ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее раскрытие, в общем, относится к популяциям клеток и растворимым медиаторам, способным подавлять активацию Т-клеток, и к применению таких популяций клеток и растворимых медиаторов для подавления активации Т-клеток, как, например, при лечении заболеваний или состояний, опосредованных функцией эффекторных Т-клеток. Данное раскрытие также относится к способам детектирования присутствия маркера у субъекта, который указывает на чувствительность субъекта к заболеваниям или состояниям, опосредованным функцией эффекторных Т-клеток.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Регуляторные Т-клетки (Treg клетки, также известные как Т-клетки-супрессоры) представляют собой субпопуляции Т-клеток, которые поддерживают иммунный гомеостаз и помогают предотвращать аутоиммунное заболевание (Sakaguchi et al., 2008; Shevach, 2006; Vignali et al., 2008). Интерес к Treg клеткам главным образом сосредоточен на прототипных Treg клетках CD4+ CD25+, которые программируются транскрипционным фактором FoxP3 (Fontenot et al., 2003; Hori et al., 2003). В полиспецифичных популяциях в состоянии покоя данные Treg клетки характеризуются у мышей и как «природные», происходящие из тимуса, и как индуцированные «адаптивные» клетки, которые подавляют активацию, пролиферацию и функции других Т-клеток (Sakaguchi et al., 2008; Shevach, 2006). Однако в человеческой крови Treg клетки CD4+ не так надежно отличаются по экспрессии FoxP3 (Roncarolo and Gregori, 2008; Allan et al., 2007; Gavin eral., 2006). Таким образом, было показано, что Т-клетки CD4+с маркерами либо наивных клеток, либо клеток памяти имеют сходные супрессорные функции, несмотря на низкую и высокую экспрессию FoxP3 соответственно (Miyara et al., 2009). Другие поверхностные маркеры человеческих Treg клеток CD4+ CD25+ FoxP3+, такие как пониженная экспрессия рецептора IL-7, CD127 (Liu et al., 2006; Seddiki et al., 2006), не являются специфичными для Treg клеток.
Помимо малочисленности специфичных маркеров поверхности клетки, механизмы, лежащие в основе подавления Treg клетками CD4+ CD25+ FoxP3+, остаются противоречивыми. У мышей было показано, что подавление ex vivo требует контакта клетка-клетка, но было приписано многим механизмам (Vignali et al., 2008; Shevach, 2009; Sakaguchi et al., 2009); даже меньше известно о функции аналогичных человеческих Treg клеток. Кроме того, в контексте разных сайтов в ткани или заболеваний были описаны другие типы как CD4+, так и CD8+Treg клеток, которые отличаются по предложенным механизмам функции подавления (Vignali et al., 2008).
Было показано, что Treg клетки, индуцированные введением аутоантигенов, защищают против некоторых аутоиммунных заболеваний в определенных животных моделях (обзор дается von Herrath and Harrison, 2003). Например, было показано, что в модели диабета типа 1 (T1D) на основе не страдающих ожирением диабетических (NOD) мышей Treg клетки CD4+, индуцированные введенными аутоантигенами островков поджелудочной железы, такими как инсулин (Bergerot et al., 1994) или декарбоксилаза 65 глутаминовой кислоты (GAD65) (Tisch et al., 1999), или перенос Treg клеток CD4+, индуцированных проинсулином (Every et al., 2006), или предположительным антигеном островков поджелудочной железы (Tang et al., 2004), защищают против аутоиммунного диабета. Однако в данных моделях Treg клетки исследовали в полиспецифичных популяциях, находящихся в состоянии покоя, а не во время ответа хозяина на конкретный антиген. Недавно были получены клоны проинсулин- и GAD65-специфичных человеческих Т-клеток CD4+, и клоны Treg различали по их супрессорной функции in vitro (Dromey et al., 2011). Было показано, что заякоренный в мембране гликопротеин CD52 поверхности клетки подвергается повышающей регуляции в данных размноженных клонах Treg CD4+. Однако механизм иммунодепрессии ранее не был охарактеризован.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения идентифицировали растворимый медиатор подавления Treg клетками. Соответственно, согласно настоящему раскрытию предложена фармацевтическая композиция, содержащая любой один или более чем один из следующих:
i) растворимый гликопротеин CD52,
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv);
vi) выделенная клетка CD52hi, способная продуцировать растворимый гликопротеин CD52;
vii) выделенная популяция клеток, содержащая множество клеток CD52hi, способных продуцировать растворимый гликопротеин CD52;
viii) среда культуры клеток или ее фракция, содержащая растворимый гликопротеин CD52, выделенный из культуры клеток, содержащей клетку по разделу vi) или популяцию клеток по разделу vii) и
ix) агент, способный увеличивать уровень экспрессии растворимого гликопротеина CD52 клеткой;
и фармацевтически приемлемый носитель.
В предпочтительном воплощении растворимый гликопротеин CD52 содержит аминокислотную последовательность, по меньшей мере на 60% идентичную аминокислотной последовательности любой одной или более чем одной из следующих: GQNDTSQTSSPS (SEQ ID NO: 3), SQNATSQSSPS (SEQ ID NO: 4), GQATTAASGTNKNSTSTKKTPLKS (SEQ ID NO: 5), GQNSTAVTTPANKAATTAAATTKAAATTATKTTTAVRKTPGKPPKA (SEQ ID NO: 6) или GNSTTPRMTTKKVKSATPA (SEQ ID N0:7), и углевод. Более предпочтительно гликопротеин содержит аминокислотную последовательность, которая по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% идентична или на 100% идентична любой одной или более чем одной из аминокислотных последовательностей, идентифицированных в SEQ ID NO: 3, 4, 5, 6 или 7.
В одном примере гликопротеин содержит аминокислотную последовательность, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% идентичную или на 100% идентичную аминокислотной последовательности SEQ ID NO: 3, которая представляет собой фрагмент человеческого растворимого CD52.
Предпочтительно любой один или более чем один из растворимого гликопротеина CD52, слитого белка, полинуклеотида, вектора, клетки, популяции клеток, культуральной среды клеток и агента присутствует в достаточном количестве для подавления функции эффекторной Т-клетки и/или иммунного ответа.
В другом воплощении растворимый гликопротеин CD52, слитый белок, полинуклеотид, вектор, клетка, популяция клеток, культуральная среда клеток и агент присутствуют в достаточном количестве, так что подавление иммунного ответа приводит к устойчивости к по меньшей мере одному антигену, такому как аутоантиген.
В другом воплощении любой один или более чем один из растворимого гликопротеина CD52, слитого белка, клетки, популяции клеток, культуральной среды клеток и агента способен подавлять функцию эффекторной Т-клетки и/или способен уменьшать иммунный ответ, такой как иммунный ответ на аутоантиген.
В одном воплощении композиция содержит один или более чем один из растворимого гликопротеина CD52, слитого белка, культуральной среды клеток или агента и приготовлена для введения через слизистую и/или чрескожного введения.
В другом воплощении композиция, кроме того, содержит инсулин и/или аутоантиген.
Согласно настоящему раскрытию также предложен слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок.
Предпочтительно растворимый гликопротеин CD52 слитого белка содержит аминокислотную последовательность, которая по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% идентична или на 100% идентична любой одной или более чем одной из аминокислотных последовательностей, идентифицированных в SEQ ID NO: 3, 4, 5, 6 или 7.
Предпочтительно слитый белок способен подавлять функцию эффекторных Т-клеток и/или способен уменьшать иммунный ответ, такой как иммунный ответ на аутоантиген. В одном воплощении слитый белок уменьшает иммунный ответ в такой степени, что он приводит к устойчивости по меньшей мере к одному антигену, такому как аутоантиген.
Второй белок может представлять собой любой белок, способный увеличивать стабильность и/или растворимость растворимого гликопротеина CD52, ускорять процесс получения растворимого гликопротеина CD52 способами генной инженерии или усиливать терапевтический эффект растворимого гликопротеина CD52. В одном примере второй белок может содержать фрагмент антитела, такой как Fc.
Предпочтительно слитый белок является растворимым.
Согласно настоящему раскрытию также предложен выделенный или рекомбинантный полинуклеотид, кодирующий раскрытый здесь слитый белок.
Согласно настоящему раскрытию также предложен вектор, содержащий раскрытый здесь полинуклеотид.
Согласно настоящему раскрытию также предложена выделенная клетка, содержащая раскрытый здесь полинуклеотид и/или вектор. Данная клетка может представлять собой клетку млекопитающего. В одном примере клетка представляет собой клетку НЕК293Т. В другом примере клетка представляет собой клетку В-лимфобласт Дауди.
Кроме того, согласно настоящему раскрытию предложен способ получения слитого белка, включающий проведение экспрессии раскрытого здесь полинуклеотида или вектора при условиях, обеспечивающих гликозилирование.
В одном воплощении условия, обеспечивающие гликозилирование, включают проведение экспрессии слитого белка в клетке-хозяине, такой как клетка млекопитающего.
Согласно настоящему раскрытию также предложено применение любого одного или более чем одного из следующих:
i) растворимый гликопротеин CD52,
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv);
vi) выделенная клетка CD52hi, способная продуцировать растворимый гликопротеин CD52;
vii) выделенная популяция клеток, содержащая множество клеток CD52hi, способных продуцировать растворимый гликопротеин CD52;
viii) среда культуры клеток или ее фракция, содержащая растворимый гликопротеин CD52, выделенный из культуры клеток, содержащей клетку по разделу vi) или популяцию клеток по разделу vii);
ix) агент, способный увеличивать уровень экспрессии растворимого гликопротеина CD52 клеткой; и
x) фармацевтическая композиция по изобретению
для подавления функции эффекторных Т-клеток и/или уменьшения иммунного ответа, такого как иммунный ответ на аутоантиген.
Согласно настоящему раскрытию также предложен способ лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса у субъекта, включающий введение терапевтически эффективного количества любого одного или более чем одного из следующих:
i) растворимый гликопротеин CD52,
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv);
vi) выделенная клетка CD52hi, способная продуцировать растворимый гликопротеин CD52;
vii) выделенная популяция клеток, содержащая множество клеток CD52hi, способных продуцировать растворимый гликопротеин CD52;
viii) среда культуры клеток или ее фракция, содержащая растворимый гликопротеин CD52, выделенный из культуры клеток, содержащей клетку по разделу vi) или популяцию клеток по разделу vii);
ix) агент, способный увеличивать уровень экспрессии растворимого гликопротеина CD52 клеткой; и
x) фармацевтическая композиция по изобретению.
В одном воплощении растворимый гликопротеин CD52, слитый белок, среду культуры клеток, агент или композицию вводят в сайт на слизистой или под кожей.
Согласно настоящему раскрытию также предложен любой один или более чем один из следующих:
i) растворимый гликопротеин CD52,
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv);
vi) выделенная клетка CD52hi, способная продуцировать растворимый гликопротеин CD52;
vii) выделенная популяция клеток, содержащая множество клеток CD52hi, способных продуцировать растворимый гликопротеин CD52;
viii) среда культуры клеток или ее фракция, содержащая растворимый гликопротеин CD52, выделенный из культуры клеток, содержащей клетку по разделу vi) или популяцию клеток по разделу vii);
ix) агент, способный увеличивать уровень экспрессии растворимого гликопротеина CD52 клеткой; и
x) фармацевтическая композиция по изобретению
для применения в лечении или предупреждении заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса.
Кроме того, согласно настоящему раскрытию предложено применение любого одного или более чем одного из следующих:
i) растворимый гликопротеин CD52,
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv);
vi) выделенная клетка CD52hi, способная продуцировать растворимый гликопротеин CD52;
vii) выделенная популяция клеток, содержащая множество клеток CD52hi, способных продуцировать растворимый гликопротеин CD52;
viii) среда культуры клеток или ее фракция, содержащая растворимый гликопротеин CD52, выделенный из культуры клеток, содержащей клетку по разделу vi) или популяцию клеток по разделу vii);
ix) агент, способный увеличивать уровень экспрессии растворимого гликопротеина CD52 клеткой; и
x) фармацевтическая композиция по изобретению в изготовлении лекарственного средства для лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса.
В одном воплощении лекарственное средство приготовлено для введения в сайт на слизистой или под кожей.
В одном примере заболевание, опосредованное функцией эффекторных Т-клеток, представляет собой аутоиммунное заболевание, такое как диабет типа I или ревматоидный артрит. В другом примере состояние, опосредованное функцией эффекторных Т-клеток, представляет собой отторжение аллотрансплантата или реакцию «трансплантат против хозяина».
Согласно настоящему раскрытию также предложен способ проведения диагностики чувствительности субъекта к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, включающий:
детектирование уровня растворимого гликопротеина CD52 в образце, взятом у субъекта; и
сравнение уровня растворимого гликопротеина CD52, детектированного в образце, взятом от субъекта, с контрольным уровнем, определенным у одного или более чем одного здорового субъекта,
в котором меньший уровень растворимого гликопротеина CD52, детектированного в образце, взятом от субъекта, по сравнению с контрольным уровнем указывает на то, что субъект имеет повышенную чувствительность к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сеписа.
Согласно настоящему раскрытию также предложен способ проведения диагностики чувствительности субъекта к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, включающий:
детектирование частоты клеток CD52hi в образце, взятом от субъекта; и
сравнение частоты клеток CD52hi, детектированных в образце, взятом от субъекта, с контрольным уровнем, определенным у одного или более чем одного здорового субъекта,
в котором меньшая частота клеток CD52hi, детектированная в образце, взятом от субъекта, по сравнению с контрольным уровнем указывает на то, что субъект имеет повышенную чувствительность к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сеписа.
Согласно настоящему раскрытию также предложен способ проведения диагностики чувствительности субъекта к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, включающий:
детектирование активности клеток CD52hi в образце, взятом от субъекта; и
сравнение активности клеток CD52hi, детектированных в образце, взятом от субъекта, с контрольным уровнем, определенным у одного или более чем одного здорового субъекта,
в котором пониженная активность клеток CD52hi, детектированная в образце, взятом от субъекта, по сравнению с контрольным уровнем, указывает на то, что субъект имеет повышенную чувствительность к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сеписа.
В одном примере частоту клеток CD52hi определяют детектированием уровня мембраносвязанного CD52 в образце путем детектирования уровня экспрессии белка CD52 в образце и/или детектирования уровня экспрессии мРНК CD52 в образце.
В одном примере образец берут у субъекта, которому вводили антиген.
В другом примере образец берут из местного сайта заболевания у субъекта.
Согласно настоящему раскрытию также предложен способ определения пригодности субъекта для включения в испытание по скринингу лекарственного средства, включающий проведение способа по изобретению и идентификацию субъекта как являющегося более подходящим для включения в испытание по скринингу лекарственного средства, если субъект имеет более низкий уровень растворимого гликопротеина CD52, меньшую частоту клеток CD52hi или пониженную активность клеток CD52hi, чем контрольный образец. Например, испытание по скринингу лекарственного средства представляет собой испытание по скринингу антидиабетического лекарственного средства.
Кроме того, согласно настоящему раскрытию также предложен способ идентификации агента, способного имитировать подавление эффекторных Т-клеток и/или подавление иммунного ответа, функцию растворимого гликопротеина CD52, включающий определение того, подавляет ли тестируемый агент функцию эффекторных Т-клеток и/или иммунный ответ.
Согласно настоящему раскрытию также предложен способ идентификации потенциального терапевтического агента для лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, включающий приведение в контакт тестируемого агента с клеткой CD52hi или популяцией клеток CD52hi и детектирование любого одного или более чем одного из уровня растворимого гликопротеина CD52, продуцируемого клеткой или популяцией клеток, частоты клеток CD52hi и/или активности клеток CD52hi и идентификацию тестируемого агента в качестве потенциального терапевтического агента для лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, если уровень растворимого гликопротеина CD52, частота клеток CD52hi и/или активность клеток CD52hi увеличивается после контакта с тестируемым агентом.
Характеристики любого описанного здесь воплощения следует принимать как относящиеся, с учетом необходимых изменений, к любым другим воплощениям, если конкретно не утверждается иное.
Настоящее раскрытие не органичивается объемом описанных здесь конкретных воплощений, которые предназначены лишь для цели приведения примера. Функционально эквивалентые продукты, композиции и способы явно находятся в пределах объема изобретения, как здесь описано.
Во всем данном описании изобретения, если конкретно не утверждается иное или контекст не требует иного, следует принимать, что ссылка на одну стадию, состав вещества, группу стадий или группу составов вещества охватывает одну и множество (т.е. одну или более чем одну) из данных стадий, составов вещества, групп стадий или групп составов вещества.
Ниже изобретение описано посредством следующих неограничивающих Примеров и со ссылкой на сопровождающие графические материалы.
КРАТКОЕ ОПИСАНИЕ СОПРОВОЖДАЮЩИХ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг 1: клоны GAD65-специфичных супрессорных Т-клеток CD4+ демонстрируют более высокую экспрессию CD52
(А) Пролиферация клона САОбб-специфичных Т-клеток в присутствии аутологичного супрессорного клона. Использованная GAD65 представляла собой рекомбинантную человеческую декарбоксилазу глутаминовой кислоты 65. Фиксированное число (25000) САйбб-специфичных клеток из клона, не являющегося Treg (3.19) сокультивировали с возрастающими количествами клеток аутологичного СА065-специфичного Treg клона (1.4) в присутствии или в отсутствие GAD65 и облученных РВМС (одноядерные клетки периферической крови) (100000) в качестве антигенпрезентирующих клеток. Поглощение 3Н-тимидина измеряли через 72 ч. Результаты (среднее плюс/минус sem (стандартная ошибка среднего) трех повторностей) являются репрезентативными для многих пар аутологичного супрессорного и несупрессорного клона, как описано ранее (Dromey et al., 2011). (Б) Активированные 6А065-специфичные супрессорные клоны имеют более высокую экспрессию CD52. Гистограммы проточной цитометрии экспрессии CD52 аутологичным ОА065-специфичным супрессорным (сплошная линия) и несупрессорным (пунктирная линия) клонами после стимуляции связанным с планшетом антителом против CD3 в течение ночи. Окрашивание изотипичным контрольным антителом показано серым. Результат является репрезентативным для 3 пар клонов от 3 индивидуумов.
Фиг. 2: высокая экспрессия CD52 является маркером активированных антигеном Т-клеток CD4+ крови с супрессорной функцией
(А) Пролиферация стимулированных столбнячным анатоксином (ТТ), сортированных FACS (флуоресцентная сортировка клеток) Т-клеток CD4+, повторно активированных ТТ, в присутствии GAD65-активированных и сортированных клеток CD4+ CD52hi или CD52lo. Активированные клетки CD4+ получали инкубированием РВМС, меченных CFSE, либо с GAD65, либо с ТТ в течение 7 суток (А1). GAD65-активированные Т-клетки CD4+ CD52hi и CD4+ CD52lo, и активированные ТТ Т-клетки CD4+ затем выделяли посредством FACS. В присутствии GAD65 пролиферация клеток, повторно активированных ТТ, подавляется GAD65-активированными клетками CD4+ CD52hi. Поглощение 3Н-тимидина измеряли в течение последних 16 ч 3-суточной культуры (А2). Результаты (среднее плюс/минус sem трех повторностей) являются репрезентативными для независимых экспериментов на клетках от 5 индивидуумов.
(Б) Секреция IFN-γ GAD65-активированными и сортированными Т-клетками CD4+ в отсутствие или в присутствии GAD65. РВМС, меченные CFSE, инкубировали с GAD65 в течение 7 суток и сортировали на Т-клетки CD4+ CD52hi и CD52lo. Сортированные клетки (5000) инкубировали в планшетах ELISpot с облученными РВМС (20000). Результаты (среднее плюс/минус sem трех повторностей) представляют многочисленные независимые эксперименты на клетках от 5 индивидуумов.
(В) Секреция IFN-γ активированными ТТ и сортированными Т-клетками CD4+ в отсутствие или в присутствии ТТ плюс/минус IL-2 (интерлейкин-2) (10 U/мл). Как и в (Б), за исключением того, что популяции CD4+ CD52hi и CD52lo сортировали из РВМС, меченных CFSE, активированных ТТ. Результаты представляют среднее плюс sem трех повторностей.
(Г) Пролиферация РВМС, исходно обедненных посредством FACS либо клетками CD4+ CD52hi, либо CD52lo до мечения CFSE и инкубации в отсутствие или в присутствии GAD65 в течение 7 суток. Результаты представляют два эксперимента.
Фиг. 3: Т-клетки CD4+ CD52hi не происходят из Т-клеток CD4+ CD25+ в состоянии покоя
Секреция IFN-γ активированными ТТ и сортированными Т-клетками CD4+ в отсутствие (незакрашенные столбики) или в присутствии (закрашенные столбики) ТТ после исходного обеднения РВМС клетками CD25hi.
Фиг. 4: активированные антигеном Т-клетки CD4+ CD52hi не различаются посредством маркеров традиционных Treg клеток CD4+ CD25+
Определенная проточной цитометрией экспрессия (A) CD25, (Б) FoxP3, (В) поверхностного и (Г) внутриклеточного CTLA-4, (Д) GITR, (Е) CD127, (Ж) CD24 и (3) CD59 на поделившихся Т-клетках CD4+ CD52hi (черная линия) и CD52lo (серая линия) после инкубации РВМС с ТТ в течение 7 суток. Окрашивание изотипическим контрольным антителом показано в виде серого закрашивания. Результаты характеризуют 5 индивидуумов.
Фиг. 5: экспрессия гена CD52 выше в Т-клетках CD4+ CD52hi относительно Т-клеток CD4+ CD52lo
Экспрессия генов в Т-клетках CD4+ CD52hi относительно CD52lo. Количественную ПЦР-ОТ (полимеразная цепная реакция, сопряженная с обратной транскрипцией) проводили на образцах РНК в тройной повторности, экстрагированной из сортированных Т-клеток CD4+ CD52hi и CD52lo, меченных CFSE, от трех индивидуумов через 7 суток после активации GAD65. Результаты выражены в виде медианы плюс интервал между квартилями.
Фиг. 6: экспрессия CD24 не выделяет Т-клетки CD4+ CD52hi с супрессорнойфункцией
Секреция IFN-γ активированными ТТ и сортированными Т-клетками CD4+ CD52lo, повторно стимулированными ТТ в присутствии стимулированных ТТ и сортированных субпопуляций CD52 и CD24. Результаты представляют собой среднее плюс sem трех повторностей.
Фиг. 7: контакт клетка-клетка не требуется для подавления Т-клетками CD4+ CD52hi
Фиг. 8: высвобождение растворимого CD52 объясняет подавление Т-клетками CD4+ CD52hi
(A) Иммуноблоттинг сред, кондиционированных GAD65-активированными Т-клетками CD4+ CD52hi или CD52lo, затем повторно активированными GAD65. РВМС, меченные CFSE, инкубировали с GAD65 в течение 7 суток и сортировали на Т-клетки CD4+ CD52hi и CD52lo. Сортированные клетки повторно активировали GAD65, и среды собирали через 24 часа. Среды 10-кратно концентрировали, фракционировали SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), переносили на PDVF мембрану и осуществляли блоттинг с поликлональным антителом кролика к CD52. Указана приблизительная молекулярная масса нативного растворимого CD52.
(Б) Среды для иммуноблоттинга, кондиционированные РВМС, активированными ТТ, плюс/минус ингибитор фосфолипазы С U73122. РВМС, меченные CFSE, инкубировали с ТТ, и среды, отобранные через 1 ч, обрабатывали так же, как и в (А).
(B) Эффект ингибитора фосфолипазы С на подавление Т-клеток CD4+ CD52hi, активированных ТТ. РВМС, меченные CFSE, инкубировали с ТТ в течение 7 суток и сортировали на Т-клетки CD4+ CD52hi и CD52lo, которые затем инкубировали совместно (по 5000 каждых) в планшетах ELISpot с облученными РВМС (20000) и ТТ плюс/минус ингибитор фосфолипазы С U73122. Результаты представляют собой среднее плюс sem трех повторностей. В отсутствие ТТ не было эффекта U73122.
(Г) Эффект антитела к углеводной группировке CD52 на подавление посредством Т-клеток CD4+ CD52hi, активированных ТТ. Методики были такими же, как и в (В), за исключением того, что клетки в анализе ELISpot инкубировали с или без ТТ и либо с 10 мкг/мл антитела против CD52 (CF1D12), либо изотипического контрольного (lgG3) моноклонального антитела. Результаты (среднее плюс/минус sem) характеризуют три независимых эксперимента.
Фиг. 9: растворимый CD52, продуцируемый из клеток Дауди, непосредственно подавляет пролиферацию и эффекторную функцию Т-клеток
(A) Иммуноблоттинг сред, кондиционированных линиями клеток. Среды 10-кратно концентрировали, фракционировали SDS-PAGE, переносили на PDVF мембрану и осуществляли блоттинг с поликлональным антителом кролика к CD52.
(Б) Подавление пролиферации Т-клеток средой, кондиционированной клетками Дауди. РВМС (200000 клеток) культивировали в течение 7 суток в IMDM (среда Дульбекко, модифицированная по способу Исков), содержащей 20% среды, кондиционированной клетками Дауди, с ТТ, и либо антитело против CD52 (CF1D12), либо изотипическое контрольное антитело (10 мкг/мл). Для обеднения растворимым CD52 среду от клеток Дауди инкубировали в течение ночи с поликлональным кроличьим антителом против CD52 (1 мкг/мл среды), с последующим осаждением белок G-сефарозой в течение 1 ч при 4°С. Результаты (среднее плюс/минус sem) характеризуют три независимых эксперимента.
Фиг. 10: конструкции ДНК для экспрессии в лентивирусном векторе
SigP - сигнальный пептид; ECD - внеклеточный домен; Strep2 - метка для очистки, кодирующая 8 аминокислот, которые связываются со Стрел-тактином, стрептавидином, сконструированным специальным способом.
Фиг. 11: растворимый слитый белок CD52 прямо подавляет пролиферацию и эффекторную функцию Т-клеток
Подавление пролиферации Т-клеток рекомбинантным CD52-Fc. РВМС (200000) культивировали с ТТ в течение 7 суток (А), и очищенные Т-клетки CD4+ (20000) с антителом против CD3 (100 нг/мл) и антителом против CD28 (200 нг/мл) в течение 48 ч (Б) с 4-кратным избытком облученных РВМС в 200 мкл круглодонных лунках в присутствии рекомбинантного CD52-Fc или контрольного белка Fc в указанных концентрациях. Поглощение 3Н-тимидина измеряли на протяжении последних 16 ч инкубации. Результаты (среднее плюс/минус sem трех повторностей) характеризуют шесть независимых экспериментов.
(B) Подавление секреции цитокинов рекомбинантными CD52-FC. Среды от РВМС, активированных ТТ, в (В) плюс/минус 3,3 мкМ белков CD52-Fc или Fc отбирали после 48 часов инкубации и анализировали на цитокины посредством мультиплексного набора шариков.
(Г) Ослабленное подавление посредством CD52-FC после отщепления N-связанного углевода. CD52-FC (20 мкг) инкубировали с ПНГазой F (пептид-N-гликозидаза F) (1000 единиц) или без нее в 20 мкл PBS в течение 1 ч при 37°С, и реакцию завершали нагреванием при 75°С в течение 10 мин. РВМС инкубировали с ТТ и обрабатывали или не обрабатывали CD52-FC (конечная концентрация 2,5 мкМ) в течение 7 суток при 37°С, и затем измеряли поглощение 3Н-тимидина так же, как и в (В). На верхней панели показано определение уменьшения размера CD52-Fc после обработки ПНГазой F посредством SDS-PAGE и окрашивания кумасси.
Фиг. 12: связывание углевода CD52 с Siglec-10 требуется для эффекторной функции растворимого CD52
(A) Подавление активации Т-клеток посредством CD52-Fc плюс/минус обработка нейраминидазой. CD52-FC (3,3 мкМ) инкубировали с нейраминидазой (1 единица) или только с буфером-носителем в 20 мкл в течение 30 мин при 37°С. РВМС затем инкубировали с ТТ плюс/минус CD52-Fc (конечная концентрация 0,33 мкМ), обработанное или необработанное нейраминидазой, в 48-луночном планшете в течение 1 ч при 37°С перед переносом неприкрепленных клеток на планшет ELISpot и проявлением на пятна IFN-γ после 24 ч при 37°С.
(Б) Экспрессия Siglec-10 на человеческих Т-клетках после активации Т-клеток. Гистограммы проточной цитометрии экспрессии Siglec-10 на Т-клетках CD4+ после инкубации РВМС с ТТ или растворимым антителом против CD3 в течение 4 суток.
(B) Подавление функции Т-клеток CD52-Fc при совместной инкубации с антителом против Siglec-10. РВМС инкубировали в планшете ELISpot с ТТ и CD52-Fc (3,4 мкМ) и разными концентрациями подвергнутого аффинной очистке антитела козы к внеклеточному домену Siglec-10 или Fc (0,34 мкМ) плюс/минус антитело перед переносом неприкрепленных клеток на планшет ELISpot на 24 ч для проявления пятен IFN-γ.
(Г) Подавление функции Т-клеток посредством CD52-FC при совместной инкубации с растворимым рекомбинантным Siglec-10-Fc. РВМС инкубировали в 48-луночном планшете с ТТ и CD52-FC (3,4 мкМ) и разными концентрациями рекомбинантного Siglec-10-Fc перед переносом неприкрепленных клеток на планшет ELISpot на 24 ч для проявления пятен IFN-γ.
(Д) Блокировка Siglec-10, но не других типов Siglec, уменьшает подавление Т-клеток посредством CD52-FC. Т-клетки CD4+ (20000) инкубировали в тройной повторности в лунках планшета ELISpot при 37°С с ТТ совместно с CD52-Fc или Fc (3,4 мкМ каждого) и антителами против человеческого Siglec (10 мкг/мл каждого) или рекомбинантным человеческим Siglec 2-Fc (20 мкг/мл), как указано. Через 20 ч лунки промывали и проявляли на пятна IFN-γ.
Фиг. 13: CD52-Fc не влияет на стимулирующую способность очищенных дендритных клеток крови в отношении Т-клеток
Сортированные FACS DC (дендритные клетки) человеческой крови CD1b/c+ предынкубировали с белком CD52-FC или Fc, дважды промывали и сокультивировали с аллогенными Т-клетками CD4+, меченными CFSE, в течение 6 суток. Частоту делящихся Т-клеток CD4+, определенных как CFSElo, определяли проточной цитометрией. Результат характеризует два независимых эксперимента с разными донорами. Аналогичные результаты были получены для плазмацитоидных DC CD304+ и для моноцитов CD14+ (данные не показаны).
Фиг. 14: перенос спленоцитов, обедненных CD52hi, индуцирует быстрое начало диабета у NOD.SCID мышей
Общие спленоциты от NOD мышей дикого типа с обеднением или имитацией обеднения клетками CD52hi внутривенно инъецировали (А) 8-недельным мышам RIP.B7/NOD.SCID (2×106 клеток) и (Б) облученным (750 рад (7,5 Дж/кг)) 8-недельным самцам NOD мышей (1,2×107 клеток). Мышей отслеживали посредством измерения глюкозы в моче дважды в неделю с использованием Diastix (Bayer), и диабет подтверждали измерением уровня глюкозы в крови, превышающего 14 мМ, в следующие друг за другом сутки. Результаты показывают процентную долю выживания во времени после переноса соответствующих популяций клеток.
Фиг. 15: Т-клетки CD3+, обедненные CD52hi, ускоряют начало диабета у облученных NOD мышей
Облученным (750 рад (7,5 Дж/кг)) 8-недельным самцам NOD мышей инъецировали 1,2×107 спленоцитов или спленоцитов CD3+, обедненных CD52hi, полученных от 10-недельных недиабетических самок NOD мышей. Мышей отслеживали посредством измерения глюкозы в моче дважды в неделю с использованием Diastix (Bayer), и диабет подтверждали измерением уровня глюкозы в крови, превышающего 14 мМ, следующие друг за другом сутки. Результаты показывают (А) процентную долю выживания во времени после переноса соответствующих популяций клеток и (Б) бальный показатель инсулита (n равно 4 на группу) через 4 недели.
Фиг. 16: частота Т-клеток CD4+ CD52hi, генерированных в ответ на стимуляцию GAD65, снижается при диабете типа 1
Доля Т-клеток CD4+ CD52hi, размноженных из РВМС в ответ на (A) GAD65 и (Б) ТТ, показана для индивидуумов в следующих категориях: Pre-T1D -подвергающиеся риску развития диабета типа 1; T1D - имеющие диабет типа 1; здоровые - не имеющие заболевания молодые взрослые с HLA (антиген лейкоцитов человека) DR3 и/или DR4; T2D - диабет типа 2. Горизонтальная планка представляет собой медиану для каждой группы. Показанные общие значения Р из диперсионного анализа определяли критерием Крускала-Уоллиса; критерий множественных сравнений Данна затем выявил значимые различия между и Pre-T1D, и T1D по сравнению со здоровыми или T2D при Р меньше 0,05.
Фиг. 17: подавление посредством клеток CD4+ CD52hi, генерированных в ответ на GAD65, снижается при доклиническом T1D
Секреция IFN-γ активированными ТТ или GAD65 и сортированными Т-клетками CD4+ в отсутствие (незакрашенные) или в присутствии (закрашеные) антигена. Результаты (среднее плюс sem трех повторов) характеризуют эксперименты на клетках от шести индивидуумов, подверженных риску диабета типа 1, с аутоантителами против островковых клеток.
Фиг. 18: лечение CD52-Fc обращает гипергликемию у NOD мышей с недавно начавшимся диабетом
Уровни глюкозы в крови у самок NOD мышей отслеживали путем еженедельного тестирования на глюкозу в моче, и диабет диагностировали у мышей с положитеьным анализом мочи по концентрации глюкозы в крови, превышающей 14 мМ. Как только подтверждали гипергликемию, мышам в.б. (внутрибрюшинно) давали 20 мкг CD52-FC или Fc, шесть доз через сутки, и затем отслеживали их концентрации глюкозы в крови дважды в неделю. Результаты показаны для двух пар мышей, которые получали либо CD52-FC (А), либо контроль в виде Fc (Б).
Фиг. 19: развитие диабета у NOD.SCID мышей после переноса от диабетических NOD мышей спленоцитов, обработанных ex vivo hCD52-Fc или Fc
Спленоциты от диабетических NOD мышей, обработанные рекомбинантным человеческим CD52-FC или Fc, инъецировали мышам NOD.SCID. Спленоциты от самок диабетических мышей выделяли и инкубировали либо с рекомбинантным hCD52-Fc, либо с белком Fc в «буфере для CD52». Клетки ресуспендировали и инъецировали самцам NOD.SCID мышей (6 на группу; см. Пример 18 Методов).
