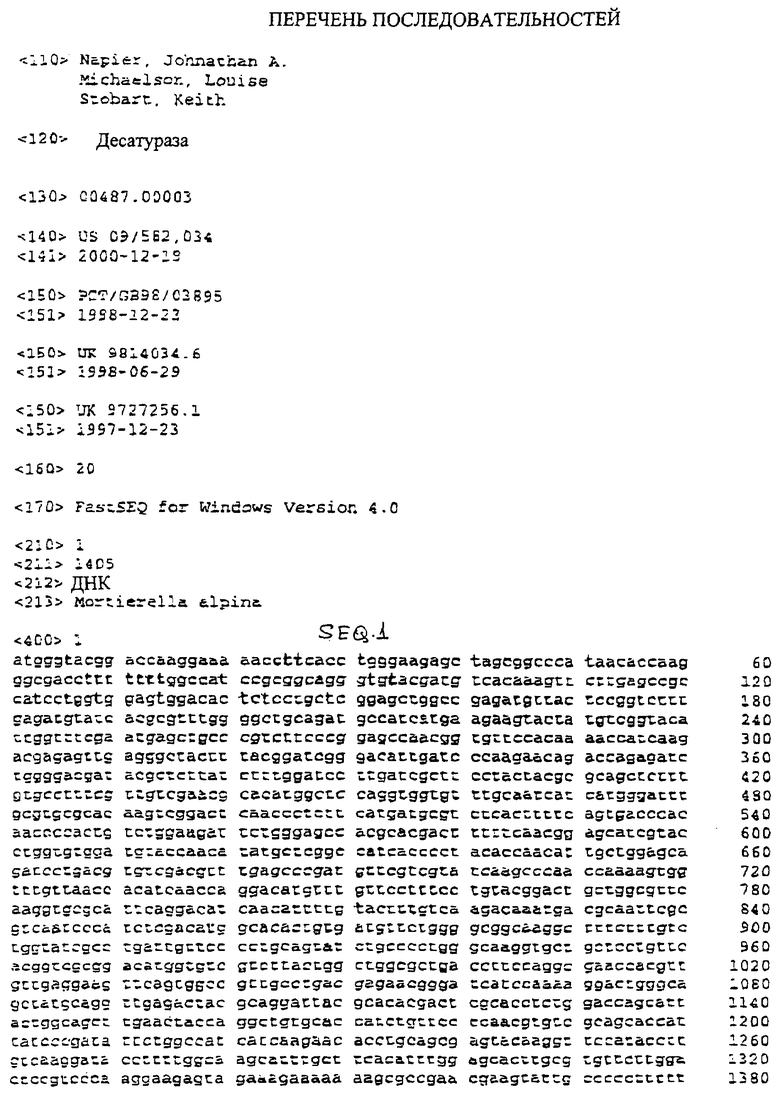
Это изобретение относится к последовательностям ДНК, кодирующим Δ5-десатуразы жирных кислот, кодируемым Δ5-десатуразам жирных кислот и применению Δ5-десатураз жирных кислот.
Полиненасыщенные жирные кислоты важны физиологически благодаря их специфическим укрепляющим здоровье активностям и биомедицински в отношении их потенциального фармацевтического применения при лечении определенных болезненных состояний.
Полиненасыщенные жирные кислоты являются предшественниками двух больших классов метаболитов: простаноидов (которые включают в себя простаглаидины и тромбоксаны) и лейкотриенов. Δ5-Десатураза жирных кислот катализирует превращение дигомо-γ-линоленовой кислоты (ДГЛК) в арахидоновую кислоту (АК) и эйкозатетраеноата (ЭТЕ) в эйкозапентаеноат (ЭПЕ), путем введения двойных связей в положение Δ5-углерода соответствующих субстратов, и существует в форме связанного с мембраной эндоплазматического ретикулума белка в его нативном состоянии.
Арахидоновая кислота имеет 20-членную углеродную цепь с 4 двойными связями и играет большое значение в метаболизме человека, так как она является предшественником для синтеза простагландинов - жирных кислот с 20-членной углеродной цепью, которые содержат 5-членное углеродное кольцо. Простагландины являются модуляторами действия гормонов, а потенциальные эффекты простагландинов включают в себя стимуляцию воспаления, регуляцию кровотока к отдельным органам, контроль ионного транспорта через некоторые мембраны и модуляцию синаптической передачи. Простагландины также потенциально применимы в качестве контрацептивов, благодаря их способности подавлять секрецию прогестерона. Поэтому умение модулировать синтез простагландинов путем контролируемых уровней экспрессии синтеза предшественников полиненасыщенных жирных кислот является очень важным как медицински, так и промышленно.
Возрастающее значение полиненасыщенных жирных кислот в пищевой и фармацевтической промышленностях привело к повышенному спросу, который превысил уровни современного производства и дополнительных источников высокого качества, необходимы дешевые полиненасыщенные жирные кислоты.
Современные промышленные источники полиненасыщенных жирных кислот включают в себя выбранные семенные растения, морскую рыбу и выбранных млекопитающих, а традиционные методы переработки для экстрагирования полиненасыщенных жирных кислот из этих источников включают в себя экстракцию растворителем, демаргаринизацию, образование аддуктов мочевины и дистилляцию. Однако представленные источники имеют недостатки, касающиеся сезонных и климатических различий как в уровнях производства, так и в качестве нехватки растительных и рыбных источников и высоких цен рафинирования низкосортных масел. Высокие цены, связанные с недостаточными уровнями производства, замедлили развитие промышленно пригодного применения полиненасыщенных жирных кислот.
