Область техники
Настоящее изобретение относится к биотехнологии, в частности к способу получения L-аминокислот методом ферментации, и более конкретно к генам, полученным из бактерии Escherichia coli. Указанные гены позволяют улучшить продукцию L-аминокислот, например L-треонина, L-валина, L-пролина, L-лейцина и L-метионина.
Предшествующий уровень техники
Традиционно L-аминокислоты в промышленном масштабе могут быть получены методом ферментации с использованием штаммов микроорганизмов, полученных из природных источников или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано множество методов увеличения продукции L-аминокислот, например, путем трансформации микроорганизма рекомбинантной ДНК (см., например, патент США 4278765). Указанные методы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот, и/или уменьшении чувствительности целевого фермента к обратному ингибированию, продуцируемой L-аминокислотой (см. , например, выложенную патентную заявку Японии 56-18596 (1981), WO 95/16042 или патенты США 5661012 и 6040160).
С другой стороны, повышенная экскреция L-аминокислот может увеличить продуктивность штамма, продуцирующего L-аминокислоту. Описан штамм бактерии, принадлежащей к роду Corynebacterium, обладающей повышенной экспрессией гена экскреции L-лизина (ген lysE) (WO 9723597 A2). Кроме того, описаны гены, кодирующие белки, способные к секреции L-цистеина, L-цистина, N-ацетилсерина или производных триазолидина (патент США 5972633).
К настоящему времени описаны несколько генов, кодирующих, как предполагается, мембранные белки, которые увеличивают продукцию L-аминокислот. Дополнительная копия гена rhtB делает бактерию более устойчивой к L-гомосерину и увеличивает продукцию L-гомосерина, L-треонина, L-аланина, L-валина и L-изолейцина (Европейская патентная заявка ЕР 994190 А2). Дополнительная копия гена rhtC делает бактерию более устойчивой к L-гомосерину и L-треонину и увеличивает продукцию L-гомосерина, L-треонина и L-лейцина (Европейская патентная заявка ЕР 1013765 А1). Дополнительные копии генов yahN, yeaS, yfiK, uyggA увеличивают продукцию L-глутаминовой кислоты, L-треонина, L-аланина, L-гистидина, L-пролина, L-аргинина, L-валина и L-изолейцина (Европейская патентная заявка ЕР 1016710 А2). И хотя нуклеотидная последовательность всего генома Escherichia coli К-12 описана (Blattner F.R., Plunkett G., Bloch C.A. et al., Science, 227, 1453-1474,1997; ftp://ftp.genetics.wisc.edu/pub/sequence/ecolim52.seq.gz), существует множество открытых рамок считывания, функция которых остается неизвестной.
Описание изобретения
Целью настоящего изобретения является увеличение продуктивности штаммов, продуцирующих L-аминокислоты, и предоставление способа получения L-аминокислот, например L-треонина, L-валина, L-пролина, L-лейцина или L-метионина, с использованием указанных штаммов.
Данная цель была достигнута путем идентификации генов, кодирующих белки, которые не вовлечены в пути биосинтеза целевых L-аминокислот, но увеличивают их продукцию. Примером такого белка может являться мембранный белок, обладающий активностью, обеспечивающей экскрецию L-аминокислот. При анализе нуклеотидной последовательности полного генома Escherichia coli были отобраны белки с 4 или более предполагаемыми трансмембранными сегментами (ТМС). В результате тщательного исследования авторы настоящего изобретения идентифицировали среди них два гена, такие как b2682 и b2683, и тщательно изучили их. Гены b2682 и b2683 были известны как предполагаемые кодирующие последовательности, которые могут кодировать белки с неизвестной функцией (номера нуклеотидов с 92 по 829 и с 819 по 1154 в нуклеотидной последовательности под номером АЕ000353 U00096 в Genbank соответственно). Ген b2683 также известен как ygaH. Также авторы настоящего изобретения установили, что при повышении активности белков, кодируемых генами b2682 и b2683, увеличивается продуктивность штаммов, продуцирующих L-аминокислоты. Таким образом было совершено настоящее изобретение.
Настоящее изобретение включает в себя следующее:
1. Бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia, в которой продукция L-аминокислоты упомянутой бактерией увеличена за счет повышения активности белков, описанных в пунктах (А) или (В) и (С) или (D), в клетке упомянутой бактерии:
(A) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 3;
(B) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 3, и который обладает активностью, придающей бактерии устойчивость к L-аминокислотам и/или их аналогам;
(C) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 5;
(D) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 5, и который обладает активностью, придающей бактерии устойчивость к L-аминокислотам и/или их аналогам;
(здесь и далее белки, описанные в вышеупомянутых пунктах (А) или (В) и (С) или (D), упоминаются как "белки согласно настоящему изобретению") (список последовательностей см. в конце описания).
2. Бактерия в соответствии с вышеупомянутой бактерией, в которой активности белков, описанных в пунктах (А) или (В) и (С) или (D), повышены путем трансформации бактерии с помощью ДНК, кодирующей белки, описанные в пунктах (А) или (В) и (С) или (D), или путем изменения регуляции экспрессии нуклеотидной последовательности в хромосоме упомянутой бактерии.
3. Бактерия в соответствии с вышеупомянутой бактерией, в которой трансформация осуществляется с использованием многокопийного вектора.
4. Способ получения L-аминокислоты, включающий выращивание бактерии в соответствии с вышеупомянутой бактерией в питательной среде и сбор из культуральной жидкости полученной и накопленной в ней L-аминокислоты.
5. Способ в соответствии с вышеупомянутым способом, в котором L-аминокислотой является L-треонин.
6. Способ в соответствии с вышеупомянутым способом, в котором у указанной бактерии повышена экспрессия треонинового оперона.
7. Способ в соответствии с вышеупомянутым способом, в котором L-аминокислотой является L-валин.
8. Способ в соответствии с вышеупомянутым способом, в котором у указанной бактерии повышена экспрессия генов ilv оперона.
9. Способ в соответствии с вышеупомянутым способом, в котором L-аминокислотой является L-пролин.
10. Способ в соответствии с вышеупомянутым способом, в котором у указанной бактерии повышена экспрессия генов биосинтеза L-пролина.
11. Способ в соответствии с вышеупомянутым способом, в котором L-аминокислотой является L-лейцин.
12. Способ в соответствии с вышеупомянутым способом, в котором у указанной бактерии повышена экспрессия генов leu оперона.
13. Способ в соответствии с вышеупомянутым способом, в котором L-аминокислотой является L-метионин.
14. Способ в соответствии с вышеупомянутым способом, в котором у указанной бактерии повышена экспрессия генов met регулона.
