Предпосылки изобретения
Биологический синтез жирных кислот в высших растениях недавно стал привлекать пристальное внимание в связи с возможностью использования растительных масел в качестве возобновляемых источников восстановленного углерода. Биологический синтез жирных кислот в растениях происходит в хлоропластах зеленой ткани или в пластидах тканей, не ведущих фотосинтез. В большинстве растений основными продуктами являются сложные эфиры ацилпереносящего белка (АСР) насыщенных кислот - пальмитиновой и стеариновой.
Δ9-стеароил-ацил-переносящий белок-десатураза (Δ9-десатураза) является пластидолокализованным, немембранно связанным, растворимым ферментом, который вводит первую двойную связь в молекулу насыщенной жирной кислоты (что приводит к соответствующим моно-ненасыщенным жирным кислотам). Недавно в тканях семян различных растений было идентифицировано несколько родственных растворимых десатураз, которые содержат жирные кислоты с необычным расположением двойных связей. Ферменты этого класса растворимых десатураз являются специфичными для конкретной длины цепи субстрата и вводят двойную связь между определенными атомами углерода, ведя счет от карбоксильного конца жирной кислоты; например, Δ9-десатураза является специфичной для стеароил-АСР, и она вводит двойную связь между девятым и десятым атомами углерода. Начальные реакции введения двойной связи у животных и грибков и последующие реакции введения двойной связи в растениях передаются определенным классом десатураз жирных кислот, которые являются интегральными мембранными белками. Поскольку у большинства растений отсутствуют другие десатуразы, которые действуют на уровне 18:0, соотношение насыщенных жирных кислот к ненасыщенным жирным кислотам в высших растениях регулируется главным образом ферментами, которые катализируют превращение насыщенных жирных кислот в моно-ненасыщенные жирные кислоты.
кДНК Δ9-десатуразы кодируют белки - предшественники, содержащие N-терминальный транзитный пептид, для нацеливания в пластиду. Для сафлорового и касторового масел остаточный транзитный пептид 33 остатка транзитного пептида отщепляется, образуя зрелую десатуразу, состоящую из 363 аминокислот с кажущейся молекулярной массой 37000 единиц Дальтона (37 кДа) на субъединицу по методу SDS-PAGE. Этот фермент возникает в виде димеров с приблизительный молекулярной массой 70 кДа. Для этой ферментативной реакции требуется молекулярный кислород, NAD(P)H- ферредоксиноксидоредуктаза и ферредоксин.
В предыдущих исследованиях было показано, что для каталитической активности как растворимых, так и связанных с мембраной Δ9-десатураз необходимо негемное железо. Сравнительно недавно при спектроскопическом анализе и сопоставлении аминокислотной последовательности было установлено, что Δ9-десатураза содержит кластер из двух атомов железа. Этот класс белков с двумя атомами железа отличается двумя местонахождениями мотива последовательности Е-Х-Х-Н, которые разделены приблизительно ста аминокислотами, и включают R2 субъединицу рибонуклеотидной редуктазы и гидроксилазный компонент монооксигеназы метана. Лучшее понимание механизма катализа ферментами ацил-АСР-десатуразы может дать возможность применения таких ферментов в производстве масла из семян растений.
Краткое описание изобретения
Настоящее изобретение относится к способу модифицирования длины цепи и специфичности расположения двойных связей растворимой десатуразы растительных жирных кислот. Более конкретно, этот способ включает модифицирование аминокислотных контактных остатков в канале связывания субстрата в растворимой десатуразе жирных кислот, которая контактирует с жирными кислотами. В предпочтительных вариантах воплощения растворимая десатураза растительных жирных кислот представляет собой ацил-АСР-десатуразу.
Аминокислотные контактные остатки, которые находятся внутри канала связывания субстрата, идентифицируют, например сначала предоставляя главную последовательность аминокислот ацил-АСР-десатуразы. Известны многие такие последовательности, а другие могут быть определены с использованием обычного экспериментирования. Затем такие аминокислотные последовательности выравнивают с главной аминокислотной последовательностью Δ9-десатуразы Ricinus communis для максимального сохранения последовательности. Затем трехмерную модель ацил-АСР-десатуразы конструируют на основе последовательности, консервативной с последовательностью Δ9-десатуразы Ricinus communis. Потом идентифицируют аминокислотные контактные остатки внутри канала связывания субстрата моделированной структуры. Затем путем замещения одного или нескольких аминокислотных контактных остатков на другой аминокислотный остаток генерируют мутантную ацил-АСР-десатуразу, имеющую модифицированную длину цепи и особенности расположения двойных связей.
В другом аспекте настоящее изобретение относится к мутантной ацил-АСР-десатуразе, которая характеризуется способностью катализировать образование двойной связи в первой жирной кислоте и во второй жирной кислоте, причем первая и вторая жирные кислоты отличаются длиной цепи. Этот мутант дополнительно характеризуется способностью к образованию двойных связей в первой жирной кислоте и во второй жирной кислоте со скоростями, которые различаются не более чем приблизительно в 4 раза.
Это изобретение также относится к композициям, таким как последовательности нуклеиновых кислот и векторы экспрессии, которые кодируют мутантную ацил-АСР-десатуразу описанного выше типа. Другие композиции настоящего изобретения включают клетки, трансформированные такими векторами экспрессии. В другом аспекте настоящее изобретение относится к химерным ацил-АСР-десатуразам, имеющим модифицированную длину цепи и особенности расположения двойных связей.
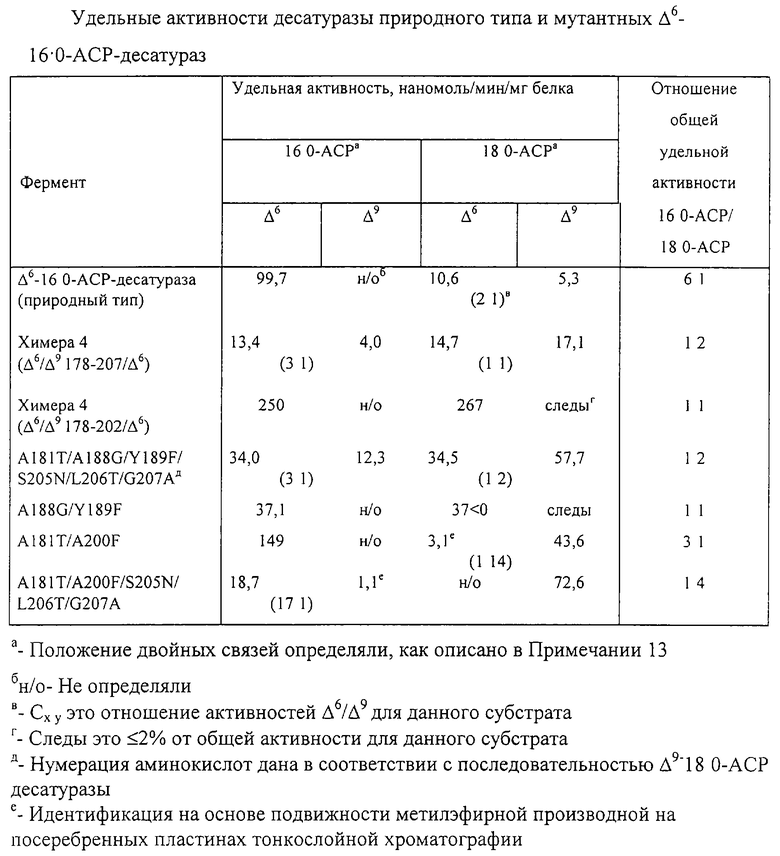
В таблице обобщены значения удельных активностей десатураз природного типа и мутантных ацил-АСР-десатураз.
Подробное описание изобретения
Настоящее изобретение частично основано на идентификации желобка, связывающего субстрат и критических контактных остатков, которые устанавливают впадину в ацил-АСР-десатуразе. Кроме того, это изобретение включает анализ влияния логически обоснованных, сайт-направленных мутаций. До этого изобретения были неизвестны относительное местоположение каналов, связывающих субстрат в ацил-АСР-десатуразах, и критические остатки, контактирующие с субстратом.
Ацил-АСР-десатуразы являются весьма консервативными, причем найдено более 70% гомологичности аминокислотной последовательности между ферментами различных семейств, такими как Δ4, Δ6 и Δ9-ацил-АСР-десатуразы. Известно, что каждая из этих десатураз катализирует образование двойных связей между атомами углерода того же самого или аналогичного субстрата - жирных кислот. Основное отличие между активностью различных ацил-АСР-десатураз заключается в расположении атомов углерода внутри субстратных жирных кислот, которые станут ненасыщенными.
Сохранение аминокислотной последовательности является даже более значительньм в конкретном семействе ацил-АСР-десатураз, таком как Δ9. На основании настоящего описания специалист в этой области химии сможет предсказать, какие контактные остатки внутри канала, связывающего субстрат, из всех ферментов Δ9-ацил-АСР-десатураз являются практически подобными, если не идентичными тем, что идентифицированы в Δ9-ацил-АСР-десатуразе, описанной ниже в Примере 1.
Высокая степень гомологичности аминокислотной последовательности между семействами ацил-АСР-десатураз, которые катализируют те же самые ферментативные реакции, и гомологичности аминокислотной последовательности между семействами ацил-АСР-десатураз, которые катализируют другие ферментативные реакции, позволяет предположить, что определенные части ферментов могут обладать аналогичными третичными структурами. Это согласуется с наблюдениями для других молекул, таких как антитела, в которых сохранение гомологичности аминокислотного остатка обычно является наибольшим для тех аминокислот, которые вовлечены в поддержание функциональной структуры рассматриваемой молекулы.
Одной такой областью структуры в ацил-АСР-десатуразах, которая сохраняется между различньми ацил-АСР-десатуразами, является канал, связывающий субстрат, описанный в последующем разделе Примеров. Описанный ниже канал, связывающий субстрат, обладает архитектурой, которая обеспечивает почти превосходную аккомодацию для субстрата - жирной кислоты. Подобное совершенное соответствие является весьма необычным, если не является беспрецедентным.
