приемлемых клеток-хозяев млекопитающих. Трансформация может быть проведена любыми известными способами введения полинуклеотидов в клетку-хозяин, включая, например, упаковку полинуклеотида в вирус (или вирусный вектор) и трансдуцирование клетку-хозяин с вирусом (или вектором) или путем трансфекции методами, известными из уровня техники, как показано в патентах США №№4,399,216, 4,912,040, 4,740,461 и 4,959,455 (эти патенты включены в данную заявку путем ссылок). Процедура трансформации, использованная здесь, зависит от хозяина, который будет трансформирован. Способы введения гетерологических полинуклеотидов в клетки млекопитающих хорошо известны из уровня техники и включают декстран-опосредованные трансфекции, осаждение фосфатом кальция, полибрен-опосредованную трансфекцию, протопластное слияние, электропорацию, капсулирование полинуклеотида(ов) в липосомы и прямую микроинъекцию ДНК в ядра.
Клеточные линии млекопитающих, присутствующие в качестве хозяев экспрессии, хорошо известны из уровня техники и включают многие иммортализованные клеточные линии, доступные в Американской коллекции типовых культур (АТСС), включая, но не ограничиваясь приведенным, клетки яичников хомячков (СНО), клетки HeLa, клетки почек детенышей хомячков (ВНK), клетки почек обезьян (COS), клетки человеческого печеночно-клеточного рака (например, Hep G2), эпителиальные клетки человеческих почек 293 и ряд других клеточных линий. Особенно преимущественные клеточные линии отбирают посредством определения клеточных линий с высокими уровнями экспрессии, которые продуцируют антитела со значительными IGF-I/II связывающими свойствами.
Анти-IGF-I/II антитела полезны для детекции IGF-I/II в пробах пациентов и, соответственно, полезны для диагностики болезненных состояний, как описано в данной заявке. Кроме того, основываясь на их способности значительно нейтрализовать активность IGF-I/II (как показано в Примерах, приведенных ниже), анти-IGF-I/II антитела имеют терапевтический эффект для лечения симптомов и состояний, возникших в результате экспрессии IGF-I/II. В конкретных воплощениях, антитела и способы, приведенные в данной заявке, относятся к лечению симптомов, возникших в результате клеточной пролиферации, вызванной IGF-I/II. Дополнительные воплощения включают использование антител и способов, описанных в данной заявке, для лечения заболеваний, включая неопластические заболевания, такие как меланома, немелкоклеточный рак легких, глиома, печеночно-клеточный рак, опухоль щитовидной железы, рак желудка, рак простаты, рак молочной железы, рак яичников, рак мочевого пузыря, рак легких, глиобластому, рак эндометрия, рак почек, рак толстой кишки, гинекологические опухоли, рак головы и шеи, рак пищевода и рак поджелудочной железы. Другими не-неопластическими болезненными состояниями могут быть акромегалия и гигантизм, псориаз, остеопороз, атеросклероз и рестеноз гладкой мускулатуры кровяных сосудов, а также диабет.
Терапевтическое введение и композиции
Воплощения в соответствии с данным изобретением включают стерильные фармацевтические композиции анти-IGF-I/II антитела, полезные для лечения заболеваний. Такие композиции могут ингибировать связывание IGF-I/II с его рецептором IGF-IR, обеспечивая, таким образом, эффективное лечение патологических состояний, где, например, ненормально повышены сыворотка или ткани IGF-I/II. Анти-IGF-I/II антитела, преимущественно, имеют соответствующую афинность для возможной нейтрализации IGF-I/II, и, преимущественно, имеют адекватную длительность действия, что позволяет уменьшить дозировку для людей. Длительный период воздействия позволит уменьшить частоту введений путем применения альтернативных маршрутов парентерального введении, таких, как подкожное или внутримышечное.
Стерильные композиции могут быть созданы, например, путем фильтрования через стерильные фильтровальные мембраны, до или после липофилизации и восстановления антитела. Обычно тело сохраняют в лиофилизованном виде или в растворе. Терапевтические композиции антител, в общем, помещают в контейнер со стерильным портом доступа, например, контейнер или виалу с внутривенным раствором, с адаптером, которые позволяет извлечь композицию, где пробка является проницаемой для иглы для подкожных вливаний.
Маршрут введения антитела соответствует известным способам, например внутривенному, интраперитонеальному, интрацеребральному, внутримышечному, интраокулярному, внутриартериальному, интратекальному, ингаляционному введению или вливанию, или введению или вливанию в пораженные ткани, или при помощи систем замедленного высвобождения, как отмечено ниже. Антитело, преимущественно, вводят непрерывно путем вливания или болюсной инъекции.
Эффективное количество антитела, которое будет введено терапевтически, зависит, например, от целей лечения, маршрута введения, и состояния пациента. Соответственно, преимущественным является, чтобы врач титровал дозировку и модифицировал маршрут введения, как это требуется для получения оптимального терапевтического эффекта. Типично, врач будет вводить антитело до достижения дозировки, обеспечивающей желательный эффект. Успех лечения легко оценить при помощи традиционных способов анализа, или способов анализа, описанных ниже в данной заявке.
Антитела, как описано в данной заявке, могут быть получены в смеси с фармацевтически приемлемым носителем. Эта терапевтическая композиция может быть введена внутривенно или через нос или через легкие, преимущественно, как жидкий или порошкообразный аэрозоль (лиофилизованный). Композиция может быть также введена парентерально или подкожно, по желанию. При системном введении терапевтическая композиция должна быть стерильной, не содержать пирогенов и находиться в парентерально приемлемом растворе с приемлемым значением pH, изотоничности и стабильности. Эти условия известны специалистам в данной области. Кратко, составы дозировок соединений, описанных в данной заявке, получают для хранения или введения путем смешения соединения желаемой степени чистоты с физиологически приемлемыми носителями, наполнителями или стабилизаторами. Эти материалы не токсичны для пациентов при дозировках и концентрациях, в которых применяются, и содержат такие буферы, как TRIS HCl, фосфат, цитрат, ацетат и другие органические соли; такие антиоксиданты, как аскорбиновая кислота; низкомолекулярные (менее, приблизительно, десяти остатков) пептиды, такие как полиаргинин, протеины, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамовая кислота, аспарагиновая кислота или аргинин; моносахариды, дисахариды и другие углеводы, включая целлюлозу или ее производные, глюкозу, маннозу или декстрины; хелаторы, такие как EDTA; сахарные спирты, такие как маннитол или сорбитол; противоионы, такие как натриевые и/или неионные поверхностно-активные вещества, такие как TWEEN, PLURONICS или полиэтиленгликоль.
Стерильные композиции для инъекций могут быть составлены в соответствии с традиционными фармацевтическими процедурами, как описано у Remington: The Science and Practice of Pharmacy (20th ed, Lippincott Williams & Wilkens Publishers (2003)). Например, желательными могут быть раствор или суспензия активного вещества в основе, такой как вода или природное растительное масло, такое как кунжутное, кокосовое или хлопковое, или синтетические жирные основы, такие как этилолеат и т.п. Буферы, консерванты, антиоксиданты и т.п. могут быть включены в соответствии с принятыми фармацевтическими процедурами.
Приемлемые примеры препаратов замедленного высвобождения включают полунепроницаемые матрицы твердых гидрофобных полимеров, содержащих полипептид, где матрицы находятся в форме профилированных изделий, пленок или микрокапсул. Примеры матриц замедленного высвобождения включают полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат), как описано у Langer et al., J. Biomed Mater. Res. (1981), 15: 167-277 and Langer, Chem. Tech., (1982) 12: 98-105, или поли(виниловый спирт)), полилактиды (пат. США №3,773,919, EP 58,481), сополимеры L-глутаминовой кислоты и гамма этил-L-глутамат (Sidman et al., Biopolymers, (1983) 22: 547-556), неразлагаемый этилен-винил ацетат (Langer et al., supra), разлагаемые сополимеры молочной и гликолевой кислот, такие, как LUPRON Depot™ (микросферы для инъекций, состоящие из сополимера молочной и гликолевой кислот и лейпролид ацетата), и поли-D-(-)-3-гидроксимасляной кислоты (EP 133,988).
В то время как такие полимеры, как этилен-винил ацетат, молочная кислота и гликолевая кислота, способны высвобождать молекулы на протяжении более 100 дней, некоторые гидрогели высвобождают протеины за более короткое время. Когда инкапсулированные протеины остаются в теле на более длительный срок, они могут денатурировать или агрегировать в результате воздействия влаги при 37°C, что приводит к потере биологической активности и возможных изменений иммуногенности. Для стабилизации протеина могут быть разработаны рациональные стратегии в зависимости от используемого механизма. Например, если обнаружено, что механизм агрегации представляет собой образование внутримолекулярных S-S связей посредством дисульфидного обмена, стабилизация может быть достигнута путем модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контроля содержания влаги, использования соответствующих добавок и разработки конкретных полимерных матриксных композиций.
Композиции замедленного высвобождения также содержат препараты кристаллов антитела, суспендированные в приемлемом составе, способном сохранять кристаллы в суспензии. Эти препараты при введении подкожно или интраперитонеально могут вызывать также эффект замедленного высвобождения. Другие композиции также содержат захваченные антитела. Липосомы, содержащие такие антитела, получают способами, известными из: патент США №DE 3,218,121; Epstein et al., Proc. Natl Acad. Sci. USA, (1985) 82: 3688-3692; Hwang et al., Proc. Natl Acad. Sci. USA, (1980) 77: 4030-4034; EP 52,322; EP 36,676; EP 88,046; EP 143,949; 142,641; японская патентная заявка 83-118008; патенты США №№4,485,045 и 4,544,545; и EP 102,324.
Дозировка этих композиций антител для данного пациента будет определена путем посещения врача, принимая во внимание различные факторы, которые, как известно, модифицируют воздействие лекарств, включая тяжесть и тип заболевания, массу тела, пол, диету, время и маршрут введения, другие лекарственные средства и релевантные клинические факторы. Терапевтически эффективные дозировки могут быть определены способами in vitro или in vivo.
Эффективное количество антител, описанных в данной заявке, для терапевтического применения будет зависеть, например, от терапевтических целей, маршрута введения и состояния пациента. Соответственно, для врача преимущественным будет титрование дозировок и модификация маршрута введения, необходимых для получения оптимального терапевтического эффекта. Типичная дневная доза может быть в диапазоне от, приблизительно, 0,001 мг/кг до 100 мг/кг или больше, в зависимости от перечисленных факторов. Типично, врач будет вводить терапевтическое антитело, пока не будет достигнута дозировка, при которой достигается желаемый эффект. Успех лечения легко оценить при помощи традиционных способов анализа или как описано в данной заявке.
Будет оценено, что введение терапевтических объектов в соответствии с композициями и способами по данной заявке будет осуществлено с приемлемыми носителями, наполнителями и другими агентами, включенными в формулировку для обеспечения улучшенного переноса, доставки, переносимости и т.п. Эти композиции включают, например, порошки, пасты, мази, желе, воски, масла, липиды (катионные или анионные), содержащие основы (такие как Lipofectin™), конъюгаты ДНК, безводные адсорбционные пасты, эмульсии типа «масло в воде» и «вода в масле», эмульсии карбовакс (полиэтиленгликоли с различной молекулярной массой), полутвердые гели, и полутвердые смеси, содержащие карбовакс. Любые из вышеуказанных смесей могут быть приемлемыми для лечения и терапии в соответствии с данным изобретением, при условии что активный ингредиент в композиции не активирован композицией, и композиция физиологически совместима и переносима при данном маршруте введения. См. также Baldrick P. "Pharmaceutical excipient development: the need for preclinical guidance." Regul. Toxicol. Pharmacol. 32(2): 210-8 (2000), Wang W. "Lyophilization and development of solid protein pharmaceuticals." Int. J. Pharm. 203 (1-2): 1-60 (2000), Charman WN "Lipids, lipophilic drugs, and oral drug delivery-some emerging concepts." J Pharm Sci.89(8): 967-78 (2000), Powell et al. "Compendium of excipients for parenteral formulations" PDA J Pharm Sci Technol. 52: 238-311 (1998), и ссылки, приведенные там для дополнительной информации относительно композиций, наполнителей и носителей, известных химикам-фармацевтам.