Способ получения L-аминокислоты включает продукцию L-треонина с использованием бактерии - продуцента L-треонина, в которой повышены активности белков согласно настоящему изобретению, например, представленных аминокислотными последовательностями, приведенными под номерами 3 и 5. Также способ получения L-аминокислоты включает продукцию L-валина с использованием бактерии - продуцента L-валина, в которой повышены активности белков согласно настоящему изобретению, например, представленных аминокислотными последовательностями, приведенными под номерами 3 и 5. Кроме того, способ получения L-аминокислоты включает продукцию L-пролина с использованием бактерии - продуцента L-пролина, в которой повышены активности белков согласно настоящему изобретению, например, представленных аминокислотными последовательностями, приведенными под номерами 3 и 5. Далее способ получения L-аминокислоты включает продукцию L-лейцина с использованием бактерии - продуцента L-лейцина, в которой повышены активности белков согласно настоящему изобретению, например, представленных аминокислотными последовательностями, приведенными под номерами 3 и 5. И, наконец, способ получения L-аминокислоты включает продукцию L-метионина с использованием бактерии - продуцента L-метионина, в которой повышены активности белков согласно настоящему изобретению, например, представленных аминокислотными последовательностями, приведенными под номерами 3 и 5.
Настоящее изобретение более детально будет описано ниже.
Вышеуказанной бактерией согласно настоящему изобретению является бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia, в которой продукция L-аминокислоты указанной бактерией увеличена за счет повышения активностей белков согласно настоящему изобретению в клетке бактерии.
Вышеуказанной бактерией согласно настоящему изобретению является бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia и обладающая повышенной активностью белков, которые увеличивают продукцию целевой L-аминокислоты. Конкретно, бактерией согласно настоящему изобретению является бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia и обладающая повышенной активностью белков согласно настоящему изобретению. Более конкретно, бактерия согласно настоящему изобретению содержит ДНК, в которой повышена экспрессия генов b2682 и b2683 на хромосоме ДНК или на плазмиде в бактерии, обладает повышенной способностью к продукции L-аминокислоты, например L-треонина, L-валина, L-пролина, L-лейцина и/или L-метионина.
К белкам согласно настоящему изобретению относятся белки, описанные в следующих пунктах (А) или (В) и (С) или (D):
(A) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 3;
(B) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 3, и который обладает активностью, придающей бактерии устойчивость к L-аминокислотам и/или их аналогам;
(C) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 5;
(D) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 5, и который обладает активностью, придающей бактерии устойчивость к L-аминокислотам и/или их аналогам.
Количество "нескольких" аминокислот различается в зависимости от положения и типа аминокислотного остатка в трехмерной структуре белка. Оно может быть от 2 до 24, предпочтительно от 2 до 12 и более предпочтительно от 2 до 5 для белка (А), и от 2 до 11, предпочтительно от 2 до 7 и более предпочтительно от 2 до 5 для белка (С) соответственно.
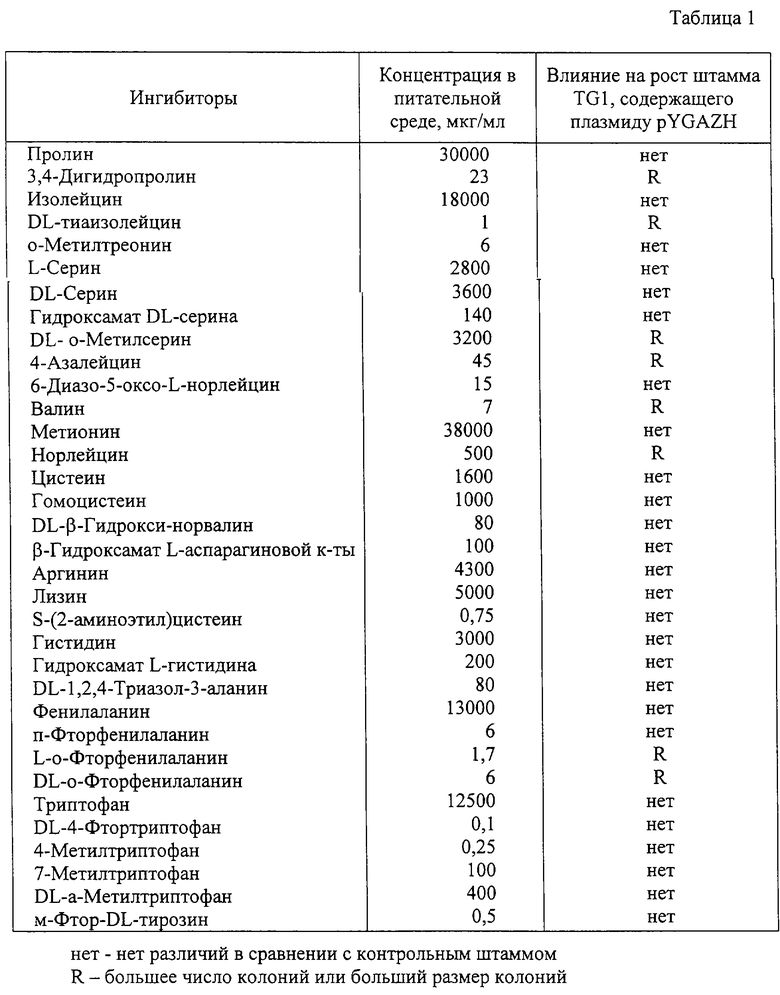
Устойчивость к L-аминокислотам и/или их аналогам означает способность бактерии к росту на минимальной питательной среде, содержащей L-аминокислоту и/или ее аналог в концентрации, при которой штамм дикого типа или родительский штамм не может расти, или способность бактерии расти с большей скоростью на питательной среде, содержащей L-аминокислоту и/или ее аналог, чем штамм дикого типа или родительский штамм. Примерами аналогов L-аминокислот являются 3,4-дигидропролин, DL-тиаизолейцин, DL-o-метилсерин, 4-азалейцин, норлейцин, L-o-фторфенилаланин и DL-o-фторфенилаланин. Упомянутая выше концентрация L-аминокислоты и ее аналога варьируется значительно (от 0,5 мкг/мл в случае DL-тиаизолейцина до 9600 мкг/мл в случае DL-o-метилсерина) в зависимости от химической структуры используемого соединения. Например, указанная концентрация составляет обычно от 7 до 70 мкг/мл, предпочтительно от 20 до 25 мкг/мл в случае 3,4-дигидропролина; обычно от 0,5 до 5,0 мкг/мл, предпочтительно от 0,9 до 1,1 мкг/мл в случае DL-тиаизолейцина; обычно от 1100 до 9600 мкг/мл, предпочтительно от 3000 до 3500 мкг/мл в случае DL-o-метилсерина; обычно от 15 до 150 мкг/мл, предпочтительно от 40 до 50 мкг/мл в случае 4-азалейцина; обычно от 150 до 1500 мкг/мл, предпочтительно от 450 до 550 мкг/мл в случае норлейцина; обычно от 0,6 до 6,0 мкг/мл, предпочтительно от 1,5 до 2,0 мкг/мл в случае L-o-фторфенилаланина; обычно от 2 до 20 мкг/мл, предпочтительно от 5 до 7 мкг/мл в случае DL-o-фторфенилаланина.
