Изобретение относится к обнаружению и количественному определению белка, связанного с репликацией ДНК (Rep), для применения в диагностике нейродегенеративных заболеваний, таких, к примеру, как рассеянный склероз (MS). В частности, изобретение касается мутантного MSBI1 белка Rep, кодируемого геномом.
Этиология рассеянного склероза (MS) еще не установлена. Поэтому существует потребность в биомаркере для MS, который можно было бы использовать для диагностики MS и/или мониторинга MS или лечения MS и/или оценки предрасположенности к MS.
Рассеянный склероз (MS) характеризуется демиелинизированными очагами MS, поражающими нервные клетки головного и спинного мозга. Симптомы MS проявляются в виде эпизодов внезапного ухудшения (рецидивы, обострения, приступы, припадки) или в виде постепенного ухудшения во времени (прогрессирующие формы). Демиелинизация запускает воспалительные процессы, которые активируют Т-клетки и высвобождение цитокинов и антител. Для диагностики MS, среди прочего, применяется нейровизуализация, анализ спинномозговой жидкости и вызванных потенциалов.
Из различных исследуемых материалов (ткани головного мозга с рассеянным склерозом (MS), бычьей сыворотки, молока) было выделено 17 разных, но частично родственных молекул ДНК (Funk, Gunst et al. 2014; Gunst, Zur Hausen et al. 2014; Lamberto, Gunst et al. 2014; Whitley, Gunst et al. 2014).
Среди этих изолятов из ткани головного мозга от пациентов с MS были выделены две молекулы ДНК, близкородственные связанному с трансмиссивной губчатой энцефалопатией (TSE) изоляту Sphinx 1.76 (1,758 п.н.; № доступа HQ444404 (Manuelidis L. 2011)). Эти изоляты - MSBI1.176 (MSBI, изолят из мозга с рассеянным склерозом) (1766 п.о.) и MSBI2.176 (1766 п.о.), которые обозначаются как “геном MSBI1” и “геном MSBI2”, соответственно. Нуклеотидная последовательность MSBI1.176 на 98% аналогична последовательности Sphinx 1.76. Большие открытые рамки считывания (ORF) этих изолятов кодируют предполагаемый белок репликации ДНК, имеющий большое сходство между ними. Другой общий признак - наличие тандемных повторов типа итеронов. Выравнивание этой области повторов указывает на вариации в остове по отдельным нуклеотидам. Эти итероноподобные повторы могут составлять сайты связывания белков Rep. Последовательности изолятов были депонированы в банке данных EMBL под номерами доступа LK931491 (MSBI1.176) и LK931492 (MSBI2.176) (Whitley C. et al. 2014), а их выравнивание описано в WO 2016/005064.
Авторы настоящего изобретения недавно обнаружили, что геном MSBI1 дикого типа демонстрирует значительную продукцию транскрибируемой РНК, а кодируемый геномом MSBI1 белок Rep экспрессируется в клетках человека. Они обнаружили, что кодируемый геномом MSBI1 и MSBI2 белок Rep дикого типа (Rep MSBI1 и Rep MSBI2) представляет собой биомаркер для анализов при скрининге на патогенность. В качестве белка, связанного с репликацией ДНК (RepB), белок Rep обладает ДНК-связывающей активностью и может быть необходим для инициации репликации эписомальных или вирусных молекул ДНК. Однако белки Rep, которые структурно близки кодируемым геномом MSBI1 и 2 Rep дикого типа, проявляют заметную способность к самоолигомеризации и агрегации.
Для диагностических скрининговых анализов очищали антигены белков Rep дикого типа Rep MSBI1 и Rep MSBI2 и хранили в денатурирующих, а также восстановительных условиях, чтобы минимизировать эффекты агрегации или деградации на белок. К сожалению, авторы изобретения наблюдали, что при предварительной очистке белка Rep MSBI1 дикого типа в неденатурирующих условиях действительно проявлялась массивная и видимая агрегация белка, которая вполне очевидно делала белок Rep недоступным для аффинной очистки. Остаточные очень небольшие количества очищенного белка Rep агрегируют в пределах очень короткого промежутка времени (несколько часов), делая белок неадекватным для дальнейших диагностических анализов. Это соответствует предыдущим исследованиям, описывающим агрегацию белков Rep, сравнимую с Rep MSBI1 (Giraldo 2007; Torreira, Moreno-Del Alamo et al. 2015).
Для диагностических скрининговых анализов стабильность и целостность белкового антигена, который используется для обнаружения антигенсвязывающих антител в образцах крови, имеет первостепенное значение для воспроизводимости и надежности таких чувствительных экспериментов. В этом отношении как сохранность белкового антигена при длительном сроке хранения, так и биофизическое поведение во время самого диагностического скрининга (стадии покрытия, блокирования, отмывки, инкубации для выявления антител) играют решающую роль для успешного анализа. Однако, учитывая наблюдаемое явление агрегации белков Rep дикого типа (MSBI1-Rep и MSBI2-Rep), существует потребность в улучшенном белке Rep, не обладающем такими же свойствами агрегации и самоолигомеризации, как белки Rep дикого типа.
Авторы изобретения обнаружили, что вышеуказанных отрицательных свойств можно избежать при помощи мутантного белка. Настоящим изобретением предусмотрены мутантные белки Rep MSBI1 и Rep MSBI2, которые содержат по меньшей мере две точечные мутации по сравнению с соответствующими белками дикого типа и являются биомаркерами для анализов при скрининге на патогенность. Синтезированные белки Rep MSBI1 и MSBI2 являются мутантами, происходящими из кодируемых геномом MSBI1 и MSBI2 белков Rep дикого типа (wt), как показано в SEQ ID. NO: 1 (MSBI1.176) и SEQ ID NO: 8 (MSBI 2.176). Синтезированные мутантные белки Rep MSBI1 и MSBI2 обеспечивают большую стабильность белка Rep и меньшую агрегацию в условиях in vitro, а также в буфере для хранения с мочевиной. Синтезированные мутантные белки Rep MSBI1 и MSBI2 являются более стабильными антигенами, чем белки Rep MSBI1 и MSBI2 дикого типа для применения в анализах при диагностическом скрининге.
Антитела против Rep применяются в качестве патогенных маркеров из-за связи патогенной активности выделенной ДНК агента (напр., MSBI1) с экспрессией белка Rep. Сыворотка пациента, содержащая повышенное количество антител против Rep, указывает на то, что соответствующий пациент определенно подвергался воздействию родственных Rep белков или же сам экспрессировал Rep на протяжении времени, достаточного для запуска специфичного к Rep иммунного ответа. В качестве мишени для человеческих антител в качестве антигена используется белок Rep. На основании количественного определения антител против Rep можно диагностировать или отслеживать острый MS, а также предрасположенность к MS. Поскольку было признано, что повышение содержания индуцированных антител против Rep или экспрессированного белка Rep в образцах указывает на возникновение и/или состояние MS, то повышение содержания антител против Rep и белка Rep, соответственно, может применяться в качестве патогенного биомаркера для диагностики MS.
Преимущественно патогенный биомаркер для MS можно детектировать в образцах крови типа образцов сыворотки или плазмы, и нет необходимости получать образцы из спинномозговой жидкости.
Итак, изобретением предусмотрен белок, связанный с репликацией ДНК (Rep), который включает (i) аминокислотную последовательность, приведенную в SEQ ID NO: 11;
(ii) фрагмент по SEQ ID NO: 11, который способен связываться с антителом против Rep, специфичным для белка, содержащего аминокислотную последовательность, приведенную в SEQ ID NO: 11; или
(iii) аминокислотную последовательность, которая на 90% или больше гомологична аминокислотной последовательности (i) или (ii) и способна связываться с антителом против Rep, специфичным для белка, содержащего аминокислотную последовательность, приведенную в SEQ ID NO: 11.
Кроме того, изобретением предусмотрен белок, связанный с репликацией ДНК (Rep), который включает (i) аминокислотную последовательность, приведенную в SEQ ID NO: 12;
(ii) фрагмент по SEQ ID NO: 12, который способен связываться с антителом против Rep, специфичным для белка, содержащего аминокислотную последовательность, приведенную в SEQ ID NO: 12; или
(iii) аминокислотную последовательность, которая на 90% или больше гомологична аминокислотной последовательности (i) или (ii) и способна связываться с антителом против Rep, специфичным для белка, содержащего аминокислотную последовательность, приведенную в SEQ ID NO: 12.
Белок Rep по настоящему изобретению может применяться практически в любом формате анализа, в котором используется известный антиген для выявления антител или клеточных иммунных ответов. Таким образом, настоящее изобретение также охватывает обнаружение опосредованных клетками, напр., опосредованных Т-клетками иммунных ответов против белка Rep.
В некоторых воплощениях изобретения предусмотрен способ диагностики нейродегенеративного заболевания у субъекта, включающий стадии:
(a) инкубирование образца от субъекта с белком Rep, как определено выше;
(b) определение количества антител в образце от субъекта, образующих иммунологический комплекс с белком Rep; и
(c) коррелирование количества антител, связавшихся с белком Rep, по сравнению с количеством в контрольном образце, с диагнозом нейродегенеративного заболевания.
В определенных воплощениях изобретения предусмотрен способ диагностики MS у субъекта, включающий стадии:
(a) инкубирование образца от субъекта с белком Rep;
(b) определение количества антител в образце от субъекта, образующих иммунологический комплекс с белком Rep; и
(c) коррелирование количества антител, связавшихся с белком Rep, по сравнению с количеством в контрольном образце, с диагнозом MS.
Повышение содержания антител против Rep в образце от субъекта по сравнению с содержанием антител против Rep в контрольном образце коррелирует с диагнозом нейродегенеративного заболевания, напр., MS, т.е. указывает на MS. В определенных воплощениях на диагноз нейродегенеративного заболевания, напр., MS или предрасположенность к нейродегенеративному заболеванию, напр. MS, указывает повышенное содержание антител против Rep по меньшей мере в 2 раза по сравнению с контрольным образцом.