Разработка и создание других лекарственных средств
В соответствии с данным изобретением и исходя из активности антител, полученных и охарактеризованных в данной заявке по отношению к IGF-I/II, разработка других терапевтических модальностей облегчена и описана специалистам в данной области. Такие модальности включают, без ограничения, продвинутые лекарственные средства на основе антител, таких, как биспецифические антитела, иммунотоксины, радиоактивно меченные средства и одиночные домены антител V, антитело-подобные связывающие агенты, основанные на других областях, чем область V, создание пептидных лекарств, генную терапию, в особенности интрателами, антисмысловую терапию и небольшие молекулы.
В связи с созданием продвинутой терапии антителами, где фиксация комплемента является желательной, возможна независимость от комплемента для убийства клеток посредством использования, например, биспецифических антител, иммунотоксинов или радиоактивных меток.
Например, могут быть получены биспецифические антитела, которые содержат (i) два антитела, одно из которых специфично по отношению к IGF-I/II, и другое антитело, специфичное к другой молекуле, которые сконъюгированы вместе, (ii) одно антитело, имеющее одну цепь, специфичную к IGF-I/II и которую цепь, специфичную ко второй молекуле, или (iii) одноцепочечное антитело, специфичность как по отношению к IGF-I/II, так и по отношению к другой молекуле. Такие биспецифические антитела могут быть получены с использованием методик, которые хорошо известны из уровня техники; например, в связи с (i) и (ii) см., например, Fanger et al. Immunol Methods 4: 72-81 (1994) и Wright and Harris, supra. И в связи с (iii) см., например, Traunecker et al. Int. J. Cancer (Suppl.) 7: 51-52 (1992). В каждом случае, вторая специфичность может быть получена при необходимости. Например, вторая специфичность может быть получена для рецепторов активации тяжелой цепи, включая, без ограничения, CD16 или CD64 (см., например, Deo et al., 18: 127 (1997)) или CD89 (см., например, Valerius et al. Blood 90: 4485-4492 (1997)).
Антитела также могут быть модифицированы таким образом, чтобы действовать как иммунотоксины, с использованием методик, известных из уровня техники. См., например, Vitetta Immunol Today 14: 252 (1993). См. также патент США №5,194,594. В связи с получением радиоактивно меченых антител, такие модифицированные антитела могут быть также легко получены с использованием методик, которые хорошо известны из уровня техники. См., например, Junghans et al. в Cancer Chemotherapy and Biotherapy 655-686 (2d edition, Chafner and Longo, eds., Lippincott Raven (1996)). См. также патенты США №№4,681,581, 4,735,210, 5,101,827, 5,102,990 (RE 35,500), 5,648,471 и 5,697,902. Каждый из иммунотоксинов и радиоактивно меченых молекул, возможно, будут убивать клетки, экспрессирующие желаемые многомерные ферментные субъединицы доменов олигомеризации. В некоторых воплощениях, обеспечивается фармацевтическая композиция, содержащая эффективное количество антитела в комбинации с фармацевтически приемлемым носителем или разбавителем.
В некоторых воплощениях, анти-IGF-I/II антитело связано с агентом (например, радиоактивным изотопом, фармацевтической композицией или токсином). Преимущественно, такие антитела могут быть использованы для лечения заболевания, такие заболевания могут относится к клеткам, экспрессирующим IGF-I/II или избыточно экспрессирующим IGF-I/II. Например, описано, что лекарства имеют фармацевтические свойства и их выбирают из группы, которая состоит из противоопухолевых, алкилирующих, антиметаболитных, антиангиогенных, апоптических, алкалоидных, СОХ-2, и антибиотических агентов и их комбинаций. Лекарство выбирают из группы, состоящей из азотистого иприта, производных этиленимина, алкилсульфонатов, нитрозомочевин, триазенов, аналогов фолиевой кислоты, антрациклинов, таксанов, ингибиторов СОХ-2, аналогов пиримидина, аналогов пурина, антиметаболитов, антибиотиков, ферментов, эпиподофилотоксинов, координационных комплексов платины, алкалоидов барвинка, замещенных мочевин, производных метилгидразина, подавителей деятельности коры надпочечников, антагонистов, эндостатина, таксолов, камптотецинов, оксаплатина, доксорубицинов и их аналогов, и их комбинаций.
Примеры токсинов также включают гелонин, эксотоксин Pseudomonas (РЕ), РЕ40, РЕ38, токсин дифтерии, рицин, абрин, альфа-токсин, сапорин, рибонуклеазу (RNase), DNase I, токсин стафилококка-A, противовирусный протеин фитолакки американской, и их производные, комбинации и модификации.
Примеры радиомеченных изотопов включают гамма-генераторы, позитрон-генераторы и рентгеновские генераторы, которые могут быть использованы для локализации и/или терапии, и бета-генераторы и альфа-генераторы, которые могут быть использованы для терапии. Радиомеченные изотопы, описанные выше в качестве полезных для диагностики, прогностики и этапирования, также полезны для лечения. Неограниченные примеры противораковых или противолейкемических агентов включают антрациклины, такие как доксорубицин (адриамицин), даунорубицин (дауномицин), идарубицин, деторубицин, карминомицин, эпирубицин, эзорубицин, и морфолино и замещенные производные, их комбинации и модификации. Иллюстративные фармацевтические агенты включают цис-платину, таксол, калихеамицин, винкристин, цитарабин (Ara-C), циклофосфамид, преднизон, даунорубицин, идарубицин, флударабин, хлорамбуцил, интерферон альфа, гидроксимочевину, темозоламид, талидомид, блеомицин и их производные, комбинации и модификации. Преимущественно, противораковыми или противолейкемическими агентами являются доксорубицин, морфолино доксорубицин или морфолинодаунорубицин.
Как будет оценено специалистом в данной области, вышеуказанные воплощения, в то время как значения аффинности могут быть важными, другие факторы могут иметь такое же или большее значение, в зависимости от конкретной функции антитела. Например, для иммунотоксина (токсина, связанного с антителом) действие связывания антитела с мишенью может быть полезным; однако, в некоторых воплощениях, желаемой целью является интернационализация токсина в клетку. Как таковые, антитела с высоким процентом интернализации могут быть желательными в этих ситуациях. Поэтому, в одном воплощении, описаны антитела с высокой эффективностью в интернализации. Высокая эффективность интернализации может быть измерена в процентах интернализованных антител и может составлять от низкого значения до значения 100%. Например, в различных воплощениях, 0,1-5, 5-10, 10-20, 20-30, 30-40, 40-45, 45-50, 50-60, 60-70, 70-80, 80-90, 90-99 и 99-100% могут составлять высокую эффективность. Как будет оценено специалистом в данной области, желаемая эффективность может отличаться в различных воплощениях, в зависимости от, например, связанного агента, количества антитела, которое может быть введено в область, побочных эффектов комплекса антитела-агент, вида (например, вида рака) и тяжести заболевания, которое будут лечить.
В других воплощениях, антитела, описанные в данной заявке, обеспечивают набор для анализа для обнаружения экспрессии IGF-I/II в тканях или клетках млекопитающих для скрининга заболевания или расстройства, связанных с изменением экспрессии IGF-I/II. Набор содержит антитело, которое связывает IGF-I/II и предназначено для индикации реакции антитела с антигеном, если он присутствует.
В некоторых воплощениях, обеспечиваются готовые изделия, содержащие контейнер, который содержит композицию анти-IGF-I/II антитела и упаковку или метку с указанием того, что композиция может быть использована для лечения заболевания, опосредованных экспрессией IGF-I/II. Преимущественно, млекопитающие, а более предпочтительно, человек, получают анти-IGF-I/II антитело.
Комбинации
Анти-IGF-I/II антитела, указанные ранее в данной заявке, могут быть использованы для лечения сами по себе или могут содержать, дополнительно, соединение в соответствии с данным изобретением, традиционную хирургию или радиотерапию или химиотерапию. Такая химиотерапия может включать одну или более таких категорий антиопухолевых агентов:
(i) антипролиферативные/антинеопластические лекарства и их комбинации, которые используют в медицинской онкологии, такие, как алкилирующие агенты (например, цисплатин, карбоплатин, циклофосфамид, азотистый иприт, мелфалан, хлорамбуцил, бусульфан и нитромочевины); антиметаболиты (например, антифолаты, как фторпиримидины, такие как 5-фторурацил и тегафур, ралтитрексед, метотрексат, цитозин арабиноза и гидроксимочевина; противоопухолевые антибиотики (например, антрациклин-подобные адриамицин, блеомицин, доксорубицин, дауномицин, эпирубицин, идарубицин, митомицин-C, дактиномицин и митрамицин); антимитотические агенты (например, такие алкалоиды барвинка, как винкристин, винбластин, виндезин и винорельбин, и такие таксоиды, как таксол и таксотер); и ингибиторы топоизомеразы (например, эпиподофилотоксины, такие как этопозид и тенипозид, амсакрин, топотекан и камптотецин);
(ii) такие цитостатические агенты, как антиэстрогены (например, тамоксифен, торемифен, ралоксифен, дролоксифен и йодоксифен), регуляторы рецепторов эстрогена (например, фулвестрант), антиандрогены (например, бикалутамид, флутамид, нилутамид и ципротерон ацетат), антагонисты LHRH или агонисты LHRH (например, госерелин, лейпрорелин и бусерилин), прогестрогены (например, мегестрол ацетат), ингибиторы ароматазы (например, анастрозол, летрозол, воразол и экземестан) и ингибиторы 5α-гредуктазы, такие как финастерид;
(iii) агенты, которые ингибируют инвазию раковых клеток (например, такие ингибиторы металлопротеазы, как маримастат ингибиторы активаторной функции рецептора урокиназного плазминогена);
(iv) ингибиторы функции фактора роста, например, такие ингибиторы включают антитела фактора роста, рецептора антитела фактора роста (например, анти-erbb2 антитело трастузумаб [Herceptin™] и анти-erbb1 антитело цетуксимаб), фарнезил, ингибиторы трансферазы, ингибиторы МЕK, ингибиторы тирозинкиназы и ингибиторы серин/треонинкиназы, например, ингибиторы семейства эпидермального фактора роста (например, такие ингибиторы EGFR семейства тирозинкиназ, как N-(3-хлор-4-фторфенил)-7-метокси-6-(3-морфолинопропокси)хиназолин-4-амин (гефитиниб, AZD1839), N-(3-этинилфенил)-6,7-бис(2-метоксиэтокси)хиназолин-4-амин (эрлотиниб, OSI-774) и 6-акриламидо-N-(3-хлор-4-фторфенил)-7-(3-морфолинопропокси)хиназолин-4-амин (CI 1033)), например ингибиторы тромбоцитного семейства факторов роста и, например, ингибиторы гепатоцитного семейства фактора роста;
(v) такие антиангиогенные агенты, которые могут ингибировать влияние фактора роста сосудистого эндотелия (например, анти-антитело фактора роста эндотелиальных клеток сосудов бевацизумаб [Avastin™], такие соединения, которые описаны в международных патентных заявках WO 97/22596, WO 97/30035, WO 97/32856 и WO 98/13354) и соединения, которые действуют по другим механизмам (например, линомид, ингибиторы интегрина αvβ3 функция, ангиостатин и ингибиторы действия ангиопоэтинов, например ангиопоэтин 1 и ангиопоэтин 2);
(vi) агенты сосудистого повреждения, такие как комбретастатин A4 и соединения, описанные в международных патентных заявках WO 99/02166, WO 00/40529, WO 00/41669, WO 01/92224, WO 02/04434 и WO 02/08213;
(vii) антисмысловая терапия, например, направленная на указанные выше мишени, такая как ISIS 2503, анти-ras антисмысловая;
(viii) подходы генной терапии, включая, например, подходы для замещения таких аберрантных генов, как аберрантный р53, или аберрантный BRCA1 или BRCA2, такие подходы GDEPT (ген-направленная ферментная пролекарственная терапия), как подходы с использованием цитозин диаминазы, тимидин киназы или бактериального нитроредуктазного фермента и подходы для увеличения переносимости пациентами химиотерапии или радиотерапии, такие как генная терапия мультилекарственной резистентности;
(ix) иммунотерапевтические подходы, включая, например, ex-vivo и in-vivo подходы для усиления иммуногенности опухолевых клеток пациента, такие как трансфекция цитокинами, такими как интерлейкин 2, интерлейкин 4 или гранулоцит-макрофаговый фактор стимуляции толстой кишки, подходы для уменьшения астении Т-клеток, подходы с использованием трансфектованных иммунных клеток, таких как цитокин-трансфектованных дендритных клеток, подходы с использованием цитокин-трансфектованных опухолевых клеточных линий и подходы с использованием анти-идиотипических антител;
(x) ингибиторы клеточного циклы, включая, например, ингибиторы CDK (например, флавопиридол) и другие ингибиторы контрольных точек клеточного цикла (например, контрольной киназы); ингибиторы киназы Аврора и других киназ, задействованных в регуляции митоза и цитокинеза (например, митотические кинезины); и ингибиторы гистон деацетилазы;
(xi) антагонисты эндотелина, включая антагонисты эндотелина A, антагонисты эндотелина B и антагонисты эндотелина A и B; например, ZD4054 и ZD1611 (WO 96 40681), атразентан и YM598; и
(xii) биотерапевтические лечебные подходы, например, с использованием пептидов или протеинов (таких, как антитела или растворимые конструкции внешних рецепторных доменов) с секвенированием рецепторных лигандов, блокированием лигандного связывания с рецептором или уменьшением передачи сигналов рецептором (например, из-за усиленной деградации рецепторов или пониженных уровней экспрессии).