К бактерии согласно настоящему изобретению также относятся бактерии, в которой активности белков согласно настоящему изобретению повышены путем трансформации бактерии с помощью ДНК, кодирующей белки, описанные в пунктах (А) или (В) и (С) или (D), или путем изменения регуляции экспрессии последовательности указанной ДНК в хромосоме упомянутой бактерии.
Упомянутая ДНК, использующаяся для модификации бактерии согласно настоящему изобретению, кодирует, как предполагается, мембранный белок. Конкретно, упомянутая ДНК кодирует белок с 4 или более трансмембранными сегментами. Такая ДНК может кодировать белки, обладающие активностью по экскреции L-аминокислот. Более конкретно, гены b2682 и b2683 являются такой ДНК. Необходимо отметить, что кодирующий участок гена b2682 в области 728-738 и кодирующий участок гена b2683 в области 1-11 перекрываются. Оба гена могут быть получены, например, с помощью ПЦР с использованием затравок с нуклеотидной последовательностью, приведенной под номерами 1 и 2, в виде единственного продукта.
Анализ последовательности полного генома Escherichia coli позволил выбрать гены, кодирующие белки с 4 и более предполагаемыми ТМС. Белки с известной функцией и транспортеры, описанные Paulsen I.T., Sliwinski M.I., Saier M.H. (J.Mol.Biol., 1998, 277, 573) и Linton K.J., Higgins C.F. (Molecular Microbiology, 1998, 28(1), 5), исключили из группы, подлежащей изучению. В результате тщательного отбора среди оставшихся генов были выбраны несколько генов, кодирующих, как предполагалось, мембранные экспортеры. И было обнаружено, что повышенная экспрессия генов b2682 и b2683 увеличивает продукцию L-аминокислот штаммами - продуцентами L-аминокислот.
К ДНК согласно настоящему изобретению относится ДНК, кодирующая белок, включающий делеции, замены, вставки или добавление одной или нескольких аминокислот в одно или несколько положений белка (А) или (С), при условии, что они не приводят к утрате активности указанного белка. Хотя количество "нескольких" аминокислот различается в зависимости от положения и типа аминокислотного остатка в трехмерной структуре белка, оно может быть от 2 до 24, предпочтительно от 2 до 12 и более предпочтительно от 2 до 5 для белка (А) и от 2 до 11, предпочтительно от 2 до 7 и более предпочтительно от 2 до 5 для белка (С) соответственно. ДНК, кодирующая практически такой же белок, как белок, описанный в пунктах (А) и (С), может быть получена, например, путем модификации нуклеотидной последовательности, кодирующей белки, описанные в пунктах (А) и (С), с использованием сайт-направленного мутагенеза таким образом, что один или несколько аминокислотных остатков будут удалены, заменены, введены или добавлены. Модифицированная подобным образом ДНК может быть получена традиционными методами, использующими обработку химическими реагентами и содержание в условиях, вызывающих мутации. К подобного рода обработкам относятся обработка ДНК, кодирующей белки согласно настоящему изобретению, с помощью гидроксиламина или обработка бактерии, содержащей ДНК, с помощью УФ-излучения или химического реагента, такого как N-метил-N'-нитро-N-нитрозогуанидин или азотистая кислота.
К ДНК согласно настоящему изобретению относятся варианты, которые могут быть найдены в различных штаммах или вариантах бактерий, принадлежащих к роду Escherichia, ввиду природного разнообразия. ДНК, кодирующая подобные варианты, может быть получена путем выделения ДНК, которая гибридизуется с генами b2682 и b2683 или частью указанных генов в жестких условиях и которая кодирует белок, увеличивающий продукцию L-аминокислот. Термин "жесткие условия", упомянутый здесь, означает условия, при которых образуются так называемые специфические гибриды, а неспецифические не образуются. Например, к жестким условиям относятся условия, при которых гибридизуются ДНК, обладающие высокой степенью гомологии, к примеру ДНК, обладающие гомологией не менее 70% друг относительно друга. В качестве варианта примером жестких условий являются условия, соответствующие условиям отмывки при гибридизации по Саузерну, например 60oС, 1xSSC, 0,1% SDS, предпочтительно O,lxSSC, 0,1% SDS. В качестве зонда для ДНК, кодирующей варианты и гибридизующейся с генами b2682 и b2683, также может быть использована часть нуклеотидной последовательности под номером 3 или номером 5 соответственно. Зонд подобного рода может быть получен в результате ПЦР с использованием в качестве затравок олигонуклеотидов, полученных на основе нуклеотидной последовательности под номером 3 или номером 5, и фрагмента ДНК, содержащего нуклеотидную последовательность под номером 3 или номером 5, в качестве матрицы. В случае, когда в качестве зонда используется фрагмент ДНК длиной около 300 пар оснований, условия отмывки при гибридизации соответствуют, например, 50oС, 2xSSC и 0,1% SDS.
Трансформация бактерии с помощью ДНК, кодирующей белок, означает введение указанной ДНК в клетку-бактерию, например, с помощью традиционных методов для того, чтобы усилить экспрессию генов, кодирующих белок согласно настоящему изобретению, и повысить активность белка в клетке-бактерии.
К методам увеличения экспрессии генов относятся методы увеличения числа копий гена. Введение гена в вектор, способный к функционированию в бактерии, принадлежащей к роду Escherichia, увеличивает число копий указанного гена. Для подобных целей могут быть предпочтительно использованы многокопийные векторы. Примерами многокопийных векторов являются pBR322, pMWl19, pUC19, pET22b и подобные им.
Кроме того, усиление экспрессии гена может быть достигнуто путем введения некоторого числа копий гена в бактериальную хромосому, например, методом гомологичной рекомбинации или подобным.
В случае, когда добиваются усиления экспрессии двух или более генов, указанные гены могут быть расположены вместе на одной и той же плазмиде или раздельно на различных плазмидах. Также допустимо, чтобы одни из генов располагались в хромосоме, а другие гены располагались на плазмиде.
С другой стороны, усиление экспрессии генов может быть достигнуто помещением ДНК согласно настоящему изобретению под контроль сильного промотора. Например, в качестве сильных промоторов известны lac промотор, trp промотор, trc промотор, PL и PR промоторы фага лямбда. Использование сильного промотора может быть совмещено с увеличением числа копий гена.