Большие усилия были затрачены на разработку альтернативных источников полиненасыщенных жирных кислот, и были проведены исследования для характеристики составных генов и кодируемых белков их биосинтеза. Инженерия биосинтеза полиненасыщенных жирных кислот, например в масличных культурах, имеет много достоинств для производства крупномасштабных количеств, например, γ-линолената (ГЛ), дигомо-γ-линолената (ДГГЛ), арахидоновой кислоты (АК), эйкозапентаеноата (ЭПЕ) и докозагексаеноата (ДГЕ). Практическая реализация этого была проиллюстрирована экспрессией гена Δ6-десатуразы бурачника в табаке, имевшей результатом продукцию ГЛ и октадекатетраеновой кислоты, 18:4 (Soyanova et.al. (1997), PNAS 94, 9411-9414), Поскольку большинство биосинтетических генов для синтеза полиненасыщенных жирных кислот становятся доступными, это предоставляет возможность для продуцирования по меньшей мере ГЛ, АК, ЭПЕ и ДГЕ в масличных культурах, а также контролирования типа собранного липида. Преимущества, которые были бы получены от таких культур включают в себя дешевый и поддерживаемый запас требуемых в крупном масштабе полиненасыщенных жирных кислот, заданный профиль полиненасыщенных жирных кислот для удовлетворения конкретных пищевых потребностей, а в тонкой химической промышленности получение необычных жирных кислот с заданными степенями и положениями ненасыщенности.
Дополнительным подходом к получению полиненасыщенных жирных кислот является утилизация биосинтетической способности низших организмов, например, водорослей, бактерий, грибов (включая фикомицетов), которые могут синтезировать весь ряд полиненасыщенных жирных кислот и могут выращиваться в промышленных масштабах. Генетическая трансформация этих организмов будет облегчать выведение сверхпродуцирующих штаммов и манипулирование полиненасыщенным профилем путем инженерии.
Были клонированы грибные Δ5- и Δ6-десатуразы жирных кислот, а их последовательности рассмотрены в WO 98/46763, WO 98/46764, WO 9846765.
Метаболизм полиненасыщенных жирных кислот имеет наибольшее значение в метаболизме человека. Эти кислоты, через эйкозаноиды, являются фундаментальными для правильного поддержания гомеостаза и сцеплены с серьезными физиологическими и патофизиологическими синдромами.
Неожиданно авторы изобретения выделили из обитающего в почве мицелиального гриба класса зигомицетов, Mortierella alpina, и охарактеризовали последовательность ДНК, кодирующую функционально активную Δ5-десатуразу жирных кислот.
Вместе с тем, авторы изобретения неожиданно выделили из нематоды, Caenorhabditis elegans, и охарактеризовали последовательность ДНК, кодирующую функционально активную Δ5-десатуразу жирных кислот. Эта последовательность ДНК, кодирующая функционально активную Δ5-десатуразу жирных кислот, как полагают, вероятно, более тесно связана с Δ5-десатуразой жирных кислот человека, чем какие-либо из до сих пор выделенных последовательностей гена Δ5-десатуразы жирных кислот.
Также как и потенциальные благоприятные эффекты для человека от полипептида, кодируемого последовательностями ДНК по изобретению, последовательности ДНК по изобретению могут облегчить клонирование эквивалентного гена человека и, таким образом, способствовать сверхпродуцированию последовательности ДНК человека и сделать возможным ее биомедицинское использование при лечении некоторых болезней человека.
Растительные и грибные десатуразы являются, главным образом, интегральными мембранными полипептидами, что затрудняет их очистку и впоследствии характеристику традиционными методами. Поэтому для лучшего изучения липидного обмена выбраны молекулярные методы, включающие в себя использование мутантов и трансгенных растений.
Согласно первому аспекту изобретения предложена выделенная животная Δ5-десатураза жирных кислот и ее функционально активные части.
Согласно второму аспекту изобретения предложена выделенная Δ5-десатураза жирных кислот С.elegans.
Согласно третьему аспекту изобретения предложена последовательность ДНК по первому или второму аспекту изобретения, включающая в себя по меньшей мере часть последовательности, представленной в SEQ.2, и эквиваленты этой последовательности или частей этой последовательности, которые кодируют функционально активную Δ5-десатуразу жирных кислот в силу вырожденности генетического кода. Предпочтительно, последовательность ДНК имеет происхождение от последовательности ДНК Caenorhabditis elegans.
Предпочтительно, ген, кодирующий Δ5-десатуразу жирных кислот, кодируемую клонированным геном, имеет длину 1341 п.н. (пар нуклеотидов). Белок с подсчитанной молекулярной массой 57 кДа имеет длину 447 аминокислот.
В качестве альтернативы, последовательность ДНК кодирует функционально активную Δ5-десатуразу жирных кислот и включает в себя по меньшей мере часть последовательности, представленной в SEQ.1, и эквиваленты этой последовательности или частей, этой последовательности, которые кодируют функционально активную Δ5-десатуразу жирных кислот в силу вырожденности генетического кода. Предпочтительно, последовательность ДНК имеет происхождение от последовательности ДНК Mortierelfa alpina.
Предпочтительно, ген, кодирующий Δ5-десатуразу жирных кислот, кодируемую клонированным геном, имеет длину 1338 п.н. Белок с подсчитанной молекулярной массой 57 кДа имеет длину 447 аминокислот.
Предпочтительно, последовательность ДНК по третьему аспекту изобретения функционально активна у млекопитающего.
Предпочтительно, последовательность ДНК экспрессируется у млекопитающего.
Предпочтительно, последовательность ДНК экспрессируется у человека.
Предпочтительно, последовательность ДНК получена путем модификации функционально активного природного гена, кодирующая Δ5-десатуразу жирных кислот.