Такое совместное лечение может быть достигнуто одновременной, последовательной или раздельной дозировки отдельных компонентов лечения. Такие продукты комбинации содержат соединения в соответствии с данным изобретением в диапазоне дозировок, описанном выше, и другие фармацевтически активные агенты в утвержденном диапазоне дозировок.
ПРИМЕРЫ
Нижеприведенные примеры, включая проведенные эксперименты и полученные результаты, приведены только для иллюстрации и не должны истолковываться как ограничивающие теории и учения, описанные в данной заявке.
ПРИМЕР 1
Иммунизация и титрование
Иммунизация
В качестве антигенов использовали рекомбинантные человеческие IGF-I и IGF-II, полученные от R&D Systems, Inc. (Minneapolis, MN Кат. №291-G1 и 292-G2, соответственно). Моноклональные антитела к IGF-I/II разрабатывали путем последовательной иммунизации мышей XenoMouse® (штаммы XenoMouse XMG2 и XMG4 (3С-1 штамм), Abgenix, Inc. Fremont, СА). Животных XenoMouse иммунизировали посредством введения всех инъекций в подушечки лап. Общий объем каждой инъекции составлял 50 мкл на мышь, 25 мкл на каждую подушечку лап. В каждой группе было иммунизировано десять (10) мышей. Каждая инъекция составляла 10 мкг IGF-I или IGF-II на мышь самих по себе или в комбинации с антигеном Keyhole Limpet Hemocyanin (KLH) в качестве носителя, как подробно описано в Таблице 2. Первую инъекцию проводили в фосфатно-солевом буфере (PBS), модифицированном по Дульбекко (DPBS) и смешивали 1:1 об. с адъювантом Titermax Gold Adjuvant (SIGMA Кат. #T2684, партия #К1599). Затем вводили всего 8-11 дополнительных буст-инъекций в течение 27-38 дней, в смеси с 25 мкг Adju-Phos (алюминий-фосфатный гель, каталожный номер 1452-250, партия #8937, HCI Biosector) и 10 мкг CpG (15 мкл адъюванта ImmunEasy Mouse Adjuvant, каталожный номер 303101; партия #11553042; Qiagen) на мышь, с последующей финальной буст-инъекцией 10 мкг антигена в не содержащем пироген DPBS, без адъюванта. Для комбинированной иммунизации (животные иммунизированы как IGF-I, так и IGF-II), второй антиген вводили в последних двух (2) буст-инъекций.
ПРИМЕР 2
ВОССТАНОВЛЕНИЕ ЛИМФОЦИТОВ, ВЫДЕЛЕНИЕ В-КЛЕТОК, СЛИЯНИЯ И ОБРАЗОВАНИЕ ГИБРИДОМ
Иммунизированных мышей иммортализовали путем смещения шейных позвонков и дренированные лимфатические узлы из каждой группы собирали и накапливали. Лимфоидные клетки диссоциировали путем измельчения в среде Игла, модифицированной Дульбекко (DMEM) для извлечения клеток из тканей, а клетки суспендировали в DMEM. Клетки пересчитывали и 0,9 мл DMEM на 100 миллионов лимфоцитов добавляли клеточные планшеты для повторной суспензии, осторожно, но полностью. Использование 100 мкл CD90+ магнитные частицы на 100 миллионов клеток, клетки были помечены путем инкубирования клеток с магнитными частицами при 4°C в течение 15 минут. Суспензию магнитно-меченых клеток, которая содержала до 108 позитивных клеток (или всего до 2×109 клеток), погружали в LS+ колонку и промывали ее DMEM. Всю вытекшую жидкость собирали в качестве СD90-негативной фракции (ожидалось, что большинство этих клеток являются B клетками).
Слияние выполняли путем смешения промытых обогащенных В клеток, полученных так, как описано выше, и несекреторных миеломных клеток P3X63Ag8.653, приобретенных у АТСС, кат. # CRL 1580 (Kearney et al., J. Immunol. 123, 1979, 1548-1550) в соотношении 1:1. Клеточную смесь осторожно гранулировали центрифугированием при 800×g. После полного удаления супернатанта, клетки обрабатывали 2-4 мл раствора проназы (CalBiochem, кат. # 53702; 0,5 мг/мл в PBS) не более 2 минут. Затем 3-5 мл раствора эмбриональной бычьей сыворотки (FBS) добавляли для прекращения ферментной активности, а суспензию доводили до общего объема в 40 мл при помощи раствора для электрослияния клеток (ECFS, 0,3 М сахарозы, Sigma, кат. # S7903, 0,1 мМ ацетата магния, Sigma, кат. # М2545, 0,1 мМ ацетата кальция, Sigma, кат. # С4705). Супернатант удаляли после центрифугирования, а клетки повторно суспендировали в 40 мл ECFS. Эту стадию промывания повторяли и клетки снова повторно суспендировали в ECFS до концентрации 2×106 клеток/мл.
Электрослияние клеток выполняли с использованием генератора слияния (модель ЕСМ2001, Genetronic, Inc., San Diego, СА). Размер использовавшейся камеры слияния составлял 2,0 мл, с использованием таких настроек приборов:
условия выравнивания: напряжение: 50 В, время: 50 сек;
разрушение мембраны при: напряжение: 3000 В, время: 30 мсек;
время удерживания после слияния: 3 сек.
После ECF клеточные суспензии осторожно удаляли из камеры слияния в стерильных условиях и переносили в стерильную пробирку, содержащую такой же объем гибридомной культуральной среды (DMEM, JRH Biosciences), 15% FBS (Hyclone), с добавлением L-глутамина, пен/стреп, OPI (оксалоацетат, пируват, коровий инсулин) (все получены от Sigma) и IL-6 (Boehringer Mannheim). Клетки инкубировали в течение 15-30 минут при 37°C, а затем центрифугировали при 400×g (1000 rpm) в течение пяти минут. Клетки осторожно повторно суспендировали в небольшом объеме гибридизационной селекционной среды (гибридомная культуральная среда с 0,5х НА (Sigma, кат. # А9666)) и объем регулировали путем добавления соответствующих количеств гибридизационной селекционной среды, исходя из конечной планшеты на 5×106 B клеток всего на планшету на 96 лунок и 200 мкл на планшету. Клетки осторожно перемешивали и при помощи пипетки добавляли в планшеты на 96 лунок и оставляли расти. На 7 или 10 день половину среды удаляли, а в клетки повторно добавляли гибридизационную селекционную среду.
ПРИМЕР 3
ОТБОР ВОЗМОЖНЫХ АНТИТЕЛ ПРИ ПОМОЩИ ELISA
Через 14 дней культивирования гибридомные супернатанты подвергали скринингу на IGF-I/II-специфичные моноклональные антитела. Планшеты ELISA (Fisher, кат.№12-565-136) покрывали 50 мкл/лунка человеческого IGF-I или IGF-II (2 мкг/мл) в покрывающем буфере (0,1 М карбонатный буфер, pH 9,6, NaHCO3 8,4 г/л), затем инкубировали при 4°C всю ночь. После инкубации планшеты промывали промывочным буфером (0,05% Tween 20 в PBS) 3 раза. Добавляли 200 мкл/лунку блокирующего буфера (0,5% BSA, 0,1% Tween 20, 0,01% тимеросала в 1x PBS) и инкубировали планшеты при комнатной температуре в течение 1 часа. После инкубации планшеты промывали промывочным буфером три раза. Добавляли 50 мкл/лунка гибридомных супернатантов и позитивный и негативный контроль и инкубировали планшеты при комнатной температуре в течение 2 часов.
После инкубации планшеты три раза промывали промывочным буфером. 100 мкл/лунка козлиного анти-huIgGFc-HRP (Caltag, кат. №H10507) для детекции антител добавляли, а планшеты инкубировали при комнатной температуре в течение 1 часа. При втором скрининге позитивные контроли в первом скрининге подвергали скринингу в два этапа, один для детекции человеческого IgG (тяжелая цепь), а остальные для детекции человеческой Ig каппа легкой цепи (козлиный анти-hIg каппа-HRP (Southern Biotechnology, кат. №2060-05) для демонстрации полностью человеческого состава как для IgG, так и для Ig каппа. После инкубации планшеты три раза промывали промывочным буфером. 100 мкл/лунка ТМВ (BioFX Lab. кат. №TMSK-0100-01) добавляли и оставляли планшеты на, приблизительно, 10 минут (пока лунки негативного контроля не начали отчетливо проявлять цвет). Затем добавляли 50 мкл/лунка стоп-раствора (ТМВ стоп-раствор (BioFX Lab. кат.№STPR-0100-01)) и считывали планшеты на планшетном ридере ELISA при 450 нм. Как показано в таблице 3, всего 1,233 лунок содержали антитела к IGF-I и -II.
Все антитела, которые связывали в анализе ELISA, подвергали количественному анализу связывания с инсулином при помощи ELISA для исключения тех, которые перекрестно реагируют с инсулином. Планшеты ELISA (Fisher, Кат. №12-565-136) покрывали 50 мкл/лунка рекомбинантного инсулина (концентрация: 1 мкг/мл; Sigma, каталог # 12643) в покрывающем буфере (0,1 М карбонатного буфера, pH 9,6, NaHCO3 8,4 г/л), затем инкубировали при 4°C всю ночь. Как подробно описано в Таблице 3, всего 1,122 антител из исходных 1233 перекрестно реагировали с инсулином.
Наконец, антитела, отобранные путем количественного скрининга, проверяли при помощи ELISA для подтверждения связывания протеинов IGF-I и IGF-II. Всего идентифицировали 683 гибридомных линий, имеющих перекрестную реактивность к мышиному IGF-I/II. Соответственно, эти гибридомные линии экспрессировали антитела, которые связывали человеческий IGF-I, человеческий IGF-II, мышиный IGF-I и мышиный IGF-II, но не связывали человеческий инсулин.
ПРИМЕР 4
ИНГИБИРОВАНИЕ IGF-I И IGF-II СВЯЗЫВАНИЯ IGF-IR
Целью данного исследования было проведение скрининга 683 анти-IGF-I/II человеческих IgG2 и IgG4 антител на стадии гибридомного супернатанта на предмет нейтрализующей активности, которую определяли путем ингибирования IGF-I и IGF-II связывания рецептора IGF-IR. Поэтому анализ связывания рецептор/лиганд проводили с клетками NIH3T3, которые избыточно экспрессировали человеческий IGF-IR рецептор, как описано ниже.