Бактерия согласно настоящему изобретению может быть получена путем введения вышеуказанных ДНК в бактерию, уже обладающую способностью к продукции L-аминокислоты. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания бактерии, уже содержащей указанные ДНК, способности к продукции L-аминокислоты. В качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии - продуценты L-треонина, принадлежащие к роду Escherichia, такие как VL2054 (ВКПМ В-8067), ВНИИГенетика 472Т23 (патент США 5631157), ВКПМ В-3996 (патенты США 5175107 и 5976843), КССМ-10132 (WО 009660 A1), КССМ-10133 (WО 009661 A1) или подобные им. Также в качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии - продуценты L-валина, принадлежащие к роду Escherichia, такие как Н-81 (ВКПМ В-8066), NRRL В-12287 и NRRL В-12288 (патент США 4391907), ВКПМ В-4411 (патент США 5658766), ВКПМ В-7707 (европейская патентная заявка ЕР 1016710 А2) или подобные им. Кроме того, в качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии - продуценты L-пролина, принадлежащие к роду Escherichia, такие как NRRL В-12403 и NRRL В-12404 (патент Великобритании GB 2075056), ВКПМ В-8012 (Российская патентная заявка 2000124295), мутанты, содержащие плазмиды, описанные в патенте Германии DE 3127361, мутанты, содержащие плазмиды, описанные в Bloom F. R. et al (The 15th Miami winter symposium, 1983, p.34) и подобные им. Также в качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии - продуценты L-лейцина, принадлежащие к роду Escherichia, такие как Н-9070 (FERM ВР-4704) и Н-9072 (FERM ВР-4706) (патент США 5744331), ВКПМ В-7386 и ВКПМ В-7388 (патент РФ 2140450), W1485atpA401/pMWdAR6, W1485lip2/pMWdAR6 и AJ12631/pMWdAR6 (Европейский патент 0872547) или подобные им. И, наконец, в качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии - продуценты L-метионина, принадлежащие к роду Escherichia, такие как АJ 11539 (NRRL В-12399), АJ 11540 (NRRL В-12400), АJ 11541 (NRRL В-12401), АJ 11542 (NRRL В-12402) (патент Великобритании GB 2075055) или подобные им.
В бактерии согласно настоящему изобретению в дальнейшем может быть усилена экспрессия одного или нескольких генов, вовлеченных в биосинтез L-аминокислот. Примерами таких генов для бактерии - продуцента L-треонина являются гены треонинового оперона, предпочтительно ген, кодирующий аспартаткиназу - гомосериндегидрогеназу, чувствительность к ингибированию L-треонином по типу обратной связи в которой утрачена (патент Японии 1-29559). Примерами таких генов для бактерии - продуцента L-валина являются гены оперона ilvGMEDA, в котором предпочтительно не экспрессируется треониндезаминаза и аттенуация которого подавлена (выложенная патентная заявка Японии 8-47397). Примерами таких генов для бактерии - продуцента L-пролина являются гены биосинтеза пролина, предпочтительно ген рrоВ, кодирующий глутаматкиназу, чувствительность к ингибированию L-пролином по типу обратной связи в которой утрачена (патент Германии DЕ 3127361). Также примерами таких генов для бактерии - продуцента L-лейцина являются гены лейцинового оперона, leu оперона, предпочтительно ген, кодирующий изопропилмалатсинтазу, чувствительность к ингибированию L-лейцином по типу обратной связи в которой утрачена (Российская патентная заявка 99114325). Также примерами таких генов для бактерии - продуцента L-метионина являются гены метионинового регулона. Указанный метиониновый регулон может содержать мутантные гены, кодирующие белки с пониженной активностью в репрессии биосинтеза аминокислоты. Примером такого гена является вариантный тип гена metJ из E.coli, кодирующий репрессор биосинтеза L-метионина, активность которого в репрессии биосинтеза L-метионина понижена (выложенная патентная заявка Японии JP 2000157267 A2).
К способам согласно настоящему изобретению относится способ продукции L-треонина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-треонина в указанной питательной среде и сбора L-треонина из культуральной жидкости. Также к способам согласно настоящему изобретению относится способ продукции L-валина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-валина в указанной питательной среде и сбора L-валина из культуральной жидкости. Кроме того, к способам согласно настоящему изобретению относится способ продукции L-пролина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-пролина в указанной питательной среде и сбора L-пролина из культуральной жидкости. Также к способам согласно настоящему изобретению относится способ продукции L-лейцина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-лейцина в указанной питательной среде и сбора L-лейцина из культуральной жидкости. И, наконец, к способам согласно настоящему изобретению относится способ продукции L-метионина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-метионина в указанной питательной среде и сбора L-метионина из культуральной жидкости.
Согласно настоящему изобретению выращивание, сбор и очистка L-аминокислоты из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием микроорганизма. Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, которые требуются микроорганизму для роста. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, и различные органические кислоты. В зависимости от степени ассимиляции используемого микроорганизма могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут использоваться различные соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов и ферментолизат микроорганизмов. В качестве минеральных добавок используются монофосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные соединения.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание, ферментация с аэрацией, при температуре от 20 до 40oС, предпочтительно от 30 до 38oС. рН питательной среды находится в пределах от 5 до 9, предпочтительно от 6,5 до 7,2. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферами. Обычно выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем целевая L-аминокислота может быть собрана и очищена методами ионообменной хроматографии, концентрирования и кристаллизации.
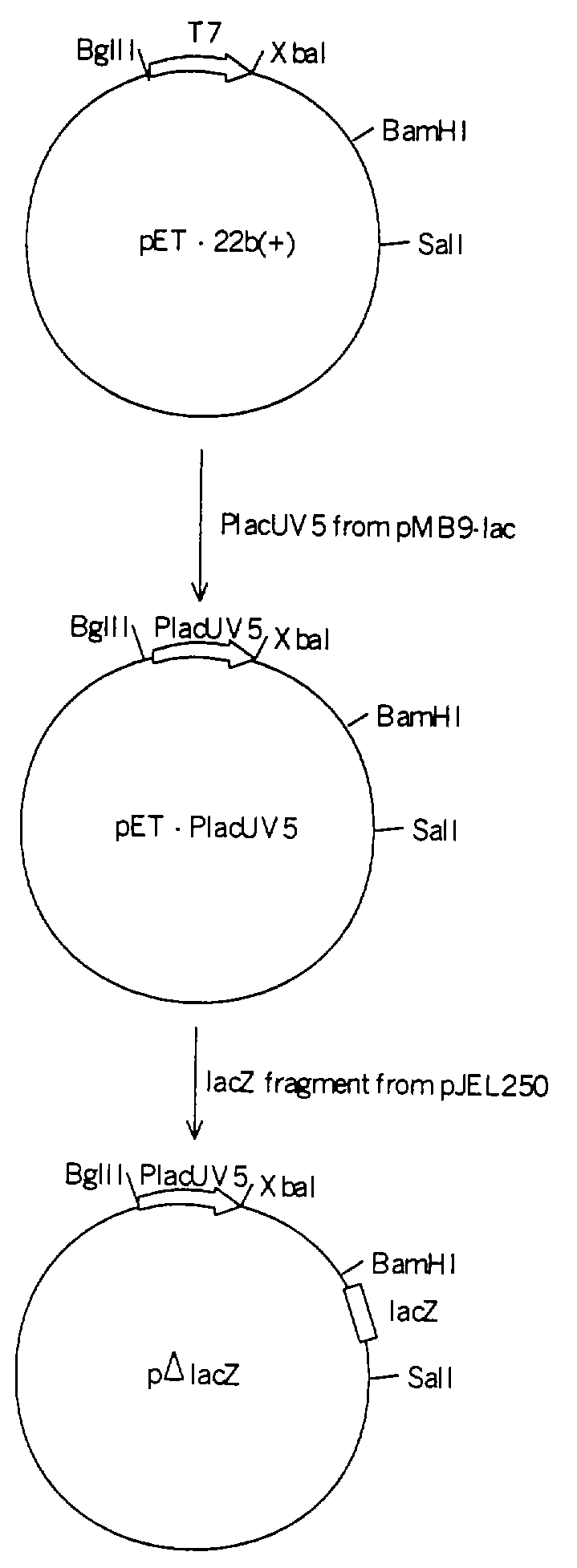
На чертеже показана схема конструирования плазмиды pΔlacZ.