В определенных воплощениях белок Rep иммобилизируют, напр., прикрепляют к подложке или носителю, после чего иммобилизованный белок Rep инкубируют с образцом от субъекта.
В других воплощениях белок Rep экспрессируют в клетках, после чего клетки инкубируют с образцом от субъекта.
В некоторых воплощениях количество антител, образующих иммунологический комплекс с белком Rep, определяют с помощью дополнительного связывающего агента, соединенного с издающим сигналы соединением, который способен связываться с антителами против Rep в иммунологическом комплексе, к примеру, помеченного детектируемой меткой вторичного антитела, предпочтительно антитела против человека.
В других воплощениях антитела в образце от субъекта иммобилизируют, после чего инкубируют с определенным количеством белка Rep.
Предпочтительно образец от субъекта и контрольный образец представляет собой образец крови типа образца сыворотки или плазмы.
Кроме того, авторы изобретения создали антитело против Rep, которое связывается с эпитопом, находящимся в пределах аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-136, 137-229 и 230-324 по SEQ ID NO: 1 или 11. Например, антитело связывается с эпитопом, входящим в SEQ ID NO: 2 или SEQ ID NO: 3.
В других воплощениях изобретения предусмотрен набор для применения при диагностике MS, включающий: (a) белок Rep MSBI1 27/154E или Rep MSBI2 27/154E, (b) дополнительный связывающий агент, соединенный с издающим сигналы соединением, к примеру, антитело против человека, соединенное с детектируемой меткой и способное связываться с антителом против Rep по изобретению, и (c) твердый матрикс, подходящий для иммобилизации белка Rep согласно (а) или антител против Rep, причем данные антитела предполагаются в образце, в частности, в образце сыворотки или плазмы.
В определенных воплощениях набор составлен для применения в иммуноанализе, к примеру, выбранном из группы, состоящей из ферментного иммуносорбентного анализа (ELISA), радиоиммуноанализа (RIA), иммуноферментного анализа (EIA), флуоресцентного иммуноанализа (FIA), люминесцентного иммуноанализа (LIA) и анализа на полосках.
Краткое описание фигур
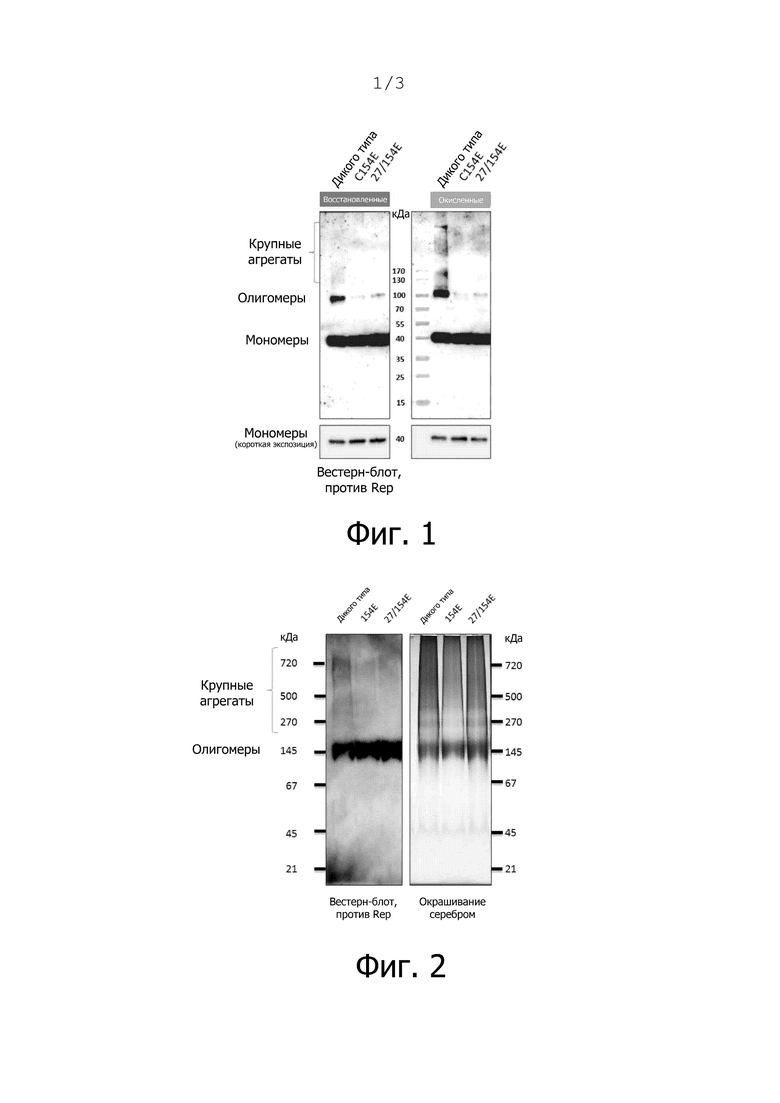
Фиг. 1. Белок Rep сверхэкспрессировали путем временной трансфекции клеток HEK293TT плазмидой pcDNA3.1(-), кодирующей либо Rep MSBI1 дикого типа (WT), либо мутантный Rep MSBI1 27/154E, в течение 72 часов. Клетки трипсинизировали, промывали в PBS и обрабатывали ультразвуком в буфере для лизиса (50 мМ трис, рН 7,6, 150 мМ NaCl, 1,5% Triton X-100, 5 мМ имидазола, 5 мМ β-меркаптоэтанола, смесь ингибиторов протеиназ ×1). После инкубации (30 мин, 4°C) и центрифугирования (30 мин, 10000 g, 4°C) проводили неденатурирующую очистку белка путем связывания белка с 1 мл уравновешенных шариков Ni-NTA (Clontech), промывки 10 колоночными объемами промывочного буфера (буфер для лизиса, содержащий 55 мМ имидазола) и элюирования (буфер для лизиса, содержащий 300 мМ имидазола). После определения белка методом NanoDrop и Bradford равные количества белка (500 нг на дорожку) кипятили в буфере Леммли с 5 мМ β-меркаптоэтанола (восстановленные) или без него (окисленные) и анализировали методом SDS-PAGE и вестерн-блоттинга с помощью антител против Rep.
Фиг. 2. Белок Rep очищали в неденатурирующих условиях, как описано выше. 2000 нг белка Rep подвергали BN-PAGE (Serva) и проводили окрашивание методом иммунодетекции против Rep либо окрашивания белка серебром.
Фиг. 3. Белок Rep выделяли из E. coli в денатурирующих условиях (см. методику ниже). По 2 мкг (вестерн-блоттинг) или 5 мкг (окрашивание Кумасси) очищенного Rep MSBI1 или мутанта Rep MSBI1 27/154E характеризовали методом иммунодетекции против Rep либо окрашиванием белков Кумасси после SDS-PAGE.
Фиг. 4. Белок Rep выделяли из E. coli в денатурирующих условиях (см. методику ниже). По 5 мкг (окрашивание Кумасси) Rep MSBI1 wt или мутанта Rep MSBI1 27/154E использовали для BN-PAGE и иммунодетекции против Rep.
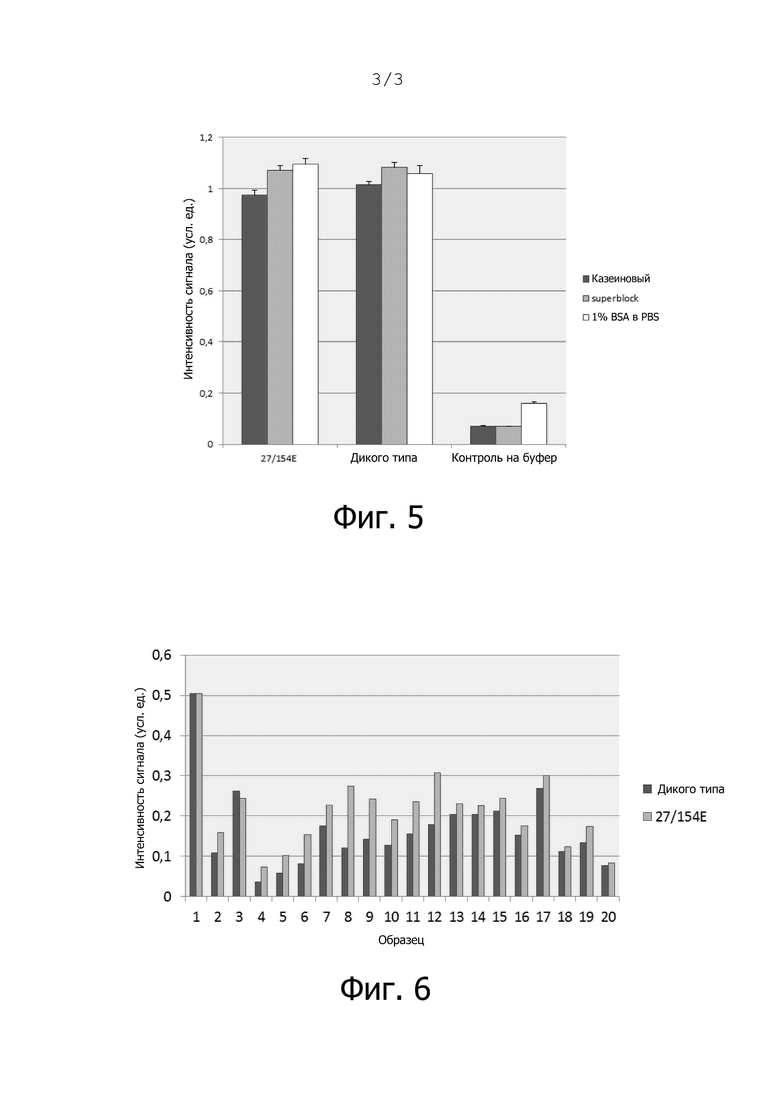
Фиг. 5. Планшеты для ELISA (Maxisorp, Thermo Fisher Scientific) покрывали либо очищенным денатурированным Rep MSBI1 wt, либо мутантным Rep MSBI1 27/154E при 4°C в течение ночи в разведении 1:1 в PBS с 8 М мочевиной (200 нг белка на лунку) в тройных пробах. Проводили блокирование в различных буферах для анализа (казеиновый, Superblock, 1% BSA в PBS) в течение 2 ч при комнатной температуре. Фиксированный белок определяли с помощью комплекта из 3 мышиных моноклональных антител против Rep (разведение 1:500, с эпитопами в N-, центральном и С-концевом домене Rep) в качестве первичных антител, после чего инкубировали с конъюгированным с HRP вторичным козьим антителом против мыши (разведение 1:5000, по 1 ч при 37°C, отмывка PBS с 0,1% Tween, субстрат TMB для ELISA от Thermo Fisher Scientific, считывание при 450 нм). Представлены необработанные интенсивности сигналов.
