Данная заявка является частичным продолжением патентных заявок США 08/133438 и 08/133439, зарегистрированных 8 октября 1993 года, обе из которых включены в настоящее описание изобретения в качестве ссылочного материала. Обе указанные патентные заявки США 08/133438 и 08/133439 являются частичным продолжением патентной заявки США 07/931943, зарегистрированной 12 августа 1992 года, которая является продолжением заявки США 07/821872 настоящего заявителя, зарегистрированной 16 января 1992 года и являющейся продолжением патентной заявки США 07/533315 настоящего заявителя, зарегистрированной 5 июня 1990 года, каждая из которых включена в настоящее описание изобретения в качестве ссылочного материала. Патентная заявка США 07/533315 является частичным продолжением патентной заявки США 06/581406, зарегистрированной 17 февраля 1984 года и являющейся частичным продолжением патентной заявки США 06/472276, зарегистрированной 4 марта 1983 года, каждая из которых включена в настоящее описание изобретения в качестве ссылочного материала. Патентная заявка США 07/533315 также является частичным продолжением патентной заявки США 07/363383 настоящих заявителей, зарегистрированной 5 июня 1989 года, по которой принято решение о выдаче патента 12 декабря 1989 года, которая является продолжением патентной заявки США 06/867633, зарегистрированной 27 мая 1986 года, являющейся продолжением патентной заявки США 06/472276, зарегистрированной 4 марта 1983 года, каждая из которых включена в данное описание в качестве ссылочного материала. Научно-исследовательские работы, результаты которых подробно изложены в материалах настоящей заявки, проведены при содействии Национального Института Здоровья и частично Центра по Медицинской Биотехнологии Мерилендского Биотехнологического Института.
Холерный вибрион (Vibrio cholerae) является неинвазивным энтеропатогеном тонкой кишки, не способным проникать через слизистую оболочку. Локальный Siga-опосредованный иммунитет слизистой кишечника выступает, таким образом, в качестве защитного механизма. Патогенный холерный вибрион (V.cholerae) серотипа 01 выделяет белковый энтеротоксин (известный также под названием холерный энтеротоксин или холероген, или холерный экзотоксин), который ответственен за размножения и выделение кишечником холерного вибриона, что приводит к возникновению диареи - клиническому симптому холерной инфекции. Гены, ответственные за продукцию холерного энтеротоксина относятся к ctx-генам (известным специалистам также под названием tox-гены или гены токсинов). Холерная диарея может иметь чрезвычайно тяжелое течение, приводя к значительной потере воды и минеральных солей организмом человека, состоянию обезвоживания, ацидозу, шоку и смерти без оказания экстренного терапевтического лечения.
Известно, что в области хромосомы V.cholerae, содержащей ctx-гены, которые ответственны за размножение, можно обнаружить участок длиной 2700 пар оснований, названный RS1 (в виде повторяющейся последовательности). Mekalanos, Cell, v.35, pp. 253-263 (1983).
В ходе создания настоящего изобретения было также установлено, что V. cholerae продуцирует второй энтеротоксин, названный как токсин зоны плотных контактов (zonula occludents toxin), о чем ранее сообщали Fasano и др., в статье " Vibrio cholerae Produces a Second enterotoxin Which Affects Interstinal Tight Junction ", Proc. Nat. Acad. Sci. (USA) 88.5242-5246 (1991).
Созданные в настоящее время холерные вакцины можно разделить на две широкие категории: вакцины, стимулирующие антитоксический иммунитет, и вакцины, предназначенные для индукции противобактериального иммунитета. Эксперименты на модельных животных подтверждают защитную роль антитоксического и/или противобактериального иммунитета. Было сделано предположение, что, если оба иммунитета действуют одновременно, возникает синергический эффект. [Holmgren, J. и др., J.Infect.Dis 136 Suppl., S105-S1122 (1977); Peterson, J. W. Infect. Immun. 25, 594 (1979); Resnick, I.G. и др., Infect. Immun. 13, 375 (1980); Svennerholm, А.-М. и др. Infect. Immun. 13, 735 (1976)]. Однако оказывается, что у людей защитный иммунитет проявляется без синергического эффекта, то есть, либо действует механизм защиты против токсинов, либо против бактериальной инфекции [Eubanks, E.R. и др., J.Infect. Immun. 15, 533 (1977); Fujita, K. и др., J. Infect. Dis. 125, 647 (1972); Holmgren, J., J. Infect. Dis., см. ранее; Lange, S. и др., Acta Path. Microbiol. Scand Sect. С 87,145 (1978); Peterson, J.W., см. ранее (1979); Pierce, N.F. и др. Infect. Immun. 37, 687 (1982); Pierce, N.F. и др., Infect. Immun. 21, 185 (1978); Pierce, N.F. и др, J. Infect. Dis. 135,888 (1977); Resnick, I.G. и др., см. ранее; Svennerholm, А.-М. и др., см. ранее].
Убитые цельноклеточные вакцины
1. Парентеральные цельноклеточные вакцины
Уже почти столетие убитые цельные холерные вибрионы используют в качестве парентеральных вакцин; эти вакцины все еще имеются на рынке. Накопленный опыт в области использования вакцин, полученных из инактивированных цельных холерных вибрионов, проанализирован Joo, I. в работе "Cholera vaccines. " в книге Cholera. (Barua D. и Burrows W., eds.), Saunders, Phyladelphia, стр. 333-355 (1974) и Feeley, J.D. и др., в тезисах доклада Cholera and Related Diarrhears, 43rd Nobel Symp., Stockholm, 1978. (O.Oucherlong, J.Holmgren, eds) Karger, Basel, стр. 204-210 (1980). Такие вакцины стимулируют образование высоких титров сывороточных антител с вибриоцидными свойствами. Они также усиливают нарастание титра Siga антитела в кишечнике против соматического O-антигена V.cholerae в условиях вакцинации населения Пакистана, а не Швеции [Svennerholm, А.-М. и др. Infect. Immun. 30., 427 (1980); Svennerholm, А.-М. и др., Scan. J. Immun. 6, 1345 (1977)]. Было выдвинуто предположение, что такая иммунная реакция на вакцину происходит у реципиентов в Пакистане потому, что они уже иммунологически устойчивы в результате их предыдущего контакта с антигеном, в то время как такой иммунной реакции не происходит у лиц, проживающих в неэндемических регионах (например, Швеция). Парентеральные убитые цельноклеточные вакцины, как свидетельствуют результаты их апробации, обладают высоким иммунным потенциалом для защиты организма от V.cholerae гомологичного серотипа, но, как правило, на срок менее одного года [Joo, I. см. ранее; Feeley,J.C., см. ранее; Svennerholm, А.-М. и др. , см. ранее,(1980); Svennerholm, A.-M. и др., см. ранее, (1977); Mosley, W. H. и др., Bull. Wld. Hlth., Org. 49, 13 (1973); Philippines Cholera Committee, Bull. Wld. Hlth. Org., 49, 381 (1973)]. Имеются некоторые данные, подтверждающие предположение о том, что парентеральная цельноклеточная вакцина на основе холерного вибриона серотипа Инаба обеспечивает превосходный непродолжительный иммунитет как к холерному вибриону серотипа Огава, так и серотипу Инаба, тогда как вакцина серотипа Огава эффективна только против серотипа Огава. Использование адъювантов в составе парентеральной цельноклеточной вакцины, позволило обеспечить защитный иммунитет примерно на 70% в течение до полутора лет (см., например, Soroso, J.S. и др., Bull. Wld.Hlth. Org., 56, 619 (1978)). Однако в месте прививки вакцины с адъювантом довольно часто возникают сильно выраженные побочные явления (включая стерильные абсцессы), что ограничивает использование таких адъювантных вакцин общепринятым методом.
2. Пероральные цельноклеточные вакцины
Убитые цельные вибрионы, вводимые перорально, усиливают выработку секреторного антитела против вибрионного антигена. [Freter, R. J., Infect. Dis., 111, 37 (1972); Freter R. и др., J, Immunol., 91 724 (1963); Ganguly, R. и др. , Bull. Wld. Hlth.Org., 52, 323 (1975)]. Результаты, полученные другими исследователями, свидетельствовали о высокой эффективности таких вакцин, однако в большинстве случаев они приводили к развитию холерной диареи после последующей провокационной пробы патогенными вибрионами [Cash, R.A. и др. J. Infect. Dis. 130, 325 (1974)].
Токсоиды
К иммуногенам, предназначенным для противохолерной профилактики путем усиления антитоксического иммунитета, относятся следующие вещества:
1) Обработанный формальдегидом холерный токсоид;
2) Обработанный глютаральдегидом холерный токсоид;
3) Очищенная В-субъединица холерного энтеротоксина и
4) Термоинактивированный холерный энтеротоксин (обработанный или необработанный формальдегидом).
1. Обработанный формальдегидом холерный токсоид
Обработка очищенного холерного энтеротоксина формальдегидом in vitro устраняет полностью его вирулентность, в результате чего получают анатоксин, проявляющий биологическую активность с незначительным токсическиv эффектом, но стимулирующий образование антитоксиновых антител после парентеральной иммунизации животных. Однако при введении первичного анатоксина такого типа, как обезьянам, так и человеку, в качестве парентеральной вакцины он становился опять частично токсикогенным, вызывая нежелательные побочные явления в месте прививки [Northrup, R.S. и др., J. Infect. Dis., v. 125, 471 (1972)]. Формалинированный холерный токсоид с адъювантом на основе окиси алюминия вводили парентерально бангладешцам-добровольцам, в том числе кормящим матерям, но без массовой апробации этой вакцины [Merson, M.H. и др., The Lancet 1,931(1980)]. Кроме того, формалинированный холерный анатоксин, полученный в присутствии глицина, был апробирован путем перорального введения, однако не было получено никаких данных, свидетельствующих об иммунологической эффективности этой вакцины [Ohtomo, N. In Proceedings of the 12 th Joint Conference on Cholera, U. S. - Japan Cooperative Medical Science Program, Sapporo (Fukumi H., Zinnaka Y., eds), pp. 286- 296 (1976); Noriki.H, In Proceedings of the 12 th Joint Conference on Cholera, U.S. - Japan Cooperative Medical Science Program, Sapporo (Fukumi H., Zinnaka Y., eds), pp. 302-310 (1976)].
2. Обработанный глютаральдегидом холерный токсоид
В настоящее время разработаны методы для крупномасштабного производства обработанного глютаральдегидом холерного анатоксина, который практически не содержит соматического антигена [Rappaport, E.S. и др. Infect, lmmun., v.l4, р. 687 (1976)] . Исследователи возлагали надежду на то, что этот антиген, полученный в чистом виде, можно будет использовать для оценки защитного механизма антитоксического иммунитета. Массовую апробацию этого анатоксина, вводимого в виде парентеральной вакцины, провели в Бангладеш в 1974 году [Curlin, G. In Proceeding of the 11 th Joint Conference on Cholera, US-Japan Cooperative Medical Science Program., pp.314-329, New Orleans, 1975)]. Этот анатоксин усиливал образование высоких титров циркулирующих антитоксиновых антител у бангладешских реципиентов. Две вспышки холеры, вызванные вибрионом Эль Тор Инаба, а затем Эль Тор Огава захватили и район, где проводились массовые испытания вакцины, что позволило сделать достаточно достоверную оценку ее эффективности. Однако защитный эффект вакцины смогли продемонстрировать только на одной возрастной группе населения, и при этом он был ограничен продолжительностью эпидемической холеры серотипа Инаба, в результате чего эффективность обработанного глютаральдегидом холерного токсоида, полученного в чистом виде в форме парентеральной вакцины, была невысокой и ниже результатов апробации парентеральных инактивированных цельноклеточных вакцин на той же самой категории населения.
Была исследована возможность использования обработанного глютаральдегидом холерного токсоида в качестве пероральной вакцины, базируясь на предположении, что указанный анатоксин, вводимый таким путем, должен быть более эффективен за счет усиления образования антител против холерного энтеротоксина [Levine, М.М. и другие, Trans. Roy. Soc. Trop. Med. Hyg. 73, 3, (1979)] . С этой целью две группы добровольцев иммунизировали путем введения трех доз по 2.0 мг или трех доз по 8.0 мг указанного анатоксина непосредственно в просвет тонкой кишки (через кишечный зонд), каждую через месяц. После чего получившие вакцину добровольцы и не иммунизированные лица, служившие в качестве контроля, приняли участие в экспериментах с контрольным инфицированием их холерным вибрионом. Ни в одном эксперименте с антигенным стимулом не отмечено достоверного ослабления симптоматики болезни и тяжести диареи у вакцинированных по сравнению с контролем. Отсутствие эффективности перорально вводимого, обезвреженного глютаральдегидом холерного энтеротоксина может быть обусловлено тем, что способность В-субъединиц связывать ганглиозид GM1 сильно снижается в результате его обезвреживания глютаральдегидом.
3. Очищенная субъединица B холерного энтеротоксина
Холерный энтеротоксин состоит из двух субъединиц, A и B, кодируемых ctx AB-опероном. A-субъединица индуцирует ферментативные изменения, которые вызывают секрецию жидкости в просвет кишечника, в то время как нетоксичная B-субъединица представляет собой иммуногенную группу, которая связывает рецептор токсина (ганглиозид GM1) на клетках эпителия кишечника [Holmgren, J. Nature, 292, 413 (1981)]. Было продемонстрировано, что очищенная B-субъединица, вводимая как перорально, так и парентерально бангладешцам, усиливает образование Siga антитоксина в жидкости кишечника, то есть результат, полученный, возможно, за счет иммунологической устойчивости в эндемическом холерном очаге [Svennerholm, A. -M. и другие. The Lancet I, 305 (1982)].
К существенным преимуществам пероральной вакцины из B-субъединицы холерного энтеротоксина, предназначенной для усиления антитоксического иммунитета, относятся ее полная безопасность (исключается вероятность обратного превращения в токсин, как это имеет место с анатоксинами), и сохранение способности связывать рецепторы энтеротоксина на энтероцитах. Результаты исследований на животных свидетельствуют, однако, что очищенная В-субъединица имеет меньший потенциал по сравнению с нативным голотоксином в усилении выработки нейтрализующих токсин антител [Pierce, N.F., см. ранее, (1982].
Следует понимать, что очищенную B-субъединицу можно использовать, если вообще существует такая необходимость, совместно, например с пероральными инактивированными вибрионными вакцинами в качестве комбинированной пероральной вакцины, предназначенной для усиления выработки как антибактериальных, так и токсин нейтрализующих антител.
4. Термоинактивированный холерный энтеротоксин
Термоинактивированный холерный энтеротоксин представляет собой анатоксин большой молекулярной массы (Мол.масса примерно 1000000), полученный в результате термообработки холерного энтеротоксина при температуре 65oC в течение по меньшей мере пяти минут [Finkelstein, R.A. и другие, J. Immunol., 107, 1043 (1971)] . Этот анатоксин обладает иммуногенной способностью, хотя все еще сохраняет менее 5% токсикогенной активности нативного холерного энтеротоксина. При более продолжительной термообработке (например, в течение 25 минут) получают менее биологически токсичный анатоксин [Germanier, R. и другие, J. lmmun., 13, 1692 (1976)], и последующая его обработка формальдегидом полностью устраняет в нем остаточную биологическую токсичность. Полученный формалинированный термоинактивированный холерный энтеротоксин обладает по меньшей мере таким же потенциалом, как и нативный энтеротоксин в усилении образования сывороточного антитоксинового антитела после иммунизации кроликов. У швейцарцев-добровольцев развивался мощный гуморальный иммунный ответ после парентеральной иммунизации их формалинированным термоинактивированным холерным энтеротоксином, получаемым в дозах по 10, 30 или 100 мкг [Germanier, R., и другие, J. Inf. Dis., 135, 512 (1977)]. Никаких выраженных побочных реакций при этом не отмечалось.
В качестве перорального антигена, термоинактивированный холерный энтеротоксин обладает большей иммуногенностью при приеме его в форме без обработки формальдегидом. У собак переносимость к необработанному формальдегидом термоинактивированному холерному энтеротоксину такая же хорошая, как и в случае пероральной вакцины; перорально вводимые дозы его (с использованием NaHCO3 в качестве носителя) до 500 мкг не вызывают диареи. Пероральный прием пяти доз его по 500 мкг в течение примерно 42 дней обеспечивает достоверное усиление защитного иммунитета у собак против пероральной пробы с патогенным V. cholerae. В группах из шести и четырех взрослых добровольцев, получавших его в дозах по 50 мкг и 200 мкг, соответственно, с использованием NaHCO3 в качестве носителя, не отмечено побочных эффектов.
Следует понимать, что термоинактивированный холерный энтеротоксин можно использовать в комбинации, например, с живыми вакцинами, убитыми вибрионами или другими родственными антигенами, которые обладают способностью стимулировать антибактериальный иммунитет, в результате чего усиливается антитоксический иммунитет, индуцируемый термоинактивированным холерным энтеротоксином.
Комбинированные вакцины
Главным преимуществом пероральной вакцины из убитых возбудителей холеры является ее безопасность. Пероральная вакцина, состоящая из смеси антигенов, предназначенная для стимуляции как антибактериального, так и антитоксического иммунитета вероятнее всего имела успех по следующим причинам: вакцины из анатоксинов, стимулирующие только антитоксический иммунитет, неэффективны, как свидетельствуют результаты исследований, для защиты от заболевания холерой у человека, хотя их можно использовать для иммунизации модельных животных. Кроме того, пероральные и парентеральные убитые цельноклеточные вакцины, не стимулирующие никакой иммунной реакции на энтеротоксин, обеспечивают существенный защитный иммунитет против холеры у человека, хотя и на короткий промежуток времени. Более того, смеси антигенов (например, неочищенный холерный энтеротоксин, или токсин вместе с липосахаридом), стимулирующие как антитоксический, так и антибактериальный иммунитет, вызывают синергический защитный эффект.
К настоящему времени проведено пока еще два широкомасштабных испытания комбинированных вакцин в условиях практической работы. В первом испытании девять добровольцев, получавших обработанный глютаральдегидом холерный энтеротоксин (в дозе по 2 мг еженедельно в течение четырех недель) вместе с убитыми холерными вибрионами Эль Тор Инаба (1010 вибрионов два раза в неделю в течение четырех недель) через месяц после иммунизации были инфицированы, наряду с шестью не иммунизированными добровольцами, служившими в качестве контроля, патогенными холерными вибрионами Эль Тор Инаба в дозе 106 вибрионов. Диарея имела место только у двух из девяти вакцинированных, против четырех из шести в контроле (эффективность вакцины составляла 67%), при этом болезнь протекала в более легкой форме у двух заболевших, принимавших вакцину. Более важным в этом эксперименте, возможно, является факт, что V.cholerae можно непосредственно культивировать из выделенных испражнений только у двух из девяти вакцинированных, против шести из шести в контроле. Это свидетельствует о том, что иммунологические механизмы ингибируют пролиферацию вибрионов. В другом исследовании, три дозы комбинированной вакцины, состоящей из B-субъединицы холерного энтеротоксина и убитого цельного вибриона холеры давали взрослым добровольцам, которые затем участвовали в эксперименте с контрольным инфицированием для оценки эффективности вакцины. Комбинированную вакцину принимали на 0, 14, и 28 день. Каждая из трех доз вакцины содержала по 0.5 мг очищенной B-субъединицы холерного энтеротоксина и 2•1011 убитого V. cholerae (5•1010 классического вибриона серотипа Инаба, 5•1010 классического вибриона серотипа Огава, и 1•1011 вибриона Эль Тор Инаба).
Группе из одиннадцати добровольцев, иммунизированных этой комбинированной вакциной вводили через месяц после приема последней дозы вакцины патогенный V. cholerae Эль Тор Инаба, наряду с семью добровольцами, служившими в качестве контроля. Диарея имела место у семи из семи добровольцев в контроле, и только у четырех из одиннадцати вакцинированных (p=0.01). Симптоматика болезни у четырех из получавших вакцину имела менее выраженный характер.
Таким образом, результаты оценочных исследований пероральных комбинированных вакцин на основе указанного анатоксина и убитых цельных клеток демонстрируют соизмеримую степень их эффективности. Эффективность таких вакцин для стимуляции защитного иммунитета, однако, относительно невысока (55-65%), и для индуцирования механизма противоинфекционной защиты необходимо проведение нескольких ревакцинаций.
Вакцины из ослабленного V. cholerae
Исследования, проведенные с участием североамериканских добровольцев, свидетельствуют о том, что как так называемые классические холерные инфекции, так и вызванные клиническим штаммом вибриона Эль Тор, стимулируют высокую степень защитного иммунитета в течение по меньшей мере трех лет [Cash, R.A. и другие, см. ранее (1974): Levine, М.М. и другие, см. ранее (1979; Levine, М.М. и другие, " Volunteers studies in development of vaccines against Cholera and enterotoxigenic Escherichia coli: обзорная статья в книге " Acute Enteric Infections in Children: New Prospects for Treatment and Prevention. (T. Holm, J. Holmgren, M.Merson, и R. Mollby, eds.) Elsevier, Amsterdam, pp.443-459 (1981); и Levine, М.М. и другие, J. Infect. Dis.,143, 818 (1981)] . На основе результатов наблюдений за состоянием этих добровольцев, можно предположить, что наиболее многообещающий подход к решению проблемы иммунологической защиты от холеры возможно будет связан с ослабленными невирулентными штаммами V. cholerae, используемыми в качестве пероральных вакцин.
1. Природные штаммы серогруппы 01 V. cholerae.
Невирулентные штаммы серогруппы 01 V. cholerae, выделенные из источников окружающей среды в Индии и Бразилии, были апробированы на добровольцах в качестве потенциальных кандидатов для получения вакцин, и полученные результаты были неутешительны. Эти штаммы не колонизовали кишечник у человека, или колонизовали в минимальной степени; реакции вибриоцидных антител на антиген были слабые, и кроме того они не вызвали защитного иммунитета в экспериментальных исследованиях с контрольной прививкой [Cash, R.A. и другие,. Infect. Immun. 10, 762 (1974); Levine М.М. и другие, J. Infect. Dis. 145, 296 (1982)] . У этих штаммов, как показали результаты измерения методом гибридизации с использованием радиоактивно-меченого ДНК-зонда, утрачен токсиновый ген [Kaper, J.B. и другие. Infect. Immun. 32, 661 (1981)].
2. Мутагенные ослабленные штаммы.
Классический Инаба 569B был подвергнут мутации с использованием нитрозогуанидина (NTG) и был выделен гипотоксикогенный мутант. [Finkelstien, R.A. и другие, J. Infect. Dis. 129, 117 (1974); Holmes, R.K. и другие, J. Clin. Invest. 55, 551 (1975). Этот мутантный штамм, M13, вводили добровольцам. Диарея не возникала, однако штамм имел слабую колониеобразующую способность. Исследования с контрольным инфицированием продемонстрировали, что некоторый защитный эффект обеспечивался за счет иммунизации многократными дозами [Woodward, E. и другие, Develop. Biol. Stand. 33,108,(1976)].
Штамм холерного вибриона Эль Тор Огава 3083 также подвергали мутации [Honda, Т. и др., Proc. Nat. Acad. Sci., вып. 76, 2052 (1979]. В результате интенсивного скрининга и анализа тысяч колоний получили один изолят, который продолжал продуцировать иммуногенную B-субъединицу, однако без обнаруживаемой продукции A-субъединицы или голотоксина. Этот единственный изолят, Texas Star-SR, отвечал указанным критериям. Штамм Texas Star-SR продуцирует нормальную или повышенную продукцию B-субъединицы, но обнаруживает отрицательную реакцию в тест-пробах на активность голотоксина или активность A- субъединицы.