Наилучший способ осуществления изобретения
Настоящее изобретение более детально описано со ссылкой на примеры. В указанных примерах аминокислоты являются аминокислотами L-конфигурации, если не указано иное.
Пример 1. Клонирование генов b2682 и b2683 на плазмиде pΔlacZ.
Для клонирования генов b2682 и b2683 был использован вектор pΔlacZ. Вектор pΔlacZ является производным вектора pET-22b(+) (Novagen, Madison, WI, USA). Вектор pET-22b(+) был обработан рестриктазами BglII и XbaI и лигирован с фрагментом полимеразной цепной реакции (ПЦР), полученным на плазмиде рМВ9-lас (Fuller F., Gene, 19, 43-54, 1982), обработанным теми же рестриктазами, и содержащим промотор PlacUV5. Для амплификации с помощью ПЦР фрагмента с промотором PlacUV5 использовались затравки, приведенные под номерами 7 и 8. В полученную плазмиду была вставлена структурная часть гена lacZ (237 пар оснований без промотора) путем клонирования SalI-BamHI фрагмента плазмиды pJEL250 (Дымакова Е. и др.. Генетика, 35, 2, 181-186, 1999). Схема конструкции вектора pΔlacZ показана на чертеже.
Исходным материалом для клонирования предполагаемых рамок считывания b2682 и b2683 из Е.coli (генов b2682 и b2683) был фрагмент ПЦР, полученный с использованием ДНК из штамма Е. coli TG1 в качестве матрицы. Для синтеза этого фрагмента были использованы две затравки, нуклеотидная последовательность которых приведена под номерами 1 и 2. ПЦР осуществлялась на "Perkin Elmer GeneAmp PCR System 2400" в следующих условиях: 40 секунд при 95oС, 40 секунд при 47oС, 40 секунд при 72oС, 30 циклов. Таким образом был получен линейный фрагмент ДНК длиной 1158 пар оснований, содержащий гены b2682 и b2683. Данный фрагмент ПЦР был обработан рестриктазами ХbaI и ВаmHI и введен в многокопийный вектор pΔlacZ, предварительно обработанный теми же рестриктазами.
Полученная плазмида, содержащая фрагмент ПЦР, была названа как pYGAZH и содержала гены b2682 и b2683 под контролем лактозного промотора (PlacUV5).
Пример 2. Влияние амплифицированных генов b2682 и b2683 на устойчивость штамма Е.coli TG1 к аминокислотам и их аналогам.
Штамм Е. coli TG1 (pYGAZH) и штамм Е.coli TG1, содержащий вектор без вставки (контрольный штамм), выращивались в течение ночи в среде LB, содержащей ампициллин (100 мкг/мл). Ночные культуры всех штаммов были разбавлены в 25 раз свежей средой LB, содержащей ампициллин (100 мкг/мл) и ИПТГ (0,5 мМ), и инкубировались в течение 2 часов при 37oС с аэрацией. Культуры в фазе логарифмического роста были разбавлены 0,9%-ным раствором хлорида натрия и около 1000 клеток были высеяны на чашки с твердой питательной средой Адамса, содержащей ампициллин (100 мкг/мл), ИПТГ (0,5 мМ) и аминокислоту или ее аналог. После 2-4 дней инкубации при 37oС определялись различия в размере колоний или их числе между штаммом TG1, содержащим гибридную плазмиду, и контрольньм штаммом TG1. Результаты экспериментов представлены в таблице 1.
Пример 3. Продукция треонина штаммом, содержащим плазмиду pYGAZH.
Штамм VL2054 - продуцент треонина был трансформирован плазмидой pYGAZH, содержащей гены b2682 и b2683 под контролем промотора PlacUV5. Полученный штамм был назван VL2054(pYGAZH). Штамм VL2054 является производным штамма ВКПМ В-3996 и содержит на хромосоме:
а) интегрированный треониновый оперон под контролем промотора PR;
б) нативный ген rhtA;
в) инактивированный хромосомный ген, кодирующий треониндегидрогеназу (ген tdh), и инактивированный ген устойчивости к канамицину (ген kan) в Тn5 (tdh::Tn5,Kans);
г) мутацию ilvA442.
Штамм VL2054 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (Россия, 113545 Москва, 1-й Дорожный проезд, 1) 30 января 2001 года под инвентарньм номером ВКПМ В-8067.
5 колоний каждого из штаммов VL2054, VL2054(pΔlacZ) в качестве контрольного штамма, содержащего плазмиду без вставки, и VL2054(pYGAZH) были суспендированы в 2 мл минимальной питательной среды ((NH4)2SО4 - 11 г/л, NaCl - 0,4 г/л, MgSО4 - 0,4 г/л, К2НРО4 - 1 г/л, FeSО4 - 10 мг/л, MnSО4 - 10 мг/л, тиамин - 0,1 мг/л, дрожжевой экстракт - 0,5 г/л, глюкоза - 40 г/л, ампициллин - 300 мг/л, если необходимо) в 20 мл пробирках и инкубировались в течение ночи с аэрацией при 32oС. 0,2 мл каждой ночной культуры были перенесены в три 20 мл пробирки с 2 мл свежей среды для ферментации с или без ИПТГ и выращивались при 32oС в течение 48 или 72 часов на роторной качалке.