Предпочтительно, модификация включает в себя модификацию химическими, физическими или биологическими способами без утраты каталитической активности фермента, который она кодирует.
Предпочтительно, модификация улучшает каталитическую активность фермента, который она кодирует.
Предпочтительно, биологическая модификация включает в себя методы рекомбинантных ДНК и методы принудительной эволюции.
Предпочтительно, метод принудительной эволюции представляет собой перетасовку ДНК.
Согласно четвертому аспекту изобретения предложен полипептид, кодируемый последовательностью ДНК по третьему аспекту изобретения.
Предпочтительно, по меньшей мере часть полипептида имеет последовательность, представленную в SEQ.3, или функционально активные эквиваленты этой последовательности или частей этой последовательности. В качестве альтернативы, по меньшей мере часть полипептида имеет последовательность, представленную в SEQ.4, или функционально активные эквиваленты этой последовательности или частей этой последовательности.
Предпочтительно, полипептид катализирует превращение дигомо-γ-линоленовой кислоты в арахидоновую кислоту.
Предпочтительно, полипептид модифицирован без утраты каталитической активности кодируемого полипептида.
Предпочтительно, полипептид модифицирован таким образом, чтобы вводить конкретную степень насыщенности субстрата в конкретное положение в молекулярной структуре субстрата.
Согласно пятому аспекту изобретения предложен вектор, содержащий последовательность ДНК любой части последовательности ДНК по третьему аспекту изобретения.
Согласно шестому аспекту изобретения предложен способ получения полиненасыщенных жирных кислот, при котором субстрат приводят в контакт с Δ5-десатуразой жирных кислот по первому или второму аспекту изобретения или с полипептидом по четвертому аспекту изобретения.
Согласно седьмому аспекту изобретения предложен способ превращения дигомо-γ-линоленовой кислоты в арахидоновую кислоту, при котором превращение катализируют Δ5-десатуразой жирных кислот по первому или второму аспекту изобретения или полипептидом по четвертому аспекту изобретения.
Согласно восьмому аспекту изобретения предложен организм, сконструированный для продуцирования высоких уровней полипептида по четвертому аспекту изобретения.
Согласно девятому аспекту изобретения предложен организм, сконструированный для продуцирования высоких уровней продукта реакции, катализируемой Δ5-десатуразой жирных кислот по первому или второму аспекту изобретения или полипептидом по четвертому аспекту изобретения.
Предпочтительно, организм сконструирован для осуществления способа по шестому или седьмому аспекту изобретения.
Предпочтительно, организм представляет собой микроорганизм.
Предпочтительно, микроорганизм выбран из водорослей, бактерий и грибов.
Предпочтительно, грибы включают в себя фикомицеты. В качестве альтернативы, микроорганизм представляет собой дрожжи.
В качестве альтернативы, организм представляет собой растение. Предпочтительно, растение выбрано из масличных растений.
Предпочтительно, масличные растения выбраны из масличного рапса, подсолнечника, злаков, включая кукурузу, табака, бобовых, включая арахис и сою, сафлора, масличной пальмы, кокосовой и других пальм, хлопчатника, кунжута, горчицы, льна, клещевины, бурачника и энотеры.
Согласно десятому аспекту изобретения предложен семенной или другой репродуктивный материал, имеющий происхождение от организма по девятому аспекту изобретения.
Предпочтительно, организм представляет собой млекопитающее.
Согласно одиннадцатому аспекту изобретения предложен мультиферментный путь, где путь включает в себя Δ5-десатуразу жирных кислот по первому или второму аспекту изобретения.
Согласно двенадцатому аспекту изобретения предложено соединение, полученное путем превращения субстрата, где указанное превращение катализируют Δ5-десатуразой жирных кислот по первому или второму аспекту изобретения.
Согласно тринадцатому аспекту изобретения предложено промежуточное соединение, полученное путем реакции, катализируемой Δ5-десатуразой жирных кислот по первому или второму аспекту изобретения.
Согласно четырнадцатому аспекту изобретения предложен продукт питания или диетическая добавка, содержащие полиненасыщенную жирную кислоту, полученную способом по шестому аспекту изобретения.
Согласно пятнадцатому аспекту изобретения предложен фармацевтический препарат, содержащий полиненасыщенную жирную кислоту, полученную способом по шестому аспекту изобретения.
Согласно шестнадцатому аспекту изобретения предложены простагландины, синтезируемые биосинтетическим путем, включающим в себя каталитическую активность Δ5-десатуразы жирных кислот по первому или второму аспекту изобретения.
Согласно семнадцатому аспекту изобретения предложен способ модуляции синтеза простагландинов путем контроля уровней экспрессии последовательности ДНК по третьему аспекту изобретения.
Согласно восемнадцатому аспекту изобретения предложен зонд, включающий в себя всю последовательность ДНК по третьему аспекту изобретения или ее часть либо эквивалентную последовательность РНК.
Согласно девятнадцатому аспекту изобретения предложен зонд, включающий в себя весь полипептид-Δ5-десатуразу жирных кислот по четвертому аспекту изобретения или его часть.
Согласно двадцатому аспекту изобретения предложен способ выделения Δ5-десатураз с использованием зонда по девятнадцатому аспекту изобретения.
Возможно, что ген по изобретению может быть трансформирован в клетки человека и использован в методах генотерапии на подходящем уровне in vivo для обеспечения в организме пациента постоянного запаса жирных кислот, превращаемых ферментами в полиненасыщенные жирные кислоты. Это могло бы быть эффективным профилактическим лечением, например, для пациентов страдающих высокими уровнями холестерина или другими медицинскими состояниями, где введение полиненасыщенных жирных кислот может иметь благоприятные эффекты, предупреждающие заболевание.
