Настоящая патентная заявка является частичным продолжением патентной заявки Заявителя в США N 06/581406, поданной 17 февраля 1984 г., которая является частичным продолжением патентной заявки США N 06/472276, поданной 4 марта 1983 г., которые включены здесь в качестве ссылки. Настоящая патентная заявка является также частичным продолжением патентной заявки Заявителя в США N 07/363383, поданной 5 июня 1989 г., по которой выдан патент 12 декабря 1989 г. , которая является продолжением патентной заявки США N 06/867633, поданной 27 мая 1986 г., которая является продолжением патентной заявки США N 06/472276, поданной 4 марта 1983 г., которые включены здесь в качестве ссылки. Исследование, проведенное в соответствии с настоящей заявкой, было поддержано национальным институтом здоровья и, частично, Медицинским Центром Биотехнологии Института Биотехнологии штата Мэрилэнд.
Предпосылки настоящего изобретения
Vibrio cholerae (V. cholerae) является нонинвазивным энтеропатогеном тонкой кишки, который не проникает через поверхность слизистой оболочки. Локальный иммунитет с участием SIgA на поверхности слизистой оболочки, таким образом, объясняется защитным механизмом. Патогенный V.cholerae 01 вырабатывает протеиновый энтеротоксин (известный также под названием энтеротоксин холеры или холераген или токсин холеры), который отвечает за индукцию обильного выделения, приводящего к водянистому поносу, клиническому следствию холерной инфекции. Генами, отвечающими за токсин холеры, являются гены ctx (известные также как tox-гены). Холерный понос может быть в высшей степени болезненным и может приводить к потере столь значительного количества воды и солей, что это приводит к обезвоживанию, ацидозису, шоку и гибели в конечном итоге, если не применять экстренной терапии. Заявитель установил, что V. cholerae продуцирует второй токсин, называемый токсином "zonula occludens" (блокировки малой зоны), описанный Fasano и др., Vibrio cholerae Produces a Second enterotoxin Which Affects Intestinal Tight Junctions, Nature (в печати, 1990 г. ). Кроме того, имеется область хромосомы, содержащая гены ctx, которая содержит несколько копий последовательности в 2700 пар оснований, называемой RSI (для повторяющейся последовательности), Mekalanos, Cell, т. 35, стр. 253-263 (1983).
Вакцины холеры, которые были разработаны, могут быть грубо разделены на две категории: вакцины, помогающие выработке антитоксичного иммунитета, и вакцины, предназначенные для того, чтобы индуцировать антибактериальный иммунитет. Эксперименты с животными моделями подтверждают защитную роль как антитоксичного, так и антибактериального иммунитета. Было предложено, что когда оба типа иммунитета работают одновременно, то можно добиться синергического эффекта [Holmgren, J. и др., J. Infect. Dis, Suppl, 136, стр. 105 - 1122 (1977); Peterson, J.W.Infect. Immun. т. 26, стр. 594 (1979); Resnick, I. G. и др., Infect. Immun. т. 13, стр. 375 (1980); Svennerholm A.-M. и др., Infect. Immun. т. 13, стр. 735 (1976)]. Однако оказалось, что защитный иммунитет у человека можно индуцировать без такого синергического эффекта, то есть при помощи либо антитоксического иммунитета, либо антибактериального иммунитета [Eubanks, E.R., и др., Infect. Immun, т. 15, стр. 533 (1977); Fujita, K. и др. Infect. Dis., т. 125, стр. 647 (1972); Holmgren, J. J.Infect. Dis., см. выше; Lange, S. и др., Acta Path. Microbiol. Scand Sect. C, т. 86, стр. 145 (1978); Peterson, J.W., см. выше (1979); Pierce, N.F. и др., Infect. Immun. т. 37, стр. 687 (1982); Pierce, N.F. и др., J. Infect. Immun., т. 21, стр. 185 (1978); Pierce. N.F. и др. J. Infect. Dis, т. 135, стр. 888 (1977); Resnick I.G. и др., см. выше; Svennerholm, A.-M. и др., см. выше].
Вакцины из убитых целых клеток
1. Парантеральные вакцины, содержащие целые клетки
Вот уже в течение почти века убитые целые клетки V.choleral использовались в качестве парентеральных вакцин; эти вакцины до сих пор производятся промышленностью. Опыт работы с парентеральными вакцинами с целыми клетками обобщен в обзоре Joo, I "Cholera vaccines" в книге "Cholera". (Barua D. и Burrows W. , ред.), изд. Saunders, Philadelphia, стр. 333-355 (1974), и Feeley, J.D. и др. в книге "Cholera and Related Diarrheas", 43rd Nobel Symp., Stokholm 1978 г. (O.Oucherlong. J. Holmgren ред.), изд. Karger, Basel, стр. 204-210 (1980). Такие вакцины стимулируют высокие титры вибриоцидальных антител сыворотки. Они также стимулируют увеличение антител SIgA кишечника к соматическому антигену O V.cholerae, когда их вводят пакистанцу, но не шведу [Svennerholm, A. -M. и др. Infect. Immun. т. 30, стр. 427) (1980); Svennerholm A. -M. и др. Scan. J. Immun. т. 6, стр. 1345- (1977)]. Было предположено, что реципиенты вакцины из Пакистана реагируют таким образом из-за того, что они уже иммунологически подготовлены благодаря предварительному антигенному контакту, в то время как люди, живущие в неэндемических областях (например, в Швеции), к этому не подготовлены. Было установлено во время полевых испытаний, что парентеральные вакцины убитых целых клеток обеспечивают существенную защиту против гомологического серотипа V.cholerae, но в общем случае в течение периода меньше года (Joo, I, см. выше; Feeley, J.C., см. выше; Svennerholm A.-M. и др., см. выше (1980); Svennerholm A.-M. и др., см выше, (1977); Mosley, W.H. и др., Bull. Wld, Hlth. Org., т. 49, стр. 13 (1973); Phillipines Clolera Committee, Bull. Wld, Hlth, Org., т. 49, стр. 381 (1973)/. Имеются некоторые причины, чтобы предположить, что парентеральная вакцина целых клеток Инаба обеспечивает хорошую кратковременную защиту против Огава, а также против холеры Инаба, в то время как вакцина Огава только против Огава.
Если использовать добавки, то имеется возможность поддерживать эффективность вакцины приблизительно на уровне 70% в течение полутора лет, используя парентеральную вакцину (см., например, Saroso, J.S. и др., Bull. Wld. Helt. Org. , т. 56, стр. 619 (1978). Однако побочные реакции, имеющие место в месте прививки вакцин с добавками (которые включают стерильные абсцессы), происходят достаточно часто и болезненно, что препятствует стандартному использованию таких вакцин с добавками.
2. Стоматические вакцины целых клеток
Убитые полные вибрины, применяемые стоматически, стимулируют появление локальных антител кишечного антивибриона [Freter, R.J.Infect. Dis., т. 111, стр. 37 (1972); Freter, R. и др., J. Immunol., т. 91, стр. 724 (1963); Ganguly, R и др. , Bull. Wld. Heth. Org. т. 52, стр. 323 (1975)]. Другие исследователи показали существенную эффективность вакцины, но большая доля вакцин приводила к поносу после последующего введения патогенных вибрионов [Cash, R.A. и др., J. Imfect. Dis., т. 130, стр. 325 (1974)].
Токсоиды
Иммунизирующие агенты, предназначенные для предотвращения холеры при помощи стимуляции антитоксичного иммунитета, включают:
1) Токсоид холеры, обработанный формальдегидом;
2) Токсоид холеры, обработанный глютаральдегидом;
3) Очищенный субблок B;
4) Прохолерагеноид (обротанный формальдегидом или без такой обработки).
1. Токсоид холеры, обработанный формальдегидом
Обработка очищенного токсина холеры ин витро формальдегидом уничтожает его токсичность, что приводит к образованию токсина, которая проявляет слабую токсичную биологическую активность, но стимулирует антитоксичные антитела после парентеральной иммунизации животных. Однако, когда первый токсоид такого типа применяли к обезьянам и человеку в форме парентеральной вакцины, то токсоид частично восстанавливал токсичность, что приводило к нежелательным локальным неблагоприятным реакциям в месте прививки [Northrup, R. S и др., J. Infect. Dis., т. 125, стр. 471 (1972)]. Токсоид холеры после обработки формалином с добавкой альбумина применяли парентерально добровольцам из Бангладеш, включая кормящих матерей, но полевых испытаний с этой вакциной не проводили [Merson, M. H. и др., Lancet, 1, стр. 931 (1980)]. Токсоид холеры, обработанный формалином, полученный в присутствии гликокола, также испытывали парентеральным способом, но такая вакцина не была эффективной [Ohtomo, N. In Proceedings of the 12th Joint Conference on Cholera, U.S. - Japan Cooperative Medical Science Program, Sapporo (Fukumi H., Zinnaka Y., ред. стр. 286-296 (1976); Noriki, H. In Proceedings of the 12th Joint Conference on Cholera, U. S. - Japan Cooperative Medical Science Program, Sapporo (Fukumi H., Zinnaka Y., ред. стр. 302-310 (1976)].
2. Токсоид холеры, обработанный глютаральдегидом
Были разработаны способы для широкомасштабного получения токсоида холеры, обработанного глютаральдегидом, который по существу свободен от загрязняющего соматического антигена [Rappaport, E.S. и др., Infect Immun., т. 14, стр. 687 (1976)] . Можно было надеяться, что этот антиген можно будет использовать для анализа "чистым" способом защитной роли отдельно антитоксичного иммунитета. Широкомасштабные полевые испытания этого токсоида, применяемого в форме парентеральной вакцины, осуществляли в Бангладеш в 1974 г. [Curlin, G. и др., в Proceeding of the 11th Joint Conference on Cholera, U. S. - Japan Cooperative Medical Science Program стр. 314-329, New Orleans (1975)] . Этот токсоид стимулировал высокие титры циркулирующих антитоксинов у реципиентов из Бангладеш. Две волны холеры, Эль Тор Инаба, затем Эль Тор Огава, расширили область испытаний, что давало возможность получить хорошую оценку эффективности вакцины. Защитный эффект оказалось возможным подтвердить только в одной возрастной группе и он был ограничен на период эпидемии Инаба так, что токсоид холеры, обработанный глютаральдегидом, применяемый отдельно в форме парентеральной вакцины, обеспечивал слабую защиту и она была существенно ниже по сравнению с аналогичными полевыми испытаниями в той же группе населения с парентеральными вакцинами убитых целых клеток.
Применение токсоида холеры, обработанного глютаральдегидом, в форме стоматической вакцины было исследовано в предположении, что токсоид, введенный таким путем, является более эффективным при стимуляции кишечного антитоксина [Le Vine M.M и др., Trans. Roy. Soc. Trop. Med. Hyg. т. 73, стр. 3 (1979)] . Две группы добровольцев были иммунизированы тремя 2,0 мг или тремя 7,0 мг дозами токсоида, введенного непосредственно в малую полость кишечника (через трубку, вставленную в кишечнике) с месячным интервалом. Вакцины и иммунизированных контрольных животных затем использовали в экспериментальных исследованиях с холерой. Ни в одном из исследований с симптомами холеры не было отмечено какого-либо существенного снижения степени или болезненности поноса как у носителей вакцины, так и у контрольных организмов. Отсутствие эффективности стоматического токсоида холеры, обработанного глютаральдегидом, может быть объяснено тем фактом, что способность субблоков B связываться с ганглиозидом GMI значительно снижается, как следствие токсоидизации с глютаральдегидом.
3. Очищенный субблок B
Энтеротоксин холеры состоит из двух субблоков, обозначаемых через A и B. Субблок A индуцирует ферментативные изменения, которые приводят к выделению жидкости, в то время как нетоксичный субблок B является иммуногенной составляющей, которая связывается с рецептором для токсина (ганглиозида GMI) на клетках эпителия кишечника [Holmgren, J. Nature, т. 292, стр. 413 (1981)]. Было показано, что очищенный субблок B, введенный стоматически или парентерально бангладешцам, стимулирует появление антитоксина SJgA в жидкости кишечника, что приписывают иммунологической "затравке" в районах эпидемии холеры [Svennerholm, A.-M. и др., Lancet 1, стр. 305 (1982)].
Основные преимущества стоматической вакцины субблока B с тем, чтобы стимулировать антитоксиновый иммунитет, включают ее полную безопасность (отсутствует потенциальная опасность превращения в токсин, которая имеется с токсоидами) и сохранение ее способности "прилипать" к рецепторам токсинов на энтероцитах. Исследования на животных наводят на мысль, что она менее сильна, чем нативный голотоксин при стимуляции антитоксина [Pierce, N.F. см. выше, (1982)].
Должно быть ясно, что очищенный субблок B можно использовать, если он используется вместе, например, со стоматическими убитыми вибрионами в форме комбинации стоматической вакцины, предназначенной для стимулирования как антибактериальных, так и антитоксичных антител.
4. Прохолерагеноид
Прохолерагеноид является токсоидом с большим молекулярным весом (приблизительно 1 000 000), который получают, когда энтеротоксин нагревают до 65oC в течение по крайней мере пяти минут [Finkelstein, R.A. и др., J. Immunol. т. 107, стр. 1043 (1971)]. Он имммуногенен, при этом сохраняет менее 5% биологической токсичной активности "родительского" токсина. Нагревание на более продолжительное время (например, на 25 минут) дает меньшую биологическую токсичность Z [Germanier, R. и др., J. Infect. Dis., т. 13, стр. 1692 (1976)] , а последующая обработка формальдегидом полностью уничтожает остаточную биологическую токсичность. Полученный в результате обработанный формальдегидом прохолерагеноид является по крайней мере столь же эффективным, как и родительский токсин при стимулировании антитоксина сыворотки после иммунизации кроликов. У швейцарских добровольцев развивались быстрые реакции антитоксина сыворотки после парентегральной иммунизации дозами в 10, 30 или 100 микрограмм обработанного формальдегидом прохолерагеноида [Germanier, R. и др., J. Infect. Dis., т. 135, стр. 512 (1977)]. Никаких заметных неблагоприятных реакций не было отмечено.
Таким образом, стоматический прохолерагеноид антигена является более иммуногенным, когда его применяют в форме без обработки формальдегидом. У собак необработанный прохолерагеноид переносился так же хорошо, как стоматическая вакцина; стоматические дозы (с NaHCO3) до 500 микрограмм не вызывают понос. Пять доз в 500 микрограмм, распределенные на 42 дня, стимулируют существенную защиту у собак против соматического заражения патогенным V.cholerae. Дозы в 50 микрограмм и 200 микрограмм NaHCO3 давали группам из шести и четырех взрослых добровольцев. Соответственно, без заметных неблагоприятных реакций.
Должно быть ясно, что прохолерагеноид может быть использован вместе с, например, убитыми вибрионами или другими близкими антигенами, чтобы стимулировать антибактериальный иммунитет так, что антитоксичный иммунитет, вызванный прохолерагеноидом, увеличивается.
Комбинированные вакцины
Основная привлекательность неживой стоматической вакцины холеры заключается в ее безопасности. Стоматическая вакцина, состоящая из комбинации антигенов, предназначенная для стимулирования как антибактериального, так и антитоксичного иммунитета, была бы наиболее подходящей для достижения успеха по следующим причинам: токсоидные вакцины, которые стимулируют чисто антитоксичный иммунитет, не демонстрируют эффективность при защите человека от холеры, хотя они могут защищать животные модели. Кроме того, стоматические или парентеральные убитые целые клетки вакцины, которые не стимулируют антитоксичный иммунитет, обеспечивают существенную защиту против холеры у человека, хотя на короткий период времени. Кроме того, комбинации антигенов (таких, как неочищенный токсин холеры или токсин плюс липополисахарид), которые стимулируют как антитоксичный, так и антибактериальный иммунитет, обеспечивают синергическую защиту.
Два исследования до сих пор были осуществлены в основном с комбинациями вакцин. Во-первых, девяти добровольцам давали стоматическим способом обработанный глютаральдегидом токсоид холеры (2 мг еженедельно в течение четырех недель) плюс убитые вибрионы Эль Тор Инаба (1010 вибрионов дважды в неделю в течение четырех недель), а затем спустя один месяц заражали 106 патогенными вибрионами Эль Тор Инаба вместе с шестью иммунизированными контрольными организмами. Понос появлялся только у двух из девяти носителей вакцины против четырех из шести контрольных участников (эффективность вакцины 67%) и заболевание было очевидно более легким у двух носителей вакцины. Более уместным, видимо, является наблюдение, состоящее в том, что V.cholerae можно непосредственно культивировать из стула только двух из девяти носителей вакцины против шести из шести контрольных участников. Это подтверждает, что иммунологические механизмы препятствуют пролиферации вибрионов.
Совсем недавно, три дозы вакцины субблок B/убитые целые клетки вводили взрослым добровольцам, которые участвовали в исследовании эффективности вакцины. Комбинированную вакцину давали на 0, 14 и 28 день. Каждая из трех доз вакцины содержала 0,5 мг очищенного субблока B и 2 • 1011 убиты V.cholerae (5 • 1010 классических Инаба, 5 • 1010 классических Огава и 1 • 1011 Эль Тор Инаба).
Группу из одиннадцати добровольцев, иммунизированных этой комбинированной вакциной, заражали спустя один месяц после введения последней дозы 106 патогенными V.cholerae Эль Тор Инаба, вместе с семью контрольными добровольцами. Понос появлялся у семи из семи контрольных участников, но только у четырех из одиннадцати носителей вакцины (p = 0,01). Состояние у четырех носителей вакцины было определенно лучше.
Таким образом, результаты со стоматическими комбинированными вакцинами токсоид/убитые целые клетки подтверждают измеренную степень эффективности. Защитная эффективность вакцин, однако, только средняя (55-65%) и необходимо несколько доз, чтобы вызвать защиту.