Фиг. 6. Планшеты для ELISA (Maxisorp, Thermo Fisher Scientific) покрывали либо очищенным денатурированным Rep MSBI1 wt, либо мутантным Rep MSBI1 27/154E при 4°C в течение ночи в разведении 1:1 в PBS с 8 М мочевиной (200 нг белка на лунку). Проводили блокирование в буфере для анализа Superblock в течение 2 ч при комнатной температуре. Проводили инкубацию сыворотки (1:500 в буфере Superblock) в течение 1 часа при 37°C. Определяли связавшиеся с Rep человеческие антитела IgG с помощью конъюгированного с HRP вторичного козьего антитела против человека (разведение 1:5000, 1 час при 37°C) в двойных пробах (отмывка PBS с 0,1% Tween, субстрат TMB для ELISA от Thermo Fisher Scientific, считывание при 450 нм, сигнал нормализован по контролю на BSA).
Изобретением предусмотрены мутантные белки Rep и методы диагностического скрининга на присутствие антител против Rep в качестве патогенных маркеров. Образцы с повышенным содержанием антител против Rep указывают на то, что соответствующий субъект определенно подвергался воздействию родственного Rep белка или же сам экспрессировал белок Rep на протяжении времени, достаточного для индукции специфичного к белку Rep иммунного ответа. С помощью таких методов скрининга можно проводить диагностику, прогнозирование и мониторинг MS на основании определения антител против Rep.
“Белок Rep” в настоящем изобретении относится к связанным с репликацией ДНК белкам (RepB). Белок Rep обладает ДНК-связывающей активностью и может быть необходим для запуска репликации эписомальных/вирусных молекул ДНК. В общем, белок Rep относится к белкам Rep из группы генома Small Sphinx (Whitley et al., 2014). В частности, белок Rep представляет собой синтезированный, кодируемый геномом белок Rep MSBI1 27/154E или кодируемый геномом белок Rep MSBI2 27/154E. Предпочтительно синтезированный белок Rep MSBI1 имеет аминокислотную последовательность, приведенную в SEQ ID NO: 11, и происходит из MSBI1.176, депонированного в банке данных EMBL с № доступа LK931491, который имеет аминокислотную последовательность, приведенную в SEQ ID NO: 1, или же синтезированный белок Rep имеет аминокислотную последовательность, приведенную в SEQ ID NO: 12, и происходит из MSBI2.176, депонированного в банке данных EMBL с № доступа LK931492, который имеет аминокислотную последовательность, приведенную в SEQ ID NO: 8.
В особенно предпочтительном воплощении белок Rep содержит N-концевую область, консервативную среди геномов Small Sphinx, состоящую в основном из аминокислот 1-229 по SEQ ID NO: 11 или 12, и C-концевую вариабельную область, специфичную для MSBI1.176, состоящую в основном из аминокислот 230-324 по SEQ ID NO: 11, или C-концевую вариабельную область, специфичную для MSBI2.176, состоящую в основном из аминокислот 230-324 по SEQ ID NO: 12. N-концевая консервативная область содержит предполагаемый первый ДНК-связывающий домен, состоящий в основном из аминокислот 1-136 по SEQ ID NO: 1, 8, 11 или 12, и второй предполагаемый ДНК-связывающий домен, состоящий в основном из аминокислот 137-229 по SEQ ID NO: 1, 8, 11 или 12.
“Белок Rep” также охватывает фрагменты и варианты белка, которые способны связывать антитело против Rep, специфичное к белку Rep, имеющему аминокислотную последовательность SEQ ID NO: 11 или 12. Предпочтительно такой фрагмент представляет собой иммуногенный фрагмент белка, имеющего аминокислотную последовательность SEQ ID NO: 11 или 12, который охватывает по меньшей мере один эпитоп для антитела против белка Rep против белка Rep по SEQ ID NO: 11 или SEQ ID NO: 12 и предпочтительно содержит как минимум 7, 8, 9, 10, 15, 20, 25 или 50 смежных аминокислот. В определенных воплощениях этот фрагмент включает или состоит в основном из домена белка Rep, к примеру, N-концевой консервативной области, C-концевой вариабельной области, первого или второго ДНК-связывающего домена. Вариант белка по SEQ ID NO: 11 или SEQ ID NO: 12 содержит одну или несколько делеций, замен или вставок аминокислот по сравнению с SEQ ID NO: 11 или SEQ ID NO: 12 и гомологичен по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 11 или SEQ ID NO: 12, причем вариант способен связывать антитело против Rep, специфичное к белку Rep, имеющему аминокислотную последовательность SEQ ID NO: 11 или SEQ ID NO: 12. Определение варианта включает, к примеру, полипептиды, содержащие один или несколько аналогов аминокислот (в том числе, к примеру, неприродные аминокислоты, пептидные нуклеиновые кислоты (PNA) и т.д.), полипептиды с замещенными связями, а также с другими модификациями, известными в данной области, как встречающиеся в природе, так и не встречающиеся в природе. Термин белок Rep включает слитые белки с гетерологичной аминокислотной последовательностью, с лидерной последовательностью или с последовательностью метки и т.п. Например, белок Rep может быть слит с последовательностью метки, к примеру, выбранной из группы, состоящей из метки His6-Tag (SEQ ID NO: 4), T7-Tag (SEQ ID NO: 5), FLAG-Tag (SEQ ID NO: 6) и Strep-II-Tag (SEQ ID NO: 7).
Синтезированный белок Rep MSBI1 (Rep MSBI1 27/154E) или белок Rep MSBI2 (Rep MSBI2 27/154E) по изобретению, включая фрагменты Rep и варианты Rep, как определено выше, можно получить классическими методами химического синтеза. Синтез может проводиться в однородном растворе или в твердой фазе. Полипептиды по настоящему изобретению также могут быть получены методами рекомбинантной ДНК. Например, был разработан Rep MSBI1 27/154E, у которого была минимизирована способность к агрегации двух аминокислотных последовательностей между остатками 25-31 (LLILLAII) и остатками 151-155 (LLICW) посредством двух отдельных точечных мутаций. Основой для клонирования служила последовательность ДНК Rep MSBI1, которая была оптимизирована по кодонам для экспрессии Rep в человеческой системе, кодирующей исходную первичную аминокислотную последовательность Rep MSBI1. Нуклеотиды, кодирующие аминокислоту 27 (L = лейцин, ДНК-кодон CTA), а также нуклеотиды, кодирующие аминокислоту 154 (C = цистеин, ДНК-кодон TGT), были заменены нуклеотидами, кодирующими аминокислоту глутаминовая кислота (E, ДНК-кодон GAG), получая в итоге конечную последовательность ДНК SEQ ID NO: 11. У этого двойного мутанта Rep чистый потенциал агрегации был минимизирован до отметки 141, находящейся в диапазоне белков, не склонных к агрегации.
Как показано в примерах (фиг. 2, 3), мутант Rep 27/154E проявлял значительное снижение (> 50%) олигомеров и более крупных агрегатов Rep по сравнению с белком Rep дикого типа. Вестерн-блоттинг также показал значительно меньше олигомеров Rep, а также меньше высокомолекулярных агрегатов (фиг. 4). Только у мутанта Rep обнаруживался дополнительный вид явно мономерного белка Rep при нативных условиях PAGE. Ясно, что интенсивность высокомолекулярных агрегатов значительно снижается у двойного мутанта 27/154E Rep. В общем, присутствие агрегатов Rep было выше у тех белков, которые подвергались очистке в неденатурирующих условиях. В любом случае, также белки, очищенные и хранящиеся в денатурирующих условиях, проявляют очень сильную агрегацию, которая значительно снижается у двойного мутанта Rep. Это представляет особый интерес, так как экспериментальные условия ELISA основаны на ренатурации поверхностных белковых антигенов, поскольку связывание антигена антителами требует нативных условий.
Белки Rep дикого типа можно получить классическими методами химического синтеза. Синтез может проводиться в однородном растворе или в твердой фазе. Полипептиды по настоящему изобретению также могут быть получены методами рекомбинантной ДНК. Пример получения и очистки белка Rep дикого типа представлен в примере 1.
“Субъект” в настоящем изобретении означает индивид или пациент из млекопитающих, включая грызунов, крупный рогатый скот, к примеру, быков, обезьян и людей. Предпочтительно субъектом является человек-пациент.
“Образец” в настоящем изобретении означает биологический образец, включая жидкие и твердые образцы. Жидкие образцы включают в себя жидкости крови, такие, к примеру, как сыворотка или плазма, и спинномозговую жидкость (CSF). Твердые образцы включают образцы тканей типа культуры тканей или образцы биопсии.
“Коррелирует с” в настоящем изобретении означает, что количество, т.е. уровень или титр антител против Rep или белка Rep, соответственно, имеет значительную корреляцию со статусом заболевания, к примеру, MS. Корреляция определяется путем выявления степени отличия между количеством, присутствующим в образце от исследуемого субъекта, и контрольным образцом. “Контрольный образец” означает один образец или среднее из различных, т.е. более двух контрольных образцов. Контроль берут у здорового индивида, у которого не было диагноза MS. С другой стороны, корреляция может быть определена теоретически путем выявления степени отличия между количеством, присутствующим в образце от исследуемого субъекта, и предварительно установленным пределом отсечения. Предел отсечения - это контрольное значение, дающее статистически значимое разделение между различными состояниями заболевания, напр., между здоровым и больным состоянием. Предел отсечения может быть определен путем статистического анализа достаточно большого комплекта тест-образцов от пациентов с историей болезни и образцов из группы здоровых испытуемых при помощи статистических тестов, известных в данной области.
