Изобретение относится к антиген-специфическим супрессирующим механизмам иммунного ответа, использующим Zot или зонулин. Конкретно, данное изобретение охватывает способ ингибирования опосредуемой антиген-представляющей клеткой антиген-специфической пролиферации лимфоцитов в зависимости от дозы введением эффективного количества Zot или зонулина.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
I. 3оны плотных контактов и актиновый цитоскелет
Зоны плотных контактов (далее в данном описании "tj") или zonula occludens (далее в данном описании "ZO") являются одним из критериев абсорбирующих и секреторных эпителиальных тканей (Madara, J. Clin. Invest., 83:1089-1094 (1989) и Madara Textbook of Secretory Diarrhea Eds. Lebenthal et al., Chapter 11, pages 125-138 (1990). В качестве барьеров между апикальным и базолатеральным отделениями они селективно регулируют пассивную диффузию ионов и растворимых в воде веществ при трансклеточном транспорте (Gumbiner, Am. J. Physiol. , 253 (Cell Physiol. 22):C749-C758 (1987)). Этот барьер сохраняет любой градиент, вызванный активностью направлений, связанных с трансклеточным путем передачи (Diamond, Physiologist, 20:10-18 (1977)).
Существует множество свидетельств того, что ZO, рассматриваемые как статические структуры, в действительности являются динамическими и легко приспосабливаются ко множеству эволюционных (Magnuson et al., Dev. Biol., 67: 214-224 (1978); Revel et al., Cold Spring Harbor Symp. Quant. Biol., 40: 443-455 (1976) и Schneeberger et al., J. Cell Sci., 32:307-324 (1978)), физиологических (Gilula et al., Dev. Biol., 50:142-168 (1976); Madara et al. , J. Membr. Biol., 100:149-164 (1987); Mazariegos et al., J. Cell Biol., 98: 1865-1877 (1984) и Sardet et al., J. Cell Biol., 80:96-117 (1979)), и патологических (Milks et al., J. Cell Biol., 103:2729-2738 (1986); Nash et al. , Lab. Invest. , 59: 531-537 (1988) и Shasby et al., Am. J. Physiol., 255 (Cell Phvsiol. 24): C731-C788 (1988)) условий. Регуляторные механизмы, которые лежат в основе этой адаптации, до сих пор еще не совсем понятны. Однако ясно, что в присутствии Са++, скопление ZO есть результат клеточных взаимодействий, которые инициируют сложный каскад биохимических событий, приводящих, в конечном счете, к образованию и модуляции организованной сети ZO-элементов, структура которой была охарактеризована только частично (Diamond, Physiologist, 20:10-18 (1977)). Определялись возможные окклюдирующие нити трансмембранных белков (Furuse et al., J. Membr. Biol., 87:141-150 (1985)).
Было идентифицировано шесть белков в цитоплазматической субмембранной бляшке, определяющей контакты мембран, но их функцию еще нужно установить (Diamond, см. выше). ZO-1 и ZO-2 существуют в виде гетеродимера (Gumbiner et al., Proc. Natl. Acad. Sci., USA, 88:3460-3464 (1991)) в устойчивом к детергенту комплексе с неохарактеризованным белком 130 kD (ZO-3). Большинство исследований с помощью иммуноэлектронной микроскопии локализовали ZO-1 точно под мембранными контактами (Stevenson et al., Molec. Cell Biochem., 83: 129-145 (1988)). Два других белка, цингулин (Citi et al., Nature (London), 333: 272-275 (1988)) и антиген 7Н6 (Zhong et al., J. Cell Biol., 120:477-483 (1993)) локализованы дальше от мембраны и еще не клонировались. Rab 13, GTR-связывающий белок малого размера также был недавно локализован в области контактов (Zahraoui et al. , J. Cell Biol., 124:101-115 (1994)). Известны другие GTR-связывающие белки малого размера, регулирующие кортикальный цитоскелет, например, rho регулирует прикрепление актина к мембране при фокальных контактах (Ridley et al., Cell, 70:389-399 (1992)), а rас регулирует индуцированные фактором роста активные перемещения в мембране клетки (Ridley et al., Cell, 70:401-410 (1992)). По аналогии с известными функциями белков бляшек в лучше охарактеризованных контактах клеток, фокальных контактах (Guan et al., Nature, 358:690-692 (1992)) и плотных соединениях (при прилипании) (Tsukita et al., J. Cell Biol., 123:1049-1053 (1993)), была высказана гипотеза, что белки бляшек, ассоциированных с tj (tj-ассоциированных бляшек), участвуют в передаче сигналов через клеточную мембрану в обоих направлениях и в регуляции связей с кортикальным актиновым цитоскелетом.
Для того чтобы выдержать различные физиологические испытания, которым подвергается эпителий, ZO должны быть способны на быстрые и координированные реакции, которые требуют наличия комплексной регуляторной системы. Точная характеристика механизмов сборки и регуляции ZO в настоящее время является целью активного исследования.
Сейчас имеется масса свидетельств того, что структурные и функциональные связи tj существуют между актиновым цитоскелетом и tj-комплексом и всасывающими клетками (Gumbiner et al., см. выше; Madara et al., см. выше и Drenchahn et al. , J. Cell Biol., 107:1037-1048 (1988)). Актиновый цитоскелет состоит из сложной сети микрофиламентов, точная геометрия которой регулируется большим числом актин-связующих белков. Примером того, как состояние фосфорилирования актин-связующего белка может регулировать связывание цитоскелета с клеточной плазматической мембраной, является миристоилированный богатый аланином субстрат С-киназы (далее в настоящем описании "MARCKS"). MARCKS представляет собой субстрат специфичной протеинкиназы С (далее в настоящем описании "РКС"), который ассоциирован с цитоплазматической поверхностью плазматической мембраны (Aderem, Elsevier Sci. Pub. (UK), pages 438-443 (1992)). В своей нефосфорилированной форме MARCKS образует поперечные сшивки с актином мембраны. Т.е., по-видимому, актиновая сеть, ассоциированная с мембраной с помощью MARCKS, является относительно жесткой (Hartwig et al. , Nature, 356:618-622 (1992)). Активированная РКС фосфорилирует MARCKS, который высвобождается из мембраны (Rosen et al., J. Exp. Med. , 172: 1211-1215 (1990) и Thelen et al., Nature, 351:320-322 (1991)). Актин, связанный с MARCKS, по-видимому, пространственно отделен от мембраны и более пластичен. Когда MARCKS дефосфорилируется, он возвращается в мембрану, где снова образует поперечные сшивки с актином (Hartwig et al., см. выше и Thelen et al., см. выше). Эти данные говорят о том, что F-актиновая сеть может быть перестроена в процессе РКС-зависимого фосфорилирования, в котором участвуют актин-связывающие белки (MARCKS является одним из них).
II. Токсины зоны плотных контактов (Zonula Occludens) ("Zot") и зонулин
Большинство возможных вакцин Vibrio cholerae, созданных делетированием гена ctxA, кодирующего холерный энтеротоксин (СТ), способны вызывать мощный гуморальный иммунный ответ, но у более половины вакцинированных наблюдается слабая диарея (Levine et al., Infect. Immun., 56(1):161-167 (1988)). Учитывая важность того, что диарея возникает в отсутствие СТ, было высказано предположение, что V. cholerae продуцирует другие энтеротоксигенные факторы, которые все еще присутствуют в штаммах делетированной сtxA-последовательности (Levine et al., см. выше). В результате был обнаружен второй токсин, токсин зоны плотных контактов (далее в настоящем описании "Zot"), вырабатываемый V. cholerae, который "вносит вклад" в остаточную диарею (Fasano et al. , Proc. Natl. Acad. Sci., USA, 8:5242-5246 (1991)). Ген zot локализован как непосредственно прилегающий к генам ctx. Высокий процент совпадений гена zot с генами ctx в штаммах V. cholerae (Johnson et al., J. Clin. Microb., 31/3: 732-733 (1993) и Karasawa et al., FEBS Microbiology Letters, 106:143-146 (1993)) говорит о синергистической роли zot в обезвоживающей диарее, типичной для холеры. Недавно ген zot также был идентифицирован в других возбудителях кишечных заболеваний (Tschape, 2nd Asian-Pacific Symposium on Typhoid fever and other Salmonellosis, 47(Abstr.) (1994)).
Ранее при испытании на слизистой оболочке подвздошной кишки кролика было найдено, что Zot повышает проницаемость кишечника за счет модуляции структуры межклеточных зон плотных контактов (Fasano et al., см. выше). Было обнаружено, что вследствие модификации перицеллюлярного пути интестинальная слизистая оболочка становится более проницаемой. Также было найдено, что Zot не влияет на одновременный активный транспорт Na+-глюкозы, не цитотоксичен и не может полностью уничтожить трансэпителиальную устойчивость (Fasano et al. , см. выше).
Позднее было обнаружено, что Zot способен обратимо открывать зоны плотных контактов в интестиальной слизистой оболочке и, следовательно, Zot при одновременном введении с терапевтическим агентом может осуществлять интестиальную доставку терапевтического агента, если применяется в составе пероральной лечебной дозы для интестиальной доставки лекарственного вещества (WO 96/37196, Патенты США 5827534 и 5665389). Также было найдено, что Zot способен обратимо открывать зоны плотных контактов в слизистой оболочке носа и, следовательно, Zot при совместном введении с терапевтическим агентом может повышать всасывание терапевтического агента через нос (WO 98/30211 и Патент США 5908825).
В Патентах США 5864014 и 5912323 рецепторы Zot из СаСо2-клеток, тканей сердца, кишечника и мозга были идентифицированы и выделены. Рецепторы Zot представляют собой первую стадию перицеллюлярного (околоклеточного) транспорта, участвующего в регуляции эпителиальной интестинальной и назальной проницаемости.
В Патенте США 5945510 были идентифицированы и очищены белки млекопитающих, которые иммунологически и функционально близки Zot и которые функционируют как физиологический модулятор зоны плотных контактов у млекопитающих. Эти белки млекопитающих, названные "зонулин", применимы для повышения всасывания терапевтических агентов при прохождении через зоны плотных контактов интестиальной и назальной слизистой оболочки, а также через зоны плотных контактов гематоэнцефалического барьера. Эти белки дополнительно характеризуются способностью связываться с рецепторами Zot.
В находящейся на рассмотрении патентной Заявке США 09/127815, поданной 3 августа 1998 г., озаглавленной "Пептидные антагонисты зонулина и способы их использования", были идентифицированы пептидные антагонисты зонулина. Указанные пептидные антагонисты связываются с рецептором Zot, однако не функционируют таким образом, чтобы модулировать открытие зон плотных контактов у млекопитающих. Пептидные антагонисты конкурентно ингибируют связывание Zot и зонулина с рецептором Zot, подавляя тем самым способность Zot и зонулина физиологически модулировать открытие зон плотных контактов млекопитающих.
III. Антиген-представляюшие (презентирующие) клетки и иммунные реакции
Подробно иммунные реакции и иммуномодуляция обсуждаются в главе 10 "Recent Advances in Immunology", Sztein et al., New Generation of Vaccines, pages 99-125.
Один из первичных механизмов защиты от инфицирующих агентов включает специфический или приобретенный (искусственный) иммунитет. В противоположность врожденному иммунитету эффекторные механизмы приобретенного иммунитета, которые включают, среди прочих, антитела, цитотоксические лимфоциты (далее в настоящем описании "CTL"), вырабатываемые Т-лимфоцитами цитокины (такие как IFN-g, IL-4 и т.д.), инициируются после экспозиции с антигенами или инфицирующими агентами, и интенсивность (т.е. иммунного ответа) возрастает при последующей экспозиции со специфическими антигенами. Эта способность "вспоминать" предшествующие экспозиции с антигенами и быстро отвечать более интенсивными иммунологическими эффекторными реакциями (иммунологическая память) составляет основу иммунопрофилактической иммунизации (вакцинации) против возбудителей инфекции. Основные виды клеток, участвующих в специфическом иммунном ответе, представляют собой Т- и В-лимфоциты.
В-лимфоциты или В-клетки образуются в костном мозге и являются предшественниками секретирующих антитела клеток (плазматических клеток). В-клетки распознают антигены (белки, углеводы или простые химические группы) с помощью иммуноглобулиновых рецепторов на клеточной мембране (Fearon et al., Science, 272: 50-53 (1996); Zeigler-Heitbroak et al., Immunol. Today, 14: 121-152 (1993) и Banchereau et al., Adv. Immunol., 52:125-262 (1992)). После стимулирования антигеном они клонально размножаются и переключают экспрессию антительного изотипа (например, IgM на IgG, IgE или IgA) под влиянием цитокинов, вырабатываемых Т-клетками, макрофагами и другими видами клеток. Генерируются соматически мутированные высокоаффинные В-клетки и отбираются антигеном в и вокруг зародышевых центров, которые образуются в лимфатических узлах, солезенке, пейеровых бляшках и более беспорядочных лимфатических агрегациях периферической лимфоидной системы (Banchereau et al., (1996), см. выше; Сlark et al., Ann. Rev. Immunol., 9:97-127 (1991) и MacLennan et al., Immunol. Today, 14:29-34 (1993)). Они являются основой В-клеточной памяти.
Т-лимфоциты или Т-клетки, в отличие от В-клеток, распознают пептиды из белковых антигенов, которые присутствуют на поверхности антиген-представляющих клеток (далее в настоящем описании "АРС") в виде конъюгатов с молекулами класса I и класса II главного комплекса гистосовместимости (МНС). Клоны Т-лимфоцитов, экспрессирующие Т-клеточные рецепторы (далее в настоящем описании "TCR") соответствующей аффинности, "запускаются" антигеном для пролиферации и эволюции в эффекторные клетки (Fearon, 91996), см. выше; Sprent et al. , Cell, 76:315-322 (1994) и Hendrick et al., Germain, Fundamental Immunology, 3rd ed., pages 629-676 (1993)). После элиминирования инфицирующего агента антиген-специфические клоны оставляют в качестве Т-клеток памяти те клетки, которые, после соответствующей экспозиции с антигеном ("облучения"), дают более мощный, более быстрый и иногда качественно отличный специфический иммунный ответ.
Существует две основные популяции Т-клеток, одна из которых экспрессирует молекулы CD4, а вторая - молекулы CD8. CD4 и CD8 являются гликопротеинами Т-клеточной поверхности, которые служат в качестве важных вспомогательных веществ (корецепторы) во время презентации антигена, связываясь с МНС-молекулами класса II и класса I, соответственно (Hendrick et al., см. выше (1993)). То есть, молекулы CD4 и CD8 играют значительную роль в стабилизации взаимодействий Т-клеток и АРС, инициированных специфическим связыванием TCR-комплекса с антигенными пептидами, ассоциированными с МНС-молекулами. Соответственно, молекулы CD4 и CD8, первоначально используемые в качестве маркеров для идентификации популяций Т-клеток с различными функциональными характеристиками, играют главную роль при класс II МНС-рестриктированной и класс I MHC-рестриктированной активации Т-клеток. Клетки CD4+ (Т-хелперная, или Th) в основном участвуют в воспалительных реакциях и способствуют продуцированию антител В-клетками, тогда как CD8+ (Т-цитотоксическая, или Те) составляют большинство CTL, первоначально участвующих в класс I МНС-рестриктированном киллинге клеток-мишеней, инфицированных патогенными организмами, включая бактерии, вирусы и паразиты (Sztein et al., J. Immunol. , 155: 3987-3993 (1995); Kaufman, Ann. Rev. Immunol., 11:129-163 (1993) и Immunol. Today, 9:168-174 (1988); Townsend et al., Cell, 44:959-968 (1986); Malik et al., Proc. Natl. Acad. Sci., USA, 88:3300-3304 (1991); Sedegah et al. , J. Immunol., 149:966-971 (1992) и Shearer et al. Immunol. Today, 17: 21-24 (1996)).
Для успешной антиген-специфической активации Т-клеток, приводящей к экспансии и дифференцировке Т-клеток (или пролиферации лимфоцитов), нужен первый сигнал, обеспечиваемый взаимодействием TCR на поверхности Т-клеток с МНС-антигенными комплексами на АРС, и второй, комплементарный, сигнал, предоставляемый растворимыми факторами, такими как IL-2, или связыванием CD28 (костимулятора) с представителями семейства В7 (например, CD80 (В7-1) или CD86 (В7-2)) на АРС (Lenschow, Ann. Rev. Immunol., 14:233-258 (1996) и Linsley et al. , Ann. Rev. Immunol., 11:191-212 (1993)). Изучение CD28/B7 костимуляторного пути и других "липких" молекул, которые помогают стабилизировать взаимодействия Т-се11-АРС (и которые, по-видимому, играют решающую роль в холинге лимфоцитов), является одной из ключевых областей, в которой в последние годы сделаны значительные успехи.