Ослабленные вакцины V.cholerae
Как классическая, так клиническая холера Эль Тор стимулируют высокую степень защитного иммунитета на по крайней мере три года у добровольцев из Северной Америки [Cash, R.A. и др., см. выше (1974); Levine, M.M. и др., см. выше (1979); Levine M.M. и др. "Volunteers studies in development of vaccines against cholera and enterotoxigenic Escherichia coli: обзор в книге "Acute Enteric Infections in Children: New Prospects for Treatment and Prevention (T. Holm, J. Holmgren, M.Merson and R. Mollby, ред.), изд. Elsevier, Amsterdam, стр. 443-459 (1981); и Levine, M.M. и др. J. Infect. Dis. т. 143, стр. 818 (1981)]. На основе этих наблюдений на добровольцах, видимо, самый многообещающий подход в направлении иммунологического контроля холеры заключается в использовании ослабленных нетоксигенных штаммов V.cholerae в качестве стоматических вакцин.
1. Естественно встречающиеся штаммы
Нетоксигенные штаммы V.cholerae 01, изолированные из окружающих источников в Индии и Бразилии, исследовали на добровольцах в качестве потенциальных кандидатов на вакцину с неутешительными результатами. Они либо не образовывали колонии в кишечнике человека, либо делали это минимально; реакции вибриоцидальных антител были слабыми и они не обеспечивали защиту в экспериментальном исследовании с заражением [Cash, R.A и др., Infect. Immun., т. 10, стр. 762 (1974); Levine M. M. и др., J. Infect. Dis., т. 145, стр. 296 (1982)]. Многие из этих штаммов не имели гена токсина, что устанавливали при помощи гибридизации с радиоактивным ДНК-зондом [Kaper, J.B. и др., Infect. Immun. т. 32, стр. 661 (1981)].
2. Мутагенизированные ослабленные штаммы
Классический штамм Инаба 569B мутагенизировали с нитрозогуанидом (НТГ) и гипотоксигенный мутант выделяли [Finlekstein, R.A и др., J. Infect. Dis., т. 129, стр. 177 (1974); Holmes, R.K. и др. J. Clin. Invect., т. 55, стр. 551 (1975)] . Этот мутантный штамм M13 вводили добровольцам. Понос не появлялся ни у одного из участников, но штамм слабо образовывал колонии. Исследование с заражением подтверждало, что некоторая защитная эффективность имела место в результате иммунизации кратными дозами [Woodward, E. и др., Develop. Biol. Stand., т. 33, стр. 108 (1976)].
Эль Тор Огава 3083 также подвергали мутагенезу [Honda, T. и др., Proc. Nat. Acad. Sci. , т. 76, стр. 2052 (1979)]. Грубые селекция и анализ тысяч колоний дали один изолят, который продолжал продуцировать иммуногенные B-субблоки, в то время как не давал обнаруживаемых A-субблоков или голотоксина. Один изолят, Техас Стар-CR, удовлетворял этим критериям. Техас Стар-CR продуцирует стандартное или увеличенное количество субблоков B, но давал отрицательный результат при анализе на активность голотоксина или активность A-субблоков.
Техас Стар-CR широко исследовали на добровольцах [см., например, Levine, M.M. и др., в "Acute Enteric", см. выше (1981)]. Группы из пяти добровольцев получали две дозы по 109 организмов через неделю и восемнадцать добровольцев получали две дозы из 2 • 1010 организмов через неделю. Некоторая степень поноса была отмечена у шестнадцати из шестидесяти восьми носителей вакцины (24%). У только одного участника был отмечен объем стула, превышающий 1,0 литр (1464 мл). В общем случае вызванный вакциной понос состоял из двух или трех небольших стулов, составляющих по объему меньше 400 мл. Организмы вакцины извлекали из сопрокультур приблизительно половины реципиентов вакцины. Там, где тонкокишечную жидкость культивировали (реципиенты доз в 108 и более организмов в вакцине), культуры были положительными у тридцати пяти из сорока шести носителей вакцины (76%). Сотни колоний Техас Стар извлекали из сопрокультур и культуры тонкокишечных жидкостей анализировали на голотоксин холеры при помощи чувствительного анализа клеток надпочечника Y-I; ни один из анализов не был положительным.
Существенный рост антитоксина сыворотки обнаруживали только у 29% носителей вакцины; однако, у 93% был отмечен существенный подъем вибриоцидальных антител сыворотки, а титры существенно близки к титрам, обнаруженным после инфекции патогенным V.cholerae. При экспериментальном изучении заражения добровольцев было установлено, что Техас Стар-CP наделяет существенной защитой против заражения как вибрионами Эль Тор Огава, так и Эль Тор Инаба. Одна или две дозы ослабленной стоматической вакцины Техас Стар-CP обеспечивают хорошую защиту против холеры Эль Тор.
Очевидно, что использование ослабленных штаммов обладает специфическими преимуществами, так как такие штаммы наделяют полученным от инфекции mimie имунитетом к холере. Однако, штаммы Техас Стар-CP имеют некоторые недостатки. Во-первых, мугагенез (например, нитрозогуанидином) индуцирует многократные мутации, не все из которых обязательно узнаваемы. Кроме того, точное генетическое повреждение, которое, как предполагается, отвечает за ослабление Техас Стар-CP, не известно. Кроме того, Техас Стар-CP может вернуться к вирулентности подобно любому патогену, мутируемому нитрозогуанидином.
Заявитель настоящей патентной заявки выделил новым способом мутанты удаления вирулентного штамма Vibrio cholerae, о котором известно, что он одновременно наделяет иммунитетом и вызывает заболевание у добровольцев. Удаления являются фрагментами эндонуклеаз растрикции. Штаммы вакцины, являющейся предметом настоящего изобретения, специфическим образом изменяли через использование техники рекомбинантной ДНК, чтобы получить лишенный вирулентности штамм, не нарушая другие компоненты, необходимые для иммунитета. Это ослабление осуществляют при помощи использования эндонуклеаз рестрикции, которые расщепляют ДНК бактерии в специфических сайтах с целью специфического удаления генов, ответственных за токсин холеры (например, гена ctx). Плазмиды, несущие ген ctx, переваривали эндонуклеазами рестрикции, чтобы удалить ген ctx, но конструкцию осуществляли так, чтобы сохранить реальные длины боковой ДНК хромосомы V.cholerae. Перенос слитого гена плазмид в V.cholerae дает лишенный вирулентности штамм V.cholerae, несущий внехромосомные копии плазмид. Последующая конъюгация с клетками, содержащими другие плазмиды, процуцировала после соответствующей селекции селектируемых маркеров плазмид штаммы V. cholerae, содержащие удаления в областях ctx. Такие нетоксигенные мутанты удаления затем могут оказаться способными образовывать колонии в тонкой кишке и стимулировать локальный защитный иммунитет, направленный против бактериальной клетки. После переходного периода колонизации вакцина должна защищать против последующей инфекции вирулентными токсигенными штаммами V.cholerae.
Гены токсина V.cholerae клонировали [Pearson, G.D.N. и др., Proc. Nat. Acad. Sci. , т. 79, стр. 2976 (1982); Kaper, J.B. и др., Amer. Soc. Microbiol. Abstr. Annu. Merting, Atlanta, Georgia, 36 (1982); Kaper, J.B. и др. Symposium on Enteric Infections in Man and in Animals: Standardization of Immunological Procedurs, Dublin, Ireland. Реферат N 2.5 (1982)]. Мутанты удаления структурного гена токсина V.cholerae выделяли, но только при помощи инфекции мутагенными вибриофагами, способными интегрироваться в случайные сайты вдоль хромосомы [Mekalanos, J.J. и др. Proc. Nat. Acad. Sci., т. 79, стр. 151 (1982)]. Рекомбинация в Vibrio cholerae описана, но ее не использовали, чтобы выделить удаления фрагментов рестрикции в генах ctx для целей вакцинации [Parker, C. и др., J. Bact., т. 112, стр. 707 (1972); Johnson, S. R и др., Molec. Gen. Genet, т. 170, стр. 93 (1979); Sublett, R.D. и др., Infect. Immun. , т. 32, стр. 1132 (1981) и Thomson, J.A. и др., J. Bact. т. 148, стр. 374 (1981)].
Краткое описание настоящего изобретения
Описана культура Vibrio cholerae, содержащая штамм Vibrio cholerae, с удаленным фрагментом эндонуклеазы рестрикции ДНК, чтобы сообщить авирулентность и сохранить способность колонизировать кишечник животного-хозяина. Удаленный ДНК-фрагмент может кодировать токсин V.cholerae или его части, такие как субблок A1. Один выделенный мутант удаления заключает удаление в гене ctx, которое определено сайтами эндонуклеазы рестрикции ACC 1.
Способ выделения таких мутантов удаления Vibrio cholerae также описан и он содержит стадии:
(a) построения первой плазмиды, содержащей боковые последовательности Vibrio cholerae одного или нескольких удаленных фрагментов эндонуклеазы рестрикции, и ген для первого селектируемого маркера чужеродного происхождения, подвергнутый лигации с вышеупомянутыми боковыми последовательностями, чтобы заместить и занять место вышеупомянутого удаленного фрагмента, причем вышеупомянутые последовательности имеют достаточную длину, чтобы промотировать обнаруживаемую ин виво рекомбинацию;
(b) скрещивания вирулентного штамма Vibrio cholerae с первым микроорганизмом, несущим первую плазмиду;
(c) селекции Vibrio cholerae, экспрессирующего первый селектируемый маркер;
d) скрещивания продукта, выбранного на стадии (c), со вторым микроорганизмом, несущим вторую плазмиду со вторым селектируемым маркером, причем вышеупомянутая вторая плазмида не совместима с первой плазмидой;
(e) селекции Vibrio cholerae, экспрессирующего как первый селектируемый маркер, так и второй селектируемый маркер.
Описана вторая культура Vibrio cholerae, содержащая штамм Vibrio cholerae с удаленным первым фрагментом эндонуклеазы рестрикции ДНК, чтобы лишить вирулентности и сохранить способность колонизировать кишечник животного хозяина, и удаленным вторым фрагментом эндонуклеазы рестрикции, кодирующим токсин блокатора малой зоны (ZOT) с тем, чтобы уменьшить остаточный понос у животного-хозяина. Первый удаленный ДНК-фрагмент может кодировать токсин V. cholerae или его части такие, как субблок A1. Один выделенный мутант удаления содержит удаление в гене ctx, которое определено сайтами эндонуклеазы рестрикции Acc 1, и удаление в гене zot. Другой выделенный мутант удаления содержит удаление в гене ctx, которое определено сайтами эндонуклеаз рестрикции Xba и Cla 1, и удаление в гене zot, которое определено сайтами эндонуклеаз рестрикции stu 1 и Acc 1.
Описан также способ выделения таких мутантов удаления, содержащий стадии:
(a) конструкции первой плазмиды, содержащей боковые последовательности Vibrio cholerae одного или нескольких удаленных фрагментов эндонуклеаз рестрикции, и ген обнаруживаемого маркера чужеродного происхождения, подвергнутый лигации с вышеупомянутыми боковыми последовательностями, чтобы заменить и занять место вышеупомянутого удаленного фрагмента, причем вышеупомянутые последовательности должны иметь достаточную длину, чтобы промотировать обнаруживаемую ин виво рекомбинацию;
(b) скрещивания вирулентного штамма Vibrio cholerae с первым микроорганизмом, несущим первую плазмиду;
(c) селекции Vibrio cholerae, экспрессирующего первый селектируемый маркер;
(d) скрещивания выбранного продукта со стадии (c) со вторым микроорганизмом, несущим вторую плазмиду со вторым селектируемым маркером, причем вышеупомянутая вторая плазмида несовместима с первой плазмидой;
(e) селекции на экспрессию Vibrio cholerae как первого селектируемого маркера, так и второго селектируемого маркера;
(f) конструирования третьей плазмиды, несущей боковые последовательности Vibrio cholerae одного или нескольких удаленных фрагментов эндонуклеазы рестрикции, гомологичных тем, что были описаны на стадии (a), но отличающихся отсутствием селектируемого маркера чужеродного происхождения;
(g) скрещивания выбранного продукта со стадии (e) с третьим микроорганизмом, несущим третью плазмиду, описанную на стадии (f);
(h) селекции Vibrio cholerae, который уже не экспрессирует первый селектируемый маркер.
Этот способ может быть использован только для штаммов ZOT минус или для получения производного ZOT минус, из которого уже удалены гены токсина холеры.
Предложена третья культура Vibrio cholerae, содержащая штамм Vibrio cholerae, с удаленной областью хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны (ZOT). Предложен также способ выделения таких мутантов удаления Vibrio cholerae, содержащий стадии:
(a) конструирования плазмиды, содержащей последовательности Vibrio cholerae, кодирующие токсин холеры и токсин блокатора малой зоны, и ген селектируемого маркера чужеродного происхождения, причем вышеупомянутая плазмида не способна к репликации вне хромосом в Vibrio cholerae;
(b) скрещивания микроорганизма, несущего вышеупомянутую плазмиду, с вирулентным штаммом Vibrio cholerae, содержащим вышеупомянутые последовательности, вставленные между боковыми идентичными копиями второй последовательности, такой как RSI-элементы достаточной длины, чтобы промотировать обнаруживаемую ин виво рекомбинацию;
(c) селекции Vibrio cholerae, экспрессирующего вышеупомянутый селектируемый маркер;
(d) выращивания выбранного продукта со стадии (c) без селективного агента;
(e) селекции Vibrio cholerae, который уже не экспрессирует селективный маркер и, следовательно, содержит удаленную область хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны.
Предложена четвертая культура Vibrio cholerae, содержащая штамм Vibrio cholerae с удаленной областью хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны, и содержащий вставленный ген стойкости к ртути и ДНК, кодирующую субблок B токсина Vibrio cholerae. Предложен также способ выделения таких мутантов удаления, содержащий стадии:
(a) конструирования плазмиды, содержащей последовательности Vibrio cholerae, кодирующие токсин холеры и токсин блокатора малой зоны, и ген селектируемого маркера чужеродной природы, причем вышеупомянутая плазмида не способна к репликации вне хромосом в Vibrio cholerae;
(b) скрещивания микроорганизма, несущего вышеупомянутую плазмиду, с вирулентным штаммом Vibrio cholerae, содержащим вышеупомянутые последовательности, кодирующие токсин холеры и токсин блокатора малой зоны, вставленные между боковыми идентичными копиями второй последовательности достаточной длины, чтобы промотировать обнаруживаемую ин виво рекомбинацию;
(c) селекции Vibrio cholerae, экспрессирующего вышеупомянутый селектируемый маркер;
(d) выращивания выбранного продукта со стадии (c) без селективного агента;
(e) селекции Vibrio cholerae, который уже не экспрессирует селективный маркер и, таким образом, уже не имеет области хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны;
(f) конструирования второй плазмиды, содержащей ген стойкости к ртути и ДНК, кодирующую субблок B токсина Vibrio cholerae, и ген второго селектируемого маркера чужеродной природы, причем вышеупомянутая плазмида не способна к репликации вне хромосом в Vibrio cholerae, и в которой последовательности имеют достаточную длину, чтобы промотировать обнаруживаемую ин виво рекомбинацию, примыкающую к вышеупомянутому гену стойкости к ртути, и ДНК, кодирующей субблок B токсина Vibrio cholerae;
(g) скрещивания микроорганизма, несущего вышеупомянутую вторую плазмиду с вышеупомянутым Vibrio cholerae, описанным на стадии (e), содержащим последовательности, гомологические вышеупомянутым последовательностям достаточной длины, чтобы промотировать обнаруживаемую ин виво рекомбинацию;
(h) селекции Vibrio cholerae, экспрессирующего вышеупомянутый второй селектируемый маркер;
(i) выращивания выбранного продукта со стадии (h) без второго селективного агента;
(j) селекции Vibrio cholerae, который уже не экспрессирует второй селективный маркер;
(k) просеивания вышеупомянутого Vibrio cholerae, описанного на стадии (j), чтобы выделить Vibrio cholerae, которые содержат ген стойкости к ртути и ДНК, кодирующую субблок B токсина Vibrio cholerae, но у которых удалена область хромосомной ДНК, кодирующая токсин холеры и токсин блокатора малой зоны.
Мутанты удаления Vibrio cholerae, являющиеся предметом настоящего изобретения, являются полезными при вакцинации против холеры.
Один штамм Vibrio cholerae, являющийся предметом настоящего изобретения, именуемый CVD 101, как ожидается, обеспечивает по существу 100% эффективность для человека против последующего заражения штаммом аналогичного серотипа. Другие штаммы Vibrio cholerae, являющиеся предметом настоящего изобретения, именуемые второй культурой и третьей культурой, такие как CVD 109, как ожидается, обеспечивают по существу 100% эффективную защиту человека против последующего инфицирования штаммом аналогичного серотипа и позволяют избежать нежелательных побочных эффектов, таких как понос и тошнота, а также судороги. Еще один штамм Vibrio cholerae, являющийся предметом настоящего изобретения, CVD 110, именуется четвертой культурой.
Краткое описание фигур
Фиг. 1. V.cholerae N 16961 (pJBK55) (Apr).
Фиг. 2. Процедура переноса скрещенного и присоединенного гена при построении V.cholerae JBK56.
Фиг. 3. V.cholerae JBK56.
Фиг. 4. Схема конструкции JBK21.
Фиг. 5. Схема конструкции JBK54.
Фиг. 6. Схема конструкции V.cholerae JBK56.
Фиг. 7. Рекомбинация ин виво при помощи переноса и исключения гена ctx.
Фиг. 8. Схема конструкции pJBK51.
Фиг. 9. Схема конструкции для pCVD14 и pCVD15.
Фиг. 10. Схема конструкции pJBK108.
Фиг. 11. Схема конструкции pJBK107.
Фиг. 12. ДНК-последовательность (вверху) сайтов Xba 1 и CIa 1, которые определяют концы удаленного фрагмента Хва 1 - CIa 1 в 550 пар оснований (по) A-субблока в Огава 395, и (внизу) соединение с CVD 101 после удаления этого фрагмента и вставки Xba 1-линкера.
Фиг. 13A и 13B. Эффект верхнего слоя культуры V. cholerae на ток подвзвдошного короткого контура (Isc) и ионную проводимость ткани (Gt). Приведены средние значения для 6 животных в каждый момент времени; прямые скобки содержат 1 стандартную ошибку а, эффект верхних слоев V.cholerae 395 на Isc (сплошные линии) и Gt (штриховые линии), в, Эффект V.cholerae 395 (сплошная линия), CVD 101 (линия с длинными штрихами) и 395NI (точечная линия) на Gt. Контрольный эксперимент со средней (линия из коротких штрихов) состоял из непривитой культурной среды.