Тот факт, что этот канал, связывающий субстрат, является высоко консервативным, может быть подтвержден посредством выравнивания по максимальной идентичности (по общепринятым методикам) аминокислотных последовательностей представителей других семейств ацил-АСР-десатураз с аминокислотной последовательностью касторовой (то есть Ricinus communis) Δ9-ацил-АСР-десатуразы, описанной ниже в Примере 1. Выведенная аминокислотная последовательность этого касторового белка была опубликована Шанклином и Сомервиллем (в Proc. Natl. Acad. Sci. USA, т. 88, с.2510 (1991)). В соответствии с этим выравниванием может быть создана трехмерная модель, в которой будет выявлен характерный канал, связывающий субстрат. Среди установленных к настоящему времени последовательностей ацил-АСР-десатуразы из различных растительных источников доступными из Генетического банка являются следующие (в квадратных скобках указаны коды доступа): [BRSACP] В. rapa; [CAHSACPD] С. tinctorius; [SMMSACPD] Simmondsia chinensis; [SOACCPDS] S. oleracea; [SSMSACPD] растительный источник - кунжут; [TAU07597] Thunbergia alata (клон pTAD2 Δ9); [TAU07605] Thunbergia alata (клон pTAD3 Δ9); [ATSTACPDS] A. thaliana; [BNAACPDES] B. napus; [BNSACPD] B. napus; [GHSACPDES] G.hinsutum; [LUSACPDE] L. usitatissimum; [RCSTEA] R. communis; [SOYSACPD] Glycine max; [SSMSACPDA] растение - кунжут; [SSMSACPDB] растение - кунжут; [TAU07552] Thunbergia alata (клон pTAD1 Δ9).
При изучении результатов молекулярного моделирования для любой из ацил-АСР-десатураз, что проведено ниже в связи с касторовой Δ9-ацил-АСР-десатуразой, могут быть идентифицированы аминокислотные остатки внутри канала, связывающего субстрат, которые ориентированы таким образом, что могут находиться в непосредственной близости к субстрату - жирной кислоте. Такие остатки называются "контактными остатками". Как выявлено ниже в результате описания экспериментального исследования, модификация контактного остатка (а в некоторых случаях - других остатков, как, например, химерных из Примера 2) может изменять длину цепи и особенности расположения двойных связей ацил-АСР-десатуразы.
Например, как показано ниже в Примере 2, был получен химерный фермент, в котором аминокислоты 172-202 в Δ6-16:0-АСР-десатуразе были заменены аминокислотами 178-207 Δ9-18:0-АСР-десатуразы. Это приводит к введению 9 новых аминокислот в канал, связывающий субстрат, Δ6-16:0-АСР-десатуразы, которые отличаются от аминокислот в соответствующих положениях в Δ6-16:0-АСР-десатуразе природного типа. Эта химера не только была способна образовать двойную связь в жирной кислоте 16:0, которая лучше всего превращается на ферменте природного типа, но также способна образовать двойную связь в кислоте 18:0 в обоих положениях, Δ6 и Δ9, и в равной степени.
Тот факт, что аминокислотные контактные остатки в канале ацил-АСР-десатуразы, связывающем субстрат, играют такую важную роль при определении длины цепи и позиционной специфичности двойных связей, обеспечивает возможность для разумного конструирования мутантных ацил-АСР-десатураз, которые обладают уникальными и полезными свойствами.
Такие новые мутантные молекулы могут быть созданы, например, путем первоначальной идентификации контактных остатков внутри канала, связывающего субстрат (как описано выше, посредством выравнивания по касторовой Δ9-аминокислотной последовательности с последующим трехмерным моделированием). Затем в рассматриваемую молекулу ацил-АСР-десатуразы могут быть введены специфические точечные мутации. Наиболее удобно это выполняется на генетическом уровне с использованием общепринятых методик, таких как сайт-направленный мутагенез.
Для введения специфического изменения (то есть замещения) аминокислотного кодона внутри таких ДНК можно использовать множество методик сайт-направленного мутагенеза. Необходимо проявлять осторожность в выборе остатка, подлежащего введению вместо существующего контактного остатка в канале, связывающем субстрат, в ацил-АСР-десатуразе природного типа. Это особенно важно в начале исследования, например, для выбора замещающих остатков, которые существенным образом не отличаются по размеру боковой цепи или заряду. Например, если внутри канала, связывающего субстрат, идентифицируется глициновый контактный остаток (отличается компактностью собственной алифатической цепи), то замещение его таким аминокислотным остатком, как аргинин (отличается наличием объемной основной боковой цепи), может привести к блокированию поступления субстратной жирной кислоты в канал, связывающий субстрат, за счет стерических затруднений. В общем, начальные замещения аминокислот в контактных остатках должны быть произведены с использованием аминокислот, имеющих аналогичные зарядовые характеристики при относительно небольших различиях в значениях объема боковой цепи. Однако при оговоренных ограничениях вполне возможно, что при замещении аминокислотой, имеющей совершенно другие свойства, чем контактный остаток природного типа, можно получить особенно эффективную мутантную ацил-АСР-десатуразу. Такая молекула будет охвачена настоящим изобретением. Приведенное выше краткое обсуждение стратегии замещения предназначается только для того, чтобы служить руководством для последовательного модифицирования ацил-АСР-десатуразы.
Таким образом, именно знание идентичности контактных остатков в пределах ацил-АСР-десатуразы позволяет специалисту в этой области химии осуществить такие модификации фермента, которые могут изменить длину цепи и особенности расположения двойных связей фермента без ингибирования его способности вести процесс ферментативного катализа. В свою очередь, эти знания зависят от способности специалиста в этой области химии идентифицировать канал, связывающий субстрат, и создать трехмерную модель.
Как уже обсуждалось, описаны нуклеотидные последовательности многих ацил-АСР-десатураз. Более того, приведены их высокие степени консервативности, рутинные эксперименты по гибридизации нуклеиновых кислот, проведенные с использованием ДНК, выделенной из рассматриваемых растений, которые, вероятно, могли бы дать ДНК, кодирующую дополнительные ацил-АСР-десатуразы.
Кроме того, как указано выше, специалист в этой области химии сможет предвидеть, что в пределах семейства Δ9-ацил-АСР-десатуразы аминокислотные контактные остатки внутри желобка, связывающего субстрат, могут быть практически подобными, если не идентичными. Аминокислотные контактные остатки, идентифицированные методом рентгеновской кристаллографии, как описано в Примере 1, являются остатками M114, L115, Т117, L118, Р179, Т181, G188 и F189. То, что модификация этих остатков в Δ9-ацил-АСР-десатуразе действительно изменяет длину цепи и особенности расположения двойных связей фермента, подтверждено в экспериментах, описанных в Примере 2. Более конкретно, в предварительном экспериментальном исследовании было выявлено, что единственное замещение аминокислоты в положении 118 (лейцин на фенилаланин) в касторовой Δ9-ацил-АСР-десатуразе приводит приблизительно к десятикратному увеличению ее активности с 16:0-АСР. Таким образом, одно аминокислотное замещение в положении контактного остатка может создать ацил-АСР-десатуразу с новыми и полезными свойствами.
До этого изобретения единственным источником вариантных ацил-АСР-десатураз была ткань растений, которые синтезируют необычные изомеры моно-ненасыщенных жирных кислот. Например, Δ4-16:0-АСР-десатуразу получали из семенного эндосперма кориандра - ткани, которая продуцирует большие количества петроселиновой кислоты (18:1Δ6), необычной ненасыщенной жирной кислоты. Настоящее изобретение обеспечивает конструирование и производство новых типов ацил-АСР-десатураз без необходимости выделения кДНК для этих ферментов из растительных источников. Кроме того, настоящее изобретение обеспечивает конструирование ацил-АСР-десатураз, которые могут катализировать синтез экономически ценных мононенасыщенных жирных кислот, которые не встречаются в природе.
В более конкретном примере это изобретение предоставляет альтернативное средство для получения петроселиновой кислоты в растениях. Эта жирная кислота имеет ряд возможных промышленных и пищевых областей применения. Единственный известный путь образования петроселиновой кислоты в растениях включает введение Δ4-нeнacыщeннocти 16:0-АСР с последующим удлинением образовавшейся 16:1Δ4-АСР, чтобы получить 18:1Δ6 (или петроселиноил)-АСР. Для этого пути требуется, наряду с другими средствами, новая ацил-АСР-десатураза и специфическая система удлинения ацил-АСР. Среди описанных ниже мутантных десатураз имеются ферменты, которые могут катализировать введение Δ6 ненасыщенности 18:0-АСР, чтобы образовать петроселиноил-АСР. Такие ферменты полезны для получения петроселиновой кислоты в трансгенных культурных растениях без необходимости в передаче дополнительных генов для пути удлинения 16:1Δ4-ACP. Этот путь представляет собой в настоящее время ограничение возможностей получения петроселиновой кислоты в трансгенных культурных растениях путем введения гена для Δ4-16:0-АСР-десатуразы.
Таким образом, мутанты, созданные путем изменения идентичности одного или нескольких контактных остатков в канале, связывающем субстрат, могут быть использованы для создания ацил-АСР-десатураз, имеющих уникальные функциональные характеристики. Такие ферменты могут быть использованы, например, для создания растительных масел, обогащенных моно-ненасыщенными жирными кислотами. Такие растительные масла важны для питания людей и могут быть использованы в качестве возобновляемых источников промышленных химических реагентов. Кроме того, возможность манипулирования предпочтениями длины цепи и положениями двойных связей этих молекул обеспечивает способ управления физическими свойствами и промышленное применение традиционных растительных масел. К тому же, на основе этого описания можно эксплуатировать разработку трансгенных культур, способных продуцировать необычные типы моно-ненасыщенных жирных кислот.
Мутанты, описанные ниже в Примере 2, проявляют определенно уникальные свойства. Например, у ацил-АСР-десатураз природного типа имеется тенденция к проявлению очень сильного предпочтения к жирным кислотам с определенной длиной цепи и расположением связей. Однако в описанных ниже опытах было показано, что аминокислотные замещения в контактных остатках внутри канала, связывающего субстрат, модифицируют это предпочтение. Например, описаны химерные мутанты, которые обладают способностью катализировать образование ненасыщенности в субстратах различной длины (например 16:0 и 18:0) со скоростями, различающимися не более чем в 4 раза.