Состав среды для ферментации:
(NH4)2SО4 - 22 г/л
NaCl - 0,8 г/л
MgSО4 - 0,8 г/л
К2НРО4 - 2,0 г/л
FeSО4 - 20 мг/л
MnSО4 - 20 мг/л
Тиамин - 0,2 мг/л
Дрожжевой экстракт - 1,0 г/л
СаСО3 - 30 г/л
Глюкоза - 80 г/л
Ампициллин - 300 мг/л, если необходимо
ИПТГ - 0,5 мМ, если необходимо
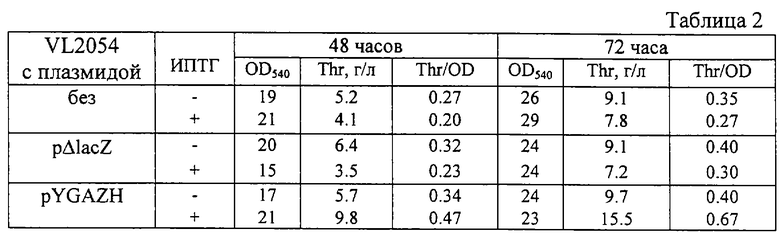
После выращивания стабильность плазмиды и оптическая плотность культуральной жидкости при 540 нм были определены традиционными методами. Накопленное в культуральной жидкости количество треонина было определено с помощью тонкослойной хроматографии (ТСХ). Состав подвижной фазы для ТСХ следующий: изопропанол - 50 мл, ацетон - 50 мл, NH4ОH (30%) - 12 мл, Н2О - 8 мл. Результаты приведены в таблице 2. Как видно, гибридная плазмида pYGAZH увеличивала накопление треонина штаммом VL2054 - продуцентом треонина.
Пример 4. Продукция валина штаммом, содержащим плазмиду pYGAZH.
Штамм Н-81 - продуцент валина был трансформирован плазмидой pYGAZH, содержащей гены b2682 и b2683 под контролем промотора PlacUV5.
Штамм Н-81 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (Россия, 113545 Москва, 1-й Дорожный проезд, 1) 30 января 2001 года под инвентарньм номером ВКПМ В-8066.
5 колоний каждого из штаммов Н-18, H-81(pΔlacZ) в качестве контрольного штамма, содержащего плазмиду без вставки, и Н-81 (pYGAZH) были суспендированы в 2 мл минимальной питательной среды ((NH4)2SО4 - 18 г/л, K2HPО4 - 1,8 г/л, MgSО4 - 1,2 г/л, тиамин - 0,1 мг/л, дрожжевой экстракт - 0,5 г/л, глюкоза - 60 г/л, ампициллин - 300 мг/л, если необходимо) в 20 мл пробирках и инкубировались в течение ночи с аэрацией при 32oС. 0,2 мл каждой ночной культуры были перенесены в три 20 мл пробирки с 2 мл свежей среды для ферментации с или без ИПТГ и выращивались при 32oС в течение 48 или 72 часов на роторной качалке.
Состав среды для ферментации:
(NH4)2SО4 - 18 г/л
К2НРО4 - 1,8 г/л
MgSО4 - 1,2 г/л
СаСО3 - 20 г/л
Тиамин - 0,1 мг/л
Глюкоза - 60 г/л
Ампициллин - 300 мг/л, если необходимо
ИПТГ - 0,5 мМ, если необходимо
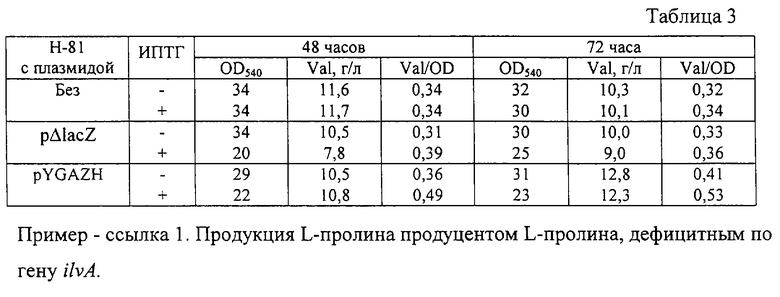
После выращивания стабильность плазмиды и оптическая плотность культуральной жидкости при 540 нм были определены традиционными методами. Накопленное в культуральной жидкости количество валина было определено с помощью тонкослойной хроматографии (ТСХ). Состав подвижной фазы для ТСХ следующий: изопропанол - 80 мл, этилацетат - 80 мл, NH4ОH (30%) - 15 мл, Н2О - 45 мл. Результаты приведены в таблице 3. Как видно, гибридная плазмида pYGAZH увеличивала накопление валина штаммом Н-81 - продуцентом валина.
Клетки природного штамма Е.coli К12 (ВКПМ В-7) были обработаны мутагеном, N-метил-N'-нитро-N-нитрозогуанидином (0,1 мг/мл), в течение 20 минут при 37oС, отмыты и помещены на минимальную агаризованную среду М9, дополненную 1,25 мг/мл триптона, 10 мг/мл L-пролина и 0,05 мг/мл хлорида 2,3,5-трифенилтетразолина. Большинство колоний, выросших после 3 дней инкубации при 37oС, были окрашены красным. Несколько колоний, не способных окислять L-пролин, были белыми. Одна из этих колоний была использована в качестве исходной для получения мутантов, устойчивых к аналогам пролина (3,4-дегидроксипролин и азетидин-2-карбоксилат), каждый из которых был добавлен в агаризованную среду М9 в концентрации 2 мг/мл.
Некоторые из выросших мутантов могли продуцировать L-пролин. Наилучший продуцент L-пролина 702 был обработан бактериофагом Р1, выращенным на клетках штамма TG1, в котором ген ilvA был разрушен путем вставки гена устойчивости (Cmr) к хлорамфениколу (Cm). Один из устойчивых к Cm трансдуктантов, 702ilvA, который стал ауксотрофным по L-изолейцину, был гораздо более эффективным продуцентом L-пролина, чем исходный прототрофный по L-изолейцину штамм 702 (таблица 4). Питательная среда для ферментации содержала 60 г/л глюкозы, 25 г/л (NH4)2SО4, 2 г/л КН2РО4, 1 г/л MgSО4, 0,1 мг/л тиамина, 50 мг/л L-изолейцина и 25 г/л мела (рН 7,2). Глюкоза и мел были стерилизованы раздельно. 2 Мл питательной среды были помещены в пробирки и инокулированы одной петлей исследуемых микроорганизмов, затем выращивание производилось при 37oС в течение 2 дней на качалке.
Штаммы 702 и 702ilvA были депонированы во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) 25 июля 2000 г. под инвентарньми номерами ВКПМ В-8011 и ВКПМ В-8012 соответственно.
Пример 5. Продукция пролина штаммом, содержащим плазмиду pYGAZH.
Штамм E. coli 702ilvA - продуцент пролина был трансформирован плазмидой pYGAZH, содержащей гены b2682 и b2683 под контролем промотора PlacUV5.
