Настоящая заявка испрашивает приоритет по заявке на предварительный патент 60/055881, содержание которой целиком включено в данную заявку в качестве ссылки.
Настоящее изобретение было отчасти выполнено за счет финансирования системой Федеральных медицинских институтов США, грант за NCI P50 СА68438. В связи с этим правительство США может обладать определенными правами на настоящее изобретение.
Область техники
Настоящее изобретение в целом относится к лечению эстрогензависимых заболеваний и расстройств, и, в частности, к способу лечения эстрогензависимых злокачественных опухолей, конкретно рака молочной железы, с применением антиэстрогеновых средств.
Предпосылки
Рецептор эстрогена человека (ER) является членом суперсемейства ядерных рецепторов транскрипционных факторов (Evans, 1988, Science, 240, 889-895). В отсутствие гормона рецептор остается в ядрах клеток-мишеней в транскрипционно неактивном состоянии. По связыванию своего лиганда ER претерпевает конформационное изменение, сопряженное с каскадом процессов, которые в конечном счете приводят к его связыванию со специфическими регуляторными сегментами генов-мишеней (O'Malley et al., 1991, Hormone Res., 47, 1-26). Конкретная регуляция транскрипции зависит от конкретной клетки и промотора ДНК-связывающего рецептора (Тоrа et а1., 1989, Cell, 59, 477-487; Tasset et al., 1990, Cell, 62, 1177-1187; McDonnell et al., 1995, Mol. Endocrinol., 9, 659-669; Tzukerman et al., 1994, Mol. Endocrinol., 8, 21-30). В этом смысле ясно, что физиологический антагонист активности ER - эстрадиол - проявляет свою биологическую активность в половой скелетной и сердечно-сосудистой системах организма (Clark & Peck, eds., 1979, "Female Sex Steroids: Receptors and Functions", Monographs Endocrinol., Springer-Verlag, New York; Chow et al. , 1992, J. Clin Invest., 89, 74-78; Eaker et al., 1993, Circulation, 88, 1999-2009).
Помимо указанных типов активностей было подтверждено функционирование эстрогена в качестве митогенного фактора в большинстве ER-позитивных клетках опухолей молочной железы. Следовательно, способы лечения, основанные на применении антиэстрогеновых средств, т.е. синтетических соединений, противоположных по действию эстрогену, должны быть клинически эффективными с точки зрения излечения или подавления прогрессирования заболевания (Jordan & Murphy, 1990, Endocrine Rev. , 11, 578-610; Parker, 1993, Breast Cancer Res. Treatment, 26, 131-137). Применение таких синтетических модуляторов активности ER и раскрытие механизма (механизмов) их действия позволит пролить свет на механизмы активности самого ER.
Одно из наиболее изученных в этом отношении соединений - тамоксифен (Jordan & Murphy, 1990, Endocrine Rev., 11, 578-610). Это соединение функционирует в качестве антагониста в большинстве ER-позитивных опухолей молочной железы, но при этом проявляет парадоксальную агонистическую активность в костной ткани и сердечно-сосудистой системе, а также неполную агонистическую активность в матке (Kedar et al., 1994, Lancet, 343, 1318-1321; Love et al. , 1992, New England J. Med., 326, 852-856; Love et al., 1991, Ann. Intern. Med. , 115, 860-864). Следовательно, активность агониста/антагониста, присущая комплексу ER-тамоксифен, зависит от типа клеток. Это принципиальное наблюдение входит в видимое противоречие с долговременными моделями, свидетельствующими, что ER существуют только в клетках, находящихся в активном или неактивном состоянии (С1ark & Peck, eds., 1979, "Female Sex Steroids: Receptors and Functions", Monographs Endocrinol., Springer-Verlag, New York). Напротив, это указывает на то, что действующие по связыванию с одним и тем же рецептором разные лиганды могут проявлять различные биологические эффекты в клетках различного типа. Установление механизма такой избирательности, по-видимому, позволит объяснить некоторые процессы, такие как возникновение резистентности к тамоксифену, выявляемую в большинстве ER-позитивных случаев рака молочной железы, при которых наблюдаются аномальные параметры сигнального пути ER (Tonetti & Jordan, 1995, Anti-Cancer Drugs, 6, 498-507).
С применением тестирования in vitro был определен вероятный механизм избирательно агонистической или антагонистической активности тамоксифена (Тоrа et al. , 1989, Cell, 59, 477-487; Tasset et al., 1990, Cell, 62, 1177-1187; McDonnell et al., 1995, Mol. Endocrinol., 9, 659-669; Tzukerman et al. , 1994, Mol. Endocrinol., 8, 21-30). Важным является то, что была выявлена индукция тамоксифеном изменения конформации ER, которое отличается от такового, индуцируемого эстрадиолом (McDonnell et al., 1995, Mol. Endocrinol. , 9, 659-669; Beekman et a1., 1993, Mol. Endocrinol., 7, 1266-1274). Более того, определение аминокислотных последовательностей в составе ER, необходимых для активации транскрипции, указывает на то, как конкретные комплексы "лиганд-рецептор" дифференцированно распознаются клеточными факторами контроля за транскрипционной активностью. Конкретно было установлено, что ER включает два активационных домена - AF-1 (фактор активаторной функции 1) и AF-2, которые обеспечивают его взаимодействие с контролирующей транскрипцию системой. Относительный вклад этих доменов AF в общую активность ER оказывается неодинаковым в разных клетках (Тоrа et al., 1989, Cell, 59, 477-487; McDonnell et al., 1995, Mol. Endocrinol., 9, 659-669; Tzukerman et al. , 1994, Mol. Endocrinol., 8, 21-30). Для эстрадиола было установлено функционирование в качестве агониста по отношению и к AF-1, и к AF-2: это проявляется в том, что он обусловливает максимальный уровень активности независимо от того, какой из доменов AF является доминантным в конкретной клеточной среде. С другой стороны, тамоксифен функционирует в качестве антагониста домена AF-2, подавляя активность рецептора ER в клетках, в которых AF-2 необходим или является доминантным активаторным фактором (Тоrа et al., 1989, Cell, 59, 477-487; McDonnell et al., 1995, Mol. Endocrinol., 9, 659-669; Tzukerman et al., 1994. Mol. Endocrinol., 8, 21-305. С другой стороны, тамоксифен активен как агонист тогда, когда только AF-1 является необходимым (McDonnell et al. , 1995. Mol. Endocrinol., 9, 659-669; Tzukerman et al., 1994, Mol. Endocrinol., 8, 21-30). Следовательно, основываясь на параметрах относительной активности системы AF-1/AF-2, было идентифицировано четыре кинетически дифференцированных группы ER-модуляторов: полные агонисты (например, эстрадиол), два разных типа частичных агонистов, представленные тамоксифеном и ралоксифеном, и полные антагонисты, представителем которых является препарат ICI182780 (McDonnell et al., 1995, Mol. Endocrinol., 9, 659-669; Tzukerman et al., 1994, Mol. Endocrinol., 8, 21-30). Эти данные позволяют объяснить наблюдаемые различия в проявлениях биологической активности некоторых модуляторов ER, исходя из кинетических параметров (т.е. различий в механизмах реакций), и указывают на то, что механизмы, в соответствии с которыми рецептор ER действует в различных тканях, неодинаковы. Интересно, что проявляемая модуляторами ER, такими как эстроген и тамоксифен, агонистическая активность в подобных системах in vitro отражает их активность в половых путях интактных животных. Эта корреляция, однако, не распространяется на костную ткань, где и эстрадиол и тамоксифен, и ралоксифен, которые проявляют различную степень агонистической активности в отношении системы AF-1/AF-2, эффективно предотвращают уменьшение массы костной ткани у модельных крыс после овариэктомии. Следовательно, за исключением являющихся стероидами полных антиэстрогенов (таких как ICI182780), все известные типы модуляторов ER, по- видимому, защищают уменьшение массы костной ткани у человека и в родственных животных моделях, в то время как они характеризуются различной степенью эстрогенподобной активности в других тканях (Chow et al., 1992, J. Clin. Invest., 89, 74-78; Love et al., 1992, New England J. Med., 326, 852-856; Draper et al., 1993, "Biochemical markers of bone and lipid metabolism in healthy postmenopausal women". In Proc. 4th Intern. Symp. Osteoporosis & Consensus Develop. Conf., С. Christiansen & B. Biis (eds.), Handelstrykkeriet, Aalborg; Wagner et al., 1996, Proc. Natl. Acad. Sci. USA, 93, 8739-8744; Black et al., 1994, J. Clin. Invest., 93, 63-69).
Сущность изобретения
Настоящее изобретение основывается на идентификации модуляторов ER, которые по кинетическим параметрам отличаются от таких модуляторов, каким является тамоксифен. Эти модуляторы находят свое применение в лечении различных эстрогензависимых заболеваний и расстройств, включая рак молочной железы. Эти модуляторы особенно важны в лечении случаев рака молочной железы, которые характеризуются резистентностью к тамоксифену, возникшей de novо, или становятся резистентными в процессе лечения.
Объекты и преимущества настоящего изобретения будут ясны из нижеследующего описания.
Краткое описание чертежей
Фигуры 1А и 1B. GW5638 является соединением, отличающимся по кинетическим параметрам от известных типов модуляторов ER. Промотор гена С3 человека (нуклеотиды [-1807]-[+58]), соединенный с геном-репортером, кодирующим люциферазу светляка, использовали для трансфекции клеток HepG2 наряду с экспрессирующей плазмидой, включающей (фиг.1А) ген рецептора эстрогена человека дикого типа (ERwt) или (фиг.1В) мутированный ген ER, в котором функция активаторного домена AF2 была нарушена (ER-TAF1), с последующим тестированием активации транскрипции в присутствии возрастающих концентраций модуляторов ER, так как это указано. Процесс трансфекции оптимизировали по параметрам эффективности и числу клеток с использованием одновременной трансфекции экспрессирующей плазмидой, включающей репортерный ген β-галактозидазы. Нормализованный ответ получали путем соотнесения единиц индуцируемого света (активность люциферазы) к активности β-галактозидазы, измеряемой в тесте на каталитическую активность. Эксперименты по трансфекции были проведены в трех повторностях. Показанные данные представляют обобщение по нескольким экспериментам, проведенным в сходных условиях.