Фиг. 14, A - D. Анализ на проницаемость агглютинина пшеничного зерна-пероксидазы хрена обыкновенного (WGA - HRP) подвздошной ткани кролика под действием верхних слоев культур различных штаммов V.cholerae, а, контрольный для среды; b, V.cholerae 395; c, V.cholerae 395N1; d, V.cholerae CVD101.
Фиг. 15, A и B. Исследование замораживанием-изломом подвздошной ткани кролика под действием верхних слоев культур V.cholerae a, Интактный ZO с многочисленными пересечениями (острые стрелки) между примыкающими нитями M, микроворсинки. b, Нарушенный ZO из подвздошной ткани под действием V.cholerae 395; ретикулюм имеет упрощенный вид из-за значительного снижения сферы действия пересечений нитей. c, Квантование сложности ZO в тканях под действием верхних слоев культур или контрольного бульона.
Фиг. 16. Обратимость вариаций Gt, индуцированных верхним слоем V.cholerae 395. Добавляли верхние слои культур V.cholerae (треугольники) и непривитую среду (квадраты) и удаляли в моменты времени, указанные стрелками.
Фиг. 17. Схема конструкции CVD109. Гены zot и ctx примыкают друг к другу на хромосоме V. cholerae и находятся в области хромосомы, которая содержит несколько копий последовательности 2 700, именуемой RSI (повторяющейся последовательности). Элементы RSI находятся по обе стороны генов zot и ctx в вирулентном штамме V. cholerae E7946 (биотип Эль Тор, серотип Огава). Гены zot и ctx показаны большой незаштрихованной или заштрихованной стрелкой. RSI-последовательности изображены меньшей сплошной стрелкой.
Фиг. 18Аи 18В. ДНК-последовательность гена zot токсина блокатора малой зоны из нуклеотидов с номерами с 1 по 1428. Буквы над ДНК-последовательностью указывают на предсказанную аминокислотную последовательность ZOT-протеина, кодируемого геном zot.
Фиг. 19. Схема конструкции плазмиды pCVD621.
Фиг. 20А - 20С. Схема конструкции плазмиды pCVD622.2B.
Фиг. 21А - 20G. ДНК-последовательность гена ctx B и гена mer, вставленных в ген hlyA.
Сокращения для сайтов эндонуклеаз рестрикции на фигурах:
A = сайт эндонуклеазы рестрикции Acc I
B = сайт эндонуклеазы рестрикции Bgl II
C = сайт эндонуклеазы рестрикции CIa I
E = сайт эндонуклеазы рестрикции Eco RI
H = сайт эндонуклеазы рестрикции Hind III
P = сайт эндонуклеазы рестрикции Pst I
S = сайт эндонуклеазы рестрикции Sal I
X = сайт эндонуклеазы рестрикции Xba I
K = сайт эндонуклеазы рестрикции Kpn I
Другие сокращения на фигурах и в других местах:
Ap = ген стойкости к ампициллину
Apr = фенотип стойкости к ампициллину
Aps = фенотип чувствительности к ампициллину
Chrom = Хромосома
Cm = ген стойкости к хлорамфениколу
CT = токсин холеры
ctx = ген токсина холеры
CTA = ген субблока A токсина холеры
ctxA = ген субблока A токсина холеры
CTB = ген субблока B токсина холеры
ctxB = ген субблока B токсина холеры
hylA = ген гемолизина
kb = килооснований
mer = ген стойкости к ртути
p = плазмида
Su = сульфонамид
Sur = фенотип стойкости к сульфонамиду
Tc = тетрациклин
Tcs = фенотип чувствительности к тетрациклину
Tp = триметоприн
Zot = ген токсина блокатора малой зоны /Zonula occludens/.
Детальное описание настоящего изобретения
Основной принцип настоящего изобретения состоит в изоляции штамма вакцины Vibrio cholerae, специфическим образом измененного при помощи техники рекомбинантой ДНК, чтобы лишить его вирулентности без нарушения других компонент, необходимых для иммунитета. Такое ослабление осуществляют при помощи переваривания эндонуклеазами рестрикции плазмид, несущих соответствующие последовательности V. choleae, чтобы специфическим образом удалить гены, кодирующие токсин холеры или его участки. В результате переноса соединенного гена таких переваренных плазмид и последующих процедур селекции ин виво рекомбинантов с вирулентными хозяевами V.cholerae, получали штаммы без генов токсина и его частей. Должно быть ясно, что способы, являющиеся предметом настоящего изобретения, применимы к изоляции других мутантов удаления вирулентного V. cholerae, и к изоляции штаммов, содержащих полностью или частично такие удаленные последовательности, снова вводимые в клетку V.cholerae.
Исходным материалом для вакцины был токсигенный штамм Vibrio cholerae N 16961, который, как было установлено, вызывает у добровольцев характерный понос и сильный защитный иммунитет к последующей инфекции (Levine M.M. и др. , Acute enterie ..., см. выше, 1981). Область бактериальной хромосомы, которая, как было установлено, отвечает за продуцирование токсина холеры, клонируют в клонирующий переносчик плазмиды pBR325, после просеивания переваривания ферментом Hind III V.cholerae с использованием зонда, лабильного к теплу гена E.coli (Kaper и др., Amer. Soc., см. выше; Kaper и др. Sumposium, см. выше). Фрагмент хромосомы V.cholerae, как было установлено, содержит все гены, необходимые для продуцирования токсина. Далее эту хромосомную область анализировали и составляли карту точных частей, содержащих гены токсинов [Kaper, J.B. и др., Lancet 11, стр. 1162(1981)]. Ферменты рестрикции использовали для того, чтобы отсекать ДНК-фрагменты, содержащие эти гены, а ДНК-фрагмент, кодирующий селектируемый маркер (например, стойкость к ампициллину), вставляли при помощи лигации. Ген стойкости к ампициллину и боковую ДНК Vibrio затем клонировали в производном от pRK290, который можно перенести из E. coli в V.cholerae. Полученную в результате плазмиду, pJBK55, переносили из t K-12 E.coli в N16961 V.cholerae при помощи конъюгации.
Полученный в результате штамм, V.cholerae N16961 (pJBK55) (Apr), содержал область в своей хромосоме, содержащей интактные гены токсина и, во внехромосомном состоянии, плазмиду, содержащую такую же область с удаленными генами токсинов и замененным геном ампициллина (см. фиг. 1). С низкой частотой, видимо в одном случае из 106 - одном случае из 108, идентичные области, окружающие хромосомные гены токсина, и внехромосомный (плазмидный) ген стойкости к ампициллину, будут меняться и "переходить" или претерпевать ин виво рекомбинацию так, что область ДНК, содержащая этот ген стойкости, вытесняет ген токсина на хромосоме (фиг. 12). Этот редкий случай выбирают при помощи тестирования смеси мутированных и немутированных клеток на отдельных клетках, которые способны служить в качестве хозяина для введения несовместимой плазмиды [Ruvkun, G. B. и др., Nature, т. 289, стр. 85 (1981)]. Плазмиды разбивали на группы, обозначенные через A-W, элементы которых не могут сосуществовать стабильно друг с другом. Например, плазмида из группы несовместимости p не может стабильно поддерживаться в той же клетке, что и другая плазмида группы P (Inc P). Таким образом, плазмиды Inc P такие, как R702, которые характеризуют стойкость к сульфонамиду, не могут поддерживаться в клетке, которая содержит другую плазмиду Inc P, такую как PRK 290, pJBK45 или P JBK55. Поэтому R702 не может поддерживаться в штамме, в котором была рекомбинирована стойкость к ампициллину в хромосоме, но не в штамме, в котором плазмида Inc P (например, pJBK55) подвергается репликации вне хромосомы. При помощи скрещивания штамма E.coli, содержащего Inc PR702 (стойкость к сульфонамиду), и p JBKK55 V.cholerae (стойкость к ампициллину), и селекции V.cholerae, которые являются стойкими как к ампициллину, так и сульфонамиду, выделяли колонии, в которых стойкость к сульфонамиду устанавливается с внехромосомным участием p702, а стойкость к ампициллину устанавливается в хромосоме через замену геном стойкости к ампициллину гена токсина (фиг. 3). Один такой штамм, именуемый как V.cholerae JBK56, выделяли и при испытании на продуцирование токсина устанавливали, что он нетоксиногенный.
Окончательную версию штамма вакцины, JBK70, получали при помощи замены стойкости к ампициллину, используемому в терапии антибиотику, стойкостью к ртути. Эту замену осуществляют при помощи клонирования гена стойкости к ртути непосредственно в ген стойкости к ампициллину p JBK55, при этом дезактивируют стойкость к ампициллину и активируют стойкость к ртути. Полученная в результате плазмида p JBK66 также несовместима с R702 и ее переносят в V.cholerae JBK56. Мутант, в котором стойкость к ртути восстановлена в хромосоме, подвергали селекции, используя плазмиду R702 Inc P, и выбирая V. cholerae, которые чувствительны к ампициллину, стойки к ртути и стойки к сульфонамиду. Спонтанное производное позже выбирали так, чтобы он включал pR702. Финальный мутант, JBK70, был нетоксиногенным и стойким только к ртути.
Штамм вакцины V.cholerae JBK70 является штаммом серотипа Инаба. Другим основным серотипом V.cholerae был серотип Огава. Можно ожидать, что вакцина, полученная из одного серотипа, будет защищать против другого серотипа (34). В том случае, когда это не так, штамм живой вакцины может быть получен из серотипа Огава и при защите добровольцев /Levine, M.M. и др., Acurt enter... , см. выше (1981)/. Точная мутация, созданная в штамме Инаба JBK56 V.cholerae, вновь создавалась в штамме E7946 при помощи непосредственного переноса области хромосомы, содержащей стойкость к ампициллину вместо гена токсина в JBK56 в E 7946, через генетическую рекомбинацию с участием P, секcфактора V. cholerae /Parker, C. и др., см. выше/. P - фактор, который отличается от плазмиды Inc P, переносили в JBK56, а затем скрещивали с мутантом стойкости к рифампину E7946. При помощи селекции мутанта, который был стойким как к ампициллину, так и рифампину, выделяли штамм вакцины, который имел серотип Огава с полностью удаленными генами токсинов.
Если антибактериальный иммунитет является недостаточным для защиты, то антитоксичную компоненту можно добавить при помощи повторного присоединения генов для продуцирования токсина B холеры, но не субблока A. Эту процедуру осуществляют при помощи клонирования гена субблока B в клонирующий вектор pMS89. Полученная в результате плазмида, pJBK51, продуцирует высокие концентрации субблока B и ее снова вводят в штамм нетоксичной вакцины V.cholerae JBK70, чтобы получить штамм ослабленной вакцины JBK70 /JBK51/, который не может продуцировать субблок A.
Штаммы вакцины, являющиеся предметом настоящего изобретения, получают среди прочего из V. cholerae N 16961, имеющего серотип Инаба. Должно быть ясно, что другие штаммы или другие биотипы и серотипы можно использовать для того, чтобы заменить N 16961, чтобы получить штаммы вакцины, имеющие специфические удаления в гене или генах ctx, или других местах вдоль хромосомы V. cholerae. Так как целью изоляции таких штаммов вакцины являются мимики, процесс инфицирования без сопутствующих патологических явлений, направленный на сайты мутагенез вирулентных штаммов, как это описано в настоящей патентной заявке, обеспечивает существенные возможности при профилактической вакцинации против холеры.
Например, заявитель получил еще один штамм вакцины V.cholerae CVD101, отличающийся удалением большей части гена субблока A в 2 копиях гена ctx. Можно ожидать, что эффективность CVD101 по существу близка к 100%, так как родительский штамм 395 обеспечивает 100% эффективность.
При конструировании CVD101 следуют в общем случае принципам, описанным выше, например, при конструкции JBK70; при этом можно ожидать, что полученный в результате CVD101 не имеет гена стойкости, который требует исправления. Последняя стадия при изоляции второго и нахождении ин виво рекомбинанта включает схему для селекции чувствительности к антибиотику, например, чувствительности к тетрациклину, тогда как родительский штамм вставляли в расположение гена A CT, гена стойкости к тетрациклину. Ясно, что такая антибиотическая чувствительности является еще одним примером селектируемого маркера.
Продуцирование штаммов вакцины может быть осуществлено при помощи самых разнообразных приемов, включая следующие: Vibrio cholerae субкультивировали из сырьевых культур в агаре инфузии клеток головного мозга /сердца /BHIA/ и выращивали при 37oC в течение ночи. Идентичность испытывают при помощи антисыворотки, специфической относительно группы и типа, а двадцать-тридцать колоний суспендировали в бульон BHI. На предварительно инкубированные пластики BHIA прививали суспензию BHI. После инкубирования в течение пяти-шести часов содержимое каждой пластинки собирали в 5 мл стерильного соляного раствора, буферированного до pH 7,2 ± 0,1. Собранные организмы подвергали центрифугированию в холоде со скоростью 750 g в течение десяти минут, снова суспендировали и промывали дважды в четырехкратном первоначальном объеме. Суспензию подвергали стандартизации под спектрофотометром и разбавляли до примерно того количества организмов, которое требуется для вакцинации (примерно 106, которое варьируется в зависимости от результатов исследования на добровольцах). Материал для репликации, количественные культуры на пластинках, получали из прививки перед и после введения веществ, чтобы подтвердить размер прививки. Финальную прививку анализировали штаммом Грама и подвергали агглютинации с гомологической антисывороткой перед введением.
Штаммы Vibrio cholerae, являющиеся предметом настоящего изобретения, могут быть применены стоматическим способом. Два грамма NaHCO3 растворяли в пяти унциях дистиллированной воды. Добровольцы выпивали четыре унции (1 унция = 28,3 г) NaHCO3/воды; одну минуту спустя добровольцы принимали вибрионы, суспендированные в оставшейся одной унции NaHCO3/воды. Добровольцев обследовали за девяносто минут до прививки и через девяносто минут после.
Что касается безопасности, то основной момент состоит в том, что штамм вакцины не возвращается к токсигенности (т.е., продуцирует интактный токсин холеры), что могло бы вызвать заболевание. Двумя основными анализами при испытании токсина являются анализ клеток надпочечников V-I [Sack, D.A., и др., Infect Immun., т. 11, стр. 334 (1975)] и анализ с использованием фермента, связывающего иммуносорбент (ЭЛИЗА) [Sack, D.A. и др., J. Clin. Micro., т. 11, стр. 35 (1980)]. Штамм вакцины /JBK70/ повторно испытывали в этих двух анализах и обнаруживали, что он отрицательный в каждом случае. Гораздо более важными, однако, являются генетические анализы, осуществляемые на присутствие генов токсина. ДНК генов токсина холеры может быть помечена радиоактивным методом и использована в качестве специфического зонда для идентификации других генов токсина холеры в штамме, следуя процедуре Саутерна, E.M., J. Mol. Bio, т. 98, стр. 503 (1975). При испытании таким методом штамм вакцины, предлагаемый в соответствии с настоящим изобретением, не содержал обнаруживаемого генетического материала, который может включать токсин холеры. Эту вакцину также испытывали на модели, молодых мышах, в соответствии с Baselski, V. и др., Infect. Immun., т. 15, стр. 704 (1977). После повторной серии прохождения (всего десять) никаких признаков накопления жидкости не было обнаружено (а значит, и признаков заболевания). Как и ожидалось, было установлено, что JBK70 колонизирует кишечник молодых мышей.
Для того чтобы избежать побочных эффектов при вакцинации штаммами, таких как понос и рвоты, судороги и другие симптомы, штаммы вакцины могут быть также лишены второго фрагмента эндонуклеазы рестрикции ДНК, кодирующей токсин блокатора малой зоны (ZOT).
Культура Vibrio chоlerae содержит штамм Vibrio chоlerae с удаленным фрагментом ДНК эндонуклеазы рестрикции, чтобы лишить вирулентности и сохранить способность колонизировать кишечник животного-хозяина, у которого можно удалить второй фрагмент эндонуклеаз рестрикции ДНК, кодирующий токсин блокатора малой зоны (ZOT), чтобы уменьшить болезненный понос у животного-хозяина. Первый удаленный ДНК-фрагмент может кодировать токсин V.cholerae или его части, такие как A1-субблок. Один выделенный мутант удаления содержит удаление в гене ctx, которое определено сайтами эндонуклеазы рестрикции Acc 1, и удаление в гене zot. Другой выделенный мутант удаления содержит удаление в гене ctx которое определено сайтами эндонуклеаз рестрикции Xba 1 и Cla 1, и удаление в гене zot, которое определено сайтами эндонуклеаз рестрикции Stu 1 и Acc 1.
Способ выделения таких мутантов удаления Vibrio chоlerae содержит стадии:
(a) конструирования первой плазмиды, содержащей боковые последовательности Vibrio chоlerae одного или нескольких удаленных фрагментов эндонуклеазы рестрикции и ген селектируемого маркера чужеродного происхождения, подвергнутый лигации с вышеупомянутыми боковыми последовательностями, чтобы заменить и занять место вышеупомянутого удаленного фрагмента, причем вышеупомянутые последовательности имеют достаточную длину, чтобы промотировать обнаруживаемую ин виво рекомбинацию;
(b) скрещивания вирулентного штамма Vibrio cholerae с первым микроорганизмом, несущим первую плазмиду;
(c) селекции Vibrio cholerae, экспрессирующего первый селектируемый маркер;
d) скрещивания продукта, выбранного на стадии (c) со вторым микроорганизмом, несущим вторую плазмиду со вторым селектируемым маркером, причем вышеупомянутая вторая плазмида не совместима с первой плазмидой;
(e) селекции Vibrio cholerae, экспрессирующего как первый селектируемый маркер, так и второй селектируемый маркер;
(f) конструирования третьей плазмиды, содержащей боковые последовательностям Vibrio cholerae одного или нескольких удаленных фрагментов эндонуклеаз рестрикции, гомологичных последовательностям, описанным на стадии (a), но отличающихся отсутствием селектируемого маркера чужеродного происхождения;
(g) скрещивания выбранного продукта со стадии (e) с третьим микроорганизмом, несущим третью плазмиду, описанную на стадии (f);
(h) селекции Vibrio cholerae, который уже не экспрессирует первый селектируемый маркер.