Фиг. 20: человеческий CD52-Fc подавляет пролиферацию мышиных специфичных в отношении овальбумина (Ova) трансгенных по TCR (рецептор Т-клетки) Т-клеток CD4 (ОТ-II)
Спленоциты (1×105) от 10-недельных самок мышей ОТ-II в течение 3 суток инкубировали в круглодонных 96-луночных планшетах в 200 мл среды RPMI-1640, содержащей 5% FCS (фетальная коровья сыворотка) и указанные концентрации белка или пептида Ova, или антитела против CD3 (клон 2С-11) и рекомбинантный человеческий белок CD52-FC или Fc. Поглощение 3Н-тимидина измеряли на протяжении последних 16 ч культуры. Результаты представляют собой среднее плюс/минус sem трех повторностей.
Фиг. 21: идентификация CD52 в образцах человеческой семенной жидкости посредством ELISA
Показано поглощение при 450 нм для растворимого CD52 в серийных разведениях образцов семенной жидкости (n равно 26).
Фиг. 22: CD52, происходящий из семенной жидкости, подавляет пролиферацию человеческих Т-клеток
Эффект на пролиферацию Т-клеток (индекс клеточных делений; CDI), рассчитанный из разведения красителя CFSE, в ответ на столбнячный анатоксин (ТТ, 5 Lfu/мл) показан для двух образцов семенной жидкости (при разведении 1:20) без иммуннообеднения или с обеднением контрольным IgG ('Octagam') или IgG против CD52 (Campath).
Фиг. 23: пролиферация Т-клеток (разведение красителя CFSE) в ответ на столбнячный анатоксин: эффект семенной жидкости плюс/минус блокирующего антитела к CD52
Эффект на пролиферацию Т-клеток (CDI, рассчитанный из разведения красителя CFSE) в ответ на столбнячный анатоксин (ТТ, 5 Lfu/мл) показан для образцов семенной жидкости (1:20) плюс/минус блокирующее антитело CF1D12 (20 мкг/мл).
Фиг. 24: hCD52-Fc подавляет секрецию IL-1β клетками ТНР1 в ответ на LPS
Клетки ТНР-1 инкубировали с разными дозами CD52-FC или контроля в виде Fc в присутствии LPS, собирали среду, и концентрацию IL-1β измеряли ELISA.
Фиг. 25: hCD52-Fc подавляет секрецию IL-1β клетками ТНР1 в ответ на Pam3CSK
Клетки ТНР-1 инкубировали с разными дозами CD52-Fc или контроля в виде Fc в присутствии Pam3CSK - агониста TLR-2, собирали среды, и концентрацию IL-1β измеряли ELISA.
Фиг. 26: hCD52-Fc (50 мкг/мл) подавляет секрецию IL-1B дифференцированными клетками ТНР1 в ответ на квасцы
Клетки ТНР-1 подвергали дифференциации с форбол-12-миристат-13-ацетатом (РМА), промывали и инкубировали с CD52-FC или контролем в виде Fc. Среду собирали, и концентрацию IL-1β измеряли ELISA.
Фиг. 27: mCD52-Fc подавляет секрецию IL-1β дендритными клетками, происходящими из костного мозга мыши в ответ на целый ряд врожденных иммунных стимулов
Дендритные клетки, происходящие из костного мозга (BMDC), от мышей С57/В6 инкубировали с мышиным CD52-FC или PBS (контроль) в присутствии LPS, CPG или Listeria monocytogenes, примировали LPS и затем стимулировали известными воспалительными агонистами - уратом мононатрия (MSU), квасцами и нигерицином. Концентрации цитокинов в средах измеряли мультиплексным анализом на цитокины.
Фиг. 28: обработка нейраминидазой A. ureafaciens отменяет подавляющий эффект mCD52-Fc на индуцированную LPS продукцию IL-1β клетками ТНР-1
Клетки ТНР-1 инкубировали с mCD52-Fc, обработанным нейраминидазой или буфером для реакции, в присутствии LPS. Среды собирали, и концентрацию IL-1β измеряли посредством ELISA.
Фиг. 29: обработка ПНГазой-F для удаления N-связанного олигосахарида отменяет подавляющий эффект hCD52-Fc на индуцированную LPS секрецию IL-1β клетками ТНР-1
Человеческий CD52-Fc (300 мкг), обработанный или не обработанный ПНГазой F для удаления N-связанного олигосахарида, использовали для обработки клеток ТНР-1 в присутствии LPS. Среды собирали, и концентрацию IL-1β измеряли ELISA.
ЛЕГЕНДА К ПЕРЕЧНЮ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 мРНК транскрипт человеческого CD52 (эталонная последовательность NCBI (Национальный центр биотехнологической информации): NM_001803.2)
SEQ ID NO: 2 Аминокислотная последовательность человеческого CD52
SEQ ID NO: 3 12-аминокислотный растворимый пептид человеческого CD52
SEQ ID NO: 4 Ортологичный растворимый пептид CD52 обезьяны
SEQ ID NO: 5 Ортологичный растворимый пептид CD52 мыши
SEQ ID NO: 6 Ортологичный растворимый пептид CD52 крысы
SEQ ID NO: 7 Ортологичный растворимый пептид CD52 собаки
SEQ ID NO: 8 F (прямой) праймер CD52
SEQ ID NO: 9 R (обратный) праймер CD52
SEQ ID NO: 10 F праймер FOXP3
SEQ ID NO: 11 R праймер FOXP3
SEQ ID NO: 12 F праймер CTLA-4
SEQ ID NO: 13 R праймер CTLA-4
SEQ ID NO: 14 F праймер GITR
SEQ ID NO: 15 R праймер GITR
SEQ ID NO: 16 F праймер CD127
SEQ ID NO: 17 R праймер CD127
SEQ ID NO: 18 прямой праймер IL-2α
SEQ ID NO: 19 обратный праймер IL-2α
SEQ ID NO: 20 прямой праймер IL-27β
SEQ ID NO: 21 обратный праймер IL-27β
SEQ ID NO: 22 прямой праймер IL-12α
SEQ ID NO: 23 обратный праймер IL-12α
SEQ ID NO: 24 праймер 1F1
SEQ ID NO: 25 праймер 1R1
SEQ ID NO: 26 праймер 1F2
SEQ ID NO: 27 праймер 1R2
SEQ ID NO: 28 праймер 2F
SEQ ID NO: 29 праймер 2R1
SEQ ID NO: 30 праймер 2R2
SEQ ID NO: 31 прямой праймер CD52
SEQ ID NO: 32 обратный праймер CD52
SEQ ID NO: 33 прямой праймер IL-2
SEQ ID NO: 34 обратный праймер IL-2
SEQ ID NO: 35 прямой праймер IL-4
SEQ ID NO: 36 обратный праймер IL-4
SEQ ID NO: 37 прямой праймер IL-10
SEQ ID NO: 38 обратный праймер IL-10
SEQ ID NO: 39 прямой праймер IL-13
SEQ ID NO: 40 обратный праймер IL-13
SEQ ID NO: 41 прямой праймер FoxP3
SEQ ID NO: 42 обратный праймер FoxP3
SEQ ID NO: 43 прямой праймер CD127
SEQ ID NO: 44 обратный праймер CD127
SEQ ID NO: 45 прямой праймер CTLA-4
SEQ ID NO: 46 обратный праймер CTLA-4
SEQ ID NO: 47 прямой праймер FASLG
SEQ ID NO: 48 обратный праймер FASLG
SEQ ID NO: 49 прямой праймер TGFM
SEQ ID NO: 50 обратный праймер TGFM
SEQ ID NO: 51 прямой праймер TGFb2
SEQ ID NO: 52 обратный праймер TGFb2
SEQ ID NO: 53 прямой праймер IFNg
SEQ ID NO: 54 обратный праймер IFNg
SEQ ID NO: 55 прямой праймер IL-12 альфа
SEQ ID NO: 56 обратный праймер IL-12 альфа
SEQ ID NO: 57 прямой праймер Ebi3
SEQ ID NO: 58 обратный праймер Ebi3
SEQ ID NO: 59 прямой праймер RARA
SEQ ID NO: 60 обратный праймер RARA
SEQ ID NO: 61 прямой праймер GITR
SEQ ID NO: 62 обратный праймер GITR
SEQ ID NO: 63 прямой праймер GRANZMB
SEQ ID NO: 64 обратный праймер GRANZMB
SEQ ID NO: 65 прямой праймер ALDH1A2
SEQ ID NO: 66 обратный праймер ALDH1A2
SEQ ID NO: 67 прямой праймер ACTIN
SEQ ID NO: 68 обратный праймер ACTIN
SEQ ID NO: 69 последовательность человеческого белка Siglec-10 (№ доступа GenBank AF310233.1)
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Общие методики и определения
Если конкретно не определено иначе, следует принимать, что все используемые здесь технические и научные термины имеют такое значение, которое обычно понятно обычному специалисту в данной области (например в области культуры клеток, молекулярной генетики, иммунологии, иммуногистохимии, химии белка и биохимии).
Если не указано иное, методики, связанные с рекомбинантным белком, культурой клеток и иммунологические методики, используемые в настоящем изобретении, представляют собой стандартные процедуры, хорошо известные специалистам в данной области. Такие методики описаны и объяснены во всей литературе в таких источниках как J. Perbal, A Practical Guide to Molecular Cloning, John Wiley and Sons (1984), J. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press (1989), T.A. Brown (editor), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991), D.M. Glover and B.D. Hames (editors), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 and 1996) и F.M. Ausubel et al., (editors), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-lnterscience (1988, включая все исправленные и дополненные издания до настоящего момента), Ed Harlow and David Lane (editors) Antibodies: A Laboratory Manual, Cold Spring Harbour Laboratory, (1988) и J.E. Coligan et al. (editors) Current Protocols in Immunology, John Wiley & Sons (включая все исправленные и дополненные издания до настоящего момента).
Следует понимать, что термин «и/или», например, «X и/или Y» означает либо «X и Y», либо «X или Y», и следует принимать, что он обеспечивает недвусмысленную поддержку обоим значениям или для одного из значений.
Термин «примерно» в том виде, как он здесь используется, если не указано противоположное, относится к плюс/минус 20%, более предпочтительно плюс/минус 10%, более предпочтительно плюс/минус 5% относительно указанного значения. Во избежание сомнений, термин «примерно» с последующим указанным значением следует интерпретировать как также охватывающий само точное указанное значение (например «примерно 10» также охватывает точно 10).
Будет понятно, что во всем данном описании слово «включать» или вариации, такие как «включает» или «включающий», подразумевает включение указанного элемента, целого числа или стадии, или группы элементов, целых чисел или стадий, но не исключение любого другого элемента, целого числа или стадии, или группы элементов, целых чисел или стадий.
Термин «иммунный ответ» в том виде, как он здесь используется, имеет его обычное значение в данной области и включает как гуморальный, так и клеточный иммунитет. Иммунный ответ может быть опосредован одним или более чем одним из: активации Т-клеток, активации В-клеток, активации природных клеток-киллеров, активации антигенпрезентирующих клеток (например, В-клеток, DC (дендритная клетка), моноцитов и/или макрофагов), продукции цитокинов, продукции хемокинов, экспрессии специфичных маркеров поверхности клетки, в частности, экспрессии костимулирующих молекул. В предпочтительном воплощении иммунный ответ, который подавляется с использованием способов по изобретению, представляет собой по меньшей мере функцию эффекторных Т-клеток посредством уменьшения выживания, активности и/или пролиферации данного типа клеток. В другом предпочтительном воплощении иммунный ответ, который подавляется с использованием способов по изобретению, представляет собой по меньшей мере одно или более чем одно из функции моноцитов, макрофагов или дендритных клеток посредством уменьшения выживания, активности и/или пролиферации одного или более чем одного из данных типов клеток. В другом предпочтительном воплощении иммунный ответ подавляется в такой степени, которая индуцирует толерантность к антигену, такому как аутоантиген.
Термин «толерантность» в том виде, как он здесь используется, относится к состоянию иммунологической неотвечаемости на конкретный антиген или группу антигенов, на которые обычно отвечает субъект. Иммунологическая толерантность достигается при условиях, при которых подавляется иммунная реакция, и она не является просто отсутствием иммунного ответа.
Термины «осуществление лечения», «лечить» или «лечение» в том виде, как они здесь используются, включают введение терапевтически эффективного количества агента, достаточного для уменьшения или устранения по меньшей мере одного симптома заболевания.
Термины «осуществление предупреждения», «предупреждать» или «предупреждение» в том виде, как они здесь используются, включают введение терапевтически эффективного количества агента, достаточного для предупреждения проявления по меньшей мере одного симптома заболевания.
Термин «осуществление подавления» в том виде, как он здесь используется, включает уменьшение на любую поддающуюся количественной оценке величину.
Термин «субъект» в том виде, как он здесь используется, относится к животному, например, к млекопитающему. В предпочтительном воплощении субъект представляет собой млекопитающее, например, человека. Другие предпочтительные воплощения включают домашний скот как, например, лошадей, крупный рогатый скот, овец и коз, а также животных-спутников, таких как кошки и собаки.
Термин «хозяин» в том виде, как он здесь используется, относится к любому организму, из которого можно выделять растворимый CD52 или в котором можно продуцировать растворимый CD52 любыми способами. Хозяин может представлять собой целый организм или может быть клеткой, полученной из него. Хозяин может представлять собой животное, например, млекопитающее. В предпочтительном воплощении хозяин представляет собой млекопитающее, например, человека. Другие предпочтительные хозяева включают мышей, крыс, обезьян, хомяков, морских свинок, кроликов и любое животное или клетку, которые могут служить в качестве подходящего хозяина, из которого можно выделять растворимый CD52 или в котором можно продуцировать растворимый CD52.
Термины «связанный», «присоединенный», «конъюгированный» или их вариации в том виде, в котором они здесь используются, используются в широком смысле для отнесения к любой форме ковалентной или нековалентной ассоциации, которая связывает один элемент с другим в течение любого периода времени.
Растворимый CD52
Согласно настоящему раскрытию впервые описано подавление иммунных клеток, таких как эффекторные Т-клетки, моноциты и дендритные клетки, фрагментом растворимого гликопротеина CD52. CD52 представляет собой поверхностный, заякоренный гликозилфосфатидилинозитолом (GPI) гликопротеин, присутствующий на большинстве лимфоидных клеток, исходно признанный в качестве мишени связывающих комплемент моноклональных антител САМРАТН, используемых терапевтически для обеднения лимфоцитами. (Treumann era/., 1995; Xia et al., 1991; Hale, 2001). мРНК транскрипт человеческого гена CD52 показан в SEQ ID NO: 1, и транслированная аминокислотная последовательность показана в SEQ ID NO: 2. Зрелый CD52, связываемый его GPI якорем, содержит только 12 аминокислот и концевой углевод, связанный с acnaparnHOM(N).
Если не утверждается иное, термины «растворимый гликопротеин CD52», «растворимый CD52», «растворимый гликопротеин» и их вариации используются здесь взаимозаменяемо.
Заякоренный в мембране CD52 можно отщеплять (например, ферментативно) с высвобождением растворимого пептидного фрагмента, содержащего аминокислотную последовательность GQNDTSQTSSPS (SEQ ID NO: 3). Раскрытый здесь растворимый гликопротеин CD52 может содержать аминокислотную последовательность, по меньшей мере на 60% идентичную аминокислотной последовательности SEQ ID NO: 3 или по меньшей мере на 60% идентичную аминокислотной последовательности других известных ортологичных последовательностей фрагментов растворимого CD52. Таким образом, настоящее раскрытие охватывает ортологичные последовательности растворимого пептидного фрагмента CD52. Такие последовательности включают последовательность обезьяны SQNATSQSSPS (SEQ ID NO: 4), последовательность мыши GQATTAASGTNKNSTSTKKTPLKS (SEQ ID NO: 5), последовательность крысы GQNSTAVTTPANKAATTAAATTKAAATTATKTTTAVRKTPGKPPKA (SEQ ID NO: 6), последовательность собаки GNSTTPRMTTKKVKSATPA (SEQ ID NO:7) и другие ортологичные последовательности, легко идентифицируемые из известных полипептидных и полинуклеотидных последовательностей CD52, но не ограничиваются ими.
Процентную долю идентичности с любой раскрытой здесь аминокислотной или полинуклеотидной последовательностью можно определять способами, известными в данной области. Например, аминокислотные и полинуклеотидные последовательности можно сравнивать вручную или с использованием инструментов сравнения и идентификации последовательности на основе компьютера, в которых используются такие алгоритмы, как BLAST (инструмент поиска основного местного выравнивания; Altschul er al., 1993); см. также www.ncbi.nlm.nih.gov/BLAST/), способ выравнивания Clustal (Higgins and Sharp, 1989) и других, в которых могут быть выбраны подходящие параметры для каждого конкретного сравнения последовательностей, как было бы понятно специалисту в данной области. Аминокислотная последовательность пептидной части раскрытого здесь гликопротеина может быть по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% идентичной или по меньшей мере на 99% идентичной любой одной или более чем одной из аминокислотных последовательностей, идентифицированных в SEQ ID NO: 3, 4, 5, 6 или 7. Например, аминокислотная последовательность пептидной части раскрытого здесь гликопротеина может быть на 100% идентичной любой из аминокислотных последовательностей, идентифицированных в SEQ ID NO: 3, 4, 5, 6 или 7.
Выделенный растворимый гликопротеин CD52 можно использовать для получения фармацевтических композиций по изобретению. Термин «выделенный» используется здесь для определения выделения растворимого гликопротеина CD52, так что он присутствует в подходящей форме для применения в фармацевтической композиции. Таким образом, раскрытый здесь гликопротеин выделяют из других компонентов клетки-хозяина или жидкости, или экспрессионной системы в той степени, которая требуется для последующего приготовления гликопротеина в виде фармацевтической композиции. Выделенный гликопротеин, следовательно, предложен в форме, которая по существу не содержит других компонентов клетки-хозяина (например, белков), которые могут мешать фармацевтическому эффекту гликопротеина. Таким образом, выделенный гликопротеин может не содержать или по существу не содержать вещества, с которым он ассоциирован в природе, такого как другие гликопротеины, полипептиды или нуклеиновые кислоты, с которыми он находится в его природном окружении или в окружении, в котором его получают (например, в культуре клеток), когда такое получение осуществляется методикой генной инженерии, которую воплощают на практике in vitro или in vivo. Растворимый гликопротеин можно выделять из клетки-хозяина или жидкости, или экспрессионной системы способами, известными в данной области.
Термин «растворимый» используется здесь для определения пептида или гликопротеина, который не связан с клеточной мембраной. Растворимый пептид или гликопротеин может иметь способность к свободному перемещению в любом растворителе или жидкости, такой как жидкость организма. Например, растворимый пептид или гликопротеин может иметь способность к циркуляции в крови.
Углевод может представлять собой любую углеводную группировку, присоединенную к фрагменту растворимого пептида CD52. Например, углевод может представлять собой любую углеводную группировку, для которой было обнаружено, что она присоединена к внеклеточной части белка CD52 у хозяина. Таким образом, углевод может представлять собой любой углевод, способный к присоединению к внеклеточной части белка CD52 посредством реакции гликозилирования, для которой известно, что она идет у хозяина.
Углеводные группировки, присутствующие на гликопротеине CD52, встречающемся в природе, можно идентифицировать известными способами, такими как способы, описанные в Schroter et al. (1999). Такие углеводные группировки можно идентифицировать у гликопротеинов CD52, присутствующих в любой клетке-хозяине, экспрессирующей CD52, и обобенно в лимфоцитах, таких как Т-клетки CD4+ или CD8+, моноцитах или клетках половых путей, таких как клетки спермы или клетки эпидидимальных протоков. Таким образом, точную структуру углеводной группировки можно определять посредством применения таких методов, как масс-спектрометрия (например, времяпролетная масс-спектрометрия с лазерной ионизацией и десорбцией из жидкой матрицы (MALDI-TOF), анионообменная хроматография на Mono-Q, анионообменная хроматография при высоком рН (HPAEC-PAD), анализ метилирования, расщепление эндо-β-галактозидазой и другими методами. N-гликаны можно отделять от встречающегося в природе гликопротеина CD52 с использованием известных расщепляющих ферментов, таких как пептид-N4-(N-ацетил-β-D-глюкозаминил)аспарагинамидаза F («ПНГаза F» из Flavobacterium meningosepticum, рекомбинантная из Escherichia coli; доступная от коммерческих поставщиков, таких как Roche). N-гликаны могут быть выделены для дальнейшей характеризации с использованием известных хроматографических методов, таких как хроматография с С8-обращенной фазой. В одном примере углевод может содержать один или более чем один би-, три- или тетраантенный сахар, который может быть сиалированным на конце. Например, углевод может содержать один или более чем один тетраантенный сахар. Сахара могут быть разветвленными или неразветвленными. Сахара могут содержать проксимальную фукозу. Таким образом, углевод может быть фукозилированными. Сахара могут содержать один или более чем один N-ацетиллактозаминный повтор. Таким образом, сахара могут содержать полилактозаминные элементы. Кроме того, сахара могут содержать маннозное ядро.
Углевод может иметь любую одну или более чем одну структуру, описанную в Treumann et а/. (1995). Таким образом, например, углевод может иметь любую из следующих структур:
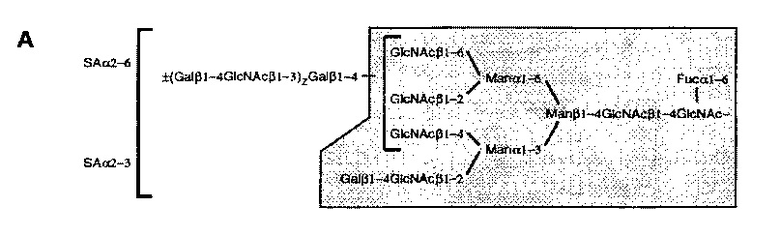
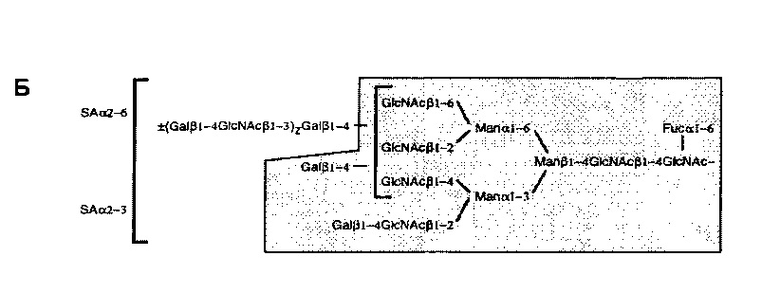
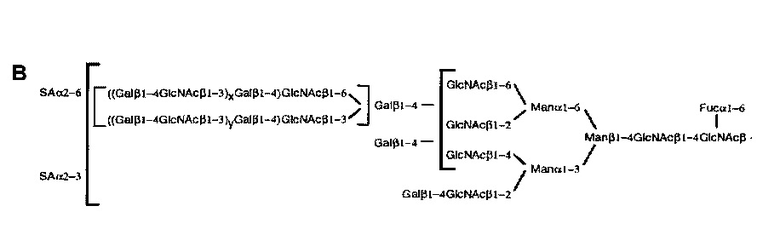
Таким образом, углевод может содержать одну или более чем одну сиаловую кислоту. Одна или более чем одна сиаловая кислота может быть расположена в любой части углевода. Например, одна или более чем одна сиаловая кислота может представлять собой терминальную сиаловую кислоту. В одном конкретном примере углевод может содержать терминальную α2-6 сиаловую кислоту. Таким образом, углевод может содержать одну или более чем одну поверхностную а2-6-сиалиллактозную группу. Одна или более чем одна сиаловая кислота может быть присоединена к галактозе в р1-4 связи с N-ацетилглюкозамином.
В настоящем раскрытии продемонстрировано то, что растворимый гликопротеин CD52 оказывает его подавляющий эффект, по меньшей мере частично, через связывание с lg-подобным лектином-10, связывающим сиаловую кислоту (Siglec-10), трансмембранным рецептором поверхности клетки и членом надсемейства иммуноглобулинов, несущим два цитоплазматических ингибирующих мотива иммунорецептора на основе тирозина (ITIM) (Munday et al., 2001; Crocker et al., 2007). Таким образом, раскрытый здесь растворимый гликопротеин может иметь способность к связыванию с Siglec-10. Например, раскрытый здесь растворимый гликопротеин может содержать углеводную группировку, способную связываться с Siglec-10. В одном примере углеводная группировка содержит одну или более чем одну поверхностную α2-6- или α2-3-сиалиллактозную группу, которая способна связываться с Siglec-10. В качестве альтернативы, углеводная группировка может содержать любые другие поверхностные группы, которые способны связываться с Siglec-10.
Раскрытый здесь растворимый гликопротеин может быть способен к связыванию с Siglec-10, происходящим из любого вида. Например, раскрытый здесь растворимый гликопротеин может быть способен к связыванию с Siglec-10, происходящим из любого вида млекопитающих. Предпочтительно раскрытый здесь растворимый гликопротеин способен к связыванию с человеческим Siglec-10. Полипептидная последовательность человеческого Siglec-10 определена в Munday et al. (2001), в № доступа GenBank AF310233.1 и в SEQ ID NO: 69.
Раскрытый здесь растворимый гликопротеин может быть способен к осуществлению сигнализации через рецептор Siglec-10. Таким образом, раскрытый здесь растворимый гликопротеин может быть способен к связыванию с Siglec-10 в любой степени, достаточной для осуществления сигнализации через рецептор Siglec-10. Таким образом, точный уровень связывания с Siglec-10 может варьировать. В данной области известны способы определения того, способен ли данный гликопротеин связываться с Siglec-10, и определения того, способен ли данный гликопротеин осуществлять сигнализацию через рецептор Siglec-10.
Другими примерами N-связанного углевода CD52, который может содержать раскрытый здесь гликопротеин, являются углеводы, происходящие или которые могут происходить из гликопротеинов CD52 лимфоцитов хозяина или гликопротеинов CD52 клеток половых путей.
Будет понятно, что из-за сложной природы многих встречающихся в природе углеводных группировок, для которых известно, что они связаны с внеклеточной белковой частью человеческого CD52, и многих вариаций данных структур, которые могут возникать из-за варьирующих картин гликозилирования, точная природа углеводной группировки, присутствующей в раскрытом здесь гликопротеине, может варьировать. Как заявлено выше, доступны способы точной идентификации конкретных углеводных группировок из встречающихся в природе гликопротеинов CD52. Кроме того, целый ряд разных углеводных группировок может быть добавлен к растворимому пептидному фрагменту CD52 путем проведения экспрессии CD52 при различных условиях гликозилирования. Например, раскрытый здесь растворимый гликопротеин можно экспрессировать в и/или выделять из клеток лимфоцитов-хозяев, моноцитов или клеток половых путей-хозяев (например, клеток спермы или клеток эпидидимальных протоков), или семенной жидкости, и, следовательно, в результате, он может содержать разные углеводные группы. Авторы данного изобретения показали, что растворимый CD52, присутствующий в человеческой семенной жидкости, аналогично растворимому CD52, высвобождаемому из лимфоцитов, таких как В-клетки Дауди, способен подавлять функцию Т-клеток и/или иммунный ответ. В качестве альтернативы, клетки-хозяева, предоставляющие другие условия гликозилирования, могут подвергаться отбору на экспрессию растворимого CD52 для того, чтобы получать альтернативные формы углевода на растворимом гликопротеине.
Углевод может быть присоединен к любой одной или более чем одной аминокислоте в пептиде, который способен иметь углеводную группировку, присоединенную к нему. Например, углевод может быть присоединен к одному или более чем одному из остатков аспарагина, серина, треонина, тирозина, гидроксилизина, гидроксипролина, фосфосерина или триптофана, если они присутствуют в аминокислотной последовательности. В одном примере углевод присоединен к остатку аспарагина (N) в пептидной части, имеющей последовательность по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% идентичную или на 100% идентичную аминокислотной последовательности, изложенной в SEQ ID NO: 3.
Согласно настоящему раскрытию также предложены варианты, мутанты, биологически активные фрагменты, модификации, аналоги и/или производные раскрытого здесь гликопротеина. Такие соединения можно идентифицировать скринингом на соединения, которые имитируют структуру и/или функцию раскрытого здесь полипептида, с использованием способов, включающих любой из раскрытых здесь способов.
Функция растворимого CD52
Раскрытый здесь гликопротеин предпочтительно способен подавлять активность («функцию») иммунных клеток, включающих лимфоциты (такие как Т-клетка) и моноциты. Например, раскрытый здесь гликопротеин способен подавлять функцию одной или более чем одной эффекторной Т-клетки, моноцита, макрофага и дендритной клетки. Эффекторные Т-клетки, моноциты, макрофаги и дендритные клетки и их функции будут известны специалисту в данной области.
Т-клетки можно легко идентифицировать по присутствию любого одного или более чем одного маркера Т-клетки, известного в данной области. Раскрытый здесь гликопротеин способен уменьшать пролиферацию Т-клеток в ответ на стимуляцию антигеном и/или способен снижать продукцию цитокинов Т-клетками (как, например, продукцию любого одного или более чем одного из IFN-γ (интерферон-гамма), IL-2 (интерлейкин-2), IL-10, IL-17, G-CSF (гранулоцитарный колониестимулирующий фактор), TNF-α (фактор некроза опухолей-альфа) и других цитокинов, для которых известно, что они секретируются активированными Т-клетками). Например, растворимый CD52 способен уменьшать продукцию IFN-γ Т-клетками.
В другом примере растворимый CD52 способен уменьшать секрецию IL-1β моноцитами, макрофагами и дендритными клетками.
Соответственно, раскрытый здесь гликопротеин способен уменьшать иммунный ответ у хозяина. Авторы данного изобретения показали, что раскрытый здесь гликопротеин способен ослаблять функцию эффекторных Т-клеток в ответ на стимуляцию любым антигеном. Подавляющая функция не зависит от конкретного антигена, использованного в стимуляции. Таким образом, раскрытый здесь гликопротеин способен ослаблять иммунный ответ на любой антиген. В одном примере антиген представляет собой аутоантиген.
Можно использовать любые известные способы определения подавления функции Т-клеток и/или иммунного ответа, такие как (но не ограничивающиеся) способами, описанными в приведенных здесь примерах. Таким образом, данные способы могут включать определение эффекта раскрытого здесь гликопротеина на пролиферацию одной или более чем одной эффекторной Т-клетки, моноцита, макрофага и дендритной клетки и/или на продукцию любого одного или более чем одного из IFN-γ, IL-2, IL-10, IL-17, G-CSF, TNF-α и других цитокинов, для которых известно, что они секретируются активированными Т-клетками, моноцитами, макрофагами или дендритными клетками.
Слитые белки
Пептидную часть раскрытого здесь гликопротеина CD52 можно, например, конъюгировать со вторым белком в виде слитого белка. Второй белок может представлять собой любой белок, способный увеличивать стабильность и/или растворимость гликопротеина, улучшать способ получения гликопротеина методами генной инженерии или улучшать терапевтический эффект гликопротеина. Таким образом, второй белок может иметь способность увеличивать период полувыведения раскрытого здесь гликопротеина.
Второй белок может иметь любую подходящую длину. В одном воплощении второй белок может быть относительно коротким. Например, второй белок может состоять из любого числа из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот. Второй белок может содержать по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9 или по меньшей мере 10 аминокислот. Второй белок также может содержать больше, чем 10 аминокислот. Например, второй белок может содержать по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30 или по меньшей мере 50 аминокислот.
В одном примере второй белок представляет собой фрагмент антитела. Подходящие фрагменты антитела включают любой фрагмент антитела, способный активировать иммунную систему. Фрагмент антитела может представлять собой область кристаллизуемого фрагмента (Fc) (которая может представлять собой одиночный полипептид) или любой один или более чем один константный домен тяжелой цепи (например, CH домены 2, 3 и/или 4) из области Fc. В одном примере второй белок представляет собой фрагмент Fc.
В другом примере второй белок может представлять собой метку для очистки. Известны многие примеры меток для очистки, и они включают (без ограничения) His метку, метку Т7, метку FLAG, S-метку, метку НА, метку с-Мус, DHFR (дегидрофолатредуктаза), хитинсвязывающий домен,
кальмодулинсвязывающий домен, целлюлозосвязывающий домен, метку Strep 2 (метка для очистки, кодирующая восемь аминокислот, которые связываются со Стрел-тактином, специально сконструированным стрептавидином (Schmidt and Skerra, 2007), и другие.
Второй белок может увеличивать растворимость экспрессируемого белка. Такие белки включают (без ограничения) NusA, тиоредоксин, малый убиквитиноподобный модификатор (SUMO), убиквитин и другие, известные в данной области.
Второй белок может увеличивать растворимость экспрессируемого белка, а также улучшать способы очистки. Такие белки включают (без ограничения) GST (глутатион-S-трансфераза), МВР, ген 10 Т7 и другие, известные в данной области.
Метку для очистки возможно можно удалять из слитого белка после его продукции. Подходящие способы удаления метки для очистки из слитого белка будут варьировать, в зависимости от конкретной использованной метки для очистки. Такие способы обычно будут известны в данной области.
Раскрытый здесь слитый белок может содержать один или более чем один из любых описанных выше вторых белков в любой комбинации. Таким образом, слитый белок может содержать фрагмент антитела (такой как Fc) и метку для очистки (такую как метка Strep 2).
Полинуклеотиды
Согласно настоящему раскрытию, кроме того, предложены выделенные или рекомбинантные полинуклеотиды, кодирующие белковый компонент растворимого гликопротеина CD52 или слитый белок. Последовательности таких полинуклеотидов можно получать из аминокислотных последовательностей белка CD52 и описанного здесь растворимого пептидного фрагмента CD52 и второго белка, содержащегося в пределах слитого белка. Раскрытые здесь полинуклеотиды также могут кодировать полноразмерный белок CD52, который, например, может быть его зрелой формой или его предшественником.
Подразумевается, что термин «выделенный полинуклеотид» означает полинуклеотид, который обычно отделяли от полинуклеотидных последовательностей, с которыми он ассоциирован или связан в его нативном состоянии. Предпочтительно выделенный полинуклеотид по меньшей мере на 60% свободен, более предпочтительно по меньшей мере на 75% свободен и более предпочтительно по меньшей мере на 90% свободен от других компонентов, с которыми он ассоциирован в природе. Кроме того, термин «полинуклеотид» используется здесь взаимозаменяемо с терминами «молекула нуклеиновой кислоты», «ген» и «мРНК».
Термин «рекомбинантный» в контексте полинуклеотида относится к полинуклеотиду при его присутствии в клетке или в бесклеточной экспрессионной системе в измененном количестве по сравнению с его нативным состоянием. В одном воплощении клетка представляет собой клетку, которая в природе не содержит данный полинуклеотид. Однако клетка может представлять собой клетку, которая содержит неэндогенный полинуклеотид, что приводит к измененному, предпочтительно повышенному уровню продукции кодируемого полипептида. Рекомбинантный полинуклеотид по изобретению включает полинуклеотиды, которые не были отделены от других компонентов трансгенной (рекомбинантной) клетки или бесклеточной экспрессионной системы, в которой он присутствует, и полинуклеотиды, продуцируемые в таких клетках или бесклеточных системах, которые затем очищают от по меньшей мере некоторых других компонентов.