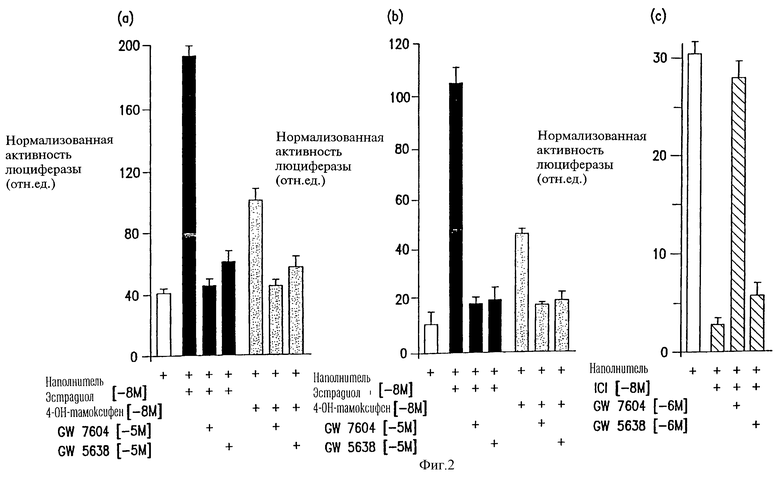
Фигура 2А-2С. GW5638 и GW7604 препятствуют агонистической активности эстрадиола, частичной агонистической активности тамоксифена и инвертивной агонистической активности ICI182780. Фигура 2А. Способность GW5638 или GW7604 подавлять агонистическую активность эстрадиола-17β в концентрации 10-8 М или частичную агонистическую активность, проявляемую тамоксифеном при концентрации 10-8 М, оценивали в клетках HepG2, трансфицированных геном ERwt. Фигура 2В. Способность GW5638 или GW7604 подавлять агонистическую активность эстрадиола-17β в концентрации 10-8 М или частичную агонистическую активность, проявляемую 4-ОН-тамоксифеном при концентрации 10-8 М, оценивали в клетках HepG2, трансфицированных мутантным геном ER-TAF1 (McDonnell et al. , 1995, Mol. Endocrinol., 9, 659-669). Фигура 2С. И GW5638, и GW7604 способны подавлять инвертивную агонистическую активность ICI182780 (далее - ICI) по отношению к ER в связи с контролем активности промотора С3 при тестировании в клетках HepG2 при указанных концентрациях. Процесс трансфекции оптимизировали по параметрам эффективности и числу клеток с использованием одновременной трансфекции экспрессирующей плазмидой, включающей репортерный ген β-галактозидазы. Нормализованный ответ получали путем соотнесения единиц индуцируемого света (активность люциферазы) к активности β-галактозидазы, измеряемой в тесте на каталитическую активность. Показаны данные выборочных тестов, в которых трансфекцию проводили в трех повторностях. Обозначения ошибок соответствуют стандартному отклонению от среднего (s.e.m.).
Фигуры 3А и 3В. GW5638 защищает крыс после овариэктомии от уменьшения массы костной ткани. Фигура 3А. Влияние GW5638 на степень минерализации кости (СМК) в поясничном отделе позвоночника (позвонки L1-L4) измеряли методом двухуровневой рентгеновской абсорбциометрии. Достоверность различий по СМК между крысами OVX и подвергавшимися воздействию животными определяли с помощью теста Даннета (*Р≤0,005). Определены диапазоны величин СМК, выявленных у крыс, которым симулировали операцию по удалению яичников (закрашенные области), и OVX (незакрашенные области). Фигура 3В. Влияние GW5638 на СМК в проксимальном метафизе большеберцовой кости крыс OVX измеряли методом количественной компьютерной томографии (ККТ). Достоверность различий величин СМК между крысами OVX и опытными животными (обозначены звездочками) была определена с помощью теста Турки-Кремера (Р≤0,05).
Фигура 4. GW5638 подавляет вызванное овариэктомией (удалением яичников) повышение уровня сывороточного холестерина. Измерения содержания сывороточного холестерина были проведены в пробах крови, взятых у группы 90-дневных овариэктомизированных крыс, которым вводили либо эстрадиол, либо GW5638, как это указано. Каждая точка указывает на среднее значение сывороточного холестерина (± s. e.m.) для контрольных крыс OVX (n=7), обработанных эстрадиолом (n=7) и GW5638 (n=7), в соответствии с указанным. Звездочками отмечены группы, которые достоверно отличаются от контрольной группы. Определен размах значения уровня сывороточного холестерина у крыс линии OVX (незакрашенные области).
Фигура 5. GW5638 не проявляет ЕR-агонистической активности в матке у неполовозрелых крыс. Группе 21-дневных крысят перорально вводили либо только наполнитель, либо GW5638, либо тамоксифен в виде единственного фактора, либо GW5638 или тамоксифен в присутствие эстрадиола. Показанные данные соответствуют средним значениям (± s.e.m.). Обозначен диапазон измерений, проведенных у крыс, которым вводили эстрадиол (закрашенные области), и у симулятивно прооперированных животных (незакрашенные области).
Фигура 6. Влияние GW5638 на нативный вес матки у крыс с удаленными яичниками. Группе симулятивно прооперированных крыс или группе 90-дневных овариэктомизированных особей вводили в течение 28 дней либо только наполнитель, либо эстрадиол, либо GW5638. Полученные результаты приведены в виде средних значений нативного веса матки (± s.e.m.) для 7 особей каждой группы. Обозначены диапазоны измерений для симулятивно прооперированных крыс (закрашенные области) и животных OVX (незакрашенные области).
Фигуры 7A-7F. Влияние GW5638 на гистологические параметры матки у овариэктомизированных крыс. Дана сравнительная гистологическая картина (при небольшом увеличении) маток у 90-дневных крыс, которые были симулятивно прооперированы (фиг. 7А), которым проводили овариэктомию (фиг.7В), проводили овариэктомию и вводили эстрадиол (фиг.7С) или проводили овариэктомию и вводили GW5638 дозами 1 мкг/кг (фиг.7D), 3 мкг/кг (фиг.7Е) или 10 мкг/кг (фиг. 7F).
Фигуры 8A-8D. Влияние GW5638 на гистологические параметры матки у овариэктомизированных крыс. Дана сравнительная гистологическая картина маток у 90-дневных крыс, которые были симулятивно прооперированы (фиг.8А), которым проводили овариэктомию (фиг.8В), проводили овариэктомию и вводили эстрадиол (фиг. 8С) или проводили овариэктомию и вводили GW5638 дозой 10 мкг/кг (фиг. 8D). Микрофотографии сделаны при увеличении 150х и затем увеличены до окончательного увеличения 600х.
Фигура 9. Влияние противоэстрогеновой обработки на опухоль молочной железы MCF-7 у мышей линии "nude". День "0" указывает на начальный день обработки, имевший место через 2 недели после инокуляции опухоли, а статистический анализ показал, что каждая обработанная группа достоверно превосходит контроль (ANOVA: Р<0,5) при отсутствии достоверных различий между двумя наивысшими дозами GW5638 и тамоксифена.
Фигура 10. Анализ зависимости от дозы.
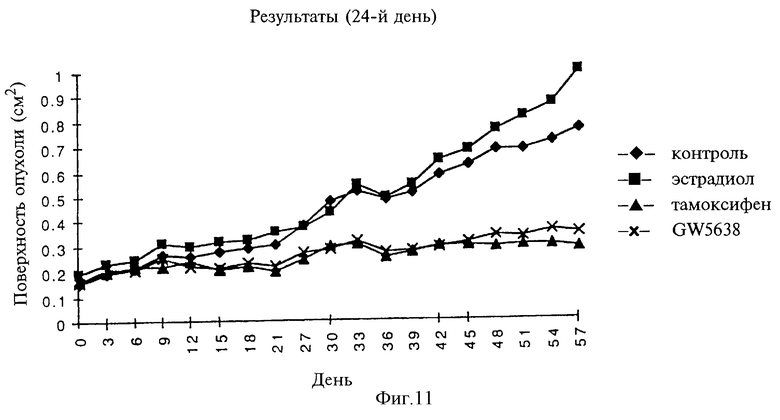
Фигура 11. Анализ LCC2.
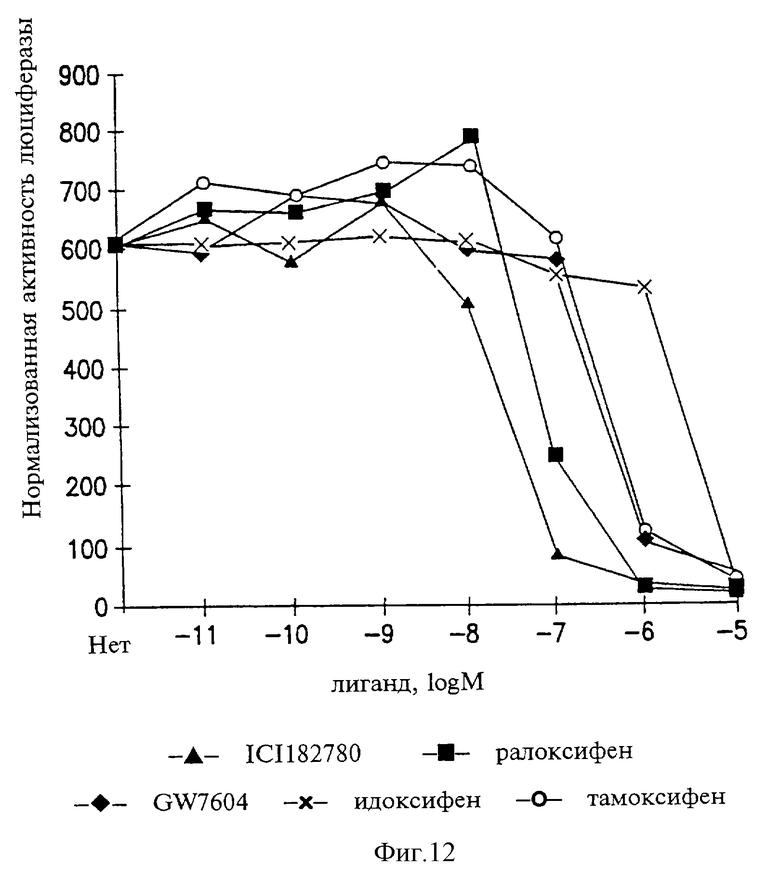
Фигура 12. Активность GW5638 как антиэстрогена в клетках опухоли молочной железы MCF-7.