Кратко, мультискриниговые фильтровочные пластины (MultiScreen 0,65 мкМ 96-луночная PVDF, Millipore, Кат. №MADV N0B 10) блокировали блокирующим буфером (PBS, содержащий 10% BSA с 0,02% NaN3) при 200 мкл/лунка всю ночь, при 4°C. -меченый IGF (Amersham Life Sciences кат. №IM172 (IGF-I) или IM238 (IGF-II)) при 100 м Ci мл и 50 нМ разбавляли до соответствующей концентрации (70 пМ конечная IGF-I и 200 пМ конечная для IGF-II) в связующем буфере (PBS содержащий 2% BSA с 0,02% NaN3). Фильтровочную пластину, покрытую блокирующим буфером, однократно промывали 200 мкл PBS, а 50 мкл анти-IGF-I/II Ab супернатанта (разбавленного в связующем буфере до конечного 25% конечного объема) предварительно инкубировали с 25 мкл -IGF в планшете MultiScreen в течение 30-60 минут во льду. Субконфлюэнт NIH3T3 мышиных фибробластов, стабильно экспрессирующих hIGF-IR (полученных от AstraZeneca) собирали трипсином и повторно суспендировали в холодном связывающем буфере при 6×106/мл и 25 мкл клеток добавляли в планшету для двухчасовой инкубации во льду. Планшету промывали четыре раза 200 мкл холодного PBS и сушили всю ночь. Добавляли двадцать пять мкл/лунка сцинтиллянта (коктейль SuperMix, Wallac/Perkin Elmer кат. №1200-439) и считывали планшеты при помощи ридера Microbeta Trilux (Wallac).
Для каждой скрининговой планшеты использовали такой контроль: отсутствие антител (весь IGF связан), контрольные нейтрализующие анти-IGF-I (#05-172, Upstate) или анти-IGF-II (#МАВ292, R&D Systems) mAbs в 50 мкл/мл (неспецифический фон) и 0,075-0.5 мкл/мл (приблизительные значения ЕС50 нейтрализующих антител), и изотоп-совпадающий контрольный человеческий IgG2 (РК 16.3.1, Abgenix, партия #360-154) или IgG4 (108.2.1, Abgenix, партия #718-53А) mAbs при концентрации 0,5 мкл/мл (приблизительные значения ЕС50 нейтрализующих антител). Дополнительное титрование контрольных нейтрализующих антител и изотоп-совпадающих контрольных человеческих антител добавляли в одну планшету на один скрининговый анализ (1/10 серийное разбавление от 50 мкг/мл (333,3 нМ)). Все контроли с антителами или без них готовили в связывающем буфере с добавлением анти-KLH человеческого IgG2 или IgG4 израсходованного супернатанта при 25% конечного объема.
Процент ингибирования определяли таким образом:
% Ингибирование=([(Среднее СРМ общего количества связанных 125I-IGF)-(Среднее СРМ связанных 125I-IGF в присутствии антитела)]/[(Среднее СРМ общего количества связанных 125I-IGF)-(Среднее СРМ связанных 125I-IGF в присутствии избытка контрольного нейтрализующего антитела∗)])×100
∗неспецифический фон определяли как СРМ клеток при избытке контрольных нейтрализующих анти-IGF Ab (50 мкг/мл, 333,3 нМ), который, как было найдено, был эквивалентен избытку холодного IGF (меньшим или равным 10% от общего СРМ)
Скрининг анти-IGF-I/II супернатанта был расщеплен на изотопы из-за присутствия радиоактивно меченных лигандных тканей. Как показано в Таблице 4, супернатанты от анти-IGF-I/II антител с IgG2 изотипом (всего 293) сначала подвергали скринингу на радиомеченные IGF-I. Предел 40% ингибирования применяли сначала для данного скрининга (т.е. были отобраны гибридомные линии, ингибирующие при 40% и выше), и было отобрано 111 попаданий для последовательного скрининга к IGF-II. Из 111 попаданий было обнаружено всего 91 линий, которые ингибируют связывание IGF-II с его рецептором при 50% пределе. Всего было отобрано 71 конечных попаданий путем отбора супернатантов, которые нейтрализуют 50% активности IGF-I и IGF-II.
Был проведен первоначальный скрининг всех супернатантов, экспрессирующих IgG4 изотипы (всего 390) в отношении радиоактивно меченных IGF-II, а 232 попадания с пределом ингибирования 50% были подвергнуты последовательному скринингу по отношению к IGF-I. Всего 90 линий были способны ингибировать связывание IGF-I с его рецептором при пределе 50%. После комбинирования попаданий для IgG2 (71) и IgG4 (90), была получена всего 161 линия, которая ингибировала IGF-I и IGF-II на 50% или больше.
В заключение, из 683 исходных супернатантов 343 (111 IgG2 и 232 IgG4, 50,2%) были отобраны из первого скрининга с IGF-I или IGF-II. Всего было получено 161 конечных попаданий (23,6% от исходных линий), способных блокировать связывание IGF-I и IGF-II с IGF-IR с общим критерием предела, составляющим 50% ингибирования.
Предел 40%
ПРИМЕР 5
ВЫСОКОЧУВСТВИТЕЛЬНЫЙ АНТИГЕННЫЙ И ОГРАНИЧЕННЫЙ АНТИГЕННЫЙ АНАЛИЗ ELISA
Для определения относительных афинностей среди 161 гибридомных линий, отобранных в Примере 4, а также концентрации антитела в супернатантах каждой линии, были проведены высокочувствительный антигенный (НА) и ограниченный антигенный (LA) анализы ELISA. В количественном НА анализе, высокие концентрации антигена и период инкубации на протяжении всей ночи ограничивали влияние афинности антител, позволяя количественное определение относительного количества антиген-специфического антитела, присутствующего в каждой пробе. Низкие концентрации антигена в LA анализе ограничивали влияние концентрации антител и приводили к классификации антител, исходя из их относительной аффинности.
Высокочувствительный антигенный количественный анализ
Планшеты ELISA покрывали относительно большим количеством антигена IGF-I или IGF-II (R&D Systems, Inc., Minneapolis, MN Кат. №291-G1 и 292-G2, соответственно) при 500 нг/мл (67 nM). Содержащие антитела гибридомные супернатанты титровали в диапазоне разбавления 1:50 - 1:12200. Контроль известными IGF-специфичными антителами (R&D Systems, Inc., Minneapolis, MN Кат. №MAB291 и МАВ292, соответственно) использовали для определения линейного диапазона анализа. Данные в линейном диапазоне использовали для получения относительной концентрации IGF-специфического антитела в каждой оттитрованной пробе. Ограниченный антигенный анализ
Микротитрованные планшеты покрывали при низких концентрациях антигенов. Пятьдесят микролитов (50 мкл) IGF-I или IGF-II при 64, 32, 16, 8, 4, и 2 нг/мл (покрытые в диапазоне от 8,5 нМ до 0,26 нМ) в 1% снятого молока/1X PBS pH 7,4/0.5% азида добавляли в каждую лунку. Планшеты инкубировали в течение 30 минут.
Планшеты промывали четыре раза (4Х) водой и 50 мкл гибридомного супернатанта, содержащего тестовые антитела, разбавленные 1:25 in 1% снятого молока/IX PBS pH 7.4/0,5% азида добавляли в лунки. Планшеты плотно оборачивали пластиковой оберткой или парафильмом и инкубировали всю ночь при встряхивании при комнатной температуре.
На следующий день все планшеты промывали пять раз (5Х) и в каждую лунку добавляли 50 мл козлиного анти-человеческого IgG Fc HRP поликлонального антитела при концентрации 0,5 мкг/мл в 1% молока, IX PBS pH 7,4. Планшеты инкубировали в течение 1 часа при комнатной температуре.
Планшеты промывали, как минимум, пять раз (5Х водопроводной водой). Пятьдесят микролитров (50) мкл субстрата HPR ТМВ добавляли в каждую лунку и инкубировали планшеты в течение 30 минут. Реакцию HRP-TMB прекращали путем добавления 50 мкл 1М фосфорной кислоты в каждую лунку. Оптическую плотность (поглощение) при 450 измеряли для каждой лунки планшеты.
Анализ данных
Значения оптической плотности тестовых антител усредняли и рассчитывали их диапазон. Антитела с наивысшими сигнальными и приемлемо низкими стандартными отклонениями отбирали в качестве антител с наибольшей афинностью к антигенам, по сравнению с реперным антителом.
Затем проводили анализ для отбора верхних антител, исходя из нейтрализации (Пример 4), эффективности (низкой концентрации антител по результатам НА ELISA и высокой эффективности лигандного связывания), афинности (LA ELISA), или по всем трем критериям. По результатам этого анализа был составлен список из 25 антител. Отдельный анализ на основе среднего % ингибирования IGF-I и -II связывания и афинности для IGF-I и IGF-II привел к получению второго списка, состоящего из 25 антител. Шестнадцать антител совпадали для обоих списков, в результате был получен окончательный список из 40 антител. Результаты LA и НА для этих 40 антител приведены в Таблице 5. Эти 40 линий были отобраны для клонирования, из них 33 были успешно клонированы.
ПРИМЕР 6
СВЯЗЫВАНИЕ АНТИТЕЛ С IGF-I И IGF-II, СВЯЗАННЫМИ С IGFBP-3
IGF-I и -II, которые циркулируют в сыворотке, главным образом связаны с IGF-связывающими протеинами (IGFBPs). Наша цель состояла в идентификации антител, которые не распознают IGFs в комплексе с IGFBPs, во избежание делеции in vivo анти-IGF антител. Следующий формат анализов был разработан для характеристики антител, которые распознают IGF-I или IGF-II если эти факторы роста связаны в комплекс IGFBP-3. Конкретно, в этом анализе проверяли способность IGF в комплексах с IGF/aHTH-IGF антителом связывать IGFBP-3.
Антитело-опосредованное блокирование улавливания IGF IGFBP-3
Был разработан анализ, в котором предварительно образовывались комплексы между IGF-I или IGF-II и IGF-специфическими антителами, описанными в вышеприведенных примерах. Способность этих комплексов связывать IGFBP-3 проверяли при помощи технологии проведения анализов AlphaScreen (PerkinElmer). В планшету на 384 лунок, 10 мкл 1:20 разбавленных гибридомных супернатантов смешивали с 10 мкл 3 нМ биотилинированных IGF-I или IGF-II и инкубировали при комнатной температуре в течение 2 часов. Добавляли покрытые стрептавидином AlphaScreen донорные гранулы и IGFBP-3-соединенные AlphaScreen акцепторные гранулы (10 мкл смеси, для 1/60 конечного разбавления гибридомных супернатантов) и продолжали инкубацию еще час. Затем пробы считывали на планшетном ридере Packard Fusion.
Три коммерчески доступных анти-IGF моноклональных антитела М23 (Cell Sciences), 05-172 (Upstate) и МАВ291 (R&D Systems) показали разную способность к ингибированию связывания IGF с IGFBP со значениями IC50 в диапазоне от низких значений нг/мл до 100 нг/мл. Ингибирование связывания IGF-I с IGFBP-3 наблюдалось у IgG посторонней мыши и человеческого IgG до 10 мкг/мл, вызвав предположение, что анти-IGF-I эффект является специфическим. Коммерчески доступные моноклональные антитела 5-166 (Upstate) и МАВ292 (R&D) проявляли значительную разницу в афинности для ингибирования взаимодействий IGF-II/IGFBP-3. Эти эксперименты показали, что анти-IGF mAbs могут блокировать связывание IGF с IGFBP-3, обеспечив анализ, который может быть использован для скрининга очищенных антител гибридомных линий. Следующая стадия состоит в оценку влияния расходуемой гибридомной среды на аналитический сигнал.
Серийный разведения гибридомной среды и анти-KLH гибридомных расходуемых супернатантов проверяли в аналитической системе. При разведении гибридомных супернатантов 1:10 при получении предварительной инкубации IGFI/II (конечное разбавление в анализе составляло 1:60), практически отсутствовало влияние среды на результаты анализа. Исходя из этих данных, гибридомные супернатанты разбавляли для предварительной инкубации IGF, обеспечивая преимущественное разведение 1/60 - конечное разбавление в анализе.