Презентация антигенов Т-клетками участвует в ряде внутриклеточных событий, включая образование антигенных пептидных фрагментов, связывание этих пептидов с МНС-молекулами с образованием устойчивых комплексов пептид-МНС и транспорт этих комплексов к поверхности клетки, где их могут распознать TCR на поверхности Т-клеток. Накоплены свидетельства существования двух основных путей процессирования и презентации антигенов ("классические пути"). Один из этих путей, "цитозольный путь", преимущественно используется для презентации пептидов, продуцируемых эндогенно в АРС, таких как вирусные белки, опухолевые антигены и аутологичные пептиды, ассоциированные с МНС-молекулами класса I (Hendrick et al., см. выше и Germain, см. выше (1993)). Презентация большого числа аутологичных пептидов, закомплексованных с класс I МНС-молекулами, является результатом неспособности АРС дифференцировать аутологичные и неаутологичные. В нормальных условиях большинство Т-клеток, отобранных для распознавания аутологичных пептидов, элиминирует во время дифференцировки Т-клеток или активно ингибируются (негативно регулируются) и, следовательно, не могут активироваться комплексами аутологичный пептид- класса I MHC. Второй "классический путь" процессирования и презентации антигена, "эндосомный путь", который преимущественно используется для презентации растворимых (экзогенных) чужеродных антигенов, связанных с МНС-молекулами класса II, включает захват антигена с помощью АРС или за счет связывания со специфическим рецептором или улавливания в жидкой фазе за счет макропиноцитоза (Lanzavecchia, Curr. Opin. Immunol., 8:348-354 (1996)). Стимуляция Т-клеток с помощью TCR была показана на 200-600 комплексах пептид/МНС для случая нуклеопротеинов гриппа (Falk et al. , Semin. Immunol., 5:81-94 (1993)). В большинстве иммунных реакций антигенные эпитопы, связанные с молекулами МНС класса I, стимулируют активацию СD8+СТL-иммунных ответов, тогда как антигенные фрагменты (эпитопы) из растворимых белков, закомплексованных с МНС-молекулами класса II, распознаются CD4+Th-клетками. Эти открытия представляют собой один из важнейших за последние несколько лет вкладов в механизмы, участвующие на ранних стадиях иммунной активации, и являются основополагающими для создания успешных вакцин.
Как указано выше, существуют два "классических" пути процессирования и презентации антигена. Путь МНС класса I обычно наиболее применим для процессирования клеточных белков, присутствующих в большинстве, если не во всех участках клетки, включая цитозоль, ядро и митохондрии (Falk et al., см. выше (1993)), для распознавания с помощью CD8+CTL. Путь МНС класса II преимущественно использует для процессирования и презентации экзогенных антигенов, таких как белки, продуцируемые внеклеточными бактериями и другими инфекционными (болезнетворными) микроорганизмами, которые могут быть представлены СD4+Тh-клетками. Молекулы МНС класса I и класса II связывают пептидные антигены за счет использования поверхностных "рецепторов" или "связывающих карманов". Однако способ процессирования и получения антигена в этих двух случаях резко отличается. Антигены класса I процессируют и получают "цитозольным путем". Конкретно, пептиды, синтезированные внутриклеточно, распадаются на малые фрагменты белков, которые затем проходят через мембрану эндоплазматического ретикулума (ER). Внутри ER антигенные фрагменты связываются с МНС-молекулами класса I, образуя комплекс, который затем переносится в аппарат Гольджи и, наконец, к клеточной поверхности, где они распознаются TCR, передающими сигнал антиген-специфической CTL-экспансии и дифференцировки, первой стадии иммунного ответа. Антигены класса II, с другой стороны процессируют и получаются "эндосомным путем". Конкретно, нативные антигены захватываются циркулирующими АРС, при этом антиген связывается со специфическим или неспецифическим рецептором. Затем антиген интернализуется АРС по механизму опосредуемого рецепторами эндоцитоза или пиноцитоза. Интернализованный антиген затем локализуется в эндосоме, мембрано-связанной везикуле, участвующей во внутриклеточном транспорте и распаде антигена. Фрагменты расщепленного пептида затем связываются с молекулами МНС класса II, образуя комплекс, который переносится через аппарат Гольджи, эндосомное отделение и к поверхности клетки, чтобы их могли распознать TCR, опять же передающие сигнал антиген-специфической Th-клеточной экспансии и дифференцировки.
АРС играют жизненно важную роль в выработке иммунного ответа. Для презентации процессированных антигенов CTL класс I-ограниченным образом, АРС должны экспрессировать МНС-молекулы класса I и иметь (возможность) способность экспрессировать на клеточной поверхности эндогенно продуцированные белки, закомплексованные с МНС-молекулами класса I. Почти все клетки, эндогенно продуцирующие вирусные, паразитные или бактериальные белки или опухолевые антигены, которые получают доступ к цитозоли, могут работать как АРС. Для презентации процессированных антигенов Th-клеткам класс II-ограниченным образом, АРС должны уметь распознавать антиген и связываться с ним с помощью (через посредство) специфических и неспецифических рецепторов конкретного антигена. Клетки, которые наиболее эфективно презентируют (представляют) антигены Th-лимфоцитам, так называемые профессиональные АРС, включают дендритные клетки (DC), макрофаги, В-лимфоциты, клетки Лангерганса и, в некоторых случаях (примерах), эндотелиальные клетки человека (Lanzavecchia, см. выше (1996)).
Полагают, что DC, которые образуются в костном мозге, являются наиболее эффективными АРС для презентации растворимых антигенов. DC захватывают антигены на периферии и мигрируют в селезенку или лимфатические узлы, где они эффективно активируют Тh-клетки, особенно "необученные" Т-клетки (Lanzavecchia, см. выше (1996) и Peters et al., Immunol. Today, 17:273-278 (1996)). Несколько уникальных особенностей позволяют DC функционировать так эффективно в качестве антиген-презентирующей клетки. А именно: они способны интернализовать растворимые антигены по нескольким механизмам, включая конститутивный макропиноцитоз, интернализацию комплексов антиген-антитело путем CD32-рецепторного связывания и интернализацию маннозилированных или фукозилированных антигенов путем маннозо-рецепторного связывания. Это позволяет DC испытать (проверить) большие количества жидкости за короткое время, аккумулируя их в лизосомном отделении, содержащей МНС-молекулы класса II и протеазы. DC также конститутивно экспрессируют ряд костимуляторов и других адгезивов, которые позитивно регулируются (активируются) провоспалительными цитокинами, такими как IL-1a, IL-1b и TNF-a, с помощью этого повышается их способность функционировать как АРС для класса II МНС-ограниченных Th-иммунных реакций.
Макрофаги и другие мононуклеарные фагоциты являются, возможно, наиболее эффективными АРС для антигенов из наиболее болезнетворных микроорганизмов, иных нежели вирусы, вследствие их способности фагоцитировать большие частицы, такие как бактерии и паразиты. В типичных условиях фаготированные микрорганизмы затем уничтожаются в фаголизосомах и расщепляются, приводя к образованию антигенных фрагментов, способных связываться с класс II МНС-молекулами для презентации Тh-клеток. Другие важные механизмы, с помощью которых макрофаги могут служить в качестве эффективных АРС, включают их способность интернализовать антигены путем связывания комплексов антиген-антитело с CD16-, CD32- и CD64-peцeптopaми. Макрофаги также интернализуют вирусные белки комплемента через рецепторы С3 и других Су-компонентов и путем стимуляции факторами роста, с помощью макропиноцитоза. Кроме того, макрофаги экспрессируют рецепторы маннозы и являются главным источником провоспалительных цитокинов, включая IL-1a, IL-1b, IL-6, IL-8, IL-12, TNF-a и TNF-b, которые вызывают мощную иммунорегуляторную активность при Т-клеточных иммунных реакциях (Sztein et al., см. выше (1997)).
В-лимфоциты являются очень эффективными АРС для растворимых антигенов для презентации Th-клеткам. Это в большой степени основано на их способности связывать и интернализовать специфичные растворимые антигены очень эффективно за счет комплекса В-клеточных рецепторов (BCR), состоящего из специфичного мембранного иммуноглобулина (mIg) и гетеродимера Iga (CD79a)-Igb (CD79b) (Falk et al., см. выше (1993)).
Клетки Лангерганса (LC), образованные из предшественников костного мозга, рассматриваются как единственные клетки в эпидермии с АРС-способностью. LC мигрируют из эпидермиса через лимфатические протоки в региональные лимфатические узлы, где они эволюционируют в DC. Интересно, что LC экспрессируют CD1, неклассическую МНС-молекулу, способную презентировать Т-клеткам, ограниченным образом, небелковые антигены, такие как микробные липидные и гликолипидные антигены.
Изобретение по данному описанию сфокусировано на антеген-специфических супрессирующих механизмах АРС-опосредуемых иммунных ответов. Изобретение порождено открытием рецептора макрофагальной поверхности, с которым Zot связывается специфическим и "насыщаемым" образом. Настоящее изобретение описывает способ применения Zot или зонулина в качестве антиген-специфических иммунорегуляторов и в качестве иммунотерапевтических средств. А именно, как Zot, так и зонулин ингибируют АРС-опосредуемую антиген-специфическую пролиферацию лимфоцитов в зависимости от дозы, не влияя на ответы, вызываемые митогеном. Эти супрессирующие механизмы иммунного ответа, по меньшей мере, частично, связаны с пониженным поглощением антигена.
Имеющиеся в настоящее время модуляторы иммунного ответа, такие как циклоспорин и стероидные соединения, оказывают общий эффект на антигенную и митогенную стимуляции иммунной системы (Reed et al., J. Immunol., 137:150-154 (1986)). Раскрываемое в данном описании изобретение имеет преимущество, делая возможной негативную регуляцию (супрессорный механизм) иммунных ответов на (конкретный) отдельный антиген, не вызывая негативных побочных эффектов, таких как повышенная чувствительность к инфекции и общая иммунная супрессия, типичные для иммуномодуляторов предыдущего уровня данной области техники.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Целью данного изобретения является предоставить способ негативной регуляции иммунного ответа хозяина-животного на некоторые антигены, посредством этого облегчая иммунотерапию. Конкретно, предметом данного изобретения является ингибирование (подавление) способности антиген-презентирующих клеток (АРС) процессировать и презентировать антигены лимфоцитам, с помощью этого подавляя пролиферацию лимфоцитов и последующие реакции иммунной системы в ответ на определенные антигены.
Еще одной целью данного изобретения является обеспечение лечения животных, пораженных аутоиммунным или иммунопатологическим заболеванием или расстройством, таким как рассеянный склероз, ревматоидный артрит, инсулинозависимый сахарный диабет, глютеновая болезнь, синдром Шегрена, системный волчий эритематоз, аутоиммунный тироидит, идиопатическая тромбоцитопеническая пурпура, гемолитическая анемия, базедова болезнь, аддисонова болезнь, аутоиммунный орхит, пернициозная анемия, васкулит, аутоиммунные коагулопатии, тяжелая миастения (myasthenia gravis), полиневрит, пузырчатка, ревмокардит, полимиозит, дерматомиозит и склеродермия, введением эффективного количества родственного Zot иммунорегулятора. В альтернативном варианте изобретения лечение животного, пораженного аутоиммунным (аутоаллергическим) и иммунопатологическим заболеванием или расстройством, может включать прием эффективного количества родственного Zot иммунорегулятора в сочетании со специфическим(-и) родственным (-и) антигеном(-ами).
Еще одной целью изобретения является создание способа терапии животного с отторжением, связанным с иммунными нарушениями после трансплантации ткани или органа, введением эффективного количества Zot-связанного иммунорегулятора. В альтернативном варианте изобретения лечение животного с вызванным иммунными нарушениями отторжением после трансплантации ткани или органа, может включать введение эффективного количества Zot-связанного иммунорегулятора в сочетании со специфическим(-и) трансплантационным(-и) антигеном (-ами).
Следующей целью данного изобретения является создание способа лечения животного, пораженного воспалительным или аллергическим заболеванием или состоянием, таким как астма, псориаз, экзематозный дерматит, саркома Капоши, рессеянный склероз, кишечное заболевание, нарушение пролиферации клеток гладких мышц, и воспалительным состоянием, связанным с микотическими, вирусными, паразитарными или бактериальными инфекциями, введением терапевтически эффективного количества Zot-связанного иммунорегулятора. В альтернативном варианте изобретения лечение животного, пораженного воспалительным или аллергическим заболеванием или расстройством, может включать введение эффективного количества Zot-связанного иммунорегулятора в сочетании со специфическим(-и) родственным(-и) воспалительному антигеном(-ами) или аллергеном(-ами).
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг. 1 даны кривые насыщения Zot-FITC для лимфоцитов и макрофагов. Данные показывают, что Zot связывается предпочтительно с моноцитами/макрофагами человека.
Фиг. 2 иллюстрирует блокаду Zot-FITC-связывания немеченым Zot. Предварительное термостатирование РВМС с немеченым Zot понижает связывание Zot-FITC как с моноцитами/макрофагами, так и с Т-лимфоцитами, примерно на 33%, что говорит, что Zot-связывание с этими клетками опосредуется рецептором. Предварительное термостатирование (преинкубация) клеток с очищенным МВР не оказывает влияния на блокаду Zot-FITC-связывания, показывая, что блокада меченым Zot есть специфический феномен.
На фиг. 3 иллюстрировано влияние Zot на пролиферацию РВМС человека, индуцированную РНА и столбнячным токсином. Данные показывают, что Zot заметно подавляет стимуляцию столбнячным токсином пролиферацию в зависимости от дозы, не оказывая влияния на пролиферацию, стимулированную РНА.
На фиг. 4 показано действие антисыворотки к анти-Zot на Zot-индуцированную супрессию пролиферации РВМС человека, вызванной столбнячным токсином. Добавление анти-Zot "реверсирует" Zot-опосредуемую супрессию индуцированной столбнячным токсином пролиферацию более чем на 50%.
На фиг. 5A-5D проиллюстрировано действие Zot на поглощение FITC-декстрана нормальными CD14+HLA-DR+ макрофагами человека. На фиг. 5А изображено поглощение FITC-декстрана в средах при 0oС (2,9%) и представлена температурная зависимость поглощения антигена. На фиг. 5В изображено поглощение FITC-декстрана в средах при 37oС (46,0%) и представлен контроль за поглощением антигена. На фиг. 5С изображено поглощение FITC-декстрана в BSA при 37oС (39,0%) и представлен негативный контроль поглощения антигена. На фиг. 5D изображено поглощение FIТС-декстрана в Zot при 37oС (19,3%). Данные указывают, что Zot понижает поглощение антигена.
На фиг. 6 показано число FITC-Zot сайтов связывания/в клетке в макрофагах и лимфоцитах человека. РВМС термостатировали (инкубировали) при увеличивающихся концентрациях Zot-FITC и анализировали проточной цитометрией. Среднюю интенсивность флуоресценции каждой популяции переводили в число Zot-связывающих сайтов/в клетке с помощью стандартной кривой, построенной с использованием набора Quantum 26-MESF. Эти данные показывают, что связывание Zot является насыщаемым феноменом, причем насыщение достигается примерно, при 0,5 пМ, и что среднее число Zot-связывающих сайтов/в клетке, примерно, в 10 раз выше в макрофагах (~106000), чем в лимфоцитах (~9000).
На фиг. 7А-7С показана кинетика связывания Zot с макрофагами и лимфоцитами человека. Исследование кинетики связывания Zot с клетками человека проводили проточной цитометрией, используя конъюгаты Zot-FITC. РВМС, меченые анти-CDS-ECD и анти-СD-14-РЕ mAb, выдерживали при 37oС во время эксперимента (12 мин), тогда как данные получали, используя устройство для сортировки жизнеспособных образцов (кинетический модуль), соединенный с проточным цитометром. Уровни фоновой FITC-флуоресценции регистрировали в течение 90-150 с (указанные стрелками) и регистрацию данных прекращали на 10-15 с для инъекции Zot-FITC (фиг. 7А и 7В). На фиг. 7С показана кинетика связывания aнти-CD-14-FITC с немечеными моноцитами/макрофагами человека. Данные представлены в виде изометрического изображения интенсивности Zot-FITC (ось у) во времени (ось х) относительно числа клеток (ось z) для клеток (пропускаемых), селектируемых по CD3 (лимфоциты) или CD14 (макрофаги). Результаты показывают, что связывание Zot с макрофагами (фиг. 7А) и лимфоцитами (фиг. 7В) человека происходит очень быстро, достигая равновесия в пределах 2 мин после добавления Zot-FITC.
На фиг. 8 изображено связывание FITC-Zot с Т- и В-лимфоцитами. РВМС термостатировали с Zot-FITC и mAb к молекулам, присутствующим в Т-(СВ3+) и B-(CD19+) лимфоцитах, и анализировали прточной цитометрией. Меченое изотопным FITC контрольное mAb (антитело) (mIg), соответствующее клеткам, пропускаемым (селектируемым) как рассеяние вперед по сравнению с боковым рассеянием лимфоцитов, также показано как индикатор неспецифического связывания. Результаты показывают, что Zot связывается как с Т-, так и с В-лимфоцитами.
На фиг. 9 показана неспособность антагонистов Zot блокировать связывание Zot-FITC. РВМС, окрашенные CD14-PE и CD3-ECD, отмывали и термостатировали в течение 15 мин при 4oС в среде AIM-V, либо одной, либо со 100-кратным избытком FZI/0 (SEQ ID N0:7), FZI/1 (SEQ ID N0:8), BSA (негативный контроль) или 4-кратным избытком немеченого Zot (позитивный контроль). Затем клетки термостатировали с Zot-FITC и анализировали проточной цитометрией. Результаты выражены в % супрессии средней интенсивности флуоресценции клеток, термостатированных с Zot-FITC в присутствии антагонистов Zot, немеченого Zot или BSA, относительно средней интенсивности флуоресценции клеток, термостатированных только в средах (произвольно взятой за 100%). Результаты показывают, что добавление антагонистов Zot FZI/0 или FZI/1 незначительно блокирует связывание Zot-FITC с CD14+ отобранными (пропущенными) макрофагами. Напротив, преинкубация (предварительное термостатирование) с немеченым Zot блокировала связывание Zot-FITC на 24-43%.
На фиг.х 10А-10В проиллюстрирована супрессия экспрессии CD14 на моноцитах/макрофагах человека. РВМС термостатировали в течение 18 ч в отсутствие или в присутствии ТТ без или с очищенным Zot или BSA, окрашивали CD14-FITC и анализировали проточной цитометрией. Результаты приведены в виде единых цветных гистограмм на клетках, пропущенных в "области моноцитов", определяемой как рассеяние вперед по сравнению с боковым рассеянием макрофагов человека. Добавление Zot вызвало заметную супрессию экспрессии CD14 в отсутствие ТТ (фиг. 10А) или в присутствии ТТ (фиг. 10В).