В некоторых воплощениях диагноз, к примеру, MS, указывается по повышению по меньшей мере в 2, 3, 4, 5, 10, 50, 100, 500 или 1000 раз количества белка, т.е. белка Rep либо антител против Rep, соответственно, в образце от субъекта по сравнению с контрольным образцом.
“Антитело против Rep” в настоящем изобретении означает антитело, которое связывается на заметном уровне с белком Rep в способах по изобретению с более сильным сродством к белку Rep по изобретению, чем не к белку Rep. Предпочтительно сродство к антигену для белка Rep по меньшей мере в 2 раза больше фонового связывания. В частности, антитело против Rep специфично к Rep MSBI1 с аминокислотной последовательностью SEQ ID NO: 1 или 11 либо к Rep MSBI2 с аминокислотной последовательностью SEQ ID NO: 8 или 12. В определенных воплощениях антитело обладает перекрестной специфичностью для Rep MSBI1 и Rep MSBI2.
Общей особенностью всех анализов является то, что белок Rep приводится в контакт с образцом, предположительно содержащим антитела против белка Rep в условиях, позволяющих белку Rep связываться с любым таким антителом, присутствующим в образце. Такими условиями обычно являются физиологическая температура, pH и ионная сила с использованием избытка белка Rep. За инкубацией белка Rep с образцом следует детектирование иммунных комплексов, состоящих из антигена. В некоторых воплощениях либо белок Rep связан с издающим сигнал соединением, к примеру, детектируемой меткой, либо для детектирования иммунного комплекса используется дополнительный связывающий агент, напр., вторичное антитело против человека, связанное с издающим сигнал соединением.
Антитела против Rep можно детектировать и определять методами на основе белка Rep в качестве белкового антигена, который служит мишенью для антител млекопитающих, напр., человека, предполагаемых в образце. Предпочтительно белок Rep очищают (напр., см. пример 1), а образцами могут быть, к примеру, образцы сыворотки или плазмы. Способы включают иммобилизацию белка Rep на матриксе с последующей инкубацией иммобилизованного белка Rep с образцами. Наконец, связавшиеся с Rep антитела из образовавшегося иммунологического комплекса между белком Rep и антителами из образцов определяют с помощью детектирующего связывающего агента, связанного с издающим сигнал соединением, напр., конъюгированного с HRP (пероксидазой хрена) вторичного детектирующего антитела, позволяющего проводить определение на основе субстрата HRP. Это издающее сигнал соединение или метка детектируются сами по себе или же могут реагировать с другим соединением с образованием детектируемого продукта.
В других воплощениях антитела против Rep определяют косвенным образом, при этом сначала антитела из образца иммобилизируют на матриксе, а затем инкубируют с определенным количеством белка Rep, причем антитела против Rep, иммобилизованные и находящиеся на матриксе, захватывают белок Rep из жидкой смеси белка с образцом, после чего проводится определение связавшегося белка Rep.
В других воплощениях белок Rep можно экспрессировать в клетках, а эти клетки инкубировать с образцом. После этого детектируют и определяют антитела против Rep из образца, связавшиеся с белком Rep, экспрессированным в клетках.
Схема иммуноанализа подвержена значительным вариациям, и в данной области известно много форматов. В методиках, к примеру, могут применяться твердые носители либо иммунопреципитация. В большинстве методов применяются связывающие агенты, связанные с издающими сигнал соединениями, к примеру, меченые антитела или меченый белок Rep; метки могут быть, к примеру, ферментативные, флуоресцентные, хемилюминесцентные, радиоактивные или же молекулы красителей. Также известны методы, в которых усиливаются сигналы от иммунного комплекса; примерами таковых являются методы, в которых используются биотин и авидин или стрептавидин, а также иммуноанализы с мечеными ферментами и иммуноферментные типа методов ELISA.
Иммуноанализ может быть в гетерогенном или гомогенном формате, стандартного или конкурентного типа. В гетерогенном формате полипептид (белок Rep или антитело против Rep) обычно связывают с твердым матриксом, подложкой или носителем, чтобы облегчить отделение образца от полипептида после инкубации. Примерами твердых подложек, которые можно использовать, являются нитроцеллюлоза (напр., в виде мембраны или лунок микропланшета), поливинилхлорид (напр., в виде листов или лунок микропланшета), полистироловый латекс (напр., в виде шариков или микропланшета), поливинилидинфторид (известный как Immunolon), диазотированная бумага, нейлоновые мембраны, активированные шарики и шарики с белком A. Твердые подложки, содержащие антигенные полипептиды, обычно промывают после отделения их от исследуемого образца и перед детектированием связавшихся антител против Rep. В данной области известны как стандартные, так и конкурентные форматы.
В гомогенном формате исследуемый образец инкубируют с белком Rep в растворе. Например, это может быть в условиях, при которых осаждаются любые образующиеся комплексы белок Rep-антитело. В данной области для этих методов известны как стандартные, так и конкурентные форматы.
В стандартном формате прямо отслеживается содержание антител против Rep в комплексах белок Rep-антитело. Это может осуществляться путем определения того, что (меченые) антиксеногенные (напр., против человеческих детерминант) антитела, которые распознают эпитоп на антителах против Rep, будут связываться вследствие образования комплекса. В конкурентном формате содержание антител против Rep в образце определяется путем отслеживания влияния конкуренции на связывание известного количества меченого антитела (или другого конкурирующего лиганда) в комплексе.
Образовавшиеся комплексы, содержащие антитело против Rep (или, в случае конкурентных методов, содержание конкурирующих антител), детектируют любым из целого ряда известных методов в зависимости от формата. Например, можно детектировать немеченые антитела против Rep в комплексе с помощью конъюгата антиксеногенного Ig в комплексе с меткой (напр., ферментной меткой, такой, к примеру, как HRP).
В формате методов иммунопреципитации или агглютинации при реакции между белком Rep и антителом против Rep образуется сетчатая структура, которая выпадает в осадок из раствора или суспензии и образует видимый слой или пленку осадка. Если в образце нет антител против Rep, то видимого осадка не образуется.
Выбранная твердая фаза может включать полимерные или стеклянные шарики, нитроцеллюлозу, микрочастицы, микролунки реакционного планшета, пробирки и магнитные шарики. Издающие сигнал соединения могут включать ферменты, люминесцентные соединения, хромогены, радиоактивные элементы и хемилюминесцентные соединения. Примеры ферментов включают щелочную фосфатазу, пероксидазу хрена (HRP) и β-галактозидазу. Примеры соединений-усилителей включают биотин, анти-биотин и авидин. Примеры веществ, связывающих соединения-усилители, включают биотин, анти-биотин и авидин.
В других воплощениях изобретения предусмотрены способы, в которых повышение содержания белка Rep в образце коррелирует с диагнозом или предрасположенностью к нейродегенеративному заболеванию, к примеру, MS, или используется для мониторинга заболевания, к примеру, MS, или мониторинга лечения болезни, к примеру, MS. В таких воплощениях белок Rep в образце детектируется с помощью антител против Rep.
Такие способы включают стадии:
(a) определение количества белка Rep в образце от субъекта с помощью антител против Rep; и
(b) коррелирование количества белка Rep, обнаруженного в образце от субъекта на стадии (a), по сравнению с количеством в контрольном образце, с диагнозом нейродегенеративного заболевания, к примеру, MS.
Примеры методов, которые можно использовать в таких способах обнаружения белка Rep в образцах сыворотки или плазмы, включают, без ограничения, иммунопреципитацию, иммунофлюоресценцию, дот-блоттинг и вестерн-блоттинг.
Например, образец сыворотки можно инкубировать с антителами против белка Rep для захвата белка Rep в образце с последующей стадией иммунопреципитации белка Rep, а затем стадией детектирования методами SDS-PAGE и вестерн-блоттинга.
В другом примере можно инкубировать мембрану для дот-блоттинга с сывороткой с последующей стадией SDS-PAGE и вестерн-блоттинга.
В другом примере разведения сыворотки из образца наносят на SDS-PAGE с последующим вестерн-блоттингом.
В других воплощениях белок Rep выявляют в образцах тканей иммуногистохимическими методами или методом иммунофлуоресцентной микроскопии.
В некоторых воплощениях для обнаружения или захвата белка Rep в образце применяются антитела против Rep.
Термин “антитело” предпочтительно относится к таким антителам, которые в основном состоят из сборных поликлональных антител с различной специфичностью к эпитопам, а также к препаратам различных моноклональных антител. В настоящем изобретении термин “антитело” (Ab) или “моноклональное антитело” (mAb) служит для обозначения интактных молекул иммуноглобулина, а также фрагментов антител (таких, к примеру, как фрагменты Fab и F(ab′)2), которые способны специфически связываться с белком Rep. Фрагменты Fab и F(ab′)2 лишены Fc-фрагмента интактных антител, быстрее выводятся из кровотока и могут иметь меньшее неспецифическое связывание с тканями по сравнению с интактными антителами. Поэтому эти фрагменты предпочтительны, а также продукты библиотек, экспрессирующих Fab или другие иммуноглобулины. Кроме того, антитела, применимые для целей настоящего изобретения, включают химерные, одноцепочечные, мультифункциональные (напр., биспецифичные) и гуманизированные антитела или человеческие антитела.
В некоторых воплощениях антитела либо их антигенсвязывающие фрагменты связаны с издающим сигнал соединением, напр., несут детектируемую метку. Антитела/фрагменты могут быть прямо или косвенно помечены детектируемой меткой, к примеру, радиоизотопом, флуоресцентным соединением, биолюминесцентным соединением, хемилюминесцентным соединением, хелатором металлов или ферментом. Рядовым специалистам в данной области должны быть известны и другие подходящие метки для связывания с антителами или же они смогут установить таковые с помощью обычных экспериментов.