Этот способ можно использовать только для штаммов ZOT минус или для продуцирования ZOT минус-производного штамма, у которого уже удалены гены токсина холеры.
Другая культура Vibrio cholerae содержит штамм Vibrio cholerae с удаленной областью хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны (ZOT). Способ изоляции таких мутантов удаления Vibrio cholerae содержит стадии:
(a) конструирование плазмиды, содержащей последовательности Vibrio cholerae, кодирующие токсин холеры и токсин блокатора малой зоны, и ген селектируемого маркера чужеродного происхождения, причем вышеупомянутая плазмида не способна к репликации вне хромосомы в Vibrio cholerae;
(b) скрещивания микроорганизма, несущего вышеупомянутую плазмиду, с вирулентным штаммом Vibrio cholerae, содержащим вышеупомянутые последовательности, чтобы промотировать обнаруживаемую ин виво рекомбинации;
(c) селекции Vibrio cholerae, экспрессирующего вышеупомянутый селектируемый маркер;
(d) выращивания выбранного продукта со стадии (c) без селективного агента;
(e) селекции Vibrio cholerae, который уже не экспрессирует селективный маркер; и, таким образом, у которого удалена область хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны. Стадия (b) может содержать: (a) скрещивание микроорганизма, несущего вышеупомянутую плазмиду, с вирулентным штаммом Vibrio cholerae, содержащим вышеупомянутые последовательности, вставленные между боковыми идентичными копиями второй последовательности, такой как RSI-элементы, достаточной длины, чтобы промотировать обнаруживаемые ин виво рекомбинации.
Мутанты удаления Vibrio cholerae, являющиеся предметом настоящего изобретения, можно использовать для вакцинации против холеры.
Здесь описан новый токсичный фактор, вырабатываемый V.cholerae, который увеличивает проницаемость малой слизистой оболочки в результате нарушения структуры межклеточных плотных соединений, или блокатор малой зоны (ZO) (межклеточных путей переноса ионов). Продуцирование этого фактора штаммом V. cholerae коррелирует с болезненными поносами у добровольцев. При помощи нарушения стандартных процессов поглощения в малой кишке через межклеточный путь этот фактор может быть ответственным за остаточный понос, вызываемый мутантами удаления ctx из V.cholerae, и может вносить вклад в болезненный понос, который отличает холеру от других, сопровождающихся поносом, заболеваний.
Были проведены исследования изменений функции кишечника, которые вызываются тремя штаммами V. cholerae, одним дикого типа и двумя ослабленными штаммами вакцины. Штамм V.cholerae 395, классический биотип, серотип Огава, является высоковирулентным штаммом, который широко исследовали на добровольцах в Центре Разработок Вакцин. Этот штамм вызывает понос со средним объемом стула в 5,5 литров (вообще говоря, от 0,3 до 44 л) у более, чем 90% добровольцев, которым через рот вводили 106 организмов [Levine, M.M., и др. Infect. Immun., т. 56, стр. 161-167 (1988)]; [Levine, M.M., "Cholera and Related Diarrheas", стр. 195-203] [изд. Karger, Basel, 1980]. Холерные поносы вызываются главным образом ферментативными воздействиями A-субблока CT на слизистую оболочку кишечника. Субблок A CT, кодируемый геном ctx, стимулирует аденилат циклазу и приводит к чистому выделению жидкости в полость кишечника [Gill, D. M. , Adv. Cyclic Nucleotide res., т. 8, стр. 85-118 (1977]. Штамм CVD 101 вакцины V.cholerae является мутантом удаления ctx 395, в котором 94% последовательности, кодирующей A1-пептид CT, удалены. Неожиданным образом, хотя CVD 101 уже не продуцирует активный CT, этот штамм вызывал (от среднего до незначительного) понос (средний объем стула 0,9 л с разбросом от 0,3 до 2,1 л) у 54% добровольцев, которым вводили этот организм. Второе производное 395, штамм 395 N1 вакцины, полученное Мекаланосом и др., Nature т. 306, стр. 551-557 (1983), не содержит примерно 77% последовательностей, кодирующих A1-пептид, как подсчитал Заявитель. В контрасте с CVD101 штамм 395N1 вызывал очень умеренный понос (объем стула 0,3 л) только у одного из 21 добровольца (P = 0,002, по сравнению с 13 из 24 добровольцев, у которых был понос после введения CVD101) [Herrington, D.A. и др., J. Exp. Med., т. 168, стр. 1487-1492 (1982)] . Так как эти штаммы были похожи по их способности колонизировать кишечник, заявитель предположил, что CVD101 продуцирует секретогенный фактор, который экспрессируется слабо или вовсе не экспрессируется штаммом 395N1, и что этот фактор отвечает за понос, который наблюдали у добровольцев, принявших CVD101.
Эти штаммы изучали с использованием ткани кишечника кролика, установленной в камерах Юссинга, классическом средстве для изучения процесса переноса через ткань кишечника. Верхние слои культур V.cholerae добавили в эти камеры, и измеряли разность потенциалов (PD) и ток в коротком контуре (Isc). PD является разностью потенциалов, измеренной на стороне слизистой оболочки относительно серозной стороны этой ткани, а Isc является количеством тока, необходимым, чтобы занулить PD. На основании этих измерений проводимость ткани (GT) рассчитывали с использованием закона Ома: Isc = PD х GT. Заявитель сначала изучал эффект верхних слоев штамма 395 дикого типа на эти параметры, используя непривитую культурную среду, добавленную в подобранную подвздошную ткань из того же животного, что и отрицательное контрольное испытание. На фиг. 13A представлены полученные изменения Isc и GT. Первоначальные пики в Isc и PD, которые появляются как в отрицательных контрольных, так и в испытываемых образцах, наиболее вероятно вызваны одновременным переносом Na и питательных веществ, содержащихся в среде. В отрицательном контрольном испытании Isc и PD возвращались к значениям начала отсчета спустя примерно один час, а затем Isc, PD и Gt оставались неизменными до завершения эксперимента. Напротив, ткани, оказавшиеся под воздействием верхнего слоя штамма 395, давали значительное увеличение Gt, достигая максимального значения спустя 2 часа инкубирования. В таких образцах Isc никогда не возвращался к началу отсчета, но период равновесного состояния для Isc отмечали в диапазоне от 40 до 60 минут. Так как Isc эквивалентно PD x GT, а отмеченное PD спустя 60 минут было аналогично начальному значению (данные не приведены), существенное увеличение Isc в ткани, обработанной 395, в этот момент времени может быть только вызвано увеличением Gt (см. фиг. 13A, время 60 мин) (12). Через 60 мин Isc начинал увеличиваться снова вместе с PD в обработанных при помощи 395 тканях. Эта вторая фаза, видимо, отражает эффекты токсина холеры на истечение ионов, так как очищенный CT увеличивает Isc с подвздошной ткани кролика только спустя время задержки не менее 40 минут. Эти данные наводят на мысль, что имюется два фактора, которые экспрессируются в V.cholerae 395, которые могут изменять перенос ионов в камерах Юссинга. Один фактор, токсин холеры, вызывает увеличение Isc и PD, начиная с примерно 60 минуты после добавления верхнего слоя культуры в то время, как второй фактор индуцирует немедленное увеличение в проводимости ткани, которое можно наблюдать в течение 20 минут после добавления верхнего слоя культуры.
Далее изучали изменение Gt, вызванное верхними слоями культур ослабленных штаммов CVD 101 и 395NI V.cholerae. CVD101 индуцировал немедленное увеличение Gt, которое нельзя было отличить от увеличения, которое отмечали с 395 (фиг. 13B). Напротив, 395N1 не индуцировал немедленного увеличения Gt; изменением Gt в обработанных 395N1 тканях было аналогично негативному контрольному бульону и существенно ниже, чем изменение, отмеченное для 395 и CVD101, в течение почти 100 минут инкубирования. Спустя это время, изменения Gt в тканях под действием 395, CVD101 и 395N1 аналогичны. Эти результаты наводят на мысль, что 395N1 продуцирует меньшие количества или менее активную форму фактора, отвечающего за такое увеличение Gt.
Вариации в трансэптелиальной проводимости отражают изменение в проницаемости ткани через межклеточное пространство, так как стойкости мембран плазмы относительно высоки. Так как ZO представляет основной барьер на таком межклеточном пути и вариация в Gt является наиболее чувствительной мерой ZO-функции, анализировали морфологические модификации ZO, вызванные верхними слоями V. cholerae 395, CVD101 и 395N1. Если маркер плотности электронов с низким молекулярным весом такой, как агглютинин пшеничных зерен - пероксидаза хрена обыкновенного (WGA-HRP), добавляли на сторону слизистой оболочки слоя эпителия, то он в общем случае не выходил за пределы ZO [Alberts, B. и др., "Molecular Biology of the Cell", 2-е изд. (1989)]. WGA-HRP добавляли на сторону слизистой оболочки ткани кишечника, обработанной верхними слоями культур 395, CVD101, 395N1 или непривитым контрольным бульоном в течение 60 минут. Как показано на фиг. 14, ткани, обработанные непривитой культурной средой, непроницаемы для WGA-HPR (фиг. 14,A), в то время как ткани, обработанные 395 и CVD101, указывают на наличие окраски в межклеточном пространстве (фиг. 14,B и 14,D). Ткани, на которые воздействовали верхние слои 395N1, не подвергались изменениям, так как межклеточное пространство оставалось достаточно узким, чтобы исключить проникновение WGA-HRP (15C). Эти результаты подтверждали и распространяли, используя электронную микроскопию трещин при замораживании, при котором количество нитей, лежащих параллельно ZO, коррелирует с трансэпителиальной электрической проводимостью. Ткани, на которые воздействовали верхние слои культур, указывали на смесь неизменного ZO (фиг. 15,A) измененного ZO с уменьшенной сложностью нитей (фиг. 15,B). Нити, лежащие перпендикулярно длинной оси ZO, оказывались в основном удаленными, что приводило к уменьшенному числу пересечений нитей. Сложность ZO, находившегося под действием верхнего слоя каждого штамма, разделяли при помощи измерения плотности пересечений нитей. Как можно видеть на фиг. 15,C, ткани, обработанные верхними слоями культур 395 или CVD101, показывали значительное снижение числа нитей и сложности ретикулюма ZO, если сравнивать с тканями, обработанными непривитым бульоном или верхними слоями 395N1.
Изменения ZO-морфологии, вызванные 395 и CVD101, параллельны увеличивающейся проводимости ткани, вызванной этими штаммами. Функция кишечного ZO заключается в том, чтобы регулировать параклеточный путь и ограничивать или предотвращать диффузию растворимых в воде молекул через межклеточное пространство снова в полость. Эта диффузия вызывается градиентами концентраций, которые создаются процессами трансэпитеального переноса. Как следствие изменения параклеточного пути, слизистая оболочка кишечника становится более проницаемой и вода, Na и Cl просачиваются в полость, что приводит к поносу. Изменение параклеточного пути, вызванное V.cholerae 395 и CVD101, является специфическим для малой кишки; замена слепой кишки кролика на подвздошную ткань не приводит к изменению Gt, вызываемому верхним слоем 395 (данные не показаны). Здесь впервые приводятся результаты о бактериальном факторе, который способен ослаблять жесткие соединения в интактной ткани кишечника, и который отражает новый механизм бактериального поноса. Как было установлено, токсин A Clostrildium difficile, грипп и везикулярный стоматит (YSY) ослабляют жесткость соединения в монослоях культуры ткани, но такая активность в интактной ткани и корреляция с поносом ранее не были известны.
Таким образом, V.cholerae 395 и CVD101 продуцируют фактор, который может отвечать за понос, отмечаемый у добровольцев, принимавших мутанты удаления V.cholerae. Понос, вызываемый такими ctx-мутантами, эквивалентен поносу, отмеченному для многих штаммов энтеротоксигенной E.coli. Этот секретогенный фактор, который Заявитель называет ZOT для токсина блокатора малой зоны, вызывает раннее увеличение Isc и проводимости ткани, которое не связано с эффектами CT на поток ионов. Это увеличение Gt связано с ослаблением жестких соединений, эффектом, который быстро обращается после удаления верхнего слоя (фиг. 16). Быстрое обращение этого эффекта находится в контрасте с продолжительными эффектами CT. Эти результаты не учитывают ранее необъясненные наблюдения Fields и др., J. Clin. Invest., т. 51, стр. 796-804 (1972), которые отметили немедленное увеличение Isc, вызванное сырыми, но не очищенными CT-препарациями, но могут объяснить результат Nishibuchi и др., Infect. Immun. , т. 40, стр. 1083-1091 (1983), которые заметили раннее накопление жидкости (FA), не связанное с запаздыванием CT-индуцированного FA у молодых мышей, которым вводили V.cholerae. Способность CT-отрицательной V.cholerae вызывать понос у добровольцев коррелирует с продуцированием ZOT двумя ослабленными штаммами, полученными от одного и того же родительского штамма; CVD101 (вызывающий понос) продуцирует ZOT в то время, как 395N1 (не вызывающий поноса) продуцирует мало ZOT-активности или вовсе ее не продуцирует.
Другая культура Vibrio cholerae содержит штамм Vibrio cholerae с удаленной областью хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны, но содержащий вставленный ген стойкости к ртути и ДНК, кодирующую субблок B токсина Vibrio cholerae. Способ выделения таких мутантов удаления, также являющийся предметом настоящего изобретения, содержит стадии:
(a) конструирования плазмиды, содержащей последовательности Vibrio cholerae, кодирующие токсин холеры и токсин блокатора малой зоны, и ген селектируемого маркера чужеродного происхождения, причем вышеупомянутая плазмида не способна к репликации вне хромосомы в Vibrio cholerae;
(b) скрещивания микроорганизма, несущего вышеупомянутую плазмиду, с вирулентным штаммом Vibrio cholerae, содержащим вышеупомянутые последовательности, кодирующие токсин холеры и токсин блокатора малой зоны, вставленные между боковыми идентичными копиями второй последовательности достаточной длины, чтобы промотировать обнаруживаемую ин виво рекомбинации;
(c) селекции Vibrio cholerae, экспрессирующего вышеупомянутый селектируемый маркер;
(d) выращивания выбранного продукта со стадии (c) без селективного агента;
(e) селекции Vibrio cholerae, который уже не экспрессирует селективный маркер и, таким образом, не имеет области хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны;
(f) конструирования второй плазмиды, содержащей ген стойкости к ртути и ДНК, кодирующую субблок B токсина Vibrio cholerae, и ген второго селектируемого маркера чужеродного происхождения, в котором вышеупомянутая плазмида не способна к репликации вне хромосомы в Vibrio cholerae, и в которой последовательности имеют достаточную длину, чтобы промотировать обнаруживаемую ин виво рекомбинацию, примыкающую к вышеупомянутому гену стойкости к ртути, и ДНК, кодирующей субблок B токсина Vibrio cholerae;
(g) скрещивания микроорганизма, несущего вышеупомянутую вторую плазмиду, с вышеупомянутым Vibrio cholerae, описанным на стадии (e), содержащим последовательности, гомологические относительно вышеупомянутых последовательностей, достаточной длины, чтобы промотировать обнаруживаемую ин виво рекомбинацию;
(h) селекции Vibrio cholerae, экспрессирующего вышеупомянутый второй селектируемый маркер;
i) выращивания выбранного продукта на стадии (h) без второго селективного агента;
(j) селекции Vibrio cholerae, который уже не экспрессирует второй селективный маркер;
(k) просеивания вышеупомянутого Vibrio cholerae, описанного на стадии (j), с целью выделения Vibrio cholerae, который содержит ген стойкости к ртути и ДНК, кодирующую субблок B токсина Vibrio cholerae, и не содержит области хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны.
Этот способ выделения мутантов удаления Vibrio cholerae, не имеющих области хромосомной ДНК, кодирующей токсин холеры и токсин блокатора малой зоны, и содержащих вставки гена стойкости к ртути и ДНК, кодирующей субблок B Vibrio cholerae, может использовать на стадии (f) боковые последовательности достаточной длины, содержащие ген, который может быть разрушен без нарушения колонизации и иммунитета Vibrio cholerae. Примером является ген гемолизина. CVD101 Vibrio cholerae конструировали в соответствии с этой процедурой, и он не содержит области хромосомной ДНК, кодирующей субблоки A и B токсина холеры и токсина блокатора малой зоны, но включает ген стойкости к ртути и ДНК, кодирующую субблок B токсина Vibrio cholerae, вставленные в сайт гена гемолизина. Другие примеры последовательностей достаточной длины содержат ген his [Hone, Microbial Pathogenesis, т. 5, стр. 407-478 (1989)] и ген nan H [Virm. J. of Bacteriology, т. 170, стр. 1495-1504 (1988)].
В примерах, которые следуют ниже, все приемы, реакции и процедуры разделения являются уже хорошо известными в этой области техники. Все ферменты, если не оговорено противное, являются доступными из одного или нескольких источников таких, как фирма New England Biolabs - Beverly, Massachusetts; Collaborative Research - Waltham; Massachusets; Miles Laboratories-Elkhart, Indiana, Boehringer Biochemicals Inc. - Indianapolis, Indiana и фирма Bethesda Research Laboratories-Rockville, Maryland, если перечислять только некоторые из них. Буферы и условия реакции для переваривания ферментами рестрикции используются те, что рекомендованы производителями для каждого фермента, если не указано противное. Частичные переваривания ферментами рестрикции осуществляли с использованием пониженных концентраций ферментов, которые должны быть предварительно определены из предварительных экспериментов для каждой порции фермента. Стандартная методология для других ферментативных реакций, разделений с использованием гелевого электрофореза и трансформаций E. coli могут быть найдены в книге "Methods in Enzymology", том 68, ред. Ray. Wu, изд. Academic Press (1979). Другой стандартной ссылкой является Maniatic, T. и др. , Molecular Cloning, изд. Cold Spring Harbor (1982). Бактерии выращивали в соответствии с приемами, описанными Миллером, Experiments in Molecular Genetics, изд. Cold Spring Harbor Laboratory (1972). Vibrio cholerae размножали в соответствии с процедурами, описанными в общем случае у Lennett, E.A. и др., ред. Manual of Clinical Microbiology, 3-e издание, American Society of Microbiology, Washington (1980). E.coli и Vibrio cholerae скрещивали в соответствии с процедурами, описанными в общем случае Johnson, Steven R. и др., J. Bact., т. 137, стр. 531 (1979); и Yokata T. и др., J. Bact., т. 109, стр. 440 (1972).