Вместе с тем, либо целые последовательности ДНК по изобретению или их части, либо целые полипептидные последовательности по изобретению или их части могли бы быть использованы в качестве зондов для поиска в исследовательских или диагностических целях.
Далее изобретение будет описано посредством единственного примера со ссылкой на сопровождающие графические материалы, SEQ.1-4 и Фиг.1-4, в которых:
SEQ.1 представляет собой последовательность кДНК, кодирующую Δ5-десатуразу жирных кислот из Mortierella alpina;
SEQ.2 представляет собой последовательность кДНК, кодирующую Δ5-десатуразу жирных кислот из С.elegans;
SEQ.3 представляет собой пептидную последовательность, полученную трансляцией генной последовательности SEQ.1;
SEQ.4 представляет собой пептидную последовательность, полученную трансляцией генной последовательности SEQ.2;
Фиг.1 представляет собой выравнивание гена, кодирующего Δ5-десатуразу жирных кислот Mortierella alpina с разными *6-десатуразами и Δ5-десатуразой;
Фиг.2 представляет собой выравнивание гена, кодирующего Δ5-десатуразу жирных кислот с Δ6-десатуразой С.elegans и грибной Δ5-десатуразой из М.alpina;
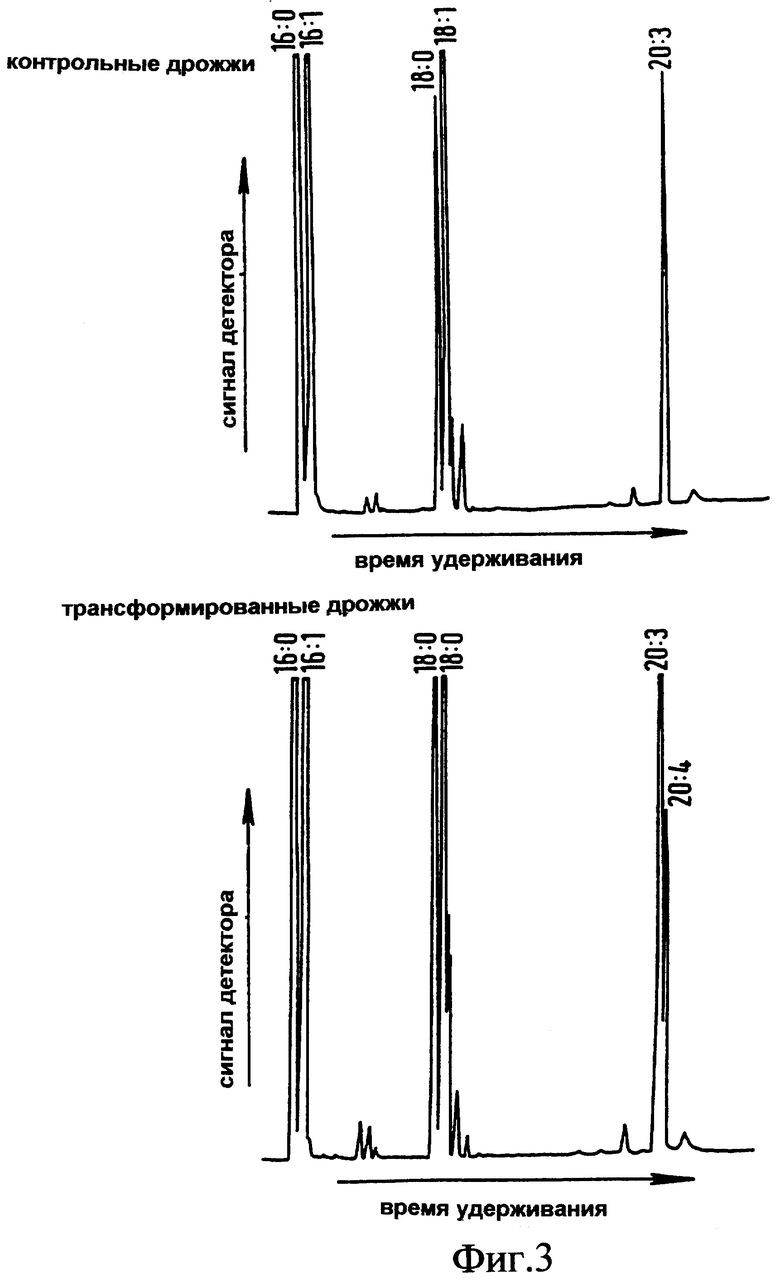
Фиг.3 представляет собой полученную с помощью газовой хроматографии хроматограмму метиловых эфиров жирных кислот из индуцированных дрожжевых клеток-трансформантов, трансформированных геном Δ5-десатуразы жирных кислот Mortierella alpina, и неиндуцированных дрожжевых клеток-трансформантов;
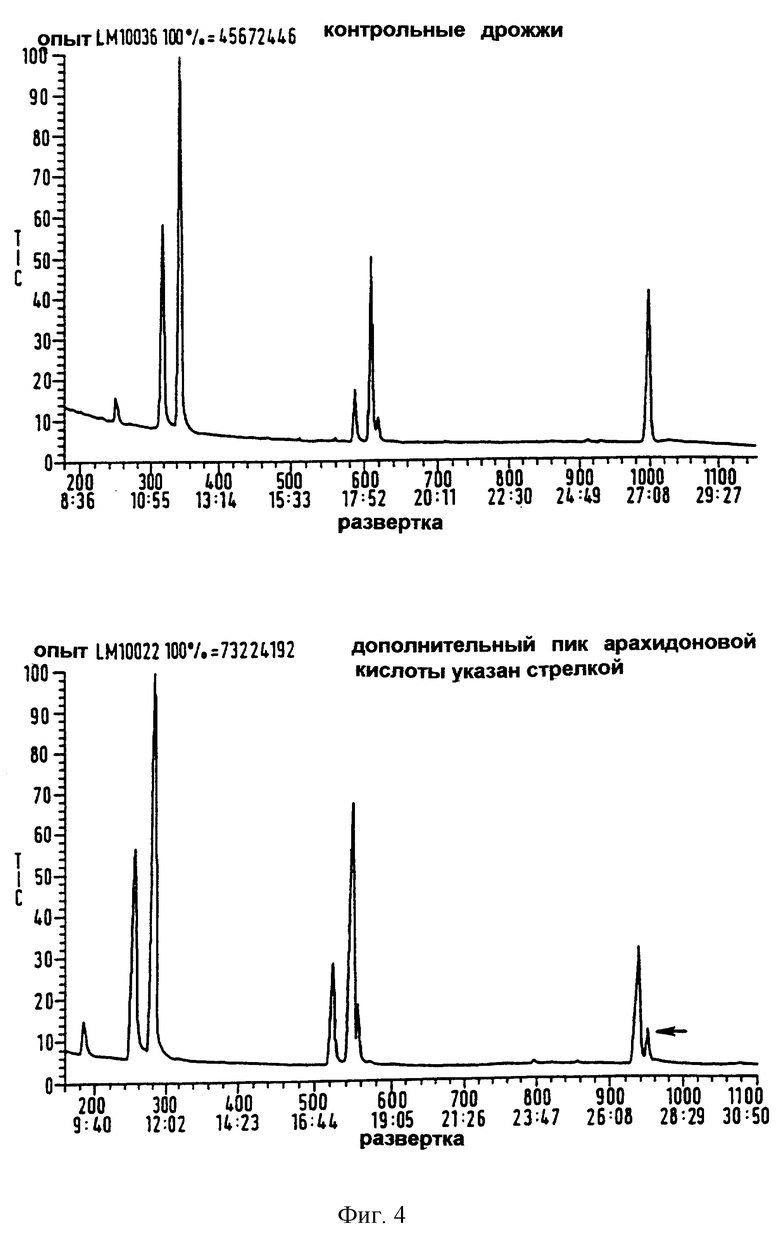
Фиг.4 представляет собой полученную с помощью газовой хроматографии хроматограмму метиловых эфиров жирных кислот из индуцированных дрожжевых клеток-трансформантов, трансформированных геном Δ5-десатуразы жирных кислот C.elegans, и неиндуцированных дрожжевых клеток-трансформантов.
Клонирование и секвенирование гена Δ5-десатуразы жирных кислот из Mortierella alpina
Последовательности ДНК по изобретению, кодирующие Δ5-десатуразы жирных кислот, клонировали, используя метод ПЦР в сочетании с матрицами из библиотеки кДНК и праймерами, сконструированными конкретным образом. Функцию последовательностей ДНК, а именно превращение дигомо-γ-линоленовой кислоты (ДГГЛК) в арахидоновую кислоту (АК) и эйкозатетраеноата (ЭТЕ) в эйкозапентаеноат (ЭПЕ), проверяли путем экспрессии соответствующих кДНК в дрожжах.