Термин «полинуклеотид» относится к олигонуклеотиду, полинуклеотиду или его любому фрагменту. Он может представлять собой ДНК или РНК геномого или синтетического происхождения, двухцепочечную или одноцепочечную, и, в сочетании с углеводом, липидами, белком или другими веществами, может осуществлять конкретную определенную здесь активность.
Раскрытые здесь полинуклеотиды, при сравнении со встречающимися в природе молекулами (такими как геномные полинуклеотиды, кодирующие CD52 или его растворимый фрагмент), могут обладать одной или более чем одной мутацией, которая представляет собой делецию, вставку или замены нуклеотидных остатков. Мутанты могут быть либо встречающимися в природе (то есть, выделенными из природного источника), либо синтетическими (например, полученными проведением сайт-направленного мутагенеза или методиками перетасовки ДНК, как описано в широком смысле Harayama (1998)). Таким образом, очевидно, что полинуклеотиды по изобретению могут быть либо встречающимися в природе, либо рекомбинантными.
Конкретную последовательность полинуклеотида можно определять из пептидной последовательности. Из-за избыточности генетического кода для кодирования того же самого пептида можно использовать разные последовательности. Кроме того, полинуклеотидная последовательность может быть конкретно изменена так, чтобы усиливать ее экспрессию в конкретной клетке-хозяине. Такой способ хорошо известен в данной области как «оптимизация кодонов». Таким образом, раскрытый здесь полинуклеотид может быть оптимизирован по кодонам для усиления экспрессии в клетке-хозяине.
Векторы
Раскрытый здесь полинуклеотид может быть вставлен в нуклеотидный вектор для того, чтобы облегчать экспрессию белкового компонента гликопротеина или слитого белка. Соответственно, согласно настоящему раскрытию предложен вектор, содержащий полинуклеотид, кодирующий белковый компонент раскрытого здесь гликопротеина или раскрытого здесь слитого белка. Данный вектор может представлять собой либо РНК, либо ДНК, либо прокариотическую, либо эукариотическую, и может представлять собой транспозон (как, например, описано в US 5792294), вирус или плазмиду.
Предпочтительно полинуклеотид, кодирующий белковый компонент гликопротеина или слитого белка связан функциональным образом с промотором, который способен экспрессировать пептид при подходящих условиях. Фраза «связанный функциональным образом» в том виде, как она здесь используется, относится к функциональной связи между двумя или более чем двумя отрезками нуклеиновой кислоты (например, ДНК). Типично она относится к функциональной связи элемента, регулирующего транскрипцию, с транскрибируемой последовательностью. Например, промотор связан функциональным образом с кодирующей последовательностью, такой как опредленный здесь полинуклеотид, если он стимулирует или модулирует транскрипцию кодирующей последовательности в подходящей клетке или бесклеточной экспрессионной системе. В общем, промоторные элементы, регулирующие транскрипцию, которые связаны с транскрибируемой последовательностью функциональным образом, являются физически смежными с транскрибируемой последовательностью, т.е. они являются цис-действующими. Однако некоторые элементы, регулирующие транскрипцию, такие как энхансеры, не обязательно должны быть физически смежными или расположенными в тесной близости с кодирующими последовательностями, транскрипцию которых они усиливают.
Вектор предпочтительно представляет собой экспрессионный вектор. Термин «экспрессионный вектор» в том виде, как он здесь используется, представляет собой вектор на основе ДНК или РНК, который способен трансформировать клетку-хозяина и осуществлять экспрессию определенной молекулы полинуклеотида. Предпочтительно экспрессионный вектор также способен реплицироваться в клетке-хозяине. Экспрессионные векторы могут быть либо прокариотическими, либо эукариотическими и типично представляют собой вирусы или плазмиды. Раскрытые здесь экспрессионные векторы включают любые векторы, которые функционируют (например, управляют экспрессией гена) в раскрытых здесь рекомбинантных клетках (включая клетки животных) или в подходящей бесклеточной системе экспрессии.
В частности, раскрытые здесь экспрессионные векторы могут содержать регуляторные последовательности, такие как последовательности контроля транскрипции, последовательности контроля трансляции, точки начала репликации и другие регуляторные последовательности, которые совместимы с рекомбинантной клеткой или бесклеточной системой экспрессии, и которые контролируют экспрессию раскрытых здесь полинуклеотидных молекул. В частности, раскрытые здесь векторы могут включать последовательности контроля транскрипции. Последовательности контроля транскрипции представляют собой последовательности, которые контролируют инициацию, элонгацию и терминацию транскрипции. Особенно важными последовательностями контроля транскрипции являются последовательности контроля транскрипции, которые контролируют инициацию транскрипции, такие как последовательности промотора, энхансера, оператора и репрессора. Подходящие последовательности контроля транскрипции включают любую последовательность контроля транскрипции, которая может функционировать по меньшей мере в одной описанной здесь рекомбинантной клетке или бесклеточной системе экспрессии. Целый ряд таких последовательностей контроля транскрипции известен специалистам в данной области.
Раскрытые здесь векторы также могут содержать (а) сигналы секреции (т.е. сигнальный отрезок последовательностей нуклеиновых кислот) для обеспечения секреции экспрессируемого белка или пептида из клетки, которая продуцирует пептид, и/или (б) последовательности слияний, которые приводят к экспрессии раскрытых здесь пептидов в виде слитых белков. Примеры подходящих сигнальных отрезков включают любой сигнальный отрезок, способный управлять секрецией раскрытого здесь гликопротеина или слитого белка. Векторы также могут включать промежуточные и/или нетранслируемые последовательности, окружающие и/или находящиеся внутри последовательности(тей) нуклеиновой кислоты, кодирующей(щих) раскрытый здесь пептид.
Полинуклеотид или вектор может экспрессироваться в клетке-хозяине или в бесклеточной экспрессионной системе для того, чтобы продуцировать раскрытый здесь гликопротеин или слитый белок. Такую экспрессию, например, можно проводить в клетке млекопитающего, бакуловирусной экспрессионной системе, экспрессионной системе грибов (которую можно выбрать так, чтобы обеспечить гликозилирование экспрессируемого белка).
Клетка-хозяин может представлять собой любую клетку, способную продуцировать раскрытый здесь гликопротеин или слитый белок. Таким образом, в одном примере клетка-хозяин способна обеспечивать гликозилирование белкового компонента раскрытого здесь гликопротеина. Подходящие клетки-хозяева могут быть легко идентифицированы квалифицированным специалистом, и они включают, например, клетки животных, такие как клетки млекопитающих. В одном примере клетка-хозяин представляет собой клетку СНО, миеломную клетку (такую как клетки мышиной миеломы NS-O или SP2-O) или клетку НЕК293Т. В другом примере клетка-хозяин представляет собой клетку В-лимфобласт Дауди (Hu er al., 2009).
Кроме того, полинуклеотид или вектор может быть введен в клетку-хозяина для введения субъекту. Таким образом, раскрытая здесь фармацевтическая композиция может содержать клетку, содержащую раскрытый здесь полинуклеотид или вектор. Клетка может представлять собой выделенную клетку. Клетка предпочтительно представляет собой рекомбинантную клетку. Таким образом, клетка предпочтительно трансфицирована раскрытым здесь полинуклеотидом или вектором. Можно использовать любую клетку-хозяина, подходящую для введения субъекту. В одном примере клетка может быть взята от субъекта, подлежащего лечению. Таким образом, клетка может быть аутологичной клеткой. Соответственно, одна или более чем одна клетка может быть взята от субъекта, полинуклеотид или вектор, как здесь раскрыто, может быть введен в клетку-субъекта, и клетку можно затем вводить тому же самому субъекту. В одном примере клетка может представлять собой лимфоцит, такой как Т-клетка, такой как Т-клетка CD4+. В данном отношении способы отбора подходящего клеточного образца от субъекта будут известны в данной области. Когда клетка, подлежащая применению, представляет собой лимфоцит, способы могут включать лимфоцитоферез. Равным образом можно использовать другие подходящие клетки-хозяева, которые не обязательно происходят от субъекта, подлежащего лечению. Экспрессия полинуклеотида или вектора в клетке предпочтительно приводит к продукции и/или секреции раскрытого здесь гликопротеина.
Трансформацию полинуклеотида в клетку-хозяина можно осуществлять любым подходящим способом, известным в данной области. Методики трансформации включают трансфекцию, электропорацию, микроинъекцию, липофекцию и адсорбцию, но не ограничиваются ими. Рекомбинантная клетка может оставаться одноклеточной или может вырастать в ткань, орган или многоклеточный организм. Трансформированные полинуклеотидные молекулы, как здесь описано, могут оставаться внехромосомными или могут интегрироваться в одном или более чем одном сайте в пределах хромосомы трансформированной (т.е. рекомбинантной) клетки так, что сохраняется их способность к экспрессии.
Подходящие клетки-хозяева для трансформации включают любую клетку, которую можно трансформировать раскрытым здесь полинуклеотидом. Клетки-хозяева либо могут иметь способность продуцировать полипептиды по настоящему изобретению эндогенно (т.е. в природе), либо могут получить способность продуцировать такие полипептиды после трансформации по меньшей мере одной полинуклеотидной молекулой, как здесь раскрыто.
Для улучшения экспрессии трансформированной полинуклеотидной молекулы можно использовать методики генной инженерии, например, манипулируя числом копий полинуклеотидной молекулы в пределах клетки-хозяина, эффективностью, с которой транскрибируются данные полинуклеотидные молекулы, эффективностью, с которой транслируются образующиеся транскрипты, и эффективностью посттрансляционных модификаций. Методики генной инженерии, полезные для увеличения экспрессии раскрытых здесь полинуклеотидных молекул, включают осуществление функциональной связи полинуклеотидных молекул с плазмидами с большим числом копий, интеграцию полинуклеотидной молекулы в одну или более чем одну хромосому клетки-хозяина, добавление в плазмиды последовательностей для стабильности вектора, замены или модификации сигналов контроля транскрипции (например, промоторов, операторов, энхансеров), замены или модификации сигналов контроля трансляции (например, сайтов связывания с рибосомой, последовательностей Шайна-Дальгарно), модификацию полинуклеотидных молекул, как здесь раскрыто, для того, чтобы они соответствовали использованию кодонов клетки-хозяина и делецию последовательностей, которые дестабилизируют транскрипты, но не ограничиваются ими.
Клетку-хозяина можно культивировать при эффективных условиях для получения гликопротеина или слитого белка. Сразу после экспрессии в клетке-хозяине гликопротеин или слитый белок можно выделять традиционными способами, известными в данной области. Таким образом, в одном воплощении выделенный гликопротеин или слитый белок, как здесь описано, продуцируется культивированием клетки, способной к экспрессии гликопротеина или слитого белка, при эффективных условиях для продукции гликопротеина или слитого белка и выделением гликопротеина или слитого белка. Эффективные условия культивирования включают эффективные среды, биореактор, температуру, рН и условия, связанные с содержанием кислорода, которые обеспечивают продукцию гликопротеина или слитого белка и, в частности, которые обеспечивают гликозилирование, но не ограничиваются ими. Термин «эффективная среда» относится к любой среде, в которой клетка культивируется с образованием раскрытого здесь гликопротеина или слитого белка. Такая среда типично содержит водную среду, имеющую источники ассимилируемого углерода, азота и фосфата и подходящие соли, минеральные вещества, металлы и другие питательные вещества, такие как витамины. Клетки-хозяева можно культивировать в традиционных ферментационных биореакторах, встряхиваемых колбах, пробирках, планшетах для микротитрования и чашках Петри. Культивирование можно проводить при температуре, рН и содержании кислорода, подходящих для рекомбинантной клетки. Такие условия культивирования находятся в пределах квалификации обычного специалиста в данной области.
Также можно использовать любую бесклеточную систему экспрессии, подходящую для экспрессии раскрытого здесь полинуклеотида. Подходящие бесклеточные системы экспрессии включают системы экспрессии, которые обеспечивают гликозилирование белкового компонента гликопротеина или слитого белка. Такие условия могут быть определены специалистом в данной области.
Раскрытый здесь гликопротеин также можно продуцировать путем индукции экспрессии CD52 в выделенной клетке-хозяине и выделения растворимого гликопротеина CD52, продуцируемого клеткой-хозяином. Таким образом, гликопротеин можно продуцировать с использованием клетки, которая продуцирует растворимый CD52 в природе. Подходящие клетки будут идентифицируемыми специалистом в данной области, и они включают (без ограничения) лимфоциты, клетки области половых путей (такие как клетки спермы), клетки В-лимфобласты Дауди (Hu et al., 2009), клетки К562 и другие. Дополнительные линии клеток, способных к продукции растворимого CD52 в природе, можно идентифицировать путем скрининга на секрецию растворимого CD52. Таким образом, можно подвергать скринингу раковые клетки на их способность секретировать растворимый CD52.
Способы получения раскрытого здесь гликопротеина из выделенной клетки-хозяина, которая продуцирует растворимый CD52 в природе, могут включать стимулирование клетки-хозяина для продукции более высоких уровней растворимого CD52. Это может достигаться, например, приведением в контакт клетки-хозяина с антигеном. Можно использовать любой антиген. В одном примере антиген представляет собой аутоантиген, такой как GAD65. В другом примере антиген представляет собой столбнячный анатоксин.
Способы получения раскрытого здесь гликопротеина из выделенной клетки-хозяина, которая продуцирует растворимый CD52 в природе, также могут включать отбор клетки, которая экспрессирует CD52 в природе, и приведение данной клетки в контакт с ферментом, способным отщеплять внеклеточную часть мембраносвязанного CD52 с высвобождением растворимого CD52. Подходящие ферменты известны в данной области и включают фосфолипазы, такие как фосфолипаза С.
Описанные здесь способы можно проводить на выделенных клетках или популяциях клеток достаточного размера для получения желательного количества растворимого CD52.
Клетки CD52hi
Согласно настоящему раскрытию также предложены выделенные клетки и популяции клеток, демонстрирующие высокие уровни экспрессии CD52. Под «высоким» подразумевается то, что уровни экспрессии CD52 являются относительно высокими по сравнению с уровнями экспрессии CD52 в данной популяции клеток. Данная популяция клеток может представлять собой, например, популяцию лимфоцитов. Популяция лимфоцитов может содержать Treg клетки и клетки, не являющиеся Treg. Кроме того, популяцию лимфоцитов возможно приводили в контакт с антигеном для того, чтобы стимулировать активность лимфоцитов. В качестве альтернативы, популяция клеток может представлять собой клетки половых путей, такие как клетки спермы. Напротив, клетки CD5210 демонстрируют относительно низкие уровни CD52 относительно данной популяции клеток.
В одном примере клетку можно определять как клетку CD52hi, если уровень экспрессии CD52 в данной клетке на 1%, 5%, 10%, 20%, 30%, 40% или 50% выше уровней экспрессии CD52 в популяции клеток. Предпочтительно клетка CD52hi имеет уровень экспрессии на 10% выше экспрессии CD52, наблюдаемой в популяции клеток.
В одном примере клетку можно определять как клетку CD52'0, если уровень экспрессии CD52 в данной клетке на 1%, 5%, 10%, 20%, 30%, 40% или 50% ниже уровней экспрессии CD52 в популяции клеток. Предпочтительно клетка CD52lo имеет уровень экспрессии на 10% ниже экспрессии CD52, наблюдаемой в популяции клеток.
Клетка CD52hi может быть выделена из популяции клеток, из которой она идентифицирована. В качестве альтернативы, популяция клеток CD52hi может быть выделена из исходной популяции клеток, из которой идентифицированы клетки CD52hi. Таким образом, раскрытые здесь популяции клеток могут быть обогащены клетками CD52hi.
Согласно настоящему раскрытию, следовательно, предложена выделенная популяция клеток, содержащая множество клеток CD52hi. Клетки CD52hi могут составлять по меньшей мере 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 95% от общей обогащенной популяции клеток.
Выделенные раскрытые здесь клетки CD52hi и популяции клеток CD52hi способны продуцировать раскрытый здесь растворимый гликопротеин CD52.
Данные выделенные клетки и популяции клеток могут быть дополнительно определены по присутствию одного или более чем одного дополнительного клеточного маркера. В одном примере клетки CD52hi представляют собой клетки CD4+ CD52hi. В качестве альтернативы, клетки CD52hi представляют собой клетки CD8+ CD52hi. Дополнительные маркеры, которые характеризуют данные клетки, включают любой один или более чем один из следующих: индуцируемый глюкокортикоидом белок, родственный рецептору фактора некроза опухоли (GITR), CD127, лиганд Fas (FasL или CD95L), рецептор сфингозин-1-фосфата (S1PR), заякоренный GPI (гликозилфосфатидилинозитол) гликопротеин CD24, CD25, FoxP3, CTLA-4 и другие маркеры в любой комбинации. Авторы данного изобретения обнаружили, что уровни экспрессии GITR, CD127, FasL, S1PR и CD24 могут быть выше в Treg клетках CD52hi по сравнению с клетками CD52lo. Данные маркеры, следовательно, можно использовать для дополнительного определения клетки CD52hi или популяции клеток CD52hi, как здесь описано.
Кроме того, функцию данной клетки можно использовать для определения клетки CD52hi или популяции клеток CD52hi, как здесь описано. Например, для идентификации клетки CD52hi или популяции клеток CD52hi можно использовать способность клетки, экспрессирующей CD52, ослаблять функцию эффекторной Т-клетки, как здесь описано.
Среда для культуры клеток
Клетки CD52hi или популяцию клеток CD52hi, как здесь описано, можно культивировать так, чтобы продуцировать среду, содержащую раскрытый здесь растворимый гликопротеин. Подходящие культуральные условия будут очевидными специалисту в данной области. Культивируемые клетки могут быть дополнительно индуцированы для увеличения их уровня экспрессии растворимого CD52 любым подходящим способом, включающим приведение данных клеток в контакт с антигеном.
Обработка клеток ex vivo
Согласно настоящему изобретению также предложена фармацевтическая композиция, содержащая клетки, предпочтительно иммунные клетки, и фармацевтически приемлемый носитель, где клетки были обработаны ex vivo любым одним или более чем одним из следующих:
i) растворимый гликопротеин CD52;
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv);
vi) выделенная клетка CD52hi, способная продуцировать растворимый гликопротеин CD52;
vii) выделенная популяция клеток, содержащая множество клеток CD52hi, способных продуцировать растворимый гликопротеин CD52;
viii) среда культуры клеток или ее фракция, содержащая растворимый гликопротеин CD52, выделенная из культуры клеток, содержащей клетку по разделу vi) или популяцию клеток по разделу vii); и
ix) агент, способный увеличивать уровень экспрессии растворимого гликопротеина CD52 клеткой.
Клетки данной композиции могут представлять собой, например, цельную кровь или ее клеточную фракцию, такую как одноядерные клетки периферической крови (РВМС).
Такие клетки, обработанные ex vivo, можно использовать в настоящем изобретении, например, для лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса.
В одном воплощении клетки являются аутологичными по отношению к субъекту, которому они будут вводиться. В другом воплощении клетки являются аллогенными.
Фармацевтические композиции
Согласно настоящему раскрытию предложена фармацевтическая композиция, содержащая любой один или более чем один из описанных здесь растворимого гликопротеина CD52, слитого белка, полинуклеотида, вектора, клетки, популяции клеток и клеточной среды и любого агента, способного увеличивать уровень экспрессии CD52 в клетке, и фармацевтически приемлемый носитель.
Фармацевтически приемлемый носитель включает носитель, подходящий для применения при ведении животным, таким как млекопитающие и по меньшей мере предпочтительно человек. В одном примере, термин «фармацевтически приемлемый» означает одобренный регулирующим агенством федерального правительства или правительства штата или перечисленный в Фармакопее США или другой общепризнанной фармакопее для применения у животных и, более конкретно, у человека. Термин «носитель» относится к разбавителю, эксципиенту или наполнителю, с которым вводится терапевтический агент.Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла, включая масла, происходящие из нефти, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное.
Терапевтические композиции можно получать смешиванием желательных соединений, имеющих подходящую степень чистоты, с возможными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences, 16th edition, Osol, A. ed. (1980)) в форме лиофилизированных композиций, водных растворов или водных суспензий. Приемлемые носители, эксципиенты или стабилизаторы предпочтительно являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают буферы, такие как Tris, HEPES, PIPES, фосфатный, цитратный и на основе других органических кислот; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензэтония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (меньше, чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG). Дополнительные примеры таких носителей включают ионообменники, квасцы, стеарат алюминия, лецитин, сывороточные белки, такие как человеческий сывороточный альбумин, буферизующие вещества, такие как глицин, сорбиновая кислота, сорбат калия, смеси частичных глицеридов насыщенных растительных жирных кислот, воду, соли или электролиты, такие как протамина сульфат, динатрия гидрофосфат, калия гидрофосфат, хлорид натрия, коллоидный диоксид кремния, трисиликат магния, поливинилпирролидон и вещества на основе целлюлозы.
Фармацевтическую композицию в том виде, как она здесь раскрыта, готовят так, чтобы она была совместима с ее намеченным путем введения. Примеры путей введения включают парентеральное (например, внутривенное, внутрикожное, подкожное, внутримышечное, внутрибрюшинное, подоболочечное), через слизистую (например, пероральное, интраназальное, щечное, вагинальное, респираторное), энтеральное (например, перорально, как, например, посредством таблеток, капсул или капель, ректально) и чрескожное (местное, например, накожное, ингаляционное, интраназальное, в виде глазных капель, вагинальное). Растворы или суспензии, используемые для парентерального, внутрикожного, энтерального или подкожного применения могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекции, физиологический раствор, нелетучие масла, полиэтиленгликоли, глицерин, пропиленгликоль и другие синтетические растворители; антибактериальные агенты, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелаторы, такие как этилендиаминтетрауксусная кислота; буферы, такие как ацетаты, цитраты или фосфаты, и агенты для корректировки тоничности, такие как хлорид натрия или декстроза. рН можно корректировать кислотами или основаниями, такими как соляная кислота или гидроксид натрия. Парентеральный препарат может быть заключен в ампулы, одноразовые шприцы или многодозовые флаконы, сделанные из стекла или пластмассы.
Чрескожная доставка осуществляется путем подвергания кожи пациента воздействию терапевтического агента в течение продолжительного периода времени. Чрескожные пластыри имеют дополнительное преимущество в обеспечении контролируемой доставки фармацевтического агента в организм (см., например, Transdermal and Topical Drug Delivery: From Theory to Clinical Practice, Williams (ed), Pharmaceutical Press, UK (2003); Transdermal Drug Delivery: Developmental Issues and Research Initiatives, Hadgraft and Guy (eds.), Marcel Dekker, Inc., (1989)). Например, простой адгезивный пластырь может быть получен из материала основы и акрилатного адгезива. Терапевтический агент и любой усилитель готовят в виде адгезивного раствора для отливки и обеспечивают его тщательное перемешивание. Отливку данного раствора осуществляют непосредственно на материале основы, и растворитель для отливки выпаривают в печи, оставляя адгезивную пленку. Для завершения системы можно присоединять покровную пленку.
В качестве альтернативы, для доставки терапевтического агента можно использовать пластырь с полиуретановой матрицей. Слои данного пластыря содержат основу, полиуретановую матрицу с лекарственным средством/усилителем, мембрану, адгезив и покровную пленку. Полиуретановую матрицу получают с использованием полиуретанового предполимера, отверждаемого при комнатной температуре. Добавление воды, спирта и комплекса в предполимер приводит к образованию клейкого прочного эластомера, отливку которого можно осуществлять непосредственно на материале основы.
В другом воплощении данного изобретения будет использоваться пластырь с гидрогелевой матрицей. Типично гидрогелевая матрица будет содержать спирт, воду, лекарственное средство и несколько гидрофильных полимеров. Эту гидрогелевую матрицу можно включать в чрескожный пластырь между основой и адгезивным слоем.
В том, что касается систем пассивной доставки, скорость высвобождения типично контролируется мембраной, помещенной между резервуаром и кожей, посредством диффузии из монолитного устройства, или самой кожей, служащей в системе доставки в качестве барьера, контролирующего скорость (см. US 4816258, 4927408, 4904475, 4588580, 4788062). Скорость доставки будет частично зависеть от природы мембраны. Например, скорость доставки через мембраны в пределах организма обычно выше, чем через кожные барьеры.
Подходящие проницаемые мембранные материалы можно выбирать на основе желательного уровня проницаемости, природы комплекса и соображений механики, относящихся к конструированию приспособления. Типичные проницаемые мембранные материалы включают широкий спектр природных и синтетических полимеров, таких как полидиметилсилоксаны (силиконовые резины), этиленвинилацетатный сополимер (EVA), полиуретаны, сополимеры полиуретана-полиэфира, полиэтилены, полиамиды, поливинилхлориды (PVC), полипропилены, поликарбонаты, политетрафторэтилены (PTFE), целлюлозные вещества, например, триацетат целлюлозы и нитрат/ацетат целлюлозы, и гидрогели, например, 2-гидроксиэтилметакрилат (НЕМА).
В приспособлении могут содержаться другие элементы, такие как другие традиционные компоненты терапевтических продуктов, в зависимости от желательных характеристик приспособления. Например, композиции согласно изобретению также могут включать один или более чем один консервант или бактериостатический агент, например, метилгидроксибензоат, пропилгидроксибензоат, хлоркрезол, бензалкония хлориды и тому подобное. Данные фармацевтические композиции также могут содержать другие активные ингредиенты, такие как противомикробные агенты, в частности, антибиотики, анестетики, анальгетики и противозудные агенты.
Согласно другому воплощению данного изобретения предложена местная доставка фармацевтической композиции. Данная схема лечения подходит либо для системного введения фармацевтического агента, либо для локализованной терапии, например, непосредственно в патологическую или пораженную заболеванием ткань. Местные препараты можно получать объединением комплекса фармацевтический агент-химический модификатор с традиционными фармацевтическими разбавителями и носителями, обычно используемыми в местных сухих, жидких композициях, композициях в виде крема и аэрозоля. Мази и кремы, например, можно готовить с водной или масляной основой с добавлением подходящих загустителей и/или гелеобразующих агентов. Такие основы могут включать воду и/или масло, как, например, жидкий парафин или растительное масло, как, например, арахисовое масло или касторовое масло. Загустители, которые можно использовать согласно природе основы, включают мягкий парафин, стеарат алюминия, цетостеариловый спирт, пропиленгликоль, полиэтиленгликоли, ланолин, гидрогенизированный ланолин, пчелиный воск и тому подобное. Лосьоны могут быть приготовлены с водной или масляной основой и, в общем, также будут включать одно или более чем одно из следующего: стабилизаторы, эмульгаторы, диспергирующие агенты, суспендирующие агенты, загустители, красители, отдушки и тому подобное. Порошки можно формовать с помощью любой подходящей порошковой основы, например, талька, лактозы, крахмала и тому подобного. Капли могут быть приготовлены с водной основой или неводной основой, также включающей один или более чем один диспергирующий агент, суспендирующий агент, солюбилизатор и тому подобное.
Лекарственные формы для местного введения включают порошки, спреи, мази, пасты, кремы, лосьоны, гели, растворы, пластыри и лекарственные формы для ингаляции. Активное соединение можно смешивать при стерильных условиях с фармацевтически приемлемым носителем и с любыми консервантами, буферами или пропеллентами, которые могут потребоваться. Мази, пасты, кремы и гели также могут содержать эксципиенты, такие как животные и растительные жиры, масла, воска, парафины, крахмал, трагакантовую камедь, производные целлюлозы, полиэтиленгликоли, силиконы, бентониты, тальк, оксид цинка или их смеси. Порошки и спреи также могут содержать эксципиенты, такие как лактоза, тальк, гидроксид алюминия, силикаты кальция, полиамидный порошок или смеси данных веществ. Спрэи могут дополнительно содержать традиционные пропелленты, такие как хлорфторуглеводороды и летучие незамещенные углеводороды, такие как бутан и пропан.
Доставка лекарственного средства через слизистую (например, через желудочно-кишечный тракт, подъязычная, щечная, назальная, легочная, вагинальная, через роговицу и мембраны глаза) обеспечивает эффективное поступление активных веществ в большой круг кровообращения и уменьшает немедленный метаболизм печенью и флорой стенки кишечника (см., например, Lee, 2001; Song et al., 2004; Hearnden et al., 2012). Лекарственные формы для доставки через слизистую (например, таблетка, суппозиторий, мазь, гель, бальзамы, кремы, пессарий, мембрана и порошок) типично поддерживают в контакте со слизистой оболочкой, и они быстро распадаются и/или растворяются с обеспечением немедленного системного поглощения.
Для доставки на щечные или подъязычные слизистые оболочки типично будет использоваться пероральная композиция, такая как пастилка, таблетка или капсула. Способ изготовления данных композиций известен в данной области и включает добавление к предварительно изготовленной таблетке комплекса фармацевтический агент-химический модификатор; холодное прессование инертного наполнителя, связующего вещества и либо комплекса фармацевтический агент-химический модификатор, либо вещества, содержащего данный комплекс (как описано в US 4806356), и инкапсулирование, но не ограничивается ими.
Пероральные композиции обычно включают инертный разбавитель или съедобный носитель. Они могут быть заключены в желатиновые капсулы или прессованы в таблетки. Для цели перорального терапевтического введения активное соединение включают с эксципиентами и используют в форме таблеток, лепешек или капсул. Пероральные композиции также получают с использованием жидкого носителя для применения в виде полосканий для рта, в которых соединение в жидком носителе применяют перорально и полощут им рот и сплевывают или проглатывают. В виде части композиции можно включать фармацевтически приемлемые связующие агенты и/или адъювантные вещества. Таблетки, пилюли, капсулы, лепешки и тому подобное могут содержать любой из следующих ингредиентов или соединений аналогичной природы: связующее вещество, такое как микрокристаллическая целлюлоза, трагакантовая камедь или желатин; эксципиент, такой как крахмал или лактоза; разрыхлитель, такой как альгиновая кислота, примогель или кукурузный крахмал; смазка, такая как стеарат магния или стероты; скользящее вещество, такое как коллоидный диоксид кремния; подсластитель, такой как сахароза или сахарин; или корригент, такой как перечная мята, метилсалицилат или корригент со вкусом апельсина.
Другой пероральной композицией является композиция, которую можно наносить с адгезивным средством, таким как целлюлозное производное, гидроксипропилцеллюлоза, на слизистую полости рта, например, как описано в US 4940587. Эта буккальная адгезивная композиция, при нанесении на слизистую щеки, обеспечивает контролируемое высвобождение комплекса фармацевтический агент-химический модификатор в рот и через слизистую оболочку щеки.
Для доставки на слизистые оболочки носа и/или дыхательной системы типично будет применяться аэрозольная композиция. Термин «аэрозоль» включает любую переносимую газом суспендированную фазу комплекса фармацевтический агент-химический модификатор, которая имеет способность к ингаляции в бронхиолы или носовые ходы. Конкретно, аэрозоль включает суспензию капелек соединений по настоящему изобретению, переносимую газом, которая может продуцироваться ингалятором или небулайзером с отмеренной дозой или туманообразующим распылителем. Аэрозоль также включает сухопорошковую композицию комплекса фармацевтический агент-химический модификатор, суспендированную в воздухе или другом газе-носителе, которую можно доставлять посредством ингаляции из ингалятора.
Для доставки через слизистую или чрескожной доставки в композиции можно использовать агенты, усиливающие проникновение, подходящие для барьеров, которые нужно преодолеть. Такие агенты, усиливающие проникновение, обычно известны в данной области и включают, например, агенты для введения через слизистую, детергенты, соли желчи и производные фузидовой кислоты.
Фармацевтические композиции, подходящие для применения в виде инъекции, включают стерильные водные растворы (в случае растворимости в воде) или дисперсии и стерильные порошки для немедленного получения стерильных инъецируемых растворов или дисперсии. Для внутривенного введения подходящие носители включают физиологический раствор, бактериостатическую воду, Cremophor™ (BASF, Parsippany, N.J.) или фосфатно-солевой буферный раствор (PBS). Во всех случаях композиция должна быть стерильной и должна быть жидкой в той степени, в которой существует возможность легкого введения шприцем. Она должна быть стабильной при условиях изготовления и хранения и должна быть защищена против загрязняющего действия микроорганизмов, таких как бактерии и грибки. Носитель представляет собой растворитель или диспергирующую среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и тому подобное) и их подходящие смеси. Правильная жидкостность поддерживается, например, посредством применения покрытия, такого как лецитин, посредством поддержания требующегося размера частиц в случае дисперсии и посредством применения поверхностно-активных веществ. Предупреждение действия микроорганизмов может достигаться разными антибактериальными и противогрибковыми агентами, например, парабенами, хлорбутанолом, фенолом, аскорбиновой кислотой и тому подобным. Во многих случаях предпочтительно включать в композицию изотоничные агенты, например, сахара, полиспирты, такие как маннит, сорбит, хлорид натрия. Пролонгированное поглощение инъецируемых композиций осуществляется включением в композицию агента, который задерживает поглощение, например, моностеарата алюминия и желатина.
Стерильные инъецируемые растворы можно получать включением активного соединения в требующем количестве в подходящем растворителе с одним или комбинацией перечисленных выше ингредиентов, по мере необходимости, с последующей стерилизацией через фильтр. Обычно дисперсии получают включением активного соединения в стерильный носитель, который содержит базовую диспергирующую среду и другие требующиеся ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных инъецируемых растворов, предпочтительными способами получения являются вакуумная сушка и лиофилизация, которые дают порошок активного ингредиента плюс любого дополнительного желательного ингредиента из их раствора, ранее подвергнутого стерилизующей фильтрации.
Понятно, что фармацевтически приемлемый носитель обозначает соединение или комбинацию соединений, входящих в фармацевтическую композицию, которые не вызывают побочных эффектов, и которые обеспечивают возможность, например, облегчать введение активного соединения для увеличения его времени жизни и/или его эффективности в организме, для увеличения его растворимости в растворе или, в качестве альтернативы, для улучшения его сохранности. Данные фармацевтически приемлемые носители хорошо известны и будут адаптированы специалистами в данной области согласно природе и способу введения выбранного активного соединения.
Фармацевтические композиции, подлежащие применению для введения in vivo, должны быть стерильными. Это легко достигается фильтрованием через мембраны для стерилизующей фильтрации до или после лиофилизации и растворения. Композицию можно хранить в лиофилизированной форме или в растворе, в случае системного введения. При ее нахождении в лиофилизированной форме, ее типично готовят в комбинации с другими ингредиентами для разведения подходящим разбавителем во время применения. Примером жидкой композиции является стерильный, прозрачный, бесцветный раствор без консервантов, которым заполнен однодозовый флакон для подкожной инъекции.