Штаммы, относящиеся к настоящему изобретению, сданы на хранение в Американское Собрание Типов Культур, расположенное в Роквилле, Мерилэнд, перед началом рассмотрения настоящей патентной заявки. Сданы на хранение штаммы V. cholerae JBK56, V. cholerae JBK70, V. cholerae N1696, V.cholerae JYK70 (pJBK51), V. cholerae Огава 395, CVD101, CVD109, V.cholerae E7946 и E.coli SM10 ламбда pir pCVD51, V. cholerae CVD110 и E. coli SY327 ламбда pir (pCVD622.2B), которым присвоены шифры хранения 39 317, 39 318, 39 315, 39 316, 39 541, 39 540, 55 057 (cданы 4 июня 1990 г.), 55 056 (сдан 4 июня 1990 г. ), 68 335 (сдан 5 июня 1990 г.), 55 188 (сдан 3 июня 1991 г.) и 68 630 (сдан 3 июня 1991 г.
Пример 1
Конструкция плазмиды, содержащей ген селектируемого маркера, вставленный, чтобы заменить гены токсинов
Плазмида JBK16 содержит PstI-BgI II-фермент в 4 ко (киллооснований - пер.) хромосомы, содержащей гены токсинов. По бокам генов токсинов расположены сайты Acc 1 и они сами содержат внутренний сайт Acc 1. JBK16 переваривали, чтобы дополнить еще Acc 1, и Acc 1-фрагменты, содержащие гены токсинов, отделяли от оставшейся части плазмиды. Оставшиеся накладывающиеся или "липкие" Acc 1-концы превращали в тупые при помощи "заполнения" фрагментом Клелоу полимеразы E.coli (т.е. однонитевую ДНК, оставшуюся после переваривания Acc 1, превращали в двухнитевую с дополняющими концами). Ген, кодирующий стойкость к ампициллину, очищали из плазмиды pREG 153 (pREG 153 является производным PREG 151 (Weiss, A. и др., J. Bact., т. 152, стр. 549-552), измененной в результате замены стойкостью к ампициллину стойкости к триметоприну и добавления последовательностей cos), а "липкие" концы "заполняли" как описано выше. Этот фрагмент далее подвергали лигации с ДНК вибриона так, чтобы гены стойкости Ap были в точности на том месте, где были только что удаленные гены токсинов, и окружены такими же последовательностями вибриона. Полученную в результате плазмиду обозначали pJBK21 (фиг. 4); она не содержала области токсина, но включала ген стойкости Ap.
Пример 2
Добавление боковых гомологических последовательностей с последующим переносом удлиненного гена в V.cholerae
Чтобы обеспечить специфическую вставку в хромосому удаления в pJBK21, к каждому концу Pct-BgI II-фрагмента из p JBK21 добавляли приблизительно 7 000 по дополнительной ДНК. (Вероятность события гомологической рекомбинации увеличивается с увеличением длины боковых гомологических последовательностей.) Чтобы обеспечить это, фрагмент в приблизительно 18 ко клонировали из хромосомы из N16961. Этот клон обозначали pJBK544 и он содержал фрагмент гена tox Pct-BgI в 4 ко, окруженный последовательностями ДНК в приблизительно 7 ко с каждой стороны (см. фиг. 5). Плазмиду pJBK 21 частично переваривали ферментом Pct 1 так, чтобы только один из сайтов Pct отсекался (дополнительный Pct-сайт добавляли внутри гена стойкости к ампициллину), а затем переваривали BgI II, чтобы выделить Pct-BgI II-фрагмент в 4 ко, содержащий область токсина удаления и область стойкости Ap. Плазмиду рJBK44, содержащую фрагмент вибриона в примерно 18 ко, частично переваривали BgI II так, чтобы только один из 4 сайтов BgI II отсекался. За таким частичным перевариванием следовало полное переваривание Pst1, а полученные в результате фрагменты разделяли при помощи электрофореза на 0,3% агарозе. Разделенные фрагменты затем подвергали очистке и анализировали, и был найден один фрагмент, который содержал все последовательности из pJBK44, за исключением фрагмента гена tox Pst-BgI в 4 ко (см. фиг. 5). Этот фрагмент, представляющий боковую ДНК, затем смешивали для лигации с Pst-BgI-фрагментом из pJBK21, содержащим стойкость к ампициллину. Полученная в результате плазмида pJBK54 содержала приблизительно 17 ко хромосомы Vibrio cholerae с геном стойкости к ампициллину, заменяющим удаленные гены токсинов.
Модифицированную хромосомную область клонировали в плазмиду, которую легко можно переносить в V.cholerae. Плазмида pRK290 (Ditta, G. и др., Proc. Nat. Acad. Sci, т. 77, стр. 7347 (1980)) принадлежит к группе P несовместимости плазмид и она обладает единственным сайтом Eco RI, в котором клонировали pJBK54 (фиг. 6). Полученную в результате плазмиду pJBK55 затем скрещивали в V.cholerae N16961, используя "сшивающую" плазмиду pKR2013, после чего получали V.cholerae N16961 (PJBK55) (Apr).
Пример 3
Рекомбинация ин виво
Мутантные гены токсинов, после переноса удлиненного гена, как это описано в Примере 2, теперь существуют вне хромосом в штамме N16961 V.cholerae (см. фиг. 1). С очень низкой частотой (видимо, от 10-6 до 10-8) гомологические боковые последовательности образуют пары оснований и скрещиваются в хромосоме (см. Фиг. 7). Это редкое событие приводит к замене удаленной области токсина на плазмиде генов ctx на хромосоме. Чтобы выделить это редкое событие, использовали явление несовместимости плазмид (Ruvkin, G.B., см. выше). Плазмиды можно разделить на группы несовместимости, обозначаемые от A до W, на основе их способности стабильно поддерживаться вместе с одной и той же клетке. Если две плазмиды не могут поддерживаться вместе с одной клетке, то они несовместимы и принадлежат одной и той же группе несовместимости, предположительно ввиду того, что они используют один и тот же механизм репликации в клетке. При помощи селективного использования стойкости к антибиотикам, содержащихся на одной плазмиде, но не на другой, имеется возможность выбрать, какая из двух несовместимых плазмид будет поддерживаться. Плазмида pJBK55 ввиду ее pRK290-происхождения принадлежит (Inc) группе P. Плазмида R702 также принадлежит Inc группе P и кодирует стойкость к канамицину, тетрациклину, сульфонамиду и стрептомицину, но не к ампициллину. При помощи скрещивания pR702 (SuR) в N16961 (pJBK55) (App) и селекции на среде, содержащей как ампициллин, так и сульфонамид, осуществляли селекцию для клеток, в которых стойкость к ампициллину была включена в хромосому, а стойкость к сульфонамиду оставалась на плазмиде R702, так как pR702 и pJBK55 несовместимы (см. фиг. 2). Полученный в результате штамм JBK56 (фиг. 3) был стойким к ампициллину и отрицательным на токсин, когда его испытывали в клетках надпочечников Y-1 и при помощи Gm1-ЭЛИЗЫ. Кроме того, когда хромосомную ДНК подвергали гибридизации с ДНК-зондами, содержащими гены токсина холеры (CT) клона, то JBK56 был отрицательным, что наводит на мысль, что гены токсина были полностью удалены.
Антибиотическую стойкость, закодированную на R702, исключали при помощи селекции спонтанно исправленного производного, не имеющего этой плазмиды (это событие появляется с частотой примерно 1 из 2 000).
Пример 4
Исключение селектируемого маркера из Примера 1
Чтобы исключить стойкость к ампициллину, конструировали производное pJBK55, в котором гены, кодирующие стойкость к ртути (Hg) из R100, клонированы в сайт Pst гена Ap, при этом в результате вставки дезактивируется стойкость к ампициллину. Это производное затем скрещивали в V.cholerae JBK56, затем при помощи pR702, а селекцию проводили как выше, для HgR, ApS V. cholerae. Финальный штамм, V.cholerae JBK70, был чувствительным ко всем испытываемым антибиотикам, стойким к ртути и, как фенотип, отрицательный относительно токсина. Его хромосомная ДНК не гиоридизировалась заметно с ДНК-зондами, содержащими CT-гены. Грубый анализ последовательности ДНК полной хромосомы показал, что JBK70 остался неизменным по сравнению с родительским штаммом N16961 за тем исключением, что удалены гены токсинов и вставлены стойкость к ртути и неактивные гены стойкости к ампициллину. Такие штаммы не могут восстанавливать токсигенность, так как гены токсинов не только подвергнуты мутации, но полностью удалены.
Пример 5
Перенос сшитого гена, чтобы обеспечить антитоксичный иммунитет
Если для синергии необходимо получить как антибактериальный, так и антитоксичный иммунитет, можно получить производное JBK70, чтобы продуцировать только субблок B токсина холеры. Чтобы добиться этого результата, получали производное токсина, которое продуцирует только B и в котором отсутствуют гены для A (фиг. 8). Hpa-II-фрагмент из pJBK16, содержащий структурный ген B, клонировали в клонирующий вектор фага, M13mp7, заменяющий сайты BamHI и Eco RI по обе стороны гена (фиг. 8). Фрагмент, окруженный теперь сайтами BamHI, клонировали в pMS9, который содержит очень сильный промотор trp. Положение генов B под контролем транскрипции сильного промотора гарантирует высокую производительность антигена B. Из проанализированных клонов приблизительно 50% не продуцировали антигена. Этот факт отражает две возможные ориентации для клонированной вставки - один раз вперед, другой раз назад. Одно производное, pJBK51, которое продуцировало субблок B, скрещивали в Vibrio cholerae JBK70, при этом устанавливали, что он продуцирует даже больше антигена B, чем родительский штамм N16961, дающий JBK70 (PJBK51). Другие мутанты "только B" конструировали с использованием других промоторов, включающих промотор PL, и они могут быть охарактеризованы в соответствующих моделях на любые существенные различия в ин виво экспрессии.
Пример 6
Колонизация кишечника молодых мышей штаммом JBK70 без возврата к токсигенности
Молодых мышей (2,0-3,5 г) отнимали от матерей и держали голодом в течение от 3 до 8 часов. Четырем из них затем прививали через рот в 1 день прививку, используя иглу для кормления животных. Прививка состояла из примерно 108 CFU/образующих колонии единиц/ /мышь JBK70 в объеме от 0,05 мл до 0,1 мл. Прививку получали в бульоне BHI, описанной Басельски, B, и др., см. выше. Прививка содержала примерно 0,01% голубого красителя Эванса. Присутствие этого красителя в желудке, заметное через стенку брюшной полости, указывало на правильную доставку прививки. Добавление голубого красителя Эванса прекращали на 2 день (см. Таблицу), чтобы избежать ингибирования JBK70.
Последующие прививки включали от мыши - к мыши (МХМ), или, в качестве альтернативы, от мыши - к пластинке - к мыши (МХРХМ), но необходимы были различные процедуры, чтобы получить прививку, сравнимую с описанием Басельски для осуществления прививки на 1-й день.
Чтобы получить прививку МХМ, кишки отделяли от желудка до анального отверстия при стерильных условиях. Кишки взвешивали, помещали в стеклянную пробирку для гомогенизации и добавляли примерно 0,5 мл бульона BHI. Cмесь подвергали гомогенизации тефлоновым пестиком до тех пор, пока ткань не становится жидкой. Полученную в результате суспензию использовали для того, чтобы привить примерно 10-8 CFU каждой молодой мыши. Ее проверяли на чистоту при помощи посева штрихом на пластинки МЕА (агар мясного экстракта). Голубой краситель Эванса не добавляли.
Чтобы получить прививку МХРХМ, стерильную петлю использовали для того, чтобы перенести клетки с MEA-пластинки в бульон BHI. В примерно 1 мл BHI добавляли примерно 1011 CFU/мл так, чтобы получить плотную суспензию. Смесь быстро перемешивали до гомогенности и 0,05-0,1 м (примерно 1010 CFU) прививали через рот каждой молодой мыши. Голубой краситель не добавляли.
При всех прививках мышей держали в химических стаканах при комнатной температуре (73-76oФ). Химические стаканы помещали в пластиковый ящик, который покрывали тонким покрывалом, чтобы поддерживать мышей при окружающей температуре примерно 78oФ.
Как показывают результаты в Таблице (см. в конце описания), имелось достаточно клеток в кишечнике, чтобы привить следующему животному, что можно проверить при помощи посева штрихом на MEA-пластинки. Таким образом, Vibrio cholerae JBK70 колонизировал кишечник молодой мыши. Кроме того, не увеличивались концентрации накапливающейся воды, так как не было отмечено существенных увеличений FA-отношения (FA-отношение, большее или равное 0,065 считается положительным накоплением жидкости). Какое-либо возвращение к токсигенности давало бы противоположные результаты.
Пример 7
Конструкция штамма V.cholerae CVD001, имеющего удаленный фрагмент рестрикции внутри гена, кодирующего субблок A
Другим классическим штаммом, выбранным для ослабления, был штамм Vibrio cholerae Огава 395 (в качестве альтернативы обозначаемый через "395"), который подобно N16961 был изучен широко на добровольцах и который сообщал хороший иммунитет [Levine, M.M. "Immunity to cholera as evaluated in Volunteers", в книге : Cholera and Related Diarrheas 43rd Nobel Suposium, Strockholm, 1978 г. (ред. O. Ouchterlong и J.Holmgren, Basel: S.Karger, стр. 195-2-3 (1980); Levine M.M. и др. Acute Enteric., см. выше (1981)]. Процедура, которую использовали для ослабления 395, по существу не отличалась от той, что использовали для N16961 (которая описана в Примерах 1-5).
Первая стадия включала клонирование и построение карты двух копий гена токсина штамма 395. В результате анализа Южных пятен обнаруживали два Hind III-фрагмента в примерно 16 и примерно 12 ко длиной, причем оба они гибридизировались с клонированными генами токсина холеры. Эти фрагменты очищали электрофорезом на агаровых гелях и клонировали в обработанную фосфатами щелочных материалов, переваренную ферментом Hind III pBR325 (фиг. 9). Полученные в результате рекомбинантные плазмиды, содержащие гены токсинов, обозначали через pCVD14 и pCVD15.
Затем для плазмид pCVD14 и pCVD15 составляли карты с использованием эндонуклеаз рестрикции. Был обнаружен фрагмент Xba I-Cla I в примерно 550 по, содержащий полную последовательность оснований A1-субблока за исключением кодонов первых 10 аминокислотных остатков A1. Этот Xba I-Cla I-фрагмент удаляли ин витро как в pCVD14, так и в pCVD15 за несколько шагов, которые показаны на фиг. 10 для pCY 15. Во-первых, в результате частичного переваривания ферментом Cla I получали популяцию линейных молекул, в которых только один из пяти сайтов Cla I отсечен. Далее концы линейных молекул делали тупыми при помощи заполнения ДНК-полимеризой. Xba I-линкеры подвергали лигации с тупыми концами сайтов Cla I, получая в результате набор молекул, в который затем добавляли фермент Xba I, чтобы срезать линкер, а затем добавляли и подвергали лигации ген стойкости к тетрациклину на Xba I-фрагменте. После трансформации в E.coli K-12 и селекции на тетрацилине анализировали содержание плазмиды в нескольких трансформантах. Обнаруживали несколько мутантов удаления, в которых были удалены один и более фрагментов Xba I-Cla I. Выбирали один мутант удаления, в котором отсутствовал только один фрагмент Xba I-Cla I в 550 по, содержащий ген A1. Этот мутант удаления, обозначаемый pCVD25, подвергали очистке, переваривали Xba I и удаляли ген стойкости к тетрациклину. Полученный в результате клон, PCVD30, был отрицательным на голотоксин, что устанавливали при помощи анализа клеток надпочечников Y-I / Sack, D.A. и др., см. выше (1980)/, но положительным на продуцирование субблока В, что устанавливали при помощи ЭЛИЗА /Sack, D.A. и др. , см. выше (1980)/, у него отсутствовали гены для A1, что показывали при помощи гибридизации с использованием меченого A1-зонда. Hind III-фрагмент из pCVD30, содержащий мутацию удаления токсина, затем клонировали в pJBK85, TC-чувствительное, CM-стойкое производное плазмиды pJBK108. Полученную в результате плазмиду обозначали pJBK108.
Отсутствие селектируемого маркера в мутации удаления токсина в pJBK108 требует модификации процедуры, ранее использованной для ослабления Эль Тор N16961. Чтобы осуществить удаление A1-генов из 395, фрагмент Hind III из pCVD15 клонировали в pJBK85, в результате чего получали pJBK88 (фиг. 11). Ген стойкости к тетрациклину на Xba I-фрагменте затем клонировали в сайт Xba внутри A1-гена и PJBK88, в результате получая pJBK107. Эту стойкость к тетрациклину затем рекомбинировали в хромосому из 395, как это ранее делали для V. cholerae pJBK56. pJBK107 (Tcr. Cmr) переносили в 395 и вводили вторую Inc P плазмиду, pR751 (Tpr). В результате селекции Tcr, Tpr, Cmr-колоний получали V.cholerae JBK113, которая содержала гены стойкости к тетрациклину в обоих копиях хромосомного гена токсина. pJBK108, содержащую мутацию удаления, затем переносили в V.cholerae JBK113. Гомологическая рекомбинация мутации удаления в хромосоме будет приводить к утрате последовательностей A1-гена, событие, которое может быть обнаружено при испытании на стойкость к тетрациклину. Ввиду того, что такая рекомбинация имеет место с очень низкой частотой, используют процедуру обогащения клеток, чувствительных к тетрациклину среди клеток, стойких к тетрациклину. Такая процедура обогащения использует тот факт, что тетрациклин является бактериостатическим антибиотиком, в то время как ампициллин и Д-циклосерин являются бактерицидными. Таким образом, культуру V.cholerae JBK113, содержащую pJBK108, выращивали 3 часа при температуре 37oC в L-бульоне, содержащем 2 микрограмма/мл тетрациклина, 50 микрограмм/мл ампициллина и 50 микрограмм/мл Д-циклосерина. В конце 3-го часа большая часть клеток, стойких к тетрациклину, гибла, а чувствительные к тетрациклину клетки обнаруживали при помощи пластинок с L-агаром и такой же пластиной с L-агаром и тетрациклином. Колонии, чувствительные к тетрациклину, зондировали на присутствие A1-генов при помощи ДНК-гибридизации. Один чувствительный к тетрациклину штамм, содержащий удаления в обеих копиях гена A1-субблока, обозначали через V.cholerae CVD101 и испытывали на продуцирование B-субблока при помощи ЭЛИЗА (Сак, см. выше), устанавливали, что V.cholerae CVD101 продуцирует антиген субблока в концентрациях, по существу эквивалентных токсигенному родительскому V.cholerae 395.