Широкомасштабные испытания вакцинного штамма Texas Star-SR проводили с участием добровольцев (см., например, Levine М.М. и др. Acute Enteric,ранее (1981). Группы из пяти добровольцев получали по две дозы 109 микроорганизмов, каждую с интервалом в неделю, и еще одна из восемнадцати добровольцев получала по две дозы 2•1010 микрорганизмов, каждую через неделю. Некоторая диарея была отмечена у 16 из 68 человек, получивших вакцину (24%). И только лишь у одного из вакцинированных общий объем стула превысил 1.0 литр (1464 мл). В типичных случаях, вызванная введением вакцины диарея сопровождалась двумя или тремя скудными жидкими стулами, не превышающими 400 мл в суммарном объеме.
Вакцинные микроорганизмы выделяли из совместных прокультур, примерно из половины реципиентов, получавших вакцину. В случае, когда культивировали содержимое тощей кишки (реципиентов, получавших вакцину в дозе 108 и более микроорганизмов), культуры обнаруживали положительную реакцию у 35 из 46 вакцинированных(76%). Сотни мутантных клонов Texas Star, выделенных из совместных прокультур и культур содержимого тощей кишки было проверено на активность субъединичного домена холерного энтеротоксина серологической тест-пробой с использованием адреноцитов и Y-1; ни в одном случае не отмечено сероположительной реакции.
Существенное нарастание в сыворотке крови нейтрализующих токсин антител обнаружено только лишь у 29% из получавших вакцину; у 93% вакцинированных отмечено значительное нарастание в сыворотке вибриоцидного антитела и титры их почти приближались к титрам, как и в случае после инфицирования патогенным V.cholerae.
В экспериментальных исследованиях с антигенным стимулом, проведенных на добровольцах, мутантный штамм Texas Star-SR, как было установлено, достоверно вызывает иммунную реакцию на антигены вибрионов как Эль-Тор Огава, так и Эль-Тор Инаба. Введение одной или двух доз пероральной вакцины из ослабленного штамма Texas Star-SR обеспечивает хороший защитный иммунитет против холеры, вызываемой вибрионом Эль-Тор.
Ясно, что использование ослабленных штаммов имеет только присущие им преимущества, поскольку такие штаммы обеспечивают имитацию иммунитета после перенесенной инфекции (поствакцинальный иммунитет). Однако, штаммы Texas Star-SR не лишены некоторых недостатков. Во-первых, мутагенез (например, с использованием нитрозогуанидина) вызывает многочисленные мутации, не все из которых обязательно распознаются. Кроме того, точный механизм генетического повреждения, который, как полагают, может быть ответственен за ослабление вирулентности Texas Star-SR, не известен. Кроме того, как и в случае с любым патогеном, претерпевшим мутацию под действием нитрозогуанидина, Texas Star-SR может восстанавливать свою вирулентность. 3. Природные 01-серологически нетипируемые штаммы V. cholerae.
Холерный вибрион серотипа 01 обычно ответственен за возникновение эпидемической холеры. Холерный вибрион, не принадлежащий к классу серотипа 01 ответственен, главным образом, за спорадические случаи гастроэнтерита и внекишечные инфекционные заболевания. Совсем недавно, однако, в печати появилось сообщение о том, что получено несколько штаммов V. cholerae, выделенных из больных во время вспышки холероподобной инфекции[Ramamurthy и другие. The Lancet, vol. 341, 703-704 (1993), содержание которого целиком приводится в данном описании в качестве ссылки]. Серологические характеристики большого количества клинических штаммов, полученных во время этой недавней эпидемии свидетельствовали о том, что эти штаммы не вызывали ни реакции агглютинации с антисывороткой 01, ни с любым из тестированных моноклональных антител, нарастаемых против фактора A, B или C V. cholerae серогруппы 01. В результате, эти вибрионы были идентифицированы как 01-серологически атипичные.
Кроме того, за исключением только лишь одного 01-серологически нетипируемого штамма в вышеуказанной тест-пробе, все остальные штаммы не серогруппы 01, испытанные во время новой вспышки в эпидемическом очаге нельзя было идентифицировать при типировании на панели из 138 антигенов, созданную для V. cholerae не 01- серогруппы в Японском Национальном Институте Здоровья, что свидетельствует о том, что штамм, ассоциируемый с этой новой вспышкой эпидемии, принадлежит к ранее нераспознаваемому типу, или появившегося недавно, не-01 серогруппы, способному вызывать эпидемическую холеру.
При анализе методом ДНК-гибридизации, все штаммы, выделенные во время этой эпидемии, скрещивались как с ctx-, так и zot- генноспецифическими зондами, но не отмечено ни одного случая гибридизации с ДНК-зондами, специфичными термоустойчивому энтеротоксину V. cholerae не серогруппы 01 (NAG-ST). Кроме того, продукция холерного энтеротоксина достоверно выявлялась методом твердофазного иммуноферментного анализа. Сообщалось, что количество энтеротоксина, продуцируемого этими новыми штаммами- изолятами, аналогично продукции энтеротоксина в клинических штаммах V.cholerae серогруппы 01. Большинство из этих штаммов, как сообщалось, были резистентны к, например, стрептомицину и фуразолидону, но чувствительны к другим традиционно используемым антибиотикам, в том числе к тетрациклину. Не сообщалось об устойчивости их к ампициллину, или его производным.
В другой недавней эпидемии, хотя общее количество случаев заболевания холеры не возросло, широкое разнообразие изолятов V. cholerae, полученных от выявленных больных с холероподобной диареей не относились к 01-серогруппе, и так же как и вышеуказанные штаммы нельзя было идентифицировать тест-пробами и с использованием стандартной панели для типирования.
Сообщалось еще об одной случае заболевания, вызванного холерным вибрионом не 01-серогруппы, мощной вспышке эпидемической холеры, которая поражала взрослое население [Albert и другие. The Lancet, vol. 341, 704 (1993), содержание которой целиком включено в данное описание в качестве ссылки]. Из мазков, взятых из прямой кишки, как сообщалось в этой публикации, приблизительно 67 процентов изолятов V. cholerae не относилось к 01-серогруппе после стандартной тест-пробы, и не один из них не типирован как V. cholerae серогруппы 01.
Сообщалось также, что V.cholerae, ответственный за эту вспышку инфекции, напоминает V. cholerae 01 серогруппы, как биохимически, так и по морфологии колоний, но в то же время, как отмечалось, он не вызывает реакции агглютинации на антисыворотку V.cholerae. Штаммы V.cholerae в тест-пробе не обнаруживали реактивности с моноклональными антителами, специфичными к фактору A V.cholerae серогруппы 01.
В этой второй эпидемии, о которой сообщалось недавно, все испытанные 01- нетипированные штаммы реагируют положительно на продукции холерного энтеротоксина в реакции агглютинации с Y-1 адреноцитами, и, как отмечено, нейтрализуются кроличьей поликлональной антисывороткой против холерного энтеротоксина.
Блот-гибридизационный тест с полимеразным усилением с использованием токсин-специфических праймеров холеры не 01-серогруппы, как также описано, вызывал амплификацию последовательности холерного энтеротоксина.
Отобранные изоляты были также проанализированы методом перевязки кроличьей подвздошной кишки и, как описано, они вызывали водянистый понос при перепетливании, аналогично, как и в случае, ассоциированном с V. cholerae 01.
Штамм V. cholerae, выделенный во время этой второй вспышки, как сообщалось, был чувствителен к определенным антибиотикам, включая тетрациклин, но был резистентен к другим вибриостатикам. Такой результат находится в противоречии с предположением о том, что преобладающие в настоящее время штаммы в данной области, большинство из которых являются изолятами, резистенты к тетрациклину.
Вследствие этих вспышек, вызвавших вспышки инфекции V. cholerae не 01-серогруппы в эпидемических очагах, имеется настоятельная потребность в получении вакцин, специфичных для этих микроорганизмов. Кроме того, было бы также желательно получить ассоциированные вакцины, которые эффективны как для 01, так и не -01 серогрупп, особенно для использования в таких регионах, где можно встретить вирулентные штаммы обеих серогрупп V. cholerae.
Краткое изложение сущности изобретения
На основе предлагаемых новых методов заявители получили и выделили мутанты вирулентного штамма Vibrio cholerae не -01 серогрупп, которые пригодны для использования в качестве вакцины для профилактики от заболевания холерой после введения вирулентного штамма V. cholerae.
Согласно изобретению заявляется авирулентный штамм Vibrio cholerae CVD112 не-01 серогруппы, используемый для вакцинации против холеры. Указанный штамм является штаммом из серогруппы 0139 и имеет генотип, включающий ctxA-, zot ace-, mer, ctxB. Указанный штамм характеризуется тем, что ДНК, кодирующую устойчивость к ртути, и ДНК, кодирующую B-субъединицу холерного энтеротоксина вставляют в сайт гемолизинового гена в хромосоме.
Указанный штамм депонирован как штамм ATCC 55752 или его производное. Согласно изобретению заявляется также авирулентный штамм Vibrio cholerae CVD112 RM не-01 серогруппы, используемый для вакцинации против холеры. Указанный штамм является штаммом из серогруппы 0139 и имеет генотип, включающий ctxA-, zot- асе-, recA-, mer, ctxB. Указанный штамм характеризуется тем, что ДНК, кодирующую устойчивость к ртути и ДНК, кодирующую B-субъединицу холерного энтеротоксина, вставляют в сайт гемолизинового гена в хромосоме.
В качестве исходного штамма для получения одного мутанта предлагаемого изобретения используют штамм V. cholerae 1837, который относится к 01-серологически атипичному штамму 0139. Заявляемые мутанты содержат делеции в области сердцевины ("ядра", кор-области) энтеротоксина V.cholerae. Эти делеции были получены с использованием эндонуклеаз рестрикции из заново идентифицированной, охарактеризованной и клонированной кор-области в ctx-локусе исходного штамма, хотя можно также использовать другие подходящие методы делеции, известные в данной области.
Для индуцирования иммуногенности, последовательности, кодирующие B субъединицу холерного энтеротоксина вновь встраивали в вакцинный штамм. Кроме того, в мутантные штаммы были встроены последовательности, кодирующие устойчивость к тяжелым металлам, например ртути, и которые обеспечивают выявление вакцинного штамма без использования потенциально терапевтически антибиотикоустойчивых маркеров.
01 серологически нетипированные вакцинные штаммы V. cholerae настоящего изобретения, таким образом, были целенаправленно трансформированы с помощью методов рекомбинантных ДНК для устранения их вирулентности, не оказывая воздействия на другие компоненты, необходимые для выработки иммунитета.
Один авирулентный штамм V.cholerae не 01-серогруппы предлагаемого изобретения является штаммом V. cholerae CVD112 (сер-, zot-, асе-, orfU-, ctxA-, ctxB, mer, HylA-, attRS1-, RS1-). Другой авирулентный штамм V. cholerae не-01 серогруппы является штаммом CVD112RM (сер-, zot-, асе-, orfU-, ctxA-, ctxB, mer, HylA-, recA-, attRS1-, RS1-).
Проведены исследования по характеристике ctx-локус вирулентного штамма V. cholerae не 01-серогруппы, в результате которых получены новые данные о том, что выделенный штамм содержит две области сердцевины (кор-области) и четыре повторяющихся последовательности (RSl-последовательности). Конструируют плазмиду, в которой есть делеция во всей, или практически во всей кор-области холерного энтеротоксина, но при этом остаются длинные фланкирующие (ограничивающие) повторяющиеся ДНК-последовательности хромосомы V. cholerae в ctx-локусе (то есть сегменте ctx-гена). В результате конъюгативного переноса гена указанной плазмиды в вирулентный V. cholerae не 01-серогруппы с последующей гомологичной рекомбинацией и второго рекомбинационного события получили V. cholerae не 01-серогруппы ДНК-последовательность только лишь с одной кор-областью и один SR1-элемент. Оставшуюся последовательность в кор-области и RSl-элемент затем удаляли с использованием вновь клонированной, а затем делированной 0139 хромосомальной ДНК.
Последовательности, кодирующие B субъединицу холерного энтеротоксина и последовательности, кодирующие устойчивость к тяжелым металлам, были затем вновь встроены в хромосому мутанта V. cholerae методом гомологичной рекомбинации.
01-Нетипированные, нетоксикогенные делеционные мутанты предлагаемого изобретения способны к колонизации тонкой кишки и стимулированию локального защитного иммунитета против внедрения бактериальной клетки. После транзиторного эпизода колонизации, вакцина вызывает защитный иммунитет против последующего заражения вирулентными токсикогенными штаммами V. cholerae не-01 серогруппы.
Изобретение также предлагает способы получения авирулентных штаммов V. cholerae не-01 серогруппы, и вакцину, полученную из этих штаммов.
В одном аспекте, данное изобретение предлагает способ получения первого авирулентного штамма V. cholerae не 01- серогруппы, содержащего делецию области сердцевины (кор-область - основная часть нуклеосомы) холерного энтеротоксина и RS1- последовательностей, но экспрессирующий B субъединицу холерного энтеротоксина и последовательности, кодирующие продукт, вызывающий устойчивость к тяжелым металлам, который включает стадии:
(а) конструирования первой плазмиды, содержащей ДНК, имеющую область сердцевины холерного энтеротоксина Vibrio cholerae и фланкирующие последовательности, достаточной длины для промотирования распознаваемой in vivo рекомбинации, лигированную с геном, кодирующим первый селектируемый чужеродный маркер, который вызывает устойчивость к избирательному фактору, при этом упомянутая первая плазмида неспособна к внехромосомной ДНК-репликации в V. cholerae;
(b) скрещивания вирулентного штамма Vibrio cholerae не 01-серогруппы с первым микроорганизмом, несущим первую плазмиду;
(с) отбора и выделения Vibrio cholerae, экспрессирующего первый селектируемый маркер;
(d) выращивания V. cholerae, выделенного на стадии (с) при отсутствии указанного избирательного фактора;
(е) скрининга V. cholerae, выращенного на стадии (d), на утрату экспрессии упомянутого первого селектируемого маркера;
(f) конструирования второй плазмиды, содержащей хромосомальные ДНК-последовательности Vibrio cholerae не 01- серогруппы, которые ограничивают локус холерного энтеротоксина, делетилированные (удаленные) из ДНК, содержащей область сердцевины холерного энтеротоксина и RSl-последовательности, лигированную со вторым селектируемым чужеродным маркером, который вызывает устойчивость ко второму избирательному фактору;
(g) скрещивания продукта, скринированного на стадии (е) со вторым микроорганизмом, несущим упомянутую вторую плазмиду; и
(h) отбора Vibrio cholerae, который экспрессирует второй селектируемый маркер;
(i) выращивания селектированного продукта стадии (h) при отсутствии второго избирательного фактора;
(j) скрининга продукта V. cholerae стадии (i) на утрату упомянутого второго селектируемого маркера; и
(k) выделения продукта, скринированного на стадии (j).
(l) конструирования третьей плазмиды, которая содержит хромосомальные последовательности V. cholerae достаточной длины для промотирования распознаваемой in vivo рекомбинации фланкирующие последовательности B-субъединицы холерного энтеротоксина, достаточные для того, чтобы вызвать иммуногенность и последовательности, кодирующие продукт, вызывающий устойчивость к тяжелым металлам, и лигирована с третьим селектируемым чужеродным маркером, при этом указанная третья плазмида неспособна к внехромосомной ДНК-репликации в V. cholerae;
(m) скрещивания скринированного продукта стадии (j) с третьим микроорганизмом, несущим указанную третью плазмиду;
(n) отбора по фенотипу Vibrio cholerae, который экспрессирует третий селектируемый маркер; и
(o) выделения селектированного продукта стадии (1).
Указанный способ получения авирулентного штамма V. cholerae не-01 серогруппы характеризуется тем, что упомянутый вирулентный штамм Vibrio cholerae не-01 серогруппы является штаммом V. cholerae 1837, который относится к 01-серологически атипичному штамму 0139. Согласно изобретению, указанный выделенный отобранный продукт на стадии (o) является штаммом
V. cholerae CVD112.
V. cholerae не 01-серогруппы, полученный в соответствии с указанным способом, представляет собой продукт с делецией всей области сердцевины холерного энтеротоксина и RS1-последовательности в ctx-локусе, но экспрессирует последовательности B субъединицы холерного энтеротоксина, достаточные чтобы вызвать иммуногенность, и последовательности, которые вызывают устойчивость к тяжелым металлам.
В другом аспекте, данное изобретение предлагает второй авирулентный V. cholerae не 01-серогруппы, который содержит делецию кор-области холерного энтеротоксина и RS1- последовательности V. cholerae не 01-серогруппы, но экспрессирует повторно вставленные последовательности B субъединицы холерного энтеротоксина, достаточные, чтобы вызвать иммуногенность, и последовательности, кодирующие продукт, вызывающий устойчивость к тяжелым металлам. Указанный мутант V. cholerae не 01-серогруппы предлагаемого изобретения представляет собой штамм минус-фенотипа, который дополнительно содержит делецию, в которой продукт recA- локуса инактивирован или утрачен (отсутствует). Предлагаемый способ включает стадии:
(а) конструирования первой плазмиды, содержащей ДНК, имеющую область сердцевины холерного энтеротоксина Vibrio cholerae и фланкирующие последовательности достаточной длины для промотирования распознаваемой in vivo рекомбинации, лигированную с геном, кодирующим первый селектируемый чужеродный маркер, который вызывает устойчивость к избирательному фактору, при этом указанная первая плазмида неспособна к внехромосомной ДНК-репликации в V. cholerae;
(b) скрещивания вирулентного штамма Vibrio cholerae не 01-серогруппы с первым микроорганизмом, несущим первую плазмиду;
(с) отбора по фенотипу и выделения Vibrio cholerae, экспрессирующего первый селектируемый маркер;
(d) выращивания V. cholerae, выделенного в стадии (с) при отсутствии указанного избирательного фактора;
(е) скрининга V. cholerae, выращенного в стадии (d), на утрату экспрессии указанного первого селектируемого маркера;
(f) конструирования второй плазмиды, содержащей хромосомальные последовательности Vibrio cholerae не 01-серогруппы, которые ограничивают локус холерного энтеротоксина, удаленные из ДНК, содержащей область сердцевины холерного энтеротоксина и RS1 - последовательности, лигированную со вторым селектируемым чужеродным маркером, который вызывает устойчивость ко второму избирательному фактору;
(g) скрещивания продукта, скринированного на стадии (е) со вторым микроорганизмом, несущим упомянутую вторую плазмиду; и
(h) отбора по фенотипу Vibrio cholerae, который экспрессирует второй селектируемый маркер;
(i) выращивания селектированного продукта стадии (h) при отсутствии второго избирательного фактора;
(j) скрининга продукта V.cholerae стадии (i) на утрату указанного второго селектируемого маркера; и
(k) выделения продукта, скринированного в стадии (j).
(l) конструирования третьей плазмиды, которая содержит хромосомальные последовательности V. cholerae достаточной длины для промотирования распознаваемой in vivo рекомбинации, которые ограничивают последовательности B-субъединицы холерного энтеротоксина, достаточные, чтобы вызвать иммуногенность, и последовательности, кодирующие продукт, вызывающий устойчивость к тяжелым металлам, и лигирована с третьим селектируемым чужеродным маркером, при этом указанная третья плазмида неспособна к внехромосомной ДНК-репликации в V. cholerae;
(m) скрещивания скринированного продукта стадии (j) с третьим микроорганизмом, несущим указанную третью плазмиду;
(n) отбора Vibrio cholerae, который экспрессирует третий селектируемый маркер;
(о) выделения селектированного продукта стадии (n);
(р) конструирования четвертой плазмиды, содержащей хромосомальные последовательности V.cholerae достаточной длины для промотирования распознаваемой рекомбинации в recA-локусе, фланкирующие последовательности с делецией в локусе recA гена для инактивирования продукта экспрессии rec A гена, и лигированную с четвертым селектируемым маркером, при этом указанная четвертая плазмида неспособна к внехромосомной ДНК- репликации в V. cholerae;
(q) скрещивания скринированного продукта на стадии (о) с четвертым микроорганизмом, несущим указанную третью плазмиду;
(r) отбора по Vibrio cholerae, который экспрессирует третий селектируемый маркер; и
(s) выделения селектированного продукта на стадии (r).
Делеционные мутанты Vibrio cholerae предлагаемого изобретения можно использовать в качестве вакцин для выработки защитного иммунитета против холеры, возбудителем которой является V.cholerae не 01-серогруппы, а также в методах, при создании противохолерных вакцин.
Один штамм Vibrio cholerae предлагаемого изобретения, обозначенный как CVD112, вызывает достоверный поствакцинальный иммунитет у человека при последующем воздействии похожего штамма не 01-серогруппы. Другой штамм Vibrio cholerae предлагаемого изобретения, обозначенный как культура CVD112 RM, может вызывать существенный поствакцинальный иммунитет у человека, в условиях прививки похожего штамма не-01 серогруппы, и также неспособен к recA-опосредованной гомологичной рекомбинации.
Изобретением также является вакцина для защиты от заболевания холерой при воздействии Vibrio cholerae не-01 серогруппы, которая, согласно изобретению, содержит авирулентный штамм CVD112 или CVD112 RM не-01 серогруппы.
Краткое описание чертежей
Фиг. 1. Штамм V. cholerae N 16961 (pJBK55) (Apr)
Фиг. 2. Методика перекрестного связывания и конъюгативного переноса гена для конструирования V. cholerae JBK56.
Фиг. 3. V. cholerae JBK56.
Фиг. 4. Схема конструирования JBK21.
Фиг. 5. Схема конструирования pJBK54.
Фиг. 6. Схема конструирования V. cholerae JBK56.
Фиг. 7. Рекомбинация in vivo путем перекрестного связывания и элиминацией ctx-гена.
Фиг. 8. Схема конструирования pJBK51.
Фиг. 9. Схема конструирования pCVD14 и pCVD15.
Фиг. 10. Схема конструирования pJBK108.
Фиг. 11. Схема конструирования pJBK107.
Фиг. 12. ДНК-последовательности и соответствующие аминокислотные последовательности (верхняя часть) сайтов рестрикции XbaI и ClaI, определяющих концы вырезанного XbaI-ClaI фрагмента (длиной 550 п.о.) A-субъединицы в серотипе Огава 395, и для (нижней части) зону сочленения в CVD101 после делеции этого фрагмента и вставки XbaI линкера.
Фиг. 13 (A и B). Влияние культурального надосадка V.cholerae на перевязку подвздошной кишки (Isc) и тканевую ионную проводимость (Gt). Показатели даны по 6 животным в каждой точке в зависимости от времени; скобки - стандартная погрешность 1A, Влияние надосадков V.cholerae 395 на Isc (сплошная линия) и Gt (пунктирные линии). B, Влияние культуральных надосадков V. cholerae 395 (сплошная линия), CVD101 (длинная пунктирная линия) и 395N1 (линия точками) на Gt. Среда, используемая в качестве контроля (короткая пунктирная линия) включает культуральную среду с неинфицированными клетками.
Фиг. 14. Количественные характеристики вариабельности зоны плотного контакта ZO в тканях под воздействием культуральных надосадков или бульонной культуры в качестве контроля.
Фиг. 15. Реверсия вариаций Gt, индуцируемых культуральным надосадком V. cholerae 395. Культуральные надосадки V. cholerae (треугольники) и неинфицированную клеточную среду (квадраты) добавляли и удаляли во время, обозначенное стрелками.