Ген Δ5-десатуразы жирных кислот из Mortierella alpina клонировали методами полимеразной цепной реакции (ПЦР), используя кДНК из Mortierella alpina в качестве матрицы и вырожденные олигонуклеотидные праймеры (ВП), сконструированные конкретным образом, как показано ниже, основанные на первом и третьем гистидиновых боксах растительных Δ12- и Δ15-десатураз, предварительно идентифицированных Шенклином (Shanklin, J, Whittle E J & Fox, BG Biochemistry. 33, 12787-12794 (1994)).
Вырожденные олигонуклеотидные праймеры (ВП),
5'-GCGAATTA(A/T)TIGGICA(T/C)GA(T/C)TG(T/C)GICA-3'
5'-GCGAATTCATIT(G/T)IGG(A/G)AAIA(G/A)(A/G)TG(A/G)TG-3'
где I представляет собой инозин, а Есо RI-сайты подчеркнуты.
ПЦР-амплификации проводили полностью традиционно в термоблоке для реакций, программированном на выполнение программы: 2 минуты при 94°С, затем 45 секунд при 94°С, 1 минута при 55°С и 1 минута при 72°С в течении 32 циклов, с последующей элонгацией при 72°С в течение дополнительных 10 минут. Продукты ПЦР-амплификации разделяли в 1%-ных агарозных гелях.
Ряд продуктов ПЦР, амплифицированных на кДНК-матрице из Mortierella alpina, включал продукт длиной 660 п.н., который очищали от геля, клонировали в векторе pGEM-T (PromegaRTM) и трансформировали в экспрессионного хозяина Escherichia coli, DH 5α.
К последовательности продукта длиной 660 п.н. сконструировали праймеры (П) и выполняли амплификацию фрагментов путем ПЦР, используя клонированный фрагмент длиной 660 п.н. в качестве матрицы и последовательность-специфичные праймеры (П), основанные на последовательности продукта длиной 660 п.н.
Delta В прямой
5'-GATGCGTCTCACTTTTCA-3'
Delta В обратный
5'-GTGGTGCACAGCCTGGTAGTT-3'
Продукты этой ПЦР-амплификации очищали от геля и использовали в качестве зондов для скрининга библиотеки кДНК Mortierella alpina. Зонд для поиска фрагмента гибридизовался с 25 из 3,5×105 проверенных фаговых клонов и один клон, как было показано путем рестрикционного анализа, имел ожидаемый размер - 1,5 т. п.н. (тысяч пар нуклеотидов). Этот клон, обозначенный L11, отобрали для дальнейшего анализа. Анализ последовательности L11 выявил открытую рамку считывания длиной 1338 п.н., кодирующую полипептид из 446 аминокислот. Когда продолжили анализировать белок и геномные базы данных, используя программу GCG8 (Devereux J. et al. Nucleic Acids. Res. 12, 387-395 (1984)), L11 показала низкую степень идентичности (20%) гену Δ6-десатуразы из вида Synechocystis PCC6803 (Фиг.1).