Фигуры 13А и 13В. Анализ влияния отдельных мутаций в ER на фармакологические параметры антиэстрогенов, показывающий существование дополнительной кинетической комплексности. фиг. 13А - ERwt (дикий тип). Фиг.13В - ER-TAF1 (мутантный вариант ER).
Фигуры 14А и 14В. Сравнительный анализ способности ряда антиэстрогеновых средств подавлять транскрипционную активность ERα (фиг.14А) и ERβ (фиг.14В).
Фигуры 15А-15С. Анализ экспрессии ER методом иммунологического Вестерн-блоттинга в клетках-мишенях после обработки их агонистами или антагонистами. Фиг.15А - клетки MCF-7. Фиг.15В - клетки Ishikawa. Фиг.15С - клетки Ishikawa, трансфицированные плазмидой pRST7ER.
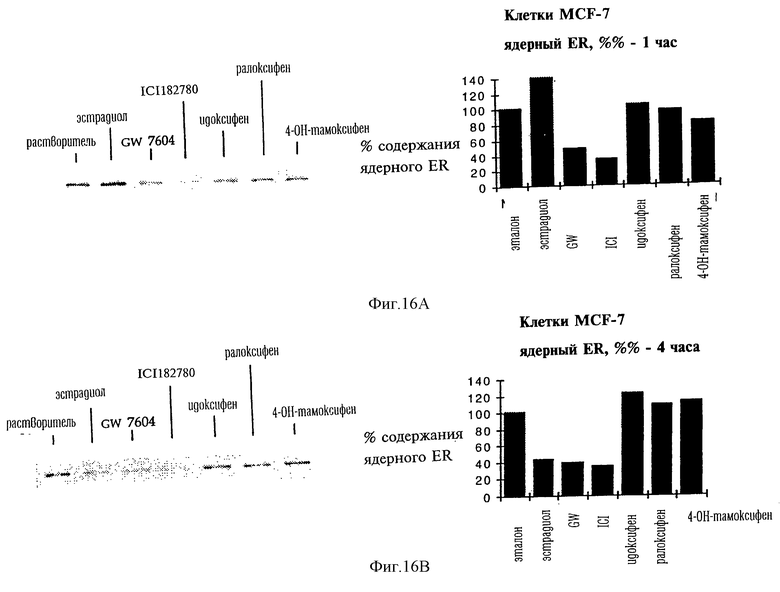
Фигуры 16А и 16В. Анализ методом иммунологического Вестерн-блоттинга эндогенной экспрессии ER в клетках MCF-7 после кратковременной обработки (фиг. 16А - 1 ч; фиг.16В - 4 ч) агонистами или антагонистами.
Фигуры 17А и 17В. Анализ методом иммунологического Вестерн-блоттинга эндогенной экспрессии ER в клетках Ishikawa после кратковременной обработки (фиг.17А - 1 ч; фиг.17В - 4 ч) агонистами или антагонистами.
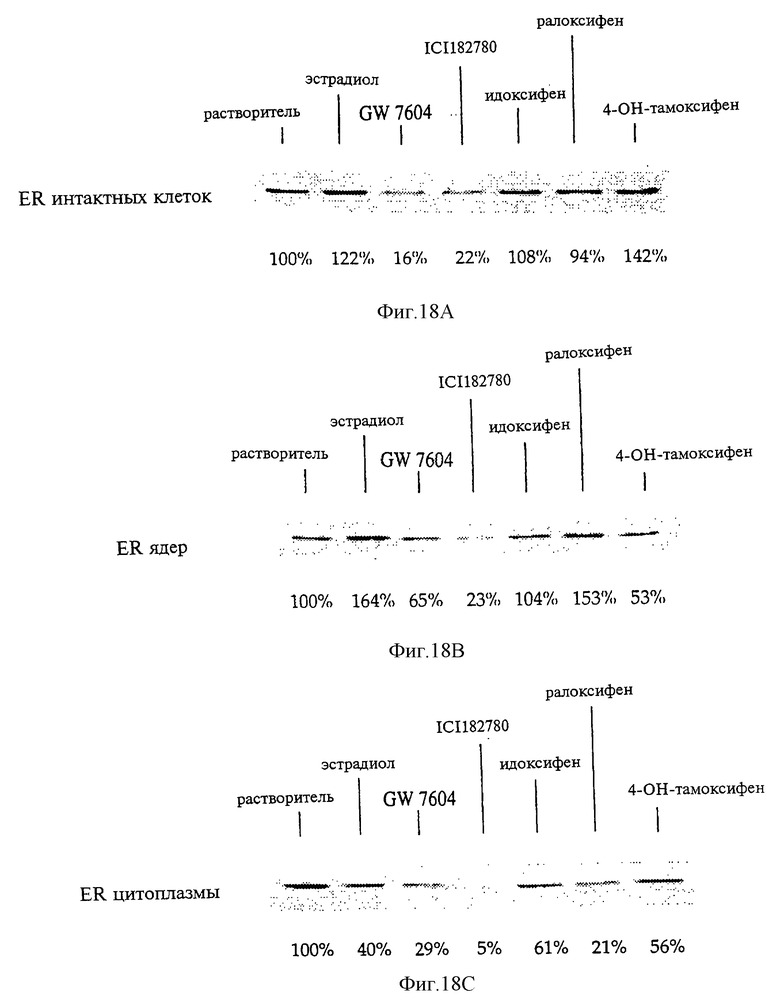
Фигуры 18А-18С. Анализ методом иммунологического Вестерн-блоттинга экспрессии эндогенного ER в интактных клеток (фиг.18А), в ядрах (фиг.18В) и цитоплазме (фиг.18С) клеток Ishikawa после кратковременной обработки агонистами или антагонистами.
Фигуры 19А-19С. Влияние на стимулированную эстрогеном пролиферацию клеток линии MCF-7. Фиг.19А - ICI 182870. Фиг.19В - GW7604. Фиг.19С - 4-ОН-тамоксифен.
Подробное описание изобретения
Настоящее изобретение относится к избирательным модуляторам рецепторов эстрогена, которые проявляют тканеспецифичную активность агонистов ЕR. Эти модуляторы по настоящему изобретению функционируют в качестве агонистов в костной ткани и в сердечно-сосудистой системе, но не в матке. Эти модуляторы отличаются по кинетическим параметрам, например, от тамоксифена и применимы в лечении опухолей, таких как опухоли молочной железы, в частности ER-позитивных вариантов рака молочной железы, характеризующихся возникшей de novo или приобретенной в ходе собственно лечения резистентности к различным модуляторам рецептора эстрогена, включая тамоксифен. Представляемые настоящим изобретением модуляторы также отличаются по кинетическим параметрам от ралоксифена, дролоксифена, идоксифена и ICI182780.
Предпочтительными модуляторами по настоящему изобретению являются производные трифенилэтилена, более предпочтительно соединения формулы I, определенные в патенте США 5681635, при том, что наиболее предпочтительными являются GW5638 и его производные, такие как GW7604. Эти соединения могут быть получены в соответствии с описанным в патенте США 5681835 и описанным Уилсоном с соавт. (Wilson et al., 1994, J. Med. Chem., 37, 1550). Эти модуляторы могут образовывать фармацевтически приемлемые соли с катионами, включая щелочные металлы, такие как натрий и калий, или щелочноземельные металлы, такие как кальций или магний.
Модуляторы по настоящему изобретению могут быть использованы в лечении и (или) профилактике ряда расстройств или состояний, таких как эстроген-стимулируемые злокачественные опухоли, включая рак матки, рак яичников, рак толстой кишки и рак молочной железы, сердечно-сосудистые заболевания (у мужчин и женщин), остеопороз (очаговая деминерализация костей) и артрит. Другие заболевания или состояния, в отношении которых (как для лечения, так и для профилактики) являются применимыми модуляторы по настоящему изобретению, включают рак предстательной железы, бесплодие (например, как фактор индукции овуляции), связанные с климаксом вазомоторные проявления (например, "горячие приливы"), вагинит, доброкачественные пролиферативные заболевания, включая эндометриоз и фиброму матки, сахарный диабет II типа, дегенерация желтого пятна, недержание мочи и болезнь Альцгеймера (познавательная способность). Далее соединения по настоящему изобретению могут быть использованы в качестве контрацептивов для женщин.
Как будет ясно из нижеследующих примеров, GW5638 и его производные являются модуляторами с уникальными кинетическими свойствами. Эти агенты, по-видимому, должны превосходить, например, тамоксифен в качестве исходных терапевтических средств и в качестве химиопрофилактических средств в отношении эстроген-стимулируемых злокачественных опухолей, в частности рака молочной железы, потому что они не обладают утеротрофной активностью. Данные средства обладают классической активностью в отношении ER и, следовательно, можно предположить, что они не будут обусловливать резистентность к себе в той же степени, что и существующие соединения. Более того, данные средства могут быть применены в лечении пациентов, которые характеризуются слабой реагируемостью на другие модуляторы рецептора эстрогенов, включая тамоксифен, идоксифен, ралоксифен и ICI182780, равно как и тех пациентов, которые сначала эффективно на них реагируют, а затем такая реагируемость исчезает. С точки зрения кинетической уникальности представляемых агентов можно ожидать, что их применение не приведет к негативным побочным эффектам, таким как глубокий тромбоз вен.