Как показано в разделе «Примеры», новый сконструированный двойной мутант Rep проявляет сравнимые показатели покрытия при ELISA при сравнении с белком Rep дикого типа. Кроме того, реактивность реагирующих с Rep антител в сыворотке людей с MS повышалась в среднем примерно на 40% по сравнению с Rep дикого типа при использовании двойного мутанта Rep 27/154E в качестве антигена при анализах ELISA (фиг. 6).
Авторы изобретения также получили (создали) антитела против белка Rep, имеющего аминокислотную последовательность SEQ ID NO:1 или SEQ ID NO:8, либо его фрагментов, способами, хорошо известными специалистам в данной области. Эти антитела против Rep применяются для связывания с несколькими или всеми видами белков Rep из группы генома Small Sphinx (антитела против Rep типа Small Sphinx или антитела против SSLRep). Такие антитела против SSLRep связываются с эпитопом в пределах консервативной N-концевой области белка Rep, состоящей из аминокислот 1-229 по SEQ ID NO:1. Применяются антитела против Rep типа анти-SSLRep, которые связываются с эпитопом в пределах SEQ ID NO:2 (аминокислоты 32-49 по SEQ ID NO: 1) или SEQ ID NO:3 (аминокислоты 197-216 по SEQ ID NO:1). Пептидные фрагменты по SEQ ID NO:2 и SEQ ID NO:3 очень консервативны среди белков Rep из группы генома Small Sphinx и явно выходят на поверхность вследствие их гидрофильности. Антитела против Rep типа анти-SSLRep могут быть получены путем иммунизации, к примеру, мышей или морских свинок пептидами, состоящими в основном из аминокислотных последовательностей, приведенных в SEQ ID NO:2 или 3; или другими иммуногенными фрагментами, предпочтительно содержащими по меньшей мере 8-15 аминокислот, происходящих из консервативной N-концевой области белка Rep из аминокислот 1-229 по SEQ ID NO:1.
Такие антитела получали, к примеру, путем иммунизации млекопитающих типа мышей или морских свинок полноразмерным белком Rep, имеющим аминокислотную последовательность SEQ ID NO:1.
Эти антитела против Rep способны обнаруживать белок Rep в пределах от пикограмм до фемтограмм в различных видах жидкостей организма, таких, к примеру, как кровь, сыворотка, спинномозговая жидкость или церебральная жидкость.
Можно использовать либо определенный вид антител против Rep, либо набор из двух или нескольких различных видов антител против Rep. При использовании набора из различных видов антител против Rep этот набор антител может содержать различные антитела против Rep, связывающиеся с разными эпитопами в различных доменах белка Rep, напр., первом ДНК-связывающем домене (напр., а.к. 1-136 по SEQ ID NO:1), втором ДНК-связывающем домене (напр., а.к. 137-229 по SEQ ID NO:2) и/или вариабельном домене (напр., а.к. 230-324 по SEQ ID NO:1), в частности, белка Rep MSBI1 (SEQ ID NO:1).
Ввиду только двух точечных мутаций в двух ДНК-связывающих доменах и сохранности одной и той же последовательности в вариабельных доменах эти антитела также распознают мутантные белки по SEQ ID NO: 11 и 12.
Для обнаружения белка Rep с помощью антител против Rep можно применять такие методы, к примеру, как, вестерн-блоттинг, иммунофлуоресцентная микроскопия или иммуногистохимические методы.
Антитела против Rep способны выявлять белок Rep в определенных клеточных локализациях. Например, антитела против Rep могут выявлять белок Rep в цитоплазме, ядерной мембране и ядре или же выявлять вкрапления в цитоплазме. Примеры таких групп антител против Rep представлены в табл. 1.
Антитела против Rep в группе A имеют эпитоп в пределах аминокислотной последовательности, приведенной в SEQ ID NO: 3 (а.к. 198-217 по SEQ ID NO: 1), и способны выявлять белки Rep и Rep MSBI1, содержащие этот консервативный эпитоп из группы генома Small Sphinx (напр., MSBI2, CMI1, CMI4). При иммунофлуоресцентном анализе такие антитела против Rep выявляют специфический профиль локализации Rep, при котором основная локализация однородно распределяется по цитоплазме и ядерной мембране; и дополнительная слабая и однородно распределенная локализация наблюдается в ядре. Примером таких антител группы A является антитело АВ01 523-1-1 (DSM ACC3327), которое использовалось в примерах в качестве антитела группы A.
Антитела против Rep в группе B имеют эпитоп в пределах аминокислотной последовательности, приведенной в SEQ ID NO: 2 (а.к. 33-50 по SEQ ID NO: 1), и способны выявлять белки Rep и Rep MSBI1, содержащие этот консервативный эпитоп из группы генома Small Sphinx (напр., MSBI2, CMI1, CMI4). При иммунофлуоресцентном анализе такие антитела против Rep специфически выявляют вкрапления (цитоплазматические скопления) белка Rep (зачастую на периферии ядерной мембраны). Примером таких антител группы B является антитело, обозначенное как AB02 304-4-1 (DSM ACC3328), которое использовалось в примерах в качестве антитела группы B.
Антитела против Rep в группе C специфически выявляют структурный эпитоп MSBI1 (SEQ ID NO: 1). При иммунофлуоресцентном анализе такие антитела против Rep выявляют специфический профиль локализации Rep, при котором основная локализация однородно распределяется по цитоплазме и ядерной мембране; и дополнительная слабая и однородно распределенная локализация наблюдается в ядре. Примером таких антител группы C является антитело MSBI1 381-6-2 (DSM ACC3329), которое использовалось в примерах в качестве антитела группы C.
Антитела против Rep в группе D специфически выявляют структурный эпитоп MSBI1 (SEQ ID NO: 1), причем антитело MSBI1 961-2-2, обозначенное как “D1”, выявляет эпитоп, приведенный в SEQ ID NO: 9 (а.к. 281-287) в С-концевом домене MSBI1. Антитело MSBI1 761-5-1 (DSM ACC3328), обозначенное как “D2”, выявляет трехмерный структурный эпитоп MSBI1, который доступен исключительно в условиях in vivo и недоступен при Вестерн-блоттинге. При иммунофлуоресцентном анализе такие антитела против Rep специфически выявляют вкрапления (цитоплазматические скопления) белка Rep (зачастую на периферии ядерной мембраны).
В некоторых воплощениях антитела против Rep из группы A, B, C или D либо комбинации антител против Rep по меньшей мере из двух разных групп A, B, C или D применяются для определения типа локализации белка Rep в пробе и коррелирует ли такая локализация белка Rep с функцией патогена. Например, присутствуют ли вкрапления. В некоторых воплощениях, т.е. способах или наборах по изобретению, по меньшей мере одно антитело против Rep, выбранное из групп A и B, комбинируют по меньшей мере с одним антителом против Rep, выбранным из групп C и D. В определенных воплощениях, т.е. способах или наборах по изобретению, антитело против Rep из группы A комбинируют по меньшей мере с одним другим антителом против Rep, выбранным из групп B, C и D. Например, антитело против Rep из группы A можно комбинировать с другими антителами против Rep из групп C и D. Такие комбинации антител против Rep из разных групп повышают четкость диагностической оценки, в частности, для диагностики MS.
Следующие антитела были депонированы в Deutsche Sammlung  Mikroorganismen und Zellkulturen (DSMZ) [Немецкая коллекция микроорганизмов и клеточных куль тур] 28 сентября 2017 г.:
Mikroorganismen und Zellkulturen (DSMZ) [Немецкая коллекция микроорганизмов и клеточных куль тур] 28 сентября 2017 г.:
антитело AB01 523-1-1 за № DSM ACC3327;
антитело AB02 304-4-1 за № DSM ACC3328;
антитело MSBI1 381-6-2 за № DSM ACC3329; и
антитело MSBI1 761-5-1 за № DSM ACC3330.
Антитело MSBI1 961-2-2 было депонировано в DSMZ 6 октября 2017 г.
В предпочтительном воплощении антитела против Rep из групп A, B, C или D либо комбинации антител против Rep по меньшей мере из двух разных групп A, B, C или D применяются для определения синтезированного белка Rep, кодируемого геномом MSBI1 (Rep MSBI1 27/154E или Rep MSBI2 27/154E).
В других воплощениях предусмотрены наборы для применения при диагностике MS. Набор может включать материал для обнаружения антител против Rep и/или белка Rep вместе с инструкциями по применению материалов при анализах для диагностики MS. Набор может содержать один или несколько из следующих компонентов: биомаркер по изобретению, то есть белок Rep или антитела против Rep, соответственно; издающее сигнал соединение; твердый матрикс для присоединения захватывающего агента; разбавитель для образцов; промывочный буфер. Издающее сигнал соединение относится к детектируемой метке, которая либо связана с дополнительным связывающим агентом, способным связываться с биомаркером по изобретению, либо прямо конъюгирована с биомаркером по изобретению.
ПРИМЕРЫ
Далее изобретение раскрывается на следующих примерах, но не ограничивается ими.
Пример 1
Очистка белка Rep MSBI1
Молекулу нуклеиновой кислоты, кодирующую полноразмерную открытую рамку считывания (ORF) Rep, идентифицированную в геноме MSBI1, клонировали в экспрессионную плазмиду (pEXP5-CT, Invitrogen), обеспечивающую экспрессию белка на основе бесклеточной системы трансляции E. coli in vitro с высоким выходом (Expressway™ Cell-Free E. coli Expression System, Invitrogen). Синтезированный белок Rep, имеющий аминокислотную последовательность SEQ ID NO: 1, в реакционной смеси для трансляции in vitro денатурировали добавлением 20 реакционных объемов 8 М мочевины в буфере для образцов рН 8,0, содержащем 100 мМ NaH2PO4, 10 мМ трис-HCl, рН 8,0, 5 мМ имидазола. Затем белок Rep очищали в денатурирующих условиях (20 мМ имидазол для промывки и 300 мМ имидазол для элюирования белка) на основе C-концевой метки His6-tag, слитой с белком Rep. Качество очистки определяли путем окрашивания белка Кумасси и вестерн-блоттинга с антителами против белка Rep. Чистоту белка Rep рассчитывали денситометрически, она составляла более 95%. Очищенный белок либо непосредственно использовали для скрининга сыворотки на основе ELISA, либо подвергали SDS-PAGE с последующим переносом путем блоттинга на нитроцеллюлозные мембраны для инкубации с сывороткой полосок мембраны с белком Rep после разделения по размерам.