5 колоний каждого из штаммов 702ilvA, 702ilvA (pΔlacZ) в качестве контрольного штамма, содержащего плазмиду без вставки, и 702ilvA(pYGAZH) были суспендированы в 2 мл минимальной питательной среды ((NH4)2SO4 - 18 г/л, К2НРО4 - 1,8 г/л, MgSО4 - 1,2 г/л, тиамин - 0,1 мг/л, дрожжевой экстракт - 0,5 г/л, глюкоза - 60 г/л, изолейцин - 50 мг/л, ампициллин - 300 мг/л, если необходимо) в 20 мл пробирках и инкубировались в течение ночи с аэрацией при 32oС. 0,2 мл каждой ночной культуры были перенесены в три 20 мл пробирки с 2 мл свежей среды для ферментации с или без ИПТГ и выращивались при 32oС в течение 40 часов на роторной качалке.
Состав среды для ферментации:
(NH4)2SО4 - 18 г/л
К2НРО4 - 1,8 г/л
MgSО4 - 1,2 г/л
СаСО3 - 20 г/л
Тиамин - 0,1 мг/л
Глюкоза - 60 г/л
Изолейцин - 50 мг/л
Ампициллин - 300 мг/л, если необходимо
ИПТГ - 0,5 мМ, если необходимо
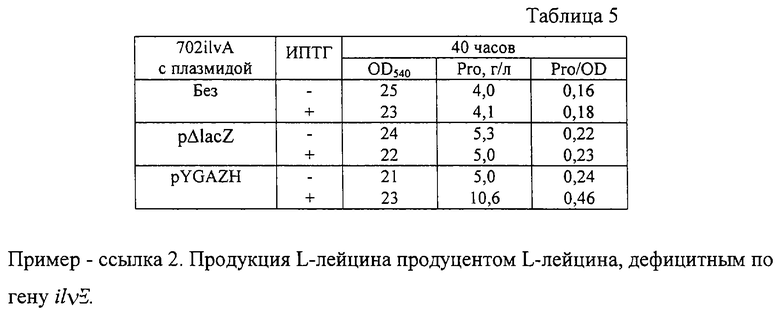
После выращивания стабильность плазмиды и оптическая плотность культуральной жидкости при 540 нм были определены традиционными методами. Накопленное в культуральной жидкости количество пролина было определено с помощью тонкослойной хроматографии (ТСХ). Состав подвижной фазы для ТСХ следующий: этанол - 80 мл, NH4OH (30%) - 5 мл, Н2О - 25 мл. Результаты приведены в таблице 5. Как видно, гибридная плазмида pYGAZH увеличивала накопление пролина штаммом 702ilvA - продуцентом пролина.
Клетки природного штамма Е.coli K12 (ВКПМ В-7) были обработаны мутагеном, N-метил-N'-нитро-N-нитрозогуанидином (0,05 мг/мл), в течение 20 минут при 37oС, отмыты 4 раза физиологическим раствором и помещены на минимальную агаризованную среду М9, дополненную 4,0 мг/мл DL-4-азалейцина. Чашки инкубировались в течение 5 дней при 37oС. Выросшие на чашках колонии были собраны и очищены путем рассева до отдельных колоний на чашках с L-агаром. Один из полученных мутантов, устойчивых к DL-4-азалейцину, был использован для индукции двойной ауксотрофности по L-изолейцину и L-валину. Было получено большое количество двойных ауксотрофов, требующих для роста L-изолейцин и L- валин. Было показано, что двойная ауксотрофность по L-изолейцину и L-валину обуславливалась мутацией в гене ilvE. Среди полученных двойных ауксотрофов был выбран наилучший продуцент L-лейцина - штамм 505, продуцирующий 1,8 г/л L-лейцина. Питательная среда для ферментации содержала 60 г/л глюкозы, 25 г/л (NH4)2SО4, 2 г/л КН2РО4, 1 г/л MgSО4, 0,1 мг/л тиамина, 100 мг/л изолейцина, 100 мг/л валина и 25 г/л мела (рН 7,2). Глюкоза и мел были стерилизованы раздельно. 2 мл питательной среды были помещены в пробирки и инокулированы одной петлей исследуемых микроорганизмов, затем выращивание производилось при 37oС в течение 2 дней на качалке.
Штамм 505 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (РФ, 113545, Москва, 1-й Дорожный проезд, 1) 14 мая 2001 г. под инвентарным номером ВКПМ В-8124.
Пример 6. Продукция лейцина штаммом, содержащим плазмиду pYGAZH.
Штамм E.coli 505 - продуцент лейцина был трансформирован плазмидой pYGAZH, содержащей гены b2682 и b2683 под контролем промотора PlacUV5.
20 колоний каждого из штаммов 505, 505 (pΔlacZ) в качестве контрольного штамма, содержащего плазмиду без вставки, и 505(pYGAZH) были перенесены петлей в 20 мл пробирки в L-бульоном с ампициллином (300 мг/л) или без него и инкубировались в течение ночи с аэрацией при 32oС. 0,1 мл каждой ночной культуры были перенесены в три 20 мл пробирки с 2 мл среды для ферментации с или без ИПТГ и выращивались при 32oС в течение 72 часов на роторной качалке.
Состав среды для ферментации:
(NH4)2SО4 - 15 г/л
К2НРО4 - 1,5 г/л
MgS04•7Н2О - 1,0 г/л
СаСО3 - 20 г/л (стерилизован отдельно)
Тиамин - 0,1 мг/л
Глюкоза - 60 г/л (стерилизована отдельно)
Изолейцин - 0,3 г/л
Валин - 0,3 г/л
Ампициллин - 150 мг/л, если необходимо
ИПТГ - 0,5 мМ, если необходимо
После выращивания стабильность плазмиды была определена традиционными методами. Накопленное в культуральной жидкости количество лейцина было определено с помощью тонкослойной хроматографии (ТСХ). Состав подвижной фазы для ТСХ следующий: изопропанол - 80 мл, этилацетат - 80 мл, NH4ОН (30%) - 25 мл, Н2О - 50 мл. Результаты приведены в таблице 6. Как видно, гибридная плазмида pYGAZH увеличивала накопление лейцина штаммом 505 - продуцентом лейцина.
В качестве исходного штамма был использован бесплазмидный штамм E.coli С600, дефицитный по треонину и валину. Сначала, путем трансдукции фагом Р1, выросшим на штамме E.coli К-12, были получены Leu+ варианты штамма E.coli C600. Затем после обработки полученных вариантов N-метил-N'-нитро-N-нитрозогуанидином (0,1 мг/мл) был выделен мутантный штамм 44, устойчивый к 8 г/л L-гомосерина. Указанный штамм 44 являлся дефицитным по L-треонину и был устойчив к высокой концентрации L-гомосерина. Штамм 44 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под инвентарным номером ВКПМ В-2175.