Фармацевтические композиции обычно помещают в контейнер, имеющий стерильный порт доступа, например, в мешок для внутривенного раствора или во флакон, имеющий пробку, прокалываемую иглой для подкожной инъекции. Композиции предпочтительно вводят парентерально, например, в виде внутривенных инъекций или инфузий, или вводят в полость тела.
Раскрытые здесь фармацевтические композиции, кроме того, могут содержать дополнительный терапевтический агент, для которого известно, что он подавляет функцию эффекторных Т-клеток и/или иммунный ответ.
В одном воплощении композиция, кроме того, содержит инсулин.
В другом воплощении композиция, кроме того, содержит аутоантиген. Примеры аутоантигенов, полезных в композициях по изобретению, включают аутоантигены, перечисленные в Таблице 1 (Lemmark, 2001), но не ограничиваются ими.
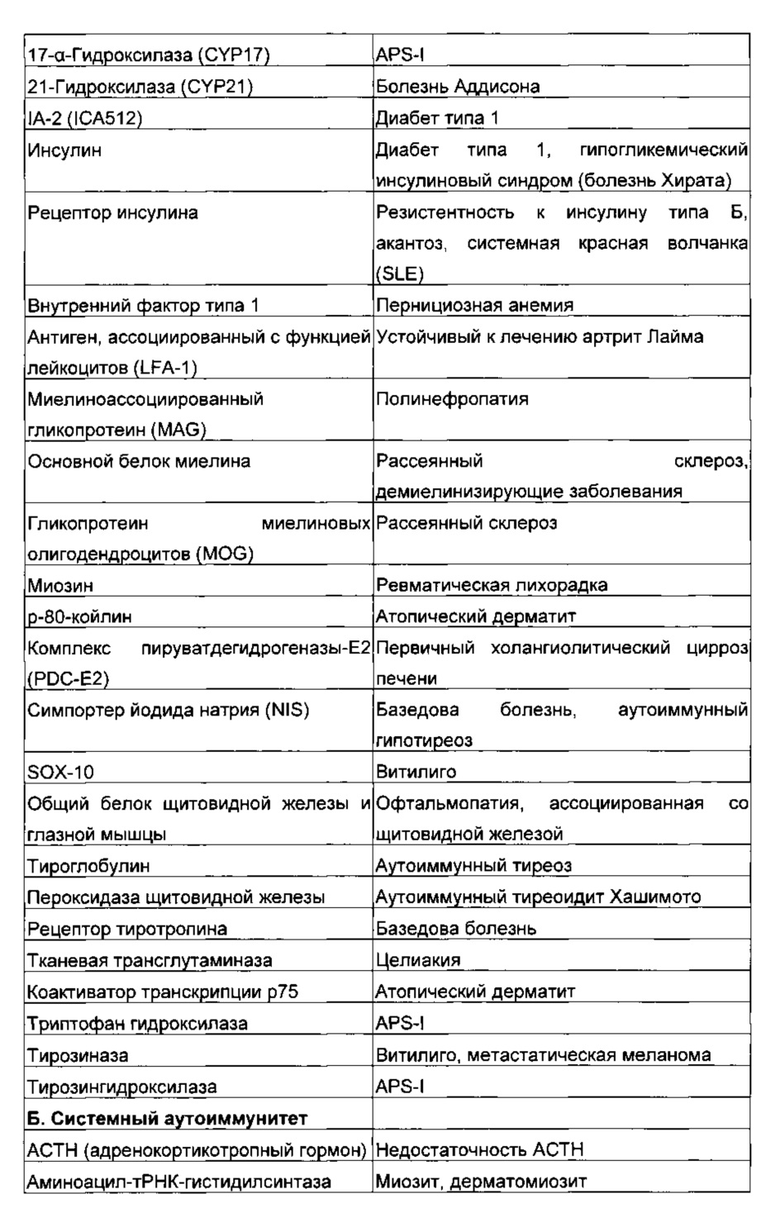

Способы лечения
Для подавления функции эффекторных Т-клеток, воспаления или сепсиса можно использовать описанные здесь растворимый гликопротеин CD52, слитый белок, полинуклеотид, вектор, клетку, популяцию клеток, среду клеток и фармацевтическую композицию и любой агент, способный увеличивать уровень экспрессии CD52 в клетке. Таким образом, описанные здесь растворимый гликопротеин CD52, слитый белок, полинуклеотид, вектор, клетку, популяцию клеток, среду клеток и фармацевтическую композицию, и любой агент, способный увеличивать уровень экспрессии CD52 в клетке можно использовать для лечения любого заболевания или состояния, опосредованного эффекторными Т-клетками, включая воспаление или сепсис.
В одном примере заболевание или состояние, опосредованное эффекторными Т-клетками, может представлять собой аутоиммунное заболевание, отторжение аллотрансплантата, реакцию «трансплантат против хозяина» или аллергическое заболевание. Термин «аутоиммунное заболевание» относится к любому заболеванию, при котором организм продуцирует иммуногенный ответ (т.е. ответ иммунной системы) на некоторый компонент его собственной ткани. Аутоиммунные заболевания можно классифицировать на заболевания, при которых поражен преимущественно один орган (например, гемолитическая анемия и антииммунный тиреоз) и заболевания, при которых процесс аутоиммунного заболевания рассеян по многим тканям (например, системная красная волчанка). Аутоиммунное заболевание может представлять собой (но не ограничивается) любым одним или более чем одним из следующих: инсулинозависимый сахарный диабет (или диабет типа 1), инсулиновый аутоиммунный синдром, ревматоидный артрит, псориатический артрит, хронический артрит Лайма, волчанка, рассеянный склероз, воспалительное заболевание кишечника, включая болезнь Крона, язвенный колит, целиакию, аутоиммунное заболевание щитовидной железы, аутоиммунный миокардит, аутоиммунный гепатит, пузырчатка, аутоиммунное заболевание против базальной мембраны трубочек (почки), семейная дилатационная кардиомиопатия, синдром Гудпасчера, синдром Шегрена, тяжелая миастения, полиэндокринная недостаточность, витилиго, периферическая нейропатия, аутоиммунный полигландулярный синдром типа I, острый гломерулонефрит, идиопатический гипопаратиреоз, начинающийся у взрослых (AOIH), полное облысение, тиреоидит Хашимото, базедова болезнь, болезнь Аддисона, хронический бериллиевый синдром, анкилозирующий спондилит, юношеский дерматомиозит, полихондрит, склеродермия, местный энтерит, дистальный илеит, гранулематозный энтерит, местный илеит и терминальный илеит, боковой амиотрофный склероз, анкилозирующий спондилит, аутоиммунная апластическая анемия, аутоиммунная гемолитическая анемия, болезнь Бехчета, целиакия, хронический активный гепатит, CREST-синдром, дерматомиозит, дилатационная кардиомиопатия, синдром эозинофилии-миалгии, буллезный приобретенный эпидермолиз (ЕВА), гигантоклеточный артериит, синдром Гудпасчера, синдром Гийена-Барре, гемохроматоз, пурпура Хенох-Шонплейна, идиопатическая IgA нефропатия, аутоиммунный инсулиновый синдром, юношеский ревматоидный артрит, синдром Ламберта-Итона, линейный IgA дерматоз, миокардит, нарколепсия, некротизирующий васкулит, синдром неонатальной волчанки (NLE), нефротический синдром, пемфигоид, пузырчатка, полимиозит, первичный склерозирующий холангит, псориаз, быстро прогрессирующий гломерулонефрит (RPGN), синдром Райтера, синдром мышечной скованности, воспалительное заболевание кишечника, остеоартрит, тиреоз и другие. В одном примере аутоиммунное заболевание представляет собой диабет типа 1. В другом примере аутоиммунное заболевание представляет собой ревматоидный артрит.В другом примере состояние представляет собой отторжение аллотрансплантата или реакцию «трансплантат против хозяина». Таким образом, раскрытые здесь способы могут включать введение любого одного или более чем одного из растворимого гликопротеина CD52, слитого белка, полинуклеотида, вектора, клетки, популяции клеток, среды клеток, агента и фармацевтической композиции в трансплантат реципиента.
Аллергическое заболевание может представлять собой (но не ограничивается) любым одним или более чем одним из пищевой аллергии, аллергии на аллерген, передающийся по воздуху, аллергии на клещей домашней пыли, аллергии на кошек, аллергии на пчелиный яд или другой аллергии.
Воспаление может возникать в ответ на травму или ненормальную стимуляцию, вызванную физическим, химическим или биологическим агентом. Воспалительная реакция может включать местные реакции и возникающие из-за них морфологические изменения, разрушение или удаление повреждающего материала, такого как инфекционный организм, и ответы, которые приводят к восстановлению и заживлению.
Воспаление возникает при воспалительных расстройствах. Термин «воспалительный», при использовании в связи с расстройством, относится к патологическому процессу, который вызван, возникает или приводит к воспалению, которое является неподходящим, или которое не проходит обычным способом. Воспалительные расстройства могут быть системными или локализованными в конкретных тканях или органах. Известно, что воспаление возникает при многих расстройствах (некоторые из них являются аутоиммунными заболеваниями), которые включают системный воспалительный ответ (SIRS); болезнь Альцгеймера (и ассоциированные состояния и симптомы, включающие хроническое нейровоспаление, активацию глии; увеличенную микроглию; образование нейритических бляшек; боковой амиотрофический склероз (ALS), артрит (и ассоциированные состояния и симптомы, включающие, но не ограничивающиеся следующими: острое воспаление суставов, артрит, индуцированный антигеном, артрит, ассоциированный с хроническим лимфоцитарным тиреоидитом, артрит, индуцированный коллагеном, юношеский артрит, ревматоидный артрит, остеоартрит, прогноз и артрит, индуцированный стрептококками, спондилоартропатии и подагрический артрит), астму (и ассоциированные состояния и симптомы, включающие следующие: бронхиальная астма, хроническое обструктивное заболевание дыхательных путей, хроническое обструктивное заболевание легких, юношеская астма и астма, связанная с родом деятельности); сердечно-сосудистые заболевания (и ассоциированные состояния и симптомы, включающие следующие: атеросклероз, аутоиммунный миокардит, хроническая сердечная гипоксия, застойная сердечная недостаточность, заболевание коронарных артерий, кардиомиопатия и дисфункция сердечных клеток, включающая: активацию гладкомышечных клеток аорты, апоптоз клеток сердца и иммуномодуляцию функции клеток сердца); диабет (и ассоциированные состояния, включающие аутоиммунный диабет, инсулинозависимый диабет (Типа 1), диабетический периодонтит, диабетическую ретинопатию и диабетическую нефропатию); желудочно-кишечные воспаления (и связанные с ними состояния и симптомы, включающие целиакию, ассоциированную остеопению, хронический колит, болезнь Крона, воспалительное заболевание кишечника и язвенный колит); язвы желудка; воспаления печени, такие как вирусный и другие типы гепатита, холестериновые желчные камни и фиброз печени; инфекцию ВИЧ (вирус иммунодефицита человека (и ассоциированные состояния, включающие дегенеративные ответы, нейродегенеративные ответы и болезнь Ходжкина, ассоциированную с ВИЧ); синдром Кавасаки (и ассоциированные заболевания и состояния, включающие кожно-слизистый синдром лимфатического узла, лимфаденопатию шейки матки, поражения коронарных артерий, отек, лихорадку, повышенный уровень лейкоцитов, умеренную анемию, шелушение кожи, высыпание, покраснение конъюнктивы, тромбоцитоз); нефропатии (и ассоциированные заболевания и состояния, включающие диабетическую нефропатию, почечное заболевание на конечном этапе, острый и хронический гломерулонефрит, острый и хронический интерстициальный нефрит, волчаночный нефрит, синдром Гудпасчера, выживание при гемодиализе и реперфузионное ишемическое поражение почки); нейродегенеративные заболевания или нейропатологические состояния (и ассоциированные заболевания и состояния, включающие острую нейродегенерацию, индукцию IL-1 при старении и нейродегенеративном заболевании, индуцированную IL-1 пластичность нейронов гипоталамуса и хроническую гиперчувствительность к стрессу, миелопатию); офтальмопатии (и ассоциированные заболевания и состояния, включающие диабетическую ретинопатию, эндокринную офтальмопатию, воспаление, ассоциированное с поражением роговицы или инфекцией, включая изъязвление роговицы и увеит), остеопороз (и ассоциированные заболевания и состояния, включающие потерю и повышенную частоту переломов альвеолярной, бедренной, лучевой, позвоночных костей или костей запястья, постменопаузную потерю кости, повышенную частоту или показатель потери кости); отит среднего уха (взрослый или детский); панкреатит или панкреатический ацинус; заболевание периодонта (и ассоциированные заболевания и состояния, включая заболевания взрослых, с началом в раннем возрасте и диабетические); легочные заболевания, включающие хроническое заболевание легких, хронический синусит, заболевание, связанное с гиалиновой оболочкой, гипоксию и легочное заболевание при SIDS (синдром внезапной смерти младенца); рестеноз коронарных или других сосудистых трансплантатов; ревматизм, включающий ревматоидный артрит, ревматические тела Ашоффа, ревматические заболевания и ревматический миокардит; тиреоидит, включающий хронический лимфоцитарный тиреоидит; инфекции мочевых путей, включающие хронический простатит, синдром хронической боли в области таза и уролитиаз; иммунологические расстройства, включающие аутоиммунные заболевания, такие как гнездная алопеция, аутоиммунный миокардит, базедова болезнь, эндокринная офтальмопатия, склероатрофический лишай, рассеянный склероз, псориаз, системная красная волчанка, системный склероз, заболевания щитовидной железы (например, зоб и тиреоидит Хашимото (лимфаденоидный зоб); поражение легкого (острое геморрагическое поражение легкого, синдром Гудпасчера, острая ишемическая реперфузия), дисфункция миокарда, вызванная загрязняющими агентами, связанными с родом деятельности, или из окружающей среды (например, синдром чувствительности к токсичному маслу, силикоз), радиационная травма и эффективность ответов по заживлению ран (например, ожогов или термических ран, хронических ран, хирургических ран и повреждений спинного мозга), септицемия, ответ острой фазы (например, лихорадочный ответ), общий воспалительный ответ, острый воспалительный-дистресс ответ, острый системный воспалительный ответ, заживление ран, адгезия, иммуно-воспалительный ответ, нейроэндокринный ответ, развтие лихорадки и устойчивость к лихорадке, ответ острой фазы, стрессовый ответ, чувствительность к заболеванию, стресс, связанный с повторными движениями, лучеплечевой бурсит, и устранение боли и болевого ответа, но не ограничивается ими.
Способы лечения могут включать введение терапевтически эффективного количества любого одного или более чем одного из описанных здесь растворимого гликопротеина CD52, слитого белка, полинуклеотида, вектора, клетки, популяции клеток, среды клеток или фармацевтической композиции, или любого агента, способного увеличивать уровень экспрессии CD52 в клетке, субъекту, нуждающемуся в этом.
«Терапевтически эффективное количество» может определять медицинский работник, и оно может варьировать от одного пациента к другому, в зависимости от таких факторов, как возраст, масса, пол и другие факторы.
Способы диагностики
На основе открытия авторов изобретения того, что растворимый CD52 является медиатором функции Treg, согласно настоящему раскрытию также предложены способы определения чувствительности субъекта к любому заболеванию или состоянию, опосредованному эффекторными Т-клетками, воспалению или сепсису, как здесь описано. Диагностические способы могут быть основаны на детекции любого одного или более чем одного из уровня растворимого CD52, частоты клеток CD52hi и функции клеток CD52hi в образце, взятом от субъекта.
Уровень растворимого CD52 можно определять любым подходящим способом, известным в данной области. Например, уровень растворимого CD52 можно определять иммуноанализом с использованием антител, которые связываются с растворимым CD52. Подходящие антитела включают гуманизированное моноклональное антитело крысы CAMPATH-1G, флуоресцентно меченые мышиные моноклональные антитела к человеческому CD52 (такие как CF1D12), кроличье поликлональное антитело к CD52 (Santa Cruz Biotechnology, Santa Cruz, США) и другие.
Частоту клеток CD52hi можно детектировать, например, посредством детектирования уровня мембраносвязанного CD52 в образце, посредством детектирования уровня экспрессии белка CD52 в образце и/или посредством детектирования уровня экспрессии мРНК CD52 в образце.
Функцию клеток CD52hi можно определять с использованием любого подходящего способа, включая любой из раскрытых здесь способов.
Диагностические способы можно проводить на любом подходящем образце, взятом от субъекта. В одном примере образец берут от субъекта-млекопитающего, такого как человеческий субъект, и он может происходить из целого ряда источников, включающих, например, одноядерные клетки периферической крови (РВМС), лейкоферез или аферез продукта крови, костный мозг, кровь спинного мозга, печень, тимус, биопсию ткани, опухоль, ткань лимфатического узла, лимфоидную ткань, ассоциированную с кишечником, ткань лимфатического узла, ассоциированного со слизистой, ткань селезенки или любую другую лимфоидную ткань, или из любого места заболевания, включающего поджелудочную железу. В предпочтительном воплощении клеточный образец поисходит из РВМС из образца крови, полученного из периферической крови субъекта.
Диагностические способы могут включать детектирование уровня любого одного или более чем одного из растворимого CD52, частоты клеток CD52hi и функции клеток CD52hi в образце, содержащем РВМС, которые приводились в контакт с антигеном. Таким образом, данные способы могут включать стадию приведения образца в контакт с антигеном. В одном примере антиген может представлять собой аутоантиген.
В одном конкретном применении раскрытых здесь диагностических способов уровень растворимого CD52, частоту клеток CD52hi и/или функцию клеток CD52hi можно определять для того, чтобы идентифицировать пригодность субъекта для поступления в испытание по скринингу лекарственного средства. Таким образом, если субъект демонстрирует меньший уровень растворимого гликопротеина CD52, меньшую частоту клеток CD52hi и/или пониженную функцию клеток CD52hi, данный субъект может быть идентифицирован как особенно подходящий для включение в испытание по скринингу в отношении лекарственного средства, намеченного к применению в лечении любого заболевания или состояния, опосредованного эффекторными Т-клетками, воспаления или сепсиса, как здесь описано. В одном примере скрининг можно проводить для того, чтобы идентифицировать предполагаемые противодиабетические лекарственные средства (в частности, лекарственные средства против диабета типа 1).
Описанные здесь диагностические способы могут, кроме того, включать стадию определения эталонного уровня растворимого CD52, частоты клеток CD52hi и/или функции клеток CD52hi из образца, взятого от одного или более чем одного здорового субъекта. В качестве альтернативы, эталонный уровень может быть задан заранее. Сравнение уровня растворимого CD52, частоты клеток CD52hi и/или функции клеток CD52hi в образце, взятом от субъекта, с контрольным уровнем может указывать на чувствительность субъекта к любому заболеванию или состоянию, опосредованному эффекторными Т-клетками, воспалению или сепсису, как здесь описано. Например, если уровень растворимого CD52, частота клеток CD52hi и/или функция клеток CD52hi в образце, взятом от субъекта, ниже, чем эталонный уровень, данный субъект может считаться более чувствительным к развитию заболевания или состояния, опосредованного эффекторными Т-клетками, воспаления или сепсиса, как здесь описано. Большее различие между уровнем в образце и эталонным уровнем может указывать на большую чувствительность субъекта к развитию заболевания или состояния, опосредованного эффекторными Т-клетками, воспаления или сепсиса, как здесь описано. Будет понятно, что точные значения, указывающие на повышенный риск развития у субъекта заболевания или состояния, опосредованного эффекторными Т-клетками, воспаления или сепсиса, будут варьировать, в зависимости от целого ряда факторов, включающих конкретное заболевание или состояния, которое диагностируется, образец, используемый для диагностики, популяцию здоровых индивидуумов, используемых для получения эталонного уровня, и других факторов, как будет понятно специалисту в данной области.
Согласно настоящему раскрытию также предложен способ скрининга агента, способного подавлять функцию эффекторных Т-клеток и/или иммунный ответ, включающий приведение в контакт описанных здесь клетки или популяции клеток (например, клетки или поуляции клеток CD52hi) с тестируемым агентом и затем детектирование уровня растворимого CD52, частоты клеток CD52hi и/или функции клеток CD52hi, в котором более высокий уровень растворимого гликопротеина CD52, большая частота клеток CD52hi и/или повышенная функция клеток CD52hi после контакта с тестируемым агентом указывает на то, что тестируемый агент может потенциально подходить для применения в качестве агента, способного подявлять функцию эффекторных Т-клеток и/или иммунный ответ.
В другом воплощении согласно настоящему раскрытию также предложен способ идентификации агента, способного имитировать подавление эффекторных Т-клеток и/или подавление иммунной системы, функции растворимого гликопротеина CD52, включающий приведение в контакт описанных здесь клетки или популяции клеток (например, клетки или популяции клеток CD52hi) с тестируемым агентом и затем детектирование уровня растворимого CD52, частоты клеток CD52hi и/или функции клеток CD52hi, в котором более низкий уровень растворимого гликопротеина CD52, меньшая частота клеток CD52hi и/или пониженная функция клеток CD52hi после контакта с тестируемым агентом указывает на то, что тестируемый агент способен имитировать подавление эффекторных Т-клеток и/или подавление иммунной системы, функции растворимого гликопротеина CD52.
Изобретение теперь будет дополнительно описано со ссылкой на следующие неограничивающие примеры.
ПРИМЕРЫ
ЭКСПЕРИМЕНТАЛЬНЫЕ МЕТОДИКИ Доноры крови
Венозную кровь, отобранную в пробирки с гепарином натрия, получали с информированным согласием и одобрением Комитета по исследовательской этике человека от 5 здоровых молодых взрослых (3 мужчин, 2 женщин) и молодого взрослого мужчины, подверженного риску T1D, для всех из которых известно, что они имеют ответы Т-клеток крови на GAD65. Все доноры были вакцинированы к столбнячному анатоксину. Одноядерные клетки периферической крови (РВМС) выделяли на Ficoll/Hypaque (Amersham Pharmacia Biotech AB, Упсала, Швеция), дважды промывали в фосфатно-солевом буферном растворе (PBS), соответствующем тоничности человека, и ресуспендировали в среде Дульбекко, модифицированной по Пскову (Gibco, Мельбурн, Австралия), содержащей 5% объединенной, инактивированной нагреванием человеческой сыворотки, 100 мМ заменимые аминокислоты, 2 мМ глутамин и 5×10-5 М 2-меркаптоэтанол (полная среда Дульбекко, модифицированная по Пскову [IMDM]).
Антитела и другие реактивы
Реактивы и поставщики были следующими: флуоресцентно меченые мышиные моноклональные антитела к человеческому CD52 (клон CF1D12) и CD24 (клон SN3) (Caltag), FoxP3, GITR, ICOS, CD25, CD127 и человеческому Siglec-10 (клон 5G6) (Biolegend, Сан-Диего, Калифорния, США); мышиный lgG3 (Caltag); поликлональное антитело кролика к CD52 (Santa Cruz Biotechnology, Санта-Круз, Калифорния, США); конъюгированное с HRP (пероксидаза хрена) лошадиное антитело против IgG кролика и антитело против мышиного IgG (Cell Signaling, Arundal, OLD, Австралия); реактив ECL (GE Healthcare, Rydalmere, NSW, Австралия), гуманизированное крысиное моноклональное антитело (CAMPATH-1G) к CD52 (Bayer Healthcare, Pymble, NSW, Австралия), мышиные моноклональные антитела к человеческому IFN-γ (Mabtech, Sydney, NSW, Австралия) и IL-10Ra (клон 37607), антитело козы против человеческого TGF-βRII и аффинно очищенное антитело козы к человеческому Siglec-10 и рекомбинантному человеческому Siglec-10-Fc (R&D Systems, Minneapolis, MN); IL-2 (доклинический репозиторий NCIBRB, Rockville, MD); синтетический человеческий пептид CD52 (GL Biochem, Шанхай); индометацин, метиловый эфир нитрон-аргинина, 1-метил^1-триптофан, SCH58261 (антагонист рецептора аденозина А2А) (Sigma-Aldrich, St. Louis, МО, США); сукцинимидиловый эфир карбоксифлуоресцеиндиацетата (CFSE) (Molecular Probes, Eugene, OR, США); нейраминидаза (С.perfringens типа V) (Sigma-Aldrich, Castle Hill, Австралия); 3Н-тимидин (ICN, Сидней, Австралия); 0,4 мкм трансвелы Corning Costar (Crown Scientific, Minto, NSW, Австралия); белок G и А-сефароза (WEHI Monoclonal Lab, Bundoora, Виктория, Австралия), (1,10-фенантролиновые) ингибиторы фосфолипазы С (U7322) и D (Sigma-Aldrich Pty. Ltd. NSW, Австралия), фосфолипаза С (Molecular Probes, Eugene, OR, США), ПНГаза F (New England Biolabs, Ipswich, MA, США), стреп-тактин сефароза (IBA GmbH Гëттинген, Германия). Столбнячный анатоксин (ТТ) был любезно предоставлен CSL (Parkville, Victoria, Австралия). Рекомбинантный GAD65, продуцированный в бакуловирусе и очищенный, как описано (Bach et а/., 1997), приобретали у Др. Peter Van Endert, Hopital Necker, Париж. Концентрация эндотоксина маточного раствора GAD65, измеренная посредством анализа с лизатом Limulus (BioWhittaker, Walkerville, MD, США), составляла 1,2 EU/мг/мл. ТТ и GAD65 использовали в концентрациях 10 львиных флоккулирующих единиц (LFU)/Mn и 5 мкг/мл соответственно, если не утверждается иное. Цитокины и растворимый рецептор α IL-2 (интерлейкин-2) (CD25) анализировали в средах посредством мультиплексных наборов шариков MAP (Abacus ALS, Брисбен, Австралия).
Статистический анализ
Повторности выражали как среднее значение плюс/минус стандартная ошибка среднего. Значимость между группами определяли непарным (2-сторонним) t-критерием Стьюдента с использованием GraphPad Prism в версии 3.0сх для Macintosh (GraphPad Software Inc., Сан-Диего, Калифорния).
Пример 1: анализ САР65-специфичных клонов Т-клеток CD4+
Способы
GAD65-специфичные клоны Т-клеток CD4+, полученные ранее и подвергнутые скринингу на GAD65-специфичную супрессорную функцию, оттаивали и культивировали, как описано (Dromey er al., 2011). Исходно супрессорные и несупрессорные клоны подвергали скринингу на поверхностные маркеры против чипа с твердофазными антителами (Medsaic Ptd Ltd, Сидней, Австралия) (Belov et al., 2003). Клоны (1×106) брали непосредственно из культуры и анализировали в состоянии покоя или после стимуляции связанным с планшетом антителом против CD3 (5 мкг/мл) в течение 24 ч. Для фенотипирования проточной цитометрией клетки окрашивали на льду подходящими концентрациями меченых антител. Объединяли окрашивание на внутриклеточный FoxP3 и внутриклеточный CTLA-4.
Результаты
Пары для скрининга автологичных супрессорных и несупрессорных клонов на различия в фенотипе поверхности с использованием чипа с антителами против CD выявили то, что активированные супрессорные клоны, как было обнаружено, постоянно имеют более высокую экспрессию CD52, причем данный результат был подтвержден проточной цитометрией (см. Фиг. 1). Таким образом, CD52 был идентифицирован как потенциальный маркер Treg клеток.
Пример 2: анализ Т-клеток крови CD4+ CD52hi
Способы
РВМС, окрашенные сукцинимидиловым эфиром карбоксифлуоресцеина (CFSE), культивировали в IMDM в 96-луночных круглодонных планшетах с или без GAD65 или ТТ в количестве 2×105 клеток в 200 мкл в повторное™ 6. Через 7 суток повторное™ объединяли, промывали в 0,1% BSA-PBS и окрашивали на льду антителами против человеческого CD4-PE, -РЕСу7 или -АРС и CD52-PE (клон CF1D12). Жизнеспособные (негативные в отношении пропидиййодида) клетки CD4+ CFSEтусклый, которые претерпели деление в ответ на GAD65, сортировали в FACSAria (BD Biosciences, North Ryde, NSW, Автралия) на фракции с экспрессией CD52 от наивысшей до самой низкой, и одиночные клетки клонировали, как описано (Dromey et al., 2011). Затем, в ответ на GAD65 или ТТ, популяции CD52hi и CD52lo, соответствующиеверхним 10% и нижним 10% экспрессии CD52 соответственно на неподелившихся клетках CD4+, сортировали для дальнейшего исследования. Данные пороги отсечения выбирали, поскольку большинство полученных GAD65-специфичных супрессорных клонов происходили из верхних 10% клеток CD52+ (см. Таблицу 1).
С использованием РВМС от того же самого донора в течение 4 последовательных недель коэффициент вариации между анализами отношения CD52hi к CD52lo в ответ на GAD65 составлял 21,8%. РВМС в состоянии покоя сортировали на клетки CD4+ CD52hi и CD52lo и также собирали несортированные клетки в качестве контроля. В отдельных экспериментах до мечения CFSE РВМС обедняли клетками CD25+ посредством селекции AutoMACS (Miltenyi Biotec); соответствующие по изотипу моноклональные антитела использовали в качестве контрольных «обеднений».
Функцию GAD65- или ТТ-активированных клеток CD4+ CD52hi и CD52lo анализировали двумя способами. Во-первых, сортированные клетки CD52hi или CD52lo сокультивировали с ТТ-активированными Т-клетками CD4+ в соотношении 1:1 (1×104/лунку) в 6 лунках 96-луночного планшета. Каждая лунка также содержала 5×104 облученных аутологичных РВМС в качестве АРС и ТТ для стимуляции пролиферации аутологичных ТТ-активированных Т-клеток CD4+. GAD65 добавляли в 3 из 6 лунок для повторной стимуляции сортированных клеток. В качестве контроля, облученные РВМС также культивировали с или без GAD65. Через 48 ч добавляли в каждую лунку 3Н тимидин (37 кБк), и клетки отбирали через 16 ч. Во-вторых, сортированные клетки CD4+ CD52hi и CD52lo (5-20000 каждых) культивировали одни или в комбинации в соотношении 1:1 в 6 повторных лунках 96-луночного планшета ELISpot (Millipore PVDF MutliScreen HTS), содержащих предварительно связанное антитело против IFN-γ. Каждая лунка также содержала четырехкратное количество облученных аутологичных РВМС в качестве АРС. GAD65 или ТТ добавляли в 3 из 6 лунок для повторной стимуляции сортированных клеток. Через 24 ч клетки удаляли посредством промывки, и пятна проявляли путем инкубирования с биотинилированным вторым антителом, с последующей инкубацией с со стрептавидином-щелочной фосфатазой и цветным реактивом BCIP/NBT. Результаты выражены в виде пятен IFN-γ/5000 клеток CD4+.
Результаты
Обнаружили, что большинство (22/29, 76%) СА065-специфичных супрессорных клонов происходили из СА065-активированных Т-клеток CD4+ с наивысшей экспессией CD52 (верхние 10%) (Таблица 1). Таким образом, супрессорные клоны, по-видимому, происходили из первичных Т-клеток крови CD4+ CD52hi, а не представляли собой артефакт условий клонирования.

*РВМС от здорового индивидуума, для которого известно, что он имеет GAD65-peaKTHBHbie Т-клетки, метили CFSE и инкубировали с GAD65 в течение 7 суток. Из каждой фракции CD52+ в лунки 96-луночных планшетов посредством FACS сортировали 240 одиночных, жизнеспособных (негативных в отношении пропидиййодида) клеток CD4+ CFSEтусклые, которые подверглись делению, и клонировали, как описано ранее (Dromey et al, 2001).
† Соответствующие экспрессии CD52 на неподелившихся Т-клетках CD4+.
Поскольку большинство GAD65-специфичных супрессорных клонов CD4+ происходили из поделившихся клеток с экспрессией CD52, соответствующей верхним 10% неподелившихся клеток CD4+, данный порог мог быть использован для определения популяции CD4+ CD52hi после активации. При повторной активации GAD65 сортированные клетки CD4+ CD52hi, но не CD52lo подавляли пролиферацию аутологичных ТТ-специфичных Т-клеток CD4+ (Фиг. 2А). Для того чтобы убедиться, что подавление было специфичным для клеток CD52hi и не обусловленным способом их отбора, GAD65-активированные клетки CD4+ сортировали на высокую экспрессию двух других GPI-заякоренных гликопротеинов - CD24 и CD59, а также на CD62L, HLA-DR, CD80 и ICOS, но данные популяции не подавляли пролиферацию ТТ-специфичных Т-клеток (данные не показаны).
Функциональные различия между сортированными Т-клетками CD4+ CD52hi и CD52lo после реактивации GAD65 также были продемонстрированы посредством анализа ELISpot. Меньшая доля клеток CD52hi, чем CD52lo секретировала IFN-γ, и добавление CD52hi к клеткам CD52lo уменьшало число клеток, секретирующих IFN-γ, в ответ на повторную активацию [сравните CD52hi плюс CD52lo (р меньше 0,002) с CD52lo плюс CD52lo клетками (р меньше 0,0002) на Фиг. 2Б]. Подавление не было уникальным для Т-клеток CD4+ CD52hi, активированных GAD65, и также наблюдалось при использовании столбнячного анатоксина (ТТ) в качестве активирующего антигена (Фиг. 2В). Поскольку ответы Т-клеток на ТТ были сильнее, в последующих исследованиях, главным образом, использовали ТТ в качестве антигена. Дополнение низкой концентрацией IL-2 (10 U/мл) увеличивало количество и CD52hi, и CD52lo клеток, секретирующих IFN-γ в ответ на повторную активацию, но не изменяло подавление клетками CD52hi (Фиг. 2В). Клетки CD4+ CD52hi, которые сортировали из неактивированных, полиспецифичных РВМС, демонстрировали слабое, обычно значимое подавление Т-клеток, активированных GAD65 или ТТ (данные не показаны). Однако после активации антигеном супрессорные клетки CD4+ CD52hi с наибольшей вероятностью происходили из предсуществующих клеток CD4+ CD52hi, поскольку обеднение данными клетками РВМС в состоянии покоя увеличивало ответ остальных Т-клеток на GAD65 (Фиг. 2Г).
Пример 3: Т-клетки CD4+ CD52hi отличаются от Treg клеток CD4+ CD25+
Способы
РВМС метили антителом против CD25α и обедняли от клеток CD25hi на колонке AutoMACS (84% по сравнению с изотипическим контрольным антителом «обеднение»). Затем клетки метили CFSE и инкубировали с ТТ в течение 7 суток перед сортировкой на клетки CD52hi и CD52lo, реактивировали ТТ и анализировали анализом ELISpot.