Пример 2
A. Характеристика белка Rep дикого типа или мутантного белка Rep в денатурирующих условиях методами SDS-PAGE и вестерн-блоттинга
Свойства очищенных антигенов Rep (дикого типа и мутанта Rep) изучали в различных сериях экспериментов. Белок Rep, который очищали в неденатурирующих (нативных) условиях из клеток линии HEK293TT человека, проявлял значительно меньший уровень олигомеров Rep (полоса в 100 кДа) и меньше высокомолекулярных агрегатов у одиночных мутантов C154E и двойных мутантов 27/154E по сравнению с диким типом при нанесении образцов как в восстановительных, так и в окислительных условиях при проведении SDS-PAGE и вестерн-блоттинга против Rep (фиг. 1).
B. Характеристика белка Rep дикого типа или мутантного белка Rep в денатурирующих условиях методом нативного PAGE
Изучение тех же неденатурированных (нативных) белков Rep методом нативного PAGE также показало значительно меньше высокомолекулярных агрегатов (размазанная высокомолекулярная полоса) у двойного мутанта по сравнению с диким типом, как при обнаружении методом вестерн-блоттинга (WB, анти-Rep), так и при высокочувствительном окрашивании общего белка (окрашивание серебром) (фиг. 2). Однако в целом при PAGE в нативных условиях олигомерный белок Rep (м.в. около 150 кДа) представляет доминирующий вид белка Rep. Это могли быть тримеры либо тетрамеры мономерного Rep (теоретический м.в. 38,2 кДа).
C. Характеристика белка Rep дикого типа или мутантного белка Rep в денатурирующих условиях методами SDS-PAGE и вестерн-блоттинга
Для стандартных методов ELISA белок Rep очищали в денатурирующих условиях из E. coli для получения с высоким выходом и лучшей стабильности белка (см. методику ниже). Для изучения агрегации денатурированного Rep дикого типа и мутантного Rep белки выдерживали 1 час в PBS, чтобы имитировать ренатурирующие условия покрытия и блокирования при ELISA, а затем подвергали SDS-PAGE либо вестерн-блоттингу с антителами против Rep или окрашиванием белков Кумасси (фиг. 3). Опять же, мутант Rep проявлял значительное снижение (> 50%) олигомеров Rep и более крупных агрегатов по сравнению с белком Rep дикого типа.
D. Характеристика белка Rep дикого типа или мутантного белка Rep в денатурирующих условиях методами BN-PAGE и вестерн-блоттинга
Изучение тех же самых образцов (по 5 мкг) методом BN-PAGE с последующим вестерн-блоттингом против Rep также показало значительно меньше олигомеров Rep, а также меньше высокомолекулярных агрегатов (фиг. 4). Дополнительная полоса по всей вероятности мономерного белка Rep обнаруживалась при PAGE в нативных условиях только у мутанта Rep.
Все результаты свидетельствуют о существовании очень стабильных олигомеров Rep (м.в. ~110 кДа), которые даже выдерживают обработку SDS при SDS-PAGE. Также при SDS-PAGE и вестерн-блоттинге выявляются очень стабильные агрегаты Rep, охватывающие еще более высокие диапазоны молекулярной массы (120-170 кДа и выше). Ясно, что интенсивность таких высокомолекулярных агрегатов значительно снижается у двойного мутанта Rep 27/154E. В общем, присутствие агрегатов Rep было выше у тех белков, которые подвергались очистке в неденатурирующих условиях. В любом случае, также белки, очищенные и хранящиеся в денатурирующих условиях, проявляют очень сильную агрегацию, которая значительно снижается у двойного мутанта Rep. Это представляет особый интерес, так как экспериментальные установки ELISA основаны на ренатурации поверхностных белковых антигенов, поскольку связывание антигена антителами требует нативных условий.
Пример 3
A. Показатели покрытия при ELISA у белка дикого типа или мутантного белка Rep
Важно, что недавно сконструированный двойной мутант Rep проявляет сравнимую эффективность покрытия при ELISA по сравнению с белком Rep дикого типа (буферы для анализа: казеиновый буфер, буфер Superblock, 1% BSA в PBS) (фиг. 5).
B. Показатели белка дикого типа или мутантного белка Rep при ELISA в сыворотке людей с MS
Кроме того, реактивность реагирующих с Rep антител в сыворотке людей с MS повышалась в среднем примерно на 40% при использовании в качестве антигена двойного мутанта Rep 27/154E по сравнению с Rep дикого типа при анализе методом ELISA (фиг. 6).
Пример 4
Очистка антигена дикого типа и мутантного белка Rep
Молекулу нуклеиновой кислоты, кодирующую полноразмерную открытую рамку считывания (ORF) Rep дикого типа, идентифицированную в геноме MSBI1, либо молекулу нуклеиновой кислоты двойного мутанта Rep 27/154E клонировали в экспрессионную плазмиду (pEXP5-CT, Invitrogen), обеспечивающую экспрессию белка в E. coli. Вкратце, химически компетентные клетки E. coli (SoluBl21, Genlantis) трансфецировали экспрессионной плазмидой с последующим клональным отбором на чашках с LB-агаром с ампициллином. Клоны с высоким уровнем экспрессии белка отбирали по пробной экспрессии белка при предварительном скрининге в небольшом объеме. При этом клональные культуры размножали в 10 мл LB Amp (37°C, на качалке) до достижения OD600 = 0,4-0,6 с последующей индукцией экспрессии белка с помощью изопропил-β-D-1-тиогалактопиранозида (IPTG, 0,66 мМ) в течение ночи при 25°C на качалке.
Экспрессию целевого белка определяли методами SDS-PAGE и вестерн-блоттинга лизатов E. coli (приготовленных в буфере  для образцов SDS-PAGE) с помощью антител против His, реагирующих с С-концевой меткой 6xHis белка Rep. Затем исходную культуру E. coli, проявлявшую наибольшую экспрессию белка Rep, дополнительно размножали в 1000 мл LB Amp и доводили до OD600 = 0,5. Экспрессию белка индуцировали IPTG (0,66 мМ) в течение ночи при 25°C на качалке. Затем клетки центрифугировали при 6000 g при 4°C, промывали PBS и хранили при -80°C порциями (10 аликвот, что соответствует по 100 мл исходной культуры E. coli, индуцированной IPTG, на 1000 мл).
для образцов SDS-PAGE) с помощью антител против His, реагирующих с С-концевой меткой 6xHis белка Rep. Затем исходную культуру E. coli, проявлявшую наибольшую экспрессию белка Rep, дополнительно размножали в 1000 мл LB Amp и доводили до OD600 = 0,5. Экспрессию белка индуцировали IPTG (0,66 мМ) в течение ночи при 25°C на качалке. Затем клетки центрифугировали при 6000 g при 4°C, промывали PBS и хранили при -80°C порциями (10 аликвот, что соответствует по 100 мл исходной культуры E. coli, индуцированной IPTG, на 1000 мл).
Синтезированный белок Rep, имеющий аминокислотную последовательность SEQ ID NO: 1 (дикий тип) или SEQ ID NO: 11 (мутант), в среде для экспрессии в E. coli денатурировали добавлением 20 реакционных объемов 8 М мочевины в буфере для образцов рН 8,0, содержащем 100 мМ NaH2PO4, 10 мМ трис-HCl, рН 8,0, 5 мМ имидазола, 5 мМ бета-меркаптоэтанола. Затем белок Rep очищали в денатурирующих условиях (55 мМ имидазол для промывки и 300 мМ имидазол для элюирования белка) на основе C-концевой метки His6-tag, слитой с белком Rep. Качество очистки определяли путем окрашивания белка Кумасси и вестерн-блоттинга с антителами против белка Rep. Чистоту белка Rep рассчитывали денситометрически, она составляла более 95%. Очищенный белок либо непосредственно использовали для скрининга сыворотки на основе ELISA, либо подвергали SDS-PAGE с последующим переносом путем блоттинга на нитроцеллюлозные мембраны для инкубации с сывороткой полосок мембраны с белком Rep после разделения по размерам.


БИБЛИОГРАФИЯ
Funk M. et al. (2014) “Isolation of protein-associated circular DNA from healthy cattle serum”. Genome Announc. 2(4).
Giraldo R. et al. (2011) “RepA-WH1 prionoid: a synthetic amyloid proteinopathy in a minimalist host”. Prion 5(2):60-64.
Giraldo R. (2007) “Defined DNA sequences promote the assembly of a bacterial protein into distinct amyloid nanostructures”. Proc. Natl. Acad. Sci. USA 104(44): 17388-17393.
Gunst K. et al. (2014) “Isolation of bacterial plasmid-related replication-associated circular DNA from a serum sample of a multiple sclerosis patient”. Genome Announc. 2(4).
Lamberto I. et al. (2014) “Mycovirus-like DNA virus sequences from cattle serum and human brain and serum samples from multiple sclerosis patients”. Genome Announc. 2(4).
Linding R., J. Schymkowitz, F. Rousseau, F. Diella and L. Serrano (2004) “A comparative study of the relationship between protein structure and beta-aggregation in globular and intrinsically disordered proteins”. J. Mol. Biol. 342(1): 345-353.
Manuelidis L. (2011) “Nuclease resistant circular DNAs co-purify with infectivity in scrapie and CJD”. J. Neurovirol. 17:131-145.
Rousseau F., J. Schymkowitz and L. Serrano (2006). “Protein aggregation and amyloidosis: confusion of the kinds?” Curr. Opin. Struct. Biol. 16(1): 118-126.
Torreira E. et al. (2015) “Amyloidogenesis of bacterial prionoid RepA-WH1 recapitulates dimer to monomer transitions of RepA in DNA replication initiation”. Structure 23(1):183-189.