Последовательности нуклеиновых кислот, кодирующие эти мутантные ацил-АСР-десатуразы, могут быть использованы для экспрессии мутантного фермента с использованием методик рекомбинантной ДНК. Например, при клонировании в подходящем векторе экспрессии мутантная ацил-АСР-десатураза может быть подвергнута экспрессии в разнообразных типах клеток, включающих, например, прокариотические и эукариотические клетки.
Векторы прокариотической экспрессии применяются, например, для приготовления больших количеств белка, кодированного рассматриваемой последовательностью ДНК. После очистки традиционными методами этот белок может быть использован для введения ненасыщенности в жирные кислоты. Кроме того, в некоторых областях применения можно использовать неочищенный продукт лизиса такой культуры прокариотических клеток.
Векторы эукариотической экспрессии используются тогда, когда важным является добавление углеводных боковых цепей (то есть гликозилирование) к белку. Боковые углеводные цепи влияют на активность белка различным образом. Например известно, что некоторые белки являются неактивными в негликозилированном состоянии. Кроме того, способность негликозилированного белка образовывать комплекс с другими белками (например, антителами или регуляторными молекулами) может быть сильно подавлена в отсутствие гликозилирования. После очистки традиционными методами мутант ацил-АСР-десатуразы, экспрессированный в эукариотической системе (например, в системе экспрессии бакуловируса), может быть использован для модифицирования длины цепи и положения двойной связи в жирной кислоте. Во многих случаях этот белок также можно использовать как часть неочищенного продукта лизиса.
Мутантные ацил-АСР-десатуразы также можно клонировать в векторе экспрессии растения. Эти векторы позволяют получить целевой белковый продукт, например мутантную ацил-АСР-десатуразу, внутри среды растительной клетки, в пределах которой остается субстратная жирная кислота. При получении фермента in situ модификация продукта может происходить до сбора клеток, что обеспечивает быструю очистку целевой жирной кислоты, имеющей подходящее расположение двойной связи, и без необходимости в дорогих стадиях производства. В некоторых случаях, в конкретном трансгенном растении, целевыми могут быть несколько мутантных ацил-АСР-десатураз, для того чтобы получить жирные кислоты, имеющие двойные связи в нескольких положениях. Растения также можно легко культивировать и выращивать в больших количествах. Во многих случаях этот белок также можно использовать как часть неочищенного продукта лизиса.
Пояснение примерами
Пример 1
Результаты и их обсуждение
Карта электронной плотности и качество модели
Трехмерную структуру рекомбинантной гомодимерной Δ9-стеароил-ацилнесущей белковой десатуразы, архетип растворимых десатураз растительных жирных кислот, которые превращают насыщенные жирные кислоты в ненасыщенные, определяют методами кристаллографии белка с разрешением 2,4 Ангстрема ( ). Шестикратно усредненная плотность электронов для основной цепи и боковых цепей хорошо разрешается для большей части полипептидной цепи. Исключениями являются первые 18 остатков на N-конце цепи, которые не разрешаются по электронной плотности и, возможно, изгибаются в кристаллической решетке. Остатки 336-347, расположенные в области петли, довольно плохо разрешены в картах электронной плотности, и также именно в этой части белковой структуры были найдены наибольшие отклонения от некристаллографической симметрии. Суммарный реальный фактор пространственной корреляции по остаткам (субъединица Вr и карта шестикратно усредненной плотности электронов 2Fo-Fc) равен 0,76. Такие критерии, как кристаллографический R-фактор (R=22,0%, Rсвоб.=28,5% с ограничениями некристаллографической симметрии), хорошая стереохимия модели (среднеквадратичная ошибка длины связи 0,008
). Шестикратно усредненная плотность электронов для основной цепи и боковых цепей хорошо разрешается для большей части полипептидной цепи. Исключениями являются первые 18 остатков на N-конце цепи, которые не разрешаются по электронной плотности и, возможно, изгибаются в кристаллической решетке. Остатки 336-347, расположенные в области петли, довольно плохо разрешены в картах электронной плотности, и также именно в этой части белковой структуры были найдены наибольшие отклонения от некристаллографической симметрии. Суммарный реальный фактор пространственной корреляции по остаткам (субъединица Вr и карта шестикратно усредненной плотности электронов 2Fo-Fc) равен 0,76. Такие критерии, как кристаллографический R-фактор (R=22,0%, Rсвоб.=28,5% с ограничениями некристаллографической симметрии), хорошая стереохимия модели (среднеквадратичная ошибка длины связи 0,008 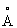 ), график Рамачандрана (только одна посторонняя область из возможных областей в субъединице, за исключением глициновых остатков) и наблюдаемый характер водородных связей, - все это указывает, что прослеживание цепи для десатуразы жирных кислот является верным. Существует очень ясная картина для пептидного атома кислорода лизина 262, остатка с неразрешенной конформацией основной цепи. Высокое среднее значение бета-фактора позволяет предположить, что эта молекула является гибкой. Наиболее упорядоченные части молекулы представляют собой области, вовлеченные во взаимодействие димера и гексамера, в то время как поверхностные петли часто имеют очень высокие значения бета-фактора.
), график Рамачандрана (только одна посторонняя область из возможных областей в субъединице, за исключением глициновых остатков) и наблюдаемый характер водородных связей, - все это указывает, что прослеживание цепи для десатуразы жирных кислот является верным. Существует очень ясная картина для пептидного атома кислорода лизина 262, остатка с неразрешенной конформацией основной цепи. Высокое среднее значение бета-фактора позволяет предположить, что эта молекула является гибкой. Наиболее упорядоченные части молекулы представляют собой области, вовлеченные во взаимодействие димера и гексамера, в то время как поверхностные петли часто имеют очень высокие значения бета-фактора.
Найдено, что главные связывающие центры для ионов дицианида золота [Au(CN)2] в кристаллах дериватизированной десатуразы близки к боковым цепям К56 и С61 на поверхности молекулы. Одним из второстепенных центров является внутренний центр, между боковыми цепями Н203 и С222, причем второй второстепенный центр находится в области, в которой вероятно расположен N-конец цепи.
Общая форма субъединицы Δ9-десатуразы представляет собой компактный цилиндр с размерами 35×35×50 и с доступной площадью поверхности 16773. Кроме петли визирной оси на самом С-конце цепи эта субъединица в основном состоит из спиральных вторичных структур, уложенных внутри одного большого домена. Девять из общих одиннадцати α-спиралей образуют антипараллельный пучок спиралей.
Часть цепи при N-конце разупорядочена, причем для первых 18 остатков не наблюдается картина электронной плотности. В следующих 15 остатках отсутствует вторичная структура, и они образуют протяженную цепь, упакованную вдоль спирального пучка со слабыми специфическими взаимодействиями, которые стабилизируют эту структуру. Первая спираль α1, состоящая из 23 остатков, начинается и заканчивается в конфигурации-310 и является настолько гибкой, что ее первая половина образует колпачок на одном конце пучка, а ее вторая часть представляет собой первую спираль пучка. Цепочка продолжается в том же направлении, образуя витки, связанные водородной связью, и спираль-310. Колпачок на другом конце пучка образуется из спиралей α2 и αΔ9 и С-терминальной визирной оси. Четыре из этих спиралей, α3, α4, α6 и α7, которые являются весьма длинными и имеют соответственно 28, 29, 30 и 31 остатков, предоставляют лиганды для центра из двух атомов железа. Хотя α3 имеет излом в спиральной структуре в середине остатка 107-108, спирали α3 и α4 являются симметричными по отношению к α6 и α7 и могут быть совмещены со среднеквадратичной ошибкой 1,39 А для 44 остатков. При таком совмещении также выравниваются атомы железа в пределах 1 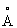 . Выравнивание соответствующих последовательностей показывает, что имеется незначительное сохранение последовательностей, кроме остатков, вовлеченных в связывание с кластером железа. Это совмещение также ориентирует часть колпачка α1 на α2. Связи между этими спиралями также приблизительно совмещаются, хотя и отсутствует детальное структурное подобие. Между α3b и α4 существует выступающая петлевая структура, стабилизированная несколькими витками с водородными связями. Пучок завершается спиралями α5, α8, α10 и α11, которые весьма искривлены.
. Выравнивание соответствующих последовательностей показывает, что имеется незначительное сохранение последовательностей, кроме остатков, вовлеченных в связывание с кластером железа. Это совмещение также ориентирует часть колпачка α1 на α2. Связи между этими спиралями также приблизительно совмещаются, хотя и отсутствует детальное структурное подобие. Между α3b и α4 существует выступающая петлевая структура, стабилизированная несколькими витками с водородными связями. Пучок завершается спиралями α5, α8, α10 и α11, которые весьма искривлены.
Имеется большое число солевых мостиков, 26, за исключением тех, что взаимодействуют с ионами железа в пределах субъединицы. Это соответствует 0,069 ионной пары на 1 остаток, что выше среднего числа ионных пар на остаток (0,04), выведенного из обзора 38 белковых структур высокого разрешения. Семь таких солевых мостиков в Δ9-десатуразе вовлекаются во взаимодействия между спиралями в пределах пучка, обеспечивая правильную взаимную упаковку и в некоторых случаях правильную ориентацию лигандов железа. Восемь пар осуществляют контакты между спиралями, а три остальные вовлечены в маятниковое связывание витков между спиралями пучка. Три пары вовлечены в контакты со спиралью-310 и петлей после α8.
Специфической чертой субъединицы Δ9-десатуразы является довольно плоская поверхность, которую образуют спирали α1, α6, α7, α10 и α11. Эта поверхность не вовлечена в контакты между субъединицами в димере, но доступна из раствора.
Димер
Поверхность раздела между субъединицами в димере занимает площадь поверхности 5826 А2 на димер, то есть 17,4% поверхности димера. Эти двукратные взаимодействия включают обширные контакты между спиралями в пучке; от α3b до той же спирали во второй субъединице, между α4 и α5, и поверх α2 и α4 до соответствующих спиралей в другой субъединице. Также имеется множество контактов между выступающей петлей между α3b и α4 и N-концом α3b и α5. Кроме того, остатки в связке между α1 и α2 осуществляют контакты с α4 и α5 во второй субъединице. Существуют три зарядовых взаимодействия в контактной области димера, в два из которых вовлечены остатки из α5. Центры из двух атомов железа изолированы в димере на расстоянии больше чем 23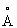 , и между ними отсутствуют прямые контакты.
, и между ними отсутствуют прямые контакты.