Пример 8
Анализ последовательности ДНК генов токсинов
Определяли полную ДНК-последовательность генов токсинов V.cholerae Инаба 62746, часть которой ранее была описана Lockman и др., J. Biol. Chem., т. 258, стр. 13722 (1983). Карта эндонуклеаз рестрикции pCVD14 и pCVD15 указывает на то, что последовательности, содержащиеся в штамме 62746, присутствуют также в генах токсинов 395. Предполагаемое соединение после удаления фрагмента Xba I-Cla I в 550 по, но с добавлением последовательности Xba I-линкера, представлено на фиг. 12. Сайт Xba I последовательности токсина холеры включает аминокислотные остатки 10 и 11 A1-структурного гена (не считая 18 аминокислотной последовательности лидера для A1). Сайт Cla этой последовательности расположен в последнем остатке из A1 в первом остатке A2.
Пример 9
Конструирование штамма V.cholerae с удаленным токсином блокатора малой зоны в CVD101
Мутант удаления zot из V.cholerae получали при помощи той же процедуры, что и мутант удаления токсина холеры CVD101, описанной в Примере 7. Ген zot содержится в рекомбинантной плазмиде pBB68. pBB68 состоит из хромосомного ДНК-фрагмента ECoRI-Рbr I из V.cholerae S69B, который содержит ген zot и гены ctx, которые имеют удаленный Xba I-Cla I-фрагмент 550 по. Фрагмент рестрикции stu I-Acc 1 в 575 пар оснований удаляли ин витро из pBB68 при помощи переваривания ферментами рестрикции stu I и Acc I, и превращали концы молекул в тупые при помощи заполнения ДНК-полимеразой. (При этом удаляют 48% из гена zot в 1199 пар оснований.) Половину этой пробы подвергали лигации на себе, получая мутант удаления. Другую половину этой пробы подвергали лигации с геном стойкости к тетрациклину (чудеродного происхождения), давая таким образом селективный маркер.
Мутант удаления zot, построенный выше ин витро, вводили в хромосому V. cholerae CVD 101, как это было ранее описано для конструкции мутанта удаления ctx CVD101. Клон, стойкий к тетрациклину, полученный выше, клонировали в Inc P плазмиду PJBK85. Эту плазмиду (Tcr, Cmr) переносили в CVD101, осуществляя селекцию по Tcr. Вводили вторую Inc P плазмиду, pR751 (Tpr). Селекция Tcr, Tpr, Cms колоний дает в результате штаммы V.cholerae, в которых Tcr-ген был рекомбинирован в гене zot.
Плазмиду, содержащую мутант удаления Stu I-Acc 1 без Tcr-гена, затем переносили в Tcr-штамм V.cholerae. Гомологическая рекомбинация мутанта удаления в хромосоме приводит к утрате последовательностей гена zot, событие, которое может быть обнаружено на основании потери Tcr. Tcs-колонии подвергали селекции и просеивали на утрату последовательностей zot при помощи ДНК-гибридизации с использованием фрагмента stu I-Acc 1 в качестве зонда.
Пример 10
Конструкция штамма V.cholerae CVD109 с удалениями фрагментов рестрикции последовательностей, кодирующих токсин V.cholerae и токсин блокатора малой зоны
Конструкция ослабленного штамма CVD109 V.cholerae включает введение клонированных последовательностей Vibrio вместе с последовательностями, кодирующими селектируемый маркер, в хромосому вирулентного штамма V.cholerae. Первоначальная ин виво рекомбинация гомологических последовательностей из рекомбинантной плазмиды в хромосоме обеспечивает селектируемый маркер в этом сайте. Второе событие ин виво рекомбинации между гомологическими боковыми последовательностями приводит к удалению действующих генов из хромосомы, конечным продуктом которого является мутация удаления.
Фиг. 17 иллюстрирует конструкцию CVD109. Гены zot и ctx примыкают друг к другу на хромосоме V.cholerae. Несколько копий ДНК-последовательности в 2 700 пар оснований, именуемой RSI (для повторяющейся последовательности) находятся по обе стороны генов zot и ctx в вирулентом штамме E7946. V.cholerae (биотип Эль Тор, Серотип Огава). На фиг. 17A последовательности zot и ctx показаны большой незаштрихованной или заштрихованной стрелками. RSI-последовательности показаны меньшей толстой стрелкой.
Рекомбинантная плазмида, pCVD51 (фиг. 17A) содержит клонирование zot/cts-последовательности (незаштрихованная стрелка), которые гомологичны хромосомным zot/ctx последовательностям (показаны заштрихованной стрелкой на фиг. 17A), и содержит селектируемый маркер, стойкость к ампициллину (Apr). Вектор плазмиды, в который клонировали последовательности Vibrio, суть PG P704 [Miller и Mekalanos, J.Bacteril., т. 170; стр. 2575-2583 (1988)]. Эта плазмида не может подвергаться репликации вне хромосом в V.cholerae, но может подвергаться репликации в разрешающих это штаммах E.coli pCVD51 скрещивали из E.coli в E7946 V.cholerae. Так как эта плазмида не может реплицировать вне хромосомы в V.cholerae, в результате селекции Apr-колонии получали штаммы, в которых целая плазмида с Apr-маркером подверглась гомологической рекомбинации в хромосоме в сайте последовательностей zot/ctx (где расположен этот сайт: в гене zot или гене ctx, неизвестно). Результат этого единственного события перехода (не двойного перехода) называется "коинтегрированной" структурой и она представлена на фиг. 17B.
RSI-последовательности, окружающие область zot/ctx, должны иметь остаточную длину, чтобы получить обнаруживаемую ин виво рекомбинацию; внутримолекулярная рекомбинация между гомологическими RSI-элементами приводит к утрате всех последовательностей между ними. Apr V.cholerae с интегрированной плазмидой выращивали без ампициллина и выбирали Ap-чувствительные (Apr) колонии. Рекомбинация RSI-элементов, окружающих zot и ctx, приводит к утрате последовательностей-посредников zot и ctx вместе с вектором плазмиды, содержащим Apr (фиг. 17C).
Ars V. cholerae колонии, полученные в результате описанных выше стадий, просеивали при помощи ДНК-гибризации на zot-последовательности. ДНК-зонд состоял из фрагмента рестрикции Stu I-Acc 1 в 575 пар оснований, полученного из клонированного гена zot. Колонии, которые не гибридизировались с этим зондом, подвергали селекции и зондировали на присутствие генов ctx при помощи ДНК-гибридизации, используя зонд гена ctx. Эти результаты гибридизации подтверждали утрату обоих генов zot и ctx. Один представительный штамм собирали и обозначали через CVD109. На фиг. 17D изображена хромосома из CVD109, из которой удалены zot и ctx последовательности, но сохранена одна копия RSI-элемента. (Плазмида изображена на фиг. 17D и она не сохраняется в финальном Aps-штамме, а приведена только с целью иллюстрации удаления второго события перехода. Этот переходной продукт спонтанно утрачивается, так как плазмида не может реплицироваться вне хромосомы в V.cholerae.)
Пример 11
Конструкция штамма CVD110 V.cholerae с удаленными фрагментами рестрикции последовательностей, кодирующих субблоки A и B токсина V.cholerae и токсина блокатора малой зоны, имеющего вставленные ген стойкости к ртути и ДНК, кодирующую субблок B токсина V.cholerae.
CVD110 конструировали непосредственно из CVD109 V.cholerae, описание которого уже было приведено. У CVD109 V.cholerae отсутствуют оба субблока A и B токсина холеры (CT), а также ген, кодирующий zot, и он является чувствительным к ртути. Фрагмент гена, который содержит ген субблока B CT (ctxB) и ген стойкости к ртути, конструировали ин витро. Затем эту конструкцию вставляли в хромосому из CVD109, обязательно в ген гемолизина. Таким образом, финальный штамм вакцины, CVD110, продуцирует B, но не продуцирует A-субблок токсина холеры, и является стойким к ртути и не продуцирует протеин HlyA дикого типа (гемолизин).
Конструкция CT B: Промоторные последовательности ctxB и ctx получали из плазмиды pCVD30, которая описана в Примере 7. Эта плазмида pCVD30 содержит удаление гена ctx A. Фрагмент в 1,4 килооснования, содержащий ген ctxB и ctx-промотор, но не ген zot, получали при помощи переваривания pCVD30 ферментами Hin PI и Hae III. Этот фрагмент обрабатывали ДНК-полимеразой T4, чтобы сделать концы этого фрагмента тупыми, а синтетические Kpn 1-линкеры подвергали лигации с этим фрагментом. Далее этот фрагмент клонировали в вектор pCVD315/Galen и др. , Advances in Research on Cholera and Related Diarrheas, том 7/Sack и др. ред./стр. 143-153 (1990)/. Вектор pCVD315 не имеет особого значения для этой цели, кроме как присутствие сайта Kpn 1. Полученную в результате плазмиду, содержащую ген ctxB, именовали pCVD621 (фиг. 19).
Гены стойкости к ртути: источником генов стойкости к ртути (mer) был тот же, что использовался для V.cholerae JBK70. Фрагмент Nc01-StuI в 4,2 ко, содержащий mer, первоначально получали из pDB 7 [Barrineau и др., J. Molecular and Appl. Genetic, (1984), том 2, стр. 601-619].
Этот фрагмент обрабатывали ДНК-полимеразой (фрагмент кленоу), чтобы получить тупые концы на этом фрагменте, а синтетические линкеры крп 1 подвергали лигации с этим фрагментом. Затем фрагмент клонировали в плазмиду pCVD43.2 (не опубликовано), которая является производной от pCVD43 [Kaper и др. , Advances in Research on Cholera and Releted Diarrheas, т.6 /Ohtomo и др. ред. /, стр. 161-167 (1688)]. pCVD43 содержит клонированные гены гемолизина (hly A) из V.cholerae без Hpa I-фрагмента в 400 по, внутреннего относительно ply A. Удаление фрагмента Hpa в 400 по делает этот ген неактивным [Kaper и др., Advances ..., стр. 6], pCVD43.2 идентичен pCVD43 за тем исключением, что синтетический крп 1-линкел был подвергнут лигации в единственном сайте Hpa I из pCVD43. Комбинированный клон mer-генов, вставленных в ген hyl A, именовали pCVD43.3.
Вставка генов ctxB и mer в CVD109: чтобы ввести эти гены в хромосому из CVD109, использовали вектор плазмиды pGP704, который описан в Примере 10. Фрагмент Cla I-Bgl II в 8,1 ко из pCVD43.3, содержащий mer и hyl A, клонировали в PG P704, чтобы получить pJMK12 (фиг. 19). pJMK12 частично переварили при помощи Kpn 1, чтобы получить популяцию линейных молекул, в которых отсечен только один из 3 сайтов Kpn 1. Фрагмент в 1,4 ко из pCVD621 (описанного выше), содержащий ген ctxB, далее повергали лигации с pJMK12, чтобы получить pCVD622.2B. Относительная позиция и ориентация вставленных генов показаны на фиг. 19.
Далее pCVD622.2B вводили в CVD109 V.cholerae при помощи конъюгации из штамма хозяина E.coli. Как описано в Примере 10, pG P704 не может реплицировать вне хромосомы в V.cholerae, но может реплицировать в позволяющих это штаммах E.coli. Так как pCVD622.2B не может реплицировать вне хромосомы в V. cholerae, в результате селекции Apr-колоний [pG P704 содержит ген, кодирующий стойкость к ампициллину] получали штаммы, в которых целая плазмида pCVD622.2B с маркером Apr гомологически подвергается рекомбинации в хромосоме в сайте гена hyl A. [Она не могла быть подвергнута рекомбинации в последовательностях ctx или zot, так как у CVD109 отсутствуют эти гены]. Результат этого единственного перехода (не двойного перехода) именуется "коинтегрированной" структурой или "меродиплоидом" (эти термины используются как эквивалентные) и представлен на фиг. 20B.
Второе событие перехода (скрещивания) может иметь место между гомологическими последовательностями hylA, окружающими интегрированную pCVD622.2B. Это второе скрещивание имеет место спонтанно и обнаруживается селекцией колоний, которые утратили Apr-фенотип. Это второе событие перехода может иметь один из двух возможных выходов в зависимости от точного сайта рекомбинации. Оба возможных выхода приводят к потере последовательностей вектора плазмиды pG P704 и они представлены на фиг. 20C и 20D. Один выход просто восстанавливает первоначальную ситуацию, т.е. штамм, идентичный CVD109, у которого отсутствуют ctx, zot и mer. Второй выход приводит к утрате последовательности pG P704, но последовательности mer и ctx, содержащиеся внутри последовательностей hly A, сохраняются. Эти два возможных выхода легко различаются при помощи ДНК-гибридизации с использованием меченых ctx-последовательностей в качестве зонда. Чтобы изолировать целевой "выход", выращивали культуру CVD109, содержащую интегрированную pCVD622.2B в L-бульоне без добавления антибиотиков. Эту культуру помещали на неселективные пластинки с L-агаром и полученные в результате колонии подвергали репликации на L-агаровых пластинках, содержащих Ap. Далее Aps-колонии подвергали гибридизации с ctx-зондом, а колонии, имеющие ctx-последовательности, изолировали. Одну такую колонию обозначали через V.cholerae CVD110. На этом штамме подтверждали при помощи ДНК-гибридизации, что он содержит последовательности ctx и mer, а также, что у них отсутствуют последовательности pG P704 и Hpa I-фрагмент в 400 по, внутренний относительно гена hly A. Кроме того, подтверждали, что CVD110 продуцирует субблок B токсина холеры при помощи ЭЛИЗА [Sack, D.A. и др., см. выше (1980)].
ДНК-последовательность вставленных генов: Точные ДНК-последовательности вставленных генов ctx и mer известны из литературы. Точный сайт гена hly A, в который эти гены вставлены, также известен. На фиг. 21 представлены ожидаемые ДНК-последовательности этих генов, показаны относительные позиции этих генов. Указаны точки начала и конца различных генов; направление стрелок указывает направление транскрипции гена. Только эти последовательности сохраняются в окончательной конструкции, поэтому, например, ctx B приведен, но часть ctx A, которая была удалена в этой конструкции, не показана.