Фиг. 16. (A, B, C и D) Схема конструирования CVD109. Zot и ctx-гены сцеплены друг с другом на хромосоме V. cholerae и находятся в области хромосомы V. cholerae, которая содержит множественные повторы ДНК-фрагмента длиной 2700 п. о. , названную элементом RSI (повторяющаяся последовательность). RSl-элементы находятся на обеих сторонах zot и ctx-генов в вирулентном штамме V. cholerae E7946 (биотипа Эль Тор, серотипа Огава). Zot и ctz-гены изображены на фигуре большой незаштрихованной или заштрихованной стрелкой. RS1- последовательности изображены на фигуре более маленькой сплошной стрелкой.
Фиг. 17 (A и B). Последовательность ДНК zot-гена зоны плотного контакта (zonula occludens) энтеротоксина, начиная от 1 до 1428 нуклеотида. Заглавные буквы сверху последовательности ДНК свидетельствуют о предсказуемой аминокислотной последовательности ZOT-белка, кодируемого zot-геном.
Фиг. 18. Схема конструирования плазмиды pCVD621 и плазмиды pCVD622.2B.
Фиг. 19 (A, B, C и D). Схема конструирования CVD110.
На фигурах сокращения, используемых для сайтов рестрикциии имеют следующие значения:
A = Сайт эндонуклеазы рестрикции AccI
B = Сайт эндонуклеазы рестрикции BgIII
C = Сайт эндонуклеазы рестрикции ClaI
E = Сайт эндонуклеазы рестрикции EcoRI
H = Сайт эндонуклеазы рестрикции HindIII
P = Сайт эндонуклеазы рестрикции PstI
S = Сайт эндонуклеазы рестрикции SalI
X = Сайт эндонуклеазы рестрикции XbaI
K = Сайт эндонуклеазы рестрикции KpnI
Другие сокращения на фигурах или в любом месте описания имеют следующие значения:
Ap = ген устойчивости к ампициллину
Apr = фенотип устойчивости к ампициллину,
Aps Фенотип, чувствительный к ампициллину
Chrom = Хромосома
Cm = ген устойчивости к хлорамфениколу
CT = холерный энтеротоксин
ctx = ген холерного энтеротоксина (холероген)
CTA = A субъединица холерного энтеротоксина
ctxA = ген A субъединицы холерного энтеротоксина
CTB = B субъединица холерного энтеротоксина
cCxB = ген B субъединицы холерного энтеротоксина
hylA = гемолизиновый ген
kb = тысяча пар оснований
mer = ген устойчивости к ртути
p = Плазмида
Su = сульфонамид
Sur = фенотип устойчивости к сульфонамиду
Tc = тетрациклин
Tcs = фенотип, чувствительный к тетрациклину
Tp = Триметоприн
zot = ген зоны плотного контакта (zonula occludens) холерного энтеротоксина
асе = ген вспомогательного белка в локусе холерного энтеротоксина
сер = потенциал кодирующей последовательности для предполагаемого, хотя еще не оформленного, кодируемого ядра.
Фиг. 20. Карта, демонстрирующая относительные положения ctx-, zot- и ace-генов, OrfU и фланкирующие последовательности RS1-элементов (RS1 показаны большими стрелками). Два блока с вертикальными полосами соответствуют двум открытым рамкам считывания, в которых первоначально локализована ACE-активность. В настоящее время считают, ACE-активность локализуется относительно открытой рамки считывания у локуса zot-гена. На фигуре показан фрагмент в составе плазмиды pCVD630.
Фиг. 21(A и B). Активность штаммов CVD110 и CVD110 V.cholerae, содержащих плазмиду pCVD630, измеренная в камере Ussing. Панель на фиг. 21A слева показывает изменения, индуцируемые в коротком потоке кругооборота (ISC), а панель на фиг. 21B справа показывает изменения разности потенциала (PD).
Фиг. 22. Последовательность ДНК длиной 2.9 т.п.о. EcoRV- фрагмента, содержащая ACE-активность (GATATC в начале и конце последовательности является сайт рестрикции EcoRV). Показана полная последовательность длиной 395 аминокислотных остатков штамма классического биотипа (SEQ ID N: 1). Ниже этой показана вставка в этом участке из штамма Е7946 Эль Top (SEQ ID N: 4)- показаны только те аминокислотные остатки Е7946, которые отличаются от последовательности 395. В случае идентичности этой последовательности для двух штаммов, показана только последовательность 395. (Пунктирные линии в остатках 236-239 свидетельствуют о том, что штамм E7946 содержит AGT-вставку, которая отсутствует в последовательности 395). Над строкой первичной ДНК-структуры показана аминокислотная последовательность (в виде только лишь одной прописной буквы), предсказанная на основе последовательности 395. Два сайта трансляции ORFs; классический 395 OrfU содержит фрагмент аминокислотных остатков с 1034 по 2218 (SEQ ID N: 3); классический 395 асе содержит фрагмент аминокислотных остатков с 2221 по 2508 (SEQ ID N: 2); Эль Top OrfU включает фрагмент от 1037 по 2221 (SEQ ID N: 6); и, Эль Top асе содержит фрагмент остатков от 2224 по 2511 (SEQ ID N: 5).
Фиг. 23. Иллюстрирует расположение локуса холерного энтеротоксина в хромосоме штамма 1837 V. cholerae не -01 серогруппы.
Фиг. 24. Иллюстрирует расположение локуса токсина холеры V. cholerae штамма 1837 (первая строка), штамма 1837.1 (вторая строка), штамма 1837.2 (третья строка) и штамма CVD112 (четвертая строка).
Фиг. 25. Иллюстрирует плазмиды pLC13, pLC14, pLC15 и pLC16.
Фиг. 26. Схема вставки последовательностей, кодирующих B субъединицу холерного энтеротоксина и последовательности, кодирующие устойчивость к ртути в ядро хромосомы в hylA-локусе штамма 1837.2 V. cholerae 0139.
Фиг. 27. Иллюстрирует рекомбинацию, которая имеет место в гесA-локусе V. cholerae в поколении штамма CVD112RN.
Фиг. 28 (A, B, C и D). Анализ на проницаемость с использованием зародышевого агглютинина пшеницы - пероксидазы хрена (WGA-HRP), измеренную на кроличьем гомогенате подвздошной кишки в условиях воздействия культуральным надосадком различных штаммов V. cholerae. A, контрольная среда; B, штамм V. cholerae 395: С, штамм V. cholerae 395N1; D, штамм V. cholerae CVD101.
Фиг. 29 (A и B). Исследование замороженной фракции из ткани подвздошной кишки кролика в условиях воздействия культуральным надосадком V.cholerae: A, интактный ZO с многочисленными пересечениями (клинообразными) между М-нитями, microvilli. B, поврежденный ZO из ткани подвздошной кишки в условиях воздействия штамма V. cholerae 395; структура ретикулина, по-видимому, упрощается за счет довольно значительного снижения частоты пересечений нитей.
Подробное раскрытие сущности изобретения
Настоящее изобретение заявляет штаммы V.cholerae, способ их получения и методы их использования. Заявляемые штаммы Vibrio cholerae, пригодные для использования в качестве вакцинных штаммов, целенаправлено трансформированы технологией рекомбинантных ДНК, для получения авирулентного штамма без существенного воздействия на другие клеточные компоненты, необходимые для обеспечения защитного иммунитета. Это ослабление было достигнуто за счет гидролиза рестриктазами плазмид, несущих соответствующие последовательности V. cholerae, со специфической полной делецией локуса генов, кодирующих холерный энтеротоксин, или его части. Конъюгативный перенос гена указанных плазмид, несущих делированные гены холерного энтеротоксина в вирулентные клетки-хозяина V. cholerae осуществляется путем генетического отбора in vivo рекомбинантов, привел к получению авирулентных штаммов, не содержащих полностью, или содержащих частично холероген. Следует понимать, что методы настоящего изобретения подходят для выделения других делеционных мутантов вирулентного V. cholerae, или для выделения штаммов, содержащих полностью или частично делетированные последовательности, которые затем повторно вставляют в клеточное ядро V. cholerae.
В качестве исходного материала для получения вакцин использовали вирулентный штамм Vibrio cholerae N 16961 серогруппы 01. Штамм N 16961, как продемонстрировано, вызывает как типичную симптоматику энтероинфекции, так и мощный защитный иммунитет против последующего заражения [Levine, М.М. и другие. Acute enteric, см. ранее, 1981]. Область хромосомы бактериальной клетки, которая как установлено, ответственна за продуцирование холерного энтеротоксина, клонировали в плазмиду pBR325, после скрининга гидролизата HindIII ДНК V. cholerae с использованием термоустойчивого зонда Е. Coli к гену энтеротоксина [Kaper и другие, Amer. Soc., см. ранее; Kaper и другие, Symposium, см. ранее]. Установлено, что хромосомальный фрагмент HindIII холерного вибриона содержит все гены, необходимые для продуцирования холерогена. Впоследствии этот участок хромосомальной ДНК был проанализирован и проведено картирование локуса генов энтеротоксина [Kaper, J.B. и другие. Lancet II, 1162 (1981)]. Для вырезки ДНК-фрагментов, содержащих эти гены, использовали рестриктирующие ферменты, а вставку фрагмента ДНК, кодирующего селектируемый маркер (например, устойчивость к ампициллину) осуществляли лигированием (сшиванием). Затем осуществляли клонирование гена устойчивости к ампициллину с фланкирующими последовательностями ДНК вибриона в производное плазмиды pRK290, которое можно использовать для переноса ДНК из Е. Coli в V. cholerae. Полученную плазмиду pJBK55 переносили из E.Coli К-12 в штамм V. cholerae N 16961 методом конъюгации.
Полученный штамм V. cholerae N 16961 (pJBK55) (Apr) содержал область, имеющую в хромосоме неповрежденный холероген а, во внехромосомальном состоянии плазмиду, содержащую эту же самую область с делетированными генами энтеротоксина и замененными на гены, кодирующие устойчивость к ампициллину (См. фиг. 1). При низкой частоте повторения, возможно при 106 до 108, идентичные области, ограничивающие хромосомальные гены холерного энтеротоксина и внехромосомальный (плазмидный) ген устойчивости к ампициллину, обычно либо совершают обмен, либо "перекрестную рекомбинацию", или претерпевают рекомбинацию in vivo так, что в области ДНК, содержащей ген устойчивости, происходит замена на ген энтеротоксина на хромосоме (фиг. 2). Это редкий результат отбирают путем тест-пробы смеси мутированных и немутированных клеток по фенотипу клеток, которые способны выступать в качестве хозяина на внедрение чужеродной плазмиды [Ruvkun, G.B. и другие. Nature, 289, 85 (1981)]. Плазмиды подразделяются на группы A-W, элементы которых не могут стабильно сосуществовать друг с другом. Например, плазмида группы несовместимости P не может стабильно удерживаться в той же самой клетке в качестве другой плазмиды группы P (Inc Р). Таким образом, Inc P-плазмиды, например как R702, которые ответственны за устойчивость к сульфонамиду, не могут сохраняться в клетке, которая имеет другую Inc P-плазмиду, например PRK 290, pJBK45, или pJBK55. Поэтому, плазмида R702 может сохраняться стабильно в штамме, ген устойчивости к ампициллину которого рекомбинирован в хромосому, а не в штамме, где Inc P-плазмида (например, pJBK55) реплицируется внехромосомально. При скрещивании штамма Е. Coli, содержащего Inc P-R702 плазмиду (сульфомидоустойчивую) и плазмиду pJBK55 V. cholerae (ампициллиноустойчивую) и проведении отбора на V. cholerae, которые являются устойчивыми как к ампициллину, так и сульфонамиду, выделяют клеточные колонии, в которых устойчивость к сульфонамиду обеспечивается вне хромосомы плазмидой р702, а к ампициллину достигается внехромосомально через замену холерогена на гена устойчивости к ампициллину (фиг. 3).
Один такой штамм, обозначенный как V. cholerae JBK56 выделяли и при испытании его в качестве продуцента энтеротоксина было обнаружено, что он авирулентен.
Конечный продукт вакцинного штамма, JBK70, производили путем замены гена устойчивости к ампициллину, терапевтически используемого антибиотика, на ген устойчивости к ртути. Эту замену осуществляли путем клонирования гена устойчивости к ртути непосредственно в локус гена ампициллиноустойчивости плазмиды pZBK55, что приводит к инактивации гена устойчивости к ампициллину и вызывается устойчивость к ртути. Полученная плазмида, pJBK66 была также несовместима с R702 и была перенесена в плазмиду pJBK56 V. cholerae.
Затем отбирали мутант, в котором ген устойчивости к ртути был рекомбинирован в хромосому, с использованием Inc P плазмиды R702 и, выбирая V. cholerae, которые были чувствительны к ампициллину и устойчивы к ртути и сульфонамиду.
Затем отобрали спонтанно образованный из pR702 делеционный мутант. Конечный мутант, JBK70, не проявлял вирулентности и имел устойчивость только лишь к ртути.
Штамм вакцины JBK70 V. cholerae - один из серотипа Инаба. Другой главный серотип V. cholerae - серотип Огава. Возлагают надежду на то, что вакцина, приготовленная из одного серотипа холерного вибриона вызовет защитный иммунитет против другого серотипа (34). Живую вибрионную вакцину можно приготовить также из серотипа Огава и испытать ее с участием добровольцев (Levine, М.М. и другие. Enteric Acute, см. ранее (1981). Точные мутации, полученные в штамме JBK56 V. cholerae серотипа Инаба затем воспроизводили в штамме E7946 путем целенаправленного переноса области хромосомы, содержащей устойчивость к ампициллину в локус холерогена в JBK56, в E7946 через генетическую рекомбинацию, опосредованную P, половым фактором V.cholerae [Parker, C. и другие, см. ранее]. P-фактор, который несовместим с Inc P-плазмидой, переносили затем в JBK56 с последующим скрещиванием с рифампинустойчивым мутантом E7946. Путем отбора мутанта, который был устойчив к ампициллину и рифампину, был получен изолят серотипа Огава с полностью делетированными генами энтеротоксина.
В случае недостаточной степени антибактериального иммунитета для защиты можно добавить антитоксический компонент введением обратно генов для продуцирования B, а не A субъединицы холерного энтеротоксина. Это достигали путем клонирования гена B-субъединицы в клонирующий вектор pMS9. Полученная плазмида, pJBK51, продуцирует высокие уровни B-субъединицы, и затем она была введена в невирулентный вакцинный штамм V. cholerae JBK70 с получением ослабленной вибрионной вакцины штамма JBK70 (pJBK51), не способной продуцировать A-субъединицу.
Штаммы вакцины настоящего изобретения получены по существу из V.cholerae N 16961, относящегося к серотипу Инаба. Следует понимать, что другие штаммы или другие биотипы и серотипы можно использовать вместо штамма N 16961 для получения вакцинных штаммов, имеющих специфические делеции в локусе ctx гена или генов, или в других участках по хромосоме V. cholerae. Поскольку этот объект-изолят из таких вакцинных штаммов должен имитировать инфекционный процесс, не вызывая патологий, то сайт-направленный мутагенез вирулентных штаммов, как описано в этой заявки, обеспечивает существенный иммунный потенциал для профилактической защиты от холерной инфекции.
Например, авторы настоящего изобретения получили другой вакцинный штамм CVD101 V. cholerae, характеризуемый делецией большей части A субъединичного гена в 2 копиях из ctx-гена. При конструировании штамма CVD101 в соответствии с общими принципами, о которых указывалось ранее, например при конструировании JBK70, полученный штамм CVD101, за исключением того, что не имел маркеров устойчивости, не требовал исправления. На заключительной стадии при выделении второго и обнаруженного in vivo рекомбинанта включали процедуру отбора на чувствительность к антибиотику, например чувствительности к тетрациклину, однако родительский штамм вставляли в локус A- субъединичного гена из CT-гена устойчивости к тетрациклину.
Необходимо понимать, что такая чувствительность к антибиотикам представляет собой другой пример селектируемого маркерного гена.
Продуцирование вакцинных штаммов можно осуществлять широким разнообразием методов, включая: Vibrio cholerae субкультивируют из маточных культур на агаре с сердечно- мозговой вытяжкой (BНIA) и инкубируют при температуре 37oC в течение ночи.
Идентичность проверяют с использованием группо- и типоспецифичной антисыворотки, и от двадцати до тридцати клеточных колоний суспендируют в бульоне из сердечно-мозговой вытяжки. Прединкубированные агаровые пластины с сердечно-мозговой вытяжкой инокулируют с BHI-суспензией. После инкубации в течение 5-6 часов с каждой пластины собирают надосадочную культуру с добавлением 5 мл стерильного физиологического раствора, забуференного pH=7,2±0,1. Собранные микроорганизмы центрифугируют в охлажденном состоянии при скорости 750 об/10 мин, ресуспендируют и два раза промывают в четырехкратном количестве от первоначального объема культуры. Полученную суспензию стандартизируют спектрофотометрическим методом и разводят для получения дозы микроорганизмов, необходимой для иммунизации (примерно 106, которая меняется в зависимости от результатов исследований с участием добровольцев). Повторный посев производят на четырехлуночном планшете из инокулята перед и после прививки для подтверждения содержания инокулята. Конечный инокулят анализируют с окрашиванием методом Грамма и агглютинируют с гомологичной антисывороткой перед приемом.
Штаммы Vibrio cholerae настоящего изобретения можно вводить перорально. Два грамма NaHCO3 растворяют в пяти унциях дистиллированной воды. Добровольцы выпивают четыре унции водного раствора NaHCO3; через одну минуту добровольцы принимают суспендированные в оставшейся унции NaHCO3/воды вибрионы. Добровольцы перед и после прививки ничего не принимают.
Что касается безопасности этого вакцинного штамма, то следует отметить, что он "не переключается обратно" до токсикогенного состояния (то есть, продуцирует интактный холерный энтеротоксин), которое может вызвать заболевание. Для определения активности энтеротоксина используют два основных метода анализа: анализ с использованием адреночувствительных VI-типа клеток [Sack, D. A. и другие. Infect. Immun. 11, 334 (1975)] и твердофазный иммуноферментный анализ (ELISA) [Sack, D.A. и другие. J. Clin. Micro., 11, 35 (1980)] . Штамм вакцины (JBK 70) был многократно проверен с использованием этих двух методов анализа и всякий раз обнаруживалась отрицательная реакция. Гораздо более существенным моментом в этих анализах, однако, является то, что они проводились на присутствие холерогенов.
В ДНК генов холерного энтеротоксина можно вводить радиоактивную метку и использовать в качестве сайт- специфического зонда для распознавания других генов холерного энтеротоксина в штамме, в соответствии с методом Саузерна, E. М. J. Mol. Bio. 98, 503 (1975). При тест-пробе с использованием этого метода, вакцинный штамм предлагаемого изобретения не содержит никакого обнаруживаемого генетического материала, который бы включал в свой состав холерный энтеротоксин. Вакцина также была испытана на модельных мышах "сосунках" в соответствии с методом Baselski, V. и другими. Infect. Immun., 15, 704 (1977). После многократного проведения (десяти) серийных проб, не отмечено никакого накопления серозной жидкости, что свидетельствует об отсутствии признаков болезни. Как и ожидалось, штамм JBK70 обладал способностью к колонизации кишечника мыши сосунка.
Чтобы избежать нежелательных побочных эффектов вакцинных штаммов, например поноса со рвотой, судорог, и другой симптоматики болезни, вакцинные штаммы могут также включать второй рестрикционный фрагмент ДНК с делецией последовательности, кодирующей токсин зоны плотного контакта (zonula occludens) токсин (ZOT).
Культура Vibrio cholerae включает штамм Vibrio cholerae, содержащий первый рестрикционный фрагмент ДНК, удаленный для обеспечения авирулентности и сохранения способности к колонизации тонкого кишечника хозяина-организма животного и второй рестрикционный фрагмент ДНК с делецией зоны плотного контакта (zonula occludens) токсин (ZOT) для ослабления остаточной диареи в хозяине-животном. Первый фрагмент ДНК с делетированной последовательностью может кодировать холероген V. cholerae или его часть, например A-субъединицу. У одного изолированного делеционного мутанта имеется делеция в локусе гена ctx, ограниченная сайтами рестрикции AccI, и делеция в локусе гена zot. Другой изолированный делеционный мутант содержит делецию в локусе ctx гена, определяемую сайтами рестрикции XbaI и ClaI, и делецию в локусе гена zot, определяемую сайтами рестрикции StuI и AccI.
Способ выделения таких делеционных мутантов Vibrio cholerae включает стадии:
(а) конструирования первой плазмиды, включающей фланкирующие ДНК последовательности Vibrio cholerae, содержащей один или несколько делетированных фрагментов в сайте эндонуклеазы рестрикции и ген для селектируемого чужеродного маркера, сшитую с указанными фланкирующими последовательностям для замены и вставки их в место упомянутого удаленного фрагмента, при этом упомянутые последовательности имеют достаточную длину для промотирования распознаваемой in vivo рекомбинации;
(b) скрещивания вирулентного штамма Vibrio cholerae с первым микроорганизмом, несущим первую плазмиду;
(с) отбора Vibrio cholerae, экспрессирующего первый селектируемый маркер;
(d) скрещивания продукта, отобранного на стадии (с) со вторым микроорганизмом, несущим вторую плазмиду со вторым селектируемым маркером, при этом указанная вторая плазмида несовместима с первой плазмидой;
(е) отбора Vibrio cholerae, экспрессирующего первый селектируемый маркер и второй селектируемый маркер;
(f) конструирования третьей плазмиды, содержащей фланкирующие ДНК-последовательности Vibrio cholerae из одного или нескольких делетированных рестриктазой фрагментов, гомологичных к описанным на стадии (а), но отличающихся отсутствием селектируемого чужеродного маркера;
(g) скрещивания отобранного продукта на стадии (е) с третьим микроорганизмом, несущим третью плазмиду, описанную в стадии (f); и
(h) отбора Vibrio cholerae, больше не экспрессирующего указанный первый селектируемый маркер.
Этот метод можно использовать только лишь для штаммов фенотипа ZOT или получения производного штамма ZOT из любого штамма с делецией генов холерного энтеротоксина.
Другая культура Vibrio cholerae включает штамм Vibrio cholerae штамм, имеющий область хромосомальной ДНК с делецией холерного энтеротоксина и токсина зоны плотного контакта (zonula occludens) токсин ZOT.
Метод изоляции таких делеционных мутантов Vibrio cholerae включает стадии (а) конструирования плазмиды, содержащей ДНК- последовательности Vibrio cholerae, кодирующие гены холерного энтеротоксина и энтеротоксина зоны плотного контакта (zonula occludens) и ген селектируемого чужеродного маркера, где упомянутая плазмида неспособна к репликации в Vibrio cholerae вне хромосомы;
(b) скрещивания микроорганизма, несущего упомянутую плазмиду с вирулентным штаммом Vibrio cholerae, содержащим указанные последовательности для промотирования распознаваемой in vivo рекомбинации:
(с) отбора на Vibrio cholerae, экспрессирующего указанный селектируемый маркер;
(d) выращивания продукта изолированного на стадии (с) при отсутствии избирательного фактора;
(е) отбора на Vibrio cholerae, не экспрессирующего больше такого селектированного маркера; и, как следствие этого, имеющего область хромосомальной ДНК, с делецией генов холерного энтеротоксина и энтеротоксина зоны плотного контакта (zonula occludens). Стадия (b) может включать манипуляции:
b) скрещивания микроорганизма, несущего упомянутую плазмиду с вирулентным штаммом Vibrio cholerae, содержащим указанные ДНК-последовательности, вставленные между фланкирующими идентичными копиями второй последовательности, например элементов RS1 последовательности достаточной длины для промотирования распознаваемой in vivo рекомбинации.