Некристаллографическая симметрия и кристаллическая упаковка
Кристаллическая асимметричная ячейка содержит три димера Δ9-десатуразы. В этих димерах субединицы соотносятся по некристаллографическим осям второго порядка, которые для одного из димеров параллельны одной из кристаллографических осей второго порядка. Под прямым углом к ней, параллельно "а", имеется некристаллографическая винтовая ось третьего порядка, относящаяся к трем димерам. Трансляция составляет одну треть длины "а", то есть она является локальной осью 31. Контакты между димерами не являются обширными и имеют тот же порядок величины, что и другие контакты кристалла, и, по-видимому, влияние контактов кристалла на его структуру является незначительным, что подтверждается наблюдаемыми малыми отклонениями от некристаллографической симметрии. Самые большие отклонения получены для остатков 336-347, где R336, Е347 и/или К346 осуществляют контакты кристалла, включая солевые мостики, в некоторых субъединицах. Как отмечалось выше, в этой области электронная плотность мала. Другая область с отклонениями от некристаллографической симметрии включает остатки от 19 до 50, которые намотаны вокруг субъединицы и также осуществляют различные контакты свободных кристаллов в субъединице. Упаковка субъединиц, соответствующих одной асимметричной ячейке, рассматривается вдоль оси третьего порядка и одной из осей второго порядка.
Центр из двух атомов железа
В предыдущих исследованиях было показано, что Δ9 -десатураза содержит четыре атома железа в одном димере, причем методами оптической и Мессбауровской спектроскопии было установлено, что эти ионы железа составляют кластеры оксо-дижелеза. Теперь эти кластеры оксо-дижелеза обнаружены в большом множестве белков, которые осуществляют как каталитические, так и некаталитические функции. Они содержат два атома железа, связанных мостиковыми либо оксо-, либо гидроксилигандами, причем их классифицируют на основе различных мотивов первичных последовательностей, предоставляющих кластерные лиганды, и на основе структурных различий, выявленных методом рентгеновской кристаллографии. Были описаны четыре класса: один содержит гемэритрин и миогемэритрин, второй содержит субъединицу R2 рибонуклеотидной редуктазы, бактериальные углеводородные гидроксилазы и Δ9-десатуразу, третий содержит рубрэритрин и четвертый содержит Fe(III)-Zn(II) фосфатазу пурпуровой кислоты (Fe(III)-Zn(II) кислую фосфатазу Strmammalian). Кроме этих растворимых белков существует отдельный класс функционально родственных интегральных мембранных белков, включающих десатуразы жирных кислот и углеводородные гидроксилазы, которые содержат центры негемного железа, активированного кислородом, которые еще должны быть структурно охарактеризованы.
Из кристаллической структуры Δ9-десатуразы следует, что этот фермент принадлежит к классу II белков с двумя атомами железа и что он содержит металлический кластер. Расстояние между ионами железа составляет 4,2 , а координационной геометрией этих ионов железа является искаженный октаэдр, в котором одна позиция лиганда является незанятой. Структура кластера обладает высокой симметрией. Роль мостиковых лигандов выполняют оба остатка Е143 из спирали α4 и Е229 из α7. Е 105 из α3а является бидентатным лигандом для одного иона железа и соответственно Е196 из α6 является бидентатным лигандом для второго иона железа. Каждый ион железа также связывается атомом азота, Nδ1 в H146 из α4 и Н232 из α7 соответственно. Ориентация лигандов железа в некоторых случаях поддерживается водородными связями с боковыми цепями; Е105 взаимодействует с Н203, Е143 с атомом N∈1 в W139, N∈2 в Н146 с боковой цепочкой D228, которая, в свою очередь, взаимодействует с боковыми цепями R145 и W62, N∈2 в Н232 с боковой цепочкой Е143, которая, в свою очередь, взаимодействует с боковой цепочкой R231. Еще дальше от одного из ионов железа обнаружен атом N∈1 в W139, который можно рассматривать как лиганд второй оболочки. Вблизи от кластера железа электронная плотность соответствует молекуле растворителя. Она находится на расстоянии от ионов железа соответственно 3,2 и 3,4
, а координационной геометрией этих ионов железа является искаженный октаэдр, в котором одна позиция лиганда является незанятой. Структура кластера обладает высокой симметрией. Роль мостиковых лигандов выполняют оба остатка Е143 из спирали α4 и Е229 из α7. Е 105 из α3а является бидентатным лигандом для одного иона железа и соответственно Е196 из α6 является бидентатным лигандом для второго иона железа. Каждый ион железа также связывается атомом азота, Nδ1 в H146 из α4 и Н232 из α7 соответственно. Ориентация лигандов железа в некоторых случаях поддерживается водородными связями с боковыми цепями; Е105 взаимодействует с Н203, Е143 с атомом N∈1 в W139, N∈2 в Н146 с боковой цепочкой D228, которая, в свою очередь, взаимодействует с боковыми цепями R145 и W62, N∈2 в Н232 с боковой цепочкой Е143, которая, в свою очередь, взаимодействует с боковой цепочкой R231. Еще дальше от одного из ионов железа обнаружен атом N∈1 в W139, который можно рассматривать как лиганд второй оболочки. Вблизи от кластера железа электронная плотность соответствует молекуле растворителя. Она находится на расстоянии от ионов железа соответственно 3,2 и 3,4  , и поэтому она не является частью первой координационной оболочки кластерного центра.
, и поэтому она не является частью первой координационной оболочки кластерного центра.
Форма десатуразы в кристаллической структуре
Наличие в Δ9-десатуразе кластера из двух атомов железа, связанных μ-оксо-мостиком в состоянии дижелезо(III), было однозначно продемонстрировано с использованием Мессбауэровской и Рамановской резонансной спектроскопии. Поэтому странно, что μ-оксо-мостик не был обнаружен на карте электронной плотности Δ9-десатуразы, поскольку использованный в экспериментах фермент находился в окисленном состоянии и в маточный раствор не добавляли восстанавливающие агенты. Кроме того, расстояние между ионами железа (4,2 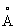 ) больше ожидаемого для кластера из двух атомов железа с неповрежденным μ-оксо-мостиком. В окисленной форме рибонуклеотидной редуктазы, содержащей μ-оксо-мостик, расстояние железо-железо составляет 3,3
) больше ожидаемого для кластера из двух атомов железа с неповрежденным μ-оксо-мостиком. В окисленной форме рибонуклеотидной редуктазы, содержащей μ-оксо-мостик, расстояние железо-железо составляет 3,3  . Наблюдаемая для Δ9-десатуразы геометрия поразительно аналогична той, что просматривается в восстановленной форме рибонуклеотидной редуктазы, в которой при химическом восстановлении R2 расстояние между ионами железа возрастает до 3,8 А, μ-оксо-мостик удаляется, и лигандное окружение становится весьма симметричньм, что видно из кристаллографии белка и Мессбауэровской спектроскопии. Можно предположить, что при воздействии рентгеновского излучения на десатуразу происходит фотохимическое восстановление металлического центра, которое сопровождается потерей μ-оксо-мостика и перестройкой лигандов. Таким образом, представленная здесь структура Δ9-десатуразы наиболее вероятно отражает восстановленную форму фермента. В кристаллической структуре Δ9-десатуразы обнаружено высокосимметричное лигандное окружение железного кластера в дижелезной форме фермента в соответствии с исследованиями резонансной Рамановской спектроскопии. Отклонения в симметрии лигандного окружения в восстановленном состоянии фермента, как было предположено ранее из температурной зависимости данных Мессбауэровской спектроскопии, возможно, обусловлены изменениями длины и углов связей в двух металлических центрах, которые слишком малы, чтобы их можно было наблюдать в картах электронной плотности при существующем разрешении.
. Наблюдаемая для Δ9-десатуразы геометрия поразительно аналогична той, что просматривается в восстановленной форме рибонуклеотидной редуктазы, в которой при химическом восстановлении R2 расстояние между ионами железа возрастает до 3,8 А, μ-оксо-мостик удаляется, и лигандное окружение становится весьма симметричньм, что видно из кристаллографии белка и Мессбауэровской спектроскопии. Можно предположить, что при воздействии рентгеновского излучения на десатуразу происходит фотохимическое восстановление металлического центра, которое сопровождается потерей μ-оксо-мостика и перестройкой лигандов. Таким образом, представленная здесь структура Δ9-десатуразы наиболее вероятно отражает восстановленную форму фермента. В кристаллической структуре Δ9-десатуразы обнаружено высокосимметричное лигандное окружение железного кластера в дижелезной форме фермента в соответствии с исследованиями резонансной Рамановской спектроскопии. Отклонения в симметрии лигандного окружения в восстановленном состоянии фермента, как было предположено ранее из температурной зависимости данных Мессбауэровской спектроскопии, возможно, обусловлены изменениями длины и углов связей в двух металлических центрах, которые слишком малы, чтобы их можно было наблюдать в картах электронной плотности при существующем разрешении.
Активный центр и взаимодействия с другими белками
Описанная здесь структура Δ9-десатуразы весьма вероятно представляет собой структуру дижелезной формы фермента, которая является результатом взаимодействия Δ9-дecaтypaзы и ферредоксина в растении. Из трехмерной структуры можно постулировать два возможных пути переноса электронов от поверхности к железному центру. Один путь простирается вдоль оси пучка спиралей и включает структурно последовательные кластеры ароматических боковых цепей W139, W135, Y236, F189, W132. Атом N∈1 в W139 находится на довольно близком расстоянии к одному из атомов железа, а атом Ne1 в W132 направлен к поверхности субъединицы, близкой к выступающей петле между α3b и α4. Эта петля и колпачковая часть α1 затем могут образовать поверхность для взаимодействия молекулы ферредоксина. Другой возможный путь переноса электронов от поверхности к железному центру включает остатки W62, D228 и H146, аналогичные тем, что предложены для R2. Этот путь приводит к образованию плоской поверхности, образованной спиралями α1, α6, α7, α10, α11.
Молекула растворителя, связанная вблизи железного центра, расположена в небольшом гидрофобном кармане, и наиболее близкими аминокислотными боковыми цепями к этой молекуле растворителя являются Т199 и W139. Аналогичная полость с треониновой боковой цепью в эквивалентном положении была обнаружена в ММО, и было предположено, что эта полость может обеспечить подходящее место для связывания молекулы кислорода. Подразумевается, что боковая цепь Т213 в МАО вовлечена в активацию кислорода таким же образом, как остаток Т252 в цитохроме Р450.
Поскольку железный центр погружен во внутреннюю часть Δ9-десатуразы, можно ожидать, что будет идентифицирована впадина для субстрата, облицованная гидрофобными остатками, соединяющая поверхность фермента с активным центром. Действительно, можно обнаружить узкий, очень глубокий канал, простирающийся от поверхности далеко внутрь белка. Этот канал подходит к дижелезному центру с той же самой стороны, что и предполагаемое место связывания кислорода. На дне этого канала обнаружена боковая цепь L115 и стенки, состоящие из остатков W139, Т192, Y111, M114, Y191, Q195, P266, T99 и Т104. Затем этот канал проходит железный кластер и продолжается по направлению к поверхности, причем в узком входе в эту впадину находятся остатки Y292, М265, F279 и S283. Общая форма субстратного канала, который изогнут в месте железного кластера, способствует связыванию продукта, олеоил-АСР, имеющего цис-конфигурацию при двойной связи.
После очистки, в карте усредненной плотности электронов 2Fo-Fc в этом канале была обнаружена сильно растянутая электронная плотность, которую нельзя приписать растворителю или атомам белка. На основе формы этой плотности и гидрофобного характера кармана можно сделать вывод, что эта электронная плотность может представлять гидрофобный ацильный конец молекулы бета-октилглюкозида. Углеводородный конец октилглюкозида мог бы хорошо соответствовать этой плотности, но плотность, соответствующая функциональной группе сахара, плохо определена. Эта предполагаемая молекула октилглюкозида ориентирована своим концом глубоко вниз в гидрофобный канал, близко к дижелезному кластеру, а углеводная функция простирается по направлению к поверхности. Низкая электронная плотность в этой части молекулы возможно указывает на локальное разупорядочение, которое является результатом менее специфических взаимодействий с ферментом.
При моделировании стеариновой кислоты в предполагаемом субстрат-связывающем кармане атом углерода С9 изображается на расстоянии примерно 5,5  от одного из ионов железа. Этот атом углерода, при котором будет формироваться двойная связь, также близок к малому карману со связанной молекулой растворителя, действительно, молекула воды является связующим мостиком на расстоянии между атомом С9 субстрата и ближайшим ионом железа. Вероятно, что в активном ферменте этот карман занят молекулой кислорода, связанной с одним или обоими атомами железа. В ходе катализа может образовываться пероксидный радикал, который способен удалять один из атомов водорода в положении С9 жирной кислоты.
от одного из ионов железа. Этот атом углерода, при котором будет формироваться двойная связь, также близок к малому карману со связанной молекулой растворителя, действительно, молекула воды является связующим мостиком на расстоянии между атомом С9 субстрата и ближайшим ионом железа. Вероятно, что в активном ферменте этот карман занят молекулой кислорода, связанной с одним или обоими атомами железа. В ходе катализа может образовываться пероксидный радикал, который способен удалять один из атомов водорода в положении С9 жирной кислоты.
Сопоставление с другими белками с двумя атомами железа
Наложение структуры Δ9-десатуразы на трехмерные структуры двух других белков с двумя атомами железа субъединицы R2 рибонуклеотидной редуктазы из Escherichia coli и α-субъединицы ММО из Methylococcus capsulate показывает, что их общие структуры довольно подобны и согласуются со среднеквадратичной ошибкой 1,90  для 144 Сα-атомов (Δ9-десатураза относительно R2) и 1,98
для 144 Сα-атомов (Δ9-десатураза относительно R2) и 1,98 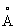 для 117 эквивалентных Сα-атомов (Δ9-десатураза относительно ММО). Весьма подобны их складки, состоящие большей частью из α-спиралей, от α1 до α8 и α10 дают свой вклад в R2 и ММО. Имеется мало сохраненных аминокислот, кроме лигандов железа, однако не может быть сомнений, что эти ферменты являются эволюционно связанными.
для 117 эквивалентных Сα-атомов (Δ9-десатураза относительно ММО). Весьма подобны их складки, состоящие большей частью из α-спиралей, от α1 до α8 и α10 дают свой вклад в R2 и ММО. Имеется мало сохраненных аминокислот, кроме лигандов железа, однако не может быть сомнений, что эти ферменты являются эволюционно связанными.
Существуют значительные различия в структуре железных центров этих трех белков. В общем, металлический центр в Δ9-десатуразе значительно более симметричен, чем в двух других белках. Однако при сопоставлении со структурой восстановленной формы R2 координационная геометрия диядерных центров железа в Δ9-десатуразе и R2 становится более подобной. Наиболее значительное отличие состоит в том, что в Δ9-десатуразе концевые карбоксилаты Е105 и Е196 соответственно действуют как бидентатные лиганды, тогда как в R2 эквивалентные боковые цепи являются монодентатными лигандами для ионов железа.
Среди этих ферментов R2 является уникальным в том смысле, что он образует стабильный радикал в положении Y122. Соответствующий остаток в Δ9-десатуразе представляет собой L150, который расположен в гидрофобном кластере, осуществляя взаимодействия упаковки в четырехспиральном пучке, связывающем кластер железа, причем отсутствуют данные, которые могли бы указать, что этот остаток необходим для каталитической активности.
Имеется мало аминокислотных остатков, которые сохраняются во всех трех ферментах. Среди них имеются лиганды к ионам металла, за исключением Е105, который замещен аспарагиновой кислотой в R2. Единственными другими неизменными остатками являются I225 и D228. Остаток I225 находится вблизи к дижелезному кластеру (ближайшее расстояние 4,6  ) на противоположной стороне субстратного канала. Боковая цепочка упакована между Н203, H146 и W62 в трехмерной структуре, и для более подробного исследования ее функции необходимо ждать результатов изучения локально направленного мутагенеза. Другой неизменный белковый остаток в трех ферментах, D228, является частью пути переноса электрона от двуядерного железного центра к поверхности белка, который предложен для R2. Этот путь в R2 проходит от одного из ионов железа через боковую цепь H118, от D237 до W48, которая расположена на поверхности белка. Эти остатки сохраняются в Δ9-десатуразе, и может быть постулирован аналогичный путь для переноса электрона, включающий структурно эквивалентные остатки H146, D228 и W62, как упоминалось ранее. Более того, для ММО также можно предположить слегка модифицированный путь для переноса электрона. В этом случае сохраняются лиганд железа (H147) и остатки аспарагиновой кислоты (D242), однако структура на поверхности является различной. Тем не менее ароматическая боковая цепочка (Y67) на поверхности находится вблизи от боковой цепи D242.
) на противоположной стороне субстратного канала. Боковая цепочка упакована между Н203, H146 и W62 в трехмерной структуре, и для более подробного исследования ее функции необходимо ждать результатов изучения локально направленного мутагенеза. Другой неизменный белковый остаток в трех ферментах, D228, является частью пути переноса электрона от двуядерного железного центра к поверхности белка, который предложен для R2. Этот путь в R2 проходит от одного из ионов железа через боковую цепь H118, от D237 до W48, которая расположена на поверхности белка. Эти остатки сохраняются в Δ9-десатуразе, и может быть постулирован аналогичный путь для переноса электрона, включающий структурно эквивалентные остатки H146, D228 и W62, как упоминалось ранее. Более того, для ММО также можно предположить слегка модифицированный путь для переноса электрона. В этом случае сохраняются лиганд железа (H147) и остатки аспарагиновой кислоты (D242), однако структура на поверхности является различной. Тем не менее ароматическая боковая цепочка (Y67) на поверхности находится вблизи от боковой цепи D242.
Большая часть других остатков, сохраненных между Δ9-десатуразой и R2, с одной стороны, и Δ9-десатуразой и ММО, с другой стороны, расположены на поверхности белка или вовлечены во взаимодействия упаковки. Консервативные остатки, общие между R2 и Δ9-десатуразой вблизи центра из двух атомов железа, представляют собой остатки W135 и W139. Хотя W135 и W139 строго сохраняются в десатуразах, соответствующие остатки W107 и W111 в R2 не являются строго консервативными. За исключением Т4 и белка Е. coli, W135 замещается фенилаланиновой или тирозиновой боковой цепочкой. Аналогично, W139 замещается глутаминовым остатком.
Материалы и методы
Очистка и кристаллизация фермента
Рекомбинантную Δ9-десатуразу экспрессируют в клетках Е. coli и очищают, как описано ранее (Фокс и др. Biochemistry, 33: 127766 (1993)). Кристаллизацию фермента осуществляют согласно публикации (Шнайдер и др., J. Mol. Biol., 225: 561 (1992)) с небольшими изменениями. Образцы фермента концентрируют до содержания 12-18 мг/мл. Аликвоту белкового раствора (7,5 мл) смешивают с таким же количеством резервуарного раствора, помещают на покровные стекла и при 20°С доводят до равновесия с 1 мл раствора источника. Резервуарный раствор содержит 0,08 моль/л какодилатного буфера с рН 5,4, 200 ммоль/л ацетата магния, 75 ммоль/л сульфата аммония, 2 ммоль/л хлорида лития, 1 ммоль/л хлорида калия, 0,2% бета-октилглюкозида и 12-15% полиэтиленгликоля (ПЭГ) 400 в качестве осадителя. Кристаллы были орторомбическими, пространственная группа P212121 с размерами ячейки а=82,2, b=147,8 и с=198,2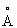 .
.
Сбор данных и приготовление производных тяжелых металлов
Попытки приготовления производных тяжелых металлов путем пропитки природных кристаллов фермента растворами различных солей тяжелых металлов в маточном растворе были не совсем успешными. В большей части опыты с пропиткой приводили к расщеплению кристаллов или к неизоморфным кристаллам, и только одна полезная производная тяжелого металла может быть приготовлена пропиткой кристаллов десатуразы в маточном растворе, содержащем 5 ммоль/л KAu(CN)2, в течение 1 недели. Данные рентгенографии для природных и дериватизированных кристаллов собирали на многожильной зонной детекторной системе UCSD (см. Хэмлин, Methods Enzymol., 114: 416 (1985)) при отделе молекулярной биологии в г. Уппсала (Швеция). Измеренные кадры были обработаны с помощью системы зонного детектора MADNES (Munich area detector NE system, Пфлюграт. Справочник пользователя. Лаборатория фирмы "Cold Spring Harbor", Нью-Йорк, США (1987)). Второй набор данных для природных кристаллов собирали на спектральной линии Х12-С при NSLS, отдел молекулярной биологии в Брукхевенской национальной лаборатории. Кадры данных собирали в виде 10 осцилляции, используя плату изображения MAR Research. Кадры данных были обработаны с помощью DENZO и масштабированы, используя SCALEPACK.
Определение фаз, построение модели и кристаллографическое усовершенствование
Большую часть кристаллографических вычислений осуществляют, используя комплект программ ССР4 (Проект компьютерного сотрудничества, № 4, Acta Crystallogr., D50: 760 (1994)). Исходный кристаллографический анализ проводят с набором данных, собранных на многожильном детекторе при разрешении до 3,1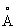 . Разностную карту Паттерсона для производной золота анализируют, используя RSPS (Кнайт, докторская диссертация, Шведский университет сельскохозяйственных наук, Уппсала, 1989). Для расчета разностных карт Фурье были использованы два центра, связанные с сильным перекрестным пиком в разностной карте Паттерсона, и были идентифицированы два новых центра. Окончательно были обнаружены 6 главных центров и 12 второстепенных центров, причем параметры тяжелого металла усовершенствовали, используя MLPHARE. Из результатов расчетов вращательной функции и положений ионов металла можно определить направление и положение операторов локальной симметрии, относящихся к шести субъединицам Δ9-десатуразы в асимметричной ячейке. Затем применяют шестикратное усреднение некристаллографической симметрии с использованием программы RAVE (Джонс, на Выходном семинаре по ССР4, 1992: Молекулярное замещение, под ред. Додсона, Гоувера и Вольфа, с. 91-105, Дэйрсбюрская лаборатория, Дэйрсбюри, Великобритания (1992)) и сферической оболочки с центром в предполагаемом положении одной субъединицы Δ9-десатуразы, для того чтобы уточнить начальные фазы SIR. Из карты электронной плотности при низком разрешении, на основе этих фаз, можно идентифицировать часть центрального четырехспирального пучка, координирующего центр из двух атомов железа и атомы железа. Координаты атомов железа были уточнены из аномальных нативных данных, а новые фазы были рассчитаны на основе производной золота и аномального вклада от атомов железа. Новая оболочка для субъединицы была выполнена с использованием метода МАМА (Клейвегт и Джонс, Acta Cryst. D50: 178 (1994)) посредством приблизительной ориентации субъединицы R2 в правильном положении для пучка спиралей.
. Разностную карту Паттерсона для производной золота анализируют, используя RSPS (Кнайт, докторская диссертация, Шведский университет сельскохозяйственных наук, Уппсала, 1989). Для расчета разностных карт Фурье были использованы два центра, связанные с сильным перекрестным пиком в разностной карте Паттерсона, и были идентифицированы два новых центра. Окончательно были обнаружены 6 главных центров и 12 второстепенных центров, причем параметры тяжелого металла усовершенствовали, используя MLPHARE. Из результатов расчетов вращательной функции и положений ионов металла можно определить направление и положение операторов локальной симметрии, относящихся к шести субъединицам Δ9-десатуразы в асимметричной ячейке. Затем применяют шестикратное усреднение некристаллографической симметрии с использованием программы RAVE (Джонс, на Выходном семинаре по ССР4, 1992: Молекулярное замещение, под ред. Додсона, Гоувера и Вольфа, с. 91-105, Дэйрсбюрская лаборатория, Дэйрсбюри, Великобритания (1992)) и сферической оболочки с центром в предполагаемом положении одной субъединицы Δ9-десатуразы, для того чтобы уточнить начальные фазы SIR. Из карты электронной плотности при низком разрешении, на основе этих фаз, можно идентифицировать часть центрального четырехспирального пучка, координирующего центр из двух атомов железа и атомы железа. Координаты атомов железа были уточнены из аномальных нативных данных, а новые фазы были рассчитаны на основе производной золота и аномального вклада от атомов железа. Новая оболочка для субъединицы была выполнена с использованием метода МАМА (Клейвегт и Джонс, Acta Cryst. D50: 178 (1994)) посредством приблизительной ориентации субъединицы R2 в правильном положении для пучка спиралей.
После некристаллографического усреднения стало возможным построение исходной модели десатуразы по карте электронной плотности. Были осуществлены циклы построения модели, ее усовершенствования в XPLOR (Брюнгер А. Acta Cryst. А45: 50 (1989)), (Брюнгер А. Справочник по X-PLOR, Йельский университет, Нью Хэвен, шт. Коннектикут (1990)), повторное определение оболочки, усовершенствования операторов симметрии с использованием IMP (Клейвегт и Джонс, Acta Cryst, D50: 171 (1994)) и усреднения, до тех пор пока в усредненных картах не появлялась новая электронная плотность. На этой стадии из карт 2Fo-Fc была построена еще одна петля, которая, по-видимому, имеет другую структуру в субъединицах.
Кристаллографическое усовершенствование осуществляли в XPLOR, используя силовое поле Энга и Хубера (Энг и Хубер, Acta Cryst. A47: 392 (1991)) и ограничения некристаллографической симметрии. Из-за низкого разрешения (3,1 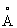 ) набора данных использовали суммарное В-значение. На этой стадии модель имела кристаллографический R-фактор, равный 26,7% при ограничениях шестикратной некристаллографической симметрии, налагаемых этим усовершенствованием. На этой стадии уточнения стал доступным новый нативный набор с разрешением до 2,4
) набора данных использовали суммарное В-значение. На этой стадии модель имела кристаллографический R-фактор, равный 26,7% при ограничениях шестикратной некристаллографической симметрии, налагаемых этим усовершенствованием. На этой стадии уточнения стал доступным новый нативный набор с разрешением до 2,4  , собранный на NSLS, и уточнение продолжалось с этим набором данных. Процесс усовершенствования прослеживался по 2,5% отражениям, которые не были включены в уточнение, но были использованы для расчета свободного значения фактора Rсв (А. Nature, 355: 472 (1992)).
, собранный на NSLS, и уточнение продолжалось с этим набором данных. Процесс усовершенствования прослеживался по 2,5% отражениям, которые не были включены в уточнение, но были использованы для расчета свободного значения фактора Rсв (А. Nature, 355: 472 (1992)).
Даже при разрешении 2,4  наблюдение до параметрического отношения составляет только около единицы, и проблема уточнения плохо выполняется. Поэтому в ходе всего процесса усовершенствования были использованы ограничения некристаллографической симметрии, для того чтобы избежать пере-аппроксимации данных по дифракции. Ограничениям не подвергались только те части структуры, для которых из карт усредненной электронной плотности было подтверждено, что они не подчиняются некристаллографической симметрии. Эти части включают остатки 19-50, 121-122, 127-129, 208-212, 241-253, 259-260, 308-319, 336-348 и некоторые боковые цепи. Электронная плотность для некоторых остатков в области 336-347 настолько низка, что их положения следует рассматривать условно, и поэтому занятость для этих атомов установлена равной нулю. Суммарное анизотропное усовершенствование снижает свободное значение фактора Rсв примерно на 2%. На этой стадии в модель были введены молекулы воды. Кроме того, были уточнены индивидуальные значения В-факторов, но при ограничении некристаллографической симметрии. Окончательная модель имела кристаллографический фактор R, равный 22,0% (Rcв=28,5%). Среднеквадратичные отклонения для ограниченных положений Сα (263 атома) субъединицы А до соответствующих частей других субъединиц составляют 0,06 и для всех атомов Сα (345 атомов) - 0,26, 0,23, 0,24, 0,32, 0,25 соответственно.
наблюдение до параметрического отношения составляет только около единицы, и проблема уточнения плохо выполняется. Поэтому в ходе всего процесса усовершенствования были использованы ограничения некристаллографической симметрии, для того чтобы избежать пере-аппроксимации данных по дифракции. Ограничениям не подвергались только те части структуры, для которых из карт усредненной электронной плотности было подтверждено, что они не подчиняются некристаллографической симметрии. Эти части включают остатки 19-50, 121-122, 127-129, 208-212, 241-253, 259-260, 308-319, 336-348 и некоторые боковые цепи. Электронная плотность для некоторых остатков в области 336-347 настолько низка, что их положения следует рассматривать условно, и поэтому занятость для этих атомов установлена равной нулю. Суммарное анизотропное усовершенствование снижает свободное значение фактора Rсв примерно на 2%. На этой стадии в модель были введены молекулы воды. Кроме того, были уточнены индивидуальные значения В-факторов, но при ограничении некристаллографической симметрии. Окончательная модель имела кристаллографический фактор R, равный 22,0% (Rcв=28,5%). Среднеквадратичные отклонения для ограниченных положений Сα (263 атома) субъединицы А до соответствующих частей других субъединиц составляют 0,06 и для всех атомов Сα (345 атомов) - 0,26, 0,23, 0,24, 0,32, 0,25 соответственно.
Эту белковую модель анализировали, используя варианты PEPFLIP и RSFIT в О (Джонс и др. Acta cryst. A47: 100 (1991)) и программу PROCHECK (Ласковский и др. J. Appl. Cryst. 26: 282 (1993)). Координаты атомов будут депонированы в Брукхэвенский банк данных белков.
Сравнения структур
Все структурные суперпозиции были осуществлены по методу наименьших квадратов, используя О (Джонс и др. Acta cryst. A47: 100 (1991)), причем они были выполнены попарно. Совмещение было выполнено посредством выбора начального набора атомов Сα, содержащих четыре отрезка полипептидной цепи (в каждом примерно по 10 остатков) от четырех спиралей, содержащих лиганды для центра с двумя атомами железа. Эту начальную ориентацию последовательно максимизировали посредством включения всех атомов Сα из атомных моделей. Остатки считались структурно эквивалентными, если они были в пределах 3,8 друг от друга и в пределах последовательного отрезка более чем от трех эквивалентных остатков.
Пример 2
Результаты и их обсуждение
Подход сочетания элементов аминокислотной последовательности из структурно родственных ферментов с различными свойствами оказался эффективным при получении характеристики субстрата и позиционных особенностей ферментов, модифицирующих жирные кислоты, таких как липоксигеназы млекопитающих и растительные ацил-АСР-тиоэстеразы. Этот подход был использован здесь для идентификации остатков, которые ответственны за различия в свойствах Δ9-18:0-АСР-десатуразы и Δ6-16:0-АСР-десатуразы, кодируемых кДНК Т. alata pTAD и рТАΔ4 соответственно. Зрелые полипептиды, кодированные этими кДНК, распределяют 65% идентичности аминокислотной последовательности. Первоначально были созданы два химерных мутанта: а) Химера 1, содержащая первые 171 аминокислоты зрелой Δ6-16:0-АСР-десатуразы, связанной с оставшимися 185 аминокислотами Δ9-18:0-ACP-десатуразы, и б) Химера 2, содержащая первые 227 аминокислот зрелой Δ9-18:0-АСР-десатуразы, связанной с оставшимися 134 аминокислотами Δ6-16:0-АСР-десатуразы. Оба фермента проявляют только детектируемую активность Δ9-18:0-АСР-десатуразы. Кроме аналогичной каталитической активности эти мутанты распределяют область перекрывания 50 остатков (остатки 178-227) Δ9-18:0-АСР-десатуразы.
Это позволяет предположить, что детерминанты длины цепи и особенностей расположения двойной связи присутствуют в этой части Δ9-18:0-АСР-десатуразы. Таким образом, была создана Химера 3, в которой остатки 172-221 Δ6-16:0-АСР-десатуразы были заменены соответствующей областью из 50 аминокислот из Δ9-18:0-АСР-десатуразы. Полученный фермент катализировал образование ненасыщенности Δ6 или Δ9 как в 16:0-АСР, так и 18:0-АСР. Почти идентичная активность была получена для Химеры 4, в которой подгруппа из 30 аминокислот этого домена (остатки 178-207 Δ9-18:0-АСР-десатуразы) была перемещена в Δ6-16:0-АСР-десатуразу. Как показано в таблице, в резком отличии от активности Δ6-16:0-АСР-десатуразы природного типа этот фермент катализирует образование ненасыщенности Δ6 и Δ9 отношении почти 3:1 и 1:1 соответственно в 16:0-АСР и 18:0-АСР. Более того, удельная активность для субстрата 18:0-АСР была почти вдвое больше, чем измеренная для 16:0-АСР. Эти результаты находятся в сильном контрасте с активностью Δ6-16:0-АСР-десатуразы природного типа. Хотя этот химерный фермент способен катализировать введение двойной связи более чем в одно положение 18:0-АСР, в то же время Δ6-16:0-АСР-десатураза природного типа обладает заметной активностью только в образовании ненасыщенности Δ6 в 16:0-АСР. Кроме того, фермент природного типа имеет в 6 раз большую активность в отношении 16:0-АСР, чем с 18:0-АСР.
Для дополнительной характеристики области 50 аминокислот Δ9-18:0-АСР-десатуразы меньшую часть этой последовательности перемещали в Δ6-l6:0-ACP-десатуразу (Химера 5). В отличие от удельной активности Δ6-16:0-АСР-десатуразы природного типа удельная активность полученного фермента была почти одинаковой, как для 16:0-, так и для 18:0-АСР. Кроме расширенной специфичности длины цепи жирных кислот, мутантная десатураза катализирует введение двойной связи почти исключительно в Δ6-положение 16:0- и 18:0-АСР. Более того, удельная активность этого фермента была более чем в два раза выше активности Δ6-16:0-АСР-десатуразы природного типа. Отчасти, это может быть следствием повышенной стабильности мутантного фермента в Е. coli (то есть мутантная десатураза имеет более высокие уровни экспрессии и проявляет повышенную растворимость чем Δ6-16:0-АСР-десатураза природного типа).
Область 178-207 Δ9-18:0-АСР-десатуразы содержит девять аминокислот, которые отличаются от аминокислот, обнаруженных в аналогичной части Δ6-16:0-АСР-десатуразы. Посредством сайт-направленного мутагенеза Δ6-16:0-АСР-десатуразы каждый из этих остатков, или индивидуально, или в сочетании, был превращен в тот, что присутствует в Δ9-18:0-АСР-десатуразе. Была получена активность, качественно аналогичная активности Химеры 4 в результате следующей мутации Δ6-16:0-АСР-десатуразы: A181T/A188G/Y189F/S205N/L206T/G207A (Примечание: нумерация аминокислот дана в соответствии с Δ9-18:0-АСР-десатуразой). Кроме того, фенотип Химеры 5 (то есть расширенная специфичность длины цепи) был качественно реализован посредством мутации A188G/Y189F в Δ6-16:0-АСР-десатуразе. В этих экспериментах также были получены мутантные десатуразы с неочевидными значениями активности. Например, мутация A181T/A200F в Δ6-16:0-АСР-десатуразе привела к получению фермента, который катализирует главным образом введение Δ9-ненасыщенности в 18:0-АСР, но с 16:0-АСР работает как Δ6-десатураза. Однако удельная активность этого фермента с 18:0-АСР была почти в 3 раза меньше активности, обнаруженной для 16:0-АСР. Более того, мутация А181T/A200F/S205N/L206T/G207A в Δ6-16:0-АСР-десатуразе дает фермент, который обладает только измеримой активностью введения Δ9-двойной связи в 18:0-АСР, и был почти в четыре раза активнее с этим субстратом, чем с 16:0-АСР. Подобно мутанту A181T/A200F этот фермент сохраняет активность введения Δ6-двойной связи в 16:0-АСР.
Изменения свойств связывания субстратов этими ферментами могут быть уменьшены, как основная причина наблюдаемых эффектов, поскольку их значения незначительно отличаются от таковых для фермента природного типа. Были оценены значения констант Михаэлиса, Km, для Δ6-16:0-АСР-десатуразы природного типа, Химеры 5 и мутанта A188G/Y189F, которые находятся в интервале от 0,2 до 0,6 мкмоль/л, как для 16:0-, так и для 18:0-АСР.
Как описано в Примере 1, была определена кристаллическая структура касторовой Δ9-18:0-АСР-десатуразы, что обеспечивает возможность интерпретации результатов, полученных для химер и мутантов, в свете окружения активного центра. В структуре субъединицы содержится очень глубокий и узкий канал, который, как оказалось, соответствует связывающему центру для стеариновокислотной части субстрата. Форма канала навязывает изогнутую конформацию алифатической цепи в том месте, где вводится двойная связь (между девятым и десятым атомами углерода), что соответствует цис-конфигурации продукта - олеиновой кислоты, причем возможная двойная связь располагается довольно близко к атому железа каталитического центра в субъединице. Таким образом канал, связывающий субстрат, накладывает жесткие ограничения на длину алифатической цепи за пределами введенной двойной связи, что может частично объяснить различия специфичности действия ферментов этого семейства. Как можно видеть, в вариантах фермента, которые могут принимать субстраты с меньшим числом атомов углерода вне двойной связи, связывающие впадины закрыты посредством замещения аминокислотами с более объемными боковыми цепями. Эти аминокислоты, вовлеченные в определение специфичности, в этой части связывающего центра представляют собой 114-115, 117-118, 179, 181 и 188-189.
В отсутствие структурной модели для комплекса фермент-субстрат-АСР трудно непосредственно проследить за детерминантами особенностей длины цепи по другую сторону двойной связи, по направлению к белку, переносящему ацил. Предполагая, что АСР в различных ферментах этого типа связывает аналогичным образом, различия аминокислотных боковых цепей в верхней части субстратного канала и при входе на поверхности субъединицы позволили бы ферментам приспособить алкильные цепи различной длины между двойной связью и фосфопантеин-простетической группой АСР. Однако аминокислоты, окружающие верхнюю часть центра связывания от двойной связи до поверхности белка, сохраняются в доступных последовательностях фермента, и наиболее вероятно, что детерминанты специфичности находятся на входе в субстратный канал и на поверхности фермента, которая взаимодействует с ацил-АСР. Здесь связывающий карман расширяется, и невозможно смоделировать фосфопантеиновую часть стеароил-АСР. Остатки 280, 283, 286 и 294 в этой области не сохраняются между различными ферментами, и они могут быть вовлечены в определение специфичности субстрата.
Из структуры связывающего центра в этой области можно дать рациональное объяснение некоторым результатам по химерам и мутантам. Все химеры и мутанты включают детерминат 179-189 (фактически остатки 179, 181, 188-189), и таким образом неудивительно обнаружить их действие на специфичность. Как Химера 1, так и Химера 2 имеют очень низкую остаточную активность, вероятно, из-за некоторых стерических затруднений при их образовании. Химера 1 имеет этот детерминант Δ9-18:0-АСР-десатуразы в глубоком кармане, а также поверхностный детерминант, специфичный для Δ9-18:0-АСР-десатуразы; только один детерминант, остатки 114-115 и 117-118, является специфичным для Δ6-16:0-АСР-десатуразы, и таким образом низкая остаточная активность этой химеры представляет собой активность Δ9-18:0-десатуразы. Химера 2 имеет полный детерминант Δ9-18:-десатуразы в области скрытого кармана и известный детерминант Δ6-16:0-АСР-десатуразы на поверхностном конце; эта химера также обладает Δ9-18:0-АСР активностью. Химеры 3 и 4 сохраняют свою активность, один из детерминантов в глубоком кармане является определителем для Δ9-18:0-АСР, остаток А181 замещается более крупной треониновой боковой цепью, но в то же время А188 замещается глицином, a Y189 - на фенилаланин, фактически создавая больше свободного пространства в глубине полости, и таким образом обеспечивая активность даже для Δ6-18:0-АСР-десатуразы. Химера 5 отличается от химеры 4 только тем, что в ней сохраняется последовательность Δ6-16:0-АСР-десатуразы для остатков 203-207. Эти остатки находятся в верхней части субстратного канала, но не осуществляют непосредственного контакта с субстратами, и трудно понять их влияние на специфичность к субстрату. Эти остатки хорошо сохраняются между известными десатуразами в этом семействе, только Δ6-16:0-АСР-десатураза имеет другую последовательность для остатков от 205 до 207, и эта область, вероятно, не является частью естественного детерминанта для специфичности к субстрату. В случае мутанта A181T/A200F снижение Δ6-16:0 активности по сравнению с ферментом природного типа согласуется со структурными изменениями в субстратном канале из-за уменьшения размера этой полости при замене А181 на треонин. Влияние A200F невозможно объяснить рационально, этот остаток находится на поверхности субъединицы и направлен в сторону от субстратного канала. Во всех секвенированных десатуразах в этом семействе, за исключением Δ6-16:0-АСР, этот остаток является фенилаланином. Из предшествующего обсуждения ясно, что активность мутанта A181T/A200F/S205N/L206T/G207A нвозможно объяснить в рамках структуры, мы не смогли найти разумное объяснение влиянию изменений в остатках 200 и 205-207.
Таким образом, было показано, что регио-специфичность и специфичность длины цепи десатуразы жирных кислот могут быть изменены посредством специальных замещений аминокислот. Детерминанты специфичности длины цепи частично наносят на карту область трехмерной структуры, которая определяет форму и размер канала, связывающего субстрат. Однако некоторые из этих остатков находятся за пределами канала, связывающего субстрат, и изменения в таких остатках могут привести к новой и полезной активности. При доступности трехмерной структуры АСР-десатуразы жирных кислот, представленные здесь успешные попытки изменения специфичности к субстратам теперь могут быть распространены на разумно сконструированные варианты ферментов, обладающих другой специфичностью к длине цепи, а также регио-специфичностью. Однако это может быть успешным только при условии, что на основе кристаллической структуры субстратного комплекса и доступности множества аминокислотных последовательностей ферментов в этом семействе мы сможем решить, что представляют собой детерминанты специфичности, расположенные на входе в субстратный канал.
Материалы и методы
Названия жирных кислот сокращены в формате x:ydz, где х - это длина цепи или число атомов углерода в жирной кислоте, у - число двойных связей и z - положение двойной связи в жирной кислоте относительно карбоксильного конца молекулы (например, олеиновая кислота или 18:1Δ9 является восемнадцатиуглеродной жирной кислотой с одной двойной связью, которая расположена при девятом атоме углерода относительно карбоксильного конца молекулы).
Получение химерных мутантов
Химерные мутанты получают путем сшивки частей кодирующей последовательности зрелых Δ6-16:0- и Δ9-18:0-АСР-десатураз Т.alata через нативные ферментные сайты рестрикции или ферментные сайты рестрикции, созданные с помощью PCR, сайт-специфичные мутации в кодирующей последовательности аминокислот 178-202 в Δ9-18:0-АСР-десатуразе (эквивалентны остаткам 172-196 в Δ6-16:0-АСР-десатуразе) вводят путем удлинения и амлификации перекрывающихся олигонуклеотидных праймеров, с использованием PCR с Pfu-полимеразой (фирма Стратаген). Мутации A181T/A188G/Y189F произведены со следующими олигонуклеотидами:
5'ATGGATCCTGGCACGGATAACAACCCGTAC3' (Праймер 1А);
5'ACGAGGTGTAGATAAATCCGAGGTACGGGTTGTTATCCG3' (Праймер 2А);
5'TATCTACACCTCGTATCAGGAGAGGGCGACA3' (Праймер 3А);
5'TTGAATTCCATGGGAAATCGCTGTCGCCCTCTCCTG3' (Праймер 4А).
Мутации A188G/Y189F вводят, используя следующие олигонуклеотиды:
5'ATGGATCCTGGCGCGGATAACAACCCGTAC3' (Праймер 1В); Праймер 2А;
Праймер 3А; Праймер 4А. Мутации A181T/A200F были созданы следующим образом:
Праймер 1А; 5'ACGAGGTGTAGATATATGCGAGGTACGGGTTGTTATCCG3' (Праймер 2В); Праймер 3А;
5'TTGAATTCCATGGGAAATGAATGTCGCCCTCTCCTG3' (Праймер 4В).
Реакции PCR проводят без добавления матрицы, используя 12,5 пикомолей Праймеров 1А или В и 4А или В и 6,25 пикомолей Праймеров 2А или В и 3А. Для первых десяти циклов PCR используют температуру отжига 37°С и температуру удлинения 72°С. После этого следуют 20 дополнительных циклов с повышенной температурой отжига, до 55°С. Продукты реакции PCR расщепляют с BamHI и EcoRI и встраивают в соответствующие сайты pBluescript II KS(-) (Стратаген), из которых определяют нуклеотидную последовательность, используя набор Секвеназы 2.0 (фирма Амершам). Затем эту плазмиду расщепляют с BamHI и EcoRI и выделенную вставку лигируют с последовательностью, кодирующей аминокислоты 1-171 зрелой Δ6-16:0-АСР-десатуразы в векторе экспрессии рЕТ3а (Новаген). Образовавшуюся конструкцию (содержащую теперь кодирующую последовательность аминокислот 1-196 мутантной или природной Δ6-16:0-АСР-десатуразы) подвергают рестрикции действием NcoI и EcoRI и лигируют с фрагментом NcoI/EcoRI, содержащим кодирующую последовательность оставшихся аминокислот (остатки 197-355) Δ6-16:0-АСР-десатуразы и частью плазмиды pET3d (нуклеотиды). Мутацию S205N/L206T/G207A создают посредством PCR амплификации кодирующей последовательности аминокислот 197-355 в Δ6-16:0-АСР-десатуразе, используя в качестве матрицы первоначальную кДНК для этого фермента в pBluescript II SK (-). 5'-олигонуклеотид
(5'TTTCCATGGGAACAAGGCTCGGCTAGCGAGGCAGAAGG3'),
содержал подходящие мутантные кодоны, а Т7 - праймер был использован в кеачестве 3'-олигонуклеотида в реакциях PCR. Продукт амплификации расщепляют с NcoI и BclI и всраивают в сайт NcoI/BamHI плазмиды рЕТ 3d. Затем фрагмент NcoI/EcoRI из этой конструкции лигируют с кодирующей последовательностью аминокислот 1-196 подходящей мутантной Δ6-16:0-ACP-дecaтypaзы (например, A181T/A200F), чтобы создать полноразмерную кодирующую последовательность. В продуктах реакций PCR секвенируют последовательности, чтобы подтвердить наличие целевых мутаций.
Продуцирование ацил-АСР-десатураз
Ацил-АСР-десатуразы природного типа и мутантные получают путем экспрессии кодирующих последовательностей в Е. coli BL21 (DE3) под контролем промотора Т7 РНК-полимеразы с использованием векторов рЕТ3а или pET3d. Рекомбинантные ферменты, активность которых описана в таблице, очищают из бактериальных культур (объемом от 6 до 9 л), культивируемых при температуре от 20 до 25°С. Очистку белка осуществляют, используя DEAE-анионно-обменную хроматографию с последующей 20HS (фирма Персептив Байосистемз) катионно-обменной хроматографией с использованием жидкостной хроматографии высокого давления Байокад Спринт (фирма Персептив Байосистемз). Мутантные десатуразы были получены с чистотой r 90%, а Δ6-16:0-АСР-десатуразу природного типа выделяли с чистотой примерно 80%. После очистки ферменты подвергают обмену в буфере, состоящем из 40 ммоль/л гидрохлорида-Трис (рН 7,5), 40 ммоль/л хлорида натрия и 10% глицерина, и хранят в аликвотах при -75°С после мгновенного замораживания в жидком азоте.
Испытание и анализ ацил-АСР-десатураз
Испытание и анализ продуктов реакции ацил-АСР-десатуразы проводят, как описано ранее (Кэйхун и др. Proc. Nat. Acad. Sci., USA. 89: 1184 (1994)) со следующими изменениями: использовали рекомбинантный растительный ферредоксин Anaebena (22 фемто-г на 1 испытание) и корень кукурузы FNR (0,4 ед/испытание) вместо ферредоксина шпината и FNR, причем количества NADPH и радиоактивной [1-14С]16:0- или 18:0-АСР, взятых на одно испытание, увеличили соответственно до 2,5 ммоль/л и 178 пикомолей (или 1,2 фемто-моль/л). Использованная в синтезе субстратов АСР представляет собой рекомбинантную ACP-I шпината. Удельная активность [1-14С]16:0- или 18:0-АСР (от фирмы Америкэн Рэйдиолэйблд Кемикалз) составляет 55 мКюри/ммоль. Активность ферментов измеряют путем определения концентрации (%) моно-ненасыщенного продукта, образовавшегося при испытании десатуразы. Распределение радиоактивности между продуктами и непрореагировавшим субстратом определяют с помощью фосфоресцирующих изображений продуктов разделения на посеребренной пластинке тонкослойной хроматографии (ТСХ) с использованием программного обеспечения Имэдж-Квант и методом счета жидкостных сцинцилляций для соскобов ТСХ.
Определение положения двойных связей.
Положение двойных связей в продуктах - мононенасыщенных жирных кислотах определяют по подвижности метилэфирных производных на посеребренных (15%) пластинках ТСХ и с использованием хромато-масс-спектрометрического (Хромасс) анализа диметилсульфидных аддуктов этих производных. Испытания образования двойных связей для Хромасс-анализов проводят с использованием немеченых 16:0-, 17:0- и 18:0-АСР в качестве субстратов и очищенных ферментов. В дополнение к результатам, приведенным в тексте, было обнаружено около 15% ненасыщенных продуктов, образовавшихся в результате реакции 17:0-АСР с Δ6-16:0-АСР-десатуразой, в виде 17:1Δ7-изомера. Остальная часть продукта представляет собой 17:1Δ6, также обнаружены следовые количества 17:1Δ9.
Изобретение относится к белковой инженерии. Аминокислотные остатки фермента, контактирующие с субстратом внутри канала связывания, идентифицируют, а затем заменяют на другие остатки, которые имеют такие же зарядовые характеристики и сходный размер боковой цепи. Замена в ацил-АСР десатуразе остатков в положениях 114, 115, 117, 118, 179, 181, 188 и 189 приводит к изменению длины цепи и специфичности расположения двойных связей в ферменте, что изменяет его активность. 7 с. и 17 з.п. ф-лы, 1 табл.
| US 5530186 А, 25.07.1996 | |||
| J | |||
| MOL | |||
| BIOLOGY, 1992, 225, 561-564 | |||
| CURRENT OPINION IN STRUCTURAL BIOLOGY, 1995, 5, 758-766 | |||
| J.BIOL | |||
| CHEMISTRY, 1994, 269, № 44, 27519-27526. |