Затем с помощью мутагенеза с использованием N-метил-N'-нитро-N-нитрозогуанидина (NTG) из штамма 44 были получены мутанты, устойчивые к аналогу метионина, норлейцину. Клетки ночной культуры штамма 44, выросшие в L-бульоне, были осаждены и ресуспендированы в физиологическом растворе (0,9% NaCl), содержащем 50 мкг/мл NTG. После обработки NTG в течение 30 минут при 37oС клетки были осаждены, отмыты 4 раза физиологическим раствором и помещены на минимальную агаризованную среду М9, содержащую 0,5 мг/мл треонина и 2,5 или 5,0 мг/мл норлейцина. Чашки инкубировались в течение 5 дней при 37oС. Выросшие на чашках колонии были собраны и очищены путем рассева до отдельных колоний на чашках с L-агаром. Наилучшим продуцентом L-метионина среди них оказался штамм 218. Выращивание нового штамма 218 в пробирках, осуществленное при 32oС в течение 3 дней, привело к накоплению в питательной среде 1,1 г/л L-метионина. В качестве питательной среды для ферментации была использована минимальная среда, содержащая 15 г/л (NH4)2SО4, 1,5 г/л КН2РО4, 1,0 г/л MgSО4, 0,1 мг/л тиамина, 40 г/л глюкозы, 0,5 г/л треонина и 20 г/л карбоната кальция. Глюкоза и мел были стерилизованы раздельно.
Штамм 218 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) 14 мая 2001 г. под инвентарным номером ВКПМ В-8125.
Затем с помощью фага Р1 в ген ррс штамма 218 была введена делеция с последующей интеграцией в полученный штамм гена русА из Bacillus subtilis (Российская патентная заявка 99121636). Полученный штамм 218русА утратил устойчивость к норлейцину. Поэтому устойчивость штамма к норлейцину была получена заново, как описано выше. Среди полученных штаммов наилучшим продуцентом L-метионина оказался штамм E.coli 73, продуцировавший 1,0 г/л L-метионина в условиях, описанных выше.
Штамм E.coli 73 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (РФ, 113545 Москва, 1-й Дорожный проезд, 1) 14 мая 2001 г. под инвентарным номером ВКПМ В-8126.
Пример 7. Продукция метионина штаммом, содержащим плазмиду pYGAZH.
Штамм E. coli 73 - продуцент метионина был трансформирован плазмидой pYGAZH, содержащей гены b2682 и b2683 под контролем промотора PlacUV5.
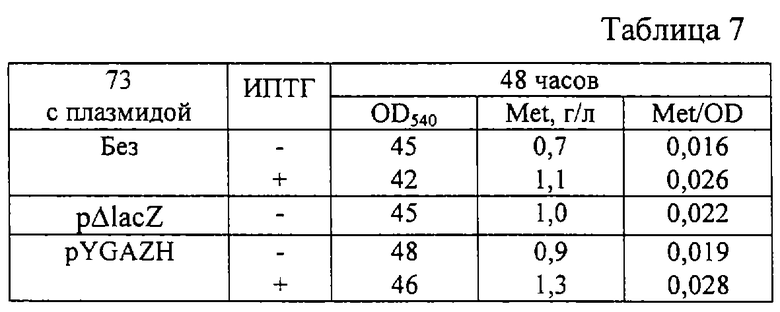
5 колоний каждого из штаммов 73, 73 (pΔlacZ) в качестве контрольного штамма, содержащего плазмиду без вставки, и 73 (pYGAZH) были суспендированы в 2 мл минимальной питательной среды ((NH4)2SО4 - 18 г/л, K2HPO4 - 1,8 г/л, MgSО4 - 1,2 г/л, тиамин - 0,1 мг/л, дрожжевой экстракт - 10 г/л, глюкоза - 60 г/л, треонин - 400 мг/л, ампициллин - 300 мг/л, если необходимо) в 20 мл пробирках и инкубировались в течение ночи с аэрацией при 32oС. 0,2 мл каждой ночной культуры были перенесены в три 20 мл пробирки с 2 мл свежей среды для ферментации с или без ИПТГ и выращивались при 32oС в течение 48 часов на роторной качалке.
Состав среды для ферментации:
(NH4)2SО4 - 18 г/л
К2НРО4 - 1,8 г/л
MgSО4 - 1,2 г/л
СаСО3 - 20 г/л
Тиамин - 0,1 мг/л
Глюкоза - 60 г/л
Треонин - 400 мг/л
Дрожжевой экстракт - 1,0 г/л
Ампициллин - 300 мг/л, если необходимо
ИПТГ - 0,5 мМ, если необходимо
После выращивания стабильность плазмиды и оптическая плотность культуральной жидкости при 540 нм были определены традиционными методами. Накопленное в культуральной жидкости количество метионина было определено с помощью тонкослойной хроматографии (ТСХ). Состав подвижной фазы для ТСХ следующий: изопропанол - 80 мл, этилацетат - 80 мл, NH4ОH (30%) - 15 мл, Н2О - 45 мл. Результаты приведены в таблице 7. Как видно, гибридная плазмида pYGAZH увеличивала накопление метионина штаммом 73 - продуцентом метионина.






Изобретение относится к биотехнологии. L-аминокислоты, такие как L-метионин, L-лейцин, L-пролин, L-валин или L-треонин, получают культивированием бактерии Escherichia coli, в которой продукция L-аминокислоты увеличена за счет повышения активности белков, кодируемых генами b2682 и b2683. После накопления L-аминокислот их извлекают из культуральной жидкости. Штаммы Е.coli трансформируют плазмидой (pYGAZH). Использование штамма Е. coli VL2054 (pYGAZH) позволяет увеличить выход L-треонина, штамма Е.coli Н-81 (pYGAZH)-L-валина, штамма Е.coli 702ilvA (pYGAZH)-L-пролина, штамма Е.coli 505 (pYGAZH)-L-лейцина, штамма Е.coli 73 (pYGAZH)-L-метионина. 6 с. и 12 з. п.ф-лы, 7 табл., 1 ил.
I. Способ получения L-аминокислот методом ферментации, включающий стадии выращивания бактерии Escherichia coli-продуцента L-аминокислоты в питательной среде и выделения L-аминокислоты из культуральной жидкости, отличающийся тем, что в качестве продуцента используют E. coli, способность к продукции L-аминокислоты которой дополнительно увеличена за счет увеличения активности белка, описанного в пункте (А) или (В) и (С) или (D): (A) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 3; (B) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 3, и который обладает активностью, придающей бактерии устойчивость к L-аминокислотам и/или их аналогам; (C) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 5; (D) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 5, и который обладает активностью, придающей бактерии устойчивость к L-аминокислотам и/или их аналогам.
| US 4278765, 14.07.1981 | |||
| US 6040160, 21.03.2000 | |||
| УСТРОЙСТВО ДЛЯ РЕГУЛИРОВАНИЯ ДВИЖЕНИЯТРАНСПОРТА | 1972 |
|
SU430430A1 |