В то время как настоящее изобретение было описано в связи с его конкретными осуществлениями, должно быть ясно, что оно может допускать дальнейшие усовершенствования, и настоящая заявка предназначена для того, чтобы включить любые вариации, использования или усовершенствования настоящего изобретения, следуя в общем случае принципам настоящего изобретения, и включить такие отправные точки от настоящего патентного описания, которые вытекают из известной или стандартной практики в этой области техники и которые могут применять отличительные свойства, указанные выше, и перечисленные в приложенных пунктах формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АВИРУЛЕНТНЫЙ ШТАММ VIBRIO CHOLERAE НЕ-01 СЕРОГРУППЫ (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ШТАММА (ВАРИАНТЫ), ВАКЦИНА | 1994 |
|
RU2140981C1 |
| СПОСОБ ПРИМЕНЕНИЯ ZOT ИЛИ ЗОНУЛИНА ДЛЯ ИНГИБИРОВАНИЯ АНТИГЕН-СПЕЦИФИЧЕСКОЙ ПРОЛИФЕРАЦИИ ЛИМФОЦИТОВ | 1999 |
|
RU2214271C2 |
| ШТАММ БАКТЕРИЙ VIBRIO CHOLERAE - ПРОДУЦЕНТ ХОЛЕРНОГО ТОКСИНА II ТИПА | 2007 |
|
RU2326941C1 |
| АВИРУЛЕНТНЫЙ ШТАММ БАКТЕРИЙ Vibrio cholerae KM 263 БИОВАРА ЭЛЬТОР СЕРОВАРА ИНАБА - ПРОДУЦЕНТ ПРОТЕКТИВНОГО О1 АНТИГЕНА | 2010 |
|
RU2425867C1 |
| АВИРУЛЕНТНЫЙ ШТАММ БАКТЕРИЙ Vibrio cholerae КМ 262 БИОВАРА ЭЛЬТОР СЕРОВАРА ОГАВА - ПРОДУЦЕНТ ПРОТЕКТИВНОГО О1 АНТИГЕНА | 2010 |
|
RU2425868C1 |
| СПОСОБ ДЕТЕКЦИИ И ОПРЕДЕЛЕНИЯ БИОТИПА, СЕРОГРУППЫ И ТОКСИГЕННОСТИ ВОЗБУДИТЕЛЯ ХОЛЕРЫ И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2007 |
|
RU2360972C1 |
| АВИРУЛЕНТНЫЙ ТЕСТ-ШТАММ БАКТЕРИЙ Vibrio cholerae БИОВАРА eltor СЕРОВАРА Ogawa (ctxA-, tcpA-, toxR-, zot-), ИСПОЛЬЗУЕМЫЙ В ИММУНОЛОГИЧЕСКИХ, ГЕНЕТИЧЕСКИХ ИССЛЕДОВАНИЯХ И УЧЕБНОМ ПРОЦЕССЕ | 2004 |
|
RU2254371C1 |
| Рекомбинантный белок для иммунизации против холеры | 2019 |
|
RU2723705C1 |
| СПОСОБ ДИФФЕРЕНЦИАЦИИ ШТАММОВ VIBRIO CHOLERAE О1 БИОВАРА ЭЛЬ ТОР С РАЗЛИЧНОЙ ЭПИДЕМИОЛОГИЧЕСКОЙ ЗНАЧИМОСТЬЮ МЕТОДОМ МУЛЬТИПЛЕКСНОЙ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2018 |
|
RU2671412C1 |
| Способ идентификации и определения патогенности штаммов Vibrio cholerae методом петлевой изотермической амплификации (LAMP) | 2023 |
|
RU2820141C1 |
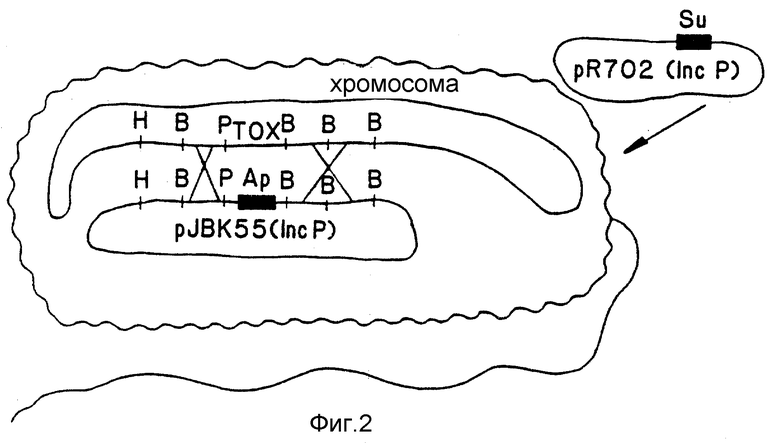
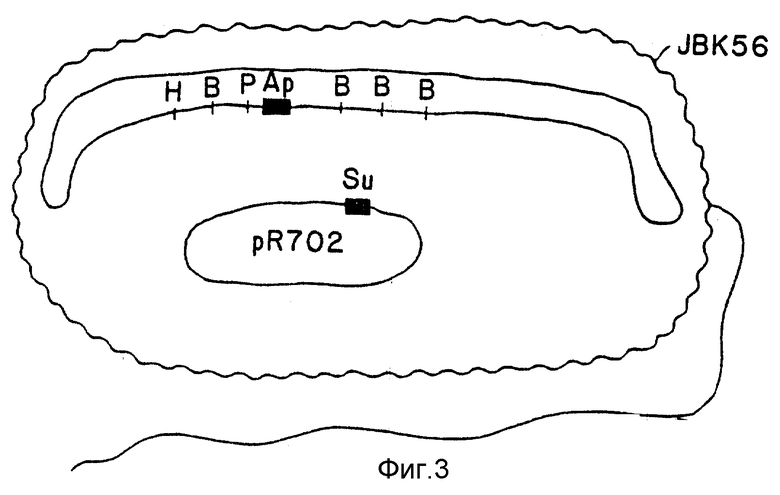
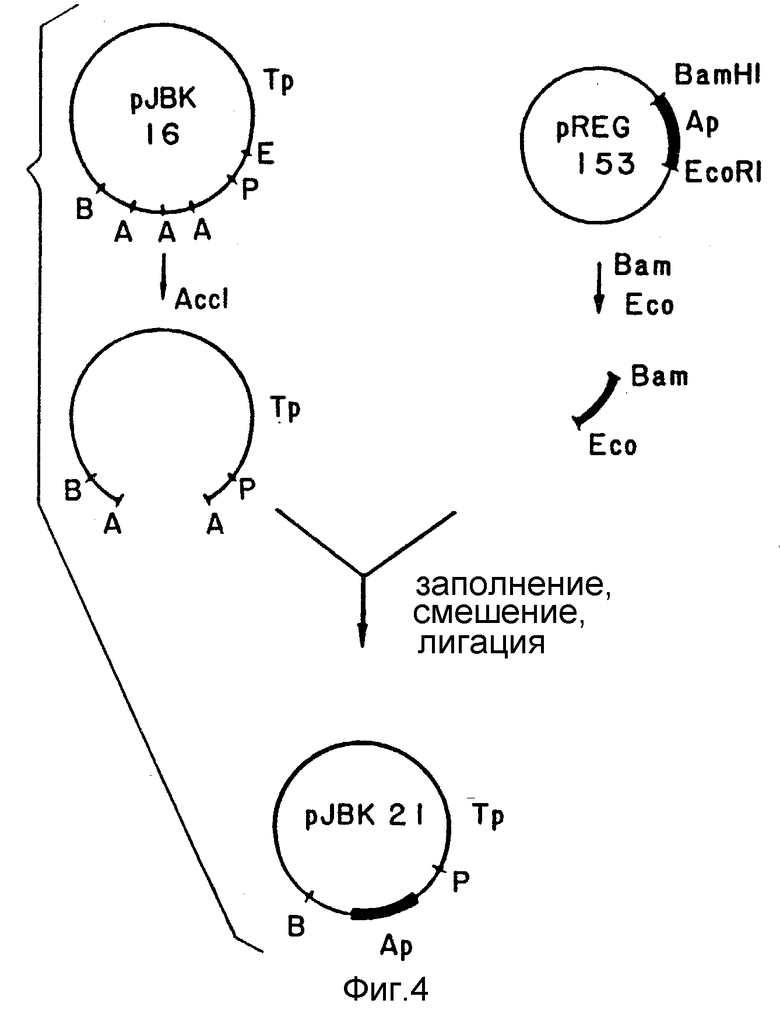
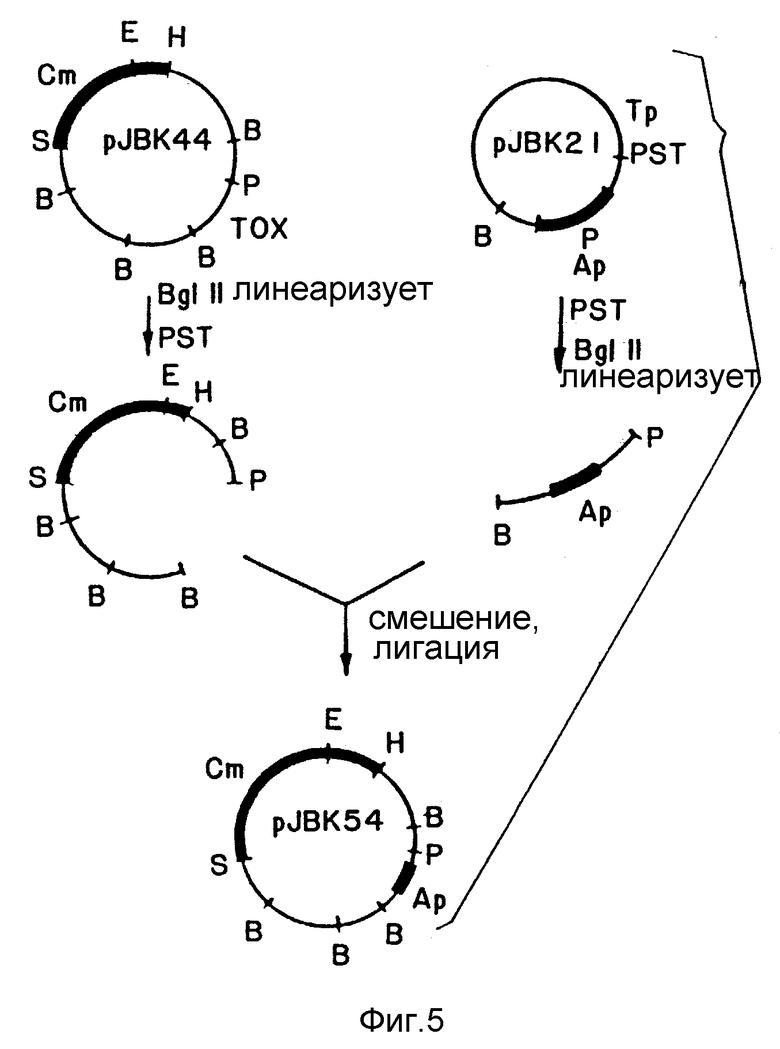
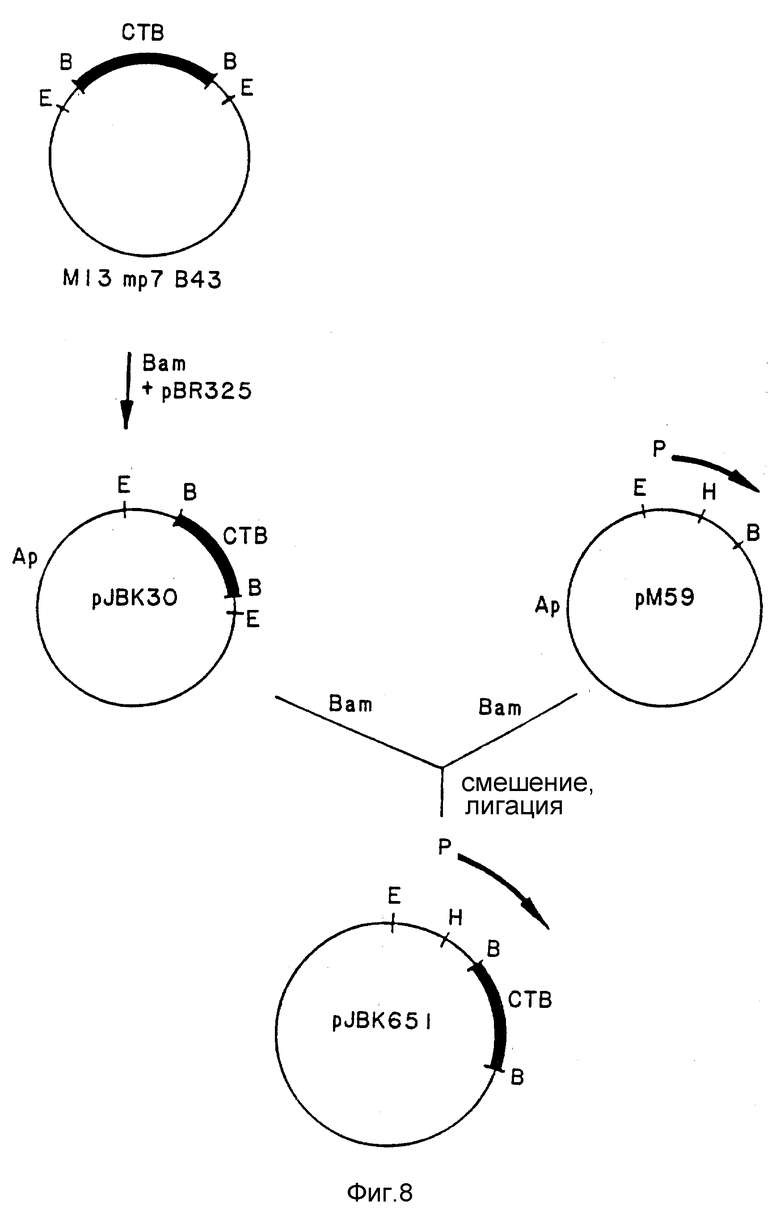
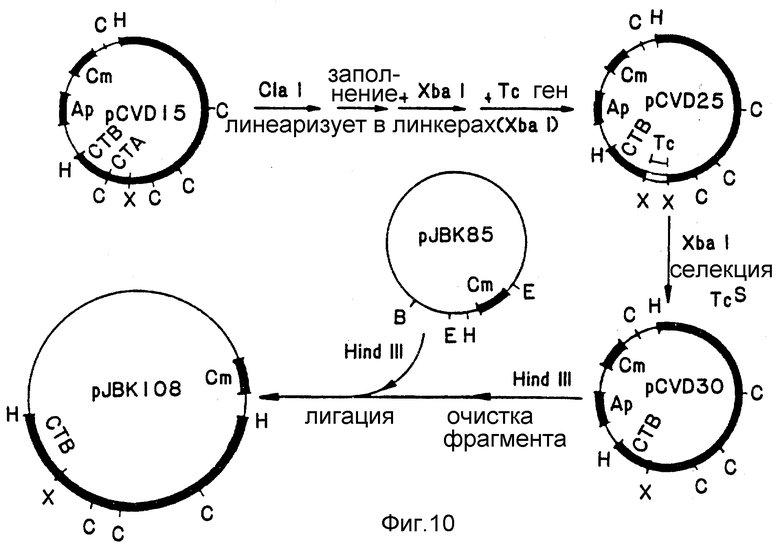
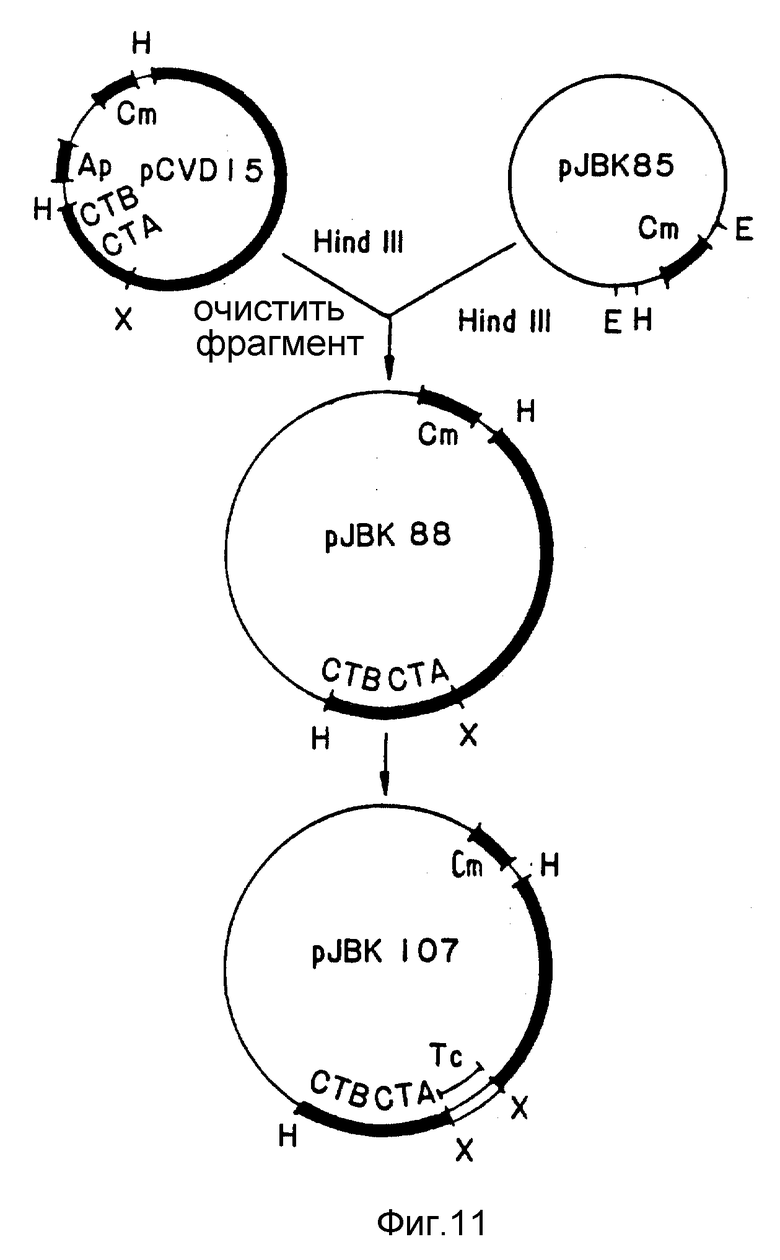
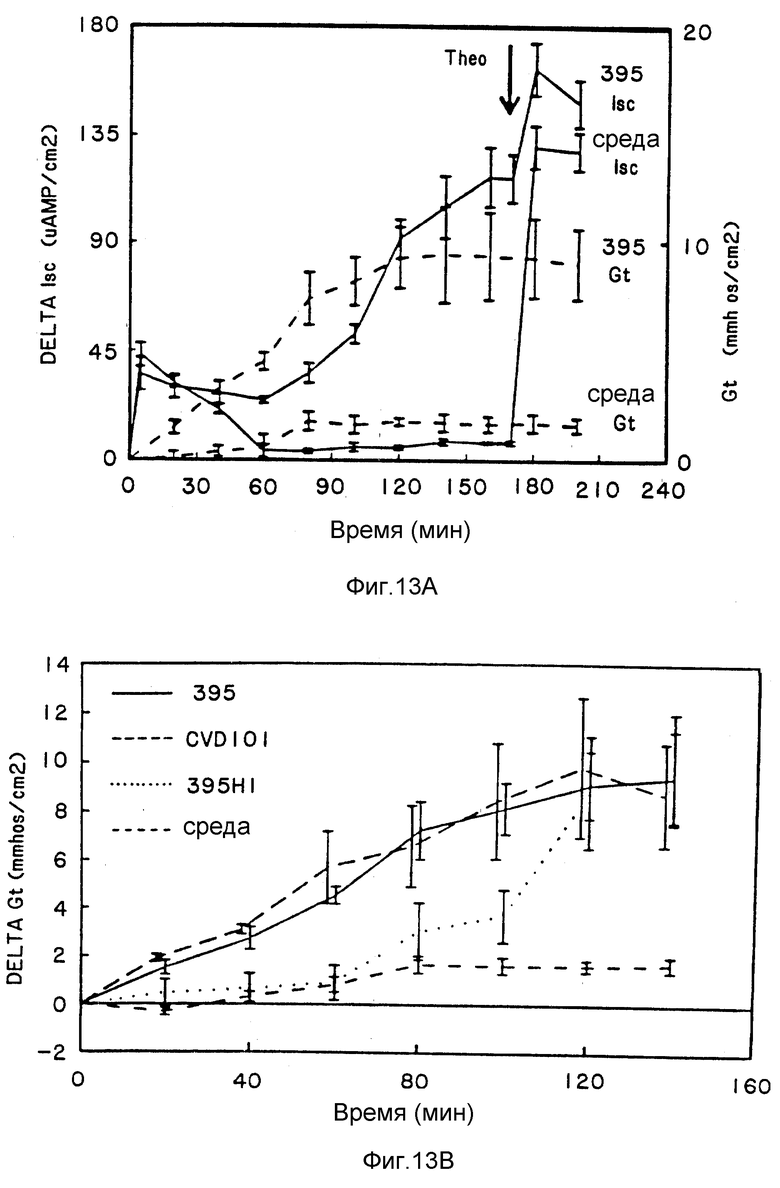
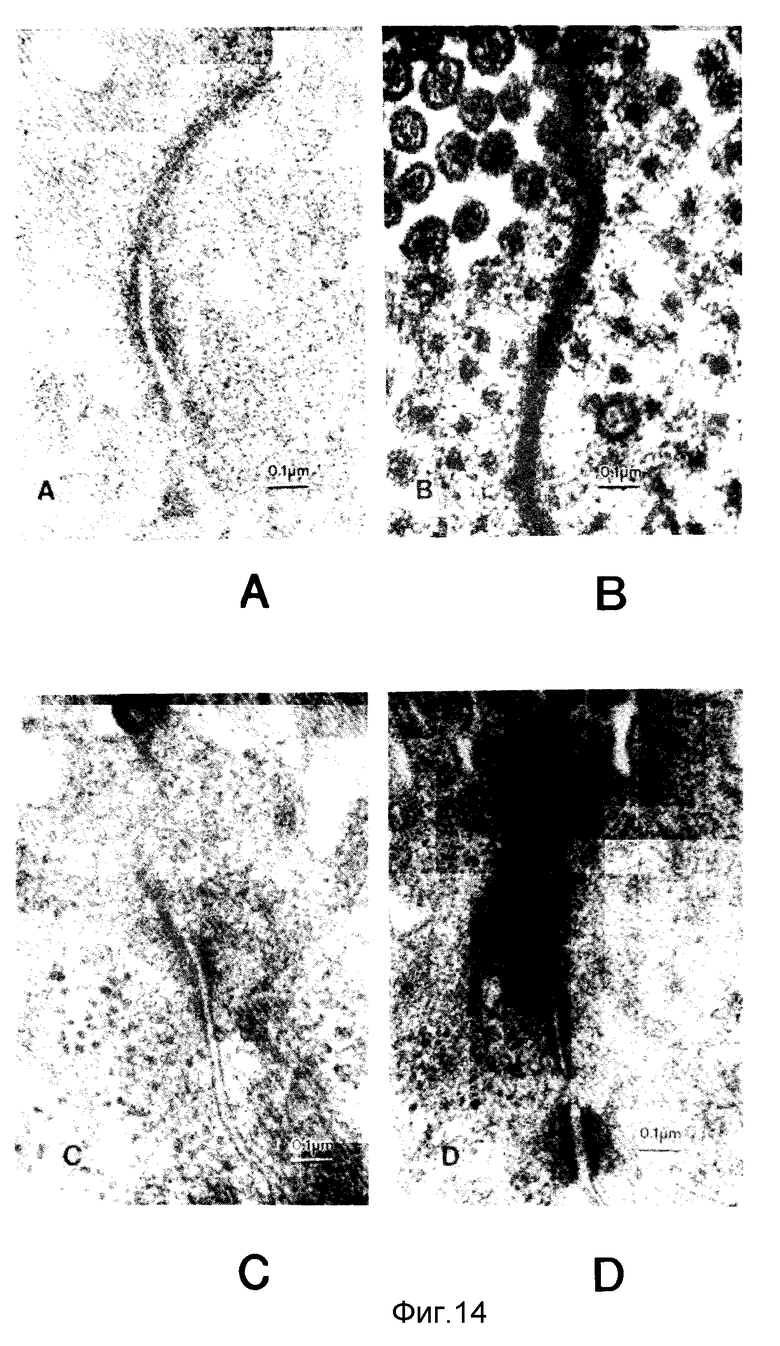
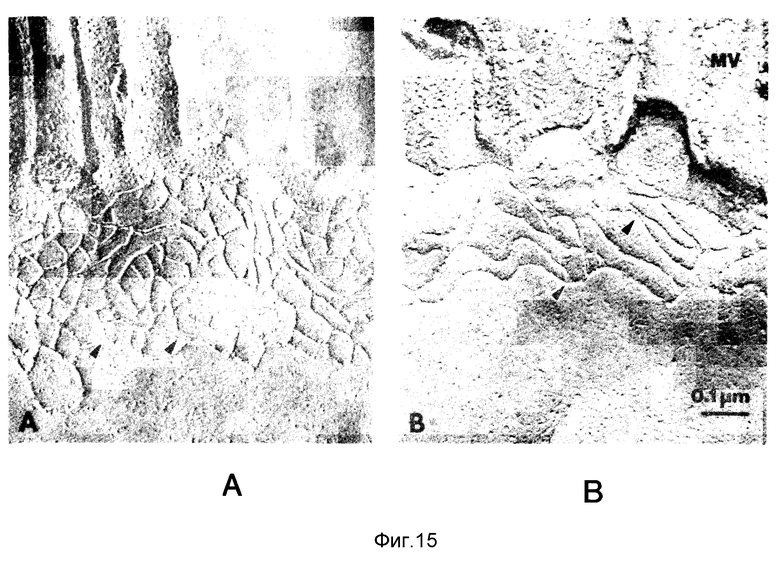
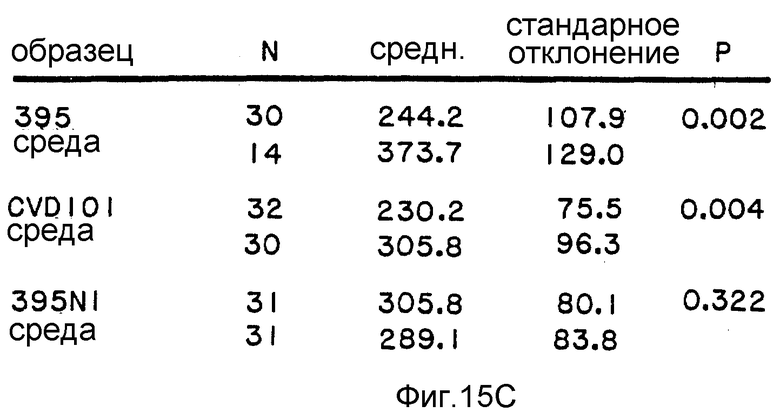
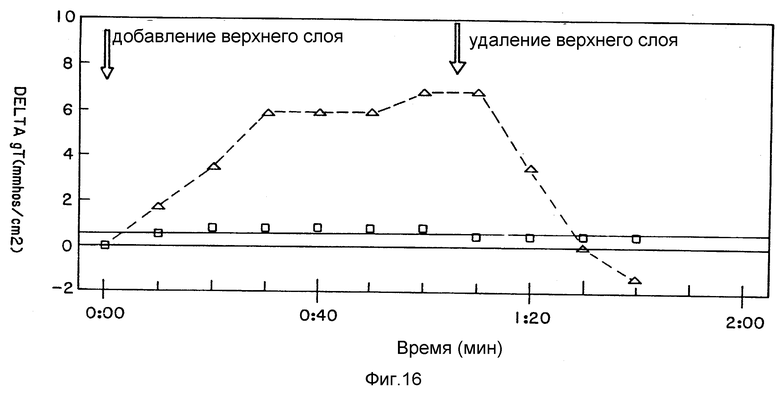
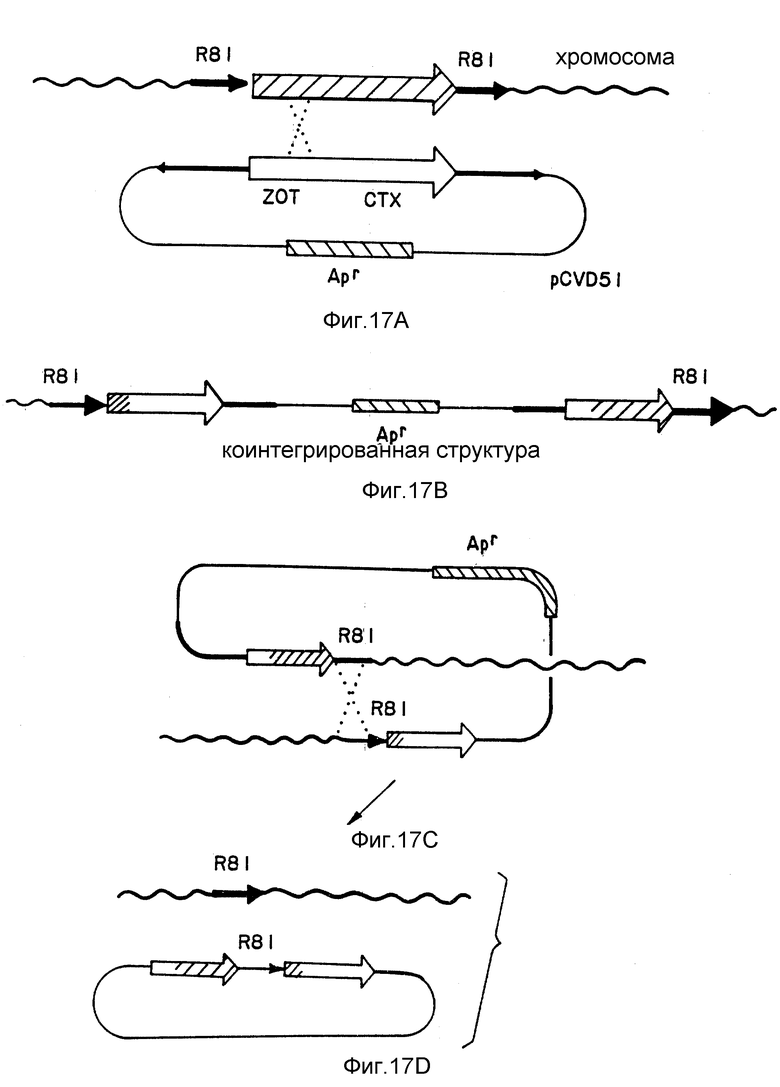

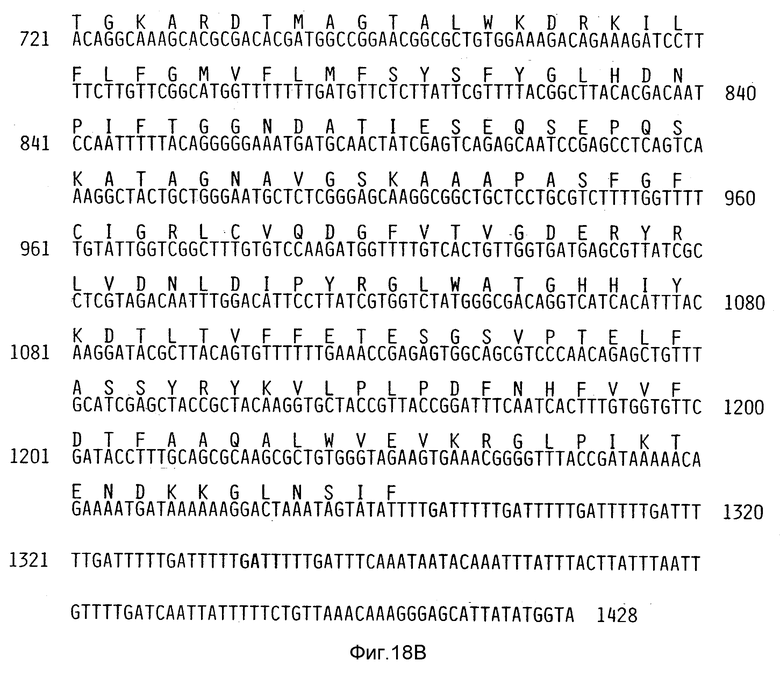
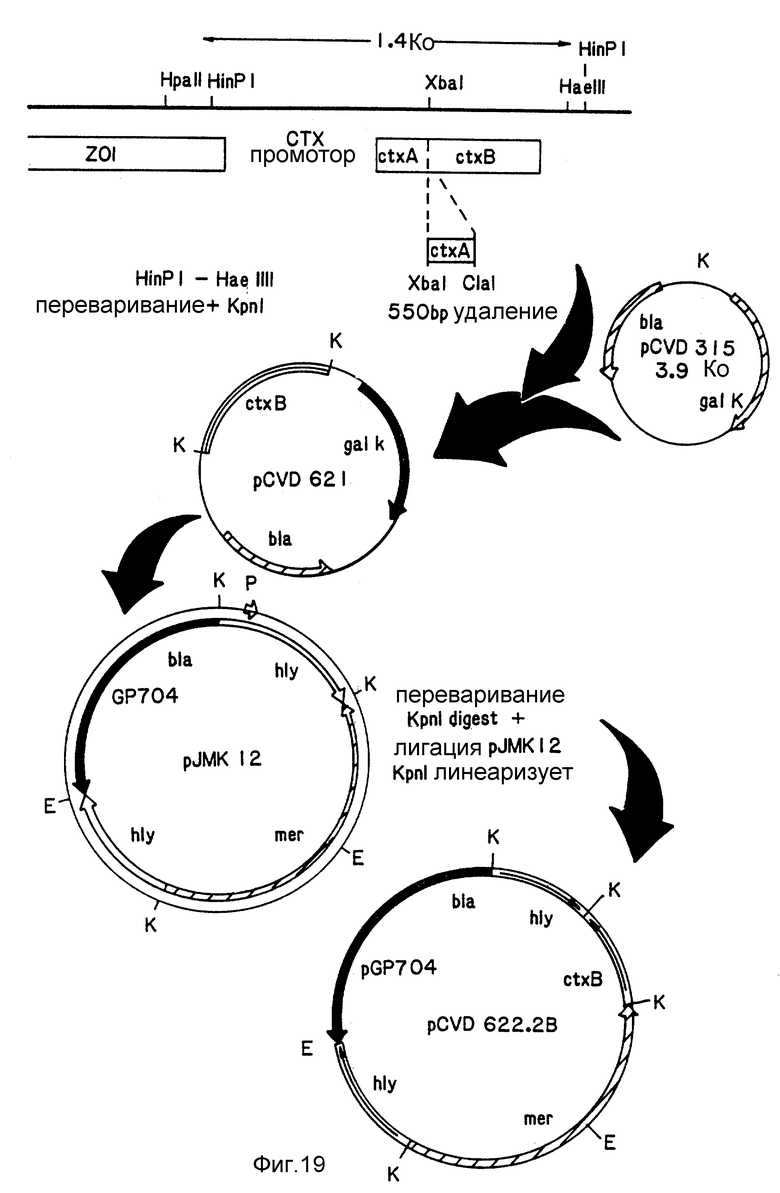







Принцип изобретения состоит в изоляции штамма вакцины Vibrio cholerae, специфическим образом измененного при помощи техники рекомбинатной ДНК, чтобы лишить его вирулентности без нарушения других компонент, необходимых для иммунитета. Такое ослабление осуществляют при помощи переваривания эндонуклеазами рестрикции плазмид, несущих соответствующие последовательности V. cholerae. В результате переноса соединенного гена таких переваренных плазмид и последующих процедур селекции in vivo рекомбинантов с вирулентными хозяевами V. cholerae получали штаммы без генов токсина и его частей. Вакцина, полученная из этого штамма, вызывает стойкий иммунитет против холеры. 2 с. и 5 з.п.ф-лы, 1 табл., 21 ил.