Результаты
После обеднения клетками CD25hi доля поделившихся клеток CD4+ CD52hi в ответ на ТТ возрастала (18,1% по сравнению с 11,8% с контрольным обеднением), но их супрессорная функция после повторной активации ТТ оставалась неизменной (Фиг. 3). Таким образом, супрессорные клетки CD4+ CD52hi, по-видимому, не происходят из популяции Т-клеток CD4+ CD25+.
Пример 4: фенотипический анализ Т-клеток CD4+ CD52hi
Способы
Определенная проточной цитометрией экспрессия (A) CD25α, (Б) FoxP3, (В) поверхностного и (Г) внутриклеточного CTLA-4, (Д) GITR, (Е) CD127, (Ж) CD24 и (3) CD59 на поделившихся Т-клетках CD4+ CD52hi (черная линия) и CD52lo (серая линия) после инкубации РВМС с ТТ в течение 7 суток. Окрашивание изотипическим контрольным антителом показано в виде серого закрашивания. Результаты представляют 5 индивидуумов.
Результаты
Treg клетки CD4+ CD25+ имеют высокую экспрессию CD25, FoxP3, CTLA-4 и белка, родственного рецептору фактора некроза опухоли, индуцируемого глюкокортикоидами (GITR), (Sakaguchi et al., 2008; Shevach, 2006) и низкую экспрессию CD127 (Seddiki et al., 2006; Liu et al., 2006). В отличие от этого, за исключением более высокой экспрессии GITR, Т-клетки CD4+ CD52hi имели аналогичную экспрессию CD25, FoxP3 и CTLA-4 и существенно более высокую экспрессию CD127 по сравнению с Т-клетками CD4+ CD52lo (Фиг. 4). Экспрессия заякоренного GPI гликопротеина, CD24, структурно родственного CD52 (Tone et al., 1999), была выше на Т-клетках CD4+ CD52hi, но это было не так для заякоренного GPI CD59 (Фиг. 4) или CD73, или для CD103, CD40, интегрина β7, ICOS и PD-1 (данные не показаны). Таким образом, Т-клетки CD4+ CD52hi представляют собой новую популяцию супрессорных клеток, которые не характеризуются экспрессией маркеров, используемых для определения человеческих Treg клеток CD4+ CD25+, и которые детектируются в контексте активации антигеном, подразумевая то, что они содействуют гомеостазу Т-клеток во время деления Т-клеток.
Пример 5: анализ экспрессии генов Т-клеток CD4+ CD52hi
Способы
Экспрессию гена CD52 и генов белков, для которых было обнаружено, что они имеют повышенную экспрессию на Т-клетках CD4+ CD52hi, исследовали количественной ПЦР-ОТ (полимеразная цепная реакция, сопряженная с обратной транскрипцией) в реальном времени. Т-клетки CD4+ CD52hi и CD52lo, меченные CFSE, от трех индивидуумов сортировали через 7 суток после активации GAD65. Общую РНК экстрагировали из клеток набором RNAeasy Mini (Quiagen, Мельбурн, Австралия), обрабатывали ДНКазой, не содержащей РНКазу (Qiagen) и количественно измеряли биоанализатором Agilent 2100. кДНК подвергали обратной транскрипции из 10 нг РНК/реакцию. Праймеры для ПЦР, сконструированные с использованием программы PrimerExpress и синтезированные Sigma-Aldrich (Castle Hill, NSW, Австралия) представляли собой:
CD52 F: САА ACT GGA СТС ТСА GGA САА A (SEQ ID NO: 8)
CD52 R: САА CTG AAG CAG AAG AGG TGG A (SEQ ID NO: 9)
FOXP3 F: ATG GTT TCT GAA GAA GGC AAA С (SEQ ID NO: 10)
FOXP3 R: GGA СТА CTT CAA GTT CCA САА CA (SEQ ID NO: 11)
CTLA-4 F: AAC СТА CAT GAT GGG GAA TGA G (SEQ ID NO: 12)
CTLA-4 R: TTA CAT AAA TCT GGG TTC CGT T (SEQ ID NO: 13)
GITR F: GGG AAA TTC AGT TTT GGC TTC (SEQ ID NO: 14)
GITR R: АСА GCG TTG TGG GTC TTG TT (SEQ ID NO: 15)
CD127 F: CCT TTT GAC CTG AGT GTC GTC T (SEQ ID NO: 16)
CD127 R: CGT CCA TTT GTT TTC АТС CTT T (SEQ ID NO: 17)
Мастер микс для ПЦР Power SYBR Green был от Applied Biosystems. Образцы кДНК в тройной повторности подвергали 40 циклам амплификации на установке ABI Prism 7900 согласно протоколу изготовителя. Экспрессию мРНК, нормированную к экспрессии эндогенного β-актина, количественно измеряли методом сравнительного критического порога (Ct) согласно формуле 2-ΔΔCt, как описано в Бюллетене 2 для пользователей от ABI (docs.appliedbiosystems.com/pebiodocs/04303859.pdf).
Результаты
Согласно анализу экспрессии посредством проточной цитометрии транскрипты CD52, CD127 и GITR имели более высокий уровень в клетках CD52hi, чем в клетах CD52lo (Фиг. 5).
Пример 6: на подавление клетками CD52hi не влияет уровень экспрессии CD24
Способы
Для того чтобы проанализировать экспрессию структурно родственного заякоренного GPI гликопротеина CD24, РВМС, меченные CFSE, инкубировали с ТТ в течение 7 суток и сортировали на Т-клетки CD4+ CD52hiCD24lo, CD52hiCD24hi, CD52loCD24lo and CD52loCD24hi. Каждую популяцию (5000 клеток) инкубировали с сортированными клетками-респодерами CD52lo (5000) и облученными РВМС (20000) и анализировали посредством анализа ELISpot. Результаты представляют собой среднее значение плюс стандартную ошибку среднего трех повторностей.
Результаты
Экспрессия заякоренного GPI гликопротеина, CD24, структурно родственного CD52 (Tone et al, 1999), была выше на Т-клетках CD4+ CD52hi. Несмотря на то, что активированные антигеном Т-клетки CD4+ CD24hi, в отличие от Т-клеток CD4+ CD52hi, не были подавляющими, было важно определить, лучше ли CD24 описывал Т-клетки CD4+ CD52hi с супрессорной функцией. РВМС, активированные ТТ, сортировали на четыре отличные популяции CD4+согласно экспрессии и CD52, и CD24, и затем тестировали на супрессорную функцию после повторной активации ТТ. Это выявило то, что на подавление клетками CD52hi не влияет экспрессия CD24 (Фиг. 6).
Пример 7: функция Treg CD52hi не требует контакта клетка-клетка
Способы
Исследовали поглощение 3Н-тимидина (имп./мин) активированными ТТ и сортированными клетками CD4+ CD52hi и CD52lo либо объединенными, либо разделенными полупроницаемым 0,4 мкм трансвелом, и повторно активированными ТТ. РВМС, меченные CFSE, инкубировали с ТТ в течение 7 суток и сортировали на клетки CD4+ CD52hi и CD52lo. Сортированные клетки (по 100000 каждых) инкубировали с облученными аутологичными РВМС (400000) и ТТ в 48-луночных планшетах; в присутствии трансвела оба компартмента содержали облученные РВМС и ТТ. Поглощение 3Н-тимидина клетками в придонном компартменте измеряли через 48 ч.
Результаты
Супрессорная функция активированных антигеном Т-клеток CD4+ CD52hi сохранялась через трансвел без контакта клетка-клетка (Фиг. 7). Таким образом, настоящее раскрытие демонстрирует то, что подавление посредством Treg CD4+ CD52hi, по меньшей мере частично, опосредовано растворимым медиатором. Как обсуждалось в Vignali et al. (2008), ингибирующие цитокины ранее исследовали в качестве возможных растворимых медиаторов подавления посредством Treg, хотя результаты и были неубедительными, и осталось общее ощущение того, что контакт клетка-клетка является важным для супрессорной функции Treg. Раскрытые здесь результаты свидетельствовали о том, что Т-клетки CD4+ CD52hi либо удаляли растворимый фактор, требующийся для функции Т-клеток-респондеров, либо продуцировали растворимый фактор, который подавлял Т-клетки-респондеры.
Пример 8: анализ IL-2 в функции Treg CD52hi
Способы
Роль IL-2 исследовали в целом ряде экспериментов, включающих применение количественной ПЦР-ОТ в реальном времени для определения уровней экспрессии. В анализе количественной ПЦР-ОТ общую РНК экстрагировали из клеток набором RNAeasy Mini (Qiagen, Мельбурн, Австралия), обрабатывали ДНКазой, не содержащей РНКазу (Qiagen) и подвергали количественной оценке биоанализатором Agilent 2100. кДНК подвергали обратной транскрипции из 10 нг РНК/реакцию. Праймеры для ПЦР, сконструированные с использованием программы PrimerExpress и синтезированные Sigma-Aldrich (Castle Hill, NSW, Австралия), представляли собой:
IL-2α ʹ5 TACAGGATGCAACTCCTGTCTT (SEQ ID NO: 18),
ʹ3 GCTCCAGTTGTAGCTGTGTTTT (SEQ ID NO: 19);
IL-27β ʹ5 GCTGTTCTCCATGGCTCCCTAC (SEQ ID NO: 20),
ʹ3 GTCGGGCTTGATGATGTGCT (SEQ ID NO: 21);
IL-12α ʹ5 CTCCAGAAGGCCAGACAAACTC (SEQ ID NO: 22),
ʹ3 CCAATGGTAAACAGGCCTCCAC (SEQ ID NO: 23).
Мастер-микс для ПЦР Power SYBR Green был от Applied Biosystems. кДНК подвергали 40 циклам амплификации на установке ABI Prism 7900 согласно протоколу изготовителя. Экспрессию мРНК, нормированную к экспрессии эндогенного β-актина, количественно оценивали методом сравнительного критического порога (Ct) согласно формуле 2-ΔΔCt, как описано в Бюллетене для пользователей 2 от ABI (docs.appliedbiosystems.com/pebiodocs/04303859.pdf).
Результаты
Потребление или деградацию IL-2 Т-клетками CD4+ CD52hi считали невероятным механизмом подавления по нескольким причинам: i) экзогенный IL-2 не преодолевал подавление (Фиг. 2В); ii) количественная ПЦР-ОТ выявила то, что экспрессия гена IL-2 была на самом деле выше в клетках CD52hi; таким образом, после 24 ч повторной активации GAD65 экспрессия мРНК IL-2α в клетках CD52hi относительно CD52lo составляла 1,54±0,15 (среднее плюс/минус sem (стандартная ошибка среднего), n=3), iii) концентрация IL-2 в среде клеток CD52hi была выше, чем для клеток CD52lo, как в состоянии покоя (89,5±4,82 относительно 64,9±3,10 пг/мл), так и после повторной активации GAD65 (138,7±4,16 относительно 82,4±1,78 пг/мл (среднее плюс/минус sem, n=3; Р=0,02, критерий Крускала-Уоллиса); iv) в средах, в которых измеряли IL-2, растворимый рецептор-a IL-2 (CD25) не был детектируемым (данные не показаны). Таким образом, полагали, что удаление IL-2 не является вероятным механизмом подавления посредством Treg CD52hi.
Пример 9: анализ других предположительных медиаторов функции Treg CD52hi
Затем обнаружили, что подавление посредством Treg не изменялось в присутствии агентов, которые блокируют действие или продукцию факторов, для которых сообщалось, что они опосредуют подавление посредством Treg клеток CD4+ (Sakaguchi et al., 2008, 2009; Shevach, 2006, 2009; Vignali et al., 2008). Они включали нейтрализующие моноклональные антитела к IL-10Rα или TGF-βRII одни или в комбинации (по 10 мкг/мл каждого), ингибитор циклооксигеназы-2 (СОХ-2) индометацин (20 мкМ) (который блокирует продукцию простагландина Е2), общий ингибитор синтазы оксида азота - N(G)-монометил-L-аргинин (800 мкМ) (который блокирует продукцию оксида азота), ингибитор индоламин-2,3-диоксигеназы (IDO) - 1-метил-dl-триптофан (200 мкМ) (который блокирует продукцию ингибирующих триптофановых метаболитов) и антагонист рецептора аденозина А2А - SCH58261 (20 мкМ) (который блокирует действие аденозина) (данные не показаны). Недавно было показано, что Treg клетками CD4+ CD25+ (которые также требовали контакта клетка-клетка для подавления) (Collison et al., 2007) секретируется новый подавляющий цитокин, IL-35, гетеродимер субъединиц IL-27β (EE3i3) и IL-12α (р35). Отсутствовала возможность непосредственно измерять IL-35, так как не были доступны антитела к IL-35 или его рецептору. Однако через 24 ч после повторной активации GAD65 экспрессия мРНК IL-27β и IL-12α была ниже в клетках CD4+ CD52hi, чем в CD52lo (0,423±0,188 относительно 1,38±0,224; среднее плюс/минус sem, n=3), указывая на малую вероятность того, что IL-35 объясняет подаление Т-клетками CD4+ CD52hi.
Пример 10: растворимый CD52 является медиатором подавления Treg CD52hi
Способы
РВМС, меченные CFSE, инкубировали с GAD65 в течение 7 суток и сортировали на Т-клетки CD4+ CD52hi и CD52lo. Сортированные клетки повторно активировали GAD65, и собирали среды через 24 ч. Среды 10-кратно концентрировали, фракционировали SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), переносили на PDVF (поливинилиденфторид) мембрану и подвергали блоттингу с поликлональным антителом кролика на CD52 для того, чтобы детектировать присутствие в средах растворимого CD52.
Затем анализировали ингибитор фосфолипазы С - U73122 в качестве потенциального ингибитора продукции растворимого CD52. РВМС, меченные CFSE, инкубировали с ТТ в течение 7 суток и сортировали на Т-клетки CD4+ CD52hi. Сортированные клетки повторно активировали ТТ, среды собирали через 24 часа и подвергали иммуноблоттингу, как описано выше. Отдельно РВМС, меченные CFSE, инкубировали с ТТ в течение 7 суток и сортировали на Т-клетки CD4+ CD52hi и CD52lo, которые затем инкубировали совместно (по 5000 клеток каждого типа) в планшетах ELISpot с облученными РВМС (20000) и ТТ плюс/минус ингибитор фосфолипазы С U73122. Результаты представляют собой среднее плюс sem трех повторностей.
Кроме того, антитело к углеводной группировке CD52 анализировали в качестве другого потенциального ингибитора подавления Т-клетками CD4+ CD52hi, активированными ТТ. Методики были такими же, как описано выше для ингибитора фосфолипазы С U73122, за исключением того, что клетки в анализе ELISpot инкубировали с или без ТТ и либо 10 мкг/мл антитела против CD52 (CF1D12), либо изотипического контрольного (lgG3) моноклонального антитела. Результаты (среднее плюс/минус sem) характеризуют три независимых эксперимента.
Результаты
Иммуноблоттинг выявил то, что CD52 присутствовал в среде Т-клеток CD4+ CD52hi, которые поделились в ответ на GAD65, и его количество увеличивалось после их повторной активации GAD65 (Фиг. 8А). Такой же результат обнаружили с ТТ в качестве антигена, и ингибитор фосфолипазы С, U73122, добавленный до повторной активации ТТ, уменьшал количество CD52 в среде (Фиг. 8Б). Кроме того, ингибирование фосфолипазы С дозозависимо обращало подавление Т-клетками CD4+ CD52hi (Фиг. 8В). Моноклональное антитело CF1D12, которое взаимодействует с концевым углеводом на пептиде CD52 (Hale, 2001), предотвращало подавление Т-клетками CD4+ CD52hi или CD52lo (Фиг. 8Г). В совокупности данные открытия указывали на то, что подавление Т-клетками CD4+ CD52hi было обусловлено растворимым CD52, высвобождаемым расщеплением фосфолипазой в ответ на стимуляцию антигеном.
Пример 11: дальнейший анализ эффекторной функции растворимого CD52
При рассмотрении более обильного источника нативного растворимого CD52 постулировали, что CD52 может спонтанно высвобождаться из некоторых линий клеток, таких как линия клеток В-лимфобластов Дауди, в которых биосинтез GPI является дефектным из-за недостаточности продукта гена PIGY (Ни et al., 2009).
Способы
Среды от сортированных клеток CD4+ CD52hi и CD52lo собирали через 24 часа после повторной активации клеток GAD65 или ТТ. Среды от линий клеток (Дауди, Раджи, Юркат и К562) собирали и 10-кратно концентрировали лиофилизацией. Образцы фракционировали SDS-PAGE и переносили на PVDF мембрану. После блокирования 5% нежирным молоком мембрану инкубировали с кроличьим поликлональным антителом к CD52 (1 мкг/мл), промывали, инкубировали с антителом козы против кроличьего IgG, конъюгированного с пероксидазой хрена, и визуализировали усиленной хемилюминисценцией.
Отдельно РВМС (200000 клеток) культивировали в течение 7 суток в кондиционированной среде IMDM, содержащей 20% клеток Дауди, с ТТ и либо антителом против CD52 (CF1D12), либо изотипическим контрольным антителом (10 мкг/мл). Для обеднения растворимым CD52, среду с клетками Дауди инкубировали в течение ночи с поликлональным антителом кролика против CD52 (1 мкг/мл среды), с последующим осаждением белок G-сефарозой в течение 1 ч при 4°С. Результаты (среднее плюс/минус sem) характеризуют три независимых эксперимента.
Результаты
Скрининг нескольких линий клеток выявил присутствие CD52 в культуральных средах клеток Дауди и К562 (Фиг. 9А). Среда от клеток Дауди подавляла пролиферацию РВМС, активированную ТТ, и подавление обращалось либо антителом CF1D12, либо иммунообеднением (подтвержденным иммуноблоттингом) CD52 (Фиг. 9Б), демонстрируя то, что подавление Т-клеток было обусловлено CD52 в среде. CD52 был растворимым и не присутствовал в экзосомах или мембранных частицах, так как на подавление не влияло центрифугирование среды при 100000 g в течение 30 мин (данные не показаны).
Пример 12: дублирование эффекторной функции растворимого CD52 посредством CD52-FC
Способы
Для дальнейшего исследования иммунодепрессивной функции растворимого CD52 зрелый CD52 поверхности клетки клонировали в лентивирусный вектор в виде слитого белка, находящегося в рамке считывания с Fc фрагментом иммуноглобулина G и С-концевой последовательностью метки Strep для очистки. Конструкцию, содержащую только Fc, клонировали в качестве контроля. Конструкции стабильно экспрессировали в клетках Дауди или временно - в клетках НЕК293Т, и растворимые рекомбинантные белки очищали из среды посредством элюции дезтиобиотином из стрептактиновой смолы.
Схема для конструирования ДНК, кодирующих слитые белки, показана на Фиг. 10. Мутировавший Fc фрагмент человеческого lgG1 (Armour et al., 2003), соединенный с последовательностью сигнального пептида (SigP) CD52, получали посредством ПЦР. Он включал гибкий линкер GGSGG, два сайта расщепления для протеаз Precission и Factor Ха между фрагментом SigP и Fc и последовательность метки Strep II для очистки (Schmidt and Skerra, 2007) на конце Fc фрагмента. Праймеры, обозначенные на Фиг. 10, использованные для получения и клонирования конструкций Fc, были следующими:
1F1: GAAGTTCTGTTCCAGGGGCCCATCGAAGGTCGTGGTG (SEQ ID NO: 24);
1R1: TCATTTTTCGAACTGCGGGTGGCTCCAGGCGCTTTTACCCGGAGACAG (SEQ ID NO: 25);
1F2: GGGGGTTCCGGGGGACTGGAAGTTCTGTTC (SEQ ID NO: 26);
1R2: CTTGATATCGAATTCTCATTTTTCGAACTG (SEQ ID NO: 27);
2F: CG CTGTTACGG ATCCCCACCATG AAG CG CTTCCTC (SEQ ID NO: 28);
2R1: TCCACCG CTACCTCCTG AGG G G CTG CTG GT (SEQ ID NO: 29);
2R2: TCCACCG CTACCTCCTG AG AGTCCAGTTTG (SEQ ID NO: 30).
Конструкцию CD52-Fc, содержащую SigP и внеклеточный домен (ECD) CD52, соединенные с фрагментом Fc, получали ПЦР. Используемыми праймерами были: 2F, 2R1, 1F2 и 1R2. ПЦР-продукты расщепляли BamHI/EcoRI и лигировали в лентивирусный вектор FTGW (Herald MJ et al., 2008). Клоны также подтверждали секвенированием. Лентивирусные частицы получали СаР04-опосредованной трансфекцией клеток НЕК293Т, высеянных в 6 см чашки с 10 мкг ДНК вектора совместно с тремя хэлперными плазмидами (pMDLRRE, pRSV-REV и pVSV-g). Среду культуры клеток, содержащую вирус, собирали через 48 ч после трансфекции и пропускали через 0,45 мкм фильтр. Один миллилитр использовали для трансдукции 1×106 клеток Дауди, выращенных в средах DME (среда Игла, модифицированная по Дульбекко), дополненных 10% FCS (фетальная телячья сыворотка), 100 мМ заменимыми аминокислотами, 2 мМ глутамином и 5×10-5 М 2-меркаптоэтанолом. Клетки подвергали скринингу на наивысшую экспрессию белка посредством внутриклеточного окрашивания и проточной цитометрии. Белки CD52-Fc или контрольный Fc очищали из среды посредством одноэтапной аффинной хроматографии на стрептактиновой смоле и элюции 2,5 мМ дезтиобиотином в 100 мМ Tris-HCI, 150 мМ NaCl, 1 мМ EDTA, рН 8,0, согласно инструкциям изготовителя. После диализа SDS-PAGE выявил одиночные окрашенные кумасси синим полосы предсказанного размера, специфичность которых подтверждали вестерн-блоттингом.
Анализ эффектов рекомбинантных слитых белков с Fc
РВМС (2×105 клеток/лунку) или очищенные Т-клетки CD4+ (5×104 клеток/лунку) в полной среде IMDM - 5% инактивированной нагреванием объединенной человеческой сыворотке инкубировали в круглодонных 96-луночных планшетах с или без 10 Lfu/мл ТТ и разными концентрациями белков CD52-FC или Fc в общем объеме 200 мкл при 37° в 5% CO2-воздухе в течение вплоть до 7 суток. Добавляли 3Н-тимидин (1 мкКи/лунку) и через еще 18 ч клетки отбирали, и измеряли радиоактивность, включенную в ДНК, посредством сцинтилляционного счета. Среду отбирали для анализа цитокинов после 48 ч инкубации. Дендритные клетки (DC) выделяли из РВМС, как описано (Mittag et al., 2011). Вкратце, РВМС сперва обогащали DC посредством обеднения с магнитными шариками клеток, меченных антителами к маркерам линий (CD3, CD19, CD56). Затем клетки окрашивали флуоресцентными антителами к HLA-DR, CD11c, CD1b/c, CD304 и CD14, и сортировали проточной цитометрией для очистки традиционных DC CD1b/c+HLA-DR+CD11c+, плазмацитоидных DC CD304+HLA-DR+CD11 с- и моноцитов CD14+CD16-CD11C+. Очищенные DC предынкубировали с белком CD52-Fc или Fc в концентрации 3,3 мкМ в течение 30 мин при 37°С и дважды промывали. Их затем серийно разводили из 6000 клеток/лунку в 96-луночном круглодонном планшете и инкубировали с меченными CFSE Т-клетками CD4+ (5×104/лунку), выделенными у другого донора. Через 6 суток измеряли ответ аллогенных Т-клеток как частоту делящихся клеток CFSElo, определенную проточной цитометрией.
Как описано выше, РВМС (200000) культивировали с ТТ в течение 7 суток и очищенными Т-клетками CD4+ (20000) с антителом против CD3 (100 нг/мл) и антителом против CD28 (200 нг/мл) с течение 48 ч, с 4-кратным избытком облученных РВМС в 200 мкл круглодонных лунках в присутствии рекомбинантного белка CD52-FC или Fc в указанных концентрациях. Поглощение 3Н-тимидина измеряли на протяжении последних 16 ч инкубации. Результаты (среднее плюс/минус sem трех повторностей) характеризуют шесть независимых эксприментов.
Среды от РВМС, активированных ТТ плюс/минус 3,3 мкМ белков CD52-Fc или Fc, отбирали после 48 часов инкубации и анализировали на цитокины посредством мультиплексного набора шариков.
CD52-Fc (20 мкг) инкубировали с или без ПНГазы F (1000 единиц) в 20 мкл PBS (фосфатно-солевой буферный раствор) в течение ночи при 37°С для того, чтобы расщепить N-связанный углевод, и реакцию завершали нагреванием при 75°С в течение 10 мин. Конкретно, ПНГаза F расщепляет олигосахариды, связанные с аспарагином, между двумя субъединицами N-ацетилглюкозамина непосредственно по соседству с остатком аспарагина с образованием усеченного углевода с одним остатком N-ацетилглюкозамина, остающимся на аспарагине. РВМС инкубировали с ТТ и обрабатывали или не обрабатывали CD52-FC (конечная концентрация 2,5 мкМ) в течение 7 суток при 37°С, и затем измеряли поглощение 3Н-тимидина, как описано выше.
Результаты
При использовании РВМС пролиферативный ответ Т-клеток на ТТ дозозависимо подавлялся CD52-Fc (Фиг. 11А), и CD52-Fc подавлял секрецию цитокинов, характеризующих разные линии Т-клеток (Фиг. 11 В). Эффект CD52-Fc на функцию Т-клеток был прямым, поскольку он подавлял пролиферацию очищенных Т-клеток CD4+ в ответ на поперечное связывание рецептора Т-клеток антителом против CD3 и костимуляцию антителом против CD28 (Фиг. 11 Б). Доказательство того, что для CD52-Fc не требуется антигенпрезентирующих клеток для подавления Т-клеток было получено посредством демонстрации того, что воздействие CD52-Fc на очищенные дендритные клетки не влияет на их способность вызывать ответ аллогенных Т-клеток (Фиг. 13).
Как показано (Фиг. 8Г), способность антитела CF1D12 блокировать подавление нативным CD52 подразумевала то, что подавление может быть опосредовано углеводной группировкой CD52. Для проверки ее роли в рекомбинантном CD52-FC, N-связанный углевод расщепляли эндогликозидазой ПНГазой F. Это снижало молекулярную массу CD52-FC от ~48 до ~30 кДа, что предсказывается, исходя из потери углевода, и уменьшало его подавляющий эффект (Фиг. 11Г), подтверждая роль углеводной группировки в опосредовании подавляющего эффекта растворимого CD52.
Пример 13: дальнейший анализ функции углевода CD52
Способы
Для дальнейшего исследования роли углеводной группировки CD52 в опосредовании подавления Т-клеток, CD52-Fc (3,3 мкМ) инкубировали с нейраминидазой (1 единица) или лишь буфером-носителем в 20 мкл в течение 30 мин при 37°С, как рекомендовано поставщиком. РВМС затем инкубировали с ТТ плюс/минус GD52-Fc (конечная концентрация 3,4 мкМ), обработанным или не обработанным нейраминидазой, в планшете ELISpot и проявляли через 24 ч при 37°С на пятна IFN-γ.
Отдельно РВМС инкубировали в планшете ELISpot с ТТ и CD52-FC (3,4 мкМ) и разными концентрациями аффинно очищенного антитела козы к внеклеточному домену Siglec-10 или Fc (3,4 мкМ) плюс/минус антитело, или разными концентрациями рекомбинантного Siglec-10-Fc до переноса неприкрепившихся клеток на планшет ELISpot на 24 ч до проявления пятен IFN-γ.
Для исследования возможности того, что растворимый CD52 может действовать через рецепторы Siglec, отличные от Siglec-10, Т-клетки CD4+ (20000) инкубировали в тройной повторности в лунках планшета ELISpot при 37°С с ТТ, совместно с CD52-FC или Fc (3,4 мкМ каждого) и антителами против человеческого Siglec (10 мкг/мл каждого) или рекомбинантным человеческим Siglec 2-Fc (20 мкг/мл), как показано на Фиг. 12Д. Через 20 ч лунки промывали и проявляли на пятна IFN-γ.
Результаты
Обработка нейраминидазой для удаления концевых сиаловых кислот уменьшала подавление посредством CD52-Fc (Фиг. 12А). Предполагается, что структура сложного полилактозамина углевода CD52 заканчивается α2-6 и возможно α2-3 сиаловыми кислотами, декорирующими галактозу, образующую β1-4 связь с N-ацетилглюкозамином (Treumann et al., 1995). Эта сиалозидная последовательность распознается человеческим связывающим сиаловую кислоту lg-подобным лектином-10 (Siglec-10), трансмембранным рецептором поверхности клетки и членом надсемейства иммуноглобулинов, несущим два цитоплазматических ингибирующих мотива иммунорецептора на основе тирозина (ITIM) (Munday et al., 2001; Crocker et al., 2007). Несмотря на то, что Siglec-10 не был детектирован на мышиных Т-клетках (Crocker et al., 2007), и некоторые другие Siglec не экспрессируются на человеческих Т-клетках (Nguyen era/., 2006), авторы изобретения обнаружили, что Siglec-10 экспрессировался на человеческих Т-клетках CD4+ и подвергался повышающей регуляции посредством активации (Фиг. 12Б). Примечательно то, что подавление функции Т-клеток посредством CD52-FC уменьшалось либо антителом к внеклеточному домену Siglec-10 (Фиг. 12 В, 12Д), либо растворимым рекомбинантным Siglec-10-Fc (Фиг. 12Г). Те же самые концентрации Siglec-10-Fc также уменьшали подавление Т-клетками CD4+ CD52hi (данные не показаны), указывая то, что и рекомбинантный и нативный CD52 распознают Siglec-10. Подавление Т-клеток посредством CD52-Fc не уменьшалось в той же самой степени антителами к другим Siglec, отличным от Siglec-10, или рекомбинантным человеческим Siglec 2-Fc. Данные открытия демонстрируют то, что подавление посредством CD52 могло бы объясняться, по меньшей мере частично, его взаимодействием с Siglec-10.
Пример 14: Т-клетки CD52hi защищают против аутоиммунного заболевания
Материалы и методы
Мыши
Мышей C57/BI6, NODLt и RIP.B7/ NODSCID размножали и содержали в Walter and Eliza Hall Institute of Medical Research. Трансгенные мыши с OVA (овальбумин)-специфичным ограниченным классом I TCR (Hogquist et al., 1993) и трансгенные мыши с OVA-специфичным ограниченным классом II TCR (Barnden et al., 1998) были описаны ранее. Репортерные мыши Foxp3GFP были предоставлены Др. Yifan Zhang.
Реактивы, антитела и проточная цитометрия
Клетки культивировали в средах RPMI, дополненных 10% FCS, 1:100 добавкой GIBCO™ GlutaMAX™-I (Invitrogen), 1:1000 2-меркаптоэтанолом (Sigma), 1:100 NEAA (gibco). Моноклональные антитела против CD52 приобретали у MBL International, клон BTG-2, конъюгированные с РЕ или немеченые. Для анализа вестерн-блоттингом использовали поликлональное антитело против CD52, приобретенное у Santa Cruz Biotechnology, Inc (sc27555). Моноклональное антитело против CD4 (L3T4, клон GK1.5) и антитело против CD8a (Ly-2, клон 53-6.7) приобретали у eBiosciences. Антитело против CD25 (клон Зс7) приобретали у BioLegend. Антитело против CD3-FITC и набор для окрашивания FoxP3 приобретали у eBioscience. Антитело против CD3 (клон 2 с11), антитело против CD28 (клон 37.51) и изотипические контрольные моноклональные антитела были от WEHI Monoclonal Antibody Lab. Анализы проточной цитометрией проводили на FACSAria с использованием программы FACS Diva. Клетки сортировали с использованием сортировщика клеток MoFlow (Cytomation, Fort Collins, CO).
Выделение клеток
Селезенки отбирали и пропускали через 70 мкм сетку, обрабатывали буфером для лизиса эритроцитов и промывали. Для активации клеток спленоциты культивировали на планшете со связанным антителом против CD3 (2 мкг/мл) плюс растворимое антитело против CD28 (1 мкг/мл) в течение 3 суток. Спленоциты OTI или ОТН инкубировали с 0,5 мкг/мл пептида OTI или 5 мкг/мл OTII в течение 4 суток до анализа. Для экспериментов по сортировке клеток наивные или активированные спленоциты из мышей C57/BI6, OTI или OTII, NODLt или Foxp3-GFP метили либо CD3-FITC (eBioscience), CD4-APC (eBioscience), CD8-APC (eBioscience) и CD52-PE (MBL International). Меченые клетки разделяли с использованием цитометра MoFlow, и чистота составляла ~95%. Выделенные клетки либо использовали для очистки РНК, для анализов пролиферации Т-клеток, либо в экспериментах in vivo.
Анализы пролиферации
Сортированные наивные или активированные Т-клетки CD4+ CD52hi или CD8+CD52hi (2×104, в экспериментах с трансвелами - 1×105) культивировали с Т-клетками CD4+CD52lo или CD8+CD52lo в соотношении 1:1 и стимулировали 1 мкг/мл растворимого антитела против CD3 (2с11) плюс (8×104, в экспериментах с трансвелами - 4×105) облученных АРС (антигенпрезентирующая клетка), обедненных Т-клетками (доза облучения 2000 рад (20 Дж/кг)). Между клетками помещали 0,4 мкМ трансвелы (Corning, вставки-трансвелы из поликарбонатной мембраны, Кат. №3413). В экспериментах по блокировке добавляли 15 мкг/мл антитела против CD52 (крысиный lgG2a, MBL International) или изотипического контроля. Анализы пролиферации проводили в течение 72 ч в 96-луночных круглодонных планшетах в конечном объеме 200 мкл среды RPMI, которая содержала 10% фетальной телячей сыворотки. В течение последних 10 часов эксперимента добавляли 1 мкКи/лунку [3Н]-тимидина, и включение тимидина измеряли сцинтилляционным счетом. Альтернативно, в качестве индикатора пролиферации использовали меченные CFSE клетки-респондеры. Наивные спленоциты или Т-клетки CD4+CD52lo или CD8+CD52lo суспендировали в теплом PBS плюс 0,1% BSA (бычий сывороточный альбумин) при количетве клеток 10×106 на мл. Добавляли 5 мкМ CFSE, и проводили быстрое ресуспендирование. Клетки инкубировали при 37 градусах в течение 5 мин перед промывкой 3 раза холодным буфером, содержащим по меньшей мере 10% BSA или FCS. Клетки-респондеры, меченные CFSE, инкубировали с Т-клетками CD4+CD52hi или CD8+CD52hi (плюс дополнительные контроли) в течение вплоть до 7 суток и анализировали с использованием FACS Aria.