С учетом уникального механизма действия модуляторов по настоящему изобретению также представляется их использование в качестве компонента терапевтических композиций, в частности предназначенных для лечения рака молочной железы. В этом смысле модуляторы по настоящему изобретению могут быть использованы в сочетании с другим средством, обладающим антиэстрогеновой активностью, с лигандом ретиноевой кислоты или Х-рецептора ретиноевой кислоты, с антипрогестиновым средством, таким как RU486, с антиандрогеном, таким как касдекс или флутамид, с витамином D (или его метаболитами), с ингибитором фарнезилтрансферазы, с агонистом PPAR-α или РРАR-γ или с ингибитором МАР-киназы.
Как указывалось выше, настоящее изобретение представляет использование заявленных модуляторов с целью профилактики, равно как и лечения идентифицированных заболеваний или симптомов. Необходимое для использования количество модулятора может варьироваться в зависимости от состояния (заболевания/расстройства), возраста и собственно состояния пациента и в конечном счете находится в ведении лечащего врача (или ветеринара в случае ветеринарного применения данных препаратов). В целом, однако, дозировки, применимые для лечения взрослого человека, обычно должны находиться в пределах от 0,001 мг/кг до примерно 100 мг/кг в день. Желательная дозировка может быть представлена в виде разовой дозы или в виде фракционированных доз, вводимых через определенные промежутки времени, например дважды, трижды, четырежды или более таких субдозировок в сутки.
Также настоящее изобретение представляет фармацевтические композиции, содержащие определенный выше модулятор или фармацевтически приемлемую его соль, наряду с одним или большим числом фармацевтически приемлемых носителей и, что необязательно, других терапевтических и (или) профилактических компонентов, включая те, которые были описаны выше.
Препараты по настоящему изобретению могут быть введены стандартным путем с целью лечения диагностированных заболеваний или расстройств, например перорально, парентерально, сублингвально, трансдермально, ректально, путем ингаляции или путем трансбуккального введения. Для трансбуккального введения композиции может быть придана форма (например, ее стандартной дозе) таблетки или лепешки, приготовленных стандартными путями. Например, таблетки и капсулы для перорального введения могут содержать традиционные наполнители, такие как связывающие агенты, наполнители, смазывающие вещества, дезинтеграторы и смачивающие агенты. Таблетки могут быть покрыты в соответствии с приемами, хорошо известными в данной области.
С другой стороны, модуляторы по настоящему изобретению могут быть приготовлены в виде жидких препаратов для перорального введения, таких как водные или масляные суспензии, растворы, эмульсии, сиропы или эликсиры. Более того, препараты, содержащие данные модуляторы, могут быть выработаны в виде сухого продукта для последующей реконституции в воде или другом подходящем наполнителе непосредственно перед использованием. Такие жидкие препараты могут содержать стандартные добавки, такие как суспендирующие агенты, эмульсификаторы, консерванты и неводные наполнители.
Такие препараты также могут быть приготовлены в виде суппозиториев, например, таких, которые содержат обычные для суппозиториев основы, например масло какао или другие глицериды. Композиции для ингаляционного введения могут быть стандартным образом приготовлены в виде раствора, суспензии или эмульсии, которые могут быть введены в виде сухого порошка или в форме аэрозоля с использованием стандартного распылительного средства, такого как дихлордифторметан или трихлорфторметан. Типичные препараты для трансдермального введения содержат стандартные водные или неводные наполнители, такие как кремы, мази, лосьоны или пасты, или приготавливаются в форме медицинского пластыря, наклейки или пленки.
Кроме того, композиции по настоящему изобретению могут быть приготовлены для парентерального введения путем инъекций или непрерывных инфузий. Препараты для инъекций могут быть приготовлены в форме суспензий, растворов или эмульсий в масляных или водных наполнителях и могут содержать такие препаративные агенты, как суспендирующие, стабилизирующие и (или) диспергирующие агенты. С другой стороны, активный компонент может быть в форме порошка, предназначенного для разведения подходящим разбавителем (например, стерилизованной, не содержащей пирогенов водой) непосредственно перед применением.
Композиция в соответствии с настоящим изобретением также может быть приготовлена в виде препарата длительного высвобождения. Такие препараты долговременного действия могут быть введены путем имплантирования (например, подкожного или внутримышечного) или путем внутримышечной инъекции. Следовательно, модуляторы по настоящему изобретению могут быть объединены при приготовлении препарата с подходящими полимерными или гидрофобными материалами (например, в виде эмульсии с подходящем масле), ионообменными смолами или в виде, например, умеренно растворимых производных или умеренно растворимых солей.
Идентификация GW5638 и GW7604 в качестве агентов, отклоняющихся по своей активности от классических агонистов, указывает на то, что соединения, которые "активируют" ER (т.е. соединения, которые обусловливают высвобождение ER от генов белков теплового шока) и которые также не обусловливают деградации ER, могут быть использованы при лечении остеопороза. Установление отсутствия разделения остеопоротической и кардиопротективной активностей GW5638 и GW7604 указывает на то, что любое соединение, которое связывается с ER и характеризуется любой из этих активностей (остеопоротической или кардиопротективной), одновременно обладает и другой ("парной" ей) активностью.
Некоторые аспекты настоящего изобретения более подробно будут описаны в нижеследующих примерах, которые не являются в чем-либо ограничивающими настоящее изобретение.
ПРИМЕРЫ
Нижеследующие подробности проведенных экспериментов приведены в конкретных примерах.
Биохимические препараты
ДНК и модифицирующие ферменты были получены от фирм Boehringer Mannheim (Indianapolis, IN), New Englands Biolabs (Beverly, MA) или Promega (Madison, WI). Основные лабораторные реагенты и экстрадиол-17β были закуплены в Sigma (St. Louis, МО). Препарат ICI182780 был безвозмездно предоставлен фирмой Zeneca Pharm. (Macclesfield, Великобритания). Ралоксифен был подарен фирмой Pfizer Pharm. (Groton, CT). 4-ОН-тамоксифен предоставлен фирмой Ligand Pharm. (San Diego, СА). Соединения GW5638 и GW76Q4 были получены в соответствии с ранее описанным (Wilson et al., 1994, J. Med. Chem., 37, 1550-1552). Антитело Н222 получено от Abbott Laboratories.
Культуры клеток и тесты с одновременной трансфекцией
Клетки HepG2 поддерживались в модифицированной культуральной среде Игла (MEM) (Life Technologies, Grand Island, NY) с добавлением 10%-ной плодной телячьей сыворотки (Life Technologies). Клетки высевали в 24-луночные планшеты (покрытые желатином) за 24 ч до трансфекции. ДНК вносили в клетки с помощью липофектиновой технологии (Life Technologies). Вкратце, трансфекция была проведена в трех повторностях с использованием 3 мкг общего количества ДНК. Для стандартной трансфекции 500 нг pCMV-βGal (нормализующий вектор с β-галактозидазным геном), 1500 нг репортерного вектора (вариатив) и 1000 нг рецепторного вектора pRST7-hER (Dana et al. , 1994, Mol. Endocrinol, 8, 1193-1207) использовали в каждой повторности. Инкубацию клеток с липофектином предваряли 3-часовым периодом, в течение которого культуральную среду удаляли, клетки промывали фосфатно-солевым буфером и затем индуцировали подходящим гормоном, разведенным в свободной от фенолового красного среде, содержащей 10% обработанного угольной пылью CS (Cyclone Inc.). Инкубацию с гормоном продолжали в течение 48 ч, после чего клетки лизировали и тестировали на люциферазную и (3-галактозидазную активность в соответствии с ранее описанным (Norris et al., 1995, J. Biol. Chem., 270, 22777-22782).
Анализ утеротрофной активности неполовозрелых крыс
21-дневные самки крыс линии Sprague-Dawley (весом 30-35 г) были получены из Harlan или Taconic Laboratories. Животных случайным образом распределяли по 5 особей в экспериментальные группы, и для каждой такой группы определяли показатель средней массы тела. Вес определяли ежедневно в течение проводимого эксперимента. GW5638 и тамоксифен готовили как раствор в 100%-ном этиловом спирте в виде базового 10-кратного раствора и хранили на холоде (-70oС) до дня непосредственного использования. В день введения препарат разводили в 0,5%-ной метилцеллюлозе, вязкость которой при 2% составила 400 сП при 25oС (т.е. 0,4 н•с/м2) Sigma, St. Louis, МО). Пероральная дозировка с помощью пищевого зонда рассчитывалась, исходя из общего объема 10 мл/кг веса тела. Эстрадиол (Sigma, St. Louis, МО) готовили в кунжутовом масле, размешивали с помощью стеклянного гомогенизатора (в растворенном виде, либо в суспензии), разделяли на аликвоты и замораживали при -70oС до применения. Дозировки для подкожного введения рассчитывали, исходя из общего количества 2 мл/кг веса тела. Скармливание (GW5638) или инъецирование (эстрадиол) животных осуществляли на протяжении 3 дней. На 4-й день животных умерщвляли путем углекислотной асфиксии, измеряли массу тела, выделяли матку, подсушивали ее промоканием и взвешивали. Данные представляли как соотношение веса матки и веса тела.
Анализ степени минерализации костей
Подготовка животных. Крыс линии Sprague-Dawley в возрасте 90 дней обездвиживали с помощью изофторана (4% - индукция анестезии, 2% - ее поддержание), подвергали овариэктомии (OVX) или оперировали симулятивно (выполняли одинаковые манипуляции за исключением удаления яичников) с последующим распределением случайным образом по группам (n=7), а затем проводили обработку с 1-го по 28-й день после операции путем перорального введения только наполнителя, эстрадиола или GW5638 в 0,5%-ной метилцеллюлозе. Животных умерщвляли путем углекислотной асфиксии, определяли вес тела, удаляли и взвешивали матку. Матку, влагалище и ткань молочной железы фиксировали в забуференном (рН 7) 10%-ном формалине. Пробы для гистологического анализа брали из средней части каждого из рогов матки. Тканевые образцы заливали в парафин, окрашивали гематоксилином и эозином и анализировали под микроскопом. Выделяли поясничные позвонки, а также левую и правую большеберцовые кости. Определяли общий уровень холестерина в крови (Roche Вiomed. Laboratories).