Делеционные мутанты Vibrio cholerae настоящего изобретения пригодны для иммунизации против холеры.
Новый токсичный фактор, раскрываемый в настоящем изобретении, полученный на основе V. cholerae, увеличивает проницаемость слизистой тонкой кишки за счет воздействия на структуру зоны внутриклеточных плотных контактов или zonula occludens(ZO) (трансклеточный путь ионного переноса). Степень продукции этого фактора, вырабатываемого V. cholerae, тесно коррелирует с результатами исследований его энтеропатогенности, проведенных с участием добровольцев. При нарушении механизма нормального обмена в тонком кишечнике через трансклеточной путь, этот фактор возможно ответственен за проявление остаточной диареи, индуцируемой делеционными мутантами ctx-гена V. cholerae и может способствовать развитию тяжелой формы холерной диареи, которая отличается от других болезней, сопровождаемых кишечными расстройствами.
Проведены исследования изменения функции тонкого кишечника в условиях воздействия трех штаммов V. cholerae, один дикого типа и два ослабленных вакцинных штамма. Штамм V. cholerae 395, классический биотип, серотип Огава, является высоковирулентным штаммом, характеристики которого были изучены в широкомасштабных исследованиях, проведенных в Центре по Разработке Вакцин с участием добровольцев. Этот штамм вызывает понос с общим средним объемом стула примерно 5,5 литра (в интервале от 0,3 до 4,4 л) у более чем 90% добровольцев, получавших его в дозе 106 микроорганизмов [Levine, М.М. и другие, Infect. lmmun. 56, 161-167 (1988)]; [Levine, M.M., Cholera and Related Diarreas, 195- 203] (Karger, Basel, 1980). Холерная диарея обусловлена преимущественно ферментативным влиянием A-субъединицы холерогена на слизистую тонкого кишечника. A-субъединица холерного энтеротоксина, кодируемая геном ctx, стимулирует аденилатциклазу и приводит к секреции жидкости плазматической сети в просвет тонкого кишечника. Gill D.M. Adv. Cyclic Nucleotide res., 8, 85-118 (1977). Вакцинный штамм V. cholerae CVD101 является мутантом штамма 395 с делецией ctx-гена, у которого были удалены 94% ДНК-последовательностей, кодирующих A1 peptide холерного энтеротоксина. Удивительным фактом было однако то, что хотя CVD101 больше не продуцировал активного холерного энтеротоксина, этот штамм вызывал диарею от легчайшей до легкой формы (средний объем стула составлял 0,9 л, варьируя в интервале от 0,3 до 2,1 л) у 54% добровольцев, получавших этот микроорганизм. У второго мутанта 395, вакцинного штамма 395 N1, полученного генной технологией Mekalanos, и другие. Nature, 306, 551-557 (1983), отсутствует, по нашим расчетам, приблизительно 77% ДНК-последовательностей, кодирующих A1-пептид. В противоположность штамму CVD101, 395N1 вызвал легчайшую диарею (объем стула 0.3 л) только лишь у 1 из 21 добровольцев (P= 0,002, по сравнению с 13 из 24 добровольцев с диареей, получавших CVD101). [Herrington, D.A. и другие. J.Exp.Med. 168, 1487-1492 (1982)] . Так как эти штаммы имели сходство по их способности к колонизации тонкого кишечника, авторы настоящего изобретения полагают, что этот штамм CVD101 продуцирует секретогенный фактор, который экспрессируется слабо, или вообще не экспрессируется при воздействии 395N1 и что этот фактор ответственен за диарею, отмечаемую у добровольцев, получавших вибрионный штамм CVD101.
Эти штаммы были изучены с использованием препарата ткани тонкой кишки кролика, помещенного в камеру Ussing, традиционного метода для изучения процессов активного транспорта через слизистую тонкого кишечника. Культуральные надосадки V. cholerae добавляли к тканевому препарату и измеряли величины разности потенциалов (PD) и тока короткого замыкания (Isc). PD означает ЭДС, измеренной на стороне слизистой относительно серозной стороны исследуемой ткани и Isc - величина тока, необходимого для установки PD в состоянии 0. В результате этих замеров была рассчитана величина электропроводности ткани (Gt) в соответствии с законом Ома: Isc = PD•Gt. Первоначально было исследовано влияние сначала культуральных надосадков штамма 395 дикого типа на эти параметры с использованием среды с неинокулированной культурой, прибавляемой к совместимой ткани подвздошной кишки этого же животного в качестве отрицательного контроля. На фиг. 13A показаны полученные вариации значений Isc и Gt. Начальные пики в Isc и PD, которые появлялись как в отрицательном контроле, так и в испытуемых образцах, наиболее вероятно обусловлены совместным транспортом Na и питательных веществ, входящих в культуральную среду. В контроле наблюдался возврат к исходным значениям величины Isc и PD примерно через час, и затем показатели Isc, PD и Gt оставались неизменными на протяжении всего оставшегося времени эксперимента. Напротив, тканевые препараты, подвергнутые воздействию культуральных надосадков штамма 395 продемонстрировали достоверное увеличение величины Gt, достигающей максимального значения после 2-часовой инкубации. В испытуемых образцах не отмечено ни одного случая возврата величины Isc к ее исходным значениям, но был отмечен период установившегося состояния в промежутке между 40 и 60 минутами. Поскольку величина Isc соответствует PD•Gt и наблюдаемые значения PD через 60 минут не отличались от исходных показателей (данные не приведены), достоверное увеличение величины Isc в тканевых препаратах, обработанных штаммом 395 в этот промежуток времени может быть обусловлен только лишь повышением величины Gt. (См. фиг. 13A на 60 мин.) (12).Через 60 мин, величина Isc стала подниматься опять с увеличением PD в тканях, обработанных штаммом 395. Эта вторая фаза по-видимому отражает влияние холерного энтеротоксина на ионные потоки, поскольку очищенный CT повышает величину Isc в ткани подвздошной кишки у кролика после времени запаздывания по меньшей мере на 40 минут. Эти данные свидетельствуют о том, что имеются два фактора, экспрессируемые штаммом V.cholerae 395, который может изменять механизм ионного транспорта в камерах Ussing. Один такой фактор, холерный энтеротоксин, вызывает достоверное увеличение величины Isc и PD, начиная приблизительно 60 минут спустя после добавления культурального надосадка, в то время как второй фактор индуцирует моментальное нарастание электропроводности ткани, которую можно наблюдать в течение 20 минут после прибавления надосадочной культуры.
Вариации Gt, индуцируемые культуральными надосадками ослабленных штаммов CVD101 и 395N1 V. cholerae были следующим аспектом исследований. Штамм CVD101 вызывал мгновенное увеличение величины Gt, которое сопоставимо с результатами, полученными в случае со штаммом 395 (фиг. 13B). В противоположность этому, штамм 395N1 не вызывал мгновенного повышения величины Gt; вариация величины Gt в образцах ткани, обработанных штаммом 395 N1 были аналогичны результатам, полученным в контроле с несовместимой бульонной культурой и достоверно ниже, чем это можно наблюдать со штаммами 395 и CVD101 в течение почти 100-минутной инкубации. После этого периода, изменение величины Gt в тканях, подвергнутых воздействию штаммов 395, CVD101 и 395N1, было аналогично. Эти результаты свидетельствуют о том, что штамм 395N1 продуцировал меньшее количество или менее активную форму фактора, ответственного за это увеличение величины Gt.
Вариации в трансэпителиальной электропроводности являются ясным отражением изменения проницаемости ткани через внутриклеточное пространство, поскольку сопротивляемость плазматической мембраны относительно высока. Вследствие того, что ZO представляет главный барьер в этом трансклеточном пути и вариации величины Gt являются наиболее точным критерием оценки функции ZO, были исследованы изменения морфологии ZO, индуцированные культуральными надосадками штаммов 395, CVD101 и 395N1 V.cholerae. При введении низкомолекулярного электронно-плотного маркера, например как агглютинин из зародышевой пшеницы и пероксидазы редьки (WGA-HRP) на сторону слизистой эпителиального пласта, он обычно не проходит за пределы зоны ZO [Alberts, B. и другие, Molecular Biology of the Cells, 2-ое издание, 1989]. WGA-HRP добавляли на сторону слизистой тонкого кишечника, обработанного культуральным надосадком штаммов 395, CVD101, 395N1 или неинокулированным бульоном в качестве контроля в течение 60 минут. Тканевые препараты, обработанные неинокулированной культуральной средой были непроницаемы для WGA- HRP, в то время как в тканевых препаратах, обработанных штаммами 395 и CVD101, отмечалось окрашивание в трансклеточном пространстве. В тканевых препаратах, подвергнутых воздействию культурального надосадка штамма 395N1 не отмечено изменений, поскольку внутриклеточное пространство осталось сомкнутым в достаточной степени, чтобы исключить прохождение в него WGA-HRP (фиг. 14). Эти результаты были подтверждены и уточнены с использованием метода электронной микроскопии замороженной фракции, где количество нитей, лежащих параллельно ZO, находятся в корреляции с данными по трансэпителиальной электропроводности. В тканевых препаратах, подвергнутых воздействию культурального надосадка наблюдалось наличие смеси из претерпевшего изменения ZO и нетрансформированного ZO, сопровождаемое упрощением строения нити. Нити, расположенные перпендикулярно относительно длинной оси ZO, оказались преимущественно потерянными, приводя тем самым к снижению количества перемычек (взаимопересечений) между нитями. Степень изменения морфологии ZO, подвергнутой воздействию культурального надосадка каждого штамма оценивали путем измерения плотности перемычек между нитями. Как видно из фиг. 14, в тканях, подвергнутых воздействию культурального надосадка штаммов 395 или CVD101, отмечено существенное снижение количества нитей и сложности ретикулярной ткани ZO по сравнению с тканями, обработанными неинокулированной бульонной средой или культуральным надосадком штамма 395N1.
Изменения морфологии ZO, индуцируемые штаммами 395 и CVD101, находятся в параллельной зависимости от увеличения показателя тканевой электропроводности, вызванной этими штаммами. Функция зоны плотного контакта ZO тонкого кишечника заключается в регулировании трансклеточного пути и ограничении или предотвращении диффузии водорастворимых молекул через внутриклеточное пространство обратно в просвет кишечника. Этот механизм диффузии запускается градиентами концентрации через процессы трансэпителиального транспорта. В результате перестройки механизма трансклеточного пути, слизистая кишечника становится все более проницаемой, и вода, Na и Cl просачиваются в просвет кишечника, приводя тем самым к возникновению диареи.
Перестройка в трансклеточном пути, индуцируемого V. cholerae 395 и CVD101 является специфичной для тонкого кишечника; замена образца ткани тонкой кишки кролика на ткань слепой кишки не привела к какому-либо изменению величины Gt, индуцируемой надосадочной жидкостью культуры штамма 395 (данные не приведены). Этот факт подтверждает впервые наличие бактериального фактора, который способен к ослаблению смыкания зоны плотного контакта в интактной ткани тонкого кишечника и может представлять собой новый механизм, вызывающий бактериальную диарею. A-субъединица экзотоксина Clostridium difficile, вирусы гриппа и везикулярного стоматита (VSV), как имеются данные, ослабляют зону плотного контакта в тканевой монослойной культуре, но нигде не сообщалось о такой активности в интактной ткани или данных о связи их с диареей.
Таким образом, штаммы 395 и CVD101 V. cholerae продуцируют фактор, который может быть ответственен за возникновение диареи, наблюдаемой у добровольцев, получавших делеционные мутанты ctx-гена V. cholerae. Понос, вызванный этими ctx-мутантами, сопоставим с результатами испытаний многочисленных штаммов энтеротоксикогенной E. Coli. Этот токсинсекретирующий фактор, который авторы настоящего изобретения назвали ZOT в качестве энтеротоксина зоны плотного контакта (zonula occludens toxin), индуцирует на начальной фазе увеличение величины Isc и тканевой электропроводности, которая не связана с влиянием CT на ионные потоки. Это увеличение величины Gt ассоциируется с ослаблением смыкания зоны плотного контакта, эффекта с быстрой реверсией в первоначальное состояние после удаления культурального надосадка (фиг. 15). Быстрая реверсия этого эффекта находится в полной противоположности долговременному воздействию холерного энтеротоксина. Эти результаты не сопоставимы с ранее сделанным необъясненным результатом наблюдений Fields, и другие, J. Clin. Invest., 51, 796-804 (1972), который отмечал моментальное увеличение величины Isc, вызываемым сырыми, и еще не очищенными препаратами холерного энтеротоксина. Эти результаты могут быть сопоставимы с результатами Nishibuchi и другими, lnfect. Immun., 40. 1083-1091 (1983), которые отмечали в ранней фазе накопление жидкости (FA), не относящегося к отсроченному CT-индуцированному FA у мышей-сосунков, которым вводили V. cholerae. Способность отрицательно реагирующего энтеротоксина V. cholerae вызывать понос у добровольцев коррелирует с результатами о продуцировании ZOT двумя ослабленными штаммами, полученными из той же самой родительской клеточной линии; штамм CVD101 (вызывающий диарею) продуцирует ZOT, в то время как штамм 395N1 (не индуцирующий диарею) вызывает продукцию ZOT с незначительной или не обнаруживаемой активностью.
Другая культура Vibrio cholerae включает штамм Vibrio cholerae штамм, имеющий область хромосомальной ДНК с делецией локуса холерного энтеротоксина и энтеротоксина зоны плотного контакта ZOT, и имеющий вставку гена устойчивости к ртути и ДНК-последовательности, кодирующей B-субъединицу энтеротоксина Vibrio cholerae. Способ изоляции таких делеционных мутантов Vibrio cholerae включает стадии:
(а) конструирования плазмиды, содержащей ДНК-последовательности Vibrio cholerae, кодирующие гены холерного энтеротоксина и энтеротоксина зоны плотного контакта (zonula occludens) и ген селектируемого чужеродного маркера, где упомянутая плазмида неспособна к репликации в Vibrio cholerae вне хромосомы;
(b) скрещивания микроорганизма, несущего упомянутую плазмиду с вирулентным штаммом Vibrio cholerae, содержащим указанные ДНК-последовательности, вставленные между фланкирующими идентичными копиями второй последовательности, имеющей достаточную длину для промотирования распознаваемый in vivo рекомбинации;
(с) отбора на Vibrio cholerae, экспрессирующего указанный селектируемый маркер;
(d) выращивания продукта изолированного на стадии (с) при отсутствии избирательного фактора;
(е) отбора на Vibrio cholerae, не экспрессирующего больше такого селектированного маркера; и, как следствие этого, имеющего область хромосомальной ДНК, с делецией генов холерного энтеротоксина и энтеротоксина зоны плотного контакта (zonula occludens).
(f) конструирования второй плазмиды, содержащей ген устойчивости к ртути и ДНК-последовательность, кодирующую B-субъединицу энтеротоксина Vibrio cholerae, и ген второго селектируемого чужеродного маркера, при этом упомянутая плазмида неспособна к репликации в Vibrio cholerae вне хромосомы, а последовательности, имеющую достаточную длину для промотирования распознаваемый in vivo рекомбинации, ограничивают область упомянутого гена устойчивости к ртути и ДНК-последовательность, кодирующую B-субъединицу энтеротоксина Vibrio cholerae;
(g) скрещивания микроорганизма, несущего упомянутую вторую плазмиду с упомянутым Vibrio cholerae, раскрываемым в стадии (е), содержащую последовательности, гомологичные к указанным последовательностям достаточной длины для промотирования распознаваемый in vivo рекомбинации;
(h) отбора Vibrio cholerae, экспрессирующего упомянутый второй селектируемый маркер;
(i) выращивания продукта, отобранного на стадии (h), при отсутствии второго селектированного фактора;
(j) отбора Vibrio cholerae, не экспрессирующего больше второго селектированного маркера; и
(k) скрининга указанного Vibrio cholerae, раскрываемого на стадии (j) на Vibrio cholerae, которые содержат ген устойчивости к ртути и ДНК-последовательность, кодирующую B-субъединицу холерного энтеротоксина и имеют область в хромосомальной ДНК с делецией гена холерного энтеротоксина и энтеротоксина зоны плотного контакта (zonula occludens).
Этот метод выделения делеционных мутантов Vibrio cholerae, имеющих область хромосомальной ДНК последовательностей, кодирующих холерный энтеротоксин, и делецией гена зоны плотного контакта, и вставкой гена устойчивости к ртути и ДНК-последовательности, кодирующей B-субъединицу Vibrio cholerae, можно использовать в стадии (f), раскрывающую фланкирующие последовательности достаточной длины, содержащие ген, который может быть удален, не оказывая при этом влияния на колонизирующую способность и иммуногенность Vibrio cholerae. В качестве примера можно привести гемолизиновый ген. Штамм CVD110 V. cholerae, сконструированный в соответствии с предлагаемым методом, имеют область хромосомального ДНК с последовательностями, кодирующими A и B-субъединицы холерного энтеротоксина и делецией энтеротоксина зоны плотного контакта (zonula occludens), и имеют также ген устойчивости к ртути и ДНК- последовательность, кодирующую B-субъединицу энтеротоксина Vibrio cholerae, вставленную в сайте гемолизинового гена. К другим примерам последовательностей достаточной длины относятся his-ген (Hone, Microbial Pathogenesis 5, 35 pp. 407-478 (1989)) и nanH-ген (Vimr, J. Bacter., 170, pp. 1495-1504 (1988)).
Заявляемый штамм CVD112, имеет область с делецией всего ядра холерного энтеротоксина, которая включает гены сер, асе, zot, orfU, и ctxAB, а также делецию RSI-элементов и attRS1-сайтов [Pearson и другие, Proc. Natl. Acad. Sci. (США), 90, pp. 3750-3754, 1993] вместе с локусом в хромосоме ctx-гена. Кроме того, этот штамм имеет последовательности, кодирующие достаточную часть B-субъединицы холерного энтеротоксина для обеспечения иммуногенности, повторно вставленные в хромосому вибриона. Вставки, дополнительно введенные в хромосому представляют собой последовательности ДНК, вызывающие устойчивость к тяжелым металлам, например ртути.
Другой штамм V. cholerae не-01 серогруппы предлагаемого изобретения, а именно CVD112 RM, имеет идентифицирующие характеристики вышеуказанного штамма не -01 серогруппы, и кроме того имеет делецию в recA-локусе Vibrio cholerae, в результате чего полученный штамм утрачивает ген в гомологичной рекомбинации.
В примерах, которые иллюстрируют далее настоящее изобретение, любые из методов, реакций, и способов выделения хорошо известны специалистам в данной области. Все ферменты, если не оговорено иначе, являются доступными из одного или нескольких коммерческих источников, например. New England BioLabs - Beverly, Штат Массачусетс; Collaborative Research - Waltham, Штат Массачусетс; Miles Laboratories -Elkhart, Штат Индиана; Boehringer Biochemicals Inc., -Индианаполис, Штат Индиана; и Bethesda Research Laboratory - Rockville, Штат Мэриленд, которые указаны здесь в качестве нескольких представителей. Буферные растворы и реакционные условия для гидролиза рестриктазами используют согласно рекомендациям, прилагаемым изготовителем для каждого фермента, если не оговорено особо. Частичный гидролиз рестрикционными ферментами осуществляют с использованием более низкой концентрации фермента, которая должна быть определена заранее из пробных экспериментов по каждой партии ферментов. Стандартная методика проведения других ферментативных реакций, методы разделения гель-электрофорезом и получение трансформантов E. Coli можно найти в Methods in Enzymology, vol.68, под редакцией Ray Wu, Academic Press (1979). Можно также обратиться к другому общеизвестному источнику - Maniatis, Т. и другие. Molecular Cloning, Cold Spring Harbor (1982). Бактериальные клетки выращивали согласно методикам, описанным Miller, в Experiments_ on Molecular Genetics, Cold Spring Harbor Laboratory (1972). Вибрионы холеры размножали согласно общим методикам, описанным Lennett, E.A. и другими, eds. Manual of Clinical Microbiology, 3-е издание, Американское Общество по Микробиологии, Вашингтон в (1980). E. Coli и V. cholerae скрещивали согласно общей методике, описанной Johnson, Steven R. и другими. J. Bact., 137, 531 (1979); и Yokata, Т. и другие, J. Bact., 109, 440 (1972).
Штаммы настоящего изобретения были депонированы в Американской Коллекции Типовых Культур, например, расположенные в Rockville, Штат Мэриленд, до подачи настоящей заявки. Депонированы штаммы V. cholerae JBK56, V. cholerae JBK70, V. cholerae N1696, V. cholerae JVK70 (pJBK51), V. cholerae 395 серотипа Огава, штаммы CVD101, CVD109 V. cholerae E7946, и E. Coli SM10 лямбда pir pCVD51, V. cholerae CVD110, и E. Coli SY327 лямбда pir (pCVD622.2B), которые имеют номер доступа АТСС 39,317, 39,318, 39,315, 39,316, 39,541, 39,540, 55,057 (депонированный 4-ого июня, 1990), 55,056 (депонированный 4-ого июня, 1990), 68,335 (депонированный 5-ого июня,1990), 55188 (депонированный 3-его июня, 1991), и 68630 (депонированный 3-его июня, 1991), соответственно.
Пример 1
Конструирование плазмиды с селектируемым маркерным геном, вставленным для замены холерогенов.
Плазмида JBK16 содержит a PstI-BgIII фрагмент длиной 4 т.п. оснований из хромосомы, содержащей гены энтеротоксина. Гены энтеротоксина рестриктированы по AccI-сайтам и содержат внутренний AccI-сайт. JBK16 была гидролизована полностью рестриктазами AccI и AccI, и полученные фрагменты, содержащие гены энтеротоксина отделяли от оставшейся плазмиды.
Перекрытие остатков плазмиды или липкие концы участка AccI превращали в тупые концы, заполняя брешь фрагментом Кленова, полученного из полимеразы E. Coli polymerase (то есть, одноцепочечную ДНК, остающуюся после гидролиза AccI, получали двуцепочечную с короткими концами). Ген, кодирующий устойчивость к ампициллину очищали от плазмиды pREG153 (pREG153 - производная плазмиды pREG151 [Weiss, A. и другие. J. Bact., 152, 549-552], модифицированной заменой гена устойчивости к триметоприну геном устойчивости к ампициллину и присоединение cos-последовательностей) и липкие концы заполняли брешью, как описано выше. Полученный фрагмент затем лигировали с ДНК вибриона в результате чего гены устойчивости к AP были расположены точно в том же самом месте, в котором находились делетированные гены энтеротоксина, фланкированные теми же самыми ДНК-последовательностями вибриона холеры. Полученная плазмида была сконструирована как pJBK21 (фиг. 4), содержащая область с делецией холерного энтеротоксина и гена устойчивости к ампициллину.
Пример 2
Введение фланкирующих гомологичных последовательностей, сопровождаемое конъюгативным переносом гена в V. cholerae.
Для обеспечения специфической вставки в хромосому из делетированной последовательности в pJBK21, фрагмент ДНК длиной приблизительно 7000 пар оснований был лигирован с каждым концом фрагмента PstI-Bg1II из плазмиды pJBK21. (Вероятность возникновения события гомологичной рекомбинации возрастает с увеличением длины фланкирующих гомологичных последовательностей). Для достижения этого, фрагмент приблизительно 18 т.п.о клонировали из хромосомы штамма N 16961. Этот клон, обозначенный как pJBK44 содержит фрагмент PstI-Bg1II-гена энтеротоксина длиной 4 т.п.о, ограниченный фланкирующей ДНК-последовательностью длиной примерно 7 т.п.о. по обоим концам (см. фиг. 5). Плазмида pJBK21 была частично гидролизована по PstI-сайту, в результате чего только лишь один из Pst-сайтов был вырезан (дополнительный Pst-сайт был вставлен в пределах локуса гена устойчивости к ампициллину) после гидролиза рестриктазой Bg1II-сайта с изоляцией Pstl-Bg1II- фрагмента длиной 4 т.п.о, содержащего область с делецией гена энтеротоксина и область устойчивости к Ap.