На фиг. 11 показано действие Zot на жизнеспособность моноцитов/макрофагов и лимфоцитов. РВМС термостатировали в течение различного времени в отсутствие или в присутствии очищенного Zot или BSA. Жизнеспособность клеток оценивали с помощью исключения пропидийиодида и проточной цитометрии. Результаты представлены как% жизнеспособных клеток, зарегистрированных в "области моноцитов" ("моноцитарной области") или в "области лимфоцитов" ("лимфоцитарной области"), определяемой как рассеяние вперед по сравнению с боковым рассеянием этих клеточных популяций. Результаты показывают, что добавление Zot влияет на жизнеспособность макрофага в относительно ранний период, тогда как действие на лимфоциты незаметно в течение, по меньшей мере, 4 дней в культуре.
На фиг. 12А-12С показана Zot-опосредованная индукция продуцирования цитокинов моноцитами/макрофагами человека. РВМС термостатировали от 6 ч до 4 дней в отсутствие или в присутствии очищенного Zot или BSA. Супернатанты собирали в указанные промежутки времени и измеряли уровни цитокинов с помощью хемилюминесцентного ELISA. Добавление Zot привело к продуцированию на высоком уровне TNF-a (фиг. 12А) и IL-10 (фиг. 12С) уже через 6 ч, пик уровней достигается через 24 ч. Наблюдалась также слабая индукция продуцирования IL-1b (фиг. 12В).
На фиг. 13А-13В проиллюстрирована Zot-опосредованная индукция продуцирования цитокинов лимфоцитами человека. РВМС термостатировали в течение 3 дней без или с ТТ в отсутствие или в присутствии очищенного Zot или BSA и измеряли уровни цитокинов с помощью хемилюминесцентного ELISA. Добавление Zot вызывало супрессию продуцирования IL-2, стимулируемую термостатированием с ТТ, тогда как в отсутствие ТТ Zot не индуцировал IL-2 на поддающемся измерению уровне (фиг. 13А). Напротив, добавление Zot последовательно индуцировало продуцирование IFN-g в отсутствие ТТ, на уровнях, аналогичных уровням, индуцируемым ТТ, и заметно увеличивал уровни IFN-g, индуцированные ТТ (фиг. 13В).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Предыдущие исследования были сосредоточены на способности Zot и зонулина физиологически модулировать открытие зон плотных контактов (zonula occludens) эпителия различных тканей млекопитающих; такая модуляция особенно применима для облегчения доставки лекарственных средств через эти мембраны. В ходе этого исследования рецепторы Zot и зонулина были идентифицированы и выделены из СаСо2 клеток, тканей сердца, кишечника и мозга. Помимо открытия Zot-рецепторов, были идентифицированы пептидные антагонисты зонулина, причем указанные пептидные антагонисты связываются с Zot-рецептором, но не функционируют так, чтобы физиологически модулировать открытие зон плотных контактов у млекопитающих (то есть, отсутствие биологической активности). Пептидные антагонисты конкурентно ингибируют связывание Zot и зонулина с Zot-рецептором, посредством чего ингибируя способность Zot и зонулина к регуляции зон плотных контактов.
В свете известного действия Zot и зонулина на трансклеточный путь (транспорт) было действительно неожиданностью обнаружить рецептор Zot на полностью дифференцированных макрофагах, выделенных из крови. Сначала было неясно, почему циркулирующие клетки, такие как макрофаги, должны иметь рецептор молекулы, ассоциированной с модуляцией клеток тканей. В поисках ответа на этот вопрос было открыто, что Zot и зонулин также способны физиологически регулировать активность макрофагов. Не желая связывать себя теорией, предполагаем, что, по-видимому, Zot блокирует рецептор на макрофаге, связываясь с рецептором макрофагальной поверхности специфическим и насыщаемым образом. Связываясь с макрофагами, Zot изменяет способность макрофагов процессировать и представлять ("презентировать") антиген лимфоцитам, в конечном счете подавляя пролиферацию лимфоцитов в ответ на антиген в зависимости от дозы и антиген-специфическим образом. Другими словами, Zot делает возможным негативную модуляцию (супрессирующие механизмы) иммунного ответа. Результаты, подробно описанные ниже, говорят о том, что эти супрессирующие механизмы, по меньшей мере, чатично связаны с пониженным поглощением антигена. По-видимому, Zot является многофункциональным белком, регулирующим иммунный ответ по двойному механизму регулирования поглощения и направленной миграции антигенов.
Токсин zonula occludens, или "Zot" вырабатывается V. cholerae. Конкретный штамм V. cholerae, из которого получен Zot, не является единственным (критическим). Примеры таких штаммов V. cholerae включают штаммы 569В, 395 и Е7946 (Levine et al., см. выше; Johnson et al., см. выше и Karasawa et al., см. выше).
Применяемый по данному описанию "Zot" относится к зрелому белку из 399 аминокислот, а также к его мутантам, которые сохраняют способность регулировать tj. Например, можно осуществить делецию N-концов аминокислот 1-8, не влияя на активность Zot, и N-концевые слитые белки Zot можно получить без изменения активности Zot. Такие мутанты можно легко получить с помощью сайт-направленного мутагенеза и скрининга на Zot-активность, как показано в данном описании.
Zot можно получать и очищать, например, используя полученные генетической инженерией (с помощью рекомбинантной ДНК) штаммы Е. coli, сверхэкспрессирующие ген zot (Baudry et al., Infect. Immun., 60:428-434 (1992)), один или слитый с другими генами, такими, как белок, связывающий мальтозу (см. ниже, пример 1), глютатион-S-трансфераза (см. ниже, пример 2) или 6-полигистидин (см. ниже, пример 2).
Употребляемый в данном описании термин "зонулин" относится к практически чистому биологически активному белку с явным молекулярным весом около 47 kDa, определенным с помощью электрофореза на SDS-полиакриламидном геле, и следующей N-концевой аминокислотной последовательностью: Asn Asp Gln Pro Pro Pro Ala Gly Val Thr Ala Туг Asp Tyr Leu Val Ile Gln (SEQ ID N0:l), или следующей N-концевой аминокислотной последовательностью: Glu Val Gln Leu Val Glu Ser Gly Gly Xaa Leu Val Gln Pro Gly Gly Ser Leu Arg Leu (SEQ ID N0:2), а также их мутанты, которые сохраняют способность связываться с рецептором Zot и регулировать tj.
Зонулин продуцируется в различных клетках и тканях млекопитающих или обнаруживается в них, например, в клетках/тканях кролика или человека. Конкретный вид клетки/ткани млекопитающих, из которого получен зонулин, не является единственным (критическим) для данного изобретения. Примеры таких видов тканей млекопитающих включают сердце, легкие, кишечник, печень, почку и поджелудочную железу.
Зонулин можно получать и очищать, например, аффинной хроматографией с применением антител против Zot, как описано в примере 3 Патента США 5945510.
Употребляемые в данном описании термины "родственный Zot иммунорегулятор" или "молекула родственного Zot иммунорегулятора" относятся к Zot или зонулину, как определяется выше. Кроме того, употребляемый в данном описании термин "ингибирование" (и "ингибировать" и "ингибирующий") относятся к негативной модуляции любого аспекта АРС-активности, которая приводит к значительному уменьшению пролиферации лимфоцитов. Примеры активности включают поглощение антигена, процессирование антигена и презентацию антигена. Термин "ингибирование" по данному описанию не обязательно подразумевает полную супрессию функции или результата. Скорее, этот термин означает частичное ингибирование.
Для терапии животного-хозяина, пораженного аутоиммунным или иммунопатологическим заболеванием или расстройством, таким как рассеянный склероз, ревматоидный артрит, инсулинозависимый сахарный диабет, глютеновая болезнь, синдром Шегрена, системный волчий эритематоз, аутоиммунный тироидит, идиопатическая тромбоцитопеническая пурпура, гемолитическая анемия, базедова болезнь, аддисонова болезнь, аутоиммунный орхит, пернициозная анемия, васкулит, аутоиммунные коагулопатии, тяжелая миастения (myasthenia gravis), полиневрит, пузырчатка, ревмокардит, полимиозит, дерматомиозит и склеродермия, родственный Zot иммунорегулятор вводят один или в комбинации со специфическим родственным аутоиммунному антигену. Примерами специфических аутоиммунных антигенов, связанных с аутоиммунными заболеваниями или нарушениями, являются глиадин (антиген, связанный с глютеновой болезнью) и миелиновый основной белок (связанный с рассеянным склерозом).
Для терапии животного-хозяина, пораженного иммунным отторжением после трансплантации ткани или органа, родственный Zot иммунорегулятор вводят один или в комбинации со специфическим трансплантационным антигеном. Трансплантационные антигены можно получать при анализе ткани или органа трансплантата на HLA.
Для терапии животного-хозяина, пораженного воспалительным или аллергическим заболеванием или нарушением, таким как астма, псориаз, экзематозный дерматит, саркома Капоши, рессеянный склероз, кишечное заболевание, нарушение пролиферации клеток гладких мышц, и воспалительным состоянием, связанным с микотическими, вирусными, паразитарными или бактериальными инфекциями, родственный Zot иммунорегулятор вводят один или в комбинации со специфическим связанным с воспалением антигеном или аллергеном. Примеры специфических воспалительных антигенов, ассоциируемых с воспалительными или аллергическими заболеваниями или нарушениями, являются пыльца и пыль (ассоциируемые с астмой), белки, обнаруженные в коровьем молоке, или их фрагменты (ассоциируемые с экзематозным дерматитом), основной миелиновый белок (ассоциируемый с рассеянным склерозом) и антигены, используемые для приготовления вакцин против отдельных вирусов, паразитов или бактерий, ассоциированных с воспалительным состоянием.
Настоящее изобретение делает возможной антиген-специфическую негативную модуляцию иммунного ответа. Как обсуждалось выше, один вариант данного изобретения включает введение эффективного количества одного из Zot-иммунорегуляторов (Zot или зонулина), одного или в сочетании со специфическим антигеном, при этом реакция лимфоцитов на указанный антиген специфически подавляется в зависимости от дозы. Из результатов, подробно представленных в данном описании, ясно, что данное изобретение ограничивается теми антигенами, процессируемыми и "презентируемыми" антиген-презентирующими (представляющими) клетками (АРС), для которых нужно "посредничество" макрофага. Изобретение относится ко всем АРС, включая макрофаги и другие мононуклеарные фагоциты, дендритные клетки, В-лимфоциты, клетки Лангерганса и эндотелиальные клетки человека. В предпочтительном варианте изобретения АРС представляет собой макрофаг.
Настоящее изобретение можно использовать как в in vivo, так и в in vitro средах. Единственно важно - это введение в животные клетки, причем указанные клетки находятся либо в живом хозяине, либо в клеточной культуре. Животные клетки определяются как ядросодержащие, не содержащие хлоропласта клетки из многоклеточных организмов или в многоклеточных организмах, таксономическое положение которых находится в царстве животных, первичная клеточная культура, культура эксплантата или трансформированная клеточная линия.
Животные-реципиенты, или животные-хозяева, используемые по данному изобретению, не являются (важными) критическими, и клетки для этой цели включают клетки во вех организмах или из всех организмов царства животных. В предпочтительном варианте изобретения используются животные семейства млекопитающих. Предпочтительными животными являются млекопитающие, а предпочтительными клетками являются клетки млекопитающих, такие как клетки человека, коровы, овцы, свиньи, кошки, буйвола, собаки, козы, лошади, осла, оленя или приматов. Наиболее предпочтительным животным является человек, наиболее предпочтительными животными клетками являются клетки человека.
Конкретные формы и способ введения не являются решающими (важными) по данному изобретению. Единственно важным является то, что как Zot-иммунорегулятор, так и антиген достигают макрофага интактными. В контексте данного изобретения Zot-иммунорегулятор может вводиться вместе с антигеном. Или же их можно вводить последовательно. Ради простоты дальнейшее обсуждение способа введения и фармацевтического препарата относится к введению и препарату антигена. Однако, ясно, что те же способы применяют к введению Zot-иммунорегулятора. Кроме того, хотя обсуждение ограничивается введением одного антигена, ясно, что можно вводить более одного антигена, при этом затем негативно модулируется иммунный ответ на более чем один антиген одновременно.
Для спешного введения требуется доставка в богатую АРС среду (окружение). Предпочтительные способы введения включают такие, которые нацелены на участки иммунной системы, такие как слизистая оболочка или лимфоидные ткани. Следовательно, предпочтительными способами введения являются назальный, внутриглазной, внутрикишечный и интравагинальный. Это не исключает того, что парентеральное введение также как внутрикожное, внутримышечное, подкожное или внутривенное, также может быть эффективным способом введения Zot или зонулина, одного или в сочетании с предпочтительным(и) антигеном (-ами).
В зависимости от конкретного способа введения лекарственная форма может быть твердой, полужидкой или жидкой. Лекарственная форма может содержать добавки, смазки, стабилизаторы, буферы, покрытия и эксципиенты, обычные в технике фармацевтических рецептур.
Что касается способа введения, то антиген можно принимать в виде доз лекарственных препаратов для перорального приема с целью доставки в тонкий кишечник. Такие лекарственные препараты для перорального приема с целью доставки в тонкий кишечник хорошо известны в технике и обычно представляют собой устойчивые в желудочной среде таблетки или капсулы (Remington's Pharmaceutical Sciences, 16th Ed. , Eds. Osol, Mack Publishing Co., Chapter 89 (1980); Digenis et al., J. Pharm. Sci., 83:915-921 (1994); Vantini et al., Clinica Terapeutica, 145: 445-451 (1993); Yoshitomi et al., Сhem. Pharm. Bull. , 40: 1902-1905 (1992); Thoma et al., Pharmazie, 46:331-336 (1991); Morishita et al., Drug Design and Delivery, 7:309-319 (1991) и Lin et al., Pharmaceutical Res., 8:919-924 (1991)).
Таблетки делают устойчивыми в желудке, добавляя, например, либо ацетатфталат целлюлозы, либо ацетаттерефталат целлюлозы.
Капсулы представляют собой твердые лекарственные формы, в которых антиген заключен в твердый или мягкий растворимый контейнер или желатиновую оболочку. Желатин, применяемый в производстве капсул, получают из коллагена гидролизом. Существует два вида желатина. Тип А, получаемый из шкуры свиней обработкой кислотой, и тип В, получаемый из костей и шкуры животных щелочной обработкой. Применение твердых желатиновых капсул позволяет, прописывая один антиген или комбинацию антигенов, выбирать точную дозировку, предположительно, лучшую для отдельного больного. Твердые желатиновые капсулы состоят из двух частей, одна в другой, таким образом полностью окружая либо только антиген, либо антиген в сочетании с иммунорегулятором. Эти капсулы заполняют, помещая антиген, либо устойчивые в желудке гранулы, содержащие антиген, в более длинном конце капсулы, а затем надевают крышку (колпачок). Твердые желатиновые капсулы делают в основном из желатина, красителей FD&C и иногда веществ, придающих непрозрачность капсуле, таких как диоксид титана. USP допускает в применяемом для этой цели желатине содержание 0,15% (вес/объем) диоксида серы для предотвращения разложения в процессе производства.
В контексте настоящего изобретения составы пероральных лекарственных форм для доставки в тонкий кишечник также включают жидкие композиции, которые содержат водные буферизующие агенты, которые предотвращают заметное инактивирование антигена жидкостью в желудке, тем самым давая возможность антигену достичь тонкого кишечника в активной форме. Примеры таких буферизующих агентов, которые можно использовать по данному изобретению, включают бикарбонатный буфер (рН 5,5-8,7, предпочтительное значение рН 7,4).
Когда пероральная лекарственная композиция является жидкой композицией, предпочтительно, чтобы композицию готовили непосредственно перед приемом, чтобы свести к минимуму проблемы стабильности. В этом случае жидкую композицию можно готовить, растворяя лиофилизированный пептидный антагонист в водном буферизующем агенте.
Также настоящим изобретением охватываются другие носители для доставки, включая, без ограничения, липосомы, "спирали" (cochleates), водорастворимые полимеры и микросферы. Композиция лекарственного вещества может дополнительно содержать адъюванты, такие как монофосфориллипид A, QS-21, ISCOMs, и цитокины.
Антиген можно также вводить в виде внутривенной лекарственной формы для доставки в системные элементы иммунной системы. Такие лекарственные формы (композиции) хорошо известны в технике, и композиции обычно содержат физиологический разбавитель, например, дистиллированную воду или 0,9% (вес/объем) NaCl. Также введение может быть парентеральным, внутрикожным, внутримышечным или подкожным и упоминалось выше. Лекарственные формы для такого введения, несомненно, содержат фармацевтически приемлемые добавки, включая физиологические буферы, разбавители или подобные соединения.
Употребляемое в данном описании выражение "эффективное количество" Zot-родственного иммунорегулятора, такого как Zot или зонулин, относится к количеству, эффективному для негативной модуляции активности указанной антиген-презентирующей клетки, тем самым эффективному для негативной модуляции опосредуемой антиген-представляющей клеткой пролиферации лимфоцитов. Конкретное количество антигена и Zot-родственного иммунорегулятора не является "критическим" и меняется в зависимости от заболевания или состояния, подвергаемого терапии, а также от возраста, веса и пола больного. Обычно, чтобы достичь такой конечной концентрации, например, в кишечнике или крови, количество Zot-родственного иммунорегулятора в однократной дозе композиции для перорального введения по данному изобретению составляет около 0,1-100 мкг, предпочтительно около 2,0-60 мкг, более предпочтительно около 20-50 мкг. Также количество антигена в разовой дозе для перорального приема обычно должно находиться в интервале около 0,01-1000 мкг, более предпочтительно около 0,1-100 мкг. Очевидно, что точная доза антигена меняется в зависимости от подвергаемого терапии заболевания или состояния, при этом предпочтительные интервалы легко определяются с помощью обычных экспериментов и в процессе оптимизации.
Нижеследующие примеры даны только с иллюстративной целью и ни в коей мере не претендуют на ограничение объема настоящего изобретения.
ПРИМЕР 1
Связывания FITC-меченого Zot с лимфоцитами и макрофагами
А. Материлы и методы
Выделение мононуклеаров периферической крови (РВМС) из крови здоровых добровольцев
РВМС выделяют центрифугированием в градиенте плотности в средах для фракционирования лимфоцитов (LSM, Organon-Teknika, Durham, NC) из крови здоровых добровольцев. Доноры являются взрослыми и они дали осознанное согласие на взятие крови. Используют РВМС свежие или аликвоты их замораживают в RPMI, содержащей 10 об.% FCS (ФТС) и 10 об.% ДМСО, используя морозильный аппарат с регулируемой линейной скоростью замораживания (1oС в мин, Planner Biomed, Salisbury, England) для сохранения жизнеспособности клеток и с тем, чтобы максимизировать регенерацию клеток. Клетки хранят в жидком азоте до использования. В некоторых опытах клетки используют непосредственно после выделения.
Приготовление очищенного ZOT-MBP
5000 мл супернатантной фракции, полученной после культивирования штамма CVD110 V. cholerae (Michalski et al., Infect. Immun., G1:4462-4468 (1993)), трансформированного плазмидой pZ14, концентрируют в 100 раз с помощью пластинчатого проточного фильтра со срезом MB 10 kDa. Конструкция pZ14, содержащая ген zot Vibrio cholera, подробно описана в, среди прочего, WO 96/37196. Образующийся супернатант затем подвергают 8% (вес/объем) SDS-PAGE. Полосы белков обнаруживают по окрашиванию геля SDS-PAGE Кумасси синим. При сравнении с контрольным супернатантом штамма CVD110, трансформированного плазмидой pTTQ181 (Amersham, Arlington Heights, IL) и обработанного тем же способом, не обнаружено никакой полосы белка, соответствующей Zot. Следовательно, даже несмотря на то, что в pZ14 ген zot помещен вслед за высокоиндуцибельным и сильным промотором tac, уровень белка в тысячекратно концентрированном супернатанте pZ14 все еще не обнаруживается при окрашивании геля SDS-PAGE Кумасси синим.
Отсюда для увеличения количества продуцированного Zot ген zot сливают в рамке (считывания) с геном мальтозусвязующего белка (далее в настоящем описании "МВР") с целью получения слитого белка MBP-ZOT.
МВР-вектор pMAL-c2 (Biolab) используют для экспрессии и очистки Zot, сливая zot-ген с геном malE E.coli. Эта конструкция использует сильный, индуцибельный tec-промотор и сигналы инициации трансляции malE, дающие высокий уровень экспрессии клонированного гена zot. Вектор pMAL-c2 имеет точную делецию сигнальной последовательности malE, которая приводит к цитоплазматической экспрессии слитого белка. Очистку МВР аффинной хроматографией используют для упрощения выделения слитого белка (Biolab).
Более конкретно, вектор pMAL-c2 линеаризуют с помощью EcoRI (которая разрезает по 3' концу гена malE), заполняют фрагментом Кленова и гидролизуют с помощью ХbаI (которая имеет единственный сайт в полилинкере pMAL-c2). orf, кодирующий ZOT, субклонируют из плазмиды рВВ241 (Baudry et al., Infect. Immun. , 60:428-434 (1992)). Плазмиду рВВ241 расщепляют с помощью BssHII, заполняют фрагментом Кленова и расщепляют с помощью ХbаI. Затем ХbaI-фрагмент с тупым концом субклонируют с pMAL-c2, получая плазмиду pLC10-c. Так как и инсерт, и вектор имеют тупой и липкий концы, правильную ориентацию получают с 3' концом malE, слитым с 5'-концом инсерта. Затем проводят электропорацию pLC10-c в штамм DH5a Е. coli. В рВВ241 BssHII-сайт рестрикции находится в zot orf. Следовательно, аминокислоты 1-8 ZOT отсутствуют в слитом белке MBP-ZOT.
Чтобы очистить слитой белок MBP-Zot, 10 мл бульона Луриа Бертани (Luria Bertani), содержащего 0,2% (вес/объем) глюкозы и 100 мкг/мл ампициллина, инкубируют с одиночной колонией, содержащей pLC10-c, термостатируют в течение ночи при 37oС при встряхивании. Культуру разбавляют 1:100 в 1,0 мл той же свежей среды и выращивают, встряхивая, при 37oС, примерно, до 1,0•108 клеток/мл. Затем добавляют 0,2 мМ IPTG (изопропилтиогалактозу) для стимуляции экспрессии MBP-Zot и культуру инкубируют при 37oС еще 3 ч. Затем бактерии осаждают центрифугированием и ресуспендируют в 20 мл охлажденного льдом "буфера для колонки", содержащего 20 мМ Tris-HCl, 0,2 М NaCl, 1,0 мМ EDTA (ЭДТК), 10 мМ 2-МЕ, 1,0 мМ NаN3. Бактериальную суспензию лизируют обработкой с помощью пресса Френча и центрифугируют в течение 30 мин при 13000 • g при 4oС. Супернатант собирают, разбавляют 1:5 буфером для колонки и загружают на 1•10 колонку с амилозной смолой (Biolabs, система очистки МВР-слияния), предварительно уравновешенной с буфером для колонки. После промывания колонки 5 объемами "колоночного" буфера слитой белок MBP-ZOT элюируют, добавляя в "колоночный" буфер 10 мл 10 мМ мальтозы. Обычно выход из 1,0 мл культуры составляет 2-3 мг белка.
Партнер МBР, слитый с ним, в очищенном слитом белке затем отщепляется с помощью 1,0 мкг Фактора Ха-протеазы (Biolabs) на 20 мкг MBP-Zot. Фактор Ха-протеаза расщепляет непосредственно перед аминоконцом Zot. Полученный при этом белок Zot подвергают электрофорезу на 8,0% (вес/объем) геле SDS-PAGE и электроэлюции с геля, применяя камеру для электропорации (Schleicher and Schuell, Keen, NH).
При испытании в камерах Уссинга (Ussing) полученный очищенный Zot стимулировал снижение Rt в зависимости от дозы при ED50 7,5•10-8 М.
Конъюгация (мечение) ZOT-MBP с флуоресцеинизотиоцианатом (ZOT-FITC)
Конъюгацию Zot-MBP c FITC (мечение) осуществляют по стандартной методике. Коротко говоря, Zot-MBP подвергают диализу против 500 мл буфера для FIТС-мечения, содержащего 0,1 М бикарбонатного буфера (например, 0,09 М NаНСО3+0,0085 М Nа2СО3), доводят до рН 9,0 концентрированной NaOH и хранят при 4oС, при 4oС за 8 ч рН повышают до 9,0. Затем добавляют по 10 мкл раствора FITC в ДМСО с концентрацией 5,0 мг/мл на каждый миллиграмм MBP-ZOT с последующей инкубацией в течение ночи в PBS при 4oС. Затем несвязанный FITC удаляют диализом в 500 мл буфера для диализа, содержащего PBS (рН 7,4), хранят при 4oС, при 4oС в течение 2 дней меняют два-три раза. Этот препарат хранят при 4oС до использования.
Связывание FITC-ZOT с РВМС человека и анализ методом проточной цитометрии
РВМС, выделенные описанным выше способом, термостатируют при повышении концентраций Zot-FITC 60 мин при 37oС в покрытых кремнием пробирках (для предупреждения связывания макрофагов со стенками аналитической пробирки) в присутствии моноклональных антител к CD14, конъюгированным с фикоэритрином (РЕ), и к CD3, конъюгированным с ECD (энергетически связанный краситель, конъюгат PE-Texas-Red). CD14 является маркером моноцитов/макрофагов человека, тогда как CD3 является маркером Т-лимфоцитов. Применение этих флуорохромов (например, FITC, РЕ и ECD) позволяет одновременно изучать связывание ZOT с моноцитами/макрофагами и Т-лимфоцитами в смешанных РВМС-популяциях методом 3-цветной проточной цитометрии. Затем клетки дважды отмывают PBS (рН 7,2), содержащей 1,0% (вес/объем) BSA и 0,1% (вес/объем) азида натрия, и сразу же анализируют проточной цитометрией, используя систему Epics Elite проточный цитометр/клеточный сортер.
В этих экспериментах в качестве контроля используют меченые флуорохромом mAb тех же изотопов, но нерелевантной специфичности. Тромбоциты, эритроциты (если таковые имеются) и клеточный дебрис исключают из анализа, устанавливая соответствующее окно в прямом направлении против параметра - 90% рассеяния света. Для каждого образца собирают данные для более чем 10000 клеток. Анализ данных осуществляют с помощью пакета аналитических программ Epics Elite (Coulter) или list-mode пакета аналитических программ WinList (Verity Software House, Topsham, ME).
Б. Результаты
Типичный опыт, иллюстрирующий анализ методом проточной цитометрии Zot-FITC-связывание с Т-лимфоцитами (СD3+) и моноцитами/макрофагами (CD14+) человека, изображен на фиг. 1. Результаты показывают, что связывание Zot-FITC, как явствует из средних уровней интенсивности флуоресценции, в несколько раз выше в моноцитах/макрофагах, чем в Т-лимфоцитах. Кроме того, добавление возрастающих количеств Zot-FITC приводит к увеличению связывания, которое начинает выравниваться после добавления 40-60 мкл Zot-FITC. Эти результаты указывают, что Zot предпочтительно связывается с моноцитами/макрофагами человека.
Далее, изучают связывание Zot-FITC с моноцитами/макрофагами и лимфоцитами в присутствии немеченого Zot с целью определения, может ли немеченый Zot блокировать связывание. Как показано на фиг. 2, предварительное термостатирование РВМС в течение 30 мин при 37oС со 100 мкл немеченого Zot с последующим добавлением ZOT-FITC (10 мкл) в течение 30 мин при 37oС, примерно на 33% уменьшает связывание Zot-FITC как с моноцитами/макрофагами, так и с Т-лимфоцитами, подтверждая, что связывание Zot с этими клетками опосредуется рецепторами. Преинкубация клеток со 100 мкл очищенного МBР с последующим добавлением ZOT-FITC (10 мкл) в течение 30 мин при 37oС не оказывает влияния на блокирование Zot-FITC-связывания, доказывая, что блокирование немеченого Zot является специфическим феноменом.
ПРИМЕР 2
Пролиферативная реакция мононуклеаров человека на митогены и антигены
А. Материалы и методы
Выделение мононуклеаров периферической крови человека (РВМС) здоровых добровольцев
РВМС выделяют центрифугированием в градиенте плотности в средах для фракционирования лимфоцитов (LSM, Organon-Teknika, Durham, NC) из крови здоровых добровольцев. В соответствии с постановлением надзирающего органа Университета штата Мериленд, Балтимор, доноры являются взрослыми и они дали осознанное согласие на взятие крови. Используют РВМС свежие или аликвоты их замораживают в RPMI, содержащей 10 об.% FCS (ФТС) и 10 об.% ДМСО, используя морозильный аппарат с регулируемой линейной скоростью замораживания (1oС в мин, Planner Biomed, Salisbury, England) для сохранения жизнеспособности клеток и с тем, чтобы максимизировать регенерацию клеток. Клетки хранят в жидком азоте до использования. В некоторых опытах клетки используют непосредственно после выделения.
Приготовление очищенного Zot
Ген zot амплифицируют методом PCR с полимеразой DeepVent (New England Biolabs), используя рВВ241-плазмидную (Baudry et al., см. выше) ДНК в качестве темплата. Используются следующие прямой и обратный праймеры: 5'-CGGGATCCCGTATGAGTATCTTT-3' (SEQ ID N0:3) и 5'-CCCAAGCTTGGGTCAAAATATACT-3' (SEQ ID N0: 4), соответственно. Хвосты 5' этих олигонуклеотидов содержат сайты рестрикции BamHI и HindIII, соответственно. Образующийся ампликон (1,2 тыс. п. о.) анализируют электрофорезом на 8,0% (вес/объем) агарозном геле и очищают от солей и свободных нуклеотидов с помощью вращающейся колонки Xtreme (Pierce).
Вышеотмеченные две рестриктазы используются затем для расщепления очищенного ампликона, и образующийся расщепленный ампликон затем встраивают в вектор pQE30 (Quiagen), который предварительно гидролизован с помощью ВатHI и HindIII, так чтобы получить плазмиду pSU113. pQE30 представляет собой вектор экспрессии, который обеспечивает экспрессию рекомбинантного белка с 6-полигистидин tag (6xHis) на высоком уровне. Продукт плазмиды pSU113, следовательно, есть слитый белок 6xHis-Zot. Затем pSU113 трансформируют в Е.coli DH5a.
Для очистки слитого белка 6xHis-Zot результирующие трансформированные Е. coli выращивают в течение ночи при 37oС в 150 мл бульона Луриа Бертани (Luria Bertani), содержащего 2,0% (вес/объем) глюкозы, 25 мкг/мл канамицина и 200 мкг/мл ампициллина так, чтобы A600 был около 1,10. Далее, 75 мл культуры, выращиваемой в течение ночи, добавляют к 1000 мл бульона Луриа Бертани, содержащего 2,0% (вес/объем) глюкозы, 25 мкг/мл канамицина и 200 мкг/мл ампициллина, термостатируют около 3 ч при 37oС при интенсивном встряхивании до A600 около 0,7-0,9. Затем добавляют IPTG до конечной концентрации 2,0 мМ, и продолжают выращивание в течение 5 ч при 37oС. Далее клетки собирают, центрифугируя при 4000 • g в течение 20 мин, клетки ресуспендируют в 5,0 мл/г сырого веса буфера А, содержащего 6,0 М GuHCl, 0,1 М фосфата натрия, и 0,01 М Tris-HCl (pH 8,0), и перемешивают в течение 1 ч при комнатной температуре. Затем смесь центрифугируют при 10000 • g в течение 30 мин при 4oС, и к результирующему супернатанту добавляют 4,0-5,0 мл/г сырого веса 50% суспензии смолы SUPERFLOW (QIAGEN) и перемешивают в течение 1 ч при комнатной температуре. Полученную в результате смолу нагружают на колонку 1,6•8,0, которую затем промывают последовательно буфером А, буфером В, содержащим 8,0 М мочевины, 0,1 М фосфата натрия и 0,01 М Tris-HCl (pH 8,0), и буфером С, содержащим 8,0 М мочевины, 0,1 М фосфата натрия и 0,01 М Tris-HCl (pH 6,3). Каждое промывание проводят до тех пор, пока A600 промывного раствора не станет менее 0,01. Слитый белок 6xHis-ZOT элюируют с колонки, используя 20 мл буфера С, содержащего 250 мМ имидазола. Затем, фракции, содержащие слитый белок 6xHis-ZOT, проверяют с помощью SDS-PAGE, используя методику, описанную Davis, Ann. N.Y. Acad. Sci., 121:404 (1964), и гель окрашивают Кумасси синим. Фракции, содержащие слитый белок 6xHis-ZOT, подвергают диализу против 8,0 М мочевины, объединяют, а затем разбавляют в 100 раз в PBS. Далее, добавляют 4,0 мл 50% суспензии смолы SUPERFLOW, перемешивание проводят в течение 2 ч при комнатной температуре и образовавшуюся смолу загружают на колонку 1,6•8,0, которую затем промывают 50 мл PBS. Слитый белок 6xHis-Zot элюируют с колонки 10 мл PBS, содержащего 250 мМ имидазола. Образующийся элюент (с элюатом) подвергают диализу против PBS, и слитый белок 6xHis-ZOT проверяют по SDS-PAGE, как описано выше.
Приготовление и очистка кроличьей антисыворотки против Zot
Для получения специфической антисыворотки экспрессируют и очищают белок глутатион-S-трансфераза (GST)-Zot.
Более конкретно, для амплификации zot orf полимеразной цепной реакцией (PCR) с применением плазмиды рВВ241 (Baudry et al., см. выше) в качестве темплатной ДНК используют олигонуклеотидные праймеры. Прямой праймер (TCATCACGGC GCGCCAGG, SEQ ID N0:5), соответствующий нуклеотидам 15-32 zot orf, и обратный праймер (GGAGGTCTAG AATCTGCCCG AT, SEQ ID N0:6), соответствующий 5'-концу ctxA orf. Следовательно, аминокислоты 1-5 ZOT отсутствуют в полученном слитом белке. Продукт амплификации встраивают в полилинкер (сайт SmaI), расположенный на конце гена GST в pGEX-2T (Pharmacia, Milwaukee, WI). pGEX-2T является вектором экспрессии слитого белка, который экспрессирует клонированный ген в виде слитого белка с GST Schistosoma japonicum. Слитый ген находится под контролем промотора tac. Вследствие индуцирования с помощью IPTG происходит депрессия и экспрессирует GST-слитый белок.
В результате электропорации из результирующей рекомбинантной плазмиды, названной pLC11, получают Е. coli DH5a. Для очистки слитого белка GST-Zot 10 мл бульона Луриа Бертани, содержащего 100 мкг/мл ампициллина, инкубируют с одиночной колонией, содержащей pLC11, и термостатируют в течение ночи при 37oС при встряхивании. Культуру разбавляют 1: 100 в 1,0 мл той же самой свежей среды и выращивают, встряхивая, при 37oС, примерно, до 1,0•108 клеток/мл. Затем добавляют 0,2 мМ IPTG для стимулирования экспрессии GST-Zot и культуру термостатируют при 37oС еще 3 ч. Затем бактерии высаживают центрифугированием, ресуспендируют в 20 мл охлажденного льдом PBS (рН 7,4) и лизируют с помощью пресса Френча. Слитый белок GST-Zot нерастворим в этих условиях, поэтому он осаждается вместе с бактериями. Далее, ресуспендируют в буфере Лэммли для лизирования, содержащем 0,00625 М 0,01 М Tris-HCl (рН 6,8), 0,2 М 2-МЕ, 2,0% (вес/объем) SDS, 0,025% (вес/объем) бромфенолового синего и 10 об.% глицерина, и подвергают электрофорезу на 8,0% (вес/объем) геле SDS-PAGE и окрашивают Кумасси синим. Полоса около 7 kDa (26 kDa GST + 44 kDa Zot), соответствующая слитому белку, подвергается электроэлюции с геля с применением электроразделительной камеры (Schleicher and Schuell, Keen, NH).
10 мкг образующегося элюированного белка (10-20 мкг) инъецируют кролику вместе с равным объемом полного адъюванта Фрейнда. Бустерные дозы вводят с неполным адъювантом Фрейнда через четыре и восемь недель. Через месяц кролика обескровливают.
Для определения получения специфических антител, 10-10 М Zot вместе с двумя слитыми белками, MBP-Zot и GST-Zot переносят на нейлоновую мембрану и термостатируют с кроличьей антисывороткой при разведении 1:5000 в течение ночи при 4oС и умеренном встряхивании. Затем фильтр промывают 15 мин 4 раза PBS, содержащей 0,05 об.% Tween 20 (далее в настоящем описании "PBS-T"), и термостатируют при разведении 1:30000 с антикроличьим IgG козы, конъюгированным с пероксидазой хрена, в течение 2 ч при комнатной температуре. Фильтр снова промывают в течение 15 мин 4 раза PBS, содержащим 0,1 об.% Tween, и определяют иммунореактивные зоны с помощью усовершенствованной хемилюминесценции (Amersham).
Иммуноблотингом было обнаружено, что кроличья антисыворотка распознает Zot, так же как слитые белки MBP-Zot и GST-Zot, но не МВР-негативный контроль.
Кроме того, для подтверждения образования соответствующих антител против Zot опыты по нейтрализации проводили в камерах Уссинга. После преинкубации с супернатантом pZ14 при 37oС в течение 60 мин Zot-специфическая антисыворотка (разведение 1:100) способна полностью нейтрализовать снижение Rt, стимулированное Zot в подвздошной кишке кролика, помещенной в камере Уссинга.
Далее, антитела против Zot очищают аффинной хроматографией на MBP-Zot-аффинной колонке. Более конкретно, аффинную колонку MBP-Zot готовят, иммобилизуя в течение ночи при комнатной температуре 1,0 мг очищенного MBP-Zot, полученного как описано выше в примере 1, на предварительно активированном геле (Aminolink, Pierce). Колонку промывают PBS и затем нагружают 2,0 мл кроличьей антисыворотки против ZOT. Через 90 мин инкубации при комнатной температуре колонку промывают 14 мл PBS и специфические антитела против Zot элюируют с колонки с помощью 4,0 мл раствора, содержащего 50 мМ глицина (рН 2,5), 150 мМ NaCl и 0,1 об.% Triton Х-100. рН 1,0 мл элюированных фракций сразу же нейтрализуют 1,0 N раствором NaOH.
Условия культивирования и анализы лимфопролиферации
РВМС (1,5•106 клеток/мл) культивируют в 1,0 мл полной среды (cRPMI), представляющей собой RPMI 1640, содержащей 10 об.% фетальной бычьей сыворотки и 50 мкг/мл гентамицина. Клетки термостатируют при 37oС, 5% СО2 в 96-луночных планшетах в отсутствие или в присутствии фитогемагглютинина (РНА, неспецифический митоген, применяемый в количестве 2,0 мкг/мл) или столбнячного токсина (ТТ, специфический антиген, в количестве 2,0 мкг/мл; Connaught, Swift Water, PA) без или с очищенным ZOT (в количестве 20 или 60 мкг/мл) или альбумина бычьей сыворотки (BSA; контрольный белок (в количестве 20 или 60 мкг/мл; Fraction V, Sigma, St. Louis, МО). В некоторых экспериментах к культурам при инициации также добавляют кроличью иммунную сыворотку или нормальную кроличью сыворотку к ZOT. Клетки культивируют 2 дня (в случае РНА) или 6 дней (в случае ТТ) и добавляют 1,0 мкКи/лунка 3H-тимидина. Клетки на планшетах собирают через 20 ч в харвестер клеток Уоллака (Wallac) (Gaithersburg, MD) и вносят тимидин, измеренный счетчиком Wallac Trilux Microbeta counter (Gaithersburg, MD).
Б. Результаты
Типичный эксперимент, показывающий влияние очищенного Zot на пролиферацию РВМС человека, индуцированную РНА и столбнячным токсином, проиллюстирован на фиг. 3. Результаты ясно указывают, что инкубация с Zot заметно подавляет вызванную столбнячным токсином пролиферацию (~85% при 60 мкг/мл), хотя оно не оказывает влияния на РНА-индуцированную пролиферацию. Кроме того, подавление ТТ-индуцированной пролиферации под действием Zot, по-видимому, зависит от дозы. Значительно более высокие уровни супрессии наблюдаются, когда ZOT добавляют в количестве 60 мкг/мл (~85%), чем при 20 мкг/мл Zot (супрессия ~56%). Добавление BSA, используемого в качестве контрольного белка, не оказывает никакого влияния ни на ТТ-, ни на РНА-стимулированную пролиферацию лимфоцитов, что демонстрирует специфичность биологической активности Zot.
Для дальнейшего изучения специфичности Zot-стимулированной супрессии ТТ-индуцированной пролиферации, РВМС термостатируют в отсутствие или в присутствии одних ТТ и ZOT, ZOT + кроличья иммунная сыворотка к ZOT (в разведении 1: 10) или ZOT + нормальная кроличья сыворотка (в разведении 1: 10). Как видно на фиг. 4, добавление кроличьей иммунной сыворотки против (к) Zot реверсирует более чем на 50% Zot-опосредованную супрессию ТТ-индуцированной пролиферации. Добавление нормальной кроличьей сыворотки не дает эффекта, что подтверждает специфичность Zot-опосредуемых эффектов. Сходным образом добавление BSA не оказывает влияния в этой системе.
ПРИМЕР 3
Влияние Zot на поглощение FITC-декстран моноцитами и макрофагами человека
А. Материлы и методы
Выделение мононуклеаров периферической крови (РВМС) человека из крови здоровых добровольцев
РВМС выделяют центрифугированием в градиенте плотности в средах для фракционирования лимфоцитов (LSM, Organon-Teknika, Durham, NC) из крови здоровых добровольцев. В соответствии с постановлением надзирающего органа Университета штата Мериленд, Балтимор, доноры являются взрослыми и они дали осознанное согласие на взятие крови. Используют РВМС свежие или аликвоты их замораживают в RPMI, содержащей 10 об.% FCS (ФТС) и 10 об.% ДМСО, используя морозильный аппарат с регулируемой линейной скоростью замораживания (1oС в мин, Planner Biomed, Salisbury, England) для сохранения жизнеспособности клеток и с тем, чтобы максимизировать регенерацию клеток. Клетки хранят в жидком азоте до использования.
Приготовление очищенного 6xHis-Zot
Слитый белок 6xHis-Zot готовят и очищают методом, описанным выше в примере 2.
Поглощение растворимого антигена
Способность поглощать растворимый антиген измеряют, используя декстран, конъюгированный с флуоресцеинизотиоцианатом (FITC) (м. в. 50700; Sigma). Свежевыделенные РВМС (500000 клеток в 0,5 мл cRPMI, содержащей 10 об.% термоинактивированной FCS и 50 мкг/мл гентамицина), инкубируют (термостатируют) в отсутствие или в присутствии очищенного Zot (40 мкг/мл) или BSA (40 мкг/мл) в течение 3 ч при 37oС в конечном объеме 0,5 мл/50 мл пробирка. Эту инкубацию осуществляют при встряхивании в покрытых кремнием пробирках для предотвращения прилипания моноцитов/макрофагов к стенкам пробирок. После такой инкубации моноклональные антитела против CD14, меченые аллофикоцианином (АРС), и против HLA-DR, меченые перидининхлорофилльным белком (РеrСР), добавляют в каждую культуру без отмывания, и клетки инкубируют в течение 30 мин при 37oС или на льду. Так как поглощение FITC-декстрана, и растворимых антигенов в целом зависит от пиноцитоза, зависящего от температуры феномена, то инкубацию при 0oС осуществляют для того, чтобы установить уровни неспецифического связывания FITC-декстрана с клетками.
В этих экспериментах для идентификации моноцитов/макрофагов используют CD14 и HLA-DR (антиген класса II главного комплекса гистосовместимости, уровень экспрессии которого повышается при активации клеток).
Затем клетки отмывают один раз ледяной PBS и немедленно переносят в систему проточный цитометр/клеточный сортер Coulter Elite Epics (Coulter Corp., Miami, FL). Анализ осуществляют, используя программное обеспечение WinList (Verity Software House, Topsham, ME). Процент клеток, которые включают FITC-декстран, получают, вычитая процент клеток, которые включают FITC-декстран при 0oС (лед), из процента клеток, которые включают FITC-декстран при 37oС.
Б. Результаты
Типичный эксперимент, показывающий действие Zot на поглощение FITC-декстрана нормальными CD14+ и HLA-DR+ моноцитами/макрофагами человека, изображен на фиг. 5A-5D. Результаты показывают, что Zot (фиг. 5D) заметно подавляет (~51-58%) поглощение FITC-декстрана моноцитами/макрофагами человека по сравнению с поглощением, наблюдаемым в клетках, инкубированных с одними средами или BSA. Не наблюдается значительной разницы между процентным содержанием клеток, которые включают FITC-декстран в средах или в присутствии BSA. Кроме того, как ожидалось, инкубация при 0oС полностью "отменяет" поглощение FITC-декстрана, подтверждая, что этот феномен зависит от температуры.
Надежно установлено, что поглощение антигена АРС, такими как моноцитами/макрофаги, является критическим событием, ведущим к активации лимфоцитов и пролиферации (Sztein et al., см. выше (1997)). Результаты показывают, что Zot препятствует поглощению FITC-декстрана, демонстрируют, что иммунорегуляторное действие Zot на ТТ-индуцированную пролиферацию опосредуется, по меньшей мере, частично, уменьшением способности моноцитов/макрофагов поглощать антиген, что приводит к изменению процессирования и презентации антигена. Это дополнительно подтверждается тем фактом, что Zot не влияет на РНА-стимулированную пролиферацию, феномен, который не требует процессирования и презентации антигена.
ПРИМЕР 4
Измерение числа сайтов связывания FITC-Zot/в клетке в моноцитах/макрофагах и лимфоцитах человека
А. Материлы и методы
Выделение мононуклеаров периферической крови человека (РВМС) здоровых добровольцев
РВМС выделяют центрифугированием в градиенте плотности в средах для фракционирования лимфоцитов (LSM, Organon-Teknika, Durham, NC) из крови здоровых добровольцев. В соответствии с постановлением надзирающего органа Университета штата Мериленд, Балтимор, доноры являются взрослыми и они дали осознанное согласие на взятие крови. Аликвоты РВМС замораживают в RPMI, содержащей 10 об.% FCS (ФТС) и 10 об.% ДМСО, используя морозильный аппарат с регулируемой линейной скоростью замораживания (1oС в мин, Planner Biomed, Salisbury, England) для сохранения жизнеспособности клеток и с тем, чтобы максимизировать регенерацию клеток. Клетки хранят в жидком азоте до использования. В некоторых опытах клетки используют непосредственно после выделения.
Приготовление очищенного 6xHis-Zot
6xHis-Zot готовят, как описано выше в примере 2.
Мечение (конъюгация) Zot (с) флуоресцеинизотиоцианатом (Zot-FITC)
Конъюгацию Zot с FITC осуществляют в соответствии со стандартными методиками. Коротко говоря, Zot диализуют против 500 мл буфера для FITC-мечения, содержащего 0,05 М борной кислоты, 0,2 М NaCl, доводят рН до 9,2 концентрированной NaOH и хранят при 4oС, при 4oС накануне удаляют свободные ионы NF4 + и повышают рН до 9,2. Затем добавляют 20 мкл 5,0 мг/мл FITC в ДМСО на каждый миллиграмм Zot с последующей инкубацией в течение 2 ч при комнатной температуре. Затем несвязанный FITC удаляют диализом в 500 мл буфера для диализа, содержащего 0,1 М Tris-HCl (рН 7,4), 0,1% (вес/объем) NаN3, 0,2 M NaCl, доведенного до рН 7,4 концентрированной NaOH, и хранят при 4oС, с двумя-тремя сменами через 2 дня при 4oС. Этот препарат хранят при 4oС до использования.
Связывание FITC-ZOT с РВМС человека и анализ методом проточной цитометрии
РВМС, выделенные описанным выше способом, термостатируют при повышении концентраций Zot-FITC 60 мин при 37oС в покрытых кремнием пробирках (для предупреждения связывания макрофагов со стенками аналитической пробирки) в присутствии моноклональных антител (mAb) к CD14, конъюгированным с фикоэритрином (РЕ), и к CD3, конъюгированным с ECD (энергетически связанный краситель, конъюгат PE-Texas-Red). CD14 используется как маркер макрофагов человека, тогда как CD3 является маркером Т-лимфоцитов. Применение этих флуорохромов (например, FITC, РЕ и ECD) позволяет одновременно изучать связывание ZOT с макрофагами и Т-лимфоцитами в смешанных РВМС-популяциях методом трехцветной проточной цитометрии. После окрашивания клетки дважды отмывают PBS (рН 7,2), содержащей 1,0% (вес/объем) BSA и 0,1% (вес/объем) азида натрия, и сразу же анализируют проточной цитометрией, используя систему Epics Elite проточный цитометр/клеточный сортер (Beckman-Coulter, Miami, FL).
В этих экспериментах в качестве контроля используют меченые флуорохромом mAb тех же изотопов, но нерелевантной специфичности. Тромбоциты, эритроциты (если таковые имеются) и клеточный дебрис исключают из анализа, устанавливая соответствующее окно в прямом направлении против параметра - 90% рассеяния света. Для каждого образца собирают данные для более чем 10000 клеток. Анализ данных осуществляют с помощью пакета аналитических программ Epics Elite (Coulter) или list-mode пакета аналитических программ WinList (Verity Software House, Topsham, ME).
Расчет сайтов связывания Zot/в клетке
Среднюю интенсивность флуоресценции каждой популяции после инкубации с Zot-FITC переводят в число сайтов связывания Zot/в клетке, используя стандартную кривую, построенную с применением набора Quantum 26 MESF (интервал 10000-50000 MESF) и программным обеспечением для калибровки QuickCal в соответствии с рекомендациями производителя (Flow Cytometry Standarts Corporation, San Juan, Puerto Rico). Стандарты флуоресценции в наборе Quantum калибруют в единицах Молекул Эквивалентных Растворимых Флуорохромов (MESF) по отношению к растворам очищенных флуоресцентных красителей. Число сайтов связывания на клетку (макрофаги или лимфоциты) выводят из числа единиц MESF Zot-FITC-инкубированных образцов (за вычетом неспецифического связывания, т. е. единиц MESF контрольного FITC-меченого мышиного IgG), коррелируемого с соотношением флуоресцеин/белок (F/P) различных партий используемого FITC-Zot.
Б. Результаты
Типичный эксперимент, иллюстрирующий анализ методом проточной цитометрии Zot-FITC-связывание с Т-лимфоцитами (CD3+) и макрофагами (CD14+) человека, изображен на фиг. 6. Результаты указывают, что связывание Zot-FITC, как явствует из повышенного числа сайтов связывания Zot/в клетке, в несколько раз выше в макрофагах, чем в CD3+ Т-лимфоцитах. Кроме того, эти данные указывают, что связывание Zot является насыщаемым феноменом, при этом насыщение достигается, примерно, при 0,5 пМ (~40 г/мл). Более того, мы наблюдали, что в условиях насыщения число сайтов связывания/на клетку составляет ~ 106000 и ~9000 Zot-связывающих сайтов/на клетку, соответственно, в макрофагах и лимфоцитах человека.
Для исследования распределения Zot-связывающих сайтов в макрофагах и лимфоцитах человека используют описанную выше методологию для определения числа сайтов связывания Zot/на клетку у нескольких добровольцев. Результаты показывают, что среднее число Zot-связывающих сайтов/на клетку, в среднем, примерно в 10 раз выше в макрофагах (среднее = 104649; интервал = 56791-142840), чем в лимфоцитах (среднее = 10684; интервал 4802-18662).
ПРИМЕР 5
Кинетика связывания Zot с моноцитами/макрофагами и лимфоцитами человека
А. Материлы и методы
Выделение мононуклеаров периферической крови человека (РВМС) из крови здоровых добровольцев
РВМС выделяют из крови здоровых добровольцев, как описано выше в примере 4.
Получение очищенного 6xHis-Zot
6xHis-Zot получают, как описано выше в примере 2.
Мечение (конъюгация) Zot (с) флуоресцеинизотиоцианатом (Zot-FITC)
Конъюгацию Zot с FITC проводят, как описано выше в примере 4.
Исследование кинетики связывания
Изучение кинетики связывания Zot с клетками человека проводят с помощью проточной цитометрии, используя конъюгаты и проточную цитометрию. Образцы пропускают в реальном времени, используя систему EPIC ELITE проточный цитометр/клеточный сортер (Beckman-Coulter, Miami, FL). РВМС окрашивают с помощью меченого ECD (энергетически связанный краситель) mAb к CD3 и с помощью меченого РЕ (фикоэритрин) mAb к CD4. Контроль для фоновой флуоресценции готовят, используя дополнительную аликвоту каждой клеточной суспензии, замещая иррелевантным mAb тех же изотопов, конъюгированным с соответствующими флуоресцентными красителями, экспериментальное mAb. PBMC, меченые mAb (антителами) к CD3 и CD4, затем отмывают и хранят до анализа (в течение 1 ч) в бане со льдом, чтобы свести к минимуму модуляцию антигенов. Перед анализом клетки уравновешивают при 37oС в течение 15-20 мин на водяной бане и выдерживают при 37oС, используя устройство для жизнеспособных образцов (кинетический модуль, Cytek, Fremont, CA), присоединенный к проточному цитометру на время эксперимента (12 мин), пока непрерывно регистрируются данные. Уровни фоновой FITC-флуоресценции снимают в течение 90-150 с и делают паузу - ~10-15 с - для инъецирования Zot-FITC (до конечной концентрации 40 мкг/мл). Регистрацию (сбор) данных возобнавляют сразу же после добавления Zot-FITC со скоростью ~300-600 клеток/с - всего 12 мин. FITC, РЕ и ECD-флуорохром возбуждает охлаждаемый воздухом аргонный лазер (эмиссия при 488 нм). Результаты рассчитывают и изображают на экране с помощью пакета аналитических программ WinList и Isocontour (Verity Software House, Topsham, ME). Данные приведены в виде изометрических изображений Zot-FITC-интенсивности (ось у) во времени (ось х) относительно числа клеток (ось z) для клеток, отсортированных по CD3 (Т-лимфоциты) или по CD4 (макрофаги).
Б. Результаты
Типичный эксперимент, иллюстрирующий кинетику Zot-FITC-связывания с Т-лимфоцитами (CD3+) и макрофагами (CD14+), показан на фиг. 7А-7С. Результаты показывают, что связывание Zot с макрофагами человека (фиг. 7А) и лимфоцитами (фиг. 7В) происходит очень быстро, достигая равновесия в течение 2 мин после добавления Zot-FITC. Чтобы сравнить время, требующееся для того, чтобы достичь максимального связывания Zot и mAb к CD14, подобные эксперименты осуществляют, используя немеченые клетки. В этих экспериментах регистрируются уровни фоновой флуоресценции FITC, как описано выше, съемку данных прерывают на ~10-15 с для инъецирования FIТС-анти-СОМ mAb и сразу же возобновляют сбор данных в целом в течение 12 мин. Результаты (фиг. 7С) указывают, что максимальные уровни связывания Zot достигаются за меньшее время (~2 мин), чем требуется для того, чтобы mAb к CD14 достигли равновесия (~5 мин).
ПРИМЕР 6
Связывание FITC-Zot с В- и Т-лимфоцитами человека
А. Материлы и методы
Выделение мононуклеаров периферической крови человека (РВМС) из крови здоровых добровольцев
РВМС выделяют из крови здоровых добровольцев, как описано выше в примере 4.
Получение очищенного 6xHis-Zot
6xHis-Zot получают, как описано выше в примере 2.
Мечение (конъюгация) Zot (с) флуоресцеинизотиоцианатом (Zot-FITC)
Конъюгацию Zot с FITC проводят, как описано выше в примере 4.
Связывание FITC-Zot с РВМС человека и анализ методом проточной цитометрии
РВМС, выделенные как описано выше, инкубируют с 40 мкг/мл Zot-FITC в течение 30 мин при 37oС в присутствии mAb к CD14, конъюгированным с РЕ, CD3, конъюгированным с Tricolor (конъюгат РЕ-Су5), и к CD19, конъюгированным с ECD (энергетически связанный краситель, PE-Texas-Red-конъюгат). CD14 представляет собой маркер макрофагов человека, CD3 является маркером Т-лимфоцитов и CD19 является маркером В-лимфоцитов. Применение этих флуорохромов (т.е. FITC, РЕ, ТС и ECD) позволяет нам одновременно изучать связывание Zot с макрофагами, Т- и В-лимфоцитами в смешанных РВМС популяциях методом четырехцветной проточной цитометрии. После окрашивания клетки дважды отмывают PBS (рН 7,2), содержащей 1,0% (вес/объем) BSA и 0,1% (вес/объем) азида натрия, и сразу же анализируют проточной цитометрией, используя систему Epics Elite проточный цитометр/клеточный сортер (Beckman-Coulter, Miami, FL). В этих экспериментах в качестве контроля используют меченые флуорохромом mAb тех же изотопов, но нерелевантной специфичности. Тромбоциты, эритроциты (если таковые имеются) и клеточный дебрис исключают из анализа, устанавливая соответствующее окно в прямом направлении против параметра - 90% рассеяния света. Для каждого образца собирают данные для более чем 10000 клеток. Анализ данных осуществляют с помощью пакета аналитических программ Epics Elite (Coulter) или list-mode пакета аналитических программ WinList (Verity Software House, Topsham, ME). Результаты показаны в виде единой цветной гистограммы интенсивности флуоресценции Zot-FITC в Т (CD3+) и В (CD19+) лимфоцитселективными популяциями.
Б. Результаты
Эксперимент, иллюстрирующий связывание Zot-FITC с Т- (CD3+) и В (CD19+) лимфоцитами, показан на фиг. 8. Результаты указывают, что Zot связывается сходным образом как с Т-, так и В-лимфоцитами. В этом эксперименте связывание Zot-FITC с макрофазами в 5-8 раз выше, чем связывание с Т- и В-лимфоцитами.
ПРИМЕР 7
Неспособность антагонистов блокировать связывание Zot-FITC
А. Материалы и методы
Выделение мононуклеаров периферической крови человека (РВМС) из крови здоровых добровольцев
РВМС выделяют из крови здоровых добровольцев, как описано выше в примере 4.
Получение очищенного 6xHis-Zot
6xHis-Zot получают, как описано выше в примере 2.
Приготовление антагонистов Zot FZI/0 и FZI/1
Пептидные антагонисты FZI/0 (Gly Gly Val Leu Val Gln Pro Gly) (SEQ ID N0: 7) и FZI/1 (Val Gly Val Leu Gly Arg Pro Gly) (SEQ ID N0:8) синтезированы химически и очищены с применением хорошо известных методик, таких, которые описаны в High Performance Liquid Chromatography of Peptides and Proteins: Separation Analysis and Conformation, Eds. Mant et al., C.R.C. Press (1991), и пептидного синтезатора, такого как Symphony (Protein Technologies, Inc.).
Мечение (конъюгация) Zot флуоресцеинизотиоцианатом (Zot-FITC)
Мечение (конъюгацию) Zot-FITC осуществляют, как указано выше в примере 6.
Условия культивирования для блокирования связывания FITC-Zot с РВМС человека и проточного цитометрического анализа
РВМС, выделенные как описано выше, окрашивают mAb (антителами) к CD14, конъюгированным с РЕ, и к CD3, конъюгированным с ECD (энергетически связанный краситель, PE-Texas-Red-конъюгат). Затем клетки отмывают и инкубируют в течение 15 мин при 4oС в 400 мкл среды AIM-V (GIBCO BRL, бессывороточная среда определенного состава, обычно используемая для культур лимфоцитов человека) с 0,2% (вес/объем) азида натрия (чтобы блокировать интернализацию/ рециклинг) в одних средах или с добавлением FZI/0 (4,0 мг/мл), FZI/1 (4,0 мг/мл), BSA (4,0 мг/мл, негативный контроль) или немеченого Zot (160 мкг/мл, позитивный контроль). Затем Zot-FITC добавляют в каждую пробирку до достижения концентрации 40 мкг/мл и инкубируют в течение 5 мин (время, требуемое до достижения равновесия), отмывают и немедленно переносят в проточный цитометр. Так, FZI/0, FZI/1 и BSA добавляют в 100-кратном избытке и немеченый Zot - в четырехкратном избытке по сравнению с концентрацией добавленного Zot-FITC. К сожалению, технические трудности при получении больших количеств препаратов очищенного немеченого Zot мешают оценке его способности блокировать связывание в случае более чем четырехкратного избытка. Тромбоциты, эритроциты (если они есть) и клеточный дебрис исключают из анализа установкой соответствующего окна рассеяния в прямом направлении против параметров 90% рассеяния света. Для каждого образца мы собираем данные более чем для 10000 клеток. Анализ данных осуществляют, используя Epics Elite пакет аналитических программ (Coulter) или WinList пакет аналитических программ (Verity Software House, Topsham, ME). Результаты изображены в виде % супрессии средней интенсивности флуоресценции клеток, инкубированных с Zot-FITC в присутствии антагонистов Zot, немеченого Zot или BSA, относительно средней интенсивности флуоресценции клеток, инкубированных в одних средах (произвольно взятой за 100%).
Б. Результаты
Типичный опыт, иллюстрирующий действие инкубации РВМС от 3 различных добровольцев с Zot-антагонистами, немеченым Zot или BSA, показан на фиг. 9. Добавление либо FZI/0, либо FZI/1-антагонистов Zot в 100-кратном избытке не блокирует в заметной степени связывания Zot-FITC с СD14+-селектированными макрофагами. Аналогично, добавление 100-кратного избытка BSA не влияет на связывание Zot-FITC. Напротив, преинкубация только с четырехкратным избытком немеченого Zot блокирует связывание Zot-FITC на 24-43%. Эти результаты подтверждают, что в связывании Zot с макрофагами человека участвует рецептор с различными сайтами связывания, которые сайты связывания Zot/зонулин-рецепторов идентифицируют в головном мозге и тканях кишечника.
ПРИМЕР 8
Zot-супрессия индуцированной столбнячным токсином (ТТ) пролиферации зависит от фактора(-ов), присутствующих в сыворотке
А. Материлы и методы
Выделение мононуклеаров периферической крови человека (РВМС) из крови здоровых добровольцев
РВМС выделяют из крови здоровых добровольцев, как описано выше в примере 4.
Получение очищенного 6xHis-Zot
6xHis-Zot получают, как описано выше в примере 2.
Условия культивирования и количественные анализы лимфопролиферации
РВМС (1,5•106 клеток/мл) культивируют в 1,0 мл либо (а) среды AIM-V, (b) RPMI 1640, содержащей 10 об.% термоинактивированной фетальной телячьей сыворотки и 50 мкг/мл гентамицина, либо (с) RPMI 1640, содержащей 10 об.% термоинактивированной АВ сыворотки и 50 мкг/мл гентамицина. Клетки термостатируют при 37oС, 5% CO2 в 96-луночных планшетах в отсутствие или в присутствии столбнячного токсина (ТТ, специфический антиген, используемый в количестве 2,0 мкг/мл; Wyeth, Marietta, PA) без или с очищенным Zot (60 мкг/мл) или альбумином бычьей сыворотки (BSA, контрольный белок, 60 мкг/мл; Fraction V, Sigma, St. Louis, МО). Клетки культивируют 6 дней и добавляют 3Н-тимидин в количестве 1,0 Ки/лунка. Через 20 ч проводят сбор с планшетов с помощью харвестера клеток Wallac (Уоллака) (Gaithersburg, MD) и включенный тимидин измеряют счетчиком Wallac Trilux Microbeta (Gaithersburg, MD).
Б. Результаты
Результаты 3-х независимых экспериментов показывают, что никакого ингибирования ТТ-индуцированной пролиферации не наблюдается, когда термостатируют РВМС в отсутствие сыворотки, например, когда в культурах используют AIM-V-среду определенного состава. Фактически в большинстве случаев наблюдают реверсию, т.е. инкубация с Zot в отсутствие сыворотки ведет к более мощному ("повышенному") пролиферативному иммунному ответу на ТТ. Напротив, инкубация с Zot приводит к заметной супрессии ТТ-индуцированной пролиферации, когда культивирование осуществляют в присутствии либо FCS, либо человеческой АВ-сыворотки. Действительно, присутствие человеческой АВ-сыворотки, по-видимому, опосредует более высокие уровни (вплоть до 90%) супрессии ТТ-индуцированной пролиферации, чем уровни, наблюдаемые в присутствии FCS.
ПРИМЕР 9
Zot-супрессия экспрессии CD14 на моноцитах/макрофагах человека
А. Материлы и методы
Выделение мононуклеаров периферической крови человека (РВМС) из крови здоровых добровольцев
РВМС выделяют из крови здоровых добровольцев, как описано выше в примере 4.
Получение очищенного 6xHis-Zot
6xHis-Zot получают, как описано выше в примере 2.
Условия культивирования и проточный цитометрический анализ
РВМС, выделенные как описано выше, инкубируют при 37oС, 5% СО2 в 24-луночных планшетах различные периоды времени (от 4 ч до 7 дней) в отсутствие или в присутствии столбнячного токсина (ТТ, специфический антиген, используемый в количестве 2,0 мкг/мл; Wyeth, Marietta, PA) без или с очищенным Zot (60 мкг/мл) или альбумином бычьей сыворотки (BSA, контрольный белок, 60 мкг/мл; Fraction V, Sigma, St. Louis, МО). Затем клетки окрашивают mAb к CD14, меченым FITC, и анализируют проточным цитометрическим анализом. Тромбоциты, эритроциты (если они есть) и клеточный дебрис исключают из анализа установкой соответствующего окна рассеяния в прямом направлении против параметров 90% рассеяния света. Для каждого образца мы собираем данные более чем для 10000 клеток. Анализ данных осуществляют, используя Epics Elite пакет аналитических программ (Coulter) или WinList пакет аналитических программ (Verity Software House, Topsham, ME). Результаты изображены в виде одноцветных гистограмм флуоресценции CD14 на клетках, селектированных в "области моноцитов", определяемой как рассеяние вперед, по сравнению с боковым рассеянием человеческих макрофагов.
Б. Результаты
Типичный эксперимент, иллюстрирующий влияние инкубации РВМС с Zot на экспрессию CD14 в макрофагах человека, дан на фиг. 10А-10В. Добавление Zot вызывает заметную супрессию экспрессии CD14 после 18 ч инкубации. Этот эффект очень проявляется либо в отсутствие (фиг. 10А), либо в присутствии (фиг. 10В) столбнячного токсина. Добавление BSA не влияет на экспрессию CD14. Кинетические исследования показывают, что Zot-индуцированная супрессия CD14-экcпpeccии наблюдается примерно в половине экспериментов уже через 4-6 ч после экспозиции с Zot, и что CD14-экcпpeccия заметно подавлена после 7 дней в культуре (последняя оцениваемая временная точка). CD14 является молекулой на поверхности макрофагов, которая ведет себя как рецептор с высокой аффинностью для LPS-LPS-связывающих белковых комплексов. Таким образом, полагают, что негативная регуляция экспрессии CD14 под действием Zot оказывает сильное влияние на активацию макрофагов и его способность эффективно инициировать опосредуемые Т-клетками иммунные реакции.
ПРИМЕР 10
Влияние Zot на жизнеспособность лимфоцитов человека и моноцитов /макрофагов
А. Материлы и методы
Выделение мононуклеаров периферической крови человека (РВМС) из крови здоровых добровольцев
РВМС выделяют из крови здоровых добровольцев, как описано выше в примере 4.
Получение очищенного 6xHis-Zot
6xHis-Zot получают, как описано выше в примере 2.
Условия культивирования и проточный цитометрический анализ
РВМС, выделенные как описано выше, инкубируют при 37oС, 5% СО2 в 24-луночных планшетах различные периоды времени (от 6 ч до 7 дней) в отсутствие или в присутствии столбнячного токсина (ТТ, специфический антиген, используемый в количестве 2,0 мкг/мл; Wyeth, Marietta, PA) без или с очищенным Zot (40 мкг/мл) или альбумином бычьей сыворотки (BSA, контрольный белок, 40 мкг/мл; Fraction V, Sigma, St. Louis, МО). Затем клетки окрашивают диализованными mAb (антителами) к CD14, меченым FITC, и отмывают. Для оценки жизнеспособности клеток пропидийиодид (PI, краситель, который легко встраивается в мертвые клетки, но исключается из жизнеспособных клеток) добавляют в суспензию клеток и сразу же анализируют образцы проточной цитометрией. Тромбоциты, эритроциты (если они есть) и клеточный дебрис исключают из анализа установкой соответствующего окна рассеяния в прямом направлении против параметров 90% рассеяния света. Для каждого образца мы собираем данные более чем для 10000 клеток. Анализ данных осуществляют, используя Epics Elite пакет аналитических программ (Coulter) или WinList пакет аналитических программ (Verity Software House, Topsham, ME). Результаты показаны как % жизнеспособных клеток, селектированных (отсортированных) в "области моноцитов" или "области лимфоцитов", определяемой на основе рассеяния в прямом направлении (вперед) по сравнению с боковым рассеянием этих популяций клеток.
Б. Результаты
Типичный эксперимент, иллюстрирующий действие инкубации РВМС с Zot на жизнеспособность макрофагов и лимфоцитов человека, показан на фиг. 11. Добавление Zot вызывает умеренное уменьшение жизнеспособности макрофагов по сравнению с контролем уже после 1 дня культивирования. Это уменьшение стало совершенно отчетливым на 4 день и практически все макрофаги погибают после 7 дней культивирования. Напротив, не наблюдается никакого различия в жизнеспособности лимфоцитов между культурами, термостатированными с Zot, и культурами, инкубированными со средами или BSA, до 4-го дня. Однако жизнеспособность лимфоцитов позднее заметно падает в присутствии Zot. Сходные результаты наблюдаются, когда ТТ добавляют в культуры в отсутствие или в присутствии Zot. Эти результаты показывают, что Zot влияют на жизнеспособность макрофагов в относительно ранний период, в то время как воздействие на лимфоциты становится явным, по меньшей мере, после 4 дней культивирования. Эти наблюдения дают дополнительную информацию относительно механизмов, которые могут лежать в основе Zot-опосредованной супрессии процессирования и презентации антигенов, указывая на раннее действие на макрофаги.
ПРИМЕР 11
Zot-опосредованная индукция продуцирования цитокинов моноцитами/макрофагами и лимфоцитами человека
А. Материлы и методы
Выделение мононуклеаров периферической крови человека (РВМС) из крови здоровых добровольцев
РВМС выделяют из крови здоровых добровольцев, как описано выше в примере 4.
Получение очищенного 6xHis-Zot
6xHis-Zot получают, как описано выше в примере 2.
Условия культивирования
РВМС, выделенные как описано выше, инкубируют при 37oС, 5% СО2 в 24-луночных планшетах различные периоды времени (от 6 ч до 4 дней) в отсутствие или в присутствии столбнячного токсина (ТТ, специфический антиген, используемый в количестве 2,0 мкг/мл; Wyeth, Marietta, PA) без или с очищенным Zot (60 мкг/мл) или альбумином бычьей сыворотки (BSA, контрольный белок, 60 мкг/мл; Fraction V, Sigma, St. Louis, МО). Для исследований, включающих продуцирование TNF-a, IL-1b и IL-10, содержимое планшетов собирают в час 6 и в дни 1, 2 и 4 в пробирки Эппендорфа на 1,5 мл и центрифугируют при 4oС в течение 10 мин при 2700 • g в охлаждаемой центрифуге Эппендорфа для удаления клеток и дебриса (осадка). Супернатанты переносят затем в новые пробирки Эппендорфа и замораживают при -70oС до проведения анализа. Супернатанты для определения цитокинов Т-лимфоцитов (например, IL-2, IL-4 и TNF-g) после стимуляции под действием ТТ без или с очищенным Zot или BSA собирают через 3 дня.
Определение цитокинов хемилюминесцентным твердофазным иммуноферментным анализом (ELISA)
Стандартный метод - хемилюминесцентный ELISA с иммобилизацией антител на твердой фазе применяют для обнаружения присутствия цитокинов в супернатантах клеточных культур. Коротко говоря, пленкой - 1,0-2,0 мкг/мл антител к цитокинам человека - покрывают 96-луночные черные непрозрачные планшеты ELISA (Corning-Costar, Cambridge, MA) - либо в PBS (рН 7,4) (Biofluids), либо в 0,1 М бикарбонате натрия (рН 8,1) в течение ночи при 4oС. Планшеты отмывают PBS (рН 7,4), содержащим 0,5 об.% Tween 20 (Sigma), и блокируют либо PBS (рН 7,4), содержащим 10 об.% FCS или 4,0% (вес/объем) BSA. После отмывания в лунки добавляют супернатанты клеточных культур или рекомбинантные цитокины человека (в качестве стандарта) и инкубируют 2 ч при комнатной температуре. После отмывания соответствующие mAb против цитокинов, конъюгированные с биотином, добавляют в лунки (45 мин при комнатной температуре), отмывают и инкубируют с авидинпероксидазой в течение 30 мин при комнатной температуре. Затем добавляют реагент ВМ хемилюменесцентного ELISA (Boehringer Mannheim, Gaithersburg, MD) и детектируют хемилюминесценцию на планшет-ридере 1450 Microbeta Trilux (Wallac, Gaithersburg, MD). Антитела против IL-1b получены от Endogen (Wobum, MA), все другие - от Pharmingen (San Diego, CA).
Б. Результаты
Типичный эксперимент, иллюстрирующий способность Zot индуцировать продуцирование цитокинов макрофагами человека, показан на фиг. 12А-12С. Добавление Zot (в отсутсвие ТТ) вызывает продуцирование значительных количеств TNF-a уже через 6 ч, достигающих максимальных уровней (пика) через 24 ч (3400-3800 пг/мл) и после этого уменьшающихся почти до фоновых уровней через 4 дня (фиг. 12А). Самые низкие уровни TNF-a наблюдаются при культивировании в средах или BSA (~1000-1100 пг/мл), возможно, в результате неспецифической активации макрофагов после прикрепления (адгезии) к полимеру. Также наблюдается слабая Zot-индукция, продуцированная IL-1b (~40-60 пг/мл), максимальные уровни достигаются через 2 дня и заметно падают на 4 день (~20 пг/мл) (фиг. 12В). При культивировании в средах или BSA не наблюдается заметных уровней IL-1b. Наконец, инкубация с Zot приводит к продуцированию на высоком уровне IL-10 через 6 ч (~1000 пг/мл); уровни продуцирования достигают максимума к концу 1 дня (~1500 пг/мл) и та же концентрация сохраняется до 4 дня (фиг. 12С). Аналогично тому, что найдено для TNF-a (описано выше), более низкие уровни IL-10 наблюдаются в культурах в средах или BSA, возможно, в результате неспецифической активации макрофагов после адгезии с полимером. Присутствие ТТ не изменяет ни кинетику, ни интенсивность иммунного ответа цитокинов, наблюдаемых после добавления Zot. Индукция мощных провоспалительных цитокинов после экспозиции с Zot дает дополнительную информацию относительно механизмов, которые могут лежать в основе опосредуемой Zot супрессии процессирования и презентации антигенов, еще раз указывая на раннее воздействие на макрофаги. Например, полагают, что Zot-опосредованная индукция продуцирования IL-10, цитокина, который, как известно, является основным ингибитором иммунного ответа типа Th1 через ингибирование цитокинов, таких как IL-2, - играет значительную роль в супрессии антиген-индуцированных лимфопролиферативных иммунных ответов, наблюдающейся, когда в культуры добавляют Zot.
Типичный эксперимент, иллюстрирующий влияние Zot на продуцирование цитокинов, образующихся в Т-клетках, показан на фиг. 13А-13В. Наблюдается, что добавление Zot подавляет продуцирование IL-2, стимуллированное инкубацией с ТТ, тогда как BSA не оказывает никакого действия (фиг. 13 А). В отсутствие ТТ Zot не индуцирует продуцирование IL-2 на поддающемся измерению уровне. Напротив, добавление Zot в отсутствие ТТ устойчиво стимулирует продуцирование IFN-g на уровне от низкого до умеренного - аналогично уровням, индуцированным под действием ТТ (фиг. 13В). Кроме того, Zot заметно повышает уровни IFN, индуцированного ТТ, тогда как BSA не оказывает влияния (фиг. 13В). Наконец, наблюдается, что добавление Zot не стимулирует продуцирование IL-4. Так как IL-2 является цитокином, который играет важную (стратегическую) роль в пролиферации лимфоцитов, полагают, что супрессия продуцирования IL-2 после антигенной стимуляции является одним из ключевых механизмов, лежащих в основе опосредуемой Zot супрессии ТТ-индуцированной пролиферации. Важно также следующее: факт, что Zot индуцирует продуцирование IFN-g в отсутствие или в присутствии ТТ, а также продуцирование многих провоспалительных цитокинов, - указывает, что Zot проявляет свое иммунорегуляторное действие на различных уровнях в ходе сложного процесса процессирования и презентации антигена, приводящего к генерации антиген-специфических иммунных реакций.
Хотя данное изобретение описано подробно и со ссылкой на его конкретные варианты, рядовому специалисту в данной области техники ясно, что различные изменения и модификации могут быть в нем сделаны без изменения сущности и объема его.



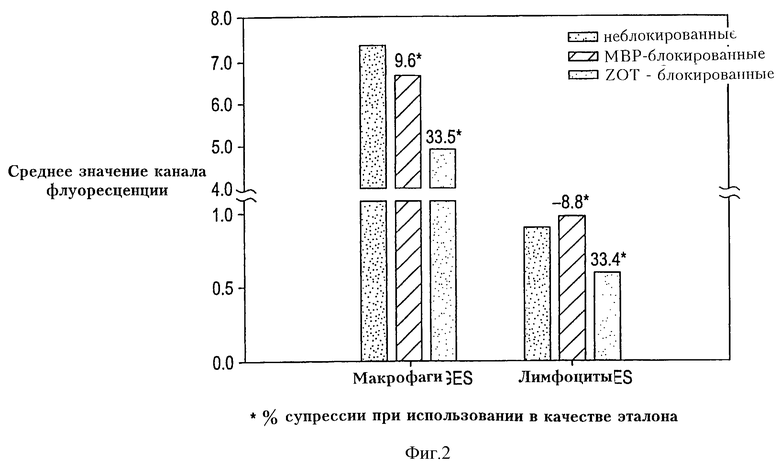
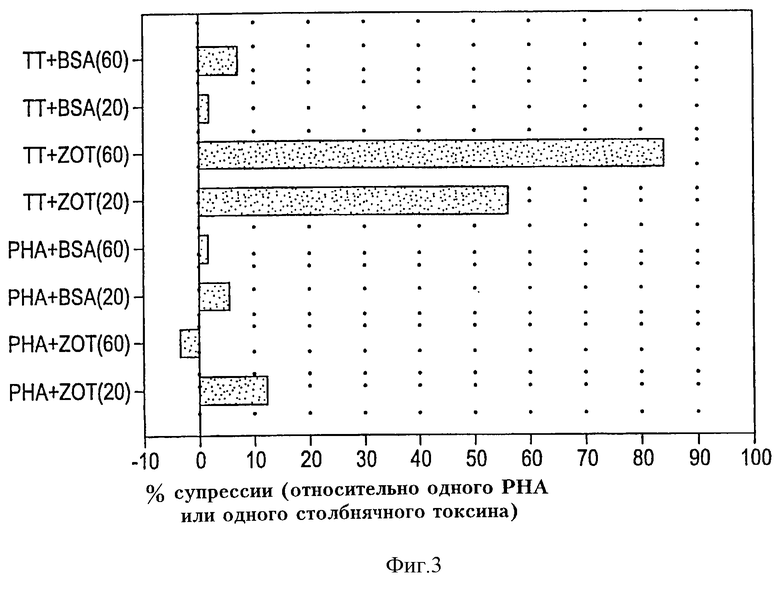
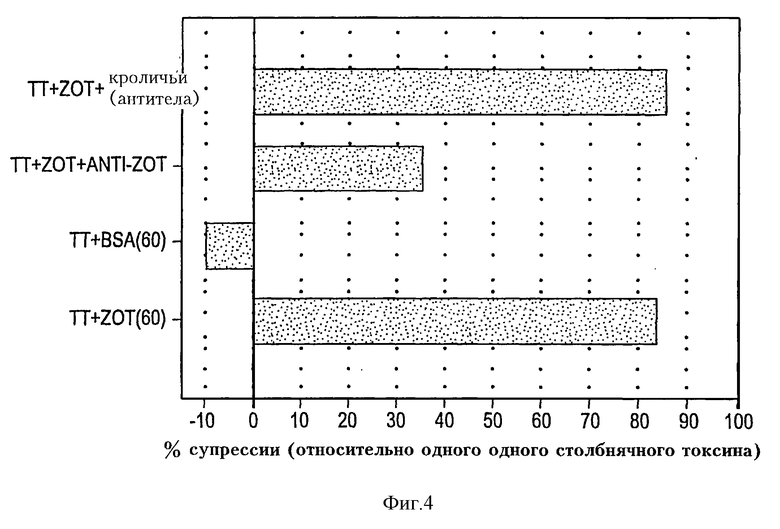
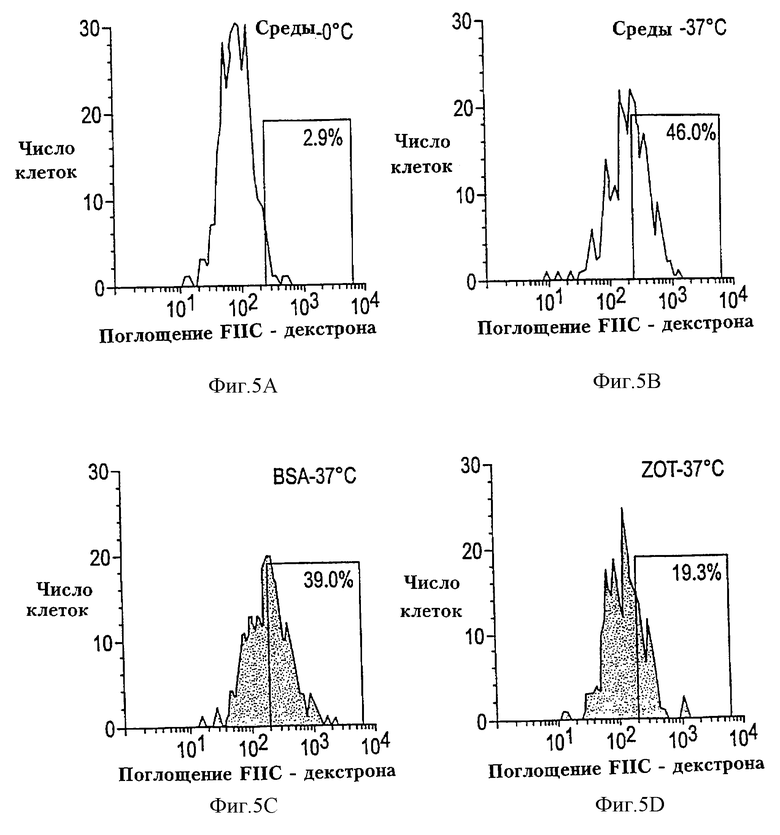
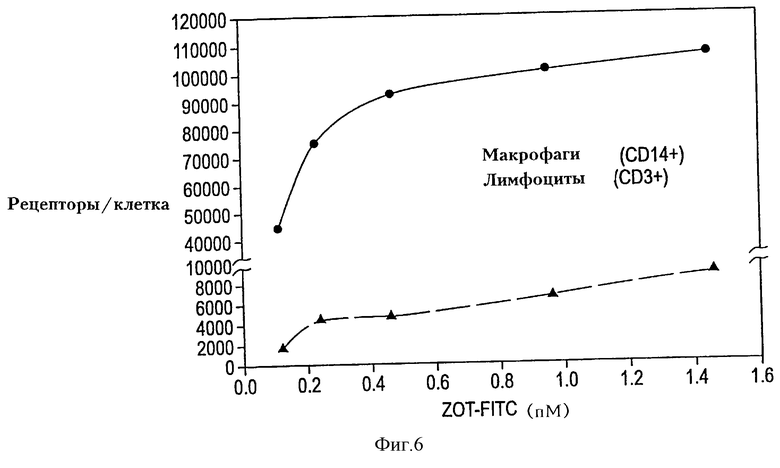
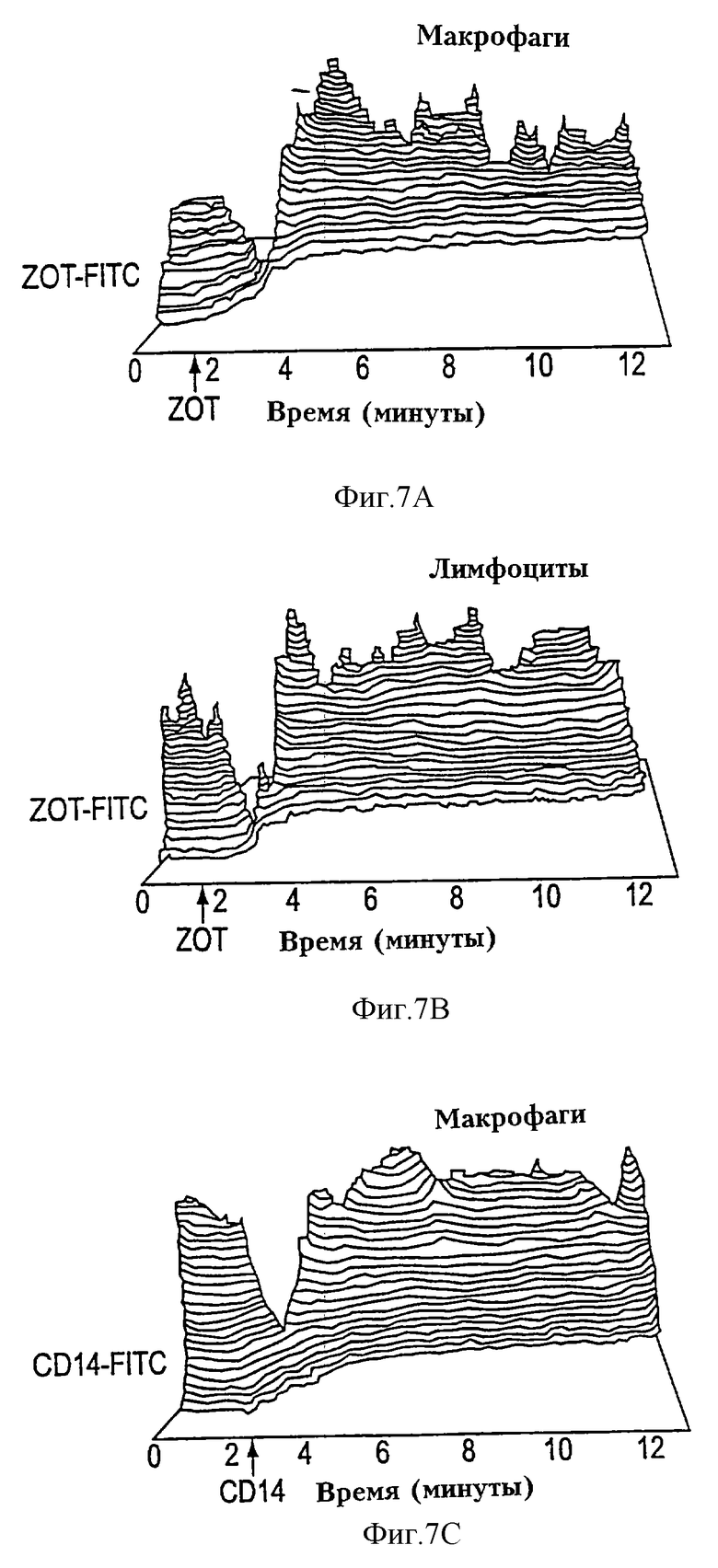
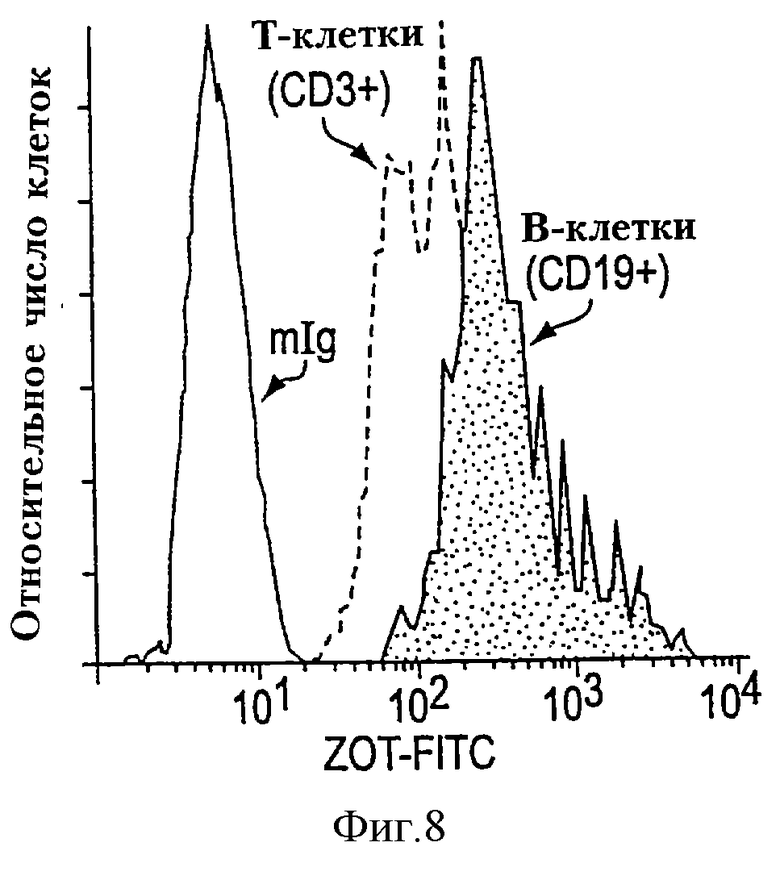
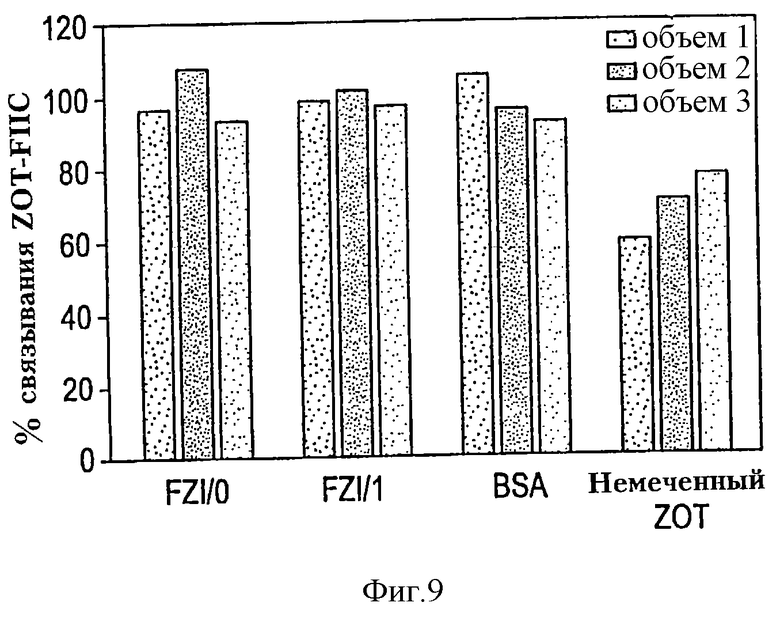
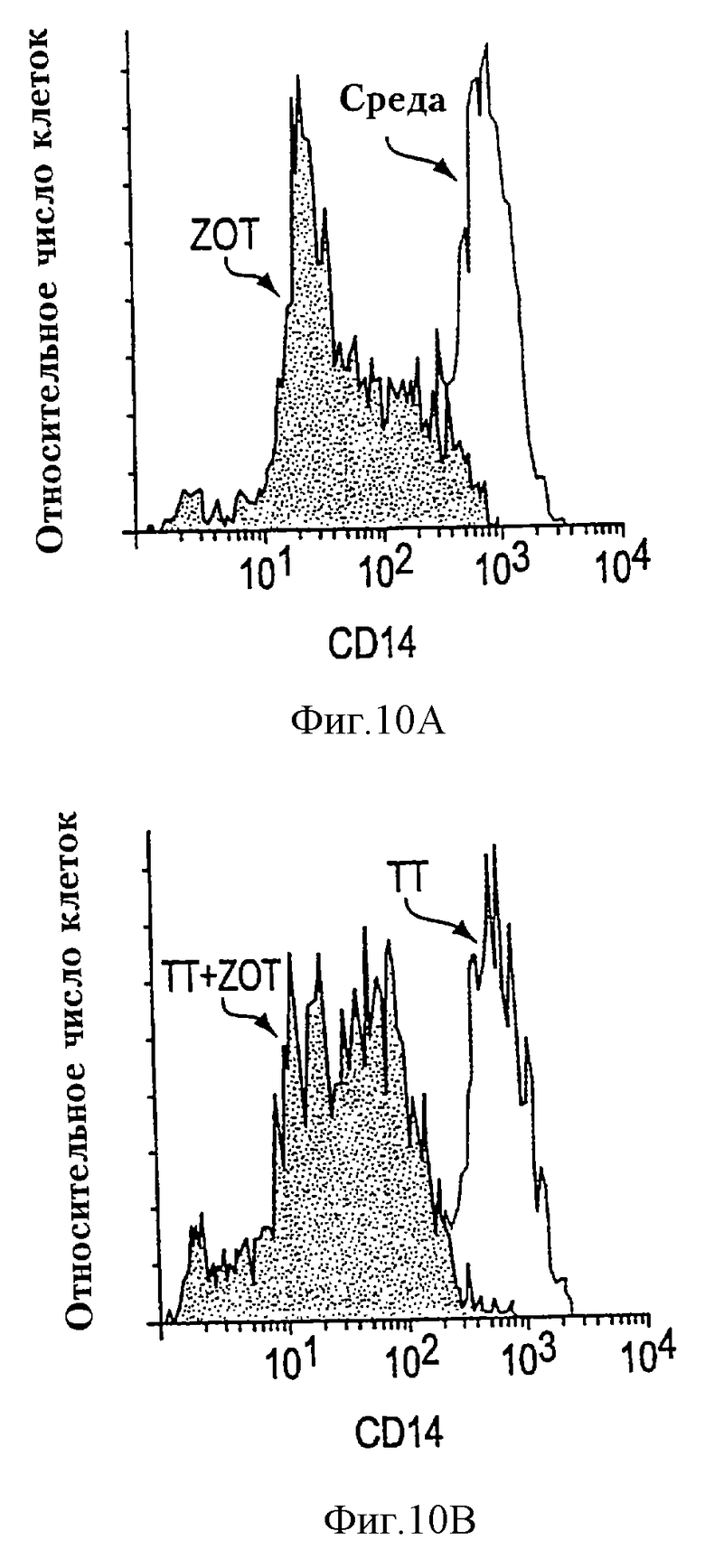
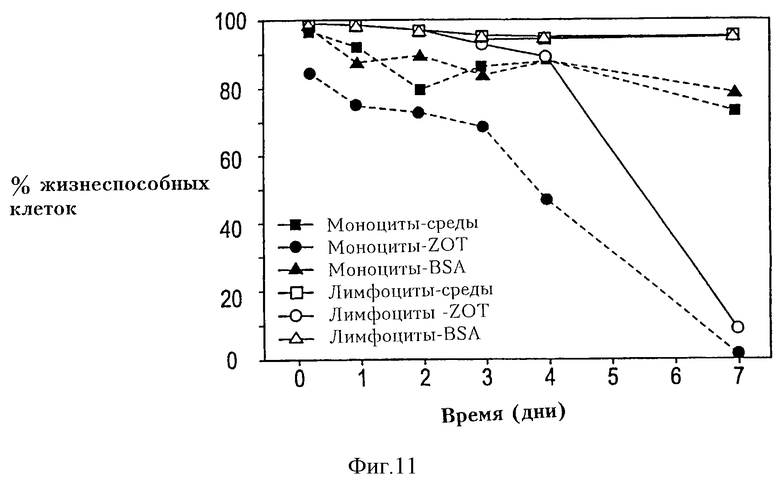
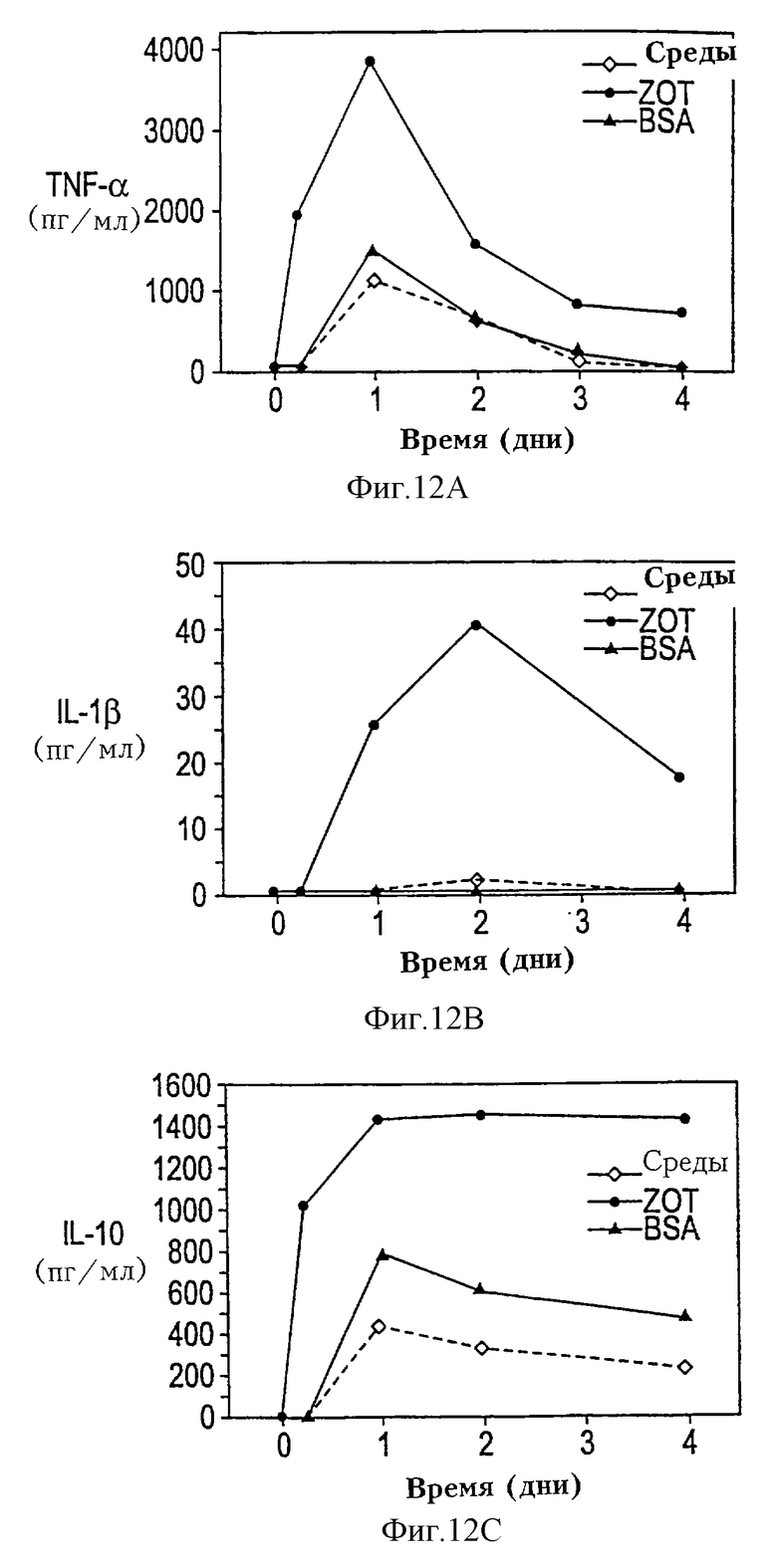
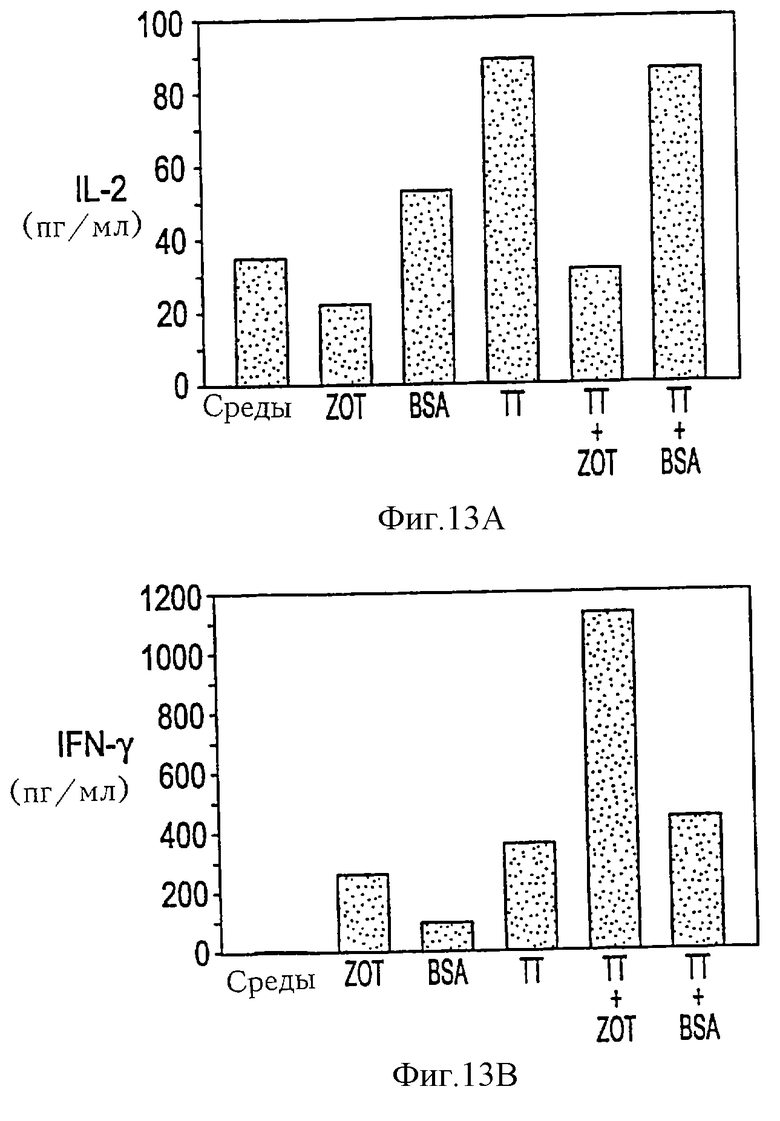
Изобретение относится к медицине, в частности к иммунологии, и касается ингибиции пролиферации лимфоцитов. Для этого используют Zot-токсин зоны плотных контактов, вырабатываемый V. cholerae, или зонулин - препарат белков млекопитающих, которые иммунологически или функционально близки Zot. Эффект ингибиции проявляется как в культуре клеток, обработанных антигеном, так и, предположительно, в условиях организма млекопитающего. Способ обеспечивает значительную ингибицию сенсибилизированных лимфоцитов за счет блокирования антигена. 4 с. и 12 з.п. ф-лы, 13 ил.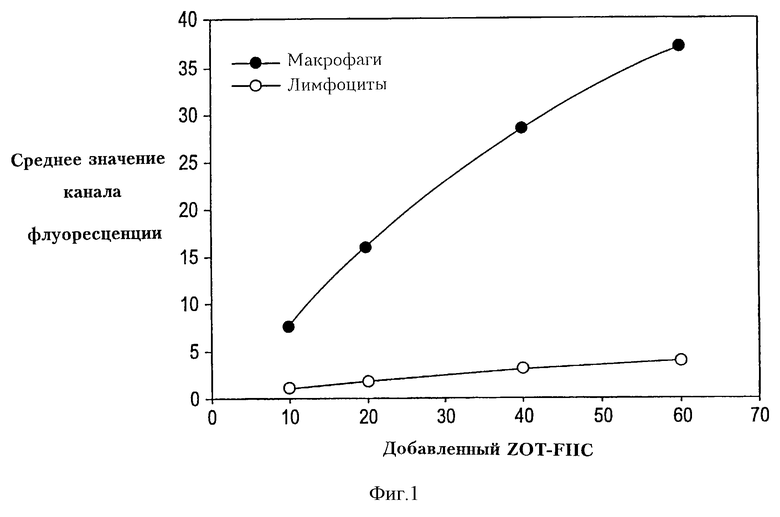
| REED et al | |||
| - in J | |||
| Immunol., 1986, 137, 150-154 | |||
| RU 96108963 А, 10.10.1998 | |||
| КУЛЬТУРА V.CHOLERAE, СПОСОБ ПОЛУЧЕНИЯ ДЕЛЕЦИОННЫХ МУТАНТОВ | 1991 |
|
RU2130970C1 |
| US 5665389 А, 09.09.1997 | |||
| US 5945510 А, 31.08.1999 | |||
| Справочник по ревматологии | |||
| /Под | |||
| ред | |||
| акад | |||
| В.А | |||
| Насоновой | |||
| - М.: Медицина, 1995, с.35-36 | |||
| FASAND et al | |||
| - in Proc | |||
| Natl | |||
| Acad | |||
| Sci., USA, 1991, 8, 5242-5246. | |||