Двухуровневая рентгеновская абсорбциометрия (DEXA). Для анализа методом DEXA использовали денситометр костей Hologic QDR-2000, оснащенный высокоинформативным программным пакетом. По умолчанию длина полосы сканирования, ее ширина, интервал и разрешение были установлены на величинах 2, 0,75, 0,01 и 0,005 дюймов соответственно. Денситометр калибровали ежедневно с использованием гидроксиапатитовой позвонковой платформы. Выделенные большеберцовые кости помещали в слой воды в 1 см так, чтобы большеберцовая и малоберцовая кости располагались в нем горизонтально. Для прижизненного тестирования крыс обездвиживали с помощью изофторана и помещали в положении "на спине" так, чтобы позвоночник оказывался параллельным длинной оси столика денситометра. Головку сканера закрепляли в положении, параллельном длинной оси столика, и проводили сканирование большеберцовой кости до места ее соединения с бедренной костью. Представляющий интерес участок большеберцовой кости анализировали с подключением программы субсегментного тестирования, фокусируя сканер на участке шириной 2 мм в 3 ми дистальнее участка нарастания.
Периферическая количественная компьютерная томография (PQCT). Компьютерную томографию проводили на установке PQCT (ХСТ-960А, Norland). Сегменты по четыре-пять миллиметров сканировали при размере воксела (объемный элемент изображения), равном Е (0,148 мм), и при шаге 0,5 мм. Сегмент размером 3-5 мм, дистальный по отношении к участку нарастания, был проанализирован с использованием режима "contmode, 2/peelmode, 5/cortmode". Были получены данные замеров по степени общей, трабекулярной и кортикальной минерализации костей. Вырезанные большеберцовые кости помещали в слой воды в 1 см таким образом, чтобы большеберцовая и малоберцовая кости располагались горизонтально, что обеспечивало гарантию вертикального сканирования костей. Крыс обездвиживали изофтораном и головку сканера располагали таким образом, чтобы изображения мест сочленения большеберцовой и бедренной костей, а также большеберцовой и малоберцовой костей могли быть точно визуализованы и использованы в качестве маркеров при проведении компьютерной томографии.
Пример 1
Идентификация новых модуляторов ER
Была разработана серия тестов in vitro, с помощью которых модуляторы ER были классифицированы в четыре кинетически дифференцированные группы (Tzukerman et al. , 1994, Mol. Endocrinol, 8, 21-30). Конкретно был разработан тест с печеночными (гепатокарциномными) клетками HepG2, в котором способность соединения регулировать транскрипционную активность промотора (С3) гена 3-го компонента комплемента, связывающегося с эстрогеном, оценивали в присутствие либо ER дикого типа (ERwt), либо мутантного рецептора (ER-TAF1), в котором функции домена AF-2 были искусственно нарушены. С использованием этих тестов явилось возможным сформировать "фингерпринты" известных модуляторов ER (McDonnell et al., 1995, Mol. Endocrino1, 9, 659-669). Хотя данные тесты не воспроизводят с полной точностью нативную среду, характерную для ER in vivo, анализ соединений в данных тестах является вполне достаточным для разделения их по группам, каждая из которых проявляет уникальный набор активностей in vivo.
Была синтезирована серия трифенилэтиленовых производных лигандов ER (Willson et а1., 1994, J. Med. Chem., 37, 1550-1552). Предварительный анализ этих соединений in vivo показал, что относительные активности этих соединений в костной ткани и в матке не являются идентичными, что, по-видимому, отражает возможные кинетические различия (Willson et al., 1994, J. Med. Chem. , 37, 1550-1552). Затем слепой анализ этих соединений был проведен в отношении рецептора дикого типа в клетках HepG2 по промотору С3, в котором было установлено, что все соединения (за исключением двух из них) по кинетическим параметрам не отличались от тамоксифена. Однако еще два соединения - GW5638 и GW7604 - продемонстрировали в существенной степени отличающиеся параметры в данной системе по сравнению с другими лигандами ER, а это явилось обоснованием для проведения дальнейших анализов. Интересно, что эти соединения по своей структуре идентичны друг другу, за исключением того, что GW7604 является 4-гидроксилированным вариантом GW5638 (табл.1). С использованием теста in vitro на конкурентное с изотопно помеченным лигандом связывание для обоих этих соединений было показано проявление высокого уровня аффинности по взаимодействий с ER. Конкретно GW5638 и GW7604 характеризовались значениями Ki 50,4 нМ (±5,4) и 15,5 нм (±1,4) соответственно. В тех же самых условиях эстрадиол-17β, как было показано, характеризуется величиной Ki на уровне 6,3 нМ (±0,4). Хотя метаболизм GW5638 не был изучен, вероятным является то, что он конвертируется в характеризующееся большей аффинностью соединение GW7604 in vivo согласно тому же механизму, по какому тамоксифен конвертируется в метаболит 4-ОН-тамоксифен, характеризующийся большей аффинностью (Jordan et al., 1977, J. Endocrinol, 75, 305-316). Сравнение агонистической активности этих соединений по отношению к представителям каждой из четырех идентифицированных групп ЕR-лигандов показано на фиг.1А. В данном тесте тамоксифен действует в качестве неполного агониста ER тогда, когда он анализируется по работе промотора С3, достигая активности на уровне 45% от таковой характеристики эстрадиола. При тестировании тем же самым способом ралоксифен и чистый антагонист ICT182789 не демонстрируют агонистической активности, но подавляют основной уровень транскрипционной активности с промотора гена С3. Недавно было установлено, что основная (конститутивная) активность промотора С3 зависит от ER, хотя и не зависит от его лиганда (Norris et al., 1996, Mol. Endocrinol, 10, 1605-1616). Поскольку и ралоксифен и ICI182780 подавляют лигандзависимую и лиганднезависимую активацию ER, они в такой клеточной среде, по видимому, действуют как "инвертивные агонисты". Однако и GS5638 и его возможный метаболит GW7604 не проявляют какой-либо агонистической или антагонистической активности по отношению к данному промотору, характеризуясь неразличающимися "фингерпринтами". Было сделано заключение, что в среде, в которой тамоксифен проявляет частичную активность агониста, в свою очередь, GW5638 и GW7604, являющиеся аналогами тамоксифена, оказываются функционально неактивными.
Хотя ралоксифен и ICI182780 проявляют себя в отношении рецептора эстрогенов дикого типа (ERwt) аналогичным образом, они различаются по кинетическим параметрам (McDonnell et al., 1995, Mol. Endocrinol, 9, 659-669; Dauvois et al., 1992, Proc. Natl. Acad. Sci. USA, 89, 4037-4041; Dauvois et al., 1993, J. Cell. Sci., 106, 1377-1388). При тестировании в отношении мутантного рецептора эстрогена (ER-TAF1), который характеризуется разрушением последовательности активационного домена AF-2, ралоксифен ведет себя так же, как и тамоксифен, проявляя активность на уровне 40% от агонистической активности эстрадиол (фиг.1В). В этом же тесте ICI182780, GW5638 и GW7604 оказываются функционально неактивными. Эти данные указывают на то, что GW5638 (и GW7604) функционирует по механизму, который отличается от ранее идентифицированных типов, смешанных по своему действию на ER агонистов и антагонистов (McDonnell et al., 1995, Mol. Endocrinol, 9, 659-669).
Одним из возможных, хотя и маловероятных объяснений этих исходных результатов, является то, что эти соединения метаболизируются (или иными путями характеризуются защищенностью от связывания с рецептором), что тем самым может объяснять их пассивность в данном тесте. Данный эксперимент был
направлен на оценку способности GW5638 и GW7604 подавлять агонистическую активность эстрадиола и тамоксифена, проявляемую в отношении вариантов ERwt и ER-TAF1 соответственно, и переключать инвертивную агонистическую активность ICI182780. Как показано на фиг.2А, эстрадиол функционировал в качестве полного агониста, а тамоксифен функционировал как частичный (неполный) агонист в отношении ERwt в ходе тестирования в отношении активности промотора гена С3 в клетках HepG2. Важным является то, что агонистическая активность, проявляемая тамоксифеном или эстрадиолом, подавлялась обоими соединениями - GW7604 и GW5638. Таким образом, эти соединения функционировали в качестве антагонистов в отношении рецептора по механизму, отличающемуся от механизма действия тамоксифена. Сходный анализ был проведен с использованием мутантного варианта ER-TAF1 вместо рецептора дикого типа (фиг.2В). Как и ожидалось, и GW5638 и GW7604 были способны подавлять индуцируемую эстрадиолом и тамоксифеном транскрипционную активность с участием ER-TAF1. Интересно, что ралоксифен подавляет неполную агонистическую активность ER-TAF1 (фиг. 1В), т.е. активность, которая подавляется обоими соединениями - GW7604 и GW5638. Обобщение полученных данных указывает на то, что GW5638 и его предполагаемый, образующийся in vivo метаболит GW7604 являются по кинетическим параметрам уникальными модуляторами ER, которые не проявляют агонистическую активность in vitro, но которые способны подавлять агонистическую активность эстрадиола, тамоксифена и ралоксифена. Хотя их параметры по данным некоторых тестов сходны с таковыми, характерными для лигандов, относимых к типу "чистых" антагонистов, тем не менее эти соединения отличаются от антагонистов-стероидов, таких как ICI182780, потому что они не проявляют активность инвертивных агонистов (фиг.1А).
С целью подтверждения того, что GW5638 и GW7604 отличаются по кинетическим параметрам от ICI 182780, проводили определение способности этих соединений переключать инвертивную агонистическую активность, проявляемую соединением ICI 182780. Результаты этого анализа показаны на фиг.2С. Конкретно было установлено, что основная активность промотора гена С3 человека подавлялась на порядок при добавлении ICI182780 и что этот эффект полностью может быть обращен одновременным добавлением GW7604 и по крайней мере отчасти обращен добавлением GW5638.