На Фиг.1 выравненные последовательности имеют следующие регистрационные номера:
S54259 Δ12-десатураза Spirulina регистрационный номер Х86736
S54809 Δ6-десатураза Spirulina регистрационный номер Х87094
S68358 предполагаемая десатураза сфинголипидов регистрационный номер Х87143
S35157 Δ6-десатураза Synechocystis регистрационный номер L 11421
PBOR6 Δ6-десатураза бурачника регистрационный номер U79010
FU2 Δ5-десатураза регистрационный номер AF054824
К тому же, хотя все три гистидиновых бокса, характерная черта десатуразных ферментов, присутствуют в транслируемой последовательности, третий гистидиновый бокс, локализованный в последовательности в положении 1159 п.н., содержит вариант QXXHH. Транслируемая последовательность также содержит на N-конце цитохром b5-подобный гем-связывающий домен, который включает в себя EHHPGG-фрагмент, несмотря на то, что эта особенность предварительно была обнаружена только на С-конце других грибных десатураз.
Саузерн-блоттинг геномной ДНК
Последовательность-специфичные праймеры, сконструированные к последовательности L11 между гистидиновыми боксами 1 и 3 последовательности L11, использовали в ПЦР-реакции для амплификации участка длиной 660 п.н. последовательности L11. Продукт ПЦР длиной 660 п.н. очищали от геля и выполняли блот-гибридизацию по Саузерну рестрицированных геномных ДНК из Mortierella alpina и Mucor circinelloides, используя в качестве зонда фрагмент длиной 660 п.н. Результаты означают, что ген, кодирующий Δ5-десатуразу жирных кислот по изобретению, представлен у Mortierella alpina уникальной копией и, по-видимому, отсутствует у Mucor circinelloides. К тому же, у Mucor circinelloides не обнаруживается Δ5-десатуразная активность.
Экспрессия клонированного гена Mortierella alpina, кодирующего Δ5-десатуразу жирных кислот
Для того, чтобы подтвердить, что последовательность L11 кодировала фермент - Δ5-десатуразу жирных кислот, кДНК субклонировали в дрожжевом экспрессирующем векторе pYES2, предоставленном Invitrogen™, под контролем промотора GAL4-полимеразы для выхода плазмиды pYES2/L11. Экспрессию L11 проверяли путем транскрипции-трансляции pYES2/L11 in vitro, используя сопряженную систему транскрипции и трансляции Promega™. Получали продукты трансляции, меченные 35S-метионином, которые разделяли электрофорезом в ПААГ (полиакриламидном геле) в присутствии SDS (додецилсульфата натрия) и визуализировали путем экспонирования с авторадиографической пленкой. Подсчитанная молекулярная масса продукта составила 55-60 кДа, а контрольная плазмида без вставки, pYES2, не способна давать какого-либо меченного продукта трансляции.
Конструкцию pYES2/L11 трансформировали в дрожжи, Saccharomyces cerevisiae, и выращивали на среде YCA, обедненной урацилом. Трансформантов отбирали благодаря присутствию селектируемого маркера URA3, который несет pYES2/L11, а экспрессию L11 индуцировали добавлением галактозы до конечной концентрации 1% мМ. Культуры выращивали в течение ночи в присутствии 0,5 мМ дигомо-γ-линолената, детергента (1%-ного тергитола, NP-40) и 2%-ной рафинозы. Аликвотные пробы отбирали в моменты времени t=0, t=4 часа и t=16 часов. Все жирные кислоты дрожжей анализировали посредством ГХ (газовой хроматографии) метиловых эфиров. Липиды из индуцированных и неиндуцированных контрольных проб трансметилировали 1М HCl в метаноле при 80°С в течение 1 часа. Метиловые эфиры жирных кислот (МЭЖК) экстрагировали в гексане. Анализ с помощью ГХ МЭЖК проводили, используя газовый хроматограф Hewlett Packard серии 58804, оснащенный присоединенной капиллярной колонкой 25 М × 0,32 мм RSL-500ВР и пламенно-ионизационным детектором.
Когда метиловые эфиры всех жирных кислот, выделенных из дрожжей, несущих плазмиду pYes2/L11 и выращенных в присутствии галактозы и дигомо-γ-линоленовой кислоты, проанализировали посредством ГХ, обнаружили дополнительный пик (см. Фиг.3). Этот дополнительный пик имел такое же время удерживания, как стандарт аутентичной арахидоновой кислоты (Sigma), показывая, что трансгенные дрожжи способны к десатурированию дигомо-γ-линоленовой кислоты в Δ5-положении. Таких пиков не было обнаружено в контрольных пробах (трансформация pYes2), Фиг.3. Идентичность дополнительного пика подтвердили посредством ГХ-МС (газовая хроматография-масс-спектрометрия) (Kratos MC80RFA, работающий при напряжении ионизации 70 эВ с диапазоном сканирования 500-40 дальтон), которая точно идентифицировала это соединение, как арахидоновую кислоту.
Это демонстрирует, что последовательность ДНК из Mortierella alpina кодирует функционально активный полипептид, вовлеченный в синтез арахидоновой кислоты в присутствии галактозы и дигомо-γ-линоленовой кислоты.