Двухцветный анализ
Т-клетки CD4+CD52hi или CD8+CD52hi окрашивали маркером клеточного деления РКН.26 (Sigma) согласно рекомендациям изготовителей. Вкратце, вплоть до 1×107 клеток ресуспендировали в разбавителе С (предоставленном в наборе) и смешивали с 2 мкМ РКН.26 в течение 4 мин при комнатной температуре. Клетки 3 раза промывали буфером, содержащим по меньшей мере 10% FCS. Отвечающие Т-клетки CD4+CD52lo или CD8+CD52lo окрашивали CFSE, как описано выше. Клетки культивировали одиночно или совместно в течение 4-6 суток.
ПЦР-ОТ в реальном времени
Общую РНК получали из сортированных Т-клеток с использованием набора RNeasy от Qiagen. кДНК синтезировали с использованием олиго-dT праймеров (Qiagen, 0,4 мкг/мл) и обратной транскриптазы M-MLV (4000 U, Applied Biosystems), следуя рекомендациям изготовителей. ПЦР-ОТ в реальном времени проводили в циклере ABI PRISM 7900 (Applied Biosystems) с использованием набора Quantitect SYBR Green PCR (Qiagen, Кат. №204143) и специфичных праймеров, оптимизированных для амплификации 100-150 п. н. фрагментов разных генов. Порог был установлен в линейной части кривой амлификации, и для каждого гена было рассчитано число циклов, необходимое для достижения порога. Относительную экспрессию мРНК определяли посредством нормирования к контрольному гену (β-актин или RPS9).
Последовательности праймеров представляют собой:



Адоптивный перенос спленоцитов, обедненных CD52hi, реципиентам NOD
Общие спленоциты, обедненные CD52hi, или спленоциты, обедненные Т-клетками CD3+, CD4+ или CD8+CD52hi, внутривенно инъецировали мышам-реципиентам. Мыши-реципиенты представляли собой либо облученных самцов мышей NOD (8-недельные самцы мышей NOD, доза облучения 750 рад (7,5 Дж/кг), за 4 часа до переноса 1-1,2×107 клеток), либо 8-недельных мышей RIP.B7/NOD.SCID, получающих 2×106 клеток. Мышей отслеживали на признаки диабета, измеряя уровень глюкозы в моче 3 раза в неделю с использованием Diastix от Bayer. Если уровень глюкозы в моче превышает 20 мМ, измеряют уровень глюкозы в крови. Мышей определяли как диабетических, если последовательные показатели уровня глюкозы в крови превышают 20 мМ концентрацию глюкозы.
Бальная оценка инсулита
Через 4 недели после адоптивного переноса клеток мышей умерщвляли. Поджелудочные железы отбирали и фиксировали в течение ночи в растворе Боуина и затем переносили в 70% этанол. Фиксированные поджелудочные железы заливали в парафиновые блоки, нарезали минимум 12 8-м км срезов, расположенных на расстоянии по меньшей мере 150 мкм друг от друга. Срезы окрашивали гематоксилином-эозином и оценивали на частоту возникновения и тяжесть инсулита посредством световой микроскопии независимо двумя исследователями. Наблюдали минимум 10 островков от каждой мыши, и подвергали бальной оценке степень инфильтрации одноядерных клеток с использованием следующего ранжирования: 0 - нет инфильтрации, 1 - инфильтрат около протоков, 2 - инфильтрат около островков, 3 - инфильтрат внутри островков, 4 - разрушение бета-клеток.
Результаты
Перенос обедненных CD52hi лимфоцитов селезенки от 8-недельных мышей NOD мышам NOD.SCID приводит к быстрому началу диабета; необедненные клетки не давали эффекта (Фиг. 14). Перенос обедненных CD52hi Т-клеток CD3+ ускорял начало диабета, но не был таким эффективным, как обеднение клетками CD52hi общих лимфоцитов (Фиг. 15). Таким образом, было показано, что лимфоциты CD52hi защищают против аутоиммунного диабета.
Пример 15: частота Т-клеток CD52hi CD4+, генерированных в ответ на стимуляцию GAD65, снижается при диабете типа 1
РВМС, окрашенные CFSE, культивировали с GAD65 или ТТ в течение 7 суток до определения частоты Т-клеток CD52hi CD4+ посредством анализа проточной цитометрией.
Результаты
Индивидуумы, имеющие или подверженные риску диабета типа 1, имеют меньшее число Т-клеток CD4+ CD52hi, чем здоровые индивидуумы, в ответ на GAD65, но не ТТ (Фиг. 16). Горизонтальная планка предствляет собой медиану для каждой группы. Общие значения Р для дисперсионного анализа определяли посредством критерия Крускала-Уоллиса; затем посредством критерия Данна для множественных сравнений выявляли значимые различия между как пред-T1D, так и T1D по сравнению со здоровыми или T2D при Р меньше 0,05.
Пример 16: подавление Т-клеток клетками CD4+ CD52hi, генерированными в ответ на GAD65, ослабевает при доклиническом диабете типа 1
Способы
РВМС, меченные CFSE, от индивидуумов с аутоантителами на островковые клетки, подверженных риску диабета типа 1, инкубировали с GAD65 в течение 7 суток и сортировали на Т-клетки CD4+ CD52hi и CD52lo согласно описанным здесь способам. Сортированные клетки (5000) инкубировали в планшетах ELISpot с облученными РВМС (20000).
Результаты
Как показано на Фиг. 17, супрессорная функция клеток CD4+ CD52hi, генерированных в ответ на GAD65, ослабевает по сравнению с супрессорной функцией клеток CD4+ CD52hi, генерированных в ответ на ТТ. Результаты характеризуют 6 субъектов, подверженных риску. Таким образом, супрессорная функция клеток CD4+ CD52hi ослабевает при доклиническом T1D.
Пример 17: растворимый CD52 кардинально снижает уровени глюкозы в крови у мышей NOD
Способы
Самок мышей NOD отслеживали путем еженедельного тестирования глюкозы в моче, и диабет диагностировали у мышей с позитивным анализом мочи по концентрации глюкозы в крови больше 14 мМ. Как только подтверждалась гипергликемия, мышам давали либо CD52-Fc, либо Fc, 20 мкг внутрибрюшинно, шесть доз через сутки, и затем отслеживали их концентрации глюкозы в крови дважды в неделю.
Результаты
Было показано, что растворимый CD52-FC снижает уровни глюкозы в крови (Фиг. 18). Как показано, введение CD52-FC имело быстрый и значимый эффект по снижению уровней глюкозы в крови, демонстрируя пригодность растворимого CD52 в качестве терапевтического средства для лечения аутоиммунных заболеваний, таких как диабет типа 1.
Пример 18: развитие диабета у мышей NOD.SCID после переноса от диабетических мышей NOD спленоцитов, обработанных ex vivo hCD52-Fc или Fc
Способы
5×106 спленоцитов от диабетических мышей NOD, обработанных rhCD52-Fc или Fc, инъецировали мышам NOD.SCID. Спленоциты от самок диабетических мышей выделяли и инкубировали либо с 50 мкг/мл рекомбинантного человеческого белка CD52-FC или Fc в течение 1,5 ч в «буфере для CD52» (физиологический раствор, буферизованный Tris, плюс 2 мМ MgCl2, CaCl2 и MnCl2 плюс 5 мМ глюкоза плюс 1% мышиная сыворотка). Клетки ресуспендировали в PBS, и 1×107 клеток инъецировали самцам мышей NOD.SCID (12 на группу).
Результаты
Обработка спленоцитов от диабетических мышей NOD ex vivo CD52-Fc приводила к увеличению выживания без диабета у мышей NOD.SCID, которым имплантировали обработанные спленоциты (Фиг. 19). Это является еще одним другим доказательством терапевтического применения растворимого CD52 для лечения аутоиммунных заболеваний, таких как диабет типа 1.
Пример 19: человеческий CD52-Fc подавляет мышиные клетки ОТ-II
Способы
Трансгенные Т-клетки CD4 с TCR, специфичным в отношении мышиного овальбумина (Ova) (ОТ-II), являются удобной моделью для тестирования иммунодепрессии, поскольку приблизительно половина Т-клеток CD4 является специфичной в отношении овальбумина, и ответы Т-клеток, следовательно, являются сильными и предсказуемыми. Спленоциты (1×105) от 10-недельных самок мышей ОТ-II инкубировали в течение 3 суток в круглодонных 96-луночных планшетах в 200 мл среды RPMI-1640, содержащей 5% FCS и указанные на Фиг. 20 концентрации белка или пептида Ova или антитело против CD3 (клон 2С-11) и рекомбинантный человеческий белок CD52-FC или Fc. Поглощение 3Н-тимидина измеряли на протяжении последних 16 ч культуры. Результаты представляют собой среднее плюс/минус sem трех повторностей.
Результаты
Как показано на Фиг. 20, CD52-FC значимо дозозависимо уменьшал пролиферацию Т-клеток в ответ на стимуляцию белком или пептидом Ova, предоставляя дополнительное доказательство терапевтического потенциала растворимого CD52 в лечении аутоиммунных заболеваний.
Пример 20: растворимый CD52. происходящий из семенной жидкости, подавляет пролиферацию человеческих Т-клеток
Способы
CD52 идентифицировали в образцах человеческой спермы с использованием следующего протокола ELISA. Сперва семенную жидкость (SF) центрифугировали при 500 g в течение 5 мин для осаждения сперматозодов, что подтверждалось микроскопической проверкой супернатанта. Антитело против человеческого CD52 (Biolegend #338202) использовали в качестве захватывающего агента (1:100 в PBS; 50 мкл/лунку в течение ночи; 4°С). Лунки промывали 3 раза в PBS-0,01% Tween, с последующими 3 промывками в PBS. Для блокирования лунок использовали раствор 5% BSA/PBS (BSA Sigma А7906) (200 мкл/лунку, 1 ч при комнатной температуре (RT)). Промывку проводили так же, как описано выше. В качестве контролей включали пустые лунки. Образцы спермы разводили в 5% BSA/PBS и добавляли по 50 мкл/лунку. Образцы инкубировали в лунках в течение 3 ч при RT. Промывку проводили так же, как описано выше. Для детекции использовали mAb-HRP Campath, разведенное 1:1000 в 5% BSA/PBS (100 мкл/лунку; 1 ч при RT). Промывку проводили так же, как описано выше. Добавляли 3,3ʹ,5,5ʹ-тетраметилбензидин (ТМВ), и считывание планшетов производили при 450 нм.
Иммунообеднение CD52 проводили согласно следующему протоколу. 200 мкл белок G-сефарозы аликвотировали в 2 пробирки Эппендорфа, с последующей промывкой х2 1 мл PBS, причем супернатант отбрасывали. 5 мг mAb Campath добавляли в одну пробирку Эппендорфа, и 5 мг «Octagam» (объединенный человеческий иммуноглобулин) - в другую. Пробирки вращали в течение 1,5 ч при 4°С, с последующими 3 промывками каждый раз 1 мл PBS. Супернатанты отбрасывали. Добавляли 500 мкл PBS, хорошо перемешивали, и образцы разделяли на равные доли в 5 пробирок Эппендорфа для каждого образца. Пробирки, содержащие Campath и Octagam, центрифугировали, и супернатанты отбрасывали. Образцы спермы добавляли в подходящие пробирки (5 разных образцов спермы), т.е. 160 мкл спермы плюс 160 мкл PBS, с последующим вращением в течение ночи при 4°С. Пробирки центрифугировали, и супернатанты собирали для применения в анализах Т-клеток в соотношении 1:20 (уже разведенные 1:2, следовательно, разводили 1:10 в анализе).