Одним из возможных объяснений выявленных кинетических различий является то, что GW5638 и GW7604 взаимодействуют с ER и подавляют способность этого рецептора взаимодействовать с ДНК. Это было установлено с использованием модифицированного ER (ER-VP16) с целью выяснения внутриядерной локализации и параметров связывания ER с ДНК в клетке после связывания с лигандом. Такой модифицированный белок во всех отношениях проявляет себя как и ER дикого типа, за исключением того, что он активирует транскрипцию путем взаимодействия участком с промотора, обозначаемым как ERE (т.е. сайт регулирования эстрогенами), вне зависимости от природы связанного на ER лиганда (McDonnell et al. , 1995, Mol. Endocrinol, 9, 659-669). С использованием этого реагента было показано, что все типы лигандов ЕВ, включая ICI182780, GW5638 и GW7604, обусловливают эффективное взаимодействие ER с ДНК-мишенью (McDonnell et a1., 1995, Mol. Endocrinol, 9, 659-669).
Таким образом, GW5638 и GW7604 взаимодействуют с ER in vivo и характеризуются такими фармакологическими свойствами, которые отличаются от таковых, известных у других модуляторов ER. Следовательно, это значит, что уникальные свойства GW5638 и GW7604 проявляются на некотором этапе после связывания их лиганда на ДНК. С учетом уникальных свойств этих соединений была проведена серия экспериментов по выяснению на моделях интактных животных активности этих соединений в отношении скелетной, сердечно-сосудистой и воспроизводительной систем.
Пример 2
Предотвращение потерь костной ткани у крыс, прошедших овариэктомию
Имеется достоверное доказательство того, что и тамоксифен и ралоксифен предотвращают уменьшение массы костной ткани, полученное в преклинических моделях постклимактерического остеопороза (Love et a1., New England J. Med., 326, 852-856; Love et al., 1991, Ann. Intern. Med., 115, 860-864; Black et al. , 1994, J. Clin. Invest., 93, 63-69). Однако механизм действия этих соединений в отношении костной ткани ранее не был определен. Судьба костной ткани у пациентов, в лечении которых используется препарат ICI182780, к настоящему времени точно не установлена, хотя данные, полученные на преклинических крысиных моделях, говорят о том, что этот препарат не является агонистом в костной ткани (Gallagher et al., 1993, Endocrinology, 133, 2787-2791). Это позволяет выдвинуть гипотезу, в соответствии с которой частичная агонистическая активность тамоксифена и ралоксифена является ключевой для защиты костной ткани (Love et a1., 1992, New England J. Med., 326, 852-856; Black et al. , 1994, J. Clin. Invest., 93, 63-69). В пользу этой гипотезы говорят данные ранее проведенных исследований, показавших, что и тамоксифен и ралоксифен могут функционировать как в равной степени эффективные агонисты в некоторых типах клеток и в ряде контекстов нуклеотидных последовательностей промоторов (McDonnell et al., 1995, Mol. Endocrinol., 9, 659-669). Однако GW5638 представляет новый "путь обращения к этой ткани". Данное соединение, которое не проявляет агонистической активности в классическом ее понимании в любом из тестов, проводимых in vitro, было протестировано по его способности подавлять уменьшение массы костной ткани у овариэктомизированных крыс. Конкретно степень минерализации костной ткани (СМК) в поясничном отделе позвоночника и в большеберцовой кости у 90-дневных крыс с удаленными яичниками оценивали после перорального введения в течение 28 дней либо эстрадиола-17β, либо повышающихся доз GW5638. Полученные результаты представлены на фиг.3А: они показывают, что существенное уменьшение массы костной ткани имеет место в поясничных позвонках у овариэктомизированных крыс (OVX) в течение 28-дневного курса по сравнению с контрольными животными, которым проводили симулятивную операцию, в то время как у экспериментальных особей OVX, которым был введен эстрадиол, СМК оставалась стабильной. По существу GW5638 продемонстрировал зависимую от дозы активность по защите костной ткани, достигая той же эффективности, что и эстрадиол, в концентрации 3 мкмоль/кг (1 мг/мл). Этот показатель близок к дозе тамоксифена, необходимой для защиты костной ткани в той же модели (Love et al., 1992, New England J. Med. , 326, 852-856; Black et al., 1994, J. Clin. Invest., 93, 63-69; Yang et al., 1996, Endocrinology, 137, 2075-2084). Обнаруженная активность по защите костной ткани не ограничивается поясничным отделом позвоночника, потому что аналогичные результаты были получены в анализе СМК в большеберцовой кости (фиг.3В). Конкретно при использовании того же самого экспериментального протокола было продемонстрировано, что GW5638 был эффективен с точки зрения поддержания общего веса костной ткани с весьма долговременным эффектом в отношении трабекулярного компартмента костей. Это представлялось интересным в свете того факта, что, как ранее было показано, эстроген в этом же компартменте обусловливает регуляцию динамического преобразования костной ткани (Gallagher et al., 1993, Endocrinology, 133, 2787-2791). При обобщении эти данные указывают на то, что GW5638, как соединение, не проявляющее ER-агонистической активности классического типа, при тестировании in vitro функционирует в качестве эффективного агониста ER в костной ткани.
Было показано, что соединения, функционирующие в качестве агонистов ER в костной ткани, такие как эстрадиол, тамоксифен и ралоксифен, также могут подавлять возрастание уровня сывороточного холестерина - эффекта, ассоциированного с удалением яичников (Love et al., 1992, Ann. Intern. Med., 115, 860-864; Black et al., 1994, J. С1in. Invest., 93, 63-69). Это наблюдение позволило предположить, что механизмы активности ER в костной ткани и в сердечно-сосудистой системе в значительной степени сходны. Хотя пока неясно, является ли выявленное подавление уровней холестерина в сыворотке достаточным для объяснения снижения уровня смертности от сердечно-сосудистых заболеваний у женщин в постклимактерический период в ходе терапии с вытеснением эстрогена, тем не менее этот эффект рассматривается в качестве маркера активности эстрогена в сердечно-сосудистой системе. С целью оценки этого эффекта была проведена оценка уровня сывороточного холестерина у овариэктомизированных крыс, которым в течение 28 дней вводили эстрадиол или GW5638. Данные, приведенные на фиг.4, указывают на то, что даже при наименьшей тестированной концентрации GW5638 оказывается столь же эффективным, что и эстрадиол, с точки зрения снижения содержания сывороточного холестерина.
Пример 3
GW5638 как утеропротективный ЕR-модулятор
Продолжая изучение тканеспецифичного характера действия GW5638, был проведен сравнительный анализ утеротрофической активности этого соединения. В исходной серии экспериментов проводили сравнение активности GW5638 и тамоксифена в матках у 21-дневных неполовозрелых крысят. В этом тесте в качестве показателя ER-агонистической активности, характерной для данной ткани, использовали сырой вес матки (фиг.5). При пероральном введении в качестве единственного агента GW5638 не проявлял сколько-нибудь существенной активности по сравнению с контролем. В частности, отмечается, что данное соединение в проведенном тесте неактивно при дозе 10 мкмоль/кг в день, что втрое выше дозы, необходимой для достижения эффекта защиты костной ткани (фиг. 3). С другой стороны, для тамоксифена выявляется уротрофическая активность (т.е. индукция разрастания матки) при дозах не более 0,1 мкмоль/кг в день. Эти исследования были продолжены с целью выяснения способности GW5638 (в отличие от тамоксифена) полностью подавлять агонистическую активность эстрадиола у этих крыс - это бы явилось подтверждением того, что данное соединение является полным антагонистом в данной ткани, по крайней мере в условиях проведенного теста.
Во второй серии экспериментов утеротрофическая активность после 28-дневного введения либо GW5638, либо эстрадиола была оценена у 90-дневных крыс с удаленными яичниками (OVX). Полученные в этом анализе результаты, приведенные на фиг.6А, указывают на то, что при дозах, которые втрое превышают дозы, необходимые для достижения эффекта защиты костной ткани, GW5638 проявляет минимальную активность по индукции разрастания ткани матки. Важным, однако, является то, что не было выявлено сколько-нибудь существенных различий в общей массе тела у овариэктомизированных крыс, которым вводили GW5638, по сравнению с особями, прошедшими симулятивную операцию. Было выявлено очень незначительное, не зависящее от дозы увеличение сырого веса матки по отношению к крысам OVX. Это соответствует данным, которые приводятся другими исследователями, анализировавшими крыс после введения ралоксифена, когда подобная активность объяснялась повышением впитыванием воды (Kedar et al., 1994, Lancet, 343, 1318-1321; Love et al., 1992, Ann. Intern. Med., 115, 860-864; Black et al., 1994, J. Clin. Invest., 93, 63-69).
В дополнение к измерениям сырого веса матки было проведено гистологическое тестирование матки, взятой у тех же животных - фиг.7A-7F (низкое увеличение) и фиг. 8A-8D (большое увеличение). В этом анализе клетки эпителия матки у крыс, которым вводили GW5638, характеризовались зависимой от дозы соединения гипертрофией, в то время как в строме наблюдалось слабое увеличение внеклеточной соединительной ткани и основного вещества. При наивысших дозах GW5638 (втрое более высоких, чем доза, необходимая для защиты костной ткани), выявленная гипертрофия эпителия сравнима с таковой, которая наблюдается в матке крыс, которым водили эстрадиол, в то время как реакция стромы и уровень инфильтрации эозинофилов были меньшими по сравнению с теми же показателями, выявляемыми при введении крысам эстрадиола (сравнить фиг.8С и 8D). При обобщении эти данные указывают на то, что GW5638 проявляет слабую ER-агонистическую активность в матке, в то время как в костной ткани это соединение действует как отчетливый агонист ER. Таким образом, GW5638 является уникальным модулятором ER, который проявляет активность агониста или антагониста ER по тканеспецифичному типу.
Пример 4
Влияние антиэстрогеновой обработки на опухоли молочной железы у мышей линии "nude"
Данный анализ был осуществлен с использованием опухолевых клеток, производных от клеточной линии MCF-7 рака молочной железы. Эта линия, позитивная по рецепторам эстрогена и прогестерона, является гормональнозависимой и чувствительна к антигормональным воздействиям. Опухолевые клетки использовали для заражения (через бока тела) мышей, гомозиготных по мутации nu/nu (бестимусные мыши, производные от линии BALB/c-Urd), подвергнутых овариэктомии. Мышам имплантировали эстрогенсодержащие гранулы медленной секреции. Ежедневно животным проводили подкожные инъекции в соответствии с приведенной ниже схемой:
Группа 1: контроль (кукурузное масло)
Группа 2: 0,3 мг GW5638
Группа 3; 0,6 мг GW5638
Группа 4: 1,0 мг GW5638
Группа 5: 1,0 мг тамоксифена
С использованием циркуля проводили измерение размеров опухолей в 2 направлениях, определяя поверхность опухоли по формуле 1/2•w/2•π. Полученные результаты показаны на фиг.9.
Пример 5
Анализ зависимости от дозы
Целью данного эксперимента явилось сравнение максимально эффективной дозы GW5638 с таковой у тамоксифена в связи со способностью подавлять рост раковых опухолей молочной железы MCF-7 у мышей линии "nude".
Десять мышей-доноров OVX были проинъецированы 5 миллионами клеток MCF-7. Развившиеся в результате опухоли трансплантировали мышам-реципиентам. Всем животным имплантировали капсулы с медленно секретируемым эстрадиолом.
Начиная с момента, когда развивающаяся опухоль становится различимой на уровне проводимых измерений, осуществляли ежедневное введение по 0,1 мг путем подкожных инъекций в соответствии со следующей схемой:
Группа 1: контроль (кукурузное масло)
Группа 2: 0,3 мг GW5638
Группа 3: 0,6 мг GW5638
Группа 4: 1,0 мг GW5638
Группа 5: 1,0 мг тамоксифена
Размер опухолей определяли с помощью циркуля, рассчитывая поверхность опухоли по следующей формуле: поверхность = 1/2•w/2•π.
Спустя 8 недель проводили межгрупповое сравнение роста опухолей. Во всех группах, которым вводили препарат, было отмечено подавление роста опухолей (на уровне статистической значимости) по сравнению с контролем. Две высшие дозы GW5638 - 0,6 мг и 1,0 мг - не отличались по своему проявлению в тест-признаке от дозы 1,0 мг тамоксифена (см. фиг.10).
Пример 6
Анализ LСС2
Линия MCF-7/LCC2 является линией клеток (полученной из Онкологического Центра Ломбарди - LCC), которая характеризуется независимостью от эстрогена, хотя и чувствительна к нему, а также является резистентной к тамоксифену. Данный эксперимент был проведен с целью определения способности GW5638 задерживать рост данной линии клеток у мышей линии "nude" по сравнению с контрольными или подвергнутыми воздействию тамоксифена опухолями.
Сорок овариэктомизированных мышей были подготовлены для внесения им этих клеток и разделены на группы следующим образом:
Группа 1: контроль
Группа 2: имплантат, секретирующий эстроген
Группа 3: 1,0 мг тамоксифена
Группа 4: 1,0 мг соединения GW5638
Контрольные животные ничего не получали, а животным групп 3 и 4 каждые 3 дня проводили инъекции по 0,1 мл кукурузного масла. Каждые три дня проводили измерение опухолей с помощью циркуля, а величину их поверхности подсчитывали по следующей формуле: поверхность = 1/2•w/2•π.
Спустя 8 недель проводили межгрупповое сравнение роста опухолей. Против ожидаемого опухоли не проявили восприимчивости к эстрогену. Кроме того, опухоли проявили чувствительность к тамоксифену, несмотря на их предварительно предсказывавшуюся резистентность к этому соединению. И тамоксифен, и GW5638 были способны в равной степени подавлять рост данного вида опухолей (см. фиг.11).
Пример 7
Функционирование GW7604 в качестве антиэстрогена в клетках MCF-7 опухоли молочной железы
Клетки MCF-7 раковой опухоли молочной железы человека были котрансфицированы по непостоянному типу с 0,9 мкг/мл экспрессирующего ER человека вектором наряду с 2 мкг/мл репортерной конструкции С3-Luc и 0,1 мкг/мл вектора pRSV-βGal, экспрессирующего β-галактозидазу (является контрольным показателем эффективности трансфекции). После трансфекции клетки инкубировали в течение 48 ч в присутствие эстрадиола-17β и повышающихся концентраций каждого из антагонистов в соответствии с указанным. Затем трансфицированные клетки тестировали на активность люциферазы и β-галактозидазы. Нормализованную активность люциферазы рассчитывали путем соотнесения в каждой точке уровня люциферазы (•104 ед.) с активностью β-галактозидазы [(А415•105)/время в мин] . В соответствии с изображенным на фиг.12 каждая точка в собранных данных представляет собой среднее по трем повторностям эксперимента значение транскрипционной активности, имеющей место в данных экспериментальных условиях. Средний коэффициент вариации для каждой из тестированных концентраций гормона был менее 10%.
Пример 8
Анализ влияния отдельных модуляторов ЕВ на фармакологические параметры антиэстрогеновых факторов как подтверждение дополнительной кинетической комплексности
Клетки гепатокарциномы человека HepG2 были по непостоянному типу трансфицированы с 0,9 мкг/мл вектора, экспрессирующего либо ER человека (pBST7ER) (см. фиг. 13А), либо мутантный ER (ER-TAF1) (см. фиг.13В), в котором была инактивирована функция домена AF-2 (ER-TAF1), наряду с 2 мкг/мл конструкции, включающей промотор гена, кодирующего 3-й компонент комплемента (С3), регулируемого эстрогенами, вместе с репортерным геном люциферазы, а также наряду с 0,1 мкг/мл вектора pRSV-β-Gal, экспрессирующего β-галактозидазу (является контрольным показателем эффективности трансфекции). После трансфекции клетки инкубировали в течение 48 часов в присутствие одного растворителя или повышающихся концентраций эстрадиола или антиэстрогенов в соответствии с указанным. После этого трансфицированные клетки тестировали на активность люциферазы и β-галактозидазы. Каждая точка в собранных данных представляет собой среднее по трем повторностям эксперимента значение транскрипционной активности, имеющей место в данных экспериментальных условиях. Средний коэффициент вариации для каждой из тестированных концентраций гормона был менее 10%. Данные, представленные на фиг.13А и 13В, указывают на то, что большинство из известных антиэстрогеновых агентов проявляют активность агонистов в отношении мутантных рецепторов эстрогена. Тот факт, что GW7604 не проявляет агонистической активности в отношении какого-либо из тестированных мутантных ER, тем самым указывает на то, что данное соединение применимо для целей лечения случаев рака молочной железы, резистентных к тамоксифену.
Пример 9
Сравнительный анализ способности различных антиэстрогенов подавлять транскрипционную активность ERα и ERβ
Клетки HeLa человека были трансфицированы либо вектором, экспрессирующим ERα (см. фиг.14А), либо вектором, экспрессирующим ERβ (cм. фиг.14В), наряду с репортерной конструкцией "ERE-TK-люцифераза", регулируемой по промоторной последовательности (ERE) с участием эстрогенов. После этого оценивали способность различных концентраций антагониста подавлять активируемую эстрадиолом (10-9) транскрипцию. Результаты эксперимента, представленные на фиг. 14А и 14В, указывают на то, что за исключением идоксифена все тестированные антиэстрогеновые соединения проявляют примерно равную активность в отношении ERα, в то время как в отношении ERβ ни ралоксифен, ни идоксифен не являются потенциальными антагонистами. Более того, полученные данные указывают на то, что GW7604 является потенциальным "пан-антагонистом" в отношении обеих изоформ рецептора эстрогена человека.
Пример 10
Анализ методом Вестерн-иммуноблоттинга экспрессии ER в клетках-мишенях после воздействия агонистов или антагонистов
Выбранные клеточные линии инкубировали в течение 48 ч в присутствии только растворителя или 10 нМ эстрадиола или антиэстрогена в соответствии с указанным. Затем приготавливали ядерные экстракты и полученные образцы разделяли с помощью электрофореза в денатурирующем ПААГ, переносили на нейлоновую мембрану и оценивали относительную экспрессию ER после проведения тестированных воздействий с помощью метода иммунологического Вестерн-блоттинга с использованием моноклонального антитела Н222, специфичного в отношении рецептора эстрогенов. На фиг.15А показано содержание эндогенного ядерного ER в клетках MCF-7 (10 мкг на 1 дорожку). На фиг.15В показано содержание эндогенного ядерного ER в клетках Ishikawa (100 мкг на 1 дорожку). На фиг.15С охарактеризованы клетки линии Ishikawa, трансфицированные по непостоянному типу 0,9 мкг/мл вектора pRST7ER: для детекции было взято по 10 мкг ядерного экстракта на 1 дорожку. Определение количественного содержания ER проводили с помощью денситометрии полученных иммуноблоттов. Полученные результаты, представленные на фиг. 15А-15С, соответствуют серии экспериментов, проведенных в одних и тех же условиях.
Пример 11
Анализ методом Вестерн-имммуноблоттинга экспрессии эндогенного ER в клетках MCF-7 после кратковременного воздействия агонистов или антагонистов
Клетки MCF-7 раковой опухоли молочной железы человека были проинкубированы в течение 1 ч (фиг.16А) или 4 ч (фиг.16В) в присутствии только растворителя или 10 нМ эстрадиола или антиэстрогена в соответствии с указанным. Были приготовлены ядерные экстракты, а полученные образцы разделяли с помощью электрофореза в денатурирующем ПААГ, переносили на нейлоновую мембрану и оценивали относительную экспрессию ER после проведения тестированных воздействий с помощью метода иммунологического Вестерн-блоттинга с использованием моноклонального антитела Н222, специфичного в отношении рецептора эстрогенов. Определение количественного содержания ER проводили с помощью денситометрии полученных иммуноблотов. Полученные результаты, представленные на фиг. 16А и 16В, соответствуют серии экспериментов, проведенных в одних и тех же условиях.