Плазмида pJBK44, содержащая фрагмент ДНК вибриона длиной приблизительно 18 т. п. о была частично гидролизована рестриктазой по сайту Bg1II, в результате чего только лишь один из 4 Bg1II- сайтов был вырезан. После этого частичного гидролиза полный гидролиз осуществляли рестриктазой PstI-сайта и полученные фрагменты отделяли методом электрофореза через 0.3% агарозном геле. Изолированные фрагменты затем очищали и анализировали, и при этом только лишь один фрагмент, как показал анализ, содержал полную последовательность pJBK44 за исключением фрагмента PstI- Bg1II-гена холерного энтеротоксина длиной 4 т. п.о. (см. фиг. 5.). Этот фрагмент, представляющий фланкирующую ДНК-последовательность, затем смешивали с лигированием с Pstl-Bg1II фрагментом из плазмиды pJBK21, содержащей ампициллиноустойчивый ген. Полученная плазмида, PJBK54, содержала хромосомальную ДНК-последовательность Vibrio длиной приблизительно 17 т.п.о с ампициллиноустойчивым геном, замененным на делетированные гены энтеротоксина. Модифицированную область в хромосомальной ДНК затем клонировали в плазмиду, которая может легко активизироваться в V. cholerae. Плазмида pRK290 [Ditta, G. И другие. Proc. Nat. Acad. Sci. 77, 7347 (1980)] относится к плазмидам группы несовместимости P и имеет всего лишь единственный EcoRI-сайт, в который клонировали плазмиду pJBK54(Фиг. 6). Полученную плазмиду pJBK55 затем скрещивали с штаммом V. cholerae N16961 с использованием конъюгативной плазмиды pRK2013. В результате получили V. cholerae N 16961 (pJBK55) (Apr-ампициллиноустойчивый фенотип).
Пример 3
Рекомбинация in vivo
Мутантные холерогены, после конъюгативного генного переноса, как описано в Примере 2, присутствовали теперь во внехромосомальном состоянии в штамме V. cholerae N 16961 (см. фиг. 1). При очень низкой частоте повторяемости (возможно от 10-6 до 10-8), гомологичные фланкирующие ДНК-последовательности образуют конъюгационную пару в хромосоме (см. фиг. 7). Этот редкий результат приводит к замене области с делетированным энтеротоксином на плазмиде на ctx-гены на хромосоме. Для отбора этого редкого результата, использовали феномен плазмидной несовместимости[Ruvkin, G.B., см.ранее]. Плазмиды можно разделить на группы несовместимости от A до W, на основе их способности удерживаться в устойчивом состоянии вместе в одной и той же клетке. Если две плазмиды не могут стабильно существовать вместе в той же самой клетке, они несовместимы и принадлежат к одной и той же группе несовместимости возможно потому, что они используют одинаковый механизм репликации в клетке. При избирательном использовании фактора антибиотикоустойчивости, присутствующего на одной плазмиде, а не на другой, можно отобрать тот тип из этих двух несовместимых плазмид, который будет совместимым. Плазмида pJBK55, вследствие того, что происходит из плазмиды pRK290, также принадлежит к группе P (Inc). R702 также принадлежит к группе P (Inc) и содержит маркер устойчивости к канамицину, тетрациклину, сульфонамиду, и стрептомицину, но не к ампициллину. При скрещивании pR702 (SuR) в штамме N16961 (pJBK55) (Aрr) и отборе на среде содержащей как ампициллин, так и сульфонамид, отобрали клетки, в которых маркер ампициллиноустойчивости вставляли в хромосому, а маркер устойчивости к сульфонамиду оставался на плазмиде R702, поскольку pR702 и pJBK55 несовместимы (см. фиг. 2). Полученный штамм JBK56 (фиг. 3) содержал маркер ампициллиноустойчивости, и энтеротоксин, обнаруживаемый отрицательную реакцию в анализе с использованием адреночувствительных Y-1 типа клеток и с помощью Gm1 в твердофазном иммуноферментном анализе (ELISA). Кроме того, когда хромосомальную ДНК гибридизировали с ДНК-зондами, содержащими клон генов холерного энтеротоксина, JBK56 обнаруживал отрицательную реакцию, что подтверждает предположение о полной делеции генов энтеротоксина.
Фактор антибиотикоустойчивости, кодируемый на R702 был элиминирован (удален) путем отбора самопроизвольно исправленной копии плазмиды, утраченной в указанной плазмиде (это событие возникало при частоте приблизительно 1 на 2000).
Пример 4
Элиминация селектируемого маркера из примера 1
Для элиминации фактора ампициллиноустойчивости, конструировали плазмиду, производную из плазмиды pJBK55, в которой гены, кодирующие устойчивость к ртути (Hgr) из R100 были клонированы в PstI-сайте гена Ap, таким образом инактивируя инсерцией фактора ампициллиноустойчивости. Эту производную плазмиду затем скрещивали в V. cholerae JBK56, в присутствии pR702 и осуществляли отбор, как описано ранее для маркера Hgr, Aps V. cholerae. Конечный продукт, штамм V. cholerae JBK70, был чувствителен ко всем проверенным антибиотикам, устойчив к ртути, и по фенотипу токсин-отрицательный. В его хромосомальной ДНК не отмечено обнаруживаемой способности к гибридизации с ДНК-зондами, содержащими гены холерного энтеротоксина. Вкратце о секвенировании ДНК полной хромосомы, JBK70, по-видимому не имеет значимых отличий от родительского штамма N16961, за исключением делеции генов энтеротоксина и вставки гена ртутной устойчивости и инактировании ампициллиноустойчивого гена. Такой штамм не реверсирует в токсикогенное состояние, потому что гены энтеротоксина претерпели не просто мутации, а были полностью утрачены (делетированы).
Пример 5
Конъюгативный перенос гена для обеспечения антитоксического иммунитета.
В случае когда, как антибактериальный иммунитет, так и антитоксический иммунитет необходимы для синергизма, из плазмиды JBK70 можно сконструировать новую плазмиду для продуцирования только лишь B-субъединицы холерного энтеротоксина. С этой целью, получали генный продукт энтеротоксина, продуцирующий только лишь B-субъединицу и в котором утрачены гены субъединицы A (фиг. 8). HpaII-фрагмент, вырезанный из плазмиды pJBK16, содержащий B структурный ген, клонировали в фаговый клонирующий вектор, М13mp7, вставленный между сайтом BamHI и EcoRI на любой стороне гена (фиг. 8). Фрагмент, фланкированный вновь BamHI-сайтами клонировали в плазмиду pMS 9, которая содержала очень активный trp-промотор. Размещение B-субъединичных генов в области регуляции транскрипции мощного промотора позволяет обеспечить высокую продукцию B-антигена. Из исследованных клонов, приблизительно 50% не синтезировало никакого антигена. Этот установленный факт отражает две возможных ориентации клонированной генной инсерции - одна транскрипционно-активная, другая - обратная. Одну плазмиду, производную из pJBK51, которая продуцировала B- субъединицу, скрещивали в штамме Vibrio cholerae JBK70, и как было обнаружено, она вырабатывала значительно большее количество B- антигена, чем родительский штамм N 16961, что привело к получению мутанта JBK70 (pJBK51). Другие только B-секретирующие мутанты получали с использованием различных промоторов, включая P1- промотор, и оценку любой существенной разницы экспресии их in vivo можно провести на соответствующих моделях.
Пример 6
Колонизация тонкого кишечника новорожденной мыши с использованием штамма JBK70 без реверсии к токсикогенности.
Мыши-сосунки (весом 2,0-3,5 г) отнимались от матерей и держались натощак в течение примерно от 3 до 8 часов. Затем четырем из них вводили инокулят на 1 день per os (через рот) в живот с помощью иглы 22-граммового микрошприца. Инокулят содержал примерно 108 CFU (колониеобразующие единицы/мышь) штамма JBK70 в объеме от 0,05 мл до 0,1 мл. Инокулят приготавливали в бульоне с сердечно-мозговой вытяжкой практически по методике, описанной Baselski, V. и другими, см. ранее. Инокулят содержал также примерно 0,01% Эванса синего. Наличие этого красителя в животе животного, как можно было видеть через стенки брюшной полости, свидетельствовало о надлежащей доставке инокулята. Введение Эванса синего прервали через день для того, чтобы исключить ингибирование активности JBK70.
Последующие инокуляции проводили методом "мышь-в-мышь"(МХМ), или в альтернативном варианте, "мышь-на-планшет-в-мышь" (МХРХМ), однако необходимы были различные процедуры для приготовления инокулята, по сравнению со схемой прививки Baselski на 1 день.
Для приготовления МХМ-инокулята, слепую кишку рассекали от живота до заднего прохода при соблюдении стерильности. Образец взвешивали, помещали в стакан с гомогенизатором, и прибавляли приблизительно 0,5 мл бульона с сердечно-мозговой вытяжкой. Полученную смесь быстро растирали до гомогенного состояния пестиком из тефлона до полного лизиса ткани. Полученную клеточную суспензию использовали для прививки приблизительно 108 CFU в каждую новорожденную мышь. Проверку на чистоту проверяли методом штриховой разводки на MEA-агаровых чашках (с мясным экстрактом), при этом не добавляли красителя Эванса синего.
Для приготовления МХРХМ-инокулята, использовали стерильную петлю для переноса клеток из MEA-агаровой чашки в бульон с сердечно-мозговой вытяжкой. Приблизительно 1011 CFU/мл добавляли к 1 мл BHI-бульона для получения густой клеточной суспензии. Смесь доводили до гомогенного состояния, и дозу 0,05-0,1 мл (приблизительно 1010 CFU) прививали per os в каждую новорожденную мышь. Краситель Эванс синий добавляли.
Для проведения всех серий прививок мышей держали в лабораторных стаканах при комнатной температуре 73-76oF (по Фаренгейту). Стаканы помещали в пластмассовую коробку, которая была неплотно закрыта для поддержания температуры немного выше комнатной, 78oF.
Как видно из результатов, в кишечнике присутствовало достаточное количество инфицированных клеток для прививки следующего животного, при исследовании клеток методом штрихования на MEA-чашках. Таким образом, штамм Vibrio cholerae JBK70 обладает способностью к колонизации тонкой кишки новорожденных мышей. Кроме того, накопление жидкого содержимого (FA) не возрастало, поскольку не отмечено существенного увеличения показателя FA (показатель FA больше или равен 0,065 - накопление серопозитивной жидкости).
Пример 7
Конструирование штамма V. cholerae CVD 101 с делецией фрагмента рестрикцией в локусе гена кодирующего A-субъединицу.
Другой классический штамм, выбранный для аттенуации, относился к штамму 395 Vibrio cholerae серотипа Огава (альтернативно обозначенный "395") который, подобно штамму N 16961, был апробирован в широкомасштабных экспериментах с участием добровольцев, и который, как установлено, вызывает устойчивый защитный иммунитет [Levine, М.М.Immunity to cholera as evaluated in volunteers, " в работе Cholera and Related Diarrheas: 43rd Nobel Symposium, Stockholm, 1978. (O. Ouchteriong & J. Holmgren, eds.) Basel: S. Karger, pp. 195-2-3 (1980); Levine, M.M и другие. Acute Enteric, см.ранее (1981)]. Используемая для аттенуации штамма 395 методика не отличалась существенно от используемой для ослабления штамма N 16961 (как описывалось ранее в Примерах 1-5). На первом этапе проводили клонирование и картирование двух копий холерного энтеротоксина штамма 395. В результате блот-гибридизации по методу Саузерна было обнаружено 2 фрагмента HindIII длиной примерно 16 и 12 п.О., каждый из которых гибридизовался с клонированными генами холерного энтеротоксина. Указанные фрагменты очищали методом элекрофореза на агарозном геле и затем клонировали в гидролизованную плазмиду pBR325 рестриктазой HindIII, обработанную щелочной фосфатазой (фиг.9). Полученные рекомбинантные плазмиды, содержащие гены энтеротоксина, обозначили как pCVD14 и pCVD15.
Плазмиды pCVD14 и pCVD15 затем картировали эндонуклеазами рестрикции. В результате картирования обнаружили фрагмент XbaI-ClaI длиной примерно 550 п. о., содержащий полную последовательность оснований субъединицы A1 за исключением кодонов первых 10 аминокислотных остатков A1. Этот XbaI-ClaI-фрагмент был вырезан in vitro из обоих плазмид pCVD14 и pCVD15 серией манипуляций, показанных на фиг. 10 на примере плазмиды pCVD15. В результате первого частичного гидролиза ClaI получили популяцию линейных ДНК-молекул, в которых был вырезан только лишь один из 5 ClaI-сайтов. В следующей стадии концы линейных молекул заполняли брешью с использованием ДНК-полимеразы для получения тупых концов. XbaI-линкеры сшивали по тупым концам ClaI-сайтов с выходом популяции молекул, к которым прибавляли XbaI-рестриктазу для удаления линкера. Затем вводили ген устойчивости к тетрациклину на XbaI-фрагменте и лигировали. После переноса генов в штамм К-12 E. Coli и отбора на устойчивость к тетрациклину, исследовали содержимое плазмиды для определения количества полученных трансформантов. Было обнаружено большое множество делеционных мутаций, которые в одном или нескольких XbaI - ClaI фрагментах содержали делеции. Был отобран один делеционный мутант, у которого отсутствовал только лишь фрагмент ClaI- XbaI, длиной 550 п.о., содержащий ген A1. Этот делеционный мутант, обозначенный как pCVD25, очищали, гидролизовали рестриктазой XbaI и повторно лигировали для удаления гена устойчивости к тетрациклину. Полученный клон, pcVD 30. обнаруживал отрицательную реакцию на голотоксин, как измерено в анализе с использованием адреночувствительных Y-1 типа клеток [Sack D.A. и другие, см. ранее, (1975)], но положительную на продукцию B-субъединицы, измеренной методом твердофазного иммуноферментного анализа ELISA [Sack D.A. и другие, см. ранее (1980)], и в котором были утрачены гены устойчивости к A1, о чем свидетельствовали результаты ДНК- гибридизации с использованием меченого A1-зонда. Фрагмент HindIII, содержащий мутацию с делеций энтеротоксина, затем клонировали в pJBK85, Tc-чувствительной, Cm-устойчивой плазмиды, производной из pJBK108. Полученную плазмиду обозначили как pJBK108.
Отсутствие селектируемого маркера в делеционной мутации энтеротоксина в PJBK108 потребовало модификации метода, используемого ранее для аттенуации штамма N16961 Эль Тор. Для выполнения элиминации Ai-генов из штамма 395, HindIII-фрагмент, вырезанный из плазмиды pCVD15 клонировали в PJBK85, в результате чего получили плазмиду pJBK88 (фиг. 11). Ген устойчивости к тетрациклину на XbaI-фрагменте затем клонировали в XbaI-сайт в области A1-гена плазмиды PJBK88 с получением производной плазмиды pJBK107. Этот маркерный ген устойчивости к тетрациклину затем рекомбинировали в хромосому штамма 395, по методике, используемой ранее для получения pJBK56 V. cholerae. PJBK107 (Tcr, Cmr) переносили в штамм 395 и затем вводили вторую Inc P-плазмиду, pR751 (Tpr). В результате отбора колоний по Tcr, Tpr, Cmr получили штамм JBKI 13 V.cholerae, который содержал тетрациклиноустойчивые гены в обеих хромосомальных копиях энтеротоксина. PJBK 108, содержащую делеционную мутацию, затем вводили в V. cholerae JBK 113. Гомологичная рекомбинация делеционной мутации в хромосоме приводит к потере последовательностей A1-гена, результат, обнаруживаемый по утрате фактора устойчивости к тетрациклину. Поскольку рекомбинация происходит даже при очень низкой частоте повторяемости, использовали процедуру обогащения тетрациклиночувствительных клеток в популяции клеток, устойчивых к тетрациклину. B этой методике обогащения принимали за основу тот факт, что тетрациклин относится к бактериостатическим антибиотикам, в то время как ампициллин и Д-циклосерин относятся к бактерицидам. В соответствии с этим, культуру штамма V. cholerae JBK113, содержащего плазмиду pJBK108 инкубировали в течение 3 часов при температуре 37oC в 1 л бульона, содержащего 2 мкг/мл тетрациклина, 50 мкг/мл ампициллина и 50 мкг/мл Д-циклосерина. К 3 часу, большая часть тетрациклиноустойчивых клеток была убита, а тетрациклиночувствительные клетки выявляли путем посева культуры на L-агар и получением реплики на L-агаре с тетрациклином. Тетрациклиночувствительные колонии затем проверяли на присутствие A1-генов методом ДНК-гибридизации. Один чувствительный к тетрациклину штамм с делецией обеих копий гена A1-субъединицы обозначили как V. cholerae CVD101 и исследовали на активность продуцирования B-субъединицы методом твердофазного иммуноферментного анализа ELISA [Sack, см. ранее]. V. cholerae CVD101, как было установлено, достоверно продуцирует B-субъединицу энтеротоксина на уровнях эквивалентных родительскому вирулентному штамму 395 V. cholerae.
Пример 8
Последовательность ДНК генов энтеротоксина.
Была определена полная ДНК последовательность генов энтеротоксина V. cholerae Инаба 62746, часть из которых сообщалась Lockman, et al. J. Biol. Chem. 258, 13722 (1983). Картирование плазмид pCVD14 и pCVD15 рестрикцией эндонуклеазой показало, что последовательности, обнаруженные в штамме 62746, также присутствуют в генах энтеротоксина штамма 395. Предсказанное скрещивание после делеции фрагмента Xba1-Cla1, длиной 550 п.о., но с введением в последовательность Xba1-линкера, показано на фиг. 12. Xba1-сайт последовательности холерного энтеротоксина охватывает 10 и 11 аминокислотных остатков структурного гена A1 (не считая 18 аминокислот главной последовательности A1).
Пример 9
Конструирование Штамма V.cholerae, содержащего делецию области плотного контакта (zonal occludens) энтеротоксина в CVD101.
Мутант с делецией zot-гена V. cholerae получали аналогичным методом, который описан ранее в примере 7 для штамма CVD101. Zot-ген содержится в рекомбинантной плазмиде pBB68. PBB68 состоит из фрагмента хромосомальной ДНК по EcoRI-PbrI-сайту, вырезанного V. cholerae 569B, который содержит zot-ген и ctx- гены, имеющие делецию XbaI-ClaI-фрагмента длиной 550 п.о. StuI- AccI-фрагмент длиной 575 пар оснований вырезают in vitro из плазмиды pBB68 при гидролизе рестриктазами StuI и AccI-сайта с получением молекул с тупыми концами путем заполнения бреши ДНК- полимеразой. (В результате происходит делеция 48% из 1199 пар оснований zot-гена). Одну половину образца лигировали с геном устойчивости к тетрациклину (чужеродного происхождения), в результате чего был получен селектируемый маркер.
Делеционный мутант zot-гена, сконструированный in vitro, как описано выше, вставляли в хромосому V. cholerae CVD101 по методике, используемой для конструирования делеционного мутанта ctx в штамме CVD101. Тетрациклиноустойчивый клон, полученный выше, клонировали в Inc P-плазмиду pJBK85. Эту плазмиду (Tcr, Cmr) переносили в штамм CVD101 с последующим отбором генотипа Tcr. Затем вводили вторую Inc P-плазмиду, pR751 (Tpr). В результате отбор колонии на Tcr, Tpr, Cmr получили штаммы V. cholerae, где произошла перекрестная рекомбинация между Tcr геном и zot-геном.
Плазмиду, содержащую делеционный мутант StuI-AccI без Tcr гена, затем переносили в штамм V. cholerae с геном Tr. Гомологичная рекомбинация делеционного мутанта в хромосому привела к потере последовательностей гена zot, результат обнаруживаемый по отсутствию колоний с Tcr. Tcr-колонии отбирали и скринировали на потерю zot-последовательностей методом ДНК-гибридизации с использованием StuI-AccI-фрагмента в качестве зонда.
Пример 10
Конструирование Штамма CVD109 V. cholerae с делетированными последовательностями, кодирующими энтеротоксины V. cholerae и энтеротоксин зоны плотного контакта Zonula Occludens.
Конструирование ослабленного V. cholerae CVD109 (АТСС # 55057) включает манипуляцию введения клонированной ДНК- последовательностей Vibrio вместе с последовательностями, кодирующими селектируемый маркер в хромосому вирулентного штамма V. cholerae. Первоначальное событие рекомбинации in vivo гомологичных последовательностей из рекомбинантной плазмиды в хромосому обеспечивает селектируемый маркер в этом сайте.
Второе событие in vivo рекомбинации между гомологичными фланкирующими последовательностями ДНК приводит к вырезанию обученных генов из хромосомы с получением конечного продукта, который является делеционным мутантом. На фиг. 16 показана схема конструирования CVD109. Zot и ctx гены расположены рядом друг с другом на хромосоме V. cholerae. Многократные копии последовательностей ДНК длиной 2700 п.о., названные как RS1-элементы (в виде повторяющейся последовательности) содержат на обоих концах гены zot и ctx в вирулентном штамме V. cholerae E7946 (Эль Тор биотип, серотип Огава). На фиг. 16A, последовательности в локусах zot и ctx генов показаны большой незаштрихованной или заштрихованной стрелкой. RS1-последовательности показаны маленькой, сплошной стрелкой. Рекомбинантная плазмида pCVD51 (фиг. 16A), содержит клонированные zot/ctx последовательности (незаштрихованная стрелка), которые гомологичны хромосомальным zot/ctx последовательностям (показанным на фиг. 16A заштрихованной стрелкой) и содержат селектируемый маркер, ампициллиноустойчивый ген (Apr). Вектор плазмиды, в который клонировали Vibrio ДНК-последовательности является плазмидой pGP704 (Miller и Mekalanos J. Bact. 170, 2575- 2583 (1988)). Эта плазмида не способна к репликации во внехромосомальном состоянии в V. cholerae, но может реплицироваться совместно с совместимыми штаммами E. Coli. PCVD51 скрещивали с микроорганизмом из E. Coli с переносом в V. cholerae E7946. Поскольку эта плазмида не способна к репликации во внехромосомальном состоянии в V. cholerae, в результате отбора колоний с ампициллиноустойчивыми клетками получили штаммы, в которых плазмида, несущая Apr маркер, гомологично рекомбинирована в хромосому в сайте zot/ctx-последовательностей. (Точный участок рекомбинации, является ли он локусом гена zot или ctx-гена, неизвестен). В результате наступления события только одной перекрестной рекомбинации, (а не двойной перекрестной рекомбинации) получают структуру так называемую совместного объединения, которая показана на фиг. 16B.
RS1 - последовательности, фланкируюшие область генов zot и ctx имеют достаточную длину для обеспечения распознаваемой in vivo рекомбинации; внутримолекулярная рекомбинация между гомологичными RS1-элементами приводит к потере всех последовательностей между ними. Штамм V. cholerae с встроенной плазмидой, содержащий маркерный ген Apr выращивали при отсутствии ампициллина и затем отбирали колонии с фактором ампициллиноустойчивости Aps. Рекомбинация RS1-элементов, фланкирующая ctx и zot привела к потере вставочных последовательностей zot и ctx наряду с вектором плазмиды, содержащей Apr (фиг. 16C).