Пролиферацию Т-клеток в ответ на антиген (ТТ) в РВМС от здоровых доноров измеряли посредством разбавления красителя CFSE (Mannering et al., 2003). Клетки, меченные CFSE (2×105/лунку, 100 мкл), культивировали в 96-луночных круглодонных планшетах в повторности 6 с одной средой или с ТТ плюс/минус mAb против CD52 - CF1D12 (конечная концентрация 20 мкг/мл). Последнее добавляли либо в 0, либо в 20 ч, причем последнее время обеспечивало инициацию активации Т-клеток, принимая во внимание то, что рецептор растворимого CD52 - Siglec-10, как было показано, подвергается повышающей регуляции посредством активации (Фиг. 12Б). Также включали и использовали неокрашенные клетки для введения поправок на проточном цитометре. Индекс клеточных делений (CDI) рассчитывали как отношение числа поделившихся клеток CD4+ CFSEтусклый на 20000 неподелившихся клеток CD4+ CFSEяркий в присутствии антигена к числу поделившихся клеток CD4+ CFSEтусклый на 20000 неподелившихся CD4+ СFSЕяркий в отсутствие антигена.
Результаты
На Фиг. 21 проиллюстрировано присутствие растворимого CD52 в 26 образцах спермы при серийных разведениях. В общем, сперма содержит высокие уровни растворимого CD52, которые титруются в интервале нескольких log разведений.
Как показано на Фиг. 22, один антиген (ТТ) кардинально увеличивал пролиферацию Т-клеток (см. столбики «Нет спермы» на Фиг. 22). Однако данный эффект значительно снижался в присутствии спермы (см. «ТТ» для образцов спермы #1 и #15 на Фиг. 22). Один цикл иммунообеднения CD52 с использованием антитела против CD52 Campath частично обращал ингибирующий эффект спермы (см. столбики «Campath + ТТ» для образцов спермы #1 и #15 на Фиг. 22). Значимого обращения не наблюдалось с контрольными образцами с иммунообеднением IgG.
Как показано на Фиг. 23, один антиген (ТТ) кардинально увеличивал пролиферацию Т-клеток (см. столбики «Нет спермы» на Фиг. 23). Добавление антитела против CD52, CF1D12, дополнительно увеличивало пролиферацию Т-клеток. Однако в присутствии спермы пролиферация Т-клеток кардинально снижалась (см. «ТТ» для образцов спермы #14, #20 и #22 на Фиг. 23). Таким образом, сперма увеличивает нестимулированную и снижает стимулированную антигеном пролиферацию. Добавление антитела против CD52, CF1D12, частично обращало ингибирующий эффект спермы.
Таким образом, растворимый CD52, происходящий из спермы, достигал такого же подавляющего эффекта на функцию эффекторных Т-клеток (проиллюстрированную в данном Примере пролиферацией Т-клеток), что и растворимый CD52, происходящий из лимфоцитов, демонстрируя то, что на раскрытом здесь растворимом гликопротеине могут присутствовать альтернативные углеводные группировки без уменьшения его ингибирующей функции.
Пример 21: эффекты CD52-FC на моноциты
Способы и результаты
Клетки ТНР-1 (линия клеток острого моноцитарного лейкоза человека) выращивали в среде RPMI-1640, дополненной 10% FCS, 2 мМ глутамином и 50 мкМ 2-меркаптоэтанолом. Клетки высевали в количестве 2×105/лунку в IMDM, содержащей 5% объединенной инактивированной нагреванием человеческой сыворотки, 100 мМ заменимые аминокислоты, 2 мМ глутамин и 50 мкМ 2-меркаптоэтанол (среда IP5), при 37°С под 5% СО2.
Клетки инкубировали с разными дозами CD52-FC или контроля в виде Fc в присутствии LPS (100 нг/мл) в течение 24 ч. Среду собирали, и концентрацию IL-1β измеряли посредством ELISA. Результаты данного эксперимента обобщены на Фиг. 24.
В другом эксперименте клетки инкубировали с разными дозами CD52-Fc или контроля в виде Fc в присутствии агониста TLR-2 Pam3CSK (100 нг/мл) в течение 24 ч. Затем отбирали среды, и концентрацию IL-1β измеряли посредством ELISA. Результаты данного эксперимента обобщены на Фиг. 25.
Клетки ТНР-1 также дифференцировались в течение 3 ч с использованием 500 нМ форбол-12-миристат-13-ацетата (РМА). Клетки затем промывали и высевали в количестве 2×105/лунку в среде IP-5 и инкубировали в течение ночи при 37°С под 5% CO2. На следующее утро среду заменяли, и клетки инкубировали с CD52-FC или контролем в виде Fc (50 мкг/мл) в присутствии квасцов (100 мкг/мл) в течение 16 часов. Среду отбирали, и концентрацию IL-1β измеряли посредством ELISA. Результаты данного эксперимента обобщены на Фиг. 26.
Костный мозг от 10-недельных мышей С57/В6 дифференцировался в течение 7 суток в гранулоцитарно-макрофагальном колониестимулирующем факторе (10 нг/мл) в среде KDS-RPMI-10% FCS. Дендритные клетки, происходящие из костного мозга (BMDC), собирали, промывали и высевали в количестве 2×104/лунку в 96-луночном планшете. Клетки инкубировали с 40 мкг/мл мышиного CD52-FC или PBS (контроль) в присутствии LPS (800 нг/мл), CPG (0,8 пМ) или Listeria monocytogenes (8×106/лунку). Кроме того, клетки примировали в течение 3 ч LPS (100 нг/мл) и затем стимулировали известными агонистами воспаления, уратом мононатрия (MSU) (150 мкг/мл), квасцами (150 мкг/мл) и нигерицином (1 мкМ). Через 24 ч среды отбирали, и концентрации цитокинов измеряли мультиплексным анализом с цитокиновым чипом. Результаты данного эксперимента с использованием IL-1β обобщены на Фиг. 27. Аналогичные результаты были получены для IL-1α, TNF-α, МСР-1, IL-6, IL-9 и IL-12 (данные не показаны).
Мышиный CD52-Fc (250 мкг) инкубировали с нейраминидазой из Arthrobacter ureafaciens (2 единицы) или реакционным буфером (250 мМ фосфат натрия, рН 6,0) при 37°С в течение ночи, и реакцию завершали нагреванием при 75°С в течение 5 минут. Клетки ТНР-1 инкубировали с mCD52-Fc (конечная концентрация 12,5 мкг/мл), обработанным нейраминидазой или реакционным буфером, в присутствии LPS (100 нг/мл) в течение 24 ч. Среды отбирали, и концентрацию IL-1β измеряли посредством ELISA. Результаты данного эксперимента обобщены на Фиг. 28.
Мышиный CD52-Fc (300 мкг) обрабатывали или не обрабатывали ПНГазой F при тех же самых условиях согласно инструкциям изготовителя (BioLabs Inc.). Удаление N-связанного олигосахарида со снижением молекулярной массы CD52-FC подтверждали SDS-PAGE и окрашиванием кумасси синим. Белковые растворы затем обессоливали диализом против чистой стерильной воды. Клетки ТНР-1 высевали в количестве 2×105/лунку в среде IP5 и инкубировали с обработанным CD52-FC (конечная концентрация 30 мкг/мл) или гликозилированным CD52-FC в присутствии 100 нг/мл LPS в течение 24 ч. Среды отбирали, и концентрацию IL-1β измеряли посредством ELISA. Результаты данного эксперимента обобщены на Фиг. 29.
Обсуждение
В ответ на ряд воспалительных стимулов CD52-Fc дозозависимо подавлял секрецию IL-1β линией человеческих моноцитов ТНР1 и дендритными клетками, происходящими из костного мозга мыши. Кроме того, как показано для Т-клеток, данный подавляющий эффект CD52-FC зависит от его олигосахаридной группировки, так как он отменялся предварительной обработкой CD52-Fc нейраминидазой с удалением концевых сиаловых кислот или РНГазой F с удалением самого N-связанного олигосахарида. Данные открытия демонстрируют то, что подавляющие эффекты CD52-Fc, показанные для Т-клеток, распространяются на другие типы клеток, которые участвуют во врожденном иммнитете и, опять-таки аналогично Т-клеткам, предпочтительно опосредованы рецептором Siglec.
Специалистам в данной области будет понятно, что в отношении изобретения могут быть сделаны многочисленные вариации и/или модификации, как показано в конкретных воплощениях, без отступления от сущности или объема изобретения, как описано в широком смысле. Настоящие воплощения, следовательно, во всех отношениях следует рассматривать как иллюстративные и не ограничивающие.
В настоящей заявке испрашивается приоритет от US 61/560254, поданной 15 ноября 2011 года, и US 61/705633, поданной 26 сентября 2012 года, все содержание обеих из которых включено сюда посредством ссылки.
Все публикации, которые здесь обсуждаются и/или на которые здесь дается ссылка, включены сюда во всей их полноте.
Любое обсуждение документов, актов, материалов, устройств, предметов или тому подобного, которое было включено в настоящее описание, осуществляется единственно с целью предоставления контекста настоящего изобретения. Его не следует принимать как допущение того, что любые или все из данных вопросов образуют часть основы предшествующего уровня техники или представляли собой обычное общее знание в области, релевантной для настоящего изобретения, в том виде, в котором оно существовало до даты приоритета каждого пункта формулы данной заявки.
ССЫЛКИ
Allan et al. (2007). Int. Immunol. 19: 345-354.
Altschul et al. (1993) J. Mol. Biol. 215: 403410
Armour (2003) Mol. Immunol. 40: 585-93.
Bach et al (1997) J Autoimmun 10:375-386.
Barnden et al. (1998) Immunol. Cells Biol. 76: 34.
Belov et al. (2003) Proteomics 3: 2147-2154.
Bergerot et al. (1994) J. Autoimmun. 7: 655-663.
Collison et al. (2007) Nature 450: 566-569.
Crocker et al. (2007) Nat. Rev. Immunol. 7: 255-266.
Dromey et al. (2011) J. Autoimmunity 36: 47-55.
Every et al. (2006) J. Immunol. 176: 4608-4615.
Fontenot etal. (2003) Nat. Immunol. 4: 330-336.
Gavin et al. (2006) Proc. Natl. Acad. Sci. U.S.A. 103: 6659-6664.
Hale (2001) J. Biol. Regul. Homeost. Agents 15: 386-91.
Harayama (1998) Trends Biotechnol. 16: 76-82.
Hearnden et al. (2012) Ad. Drug Deliver. Rev. 64:16-28.
Herald etal. (2008) Proc. Natl. Acad. Sci. U.S.A. 105: 18507-18512.
Higgins and Sharp (1989) CABIOS. 5: 151-153.
Hogquist et al. (1993) J. Exp.Med. 177: 1469.
Hori etal. (2003) Science 299: 1057-1061.
Hu et al. (2009) Exp.Hematol. 237: 423-434.
Lee (2001) J. Natl. Inst. Monogr. (2001) 29: 41-44.
Lemmark (2001) J Clin Invest 108:1091-1096.
Liu et al. (2006) J Exp Med 203: 1701-1711.
Mannering et al. (2003) J. Immunol. Meth. 283:173-83.
Mittag et al. (2011) J. Immunol. 186: 6207-17.
Miyara et al. (2009) Immunity 30: 899-911.
Munday et al. (2001) Biochem. J. 355: 489-497.