Клонирование и секвенирование гена Δ5-десатуразы жирных кислот C.elegans
Предварительно авторы изобретения идентифицировали грибные Δ5- и Δ6-десатуразы жирных кислот, которые отличались от предварительно идентифицированных микросомальных десатураз как из растительных, так и из животных видов. Это отличие было вызвано присутствием N-концевого удлиняющего сегмента, который демонстрировал гомологию с электронодонорным белком-цитохромом b5.
Во время характеристики грибной Δ5-десатуразы жирных кислот Mortierella alpina и Δ6-десатуразы жирных кислот С. afegans (находится в космиде W08D2 (регистрационный номер Z70271)), авторы изобретения идентифицировали родственную последовательность в космиде T13F2.1 (регистрационный номер Z81122), также содержащей ДНК С.elegans, которая, вероятно, кодирует десатуразу жирных кислот.
Анализ последовательностей с использованием программы Genefinder (Wilson, R. et al (1994) Nature, 368, 32-38)) показал, что космиды W08D2 и T13F2 содержат перекрывающиеся участки. Вместе с тем, было установлено, что космида T13F2, содержит открытую рамку считывания (ОРС), обозначенную T13F2.1, которая содержит N-концевой домен цитохрома b5 (определенный диагностическим фрагментом His-Pro-Gly-Gly), а также три гистидиновых бокса - характерную черту всех микросомальных десатураз. Кроме того, эта предполагаемая десатураза содержала вариантный третий гистидиновый бокс с Н→Q заменой вместо первого гистидина в фрагменте His-X-X-His-His. Эта глутаматная замена присутствует как в растительных, так и в животных Δ6-десатуразах жирных кислот, и в грибной Δ5-десатуразе жирных кислот из М.alpina.
Перекрывание между космидами T13F2 и W08D2 сделало возможным установление пространственной близости ОРС предполагаемой десатуразы T13F2.1 с Δ6-десатуразой жирных кислот, показывая, что две последовательности расположены тандемно на IV хромосоме, отделены 990 основаниями от предсказанного стоп-кодона T13F2.1 до инициирующего метионинового триплета Δ6-десатуразы жирных кислот.
Поскольку анализ последовательности предсказал, что ОРС T13F2.1 разделена рядом интронов, гетерологичная функциональная экспрессия геномной ДНК невозможна. Поэтому полимеразную цепную реакцию использовали для амплификации клона, содержащего неполную кДНК, соответствующую большому предсказанному экзону на 5'-конце ОРС T13F2.1 с применением следующих праймеров: СЕ прямого и СЕ обратного
СЕ прямой
5' - ATGGTATTACGAGAGCAAGA-3'
СЕ обратный
5' -TCTGGGATCTCTGGTTCTTG-3'
После начальной денатурации при 94°С в течение 2 минут проводили амплификацию в 32 цикла: 45 секунд при 94°С, 1 минута при 55°С и 1 минута при 72°С, с последующей заключительной элонгацией при 72°С в течение дополнительных 10 минут.
Фрагмент ДНК правильно предсказанного размера (который визуализировали в 1%-ном агарозном геле) амплифицировали, бэнд на геле вырезали, ДНК очищали и лигировали непосредственно с pGEM-T (Promega), и полученную в результате плазмиду трансформировали в клетки E.coli DH 5α. Плазмидную ДНК очищали для секвенирования, используя набор Qiagen QIAprep miniprep, а нуклеотидную последовательность вставки определяли путем автоматизированного секвенирования, используя ДНК-секвенатор ABI-377.
Для того, чтобы выделить полный кодирующий участок, соответствующий OPC T13F2.1, этот выделенный ПЦР-амплифицированный фрагмент длиной 233 п.н. использовали для скрининга библиотеки кДНК смешанной стадии С.elegans, которую сконструировали в λiZapll Prof. Yuji Kohara-Mishima, Япония. Скрининг проводили, используя стандартные методы (Sambrook et al (1989) Molecular Cloning. A Laboratory Manual), с применением в качестве зонда клонированный продукт ПЦР. Фрагменты ДНК метили радиоактивным изотопом α[32Р] дЦТФ (дезоксицитидинтрифосфат), используя готовую реакционную смесь для мечения ДНК (Pharmacia). Из 1,4×105 бляшкообразующих единиц, проверенных на гибридизацию с фрагментом длиной 233 п.н. 5 бляшек дали положительные ответы и были вырезаны из чашек с агаром и элюированы в SM-буфер. Полученные в результате фаговые суспензии проверяли на присутствие T13F2.1 путем ПЦР-амплификации, используя СЕ прямой и СЕ обратный. Один клон, обозначенный L4, очищали 2 дополнительными циклами культивирования и скрининга путем гибридизации при 65°С, используя фрагмент длиной 233 п.н., выделенный посредством ПЦР. Плазмиду L4 выделяли из λ-клона L4 путем эксцизии и секвенировали обе цепи кДНК-вставки, используя ДНК-секвенатор ABI-377 Perkin-Elmer.
Полученная в результате последовательность ДНК представлена в SEQ.2, и предсказанная аминокислотная последовательность представлена в SEQ.4.
Функциональный анализ L4 в дрожжах
Полный кодирующий участок L4 (кодирующий 447 аминокислот) амплифицировали путем ПЦР, используя показанные ниже праймеры YCED прямой и YCED обратный, которые также включали фланкирующие HindIII- и BamHI-сайты рестрикции:
YCED прямой
5'-GCGAAGCTTAAAATGGTATTACGAGAGCAAGAGC-3'
(отжиг с инициирующим метионином показан жирным шрифтом, сайт рестрикции HindIII подчеркнут)
YCED обратный
5'-GCGGGATCCAATCTAGGCAATCTTTTTAGTCAA-3'
(отжиг с комплементом стоп-кодона показан жирным шрифтом, сайт рестрикции BamHI подчеркнут).