Whitley C. et al. (2014) “Novel replication-competent circular DNA molecules from healthy cattle serum and milk and multiple sclerosis-affected human brain tissue”. Genome Announc. 2(4).
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДОЙЧЕС КРЕБСФОРШУНГСЦЕНТРУМ
<120> Mutantis Rep-Protein
<130> K 3635EP
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 324
<212> PRT
<213> homo sapiens
<400> 1
Met Ser Asp Leu Ile Val Lys Asp Asn Ala Leu Met Asn Ala Ser Tyr
1 5 10 15
Asn Leu Ala Leu Val Glu Gln Arg Leu Ile Leu Leu Ala Ile Ile Glu
20 25 30
Ala Arg Glu Thr Gly Lys Gly Ile Asn Ala Asn Asp Pro Leu Thr Val
35 40 45
His Ala Ser Ser Tyr Ile Asn Gln Phe Asn Val Glu Arg His Thr Ala
50 55 60
Tyr Gln Ala Leu Lys Asp Ala Cys Lys Asp Leu Phe Ala Arg Gln Phe
65 70 75 80
Ser Tyr Gln Glu Lys Arg Glu Arg Gly Arg Ile Asn Ile Thr Ser Arg
85 90 95
Trp Val Ser Gln Ile Gly Tyr Met Asp Asp Thr Ala Thr Val Glu Ile
100 105 110
Ile Phe Ala Pro Ala Val Val Pro Leu Ile Thr Arg Leu Glu Glu Gln
115 120 125
Phe Thr Gln Tyr Asp Ile Glu Gln Ile Ser Gly Leu Ser Ser Ala Tyr
130 135 140
Ala Val Arg Met Tyr Glu Leu Leu Ile Cys Trp Arg Ser Thr Gly Lys
145 150 155 160
Thr Pro Ile Ile Glu Leu Asp Glu Phe Arg Lys Arg Ile Gly Val Leu
165 170 175
Asp Thr Glu Tyr Thr Arg Thr Asp Asn Leu Lys Met Arg Val Ile Glu
180 185 190
Leu Ala Leu Lys Gln Ile Asn Glu His Thr Asp Ile Thr Ala Ser Tyr
195 200 205
Glu Gln His Lys Lys Gly Arg Val Ile Thr Gly Phe Ser Phe Lys Phe
210 215 220
Lys His Lys Lys Gln Asn Ser Asp Lys Thr Pro Lys Asn Ser Asp Ser
225 230 235 240
Ser Pro Arg Ile Val Lys His Ser Gln Ile Pro Thr Asn Ile Val Lys
245 250 255
Gln Pro Glu Asn Ala Lys Met Ser Asp Leu Glu His Arg Ala Ser Arg
260 265 270
Val Thr Gly Glu Ile Met Arg Asn Arg Leu Ser Asp Arg Phe Lys Gln
275 280 285
Gly Asp Glu Ser Ala Ile Asp Met Met Lys Arg Ile Gln Ser Glu Ile
290 295 300
Ile Thr Asp Ala Ile Ala Asp Gln Trp Glu Ser Lys Leu Glu Glu Phe
305 310 315 320
Gly Val Val Phe
<210> 2
<211> 18
<212> PRT
<213> homo sapiens
<400> 2
Glu Ala Arg Glu Thr Gly Lys Gly Ile Asn Ala Asn Asp Pro Leu Thr
1 5 10 15
Val His
<210> 3
<211> 21
<212> PRT
<213> homo sapiens
<400> 3
Lys Gln Ile Asn Glu His Thr Asp Ile Thr Ala Ser Tyr Glu Gln His
1 5 10 15
Lys Lys Gly Arg Thr
20
<210> 4
<211> 8
<212> PRT
<213> artificial sequence
<220>
<223> His-Tag
<400> 4
Gly Ala His His His His His His
1 5
<210> 5
<211> 11
<212> PRT
<213> artificial sequence
<220>
<223> T7 Tag
<400> 5
Met Ala Ser Met Thr Gly Gly Gln Gln Met Gly
1 5 10
<210> 6
<211> 8
<212> PRT
<213> artificial sequence
<220>
<223> Flag Tag
<400> 6
Asp Tyr Lys Asp Asp Asp Asp Lys
1 5
<210> 7
<211> 8
<212> PRT
<213> artificial sequence
<220>
<223> Strep II Tag
<400> 7
Trp Ser His Pro Gln Phe Glu Lys
1 5
<210> 8
<211> 319
<212> PRT
<213> homo sapiens
<400> 8
Met Ser Lys Leu Val Val Lys Asp Asn Ala Leu Met Asn Ala Ser Tyr
1 5 10 15
Asn Leu Asp Leu Val Glu Gln Arg Leu Ile Leu Leu Ala Ile Ile Glu
20 25 30
Ala Arg Glu Ser Gly Lys Gly Ile Asn Ala Asn Asp Pro Leu Thr Val
35 40 45
His Ala Glu Ser Tyr Ile Asn Gln Phe Gly Val His Arg Val Thr Ala
50 55 60
Tyr Gln Ala Leu Lys Asp Ala Cys Asp Asn Leu Phe Ala Arg Gln Phe
65 70 75 80
Ser Tyr Gln Ser Lys Ser Glu Lys Gly Asn Ile Gln Asn His Arg Ser
85 90 95
Arg Trp Val Ser Glu Ile Ile Tyr Ile Asp Thr Glu Ala Thr Val Lys
100 105 110
Ile Ile Phe Ala Pro Ala Ile Val Pro Leu Ile Thr Arg Leu Glu Glu
115 120 125
Gln Phe Thr Lys Tyr Asp Ile Glu Gln Ile Ser Asp Leu Ser Ser Ala
130 135 140
Tyr Ala Ile Arg Leu Tyr Glu Leu Leu Ile Cys Trp Arg Ser Thr Gly
145 150 155 160
Lys Thr Pro Ile Ile Gly Leu Gly Glu Phe Arg Asn Arg Val Gly Val
165 170 175
Leu Asp Ser Glu Tyr His Arg Ile Ala His Leu Lys Glu Arg Val Ile
180 185 190
Glu His Ser Ile Lys Gln Ile Asn Glu His Thr Asp Ile Thr Ala Thr
195 200 205
Tyr Glu Gln His Lys Lys Gly Arg Thr Ile Thr Gly Phe Ser Phe Lys
210 215 220
Phe Lys Gln Lys Lys Pro Lys Gln Ala Glu Ile Ala Thr Glu Thr Pro
225 230 235 240
Lys Thr Ala Thr Asn Asp Pro Asp Thr Thr Lys Pro Leu Thr Glu Pro
245 250 255
Gln Ile Ala Lys Tyr Ser Met Ile Leu Cys Lys Leu Gly Ser Ile Ser
260 265 270
Asp Leu Ser Asn Phe Pro Asp Tyr Pro Ala Phe Ala Asn Trp Ile Gly
275 280 285
Asn Ile Leu Arg Asn Pro Glu Lys Ala Asp Glu Gln Ile Ala Lys Arg
290 295 300
Ile Phe Thr Ala Leu Lys Thr Glu Thr Asp Tyr Ser Lys Lys Asn
305 310 315
<210> 9
<211> 7
<212> PRT
<213> homo sapiens
<400> 9
Asn Arg Leu Ser Asp Arg Phe
1 5
<210> 10
<211> 999
<212> DNA
<213> artificial sequence
<220>
<223> MSBI1 Rep 27/154E mutant
<400> 10
atgagcgacc tgatcgtgaa agacaatgcc ctgatgaacg cctcctacaa cctggcactg 60
gtcgaacaga gactgattga gctggctatc atcgaggcaa gggagaccgg caagggcatc 120
aacgccaatg accccctgac agtgcacgcc agctcctaca tcaaccagtt taatgtggag 180
cgccacaccg cctatcaggc cctgaaggac gcctgcaagg atctgtttgc ccggcagttc 240
agctaccagg agaagcggga gagaggcagg atcaacatca caagcagatg ggtgtcccag 300
atcggctata tggacgatac cgccacagtg gagatcatct ttgcaccagc agtggtgcct 360
ctgatcacca ggctggagga gcagttcaca cagtacgaca tcgagcagat ctccggactg 420
tctagcgcct acgccgtgcg catgtatgag ctgctgatcg agtggcggtc taccggcaag 480
acacctatca tcgagctgga tgagttccgc aagcggatcg gcgtgctgga caccgagtac 540
accagaacag ataacctgaa gatgagagtg atcgagctgg ccctgaagca gatcaatgag 600
cacaccgata tcacagcctc ttatgagcag cacaagaagg gccgcgtgat caccggcttc 660
agctttaagt tcaagcacaa gaagcagaac tctgacaaga caccaaagaa tagcgattcc 720
tctccccgga tcgtgaagca cagccagatc cctaccaaca tcgtgaagca gccagagaat 780
gccaagatgt ccgacctgga gcacagggca tctagggtga caggcgagat catgagaaat 840
aggctgagcg atcggttcaa gcagggcgac gagtccgcca tcgatatgat gaagagaatc 900
cagtccgaga tcatcaccga cgccatcgcc gatcagtggg aatctaaact ggaagagttt 960
ggagtcgtgt ttggagcaca tcaccatcat catcactga 999
<210> 11
<211> 324
<212> PRT
<213> artificial sequence
<220>
<223> MSBI1 Rep 27/154E mutant
<400> 11
Met Ser Asp Leu Ile Val Lys Asp Asn Ala Leu Met Asn Ala Ser Tyr
1 5 10 15
Asn Leu Ala Leu Val Glu Gln Arg Leu Ile Glu Leu Ala Ile Ile Glu
20 25 30
Ala Arg Glu Thr Gly Lys Gly Ile Asn Ala Asn Asp Pro Leu Thr Val
35 40 45
His Ala Ser Ser Tyr Ile Asn Gln Phe Asn Val Glu Arg His Thr Ala
50 55 60
Tyr Gln Ala Leu Lys Asp Ala Cys Lys Asp Leu Phe Ala Arg Gln Phe
65 70 75 80
Ser Tyr Gln Glu Lys Arg Glu Arg Gly Arg Ile Asn Ile Thr Ser Arg
85 90 95
Trp Val Ser Gln Ile Gly Tyr Met Asp Asp Thr Ala Thr Val Glu Ile
100 105 110
Ile Phe Ala Pro Ala Val Val Pro Leu Ile Thr Arg Leu Glu Glu Gln
115 120 125
Phe Thr Gln Tyr Asp Ile Glu Gln Ile Ser Gly Leu Ser Ser Ala Tyr
130 135 140
Ala Val Arg Met Tyr Glu Leu Leu Ile Glu Trp Arg Ser Thr Gly Lys
145 150 155 160
Thr Pro Ile Ile Glu Leu Asp Glu Phe Arg Lys Arg Ile Gly Val Leu
165 170 175
Asp Thr Glu Tyr Thr Arg Thr Asp Asn Leu Lys Met Arg Val Ile Glu
180 185 190
Leu Ala Leu Lys Gln Ile Asn Glu His Thr Asp Ile Thr Ala Ser Tyr
195 200 205
Glu Gln His Lys Lys Gly Arg Val Ile Thr Gly Phe Ser Phe Lys Phe
210 215 220
Lys His Lys Lys Gln Asn Ser Asp Lys Thr Pro Lys Asn Ser Asp Ser
225 230 235 240
Ser Pro Arg Ile Val Lys His Ser Gln Ile Pro Thr Asn Ile Val Lys
245 250 255
Gln Pro Glu Asn Ala Lys Met Ser Asp Leu Glu His Arg Ala Ser Arg
260 265 270
Val Thr Gly Glu Ile Met Arg Asn Arg Leu Ser Asp Arg Phe Lys Gln
275 280 285
Gly Asp Glu Ser Ala Ile Asp Met Met Lys Arg Ile Gln Ser Glu Ile
290 295 300
Ile Thr Asp Ala Ile Ala Asp Gln Trp Glu Ser Lys Leu Glu Glu Phe
305 310 315 320
Gly Val Val Phe
<210> 12
<211> 319
<212> PRT
<213> artificial sequence
<220>
<223> MSBI2 Rep 27/154E mutant
<400> 12
Met Ser Lys Leu Val Val Lys Asp Asn Ala Leu Met Asn Ala Ser Tyr
1 5 10 15
Asn Leu Asp Leu Val Glu Gln Arg Leu Ile Glu Leu Ala Ile Ile Glu
20 25 30
Ala Arg Glu Ser Gly Lys Gly Ile Asn Ala Asn Asp Pro Leu Thr Val
35 40 45
His Ala Glu Ser Tyr Ile Asn Gln Phe Gly Val His Arg Val Thr Ala
50 55 60
Tyr Gln Ala Leu Lys Asp Ala Cys Asp Asn Leu Phe Ala Arg Gln Phe
65 70 75 80
Ser Tyr Gln Ser Lys Ser Glu Lys Gly Asn Ile Gln Asn His Arg Ser
85 90 95
Arg Trp Val Ser Glu Ile Ile Tyr Ile Asp Thr Glu Ala Thr Val Lys
100 105 110
Ile Ile Phe Ala Pro Ala Ile Val Pro Leu Ile Thr Arg Leu Glu Glu
115 120 125
Gln Phe Thr Lys Tyr Asp Ile Glu Gln Ile Ser Asp Leu Ser Ser Ala
130 135 140
Tyr Ala Ile Arg Leu Tyr Glu Leu Leu Ile Glu Trp Arg Ser Thr Gly
145 150 155 160
Lys Thr Pro Ile Ile Gly Leu Gly Glu Phe Arg Asn Arg Val Gly Val
165 170 175
Leu Asp Ser Glu Tyr His Arg Ile Ala His Leu Lys Glu Arg Val Ile
180 185 190
Glu His Ser Ile Lys Gln Ile Asn Glu His Thr Asp Ile Thr Ala Thr
195 200 205
Tyr Glu Gln His Lys Lys Gly Arg Thr Ile Thr Gly Phe Ser Phe Lys
210 215 220
Phe Lys Gln Lys Lys Pro Lys Gln Ala Glu Ile Ala Thr Glu Thr Pro
225 230 235 240
Lys Thr Ala Thr Asn Asp Pro Asp Thr Thr Lys Pro Leu Thr Glu Pro
245 250 255
Gln Ile Ala Lys Tyr Ser Met Ile Leu Cys Lys Leu Gly Ser Ile Ser
260 265 270
Asp Leu Ser Asn Phe Pro Asp Tyr Pro Ala Phe Ala Asn Trp Ile Gly
275 280 285
Asn Ile Leu Arg Asn Pro Glu Lys Ala Asp Glu Gln Ile Ala Lys Arg
290 295 300
Ile Phe Thr Ala Leu Lys Thr Glu Thr Asp Tyr Ser Lys Lys Asn
305 310 315
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| REP белок в качестве белкового антигена для диагностических анализов | 2017 |
|
RU2769568C2 |
| ЛЕЧЕНИЕ И ПРОФИЛАКТИКА АЛЛЕРГИИ НА КЛЕЩЕЙ ДОМАШНЕЙ ПЫЛИ | 2019 |
|
RU2815386C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| АНТИТЕЛА ПРОТИВ ТАУ-БЕЛКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2732122C2 |
| МОДУЛЯЦИЯ АКТИВНОСТИ REP БЕЛКА ПРИ ПОЛУЧЕНИИ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (ЗКДНК) | 2020 |
|
RU2812850C2 |
| НЕВИРУСНЫЙ ИММУНОТАРГЕТИНГ | 2020 |
|
RU2833379C2 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2760301C1 |
| СТАБИЛЬНЫЕ СОСТАВЫ АНТИТЕЛ ПРОТИВ TIGIT, ОТДЕЛЬНО И В КОМБИНАЦИИ С АНТИТЕЛАМИ ПРОТИВ РЕЦЕПТОРА 1 ПРОГРАММИРУЕМОЙ СМЕРТИ (PD-1), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2820576C2 |
| НОВЫЙ МОДИФИЦИРОВАННЫЙ СЛИТЫЙ БЕЛОК FC-ФРАГМЕНТА ИММУНОГЛОБУЛИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2800919C2 |
| СУБЪЕДИНИЧНАЯ ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ ИНФЕКЦИИ ДЫХАТЕЛЬНЫХ ПУТЕЙ | 2020 |
|
RU2811991C2 |

Настоящее изобретение относится к области биотехнологии, конкретно к получению мутантного белка, связанного с репликацией ДНК (Rep), и может быть использовано для диагностики рассеянного склероза (MS). Синтезированные одиночный мутант C154E и двойной мутант 27/154E белка Rep характеризуются повышенной стабильностью и меньшей агрегацией по сравнению с белком Rep дикого типа в условиях in vitro, а также в буфере для хранения с мочевиной. Изобретение позволяет получить более стабильные антигены, чем белки Rep MSBI1 и MSBI2 дикого типа, для применения в анализах при диагностическом скрининге MS. 3 н. и 7 з.п. ф-лы, 6 ил., 1 табл., 4 пр.
1. Белок, связанный с репликацией ДНК (Rep), включающий аминокислотную последовательность, приведенную в SEQ ID NO: 11 или 12.
2. Способ диагностики MS у субъекта, включающий стадии:
(a) инкубирование образца от субъекта с белком Rep, определенным в п. 1;
(b) определение количества антител в образце от субъекта, образующих иммунологический комплекс с белком Rep; и
(c) коррелирование количества антител, связавшихся с белком Rep в образце от субъекта, по сравнению с количеством в контрольном образце, с диагнозом MS.
3. Способ по п. 2, при этом на стадии (a) белок Rep иммобилизируют, после чего иммобилизованный белок Rep инкубируют с образцом от субъекта.
4. Способ по п. 2, при этом на стадии (a) белок Rep экспрессируют в клетках, после чего клетки инкубируют с образцом от субъекта.
5. Способ по п. 2, при этом на стадии (b) количество антител, образующих иммунологический комплекс с белком Rep, определяют с помощью детектирующего связывающего агента, соединенного с издающим сигналы соединением.
6. Способ по п. 2, при этом на стадии (a) антитела в образце от субъекта иммобилизируют, после чего инкубируют с определенным количеством белка Rep.
7. Способ по любому из пп. 2-6, при этом образец от субъекта представляет собой образец сыворотки или плазмы.
8. Способ по любому из пп. 2-7, при этом на MS или предрасположенность к MS указывает повышенное содержание антител против Rep в образце от субъекта по меньшей мере в 2 раза по сравнению с контрольным образцом.
9. Набор для применения при диагностике MS, включающий:
(a) белок Rep, причем белок Rep включает аминокислотную последовательность, приведенную в SEQ ID NO: 11 или 12;
(b) антитело против человека, соединенное с детектируемой меткой и способное связываться с антителом против Rep, специфичным для белка, имеющего аминокислотную последовательность по SEQ ID NO: 11 или 12; и
(c) твердый матрикс, подходящий для иммобилизации белка Rep согласно (a).
10. Набор по п. 9 для применения при анализе, выбранном из группы, состоящей из ферментного иммуносорбентного анализа (ELISA), радиоиммуноанализа (RIA), иммуноферментного анализа (EIA), флуоресцентного иммуноанализа (FIA), люминесцентного иммуноанализа (LIA) и анализа на полосках.
| EP 2966176 A1, 13.01.2016 | |||
| WHITLEY C | |||
| ET AL., Novel Replication-Competent Circular DNA Molecules from Healthy Cattle Serum and Milk and Multiple Sclerosis-Affected Human Brain Tissue, GENOME ANNOUNCEMENTS, 2014, v | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Приспособление для усиления тяги в дымоходах и вытяжных каналах | 1925 |
|
SU849A1 |
| СПОСОБ ДИАГНОСТИКИ РАССЕЯННОГО СКЛЕРОЗА | 2003 |
|
RU2369874C2 |