Ngyuen et al. (2006) Proc. Natl. Acad. Sci. 103: 7765-7770.
Roncarolo and Gregori (2008) Eur. J. Immunol. 38: 925-927.
Sakaguchi et al. (2008) Cell 133: 775-787.
Sakaguchi et al. (2009) Int. Immunol. 21:1105-1111.
Schmidt and Skerra (2007) Nat. Protoc. 2: 1528-35.
Schroter et al (1999) J. Biol. Chem. 274: 29862-29873.
Seddiki et al. (2006) J. Exp.Med. 203: 1693-1700.
Shevach (2006) Immunity 25: 195-201.
Shevach (2009) Immunity 30: 636-645.
Song et al. (2004) Crit. Rev. Ther. Drug Carrier Syst. 21:195-256.
Tang et al. (2004) J. Exp.Med. 199: 1455-1465.
Tisch etal. (1999) J. Immunol. 163: 1178-1187.
Tone et al. (1999) Biochim. Biophys. Acta. 1446: 334-340.
Treumann et al. (1995) J. Biol. Chem. 270: 6088-6099.
Vignali et al. (2008) Nat. Rev. Immunol. 8: 523-532.
von Herrath and Harrison (2003) Nat. Rev. Immunol. 3: 223-232.
Xia et al. (1991) Eur. J. Immunol. 21: 1677-1684.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| БИОЛОГИЧЕСКИ АКТИВИРУЕМЫЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА НА ОСНОВЕ ЦИТОКИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2819307C2 |
| МНОГОЦЕПОЧЕЧНЫЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2013 |
|
RU2663725C2 |
| ПОЛИПЕПТИДЫ И ПОЛИНУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ИММУНОЛОГИЧЕСКИХ НАРУШЕНИЙ И РАКА | 2012 |
|
RU2623161C2 |
| ПОЛИПЕПТИДЫ ТРАНСПОЗАЗЫ И ИХ ПРИМЕНЕНИЕ | 2016 |
|
RU2735700C2 |
| СПОСОБЫ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ФИБРОЗА АНТАГОНИСТАМИ IL-21/IL-21R | 2006 |
|
RU2419450C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ СВЯЗАННЫХ С BCMA РАКОВЫХ ЗАБОЛЕВАНИЙ И АУТОИММУННЫХ РАССТРОЙСТВ | 2018 |
|
RU2799762C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2708311C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ АЛЛОГЕННЫХ T-КЛЕТОК С CAR | 2020 |
|
RU2828787C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2799573C2 |
















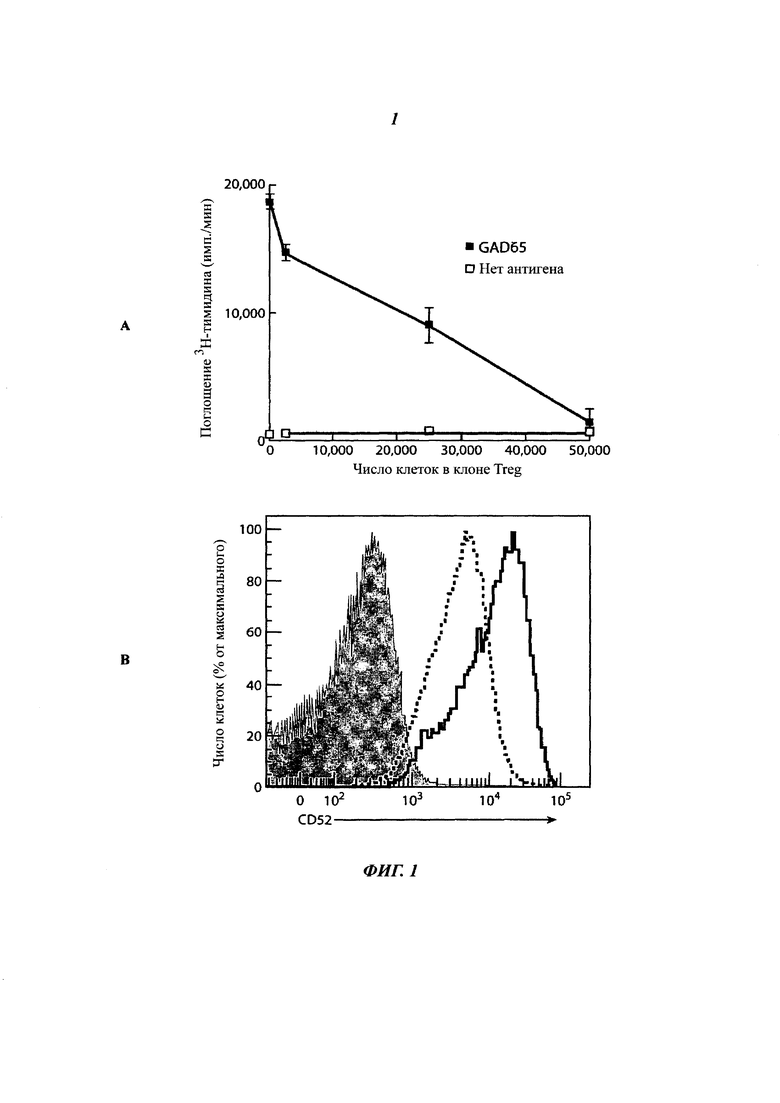
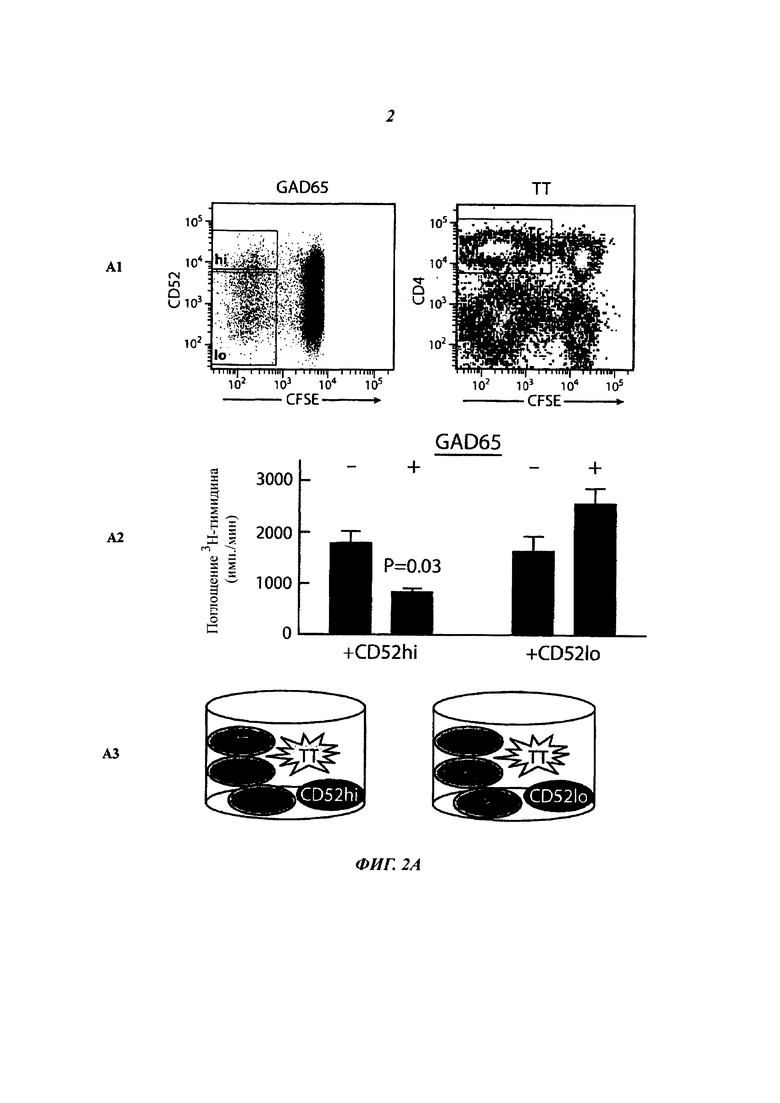
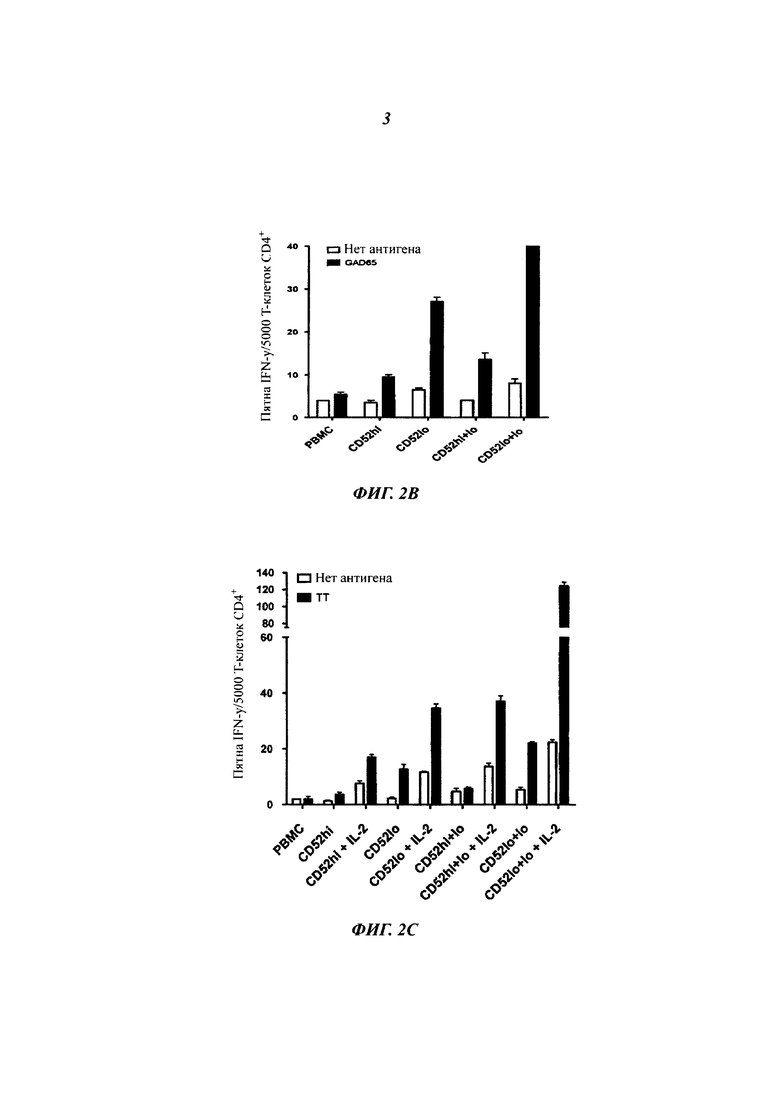
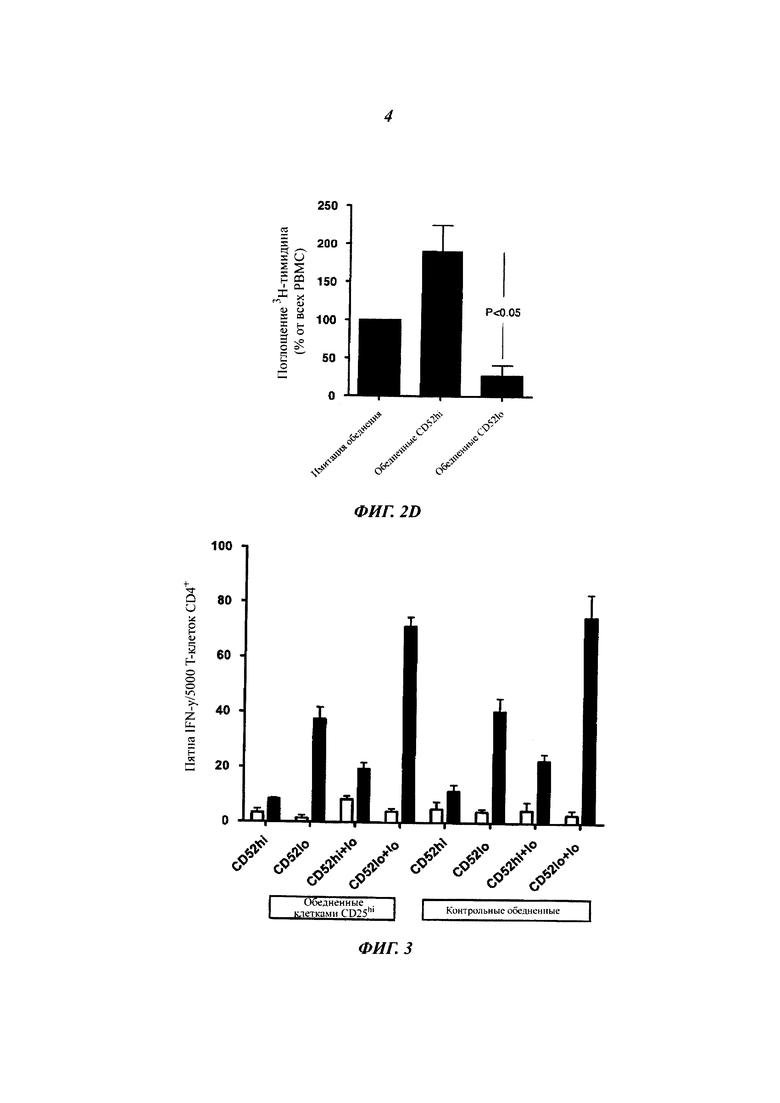
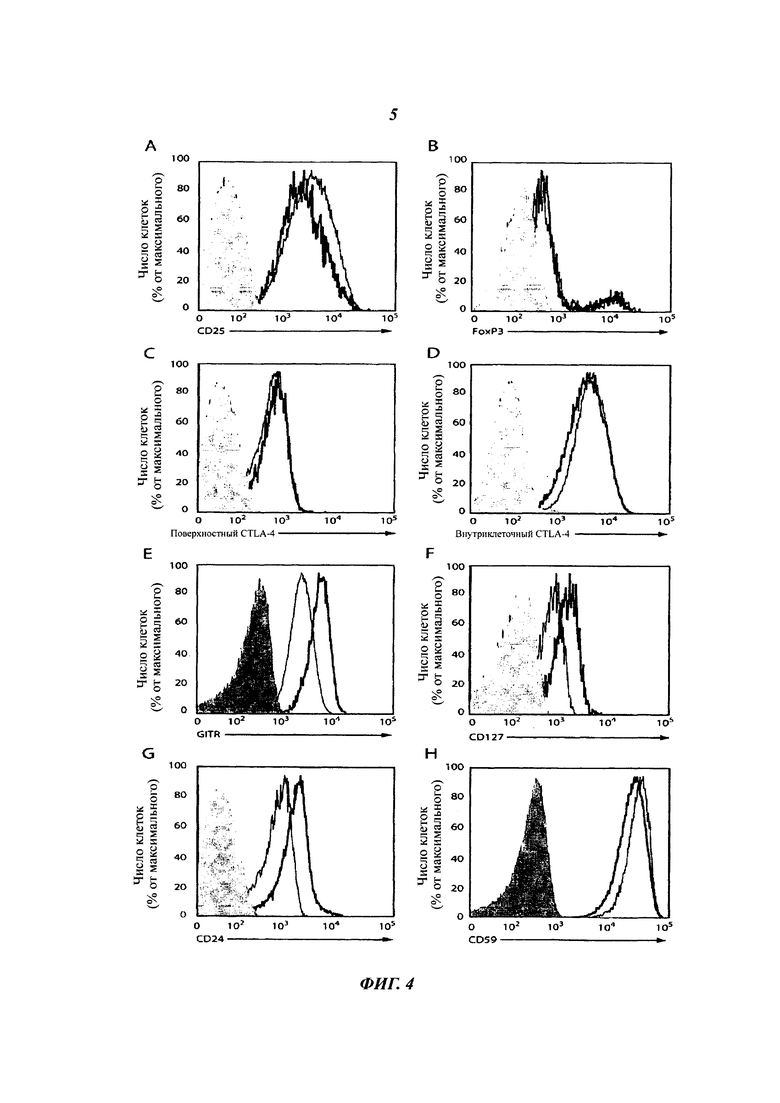
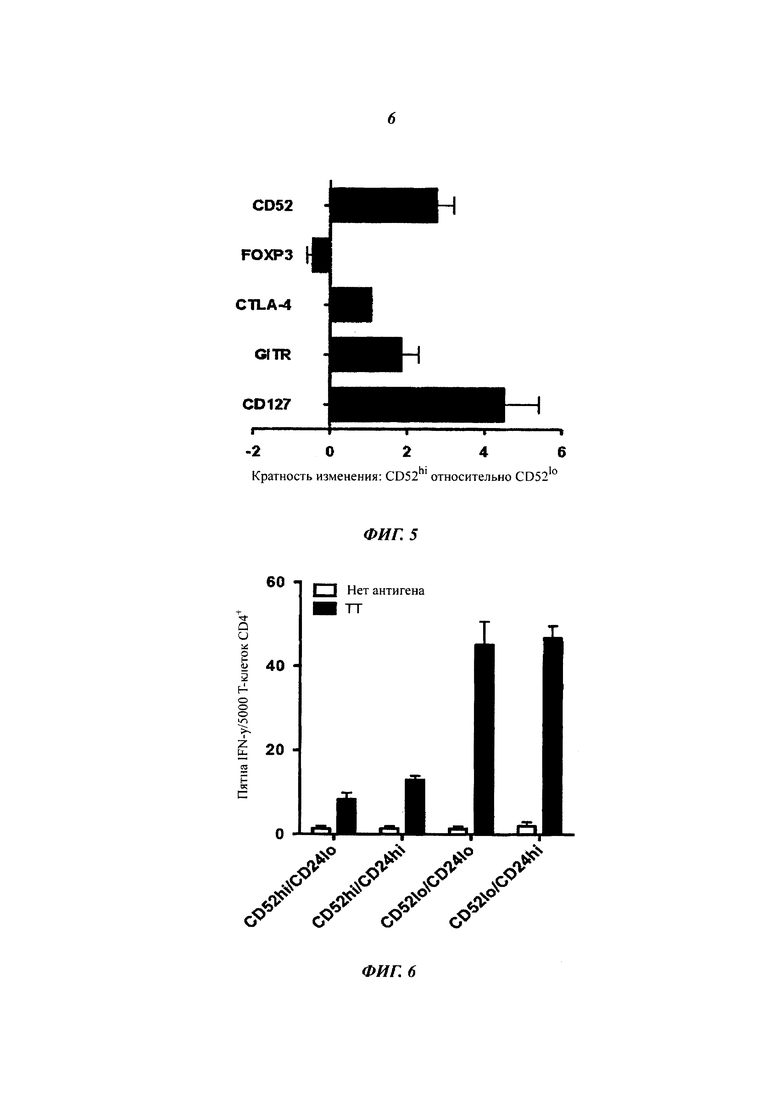
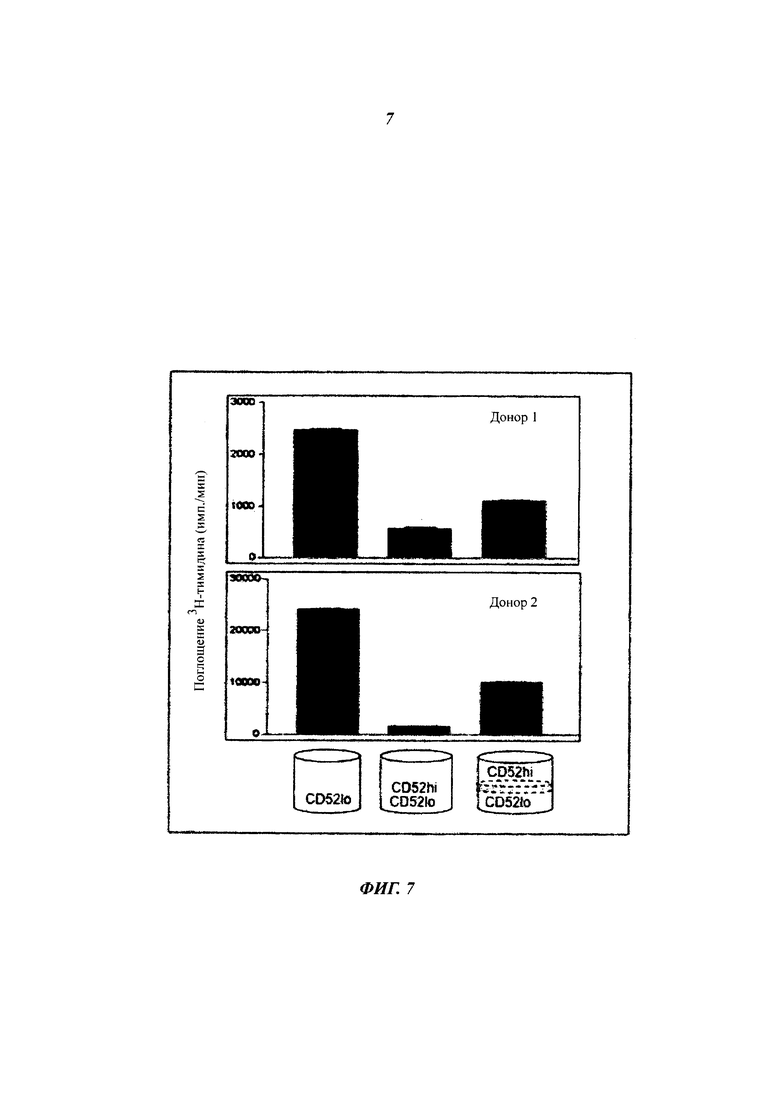
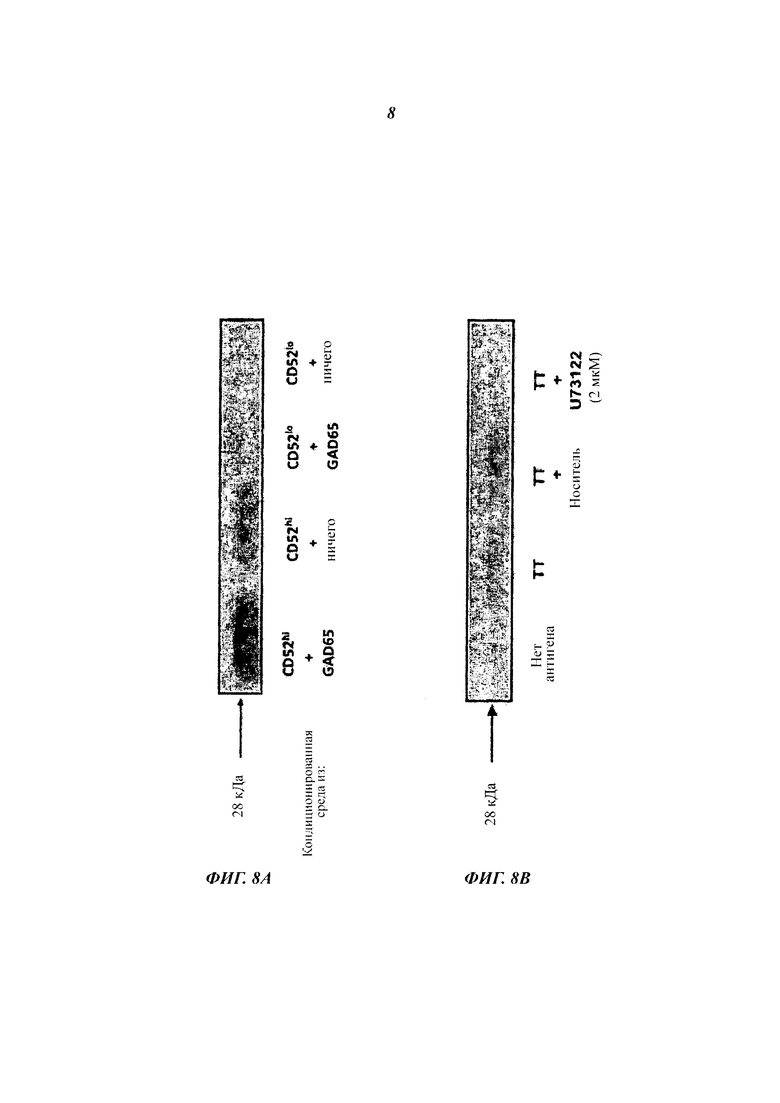
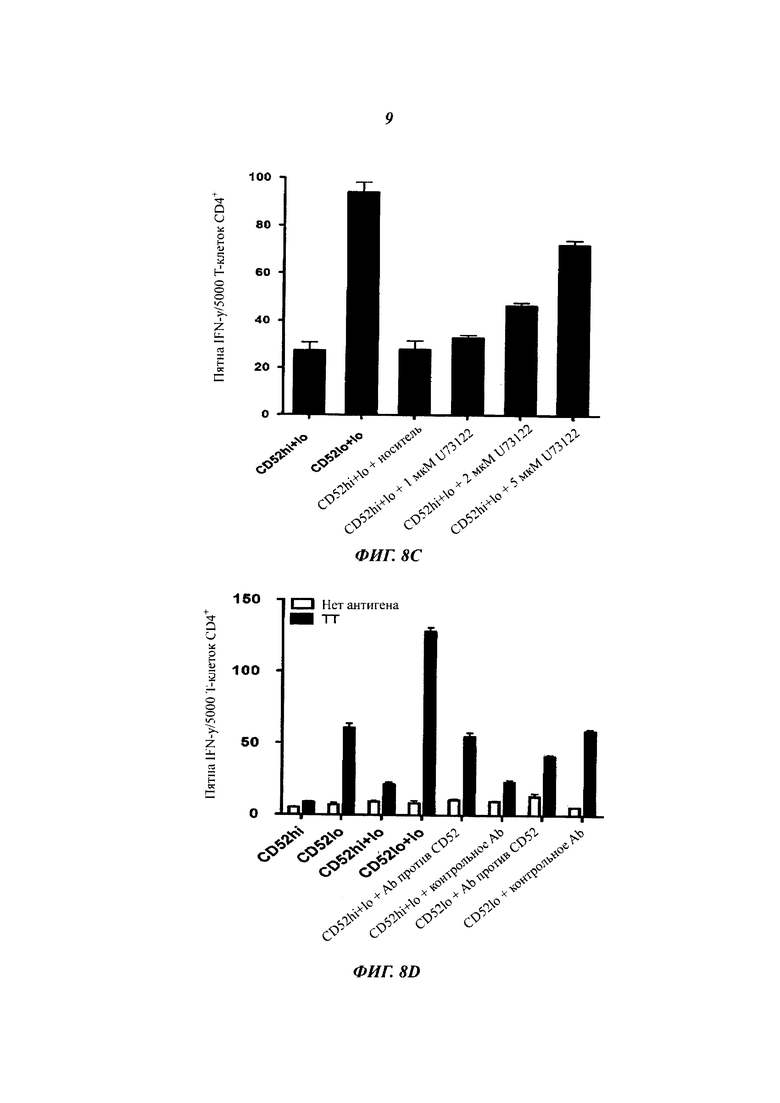
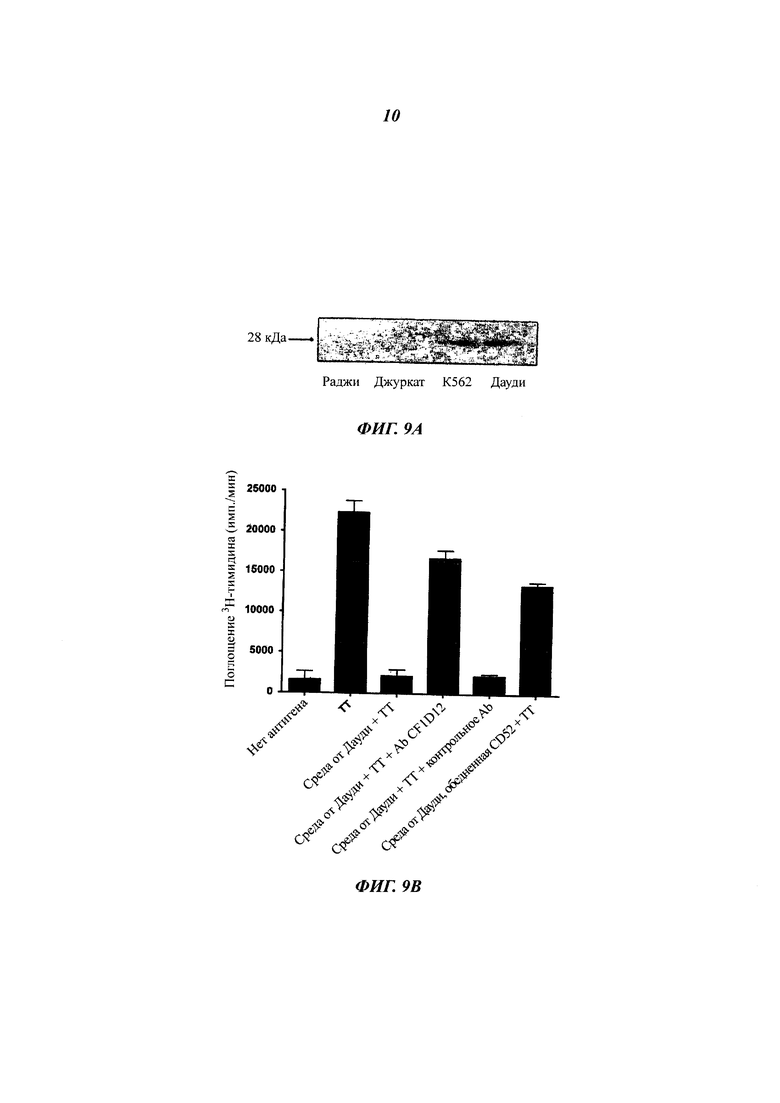
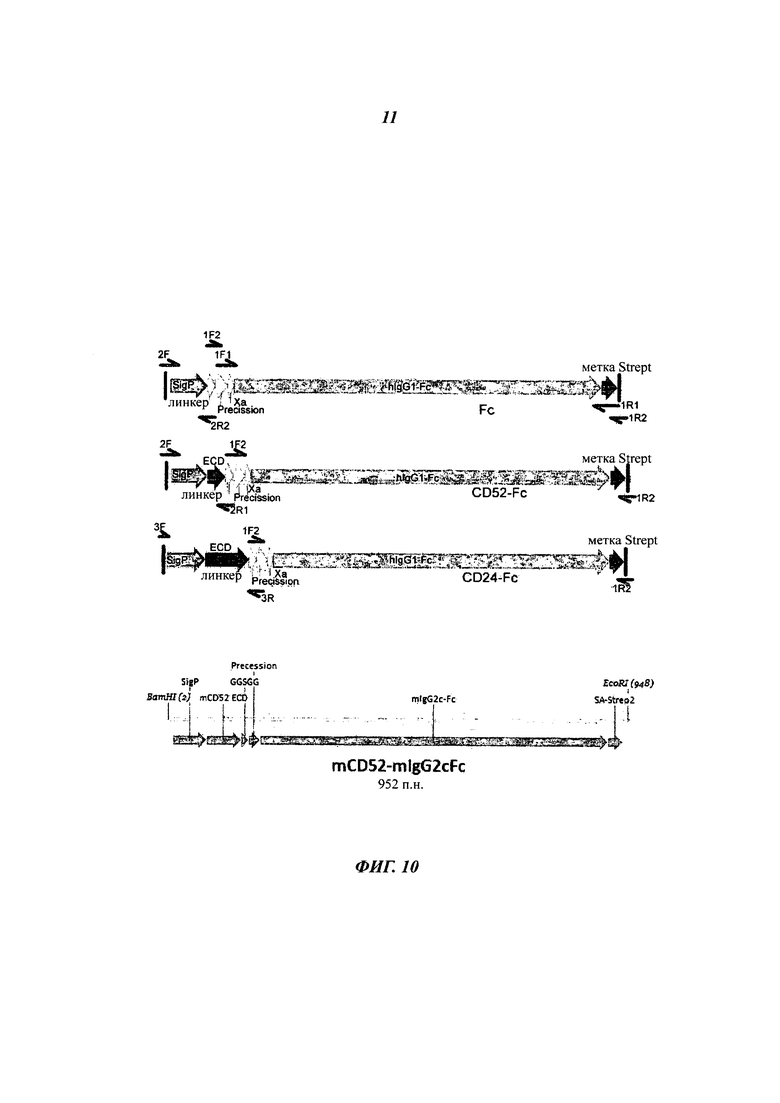
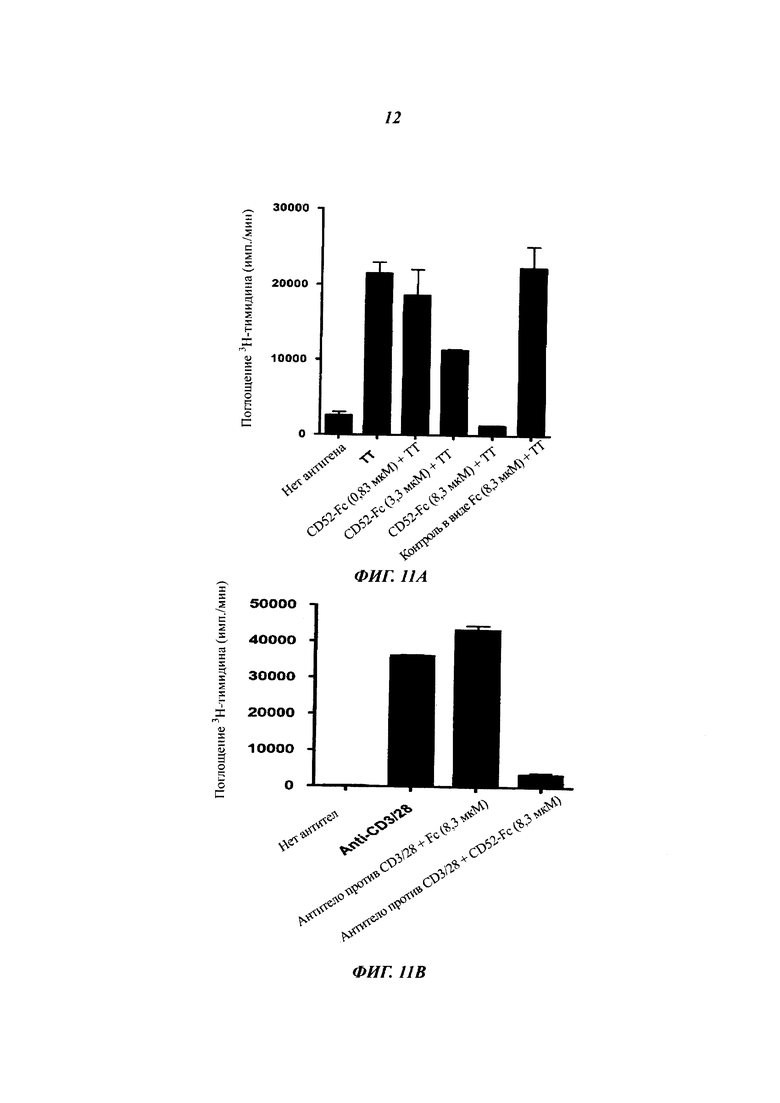
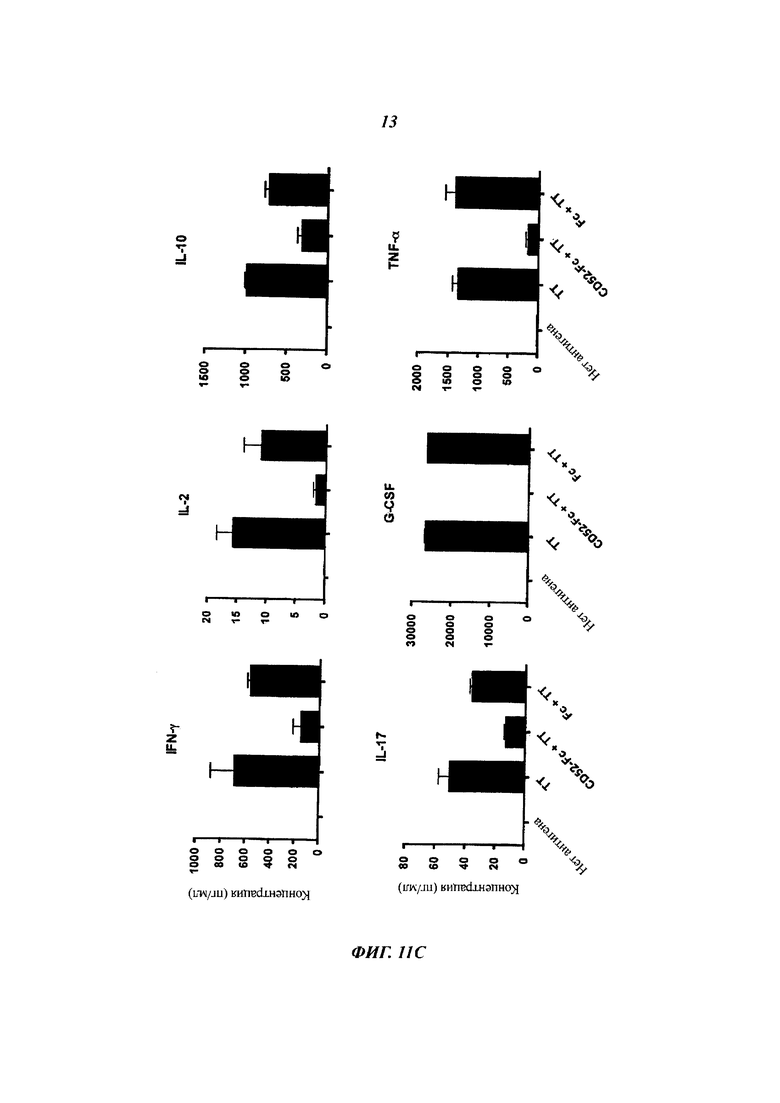
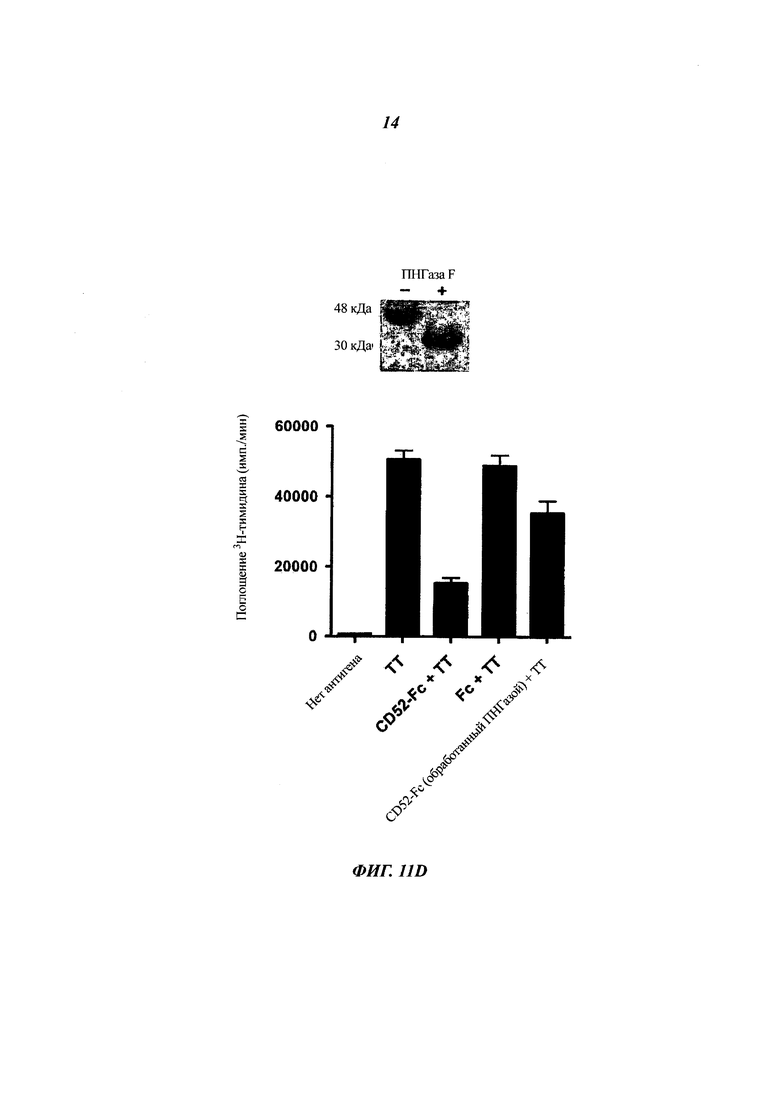
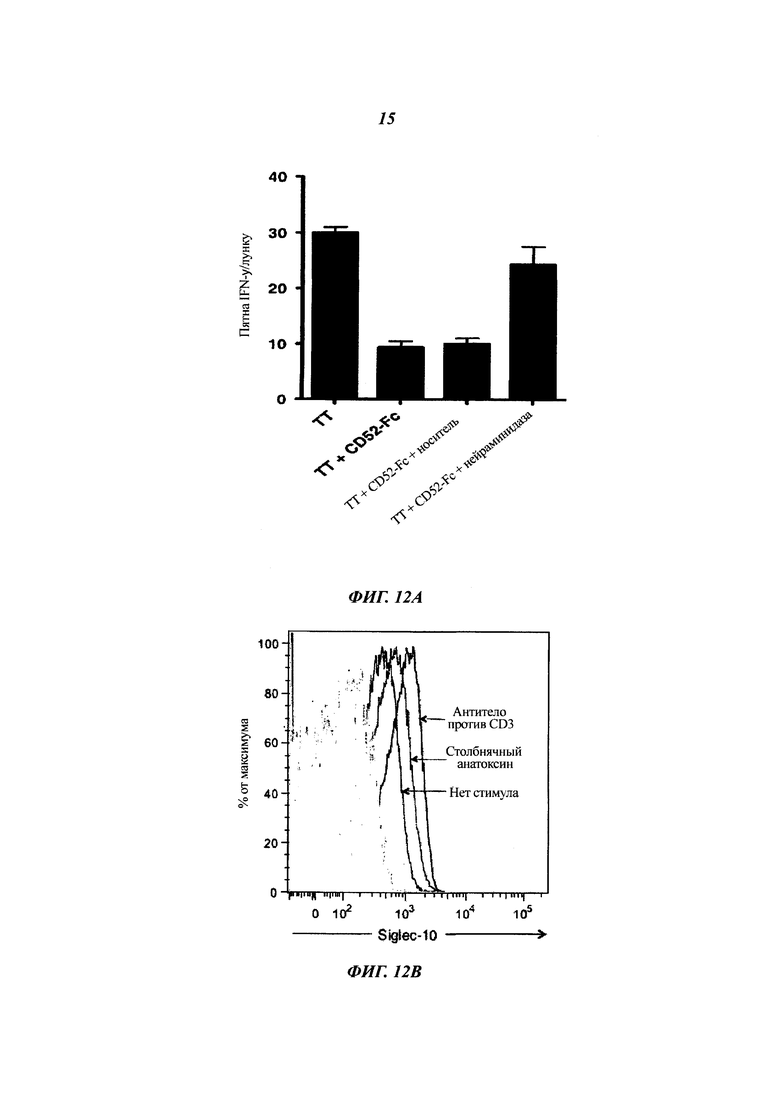
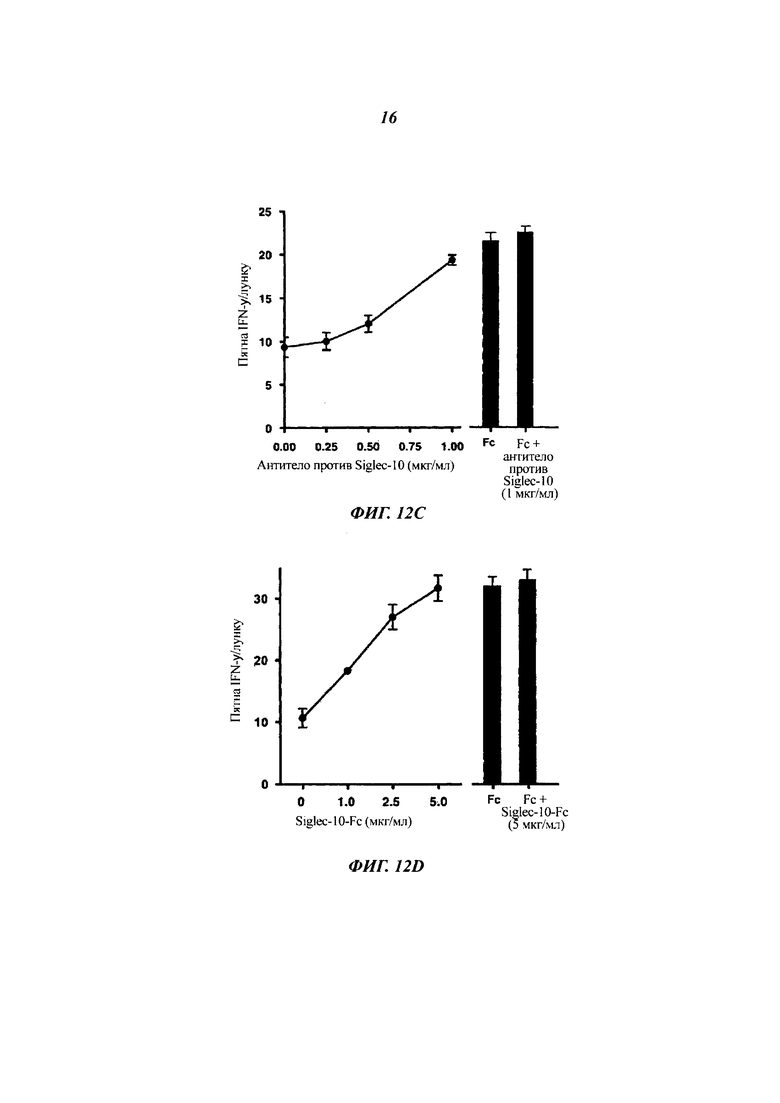
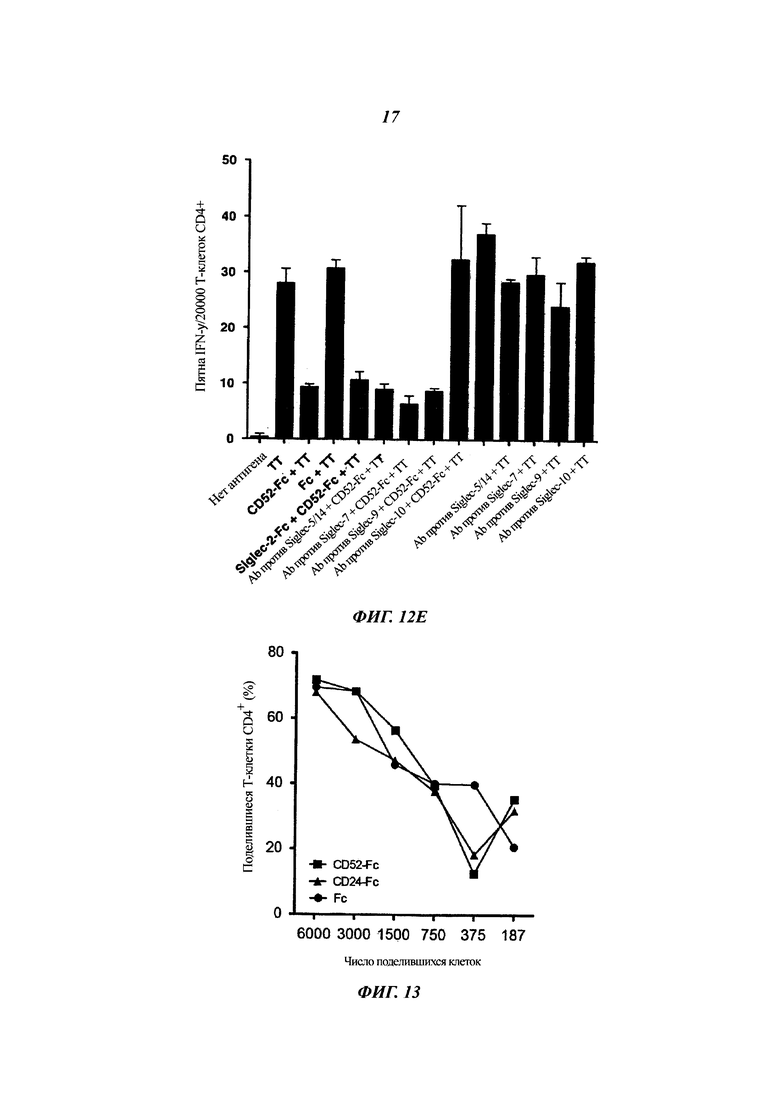
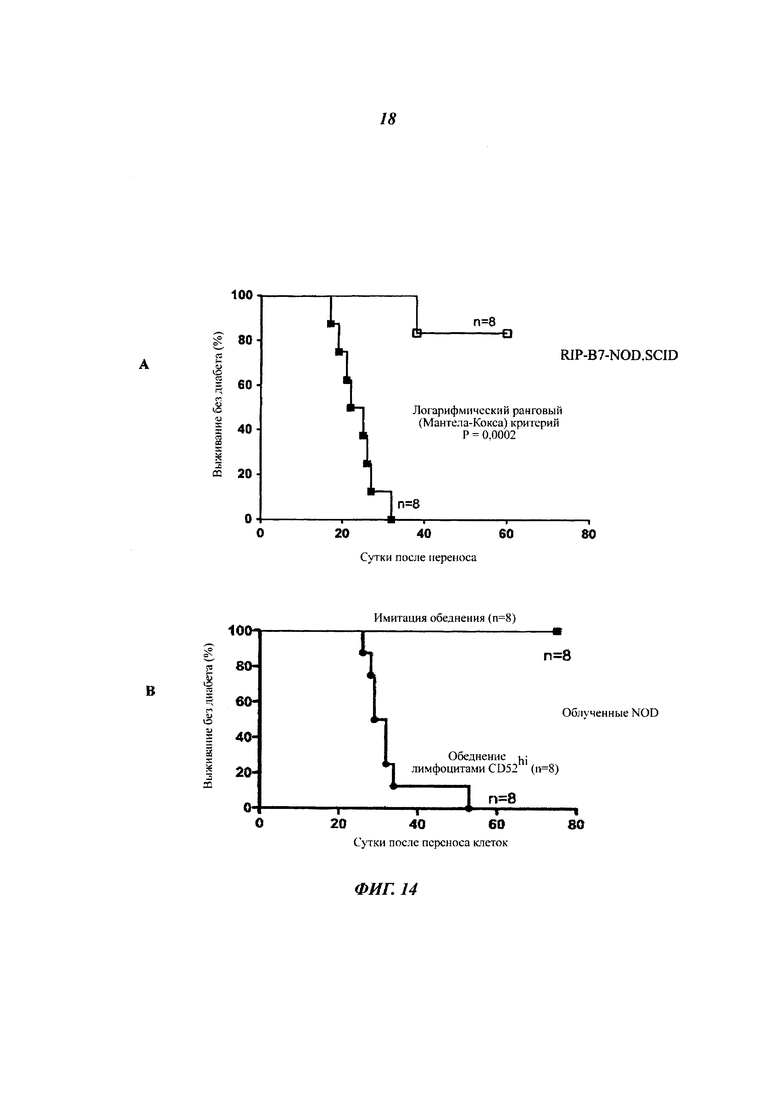
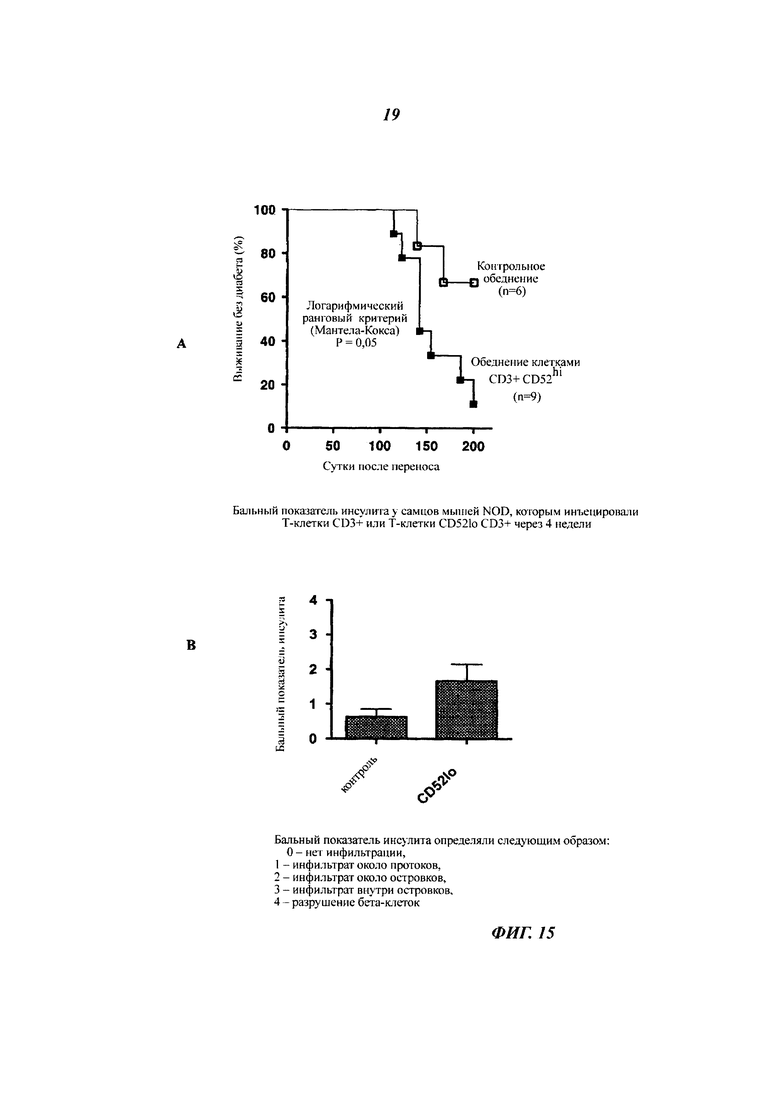
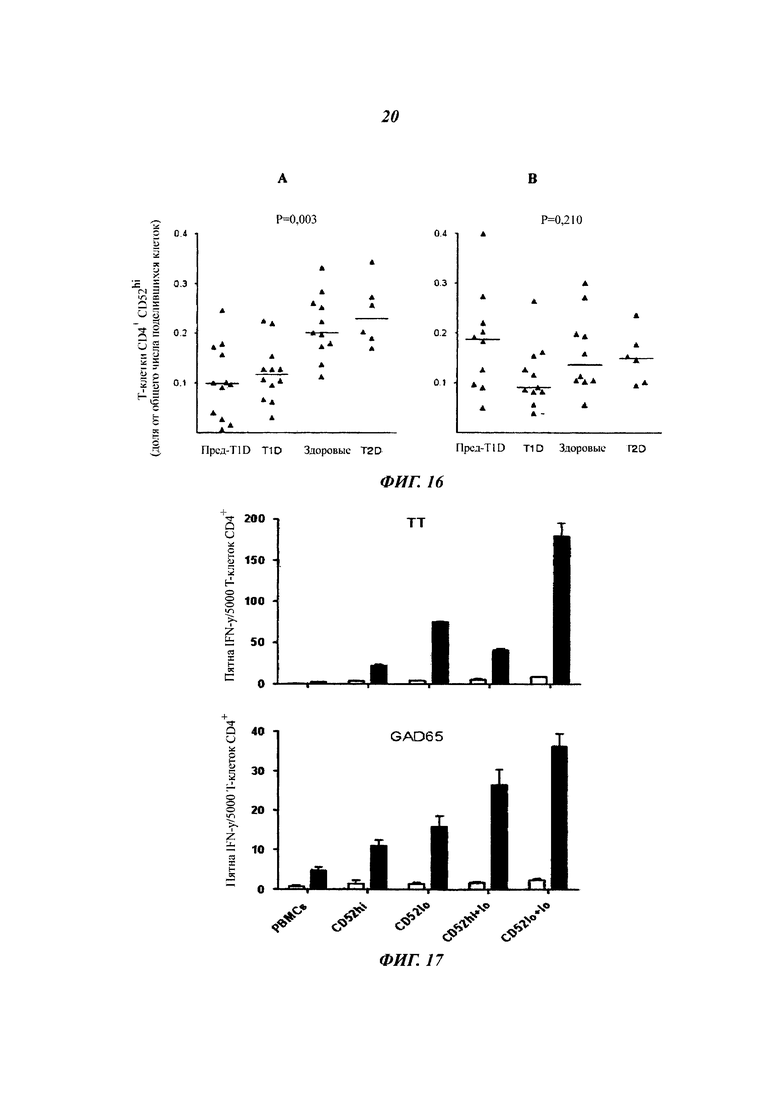
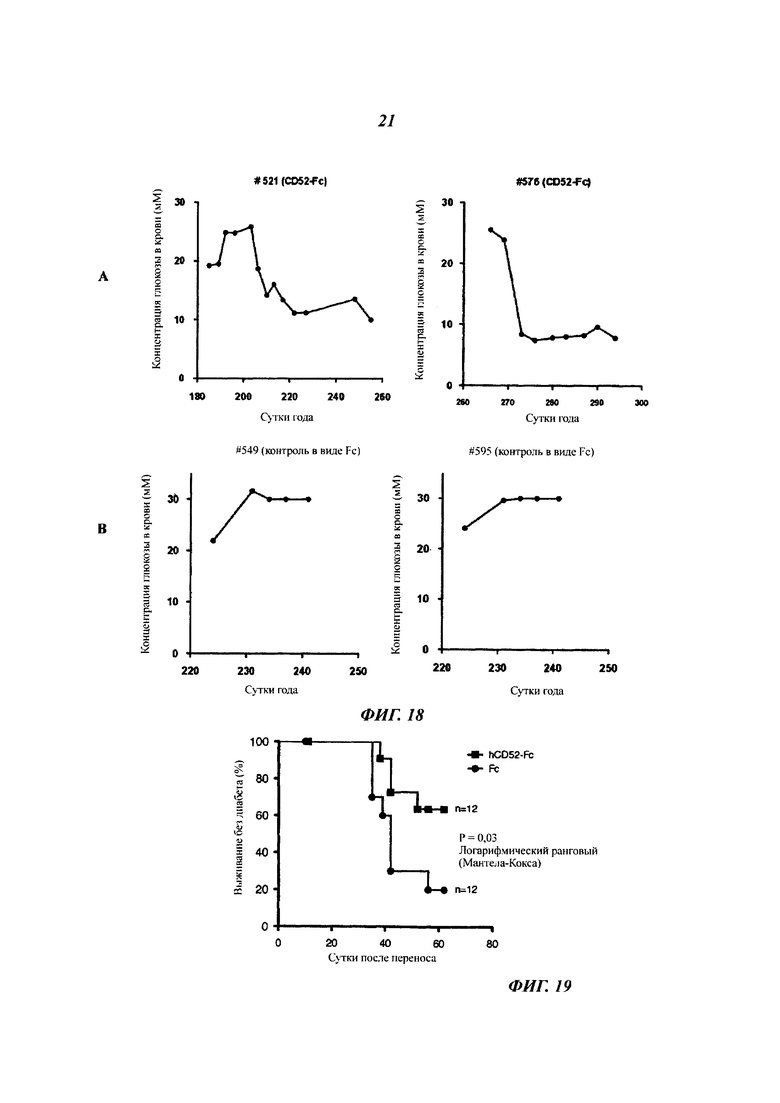
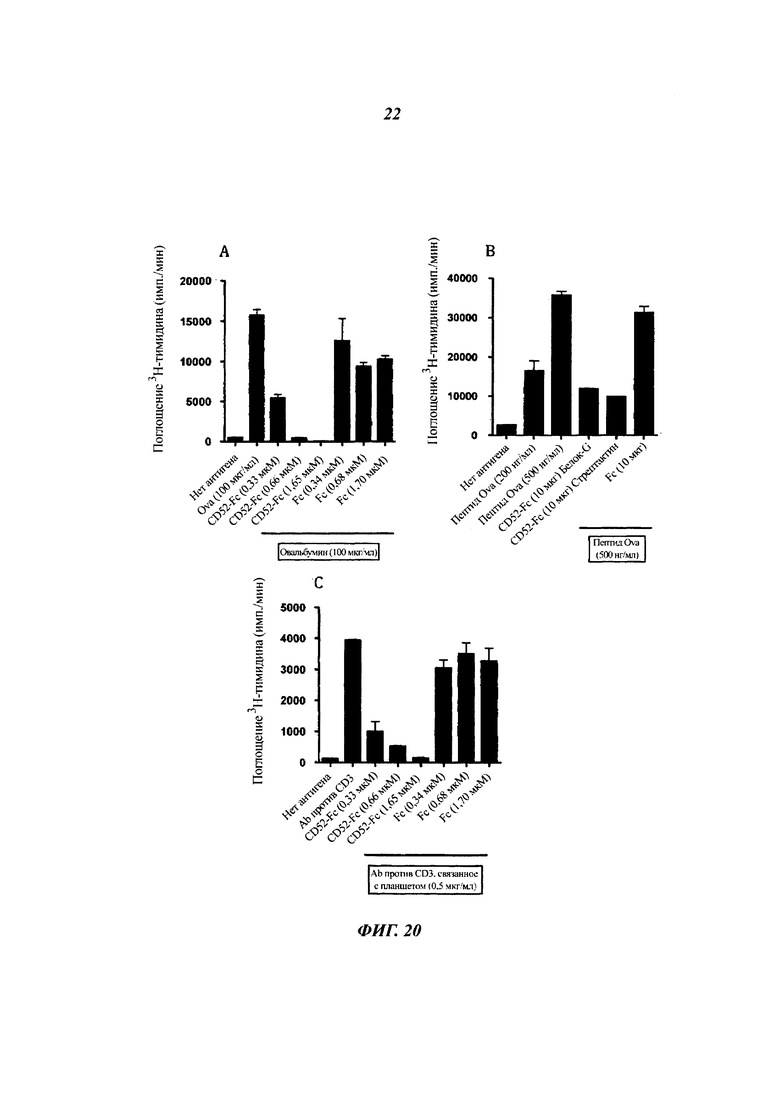
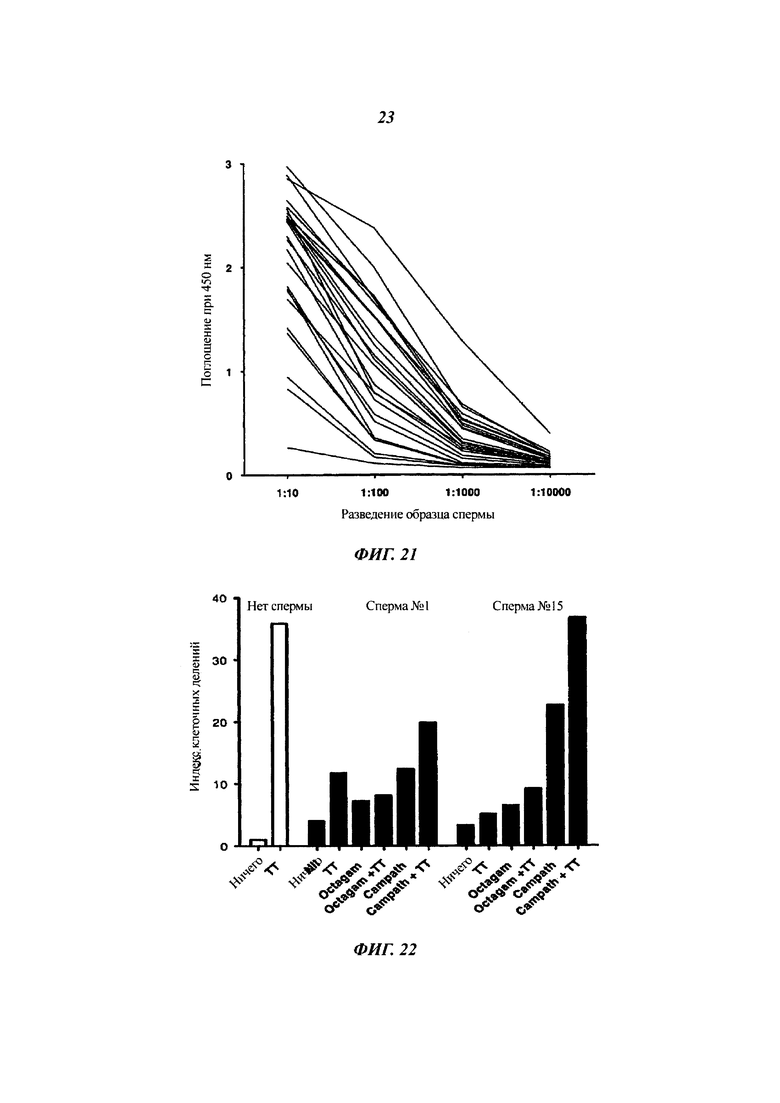
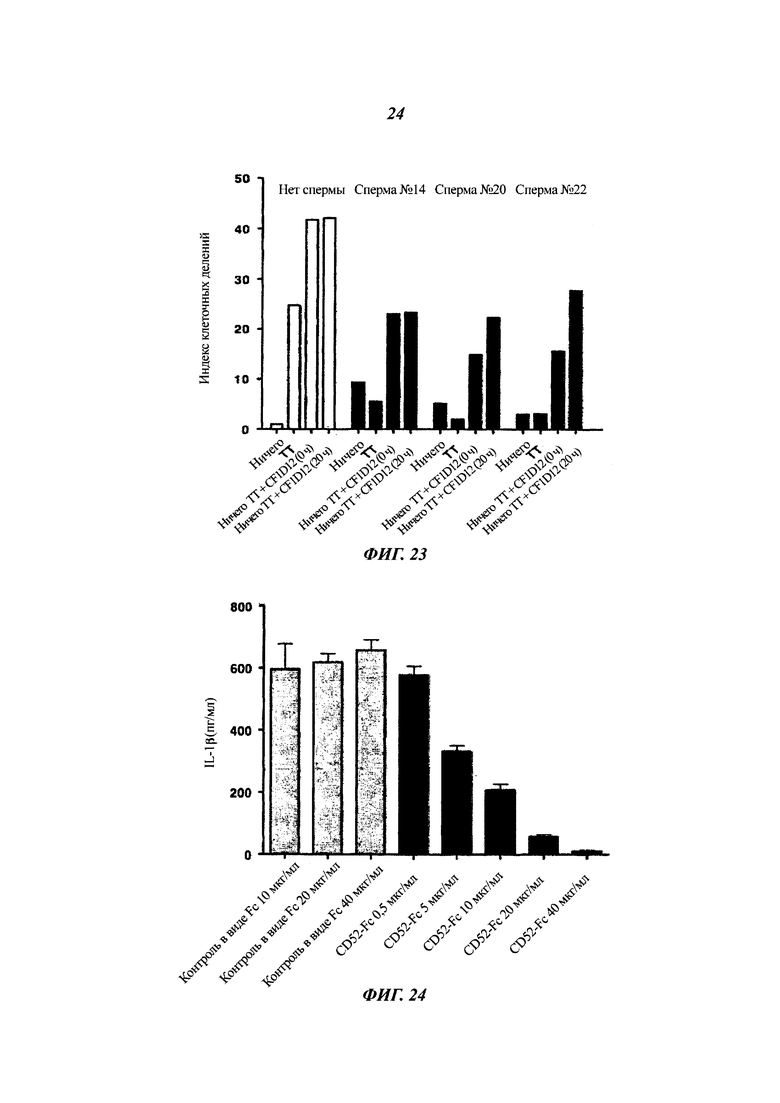
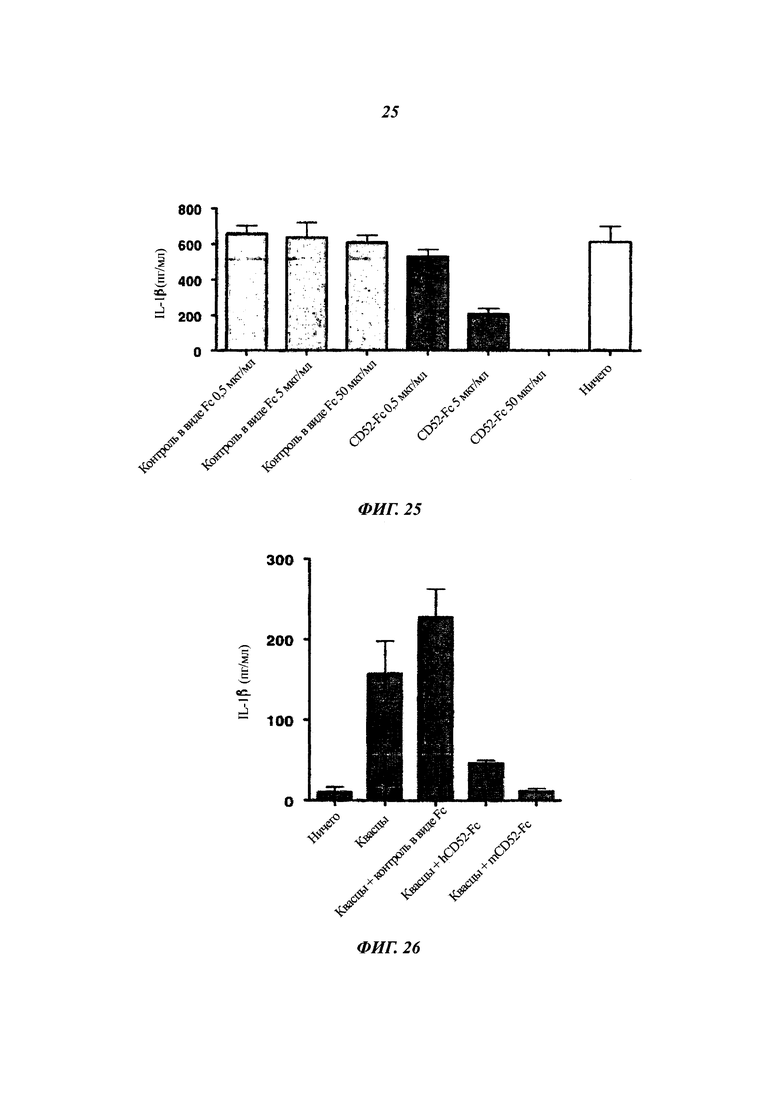
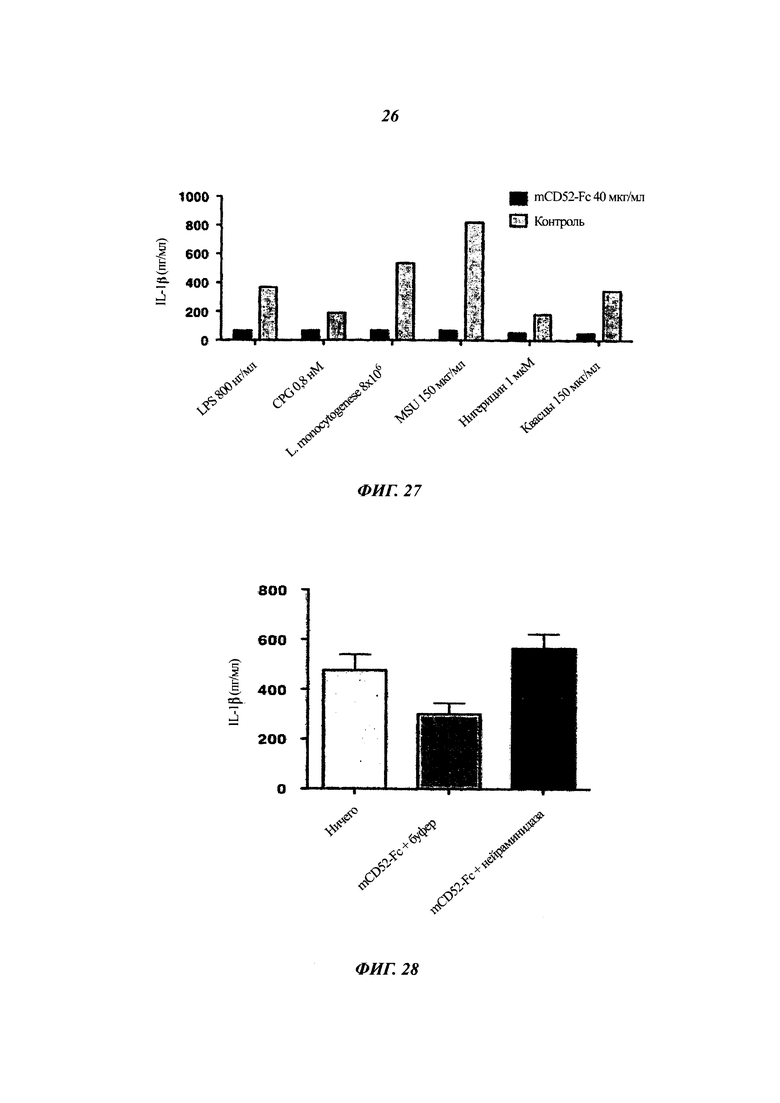
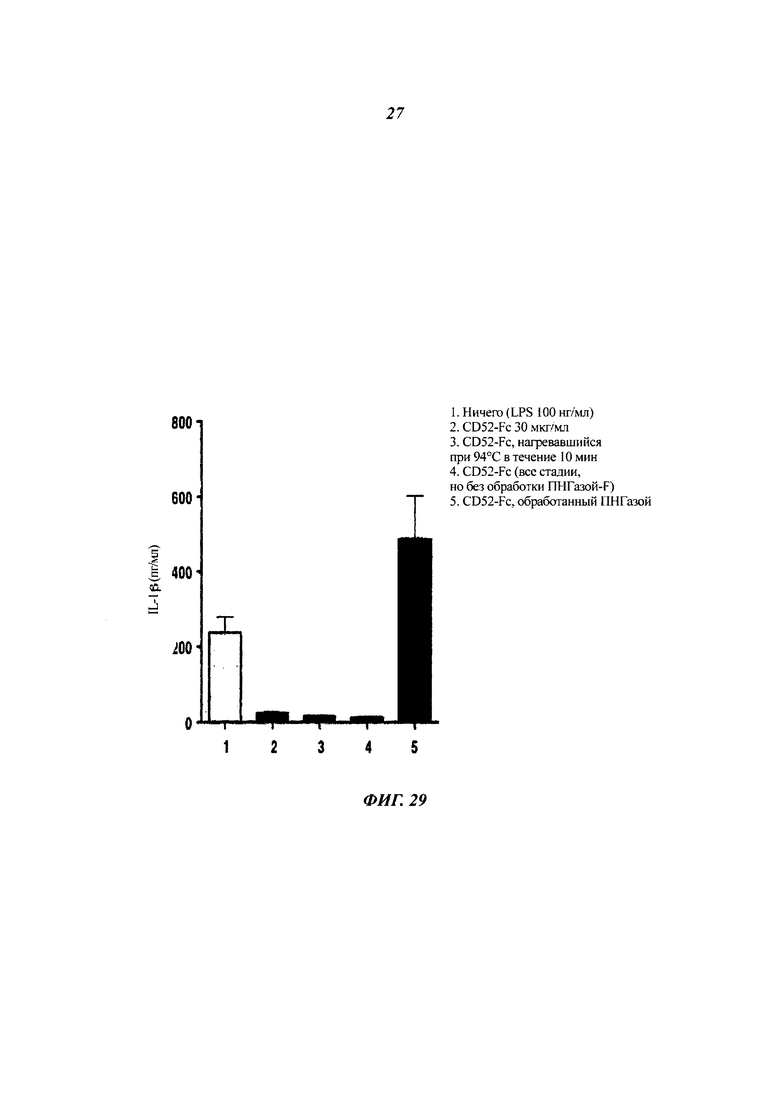
Группа изобретений относится к медицине и касается фармацевтической композиции для лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, содержащей любой один или более чем один из следующих: i) растворимый гликопротеин CD52, который содержит аминокислотную последовательность, по меньшей мере на 95% идентичную аминокислотной последовательности GQNDTSQTSSPS (SEQ ID NO: 3), и углевод, ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок; iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii); iv) вектор, содержащий полинуклеотид по разделу iii); v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv); и фармацевтически приемлемый носитель. Группа изобретений также касается способа лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса у субъекта, включающего введение субъекту терапевтически эффективного количества указанной композиции; способа проведения диагностики чувствительности субъекта к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса. Группа изобретений обеспечивает подавление функции эффекторных Т-клеток. 5 н. и 18 з.п. ф-лы, 21 пр., 29 ил., 1 табл.
1. Фармацевтическая композиция для лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, содержащая любой один или более чем один из следующих:
i) растворимый гликопротеин CD52, который содержит аминокислотную последовательность, по меньшей мере на 95% идентичную аминокислотной последовательности GQNDTSQTSSPS (SEQ ID NO: 3), и углевод,
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv);
и фармацевтически приемлемый носитель.
2. Фармацевтическая композиция по п. 1, в которой любой один или более чем один из растворимого гликопротеина CD52, слитого белка, полинуклеотида, вектора и клетки присутствует в достаточном количестве для подавления функции эффекторной Т-клетки и/или иммунного ответа.
3. Фармацевтическая композиция по п. 1, в которой углеводная часть растворимого гликопротеина CD52 связана с одним или более чем одним из остатков аспарагина, серина, треонина, тирозина, гидроксилизина, гидроксипролина, фосфосерина или триптофана, если они присутствуют в аминокислотной последовательности.
4. Фармацевтическая композиция по п. 1, в которой углеводная часть растворимого гликопротеина CD52 связана с остатком аспарагина (N) в SEQ ID NO: 3.
5. Фармацевтическая композиция по п. 1, в которой углеводная часть растворимого гликопротеина CD52 представляет собой любой углевод, для которого известно, что он присоединен к внеклеточной части растворимого гликопротеина CD52 в клетке-хозяине, такой как лимфоцит-хозяин или клетка половых путей-хозяин.
6. Фармацевтическая композиция по п. 1, в которой углеводная часть растворимого гликопротеина CD52 содержит один или более чем один би-, три- или тетраантенный сахар, который может быть сиалированным на конце.
7. Фармацевтическая композиция по п. 1, в которой любой один или более чем один из растворимого гликопротеина CD52, слитого белка, клетки, популяции клеток, среды культуры клеток и агента способен подавлять функцию эффекторной Т-клетки и/или способен уменьшать иммунный ответ.
8. Фармацевтическая композиция по п. 7, где иммунный ответ представляет собой иммунный ответ на аутоантиген.
9. Фармацевтическая композиция по п. 1, в которой слитый белок содержит второй белок, который содержит фрагмент антитела.
10. Фармацевтическая композиция по п. 9, в которой фрагмент антитела представляет собой Fc.
11. Фармацевтическая композиция по п. 1, где композиция содержит один или более чем один из растворимого гликопротеина CD52, слитого белка, среды культуры клеток или агента, и она приготовлена для введения через слизистую и/или чрескожного введения.
12. Фармацевтическая композиция по любому из пп. 1-11, которая, кроме того, содержит инсулин и/или аутоантиген.
13. Применение любого одного или более чем одного из следующих:
i) растворимый гликопротеин CD52, как определено в п. 1;
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv); и
vi) фармацевтическая композиция по любому из пп. 1-11,
для подавления функции эффекторных Т-клеток и/или для уменьшения иммунного ответа, такого как иммунный ответ на аутоантиген.
14. Способ лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса у субъекта, включающий введение субъекту терапевтически эффективного количества любого одного или более чем одного из следующих:
i) растворимый гликопротеин CD52, как определено в п. 1;
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv); и
vi) фармацевтическая композиция по любому из пп. 1-11.
15. Способ по п. 14, где заболевание, опосредованное функцией эффекторных Т-клеток, представляет собой аутоиммунное заболевание, такое как диабет типа I или ревматоидный артрит либо отторжение аллотрансплантата или реакцию «трансплантат против хозяина».
16. Способ по п. 14, где растворимый гликопротеин CD52, слитый белок, среду культуры клеток, агент или композицию вводят в сайт на слизистой или чрескожный сайт.
17. Применение любого одного или более чем одного из следующих:
i) растворимый гликопротеин CD52, как определено в п. 1;
ii) слитый белок, содержащий растворимый гликопротеин CD52 в качестве первого белка и второй белок;
iii) полинуклеотид, кодирующий пептидную часть растворимого гликопротеина CD52 по разделу i) или слитый белок по разделу ii);
iv) вектор, содержащий полинуклеотид по разделу iii);
v) выделенная клетка, содержащая полинуклеотид по разделу iii) или вектор по разделу iv); и
vi) фармацевтическая композиция по любому из пп. 1-11,
в изготовлении лекарственного средства для лечения или предупреждения заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса.
18. Применение по п. 17, где заболевание, опосредованное функцией эффекторных Т-клеток, представляет собой аутоиммунное заболевание, такое как диабет типа I или ревматоидный артрит.
19. Применение по п. 17, где состояние, опосредованное функцией эффекторных Т-клеток, представляет собой отторжение аллотрансплантата или реакцию «трансплантат против хозяина».
20. Применение по п. 17, при котором лекарственное средство готовят для введения в сайт на слизистой или чрескожный сайт.
21. Способ проведения диагностики чувствительности субъекта к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сепсиса, включающий:
детектирование уровня растворимого гликопротеина CD52 в образце, взятом у субъекта; и
сравнение уровня растворимого гликопротеина CD52, детектированного в образце, взятом от субъекта, с контрольным уровнем, определенным у одного или более чем одного здорового субъекта,
в котором меньший уровень растворимого гликопротеина CD52, детектированного в образце, взятом от субъекта, по сравнению с контрольным уровнем указывает на то, что субъект имеет повышенную чувствительность к развитию заболевания или состояния, опосредованного функцией эффекторных Т-клеток, воспаления или сеписа.
22. Способ по п. 21, в котором образец берут от субъекта, которому был введен антиген.
23. Способ по п. 22, в котором образец берут из места локального заболевания у субъекта.