Колонии V.cholerae с фактором Aps, выращенные на вышеуказанных стадиях, скринировали методом ДНК-гибридизации на zot-последовательности. ДНК-зонд содержал фрагмент с сайтом рестрикции Stul-AccI длиной 575 п.о., образованный из клонированного zot-гена. Отбирали колонии, которые не гибридизовались с указанным ДНК-зондом, и затем исследовали на присутствие ctx-генов методом ДНК-гибридизации с использованием зонда с ctx-геном. Результаты гибридизации подтверждали потерю локусов генов zot и ctx. Один типичный представитель штамма был спасен и обозначен как штамм CVD109. На фиг. 160 изображена хромосома клеток линии CVD109, в которой делетированы zot и ctx-последовательности, но сохранена одна копия RS1-элемента. (Плазмида, показанная на фиг. 160, утрачена в конечном штамме, содержащем Aps, но изображена только в качестве иллюстрации второго события перекрестной рекомбинации. Этот промежуточный продукт потерян самопроизвольно, поскольку эта плазмида не способна к репликации во внехромосомальном состоянии в V. cholerae).
Пример 11
Конструирование Штамма CVD110-а V. cholerae с делециями последовательностей, кодирующих A и B субъединицы энтеротоксина V. cholerae и энтеротоксина зоны плотного контакта (zonula occludens), и вставка гена устойчивости к ртути и ДНК-последовательности, кодирующей B-субъединицы энтеротоксина V. cholerae.
CVD110 (АТСС # 55188) конструировали непосредственно из V. cholerae CVD109, который были описан ранее. У штамма V. cholerae CVD109 делетированы как A-, так и B-субъединица холерного энтеротоксина (CT), а также ген zot и ген устойчивости к ртути. Фрагмент гена, содержащий маркерный ген B-субъединицы холерного энтеротоксина (ctxB) и маркер устойчивости к ртути конструировали in vitro. Эту конструкцию затем вставляли в хромосому штамма CVD109, причем целенаправленно в гемолизиновый ген. Таким образом, конечный вакцинный штамм CVD110, вызывает продукцию B-, но не A-субъединицы холерного энтеротоксина, устойчив к ртути и не продуцирует тип HlyA-белка (гемолизин).
Конструирование B-субъединицы холерного энтеротоксина: ctxB и ctx промоторные фрагменты получали из плазмиды pCVD30, которая описана в примере 7. Эта плазмида pCVD30 содержит делецию ctxA-гена. Фрагмент длиной 1.4 т.п.о, содержащий ctxB-ген и ctx-промотор, но утрачен zot-ген, была получена гидролизом pCVD30 с рестриктазой HinP1 и HaeIII. Полученный фрагмент обрабатывали с T4 ДНК-полимеразой для получения его с тупыми концами и синтетические KpnI линкерные последовательности сшивали с указанным фрагментом. Затем фрагмент клонировали в вектор плазмиды pCVD315[Galen, и другие. Advances in Researchon Cholera and Related Diarrheas, vol.7(Sack и другие, eds.) стр. l43-153 (1990)]. Вектор плазмиды pCVD315 не имеет никаких значимых различий от используемых для этой цели векторов кроме того, что содержит KpnI-сайт. Полученную векторную плазмиду, содержащую ctxB-ген обозначили как pCVD621 (фиг. 18).
Гены устойчивости к ртути: в качестве гена устойчивости ртути (mer) использовали маркер, аналогичный при конструировании V.cholerae JBK70. NcoI-StuI фрагмент длиной 4,2 т.п.о, содержащий ген mer был получен из плазмиды pDB7[Barrineau и другие, J. Molecular and Applied Genetics (1984) vol. 2, стр. 601, 619]. Фрагмент обрабатывали ДНК-полимеразой (фрагмент Кленова) для получения его с тупыми концами и синтетические KpnI линкерные последовательности сшивали с указанным фрагментом. Затем фрагмент клонировали в вектор плазмиды pCVD43.2 (источник неизвестен), которая является производным плазмиды pCVD43 [Kaper и другие. Advances in Researchon Cholera and Related Diarrheas, vol. 6 (Ohtomo и другие, eds.) стр. 161-167 (1988)]. PCVD43 содержит клонированные гемолизиновые гены (hlyA) V.cholerae с делецией HpaI-фрагмента длиной 400 п.о. внутри локуса hlyA. Делеция HpaI- фрагмента длиной 400 п. о. приводит к инактивации указанного [Kaper и другие. Advance, см. ранее, vol. 6] . Плазмида pCVD43.2 идентична pCVD43 за исключением того, что синтетический KpnI-линкер лигирован к только лишь одному HpaI-сайту плазмиды pCVD43. Объединенный клон mer-генов, вставленных в локус hylA гена обозначили как pCVD43.3.
Вставка ctxB и mer генов в CVD109: для переноса указанных генов в хромосому CVD109 использовали вектор плазмиды pGP704, который описан в примере 10. ClaI-Bg1II-фрагмент из плазмиды pCVD43.3, длиной 8.1 п.о, содержащий mer и hylA-гены клонировали в плазмиду pGP704 с получением плазмиды pJMK12 (фиг. 18). pJMK12 была частично гидролизована рестриктазой KpnI с выходом популяции линейных молекул, в которых вырезан только один из 3 KpnI-сайтов. Фрагмент плазмиды pCVD621, длиной 1,4 п.о. (описанный выше), содержащий ctxB-ген, затем лигировали с плазмидой pJMK12 с получением плазмиды pCVD622.2B. Относительное положение и ориентация вставленных генов показаны на фиг.18.
Плазмиду PCVD622.2B затем переносили в V.cholerae CVD109 путем конъюгации из клеток-хозяина E.coli. Как уже указывалось в примере 10, pGP704 не способна к репликации во внехромосомальном состоянии в V.cholerae, но способна к репликации в дозволяющих клетках E.Coli. Поскольку pCVD622.2B не способна к репликации во внехромосомальном состоянии в V. cholerae, в результате отбора колоний с ампициллиноустойчивыми клетками [pGP 704 содержит ген, кодирующий устойчивость к ампициллину] получили штаммы, в которых плазмида pCVD622.2B, несущая Apr маркер, гомологично рекомбинирована в хромосому в сайте hylA-гена. [Она не способна к рекомбинации между ctx или zot-последовательностями ввиду утраты этих генов]. В результате события единичной перекрестной рекомбинации, (не двойной перекрестной рекомбинации) получают так называемую совместно интегрированную структуру или "меродиплоид" (эти термины используют попеременно), которая показана на фиг. 19B.
Второе событие перекрестной рекомбинации может иметь место между гомологичными hylA-последовательностями, фланкирующими объединенными вместе плазмидами pCVD-622.2B. Это второе событие перекрестной рекомбинации происходит самопроизвольно и обнаруживается путем отбора колоний, у которых потерян фенотип Apr. Это второе событие перекрестной рекомбинации может иметь один из двух возможных исходов, в зависимости от точного сайта рекомбинации. Оба возможных результата приводят к потере последовательностей плазмидного вектора pGP704, о чем свидетельствуют фиг. 19C и 19D. Один результат просто восстанавливает первоначальную ситуацию, то есть, штамм, аналогичный CVD109, у которого утрачены гены ctx, zot, и mer. Второй исход события возможно приведет к потере последовательностей pGP704, однако mer и ctx-последовательности в пределах области hlyA-последовательностей не утрачены. Два возможных исхода события легко обнаруживаются методом ДНК- гибридизации с использованием радиоактивно-меченой ctx- последовательности в качестве зонда. Для изоляции целевого рекомбинанта, культуру клеток CVD109, содержащую совместно интегрированные плазмиды pCVD-622.2B выращивали в L-бульоне без добавления антибиотиков. Эти культуры засевали на L-агаровых чашках без селективного маркера и выращенные колонии реплицировали на ампициллинсодержащих агаровых чашках. Ампициллиночувствительные колонии затем гибридизировали с ctx-зондом и выделяли колонии с ctx-последовательностями. Одна такая колония была обозначена как V.cholerae CVD110. Этот штамм, как подтверждает анализ методом ДНК-гибридизации, содержит ctx- и mer-последовательности, а также потерю фрагмента pGP704 и HpaI-фрагмента длиной 400 п.о. в пределах области hlyA-гена. Штамм V. cholerae CVD110, как также подтверждают результаты твердофазного иммуноферментного анализа, вызывает продукцию B-субъединицы холерного энтеротоксина [Sack, D.A. и другие, см. ранее (1980].
Последовательность ДНК вставленных генов: точные последовательности ДНК вставленного ctx и mer гены известны из литературы. Точное местоположение сайта hlyA-гена, в который указанные гены вставлены, также известен.
Пример 12
Описания ACE (примесного энтеротоксина холеры)
Как указывалось ранее (пример 10), гены zot и ctx расположены в области с ДНК-последовательностью длиной 4.3 т.п.о., которая у большинства штаммов вибриона холеры Эль Тор, фланкирована копиями RS1-последовательностей. Что касается вакцинного штамма CVD110, полная ДНК-последовательность в этой области удалена. Что касается локуса zot- или ctx-гена, то имеются последовательности ДНК, кодирующие третий энтеротоксин, ACE. На фиг. 20, показана карта этой области. Фрагмент EcoRV, длиной 2,9 т.п. (SED ID N: 1) клонировали в вектор плазмиды pCVD315 [Galen et al. Advances in Research on Cholera and Related Diahrreas, выпуск 7, (Sack et al., переработанное), стр. 143-153 (1990) 1 с получением клона pCVD630(показан на фиг. 20). Плазмиду pCVD630 затем переносили в штамм CVD110 V. cholerae и активность этого штамма измеряли в камере Ussing(камеры Ussing, описанные ранее). Культуральные надосадки штаммов CVD110 и CVD110, содержащие pCVD630 проверяли по методике, описанной ранее. На фиг. 21A приведены примеры полученных фракций культуральных надосадков, которые содержали ДНК-молекулы массой менее 10 килодальтон и фракции, содержащие ДНК-молекулы массой более 10 килодальтон. По существу, никакой активности Ussing не отмечено у фракций < 10 килодальтон. Во фракции > 10 килодальтон CVD110 вызывал некоторые изменения показателей тока короткого замыкания (Isc) но не больше, чем в отрицательном контроле, PBS (забуференный фосфатом физиологический раствор). Однако культуральные надосадки CVD110 (pCVD630) не вызывали существенного роста величины Isc. Это различие было статистически достоверно (p< 0.02) по сравнению с результатами, полученными только лишь у штамма CVD110. Как указывалось ранее, увеличение величины Isc может быть обусловлено как увеличением разности потенциалов (PD), так и увеличением электропроводности ткани (Gt). Увеличение величины Isc, индуцируемое генами, содержащимися на pCVD620 вызвано повышением PD, о чем свидетельствуют результаты, приведенные на фиг. 21B.
Таким образом, область длиной 4.3 т.п.о., охваченная RS1- элементами в штаммах Эль Тор, содержит 3 условных энтеротоксина. Такой же участок длиной 4,3 т. п.о. присутствует в классических штаммах, однако RS1-повторы найдены только на одной стороне. Каждый их этих трех энтеротоксинов может вызывать увеличение величины Isc в камере Ussing при использовании образцов ткани подвздошной кишки кролика, при этом их активность сопоставима с результатом их способности вызывать диарею у человека. Два токсина, холерный энтеротоксин и ACE, воздействуют по мере увеличения величины PD, в то время как третий, ZOT начинает воздействие при увеличении тканевой электропроводности. Целесообразно исключить все эти три активных фактора из ослабленных вакцинных штаммов V. cholerae для предотвращения реактивности, которая наблюдается у штамма CVD101 и других ослабленных вакцинных штаммов. Штамм CVD110, у которого отсутствуют все эти три активности, как видно из фиг. 21, вызывает изменения в камере Ussing, сопоставимые с результатами вариаций, индуцируемых PBS (то есть, по существу не происходит каких-либо изменений).
Последовательность ДНК фрагмента, распознаваемого EcoRV, длиной 2,9. т. п. o (SED ID N: 1) в составе плазмиды pCVD620 показана на фиг. 22. Имеются две открытых рамки считывания (ORFs) с моментальной транскрипцией zot-гена в 3'-5' направлении. Меньшая открытая рамка считывания ORF с моментальной транскрипцией zot-гена в обратном направлении представляет собой нуклеотидную последовательность, длиной 297 п.о., с кодоном (ACE), которая может потенциально кодировать белок размером 11 килодальтон и большая ORF с моментальной транскрипцией гена в обратном направлении из ORF, длиной 297 п.о., представляет собой нуклеотидную последовательность, длиной 1185 п.о., с кодоном (OrfU), которая может потенциально кодировать белок массой 44 килодальтон. Активность энтеротоксина Ace, как полагают, локализована относительно открытой рамки считывания длиной 297 п.о.
Пример 13
Конструирования V. cholerae CVD112 - вакцинного штамма V. cholerae 0139 серогруппы.
Штаммы V. cholerae подразделяются на многочисленное количество групп 0. Штаммы 0 группы 1 вызывают холеру с потенциалом для широко распространенной передачи инфекции в очаги пандемии. V. cholerae из других 0 групп, так называемый не-01 V.cholerae, может вызвать заболевание, но до недавнего времени, не приводил к эпидемической холере. В последнее время появились сообщения о десятках тысяч случаев инфицирования именно этим V.cholerae, не относящимся к группе 01. Полагают, что имеющиеся в настоящее время вакцинные штаммы на основе вибриона холеры не-01 серогруппы могут оказаться неэффективными для выработки иммунитета против вибрионов не -01 серогрупп. Новый клинический штамм обозначили как новую серогруппу 0139.
В качестве исходного штамма для получения вакцинного штамма V. cholerae штамм не-01 серогруппы использовали V.cholerae 1837. Этот штамм выбран из серотипа 0139 и выделен из заболевшего холерой пациента в Бангладеш. На фиг. 23 показано расположение в хромосоме локуса холерного энтеротоксина в штамме 1837.
Плазмиду pCVD51 переносили в штамм 1837 по методике, описанной в примере 10. Эта плазмида претерпела рекомбинацию в бактериальной хромосоме, и второе событие рекомбинации вызвало потерю одной копии генов энтеротоксина и 3 копий RS1-элементов. Полученный штамм V.cholerae 1837.1, показан на второй строке фиг. 24. Для простоты, на фиг. 24, последовательности генов сер, orfU, асе, zot, ctxAB указываются вместе как "ядро" (сердцевина, кор-часть) энтеротоксина V.cholerae. Локус хромосомальной ДНК V. cholerae включает один или несколько кор-участков и ассоциированные с ними RS1-элементы.
Остальные последовательности хромосомальной ДНК штамма 1937.1 были удалены следующим образом. Получали хромосомальную ДНК из штамма 1837.1, которая частично гидролизована рестрикционной эндонуклеазой Sau3a с вырезкой фрагментов длиной примерно от 30 до 40 т.п.о. Эти фрагменты клонировали в векторе космидной плазмиды pHC79 (Hohn и Collins, Gene, 11, 291-298, 1980) и полученные клоны скринировали после переноса в E. Coli методом ДНК-гибридизации с использованием последовательностей ctx-гена в качестве зонда. Клон, содержащий cxt-гены на вставке длиной приблизительно 30 т.п.о. был идентифицирован и изолирован. Полученная плазмида была частично картирована. Изолированную конструкцию обозначили на фиг. 25 как pLC13. Эту плазмиду гидролизовали рестриктазой EcoRI и вновь сшивали, в результате чего размер вставки уменьшился в длину примерно до 25 т.п.о. Полученная плазмида была обозначена как pLC14. Плазмиду pLC14 частично гидролизовали рестриктазой HindIII и затем вновь сшивали для удаления распознаваемого HindIII фрагмента длиной примерно 14,6 т.п.о, содержащего кор-область, RS 1 -последовательности и несеквенированный фрагмент ДНК V. cholerae, длиной примерно 7,4 т.п.о. Полученную плазмиду с делецией кор-области и RS1-последовательности обозначили как pLC15. Аналогичный распознаваемый HindIII фрагмент был вырезан в процессе конструирования ослабленного штамма по методике, описанной Pearson и другими, P.N.A.S. (US), 90, 3750-3754, 1993. Полученную плазмиду pLC15 затем гидролизовали рестриктазами Sal1 и EcoRI, и ДНК вибриона клонировали в киллерную векторную плазмиду pGP704 по методике, описанной в примере 10. Полученную плазмиду обозначили как pLC16.
Киллерную векторную плазмиду pLC16, содержащую последовательности хромосомальной ДНК штамма 1837.1 V. cholerae, которые охватывают ctx-локус, переносили в штамм 1837.1 V. cholerae по методике, описанной в примере 10. Распознавание гомологичной in vivo рекомбинации векторных последовательностей в хромосому проводили скринингом на ампициллиноустойчивых колониях, поскольку фактор устойчивости к ампициллину кодируется вектор- опосредованными ДНК-последовательностями.
Ампициллиноустойчивый V. cholerae, полученный в соответствии с этим методом, содержит плазмидные ДНК-последовательности, интегрированные в бактериальную хромосому, затем выращивали при отсутствии ампициллина, отбирали ампициллиночувствительные колонии. Второе событие рекомбинации привело к потере фенотипа ctxA, ctxB, zot, асе, orfU, сер и RS1-элементов наряду с плазмидным вектором, кодирующим фактор ампициллиноустойчивости. Этот конечный штамм обозначили как 1837.2. На фиг. 24, третья строка, показана в схематическом виде хромосома вибриона в штамме 1837.2.
Ген, кодирующий B-субъединицу холерного энтеротоксина вместе с геном, кодирующим фактор устойчивости к ртути, переносили в штамм 1837.2 по методике, описанной выше в примере 11. В качестве киллерного вектора pCVD622.2B, использовали тот же самый вектор, который использовали для получения штамма CVD110. Событие, приводящее к вставке последовательностей ДНК B-субъединицы холерного энтеротоксина изображено на фиг. 26. Конечный продукт, штамм CVD112 содержит делеции всех генов ctx, zot, асе, сер и RS1-элементов, экспрессирует только B-субъединицу холерного энтеротоксина и фактор устойчивости к ртути.
Получение препарата штаммов CVD112 и Al1837 для исследования возможности использования в качестве вакцины.
CVD112 и Al1837 штаммы V. cholerae хранили в TSB, содержащем 18% глицерина при температуре -70oC. Перед введением добровольцам, содержимое ампулы соответствующего штамма оттаивали и наносили штрихами на кровяной агар, TCBS, и BI-агар. После инкубации в течение 24 часов при температуре 37oC, выбирали колонии клеток, которые агглютинировали соответствующие сыворотки, и засевали на BHI-агаре, затем инкубировали при температуре 37oC в течение приблизительно 20 часов. Штамм V. cholerae затем вновь засевали, и инкубировали в течение 5 часов при температуре 37oC и полученный урожай клеток собирали в стерильный физиологический раствор. Концентрацию определяли по оптической плотности растворов, которые сравнивали с известными стандартами. Затем проводили количественный анализ методом переноса реплики до и после введения в организм добровольцев.
Клинические испытания вакцин.
Была отобрана группа из взрослых добровольцев в возрасте от 18 до 40 лет, с которой была проведена беседа и получено их письменное согласие. Добровольцев тщательно обследовали для гарантии того, что они находятся в превосходном физическом и психическом состоянии. Группу из 14 добровольцев госпитализировали и дали возможность адаптироваться к окружающей обстановке в течение двух дней, во время которых была закончено обследование и взяты образцы крови в качестве референс-образцов для определения уровня образования антител.
Добровольцев обследовали двойным слепым методом на предмет получения одной из следующих доз вакцины на третий день:
(i) CVD112 в дозе 5•108 cfu с использованием буфера (n=6)
(ii) CVD112 в дозе 5•106 cfu с использованием буфера (n=6)
(iii) только лишь буферный раствор
Два вакцинированных добровольца, получавшие только буферный раствор, находились вместе с реципиентами, получавшими вакцину для определения возможности или невозможности переноса инфекции от одного лица другому в условиях эксперимента. За вакцинированными вели постоянное наблюдение в течение 5 дней, а затем провели курс лечения тетрациклином в течение пяти дней.
Через примерно 5 недель, добровольцев заражали повторно патогенным штаммом Al1837 V. cholerae серогруппы 0139 для оценки эффективности вакцины против гомологичного антигена. Провокационная доза содержала приблизительно 106 cfu штамма Al1837 V. cholerae 0139. За добровольцами вели постоянное наблюдение в течение приблизительно 120 часов после провокации. Все добровольцы получали тетрациклин (в дозе 500 мг каждые 6 часов в течение 5 дней), начиная с 5 дня после иммунизации и с 4 дня после контрольного инфицирования для исключения последействия вакцинных штаммов и провокации.
Восьми вакцинированным проводили контрольное заражение, при этом одна группа из четырех человек получала дозу 5•108, а другая группа из четырех человек дозу 5•106 Группа из пятнадцати невакцинированных добровольцев, служившая в качестве контроля, также получала дозу провокационной пробы. У одного из восьми получавших вакцину (13%) и двенадцати человек в контроле (80%) была отмечена диарея после провокации, в результате эффективность вакцины составила 84%. Один из вакцинированных, у которого был отмечен понос, получил вакцинный штамм CVD112 в дозе 5•106.
У трех (38%) из восьми вакцинированных и 14 (93%) в контроле не обнаружено выделения штамма, используемого для инфицирования, хотя пиковая экскреция была в 100 раз ниже, чем у вакцинированных, свидетельствуя тем самым о том, что вакцинный штамм эффективно размножался в кишечнике вакцинированных.
Эти результаты являются наглядным доказательством безопасности и эффективности штамма V. cholerae CVD112 для использования в качестве вакцины, чтобы вызвать поствакцинальный иммунитет против холеры, вызванной вибрионными штаммами не 01- серогруппы 0139 серотипа.
Пример 14
Конструирование V.cholerae CVD112 RM-ослабленного, recA- мутанта V. cholerae серогруппы 0139.
Штамм V. cholerae CVD112 является исходным штаммом для конструирования ослабленного штамма V. cholerae, rec-, 0139- серогруппы. Мутация recA-гена, как полагают, необходима для снижения уже и так малой вероятности реверсии вирулентности ослабленного штамма через рекомбинацию ctx-генами дикого типа in vivo. RecA-мутант конструировали по аналогичной методике, описанной Ketley и другими, Infect. Immun., 58 (5), 1481-1484, 1980 и Goldberg и другими, J.Bacter, 165 (3), 715-722, 1986.
Плазмиду pCVD831, несущую NarI-NheI-фрагмент длиной 7 т.п.о. с recA-геном штамма N16961 V. cholerae Эль Тор, клонировали в плазмиду-хозяин pCVD316 с широким спектром действия, которая является производной описанной ранее IncP-плазмиды pRK290. Плазмиду PCVD831 гидролизовали рестриктазами XbaI и PvuII для вырезки фрагмента длиной приблизительно 50 п.о. в пределах ДНК-последовательностей, кодирующих ген recA. Концы рестриктированного фрагмента ДНК затем заполняли брешью для получения его с тупыми концами, и полученную плазмиду повторно сшивали. Удаление фрагмента длиной приблизительно 50 п.о. обеспечивает инактивацию гесA-белка. Полученную плазмиду обозначили как pCVD832.