Шестьсот восемьдесят три израсходованных супернатантов позитивных по отношению к IGF-I и IGF-II связыванию проверяли на предмет их способности ингибировать связывание IGF с IGFBP-3. Ингибирование свыше 50% для IGF-I и свыше 60% для IGF-II использовали в качестве предельного критерия. Итоговые результаты скрининга с использованием пределов показаны в Таблице 6.
В конкурентном анализе IGFBP с использованием анализа AlphaScreen идентифицировано 87 проб, которые ингибируют связывание IGF-I с IGFBP-3 и 129 проб, которые ингибируют связывание IGF-II с IGFBP-3 среди 683 протестированных супернатантов. Пятьдесят одна проба проявила двойную конкуренцию IGF-I и IGF-II. Однако для более осторожного восстановления функции или поведения антител in vivo, где IGF и IGFBP комплексообразование может быть выполнено в большей степени, были проведены дополнительные анализы, как описано в Примере 8.
ПРИМЕР 7
ОПРЕДЕЛЕНИЕ АФИННОСТИ АНТИ-IGF-I И IGF-II АНТИТЕЛ ПРИ ПОМОЩИ АНАЛИЗА BIACORE (СКРИНИНГ НИЗКОГО РАЗРЕШЕНИЯ)
Скрининг низкого разрешения 34 очищенных моноклональных антител
Поверхностный плазмонный резонанс (SPR), или Biacore, использовали для измерения афинности антитела к антигену. Для этого получали поверхность козлиного анти-человеческого антитела высокой плотности над СМ5 чипом Biacore с использованием стандартного аминного сочетания. Все mAbs разбавляли до, приблизительно, 20 мкл/мл в HBS-P подвижном буфере, содержащем 100 мкл/мл BSA. Каждый mAb улавливали на отдельную поверхность, время контакта составляло 30 секунд при 10 мкл/мин, и промывали в течение 5 минут для стабилизации фоновой линии mAb.
IGF-I вводили путем инъекций 335,3 нМ во все поверхности при 23°C в течение 120 секунд, с последующей 5-минутной диссоциацией с использованием скорости потока 100 мкл/мин. Пробы готовили в HBS-P подвижном буфере, описанном выше. Поверхности регенерировали после каждого цикла улавливания/инъекции с одним 15-секундным импульсом 146 мМ фосфорной кислоты (pH 1,5). Аналогичные циклы улавливания/инъекции повторяли для каждого антитела с 114,7 нМ IGF-II. Бездрейфовые данные связывания для 34 mAbs получали путем вычитания сигнала из контрольной проточной ячейки и вычитания дрейфа фоновой линии буфера, впрыскиваемого перед каждой инъекцией антигена. Данные глобально совпадали с 1:1 моделью взаимодействия с использованием CLAMP для определения кинетики связывания (David G. Myszka and Thomas Morton (1998) "CLAMP©: a biosensor kinetic data analysis program", TIBS 23, 149-150). Коэффициент массопереноса использовали при подгонке данных. Кинетический анализ показал связывание IGF-I и IGF-II при 25°C, приведенное в Таблице 7 ниже. mAbs ранжировали в зависимости от афинности: от высокой к низкой.
Данные связывания IGF-I
Большинство mAbs достаточно хорошо соответствовали модели 1:1. MAbs 4.90.2 и 4.141.1 характеризовались чрезвычайно сложными данными. Эти mAbs отмечены звездочкой в Таблице 7, потому что значимые кинетические константы не могут быть установлены из модели подгонки 1:1. Последняя фаза диссоциации является очень медленной из-за этих mAbs (как минимум, 1×10-5 с-1), что может сделать эти два mAbs полезными в качестве терапевтических соединений.
Данные связывания IGF-II
Most Большинство mAbs достаточно хорошо соответствовали модели 1:1. Константа диссоциации для mAb 7.159.2 была постоянной при 1×10-5 с-1, ввиду отсутствия достаточных данных о распаде для адекватной оценки kd.
Анализы Biacore низкого разрешения в данном примере разработаны как полукачественный классификационный подход. Для получения более точной информации о характеристических константах скорости и аффинностях отдельных mAbs, были проведены анализы Biacore высокого разрешения, как описано в Примере 8.
ПРИМЕР 8
ОПРЕДЕЛЕНИЕ АФИННОСТИ АНТИ-IGF-I И IGF-II АНТИТЕЛ ПРИ ПОМОЩИ АНАЛИЗА BIACORE (СКРИНИНГ ВЫСОКОГО РАЗРЕШЕНИЯ)
Анализ Biacore высокого разрешения был проведен для дальнейшего измерения афинности антитела к антигену. Каждую mAbs 7.159.2, 7.234.2, 7.34.1, 7.251.3, и 7.160.2 улавливали, и каждый из антигенов IGF-I и IGF-II были впрыснуты в диапазоне концентраций. Полученные константы связывания приведены в Таблице 8.
Таким образом, воплощения в соответствии с данным изобретением могут включать антитело, которое предпочтительно связывает IGF-II, но перекрестно реагирует с IGF-I, связывая IGF-II с афинностью, большей, чем афинность к IGF-I. Например, антитело может связывать IGF-II с афинностью, которая в 2,5 раза превышает афинность к IGF-I. В некоторых воплощениях, антитело может связывать IGF-II с афинностью, которая в, как минимум, 5, как минимум, 10, как минимум, 25, как минимум, 50 или, как минимум, 150 раз превышает афинность к IGF-I.
Скрининг предварительно образованных комплексов IGF-I/GFBP-3
Конкурентный анализ IGFBP, описанный в Примере 6, идентифицировал 87 проб, которые ингибируют связывание IGF-I с IGFBP-3 и 129 проб, которые ингибируют связывание IGF-II с IGFBP-3 среди 683 протестированных супернатантов. Пятьдесят одна проба продемонстрировала двойную конкуренцию IGF-I и IGF-II. Однако для более тщательного восстановления функции или поведения антител in vivo, где комплексообразование IGF и IGFBP может быть проведено в большей степени, были выполнены анализы Biacore отобранных антител.
Шесть отобранных антител подвергали скринингу для определения того, связывают ли они IGF-I ил IGF-II в комплекс IGFBP. Все шесть отобранных mAbs (7.159.2, 7.146.3, 7.34.1, 7.251.3, 7.58.3 и постороннее контрольное антитело АВХ-МА1) ковалентно иммобилизировали с высокой поверхностной площадью (5,400-12,800 RU) на двух чипах СМ5 Biacore с использованием стандартного аминного связывания при помощи прибора Biacore 2000. на каждом чипе СМ5 активировали и блокировали одну проточную кювету (отсутствие иммобилизированных mAb) для использования в качестве контрольной поверхности.
Затем IGF-I и IGFBP-3 смешивали вместе с буферным солевым раствором Hepes, pH 7,4, 0,005% Р-20, 100 мкг/мл BSA (HBS-P), для получения конечного раствора при 193 нМ и 454 нМ, соответственно. IGF-II и IGFBP-3 смешивали вместе для получения конечного раствора при 192 нМ и 455 нМ, соответственно. В таких условиях IGF-I и IGF-II на 99,97% образовали комплекс с IGFBP-3. В данных условиях равновесие было достигнуто в течение минут. Растворы связанных в комплекс IGF-I/IGFBP-3 и IGF-II/IGFBP-3 пропускали через различные mAb поверхности при 40 мкл/мин при 23°С, в течение 180 секунд с последующей диссоциацией в течение 120 секунд. Свободные IGF-I и IGF-II затем пропускали через каждую поверхность при 193 нМ и 192 нМ, соответственно, a IGFBP-3 пропускали через каждую поверхность при 454 нМ. Поверхности регенерировали в течение 20 секундного импульса 10 мМ глицина, pH 2,0.
Сенсограммы обрабатывали при помощи программы Scrubber путем вычитания изменения объемного индекса преломления и любого сигнала неспецифического связывания аналита на бесцветной поверхности от поверхностей и/с иммобилизированной mAb. После коррекционных вычитаний бесцветных поверхностей, сенсограммы сравнивали снова путем вычитания средней сенсограммы буферных инъекций в конкретной проточной ячейке. Такое "двойное сравнение" скорректировало сенсограммы связывания mAb в том, что касается каких-либо систематических ошибок, имеющихся в конкретной проточной ячейке.
Образовавшие комплекс и свободные IGF-I/IGFBP-3 и IGF-II/IGFBP-3 связывались довольно слабо со связанными антителами, приблизительная оценка связывания неспецифического связывающего взаимодействия составляла КD>1 мкМ для всех шести mAbs, включая негативный контроль АВХ-МА1 (см. Таблицу 9). Однако с АВХ-МА1 связывание IGF-I/II было слабым и указывало на то, что неспецифическое связывающее взаимодействие имело место во всех трех аналитах. Очевидно, комплексы IGF/IGFBP-3 связывались со всеми этими mAbs, a IGFBP-3 сами по себе - нет. Однако из-за того, что как IGF-I, так и IGF-II и IGBP-3 сами по себе специфически связываются с этими mAbs, когда они связаны друг с другом, это приводит к даже более "клейкому" неспецифически связывающему протеиновому комплексу, что объясняется большим связывающим сигналом для комплекса. Комплексы IGF-I/II/IGFBP-3 и IGFBP-3 в значительной степени связаны с контрольной поверхностью, что также указывает на неспецифичность этих двух протеинов. Однако на сенсограммах, приведенных ниже, это фоновое связывание вычитают из первого репера во время обработки данных, как описано выше.
Эксперимент предполагает, что хотя ранее было показано, 51 проба ингибировала связывание IGF-I/II с IGFBP3 (Пример 6), антитела могут также связывать комплекс IGF/IGFBP in vitro.
ПРИМЕР 9
ОПРЕДЕЛЕНИЕ АФИННОСТИ АНТИ-ИНСУЛИНОВОГО АНТИТЕЛА ПРИ ПОМОЩИ АНАЛИЗА BIACORE (СКРИНИНГ НИЗКОГО РАЗРЕШЕНИЯ)
Перекрестную реактивность антител по отношению к IGF-I/II дополнительно путем измерения афинности mAbs по отношению к человеческому инсулину. IGF-I/II антитела иммобилизовали на чипах СМ5 Biacore, инсулин в раствор впрыскивали для определения константы ассоциации и константы диссоциации. Пять mAbs, включая 7.234.2, 7.34.1, 7.159.2, 7.160.2 и 7.251.3, были протестированы в этом эксперименте. Инсулин разбавляли до 502 нМ в подвижном буфере и вводили во все улавливающие поверхности.
При 502 нМ инсулина связывания инсулина с какими-либо mAbs не наблюдалось. Эти результаты предполагают отсутствие очевидной перекрестной реактивности IGF-I/II mAbs и инсулина.
ПРИМЕР 10
СВЯЗЫВАНИЕ АНТИТЕЛ
Эпитопное связывание проводили в целях определения того, какие из анти-IGF-I/II антител могут перекрестно конкурировать друг с другом, и, таким образом, возможно, связывают одинаковый эпитоп GF-I/II. Процесс связывания описан в патентной заявке США №20030175760, а также в Jia et al., J. Immunol. Methods (2004), 288: 91-98. обе из которых включены в данную заявку полностью путем ссылок. Кратко, гранулы Luminex связывали с мышиными анти-huIgG (Pharmingen #555784) следуя протоколу связывания протеинов, приведенному на веб-сайте Luminex. Предварительно связанные гранулы получали для связывания изначально неизвестного антитела в соответствии со следующей процедурой, защитив гранулы от света. Для каждого отдельного супернатанта использовали отдельные пробирки. Необходимый объем супернатантов рассчитывали при помощи следующей формулы: (nХ2+10)×50 мкл (где n=общее количество проб). В данном анализе использовали концентрацию 0,1 мкл/мл. Гранулы осторожно перемешивали на вихревой мешалке и разбавляли в гранул/мл.
Пробы инкубировали во встряхивателе в темноте при комнатной температуре всю ночь.
Фильтровочную пластину предварительно увлажняли путем добавления 200 мкл промывного буфера на лунку, затем буфер удаляли. 50 мкл каждой гранулы затем добавляли в каждую лунку фильтровочной пластины. Пробы промывали однократно путем добавления 100 мкл/лунка промывочным буфером и его удаления. Антиген и контроли добавляли на фильтровочную пластину при 50 мкл/лунка. Пластину покрывали, инкубировали в темноте в течение 1 часа, а затем промывали пробы, инкубировали в темноте в течение 1 часа во встряхивателе и промывали пробы 3 раза. Второе неизвестное антитело добавляли затем при 50 мкл/лунка. Концентрацию 0,1 мкг/мл использовали для первичного антитела. Затем пластину инкубировали в темноте в течение 2 часов при комнатной температуре во встряхивателе, а затем пробы промывали 3 раза. Добавляли 50 мкл/лунка биотинилированного мышиного античеловеческого IgG (Pharmingen #555785) разбавленного 1:500, и пробы инкубировали в темноте в течение 1 часа с перемешиванием при комнатной температуре.
Пробы промывали 3 раза. 50 мкл/лунка стрептавидин-РЕ при 1:1000 разведении добавляли, и инкубировали пробы в темноте в течение 15 минут при встряхивании при комнатной температуре. После проведения двух циклов промывания на Luminex100, пробы промывали 3 раза. Содержание каждой лунки повторно суспендировали в 80 мкл блокирующего буфера. Пробы осторожно перемешивали с неоднократным пипетированием для повторного суспендирования гранул. Затем пробы анализировали при помощи Luminex100. Результаты представлены ниже в Таблице 10.
ПРИМЕР 11
СТРУКТУРНЫЙ АНАЛИЗ АНТИ-IGF-I/II АНТИТЕЛ
Вариабельные области тяжелых цепей и вариабельные области легких цепей секвентировали для определения их ДНК последовательностей. Полная информация о последовательностях анти-IGF-I/II антител обеспечивается в списке нуклеотидных и аминокислотных последовательностей для каждой комбинации гамма и каппа цепей. Вариабельные области тяжелых цепей были проанализированы для определения в VH семействе, последовательности D-области и последовательности J-области. Затем последовательности транслировали для определения первичной аминокислотной последовательность и сравнения последовательностей VH, D и J-области зародышевых линий для оценки соматических гипермутаций.
Выравнивание последовательностей этих антител с их генами зародышевых линий показано в таблицах, приведенных ниже. Таблица 11 представляет собой таблицу сравнения областей тяжелой цепи антитела и областей тяжелой цепи их родственной зародышевой линии. Таблица 12 представляет собой таблицу сравнения областей каппа легкой цепи антитела с областями легкой цепи их родственной зародышевой линии. Мутации от зародышевой линии показаны как новая аминокислота.
Вариабельные (V) области иммуноглобулиновых цепей кодируются множественными сегментами ДНК зародышевой линии, присоединенные к функциональным вариабельным регионам (VHDJH или VKJK) во время В-клеточного онтогенеза. Молекулярные и генетические расхождения реакций антитела на IGF-I/II были подробно изучены. Эти анализы выявили несколько характеристик, специфических для анти-IGF-I/II антител.
Анализ пяти отдельных антител, специфических к IGF-I/II привели к определению того, что антитела были получены из трех различных генов VH зародышевых линий, четыре из которых принадлежали к семейству VH4, а 2 антитела были получены из сегмента VH4-39 гена. В Таблицах 11 и 12 приведены результаты данного анализа.
Должно быть оценено, что аминокислотные последовательности среди дочерних клонов, собранных от каждой гибридомы, являются идентичными. Например, последовательности тяжелых и легких цепей mAb 7.159.2 идентичны последовательностям, приведенным в Таблицах 11 и 12 для mAb 7.159.1.
CDR1 тяжелых цепей антител в соответствии с данным изобретением имеют последовательность, описанную в Таблице 11. CDR1, которые описаны в Таблице 11, подпадают под определение Кабата. Альтернативно, CDR1 могут быть определены с использованием альтернативного определения, таки образом, чтобы включить пять последних остатков последовательности FR1. Например, для антитела 7.159.1 FR1 последовательностью является QVQLVQSGAEVKKPGASVKVSCKAS (SEQ ID №: 93), a CDR1 последовательностью является GYTFTSYDIN (SEQ ID №: 94); для антитела 7.158.1 FR1 последовательностью является QLQLQESGPGLVKPSETLSLTCTVS (SEQ ID №: 95), a CDR1 последовательностью является GGSIRSSSYYWG (SEQ ID №: 96); для антитела 7.234.1 FR1 последовательностью является QLQLQESGPGLVKPSETLSLTCTVS (SEQ ID №: 97) и CDR1 последовательностью является GGSINSSSNYWG (SEQ ID №: 98); для антитела 7.34.1 FR1 последовательностью является QVQLQESGPGLVKPSETLSLTCTVS (SEQ ID №: 99) и CDR1 последовательностью является GGSISSYYWS (SEQ ID №: 100); а для антитела 7.251.3 FR1 последовательностью является QVQLQESGPGLVKPSETLSLTCTVS (SEQ ID №: 101), a CDR1 последовательностью является GGSISSYYWS (SEQ ID №: 102).
Также должно быть оценено, что в случаях, если конкретное антитело отличается от последовательности его соответствующей зародышевой линии на аминокислотном уровне, то последовательность антитела может мутировать назад в последовательность зародышевой линии. Такие корреляционные мутации могут происходить в одной, двух, трех или более положениях, или комбинации мутированных положений, с использованием стандартных биологических методик. В качестве неограничивающего примера, в Таблице 12 показана последовательность легкой цепи mAb 7.34.1 (SEQ ID №: 12), которая отличается от соответствующей последовательности зародышевой линии (SEQ ID №: 80) посредством мутации Pro в Ala (мутация 1) в области FR1 и посредством мутации Phe в Leu (мутация 2) в области FR2. Таким образом, аминокислотная или нуклеиновокислотная последовательность, кодирующая легкую цепь mAb 7.34.1, может быть модифицирована с мутацией изменения 1 с получением последовательности зародышевой линии в месте мутации 1. Кроме того, аминокислотная или нуклеиновокислотная последовательность, кодирующая легкую цепь mAb 7.34.1, может быть модифицирована с мутацией изменения 2 с получением последовательности зародышевой линии в месте мутации 2. Кроме того, аминокислотная или нуклеиновокислотная последовательность, кодирующая легкую цепь mAb 7.34.1, может быть модифицирована с обеими с мутациями изменения 1 и 2 с получением последовательности зародышевой линии в обоих местах мутации 1 и 2.
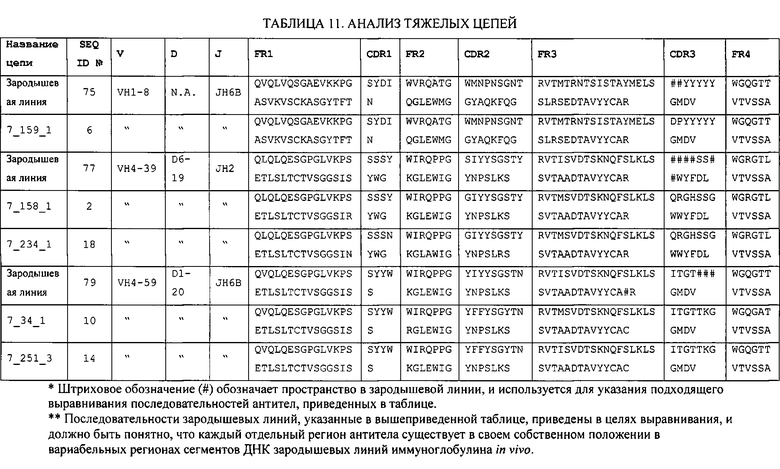
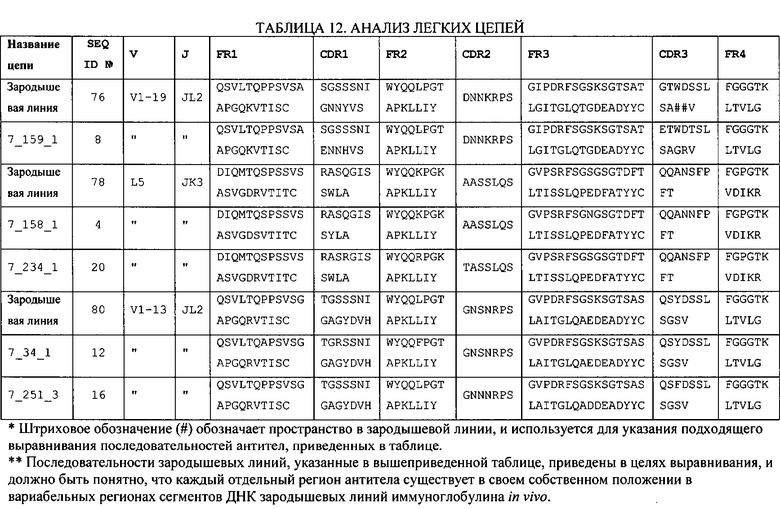
ПРИМЕР 12
ИНГИБИРОВАНИЕ IGF-I И IGF-II-ВЫЗВАННОГО ФОСФОРИЛИРОВАНИЯ hIGF-IR, КОТОРЫЙ ЭПИТОПНО ЭКСПРЕССИРУЕТСЯ В КЛЕТКАХ NIH3T3
IGF лиганды вызывают их пролиферацию и функцию антиапоптоза путем активации действия рецептора тирозин киназы в рецепторе IGF-IR. Для оценки способности анти IGF-I/II антител ингибировать IGF-вызываемое фосфорилирование IGF-IR, клетки NIH3T3, которые эктопически экспрессируют hIGF-IR, использовали в следующем анализе.
Клетки NIH3T3, которые эктопически экспрессируют человеческий IGF-IR, высевали в планшету на 96 лунок при плотности 10000 клеток на лунку и инкубировали всю ночь в «голодно» среде (1% FBS в смеси с углем). На следующий день среду для выращивания удаляли, лунки осторожно промывали два раза PBS и 100 мкл среды, не содержащей сыворотки (0% FBS), добавляли для умерщвления клеток. Через 1-2 часа 100 мкл среды, не содержащей сыворотки, с 0,05% BSA, содержащим IGF-I (10 нг/мл) или IGF-II (10 нМ), предварительно инкубировали в течение 60 минут при 37°C при различных концентрациях антител и добавляли затем к клеткам в тройном объеме. Стимулирование проводили в течение 10 минут при 37°C, после чего среду удаляли и 100 мкл 3,7% формальдегиды в PBS/3%BSA добавляли в каждую лунку и инкубировали при комнатной температуре в течение 20 минут. Затем клетки промывали 2Х PBS и в каждую лунку добавляли 100 мкл проницаемого буфера (0,1% Triton-X в 3% BSA/PBS). Смесь инкубировали при комнатной температуре в течение 10 минут, удаляли и 100 мкл 0,6% раствора перекиси водорода в PBS/3% BSA добавляли для инактивации любой эндогенной пероксидазной активности. Через 20 минут инкубирования при комнатной температуре клетки промывали 3Х PBS/0,1% Tween-20 и блокировали путем добавления 100 мкл 10% FBS в PBS/0,1% Tween-20 при комнатной температуре в течение 1 часа. Затем блокирующий буфер удаляли и 50 мкл анти-фосфо-IGFIR антитела при 1 мкг/мл (кат. #44-804, BioSource) добавляли в каждую лунку в 10% FBS/PBS-T. Через 2 часа инкубирования при комнатной температуре, клетки промывали 3Х с замачиванием PBST в течение 5 минут перед каждым промыванием. После промываний 50 мкл/лунка козлиного анти-кроличьего IgGFc-HRP вторичного антитела разбавляли 1:250 в блокирующем буфере и добавляли в каждую лунку. Через 1 час инкубации при комнатной температуре клетки промывали 3Х в течение 5 минут PBST, как описано ранее, и высушивали. Затем добавляли 50 мкл реагента ECL (DuoLux) и немедленно считывали RLU (относительные световые единицы).
Скрининг был проведен для 32 (тридцати двух) линий антител, и для каждого антигена проводили два независимых анализа. Результаты для верхних десяти антител приведены в Таблице 13 ниже.
ПРИМЕР 13
ИНГИБИРОВАНИЕ IGF-I И IGF-II-ВЫЗВАННОЙ ПРОЛИФЕРАЦИИ КЛЕТОК NIH3T3, ТРАНСФЕКТОВАННЫХ С HIGF-IR
Как обсуждалось выше, одним из критериев нейтрализации IGF-I/II антител является способность к ингибированию пролиферации, вызванной IGF. Для оценки способности антител ингибировать вызванную IGF пролиферацию, человеческие опухолевые клетки поджелудочной железы ВхРС3, которые экспрессируют эндогенный hIGF-IR, использовали в следующем анализе.
Клетки ВхРС3, которые эктопически экспрессируют hIGF-IR, высевали в планшете на 96 лунок при плотности 5000 клеток на лунку и культивировали всю ночь в стандартной среде для выращивания (1% FBS в смеси с углем). На следующий день среду для выращивания убирали, лунки осторожно дважды промывали средой, не содержащей сыворотки, и добавляли 100 мкл среды, не содержащей сыворотки, для умерщвления клеток. 100 мкл «голодной» среды, содержащей 15 нг/мл IGFI или 50 нг/мл IGFII, предварительно инкубировали в течение 30 минут при 37°C при различных концентрациях антител и добавляли к клеткам в двойном или тройном объеме. После 20 часов инкубации клетки подвергали воздействию импульсов BrdU в течение 2 часов и количественно определяли степень включения (пролиферации) при помощи набора для клеточной пролиферации ELISA от Roche (Roche, кат. #1647229).
Скрининг был проведен для 32 линий антител, и для каждого антигена проводили два или три независимых анализа. Результаты для верхних 10 антител приведены в Таблице 14 ниже.
ПРИМЕР 14
ИНГИБИРОВАНИЕ IGF-I И IGF-II-ИНДУЦИРОВАННОГО ФОСФОРИЛИРОВАНИЯ hIGF-IR, КОТОРЫЙ ЭКСПРЕССИРУЕТСЯ В КЛЕТКАХ ЧЕЛОВЕЧЕСКОЙ ОПУХОЛИ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ ВхРС3
IGF-I/II вызывают их пролиферацию и анти-апоптозную функцию путем активации действия рецептора тирозин киназы IGF-IR. Для оценки способности антител ингибировать IGF-вызываемое фосфорилирование IGF-IR, ВхРС3 человеческие опухолевые клетки поджелудочной железы, которые экспрессируют эндогенный hIGF-IR, использовали в следующем анализе.
Клетки ВхРС3 высевали в планшете на 96 лунок при плотности 55 ООО клеток на лунку и инкубировали всю ночь в обычной среде для выращивания. На следующий день, среду для выращивания удаляли, лунки осторожно промывала два раза средой, не содержащей сыворотки, и 100 мкл среды, не содержащей сыворотки, добавляли для умерщвления клеток. Через 24 часа среду удаляли и клетки осторожно промывали однократно средой, не содержащей сыворотки. Среду, не содержащую сыворотки, с 0,05% BSA, содержащим IGF-I (20 нг/мл) или IGF-II (75 нг/мл), предварительно инкубировали в течение 30 минут при 37°C при различных концентрациях антител и 100 мкл добавляли затем к клеткам в тройном объеме. Планшеты инкубировали в течение 15 минут при 37°C и последовательно промывали холодным PBS. 100 мкл лизисного буфера добавляли к лункам и инкубировали планшету в течение 30 минут при 4°C. Лизаты центробежно вращали при 2000 rpm в течение 10 минут при 4°C, а супернатанты собирали. IGF-IR фосфорилирование количественно определяли при помощи набора Duoset человеческого фосфо-IGF-IR ELISA (R&D Systems, Кат. №DYC1770).
Скрининг был проведен для десяти линий антител, и для каждого антигена проводили независимый анализ. Результаты приведены в Таблице 15 ниже.
ПРИМЕР 15
ИНГИБИРОВАНИЕ IGF-I И IGF-II-ВЫЗВАННОЙ ПРОЛИФЕРАЦИИ ЧЕЛОВЕЧЕСКОЙ ОПУХОЛИ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ ВхРС3
Как обсуждалось выше, одним из критериев нейтрализации IGF антител является способность к ингибированию пролиферации, вызванной IGF. Для оценки способности антител ингибировать вызванную IGF пролиферацию человеческие опухолевые клетки поджелудочной железы ВхРС3, которые экспрессируют эндогенный hIGF-IR, использовали в следующем анализе.
Клетки ВхРС3 высевали в планшете на 96 лунок при плотности 2000 клеток на лунку и культивировали всю ночь в стандартной среде для выращивания. На следующий день среду для выращивания убирали, лунки осторожно дважды промывали средой, не содержащей сыворотки, и добавляли 100 мкл среды, не содержащей сыворотки, с 10 мкл/мл трансферрина и 0,1% BSA (аналитическая среда) для умерщвления клеток. Через 24 часа среду удаляли, клетки осторожно однократно промывали сывороткой и добавляли 100 мкл аналитической среды, содержащей 20 нг/мл IGF предварительно инкубированного в течение 30 минут при 37°C при различных концентрациях антител, добавляли к клеткам в двойном или тройном объеме. Планшеты инкубировали в течение 3 дней и пролиферацию определяли количественно с использованием реагента CellTiter-Glo (Promega).
Скрининг был проведен для десяти линий антител, и для каждого антигена проводили два или три независимых анализа. Результаты приведены в Таблице 16 ниже. На основании функциональных данных, приведенных ниже, и данных Примера 14 были отобраны четыре наилучших антитела. Данные анализа пролиферации, вызванной IGF-I, были исключены из критериев отбора из-за высокой наблюдаемой вариабельности анализа.
ПРИМЕР 16
ОПРЕДЕЛЕНИЕ ПЕРЕКРЕСТНОЙ РЕАКТИВНОСТИ К МЫШИНОМУ IGF-I, IGF-II И ИНСУЛИНУ
Одна из задач состояла в разработке антител, специфичных по отношению к IGF-I и IGF-II, но не имеющих перекрестной реактивности по отношению к инсулину. Для выполнения следующих экспериментов на животных, антитела должны были также иметь перекрестную реактивность по отношению к мышиному IGF-I/II, но не к мышиному инсулину. Соответственно, были проведены анализы ELISA для определения способности отобранных антител перекрестно реагировать с мышиным и IGFs или инсулином.
Как показано в Таблице 17, пять из верхних десяти антител тестировали на перекрестную реактивность с мышиным или крысиным инсулином при помощи ELISA. Анализы ELISA показали, что эти антитела не имели перекрестной реактивности с мышиным или крысиным инсулином, по сравнению с негативным контрольным антителом РК16.3.1, и в отличие от позитивного контрольного антикрысиного инсулинового антитела.
ПРИМЕР 17
ИНГИБИРОВАНИЕ ВЫЗВАННОГО IGF-I И IGF-II МЫШЕЙ ФОСФОРИЛИРОВАНИЯ ЧЕЛОВЕЧЕСКОГО IGF-IR, КОТОРЫЙ ЭКТОПИЧЕСКИ ЭКСПРЕССИРУЕТСЯ В КЛЕТКАХ NIH3T3
Моноклональные антитела, имеющие перекрестную реактивность с мышиными IGF-I и IGF-II, дополнительно тестировали для того, чтобы определить степень ингибирования ими вызванного IGF фосфорилирования IGF-IR. Этот анализ выполняли так, как ранее было описано для анализа с использованием клеток NIH3T3, которые эктопически экспрессируют рецептор hIGF-IR. Результаты исследований приведены в Таблице 18.
Клетки NIH3T3, которые эктопически экспрессируют человеческие IGF-IR, высевали в планшете на 96 лунок при плотности 10000 клеток на лунку и инкубировали всю ночь в «голодной» среде (1% FBS, обесцвеченной углем). На следующий день среду для выращивания удаляли, лунки осторожно два раза промывали PBS и 100 мкл среды, не содержащей сыворотки (0% FBS), добавляли для того, чтобы умертвить клетки. Через 1-2 часа 100 мкл среды, не содержащей сыворотки с 0,05% BSA, содержащей мышиные IGF-I (10 нМ) или IGF-II (10 нМ) (R&D Systems, Inc., Minneapolis, MN Кат.№791-MG и 792-MG соответственно), которые были предварительно инкубированы в течение 60 минут при 37°C при различных концентрациях антител, добавляли к клеткам в тройном объеме. Стимулирование проводили в течение 10 минут при 37°C, после стимулирования среды удаляли и 100 мкл 3,7% формальдегида в PBS/3% BSA добавляли в каждую лунку и инкубировали при комнатной температуре в течение 20 минут. Затем клетки промывали 2Х PBS и 100 мкл проницаемого буфера (0,1% Triton-X в 3% BSA/PBS) добавляли в каждую лунку. Это все инкубировали при комнатной температуре в течение 10 минут, удаляли и 100 мкл 0,6% перекиси водорода в PBS/3% BSA добавляли для инактивации эндогенной пероксидазной активности. Через 20 минут инкубации при комнатной температуре, клетки промывали 3Х PBS/0,1% Tween-20 и блокировали путем добавления 100 мкл 10% FBS в PBS/0,1% Tween-20 при комнатной температуре в течение 1 часа. Затем блокирующие раствор удаляли и 50 мкл анти-фосфо-IGFIR антитела при 1 мкг/мл (кат. #44-804, BioSource) добавляли в каждую лунку в 10% FBS/PBS-T. Через 2 часа инкубации при комнатной температуре клетки промывали 3Х PBST, замачивая на 5 минут между каждым промыванием. После промываний 50 мкл/лунка козлиным антикроличьим IgGFc-HRP второе антитело, разбавленное 1:250 в блокирующем буфере, добавляли в каждую лунку. Через 1 час инкубирования при комнатной температуре клетки промывали 3Х в течение 5 минут PBST, как описано выше, и высушивали. Затем добавляли 50 мкл каждого ECL реагента (DuoLux) и немедленно считывали RLU.
ПРИМЕР 18
ИНГИБИРОВАНИЕ РОСТА КЛЕТОК NIH3T3, ЭКСПРЕССИРУЮШИХ IGF-II И IGF-IR IN VIVO У «ГОЛЫХ» МЫШЕЙ
Для оценки способности антител ингибировать вызванную IGF-II пролиферацию in vivo, были проведены следующие эксперименты.
«Голым» мышам женского рода в возрасте 6-8 недель (поставляются Charles River Laboratories, Wilmington, MA, USA) имплантировали подкожно 5x106 клеток клона 32 (клетки NIH3T3, эктопически избыточно экспрессирующие человеческие IGF-I и человеческие IGF-1R). Клетки суспендировали в PBS в общем прививочном объеме 330 мкл. Опухоли оставляли расти до 100-200 мм3 до лечения моноклональными антителами 7.159.2, 7.34.1 и 7.251.3. Антитела или IgG2 изотипные контрольные антитела, суспендированные в PBS, вводили интраперитонеально в рандомизированные группы, состоящие из 9 или 12 мышей, два раза в неделю на протяжении 4 недель при 5 или 50 мг/кг, начиная со Дня 22. PBS вводили как контроль основы еще одной группе, состоящей из 11 мышей еженедельно в течение 4 недель, начиная со Дня 22. Размер опухоли и массу тела измеряли 2-3 раза в неделю.
Наблюдалось значительное ингибирование роста опухолей, при этом ни в одной из групп не наблюдалось значительного снижения массы тела. Антитела 7.159.2, 7.34.1 и 7.251.3 в значительной степени ингибировали рост опухолей с клоном 32 при 5 и 50 мг/кг/неделя.
ПРИМЕР 19
ИНГИБИРОВАНИЕ РОСТА КЛЕТОК NIH3T3, ЭКСПРЕССИРУЮЩИХ IGF-I И IGF-IR IN VIVO У «ГОЛЫХ» МЫШЕЙ
В предыдущем примере, было показано, что антитела ингибируют вызванную IGF-II пролиферацию in vivo. Для того чтобы оценить антитела на предмет их способности ингибировать вызванную IGF-I пролиферацию in vivo, были проведены следующие эксперименты.