(а) конструирование первой плазмиды, содержащей последовательности ДНК V.cholerae, кодирующие холерный токсин и токсин Zonula occludens, и ген, кодирующий селективный маркер гетерологичного происхождения, причем указанная плазмида не способна реплицироваться вне хромосомы в клетках Vibrio cholerae; введение указанной плазмиды в подходящий микроорганизм;
(б) скрещивание микроорганизма, несущего введенную первую плазмиду, с вирулентным штаммом Vibrio cholerae, содержащим указанные в (а) последовательности, кодирующие холерный токсин и токсин Zonula occludens, причем данные последовательности в вирулентном штамме фланкированы повторяющимися RSI-последовательностями;
(в) отбор штамма Vibrio cholerae, полученного в результате скрещивания на стадии (б) по экспрессии селективного маркера и содержащего плазмиду, указанную в (а), которая встроена путем гомологичной рекомбинации в хромосому данного штамма по сайту указанных в (а) последовательностей;
(г) выращивание штамма, полученного на стадии (в) в отсутствии селективного агента;
(д) отбор штамма V.cholerae, не экспрессирующего селективный маркер, у которого последовательности, кодирующие холерный токсин и токсин Zonula occludens между повторяющимися RSI-последовательностями, утрачены из хромосомы;
(е) конструирование второй плазмиды, содержащей ген резистентности к ртути и фрагмент ДНК, кодирующий β-субъединицу токсина V.cholerae, ген второго селективного маркера гетерологического происхождения, причем указанная плазмида не способна реплицироваться вне хромосомы в клетках V.cholerae, указанные последовательности ДНК, кодирующие резистентность к ртути и β-субъединицу холерного токсина, фланкированы последовательностями гена гемолизина (hly A) V.cholerae, при этом последовательности hly A имеют достаточную длину для обеспечения обнаруживаемой рекомбинации in Vivo и введение указанной плазмиды в подходящий микроорганизм;
(ж) скрещивание микроорганизма, несущего введенную вторую плазмиду со штаммом V.cholerae, отобранным на стадии (g), содержащим последовательности, гомологичные указанным последовательностям hly A, имеющие достаточную длину для обеспечения обнаруживаемой рекомбинации in Vivo;
(и) отбор штамма Vibrio cholerae, экспрессирующего второй селективный маркер;
(к) выращивание штамма, полученного на стадии (и) в отсутствии второго селективного агента;
(л) селекцию штамма Vibrio cholerae, не экспрессирующего второй селективный маркер;
(м) отбор штамма Vibrio cholerae, указанного на стадии (л) по наличию гена резистентности к ртути и фрагменту ДНК, кодирующему β-субъединицу токсина Vibrio cholerae, и по делеции последовательностей хромосомной ДНК, кодирующей холерный токсин и токсин Zonula occludens.
| US 4882278, 21.11.89 | |||
| M.M.Levine et al | |||
| Safety Immunogenicity and Efficacy of Recombinant Live oral Cholera Vaccines | |||
| Клапанный регулятор для паровозов | 1919 |
|
SU103A1 |
| Lancet, issued 27 august, 1988, p.467-470. | |||