Плазмиду pCVD832 переносили в штамм V. cholerae CVD112, и в результате гомологичной рекомбинации происходит замена нативного гесA-гена гесA-мутантом. Результат события гомологичной рекомбинации определяют методом скрининга на фенотип, чувствительный к метилметансульфонату (MMS), в соответствии с методикой, описанной Kettler и другими, см. ранее (1980). Штамм, полученный в результате этих манипуляций, имеет делецию всех ctx-, сер-, orfU-, ace-локусов и RS1- элементов и инактивированную мутацию в хромосомальном rec-A локусе (то есть, представляет собой recA--штамм). Результат события перекрестной рекомбинации, который происходит в recA-локусе V. cholerae в поколении штамма CVD112 RM показан схематически на фиг. 27.
Штаммы V. cholerae CVD112 и CVD112 RM можно эффективно использовать в качестве вакцины, вызывающей защитный иммунитет против вирулентного штамма серотипа 0139 и других, не относящихся к серогруппе 01 штаммов после последующего инфицирования этими вирулентными штаммами.
Штаммы CVD112 и CVD112 RM пригодны также для использования с одним или несколькими другими вакцинными шаммами V. cholerae, анатоксинами, термоинактивированным холерным энтеротоксином и т. д. , в виде комбинированной (ассоциированной) вакцины для защиты против как вирулентных штаммов 01 серогруппы, так и вирулентных штаммов не-01 серогруппы, например штаммы V. cholerae серотипа Огава или Инаба.
Совсем недавно появились сообщения о том, что локус гена сер, который кодирует закодированные в хромосоме ядра клетки, может действовать как дополнительный фактор колонизации. Однако, продукт cep-гена, как полагают, не оказывает существенного влияния на способность штаммов Vibrio cholerae предлагаемого изобретения к колонизации тонкого кишечника организма-хозяина, в частности кишечника у человека. Однако, при желании, ДНК-последовательности, включая ген сер, или любые другие пригодные факторы колонизации, можно вставить опять в хромосому V.cholerae, в соответствии с предлагаемым изобретением, следуя методике и методам, раскрываемым в настоящем описании. В качестве примера, при повторном встраивании ДНК-последовательностей, кодирующих B-субъединицу холерного энтеротоксина и ДНК-последовательности, кодирующие фактор устойчивости к ртути, в хромосому в локусе гемолизинового гена, специалист может без затруднения перенести интересующий ген, активно связанный с сигналом экспрессии в плазмиду, которую используют в методике рекомбинации.
Штамм, полученный в результате такой перекрестной рекомбинации синтезирует B-субъединицу холерного энтеротоксина, то есть белка, вызывающего устойчивость к ртути и другим тяжелым металлам, и дополнительный ген рассматриваемый в данном описании.
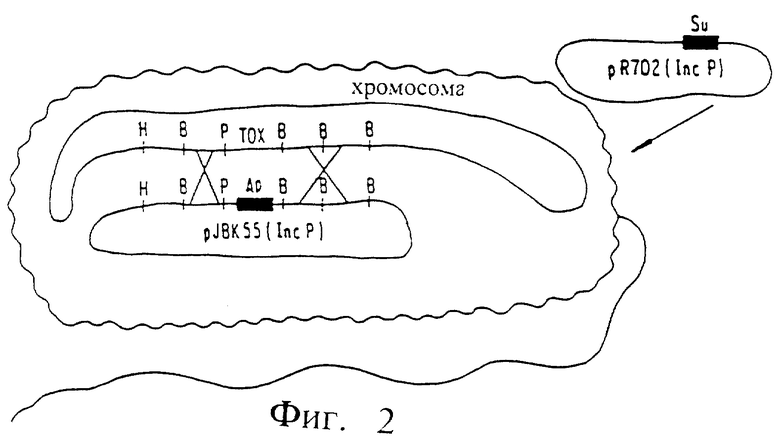
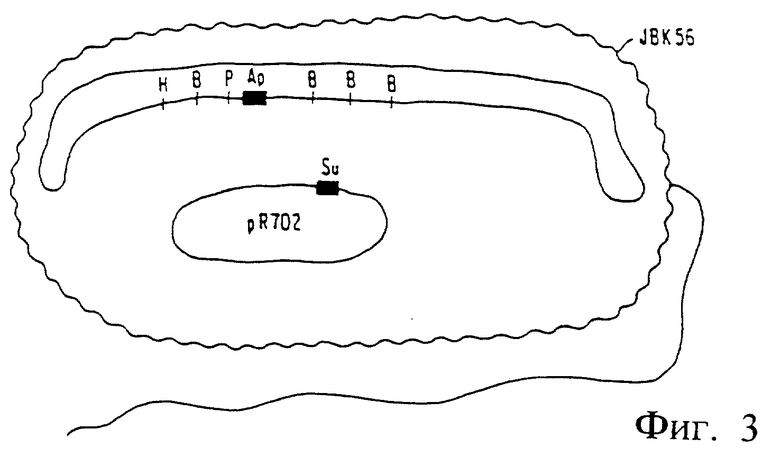
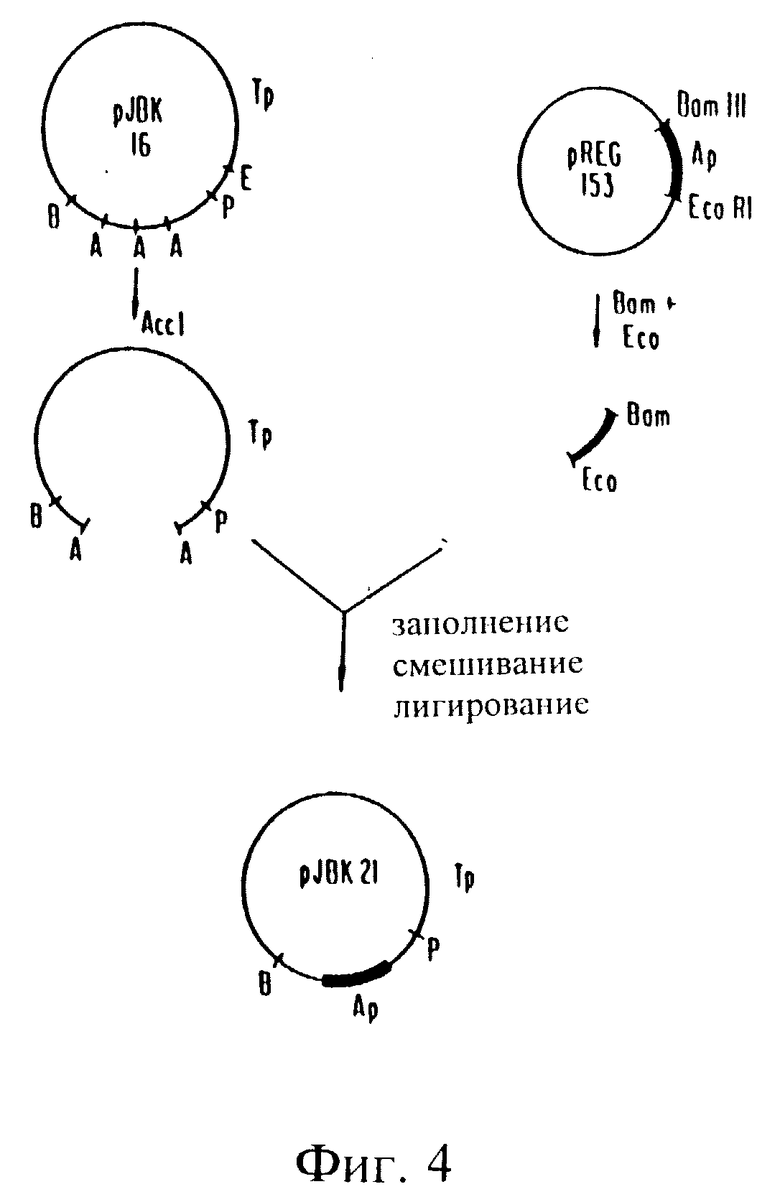
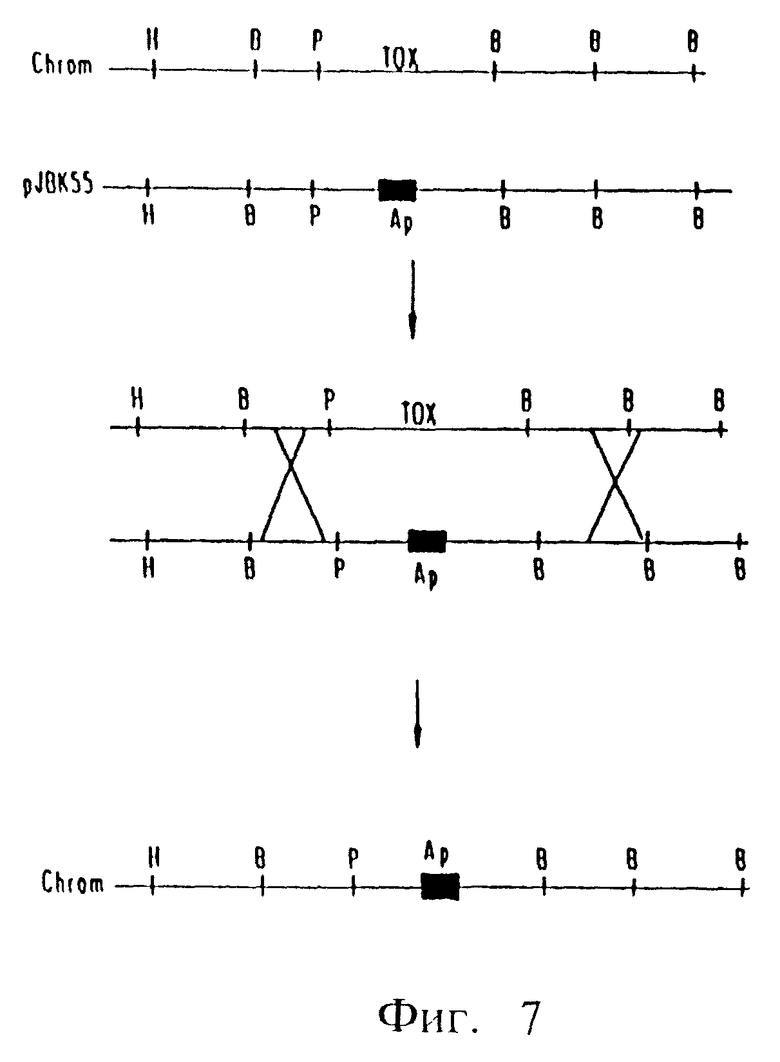
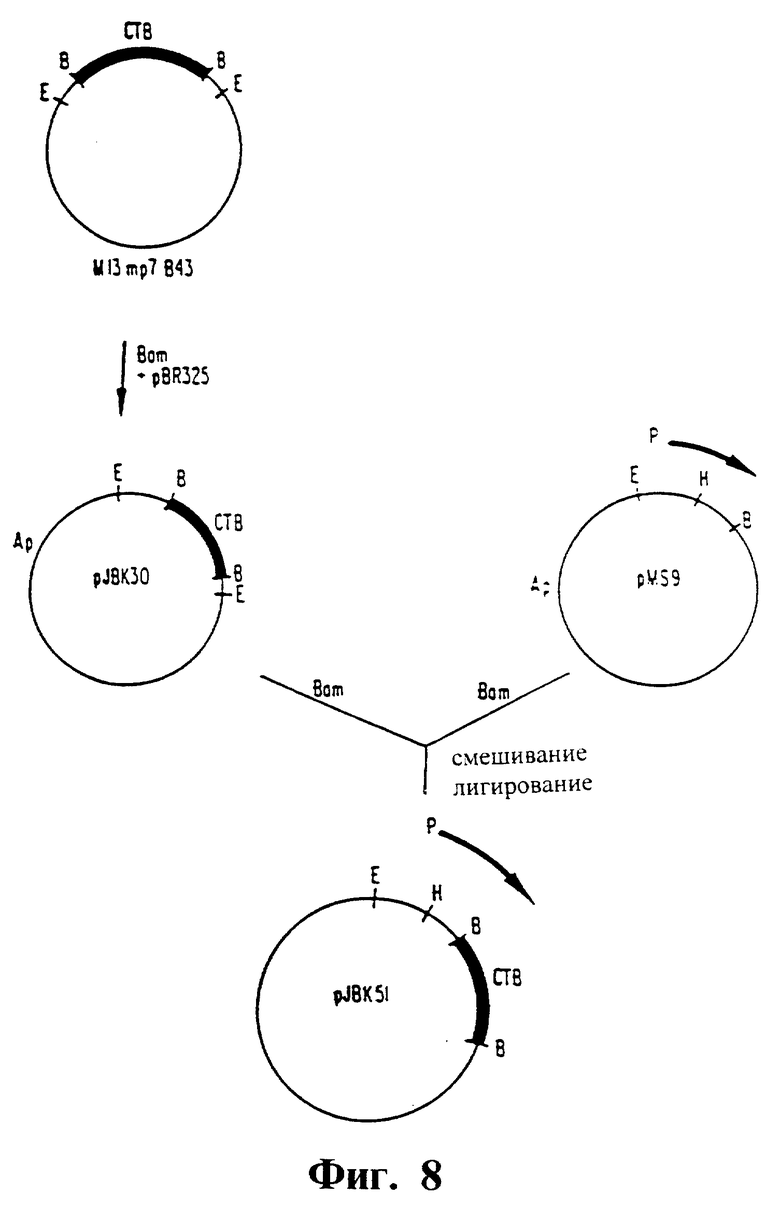
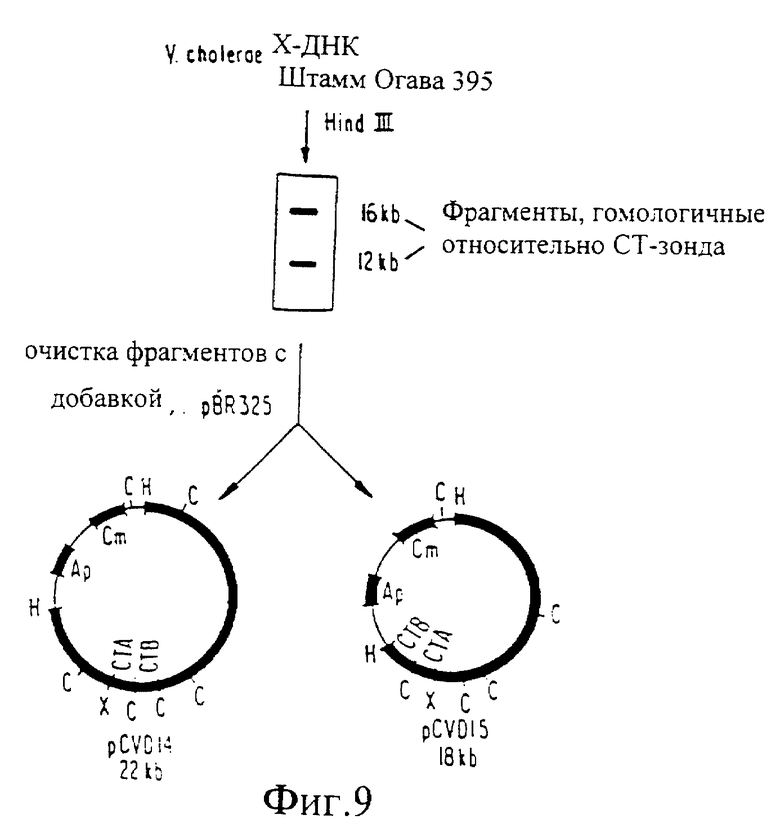
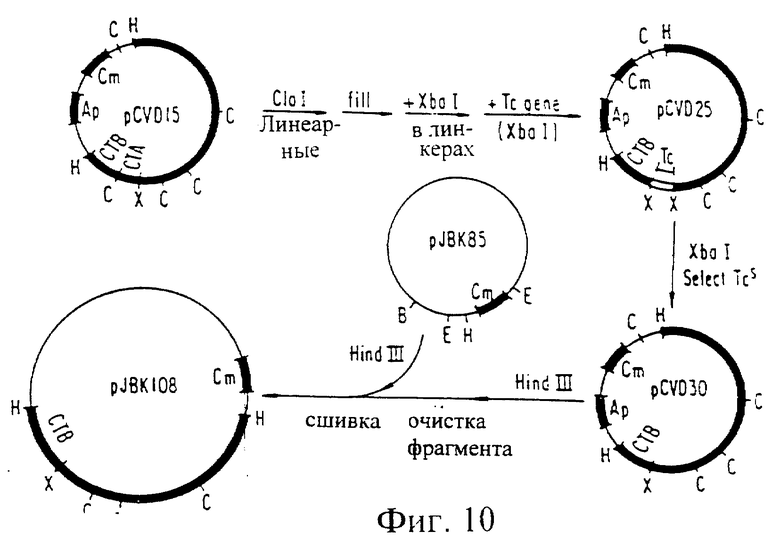
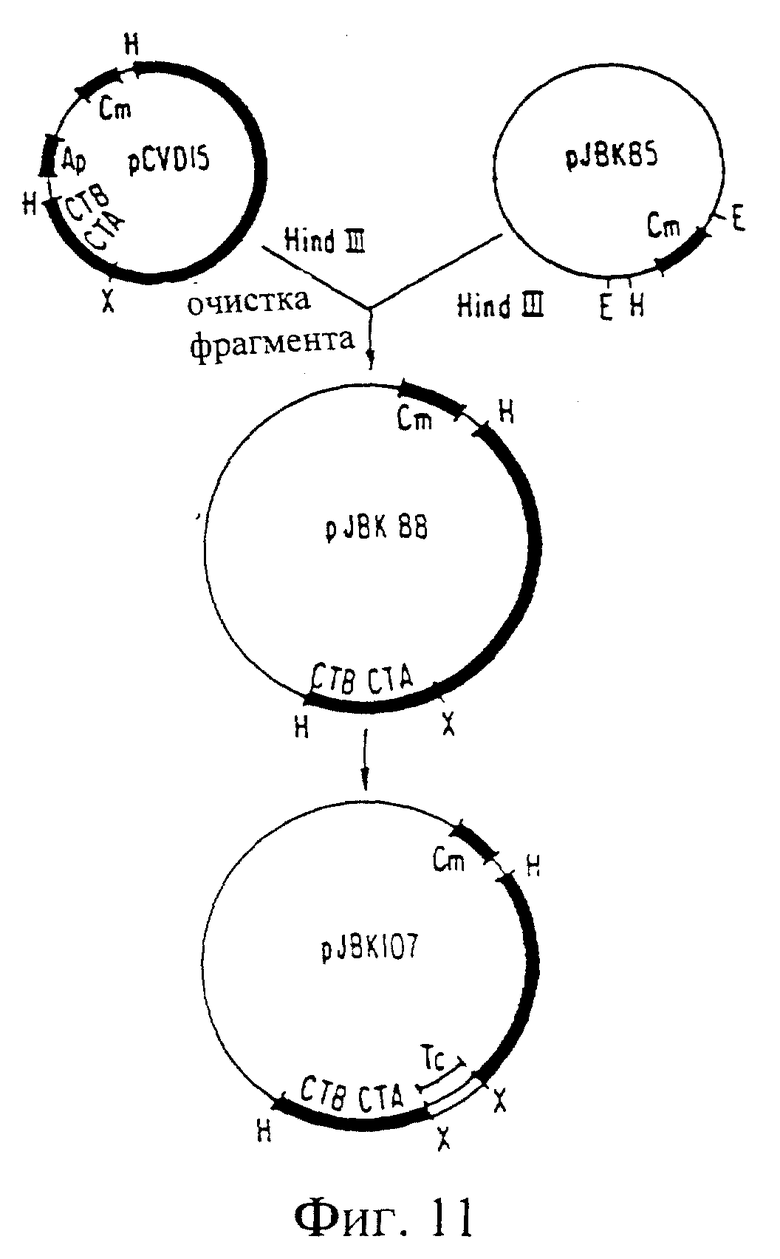
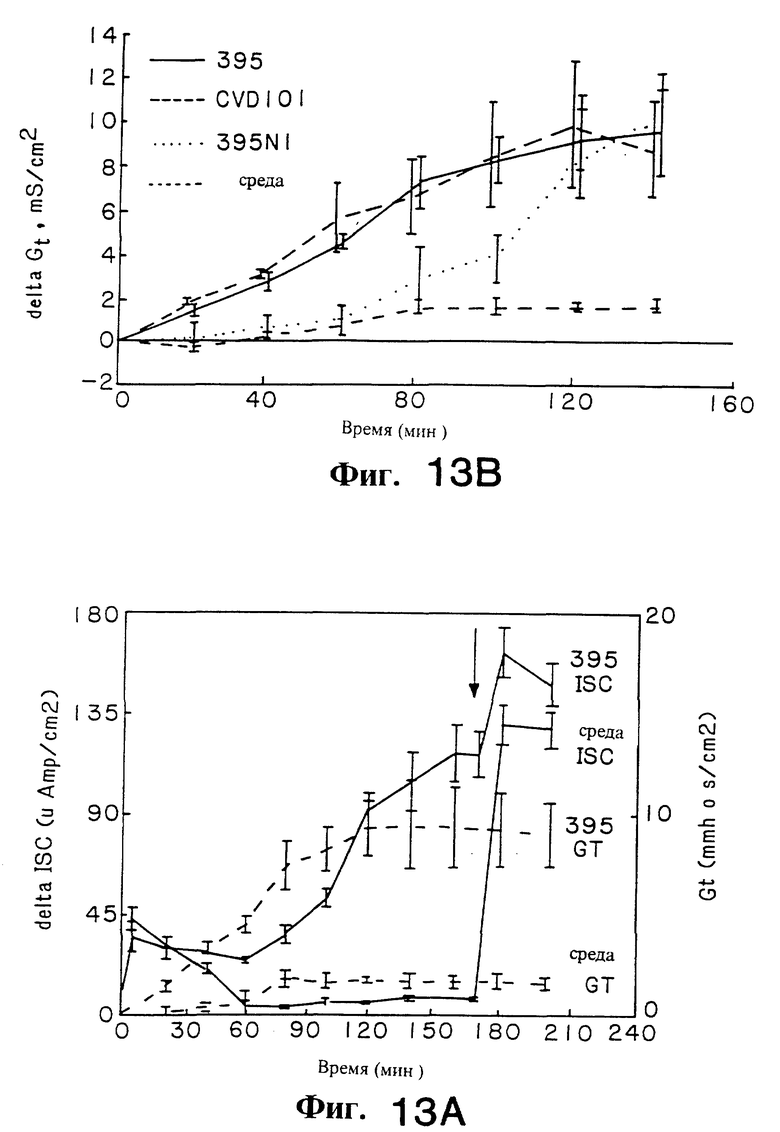
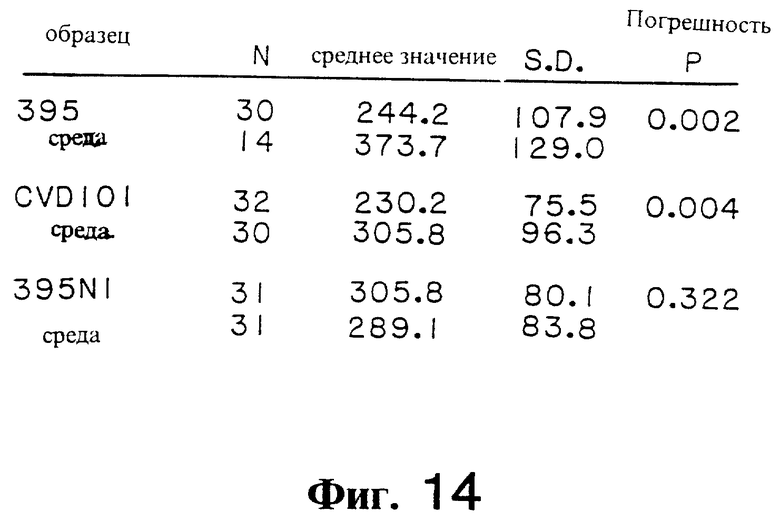
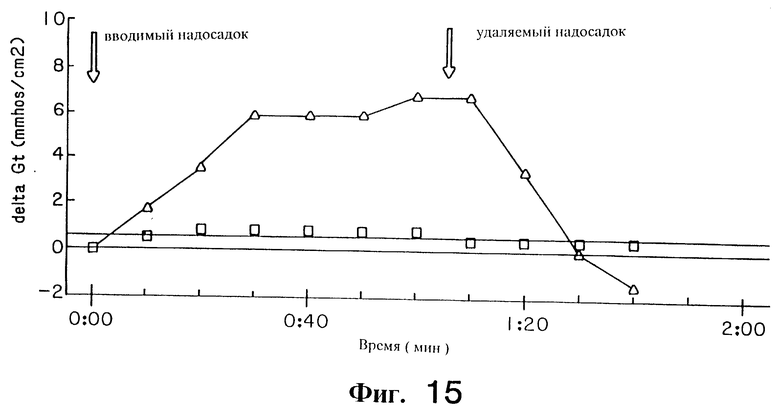
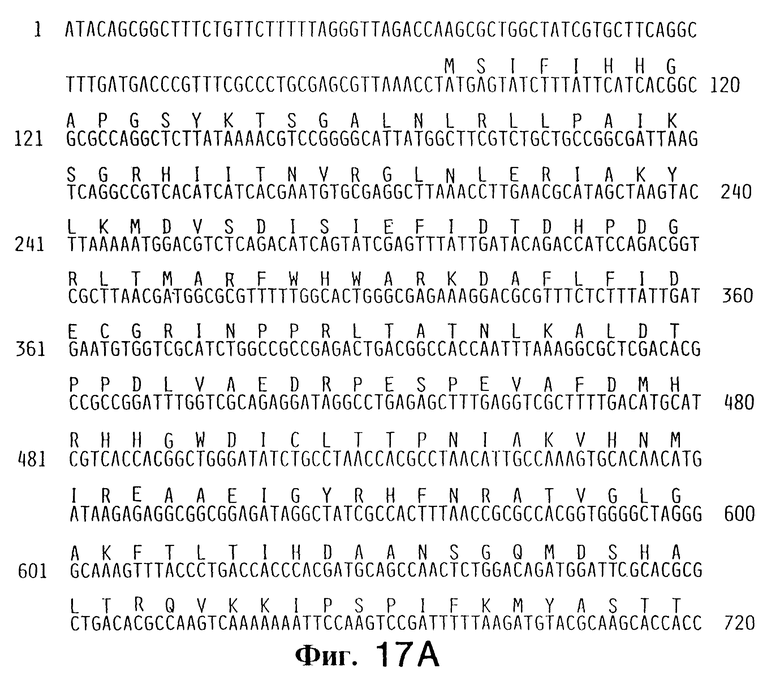
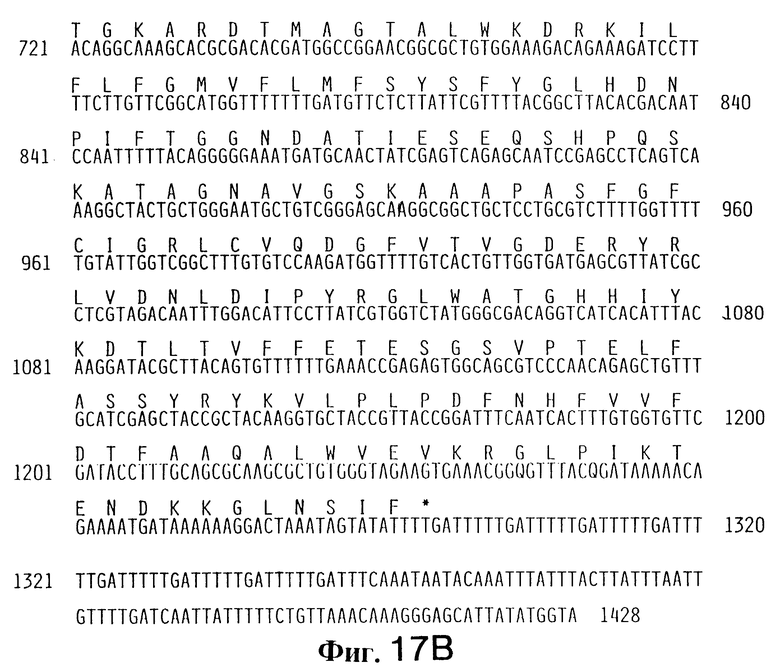
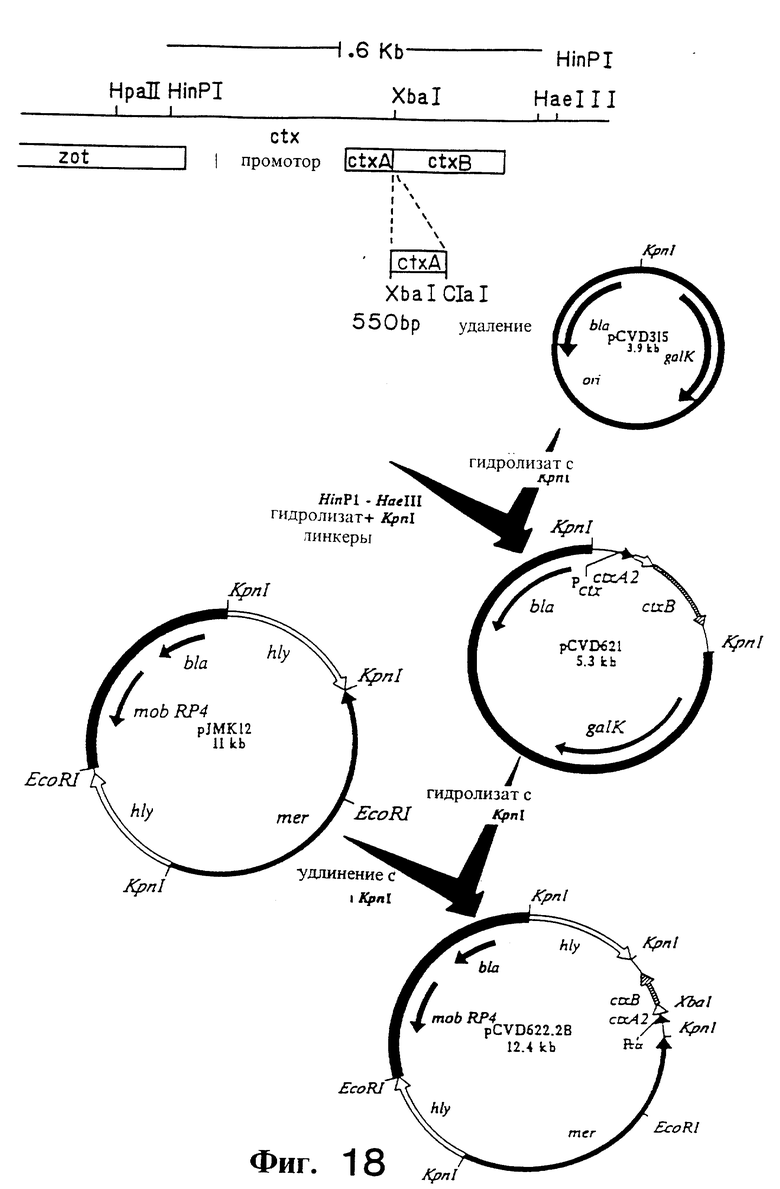
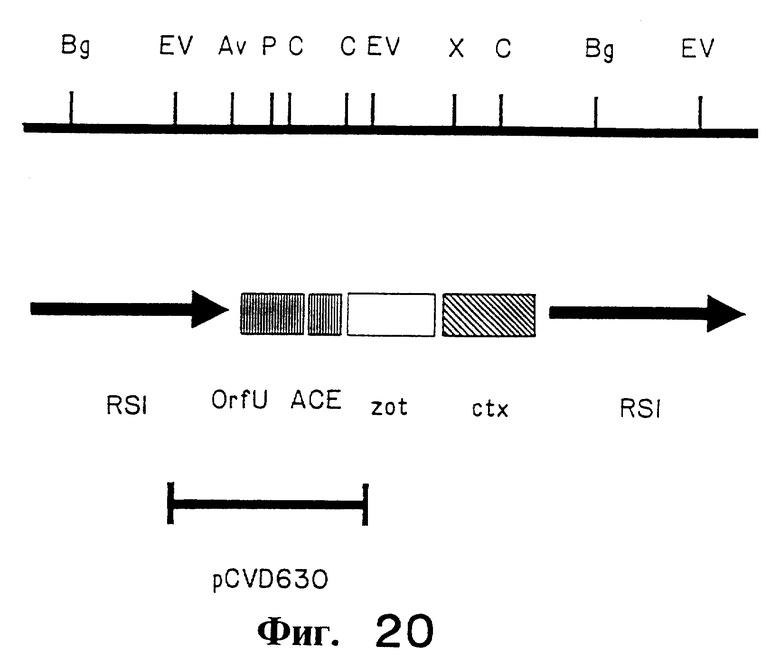
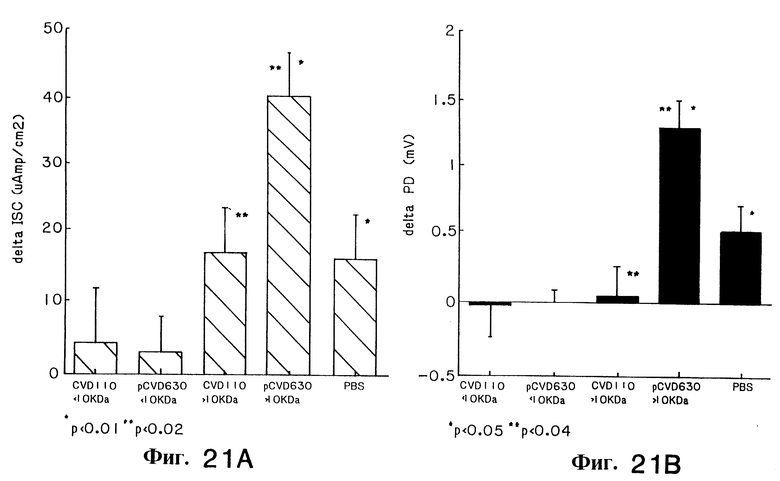





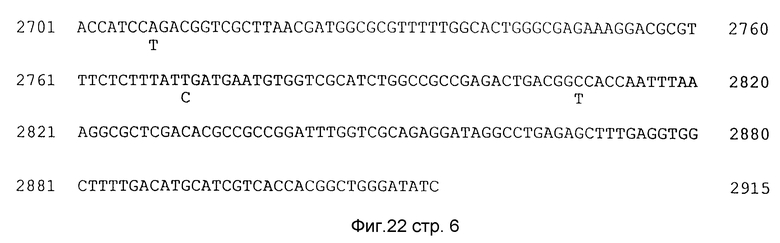
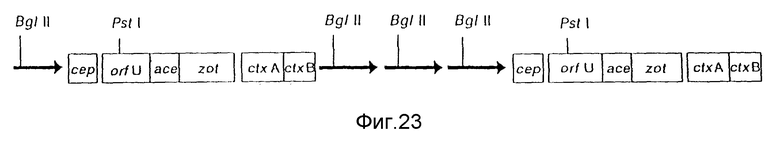
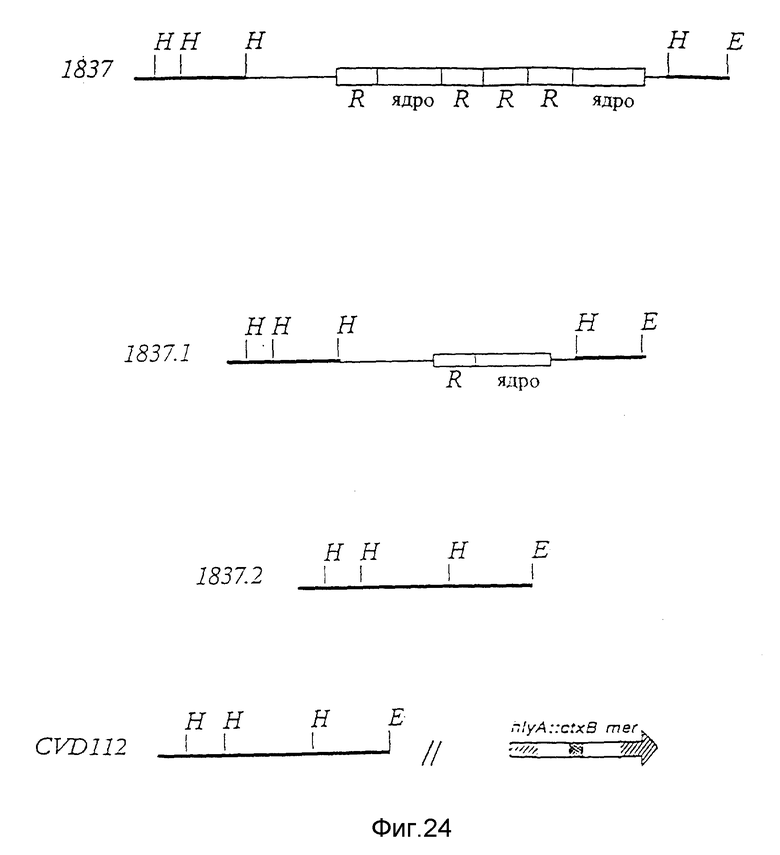
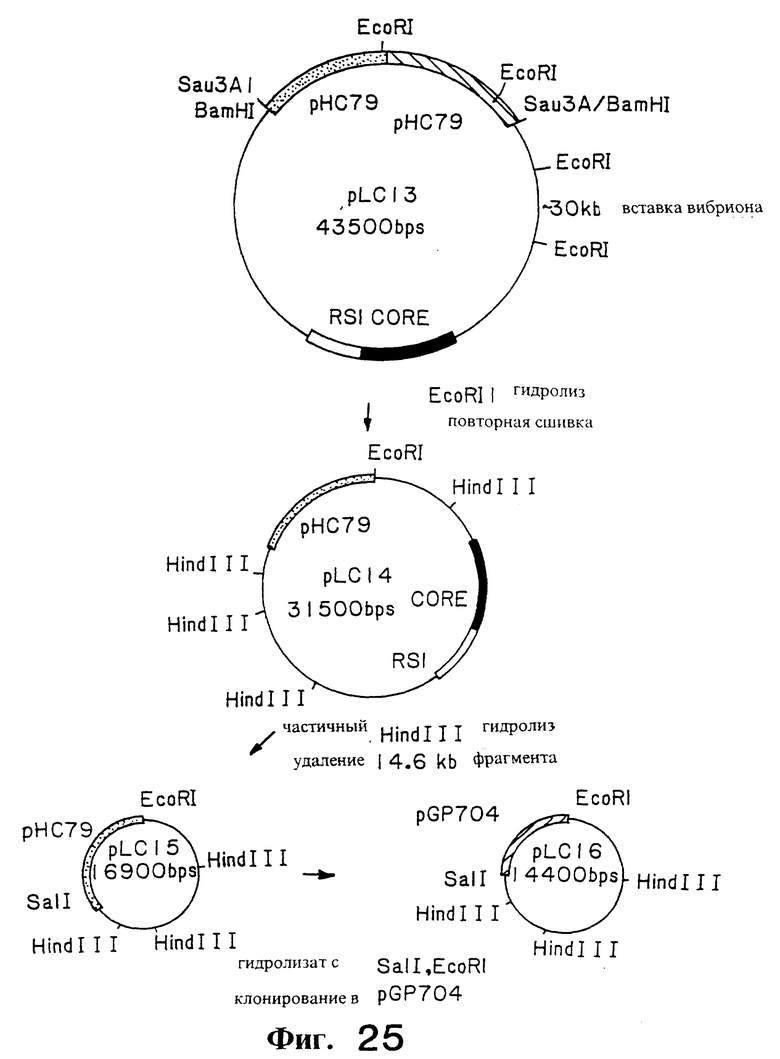
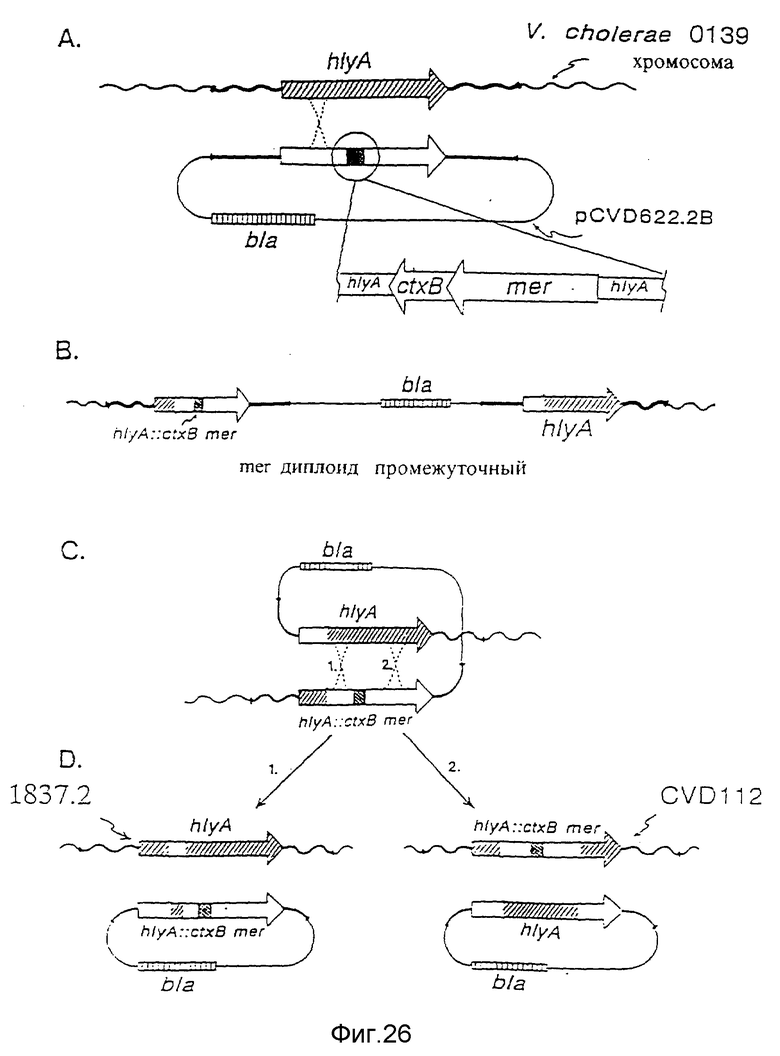
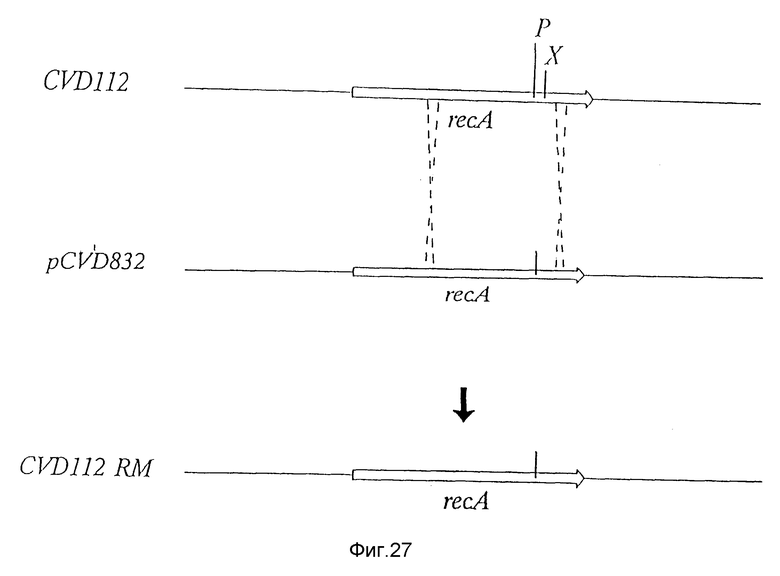
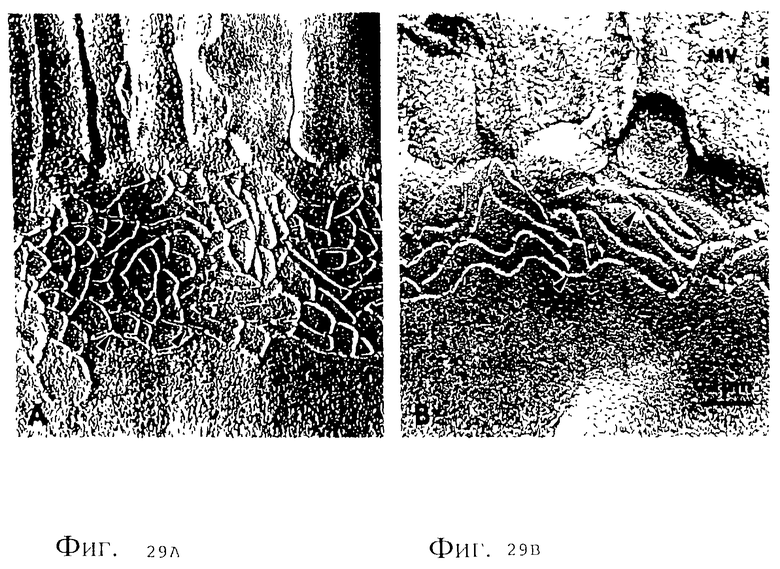
Изобретение относится к авирулентным штаммам Vibrio cholerae (CVD 112 и CVD 112RM) не-01 серогруппы, у которых отсутствует RS1-последовательность и коровая (сердцевинная) последовательность холерного энтероксина. Штаммы содержат последовательности, кодирующие устойчивость к ртути, и последовательности, кодирующие В-субъединицу холерного энтеротоксина, повторно вставленную в хромосому. Изобретение также относится к способам получения авирулентных штаммов Vibrio cholerae (CVD 112 и CVD 112RM) не-01 серогруппы. Авирулентные штаммы Vibrio cholerae CVD 112 и CVD 112RM не-01 серогруппы применяются в производстве вакцины против холеры. 5 с. и 11 з.п. ф-лы, 40 ил.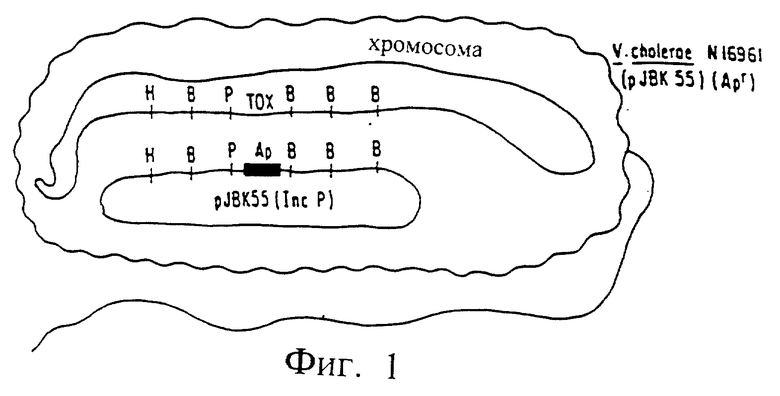
| Аналогов в научно-технической и патентной литературе не обнаружено. |