«Голым» мышам женского рода (штамм Alderley Park, полученный от штамма швейцарской «голой» мыши, который поставляется AstraZeneca) имплантировали 5×106 жизнеспособных Р12 клеток [клетки NIH3T3, эктопически избыточно экспрессирующие человеческие IGF-I и человеческие IGF-IR (Pietrzkowski et al., Cell Growth & Differentiation, 3, 199-205, 1992)] подкожно в левый бок. Клетки суспендировали в PBS в общем прививочном объеме 0,1 мл. Две группы животных (в каждой n=10) дозировали два раза в неделю, начиная с дня имплантирования клеток NIH3T3 mAb 7.159.2 при 1,0 мг на мышь, или эквивалентным объемом основы PBS (0,3 мл) по тому же графику. Все дозы давали интраперитонеально (i.p.). Массы тела животных измеряли каждый день и после установления массы, измерения опухолей проводили два раза в неделю с использованием циркулей. Объем всех измеряемых опухолей был рассчитан на основе измерений циркулем, предполагая яйцевидную форму.
Значительное ингибирование роста опухолей наблюдалось при использовании mAb 7.159.2 после интраперитонеального введения дважды в неделю 1,0 мг антитела/мышь. Ни в одной группе животных не наблюдалось значительной потери массы.
ПРИМЕР 20
ИНГИБИРОВАНИЕ РОСТА ОПУХОЛЕВЫХ КЛЕТОК У ПАЦИЕНТОВ-ЛЮДЕЙ
Группы людей с раковыми заболеваниями, у которых продиагностировали рак поджелудочной железы, произвольно разделили на лечебные группы. Каждую группу пациентов лечили еженедельными внутривенными инъекциями mAb 7.159.2, 7.34.1 или 7.251.3, описанными в данной заявке. Каждому из пациентов давали дозу эффективного количества антитела в диапазоне от 50 мг/кг до 2,250 мг/кг в течение 4-8 недель. Контрольная группа получала только стандартное химиотерапевтическое лечение
Периодически во время и после лечения массу опухоли оценивали при помощи магнитно-резонансной интроскопии (MRI). Было найдено, что пациенты, которые получали еженедельное лечение антителами mAb 7.159.2, 7.34.1 или 7.251.3, показывали значительное сокращение размеров опухолей, по сравнению с пациентами, не получившими такого лечения. У некоторых пациентов, получающих лечение, опухоли более не обнаруживались. Наоборот, в контрольной группе размер опухолей возрастал или не изменялся.
ПРИМЕР 21
ИНГИБИРОВАНИЕ РОСТА ОПУХОЛЕВЫХ КЛЕТОК У ПАЦИЕНТОВ-ЛЮДЕЙ
У пациента-мужчины была продиагностирована злокачественная опухоль. Пациента лечили путем введения раз в неделю внутривенных инъекций mAb 7.159.2 в течение 8 недель. Периодически во время и после лечения массу опухоли оценивали при помощи магнитно-резонансной интроскопии (MRI). Было найдено значительное уменьшение размера опухоли.
ПРИМЕР 22
ЛЕЧЕНИЕ АКРОМЕГАЛИИ У ПАЦИЕНТОВ-ЛЮДЕЙ
У взрослого мужчины была продиагностирована акромегалия. Пациента лечили путем введения два раза в неделю внутривенных инъекций mAb 7.34.1 в течение 2 лет. В результате у пациента наблюдалось значительное уменьшение симптомов акромегалии.
ПРИМЕР 23
ЛЕЧЕНИЕ ПСОРИАЗА У ПАЦИЕНТОВ-ЛЮДЕЙ
У взрослой женщины был продиагностирован тяжелый псориаз. Пациентку лечили путем введения два раза в неделю внутривенных инъекций mAb 7.251.3 в течение 3 недель. В результате у пациентки наблюдалось значительное уменьшение симптомов псориаза.
ПРИМЕР 24
ЛЕЧЕНИЕ ОСТЕОПОРОЗА У ПАЦИЕНТОВ-ЛЮДЕЙ
У взрослой женщины был продиагностирован остеопороз. Пациентку лечили путем введения два раза в неделю внутривенных инъекций mAb 7.159.2 в течение года. В результате это привело к значительному снижению потери костной массы.
ПРИМЕР 25
ЛЕЧЕНИЕ АТЕРОСКЛЕРОЗА У ПАЦИЕНТОВ-ЛЮДЕЙ
У взрослого мужчины был продиагностирован атеросклероз. Пациента лечили путем введения два раза в неделю внутривенных инъекций mAb 7.34.1 в течение года. В результате у пациента наблюдалось уменьшение симптомов атеросклероза, таких как инфаркт.
ПРИМЕР 26
ЛЕЧЕНИЕ РЕСТЕНОЗА У ПАЦИЕНТОВ-ЛЮДЕЙ
Взрослая женщина получала ангиопластику для облегчения состояния заблокированной артерии. После процедуры ангиопластики пациентку лечили путем введения два раза в неделю внутривенных инъекций mAb 7.251.3 в течение года. В результате у пациентки не произошло рестеноза артерии, которую лечили.
ПРИМЕР 27
ЛЕЧЕНИЕ ДИАБЕТА У ПАЦИЕНТОВ-ЛЮДЕЙ
У взрослой женщины был продиагностирован диабет. Пациентку лечили путем введения два раза в неделю внутривенных инъекций mAb 7.159.2 в течение года. В результате симптомы диабета были уменьшены.
ПОСЛЕДОВАТЕЛЬНОСТИ
Субклонированные гибридомы были секвенированы для определения их первичной структуры, как на уровне нуклеотидов, так и на уровне аминокислот, как для генов вариабельной области тяжелой и легкой цепей. Нуклеотидные и полипептидные последовательности вариабельных регионов моноклональных антител к IGF-I и IGF-II, как перечислено в Таблице 1, приведены в Перечне последовательностей.
ВКЛЮЧЕНИЕ ПУТЕМ ССЫЛОК
Все ссылки, приведенные в данной заявке, включая патенты, патентные заявки, статьи, учебники и т.п., и все процитированные в них ссылки, в той степени, в которой они приведены, целиком включены в данную заявку путем ссылок.
ЭКВИВАЛЕНТЫ
Вышеупомянутые письменные характеристики считаются достаточными для того, чтобы специалист в данной области мог реализовать изобретение. В вышеприведенных Описании и Примерах достаточно подробно описаны предпочтительные воплощения данного изобретения и описаны наилучшие режимы, предусмотренные изобретателями. Однако будет оценено, что вне зависимости от того, насколько подробно описано все вышеизложенное, данное изобретение может быть реализовано многими способами и может быть истолковано в соответствии с формулой, которая прилагается, и любыми ее эквивалентами.
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗЫВАЮЩИЕ ПРОТЕИНЫ, СПЕЦИФИЧНЫЕ ПО ОТНОШЕНИЮ К ИНСУЛИН-ПОДОБНЫМ ФАКТОРАМ РОСТА, И ИХ ИСПОЛЬЗОВАНИЕ | 2006 |
|
RU2492185C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2011 |
|
RU2573588C2 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-2 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2745451C1 |
| АНТИТЕЛА К HER3 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2560583C2 |
| АНТИ-GARP-TGF-β-АНТИТЕЛА | 2018 |
|
RU2767784C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К BTN2, И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2821703C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ПРОТИВ СПЕЦИФИЧЕСКОГО МЕМБРАННОГО АНТИГЕНА ПРОСТАТЫ (PSMA) И СПОСОБ ИНГИБИРОВАНИЯ РОСТА КЛЕТОК, ЭКСПРЕССИРУЮЩИХ PSMA | 2006 |
|
RU2421466C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2607038C2 |
| Антитело против PD-1 и его применение | 2017 |
|
RU2739610C1 |
| АНТИТЕЛА ПРОТИВ DLL4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2415869C2 |





























Настоящее изобретение относится к области иммунологии. Предложено полностью человеческое моноклональное антитело, которое связывает инсулиноподобный фактор роста-II (IGF-II) и имеет перекрестную реактивность к IGF-I, а также его антигенсвязывающий фрагмент. Рассмотрены молекула нуклеиновой кислоты, кодирующая антитело по изобретению, вектор и клетка-хозяин для экспрессии антитела по изобретению. Описана фармацевтическая композиция, а также конъюгаты для лечения и диагностики злокачественной опухоли, применение антитела по изобретению в получении лекарственного средства и способ определения уровня IGF-II и IGF-I в пробе пациента. Настоящее изобретение может найти дальнейшее применение в терапии рака. 8 н. и 8 з.п. ф-лы, 27 пр., 18 табл.
1. Полностью человеческое антитело или его антигенсвязывающий фрагмент, которые связывают инсулиноподобный фактор роста-II (IGF-II), имеют перекрестную реактивность к инсулиноподобному фактору роста I (IGF-I) и нейтрализуют активность IGF-I и IGF-II, где указанное антитело или его связывающий фрагмент включают полипептид тяжелой цепи, имеющий последовательность SEQ ID №: 14, и полипептид легкой цепи, имеющий последовательность SEQ ID №: 16.
2. Антитело или его антигенсвязывающий фрагмент по п.1, где указанное антитело является полностью человеческого моноклональным антителом.
3. Антитело или его антигенсвязывающий фрагмент по п.1, где указанный антигенсвязывающий фрагмент является антигенсвязывающим фрагментом полностью человеческого моноклонального антитела.
4. Антитело или его антигенсвязывающий фрагмент по п.3, где указанный антигенсвязывающий фрагмент выбирается из группы, состоящей из Fab, Fab′ или F(ab′)2 и Fv.
5. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-4, где указанное антитело является моноклональным антителом 7.251.3 (продуцируемым клеточной линией, которая имеет регистрационный номер АТСС № РТА-7422).
6. Композиция для лечения злокачественной опухоли, включающая фармацевтически приемлемый носитель и эффективное количество антитела или антигенсвязывающего фрагмента по любому из пп.1-5.
7. Молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп.1-5.
8. Вектор, предназначенный для экспрессии антитела или его антигенсвязывающего фрагмента по любому из пп.1-5 и содержащий молекулу нуклеиновой кислоты по п.7.
9. Клетка-хозяин для получения антитела или его антигенсвязывающего фрагмента по п.1-5, содержащая вектор по п.8.
10. Человеческое моноклональное антитело по любому из пп.1-5, где указанное антитело не связывает специфически IGF-II или IGF-I протеины, если указанные протеины связаны с инсулиновыми связывающими протеинами фактора роста.
11. Способ определения уровня инсулиноподобного фактора роста-II (IGF-II) и инсулиноподобного фактора роста I (IGF-I) в пробе пациента, который включает:
контактирование указанной пробы пациента с антителом или его антигенсвязывающим фрагментом по любому из пп.1-5; и
определение уровня IGF-I и IGF-II в указанной пробе.
12. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-5 для получения лекарственного средства для лечения злокачественной опухоли.
13. Применение по п.12, где указанную злокачественную опухоль выбирают из группы, состоящей из меланомы, немелкоклеточного рака легких, глиомы, печеночно-клеточного рака, опухоли щитовидной железы, рака желудка, рака простаты, рака молочной железы, рака яичников, рака мочевого пузыря, рака легких, глиобластомы, рака эндометрия, рака почек, рака толстой кишки, рака поджелудочной железы и эпидермоидной карциномы.
14. Применение по п.12 или 13, где указанное лекарственное средство предназначено для использования в комбинации со вторым антинеопластическим агентом, который выбирают из группы, состоящей из антитела, химиотерапевтического агента и радиоактивного лекарственного средства.
15. Конъюгат для лечения злокачественной опухоли, состоящий из антитела или антигенсвязывающего фрагмента по любому из пп.1-5 и терапевтического агента.
16. Конъюгат по п.15, где терапевтический агент представляет собой токсин; радиоактивный изотоп или фармацевтическую композицию.
| EP 1505075 A1, 09.02.2005 | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| ENJOH T | |||
| et al., "Characterization of new monoclonal antibodies to human insulin-like growth factor-II and their application in western immunoblot analysis." J Clin Endocrinol Metab | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| VAN WYK JJ, BRUTON ET., "Molecular basis for species and | |||