Пример 12
Анализ методом Вестерн-иммуноблоттинга экспрессии эндогенного ER в клетках линии Ishikawa (культивируемые клетки матки) после кратковременного воздействия агонистов или антагонистов
Клетки Ishikawa аденокарциномы эндометрия человека инкубировали в течение 1 ч (фиг.17А) или 4 ч (фиг.17В) в присутствии только растворителя или 10 нМ эстрадиола или антиэстрогена в соответствии с указанным. Были приготовлены ядерные экстракты, а полученные образцы разделяли с помощью электрофореза в денатурирующем ПААГ, переносили на нейлоновую мембрану и оценивали относительную экспрессию ER после проведения тестированных воздействий с помощью метода иммунологического Вестерн-блоттинга с использованием моноклонального антитела Н222, специфичного в отношении рецептора эстрогенов. Определение количественного содержания ER проводили с помощью денситометрии полученных иммуноблотов. Полученные результаты, представленные на фиг.17А и 17В, соответствуют серии экспериментов, проведенных в одних и тех же условиях.
Пример 13
Анализ методом Вестeрн-иммуноблоттинга экспрессии эндогенного ЕR в интактных клетках, в ядрах и цитоплазме клеток Ishikawa после кратковременного воздействия агонистов или антагонистов
Клетки Ishikawa аденокарциномы эндометрия человека инкубировали в течение 3 ч в присутствии только растворителя или 10 нМ эстрадиола или антиэстрогена. Были приготовлены интактные клетки (фиг.18А), ядерные (фиг.18В) и цитоплазматические (фиг.18С) экстракты, а полученные образцы разделяли с помощью электрофореза в денатурирующем ПААГ, переносили на нейлоновую мембрану и оценивали относительную экспрессию ER после проведения тестированных воздействий с помощью метода иммунологического Вестерн-блоттинга с использованием моноклонального антитела Н222, специфичного в отношении рецептора эстрогенов. Определение количественного содержания ER проводили с помощью денситометрии полученных иммуноблотов. Полученные результаты, представленные на фиг.18А-18С, соответствуют серии экспериментов, проведенных в одних и тех же условиях.
Пример 14
Подавление соединением GW7604 стимулированной эстрогеном пролиферации клеток МСF-7
Целью данного эксперимента было определение способности GW5638 подавлять стимулированную эстрогенами пролиферацию клеток линии MCF-7 опухоли молочной железы in vivo.
Экспериментальный протокол: высев по 25-50 тысяч клеток в расчете на 1 лунку 24-луночного планшета. Клетки высевали в культуральную среду, свободную от фенолового красного. После присоединения клетки стимулируют либо только антиэстрогеном, либо эстрогеном и антиэстрогеном. Время индукции варьировалось в пределах 12-48 ч, в зависимости от конкретного эксперимента.
Добавление 4λ (4 мкКи) тимидина, [метил-3Н] в каждую лунку.
Инкубация в течение 2-4 ч при 37С.
Отбор среды с помощью аспирации и двукратная промывка охлажденным на льду фосфатно-солевым буфером.
Однократная промывка охлажденной на льду 10%-ной трихлоруксусной кислотой (ТХУК).
Добавление 2 мл 10%-ной ТХУК в каждую лунку.
Инкубация в течение 1-2 ч при 4oС.
Однократная промывка в ТХУК.
Добавление 1 мл 0/2 N NаОН.
Перенос содержимого каждой лунки в сцинтилляционный счетчик, содержащий 2 мл сцинцилляционной жидкости.
Перемешивание и подсчет [3Н].
Полученные результаты должны показать способность различных соединений подавлять индуцированную эстрогеном и основную (т.е. конститутивную) пролиферацию клеток MCF-7 (см. фиг.19А-19С).
Все цитировавшиеся выше материалы включены в данный текст для сведения в виде библиографических ссылок.
Для ознакомившегося с представленной заявкой специалиста в данной области техники должно быть ясно, что различные изменения в общей форме и отдельных деталях могут быть осуществлены без отклонения от реального масштаба настоящего изобретения.















Изобретение относится к медицине и может быть использовано для лечения эстрогензависимых злокачественных опухолей. Способ представляет собой введение указанному млекопитающему профилактически или терапевтически эффективного количества соединения формулы 1. Предложенный способ позволяет повысить эффективность лечения за счет преодоления резистентности злокачественной опухоли применением антиэстрогенных препаратов 2 с. и 11 з.п.ф-лы, 19 ил.
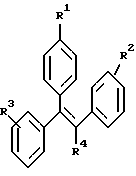
где R1 представлен - (СН2)n CR5= CR6R7, -(СН2)mC(X)NR8R9 или
R2 и R3 независимо представлены Н, -СН3, -ОН, -ОСН3, -ОСН2СН3 или -СН(СН3)2;
R4 представлен -CN, -NO2, -СН3, -СН2 СН3, -СН2СН2-Y или -Y;
R5 или R6 независимо представлены Н, -С1-4-алкилом, -С2-4-алкенилом, -С2-4-алкинилом, -Х-С1-3-алкилом, -Х-С2-4-алкенилом, -Х-С2-4-алкинилом, или -Y;
R7 представлен -CN, -С1-4-алкил-ОН, -C(O)NR10R11, C(O)NR12R13, -C1-4-алкил-NR10R11, -C(O)R12, -C(O)OR12, -C(O)NR12OR13, -C(O)NHC(O)R12, -C(O)NHCH2R12, -C(NH2)(NOR12), -S(O)R12, -S(O)(O)(OR12), -S(O)(O)(NHCO2R12), РО3R12, P(O)(NR12R13)(NR12R13), P(O)(NR12R13)(OR14), -CONR12(CH2)qOCH3, -CONR12(CH2)q NR8R9-оксадиазол, замещенный метилом;
R8 и R9 независимо представлены Н, -С1-7-алкилом, -С3-7-циклоалкилом, -O-C1-7-алкилом, -С1-7-алкил-Y или фенилом;
R10 и R11 независимо представлены метилом или этилом или вместе образуют морфолиногруппу, связавшись через атом азота;
R12, R13 и R14 независимо представлены Н, -C1-12-алкилом, -C2-12-алкенилом, -C2-12-алкинилом, -O-С1-12-алкилом, -O-С2-12-алкенилом, -O-С2-12 -алкинилом, -С3-7-циклоалкенилом, -С3-7-циклоалкинилом, линейным или циклическим гетероалкилом, арилом, гетероарилом или -Y;
Х представлен кислородом или серой;
Y представлен галогеном;
n является целым числом, выбранным из 0, 1 или 2;
m является целым числом 1 или 2;
р является целым числом от 1 до 4;
q является целым числом от 1 до 12,
или его фармацевтически приемлемой соли, причем указанная злокачественная опухоль является резистентной к отличному от указанного соединения формулы l модулятору рецептора эстрогена.
где R1 представлен -(СН2)nCR5=CR6R7, -(СН2)mC(X)NR8R9 или
R2 и R3 независимо представлены Н, -СН3, -ОН, -ОСН3, -ОСН2СН3 или -СН(СН3)2;
R4 представлен -CN, -NO2, -СН3, -СН2СН3, -СН2СН2-Y или -Y;
R5 или R6 независимо представлены Н, -С1-4-алкилом, -С2-4-алкенилом, -С2-4-алкинилом, -Х-С1-3-алкилом, -Х- С2-4-алкенилом, -Х-С2-4-алкинилом или -Y;
R7 представлен -CN, -С1-4-алкил-ОН, -C(O)NR10R11, C(O)NR12R13, -С1-4-алкил-NR10R11, -C(O)R12, -C(O)OR12, -C(O)NR12OR13, -C(O)NHC(O)R12, -C(O)NHCH2R12, -C(NH2)(NOR12), -S(O)R12, -S(O)(O)(OR12), -S(O)(O)(NHCO2R12), РО3R12, P(O)(NR12R13)(NR12R13), P(O)(NR12 R13)(OR14), -CONR12(CH2)qOCH3,
-CONR12(CH2)qNR8R9-оксадиазол, замещенный метилом;
R8 и R9 независимо представлены Н, -С1-7-алкилом, -С3-7-циклоалкилом, -O-С1-7-алкилом, -С 1-7-алкил-Y или фенилом;
R10 и R11 независимо представлены метилом или этилом или вместе образуют морфолиногруппу, связавшись через атом азота;
R12, R13 и R14 независимо представлены Н, -С1-12-алкилом, -C2-12-алкенилом, -C2-12-алкинилом, -O-C1-12-алкилом, -O-С2-12-алкенилом, -O-С2-12-алкинилом, -С3-7-циклоалкенилом, -С3-7-циклоалкинилом, линейным или циклическим гетероалкилом, арилом, гетероарилом или -Y;
Х представлен кислородом или серой;
Y представлен галогеном;
n является целым числом, выбранным из 0, 1 или 2;
m является целым числом 1 или 2;
р является целым числом от 1 до 4;
q является целым числом от 1 до 12,
или его фармацевтически приемлемой соли.
| СПОСОБ ПОЛУЧЕНИЯ ЗАМЕЩЕННОГО ИМИДАЗОЛА | 1992 |
|
RU2045519C1 |
| RU 94029664 A1, 10.08.1996 | |||
| RU 94031162 A1, 20.04.1996 | |||
| US 5410080, 25.04.1995 | |||
| WO 9640098, 19.12.1996 | |||
| US 5393785, 28.02.1995 | |||
| Kudoh M | |||
| et al | |||
| Strategy of drug development for hormone-dependent tumor, Gan To Kagaku Ryoho 1996 May; 23(6):668-72. | |||