Амплифицированный продукт ПЦР, содержащий полный кодирующий участок L4, лигировали с дрожжевым экспрессирующим вектором pYES2 (Invitrogen), вниз от промотора GAL1, используя сайты рестрикции HindIII и BamHI (ферменты предоставлены Boehringer Mannheim). Полученную в результате конструкцию, обозначенную pYES2/L4, трансформировали в клетки E.coli и правильность полученной с помощью ПЦР вставки в плазмиду pYES2/L4 подтвердили in vitro посредством сопряженной транскрипции-трансляции, используя TNT-систему (Promega). Полученные в результате продукты трансляции метили 35S-метионином, разделяли электрофорезом в ПААГ в присутствии SDS и визуализировали посредством авторадиографии.
Продукт трансляции, полученный с pYES2/L4, имел молекулярную массу приблизительно Мr=57 000, тогда как контрольный вектор, pYES2 без вставки, не давал продукта трансляции.
Для функционального анализа кодирующего участка L4 рекомбинантную плазмиду трансформировали в клетки S.cerevisiae DBY746 методом обработки ацетатом лития (Elble R. (1992) Bio Techniques 13 18-20). Клетки культивировали в течение ночи в среде, содержащей рафинозу в качестве источника углерода, обогащенную добавлением либо линолевой кислоты (18:2Δ9,12), либо дигомо-γ-линоленовой кислоты (С20:33Δ8,11,14) в присутствии 1%-ного тергитола (как описано Napier et al (1998) Biochem. J. 330 611-614). Эти жирные кислоты отсутствуют у S.cerewsiae, но служат в качестве специфических субстратов либо для Δ6-, либо для Δ5-десатуразы, соответственно. Экспрессию кодирующего участка L4 с промотора GAL1 вектора индуцировали добавлением галактозы до 1%. Выращивание культур продолжалось в течение 16 часов до отбора аликвотных проб для анализа жирных кислот посредством ГХ. Все жирные кислоты, экстрагированные из дрожжевых культур, анализировали посредством газовой хроматографии (ГХ) метиловых эфиров. Липиды трансметилировали 1М HCl в метаноле при 80°С в течение 1 часа, затем метиловые эфиры жирных кислот (МЭЖК) экстрагировали в гексане. Анализ с помощью ГХ МЭЖК проводили, используя газовый хроматограф Hewlett Packard серии 5880А, оснащенный присоединенной капиллярной колонкой 25 М × 0,32 мм RSL-500 ВР и пламенно-ионизационным детектором. Жирные кислоты идентифицировали путем сравнения с временем удерживания стандартов МЭЖК (Sigma). Относительное процентное содержание жирных кислот подсчитывали исходя из площадей под пиками. Арахидоновую кислоту идентифицировали посредством ГХ-МС, используя Krats MC80RFA, работающий при напряжении ионизации 70 эВ с диапазоном сканирования 500-40 дальтон. Фиг.4 показывает результат анализа с помощью ГХ метиловых эфиров жирных кислот трансформированных штаммов дрожжей. Дополнительный пик выявляется на развертке, полученной с индуцированной pYES2/L4, выращенной в присутствии дигомо-γ-линоленовой кислоты, по сравнению с контролем, содержащим пустой вектор. Этот пик также отсутствовал у неиндуцированных культур, выращенных на среде с дигомо-γ-линоленовой кислотой и, к тому же, важно отметить, что pYES2/L4, выращенная в присутствии линолевой кислоты, не способна накапливать какие-либо новые пики, показывая что эта жирная кислота не является субстратом для фермента, кодируемого кДНК С.elegans. Время удерживания дополнительного пика идентично времени удерживания аутентичного стандарта метил-арахидоновой кислоты. Жирную кислоту, полученную из дигомо-γ-линоленовой кислоты, дополнительно характеризовали посредством ГХ-МС (газовой хроматографии-масс-спектрометрии) и идентифицировали, как арахидоновую кислоту. Следовательно, результаты показывают, что трансформированные плазмидой pYES2/L4 дрожжевые клетки приобрели функциональную Δ5-десатуразную активность и были способны к синтезу арахидоновой кислоты из дигомо-γ-линоленовой кислоты, выступающей в качестве субстрата. Δ5-Десатураза в трансформированных дрожжах, по-видимому, являлась эффективным катализатором.
Это демонстрирует, что последовательность ДНК из С.elegans кодирует функционально активный полипептид, вовлеченный в синтез арахидоновой кислоты в присутствии галактозы и дигомо-γ-линолената.






Изобретение относится к области биотехнологии, в частности к генной инженерии, и может быть использовано в фармацевтической и пищевой промышленности для получения полиненасыщенных жирных кислот. Выделена и охарактеризована последовательность ДНК (1341 п.н.), кодирующая Δ5-десатуразу жирных кислот (447 аминокислотных остатков, 57 кДа) нематоды Caenorhabditis elegans. Путем экспрессии полученной ДНК - последовательности в бактериальных и дрожжевых клетках получена биологически активная рекомбинантная форма фермента, способная катализировать превращение дигомо-γ-линоленовой кислоты в арахидоновую кислоту и эйкозатетраеноата в эйкозапентаеноат. Применение изобретения обеспечивает возможность масштабирования процесса получения полиненасыщенных жирных кислот. 10 с. и 5 з.п. ф-лы, 4 ил.
| R.WILSONRET A.L | |||
| Nature | |||
| Полу генеративная топка для сжигания влажного торфа | 1921 |
|
SU368A1 |
| US 5057419, 15.10.1991 | |||
| DEBORAH L | |||
| et al | |||
| Biochemistry | |||
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |

